
A human monoclonal IgG1 potently neutralizing the pro-inflammatory cytokine GM-CSF
Upload
independentCategory
view
1download
0
Recombinant Human GM-CSF in E. coli 103
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
RESEARCH
103
Molecular Biotechnology © 2005 Humana Press Inc. All rights of any nature whatsoever reserved. 1073–6085/2005/30:2/103–116/$30.00
Abstract
Combined Effect of Protein Fusion and Signal Sequence GreatlyEnhances the Production of Recombinant Human GM-CSF
in Escherichia coli
Palash Bhattacharya, Gaurav Pandey, Poonam Srivastava,and Krishna Jyoti Mukherjee*
*Author to whom all correspondence and reprint requests should be addressed. Center for Biotechnology, Jawaharlal Nehru University, NewDehli, Delhi, 110067, India. E-mail: [email protected].
Granulocyte-macrophage colony-stimulating factor (GM-CSF) is a hematopoietic growth factor, that hasbeen used as a therapeutic agent in facilitating bone marrow and stem cell transplantation and in otherclinical cases like neutropenia. Although biologically active recombinant GM-CSF has been successfullyproduced in Escherichia coli, the reported levels are extremely poor. In this study we looked into the pos-sible reasons for poor expression and found that protein toxicity coupled with protease-based degradationwas the principal reason for low productivity. To overcome this problem we attached a signal sequence, aswell as an amino-terminal His-tag fusion to the GM-CSF gene. This combination had a dramatic effect onexpression levels, which increased from 0.8 µg/mL in the control to 40 µg/mL. When a larger fusion part-ner, such as the Maltose-binding protein (MBP-tag), was used the expression levels increased further to69.5 µg/mL, which along with the MBP-tag represented approx 12% of the total cellular protein.
Index Entries: GM-CSF; E. coli; signal sequence; protein fusion; high cell density.
1. IntroductionGranulocyte-macrophage colony-stimulating
factor (GM-CSF) is a hematopoietic factor, in-volved in the growth and maturation of granulo-cyte-macrophage populations (1). It plays a vitalrole in hematopoiesis by inducing distinct path-ways of development from a common progeni-tor. Recent therapeutic trials with GM-CSF havedemonstrated its use in chemotherapy and leuke-mia treatment (2). Recombinant GM-CSF hasbeen successfully used in many ways, like ame-lioration of acute and chronic states of neutro-penia after chemotherapy or bone marrowtransplantation (3,4) and in adjunctive therapywith antimicrobials (5–12). Its therapeutic rolein AIDS may also be useful to reduce myelode-pression caused by antiviral treatment and che-
motherapy for associated malignancies like Kaposi’ssarcoma (13).
Recombinant human GM-CSF (rhuGM-CSF)has been produced in Escherichia coli, althoughhigh-level production in its biologically activeform is affected by the poor stability of the proteinin the cytoplasm (14,15). In one study, rhuGM-CSF was transported to the periplasm using SP1and SP3 signal peptides. The absolute quantity ofthe cytokine produced was only 0.45 µg/mL/Uoptical density (O.D.) of the culture (14). Somereports claim that rhuGM-CSF is bacteriocidalwhen expressed in the cytoplasm and can only beisolated as an active species after transport to theperiplasm (15). Other reports claim that rhuGM-CSF has been produced as inclusion bodies in highamounts primarily as a fusion protein (16–18). A
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM103

104 Bhattacharya et al.
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
minor improvement in the production level hasbeen achieved by the supplementation of a tRNAfor a rare codon that codes for arginine (19). Thelevel of expression reported for all these studiesis, however, uniformly low.
Low expression of a recombinant protein canbe caused by many factors like plasmid instabil-ity, RNA instability, codon bias, protein toxicityto the host, and protease-based degradation. Westudied all these factors and we show that toxicityand degradation are coupled to limit rhuGM-CSFexpression. It is well established that transporting aprotein to the periplasm can reduce the toxicity ofthe protein toward its host (20), whereas a proteinfusion can reduce the rates of degradation broughtabout by the presence of specific amino acid resi-dues at the amino-terminal end (21,22). To thisend various signal peptides and fusion partnerswere used and a dramatic increase in the expres-sion of rhuGM-CSF was observed, along with asignificant improvement in the cellular health ofthe host.
2. Materials and Methods2.1. Host Strain and Plasmid Vectors
The E. coli BL21 (DE3) strain was used for allwork done in this study except the study on codonbias for which the E. coli strains BL21 (DE3)CodonPlus(RIL) and BL21 (DE3)Codon Plus(RP)(Stratagene) were used. The former strain carriesextra copies of argU, ileY, and leuW tRNA geneson a 3.5-kb plasmid with a chloramphenicolmarker. The latter strain contains extra copies ofthe argU and proL genes on the same plasmidbackbone. The plasmids used were pRSETA(Invitrogen, Carlsbad, CA), pET14b, pET22b(Novagen), and pMALp2X (NEB).
2.2. PCR Primers and TemplatesThe mRNA sequence of human GM-CSF
was obtained from NCBI (accession numberNM_000758). Two oligonucleotide primers CCTGCAGCCATATGGCACCCGCCCGCTCG andGGGGATCCTCACTCCTGGACTGG bearingNdeI and BamHI restriction sites, respectively(from Microsynth, Switzerland) were initially de-signed and used to amplify the mature humanGM-CSF cDNA (0.4 kb). Peripheral blood mono-
nuclear cells (PBMCs) were isolated from venousblood and cultured in RPMI 1640 with 10% fetalcalf serum (FCS) and 10 mg/mL phytohema-glutinin (PHA). The cells were harvested after 48h and total RNA isolated (High Pure RNA Isola-tion Kit, Boehringer, Mannheim). This total RNAwas used to amplify the hGM-CSF cDNA by re-verse transcriptase–polymerase chain reaction(RT-PCR) (C-therm Polymerase One Step RT-PCR System, Roche). This cDNA was cloned intoa pRSETA vector (Invitrogen) between the NdeIand BamHI site to form the plasmid pRSET-GM-CSF. The same fragment was cloned between theNdeI and BamHI sites of the plasmid pET14b(Novagen) to obtain the plasmid pET14b-GM-CSF. An NcoI/BamHI fragment from the plasmidpET14b-GM-CSF was cloned in the plasmidpET22b (Novagen) to give the plasmid pET22b-GM-CSF. In addition, the primers GCGAATTCGCACCCGCCCGCTCG and GGGGATCCTCACTCCTGGACTGG were used to PCR am-plify this cDNA and clone it in the plasmidpMALp2X between the EcoRI and BamHI sitesto get the plasmid pMAL-GM-CSF.
2.3. Shake Flask StudiesSingle colonies were inoculated in 10 mL of
LB medium in 100-mL conical flasks with 100µg/mL ampicillin and grown overnight at 37°Cwith shaking at 200 rpm. One milliliter each ofthe overnight cultures was used to inoculate 100mL of LB medium in 500-mL conical flasks withthe same antibiotic concentration and grown at37°C with shaking at 200 rpm. At an OD600 of 0.6the cultures were induced with isopropyl-β-D-thiogalactopyranoside (IPTG). Samples were col-lected every hour for analysis.
2.4. Plasmid Stability DeterminationThe cell culture samples taken at different time
points were appropriately diluted and aliquotswere plated on LB agar plates without antibiotics.Normally a dilution of 104 in saline was done and100 µL of this was plated. On average, 100 iso-lated colonies were picked up from the LB agarplate with toothpicks and transferred on to LB–ampicillin plates. The fraction of the colonies thatdid not grow on the antibiotic plate gave the frac-
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM104

Recombinant Human GM-CSF in E. coli 105
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
tion of nonrecombinants in the cell population.The fraction that grew gave the percentage ofplasmid stability.
2.5. Preparation of Periplasmic FractionPellets from 100-mL cultures (reagents were
scaled down proportionately when preparing fromlesser volumes) were resuspended in 10 mL of30 mM Tris-HCI (pH 8.0), 20% sucrose, and 1 mMethylenediaminetetraacetic acid (EDTA) and in-cubated on ice for 10 min. The samples were centri-fuged at 8000g at 4°C for 10 min, the supernatantswere discarded and the pellets were resuspendedin 5 mL of ice cold MgSO4 (5 mM). Samples wereincubated on ice for 10 min and centrifuged at10,000g at 4°C for 10 min. The supernatants,which constituted the periplasmic fraction, werecollected.
2.6. Western BlotThe proteins were separated on a 15% sodium
dodecyl sulfate-polyacrimide gel electrophore-sis (SDS-PAGE) and transblotted onto a polyvinyldifluoride (PVDF) membrane. After the transfer,the membranes were incubated in PBST (10 mMphosphate-buffered saline with 0.05% Tween-20)for 90 min. After a brief wash with PBST, themembrane was incubated with anti-huGM-CSFpolyclonal antibodies (goat IgG, 0.1 µg/mL) fol-lowed by three 10-min washes in PBST. Themembrane was further incubated in rabbit antigoatIgG conjugated to horseradish peroxidase (HRPO)and finally the immunoreactive bands were visu-alized with diamino-benzidine tetrahydrochloride–H2O2 solution.
2.7. Enzyme-Linked Immunosorbent AssayThe 96-well micro enzyme-linked immuno-
sorbent assay (ELISA) plates were coated withmouse anti-huGM-CSF antibody (R&D Systems)at 2 µg/mL concentration in coating buffer (phos-phate buffered solution [PBS]) overnight at roomtemperature. The coated plates were washed andblocked with 1% bovine serum albumin (BSA;250 µL/well). Samples (diluted in 0.2% BSA inPBST) were added to all wells except for the con-trol wells and incubated. Biotinylated mouseanti-huGM-CSF antibody (R&D Systems) at a
concentration of 0.5 µg/mL was used for detec-tion. Streptavidin-HRPO was added and finallycolor was developed with TMB-H2O2 solutionfollowed by addition of 2% H2SO4. Absorbancewas read at 450 nm. All steps were done at roomtemperature.
2.8. BioassayBioassay of rhuGM-CSF with HL-60 cells has
been described previously (23). A modified pro-tocol was used in this study where rhuGM-CSFpresent in supernates was appropriately diluted(1–5 ng/mL, estimated by ELISA) in culture me-dium on a 96-well flat-bottomed microplate in avolume of 50 µL. As negative control, culturemedium alone was used. The HL-60 cells wereresuspended in culture medium and adjusted to 2× 105 cells/mL. Of this cell suspension 50 µLwere added to the prediluted rhuGM-CSF samplesin each microwell (1 × 104 cells per microwell)and incubated for 48 h at 37°C and 5% CO2 10 µLMTT solution was added consequently (5 mg/mL), to each well of the microplate and incubatedfor 4 h at 37°C and 5% CO2. The reaction wasterminated by adding 100 mL of SDS/HCI solu-tion (10% SDS in 10mM HCI). The microplatewas incubated overnight and evaluated on anELISA reader by using 550 nm and 690 nm astest and reference wavelength, respectively.
3. Results3.1. Low Expression of rhuGM-CSFin E. coli Correlates With High PlasmidInstability and High Metabolic Burden
Human GM-CSF cDNA was synthesized byRT-PCR techniques using total RNA from pe-ripheral blood cells (described in Subheading 2.).The amplified fragment of 390 bp was cloned inthe plasmid pRSETA under the control of anIPTG inducible T7 promoter (Fig. 1A). The result-ing plasmid pRSET-GM-CSF was used to studyrhuGM-CSF expression in E. coli BL21 (DE3)cells. As expected, the expression of rhuGM-CSFwas poor. With an IPTG concentration of 0.5 mM,a maximum of 0.5 µg/mL of rhuGM-CSF wasobtained in shake flask cultures after 1 h of induc-tion, which rapidly fell to negligible values by 4 h(Fig. 2A). When 1mM IPTG was used the activ-
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM105

106 Bhattacharya et al.
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
Fig. 1. Schematic representation of the various plasmids used in this study. (A) pRSET-granulocyte-macrophagecolony-stimulating factor (GM-CSF); (B) pET14b-GM-CSF; (C) pET22b-GM-CSF; and (D) pMAL-GM-CSF.
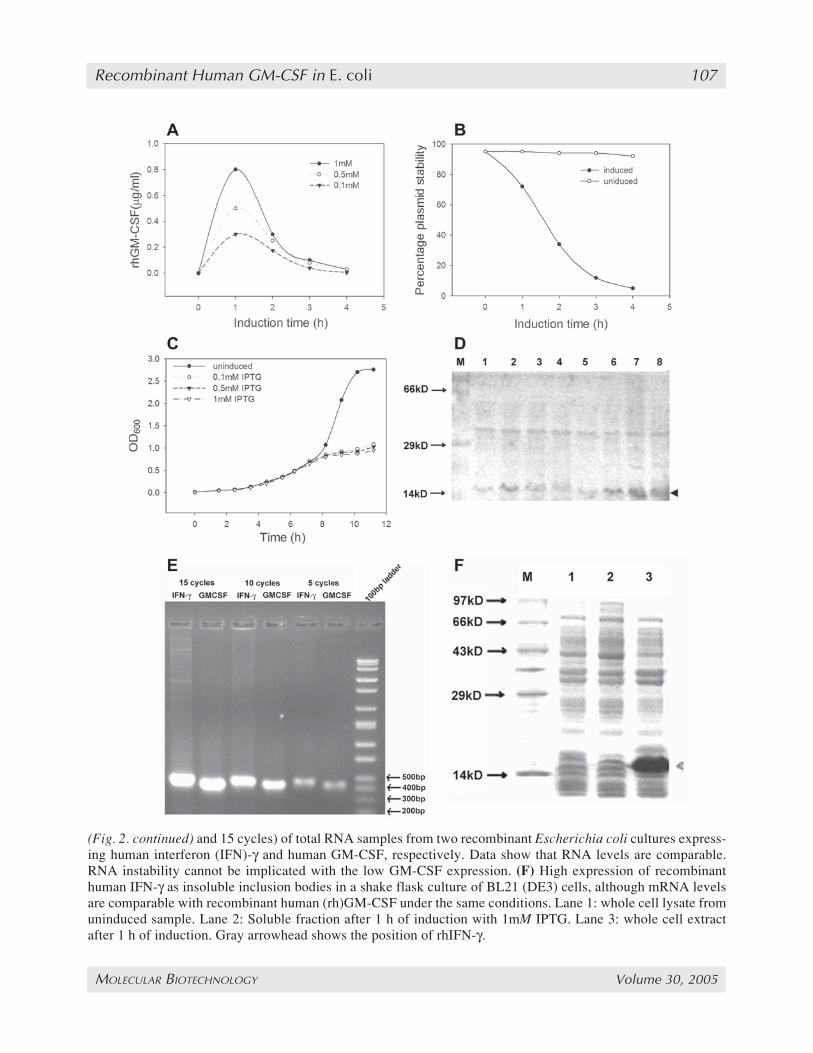
Fig. 2. (opposite page) Low granulocyte-macrophage colony-stimulating factor (GM-CSF) production corre-lates with plasmid instability but not with RNA instability. (A) Production of recombinant GM-CSF over time.(B) Percentage plasmid carrying cells in an induced culture compared to an uninduced culture. Graph shows rapidreduction in plasmid stability over time after induction. (C) Growth curve of induced cultures (with differentisopropyl-β-D-thiogalactopyranoside [IPTG] concentrations) compared to an uninduced culture. The induced cul-ture has a severe slowing down in the growth curve immediately after induction showing the toxic effects of GM-CSF on the cells. (D) Sodium dodecyl sulfate-polyacrimide gel electrophoresis comparing induction of GM-CSFexpression in cultures grown at 5X ampicillin compared to ones grown with 1X ampicillin. Lanes 1–4: cell lysatesof hourly samples from cultures grown with 1X ampicillin starting from the point of induction. Lanes 5–8: Celllysates from cultures grown with 5X ampicillin. Data shows steady buildup of GM-CSF in cultures grown in 5Xampicillin as compared to the reduction of GM-CSF levels after 1 h of induction in cultures grown in 1X ampicil-lin. Arrowhead shows the position of GM-CSF. (E) Reverse transcription polymerase chain reaction (with 5, 10,
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM106
Recombinant Human GM-CSF in E. coli 107
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
(Fig. 2. continued) and 15 cycles) of total RNA samples from two recombinant Escherichia coli cultures express-ing human interferon (IFN)-γ and human GM-CSF, respectively. Data show that RNA levels are comparable.RNA instability cannot be implicated with the low GM-CSF expression. (F) High expression of recombinanthuman IFN-γ as insoluble inclusion bodies in a shake flask culture of BL21 (DE3) cells, although mRNA levelsare comparable with recombinant human (rh)GM-CSF under the same conditions. Lane 1: whole cell lysate fromuninduced sample. Lane 2: Soluble fraction after 1 h of induction with 1mM IPTG. Lane 3: whole cell extractafter 1 h of induction. Gray arrowhead shows the position of rhIFN-γ.
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM107

108 Bhattacharya et al.
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
ity increased marginally to 0.8 µg/mL. Plasmidstability was measured at each time interval afterinduction. The values were compared to a controlculture grown without induction. At the point ofinduction, 95% of cells contained the plasmidwhich decreased to 10% in 4 h, whereas for thenoninduced culture no such decrease was observedwith plasmid-carrying cells being present after 4 h(Fig. 2B). This rapid drop in stability demonstratesthe metabolic burden associated with the plasmid-carrying cells in comparison to the plasmid-freecells. Efforts were made to increase plasmid sta-bility by growing the culture at 5X concentrationof ampicillin. This led to a greater stability, as wasevident from the continuous buildup of the recom-binant protein over time (Fig. 2D). However, theexpression values were not remarkably high witha maximum of around 4 µg/mL of rhuGM-CSFproduced after 4 h of induction with 1mM IPTG.
When the pRSET-GM-CSF carrying cells weregrown on LB plates, the cellular health was ob-served to be very poor. The cells grew slowly andhad smaller colony sizes than normal E. coli. Theywere extremely transparent, thin-layered, and alsoirregular in shape compared to the round, firm,white colonies typical of E. coli. In additionthey appeared to be granular in texture and thisgranularity increased with time of incubation. Theinduced E. coli cultures grew poorly, hardly reach-ing an OD600 of 1 after 4 h of induction, whereasthe noninduced cultures reached an OD600 of 2.76(Fig. 2C). The noninduced culture had a specificgrowth rate (µ) of 0.47 h–1, whereas the induced cul-tures had an average µ of 0.09 h–1 after induction.
3.2. Low Expression of rhuGM-CSF Is NotOwing to mRNA Instability
To determine whether rhuGM-CSF levels cor-related with higher mRNA instability and hencelow mRNA levels, RT-PCR was done at differentcycle numbers from total RNA isolated from in-duced cells 1 h after induction. Because weneeded to use a control to measure the relativeamount of mRNA, we chose a recombinant hu-man interferon (rhuIFN)-γ clone that had the samehost vector background. Moreover rhuIFN-γ isproduced in extremely large quantities by the
cells, reaching values corresponding to 40% oftotal cellular protein. Two cultures of BL21(DE3), one carrying pRSET-GM-CSF and theother carrying pRSET-IFN-γ, were induced andtotal RNA isolated after 1 h of induction. Equalamounts of total RNA (100 ng) was used to set upa reverse transcription reaction. Equal volumes ofthe RT mixture were used to set up PCR reactionsfor 5, 10, 15, and 20 cycles with both samples(Fig. 2E). In all cases it was found that the PCRproduct for rhulFN-γ (432 bp) was marginallyhigher than that of rhuGM-CSF (406 bp). Thismarginal difference is not commensurate with thehuge difference in protein expression, which was0.8 µg/mL and 40 µg/mL for rhuGM-CSF andrhuIFNγ, respectively, as seen on SDS-PAGE(Fig. 2F) and measured by ELISA. Thus, mRNAlevels may be affecting rhuGM-CSF productionmarginally but cannot possibly be the major rea-son for its low expression.
3.3. Codon Bias Does Not Contributeto Low rhuGM-CSF Expression
Excessive rare codon usage in the target genehas also been a cause for low level expression ofmany eukaryotic proteins (24,25). In an earlierreport, codon bias was stated as one of the rea-sons affecting GM-CSF production (19). In par-ticular, a single rare arginine codon (AGA) hasbeen found to be crucial. To verify this, the plas-mid pRSET-GM-CSF was used to express rhuGM-CSF in two strains—BL21-(DE3)CodonPlus-RILand BL21-(DE3)CodonPlus-RP—that have beendevised to express genes that are codon compro-mised. However, contrary to expectations, the twostrains did not show any increase in the level ofexpression of rhuGM-CSF compared with theoriginal BL21 (DE3) strain (Table 1). This dem-onstrates that codon bias was not a factor affect-ing the expression of rhuGM-CSF.
3.4. An Amino-Terminal 6-His tagIncreases rhuGM-CSF Production
The GM-CSF cDNA was subcloned in the vec-tor pET14b immediately downstream of a 6x-Hissequence. The resulting vector, pET14b-GM-CSF(see Fig. 1B), was used to drive expression of the
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM108

Recombinant Human GM-CSF in E. coli 109
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
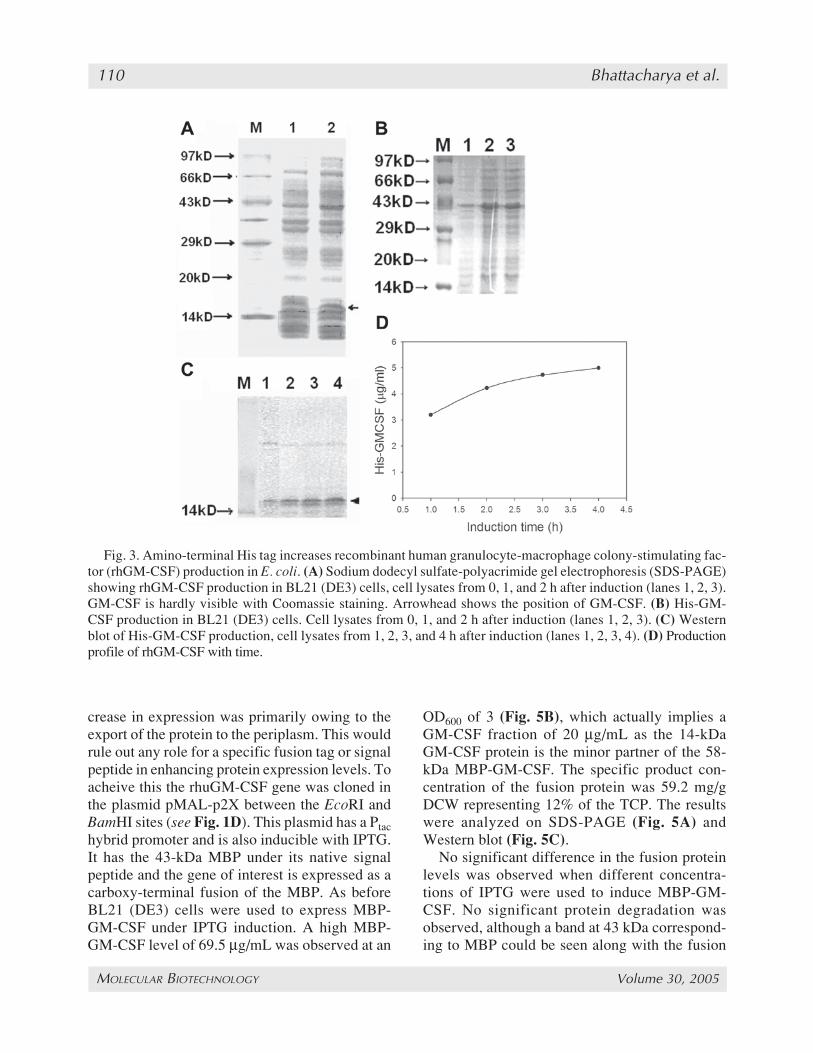
recombinant HisGM-CSF gene in E. coli BL21(DE3) cells using the vector’s IPTG inducible T7promoter. The 6x-His sequence along with thethrombin cleavage adds 29 amino acids to the fi-nal protein, originally of 14.5 kDa, thereby in-creasing its molecular weight to about 16.5 kDa.The results were compared to the expression ofrhuGM-CSF without the His-tag fusion. Themaximum concentration of soluble HisGM-CSFas measured by sandwich ELISA was 5 µg/mL(specific activity of 7.5 mg/g dry cell weight(DCW) or 1.5% of total cellular protein [TCP]) 4h after induction with 1mM IPTG (Fig. 3D),which was sixfold higher than the control (seeFig. 2A). The time course of induction was alsocompared. For the control plasmid pRSET-GM-CSF, the expression of soluble rhuGM-CSF in-creased to a maximum of 0.8 µg/mL (specificactivity 2.05 mg/g DCW or 0.4% of TCP) 1 h af-ter induction with 1mM IPTG and then decreasedrapidly in the next 3 h until it reached baseline. Inthe case of HisGM-CSF, however, this amount keptincreasing until it reached a maximum after 4 h.
Thus, the addition of the 6-His tag increasedthe rhuGM-CSF production and also sloweddown the rate of degradation of the product. Theexpression pattern was further confirmed usingSDS-PAGE (Fig. 3A,B) and Western blot (Fig. 3C).
3.5. Periplasmic Secretion FurtherEnhances HisGM-CSF Production
Periplasmic production of rhuGM-CSF in E.coli has been done with a variety of signal pep-tides and it has been shown that higher rhuGM-CSF expression is obtained (14,15,26). To checkwhether the use of a signal peptide further en-
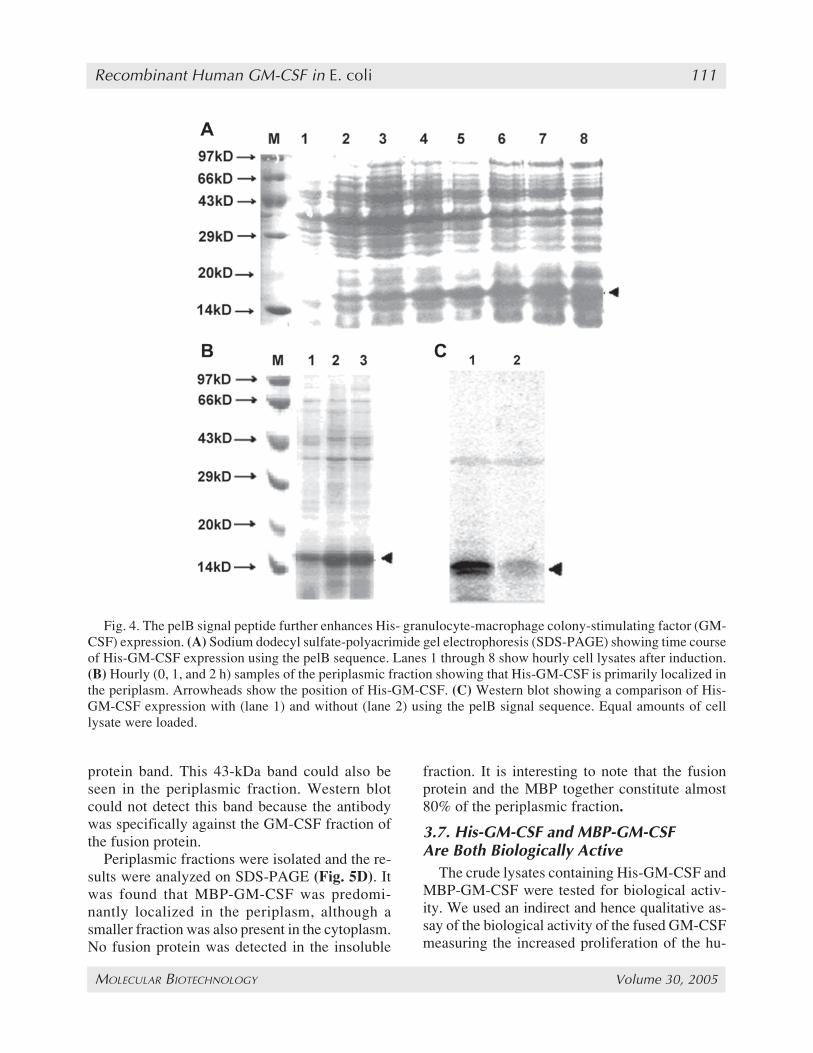
hances rhuGM-CSF expression, the rhuGM-CSFgene was excised from the plasmid pET14b-GM-CSF along with the 6-His sequence and clonedin-frame downstream of the pelB signal sequenceusing the NcoI and BamHI sites of the vectorpET22b. The resulting plasmid was called pET22b-GM-CSF (see Fig. 1C). BL21 (DE3) cells trans-formed with this plasmid were used to studyHisGM-CSF expression. Three different IPTGconcentrations, 0.1 mM, 0.5 mM, and 1 mM ofIPTG were used for induction. Although no sig-nificant difference in HisGM-CSF levels was ob-served between the different inducer concentrations,the overall production of HisGM-CSF increasedremarkably. At an OD600 of 2, the activity as mea-sured by ELISA was 40 µg/mL (a 50-fold in-crease over control), which corresponds to aspecific activity of 51.3 mg/g DCW. This impliesthat HisGM-CSF now represented more than 10%of the TCP. The results were also analyzed by SDS-PAGE and Western blot (Fig. 4A,C).
To determine whether the signal peptide wasbeing processed correctly, the periplasmic frac-tion was isolated and observed on SDS-PAGE(Fig. 4B). It was seen that most of the HisGM-CSF accumulated in the periplasm, which impliedproper processing of the signal peptide. The ac-cumulation of the protein until 7 h after inductionalso indicated enhanced protein stability. No deg-radation of the product was seen.
3.6. The Combined Effect of MBP Fusionand Signal Peptide Also EnhancesrhuGM-CSF Expression
A different combination of protein fusion andsignal peptide was used to check whether the in-
Table 1Codon Bias Does Not Contribute to Low rhuGM-CSF Expression
Expression level after 1 h of inductionStrain Rare codons supplemented at 1 mM IPTG
BL21 (DE3) None 0.8 µg/mLBL21 (DE3) CodonPlus(RIL) AGA, AGG (arg);AUA (ile); CUA (leu) 0.35 µg/mLBL21 (DE3)CodonPlus(RP) AGA, AGG (arg); CCC (pro) 0.5 µg/mL
IPTG, isopropyl-β-D-thiogalactopyranoside
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM109
110 Bhattacharya et al.
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
Fig. 3. Amino-terminal His tag increases recombinant human granulocyte-macrophage colony-stimulating fac-tor (rhGM-CSF) production in E. coli. (A) Sodium dodecyl sulfate-polyacrimide gel electrophoresis (SDS-PAGE)showing rhGM-CSF production in BL21 (DE3) cells, cell lysates from 0, 1, and 2 h after induction (lanes 1, 2, 3).GM-CSF is hardly visible with Coomassie staining. Arrowhead shows the position of GM-CSF. (B) His-GM-CSF production in BL21 (DE3) cells. Cell lysates from 0, 1, and 2 h after induction (lanes 1, 2, 3). (C) Westernblot of His-GM-CSF production, cell lysates from 1, 2, 3, and 4 h after induction (lanes 1, 2, 3, 4). (D) Productionprofile of rhGM-CSF with time.
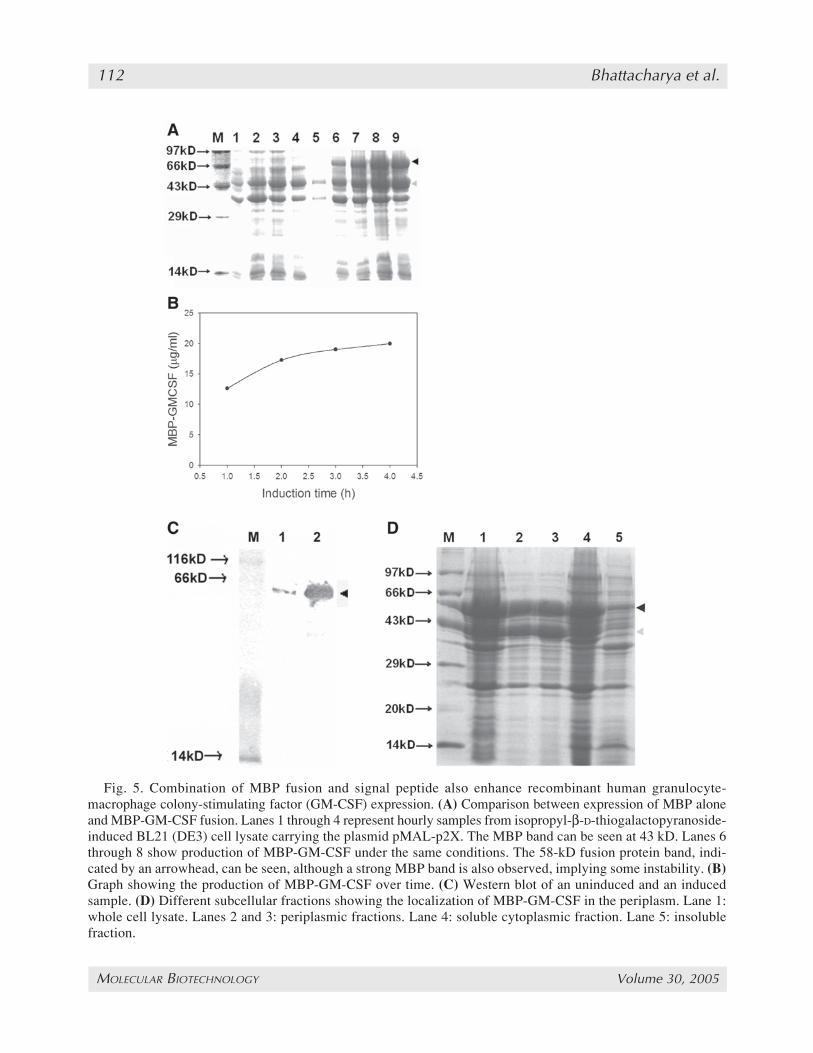
crease in expression was primarily owing to theexport of the protein to the periplasm. This wouldrule out any role for a specific fusion tag or signalpeptide in enhancing protein expression levels. Toacheive this the rhuGM-CSF gene was cloned inthe plasmid pMAL-p2X between the EcoRI andBamHI sites (see Fig. 1D). This plasmid has a Ptachybrid promoter and is also inducible with IPTG.It has the 43-kDa MBP under its native signalpeptide and the gene of interest is expressed as acarboxy-terminal fusion of the MBP. As beforeBL21 (DE3) cells were used to express MBP-GM-CSF under IPTG induction. A high MBP-GM-CSF level of 69.5 µg/mL was observed at an
OD600 of 3 (Fig. 5B), which actually implies aGM-CSF fraction of 20 µg/mL as the 14-kDaGM-CSF protein is the minor partner of the 58-kDa MBP-GM-CSF. The specific product con-centration of the fusion protein was 59.2 mg/gDCW representing 12% of the TCP. The resultswere analyzed on SDS-PAGE (Fig. 5A) andWestern blot (Fig. 5C).
No significant difference in the fusion proteinlevels was observed when different concentra-tions of IPTG were used to induce MBP-GM-CSF. No significant protein degradation wasobserved, although a band at 43 kDa correspond-ing to MBP could be seen along with the fusion
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM110
Recombinant Human GM-CSF in E. coli 111
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
Fig. 4. The pelB signal peptide further enhances His- granulocyte-macrophage colony-stimulating factor (GM-CSF) expression. (A) Sodium dodecyl sulfate-polyacrimide gel electrophoresis (SDS-PAGE) showing time courseof His-GM-CSF expression using the pelB sequence. Lanes 1 through 8 show hourly cell lysates after induction.(B) Hourly (0, 1, and 2 h) samples of the periplasmic fraction showing that His-GM-CSF is primarily localized inthe periplasm. Arrowheads show the position of His-GM-CSF. (C) Western blot showing a comparison of His-GM-CSF expression with (lane 1) and without (lane 2) using the pelB signal sequence. Equal amounts of celllysate were loaded.
protein band. This 43-kDa band could also beseen in the periplasmic fraction. Western blotcould not detect this band because the antibodywas specifically against the GM-CSF fraction ofthe fusion protein.
Periplasmic fractions were isolated and the re-sults were analyzed on SDS-PAGE (Fig. 5D). Itwas found that MBP-GM-CSF was predomi-nantly localized in the periplasm, although asmaller fraction was also present in the cytoplasm.No fusion protein was detected in the insoluble
fraction. It is interesting to note that the fusionprotein and the MBP together constitute almost80% of the periplasmic fraction.
3.7. His-GM-CSF and MBP-GM-CSFAre Both Biologically Active
The crude lysates containing His-GM-CSF andMBP-GM-CSF were tested for biological activ-ity. We used an indirect and hence qualitative as-say of the biological activity of the fused GM-CSFmeasuring the increased proliferation of the hu-
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM111
112 Bhattacharya et al.
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
Fig. 5. Combination of MBP fusion and signal peptide also enhance recombinant human granulocyte-macrophage colony-stimulating factor (GM-CSF) expression. (A) Comparison between expression of MBP aloneand MBP-GM-CSF fusion. Lanes 1 through 4 represent hourly samples from isopropyl-β-D-thiogalactopyranoside-induced BL21 (DE3) cell lysate carrying the plasmid pMAL-p2X. The MBP band can be seen at 43 kD. Lanes 6through 8 show production of MBP-GM-CSF under the same conditions. The 58-kD fusion protein band, indi-cated by an arrowhead, can be seen, although a strong MBP band is also observed, implying some instability. (B)Graph showing the production of MBP-GM-CSF over time. (C) Western blot of an uninduced and an inducedsample. (D) Different subcellular fractions showing the localization of MBP-GM-CSF in the periplasm. Lane 1:whole cell lysate. Lanes 2 and 3: periplasmic fractions. Lane 4: soluble cytoplasmic fraction. Lane 5: insolublefraction.
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM112
Recombinant Human GM-CSF in E. coli 113
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
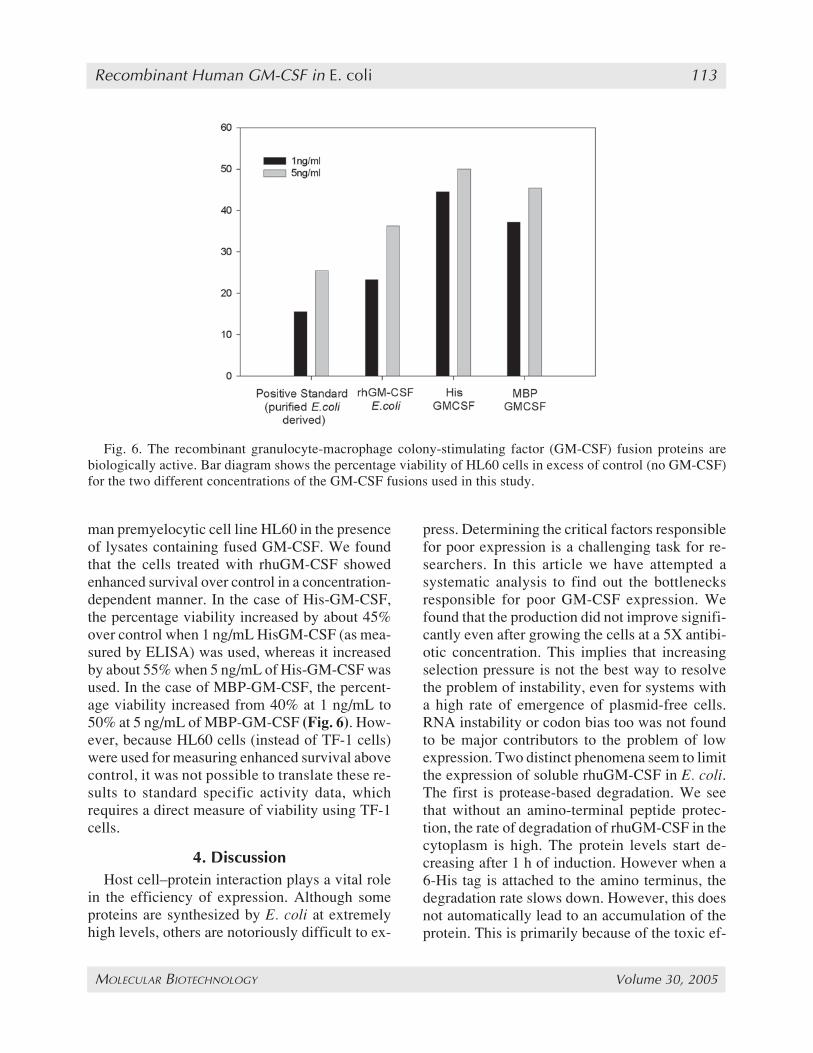
man premyelocytic cell line HL60 in the presenceof lysates containing fused GM-CSF. We foundthat the cells treated with rhuGM-CSF showedenhanced survival over control in a concentration-dependent manner. In the case of His-GM-CSF,the percentage viability increased by about 45%over control when 1 ng/mL HisGM-CSF (as mea-sured by ELISA) was used, whereas it increasedby about 55% when 5 ng/mL of His-GM-CSF wasused. In the case of MBP-GM-CSF, the percent-age viability increased from 40% at 1 ng/mL to50% at 5 ng/mL of MBP-GM-CSF (Fig. 6). How-ever, because HL60 cells (instead of TF-1 cells)were used for measuring enhanced survival abovecontrol, it was not possible to translate these re-sults to standard specific activity data, whichrequires a direct measure of viability using TF-1cells.
4. DiscussionHost cell–protein interaction plays a vital role
in the efficiency of expression. Although someproteins are synthesized by E. coli at extremelyhigh levels, others are notoriously difficult to ex-
press. Determining the critical factors responsiblefor poor expression is a challenging task for re-searchers. In this article we have attempted asystematic analysis to find out the bottlenecksresponsible for poor GM-CSF expression. Wefound that the production did not improve signifi-cantly even after growing the cells at a 5X antibi-otic concentration. This implies that increasingselection pressure is not the best way to resolvethe problem of instability, even for systems witha high rate of emergence of plasmid-free cells.RNA instability or codon bias too was not foundto be major contributors to the problem of lowexpression. Two distinct phenomena seem to limitthe expression of soluble rhuGM-CSF in E. coli.The first is protease-based degradation. We seethat without an amino-terminal peptide protec-tion, the rate of degradation of rhuGM-CSF in thecytoplasm is high. The protein levels start de-creasing after 1 h of induction. However when a6-His tag is attached to the amino terminus, thedegradation rate slows down. However, this doesnot automatically lead to an accumulation of theprotein. This is primarily because of the toxic ef-
Fig. 6. The recombinant granulocyte-macrophage colony-stimulating factor (GM-CSF) fusion proteins arebiologically active. Bar diagram shows the percentage viability of HL60 cells in excess of control (no GM-CSF)for the two different concentrations of the GM-CSF fusions used in this study.
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM113

114 Bhattacharya et al.
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
fect of GM-CSF on the host cells. Thus at onelevel, the production is inhibited by a feedbackmechanism and at another level the product isquickly degraded. The attachment of the pelB sig-nal peptide solves this problem by transportingthe protein to the periplasm, thus reducing thetoxic effect that it would have had on the cells byits cytoplasmic presence. The combination ofthese two strategies leads to greatly enhanced pro-duction levels. We were thus able to increase GM-CSF production from 0.8 µg/mL to approx 40 µg/Lusing this strategy. These results compare favorablywith reports from earlier investigators, where ex-pression levels for the soluble protein were typi-cally in the range of 0.45 mg/LOD600 units–1 (14)to approx 1.63 mg/L (27) using secretory systems.In many earlier reports of heterologous HuGM-CSFexpression only the final purified HuGM-CSF bio-activities have been reported, which translate tolow expression levels if one were to assume rea-sonable purification yields and standard bioactiv-ity of the expressed protein.
This strategy of coupling an amino-terminal tagand a signal peptide is not specific to the fusionpartners and the signal sequence. The same effectscan be observed with MBP as a fusion partner andthe MBP native signal peptide. The production ofMBP alongside the fusion has been observed byearlier investigators (28–30). The total amount offusion protein produced was higher in the case ofthe MBP fusion than the 6-His fusion, althoughthe latter was under the stronger T7 promoter,clearly demonstrating that promoter strength, andhence transcription rates, are not rate controlling.The possible reason for this could be the efficientscaffolding provided to rhuGM-CSF by the largerfusion partner leading to a more efficient reduc-tion in the toxic effect of GM-CSF. The bettergrowth rates observed with the MBP fusion pointto this possibility. However, on the whole, itseems to be more economic and efficient to use asmall fusion partner, as expression is on a one-to-one molar ratio and a larger fusion partner con-tributes negatively to the final yields of therecombinant protein. This strategy of using a fu-sion protein along with a signal peptide could possi-bly be used to overexpress a range of recombinant
proteins that are difficult to express in theirsoluble form because of their toxicity.
An additional advantage of periplasmic expres-sion is the presence of fewer contaminating pro-teins than in the cytoplasm. Amino-terminal tagslike 6-His and MBP also allow quick purificationof the rhuGM-CSF in a single step using affinitychromatography.
References1. Metcalf, D., Begley, C. G., Johnson, G. R., et al.
(1986) Biologic properties in vitro of a recombinanthuman granulocyte-macrophage colony-stimulatingfactor. Blood 67, 37–45.
2. Armitage, J. (1998) Emerging applications of recom-binant human granulocyte-macrophage colony-stimu-lating factor. Blood 92, 4491–4508.
3. Greenberg, P., Advani, R., Keating, A., ET AL.(1996) GM-CSF accelerates neutrophil recovery af-ter autologous hematopoietic stem cell transplanta-tion. Bone Marrow Transpl. 18, 1057–1064.
4. Anderson, J. E. and Appelbaum, F. R. (1994) He-matopoietic effects and clinical uses of granulocyte-macrophage colony-stimulating factor and PIXY321.Curr. Opin. Hematol. 1, 203–209.
5. Smith, T. and Grossberg, H. (1990) Successful use ofgranulocyte-macrophage colony-stimulating factor inpatients with acute lymphocytic leukemia. Am. J.Med. 89, 384–386.
6. Nemunaitis, J., Gordon, A., Cox, J., ET AL. (1994)Phase II pilot trial comparing neutrophil and mono-cyte function by microbicidal assay in oncology pa-tients receiving rhG-CSF, rHuGM-CSF or no cytokineafter cytotoxic chemotherapy. Blood 84, 135.
7. Rowe, J. M., Rubin, A., Mazza, J. J., et al. (1996)Incidence of infections in adult patients (>55 years)with acute myeloid leukemia treated with yeast-de-rived GM-CSF (sargramostim): results of a double-blind prospective study by the Eastern CooperativeOncology Group, in Acute Leukemias V: Experimen-tal Approaches and Management of Refractory Dis-ease (Hiddemann W., ed.). Springer-Verlag, Berlin,Germany, pp. 178.
8. Bodey, G. P., Anaissie, E., Gutterman, J., and Vadhan-Raj, S. (1993) Role of granulocyte-macrophage colony-stimulating factor as adjuvant therapy of fungal infectionin patients with cancer. Clin. Infect. Dis. 17, 705–707.
9. Dranoff, G., Jaffee, E., Lazenby, A., et al. (1993)Vaccination with irradiated tumor cells engineered tosecrete murine granulocyte-macrophage colony-stimulating factor stimulates potent, specific, andlong-lasting anti-tumor immunity. Proc. Natl. Acad.Sci. 90, 3539–3543.
10. Spitler, L. E., Grossbard, M. L., Ernstoff, M. S.,et al.(1997) Adjuvant therapy of stage III and IV malig-
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM114

Recombinant Human GM-CSF in E. coli 115
MOLECULAR BIOTECHNOLOGY Volume 30, 2005
nant melanoma using yeast derived, GM-CSF. Mela-noma Res. 7, 160.
11. Chachoua, A., Oratz, R., Liebes, L., et al. (1994) PhaseIB trial of granulocyte-macrophage colony-stimulatingfactor combined with murine monoclonal antibody R24in patients with metastatic melanoma. J. Immunother.Emphasis Tumor Immunol, 16, 132–141.
12. Yu, A. L., Batova, A., Alvarado, C., Rao, V. J., andCastleberry, R. P. (1997) Usefulness of a chimericanti-GD2 (ch14.18) and GM-CSF for refractory neu-roblastoma: a POG phase II study. Proc. Am. Soc.Clin. Oncol. 16, 513a.
13. Costello, R. T. (1993) Therapeutic use of granulo-cyte-macrophage colony-stimulating factor (GM-CSF). A review of recent experience. Acta Oncol. 32,403–408.
14. Berges, H., Joseph-Liauzun, E., and Fayet, O. (1996)Combined effects of the signal sequence and the majorchaperone proteins on the export of human cytokines inEscherichia coli. Appl. Environ. Microbiol. 62, 55–60.
15. Lundell, D., Lunn, C., Greenberg, R., et al. (1990)Exploiting the cell membrane for the production ofheterologous proteins in Escherichia coli. Biotechnol.Appl. Biochem. 12, 567–578.
16. Gronski, P., Badziong, W., Habermann, P., et al.(1988) E. coli derived human granulocyte-macroph-age colony-stimulating factor (rhGM-CSF) availablefor clinical trials. Behring. Inst. Mitt. 83, 246–249.
17. Wang, J., Zou, M., Huang, B., Zhang, M., and Mao,N. (1995) Molecular cloning and expression of hu-man granulocyte-macrophage colony stimulating fac-tor in Escherichia coli system. Chin. J. Biotechnol.11, 45–81.
18. Ling, M., Zou, M., Xu, M., Wang, J., and Ma, X.(1995) Purification of recombinant human granulo-cyte-macrophage colony stimulating factor expressedin Escherichia coli. Chin. J. Biotechnol. 11, 157–162.
19. Hua, Z., Wang, H., Chen, D., Chen, Y., and Zhu, D.(1994) Enhancement of expression of human granu-locyte-macrophage colony stimulating factor by argUgene product in Escherichia coli. Biochem Mol BiolInt. 32, 537–543.
20. Pluckthun, A. (1992) Mono- and bivalent antibody frag-ments produced in Escherichia coli: engineering, fold-ing and antigen binding. Immunol. Rev. 130, 151–188.
21. Bachmair, A., Finley, D., and Varshavsky, A. (1986)In vivo half-life of a protein is a function of its amino-terminal residue. Science 234, 179–186.
22. Nygren, P. A., Stahl, S., and Uhlen, M. (1994) Engi-neering proteins to facilitate bioprocessing. TrendsBiotechnol. 12, 184–188.
23. Ujihara, M., Nomura, K., Yamada, O., Shibata, N.,Kobayashi, M., and Takano, K. (2001) Granulocyte-macrophage colony-stimulating factor ensures mac-rophage survival and generation of the superoxideanion: a study using a monocytic-differentiated HL60subline. Free Radic. Biol. Med. 31, 1396–1404.
24. Sorensen, M. A., Kurland, C. G., and Pedersen, S.(1989) Codon usage determines translation rate inEscherichia coli. J. Mol. Biol. 207, 365–377.
25. Zhang, S. P., Zubay, G., and Goldman, E. (1991)Low-usage codons in Escherichia coli, yeast, fruit flyand primates. Gene 105, 61–72.
26. Petrovskaia, L. E., Kriukov, E. A., Iakimov, S. A., et al.(1995) Effect of the topography of the signal peptidasesite on the effectiveness of secretion of recombinanthuman granulocyte-macrophage colony-stimulatingfactor into Escherichia coli periplasm. Bioorg. Khim.21, 912–919.
27. Hua, Z., Jie, L., and Zhu, D. (1994) Expression of abiologically active human granulocyte-macrophagecolony stimulating factor fusion protein in Escheri-chia coli. Biochem. Mol. Bio. Int. 34, 621–626.
28. Eliseev, R., Alexandrov, A., and Gunter, T. (2004)High-yield expression and purification of p18 formof Bax as an MBP fusion protein. Protein Exp. Puri-fication 35, 206–209.
29. Pryor, K. D. and Leiting, B. (1997) High-level ex-pression of soluble protein in Escherichia coli usinga His6-Tag and maltose-binding protein double-affin-ity fusion system. Protein Exp. Purification 10, 309–319.
30. Guan, M., Su, B., Ye, C., and Yuan L. (2002) Pro-duction of extracellular domain of human tissue fac-tor using maltose-binding protein fusion system.Protein Exp. Purification 26, 229–234.
103_116_Mukherjee_MB04_0083_F 4/25/05, 11:19 AM115
Copyright © 2022 FDOKUMEN




















