
Comparative biochemistry of mouse and chick secondary-palate development in vivo and in vitro with...
15
Archs oral Bid. Vol. 36, No. 6, pp. 457471, 1991 0003-9969/91 $3.00 + 0.00 Printed in Great Britain. All rights reserved Copyright 0 1991 Pergamon Press plc COMPARATIVE BIOCHEMISTRY OF MOUSE AND CHICK SECONDARY-PALATE DEVELOPMENT IN VW0 AND IN VITRO WITH PARTICULAR EMPHASIS ON EXTRACELLULAR MATRIX MOLECULES AND THE EFFECTS OF GROWTH FACTORS ON THEIR SYNTHESIS DAVID M. FOREMAN, PAUL M. SHARPE and MARK W. J. FERGUSON* Animal and Human Reproduction, Development and Growth Research Group, Department of Cell and Structural Biology, University of Manchester, Coupland III Building, Manchester Ml3 9PL, U.K. (Accepted 8 January 1991) Sammary-A biochemical study analysing the wet weight, dry weight, water, protein and DNA content, collagen and GAG composition of all stages of the developing secondary palate in uiuo and in vitro was undertaken to investigate differences between a species in which the palatal shelves elevate (mouse) and one in which they do not (chick). The effects of EGF, bFGF, PDGF and TGF-/I, on collagen and GAG synthesis by cultured mouse and chick palatal shelves of different embryonic stages were also studied. The total GAG content of developing mouse palatal shelves decreased with developmental time; heparan sulphate proteoglycan formed the major species in early palates but hyaluronan was the major species in mid-late palates. There was a peak of hyaluronan synthesis in embryonic palatal shelves in vitro at day 13 (T.21), i.e. immediately before shelf elevation. By contrast the total GAG content of chick palates increased with development; chondroitin-6-sulphate formed the major GAG species and there was no peak in hyaluronan synthesis. The water content of developing murine palates rose rapidly at day 14 (T.22) i.e. the time of shelf elevation. No such peak was seen in the chick, where the water content rose exponentially with developmental time. Mouse palates synthesized chondroitin&sulphate and novel proteins around the time of shelf elevation; chick palates synthesized chondroitin-6-sulphate and no novel proteins at any developmental stage. Collagen synthesis also peaked in oitro in T.21 murine palates. EGF markedly stimulated murine palatal collagens and GAG synthesis between stages T.20-T.22, but had no effect thereafter. Basic FGF had similar but smaller stage-related effects. PDGF had no effect on mouse palatal collagen and GAG synthesis whilst TGF-8, inhibited GAG synthesis at T.21. The ratios of collagens I, III and V produced by mouse palates were unaltered by the growth factors. All the growth factors had no effect on chick palatal collagen synthesis at any stage and minimal effect on GAG synthesis; TGF-8, stimulated it in early but inhibited it in mid- to late-stage chick palates. These data indicate that extracellular matrix molecule metabolism within the palate is markedly different in the two species studied and suggest that the differing profiles of such molecules may be regulated at certain developmental stages by specific growth factors. Key words: palate development, extracellular matrix, growth factors. INTRODUCTION In all vertebrates the secondary palate arises as bilateral outgrowths from the maxillary processes. Mammalian palatal shelves initially grow vertically down the sides of the tongue [Plate Figs l(A) and (B)], but elevate at a precise time to a horizontal position above the dorsum of the tongue and fuse with each other [Plate Fig. l(C)] to form an intact secondary palate (Ferguson, 1988). Palatal shelf elevation is thought to be the result of an intrinsic shelf-elevating force chiefly generated by the pro- *To whom correspondence should be addressed. Abbreviations: bFGF, basic fibroblast growth factor; DMEM, Dulbecco’s minimal essential medium; EGF, epidermal growth factor; GAG, glycosaminoglycans; GuHCl, guanidinium hydrochloride; PDGF, platelet- derived growth factor; SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis; TGF-/I, , transforming growth factor-b,. gressive accumulation and hydration of hyaluronan, the force being directed along collagen fibres (Brinkley and Morris-Wiman, 1987). In birds, palatal shelves arise from the maxillary processes, but instead of growing vertically (as in mammals) they initially develop and grow horizontally above the dorsum of the tongue [Plate Figs l(D) and (E)]. The shelves contact, but do not fuse [Plate Fig. l(C)], so that birds have a naturally cleft palate (Koch and Smiley, 1981). These fundamental differences in palatogenesis between aves and mammals, and the apparent central role of extracellular matrix com- ponents in shelf elevation in mammals, prompted us to compare the biochemical changes leading to palate development in chick embryos with those of mouse embryos. Several studies have implicated EGF as playing a part in palatogenesis (Hassell, 1975; Tyler and Pratt, 1980) possibly via an effect on biosynthesis of extra- cellular matrix (Pratt et al., 1980; Silver, Murray and Pratt, 1984; Turley, Hollenberg and Pratt, 1985). AOB M/&-D 457
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Comparative biochemistry of mouse and chick secondary-palate development in vivo and in vitro with...

Archs oral Bid. Vol. 36, No. 6, pp. 457471, 1991 0003-9969/91 $3.00 + 0.00 Printed in Great Britain. All rights reserved Copyright 0 1991 Pergamon Press plc
COMPARATIVE BIOCHEMISTRY OF MOUSE AND CHICK SECONDARY-PALATE DEVELOPMENT IN VW0 AND
IN VITRO WITH PARTICULAR EMPHASIS ON EXTRACELLULAR MATRIX MOLECULES AND THE
EFFECTS OF GROWTH FACTORS ON THEIR SYNTHESIS
DAVID M. FOREMAN, PAUL M. SHARPE and MARK W. J. FERGUSON*
Animal and Human Reproduction, Development and Growth Research Group, Department of Cell and Structural Biology, University of Manchester, Coupland III Building, Manchester Ml3 9PL, U.K.
(Accepted 8 January 1991)
Sammary-A biochemical study analysing the wet weight, dry weight, water, protein and DNA content, collagen and GAG composition of all stages of the developing secondary palate in uiuo and in vitro was undertaken to investigate differences between a species in which the palatal shelves elevate (mouse) and one in which they do not (chick). The effects of EGF, bFGF, PDGF and TGF-/I, on collagen and GAG synthesis by cultured mouse and chick palatal shelves of different embryonic stages were also studied.
The total GAG content of developing mouse palatal shelves decreased with developmental time; heparan sulphate proteoglycan formed the major species in early palates but hyaluronan was the major species in mid-late palates. There was a peak of hyaluronan synthesis in embryonic palatal shelves in vitro at day 13 (T.21), i.e. immediately before shelf elevation. By contrast the total GAG content of chick palates increased with development; chondroitin-6-sulphate formed the major GAG species and there was no peak in hyaluronan synthesis. The water content of developing murine palates rose rapidly at day 14 (T.22) i.e. the time of shelf elevation. No such peak was seen in the chick, where the water content rose exponentially with developmental time. Mouse palates synthesized chondroitin&sulphate and novel proteins around the time of shelf elevation; chick palates synthesized chondroitin-6-sulphate and no novel proteins at any developmental stage. Collagen synthesis also peaked in oitro in T.21 murine palates.
EGF markedly stimulated murine palatal collagens and GAG synthesis between stages T.20-T.22, but had no effect thereafter. Basic FGF had similar but smaller stage-related effects. PDGF had no effect on mouse palatal collagen and GAG synthesis whilst TGF-8, inhibited GAG synthesis at T.21. The ratios of collagens I, III and V produced by mouse palates were unaltered by the growth factors. All the growth factors had no effect on chick palatal collagen synthesis at any stage and minimal effect on GAG synthesis; TGF-8, stimulated it in early but inhibited it in mid- to late-stage chick palates.
These data indicate that extracellular matrix molecule metabolism within the palate is markedly different in the two species studied and suggest that the differing profiles of such molecules may be regulated at certain developmental stages by specific growth factors.
Key words: palate development, extracellular matrix, growth factors.
INTRODUCTION
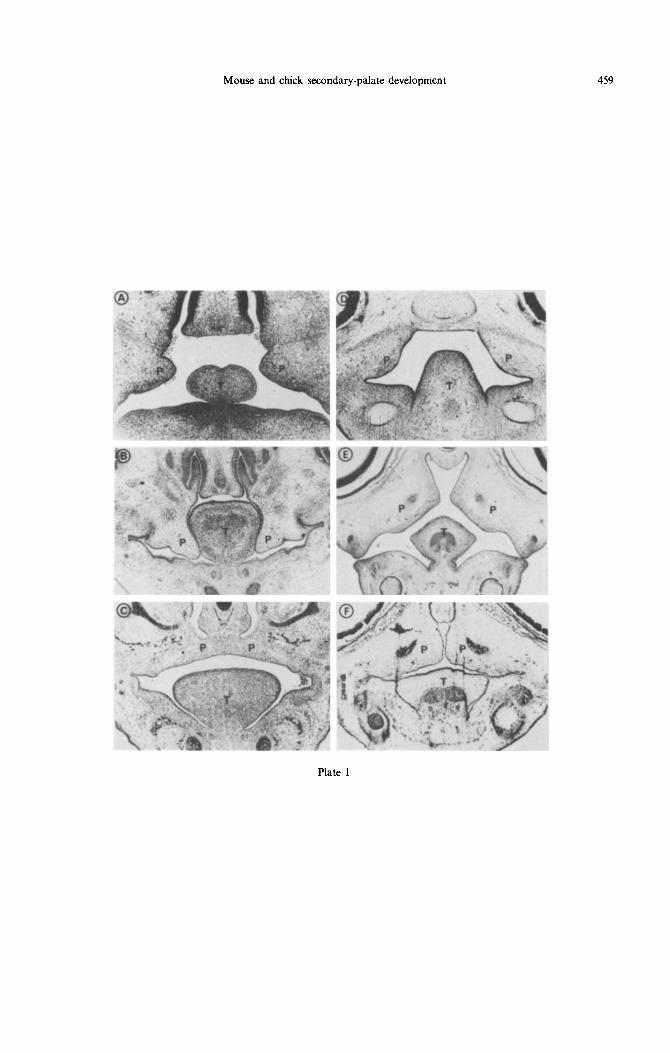
In all vertebrates the secondary palate arises as bilateral outgrowths from the maxillary processes. Mammalian palatal shelves initially grow vertically down the sides of the tongue [Plate Figs l(A) and (B)], but elevate at a precise time to a horizontal position above the dorsum of the tongue and fuse with each other [Plate Fig. l(C)] to form an intact secondary palate (Ferguson, 1988). Palatal shelf elevation is thought to be the result of an intrinsic shelf-elevating force chiefly generated by the pro-
*To whom correspondence should be addressed. Abbreviations: bFGF, basic fibroblast growth factor;
DMEM, Dulbecco’s minimal essential medium; EGF, epidermal growth factor; GAG, glycosaminoglycans; GuHCl, guanidinium hydrochloride; PDGF, platelet- derived growth factor; SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis; TGF-/I, , transforming growth factor-b,.
gressive accumulation and hydration of hyaluronan, the force being directed along collagen fibres (Brinkley and Morris-Wiman, 1987). In birds, palatal shelves arise from the maxillary processes, but instead of growing vertically (as in mammals) they initially develop and grow horizontally above the dorsum of the tongue [Plate Figs l(D) and (E)]. The shelves contact, but do not fuse [Plate Fig. l(C)], so that birds have a naturally cleft palate (Koch and Smiley, 1981). These fundamental differences in palatogenesis between aves and mammals, and the apparent central role of extracellular matrix com- ponents in shelf elevation in mammals, prompted us to compare the biochemical changes leading to palate development in chick embryos with those of mouse embryos.
Several studies have implicated EGF as playing a part in palatogenesis (Hassell, 1975; Tyler and Pratt, 1980) possibly via an effect on biosynthesis of extra- cellular matrix (Pratt et al., 1980; Silver, Murray and Pratt, 1984; Turley, Hollenberg and Pratt, 1985).
AOB M/&-D 457

458 DAVID M. FOREMAN et al.
Furthermore, other soluble growth factors have been identified as potent modulators of that biosynthesis (Narayanan and Page, 1983; Rizzino, 1988; Tseng et al., 1982). Consequently, we have examined the effects of a range of growth factors on the production of extracellular matrix by developing chick and mouse embryonic palates in organ culture.
The conventional system of palatal organ culture using Millipore filters on metal grids within Falcon culture dishes (Ferguson, Honig and Slavkin, 1984) was not used because preliminary experiments (data not shown) indicated that exogenous growth factors in the media had few effects upon the developing shelves under these conditions, possibly due to bind- ing to the Millipore filters used as the culture sub- stratum. Submerged cultures using a gelatin-coated, plastic substratum were used as an alternative. It has been shown previously that palatal shelves from mouse, chicken and alligator embryos undergo nor- mal differentiation in serum-free medium (Ferguson et al., 1984); however, this differentiation is delayed by approx. 24 h compared to the in uiuo timetable. Thus it is impossible to define precisely to which in uiuo stage our in vitro cultures correspond; for this reason all references to in vitro organ cultures refer to the developmental stage at time of explant, and the length of the culture period, to avoid confusion,
MATERIALS AND METHODS
Tissue source
Mice (Manchester strain) were mated overnight and the day of finding a vaginal plug designated day zero of embryonic development. On embryonic days 12, 13, 14 and 15 female mice were killed by an ether overdose and the embryos aseptically removed. Chick eggs were incubated and the embryos killed by decapitation on embryonic days 6-10. Murine and chick embryos were staged on the basis of external morphological features (Hamburger and Hamilton, 1951; Theiler, 1972) and, after transfer to Hanks balanced salt solution (Gibco), palates were dissected from Theiler (T) stage 20-23 (mouse) and Hamburger and Hamilton (H&H) stage 30-35 (chick) embryos plate Figs l(AHF)]. Each assay was done with 10 palates, in triplicate (from three separate litters in the case of the mouse), per developmental stage.
Wet weight, dry weight and water content
Palatal shelves were placed on pre-weighed plastic film (Parafilm) (routinely 10 palates/film), blotted to remove surface liquid and weighed on an analytical balance. The shelves plus plastic film were immedi- ately lyophilized and reweighed.
Protein and DNA content
Palatal shelves were homogenized in 2 M NaCl, 50 mM phosphate buffer, pH 7.4. Protease inhibitors were added to the final concentrations indicated: phenylmethanesulphonyl fluoride (2 mM), N-ethyl-
maleimide (10 mM), 6-aminohexanoic acid (25 mM) and EDTA (25 mM), and the extracts sonicated for 30 s. Subsamples were analysed for protein and DNA content.
Protein content was determined according to the method of Bradford (1976) using Coomassie protein assay reagent (Pierce) and bovine serum albumin to construct a standard curve. Proteins were analysed by SDS-PAGE according to the method of Laemmli (1970) using slab gels consisting of 6.5% separating gels and 3% stacking gels, and the proteins analysed by densitometry. To study collagens, palatal shelves were homogenized in 0.5 M acetic acid. Collagen was solubilized by limited digestion for 16 h at 4°C with 100 pg/ml of pepsin. The samples were lyophilized and the collagen types separated by SDS-PAGE. In order to distinguish between collagen types I and III, the interrupted reduction method described by Sykes et al. (1976) was used. Collagens were stained with Coomassie brilliant blue and the proportions of the major collagen types analysed by laser densitometry.
DNA content was determined according to the method of Labarca and Paigen (1980) using Hoechst No. 33258 fluorescent dye and calf thymus DNA (Sigma) to construct a standard curve.
Analysis of GAGS
Proteoglycans were extracted from murine and chick palatal shelves using 4 M guanidinium hydro- chloride (4 M GuHCl), 25 mM tris-containing protease inhibitors, pH 7.8 at 4°C for 18 h. GAGS were recovered from the extracts as described by Chen et al. (1989) and the GAG types separated by electrophoresis on Titan III zip-zone cellulose acetate plates (Helena Laboratories, Gateshead) as described by Cappelletti, Del Rosso and Chiarugi (1979). Quantification of unknowns was by comparison with the intensity of staining for 200 ng of standard run simultaneously and analysed using an LKB laser densitometer and integrator.
Analysis of the production of extracellular matrix molecules by palatal shelves in culture
Organ culture of palatal shelves. Palatal shelves from each developmental stage (T.2cT.23 and H&H 30-35 for mouse and chick embryos, respectively) were transferred to 35 mm plastic tissue-culture dishes (Nunc) pre-coated with an air-dried film of 1% gelatin in sterile water. Routinely 10 palates were used per dish. The shelves were cultured for the least time needed to facilitate attachment (4-6 h) in standard medium: DMEM and Hams F12 medium (DMEM/F12, 1: 1 Vol) containing 2 mM glutamine and 1% penicillin-streptomycin (Gibco), sup- plemented with 2.5% donor calf serum (Imperial Laboratories, Andover) in a 5% CO, in air, 37°C environment. The attached palates were washed 3 x 5 min in standard medium with serum omitted (serum- free medium) and incubated a further 12 h in serum- free medium before adding experimental medium.
Plate 1 Fig. 1. Coronal sections (7 pm) showing the position of the tongue (T) and palatal shelves (P) in paraffin-embedded tissue from murine (A-C) and chick (D-F) embryos. Sections represent: (A) Theiler
(T) stage 20; (B) T.21; (C) T.23; (D) H&H stage 31; (E) H&H 33; (F) H&H 35.
Mouse and chick secondary-palate development 459
Plate 1
460
205 -
716 -
66 -
45-
DAVID M. FOREMAN et al.
Plate 2

Mouse and chick secondary-palate development 461
Table 1. Composition of mouse and chick palatal processes over the period of palatogenesis
Developmental Dry weight $S
Protein DNA stage (mg) 01g) 019)
Mouse T.20 0.020” + 0.014 0.192” + 0.028 11.6” + 4.79 6.5” f 1.93 T.21 O.lOEb 0.494s 38.4b 10.2’ T.22 0.09Eb 0.823’ 78.7’ 20.3b T.23 0.255’ 1.190d 153.0d 38.4’
Chick H&H 31 0.320” f 0.024 1.042’ f 0. t 10 20.0” f 2.41 3.8” i 0.31 H&H 33 0.585b 1.825b 33.0b 5.3b H&H 35 0.670’ 7.351’ 70.0’ 9.3’
Each value is the mean of three determinations + SE of difference. “,b,c,dValues in vertical columns with different superscripts, for each species, differed significantly,
p < 0.05.
Experimental media. Palatal shelves were incubated for 24 h in serum-free medium in the presence or ab- sence of EGF (10 ng/ml; Flow, Herts), bFGF (10 ng/ ml; British Biotechnology, Oxford), PDGF (10 ng/ ml; British Biotechnology, Oxford) or TGF-/I, (10 ng/ml; British Biotechnology, Oxford).
Analysis of newly synthesized proteins
To radiolabel proteins, 20 pCi/ml L[~ HI-proline was included for the final 6 h of culture. After incubation, culture medium was dialysed extensively against 0.05 M acetic acid and non-dialysable material detected by scintillation counting. Cell- associated proteins were extracted with GuHCl as described previously. Extracts were dialysed, counted and lyophilized. Collagen synthesis was assayed by the method of Peterkofsky and Diegelmann (1971). Non-collagenous proteins were removed by limited digestion with pepsin for 16 h at 4°C. The newly synthesized collagens and proteins were analysed by SDS-PAGE, followed by autoradiography, as de- scribed by Bonner and Laskey (1974). Samples were run as the relative number of counts incorporated per pg DNA.
Analysis of GAGS
Palates were incubated for 24 h in serum-free medium in the presence or absence of growth factors, as described above. [3H]-glucosamine hydrochloride (20 pCi/ml) was included for the last 16 h of incubation.
After the incubation period, medium was removed and the cell/matrix-associated GAGS extracted using 4 M GuHCl. Palate extracts and the medium were dialysed against papain buffer and digested with papain as described previously.
GAGS were analysed by cellulose acetate chroma- tography as described previously. Individual GAG bands were carefully scraped into vials and tritium incorporation determined by scintillation counting.
Size class of hyaluronan was determined by Sepharose CL-2B chromatography as described by Chen et al. (1989). Fractions (0.1 ml) were collected
and each assayed for radioactivity. Blue dextran 2000 was used to determine the void volume (V,) and [3H]-glucosamine (approx. 0.05 PCi) to determine the total volume (V,) of the column.
Statistical treatment
Samples were analysed by analysis of variance followed by a protected t-test. Means were con- sidered to differ significantly at p < 0.05.
RESULTS
Wet weight, dry weight and water content
As the size of the palatal shelves of the murine embryo increased during development the wet weight and water content of the shelves at each develop- mental stage showed a corresponding increase (Table 1). However, during the critical period of shelf elevation (T.21 and T.22), the dry weight of the shelves remained constant. Consequently a peak of water/dry weight was observed during the period of shelf reorientation (Table 2). Chick palates showed an exponential increase in both wet weight and water content, although dry weight increased less dramatically (Table 1). No peak in water content was apparent during chick palatogenesis.
Table 2. Water/dry weight, GAG/dry weight and pro- tein: DNA ratios during mouse and chick palatogenesis
Developmental H,O (mg)/ GAG @g)/ Protein @g): stage mg dry wt mg dry wt DNA (pg)
T.20 T.21 T.22 T.23
H&H 30 H&H 31 H&H 32 H&H 33 H&H 34 H&H 35
Mouse 9.6 271 1.8 4.6 120 3.7 8.4 120 3.9 4.7 70 4.0
Chick 3.2 241 3.4 3.3 233 5.3 3.0 224 6.1 3.1 214 6.1 4.4 287 6.2
11.0 504 7.5
Plate 2
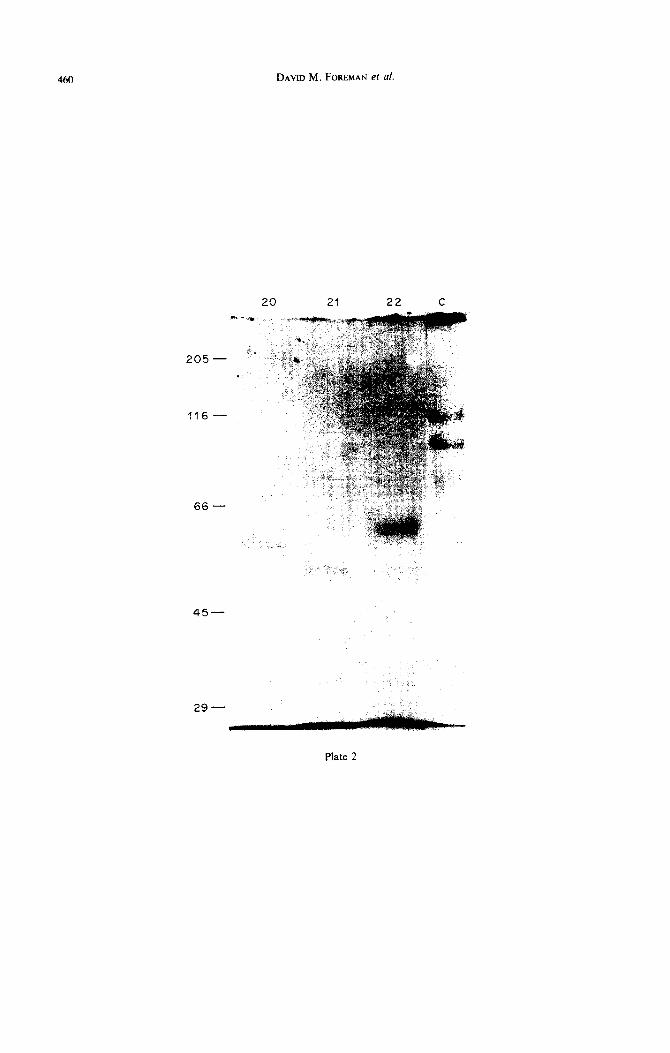
Fig. 2. SDS-PAGE (6.5%) protiles of proteins obtained from stage T.20, T.21 and T.22 murine palates. Proteins present in the GuHCl extracts were separated under reducing conditions and stained with
Coomassie blue. Lane C: type I collagen standard.

462 DAVID M. FOREMAN et al.
Protein and DNA content Palatal GAGS
In vivo. The protein and DNA content of mouse and chick palatal shelves increased with developmental maturity (Table 1). Both species showed an initial increase in the palatal pro- tein : DNA ratio, which then remained constant over the period of shelf reorientation in the mouse and growth in the chick (Table 2). In chick a further increase in this ratio was observed towards the end of palatogenesis.
The majority of palatal proteins separated by SDS-PAGE migrated to positions of low molecular weight. Comparison of the proteins separated by SDS-PAGE revealed some differences in the low molecular-weight proteins obtained from mouse palates of various stages, with the disappearance of a protein of 50 kD from T.20 palates followed by the appearance of a protein of 55 kD at the time of shelf reorientation (Plate Fig. 2). The nature of these proteins remains to be determined. No differences in the protein profile obtained from different chick palates of different embryonic stages were found (data not shown). Type I collagen was the only type detectable by pepsin digestion in both species throughout palatogenesis.
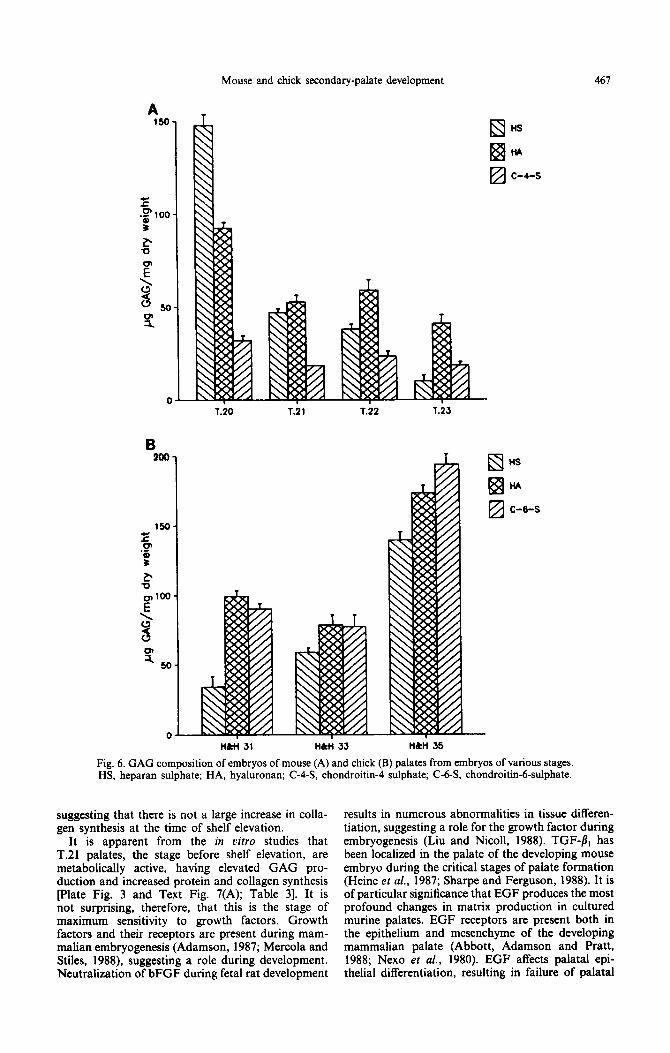
Three main classes of GAG were found in the developing palate of both chick and mouse, namely hyaluronan, heparan sulphate, and chondroitin sulphate. The type of chondroitin differed between species, mouse palates contained chondroitin-rl- sulphate and chick chondroitin-6-sulphate pext Figs 6(A) and (B)]. At the earliest stages of mouse palatogenesis (T.20), heparan sulphate was the pre- dominant GAG type, but levels of this decreased rapidly between T.20 and T.21 and then gradually over the rest of development [Text Fig. 6(A)]. By contrast, levels of chondroitin-4-sulphate remained relatively constant. After an initial decrease between stages T.20 and T.21, the amount of hyaluronan/dry weight in murine palates in vivo remained constant, with hyaluronan becoming the predominant GAG type as development progressed [Text Fig. 6(A)].
In vitro. Separation of the [j HI-proline-labelled proteins obtained from murine palates showed no change in the SDS-PAGE profiles from the various embryos (Plate Fig. 3). However, a peak of protein production was present at T.21 (Table 3). Type I collagen, together with significant amounts of colla- gen types III and V, were the major types produced by murine palates in vitro (Plate Fig. 4). The ratio of collagen types did not alter throughout development (data not shown).
Of the growth factors studied, only EGF signifi- cantly altered i3 HI-proline incorporation in murine palates in vitro (Text Fig. 5). EGF increased protein production in early- and mid- (T.20-T.22) stage murine palates but had no effect on late-stage (T.23) palates (Text Fig. 5). None of the growth factors had any significant effect on the ratio of collagen isotypes (data not shown).
The distribution of GAGS during chick palatogen- esis was totally different from that in mouse. Total GAG/dry weight in chick palatal processes remained constant until a large increase in the mature shelves (Table 2). By contrast the amount of GAG decreased during murine palatogenesis (Table 2). In addition, heparan sulphate, the major GAG in early murine palates, was the least abundant class at the early stages of chick palate development (H&H 30-31) [Text Figs 6(A) and (B)]. Levels of heparan sulphate rose markedly during chick palate development [Text Fig. 6(B)] but fell during murine palate development [Text Fig. 6(A)]. Chondroitin sulphate, the least abundant GAG type in mouse palates (5-15% of total GAG), accounted for approx. 40% of GAG in the chick palates [Text Figs 6(A) and (B)]. Further- more, in the mature palate, hyaluronan accounted for only approx. 30% of the total GAG in the chick compared to approx. 60% in the mouse [Text Fig. 6(B)].
The rate of proline incorporation in early- and mid-stage chick palatal shelves in vitro remained relatively constant but was elevated in later stages (Table 3). This increase was greater in collagen than non-collagenous protein. Separation of [3H]- proline-labelled chick palatal proteins showed no differences between developmental stages. Chick palates also synthesized collagens types I, III and V, although the proportion of type V collagen was less than in the mouse (Plate Fig. 4). As in murine palates, the ratio of the collagen types did not vary throughout development. None of the growth factors studied significantly affected [‘HI- proline incorporation by chick palates in organ culture (data not shown).
Palatal shelves in culture synthesized the GAG types found in uivo but in different relative pro- portions. In the murine palate, hyaluronan was the most abundant GAG synthesized, with a peak of production at T.21 [Text Fig. 7(A)]. A similar peak in production of chondroitin-4-sulphate was observed. Heparan sulphate was produced in the least amounts and rates of its production decreased with developmental maturity.
In the chick palate, chondroitin-6-sulphate was the most abundant GAG synthesized and production rates increased at later developmental stages [Text Fig. 7(B)]. A similar trend was seen with hyaluronan. By contrast, levels of heparan sulphate fell in the more mature chick palates [Text Fig. 7(B)].
Analysis of the size class of hyaluronan produced by murine palates in vitro revealed the existence of a very high molecular-weight compound (Mr > 2 x 106), which was eluted at the void volume, together with diverse lower molecular-weight species
Plate 3
Fig. 3. Fluorograms of [‘H]-pro1ine-1abeI1ed proteins synthesized by murine palatal shelves of different stages in culture. Track 1 and 6; T.20 GuHCl extract, Track 2 and 7; T-21 GuHCl extract, Track 3 and 8; T.22 GuHCl extract, Track 4 and 9; T.23 GuHCl extract, Track 5; [3H]-acctyiated rat tail tendon
collagen. Tracks 1-5 were unreduced, Tracks 6-9 were reduced.

Mouse and chick secondary-palate development
123456 7 8 9
463
al (I)
Plate 3

DAVID M. FOREMAN et al.
Plate 4

Mouse and chick secondary-palate development 465
Table 3. Production of collagenous and non-collagenous proteins by various stage mouse and chick palates in culture
Developmental Collagen stage NCP CP (%)
Mouse T.20 T.21 T.22 T.23
35.1’ f 2.25 8.3’ f 0.51 4.2 43.6b 10.9b 4.4 24.Y 6.2’ 4.4 17.9d 4.4d 4.3
Chick H&H 30 18.2” f 1.26 4.6’ k 0.39 4.5 H&H 31 19.3”b 4.8”b 4.4 H&H 32 18.5’ 4.6” 4.4 H&H 33 19.0” Qb 5.3 H&H 34 22.6b 7.5’ 5.8 H&H 35 22.7b 9.3d 7.1
Incorporation of [‘HI-proline (dpm x IO-‘/pgDNA). Each value is the mean of three determinations k SE of
difference. Values in vertical columns with different superscripts, for each species, differ significantly, p < 0.05. NCP, non-collagenous protein, CP, collagenous protein. To correct for the enriched proline content of collagen, % collagen was determined as follows (%): (dpm CP x 100)/([5.4 x NCP] + CP).
rext Fig. 8(A)]. By contrast, chick palates in vitro produced only high molecular-weight hyahtronan pext Fig. 8(B)].
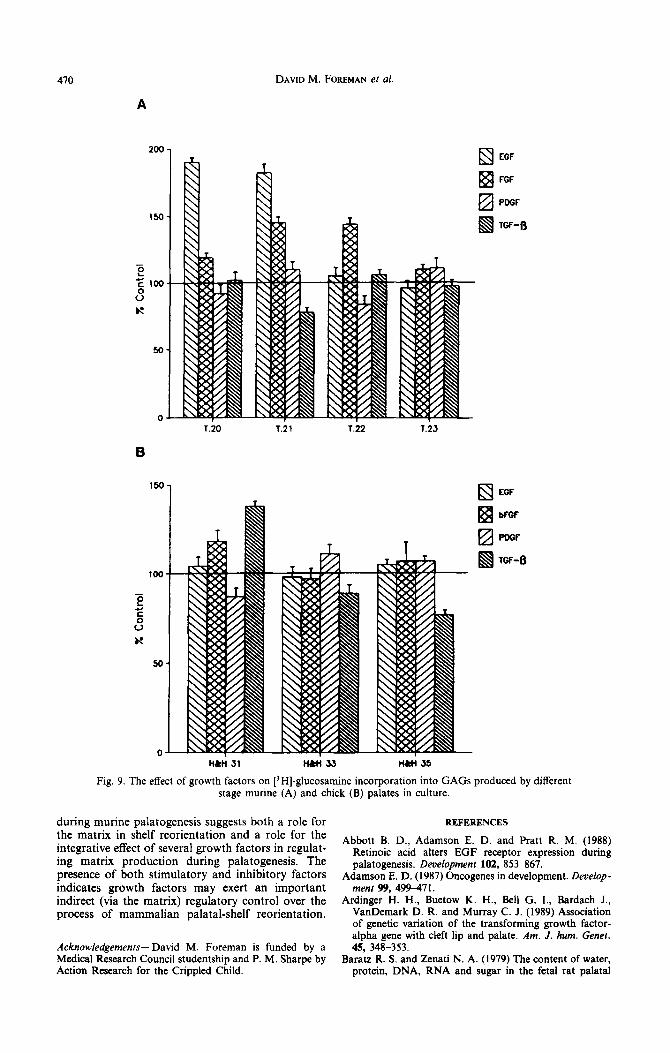
The incorporation of [3H]-glucosamine into the GAGS of the murine palate could be modulated by the addition of growth factors to the culture medium [Text Fig. 9(A)]. EGF caused a significant increase in GAG accumulation in early-stage (T.20 and T.21) palates, but had no effect on later stages [Text Fig. 9(A)]. bFGF induced a large rise in total GAG production at stages T.21 and T.22, had a smaller effect at T.20, and was ineffective on T.23 palates [Fig. 9(A)]. The increases in GAGS caused by EGF and bFGF were due to an increase in all three GAG types (data not shown). PDGF had no significant effect at any stage. TGF-/? caused a significcant reduction in [3H]-glucosamine incorporation only at T.21 [Text Fig. 9(A)].
Chick palatal shelves were relatively insensitive to the effects of growth factors. In early-stage palates (H&H 31), only TGF-fl caused a significant increase in GAG production whereas at later stages (H&H 33, 35) TGF-fi decreased GAG production [Text Fig. 9(B)].
DISCUSSION
There are many theories concerning the mechanism of mammalian palatal-shelf elevation (recently reviewed by Ferguson, 1988). These theories are not mutually exclusive and shelf elevation is probably the result of a co-ordinate interaction of several factors and may even occur by different principal mechan- isms along the length of the shelf. The predominant
Plate 4
putative mechanism is the production of an ‘internal shelf force’ caused by the hydration of palatal GAGS (Ferguson, 1988), the resultant turgor pressure being directed along collagen fibres and restrained by the covering epithelia. We therefore proposed to compare the biochemistry, in terms of GAG and protein (collagen) metabolism, in embryonic mouse palatal shelves, which elevate, and chick palatal shelves, which do not.
Unfortunately, it became clear that, rather than showing many similarities and relatively few differences, chick palatal shelves have a completely different biochemical composition to their murine counterparts. For example, chick palates have a relatively lower DNA and protein but higher GAG content than mouse (Table 1). Furthermore, the GAG composition of the chick palatal shelf differed markedly from that of the mouse [Text Figs 6(A) and (B)]. In addition, [‘HI-glucosamine incorporation by chick palates in uitro did not show a peak as in the mouse but rather increased gradually with develop- mental age [Text Figs 7(A) and (B)]. Interestingly, the type of chondroitin sulphate present differed between the two species, the 6-sulphate being found in chick, with the 4-sulphate present in mouse. These findings differ from those of both Larsson (1962) and Knud- sen, Bulleit and Zimmerman (1985), who reported the presence of both chondroitin4- and -6-sulphate in developing murine palates. However, their findings were based on histochemical techniques rather than a complete separation of the GAG types by chroma- tography. The difference in chondroitin types be- tween mouse and chick may be a species-specific difference, or could reflect the fact that chondrogen- esis occurs in the chick palatal shelf towards the end of palatogenesis and chondroitin-6-sulphate is present as a marker for cartilage formation. The increase in protein and GAG content in vivo and the increased protein and collagen synthesis in vitro in mature chick palates are probably attributable to osteogenesis and chondrogenesis.
Differences between the species were also observed in the size of hyaluronan produced by the palatal shelves in oitro [Text Figs 8(A) and (B)]. Mouse palates produced high molecular-weight hyaluronan together with diverse lower molecular-weight forms, whereas chick palates yielded only high molecular- weight forms. Whether the lower molecular-weight forms found in mouse palates are derived from the higher molecular-weight forms via degradation within the palate is unclear, but different size classes of hyaluronan are known to have different effects on cell differentiation (Caplan, 1987) and may therefore contribute toward the different developmental fates within the chick and mouse palatal shelves.
Novel synthesis of an unidentified low molecular- weight protein occurred at the time of shelf reorienta- tion in the murine palate and not the chick (Plate Fig. 2).
Fig. 4. Fluorogram of the [3H]-proline-labelled collagens derived from chick and mouse palates in culture. Proteins extracted with GuHCl were digested with pepsin and the collagen types separated by interrupted
reduction on 8% polyacrylamide gels. Track 1; chick (H&H 33) Track 2; mouse (T.21).

466 DAVID M. FOREMAN et al.
EGF
Iz! FGF
I!zI PDGF
TGF-8
Fig. 5. The
T.iO Til T.iZ 7.23
effect of growth factors on [3H]-proline incorporation into proteins synthesized stage murine palates in culture.
by different
These extreme differencess in metabolism between mouse and chick palates caution against a narrow mechanistic interpretation of the data in terms of shelf elevation: species-specific as well as morphogen- etic-specific differences are likely.
Pratt et al. (1973) reported that hyaluronan com- prised approx. 60% of the total GAG in the develop- ing rat palate and found a peak in GAG production both in vivo and in vitro during the period of shelf rotation and fusion. Knudsen et al. (1985) identified hyaluronan as the predominant GAG class in murine palates and also showed the presence of both chon- droitin-4- and -6-sulphate. Our work has confirmed that hyaluronan is the predominant GAG class in mid and mature murine palatal shelves, but has further demonstrated that heparan sulphate is the major GAG type in early (stage T.20) palates [Text Fig. 6(A)]. As heparan sulphate is concentrated in basement membranes, its preponderance at the onset of palatogenesis may simply reflect the increased ratio of epithelium to mesenchyme found in the small, immature, palatal shelf. However, in the early-stage chick palate, heparan sulphate was the minor GAG type, the amounts of which, in contrast to the mouse, increased as development proceeded [Text Fig. 6(B)]. Mesenchymal heparan sulphate in the murine palate is predominantly pericellular in distribution (Thomas B. and Ferguson M. W. J., unpublished observation) and may serve to effect relatively strong cellLcel1 adhesion. The decreasing amount of heparan sul- phate in the mouse palate correlates with an increase in mesenchyme cell separation as the turgor-like shelf force increases, whereas in the chick no such changes occur and heparan sulphate may contribute to keeping the cells in close opposition.
After an initial large decrease, the relative amount of hyaluronan in murine palates remained fairly constant. In contrast to the findings of Pratt et al. (1973) no peak in hyaluronan was detected in vivo (there was a small rise at T.22) although when
cultured in vitro, increased [‘HI-glucosamine incor- poration into hyaluronan and chondroitin sulphate was found at stage T.21 [Text Fig. 7(A)], the stage before shelf elevation. Indeed, in murine palates in vivo, total GAG/dry weight decreased with developmental maturity (Table 2). It is difficult, therefore, to correlate the increase in hydration observed in the murine palate at the time of shelf reorientation (T.22) with an increase in palatal GAG content, although this does not exclude a regionally specific accumulation of hyaluronan contributing to shelf elevation as postulated by Ferguson (1978) Knudsen et al. (1985) and Brinkley and Morris-Wiman (1987).
Baratz and Zenati (1979) reported that the water content of rat palatal shelves decreased throughout development, casting doubt on the hydration theory of shelf elevation. We have now examined the relative degree of hydration in the murine palate over time and found that, after an initial decrease, the water content of the shelf increased at the time of shelf reorientation (T.22), decreasing again after shelf fusion (Table 2). These findings are consistent with the hydration hypothesis proposed by Pratt et al. (1973).
Collagens are thought to play an important role during palatal-shelf elevation because inhibition of collagen cross-linking results in cleft palate (Pratt and King, 1972; Steffek, Verrusio and Watkins, 1972). Pratt and King (1971) found a significant increase in the collagen content of the rat palate during the time of shelf reorientation and fusion. We here found a similar peak of collagen and protein synthesis in cultured day-13 murine palates (Plate Fig. 3; Table 3). Interestingly, both mouse and chick palates synthesized significant amounts of types III and V collagen in vitro, which were not detected in vivo (Plate Fig. 4). In viva, after an initial increase, the palatal protein : DNA ratio remained constant over the period of shelf elevation and fusion (Table 2),
Mouse and chick secondary-palate development 467
HS
HA
c-4-s
69 I+3
Eh Ei c-e-s
Fig. 6. GAG composition of embryos of mouse (A) and chick (B) palates from embryos of various stages HS, heparan sulphate; HA, hyaluronan; C-4-S, chondroitin-4 sulphate; C-63, chondroitin-6-sulphate.
suggesting that there is not a large increase in colla- gen synthesis at the time of shelf elevation.
It is apparent from the in vitro studies that T.21 palates, the stage before shelf elevation, are metabolically active, having elevated GAG pro- duction and increased protein and collagen synthesis [Plate Fig. 3 and Text Fig. 7(A); Table 31. It is not surprising, therefore, that this is the stage of maximum sensitivity to growth factors. Growth factors and their receptors are present during mam- malian embryogenesis (Adamson, 1987; Mercola and Stiles, 1988), suggesting a role during development. Neutralization of bFGF during fetal rat development
results in numerous abnormalities in tissue differen- tiation, suggesting a role for the growth factor during embryogenesis (Liu and Nicoll, 1988). TGF-fi, has been localized in the palate of the developing mouse embryo during the critical stages of palate formation (Heine et al., 1987; Sharpe and Ferguson, 1988). It is of particular significance that EGF produces the most profound changes in matrix production in cultured murine palates. EGF receptors are present both in the epithelium and mesenchyme of the developing mammalian palate (Abbott, Adamson and Pratt, 1988; Nexo et al., 1980). EGF affects palatal epi- thelial differentiation, resulting in failure of palatal

468 DAVID M. FOREMAN et al.
T.20 T.21 7.22 7.23
0-l 1 I
H&H 31 Ii&H 33 ll&tl 35
Fig. 7. [3H]-glucosamine incorporation into GAGS (see Text Fig. 6) produced by cultured mouse (A) and chick (B) palates from various stage embryos. Results are expressed as d.p.m./pgDNA.
fusion in vitro (Hassell, 1975; Tyler and Pratt, 1980). Furthermore, Ardinger et al. (1989) have recently shown a significant association between mutation in the TGF-c( (a homologue of EGF) gene and predisposition to cleft lip and palate in man.
When considering the response of palatal shelves to added growth factors in vitro, it has to be borne in mind that endogenous concentrations of such factors are unknown, and will in any case presumably vary with developmental time, so that the amount of factor added may vary markedly from levels found in oioo. The response of murine palates to growth factors was found to he developmentally regulated. EGF increased [3H]-glucosamine incorporation into
GAGS [Text Fig. 9(A)] and [3H]-proline incorpor- ation into proteins (Text Fig. 5) in early-stage palates. In contrast to the work of Silver, Murray and Pratt (1984), who noticed a significant increase in type V collagen synthesis on addition of EGF to cultured murine palates, we found EGF did not cause a shift in collagen isotypes, but rather resulted in a broad- spectrum increase in all collagens synthesized (data not shown). The effect of EGF decreased with devel- opmental maturity, in later-stage palates (T.22-T.23) the growth factor had no effect. This difference in response with developmental time suggests a develop- mental regulation of receptor numbers and/or affinity in the embryonic murine palate: such decreases have

Mouse and chick secondary-palate development 469
A
5000 -e- control jc HAase Dig
0 _p-
0 10 20 30 40 50
Fraction
4 control SC HAase Dig
6000.
3000.
2000.
0 IO 20 30 40 50
Fraction
Fig. 8. Sepharose 2B chromatography of [3H]-glucosarnine lahelled material obtained from T.21 murine (A) and H&H 33 chick palates (B). Duplicate samples were run as controls and Strepromyces
hyaluronidase (HAase Dig) digested.
been reported for the medial-edge epithelia (Abbott from other systems (Wrann, Fox and Ross, 1980) et al., 1988). These results differ from those of Turley indicating that PDGF can induce the expression et al. (1985), who found that EGF stimulated GAG of EGF receptors, thus perhaps masking the production at all stages of palatogenesis. However, developmental regulation described here. bFGF also our experiments were conducted in serum-free increased GAG production [Text Fig. 9(A)] but its medium, whereas the medium used by Turley et al. maximum effect occurred during mid palatogenesis (1985) was supplemented with 10% fetal calf serum. (T.21 and T.22). By contrast, TGF-8, decreased It is known that the effects of growth factors can be GAG production at the time of shelf reorientation. profoundly altered by the presence of other growth Chick palatal shelves were less responsive to growth factors (Edwards et al., 1987), thus addition of 10% factors than mouse palates, only TGF-fi, producing fetal calf serum to their palate cultures suggests that any significant response [Text Fig. 9(B)]. Interest- Turley and co-workers may have been observing the ingly, the response of chick palates to TGF-& was effects of an interaction of EGF with other growth also developmentally regulated. The developmental factors present in the serum. Of interest are data timing of growth-factor effects on matrix production
470 DAVID M. FOREMAN et al.
A
B
0
69 EGF
•!j FGF
Ezl PDGF
IQ TGF-fi
q EGF
Fig. 9. The effect of growth factors on [‘HI-glucosamine incorporation into GAGS produced by different stage murine (A) and chick (B) palates in culture.
during murine palatogenesis suggests both a role for the matrix in shelf reorientation and a role for the integrative effect of several growth factors in regulat- ing matrix production during palatogenesis. The presence of both stimulatory and inhibitory factors indicates growth factors may exert an important indirect (via the matrix) regulatory control over the process of mammalian palatal-shelf reorientation.
Acknowledgements-David M. Foreman is funded by a Medical Research Council studentship and P. M. Sharpe by Action Research for the Crippled Child.
REFERENCES
Abbott B. D., Adamson E. D. and Pratt R. M. (1988) Retinoic acid alters EGF receptor expression during palatogenesis. Development 102, 853-867.
Adamson E. D. (1987) Oncogenes in development. Deuelop-
ment 99, 499-47 1. Ardinger H. H., Buetow K. H., Bell G. I., Bardach J.,
VanDemark D. R. and Murray C. J. (1989) Association of genetic variation of the transforming growth factor- alpha gene with cleft lip and palate. Am. J. hum. Genet. 45, 348-353.
Baratz R. S. and Zenati N. A. (1979) The content of water, protein, DNA, RNA and sugar in the fetal rat palatal

Mouse and chick secon .dary-palate development 471
Liu L. and Nicoll C. S. (1988) Evidence for a role of basic fibroblast growth factor in rat embryonic growth and differentiation. Endocrinology 123, 2027-203 1.
Mercola M. and Stiles C. D. (1988) Growth factor super- families and mammalian embryogenesis. Deuelopmen~ 102, 369-376.
Narayanan A. S. and Page R. C. (1983) Biosynthesis and regulation of type V collagen in diploid human fibro- blasts. 1. bioi. Chem. 258, 11694-I 1699.
Nexo E., Hollenberg M. D., Figueroa A. and Pratt R. M. (1980) Detection of epidetmal growth factor-urogastrone and its receptor during fetal mouse development. Proc. natn. Acad. Sci., U.S.A. 77, 2782-2735.
Peterkofsky B. and Diegelmann R. (1971) Use of a mixture of proteinase-free collagenases for the specific assay of radioactive collagen in the presence of other proteins. Biochemistry 10, 988-994.
Pratt R. M. and King C. T. G. (1971) Collagen synthesis in the secondary palate of the developing rat. Archs oral Bioi. 16, 1181-1185.
Pratt R. M. and King C. T. G. (1972) Inhibition of collagen cross-linking associated with B-aminopropionitrile- induced cleft palate in the rat. Deui Biol. 27, 322-328.
Pratt R. M., Goggins J. F., Wilk A. L. and King C. T. G. (1973) Acid mucopolysaccharide synthesis in the sec- ondary palate of the developing rat at the time of rotation and fusion. Devl Biol. 32, 230-237.
Pratt R. M., Yoneda T., Silver M. H. and Salomon D. S. (1980) Involvement of glucocorticoids and epidermal growth factor in secondary palatal development. In Current Research Trends in Prenatal Craniofacial Deuelopmenf (Eds Pratt R. M. and Christiansen R. L.), pp. 235-252. Elsevier, North Holland.
Rizzino A. (1988) Transforming growth factor-j?: Multiple effects on cell differentiation and extracellular matrices. Devl Biol. 130, 411422.
Sharpe P. M. and Ferguson M. W. J. (1988) Mesenchymal influences on epithelial differentiation in developing sys- tems. J. Cell Sci. Suppl. 10, 195-230.
Silver M. H., Murray J. C. and Pratt R. M. (1984) Epider- ma1 growth factor stimulates type-V collagen synthesis in cultured murine palatal shelves. Differenriation 27, 205-208.
Steffeck A. J., Verrusio A. C. and Watkins C. A. (1972) Cleft palate in rodents after maternal treatment with various lathyrogenic agents. Teratology 5, 3340.
Sykes B., Puddle B., Frances M. and Smith R. (1976) The estimation of two collagens of human dermis by interrupted gel electrophoresis. Biochem. biophys. Res. Commun. 72, 1472-1480.
Theiler K. (1972) The House Mouse. Development and Normal Stages from Fertilisation to 4 Weeks of Age. Springer, Berlin:
processes during reorientation and fusion. Archs oral Biol. 24, 607-611.
Bonner W. M. and Laskey R. A. (1974) A film detection method for tritium-labelled proteins and nucleic acids in polyacrylamide gels. Eur. J. Biochem. 46, 83-88.
Bradford M. M. (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. A. Biochem. 72, 248-254.
Brinkley L. L. and Morris-Wiman J. (1987) Computer- assisted analysis of hyaluronate distribution during mor- phogenesis of the mouse secondary palate. Development 100, 629635.
Caplan A. I. (1987) The extracellular matrix is instructive. BioEssays 5, 129-132.
Cappelletti R., Del Rosso M. and Chiarugi V. P. (1979) A new electrophoretic method for the complete separation of all known animal glycosaminoglycans in a mono- dimensional run. Biochemistry 99, 3 1 l-3 15.
Chen W. Y. J., Grant M. E., Schor A. M. and Schor S. L. (1989) Differences between adult and foetal fibroblasts in the regulation of hyaluronate synthesis: correlation with migratory activity. J. Cell Sci. 94, 577-584.
Edwards D. R., Murphy G., Reynolds J. J., Whitam S. E., Docherty A. J. P., Angel P. and Heath J. K. (1987) Transforming growth factor beta modulates the ex- pression of collagenase and metalloproteinase inhibitor. EMBU Jl6, 1899-1904.
Ferguson M. W. J. (1978) Palatal shelf elevation in the Wistar rat fetus. J. Anal. 125, 555-577.
Ferguson M. W. J. (1988) Palate development. In Craniofacial Development, Development 103 supple- ment (Eds Thorogood P. and Tickle C.), pp.4160. Company of Biologists, Cambridge.
Ferguson M. W. J., Honig L. S. and Slavkin H. C. (1984) Differentiation of cultured palatal shelves from alligator, chick and mouse embryos. Anat. Rec. 209, 23 l-249.
Hamburger V. and Hamilton H. L. (1951) A series of normal stages in the development of the chick embryo. J. Morph. 88, 49-92.
Hassell J. R. (1975) The development of rat palatal shelves in vitro. An ultrastructural analysis of the inhibition of epithelial cell death and palate fusion by the epidermal growth factor. Devl Biol. 45, 90-102.
Heine U. I., Munoz E. F., Flanders K. C., Ellingsworth L. P., Lam P., Thompson N. C., Roberts A. B. and Spom M. B. (1987) The role of transforming growth factor beta in the development of the mouse embryo. J. Cell Biol. 105, 2861-2867.
Knudsen T. B., Bulleit R. F. and Zimmerman E. F. (1985) Histochemical localisation of glycosaminoglycans during morphogenesis of the secondary palate in mice. Anat. Embryol. 173, 137-142.
Koch W. E. and Smiley G. R. (1981) In-uiuo and in-vitro studies of the development of the avian secondary palate. Archs oral Biol. 26, 181-187.
Labarca C. and Paigen K. (1980) A simple rapid and sensitive DNA assay procedure. Anaiyr. Biochem. 102, 344-352.
Laemmli U. K. (1970) Clearage of proteins during the assembly of the head of bacteriophage T4. Nature 227, 68&685.
Larsson K. S. (1962) Studies on the closure of the secondary palate. Autoradiographic and histochemical studies in the normal mouse embryo. Acta morph. neeri-stand. 4, 349-367.
Tsene S. C. G.. Savion N.. Stern R. and Gosoodarowicz D. (1982) Fib&blast growth factor modulates synthesis of collagen in cultured vascular endothelial cells. J. Cell Bioi. 122, 3555360.
Turley E. A., Hollenberg M. D. and Pratt R. M. (1985) Effect of epidermal growth factor/urogastrone on glycosaminoglycan synthesis and accumulation in uirro in the developing mouse palate. Dzfirenriarion X&279-285.
Tyler M. S. and Pratt R. M. (1980) Effect of epidermal growth factor on secondary palatal epithelium in oitro: tissue isolation and recombination studies. J. Embryol. exp. Morph. 58, 93-106.
Wrann M., Fox C. F. and Ross R. (1980) Modulation of epidermal growth factor receptors on 3T3 cells by platelet-derived growth factor. Science 210, 1363-1365.




















