Effect of Neofabraea Alba on Bark and Wood Anatomy of Fraxinus Spp

IAWA Journal, Vol. 26 (4), 2005: 477–487
COMPARATIVE BARK ANATOMY OF ROOT AND STEM INSTYRAX CAMPORUM (STYRACACEAE)
Silvia Rodrigues Machado1, Carmen Regina Marcati2, Berta Lange de Morretes3 & Veronica Angyalossy3
SUMMARY
The bark of Styrax camporum Pohl (Styracaceae) differs anatomically in the root and stem. Roots have layered secondary phloem; short sieve tubes with simple, transverse or more or less inclined sieve plates; fibres in tangential bands; astrosclereids; wide rays, and a poorly developed periderm. Stems have non-layered secondary phloem; longer sieve tubes with compound, scalariform, inclined sieve plates; sclerified cells and brachysclereids; a developed periderm, and a non-persistent rhytidome. Prismatic crystals, starch grains, phenolic compounds and lipidic contents were observed in root and stem bark cells. The differences between the secondary phloem of root and stem are discussed.
Key words: Bark anatomy, root, secondary phloem, stem, Styracaceae, Styrax camporum.
INTRODUCTION
Styracaceae occur in relatively warm parts of the world, such as the Mediterranean re-gion, eastern Asia, the Malay Archipelago and parts of North and South America, and one genus in tropical Africa. This family is best known by the indumentum of stellate or scale-like hairs and for its production of benzoin (gum benzoin) and storax (Metcalfe & Chalk 1950; Hutchinson 1973; Spongberg 1976). Styrax L. is by far the largest and most widespread of the 11 genera in the Styraca-ceae, with about 130 of the c. 160 species in the family. It has a widespread but disjunct distribution, occurring in the Americas, eastern Asia, and the Mediterranean region. Over half the species of the genus occur in South America, where they are distributed among a wide array of habitats, including the lowland and montane rain forest, sub-paramo, tepui scrub, restinga, rocky grasslands, and cerrados (Nakajima & Monteiro 1986; Fritsch 2001). Some Brazilian species of Styrax, for example S. ferrugineus and S. camporum, produce resin when the bark is wounded (Mors & Rizzini 1966). According to Metcalfe and Chalk (1950), the resin Storax and Gum Benzoin obtained
1) Departamento de Botânica, Instituto de Biociências, Universidade Estadual Paulista, Botucatu SP, CP 510, CEP 18618-000, Brazil [E-mail: [email protected]].
2) Departamento de Recursos Naturais – Ciências Florestais, Universidade Estadual Paulista, Botucatu SP, CP 237, CEP 18603-970, Brazil.
3) Departamento de Botânica, Universidade de São Paulo, São Paulo SP, CP 11461, CEP 05421-970, Brazil.
Associate Editor: Alex Wiedenhoeft

IAWA Journal, Vol. 26 (4), 2005478 479Machado et al. — Bark anatomy of Styrax camporum
from Styrax officinale and S. benzoin, respectively, are secreted chiefly in pathological intercellular spaces of the phloem and wood of the stem. Styrax camporum Pohl, known as “estoraque” or “beijoeiro”, is a widely distributed small tree or shrub in the cerrado vegetation of south-eastern Brazil (Nakajima & Mon-teiro 1986). Cerrados, savanna-like ecosystems, are characterized by seasonal rainfall with dry cool winters (Franco 2002). In the dry season, fire is common in cerrados, as it is for most savanna ecosystems. The woody vegetation of cerrados is adapted to fire (Coutinho 1990), and the bark of S. camporum represents an effective protection for the cambium (Miranda et al. 2002). Although bark is of great importance both for resin production (Fahn 1979; Nair 1995) and as a protective barrier, very little information is available about bark anatomy in Styrax. General information about stem bark from several Styracaceae genera, including periderm origin and secondary phloem structure, were given by Metcalfe and Chalk (1950). In 1987, Cutler et al. described the bark anatomy of the roots of many species in their manual on root identification and included 4 species of Styrax. This paper describes the bark anatomy of root and stem of S. camporum and relates the results to the environmental conditions of the cerrado.
MATERIALS AND METHODS
Bark samples were obtained from adult plants of Styrax camporum from the cerrado near Botucatu, State of São Paulo, Brazil, (22° 55 ̓S, 48° 30 ̓W). The vouchers and sam-ples of root and stem wood of one specimen are deposited, respectively, in the Her-barium (SPF 42.588) and the Wood Collection (SPFw 302-root and 303-stem) of the Botany Department, University of São Paulo. Root samples were taken 1 m from the stem base, and have a diameter of 2 cm. Stem samples were taken below the first branch, at a distance of 50 cm from the base of the stem, and have a diameter of 5 cm. Bark samples were fixed in FAA 50, dehydrated in alcohol and subsamples were embedded in paraffin (Johansen 1940). Transverse, radial, and tangential sections (8–10 μm) were double-stained with safranin-fast-green and mounted in Permount synthetic medium. Some fixed, unembedded samples were cut on a sliding microtome and the sections (12–15 μm) stained with 1% aqueous solution of safranin and astra blue (Roeser 1972). Specimens from both root and stem were macerated in a mixture of equal volumes of acetic acid and hydrogen peroxide at 60 °C (Johansen 1940) for 12 to 24 hours. The material was stained with 1% aque-ous solution of safranin and astra blue and mounted in glycerine. Freehand sections of fresh material were treated with Sudan IV for identification of lipids and with ferric chloride for the identification of phenolic compounds (Jensen 1962). Bark descriptions follow Trockenbrodt (1990), Angyalossy-Alfonso and Richter (1991) and Richter et al. (1996). Quantitative data are based on 30 individual counts; the statistical requirements for minimum numbers of measurements were fulfilled: N = (t-value)2 . (sample variance) / (accuracy of 10% × sample mean)2, following Freese (1967) and Eckblad (1991). The numerical values given in the descriptions are expressed as minimum - mean - maximum.

IAWA Journal, Vol. 26 (4), 2005478 479Machado et al. — Bark anatomy of Styrax camporum
RESULTS
RootGeneral description — Bark reddish brown externally and dark reddish internally; surface smooth; thickness variable (0.2–0.4 cm) in relation to root diameter/age; bal-samic odor. Periderm: developed (Fig. 1 & 2), shallow and with straight course; phel-lem layered, composed of 3–10 layers of radially flattened thin-walled cells (Fig. 2); phelloderm composed of 1–3 rows of radially flattened thin-walled cells; oil contents were detected in phelloderm cells and phenolic contens in all periderm cells. Secondary phloem: regularly layered from cambium toward periphery in the transverse section; layers formed by multiseriate tangential bands of parenchyma and conducting cells (thin-walled sieve tube elements and companion cells) regularly alternating with tan-gential rows of thick-walled cells (sclerenchyma) (Fig. 1 & 3). These rows are crossed by phloem rays. Axial parenchyma: arranged in uniseriate tangential bands and with intercellular spaces in the innermost region (Fig. 4); cells become dilated towards out-ermost region of the phloem. Sieve tube elements: solitary or in groups of 4–5 (Fig. 4 & 14), interspersed with parenchyma cells and radially arranged; irregular in outline, about 26–33–46 μm in diameter and 105–297–462 μm in length; sieve plates hori-zontal to inclined, predominantly simple (Fig. 14 & 15); rarely compound sieve plates were observed with 3–6 sieve areas in a scalariform pattern; lateral sieve areas poorly developed. Rays: uniseriate (including locally biseriate rays) to 3–4 cells wide (Fig. 7); 32–48–73 μm in width, 10–16–26 cells and 305–636–976 μm in height; heterocellular with more than 4 rows of upright and/or square marginal cells; dilated towards the periphery due to anticlinal cell division and/or tangential cell expansion (Fig. 1 & 3); when in contact with fibres in the outer phloem, these cells accumulate phenolic com-pounds, develop a thick lignified secondary wall and show a pronounced intrusive growth (Fig. 6). Fibres: arranged in regularly spaced tangential bands (Fig. 1, 3 & 16); 1098–1311–1723 μm in length and 32 μm in diameter; fibre walls with short projections bearing crystalliferous cells (Fig. 17 & 18). Sclereids: astrosclereids (Fig. 19) occur near the phloem fibres and differentiate from parenchyma cells; these cells are filled with phenolic compounds before becoming sclerified and undergoing intrusive growth (Fig. 5). Inorganic contents: prismatic crystals isolated in chambered axial parenchyma cells and in square and/or upright subdivided ray cells (Fig. 6 & 7). Organic contents: starch grains in parenchyma cells; phenolic compounds in parenchyma cells (Fig. 5) and sclereids (Fig. 6); lipids in procumbent idioblastic ray cells (Fig. 7).
StemGeneral description — Bark grey-brown externally and dark reddish internally; surface scaly; thickness variable (0.1–0.3 cm); balsamic odor. Periderm: well-developed and undulating (Fig. 8 & 9); phellem composed of 20–30 layers of radially flattened cells with U-shaped thickenings; phelloderm poorly developed, composed of 2–3 rows of thin-walled cells. Rhytidome non-persistent, encompassing 4–5 periderm layers and including parts of the collapsed phloem. Phenolic compounds in all cells of phellem and phelloderm. Lenticels present. Secondary phloem: non-layered; small groups of scler-

IAWA Journal, Vol. 26 (4), 2005480 481Machado et al. — Bark anatomy of Styrax camporum
enchyma cells interspersed with parenchyma cells (Fig. 8, 9 &10). Axial parenchyma: rectangular thin-walled cells, without intercellular spaces, in the innermost region of bark (Fig. 9); in the outer portion the parenchyma cells are tangentially enlarged and some cells become sclerified. Sieve tube elements: solitary or in groups of two, inter-spersed with parenchyma cells; about 25–27–30 μm in diameter and 340–390–480 μm in length; sieve plates inclined, compound, with 3–8 sieve areas in a scalariform pattern; lateral sieve areas prominent (Fig. 20). Rays: uniseriate to locally biseriate, rarely to 3 cells wide (Fig. 11); about 15–27–37 μm in width, 11–17–29 cells and 243–498–780 μm in height; heterocellular, with more than 4 rows of upright and/or square margi-nal cells (Fig. 10); outer region of the phloem similar to root phloem with regard to ray cell dilatation and sclerification (Fig. 8 & 9). Sclereids: solitary or in groups, numerous in the outer, dilated portion of the phloem (Fig. 8, 9 & 10). Brachysclereids with thick, concentric wall lamellae and numerous pit canals (Fig. 21 & 22); developed from en-larged parenchyma cells. Fibers: absent. Sclerified cells: clusters of axially elongated fibre-like cells (Fig. 23) with thick concentric wall lamellae when seen in transverse section (Fig. 8–10); 497–843–1050 μm in length and about 100 μm in diameter; dif-ferentiated in the innermost phloem region near the cambium where they are arranged in tangential bands (Fig. 8 & 9). Inorganic contents: prismatic crystals isolated in square and/or upright subdivided ray cells (Fig. 11 & 13). Organic contents: lipids in ray cells (Fig. 12); starch grains and phenolic compounds in parenchyma cells (Fig. 13).
→Fig. 1–7. Root bark. – 1: Transverse section, diagram showing original periderm and layered sec-ondary phloem (parenchyma and conducting cells regularly alternating with tangential bands of sclerenchyma). – 2: Radial section, original periderm. – 3–6: Transverse sections. – 3: Layered secondary phloem and dilated rays. – 4. Clusters of sieve-tube elements in non-collapsed second-ary phloem with simple, transverse sieve plates (sp). Companion cells (cc). Intercellular spaces (arrows). – 5: Intrusive growth in axial parenchyma cells filled with phenolic compounds. Note spaces between parenchyma cells. – 6: Sclereids and prismatic crystals (arrow) in dilated rays. – 7: Tangential section, oil content (arrow) and prismatic crystals (arrowhead) in rays.
Symbols in 1: periderm, phellogen, parenchyma, sieve tubes and companion cells, sclereids, fibres, cambial zone.
Scale bar in 1 = 1000 μm; in 2 = 500 μm; in 3 = 200 μm; in 4, 5 & 7 = 50 μm; in 6 = 100 μm.
→ →Fig. 8–13. Stem bark. 8 & 9: Transverse sections. – 8: Diagram showing periderm developed with rhytidome, non-stratified secondary phloem. – 9: Sclerified cells near cambium and sclereids in dilated rays. – 10: Radial section, note the isodiametric sclereids in the external secondary phloem and axially elongated sclerified cell in the inner secondary phloem. – 11–13: Tangential sections. – 11: Uni- and multiseriate rays in the inner secondary phloem. Crystals (arrow) in subdivided ray cells. – 12: Lipids in ray cells (arrowhead). – 13: Crystals (arrowhead) and starch grains (arrow) in ray cells. Note the sclerification of the parenchyma cell.
Symbols in 8: periderm, phellogen, parenchyma, sieve tubes and companion cells, sclereids, “sclerified cells”, cambial zone.
Scale bar in 8 = 1000 μm; in 9–11 & 13 = 200 μm; in 12 = 100 μm.

IAWA Journal, Vol. 26 (4), 2005480 481Machado et al. — Bark anatomy of Styrax camporum
For legends, see page 480.

IAWA Journal, Vol. 26 (4), 2005482 483Machado et al. — Bark anatomy of Styrax camporum
For legends, see page 480.
IAWA Journal, Vol. 26 (4), 2005482 483Machado et al. — Bark anatomy of Styrax camporum
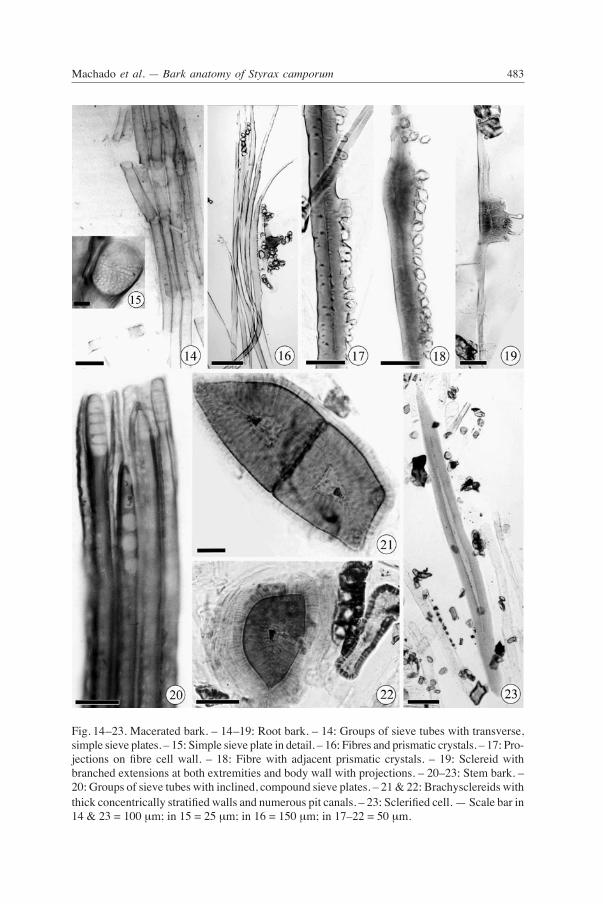
Fig. 14–23. Macerated bark. – 14–19: Root bark. – 14: Groups of sieve tubes with transverse, simple sieve plates. – 15: Simple sieve plate in detail. – 16: Fibres and prismatic crystals. – 17: Pro-jections on fibre cell wall. – 18: Fibre with adjacent prismatic crystals. – 19: Sclereid with branched extensions at both extremities and body wall with projections. – 20–23: Stem bark. – 20: Groups of sieve tubes with inclined, compound sieve plates. – 21 & 22: Brachysclereids with thick concentrically stratified walls and numerous pit canals. – 23: Sclerified cell. — Scale bar in 14 & 23 = 100 μm; in 15 = 25 μm; in 16 = 150 μm; in 17–22 = 50 μm.

IAWA Journal, Vol. 26 (4), 2005484 485Machado et al. — Bark anatomy of Styrax camporum
DISCUSSION
As is evident from the descriptions, the bark of Styrax camporum differs anatomically in the root and stem; these differences are related to the function of the bark in each organ and are further related to the ecological conditions in which the species grows. With regard to periderm development, in the root there is only the original periderm while in the stem there are several periderms. These differences between the extent of periderm development in the root and stem follow normal expectations. The multiple periderms and dead secondary phloem sandwiched between them, constituting a rhy-tidome in the stem, provides protection against extreme temperatures (Miranda et al. 2002). Solereder (1908) and Cutler et al. (1987) already described the presence of numer-ous crystals in the root and stem bark of some species of Styrax, and we also observed this feature in S. camporum. We also observed that parenchyma cells with phenolic compounds had walls thicker than those of adjacent cells without phenolic compounds. According to Roth (1981) there is a strong correlation between the presence of phe-nolic compounds in parenchyma cells and their transformation into sclereids. The lipid contents in ray and phelloderm cells may be associated with resin secretion in the bark. Future studies will reveal their chemical nature and mode of secretion. Sclerenchyma is a common component of root and stem secondary phloem in S. cam-porum. The most remarkable difference between the organs is the occurrence of elon-gated sclerified cells in the stem. These cells differ from normal phloem fibres (observed in the root bark) in that they have very thick walls formed of concentric lamellae, are well-pitted, and the ovoid shape in transverse section. These sclerified cells fit neither the definitions of fibres nor sclereids according to Trockenbrodt (1990). This cell type was already described in the secondary phloem of S. camporum as “fibre-like scleren-chymatous cells” of the nature of rod cells, and having abundant pits (Solereder 1908). Sclerified cells was a term used to denote similar elements in the stem bark of Buchena-via and Terminalia (Combretaceae) (Angyalossy-Alfonso & Richter 1991) and Delar-brea paradoxa (Araliaceae) (Kolalite et al. 2003). In the stem of S. camporum, sclerified cells differentiate near the cambium in the non-collapsed phloem. In the secondary phloem of S. camporum, the most important difference between sclereids and sclerified cells is related to the period in which they are formed. The scleri-fied cells are completely differentiated near the cambium as an important component of the bark pattern. The sclereids develop in dilatating zones from already differentiated parenchyma cells and, therefore, are of secondary origin. According to Roth (1981), the secondary formation of sclereids is more or less a matter of coincidence of certain unpredictable factors induced by dilatation processes. Although Kolalite et al. (2003) stated that sclerified cells are differentiated from the “fusiform axial phloem parenchyma cells”, only an ontogenetic study would confirm their origin in S. camporum. The shape and localization of sclereids is considerably different between root and stem secondary phloem of S. camporum. Groups of astrosclereids are present in the innermost phloem region of the root, which has intercellular spaces. These sclereids dif-ferentiate from axial parenchyma cells adjacent to the phloem fibres. These cells show

IAWA Journal, Vol. 26 (4), 2005484 485Machado et al. — Bark anatomy of Styrax camporum
tip growth and their branches enter intercellular spaces (intrusive growth). In the stem bark, brachysclereids occur in the dilated rays in the outer phloem region and differenti-ate from ray cells that are adjacent to the sclerified cells. Sclerification of ray cells in such zones has been reported in other taxa (Chattaway 1955, 1959; Zahur 1959; Para-meswaran & Liese 1970; Richter 1981, 1990; Angyalossy-Alfonso & Richter 1991). The occurrence of alternation of parenchyma and conductive cells and sclerenchyma in the root secondary phloem may be regarded as an adaptation to the soil conditions of the cerrado, as the soil is compact and dry for the first few meters of depth (Ferri 1979; Oliveira & Marquis 2002). We hypothesize that the continuous tangential bands of sclerenchyma cells may serve as a protective mechanical barrier against the collapse of living cells (parenchyma and sieve tubes) in this harsh soil environment. These tangential bands of fibres may also indicate growth increments in root secondary phloem; further studies may clarify this point. The larger intercellular spaces in the root parenchyma tissue may play an important role in aeration, as there are no lenticels in the root to provide this function. In S. camporum we have observed parallels between vessel-element morphology and sieve-tube-element morphology, such as simple and multiple perforation plates in root and stem wood, respectively (Machado et al. 1997), and simple and compound sieve plates in root and stem bark, respectively. Root secondary phloem characteristics such as shorter and wider sieve tube elements, simple sieve plates with wider sieve pores, and lateral sieve areas poorly developed may be considered as potential indicators of specialization according to Esau and Gifford Jr. (1953), Zahur (1959), and Esau (1979). However, according to Carlquist (1975) sieve tube element morphology seems to be related to conductive efficiency and to the physiology in particular organs, and cannot be explained on the basis of differential evolution rates. According to the same author, some species will evolve simple sieve plates to accommodate volume and rapidity of conduction. So, the occurrence of simple sieve plates in the root secondary phloem of S. camporum may be regarded as an adaptation to the environmental conditions of the cerrado where root growth rate is significantly higher than the stem growth rate (Machado et al. 1997; Felipe 2002). Further comparative studies in Styrax species growing in different habitats may reveal if the parallels between secondary xylem and phloem observed in S. camporum are widespread. The presence of wider rays in the root secondary phloem is presumably related to the transport efficiency between phloem and xylem, but also might be implicated in the storage of photosynthates. Since there is periodic fire in the cerrado, wide rays could hold more starch and lipids and thus provide a much-needed reservoir of carbon to stimulate growth after the shoot might be damaged (leaf loss, partial stem or twig death) by fire. In addition, Alves and Angyalossy-Alfonso (2002) considered wide rays of great importance in the supply of metabolites to the cambium tissue early in the growing season. Finally, additional comparative studies of root and stem bark anatomy of a larger num-ber of species of Styrax from other environments are necessary to determine if such differences between the organs are widespread and of taxonomic and adaptive signifi-cance.

IAWA Journal, Vol. 26 (4), 2005486 487Machado et al. — Bark anatomy of Styrax camporum
ACKNOWLEDGEMENTS
The authors are grateful to Dr. Dirk Koedam and to Dr. Regis B. Miller for useful comments on the manuscript, to the reviewers of the IAWA for providing helpful comments during the revision of the manuscript, to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq - Proc. 30241/2002-0) and Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP: BIOTA Pro-gram - Proc. 00/12469-3) for financial support.
REFERENCES
Alves, E.S. & V. Angyalossy-Alfonso. 2002. Ecological trends in the wood anatomy of some Brazilian species. 2. Axial parenchyma, rays and fibres. IAWA J. 23: 391–418.
Angyalossy-Alfonso, V. & H.G. Richter. 1991. Wood and bark anatomy of Buchenavia Eichl. (Combretaceae). IAWA Bull. n.s. 12: 123–141.
Carlquist, S. 1975. Ecological strategies of xylem evolution. Univ. of California Press, London.Chattaway, M.M. 1955. The anatomy of bark. III. Enlarged fibres in the blood woods (Eucalyptus spp.). Aust. J. Bot. 3: 28–38.Chattaway, M.M. 1959. The anatomy of bark. VII. Species of Eugenia. Trop. Woods 111: 1–14.Coutinho, L.M. 1990. Fire in the ecology of the Brazilian Cerrado. In: J.G. Goldammer (ed.), Fire in the tropical biota; Ecosystem processes and global challenges: 82–105. Springer Verlag, Berlin.Cutler, D.F., P.J. Rudall, P.E. Gasson & R.M.O. Gale. 1987. Root identification manual of trees
and shrubs; a guide to the anatomy of trees and shrubs hardy in Britain and Northern Europe. Chapman & Hall, London.
Eckblad, J.W. 1991. How many samples should be taken? BioScience 41: 346–348.Esau, K. 1979. Phloem. In: C.R. Metcalfe & L. Chalk (eds.), Anatomy of the dicotyledons. Vol. I.
Ed. 2. Oxford University Press, Oxford.Esau, K. & E.M. Gifford Jr. 1953. Comparative structure and possible trends of specialization
of the phloem. Amer. J. Bot. 40: 9–19.Fahn, A. 1979. Secretory tissues in plants. Academic Press, London.Felipe, G.M. 2002. Desenvolvimento de plantas do cerrado: uma experiência pessoal. In: A.L.
Klein (ed.), Eugen Warming e o cerrado brasileiro; um século depois. Ed. UNESP, São Paulo.
Ferri, M.G. 1979. Transpiração nos principais ecossistemas brasileiros e em espécies cultivadas no Brasil. In: M.G. Ferri (ed.), Fisiologia vegetal: 25–73. Edusp, São Paulo.
Franco, A.C. 2002. Ecophysiology of woody plants. In: P.S. Oliveira & R.J. Marquis (eds.), The cerrados of Brazil: 178–197. Columbia University Press, New York.
Freese, F. 1967. Elementary statistical methods for foresters. U.S. Government Printing Office, Washington.
Fritsch, P.W. 2001. Phylogeny and biogeography of the flowering plant genus Styrax (Styracaceae) based on chloroplast DNA restriction sites and DNA sequences of the internal transcribed spacer region. Molec. Phylogen. & Evol. 19: 387–408.
Hutchinson, J. 1973. The families of flowering plants. Ed. 3. Clarendon, Oxford.Jensen, W.A. 1962. Botanical histochemistry. W.H. Freeman, San Francisco.Johansen, D.A. 1940. Plant microtechnique. McGraw-Hill Book Co., New York.Kolalite, M.R., A.A. Oskolski, H.G. Richter & U. Schmitt. 2003. Bark anatomy and intercellular
canals in the stem of Delarbrea paradoxa (Araliaceae). IAWA J. 24: 139–154.Machado, S.R., V. Angyalossy-Alfonso & B.L. de Morretes. 1997. Comparative wood anatomy
of root and stem wood in Styrax camporum (Styracaceae). IAWA J. 18: 13–25.

IAWA Journal, Vol. 26 (4), 2005486 487Machado et al. — Bark anatomy of Styrax camporum
Metcalfe, C.R. & L. Chalk. 1950. Anatomy of the Dicotyledons. Clarendon Press, Oxford.Miranda, H.S., M.M.C. Bustamante & A.C. Miranda. 2002. The fire factor. In: P.S. Oliveira & R.J. Marquis (eds.), The cerrados of Brazil: 51–68. Columbia University Press, New York.Mors, W.B. & C.T. Rizzini. 1966. Useful plants of Brazil. Holden-Day Inc., Amsterdam.Nair, M.N.B. 1995. Some notes on gum and resin ducts and cavities in angiosperms. In: M. Iqbal
(ed.), The cambial derivatives: 317–340. Gebr. Borntraeger, Berlin, Stuttgart.Nakajima, J.N. & R. Monteiro. 1986. Estudos fitogeográficos com espécies de Styrax L. (Styraca-
ceae) dos cerrados brasileiros. Eugeniana 12: 3–10.Oliveira, P.S. & R.J. Marquis. 2002. The cerrados of Brazil. Columbia University Press, New
York.Parameswaran, N. & W. Liese 1970. Mikroskopie der Rinde tropischer Holzarten. Umschau Ver-
lag, Frankfurt.Richter, H.G. 1981. Wood and bark anatomy of Lauraceae. I. Aniba Aublet. IAWA Bull. n.s. 2:
79–87.Richter, H.G. 1990. Wood and bark anatomy of Lauraceae. III. Aspidostemon Rohwer & Richter.
IAWA Bull. n.s. 11: 47–56.Richter, H.G., S.C. Mazzoni-Viveiros, E.S. Alves, A.E. Luchi & C.G. Costa. 1996. Padronização
de critérios para a descrição anatômica da casca: lista de características e glossário de termos. IF Série Registros 16: 1–25.
Roeser, K.R. 1972. Die Nadel der Schwarzkiefer-Massenprodukt und Kunstwerk der Natur. Mikrokosmos 61: 33–36.
Roth, I. 1981. Structural patterns of tropical barks. In: H.J. Braun, S. Carlquist, P. Ozenda & I. Roth (eds.), Handbuch der Pflanzenanatomie. Gebr. Borntraeger, Berlin.
Solereder, H. 1908. Systematic anatomy of the dicotyledons. Transl. by L.A. Boodle & F.E. Fritsch. Clarendon Press, Oxford.
Spongberg, S.A. 1976. Styracaceae hardy in temperate North America. J. Arnold Arbor. 57: 54–73.
Trockenbrodt, M. 1990. Survey and discussion of the terminology used in bark anatomy. IAWA Bull. n.s. 11: 141–166.
Zahur, M.S. 1959. Comparative study of secondary phloem of 423 species of woody dicotyledons belonging to 85 families. Cornell Univ. Agron. Exp. Stn. Mem. 358.
Copyright © 2022 FDOKUMEN