
A census of RND superfamily proteins in the Burkholderia genus

R E S E A R C H A R T I C L E
Comparative invivoand invitro analysesof putativevirulencefactorsofBurkholderia pseudomallei using lipopolysaccharide,capsuleand£agellinmutantsChanthiwa Wikraiphat1, Jaruek Charoensap1, Pongsak Utaisincharoen1, Surasak Wongratanacheewin2,Suwimol Taweechaisupapong3, Donald E. Woods4, Jan G.M. Bolscher5 & Stitaya Sirisinha1
1Department of Microbiology, Faculty of Science, Mahidol University, Bangkok, Thailand; 2Department of Microbiology, Faculty of Medicine, Khon Kaen
University, Khon Kaen, Thailand; 3Department of Oral Diagnosis, Faculty of Dentistry, Khon Kaen University, Khon Kaen, Thailand; 4Department of
Microbiology and Infectious Diseases, University of Calgary Health Sciences Center, Calgary, Alberta, Canada; and 5Department of Oral Biochemistry,
Academic Centre for Dentistry, VU Medical Center, Amsterdam, The Netherlands
Correspondence: Stitaya Sirisinha,
Department of Microbiology, Faculty of
Science, Mahidol University, Bangkok,
Thailand. Tel.: 1662 201 5675; fax: 1662
644 5411; e-mail: [email protected]
Received 19 February 2009; revised 14 May
2009; accepted 23 May 2009.
DOI:10.1111/j.1574-695X.2009.00574.x
Editor: Richard Marconi
Keywords
Burkholderia pseudomallei; polymorphonuclear
cell; macrophage; capsule; lipopolysaccharide;
flagella.
Abstract
Burkholderia pseudomallei is a gram-negative bacillus that is the causative agent of
melioidosis. We evaluated host–pathogen interaction at different levels using three
separate B. pseudomallei mutants generated by insertional inactivation. One of
these mutants is defective in the production of the polysaccharide side chains
associated with lipopolysaccharide; one does not produce the capsular polysac-
charide with the structure -3)-2-O-acetyl-6-deoxy-b-D-manno-heptopyranose-(1-;
and the third mutant does not produce flagellin. We compared the in vivo
virulence in BALB/c mice, the in vitro fate of intracellular survival inside human
polymorphonuclear cells (PMNs) and macrophages (Mfs) and the susceptibility
to killing by 30% normal human serum, reactive nitrogen and oxygen intermedi-
ates and antimicrobial peptides with that of their wild-type counterpart. The
lipopolysaccharide and capsule mutants demonstrated a marked reduction in
virulence for BALB/c mice, but the flagellin mutant was only slightly less virulent
than the parent strain. The results from the BALB/c mice experiments correlated
with survival in Mfs. The lipopolysaccharide and capsule mutants were also more
susceptible to killing by antimicrobial agents. All bacteria were equally susceptible
to killing by PMNs. Altogether, the data suggest that lipopolysaccharide and
capsule and, to a much lesser extent, flagella, are most likely associated with the
virulence of this bacterium and highlight the importance of intracellular killing by
PMNs and Mfs in disease pathogenesis.
Introduction
Burkholderia pseudomallei is a facultative gram-negative
bacillus that is the causative agent of a potentially fatal
disease known as melioidosis (White, 2003; Wiersinga et al.,
2006). Clinical manifestations vary from asymptomatic
infection to septic shock. The latter can have a mortality
rate as high as 80% despite appropriate antibiotic treatment.
The pathogenesis of melioidosis remains to be clarified,
but different lines of evidence suggest that innate immunity
is important in determining the outcome of infection
(Wiersinga et al., 2006). It was shown, for example, that
interferon-g (IFN-g) played an essential role in resistance in
a murine model of infection (Santanirand et al., 1999).
Enhancing IFN-g production before infection via CpG
oligodeoxynucleotide administration protected animals
against fatal challenge (Wongratanacheewin et al., 2004).
Macrophages (Mfs) preactivated with IFN-g could readily
kill B. pseudomallei (Miyagi et al., 1997). More recently,
the role of polymorphonuclear cells (PMNs) has attracted
the attention of many groups of investigators (Easton
et al., 2007; Wiersinga et al., 2007, 2008a, b), but attempts
to activate these cells with granulocyte colony-stimulating
factor in patients with severe sepsis due to melioidosis have
given rise to contradictory and questionable results (Cheng
et al., 2007).
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
FEMS Immunol Med Microbiol ]] (2009) 1–7 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
F E M S I M 5 7 4 B Dispatch: 8.6.09 Journal: FEMSIM CE: xx
Journal Name Manuscript No. Author Received: No. of pages: 7 Op: Chris/BinduFEMSIM 574(BW
UK
FE
MSI
M 5
74.P
DF
08-J
un-0
9 14
:5 2
5235
1 B
ytes
7 P
AG
ES
n op
erat
or=
jnm
.Chr
istin
a)

Although considerable attention has been paid to identi-
fying and characterizing the virulence factors of B. pseudo-
mallei, no definitive conclusions have been reached. Several
putative virulence factors have been implicated in disease
due to B. pseudomallei including lipopolysaccharide, the
capsular polysaccharide with the structure -3)-2-O-acetyl-6-
deoxy-b-D-manno-heptopyranose-(1- (hereafter referred to
as the mannoheptan capsule), flagella, the type III secretion
system, phospholipase C and several other toxins (Cheng &
Currie, 2005; Wiersinga et al., 2006). In the present study, we
used three B. pseudomallei mutants that we have developed
to compare their virulence in murine infection models, their
interaction with human PMNs and Mfs, as well as their
susceptibility to killing by antimicrobial agents in order to
obtain more informative data regarding virulence and
pathogenesis. The results suggest that lipopolysaccharide
and the mannoheptan capsule, but not flagella, are relevant
to the virulence of this bacterium.
Materials and methods
Bacterial cultivation and characterization
The lipopolysaccharide mutant SRM117 (DeShazer et al.,
1998), the mannoheptan capsule mutant SR1015 (mutated
in the wchB gene, which encodes for a protein with
homology to a glycosyltransferase, Reckseidler et al., 2001)
and flagellin mutant MM35 (DeShazer et al., 1997) were
generated from wild-type B. pseudomallei strain 1026b by
insertional mutagenesis. With regard to the insertional
mutagenesis, all the mutant strains contained a tetracycline
resistance cassette (Tcr) to indicate their insertional inacti-
vation. All strains were cultured at 37 1C in Luria–Bertani
(LB) medium for 16–18 h to obtain bacteria in the late
logarithmic phase of growth before harvesting and use in the
study. If not indicated otherwise, the medium for culturing
the mutants was always supplemented with tetracycline at a
concentration of 50mg mL�1 to prevent possible reversion
to the wild type. Under these conditions of growth, the
doubling times for all strains were calculated and found to
be c. 48 min. The number of viable bacteria, expressed as
CFU, was determined by serial dilution and the pour-plating
technique performed in duplicate using trypticase soy agar
(TSA) in the absence of an antibiotic. The presence or
absence of lipopolysaccharide, capsule or flagella in the
stock cultures (Table 1) was confirmed as described
(DeShazer et al., 1997; Anuntagool et al., 2000; Sirisinha
et al., 2000). Biofilm production was also determined (Tawee-
chaisupapong et al., 2005). Lethal dose 50% (LD50) determi-
nations in BALB/c mice for all four strains were performed as
described earlier (Wongratanacheewin et al., 2004).
M/ and PMN preparations
Mfs and PMNs were isolated from heparinized whole blood
of healthy human volunteers by Ficoll–Hypaque density
gradient separation (ethical clearance was provided by the
Ethical Committee of the Ramathibodi Hospital, Mahidol
University, Bangkok, Thailand, clearance number 2549/
452). In brief, peripheral blood mononuclear cells were
harvested from the fluid at the interface of the separation
tube, and monocytes were purified by positive selection of
CD141 cells using a magnetic cell sorter system (MACS@
Miltenyi Biotec, GmbH, Germany). The Mfs were gener-
ated from the purified monocytes by culturing the latter in
Iscove’s modified Dulbecco’s medium supplemented with
1% L-glutamine and 10% heat-inactivated human AB serum
(GemCellTM, West Sacramento, CA) at 37 1C for 5–7 days in
a humidified 5% CO2 incubator. The fluid in the lower layer
from the Ficoll–Hypaque separation tube was used for PMN
preparation (Watson et al., 1992). The PMNs were then
purified using 3% Dextran T500 (GE Healthcare, BioS-
ciences, Uppsala, Sweden). Residual contaminating red
blood cells were lysed in a hypotonic solution. The PMNs
were resuspended in DMEM culture medium supplemented
with 10% heat-inactivated fetal bovine serum (HyClone,
Logan, UT) and 1% L-glutamine and used immediately.
Survival and replication of B. pseudomallei wildtype and mutants in PMNs and M/s
PMNs (2� 106 cells) and Mfs (1� 105 cells) were infected
with the bacteria as described previously (Kespichayawatta-
na et al., 2000), using a multiplicity of infection of 2 : 1 and
1 : 1, respectively. Briefly, after 2 h of infection, extracellular
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
Table 1. Phenotypic characteristics of Burkholderia pseudomallei wild type and mutants
Bacterial
strains
LD50 (CFU)
(BALB/c, IP)
Biofilm formation
(OD630 nm) 200 kDa
Reactive
with mAb to
lipopolysaccharide Motile
Wild type 104 0.807 1 1 1
Capsule mutant 4 109 1.296 � 1 1
Lipopolysaccharide mutant 108 2.461 1 � 1
Flagellin mutant 3.16�104 0.756 1 1 �
1, produces capsule or lipopolysaccharide, motile; � , does not produce capsule or lipopolysaccharide, nonmotile; IP, intraperitoneal challenge.
FEMS Immunol Med Microbiol ]] (2009) 1–7c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
2 C. Wikraiphat et al.
FEMSIM 574(BW
UK
FE
MSI
M 5
74.P
DF
08-J
un-0
9 14
:5 2
5235
1 B
ytes
7 P
AG
ES
n op
erat
or=
jnm
.Chr
istin
a)

bacteria were killed by adding kanamycin to the cell culture
to a final concentration of 250 mg mL�1. Two hours later, the
antibiotic concentration was changed to 20 mg mL�1. In
addition to the kanamycin, for experiments using the
mutants, tetracycline at a concentration of 50mg mL�1 was
also present. The number of viable intracellular bacteria
at different time intervals was determined by lysing the
infected cells with 0.1% Triton X-100, and the number of
bacteria in the lysate was quantified as described above. At
the end of all experiments, the viability of the infected cells
was determined by trypan blue staining, and experiments
with viability o 90% were discarded. Data were analyzed
using a paired-samples t-test. P-values o 0.05 were consid-
ered statistically significant.
Host-cell free killing assays
The sensitivity of the three mutants and the wild type to
killing by 30% fresh pooled normal human serum (NHS)
and of human neutrophil peptide-2 (HNP-2) was deter-
mined as described, respectively, by DeShazer et al. (1998)
and West et al. (2005). In brief, 106 CFU mL�1 of late log-
phase bacteria were incubated in LB broth containing 30%
fresh or heat-inactivated pooled human serum (obtained
from donors living in a nonendemic area of infection),
10 mg mL�1 of HNP-2 or in phosphate-buffered saline for 2 h
before residual bacteria were determined. In addition to
HNP-2, the killing activities of other antimicrobial peptides,
namely, cathelicidin (LL-37), histatins (histatin5 and its
variant, dhvar5) and lactoferrin peptides (LFcin 17-30
and LFampin 268-284), were also investigated. These pep-
tides were synthesized as described (Den Hertog et al., 2004;
van der Kraan et al., 2004). Bacteria (1� 107 CFU mL�1) in
1 mM potassium phosphate buffer (PPB), pH 7.0, were
added to equal volumes of the antimicrobial peptide solu-
tions. The final concentration of all the peptides was
100 mM. A bacterial suspension in PPB without a peptide
served as a control. After incubation at 37 1C for 1 h, the
number of residual bacteria was determined and the percen-
tage of killing was calculated using the formula [1� (CFU
sample/CFU control)]� 100%. Differences in susceptibility
to each antimicrobial peptide of each mutant compared
with that of the wild type were analyzed using Student’s t-
test. Statistical significance was considered when the P-value
was o 0.5.
Susceptibility to killing by reactive nitrogen(RNI) and oxygen (ROI) intermediates
The sensitivity of the bacteria to RNI killing was assessed by
culturing them in different concentrations of nitric oxide
(NO)-generating compounds, S-nitrosoglutathione (GSNO)
or 3-morpholinosydnonimine hydrochloride (SIN-1) (Chak-
ravortty et al., 2002). After 4 h of incubation, residual bacteria
were determined by serial dilution and plating on TSA. The
concentrations that killed 50% of the bacteria were then
calculated and expressed as lethal concentration 50%. H2O2
was used to test their sensitivity to ROI. For this test, the
bacteria were similarly cultured in LB broth that contained
different concentrations of H2O2, and the number of residual
bacteria was determined after 2 h of incubation.
Results
Virulence of B. pseudomallei wild type andmutants for BALB/c mice
The LD50 values of the wild type and mutants in BALB/c
mice are shown in Table 1. The virulence of lipopolysacchar-
ide and capsule mutants was markedly reduced compared
with that of the wild-type counterpart or the flagellin
mutant. Judging from these values, the capsule appeared to
be the most important virulence factor in this animal model,
as its LD50 value decreased by 4 5 log10 compared with that
of the wild type, and this was followed by that of the
lipopolysaccharide mutant.
Intracellular survival and replication ofB. pseudomallei in human PMNs and M/s
The fates of B. pseudomallei wild type and the three mutants
after being internalized by human PMNs and Mfs were
followed for c. 16–18 h when the experiments were termi-
nated and host cells were still viable. Inside the PMNs, both
the wild type and the mutants displayed similar survival
profiles (Fig. 1). The absolute numbers of intracellular
bacteria, expressed as CFU of viable count, gradually
declined during these time intervals, and at the end of the
experiments there was a reduction of between 1 and 2 log10
of viable intracellular bacterial counts for all strains com-
pared with their initial 3-h time points. However, unlike the
results with the PMNs, the Mfs could not suppress the
growth of the B. pseudomallei wild type and at the end of the
experiment the absolute numbers of intracellular bacteria
increased by at least 1 log10. Its doubling time in the Mfs
was about 1 h 50 min, a value similar to that obtained with
other wild-type strains that we have tested (Charoensap
et al., 2009). The profile of the intracellular lipopolysacchar-
ide mutant, on the other hand, was markedly different from
that of the wild type, and by the end of the experiment the
residual numbers of the lipopolysaccharide mutant were
significantly lower than those of the wild type by 4 2 log10
(P = 0.036). Hence, the oligosaccharide component of the
lipopolysaccharide appeared to be one of the components of
B. pseudomallei that conferred resistance against intracellu-
lar killing by human Mfs.
It is apparent from Fig. 1b that the numbers of the capsule
mutant at the 16-h time point were markedly different from
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
FEMS Immunol Med Microbiol ]] (2009) 1–7 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
3Analysis of putative virulence factors of B. pseudomallei
FEMSIM 574(BW
UK
FE
MSI
M 5
74.P
DF
08-J
un-0
9 14
:5 2
5235
1 B
ytes
7 P
AG
ES
n op
erat
or=
jnm
.Chr
istin
a)
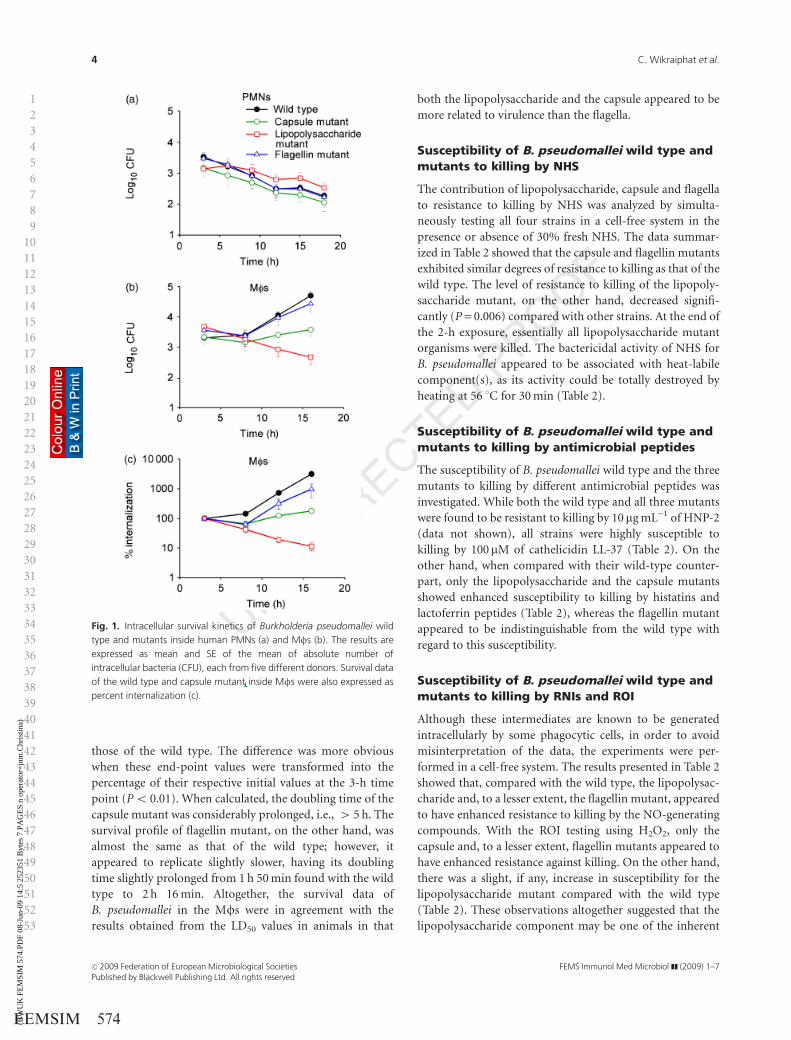
those of the wild type. The difference was more obvious
when these end-point values were transformed into the
percentage of their respective initial values at the 3-h time
point (Po 0.01). When calculated, the doubling time of the
capsule mutant was considerably prolonged, i.e., 4 5 h. The
survival profile of flagellin mutant, on the other hand, was
almost the same as that of the wild type; however, it
appeared to replicate slightly slower, having its doubling
time slightly prolonged from 1 h 50 min found with the wild
type to 2 h 16 min. Altogether, the survival data of
B. pseudomallei in the Mfs were in agreement with the
results obtained from the LD50 values in animals in that
both the lipopolysaccharide and the capsule appeared to be
more related to virulence than the flagella.
Susceptibility of B. pseudomallei wild type andmutants to killing by NHS
The contribution of lipopolysaccharide, capsule and flagella
to resistance to killing by NHS was analyzed by simulta-
neously testing all four strains in a cell-free system in the
presence or absence of 30% fresh NHS. The data summar-
ized in Table 2 showed that the capsule and flagellin mutants
exhibited similar degrees of resistance to killing as that of the
wild type. The level of resistance to killing of the lipopoly-
saccharide mutant, on the other hand, decreased signifi-
cantly (P = 0.006) compared with other strains. At the end of
the 2-h exposure, essentially all lipopolysaccharide mutant
organisms were killed. The bactericidal activity of NHS for
B. pseudomallei appeared to be associated with heat-labile
component(s), as its activity could be totally destroyed by
heating at 56 1C for 30 min (Table 2).
Susceptibility of B. pseudomallei wild type andmutants to killing by antimicrobial peptides
The susceptibility of B. pseudomallei wild type and the three
mutants to killing by different antimicrobial peptides was
investigated. While both the wild type and all three mutants
were found to be resistant to killing by 10 mg mL�1 of HNP-2
(data not shown), all strains were highly susceptible to
killing by 100 mM of cathelicidin LL-37 (Table 2). On the
other hand, when compared with their wild-type counter-
part, only the lipopolysaccharide and the capsule mutants
showed enhanced susceptibility to killing by histatins and
lactoferrin peptides (Table 2), whereas the flagellin mutant
appeared to be indistinguishable from the wild type with
regard to this susceptibility.
Susceptibility of B. pseudomallei wild type andmutants to killing by RNIs and ROI
Although these intermediates are known to be generated
intracellularly by some phagocytic cells, in order to avoid
misinterpretation of the data, the experiments were per-
formed in a cell-free system. The results presented in Table 2
showed that, compared with the wild type, the lipopolysac-
charide and, to a lesser extent, the flagellin mutant, appeared
to have enhanced resistance to killing by the NO-generating
compounds. With the ROI testing using H2O2, only the
capsule and, to a lesser extent, flagellin mutants appeared to
have enhanced resistance against killing. On the other hand,
there was a slight, if any, increase in susceptibility for the
lipopolysaccharide mutant compared with the wild type
(Table 2). These observations altogether suggested that the
lipopolysaccharide component may be one of the inherent
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
Fig. 1. Intracellular survival kinetics of Burkholderia pseudomallei wild
type and mutants inside human PMNs (a) and Mfs (b). The results are
expressed as mean and SE of the mean of absolute number of
intracellular bacteria (CFU), each from five different donors. Survival data
of the wild type and capsule mutant inside Mfs were also expressed as
percent internalization (c).
FEMS Immunol Med Microbiol ]] (2009) 1–7c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
4 C. Wikraiphat et al.
FEMSIM 574(BW
UK
FE
MSI
M 5
74.P
DF
08-J
un-0
9 14
:5 2
5235
1 B
ytes
7 P
AG
ES
n op
erat
or=
jnm
.Chr
istin
a)

properties associated with the resistance and susceptibility
of B. pseudomallei to killing by NOI and ROI and, as a
consequence, may influence the outcome of the organism’s
intracellular fate in the phagocytes. The capsule and, to a
lesser extent, the flagellin mutants appeared to be more
resistant to ROI killing than the wild type. The latter also
had a considerable delay in stimulating respiratory burst in
human PMNs compared with the wild type and the other
two mutants, judging from a chemiluminescence assay using
200 mM luminol (data not shown).
Discussion
In the present study, we attempted to provide additional
evidence to further identify and verify the possible role of
three putative virulence factors, namely, lipopolysaccharide,
capsule and flagella, in disease caused by B. pseudomallei.
This was done by comparative analyses of the three respec-
tive mutants with their wild-type counterpart in inducing
disease in susceptible BALB/c mice, their intracellular fates
in human PMNs and Mfs and their susceptibility to killing
by serum, antimicrobial peptides, ROI and RNIs. The results
from our studies strongly suggest that the lipopolysacchar-
ide and capsule, and to a much lesser extent flagella, are
associated with the virulence of this bacterium. Moreover,
because both the lipopolysaccharide and the capsule mu-
tants demonstrated noticeably higher biofilm production
than either the flagellin mutant or the wild type (Table 1), it
appears that biofilms do not play a significant role in the
disease process. This conclusion is consistent with our
previous report using a B. pseudomallei wild type possessing
different biofilm contents and a biofilm mutant (Taweechai-
supapong et al., 2005).
The observation that the lipopolysaccharide represented
one of the virulence components of B. pseudomallei is not
really unexpected (Table 1 and Fig. 1) because it was
suggested previously by our own group (DeShazer et al.,
1997; Archaroen et al., 2007 Q1). In one of these reports, we
showed that compared with the wild type, this same
lipopolysaccharide mutant was more susceptible to killing
during the early phase of infection by a mouse Mf cell line
RAW264.7 and the data obtained suggested that this may be
related to its ability to activate IFN-b and iNOS, both of
which are known to upregulate NO production that killed
this intracellular bacterium (Arjcharoen et al., 2007). It
could be argued, however, that in that report a mouse cell
line was used and so the conclusion must be interpreted
with caution. Therefore, in the present study, we similarly
tested host–bacterial interactions using freshly isolated
human phagocytic cells infected with B. pseudomallei wild
type and three different mutants. With the human system,
we found again that the lipopolysaccharide mutant was
more susceptible to killing by the Mfs than its wild-type
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53 Tab
le2.
Leth
alco
nce
ntr
atio
n50%
(LC
50)va
lues
and
susc
eptibili
tyle
vels
(with�
SD)of
Burk
hold
eria
pse
udom
alle
iwild
type
and
muta
nts
toki
lling
by
seru
man
ddiffe
rent
antim
icro
bia
lagen
ts
Bac
teria
Res
idual
bac
teria
(CFU
)%
Kill
ed(n
=6)
LC50
30%
NH
S
30%
hea
t-in
activa
ted
NH
SPB
S
His
tatins
(100mM
)La
ctofe
rrin
s(1
00mM
)C
ather
icid
in
(100mM
)RN
Is(m
M)
RO
I(mM
)
(n=
4)
His
tatin
5D
hva
r5LF
cin
17-3
0
Lfam
pin
268-2
84
LL-3
7SI
N-1
GSN
OH
2O
2
Wild
type
1.1
4�
10
61.2
3�
10
61.5
8�
10
658.0�
6.9
39.3�
9.2
25.2�
7.9
20.7�
18.9
99.9�
0.1
00.6
52.7
536�
6
Cap
sule
muta
nt
1.1
1�
10
61.1
8�
10
61.4
5�
10
678.1�
6.1�
74.7�
5.9�
71.1�
9.4�
80.5�
7.5�
99.5�
0.0
6�
0.5
52.7
273�
14
Lipopoly
sacc
har
ide
muta
nt
41
1.2
2�
10
61.5
3�
10
658.1�
9.7
80.0�
9.5�
60.2�
18.1�
57.8�
19.9�
99.9�
0.0
33.0
53.5
22�
3
Flag
ellin
muta
nt
1.4
5�
10
61.1
9�
10
61.8
1�
10
640.7�
21.8
51.9�
11.1
38.9�
13.8
16.8�
19.3
99.7�
1.0
21.8
53.6
555�
13
� Po
0.0
1co
mpar
edw
ith
stra
in1026b.
PBS,
phosp
hat
e-buff
ered
salin
e.
FEMS Immunol Med Microbiol ]] (2009) 1–7 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
5Analysis of putative virulence factors of B. pseudomallei
FEMSIM 574(BW
UK
FE
MSI
M 5
74.P
DF
08-J
un-0
9 14
:5 2
5235
1 B
ytes
7 P
AG
ES
n op
erat
or=
jnm
.Chr
istin
a)

counterpart during the entire period of observation (Fig. 1).
We would like to suggest that the decreased virulence of the
lipopolysaccharide mutant in our in vivo mouse model
(Table 1) is a combined result of their intracellular fates in
Mfs, together with enhanced susceptibility to killing by
serum, antimicrobial agents and, to a lesser extent, by ROI
(Table 2). The data presented also allow us to conclude that
the capsule is another important virulence factor for this
bacterium. Its ability to confer some degree of resistance
against intracellular killing by human Mfs in particular
sheds additional light on its possible mechanism of patho-
genicity. The association of the capsule with virulence had
been reported previously in another animal model (Reck-
seidler et al., 2001), but it was then suggested that this is
probably attributable to the fact that the capsule is known to
interfere with complement activation and subsequently
phagocytosis of B. pseudomallei (Reckseidler et al., 2005).
In addition to these possible mechanisms, our data in Fig. 1,
showing that the capsule mutant survived rather poorly in
the Mfs when compared with the wild type, provide
another possible mechanism of resistance against killing. It
should be mentioned that the slight increase in the number
of viable capsule-mutant organisms in the lysate at the later
time points (Fig. 1) was unlikely to be caused by a premature
release of bacteria from dying infected cells into the super-
natant fluids because the viability of these infected cells was
still higher than 90% when the experiment was terminated.
The ability of the capsule to confer enhanced resistance
against intracellular killing may represent a novel mechan-
ism of capsule function, and this phenomenon has never
been reported before for B. pseudomallei. This observation
is, however, in accordance with the results reported recently
for Neisseria meningitidis (Spinosa et al., 2007). It was
demonstrated further in that model that the presence of the
capsule correlated with its enhanced resistance to killing by
defensins, cathelicidins, protegerins and polymyxin B, a
finding that is also similar to our data on the susceptibility
of Burkholderia to killing by the antimicrobial peptides used
in our study (Table 2). Moreover, the enhanced resistance of
the capsule mutant to ROI killing as noted in the table may
contribute to its prolonged survival in the Mfs. The
importance of Mfs in host defense against B. pseudomallei
infection was convincingly demonstrated with the data
showing that Mf depletion by dichloromethylene bipho-
sphonate-containing liposomes rendered mice more suscep-
tible to B. pseudomalleiQ2 infection (Breitbach et al., 2006).
Consistent with the animal study reported earlier (Miyagi
et al., 1997), we have recently demonstrated in the human
system that although B. pseudomallei was able to survive
intracellular killing inside the Mfs, the killing capacity
could be readily enhanced after activation by IFN-g (Char-
oensap et al., 2009). On the other hand, the role of PMNs
should not be overlooked as it was shown earlier that they
were needed in defense against B. pseudomallei infection in a
mouse model (Easton et al., 2007; Wiersinga et al., 2008a, b).
Our results (Fig. 1), demonstrating the efficacy of PMNs in
killing B. pseudomallei in vitro, supported its potential role
in host defense against B. pseudomallei infection.
Lastly, it should be mentioned that the role of flagella is
yet to be established. For example, although we recently
reported that flagella were needed for the invasion of
B. pseudomallei into nonphagocytic cells (Chuaygud et al.,
2008), it is inconsistent with the data in the present study
using primary human phagocytic cell cultures. It is therefore
possible to attribute the difference to the types of cells used
in the two studies. Our in vivo data using flagellin mutant
bacteria in BALB/c mice (Table 1) were also different from
what was reported earlier by Chua et al. (2003). The
discrepancy between our results and those of the previous
results may be explained by a difference in the route of
bacterial challenge. In their study, they clearly demonstrated
that flagella were a virulence determinant as their flagellin-
defective mutant was avirulent for BALB/c mice when
deposited intranasally. When challenged by the intraperito-
neal route, however, as shown in the current studies, flagella
are not required for virulence. In summary, while we
showed here, using in vitro and in vivo experiments,
that lipopolysaccharide and the capsular polysaccharide
with the structure -3)-2-O-acetyl-6-deoxy-b-D-manno-
heptopyranose-(1- are definitely important virulence factors
for B. pseudomallei, the role of flagellin remains unsettled
and needs to be investigated further.
Acknowledgements
This work was supported by grants from the National
Science and Technology Development Agency (Thailand),
the Royal Golden Jubilee PhD Program of the Thailand
Research Fund, the Institutional Strengthening Program,
Faculty of Science, and the RA Scholarship, Faculty of
Graduated Studies, Mahidol University.
References
Anuntagool N, Naigowit P, Wuthiekanun V, White NJ & Sirisinha
S (2000) Monoclonal antibody-based rapid identification of
Burkholderia pseudomallei in blood culture fluid from patients
with community-acquired septicemia. J Med Microbiol 49:
1075–1078.
Arjcharoen S, Wikraiphat C, Pudla M, Limposuwan K, Woods
DE, Sirisinha S & Utaisincharoen P (2007) Fate of a
Burkholderia pseudomallei lipopolysaccharide mutant in the
mouse macrophage cell line RAW 264.7: possible role for the
O-antigenic polysaccharide moiety of lipopolysaccharide in
internalization and intracellular survival. Infect Immun 75:
4298–4304.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
FEMS Immunol Med Microbiol ]] (2009) 1–7c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
6 C. Wikraiphat et al.
FEMSIM 574(BW
UK
FE
MSI
M 5
74.P
DF
08-J
un-0
9 14
:5 2
5235
1 B
ytes
7 P
AG
ES
n op
erat
or=
jnm
.Chr
istin
a)

Breitbach K, Klocke S, Tschernig T, van Rooijen N, Baumann U &
Steinmetz I (2006) Role of inducible nitric oxide synthase and
NADPH oxidase in early control of Burkholderia pseudomallei
infection in mice. Infect Immun 74: 6300–6309.
Chakravortty D, Hansen-Wester I & Hensel M (2002) Salmonella
pathogenicity island 2 mediates protection of intracellular
Salmonella from reactive nitrogen intermediates. J Exp Med
195: 1155–1166.
Charoensap J, Utaisincharoen P, Engering A & Sirisinha S (2009)
Differential intracellular fate of Burkholderia pseudomallei
844 and Burkholderia thailandensis UE5 in human mono-
cyte-derived dendritic cells and macrophages. BMC Immunol
10: 20.
Cheng AC & Currie BJ (2005) Melioidosis: epidemiology,
pathophysiology, and management. Clin Microbiol Rev 18:
383–416.
Cheng AC, Limmathurotsakul D, Chierakul W et al. (2007) A
randomized controlled trial of granulocyte colony-stimulating
factor for the treatment of severe sepsis due to melioidosis in
Thailand. Clin Infect Dis 45: 308–314.
Chua KL, Chan YY & Gan YH (2003) Flagella are virulence
determinants of Burkholderia pseudomallei. Infect Immun 71:
1622–1629.
Chuaygud T, Tungpradabkul S, Sirisinha S, Chua KL &
Utaisincharoen P (2008) A role of Burkholderia pseudomallei
flagella as a virulent factor. Tran R Soc Trop Med Hyg 102
(suppl 1): S140–S144.
Den Hertog AL, Wong Fong Sang HW, Kraayenhof R, Bolscher
JG, Van’t Hof W, Veerman EC & Nieuw Amerongen AV (2004)
Interactions of histatin 5 and histatin 5-derived peptides with
liposome membranes: surface effects, translocation and
permeabilization. Biochem J 379: 665–672.
DeShazer D, Brett PJ, Carlyon R & Woods DE (1997) Mutagenesis
of Burkholderia pseudomallei with Tn5-OT182: isolation of
motility mutants and molecular characterization of the
flagellin structural gene. J Bacteriol 179: 2116–2125.
DeShazer D, Brett PJ & Woods DE (1998) The type II O-antigenic
polysaccharide moiety of Burkholderia pseudomallei
lipopolysaccharide is required for serum resistance and
virulence. Mol Microbiol 30: 1081–1100.
Easton A, Haque A, Chu K, Lukaszewski R & Bancroft GJ (2007)
A critical role for neutrophils in resistance to experimental
infection with Burkholderia pseudomallei. J Infect Dis 195:
99–107.
Kespichayawattana W, Rattanachetkul S, Wanun T,
Utaisincharoen P & Sirisinha S (2000) Burkholderia
pseudomallei induces cell fusion and actin-associated
membrane protrusion: a possible mechanism for cell-to-cell
spreading. Infect Immun 68: 5377–5384.
Miyagi K, Kawakami K & Saito A (1997) Role of reactive nitrogen
and oxygen intermediates in gamma interferon-stimulated
murine macrophage bactericidal activity against Burkholderia
pseudomallei. Infect Immun 65: 4108–4113.
Reckseidler SL, DeShazer D, Sokol PA & Woods DE (2001)
Detection of bacterial virulence genes by subtractive
hybridization: identification of capsular polysaccharide of
Burkholderia pseudomallei as a major virulence determinant.
Infect Immun 69: 34–44.
Reckseidler SL, DeVinney R & Woods DE (2005) The capsular
polysaccharide of Burkholderia pseudomallei contributes to
survival in serum by reducing complement factor C3b
deposition. Infect Immun 73: 1106–1115.
Santanirand P, Harley VS, Dance DA, Drasar BS & Bancroft GJ
(1999) Obligatory role of gamma interferon for host survival
in a murine model of infection with Burkholderia
pseudomallei. Infect Immun 67: 3593–3600.
Sirisinha S, Anuntagool N, Dharakul T, Ekpo P,
Wongratanacheewin S, Naikowit P, Petchclai B, Thamlikikul V
& Suputtamongkul Y (2000) Recent developments in
laboratory diagnosis of melioidosis. Acta Trop 74: 235–245.
Spinosa MR, Prodida C, Tala A, Cogli L, Alifano P & Bucci C
(2007) The Neisseria meningitidis capsule is important for
intracellular survival in human cells. Infect Immun 75:
3594–3603.
Taweechaisupapong S, Kaewpa C, Arunyanart C, Kanla P,
Homchampa P, Sirisinha S, Proungvitaya T &
Wongratanacheewin S (2005) Virulence of Burkholderia
pseudomallei does not correlate with biofilm formation.
Microb Pathogenesis 39: 77–85.
van der Kraan MI, Groenink J, Nazmi K, Veerman EC, Bolscher
JG & Nieuw Amerongen AV (2004) Lactoferrampin: a novel
antimicrobial peptide in the N1-domain of bovine lactoferrin.
Peptides 25: 177–183.
Watson F, Robinson JJ & Edwards SW (1992) Neutrophil
function in whole blood and after purification: changes in
receptor expression, oxidase activity and responsiveness to
cytokines. Bioscience Rep 12: 123–133.
West NP, Sansonetti P, Mounier J et al. (2005) Optimization of
virulence functions through glucosylation of Shigella LPS.
Science 307: 1313–1317.
White NJ (2003) Melioidosis. Lancet 361: 1715–1722.
Wiersinga WJ, van der Poll T, White NJ, Day NP & Peacock SJ
(2006) Melioidosis: insights into the pathogenicity of
Burkholderia pseudomallei. Nat Rev Microbiol 4: 272–282.
Wiersinga WJ, van’t Veer C, Wieland CW, Gibot S, Hooibrink B,
Day NP, Peacock S & van der Poll T (2007) Expression profile
and function of triggering receptor expressed on myeloid cells
(TREM)-1 in melioidosis. J Infect Dis 196: 1707–1716.
Wiersinga WJ, Wieland CW, Roelofs JJ & van der Poll T (2008a)
MyD88 dependent signaling contributes to protective host
defense against Burkholderia pseudomallei. PLoS ONE 3: e3494.
Wiersinga WJ, de Vos AF, de Beer R, Wieland CW, Roelofs JJ,
Woods DE & van der Poll T (2008b) Inflammatory patterns
induced by different Burkholderia species in mice. Cell
Microbiol 10: 81–87.
Wongratanacheewin S, Kespichayawattana W, Intachote P,
Pichyangkul S, Sermswan RW, Krieg AM & Sirisinha S (2004)
Immunostimulatory CpG oligodeoxynucleotide confers
protection in a murine model of infection with Burkholderia
pseudomallei. Infect Immun 72: 4494–4502.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
FEMS Immunol Med Microbiol ]] (2009) 1–7 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
7Analysis of putative virulence factors of B. pseudomallei
FEMSIM 574(BW
UK
FE
MSI
M 5
74.P
DF
08-J
un-0
9 14
:5 2
5235
1 B
ytes
7 P
AG
ES
n op
erat
or=
jnm
.Chr
istin
a)

Author Query Form
_______________________________________________________
_______________________________________________________
Dear Author,
During the copy-editing of your paper, the following queries arose. Please respond to these by marking up your proofs with the necessary changes/additions. Please write your answers clearly on the query sheet if there is insufficient space on the page proofs. If returning the proof by fax do not write too close to the paper's edge. Please remember that illegible mark-ups may delay publication.
Journal FEMSIMArticle 574
Query No. Description Author Response
Q1 Author: Archaroen et al. (2007) has not been included in the Reference List, please supply full publication details.
Q2
Author: Spelling of author name Breithbach has been changed to Breitbach to match the spelling in the Reference List for the reference Breitbach et al. (2006). Please confirm that this is correct.

Page 1 of 3
USING E-ANNOTATION TOOLS FOR ELECTRONIC PROOF CORRECTION Required Software Adobe Acrobat Professional or Acrobat Reader (version 7.0 or above) is required to e-annotate PDFs. Acrobat 8 Reader is a free download: http://www.adobe.com/products/acrobat/readstep2.html Once you have Acrobat Reader 8 on your PC and open the proof, you will see the Commenting Toolbar (if it does not appear automatically go to Tools>Commenting>Commenting Toolbar). The Commenting Toolbar looks like this:
If you experience problems annotating files in Adobe Acrobat Reader 9 then you may need to change a preference setting in order to edit. In the “Documents” category under “Edit – Preferences”, please select the category ‘Documents’ and change the setting “PDF/A mode:” to “Never”.
Note Tool — For making notes at specific points in the text Marks a point on the paper where a note or question needs to be addressed.
Replacement text tool — For deleting one word/section of text and replacing it Strikes red line through text and opens up a replacement text box.
Cross out text tool — For deleting text when there is nothing to replace selection Strikes through text in a red line.
How to use it: 1. Right click into area of either inserted
text or relevance to note 2. Select Add Note and a yellow speech
bubble symbol and text box will appear 3. Type comment into the text box 4. Click the X in the top right hand corner
of the note box to close.
How to use it: 1. Select cursor from toolbar 2. Highlight word or sentence 3. Right click 4. Select Replace Text (Comment) option 5. Type replacement text in blue box 6. Click outside of the blue box to close
How to use it: 1. Select cursor from toolbar 2. Highlight word or sentence 3. Right click 4. Select Cross Out Text

Page 2 of 3
Approved tool — For approving a proof and that no corrections at all are required.
Highlight tool — For highlighting selection that should be changed to bold or italic. Highlights text in yellow and opens up a text box.
Attach File Tool — For inserting large amounts of text or replacement figures as a files. Inserts symbol and speech bubble where a file has been inserted.
Pencil tool — For circling parts of figures or making freeform marks Creates freeform shapes with a pencil tool. Particularly with graphics within the proof it may be useful to use the Drawing Markups toolbar. These tools allow you to draw circles, lines and comment on these marks.
How to use it: 1. Click on the Stamp Tool in the toolbar 2. Select the Approved rubber stamp from
the ‘standard business’ selection 3. Click on the text where you want to rubber
stamp to appear (usually first page)
How to use it: 1. Select Highlighter Tool from the
commenting toolbar 2. Highlight the desired text 3. Add a note detailing the required change
How to use it: 1. Select Tools > Drawing Markups > Pencil Tool 2. Draw with the cursor 3. Multiple pieces of pencil annotation can be grouped together 4. Once finished, move the cursor over the shape until an arrowhead appears
and right click 5. Select Open Pop-Up Note and type in a details of required change 6. Click the X in the top right hand corner of the note box to close.
How to use it: 1. Click on paperclip icon in the commenting toolbar 2. Click where you want to insert the attachment 3. Select the saved file from your PC/network 4. Select appearance of icon (paperclip, graph, attachment or
tag) and close
Page 3 of 3
Help For further information on how to annotate proofs click on the Help button to activate a list of instructions:
Copyright © 2022 FDOKUMEN




















