
Community evolution of Neogene micromammals from Langebaanweg ‘E’ Quarry and other west coast...
21
Community evolution of Neogene micromammals from Langebaanweg ‘E’ Quarry and other west coast fossil sites, south-western Cape, South Africa Thalassa Matthews a, ⁎ , Christiane Denys b , John E. Parkington a a Department of Archaeology, University of Cape Town, Rondebosch, 7701, South Africa b M.N.H.N. Department Systematic and Evolution-CP51-FRE CNRS 2695 “Origin, structure and evolution of biodiversity” 55, rue Buffon-F-75005 Paris, France Received 15 July 2005; received in revised form 26 July 2006; accepted 28 August 2006 Abstract This paper provides an overview of the micromammalian palaeocommunities found in fossil bearing palaeontological and archaeological west coast sites dating from the Mio-Pliocene, as represented by the site of Langebaanweg ‘E’ Quarry, and other sites dating from the late Middle Pleistocene, until the late Holocene. Recent excavations at Langebaanweg have resulted in the addition of more murid genera to previously compiled faunal lists. An updated faunal list is presented, and a comparison is made on a generic level between the micromammals from Langebaanweg, and the considerably younger west coast fossil sites of Elands Bay Cave, Steenbokfontein Cave, the Saldanha Bay Yacht Club site and Hoedjiespunt 1, and some modern owl pellet collections. The palaeobiogeographical and palaeoenvironmental significance of the different west coast micromammal communities, and the evolution of the west coast rodent community is examined. Nine of the micromammal genera found at LBW are present in some, or all, of the west coast fossil sites dating from the late Middle Pleistocene until the Holocene. This indicates the endurance of many of the genera present at LBW, and in the Kalahari South West Arid and Namib Regions, from the Mio-Pliocene, up until the present. © 2006 Elsevier B.V. All rights reserved. Keywords: Langebaanweg; Micromammals; Mio-Pliocene; Cape Province; South Africa; West coast 1. Introduction The site of Langebaanweg ‘E’ Quarry (LBW) was discovered during phosphate mining in the Langebaan- weg area, which began in 1943. Langebaanweg's importance as a fossil site was, however, realised by local and international scientists only in 1958 (Singer, 1961; Hendey, 1982). LBW offers one of the largest collections of Mio-Pliocene fossils in Africa, and contains an extremely rich and diverse range of over 230 vertebrate and invertebrate taxa, including numer- ous micromammal bones and teeth (Hendey, 1974, 1981). The site provides an insight into the transitional fauna which inhabited the southwestern Cape in the Mio-Pliocene, a period when many relict Miocene taxa are found together with genera which inhabit the southwestern Cape, and southern Africa, today. LBW is also the site where the earliest recorded fynbos pollen has been recovered (Scott, 1995), and Cowling and Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332 – 352 www.elsevier.com/locate/palaeo ⁎ Corresponding author. Fax: +27 21 789 2814. E-mail address: [email protected] (T. Matthews). 0031-0182/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2006.08.015
Transcript of Community evolution of Neogene micromammals from Langebaanweg ‘E’ Quarry and other west coast...

alaeoecology 245 (2007) 332–352www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, P
Community evolution of Neogene micromammals fromLangebaanweg ‘E’ Quarry and other west coastfossil sites, south-western Cape, South Africa
Thalassa Matthews a,⁎, Christiane Denys b, John E. Parkington a
a Department of Archaeology, University of Cape Town, Rondebosch, 7701, South Africab M.N.H.N. Department Systematic and Evolution-CP51-FRE CNRS 2695 “Origin, structure and evolution of biodiversity” 55,
rue Buffon-F-75005 Paris, France
Received 15 July 2005; received in revised form 26 July 2006; accepted 28 August 2006
Abstract
This paper provides an overview of the micromammalian palaeocommunities found in fossil bearing palaeontological andarchaeological west coast sites dating from the Mio-Pliocene, as represented by the site of Langebaanweg ‘E’ Quarry, and othersites dating from the late Middle Pleistocene, until the late Holocene. Recent excavations at Langebaanweg have resulted in theaddition of more murid genera to previously compiled faunal lists. An updated faunal list is presented, and a comparison is made ona generic level between the micromammals from Langebaanweg, and the considerably younger west coast fossil sites of ElandsBay Cave, Steenbokfontein Cave, the Saldanha Bay Yacht Club site and Hoedjiespunt 1, and some modern owl pellet collections.The palaeobiogeographical and palaeoenvironmental significance of the different west coast micromammal communities, and theevolution of the west coast rodent community is examined. Nine of the micromammal genera found at LBWare present in some, orall, of the west coast fossil sites dating from the late Middle Pleistocene until the Holocene. This indicates the endurance of many ofthe genera present at LBW, and in the Kalahari South West Arid and Namib Regions, from the Mio-Pliocene, up until the present.© 2006 Elsevier B.V. All rights reserved.
Keywords: Langebaanweg; Micromammals; Mio-Pliocene; Cape Province; South Africa; West coast
1. Introduction
The site of Langebaanweg ‘E’ Quarry (LBW) wasdiscovered during phosphate mining in the Langebaan-weg area, which began in 1943. Langebaanweg'simportance as a fossil site was, however, realised bylocal and international scientists only in 1958 (Singer,1961; Hendey, 1982). LBW offers one of the largest
⁎ Corresponding author. Fax: +27 21 789 2814.E-mail address: [email protected] (T. Matthews).
0031-0182/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2006.08.015
collections of Mio-Pliocene fossils in Africa, andcontains an extremely rich and diverse range of over230 vertebrate and invertebrate taxa, including numer-ous micromammal bones and teeth (Hendey, 1974,1981). The site provides an insight into the transitionalfauna which inhabited the southwestern Cape in theMio-Pliocene, a period when many relict Miocene taxaare found together with genera which inhabit thesouthwestern Cape, and southern Africa, today. LBWis also the site where the earliest recorded fynbos pollenhas been recovered (Scott, 1995), and Cowling and

333T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
Richardson (1995) note that the abundance of fynbosforms at this time suggests the start of the rise topredominance of the fynbos flora in the Cape Province.The fossils found within the LBW sediments enable usto ascertain how the environmental and climatic changestaking places during the Mio-Pliocene affected theevolution of fauna in the area.
LBW is the only site in the western Cape Provincerepresenting the Mio-Pliocene transition, a slice of timewhen modern murid genera were emerging, as indicatedby both palaeontological and molecular phylogenystudies (Denys, 1996, 1999). LBW is therefore a crucialsite for our understanding of the evolution and dispersionof modern rodent taxa from southern Africa. Similaritiesin terms of genera held in common between the LaetoliBed rodent faunas with those from Makapansgat and
Fig. 1. The locality of the w
Jägersquelle have been noted (Denys et al., 1987, 1999).A comparison of rodent genera common to LBW andother southern African sites is presented further on, andprovides information on the evolution of genera inSouthern Africa.
A comparison between various west coast fossil sitesincluding Langebaanweg and several other sites fromthe west coast area dating from the terminal Pleistoceneand into the Holocene was undertaken in order to tracechanges in west coast micromammal communities, andin changing palaeoenvironmental and biogeographicalconditions on the west coast, from the Mio-Pliocene,until the present day. The study also provided insightinto the development of endemism in southern Africaand the evolution of typically southern African muridand bathyergid species.
est coast fossil sites.

334 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
1.1. Background to the site of Langebaanweg
Today, LBW and the other west coast fossil sites aresituated close to, or along, the southwest coast of SouthAfrica (Fig. 1) in an area dominated by the fynbos of theStrandveld and Sandveld, which supports a well knownmicromammal fauna (De Graaff, 1981; Skinner andSmithers, 1990; Stuart and Stuart, 2001). The open,scrub vegetation of the Strandveld fynbos includeswoody members such as Rhus, Euclea, Chrysanthe-moides and Olea, and generally grows in sandycalcareous soil (Rutherford andWestfall, 1986;Manningand Goldblatt, 1996). The most characteristic vegetationtype on the west coast is the Sandveld, which is domi-nated by restionaceous genera and small shrubs such asPhylica stipularis, Staavia radiata, Stoebe plumosaand Metalasia densa (Manning and Goldblatt, 1996).
The Langebaanweg ‘E’ Quarry sediments form partof the Varswater Formation which is part of the SandveldGroup. The Varswater Formation is underlain by theMiddle Miocene Elandsfontyn Formation, or by Neo-proterozoic to Cambrian bedrock (Roberts, in press).Overlying the Varswater formation is the calcareousaeolianite of the Langebaan Formation, or the quartzoseaeolian sands of the Springfontein Formation (Dingleet al., 1979; Roberts, in press). The Varswater Formationsuccession consists of four members, the oldest of whichis the Langeenheid Sandy Clay Member Konings VleiGravel Member. Above the Konings Vlei GravelMember is the Langeberg Quartzose Sand Member(LQSM), which in turn is overlain by the youngerMuishhond Pelletal Phosphate Member (MPPM). TheLQSM and the MPPM are the main fossil bearingdeposits of the formation. The latter contained thecommercially exploitable phosphate ore mined at LBW.The LQSM represents a number of different depositionalenvironments, and although some sub-aqueous deposi-tion of fossils does appear to have taken place, themajority of fossils are thought to have been accumulatedon a flood plain (Hendey, 1981). TheMPPM deposits arethought to represent sediments accumulated on river-banks, and in river channels (Hendey, 1981).
The MPPM and LQSM represent two separate,depositional events but the time period over which thesetwo horizons were deposited, and the age differencebetween them is uncertain as the stratigraphic relation-ship between the two horizons is complicated andpoorly understood. The MPPM has been divided intothe fossiliferous bed 3a, which consisted of a northerlyand southerly part, namely bed 3aN and bed 3aS, as wellas other undifferentiated, largely non-fossil bearing beds(Hendey, 1981). Bed 3aS is thought to have been laid
down during a northwards shift of the river's course, andbed 3aN by yet another, later, northwards shift of theriver. Bed 3aN has been the main focus of recentresearch into the ungulates at LBW (Franz-Odendaal,2002) and is less problematic than bed 3aS in that thereis no evidence to suggest that bed 3aN contains materialderived from the LQSM, a possibility which exists in thecase of bed 3aS (Hendey, 1976).
The West Coast Fossil Park was established at thesite of the Langebaanweg phosphate mine, and, underthe direction of the park, excavations were resumed in“E” quarry in 1998 in an area which is thought torepresent an extension of the river channel deposits ofbed 3aN of the MPPM.
1.2. Previous research at Langebaanweg
Research carried out on the murids and mole ratsfrom LBW in the 1970's up until the 1990's generallyconcentrated on the identification and description ofnew species (Pocock, 1976, 1987; Denys, 1990a, 1991,1994a,b, 1998). Langebaanweg is the site of first oc-currence in South Africa of a number of taxa; includingtwo species of Bathyergidae and a number of othergenera including Aethomys, Dendromus, Mystromys,Acomys, Euryotomys, Rhabdomys, Zelotomys andThallomys (Pocock, 1976, 1987; Denys, 1990a, 1991,1994a,b, 1998; Matthews, 2004).
Previous palaeoecological work at the site hasmainly been based on the larger mammals. For ex-ample, the fact that bed 3aS (MPPM) is dominated bygrazing alcephalines, and bed 3aN (MPPM) by brows-ing giraffids, has led to the suggestion that grasslandincreased at the expense of forest and bush (Klein,1981). It has also been suggested, based on dentalabnormalities (enamel hypoplasia) on the teeth ofmany of the Sivatheres and a few of the other ungulatespecies from the MPPM, that grassland increased atthe expense of woodland, causing nutritional stress(Hendey, 1981). The most recent research on potentialcauses of hypoplasia on several species at LBW wascarried out by Franz-Odendaal's (2002) study of serialδ18O isotopes along the crowns of molars of Si-vatherium hendeyi. Franz-Odendaal (2002) concludedthat normal seasonal cycles at LBW were interspersedwith periods of reduced rainfall and drought.
Until the research carried out by Franz-Odendaal(2002), it was thought that C4 grasslands may haveextended as far south as LBW during the Early Pliocene,and the climatic regime present at LBW was uncertain.It was thought that a summer rainfall pattern prevailed atLBW during the time of deposition of the LQSM and

335T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
MPPM, although the suggestion was made that thispattern may have begun to change towards the currentwinter rainfall regime which prevails today in theWestern Cape (Hendey, 1976, 1981). Franz-Odendaal's(2002) research indicated that the expected grazers (e.g.alcelaphine, hippopotamus, and rhinoceros), as well asbrowsing species, showed δ13C values which indicatedthat LBW was a C3 dominated environment. Thepresence of cool growing C3 grasses at LBW duringthe deposition of the Varswater sediments indicates thatthe present-day climatic regime of winter wet/summerdry was established at this time (Franz-Odendaal, 2002).Certain other modern climatic/environmental features ofthe west coast, such as the establishment of the Agulhascurrent and the Benguela Upwelling System, alsooccurred during the Mio-Pliocene around 5 Ma (Siesser,1980; Hendey, 1983) and probably led to the earlydevelopment of a Mediterranean climate, which wasessentially modern. Ward and Corbett (1990) place theestablishment of the Benguela current somewhat earlier,at 10–7 Ma. The effect of the cold, upwelling Benguelacurrent and the persistent, strong South Atlantic high-pressure system (a stable anticyclone which lies abovethe Atlantic Ocean at 30° S) are two of the main causesfor the present aridity along the south-western coast, theNamib and the dryness of the interior of the sub-continent (Siesser, 1978; Coetzee, 1978, 1980; Coetzeeand Rogers, 1982).
2. Material
Although one of the authors, namely CD, made abroad study of the different micromammal taxa at LBW,only a sample of the available micromammal materialfrom the LQSM and MPPM was studied. Previousstudies did not look at the taphonomy of themicromammals and no palaeoecological analysis ofthe micromammal community was carried out on thesite. This paper presents some of the results from ataxonomic and taphonomic study of the micromammalsrecovered from the recent excavations of bed 3aN(MPPM), and most of the sorted murid and bathyergidmaterial recovered from the LQSM during miningoperations.
The LBWmicromammals recovered from the LQSMand MPPM sediments retrieved during mining at LBWfrequently comprise assemblages whose exact prove-nance, and method of recovery (sieve size is particularrelevant if all micromammal fossils are to be recoveredfrom sediments), is unknown. Many fossils were surfacecollected, and generally, no concerted effort was madeto recover all the micromammals from an area.
The recently excavated micromammal assemblages,listed separately from the other MPPM micromammalsin Table 1 as ‘MPPM (F)’ come from the excavation areaopened in 1998. These sediments were excavated usingmethods in which particular attention was paid to therecovery of micromammals. The recently excavatedhorizons of the MPPM will be referred to as the MPPM(F) from this point onwards, in order to differentiatethese fossils from those recovered from the MPPMduring mining. The micromammals from two adjacentsquares in the recent excavation area (Unit F10 and UnitF11) were analysed. For the purposes of presentation ofresults, they have frequently been added together in thefollowing discussion.
The LQSM units mentioned in this article come fromseveral areas within the LQSM. The majority of thesecame from the East stream area. East stream (ES) wasthe name given to a modern, narrow channel thatdeveloped along the east wall of the mine as a result ofmining below the water table (Hendey, pers. comm.).All the fossils from this area were retrieved from thefine, white sand characteristic of the LQSM — nostratification was observed. East stream formed part ofthe LQSM sediments which are thought to have beenpartly subaerially and partly subaqueously deposited, onthe floodplain of a river (Hendey, 1981). The unitswhich contained relatively large micromammal assem-blages were Dump 2, the Elephant site, and Square 1(both the latter units were found within the east streamarea). Dump 2 was the LQSM unit which provided thelargest assemblage of micromammals. The micromam-mals from this unit were recovered by sieving ofsediments from a mine dump and are thus likely tocontain micromammals from different horizons andareas within the site. The Elephant site (a relativelysmall area of a few m2 which surrounded some elephantfossils) as well as the unit Square 1, represent relativelydiscrete areas within the East stream area of the LQSM.Tex's Pit 1 and Tex's Pit 4 represent micromammalsrecovered from actual excavations. These pits, whichwere excavated in the East stream area, were excava-tions which were usually centered around a surface find(Hendey pers. comm.). The largest of these pits wasprobably no more than a few metres square and a metredeep (Hendey pers. comm.). The micromammals fromthe unit labelled ‘peat bed’ come from a peat bed whichis described by Hendey (1981) as a salt-marsh deposit.All the micromammal fossils from this horizon showeda dark brown discolouration.
The MPPM (F) units are believed to come from apalaeo-river channel and appear to have been transportedover short distances in a medium–low or low energy

336 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
environment (Matthews, 2004). The generally low fre-quency of bones showing evidence of water-rounding,together with a lack of the degree of breakage associ-ated with prolonged water transport, suggests that theywere not exposed to extended periods in the waterbefore being buried well below the ground's surface.The LQSM units investigated in this thesis, with theexception of PB, are thought to have accumulated in afloodplain environment. The cranial bones from theLQSM units show less cranial breakage than those fromtheMPPM (F), despite the fact that the recovery methodsused in their recovery were often potentially destructive.Like the MPPM (F) assemblages, the LQSM unitsappear to have been buried relatively soon after depo-sition, and do not appear to have been exposed to pro-longed or high energy alluvial transport, or any verydestructive post-depositional forces. Generally, preser-vation in the LQSM units is better than in theMPPM (F).A taphonomic analysis done on this site is detailed inMatthews (2004).
In order to trace changes in west coast micromammalcommunities over time, several other micromammal-bearing west coast fossil sites have been included in thispaper, namely the Saldanha BayYacht Club site (Manthi,2002), Elands Bay Cave (Matthews, 1998, 1999),Steenbokfontein (Avery, 1999), and Hoedjiespunt 1(HDP1) (Matthews, 2004; Matthews et al., 2005), detailsof which are given in the following section.
Data from Bolt's Farm (Sénégas and Avery, 1998;Sénégas, 2000) and the Namibian faunas (Pickford andMein, 1988; Pickford et al., 1994; Senut et al., 1992), aswell as a compilation of southern African Pliocene andPleistocene fossil sites by Denys (1999), are included inthis paper. Also included is an inventory of Capemicromammals published by Avery (1982), whichincludes faunal lists of micromammals from modernowl pellet assemblages from the west coast (collectedfrom the West Coast National Park and Steenbokfonteinfarm), and the general biogeographical analysis ofRautenbach (1978).
2.1. Background to the other west coast fossil sites
Fig. 1 shows the location of the west coast fossil sites.The Saldanha Bay Yacht Club site (SBYC) repre-
sents a rich micromammal assemblage accumulated in asolution cavity in Langebaan Formation aeolianites. Thesediments have been radiocarbon dated to 15540±70 yrB.P (Manthi, 2002). A complete taphonomic analysishas indicated that the assemblage was accumulated byan owl, the barn owl being the most likely candidate(Manthi, 2002).
The two other sites, namely Elands Bay Cave (EBC)and Steenbokfontein Cave (STBKC), are cave siteswhich contain archaeological deposits which range inage from the Terminal Pleistocene to the Late Holocene.EBC was excavated in units defined by differences instratigraphy and micromammals were found in unitswhich covered the period from 13260 BP to 300 BP. Thebarn owl (Tyto alba afinis) was found to have beenresponsible for the deposition of the micromammals insome horizons of the site, and the spotted eagle owl(Bubo africanus), an unidentified viverrid, and a gianteagle owl (Bubo lacteus) were identified as potentialcontributors to the micromammal assemblages invarious other depositional units (Matthews, 1998, 1999).
The archaeological cave site of Steenbokfontein is setin a prominent sandstone outcrop which faces towardsthe sea, which lies some 3 km away, and is surroundedby relatively flat terrain, and low sand dunes. Themicromammalian sample studied was taken from thefive uppermost layers. Layer 0 at the very top has notbeen dated, but layer 1 to 4b have produced datesbetween 2200 and 6000 BP, and layer 4b yields datesfrom 3990 BP to 6070 BP (Avery, 1999). Although ataphonomic analysis of the STBKC micromammals hasnot been done, the great density of the micromammalfossils and the extremely low degree of breakage of themicromammals observed at this site strongly suggeststhat a barn owl was responsible for their accumulation(Jerardino, pers. comm.). The modern barn owl pelletassemblages seen in Figs. 3 and 4 were collected on thesame rocky outcrop.
Hoedjiespunt 1 (HDP1) consists of a fossil hyaena lairwhich contained a rich assortment of ungulate andcarnivore fossils. On the basis of stratigraphic observations,and the dates obtained, the site is thought to date to sometime between 200000 to 300000 yr ago (Stynder et al.,2001). A taphonomic investigation indicated that the mi-cromammal assemblages recovered from the site wereaccumulated by a barn owl. The taphonomic analysis doneon this site is detailed in Matthews et al. (in press-a), and(Matthews, 2004). The results of a palaeoecological studyof the micromammal assemblages may be found in(Matthews, 2004) and (Matthews et al., 2005).
3. Methods
In order to identify the predator/s or agent/sresponsible for a fossil accumulation, and assess post-depositional alteration of a fossil assemblage, it hasbecome customary for taphonomic analyses of micro-mammal assemblages to look at cranial and post-cranialbreakage, skeletal element abundance, digestion of

Table 1The Rodent faunal list at Langebaanweg (MPPM faunal list after Hendey, 1981; Pocock, 1976; Denys, 1990a,b, 1991, 1994a,b, 1998; MPPM (F)faunal list after Matthews, 2004)
Rodentia LQSM MPPM MPPM (F) (New data)
HystricidaeGen. and sp. not det. A (Hendey, 1981) ✓ X XGen. and sp. not det. B (Hendey, 1981) X ✓ X
BathyergidaeCryptomys broomi (Denys, 1998) ✓ ✓ XBathyergus hendeyi (Denys, 1998) ✓ ✓ ✓
MyoxidaeGraphiurus sp. (Hendey, 1981a) ✓ ✓ X
MuridaeDendromurinaeDendromus averyi (Denys, 1994a) ✓ ✓ ✓Dendromus darti (Denys, 1994a) ✓ ✓ XDelanymyinaeStenodontomys saldhanae (Denys, 1994b) ✓ ✓ ✓MystromyinaeMystromys pocockei (Denys, 1991) ✓ ✓ ✓Mystromys hausleitneri (Denys, 1991) ✓ ✓ ✓Deomyinae Acomys mabele (Denys, 1990b) ✓ ✓ ✓Acomys sp. (large) (indet.) M1 ✓ – XGerbillinaeDesmodillus sp. nov. ✓ ✓ ✓Murinae ✓ ✓ ✓Euryotomys pelomyoides (Pocock, 1976)Aethomys adamanticola (Denys, 1990a) ✓ ✓ ✓Aethomys modernis (Denys, 1990a) ✓ ✓ ✓
Recently identified speciesAethomys modernis-like (indet.) M1 X X ✓Aethomys adamanticola-like (indet.) M1 X X ✓Aethomys sp. 1 (indet.) M1 ✓ X ✓Aethomys sp. 3 (indet.) M1 X X ✓Aethomys sp. 4 (indet.) M1 X X ✓Aethomys intermed. (indet.) M1 X X ✓Rhabdomys intermed. (indet.) M1 – X ✓Rhabdomys sp. 1 (indet.) M1 ✓ X XRhabdomys sp. 2 (indet.) M1 X X ✓Mus or Acomys-like sp. (indet.) M1 and M1 # X #Small Aethomys or small Rhabdomys (indet.) M1 # X #Thallomys sp. (indet.) M1 X X ✓Zelotomys-like sp. (indet.) M1,2 – X #Zelotomys sp. (indet.) M1 X X ✓
Key: ✓=species present, X = species absent, # = more than one indet. species is included in this category.
337T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
bones and teeth, and to record taphonomic features ofthe assemblages which provide information on theprocesses which have affected the fossils since deposi-tion (e.g. Andrews, 1990; Fernandez-Jalvo andAndrews, 1992; Denys, 1997; Matthews, 1999, 2000;Manthi, 2002). A taphonomic analysis was thereforecarried out on the LBW micromammal assemblages, aswell as those from the site of Hoedjiespunt1 (HDP1)(Matthews et al., in press-a), following the methodol-
ogies established by Andrews (1990) and Fernandez-Jalvo and Andrews (1992). Taxonomic identificationwas accomplished using the teeth and cranial bones.These results are detailed in Matthews (2004). The samegeneral analytical techniques were employed in theanalysis of the micromammal material from Hoedjie-spunt 1 (Matthews, 2004; Matthews et al., 2005), ElandsBay Cave (Matthews, 1998, 1999) and the SaldanhaBay Yacht Club site (Manthi, 2002).
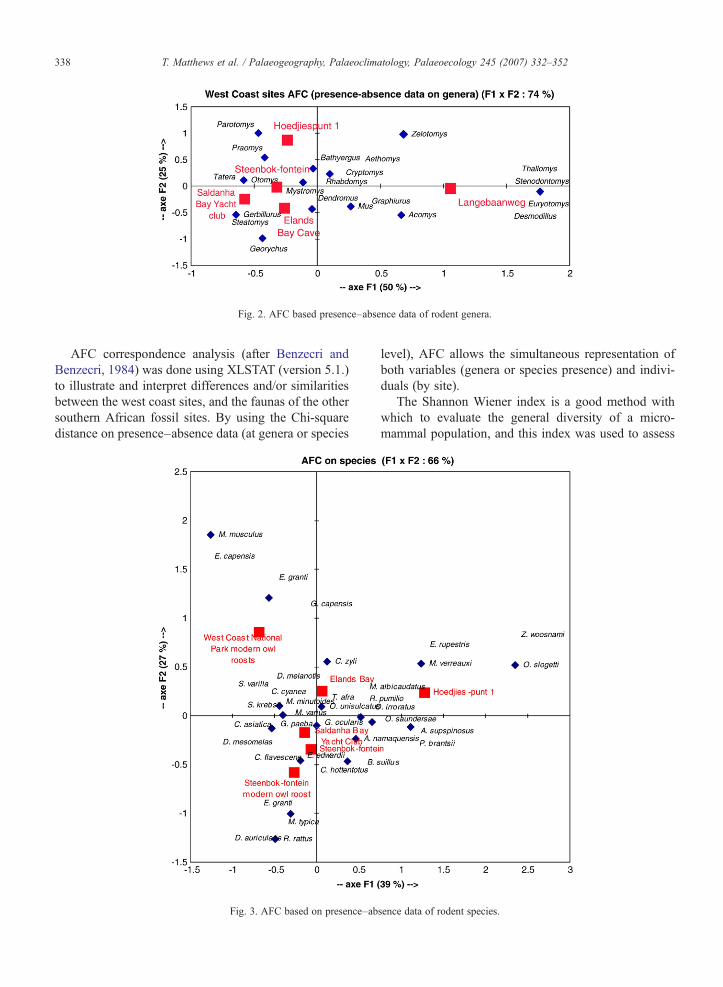
Fig. 2. AFC based presence–absence data of rodent genera.
338 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
AFC correspondence analysis (after Benzecri andBenzecri, 1984) was done using XLSTAT (version 5.1.)to illustrate and interpret differences and/or similaritiesbetween the west coast sites, and the faunas of the othersouthern African fossil sites. By using the Chi-squaredistance on presence–absence data (at genera or species
Fig. 3. AFC based on presence–ab
level), AFC allows the simultaneous representation ofboth variables (genera or species presence) and indivi-duals (by site).
The Shannon Wiener index is a good method withwhich to evaluate the general diversity of a micro-mammal population, and this index was used to assess
sence data of rodent species.

Fig. 4. AFC based on presence–absence data of rodent genera from the west coast fossil sites and the biotic zones (after Rautenbach, 1978). Key tobiotic zones: NAM = Namid, SS = Savanna grasslands, SWA = South West Arid, FOR = forest, SWC = South West Cape Fynbos.
Table 2Rodent species richness in the LQSM and MPPM (F) (after Matthews,2004, Table 8.1, page 143)
Unit Species richness NISP
LQSM unitsDump 2 ±13 417Elephant site ±11 104
339T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
general diversity at the fossil sites. General diversitytakes into account both the number of taxa present, andthe relative frequency (evenness of representation) ofeach taxon (Cruz-Uribe, 1988). The Shannon Wienerindex is calculated using the formula:
H ¼ −X
PilogPi;
where Pi=n /N, that is, the proportion of the totalsample represented by each species.
The systematic classification used in this paper followsthe revised systematic checklist of Bronner et al. (2003).
Square 1 7 49Tex's Pit 1 7 27Peat Bed 4 13Tex's Pit 4 3 5East stream (bed 2) 3 3LQSM (all units combined) ±16 619
MPPM (F) unitsF10 ±19 131F11 13 66Unit F10 and F11 combined ±20–22 197
4. Results
4.1. The Langebaanweg Rodentia
Table 1 presents a list of the various rodents found atLBW. Species identified during previous research on theMPPM and LQSM micromammals from the area areshown separately from the new species identified by
Matthews (2004) which are noted as ‘new data’. Theterm ‘indet.’ (indeterminate) following the genus nameof the new species listed in Table 1 is used to indicate

Table 3Rodent diversity in the LQSM and MPPM (F) as indicated by the Shannon Wiener index of general diversity (H ) (after Matthews, 2004, Table 8.2,page 145)
LQSM units MPPM (F ) units
Dump 2 Elephantsite
Square 1 PeatBed Tex's Pit 1 Tex's Pit 4 East stream(bed 2)
F10 F11 F10 andF11
Shannon–Wiener index of diversity (H ) 0.61 0.78 0.69 0.47 0.65 0.46 0.48 0.78 0.71 0.79
340 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
that these species were identified to genus only. Thenew, indeterminate species are noted as being identifiedfrom an upper (M1) or lower (M1) first molar, or both.The majority of these previously unrecorded species comefrom the recently excavated MPPM (F) horizons at LBW.
Further research aimed at increasing the sample sizeof many species currently represented by as little as onemolar is underway, and should confirm the exactnumber of genera represented at LBW. Some fourteengenera have been identified at LBW to date. As may beseen in Table 1, there are a number of multi-specificgenera, a relatively unusual feature. The Dendromurinaeand Mystromyinae are represented by two species,Aethomys by what may be three species, and Rhabd-omys by two (Matthews, 2004).
4.2. A comparison between the Langebaanweg muridsand some southern African fossil sites
The only species that LBW holds in common withother Early to Middle Pliocene fossil sites in southernAfrica are Mystromys hausleitsneri, and possiblyMystromys pocockei. The latter is an extinct species ofMystromys, which was described from LBW (Denys,1991). A dwarf species of Mystromys has been found atMakapansgat (in the Makapansgat rodent corner in situ,and the Makapansgat limework dumps) and is as yetundescribed, though it could prove to be the same as M.pocockei (Denys, 1991). At LBW, M. pocockei wasfound in conjunction with the larger species, M.hausleitneri, which is also found at Makapansgat,Sterkfontein Valley, and at the younger Late Plioceneto Early Pleistocene sites of Kromdraai, and theSchurweberg deposits (De Graaff, 1961; Denys, 1991;Avery, 2000a). The presence of M. hausleitneri at LBWindicates that this species survived and flourished over aconsiderable period of time as it is found at Sterkfonteinat 2.8 Ma (De Graaff, 1960; Avery, 2000b) where itdominates the fossil assemblages, together with Pa-laeotomys gracilis (De Graaff, 1960). Today, Mystr-omys albicaudatus is the only living representative ofthe Mystromyinae.
The genus Euryotomys, which was first identified atLBW (Pocock, 1976), has been found at only one otherfossil site, namely the Waypoint 160 deposits fromBolt's farm, which are thought to date to ∼4–5 Ma(Sénégas and Avery, 1998). The phylogenic relationshipbetween Euryotomys and the Otomyinae is controver-sial, with one school of thought suggesting that Eur-yotomys is an ancestor of the Otomyinae (Sénégas andAvery, 1998; Sénégas, 2001), while Denys (1989)suggests on the basis of morphological and biogeo-graphical evidence that E. pelomyoides is not anantecedent of the Otomyinae, but may rather share acommon murid ancestor with Saidomys afarensis.
The presence of Acomys mabele at LBW is evidencethat this genus was quite distinct by 5 million years ago(Denys, 1990b). The two new species of Aethomys fromLBW, identified and described by Denys (1990a),represent the oldest Aethomys species found to date inAfrica. Mus has yet to be positively identified anddescribed at LBW as what was initially identified asMus, was later found to be Acomys (Denys, 1990b).The Gerbillinae are represented by only one species atLBW, an undescribed Desmodillus sp., and there areapproximately six murine genera.
Of all the rodent genera making a first appearance inthe South African fossil record at LBW, that is, thebathyergid Bathyergus, and the murids, Aethomys,Mystromys, Acomys, Euryotomys, Rhabdomys, Zelot-omys and Thallomys, only Euryotomys is extinct. Theonly other extinct genus found at LBW is Stenodont-omys. Stenodontomys survived for a remarkably longperiod, as indicated by the fact that this species has beenfound in Late Miocene breccias from the OtaviMountains, at LBW, the site of Nosib1 (Ca 3 Ma.), andin deposits at Berg Aukas, Aigamas II, and Uisib I (alsoin the Otavi Mountains), dating to 1.5 Ma (Senut et al.,1992; Pickford et al., 1994; Denys, 1999). Stenodont-omysmakes a first appearance in breccias from the Otavimountains in Namibia in what Pickford et al. (1994) referto as ‘Late Miocene’ deposits. These breccias contain amixture of Miocene and Early Pliocene micromammalspecies.

Table 4Composition of the micromammalian fauna of the west coast fossil sites
Hoedjies-punt 1(HDP1)
Saldanha BayYacht Club(SBYC⁎)
Elands BayCave (EBC⁎⁎)
Steenbok-fonteinCave (STBKC⁎⁎⁎)
Steenbok-fontein owlroost samples⁎⁎⁎
West Coast National Parkowl roost samples⁎⁎⁎⁎
Age of site 200000–300000 BP
∼15540 BP 13600–300 BP 6000–2200 BP Modern Modern
ChyrsochloridaeC. zyli √C. asiatica √ √ √ √ √E. granti √ √E. capensis √
MolossidaeT. aegyptiaca √
RhinolophidaeR. clivosus √ √
SoricidaeC. cyanea √ cf. C. cyanea √ √ √ √C. flavescens √ √ √ √M. varius √ √ √ √ √ √S. varilla √ √ √ √ √
MacroscelididaeE. granti √ √E. edwardii √ √ √ √E. rupestris √ √
MuridaeD. melanotis √ √ √ √ √D. mesomelas √ √ √ √S. krebsii √ √ √ √ √G. paeba √ √ √ √ √T. afra √ √ √ √ √ √D. auricularis √M. albicaudatus √ √ √ √A. subspinosus √ √ √M. verreauxi √ √R. pumilio √ √ √ √ √ √A. namaquensis √ √ √ √Z. woosnami √G. ocularisM. minutoides √ √ √ √G. ocularis √ √O. irroratus √ √ √ √ √ √O. saundersae √ √ √ √O. unisulcatus √ √ √ √ √ √O. slogetti √P. brantsii √ √R. rattus √M. typica √ √M. musculus √
BathyergidaeB. suillus √ √ √ √C. hottentotus √ √ √ √ √G. capensis √ √
(continued on next page)
341T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352

Table 4 (continued )
Hoedjies-punt 1(HDP1)
Saldanha BayYacht Club(SBYC⁎)
Elands BayCave (EBC⁎⁎)
Steenbok-fonteinCave (STBKC⁎⁎⁎)
Steenbok-fontein owlroost samples⁎⁎⁎
West Coast National Parkowl roost samples⁎⁎⁎⁎
Age of site 200000–300000 BP
∼15540 BP 13600–300 BP 6000–2200 BP Modern Modern
MNI 52 804 315 3457 1351 5971H 0.91 2.05 0.75 (barn owl
assemblages only)2.31 2.21 1.78 (average of 13 roost
assemblages)Range – 1.95–2.16 0.51–1.94 2.11–2.43 1.95–2.48 1.41–2.25
After ⁎Matthews, 2004, Table 13.1, page 220, ⁎⁎Manthi, 2002, Table 7.2.2 and Table 7.2.3, page 80–81, ⁎⁎⁎ Avery, in press, ⁎⁎⁎⁎ Avery, 1999,Table 4, page 155, *****Avery, 1992, Table 3, page 391.
342 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
Many genera which make a first appearance at LBWappear to have spread northwards (e.g. Zelotomys,Rhabdomys, Aethomys) (Denys, 1999), and there is noclear evidence of immigration into southern Africa atthis time by central or eastern African species. Somegenera, such as Bathyergus and Acomys, have remainedin the area, and may have evolved into the extant,endemic Bathyergus suillus and Acomys subspinosus.
4.3. Rodent species richness and diversity at LBW
Table 2 shows rodent species richness (the number ofdifferent species) for the LQSM and the two MPPM (F)units, Unit F10 and Unit F11. The number of individualspecimens (NISP) indicates the total number of M1 andM1 molars, both isolated and in situ teeth.
The number of species found in Dump 2, the Elephantsite, and the MPPM (F) units, F10 and F11, is an under-estimate as a small number of new, undescribed specieshave been put together under a general heading, such as‘small Aethomys or small Rhabdomys (indet.)’. Thesoricid and macroscelid populations from LBW have notbeen studied in any detail to date, and these taxa are thusunfortunately missing from the micromammalian faunallist.
Table 3 gives the Shannon Wiener index of diversity(H ) for various units from different areas of the LQSM,and for the MPPM (F) units, F10 and F11, both sin-gularly and combined.
Using the Shannon Wiener index to calculatediversity presents a somewhat different picture to whena simple count of the number of species is used. Whendiversity is quantified using this index, the Elephant siteis seen to show the same diversity as the MPPM (F) unit,F10, and the diversity in F11 is seen to be relatively closeto that of Square 1. The units of unsatisfactorily smallsample size, namely Peat Bed, East Stream, Tex's Pit 1and 4, and East stream (bed 2) show a generally lowdiversity. Dump 2 shows a diversity that is considerablylower than that of the MPPM (F) units and other LQSM
units such as the Elephant site and Square 1 which comefrom more discrete areas. The Shannon–Wiener indexindicates that Dump 2 shows a surprisingly low diversityfor an assemblage of large sample size that is thought tocontain a mixture of assemblages from different areas.There are a number of possible explanations for the lowdiversity shown by Dump 2, which provided one of thelargest fossil assemblages, and one of the most likely isthat the methods used during recovery have affectedspecies richness in this assemblage. Hendey (pers.comm.) noted that micromammals are likely to beunder-represented in the dump samples owing to themesh size of the screens used, and the interests of theperson(s) doing the screening, as the recovery of largemammal bones tended to be the main focus of sievingactivities. It would appear that the low diversity in Dump2 may be an artefact of recovery. It is perhaps significantthat a higher species diversity is observed in the unitswhich came from a relatively small, discrete area ofexcavation, where there may have been a more concertedeffort to recover the micromammals within an area. Aninvestigation of the species diversity in the MPPMdeposits recovered during mining is currently underway,and should greatly contribute towards assessing whetherdifferences in species diversity between the LQSM andMPPM are an artefact of the recovery methods used, orwhether they reflect a real difference in species diversitybetween the two members.
4.4. An overview of the micromammalian fauna of theother west coast fossil sites
Table 4 shows the composition and diversity of themicromammal faunas of the other west coast fossil sitesand the assemblages from modern owl pellet collectionsfrom the West Coast National Park and Steenbokfonteinfarm.
The Steenbokfontein cave site and the Steenbokfon-tein modern owl roost show the highest general diversity(H ). EBC shows the lowest general diversity, but this

343T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
figure includes only those depositional units depositedby a barn owl. The Shannon Weiner Index for the otherassemblages, which appear to have been accumulatedby predators other than the barn owl, averages 1.67(Matthews, 1998; Avery, in press). The general diversityof HDP1 is relatively low, but not as low as the owl-accumulated EBC assemblages. HDP1 sample size issmall and this factor may well have led to a decrease indiversity. The small sample size precludes any definiteconclusions being reached until a larger sample size isanalysed. As things stand, HDP1 shows a relatively, butnot anomalously, low diversity relative to the otherfossil sites. The fossil sites of the Saldanha Bay YachtClub site and Steenbokfontein Cave show a diversityvery similar to that of the two modern owl roost studies.
A relativelywide range of 9–17micromammal specieswas obtained per roost site from the thirteen owl roostsites at different areas in the WCNP-modern, givingan average of 12.2 species. It is interesting that speciesdiversity in the Steenbokfontein area, which is currentlybeing farmed and represents an environmentally dis-turbed area, is higher than that in the WCNP-modernassemblages, which were collected from a nature reserve.This may have something to do with the fact that barnowls have a narrower and more specialised diet in moreproductive habitats, and awider spectrumdiet in areas thatare more arid and have lower micromammal densities(Taylor, 1994).
Rattus rattus andMus musculus which are found onlyin the modern owl pellet assemblages in Table 4, arecommensal, introduced species with a world-widedistribution (Stuart and Stuart, 2001). Desmodillusauricularis, a gerbillid species which is currentlywidespread in the South West Arid zone and along thewest coast, is missing from all the Pleistocene/Holocenefossil sites, suggesting that the distribution of this speciesduring the late Middle Pleistocene – Holocene may nothave extended as far south as it does today. Furtherresearch will serve to confirm or refute this. The problemof interpreting absence data of species from the fossil sitesis that absence from the fossil assemblage need not reflectabsence in the area, but may have a taphonomic causerelating to the agent/s of accumulation of the assemblage.
The Bathyergids are found in low frequencies in allthe fossil sites. Their subterranean lifestyle and, in thecase of Bathyergus suillus, large body size, preventsthem from being taken by owls in large quantities. Allmole rats eat geophytes, although the genus Bathyergusalso eats some aerial vegetation (Bennett and Faulkes,2000). The presence of mole rats may thus be taken asbeing indicative of the presence of geophytes in the areaat the time of formation of all the west coast fossil sites,
and indeed at LBW, where mole rats were found inunusually large quantities (Matthews et al., in press-b).
The Otomyinae are well represented in all the westcoast sites, with Parotomys occurring at STBKC andHDP1. The presence of this species at STBKC andHDP1 clearly indicates that Parotomys has extended itsdistribution further southwards at various periods in thepast as it is presently found slightly to the north of theSaldanha area (Stuart and Stuart, 2001).
Steatomys krebsii, which is found in all the otherwest coast fossil sites and the modern pellet assem-blages, is absent from HDP1. The reason for this isunclear as this murid appears to be restricted to open,sandy, grass-covered substrates (De Graaff, 1981;Stuart and Stuart, 2001) and these microhabitatsappear to have been present at HDP1, as indicatedby the presence of species such as Bathyergus suillus,Zelotomys woosnami and Tatera afra. For reasonsmentioned above, only the repeated absence of thisspecies in other fossil sites of a similar age mayprovide definite evidence for shifts in the distributionof species over time, or for palaeoclimatic change.
The genera Mystromys, Rhabdomys, Bathyergus andCryptomys are found in all the west coast fossil sites.Dendromus is found in all the sites but HDP1, andAethomys appears in all the fossil assemblages, with theexception of SBYC. Acomys is represented at LBW byA. mabele and at HDP1 and EBC by the endemic A.subspinosus. Graphiurus was reported to appear atLBW (Hendey, 1981), but recent research has notconfirmed the presence of this genus. Graphiurus wasfound in the Terminal–Pleistocene/Holocene where it isfound at EBC and STBKC.
Certain species common to the other fossil west coastsites, such as Crocidura asiatica, Crocidura flavescens,Suncus varilla, Elephantulus edwardii, Dendromusmelanotis, Steatomys krebsii and Gerbillurus paeba,are missing from HDP1. All four endemic micromam-mal species of the south-western Cape are present atHDP1 (that is the mole rat B. suillus, and the murids,Tatera afra, Myomyscus verreauxi and Acomys sub-spinosus), however, suggesting that these species werewell established in the area by the late MiddlePleistocene. Other species, such as O. slogetti and Z.woosnami, make an appearance only at HDP1. Otomysslogetti is today associated with rocky habitats at highaltitudes such as the Drakensberg (Stuart and Stuart,2001), however, the presence of this species at HDP1 onthe west coast, and in Sterkfontein (Northern province)in horizons dating to approximately 1.5 Ma (Avery,2000b), indicates that this species enjoyed a much widerdistribution in the past.
344 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
The microhabitats indicated by the HDP1 micro-mammals include open, scrub vegetation, and open,sandy areas, and species adapted to arid and semi-aridconditions dominate the assemblage (Matthews, 2004).Three of the species found at HDP1, namely Zelot-omys woosnami, Parotomys brantsii, and Elephantulusrupestris, have a current distribution which lies north-wards of the Saldanha area. Parotomys brantsii iscurrently found slightly further to the north on the westcoast than Saldanha Bay, just north of St Helena Bay,and is noted as being predominantly associated withdry, sandy substrates in the more arid areas of thesubregion (Skinner and Smithers, 1990). The other twospecies, that is E. rupestris and Z. woosnami, arefound considerably further to the north today. E.rupestris has a current distribution which lies some250 km to the North on the west coast in theSpringbok area and is associated with rocky, aridhabitats. Zelotomys woosnami is also currently foundseveral hundred kilometers further northwards, inNamibia, Botswana and the northern Cape Province,
Fig. 5. AFC based on presence–absence data of rodent ge
and is noted as prefering semi-arid environments withan annual rainfall of 200–500 mm (De Graaff, 1981;Skinner and Smithers, 1990). The other fossil sitescontain species that are found in, or close to the area ofthe fossil sites today. HDP1 is thus anomalous inhaving two species which are currently found hundredsof kilometers away.
5. Results of community analyses (AFCand similaritypatterns)
5.1. The west coast fossil sites: a comparison by genera
Fig. 2 displays plan F1xF2 of the AFC performedon the west coast fossil sites, using presence–absencedata of rodent genera (see Appendix A).
LBW has a strong contribution to axis 1 due to thepresence of extinct genera (Euryotomys and Stenodont-omys) not found in other, younger sites. LBW is alsodifferentiated from the other sites by the presence ofThallomys, the tree rat (usually associated with acacia
nera and the biotic zones (after Rautenbach, 1978).

345T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
trees), which is not found in any of the other fossil sites.Axis 2 shows a difference between EBC on one side andHDP1 on the other. HDP1 is characterized by the presenceof genera such as Zelotomys woosnami and Elephantulusrupestriswhich are found today much further to the northin arid areas, as discussed in the previous section.
Langebaangweg is excluded from the analysis inFig. 3, and compares the west coast fossil sites with themodern owl roost assemblages from the West CoastNational Park, and Steenbokfontein farm. The analysis isbased on species presence–absence (see Appendix A).
As Fig. 3 indicates, a comparison of modern, andPleistocene–Holocene faunas shows that the fossilmicromammal assemblages of EBC, STBKC, andSBYC are very similar to those from modern owlpellet assemblages. Differences in the HDP1 micro-mammalian fauna are clearly indicated by the highcontribution along axis 1. A taxonomic habitat indexanalysis of the HDP1 micromammals showed a heavyloading on the Arid Biotic Zone (Matthews et al.,2005). This, together with the fact that HDP1 ismissing many characteristic west coast species, andcontains species such as Zelotomys woosnami andElephantulus rupestris which are adapted to arid areas,suggests that conditions on the west coast in the late
Fig. 6. AFC based on presence–absence data of rodent genera from the SoMember (Western Cape, S. A.); LQSM = Langeberg Quartzose Sand Memb(Northern province, S. A.); MRCIS = Makapansgat rodent corner in situ, ((Northern province, S. A.); NGA = Ngamiland (Botswana); JAG = Jägersdeposits (Northern province, S. A.).
Middle Pleistocene may have been relatively more aridthan today (see Matthews, 2004; Matthews et al., 2005,for a detailed discussion on this conclusion). The roleplayed by the predator (identified as a barn owl)responsible for the accumulation of HDP1 assemblagesremains to some extent uncertain, and it is possible thatthe emphasis on arid and semi-arid adapted specieswas influenced by the owl hunting in more open/aridareas. The AFC results do, however, support thesuggestion that the environment at HDP1 was differentto that which prevailed during the deposition of theother west coast fossil sites, and that of the modernpellet assemblages. All four endemic micromammalspecies of the south-western Cape are present at HDP1(that is the mole rat B. suillus, and the murids, Tateraafra, Myomyscus verreauxi and Acomys subspinosus),suggesting that these species were well established inthe area by the late Middle Pleistocene.
A comparison with the modern biotic zones fromRautenbach (1978) in Fig. 4 enables us to compare thewest coast micromammal populations over time with thevarious biomes (see Appendix A).
Fig. 4 illustrates that LBW and HDP1 are differentfrom the other west coast fossil sites, and also frommodern Namib and SWArid micromammal populations.
uth African fossil sites. Key: MPPM = Muishond Pelletal Phosphateer (Western Cape, S. A.); EXQR = Makapansgat exit quarry red mudNorthern province, S. A.); MLWD = Makapansgat limework dumpsquelle (Namibia); NOS = Nosib (Namibia); WP160 = Waypoint 160

Fig. 7. The number of Murinae/Gerbillinae genera in southern Africansites (after Pickford and Mein, 1988; Avery, 1992, 1999, in press;Denys, 1999; Manthi, 2002; Matthews, 2004). Key: LBW =Langebaanweg, MKLW = Makapansgat limework dumps, MRCISand EXQR = Makapansgat rodent corner in situ and Makapansgat exitquarry red mud, NGA = Ngamiland, JAG — Jägersquelle, NOS =Nosib, HDP1 = Hoedjiepunt 1, SBYC = Saldanha Bay Yacht Club site,EBC = Elands Bay Cave, STBKC = Steenbokfontein Cave.
346 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
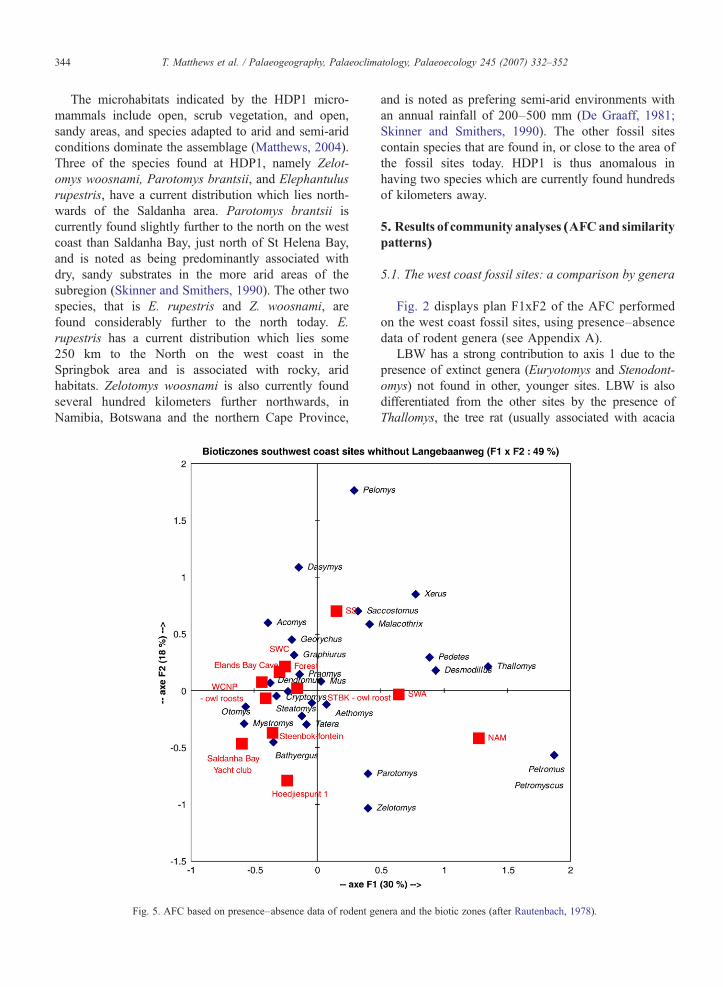
Fig. 5 shows the same AFC as Fig. 4, minus the site ofLBW.
When LBW is removed from the analysis, it becomesevident that the west coast fossil sites have few affinitieswith the more arid regions of South Africa, such as theNamib, SW arid and Savanna grasslands. Most of themlie closer to the coastal forest and South West Cape(fynbos) biotic zones, especially EBC, the WCNP owlroost, and the STBKC and Steenbokfontein modernassemblages. HDP1 and SBYC may represent slightlydistinct environments.
5.2. Comparison of the Mio-Pliocene sites of southernAfrican
Fig. 6 illustrates differences in the rodent assem-blages from the early Pliocene fossil faunas of theNamib (Jägersquelle and Nosib) and Ngamiland inBotswana (Pickford and Mein, 1988; Senut et al., 1992;Pickford et al., 1994), the new Waypoint160 assem-blages from Bolt's Farm (Senegas, 2000), the otherSterkfontein Valley sites (Denys, 1987, 1999), andLangebaanweg (represented by the LQSM and MPPM)(see Appendix B).
LBW and WP160 are relatively close due to the factthat they hold the extinct genus Euryotomys in common.There are some similarities between LBWand the sites ofJägersquelle and Nosib which are situated today inNamib region, but the absence of Gerbillurus, and Sac-costomus (rodents of Woodland Acacia–Mopane savan-nas) at LBW indicates a difference in the environment ofthe two regions.Malacothrix, which currently has a wide
distribution throughout the Namib and SWArid regionsextending into the Cape Macchia (fynbos) and Savannaregions is also notably absent from LBW, but present inthe Namibian sites, and Ngamiland, as well as in thefossil STBKC deposits, and the modern Steenbokfonteinpellet assemblages.
The relatively younger caves sites of Makapansgatand Ngamiland are characterized by a higher muriddiversity and the presence of different Otomyinae spe-cies. This is probably related to the existence of morehumid conditions, or alternately, may be a taphonomicartefact of time-averaging (Denys, 1999).
5.3. The Palaeoecology of Langebaanweg and the westcoast fossil sites
The ratio of Murinae to Gerbillinae has been used asan indicator of aridity by several authors as the numberof gerbillids are known to increase, and the number ofMurinae decrease, with an increase in aridity (Denys,1996; Fernandez-Jalvo et al., 1998; Denys, 1999). Thisis clearly indicated by the high number of gerbillid,relative to murid, species found in the Sahara, Sahelianand Sudanian Regions today (Denys, 1999). Thepresence of only one gerbillid species at LBW indicatesthat open, and relatively arid, areas existed, but does notindicate general aridity in the area. Fig. 7 compares thenumber of Murinae to Gerbillinae in the southernAfrican fossil sites.
The Gerbillinae are represented by an undescribedDesmodillus species at LBW, and there are approxi-mately six murine genera. The Ngamiland caves ofBotswana, dated to ∼3 Ma, yielded a large number ofgerbillids and Pickford and Mein (1988) list fivedifferent species, and three different genera, foundtogether with four murine genera. The Namibian sitesshow slightly less murine diversity, and in the case ofNosib, a slightly higher Gerbillinae/Murinae ratio. Interms of the South African Early–Middle Pliocene fossilsites, one gerbillid genus is found at Makapansgatrodent corner in situ and at Makapansgat exit quarry redmud, together with eight murine genera. Makapansgatlimework dumps contains no gerbillid genera, but ninemurine genera are represented. Langebaanweg and theMakapansgat sites thus show a particularly high numberof murine genera, both sites contain only one gerbillid—but not the same genus. Desmodillus is today restrictedto SW arid zone, while Tatera is a more ubiquitoussavanna marker found all over tropical Africa, except inrainforest zones (Denys, 1999).
LBW reflects a less arid environment than theNamibian fossil sites as three gerbillid genera were

347T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
represented at Jägersquelle, and two at Nosib (Denys,1999). The Namibian sites show a similar number ofmurine genera to all the other west coast fossil sites(SBYC excepted), and the number of gerbillid speciesfluctuates in a relatively even pattern from one to two,with the exception of Ngamiland which has rather morewith three genera. SBYC stands out as being the onewest coast fossil site which has only one murine, andtwo gerbillid genera.
6. Discussion
As illustrated by Fig. 6, Nosib and Jägersquelle shareseveral genera with LBW, and also show affinities withthe Makapansgat fossil sites. LBW shares more generawith Nosib than with Jägersquelle. Jägersquelle sharesmore genera with Nosib than the individual Makapans-gat sites, which cluster relatively closely together.Ngamiland remains distinct from the South Africanfossil sites. The Waypoint 160 deposits share the genusEuryotomys with LBW, but show some separation fromthe other southern African fossil sites.
The discovery of more Namibian and Botswana-based sites is eagerly awaited as it will add greatly tounderstanding the evolution of the SWArid and Namibzones and their associated micromammalian faunas, andwill hopefully contribute to our understanding of thedifferences between the faunal assemblages of Nosiband Jägersquelle, and the relationship of Namibianfaunas to eastern and southern African sites during theMiocene and Pliocene.
There appears to be a trend to share genera betweenwhat is today the SWArid and Namib regions with thatof the Cape Macchia (fynbos) from the periodrepresented by the LBW fossil deposits (∼5 Ma) untilthe present. Currently, micromammal genera shared bythe Cape, and Kalahari SWArid Regions today include;Tatera, Gerbillurus, Saccostomus, Dendromus, Otomys,Mus, Aethomys, Rhabdomys, Graphiurus, Cryptomysand Georychus (Denys, 1999). Genera shared by theCape, and Namib Regions includes Tatera, Gerbillurus,Saccostomus, Otomys, Mus, Aethomys, and Rhabdomys(Denys, 1999). All of the above-mentioned genera wererecovered from Namibia in Otavi Mountain breccias ofPost-Miocene age (Pickford et al., 1994), and Dendro-mus, Aethomys, Rhabdomys, Cryptomys and possiblyGraphiurus, are found at LBW.
Despite the similarities mentioned above, the micro-mammalian assemblages of LBW are differentiated byAFC analysis from the other, younger, west coast fossilsites, and also from modern Namib and SW Aridassemblages in Figs. 2 and 4. This is not surprising as,
although the earliest recovered fynbos pollen from thewestern Cape region comes from LBW, the largemammals contain many grazing and browsing species,indicating the presence of grasslands, woodland andriverine forest (Hendey, 1983), rather than the semi-arid or arid environments found along the west coast,and in the SW Arid and Namib regions today. Thepresence of extant genera, but extinct species, at LBWsuggests that the micromammals adapted over time tochanging conditions, which included the gradualdisappearance of woodland and forest concomitantwith an increase in grasslands, together with thedevelopment of the Cape Macchia biotic zone. Boththe fynbos and micromammal genera present at LBWhave families resident in the west coast area today, andfurther north in the Namib and South West AridRegions. This suggests the endurance and continuationof certain aspects in the environment of the west coastfrom the Mio-Pliocene, until the present, and lendssupport to the suggestion that the resident micromam-mal population evolved along with the fynbos character-istic of the Cape Macchia biotic zone. The large mole ratpopulation at LBW provides evidence that geophytes, acommon component of west coast vegetation today, werewell-established in the area around 5 Ma (Matthews,2004; Matthews et al., in press-b). Lovegrove and Jarvis(1986) have suggested that some of the Iridaceae co-evolved with mole rats. Another example of the co-dependency of micromammals and fynbos is the murid,Myomyscus verreauxi, an endemic of the southwesternCape, which is thought to be dependent on proteas forfood, while, conversely, some ground proteas depend onthis murid for pollination (David, 1978). Clearly therelationship between many fynbos and micromammalspecies is a long-standing one.
7. Conclusion
To summarise, nine micromammal genera found atLBW are present in some, or all, of the west coast fossilsites dating from the late Middle Pleistocene until theHolocene. These are: Aethomys, Rhabdomys, Mystr-omys, Dendromus, Acomys, Zelotomys, Cryptomys,Bathyergus and Graphiurus. These genera (with theexception of Zelotomys) are still found today in the CapeProvince, while Acomys subspinosus, and Bathyergussuillus are endemic to the Cape region. This indicates theendurance of many of the genera present at LBW, and inthe SW Arid and Namib Regions, from the Mio-Pliocene, up until the present. This provides goodsupport for Denys's (1999) suggestion that regionaldifferentiation of the southwest Cape Province and the

348 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
South West Arid biomes in terms of rodent taxa tookplace in South Africa between 6 and 4Ma. This is also inaccordance with the low evolutionary rates observed intropical Africa and particularly in South Africa, asdescribed by Denys and Jaeger (1986) and Sénégas(2000). The persistence of so many genera over severalmillion years on the west coast suggests a certainstability in that no radical climatic shift is indicated andthere does not appear to have been much disturbance oflocal genera by immigrant colonization, but ratherimmigration of southern species northwards. As men-tioned earlier, the climatic features controlling thepresent aridity along the south-western coast, theNamib and the dryness of the interior, appear to havebeen present, or developing, in the Early Pliocene, and awinter rainfall pattern was already established in theregion.
The general pattern observed in the west coast fossilsites and in the modern barn owl pellet assemblages,with the exception of HDP1, is that the Soricidae,Otomyinae and Gerbillidae (either T. afra or G. paeba)generally dominate the assemblages (Avery, 1992,1999, in press; Manthi, 2002). In the fossil sites ofEBC, STBKC and SBYC, two species of gerbillid (T.afra and G. paeba), four soricid species, and threespecies from the Otomyinae are always represented,although one particular gerbillid or Otomys speciesusually dominates in terms of overall numbers. As Fig.4 illustrates, the micromammalian faunas of the westcoast fossil sites of SBYC, EBC and STBKC (and, aswould be expected, the modern owl pellet collections)show many similarities with each other, and with theCape Macchia (fynbos) biotic zone, but not the Namiband SW Arid micromammal populations, suggestingthat the west coast has had a distinct micromammalianfauna from the Late Pleistocene, onwards.
A(
DSMGTDM
The HDP1 fauna is characterized by a micromammalianfauna which separates it from that of the other west coastsites, and the modern owl assemblages. HDP1 alsodiffers from the micromammalian faunas of the modernNamib and SW Arid biotic zones. The presence ofseveral arid and semi-arid adapted species and theappearance of certain species which today have a verydifferent distribution distinguish this fossil site from theothers, although the effect that the owl responsible forthe accumulation had on prey species selection isuncertain. The AFC results support the conclusion thatthe environment at HDP1 was different to that whichprevailed during the deposition of the other west coastfossil sites. More west coast sites dating to the Early,Middle and Late Middle Pleistocene need to be analysedin order to confirm and elucidate the climatic andenvironmental differences suggested by the HDP1assemblages.
Acknowledgements
Thanks are due to the many people at the Iziko SouthAfrican Museum, and the Department of Archaeology,University of Cape Town, for support received duringthis research project. Many thanks to Margaret Avery,Derek Ohland and Pippa Haarhof for providing accessto the Langebaanweg material, and to Sheena Kaal,Kerwin van Willingh and Noel Fouten for accessioninghundreds of micromammal fossils, and for alwayshelping out when there was a problem. Financialsupport for this study was received from the NationalResearch Foundation of South Africa. The NSF-RHOI(F.C. Howell and T. White) program also providedsupport through the framework of the TaphonomyAnalytical Working Group. Finally, thank you to thereferees for their constructive comments.
ppendix A. Rodent genera in the west coast fossil sites, modern owl roost assemblages, and the biotic zonesafter Rautenbach, 1978)
The west coast fossil sites and modern owl roost assemblages
The biotic zonesLBW
HDP1 SBYC EBC STBKC STBK–owl roostWCNP–owl roosts
NAM
SWA SWC SS Forestendromus
1 0 1 1 1 1 1 0 1 1 1 1 teatomys 0 0 1 1 1 1 1 0 1 0 1 0 alacothrix 0 0 0 0 0 1 0 0 1 0 1 0 erbillurus 0 0 1 1 1 1 1 1 1 1 1 0 atera 0 1 1 1 1 1 1 1 1 1 0 esmodillus 1 0 0 0 0 1 0 1 1 0 1 0 ystromys 1 1 1 1 1 0 0 0 0 1 1 0
349T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
(continued )Appendix A (continued )
1
The west coast fossil sites and modern owl roost assemblages
1 1 1 1
The biotic zones
LBW
HDP1 SBYC EBC STBKC STBK–owl roostWCNP–owl roosts
1
NAM
2
SWA
1
(c
SWC
2
ontinued
SS
1
on next
Forest
Acomys
1 0 0 1 0 1 0 0 0 1 1 1 Praomys 0 1 0 1 0 0 0 0 1 1 1 1 Rhabdomys 1 1 1 1 1 1 1 1 1 1 1 1 Mus 1 0 0 1 1 1 1 1 1 1 1 1 Aethomys 1 1 0 1 1 1 0 1 1 1 1 1 Dasymys 0 0 0 0 0 0 0 0 0 1 1 0 Zelotomys 1 1 0 0 0 0 0 0 1 0 0 0 Thallomys 1 0 0 0 0 0 0 1 1 0 1 0 Graphiurus 1 0 0 1 1 0 0 0 1 1 1 1 Otomys 0 1 1 1 1 1 1 0 0 1 1 1 Parotomys 0 1 0 0 1 0 0 1 1 1 0 0 Euryotomys 1 0 0 0 0 0 0 0 0 0 0 0 Stenodontomys 1 0 0 0 0 0 0 0 0 0 0 0 Bathyergus 1 1 1 0 1 1 0 0 1 1 0 1 Cryptomys 1 1 0 1 1 1 0 0 1 1 1 1 Georychus 0 0 0 1 0 0 1 0 1 1 1 1 Petromyscus 1 0 0 0 0 0 0 1 1 0 0 0 Saccostomus 0 0 0 0 0 0 0 0 1 1 1 0 Pedetes 0 0 0 0 0 0 0 1 1 0 1 1 Xerus 0 0 0 0 0 0 0 0 1 0 1 0 Petromus 0 0 0 0 0 0 0 1 1 0 0 0 No. of genera 5 0 8 4 3 3 8 1 2 7 2 2Key to west coast fossil sites: HDP1 = Hoedjiepunt 1, SBYC = Saldanha Bay Yacht Club site, EBC = Elands Bay Cave, STBKC = SteenbokfonteinCave.Key to the biotic zones (after Rautenbach, 1978): NAM = Namib, SS = Savanna grasslands, SWA = South West Arid, FOR = forest, SWC = SouthWest Cape Fynbos.
Appendix B. Rodent genera in southern African Pliocene sites
Langebaanweg ‘E’Quarry
Makapansgat
LQSM
MPPM WP160 EXQR MRCIS MLWD NGA JAG NOSAge
∼5 Ma ∼4 Ma ∼3.3–3.7 Ma ∼3Ma ∼3 Ma Plio-PleistoceneZelotomys
0 1 0 0 0 0 1 1 1 Mystromys 1 1 1 1 1 1 0 0 1 Stenodontomys 1 1 0 1 1 0 0 1 1 Proodontomys 0 0 0 1 1 1 0 0 0 Otomys 0 0 0 1 1 1 1 0 1 Myotomys 0 0 0 1 1 1 0 0 0 Prototomys 0 0 0 0 0 1 1 0 0 Cryptomys 1 1 1 1 1 1 0 0 1 Gypsorychus 0 0 0 0 0 1 0 0 0 Bathyergus 1 1 0 0 0 0 0 0 0 Heterocephalus 0 0 0 0 0 0 0 0 0 Euryotomys 1 1 1 0 0 0 0 0 0 Georychus 0 0 0 0 0 0 1 0 0 Graphiurus 1 1 0 0 0 0 0 1 1 Gerbillinae, indet. 0 0 1 0 0 0 0 0 0 Gerbillurus 0 0 0 0 0 0 0 1 1 Tatera 0 0 0 0 1 0 0 1 0 Desmodillus 1 1 0 0 0 0 0 1 0 Taterillus 0 0 0 1 1 0 1 0 0page)

Appendix B (continued )
350 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
Saccostomus
1
0
1
0
01
0
1
0
1
0
1
0
1
1
1
0
Dendromus 1 1 1 1 1 1 1 1 1 Steatomys 0 0 0 1 1 1 0 1 1 Malacothrix 0 0 0 1 1 1 1 1 1 Thallomys 0 1 0 0 0 0 0 1 0 Mastomys 0 0 0 0 1 1 0 0 0 Aethomys 1 1 1 1 1 1 0 0 0 Acomys 1 1 1 1 1 1 0 1 0 Dasymys 0 0 0 1 1 1 0 0 1 Grammomys 0 0 0 0 1 1 0 0 0 Mus 1 0 0 1 1 1 1 1 1 Myomyscus 0 0 0 0 1 1 1 0 0 Pelomys 0 0 0 0 0 1 1 0 0 Rhabdomys 1 1 0 1 1 1 0 0 1 Boltimys 0 0 1 0 0 0 0 0 0 Diversity 2 3 8 5 9 9 0 1 3(Faunal lists after Denys, 1987, 1999, Sénégas, 2000, Sénégas and Avery, 1998; Pickford et al., 1994; Pickford and Mein, 1988; Senut et al., 1992).Key: LQSM = Langebaanweg, Langeberg Quartzose Sand Member (Western Cape, S. A.).MPPM = Muishondfontein Pelletal Phosphate Member (Western Cape, S. A.).WP160 = Waypoint 160 deposits, Bolt's Farm (Northern province, S. A.).EXQR = Makapansgat exit quarry red mud (Northern province, S. A.).MRCIS=Makapansgat rodent corner in situ (Northern province, S. A.).MLWD = Makapansgat limework dumps (Northern province, S. A.).NGA = Ngamiland (Botswana).JAG = Jagersquelle (Namibia).NOS = Nosib1 and Nosib2 (Namibia).
References
Andrews, P., 1990. Owls, Caves and Fossils. Natural History MuseumPublications, London.
Avery, D.M., 1982. Micromammals as palaeoenvironmental indicatorsand an interpretation of the Late Quaternary in the southern CapeProvince South Africa. Ann. S. Afr. Mus. 85, 183–374.
Avery, D.M., 1992.Micromammals collected by barn owls. J. Zool. 38,385–397.
Avery, D.M., 1999. Holocene coastal environments in the WesternCape Province, South Africa: micromammalian evidence fromSteenbokfontein. Arch. Zool. 10, 163–180.
Avery, D.M., 2000a. Micromammals. In: Partridge, T.C., Maud, R.R.(Eds.), The Cenozoic of South Africa. Oxford monographs ongeology and geophysics, vol. 40. Oxford University Press, Oxford,New York, pp. 305–338.
Avery, D.M., 2000b. Notes on the systematics of micromammals fromSterkfontein, Gauteng, South Africa. Palaeontol. Afr. 36, 83–90.
Avery, D.M., in press. Micromammalian evidence for the LatePleistocene and Holocene environments of Elands Bay Cave. In:Parkington, J. (Ed.), Elands Bay Cave: A view on the past.
Bennett, N.C., Faulkes, C.G., 2000. African Mole Rats: Ecology andEusociality. Cambridge University Press, United Kingdom.
Benzecri, J.P., Benzecri, F., 1984. La pratique de l'analyse desdonnées. Exposé élémentaire, vol. I. Dunod Ed, Paris.
Bronner, G.N., Hoffmann, M., Taylor, P.J., Chimimba, C.T., Best, P.B.,Matthee, C.A., Robinson, T.J., 2003. A revised systematicchecklist of the extant mammals of the southern African subregion.Durban Mus. Novit. 28, 56–95.
Coetzee, J.A., 1978. Late Cainozoic palaeoenvironments of southernAfrica. In: Van Zinderen Bakker Sr., E.M. (Ed.), Antarctic GlacialHistory and World Palaeoenvironments. A.A. Balkema, Rotterdam,pp. 25–32.
Coetzee, J.A., 1980. Tertiary environmental changes along the south-western African coast. Palaeontol. Afr. 23, 97:203.
Coetzee, J.A., Rogers, J., 1982. Palynological and lithological evidencefor the Miocene Palaeoenvironment in the Saldhana region (SouthAfrica). Palaeogeogr. Palaeoclimatol. Palaeocol. 39, 71–85.
Cowling, R., Richardson, D., 1995. Fynbos: South Africa's UniqueFloral Kingdom. Fernwood Press in association with the Institutefor Plant Conservation, Cape Town.
Cruz-Uribe, K., 1988. The use and meaning of species diversity andrichness in archaeological faunas. J. Archaeol. Sci. 15, 179–196.
David, J.H.M., 1978. A survey of vertebrate fauna on the Rooiberg,Ladismith, Cape. Unpubl. Rep., Zoology Dept. University of CapeTown, South Africa.
De Graaff, G., 1960. A preliminary investigation of the mammalianmicrofauna in Pleistocene deposits of caves in the Transvaalsystem. Palaeontol. Afr. 7, 79–118.
De Graaff, G., 1961. On the fossil mammalian microfauna collected atKromdraai by Draper in 1895. S. Afr. J. Sci. 56, 259–260.
De Graaff, G., 1981. The Rodents of Southern Africa. Butterworths,Durban.
Denys, C., 1987. Rodentia and Lagomorpha. Fossil rodents (other thanPedetidae) from Laetoli. In: Leakey, M.D., Harris, J.M. (Eds.), Laetoli,a Pliocene Site in Tanzania. Clarendon Press, Oxford, pp. 118–170.
Denys, C., 1989. Phylogenetic affinites of the oldest East AfricanOtomys (Rodentia, Mammalia) from Olduvai Bed I. N. Jb. Geol.Palaont. Mh. 12, 705–725.
Denys, C., 1990a. Deux nouvelles espèces d'aethomys (Rodentia,Muridae) à Langebaanweg (Pliocène, Afrique du Sud); Implicationsphylogénétiques et paléoécologiques. Annal. Pal. (Vert-Invert) 76,41–69.
Denys, C., 1990b. The oldest Acomys (Rodentia, Muridae) from theLower Pliocene of South Africa and the problem of its muridaffinities. Palaeontol. Afr. 210, 79–91.

351T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
Denys, C., 1991. Un nouveau rongeur Mystromys pocockei sp. nov.(Crietinae) du Pliocene inferieur de Langebaanweg (Region duCap, Afrique du Sud). C. R. Acad. Sci. Paris 313, 1335–1341.
Denys, C., 1994a. Nouvelles espèces de Dendromus (Rongeurs,Muroidea) à Langebaanweg (Pliocene, Afrique du Sud) consequencesstratigraphiques et paléoécologiques. Palaeovertebrata 23, 153–176.
Denys, C., 1994b. Affinités systématiques de Stenodontomys(Mammalia, Rodentia) rongeur du Pliocène de Langebaanweg(Afrique du Sud). C. R. Acad. Sci. Paris 318, 411–416.
Denys, C., 1996. Olduvai rodent faunas: palaeoecological and palaeobio-geographical affinities: a comparison between East and South AfricanPlio-pleistocene faunas. Darmst. Beitr. Natur. 6, 247–261.
Denys, C., 1997. Rodent faunal lists in karstic and open-air sites ofAfrica: an attempt to evaluate predation and fossilisation biases onpaleodiversity. Cuard. Geol. Ib. 23, 73–94.
Denys, C., 1998. Phylogenetic implications of the existence of twomoderngenera of Bathyergidae (Mammalia, Rodentia) in the Pliocene site ofLangebaanweg (South Africa). Ann. S. Afr. Mus. 105, 265–286.
Denys, C., 1999. Of mice and men: evolution in East and South Africaduring Plio-Pleistocene times. In: Bromage, T.G., Schrenk, F.(Eds.), African Biogeography, Climate Change and HumanEvolution. Oxford University Press, New York, pp. 216–226.
Denys, C., Jaeger, J.J., 1986. A biostratigraphic problem: the case ofthe East African Plio-Pleistocene rodent faunas. Mod. Geol. 10,215–233.
Denys, C., Chorowicz, J., Tiercelin, J.J., 1987. Tectonic and environ-mental control on rodent diversity. In: Frostick, L.E., et al. (Ed.),Sedimentation in theAfricanRifts. . Geol. Soc. Sp. Pap. Publ., vol. 25.Geological Society, London, pp. 363–372.
Dingle, R.V., Lord, A.R., Hendey, Q.B., 1979. New sections in theVarswater Formation (Neogene) of Langebaan road, south-westernCape, South Africa. Ann. S. Afr. Mus. 78, 81–92.
Fernandez-Jalvo, Y., Andrews, P., 1992. Small mammal taphonomy ofGran Dolina, Atapuerca (Burgos), Spain. J. Archaeol. Sci. 19,407–428.
Fernandez-Jalvo, Y., Denys, C., Andrews, P.,Williams, T., Dauphin, Y.,Humphreys, L., 1998. Taphonomy and the palaeoecology ofOlduvai Bed-I (Pleistocene, Tanzania). J. Hum. Evol. 34, 137–172.
Franz-Odendaal, T., 2002. Analysis of dental pathologies in thePliocene herbivores of Langebaanweg and their palaeoenviron-mental implications. Unpublished Ph.D. thesis, University of CapeTown. Cape Town, South Africa.
Hendey, Q.B., 1974. The late Cenozoic carnivora of the south-westernCape Province. Ann. S. Afr. Mus. 63, 1–369.
Hendey, Q.B., 1976. The Pliocene occurrrences in ‘E’ Quarry,Langebaanweg, South Africa. Ann. S. Afr. Mus. 69, 215–247.
Hendey, Q.B., 1981. Palaeoecology of the Late Tertiary fossil occurrencesin ‘E’ quarry, Langebaanweg South Africa, and a reinterpretation oftheir geological context. Ann. S. Afr. Mus. 84, 1–104.
Hendey, Q.B., 1982. Langebaanweg: A Record Of Past Life. SouthAfrican Museum, Cape Town.
Hendey, Q.B., 1983. Cenozoic geology and palaeoecology of thefynbos region. In: Deacon, H.J., Hendey, Q.B., Lambrechts, J.J.N.(Eds.), Fynbos Palaeoecology: A Preliminary Synthesis. SouthAfrican National Scientific Programmes Report no 75. Mills Litho,Cape Town, pp. 35–60.
Klein, R.G., 1981. Ungulate mortality and sedimentary facies in theLate Tertiary Varswater Formation, Langebaanweg, south-westernCape Province, South Africa. Ann. S. Afr. Mus. 84, 233–254.
Lovegrove, B.G., Jarvis, J.U.M., 1986. Co-evolution between molerats (Bathyergidae) and a geophyte, Micranthus (Iridaceae).Cimbebasia 8, 79–85.
Manning, J., Goldblatt, P., 1996. West coast: South African wildflower guide 7. A joint publication from The Botanical society ofSouth Africa, the Darling Wild Flower Society, and the NationalBotanical Institute, Kirstenbosch, South Africa.
Manthi, F.K., 2002. The taphonomy of a micromammalian faunalassemblage from the Saldanha Bay Yacht Club: A contribution tothe study of the South African west coast palaeoenvironments.Unpublished Ma. thesis, University of Cape Town. Cape Town,South Africa.
Matthews, T., 1998. A taphonomic investigation of the agency ofmicrofaunal accumulation at Elands Bay Cave. Unpublished Ma.thesis, University of Cape Town. Cape Town, South Africa.
Matthews, T., 1999. Taphonomy and the micromammals from ElandsBay Cave. S. Afr. Archaeol. Bull. 170, 133–140.
Matthews, T., 2000. Predators, prey and the palaeoenvironment. S. Afr.J. Sci. 96, 23–24.
Matthews, T., 2004. The taxonomy and taphonomy of Mio-Plioceneand Middle Pleistocene micromammals, west coast, South Africa.Unpublished Ph.D thesis, University of Cape Town. Cape Town,South Africa.
Matthews, T., Denys, C., Parkington, J., 2005. The palaeoecology of themicromammals from the lateMiddle Pleistocene site ofHoedjiespunt1 (Cape Province, South Africa). J. Hum. Evol. 49, 432–451.
Matthews, T., Denys, C., and Parkington J., in press. The taphonomyof the micromammals from the late Middle Pleistocene site ofHoedjiespunt 1 (Cape Province, South Africa).
Matthews, T., Denys, C. and Parkington J., in press. An analysis of themole rats (Mammalia: Rodentia) fromLangebaanweg (Mio-Pliocene,South Africa).
Pickford, M., Mein, P., 1988. The discover of fossiliferous Plio-Pleistocene cave fillings in Ngamiland, Botswanna. C. R. Acad.Sci. Paris 307, 1681–1686.
Pickford, M., Mein, P., Senut, B., 1994. Fossiliferous Neogene karstfillings inAngola, Botswana andNamibia. S.Afr. J. Sci. 90, 228–231.
Pocock, T.N., 1976. Pliocene mammalian microfauna from Langebaan-weg: a new fossil genus linking the Otomyinae with the Murinae. S.Afr. J. Sci. 72, 58–60.
Pocock, T.N., 1987. Plio-pleistocene fossil mammalian microfauna of sou-thern Africa: a preliminary report including description of two newfossil muroid genera (Mammalia:Rodentia). Palaeontol. Afr. 26,69–91.
Rautenbach, I.L., 1978. A numerical re-appraisal of the southernAfrican biotic zones. Bull. Carnegie Mus. Nat. Hist. 6, 175–187.
Roberts, D.L., in press. Lithostratigraphy of the VarswaterFormation (Including the Langeenheid Sandy Clay, KoningsVlei Gravel, Langeberg Quartz Sand and Muishond FonteinPelletal Phosphorite Members). South African Committee forStratigraphy.
Rutherford, M.C., Westfall, R.H., 1986. Biomes of southern Africa-anobjective categorization. Memoirs of the Botanical Survey ofSouth Africa. No. 54 Botanical Research Institute. Dept. Agric.and Water Supply, South Africa.
Scott, L., 1995. Pollen evidence for vegetational and climatic changein Southern Africa during the Neogene and Quaternary. In: Vrba,E.S., Denton, G.H., Partridge, T.C., Burckle, L.H. (Eds.),Paleoclimate and Evolution with Emphasis on Human Origins.Yale University Press, Yale, pp. 65–76.
Sénégas, F., 2000. Les faunes de rongeurs (Mammalia) plio-pléistocènes de la province de Gauteng (Afrique du Sud) : misesau point et apports systématiques, biochronologiques et précisionspaléoenvironnementales. Non publiée Thèse Doctorat, UniversitéMontpellier II et EPHE, Montpellier, France.

352 T. Matthews et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 245 (2007) 332–352
Senegas, F., 2000. Les faunes de rongeurs (Mammalia) plio-pléistocènes de la province de Gauteng (Afrique du Sud) : misesau point et apports systématiques, biochronologiques et précisionspaléoenvironnementales. Non publiée Thèse Doctorat, UniversitéMontpellier II et EPHE. Montpellier, France.
Sénégas, F., 2001. Interpretation of the dental pattern of the SouthAfricanfossil Euryotomys (Rodentia, Murinae) and origin of Otomyinedental morphology. In: Denys, C., Granjon, L., Poulet, A. (Eds.),African Small Mammals. IRD, Paris, pp. 151–160.
Sénégas, F., Avery, D.M., 1998. New evidence for the murine originsof the Otomyinae (Mammalia, Rodentia) and the age of Bolt'sFarm (South Africa). S. Afr. J. Sci. 94, 503–507.
Senut, B., Pickford, M., Mein, P., Conroy, G., Van Couvering, J., 1992.Discovery of twelve new Late Cainozoic fossiliferous sites inpalaeokarsts of the Otavi Mountains, Namibia. C. R. Acad. Sci.,Paris 314, 727–733.
Siesser, W.G., 1978. Aridification of the Namib Desert: evidence fromoceanic cores. In: Van Zinderen Bakker Sr. (Ed.), Antarctic Glacial
History and World Palaeoenvironments. A.A. Balkema, Rotter-dam, pp. 105–114.
Siesser, W.G., 1980. Late Miocene origin of the Benguela upwellingsystem off northern Namibia. Science 208, 283–285.
Singer, R., 1961. The new fossil sites at Langebaanweg, South Africa.Curr. Anthropol. 2, 385–387.
Skinner, J.D., Smithers, R.H.N., 1990. The Mammals of the SouthernAfrican Subregion. University of Pretoria, Pretoria, South Africa.
Stuart, C., Stuart, T., 2001. Field Guide to Mammals of SouthernAfrica. Struik Publishers, Cape Town.
Stynder, D.D., Moggi-Cecchi, J.M., Berger, L.R., Parkington, J.E.,2001. Human mandibular incisors from the late Middle Pleistocenelocality of HDP1, South Africa. J. Hum. Evol. 41, 369–383.
Taylor, I., 1994. Barn Owls — Predator–Prey Relationships andConservation. Cambridge University Press, Great Britain.
Ward, J.D., Corbett, I., 1990. Towards an age for the Namib. In: Seely,M.K. (Ed.), Namib Ecology: 25 Years of Namib Research.Transvaal Museum, Pretoria, pp. 17–26.












![The BRICS and the South Atlantic: Emerging arena for South–South cooperation [South African Journal of International Affairs, 2014]](https://static.fdokumen.com/doc/165x107/631fe47418429976e405f443/the-brics-and-the-south-atlantic-emerging-arena-for-southsouth-cooperation-south.jpg)







