Color Atlas of Humana Anatomy
420
At a Glance Introduction Basic Elements of the Nervous System Spinal Cord and Spinal Nerves Brain Stem and Cranial Nerves Cerebellum Diencephalon Telencephalon Cerebrovascular and Ventricular Systems Autonomic Nervous System Functional Systems The Eye The Ear Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme All rights reserved. Usage subject to terms and conditions of license.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Color Atlas of Humana Anatomy

I
At a Glance
Introduction
Basic Elements of theNervous System
Spinal Cord and SpinalNerves
Brain Stem and CranialNerves
Cerebellum
Diencephalon
Telencephalon
Cerebrovascular andVentricular Systems
Autonomic Nervous System
Functional Systems
The Eye
The Ear
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

III
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

IV
Color Atlas and Textbookof Human Anatomyin 3 volumes
Volume 1: Locomotor Systemby Werner Platzer
Volume 2: Internal Organsby Helmut Leonhardt
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

V
Thieme
Stuttgart · New York
Volume 3
Nervous System andSensory Organs
byWerner Kahle, M.D.Professor Emeritus
Institute of Neurology
University of Frankfurt/Main
Frankfurt/Main, Germany
Michael Frotscher, M.D.Professor
Anatomical Institute I
University of Freiburg
Freiburg, Germany
5th revised edition
179 color plates
Illustrations by Gerhard Spitzer
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

VI
Library of Congress Cataloging-in-PublicationDatais available from the publischer
1st German edition 19762nd German edition 19783rd German edition 19794th German edition 19825th German edition 19866th German edition 19917th German edition 2001
1st English edition 19782nd English edition 19843rd English edition 19864th English edition 1993
1st Dutch edition 19782nd Dutch edition 19813rd Dutch edition 19904th Dutch edition 20011st French edition 19792nd French edition 19931st Greek edition 19851st Hungarian edition 19961st Indonesian edition 19831st Italian edition 19792nd Italian edition 19873rd Italian edition 20011st Japanese edition 19792nd Japanese edition 19813rd Japanese edition 19844th Japanese edition 19901st Polish edition 19981st Serbo-Croatian edition 19911st Spanish edition 19772nd Spanish edition 19881st Turkish edition 1987
This book is an authorized and revised transla-tion of the 7th German edition published andcopyrighted 2001 by Georg Thieme Verlag,Stuttgart, Germany.Title of the German edition: Taschenatlas derAnatomie, Band 3: Nervensystem und Sinnes-organe
Translated byUrsula Vielkind, Ph. D., C. Tran.,Dundas, Ontario, Canada
Some of the product names, patents and regis-tered designs referred to in this book are in factregistered trademarks or proprietary nameseven though specific reference to this fact is notalways made in the text. Therefore, the appear-ance of a name without designation as pro-prietary is not to be construed as a representa-tion by the publisher that it is in the publicdomain.
This book, including all parts thereof, is le-gally protected by copyright. Any use, exploita-tion or commercialization outside the narrowlimits set by copyright legislation, without thepublisher’s consent, is illegal and liable to pros-ecution. This applies in particular to photostatreproduction, copying, mimeographing or du-plication of any kind, translating, preparation ofmicrofilms, and electronic data processing andstorage.
! 2003 Georg Thieme VerlagRüdigerstraße 14, D-70469 Stuttgart, Germanyhttp://www.thieme.deThieme New York, 333 Seventh Avenue,New York, N.Y. 10001 U.S.A.http://www.thieme.com
Cover design: Cyclus, Stuttgart
Typesetting by Druckhaus Götz GmbH,71636 Ludwigsburg
Printed in Germany by Appl, Wemding
ISBN 3-13-533505-4 (GTV)
ISBN 1-58890-064-9 (TNY) 1 2 3 4 5
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

VII
Preface to the 5th Edition of Volume 3
The number of students aswell as colleagues in the fieldwho have learned neuroanatomy ac-cording to volume 3 of the color atlas has been steadily increasing. Kahle’s textbook hasproved its worth. What should one do after taking on the job of carrying on with this textbook, other than leaving as much as possible as it is? However, the rapid growth in ourknowledge of neuroscience does not permit this. In just the last few years many new dis-coveries have beenmade that have shaped thewaywe view the structure and function of thenervous system. There was a need for updating and supplementing this knowledge. Hence,new sections have been added; for example, a section onmodernmethods of neuroanatomy,a section on neurotransmitter receptors, and an introduction to modern imaging proceduresfrequently used in the hospital. The Clinical Notes have been preserved and supplemented inorder to provide a link to the clinical setting. The purposewas to provide the student not onlywith a solid knowledge of neuroanatomy but alsowith an important foundation of interdisci-plinary neurocience. Furthermore, the student is introduced to the clinical aspects of thosefields in which neuroanatomy plays an important role. I sincerely hope that the use of mod-ernmulticolor printing hasmade it possible to present thingsmore clearly and in amore uni-formway. Thus, sensory pathways are now always presented in blue, motor pathways in red,paraympathetic fibers in green, and sympathetic fibers in yellow.
I wish to thank first and foremost Professor Gerhard Spitzer and Stephan Spitzer who tookcharge of the grapic design of the color atlas and provided their enormous experience also forthe present edition. I thank Professor Jürgen Hennig and his co-workers at the radiodiagnos-tic division of the Medical School of the Albert Ludwig University of Freiburg, Germany, fortheir help with the new section on imaging procedures. Last but not least, I would like tothank Dr. André Diesel who took great care in screening the text for lack of clarity and whocontributed significantly to the color scheme of the figures, al well as my secretary, Mrs.Regina Hummel, for her help with making the many corrections. My thanks go also to Mrs.Marianne Mauch and Dr. Jürgen Lüthje at Thieme Verlag, Stuttgart, for their generous adviceand their patience.
Michael FrotscherFall 2002
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

IX
Contents
The Nervous System . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
The Nervous System—An OverallView . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2Development and Subdivision . . . . 2Functional Circuits . . . . . . . . . . . . . . . . 2Position of the Nervous System inthe Body . . . . . . . . . . . . . . . . . . . . . . . . . 4
Development and Structure of theBrain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6Development of the Brain . . . . . . . . . 6Anatomy of the Brain . . . . . . . . . . . . . 8Evolution of the Brain . . . . . . . . . . . . . 14
Basic Elements of the Nervous System . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
The Nerve Cell . . . . . . . . . . . . . . . . . . . . . 18Methods in Neuroanatomy . . . . . . . . 20Ultrastructure of the Nerve Cell . . . 22
The Synapse . . . . . . . . . . . . . . . . . . . . . . . 24Localization . . . . . . . . . . . . . . . . . . . . . . 24Structure . . . . . . . . . . . . . . . . . . . . . . . . 24Function . . . . . . . . . . . . . . . . . . . . . . . . . 24Types of Synapses . . . . . . . . . . . . . . . . 26Neurotransmitters . . . . . . . . . . . . . . . . 26Axonal Transport . . . . . . . . . . . . . . . . . 28Transmitter Receptors . . . . . . . . . . . . 30Synaptic Transmission . . . . . . . . . . . . 30
Neuronal Systems . . . . . . . . . . . . . . . . . . 32Neuronal Circuits . . . . . . . . . . . . . . . . . 34
The Nerve Fiber . . . . . . . . . . . . . . . . . . . . 36Ultrastructure of the MyelinSheath . . . . . . . . . . . . . . . . . . . . . . . . . . 36Development of the Myelin Sheathin the PNS . . . . . . . . . . . . . . . . . . . . . . . . 38Development of UnmyelinatedNerve Fibers . . . . . . . . . . . . . . . . . . . . . 38Structure of the Myelin Sheath inthe CNS . . . . . . . . . . . . . . . . . . . . . . . . . . 38Peripheral Nerve . . . . . . . . . . . . . . . . . 40
Neuroglia . . . . . . . . . . . . . . . . . . . . . . . . . . 42
Blood Vessels . . . . . . . . . . . . . . . . . . . . . . 44
Spinal Cord and Spinal Nerves . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
Overview . . . . . . . . . . . . . . . . . . . . . . . . . . 48
The Spinal Cord . . . . . . . . . . . . . . . . . . . . 50Structure . . . . . . . . . . . . . . . . . . . . . . . . 50Reflex Arcs . . . . . . . . . . . . . . . . . . . . . . . 50Gray Substance and IntrinsicSystem . . . . . . . . . . . . . . . . . . . . . . . . . . 52Cross Sections of the Spinal Cord . . 54Ascending Pathways . . . . . . . . . . . . . . 56
Descending Pathways . . . . . . . . . . . . . 58Visualization of Pathways . . . . . . . . . 58Blood Vessels of the Spinal Cord . . . 60Spinal Ganglion and PosteriorRoot . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62Spinal Meninges . . . . . . . . . . . . . . . . . . 64Segmental Innervation . . . . . . . . . . . . 66Spinal Cord Syndromes . . . . . . . . . . . 68
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

X
Peripheral Nerves . . . . . . . . . . . . . . . . . . 70Nerve Plexusus . . . . . . . . . . . . . . . . . . . 70Cervical Plexus (C1–C4) . . . . . . . . . . 72Posterior Branches (C1–C8) . . . . . . . 72Brachial Plexus (C5–T1) . . . . . . . . . . 74Supraclavicular Part . . . . . . . . . . . . . . 74Infraclavicular Part . . . . . . . . . . . . . . . 74
Nerves of the Trunk . . . . . . . . . . . . . . . 84Posterior Branches . . . . . . . . . . . . . . . . 84Anterior Branches . . . . . . . . . . . . . . . . 84Lumbosacral Plexus . . . . . . . . . . . . . . . 86Lumbar Plexus . . . . . . . . . . . . . . . . . . . 86Sacral Plexus . . . . . . . . . . . . . . . . . . . . . 90
Otic Ganglion . . . . . . . . . . . . . . . . . . . . 130Submandibular Ganglion . . . . . . . . . . 130
Midbrain . . . . . . . . . . . . . . . . . . . . . . . . . . 132Structure . . . . . . . . . . . . . . . . . . . . . . . . 132Cross Section Through the InferiorColliculi of the Midbrain . . . . . . . . . . 132Cross Section Through the SuperiorColliculi of the Midbrain . . . . . . . . . . 134Cross Section Through the PretectalRegion of the Midbrain . . . . . . . . . . . . 134Red Nucleus and Substantia Nigra . 136
Eye-Muscle Nerves (Cranial NervesIII, IV, and VI) . . . . . . . . . . . . . . . . . . . . . . 138
Abducens Nerve . . . . . . . . . . . . . . . . . . 138Trochlear Nerve . . . . . . . . . . . . . . . . . . 138Oculomotor Nerve . . . . . . . . . . . . . . . . 138
Long Pathways . . . . . . . . . . . . . . . . . . . . . 140Corticospinal Tract and Cortico-nuclear Fibers . . . . . . . . . . . . . . . . . . . . 140Medial Lemniscus . . . . . . . . . . . . . . . . 140Medial Longitudinal Fasciculus . . . . 142Internuclear Connections of theTrigeminal Nuclei . . . . . . . . . . . . . . . . 142Central Tegmental Tract . . . . . . . . . . . 144Posterior Longitudinal Fasciculus . . 144
Reticular Formation . . . . . . . . . . . . . . . 146
Histochemistry of the Brain Stem . . 148
Brain Stem and Cranial Nerves . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99
Overview . . . . . . . . . . . . . . . . . . . . . . . . . . 100Longitudinal Organization . . . . . . . . . 102Cranial Nerves . . . . . . . . . . . . . . . . . . . . 102Base of the Skull . . . . . . . . . . . . . . . . . . 104
Cranial Nerve Nuclei . . . . . . . . . . . . . . . 106
Medulla Oblongata . . . . . . . . . . . . . . . . 108Cross Section at the Level of theHypoglossal Nerve . . . . . . . . . . . . . . . . 108Cross Section at the Level of theVagus Nerve . . . . . . . . . . . . . . . . . . . . . 108
Pons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110Cross Section at the Level of theGenu of the Facial Nerve . . . . . . . . . . 110Cross Section at the Level of theTrigeminal Nerve . . . . . . . . . . . . . . . . . 110
Cranial Nerves (V, VII – XII) . . . . . . . . . 112Hypoglossal Nerve . . . . . . . . . . . . . . . . 112Accessory Nerve . . . . . . . . . . . . . . . . . . 112Vagus Nerve . . . . . . . . . . . . . . . . . . . . . 114Glossopharyngeal Nerve . . . . . . . . . . 118Vestibulocochlear Nerve . . . . . . . . . . 120Facial Nerve . . . . . . . . . . . . . . . . . . . . . . 122Trigeminal Nerve . . . . . . . . . . . . . . . . . 124
Parasympathetic Ganglia . . . . . . . . . . . 128Ciliary Ganglion . . . . . . . . . . . . . . . . . . 128Pterygopalatine Ganglion . . . . . . . . . 128
Contents
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

XIContents
Cerebellum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 151
Structure . . . . . . . . . . . . . . . . . . . . . . . . . . 152Subdivision . . . . . . . . . . . . . . . . . . . . . . 152Cerebellar Peduncles and Nuclei . . 154Cerebellar Cortex . . . . . . . . . . . . . . . . . 156Neuronal Circuits . . . . . . . . . . . . . . . . . 160
Functional Organization . . . . . . . . . . . 162Fiber Projection . . . . . . . . . . . . . . . . . . 162Results of ExperimentalStimulation . . . . . . . . . . . . . . . . . . . . . . 162
Pathways . . . . . . . . . . . . . . . . . . . . . . . . . . 164Inferior Cerebellar Peduncle(Restiform Body) . . . . . . . . . . . . . . . . . 164Middle Cerebellar Peduncle(Brachium Pontis) . . . . . . . . . . . . . . . . 166Superior Cerebellar Peduncle(Brachium conjunctivum) . . . . . . . . . 166
Diencephalon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169
Development of the Prosen-cephalon . . . . . . . . . . . . . . . . . . . . . . . . . . 170
Telodiencephalic Boundary . . . . . . . . 170
Structure . . . . . . . . . . . . . . . . . . . . . . . . . . 172Subdivision . . . . . . . . . . . . . . . . . . . . . . 172Frontal Section at the Levelof the Optic Chasm . . . . . . . . . . . . . . . 172Frontal Section through theTuber Cinereum . . . . . . . . . . . . . . . . . . 174Frontal Section at the Levelof the Mamillary Bodies . . . . . . . . . . . 174
Epithalamus . . . . . . . . . . . . . . . . . . . . . . . 176Habenula . . . . . . . . . . . . . . . . . . . . . . . . 176Pineal Gland . . . . . . . . . . . . . . . . . . . . . 176
Dorsal Thalamus . . . . . . . . . . . . . . . . . . . 178Specific Thalamic Nuclei . . . . . . . . . . 178Nonspecific Thalamic Nuclei . . . . . . 180Anterior Nuclear Group . . . . . . . . . . . 182Medial Nuclear Group . . . . . . . . . . . . 182Centromedian Nucleus . . . . . . . . . . . . 182Lateral Nuclear Group . . . . . . . . . . . . . 184Ventral Nuclear Group . . . . . . . . . . . . 184Lateral Geniculate Body . . . . . . . . . . . 186Medial Geniculate Body . . . . . . . . . . . 186Pulvinar . . . . . . . . . . . . . . . . . . . . . . . . . 186
Frontal Section Through the RostralThalamus . . . . . . . . . . . . . . . . . . . . . . . . 188Frontal Section Through the CaudalThalamus . . . . . . . . . . . . . . . . . . . . . . . . 190
Subthalamus . . . . . . . . . . . . . . . . . . . . . . 192Subdivision . . . . . . . . . . . . . . . . . . . . . . 192Responses to Stimulation of theSubthalamus . . . . . . . . . . . . . . . . . . . . . 192
Hypothalamus . . . . . . . . . . . . . . . . . . . . . 194Poorly Myelinated Hypothalamus . 194Richly Myelinated Hypothalamus . 194Vascular Supply . . . . . . . . . . . . . . . . . . 196Fiber Connections of the PoorlyMyelinated Hypothalamus . . . . . . . . 196Fiber Connections of the RichlyMyelinated Hypothalamus . . . . . . . . 196Functional Topography of theHypothalamus . . . . . . . . . . . . . . . . . . . 198
Hypothalamus and Hypophysis . . . . . 200Development and Subdivision ofthe Hypophysis . . . . . . . . . . . . . . . . . . . 200Infundibulum . . . . . . . . . . . . . . . . . . . . 200Blood Vessels of the Hypophysis . . 200Neuroendocrine System . . . . . . . . . . . 202
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

XII
Telencephalon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 207
Overview . . . . . . . . . . . . . . . . . . . . . . . . . . 208Subdivision of the Hemisphere . . . . 208Rotation of the Hemisphere . . . . . . . 208Evolution . . . . . . . . . . . . . . . . . . . . . . . . 210Cerebral Lobes . . . . . . . . . . . . . . . . . . . 212
Sections Through the Telen-cephalon . . . . . . . . . . . . . . . . . . . . . . . . . . 214Frontal Sections . . . . . . . . . . . . . . . . . . 214Horizontal Sections . . . . . . . . . . . . . . . 220
Paleocortex and Amygdaloid Body . 224Paleocortex . . . . . . . . . . . . . . . . . . . . . . 224Amygdaloid Body . . . . . . . . . . . . . . . . . 226Fiber Connections . . . . . . . . . . . . . . . . 228
Archicortex . . . . . . . . . . . . . . . . . . . . . . . . 230Subdivision and Functional Signifi-cance . . . . . . . . . . . . . . . . . . . . . . . . . . . . 230Ammon’s Horn . . . . . . . . . . . . . . . . . . . 232Fiber Connections . . . . . . . . . . . . . . . . 232Hippocampal Cortex . . . . . . . . . . . . . . 234
Neostriatum . . . . . . . . . . . . . . . . . . . . . . . 236
Insula . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 238
Neocortex . . . . . . . . . . . . . . . . . . . . . . . . . 240Cortical Layers . . . . . . . . . . . . . . . . . . . 240Vertical Columns . . . . . . . . . . . . . . . . . 240Cell Types of the Neocortex . . . . . . . 242The Module Concept . . . . . . . . . . . . . . 242Cortical Areas . . . . . . . . . . . . . . . . . . . . 244Frontal Lobe . . . . . . . . . . . . . . . . . . . . . . 246Parietal Lobe . . . . . . . . . . . . . . . . . . . . . 250Temporal Lobe . . . . . . . . . . . . . . . . . . . 252Occipital Lobe . . . . . . . . . . . . . . . . . . . . 254Fiber Tracts . . . . . . . . . . . . . . . . . . . . . . 258Hemispheric Asymmetry . . . . . . . . . . 262
Imaging Procedures . . . . . . . . . . . . . . . 264Contrast Radiography . . . . . . . . . . . . . 264Computed Tomography . . . . . . . . . . . 264Magnetic Resonance Imaging . . . . . . 266PET and SPECT . . . . . . . . . . . . . . . . . . . 266
Cerebrovascular and Ventricular Systems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 269
Cerebrovascular System . . . . . . . . . . . . 270Arteries . . . . . . . . . . . . . . . . . . . . . . . . . . . . 270Internal Carotid Artery . . . . . . . . . . . 272Areas of Blood Supply . . . . . . . . . . . . 274
Veins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 276Superficial Cerebral Veins . . . . . . . . . 276Deep Cerebral Veins . . . . . . . . . . . . . . 278
Cerebrospinal Fluid Spaces . . . . . . . . . 280Overview . . . . . . . . . . . . . . . . . . . . . . . . 280Choroid Plexus . . . . . . . . . . . . . . . . . . . 282Ependyma . . . . . . . . . . . . . . . . . . . . . . . 284Circumventricular Organs . . . . . . . . . 286
Meninges . . . . . . . . . . . . . . . . . . . . . . . . . . 288Dura Mater . . . . . . . . . . . . . . . . . . . . . . . 288Arachnoidea Mater . . . . . . . . . . . . . . . 288Pia Mater . . . . . . . . . . . . . . . . . . . . . . . . 288
Contents
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

XIIIContents
Autonomic Nervous System . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 291
Overview . . . . . . . . . . . . . . . . . . . . . . . . . . 292Central Autonomic System . . . . . . . . 292Peripheral Autonomic System . . . . . 294Adrenergic and CholinergicSystems . . . . . . . . . . . . . . . . . . . . . . . . . . 294Neuronal Circuit . . . . . . . . . . . . . . . . . . 296
Sympathetic Trunk . . . . . . . . . . . . . . . . 296Cervical and Upper ThoracicSegments . . . . . . . . . . . . . . . . . . . . . . . . 296
Lower Thoracic and AbdominalSegments . . . . . . . . . . . . . . . . . . . . . . . . 298Innervation of the Skin . . . . . . . . . . . . 298
Autonomic Periphery . . . . . . . . . . . . . . 300Efferent Fibers . . . . . . . . . . . . . . . . . . . . 300Afferent Fibers . . . . . . . . . . . . . . . . . . . 300Intramural Plexus . . . . . . . . . . . . . . . . 300Autonomic Neurons . . . . . . . . . . . . . . 302
Functional Systems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 305
Brain Function . . . . . . . . . . . . . . . . . . . . . 306
Motor Systems . . . . . . . . . . . . . . . . . . . . . 308Corticospinal Tract . . . . . . . . . . . . . . . . 308Extrapyramidal Motor System . . . . . 310Motor End Plate . . . . . . . . . . . . . . . . . . 312Tendon Organ . . . . . . . . . . . . . . . . . . . . 312Muscle Spindle . . . . . . . . . . . . . . . . . . . 314Common Terminal MotorPathway . . . . . . . . . . . . . . . . . . . . . . . . . 316
Sensory Systems . . . . . . . . . . . . . . . . . . . 318Cutaneous Sensory Organs . . . . . . . . 318Pathway of the EpicriticSensibility . . . . . . . . . . . . . . . . . . . . . . . 322
Pathway of the ProtopathicSensibility . . . . . . . . . . . . . . . . . . . . . . . 324Gustatory Organ . . . . . . . . . . . . . . . . . . 326Olfactory Organ . . . . . . . . . . . . . . . . . . 330
Limbic System . . . . . . . . . . . . . . . . . . . . . 332Overview . . . . . . . . . . . . . . . . . . . . . . . . 332Cingulate Gyrus . . . . . . . . . . . . . . . . . . 334Septal Area . . . . . . . . . . . . . . . . . . . . . . . 334
Sensory Organs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 337
The Eye . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 337
Structure . . . . . . . . . . . . . . . . . . . . . . . . . . 338Eyelids, Lacrimal Apparatus, andOrbital Cavity . . . . . . . . . . . . . . . . . . . . 338Muscles of the Eyeball . . . . . . . . . . . . 340The Eyeball, Overview . . . . . . . . . . . . 342Anterior Part of the Eye . . . . . . . . . . . 344Vascular Supply . . . . . . . . . . . . . . . . . . 346Fundus of the Eye . . . . . . . . . . . . . . . . . 346Retina . . . . . . . . . . . . . . . . . . . . . . . . . . . 348
Optic Nerve . . . . . . . . . . . . . . . . . . . . . . 350Photoreceptors . . . . . . . . . . . . . . . . . . . 352
Visual Pathway and OcularReflexes . . . . . . . . . . . . . . . . . . . . . . . . . . . 354
Visual Pathway . . . . . . . . . . . . . . . . . . . 354Topographic Organization of theVisual Pathway . . . . . . . . . . . . . . . . . . . 356Ocular Reflexes . . . . . . . . . . . . . . . . . . . 358
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

XIV
The Ear . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 361
Structure . . . . . . . . . . . . . . . . . . . . . . . . . . 362Overview . . . . . . . . . . . . . . . . . . . . . . . . 362Outer Ear . . . . . . . . . . . . . . . . . . . . . . . . 362Middle Ear . . . . . . . . . . . . . . . . . . . . . . . 364Inner Ear . . . . . . . . . . . . . . . . . . . . . . . . . 368Cochlea . . . . . . . . . . . . . . . . . . . . . . . . . . 370Organ of Corti . . . . . . . . . . . . . . . . . . . . 372
Organ of Balance . . . . . . . . . . . . . . . . . 374Vestibular Sensory Cells . . . . . . . . . . . 376
Auditory Pathway and VestibularPathways . . . . . . . . . . . . . . . . . . . . . . . . . . 378
Auditory Pathway . . . . . . . . . . . . . . . . 378Vestibular Pathways . . . . . . . . . . . . . . 382
Further Reading . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 384
Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 390
Contents
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

The NervousSystem
Introduction
The Nervous System—An OverallView 2
Development and Structure of theBrain 6
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

2Introd
uction
Introduction
The Nervous System—AnOverall View
Development and Subdivision(A–D)
The nervous system serves informationprocessing. In the most primitive forms oforganization (A), this function is assumedby the sensory cells (A–C1) themselves.These cells are excited by stimuli comingfrom the environment; the excitation isconducted to amuscle cell (A–C2) througha cellular projection, or process. The simplestresponse to environmental stimuli isachieved in this way. (In humans, sensorycells that still have processes of their ownare only found in the olfactory epithelium.)In more differentiated organisms (B), an ad-ditional cell is interposed between thesensory cell and the muscle cell – the nervecell, or neuron (BC3) which takes on thetransmission of messages. This cell cantransmit the excitation to several musclecells or to additional nerve cells, thus form-ing a neural network (C). A diffuse networkof this type also runs through the humanbody and innervates all intestinal organs,blood vessels, and glands. It is called the au-tonomic (visceral, or vegetative) nervoussystem (ANS), and consists of two com-ponents which often have opposing func-tions: the sympathetic nervous system and theparasympathetic nervous system. The interac-tion of these two systems keeps the interiororganization of the organism constant.
In vertebrates, the somatic nervous systemdeveloped in addition to the autonomicnervous system; it consists of the centralnervous system (CNS; brain and spinal cord),and the peripheral nervous system (PNS; thenerves of head, trunk, and limbs). It is re-sponsible for conscious perception, for vol-untary movement, and for the processing ofinformation (integration). Note that mosttextbooks include the peripheral nerves ofthe autonomic nervous system in the PNS.
The CNS develops from the neural plate (D4)of the ectoderm which then transforms intothe neural groove (D5) and further into the
neural tube (D6). The neural tube finallydifferentiates into the spinal cord (D7) andthe brain (D8).
Functional Circuits (E, F)
The nervous system, the remaining or-ganism, and the environment are function-ally linked with each other. Stimuli from theenvironment (exteroceptive stimuli) (E9) areconducted by sensory cells (E10) viasensory (afferent) nerves (E11) to the CNS(E12). In response, there is a command fromthe CNS via motor (efferent) nerves (E13)to themuscles (E14). For control and regula-tion of the muscular response (E15), there isinternal feedback from sensory cells in themuscles via sensory nerves (E16) to theCNS. This afferent tract does not transmitenvironmental stimuli but stimuli fromwithin the body (proprioceptive stimuli). Wetherefore distinguish between exterocep-tive and proprioceptive sensitivities.
However, the organism does not only re-spond to the environment; it also influencesit spontaneously. In this case, too, there is acorresponding functional circuit: the action(F17) started by the brain via efferent nerves(F13) is registered by sensory organs (F10),which return the corresponding informa-tion via afferent nerves (F11) to the CNS(F12) (reafference, or external feedback). De-pending on whether or not the result meetsthe desired target, the CNS sends out furtherstimulating or inhibiting signals (F13).Nervous activity is based on a vast numberof such functional circuits.
In the same way as we distinguish betweenexteroceptive sensitivity (skin and mucosa)and proprioceptive sensitivity (receptors inmuscles and tendons, autonomic sensorysupply of the intestines), we can subdividethe motor system into an environment-oriented ecotropic somatomotor system(striated, voluntary muscles) and an idio-tropic visceromotor system (smooth intestinalmuscles).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

3
Introd
uction
121110
9
15 13
14
16
14 13 12
17
11
10
1
2
4
5
6
7 8
7 8
78
2
3
11
3
Development of the Nervous System, Functional Circuits
A–C Models of primitive nervous systems (according to Parker and Bethe)
A Sensory cell withprocess to amuscle cell C Diffuse neural network
D Embryonic developmentof the central nervous system:spinal cord on the left, brain onthe right
E Functional circuit: response of anorganism to environmentalstimuli
F Functional circuit: influence of anorganism on its environment
B Nerve cell connectinga sensory cell and amuscle cell
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

4Introd
uction
Introduction: The Nervous System—An Overall View
Position of the Nervous System inthe Body (A, B)
The central nervous system (CNS) isdivided into the brain, encephalon (A1), andthe spinal cord (SC), medulla spinalis (A2).The brain in the cranial cavity is surroundedby a bony capsule; the spinal cord in thevertebral canal is enclosed by the bonyvertebral column. Both are covered bymeninges that enclose a cavity filled with afluid, the cerebrospinal fluid. Thus, the CNS isprotected from all sides by bony walls andthe cushioning effect of a fluid (fluid cush-ion).
The peripheral nervous system (PNS) in-cludes the cranial nerves, which emergethrough holes (foramina) in the base of theskull, and the spinal nerves, which emergethrough spaces between the vertebrae (in-tervertebral foramina) (A3). The peripheralnerves extend to muscles and skin areas.They form nerve plexuses before enteringthe limbs: the brachial plexus (A4) and thelumbosacral plexus (A5) in which the fibers ofthe spinal nerves intermingle; as a result,the nerves of the limbs contain portions ofdifferent spinal nerves (see pp. 70 and 86).At the entry points of the afferent nervefibers lie ganglia (A6); these are small ovalbodies containing sensory neurons.
When describing brain structures, termslike “top,” “bottom,” “front,” and “back” areinaccurate, because we have to distinguishbetween different axes of the brain (B).Owing to the upright posture of humans,the neural tube is bent; the axis of the spinalcord runs almost vertically, while the axis ofthe forebrain (Forel’s axis, orange) runs hori-zontally; the axis of the lower brain divi-sions (Meinert’s axis, violet) runs obliquely.The positional terms relate to theses axes:the anterior end of the axis is called oral orrostral (os, mouth; rostrum, beak), the pos-terior end is called caudal (cauda, tail), theunderside is called basal or ventral (venter,abdomen), and the upper side is called dor-sal (dorsum, back).
The lower brain divisions, which mergeinto the spinal cord, are collectively called
the brain stem (light gray) (B7). The anteriordivision is called the forebrain (gray) (B8).
The divisions of the brain stem, or encephalictrunk, have a common structural plan (con-sisting of basal plate and alar plate, like thespinal cord, see p. 13, C). Genuine peripheralnerves emerge from these divisions, as theydo from the spinal cord. Like the spinal cord,they are supported by the chorda dorsalisduring embryonic development. All thesefeatures distinguish the brain stem from theforebrain. The subdivision chosen herediffers from the other classifications inwhich the diencephalon is viewed as part ofthe brain stem.
The forebrain, prosencephalon, consists oftwo parts, the diencephalon and the telen-cephalon or cerebrum. In the mature brain,the telencephalon forms the two hemi-spheres (cerebral hemispheres). The dien-cephalon lies between the two hemi-spheres.
A9 Cerebellum.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

5
Introd
uction
4
4
7
8
1
9
2
3
55
6
Position of the Nervous System
A Position of the central nervous system in the body
B Axes of the brain:median section through the brain
dorsal
caudal oral (rostral)
oral (rostral)
ventraldorsal
caudalventral
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

6Introd
uction
Introduction
Development and Structureof the Brain
Development of the Brain (A–E)
The closure of the neural groove into theneural tube begins at the level of the uppercervical cord. From here, further closureruns in the oral direction up to the rostralend of the brain (oral neuropore, later theterminal lamina) and in the caudal directionup to the end of the spinal cord. Furtherdevelopmental events in the CNS proceed inthe same directions. Thus, the brain’s divi-sions do not mature simultaneously but atintervals (heterochronous maturation).
The neural tube in the head region expandsinto several vesicles (p. 171, A). The rostralvesicle is the future forebrain, prosen-cephalon (yellow and red); the caudal ves-icles are the future brain stem, encephalictrunk (blue). Two curvatures of the neuraltube appear at this time: the cephalicflexure (A1) and the cervical flexure (A2).Although the brain stem still shows a uni-form structure at this early stage, the futuredivisions can already be identified: medullaoblongata (elongated cord) (A–D3), pons(bridge of Varolius) (A–D4), cerebellum(A–D5, dark blue), andmesencephalon (mid-brain) (A–C6, green). The brain stem isdevelopmentally ahead of the prosen-cephalon; during the second month ofhuman development, the telencephalon isstill a thin-walled vesicle (A), whereas neu-rons have already differentiated in the brainstem (emergence of cranial nerves) (A7). Theoptic vesicle develops from the diencephalon(AB8, red) (p. 343, A) and forms the opticcup (A9). Anterior to it lies the telencephalicvesicle (telencephalon) (A–D10, yellow); ini-tially, its anlage is unpaired (impar telen-cephalon), but it soon expands on both sidesto form the two cerebral hemispheres.
During the third month, the prosen-cephalon enlarges (B). Telencephalon anddiencephalon become separated by thetelodiencephalic sulcus (B11). The anlage ofthe olfactory bulb (B–D12) has formed atthe hemispheric vesicle, and the pituitary
anlage (B13) (p. 201B) and the mamillaryeminence (B14) have formed at the base ofthe diencephalon. A deep transverse sulcus(B15) is formed between the cerebellar an-lage and themedulla oblongata as a result ofthe pontine flexure; the underside of thecerebellum comes to lie in apposition to themembrane-thin dorsal wall of the medulla(p. 283, E).
During the fourth month, the cerebralhemispheres begin to overgrow the otherparts of the brain (C). The telencephalon,which initially lagged behind all other braindivisions in its development, now exhibitsthe most intense growth (p. 170, A). Thecenter of the lateral surface of each hemi-sphere lags behind in growth and later be-comes overlain with parts. This is the insula(CD16). During the sixth month, the insulastill lies free (D). The first grooves and con-volutions appear on the previously smoothsurfaces of the hemispheres. The initiallythin walls of neural tube and brain vesicleshave thickened during development. Theycontain the neurons and nerve tracts thatmake up the brain substance proper. (Fordevelopment of cerebral hemispheres, seep. 208.)
Within the anterior wall of the impar telen-cephalon, nerve fibers run from one hemi-sphere to the other. The commissural sys-tems, which connect the two hemispheres,develop in this segment of the thickenedwall, or commissural plate. The largest ofthem is the corpus callosum (E). The hemi-spheres grow mainly in the caudal direc-tion; in parallel with their increase in size,the corpus callosum also expands in thecaudal direction during its developmentand finally overlies the diencephalon.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

7
Introd
uction
16
10
10
16
12
43
1
6
5
4
3
2
8
10
79
6
6
5
14
15
5
3
11
10
8
12
13 4
3
12 4
5
Development of the Brain
A–D The brain in human embryosof different crown-rumplengths (CRL)
A In an embryo of 10 mm CRL B In an embryo of27 mm CRL
C In an embryo of 53 mm CRL
D In a fetus of 33 cm CRLE Development of the corpus callosum
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

8Introd
uction
Introduction: Development and Structure of the Brain
Anatomy of the Brain (A–E)
Overview
The individual subdivisions of the braincontain cavities or ventricles of variousshapes andwidths. The primary cavity of theneural tube and cerebral vesicle becomesmuch narrower as the walls thicken. In thespinal cord of lower vertebrates, it survivesas the central canal. In the human spinalcord, it becomes completely occluded (ob-literated). In a cross section, only a few cellsof the former lining of the spinal cord markthe site of the early central canal (A1). In thebrain, the cavity survives and forms theventricular system (p. 280) which is filledwith a clear fluid, the cerebrospinal fluid.The fourth ventricle (AD2) is located in thesegment of the medulla oblongata and thepons. After a narrowing of the cavity in themidbrain, the third ventricle (CD3) lies in thediencephalon. A passage on both sides of itslateral walls, the interventricular foramen(foramen of Monro) (C–E4), opens into thelateral ventricles (CE5) (first and second ven-tricles) of both cerebral hemispheres.
In frontal sections through the hemispheres(C), the lateral ventricles are seen twice; theyhave a curved appearance (E). This shape iscaused by the crescent-shaped growth ofthe hemispheres (rotation of hemispheres,p. 208, C) which do not expand equally in alldirections during development. In themiddle of the semicircle is the insula. It liesdeep in the lateral wall of the hemisphereon the floor of the lateral fossa (C6) and isoverlain by the adjacent parts, the opercula(C7), so that the surface of the hemisphereshows only a deep groove, the lateral sulcus(lateral fissure, fissure of Sylvius) (BC8). Eachhemisphere is subdivided into several cere-bral lobes (B) (p. 212): frontal lobe (B9),parietal lobe (B10), occipital lobe (B11), andtemporal lobe (B12).
The diencephalon (dark gray in C, D) andbrain stem essentially become overlain bythe cerebral hemispheres, thus renderedvisible only at the base of the brain or in alongitudinal section through the brain. In amedian section (D), the subdivisions of the
brain stem can be recognized: medulla ob-longata (D13), pons (D14), mesencephalon(D15), and cerebellum (D16). The fourth ven-tricle (D2) is seen in its longitudinal dimen-sion. On its tentlike roof rests the cerebel-lum. The third ventricle (D3) is opened in itsentire width. In its rostral section, the inter-ventricular foramen (D4) leads into thelateral ventricle. Above the third ventriclelies the corpus callosum (D17); this fiberplate, seen here in cross section, connectsthe two hemispheres.
Weight of the Brain
The average weight of the human brainranges between 1250g and 1600g. It is re-lated to body weight: a heavier person usu-ally has a heavier brain. The average weightof a male brain is 1350 g, that of a femalebrain 1250 g. By the age of 20, the brain issupposed to have reached its maximumweight. In old age, the brain usually losesweight owing to age-related atrophy. Theweight of the brain does not indicate the in-telligence of a person. Examination of thebrains of prominent people (“elite brains”)yielded the usual variations.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

9
Introd
uction
9
8
10
17
11
12
12 2
5 4
7687
53
4 5
4
3
2 13
1615
14
17
9
8
10
17
11
12
12 2
5 4
7687
53
4 5
4
3
2 13
1615
14
17
9
8
10
17
11
12
12 2
5 4
7687
53
4 5
4
3
2 13
1615
14
17
Anatomy of the Brain, Overview
A Sections through the spinal cord and brainstem, all on the same scale
Spinal cordMedulla oblongata
Pons Midbrain
B Lateral view of the brain, diagram
C Frontal section through the brain,diagram
D Longitudinal median sectionthrough the brain, diagram
E Longitudinal paramedian sectionthrough the brain, diagram
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

10Introd
uction
Introduction: Development and Structure of the Brain
Lateral and Dorsal Views (A, B)
The two cerebral hemispheres overlie allother parts of the brain; only the cerebellum(A1) and the brain stem (A2) are visible. Thesurface of the cerebral hemisphere ischaracterized by a large number of grooves,or sulci, and convolutions, or gyri. Beneaththe surface of the relief of gyri lies the cere-bral cortex, the highest nervous organ: con-sciousness, memory, thought processes,and voluntary activities all depend on theintegrity of the cortex. The expansion of thecerebral cortex is increased through the for-mation of sulci and gyri. Only one-third ofthe cortex lies on the surface, while two-thirds lie in the depth of the gyri. As shownby the dorsal view (B), the hemispheres areseparated by a deep groove, the longitudinalcerebral fissure (B3). On the lateral surface ofthe hemisphere lies the lateral sulcus (sulcusof Sylvius) (A4). A frontal section (pp. 9, 215,and 217) clearly shows that this is not asimple sulcus but a deep pit, the lateral fossa.
The anterior pole of the hemisphere iscalled the frontal pole (A5), the posteriorone is called the occipital pole (A6). Thecerebral hemisphere is subdivided intoseveral lobes: the frontal lobe (A7) and theparietal lobe (A9), which are separated by thecentral sulcus (A8), the occipital lobe (A10),and the temporal lobe (A11). The central sul-cus separates the precentral gyrus (A12) (re-gion of voluntary movement) from the post-central gyrus (A13) (region of sensitivity).Both together constitute the central region.
Median Section (C)
Between the hemispheres lies the dien-cephalon (C14); the corpus callosum (C15)above it connects the two hemispheres. Thecorpus callosum forms a fiber plate; its oralcurvature encloses a thin wall segment ofthe hemisphere, the septum pellucidum (C16)(p. 221, B18). The third ventricle (C17) isopened. The adhesion of its two walls formsthe interthalamic adhesion (C18). The fornix(C19) forms an arch above it. In the anteriorwall of the third ventricle lies the anteriorcommissure (C20) (containing the crossingfibers of the olfactory brain); at its base lie
the decussation of the optic nerve, or opticchiasm (C21), the hypophysis (C22), and thepaired mamillary bodies (C23); in the caudalwall lies the pineal gland, or epiphysis (C24).
The third ventricle is connected with thelateral ventricle of the hemisphere throughthe interventricular foramen (foramen ofMonro) (C25); it turns caudally into the cere-bral aqueduct (aqueduct of Sylvius) (C26)which passes through the midbrain andwidens like a tent to form the fourth ven-tricle (C27) underneath the cerebellum. Onthe cut surface of the cerebellum (C28), thesulci and gyri form the arbor vitae (“tree oflife”). Rostral to the cerebellum lies thequadrigeminal plate, or tectal lamina (C29),of the midbrain (a relay station for optic andacoustic tracts). The pons (C30) bulges at thebase of the brain stem and turns into theelongated cord, or medulla oblongata (C31),which turns into the spinal cord.
C32 Choroid plexus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

11
Introd
uction
9
10
6
12
3
7
14
9
24
29
2827
11
5
78
12
4
13
23
30
22
2120
31
26
17
18
19
32
15
25
16
Anatomy of the Brain, Lateral View and Median Section
A Lateral view of the brain
B Dorsal view
C Median section through the brain,medial surface of the right hemisphere
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

12Introd
uction
Introduction: Development and Structure of the Brain
Base of the Brain (A)
The basal aspect of the brain provides anoverview of the brain stem, the ventral sur-faces of frontal lobe (A1) and temporal lobe(A2), and the base of the diencephalon. Thelongitudinal cerebral fissure (A3) separatesthe two frontal lobes; at the basal surface ofeach hemisphere lies the olfactory lobewiththe olfactory bulb (A4) and the olfactory tract(A5). The tract divides in the olfactory trigone(A6) into two olfactory striae which borderthe anterior perforated substance (A7); the lat-ter is perforated by entering blood vessels.At the optic chiasm (A8), or decussation ofthe optic nerves (A9), the base of the dien-cephalon begins with the hypophysis (A10)and the mamillary bodies (A11). The pons(A12) bulges caudally and is followed by themedulla oblongata (A13). Numerous cranialnerves emerge from the brain stem. Thecerebellum is divided into themedial, deep-lying vermis of the cerebellum (A14) and thetwo cerebellar hemispheres (A15).
White and Gray Matter (B)
Upon dissecting the brain into slices, thewhite and gray matter, substantia alba etgrisea, become visible on the cut surfaces.The gray matter represents a concentrationof neurons and the white matter the fibertracts, or neuronal processes, which appearlight because of their white envelope, themyelin sheath. In the spinal cord (B16), thegray matter lies in the center and is en-closed by the bordering white matter (as-cending and descending fiber tracts). In thebrain stem (B17) and diencephalon, the dis-tribution of gray and white matter varies.The gray areas are called nuclei. In the telen-cephalon (B18), the gray matter lies at theouter margin and forms the cortex, whilethe white matter lies inside. Thus, the dis-tribution here is the reverse of that in thespinal cord.
The arrangement in the spinal cord repre-sents a primitive state; it still exists in fishand amphibians where the neurons are in aperiventricular position even in the telen-cephalon. The cerebral cortex representsthe highest level of organization, which isfully developed only in mammals.
Subdivision into Longitudinal Zones (C)
During development, the neural tube is sub-divided into longitudinal zones. The ventralhalf of the lateral wall, which differentiatesearly, is called the basal plate (C19) and rep-resents the origin of motor neurons. The dor-sal half, which develops later, is called thealar plate (C20) and represents the origin ofsensory neurons. Between alar and basalplates lies a segment (C21) from which au-tonomic neurons originate. Thus, a struc-tural plan of the CNS can be recognized inthe spinal cord and brain stem, knowledgeof which will aid in understanding the or-ganization of various parts of the brain.
The derivatives of basal and alar plates aredifficult to identify in diencephalon and tel-encephalon. Many authors therefore rejectsuch a classification of the forebrain.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

13
Introd
uction
5
4
3
98
1011
13
12
2
6
1
7
15
14
16
20
21
19
17 18
Base of the Brain, White and Gray Matter, Subdivision into Lateral Zones
A Basal view of the brain
B Distribution of white and gray matter
C Longitudinal zones of the CNS
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

14Introd
uction
Introduction: Development and Structure of the Brain
Evolution of the Brain (A–C)
In the course of evolution, the vertebratebrain developed into the organ of human in-telligence. Since the ancestors are extinct,the developmental sequence can only be re-constructed by means of species that haveretained a primitive brain structure. In am-phibians and reptiles, the telencephalon(A1) appears as an appendix to the large ol-factory bulb (A2); mesencephalon (A3) anddiencephalon (A4) lie free at the surface. Al-ready in primitive mammals (such as thehedgehog), however, the telencephalon ex-pands over the rostral parts of the brainstem; in lemurs, it completely overlays thediencephalon and mesencephalon. Thus,the phylogenetic development of the brainessentially consists of a progressive enlarge-ment of the telencephalon and a transfer ofthe highest integrative functions to this partof the brain. This is called telencephaliza-tion. Ancient primitive structures are stillretained in the human brain and are inter-mingled with new, highly differentiatedstructures. Therefore, when we talk aboutnew and old components of the humanbrain, we refer to the brain’s evolution. Thebrain is neither a computer nor a thinkingmachine constructed according to rationalprinciples; it is an organ that has evolved incountless variations over millions of years.
We can follow the morphological evolu-tion of the human brain by means of castsmade of fossil cranial cavities (B, C). Thepositive cast of the cranial cavity (en-docranial cast) is a rough replica of theshape of the brain. When comparing thecasts, the enlargement of the frontal andtemporal lobes is striking. The changes fromHomopekinensis via Neanderthal, the inventorof sharp flint knifes, to Cro-Magnon (B), thecreator of cave paintings, are obvious.However, there are no appreciable differ-ences between Cro-Magnon and present-dayhumans (C).
During phylogenesis and ontogenesis, the in-dividual brain divisions develop at differenttimes. The parts serving the elementaryvital functions develop early and are alreadyformed in primitive vertebrates. The brain
divisions for higher, more differentiatedfunctions develop only late in higher mam-mals. During their expansion, they push theearly-developed brain parts into a deeperlocation and bulge outward (they becomeprominent).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

15
Introd
uction
2
2
3
4
4
1
13 312
4
1
2
Evolution of the Brain
A Evolution of the vertebrate brain
Frog Crocodile
Hedgehog
Lemur (bush baby)
B Endocranial casts of a gorilla and of fossil hominids
Gorilla Homo pekinensis
Neanderthal Cro-Magnon
C Endocranial casts of Homo sapiens, lateral view and basal view
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

16Introd
uction
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Basic Elements of theNervous System
The Nerve Cell 18The Synapse 24Neuronal Systems 32The Nerve Fiber 36Neuroglia 42Blood Vessels 44
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

18Ba
sicElem
ents
Basic Elements of the Nervous System
The Nerve Cell
The nervous tissue consists of nerve cellsand glial cells which originate from the ec-toderm (the latter are supporting and cover-ing cells). Blood vessels and meninges donot belong to the nervous tissue; they are ofmesodermal origin. The nerve cell (gan-glion cell or neuron) is the functional unitof the nervous system. In its mature state, itis no longer able to divide, thus making pro-liferation and the replacement of old cellsimpossible. Very few nerve cells are formedafter birth.
A neuron consists of the cell body, the peri-karyon (A1), the processes, dendrites (A2),and one main process, the axon or neurite(A–D3).
The perikaryon is the trophic center of thecell, and processes that become separatedfrom it degenerate. It contains the cell nu-cleus (A4) with a large, chromatin-richnucleolus (A5) to which the Barr body (sexchromatin) (A6) is attached in females.
The dendrites enlarge the cell surface bybranching. The processes of other neuronsend here: the dendrites are the sites wherenerve impulses are received. The processes ofother neurons often end at small dendriticappendices, spines (thorns), which give thedendrites a rough appearance (D).
The axon conducts the nerve impulse andbegins with the axon hillock (AD7), the sitewhere nerve impulses are generated. At a cer-tain distance from the perikaryon (initialsegment) it becomes covered by the myelinsheath (A8), which consists of a lipid-con-taining substance (myelin). The axon givesoff branches (axon collaterals) (A9) and fi-nally ramifies in the terminal area (A10) toend with small end-feet (axon terminals, orboutons) on nerve cells or muscle cells. Thebouton forms a synapse with the surfacemembrane of the next cell in line; it is herethat impulse transmission to the other celltakes place.
Depending on the number of processes, wedistinguish between unipolar, bipolar, ormultipolar neurons. Most neurons are multi-
polar. Some have short axons (interneurons),others have axons more than 1 m long (pro-jection neurons).
A neuron cannot be visualized in its entiretyby applying just one staining method. Thedifferent methods yield only partial imagesof neurons. The cellular stain (Nissl’smethod) shows nucleus and perikaryon(B –D). The latter, including the bases of thedendrites, is filled with clumps (Nissl sub-stance, tigroid bodies) and may contain pig-ments (melanin, lipofuscin) (D11). The axonhillock is free of Nissl bodies. The Nissl sub-stance is the light-microscopic equivalent ofa well-developed rough endoplasmatic reti-culum. Motor neurons possess a large peri-karyon with coarse Nissl bodies, whilesensory neurons are smaller and often con-tain only Nissl granules.
Impregnation with silver (Golgi’smethod) stains the entire cell including allneuronal processes; the cell appears as abrown-black silhouette (B–D). Other im-pregnation methods selectively stain theaxon terminals (E), or the neurofibrils (F)running in parallel bundles through peri-karyon and axon.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

19
BasicElem
ents
6
5
2
2
1
4
7
3
8
9
9
10
3
3
33
11
7 3
The Nerve Cell: Structure and Staining Patterns
A Neuron, diagram
B–D Equivalent images of nervecells: cellular stain (Nissl) andsilver impregnation (Golgi)
B Nerve cell in thebrain stem
C Nerve cell inthe anterior hornof the spinal cord
D Pyramidal cellin the cerebralcortex
E Impregnation ofboutons (synapses)
F Impregnation ofneurofibrils
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

20Ba
sicElem
ents
Basic Elements of the Nervous System: The Nerve Cell
Methods in Neuroanatomy (A–E)
The availability of methods for studying thestructure and function of cells, tissues, andorgans is often the limiting factor in ex-panding our knowledge. Certain terms andinterpretations can only be understood ifthe background of the method used isknown. Therefore, the methods commonlyused in neuroanatomy are presented herebriefly.
Nerve cells and glial cells can be demon-strated in thin histological sections byvarious histological techniques. The Nisslmethod has proven helpful because of excel-lent visualization of the rough endoplasmicreticulum (p. 18), which is abundant innerve cells. However, the different types ofnerve cells are essentially characterized bytheir long processes, the dendrites and theaxon, which are not stained by the Nisslmethod. For demonstration of as many ofthese processes as possible, thick sections(200µm) are required. By using silver im-pregnation (Golgi method, p. 18), individualnerve cells with a large number of processescan be demonstrated in such thick sections.Recently, however, this 100-year-old, effec-tive method has taken a back seat, becauseit is now possible to stain individual nervecells by filling them with a dye using rec-ording electrodes (A). The advantage of thistechnique is that electrical signals can berecorded from the neuron in question at thesame time. In addition to visualization bylight microscopy, the intracellularly stainedor Golgi-impregnated nerve cells can sub-sequently be examined by electron micros-copy to show the synaptic contacts of theseneurons.
An important characteristic of nerve cells istheir specific neurotransmitter ormessengersubstance by which communication withother nerve cells is achieved. By means ofimmunocytochemistry and the use of anti-bodies against the messenger substancesthemselves, or against neurotransmitter-synthesizing enzymes, it is possible to visual-ize nerve cells that produce a specific trans-mitter (B). Again, these immunocytochemi-cally stained nerve cells and their processes
can subsequently be examined by electronmicroscopy.
The longest processes of nerve cells, theaxons (which can be up to 1m long inhumans), cannot be traced to their targetarea in histological sections. In order todemonstrate the axonal projections of neu-rons to different brain regions, axonal trans-port (p. 28, D) is utilized. By means of anter-ograde and retrograde axonal transport,substances are transported from the nervecell body to the axon terminal and from theaxon terminal back to the nerve cell body.Very long fiber connections can be visual-ized (C–E) by means of tracers (e.g.,fluorescent dyes) that are injected eitherinto the target area or into the region con-taining the cell bodies of the correspondingpopulation of neurons; the tracers are thentaken up by the axon terminals or by the cellbodies of the projection neurons, respec-tively. When using retrograde transport (C),the tracer is injected into the assumed tar-get area. If the assumed connecting tractsexist, the tracer will accumulate in the cellbodies. By means of retrograde transportand the use of different fluorescent dyes (D),different projection zones of one and thesame neuron can be demonstrated. Whenusing anterograde transport (E), the tracer isinjected into the region of the cell bodies ofprojecting neurons. Labeled axon terminalswill be visible in the assumed target zone ifthe labeled neurons indeed project to thisarea.
Tissue cultures of nerve cells are being em-ployed to an increasing extent for studyingthe processes of development and re-generation of nerve cells, and also for study-ing the effects of pharmaceuticals.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

21
BasicElem
ents
Methods in Neuroanatomy
A Visualization of a neuron bymeans of an intracellularly in-jected marker
B Immunocytochemical visualization of acholinergic neuron using an antibodyagainst choline acetyltransferase
C–E Visualization of projections bymeans of retrograde and an-terograde axonal transport oftracers
C Retrograde transport
D Retrograde transport from differ-ent projection zones of a neuron
E Anterograde transport to differentprojection zones of a neuron
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

22Ba
sicElem
ents
Basic Elements of the Nervous System: The Nerve Cell
Ultrastructure of the Nerve Cell(A–C)
Electronmicrographs show the cell nucleus(A–C1) to be enclosed by a double-layeredmembrane (A2). It contains the nuclear pores(BC3) that probably open only temporarily.The karyoplasm of the nucleus containsfinely dispersed chromatin granules, whichconsist of DNA and proteins. The nucleolus(A–C4), a spongiform area of the nucleusmade up of a dense granular componentand a loose filamentous component, con-sists of RNA and proteins.
In the cytoplasm, the Nissl bodies appear asrough endoplasmic reticulum (A–C5), a lamel-lar system of membranes that enclose flat-tened, intercommunicating cisternae (BC6).Attached to the cytoplasmic side of themembranes are the protein-synthesizing ri-bosomes (BC7). To maintain the long axon(up to 1m long), it is essential that the cellhas an extremely high rate of protein syn-thesis (structural metabolism). Ribosome-free membranes form the agranular orsmooth endoplasmic reticulum (C8). The roughendoplasmic reticulum communicates withthe perinuclear space (BC9) and with themarginal cisternae (A10) below the cell sur-face. Marginal cisternae are often found atsites where boutons or glial cell processesare attached. The cytoplasm is crossed byneurofilaments and neurotubules (A–C11)that are arranged into long parallel bundlesinside the axon. The neurotubules corre-spond to the microtubules of other cells.
The transport of substances takes placealong neurofilaments and neurotubules(p. 28, D). Neurofibrils are the light-micro-scopic equivalent of densely packed neu-rotubules.
The neuron contains a large number ofmito-chondria (A–C12). These are enclosed in adouble membrane; the inner membraneshows projections (cristae) (C13) into theinner space (matrix). The mitochondria areof various shapes (short and plump in theperikaryon, long and slender in the den-drites and the axon) and move constantlyalong fixed cytoplasmic paths between the
Nissl bodies. The mitochondria are the siteof cellular respiration and, hence, of energygeneration. Numerous enzymes are local-ized in the inner membrane and in thematrix, among others the enzymes of thecitric acid cycle and respiratory-chain (oxida-tive) phosphorylation.
The Golgi complex consists of a number ofdictyosomes (A–C14), which are stacks offlattened, noncommunicating cisternae.The dictyosome has a forming side (cis face)(C15) and a maturing side (trans face) (C16).The forming side receives transport vesiclesfrom the endoplasmic reticulum. At themargins of the maturing side, Golgi vesiclesare formed by budding. The Golgi complexis mainly involved in the modification (e.g.,glycosylation, phosphorylation) of proteinsfrom the endoplasmic reticulum.
The numerous lysosomes (A–C17) containvarious enzymes (e.g., esterases, proteases)and are mainly involved in intracellulardigestion.
A18 Pigment.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

23
BasicElem
ents
Acidesterases
2
10
12
14
1714 11
5
17
6
7
11
18 5
14
9
1
34
1216
5
159
3
14
12 1
413
17 8 11
6
7
Ultrastructure of the Nerve Cell, Function of Organelles
A Electron-microscopic viewof a nerve cell (diagram)
B Detail of A
C Function ofcell organelles
Proteinsynthesis
RNA
Oxidation
ATPDNA
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

24Ba
sicElem
ents
Basic Elements of the Nervous System
The Synapse
The axon ends with numerous small knob-like swellings, the axon terminals or boutons.Together with the apposed membrane ofthe next neuron, the bouton forms the syn-apse where excitation is transmitted fromone neuron to another.
The synapse consists of the presynaptic com-ponent (bouton) (AB1) with the presynapticmembrane (BC2), the synaptic cleft (B3), andthe postsynaptic component with the postsyn-aptic membrane (BC4) of the next neuron.The bouton is free of neurofilaments andneurotubules but contains mitochondriaand small, mainly clear vesicles (BC5) whichare clustered near the presynaptic mem-brane (active zone). The synaptic cleft con-tains filamentous material and communi-cates with the extracellular space. The pre-synaptic and postsynaptic membranes ex-hibit dense zones of apposition, which re-semble those found at various cell junctions(zonulae or maculae adherentes, adherentjunctions or desmosomes). In asymmetricsynapses (see below), the density of thepostsynaptic membrane (B6) is more prom-inent than the presynaptic density.
Synapses can be classified according to theirlocalization, their structure, and their func-tion, or according to the neurotransmittersubstances they contain.
Localization (A)
The boutons may be apposed to dendrites(AC7) of the receptor neuron (axodendriticsynapses) (A8, C), to small projections of thedendritic membrane, spines (axospinous syn-apses) (A9), to the perikaryon (axosomaticsynapses) (A10), or to the initial segment ofthe axon (axoaxonal synapses) (A11). Largeneurons are occupied by thousands of bou-tons.
Structure (B)
Depending on the width of the synapticcleft and the properties of the apposingmembranes, two types of synapses, type I
and type II, can be distinguished accordingto Gray. In type I synapses, the synaptic cleftis wider and the density of the postsynapticmembrane is more pronounced (asymmetricsynapse). In type II synapses, the synapticcleft is narrower and the postsynaptic den-sity is about the same as the presynapticdensity (symmetric synapse).
Function (C)
There are excitatory and inhibitory synapses.The majority of the excitatory synapses arefound at the dendrites, often at the heads ofthe spines (A9). Most of the inhibitory syn-apses are found at the perikaryon or at theaxon hillock, where excitation is generatedand can be most effectively suppressed.While synaptic vesicles are usually round,some boutons contain oval or elongatedvesicles (C12). They are characteristic of in-hibitory synapses. Asymmetric synapses(type I) are often excitatory, whereas sym-metric synapses (type II) are mostly inhibi-tory.
C13Mitochondria.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

25
BasicElem
ents
9
9
11
7
8
10
17
5
1
62 3 3
2
4
13
5
7
2 4
2
4
12
Types of Synapses
A Electron-microscopic view of a dendrite(left) and a nerve cell (right)with synapses (according to Bak)
B Synapses,Gray type I (left) and type II (right)
C Electron microscopic view of a cross section of a dendrite with surrounding synapses(diagram according to Uchizono)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

26Ba
sicElem
ents
Basic Elements of the Nervous System: The Synapse
Types of Synapses (A, B)
There are numerous variations on thesimple form of synapses. The synaptic con-tact between parallel axons and dendrites iscalled parallel contact or bouton en passant(A1). Many dendrites have thornlike projec-tions (spines) that form a spinous synapse(A2) with the bouton. On the apical den-drites of some pyramidal cells, the terminalswelling of the axon encloses the entirespine, which may be relatively large andbranched, bearing numerous synaptic con-tacts (complex synapse) (B). Several axonsand dendrites can join to form glomerulus-like complexes in which the different synap-tic elements are closely intertwined. Theyprobably affect each other in terms of fine-tuning (modulating) the transmission ofimpulses.
Each brain division has characteristic formsof synapses. Gray type I and II synapses arepredominantly found in the cerebral cortex,glomerulus-like complexes are found in thecerebellar cortex, in the thalamus, and inthe spinal cord.
Electrical synapses
Adjacent cells can communicate throughpores (tunnel proteins), called gap junctions.Cells linked by gap junctions are electricallycoupled; this facilitates the transmission ofimpulses from one cell to another (e.g., insmooth muscles, p. 303, B8). Gap junctionsin neurons are therefore also called electri-cal synapses in contradistinction to thechemical synapses, which release neu-rotransmitters. Electrical coupling via gapjunctions occurs not only between neuronsbut also between glial cells.
Neurotransmitters (C, D)
Transmission of impulses at the chemicalsynapses is mediated by neurotransmitters.The most widely distributed transmittersubstances in the nervous system areacetylcholine (ACh), glutamate, gamma-aminobutyric acid (GABA), and glycine.Glutamate is the most common excitatorytransmitter, GABA is a transmitter of the in-
hibitory synapses in the brain, and glycine isan inhibitory transmitter in the spinal cord.
The catecholamines norepinephrine (NE)and dopamine (DA) also act as transmitters,and so does serotonin (5-HT). Many neu-ropeptides act not only as hormones in thebloodstream but also as transmitters in thesynapses (e.g., neurotensin, cholecys-tokinin, somatostatin).
The transmitters are produced in the peri-karyon and stored in the vesicles of the axonterminals. Often only the enzymes requiredfor transmitter synthesis are produced inthe perikaryon, while the transmitter sub-stances themselves are synthesized in theboutons. The small and clear vesicles arethought to carry glutamate and ACh, theelongated vesicles of the inhibitory synapsescarry GABA, while norepinephrine anddopamine are present in the granular ves-icles (C).
Most vesicles are located near the presyn-aptic membrane, the density of which canbe demonstrated by special procedures as agrid with hexagonal spaces (D3). The ves-icles pass through these spaces to reach thepresynaptic membrane and, upon excita-tion, empty their content into the synapticcleft by fusing with the presynaptic mem-brane (omega figure) (D4). The transmittersubstances are delivered in certain quanta,the morphological equivalents of which arethe vesicles. Some of the transmittermolecules return into the bouton by reup-take (D5).
D6 Axonal filaments.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

27
BasicElem
ents
2
1
5
6
4
3
Types of Synapses, Neurotransmitters
A Parallel contact (1) and spinous synapse (2)
B Complex synapse
C Different types ofsynaptic vesicles
D Model of a synapse (according to Akert, Pfenniger, Sandri and Moor)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

28Ba
sicElem
ents
Basic Elements of the Nervous System: The Synapse
Neurotransmitters (continued)(A–C)
Many neurons, perhaps most of them, pro-duce more than one transmitter substance.Nevertheless, they are classified accordingto their functionally most important neu-rotransmitter as glutamatergic, cholinergic,catecholaminergic (noradrenergic anddopaminergic), serotoninergic, and pepti-dergic neurons. The catecholaminergic andserotoninergic neurons can be identifiedby fluorescence microscopy because theirtransmitters show a green-yellow fluores-cence following exposure to formalin vapor(A, B). It is thus possible to trace the axonand to recognize the perikaryon and theoutline of its nonfluorescent nucleus. Thefluorescence is very faint in the axon, moredistinct in the perikaryon, and most intensein the axon terminals. It is here that thehighest concentration of transmitters oc-curs. Cholinergic neurons can be demon-strated by a histochemical assay for acetyl-cholinesterase, the enzyme required for thedegradation of acetylcholine. Since thisenzyme is also produced by noncholinergicneurons, the proper assay is by immunocy-tochemistry using antibodies against cholineacetyltransferase, the acetylcholine-synthe-sizing enzyme. Other transmitters and neu-ropeptides can also be demonstrated by im-munocytochemistry (C). It has been shownby double-labeling that many neuropep-tides are produced together with classicalneurotransmitters within the same neuron.So far, the functional significance of cotrans-mission, i.e., the release of different trans-mitters by the same neuron, has been stud-ied in detail only on some neurons of the au-tonomic nervous system.
Axonal Transport (D, E)
The transmitter substances or their synthe-sizing enzymes are produced in the peri-karyon and must be transported to the axonterminal. The microtubules of the neuron,neurotubules (D1), play a key role in thistransport mechanism. If they are destroyedby applying the mitotic poison colchicine,the intra-axonal transport stops. This rapid
transport of material is energy-dependentand takes place in vesicles that are movedalong the microtubules by motor proteins.The retrograde transport (in the direction ofthe cell body and toward the minus end ofthe microtubules) is mediated by dynein(D2), while the anterograde transport (inthe direction of the axon terminal andtoward the plus end of the microtubules) ismediated by kinesin (D3). The transportingvesicles are endowed with several motorproteins, the ATP-binding heads of whichinteract with the surface of the microtubulein an alternating and reversible fashion. Thisresults in ATP being hydrolyzed, and the re-leased energy is converted into molecularmovement that causes the vesicles to rollalong the microtubules in the target direc-tion. The velocity of the rapid intra-axonaltransport has been calculated at 200–400mm per day. Proteins, viruses, and tox-ins reach the perikaryon by retrogradetransport from the axon terminals.
In addition to the rapid intra-axonal trans-port, there is also a continuous flow of axo-plasm which is much slower, namely,1–5mm per day. It can be demonstrated byligating a single axon (E); proximal to theconstricted site, the axoplasm is held backand the axon shows swelling.
The anterograde and retrograde transportmechanisms are used in neuroanatomy tostudy connecting tracts (see p. 20).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

29
BasicElem
ents
2
3
1
Neurotransmitters, Axonal Transport
A, B Catecholaminergic neurons in the brain stem,fluorescence microscopic views (according to Dahlström and Fuxe)
C Peptidergic neuron,immunoperoxidase reaction(according to Star, Stumpf,et al.)
D Molecular motors (dynein and kinesin) of vesicletransport along neurotubules
(according to Wehnert and Gehring)
E Blocking the flow of axoplasm by axon ligation (according to Weiss and Hiscoe)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

30Ba
sicElem
ents
Basic Elements of the Nervous System: The Synapse
Transmitter Receptors (A, B)
There are two categories of neurotransmit-ter receptors: ligand-gated ion channels andtransmitter receptors coupled to an intra-cellular guanosine triphosphate-(GTP-)binding protein (G protein).
Ligand-gated Ion Channels
Ligand-gated ion channels consist of differ-ent subunits (A1) that are inserted into thecell membrane (A2). Binding of the neu-rotransmitter to the specific receptor causesthe channel to become permeable to certainions (B).
Excitatory amino acid receptors. Recep-tors for the excitatory transmitter glutamateare classified according to the synthetic li-gands binding to them. There are threetypes of glutamate-gated ion channels: theAMPA (aminohydroxymethylisoxazole-propionic acid) receptor (C3), the NMDA (N-methyl-D-aspartate) receptor (C4), and thekainate receptor. Binding to the AMPA re-ceptor causes an influx of sodium ions, thusleading to depolarization of the cell. Simi-larly, activation of the NMDA receptorcauses an influx of both Na+ and Ca2+. Underconditions of resting potential, the NMDAreceptor is blocked by magnesium; themagnesium blockade is lifted by depolariza-tion (through AMPA receptors). This tem-poral shift in activities of the AMPA andNMDA receptors results in a graduated re-sponse of the postsynaptic neurons to theneurotransmitter glutamate.
Inhibitory GABA and glycine receptors.GABA is the most common inhibitory trans-mitter in the brain, and glycine in the spinalcord. Both receptors are ligand-gated ionchannels that cause the influx of chlorideions when activated. The cell thus becomeshyperpolarized and inhibited.
Ligand-gated ion channels include the exci-tatory, cation-permeable nicotinic acetyl-choline receptor and the serotonin (5-HT) re-ceptor.
Receptors Coupled to G Protein
Most neurotransmitters do not bind to lig-and-gated channels but to receptorscoupled to G protein. The main differencebetween the two types of receptors is in thespeed of the synaptic response. In the case ofligand-gated ion channels, the activationcauses a rapid synaptic potential lastingonly for milliseconds. Activation of G pro-tein-coupled receptors results in responsesthat last seconds or minutes. G proteinsregulate enzymes that produce intracellularmessenger substances. These have an effecton ion channels or, via regulatory proteins,on the expression of genes.
Synaptic Transmission (C)
The synaptic transmission is essentiallycharacterized by three processes:
1 Conversion of the action potential arriv-ing at the axon terminal into a chemicalsignal. Depolarization results in theopening of calcium channels (C5) and inthe influx of calcium, which, mediated bycertain proteins, causes fusion of synapticvesicles (C6) with the presynaptic mem-brane and release of the transmitter intothe cleft (C7).
2 The released transmitter binds to specificreceptors.
3 In the case of ligand-gated ion channels,this results in their opening for certainions. In the case of glutamatergic recep-tors, the influx of Na+ or Ca2+ causesdepolarization of the postsynaptic mem-brane (excitatory postsynaptic potential,EPSP). In the case of GABA and glycine re-ceptors, the influx of Cl– causes hyper-polarization of the postsynaptic mem-brane (inhibitory postsynaptic potential,IPSP). The activation of G protein-coupledreceptors results in a long-lasting re-sponse that may finally lead to a changein gene expression in the postsynapticneuron.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

31
BasicElem
ents
2
1
5
3
4
6
7
2
1
5
3
4
6
7
Transmitter Receptors, Synaptic Transmission
A, B Ligand-gated ion channel (adapted from Kandel, Schwartz and Jessel)
A Structure of the nicotinic acetyl-choline receptor
B Binding of acetylcholine (ACh)opens the channel for Na+ and K+
C Synaptic transmission at a glutamatergic synapse
Mg2 +
Ca2 +
Glutamate
!"
!
#$
ACh Na+
K+
K+
Na+
K+
Na+
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

32Ba
sicElem
ents
Basic Elements of the Nervous System
Neuronal Systems
Groups of neurons that have the same trans-mitter and the axons of which often formdense fiber bundles are described accordingto their transmitter substance as cholinergic,noradrenergic, dopaminergic, serotoninergic,GABAergic, or peptidergic systems. The im-pulse can be transmitted to neurons of thesame type or to neurons with a differentneurotransmitter. In the sympatheticnervous system, for example, the neurons inthe spinal cord are cholinergic; however,transmission in the peripheral gangliaswitches to noradrenergic neurons (p. 297,C).
Noradrenergic, dopaminergic, and sero-toninergic neurons are located in the brainstem. Noradrenergic neurons form thelocus caeruleus (A1) (p. 100, B28; p. 132,D18) and cell groups in the lateral part of thereticular formation of themedulla oblongataand the pons; their fibers project to the hy-pothalamus, to the limbic system, diffuselyinto the neocortex, and to the anterior hornand lateral horn of the spinal cord. Sero-toninergic neurons lie in the raphe nuclei(A2) (p. 108, B28), especially in the posteriorraphe nucleus (A3); their fibers project tothe hypothalamus, to the olfactoryepithelium, and to the limbic system.Dopaminergic neurons make up the com-pact part of the substantia nigra (A4) (p. 134,A17; p. 136, AB1) from where the nigrostri-atal fibers extend to the striatum.
Peptidergic neurons are found in phylo-genetically older brain regions, namely, inthe central gray of the midbrain (A5), in thereticular formation (A6), in the hypo-thalamus (A7), in the olfactory bulb, in thehabenular nucleus (A8), interpeduncular nu-cleus (A9), and solitary nucleus (A10).Numerous peptidergic neurons have alsobeen demonstrated in the cerebral cortex, inthe thalamus, in the striatum, and in thecerebellum. The significance of the differentpeptides is still largely unclear. It is assumedthat they act as cotransmitters and have amodulating function. Many of these pep-tides are found in other organs as well, such
as the digestive system (e.g., vasoactiveintestinal polypeptide, somatostatin,cholecystokinin).
Glutamate is often the transmitter of pro-jection neurons with long axons. Gluta-matergic neurons are, for example, the pro-jection neurons of the cerebral cortex, the pyra-midal cells (p. 240, C; p. 242, A1 and B11).GABAergic inhibitory neurons are oftenclassified according to the target structureson which they form inhibitory synapses.GABAergic basket cells, which form syn-apses with cell bodies, are distinguishedfrom axo-axonal cells. The latter develop in-hibitory synapses at the beginning of theaxon (initial segment) of a projection neu-ron. GABAergic neurons often form local cir-cuits (interneurons). They often contain pep-tides (see above) and calcium-binding pro-teins apart from GABA as the classic trans-mitter.
Cholinergic neurons are found in the brainstem and also in the basal forebrain. As in thecase of catecholaminergic neurons, far-reaching projections originate from circum-scribed groups of cells, for example, in thebasal nucleus (B11) and in certain septal nu-clei (B12) that supply, via fibers in the cingu-late gyrus (limbic gyrus) (B13) and in thefornix (B14), respectively, large regions ofthe neocortex and the hippocampus (B15).These ascending cholinergic projectionsfrom the basal forebrain are thought to beassociated with processes of learning andmemory. They are affected in Alzheimer’s dis-ease which is accompanied by disturbedlearning and memory.
Synthesis, degradation, and storage oftransmitter substances can be influenced bypharmaceuticals. An excess or deficiency intransmitters can be created in the nervecells, leading to changes in motor or mentalactivity. Changing transmitter synthesis anddegradation is not the only way neurophar-maceuticals can influence the synaptictransmission; they can also act on the re-ceptors as transmitter agonists or antago-nists.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

33
BasicElem
ents
8
547
9
1
6
3
2
10
13 14
12 11 15
Neuronal Systems
A Monoaminergic and peptidergicgroups of neurons in the brain
Dopaminergic neuronsNoradrenergic neuronsSerotoninergic neuronsPeptidergic neurons
B Cholinergic groups of neurons in the basalforebrain, including the cholinergic innervationof cerebral cortex and hippocampus
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

34Ba
sicElem
ents
Basic Elements of the Nervous System: Neuronal Systems
Neuronal Circuits (A–D)
The nerve cells and their processes form anetwork (A) that is not a continuumof nervefibers (continuity theory) but consists ofcountless individual elements, the neurons(neuron theory). As the basic building blockof the nervous system, the neuron repre-sents a structural, genetic, trophic, andfunctional unit.
The neurons in the network are intercon-nected in a specific way (neuronal circuits).The connections for the inhibition of excita-tion are as important as those for the trans-mission of excitation, for they are the onesthrough which the continuous influx of im-pulses is restricted and selected: importantsignals are transmitted, unimportant onesare suppressed.
Postsynaptic inhibition does not inhibitsynaptic transmission but subsequent dis-charge of the postsynaptic neuron.
Inhibitory GABAergic neurons can be inte-grated into the neuronal circuit in differentways. In the case of recurrent (feedback) inhi-bition (B), an axon collateral of the exci-tatory projection neuron (green) activatesthe inhibitory cell (red), which in turn in-hibits the projection neuron via a recurrentcollateral. In the case of feed-forward inhibi-tion (C), the inhibitory interneuron is not ac-tivated by a recurrent collateral of the exci-tatory cell but by excitatory afferents fromanother brain region. The effect on the pro-jection neuron, however, is the same; acti-vation of the inhibitory GABAergic neuronsleads to inhibition of the projection neu-rons. In case of disinhibition (D), an inhibi-tory interneuron is again activated by an ex-citatory afferent. The target cell of this inter-neuron, however, is another inhibitory in-terneuron. Activation of the first inter-neuron through the excitatory afferenttherefore means an increased inhibition ofthe second interneuron, which now cannotexert an inhibitory effect on the next projec-tion neuron. The inhibitory effect is re-moved (disinhibition).
A single nerve cell in the brain receives alarge number of connections (convergence).
One pyramidal cell of the cerebral cortexcan establish over 10000 synaptic connec-tions with other nerve cells. In turn, such acell itself creates numerous connectionswith many other nerve cells by numerousaxon collaterals (divergence). The spatial andtemporal summation of excitatory and in-hibitory inputs of a cell decides at a givenmomentwhether the cell is depolarized andgenerates an action potential, which thenruns along the axon and leads to excitationof subsequent neurons in the series. If theinhibitory inputs predominate, the dis-charge of neurons does not take place.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
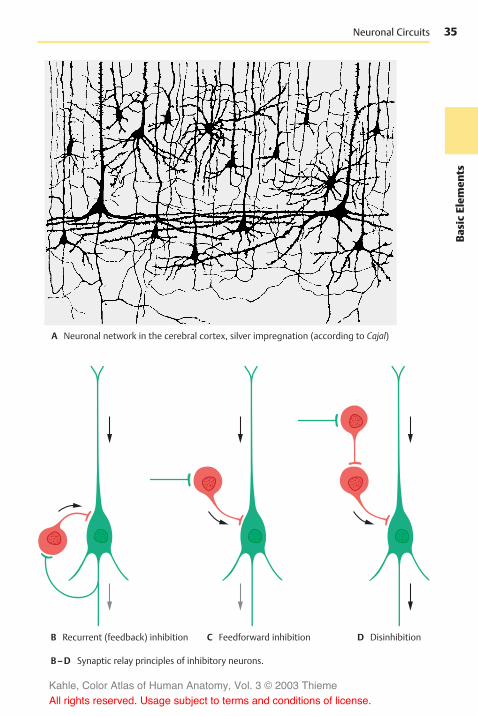
35
BasicElem
ents
Neuronal Circuits
A Neuronal network in the cerebral cortex, silver impregnation (according to Cajal)
B Recurrent (feedback) inhibition C Feedforward inhibition D Disinhibition
B–D Synaptic relay principles of inhibitory neurons.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

36Ba
sicElem
ents
Basic Elements of the Nervous System
The Nerve Fiber (A–G)
The axon (AFG1) is surrounded by a sheath:in unmyelinated nerve fibers by the cyto-plasm of the sheath cells, and in myelinatednerve fibers by the myelin sheath (ABG2).Axon and sheath together are called thenerve fiber. The myelin sheath begins be-hind the initial segment of the axon andends just before the terminal ramification. Itconsists ofmyelin, a lipoprotein produced bythe sheath cells. The sheath cells in the CNSare oligodendrocytes; in the peripheralnerves they are Schwann cells, which origi-nate from the neural crest (p. 62, C2). Themyelin sheath of fresh, unfixed nerve fibersappears highly refractile and without struc-ture. Its lipid content makes it birefringentin polarized light. The lipids are removedupon fixation, and the denatured proteinscaffold remains as a gridlike structure(neurokeratin) (D3).
At regular intervals (1–3mm), the myelinsheath is interrupted by deep constrictions,the nodes of Ranvier (AB4 F). The segment be-tween two nodes of Ranvier in peripheralnerves, the internode or interannular segment(F), corresponds to the expansion of onesheath cell. The cell nucleus (ADF5) andperinuclear cytoplasm form a slight bulgeon themyelin sheath in themiddle of the in-ternode. Cytoplasm is also contained in ob-lique indentations, the Schmidt–Lantermanincisures (C, F6) (see also p. 40, A4). Themar-gins of the sheath cells define the node ofRanvier at which axon collaterals (E) maybranch off or synapses may occur.
Ultrastructure of the Myelin Sheath(G)
The electron micrograph shows the axonenclosed by a plasma membrane, the ax-olemma; it is surrounded by a series of regu-larly spaced, concentric dark and light lines(period lines). The width of each lamellafrom one dark line to the next measures120Å on average (1Å=0.1 nm), with thedark line taking up 30Å and the light line90Å. As seen at higher magnification, thelight lines are subdivided by a thin irregular
line resembling a string of pearls (G7). Wethus distinguish a dense major period lineand a fainter intraperiod line. Studies usingpolarized light and X-rays have shown thatthe myelin sheath is made up of alternatinglayers of protein and lipid molecules. Ac-cordingly, the dark lines (major period lineand intraperiod line) are regarded as layersof protein molecules and the light lines aslayers of lipid molecules.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

37
BasicElem
ents
52
4
5
5
1
6
7
4
2
1
4
3
2
1
4
5
Myelin Sheath
A Nerve fiber (according to von Möllendorff)
B Node of Ranvier, osmium stain (diagram) C Schmidt–Lanterman incisures
D Perikaryon of a Schwann cell E Axonal branching
F Internode (according to Cajal)
G Electron-microscopicviews of themyelin sheath
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

38Ba
sicElem
ents
Basic Elements of the Nervous System: The Nerve Fiber
Development of the Myelin Sheathin the PNS (A)
The development of the myelin sheath af-fords an insight into the structure of its spi-raling lamellae. The cell body of the Schwanncell (A1) forms a groove into which the axon(A2) becomes embedded. The groove deep-ens and its margins approach each otherand finally meet. In this way, a duplicationof the cell membrane is formed, themesaxon(A3), which wraps around the axon like aspiral as the Schwann cell migrates aroundthe encircled axon.
The term mesaxon is based on the termmesenterium, a thin duplication that isformed as a suspension band by the peri-toneum and encloses the intestine. In a sim-ilar way, the Schwann cell forms a duplica-tion and envelops the axon. Like all plasmamembranes, the cell membrane of theSchwann cell consists of an outer and aninner dense layer of protein and a light lipidlayer between them. Upon membrane du-plication, the two outer protein layers comeinto apposition first and fuse to form the in-traperiod line (A4). Thus, the six-layeredmembrane duplication becomes the five-layered myelin lamella. With further encir-cling, the inner protein layers of the cellmembrane make contact as well and fuse toform the dense major period line (A5). At theend of the process, the start of the duplica-tion lies inside the myelin sheath, the inter-nal mesaxon (AB6), while the end lies out-side, the external mesaxon (7 in A, B).
Development of UnmyelinatedNerve Fibers (A)
Unmyelinated nerve fibers (A8) are en-veloped by sheath cells, each of which en-circles several axons. The margins of thegrooves may also form a membrane dupli-cation (mesaxon) but without fusion of themembrane layers.
Structure of the Myelin Sheath inthe CNS (B, C)
The myelin sheath in the CNS (B) shows dis-tinct differences as compared to the myelinsheath of the peripheral nerves (see p. 41, B).Whereas the Schwann cell in the PNSmyeli-nates only one axon, an oligodendrocyte (B9)in the CNSmyelinates several axons andwilllater remain connected with several inter-nodes via cytoplasmic bridges. The extentand shape of the cell becomes clear whenvisualizing the internodes as being unfurled(C). The mechanism of the myelinationprocess is unknown. The external mesaxonforms an external bulge (B10) starting fromthe cytoplasmic bridge. Themyelin lamellaeterminate at the paranodal region (B11)(node of Ranvier). As seen in the longitudi-nal section, the innermost lamella termi-nates first and the outermost lamella coversthe remaining endings, terminating directlyat the node of Ranvier. At the ends of thelamellae, the dense major period lineswiden into pockets filled with cytoplasm(B12). The axon of the central nerve fiber iscompletely exposed in the area of the nodeof Ranvier. There are no Schmidt–Lanter-man incisures in the CNS.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

39
BasicElem
ents
7
6
5
4
3
82
9
1
10
7
6
11
12
Myelin Sheath
A Development of the myelinsheath (according to Hamilton,Boyd and Mossman)
B Central nerve fiber,electron-microscopicdiagram (accordingto Bunge)
C Oligodendrocyte with myelinlamellae (according to Bunge)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

40Ba
sicElem
ents
Basic Elements of the Nervous System: The Nerve Fiber
Peripheral Nerve (A–D)
The myelin sheath of peripheral nervefibers is surrounded by the cytoplasm of theSchwann cell (A1). The outer cell membraneborders on a basal lamina (AB2), which en-velops the entire peripheral nerve fiber. Thenucleus of the Schwann cell (A3) is depictedin cross section. The Schmidt–Lantermanclefts (A4) are depicted in longitudinal sec-tion as cytoplasmic crevices of themajor pe-riod lines. In the three-dimensional recon-struction, they appear as spirals in whichthe cytoplasm communicates between theinside and outside. At the node of Ranvier(B5), the Schwann cell processes (AB6) slideover the paranodal region and over the axon(ABD7). They interdigitate and thus form adense envelope around the node of Ranvier.
Differences between the structures of themyelin sheaths in CNS and PNS are il-lustrated in B.
There is a regular relationship between thecircumference of the axon, the thickness ofits myelin sheath, the distance between thenodes of Ranvier, and the conductionvelocity of a nerve fiber. The larger the cir-cumference of an axon, the thicker the en-closing myelin sheath and the longer the in-ternodes. When myelinated nerve fibers arestill growing (e.g., in the nerves of thelimbs), the internodes are growing inlength. The longer the internodes, the fasterthe conduction velocity of the fiber.We dis-tinguish between myelinated, poorly myeli-nated, and unmyelinated nerve fibers, alsoreferred to as A, B, and C fibers. The myeli-nated A fibers have an axonal diameter of3–20µm and a conduction velocity of up to120m/s; the poorly myelinated B fibers are upto 3µm in diameter and have a conductionvelocity of up to 15m/s. Conduction velocityis the slowest in the unmyelinated C fibers (upto 2m/s); we are dealing here with a con-tinuous spread of excitation. By contrast,conduction in myelinated nerves is salta-tory, that is, it takes place in jumps. Themorphological basis of saltatory conductionis the alternation of myelinated internodesand unmyelinated nodes of Ranvier; thecurrent inside the axon jumps from one
node to the next, and the current circuit isclosed each time at the nodes throughchanges in the permeability of the ax-olemma (triggered by voltage-gated ionchannels). This mode of conduction is muchfaster and requires less energy than the con-tinuous spread of excitation.
The peripheral nerve fiber is surrounded bylongitudinal collagenous connective-tissuefibrils; together with the basal membrane,they form the endoneural sheath. Thenerve fibers are embedded in a loose con-nective tissue, the endoneurium (D8). A vari-able number of nerve fibers is collected intobundles or fascicles (C10) by the perineurium(CD9) which consists mainly of circularfibers. The innermost layer of the per-ineurium is formed by endothelial cells thatenclose the endoneural space in several thinlayers. The perineural endothelial cellspossess a basal membrane at their per-ineural and endoneural surfaces and arejoined together by zonulae occludentes(tight junctions). They represent a barrierbetween nerve and surrounding tissue, sim-ilarly to the endothelial cells of cerebralcapillaries (p. 45 E). The mechanicalstrength of the peripheral nerve is based onits content of circular elastic fibers. In thenerves of the limbs, the perineurium is rein-forced in the joint regions. The epineurium(CD11) borders on the perineurium; itsinner layers form concentric lamellae aswell. They change into loose connectivetissue containing fat cells (D12), blood ves-sels, and lymph vessels.
D13 Cell nuclei of Schwann cells.D14 Capillaries.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

41
BasicElem
ents5
7
6
3
10
21
4
67
9
13 7 8
11
9
14
11
12 12
2
Myelin Sheath and Endoneural Sheath of Peripheral Nerves
A Peripheral nerve fiber,electron-microscopicdiagram (accordingto Schröder)
B Node of Ranvier of a peripheralnerve fiber (top half) and of a cen-tral nerve fiber (bottom half)(according to Bunge)
PNS
CNS
C Peripheral nerve, cross section
D Detail of C
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

42Ba
sicElem
ents
Basic Elements of the Nervous System
Neuroglia
The neuroglia (glia, glue) is the supportingand covering tissue of the CNS and has all thefunctions of connective tissue: support,metabolite exchange, and, in pathologicalprocesses, digestion of degenerating cells(phagocytosis) and scar formation. It is ofectodermal origin. After Nissl staining, onlycell nuclei and cytoplasm are visible; visual-ization of cellular processes is achieved onlyby special impregnation methods and byimmunocytochemistry. We distinguishthree different types of neuroglia: astroglia(macroglia), oligodendroglia, and microglia(A).
Astrocytes have a large, clear cell nucleusand numerous processes which give the cella starlike appearance (A, C). There are proto-plasmic astrocytes with few processes (usu-ally present in graymatter) and fibrous astro-cytes with numerous long processes (pre-dominantly present in white matter). Thelatter produce fibers and contain glial fila-ments in cell body and processes. They formglial scars after damage to the brain tissue.Astrocytes are regarded as supporting el-ements, since they form a three-dimen-sional scaffold. On the outer surface of thebrain, the scaffold thickens to form a densefiber felt, the glial limiting membrane, whichforms the outer limit of the ectodermaltissue against the mesenchymal meninges.Astrocytes extend processes to blood ves-sels and play a role in metabolite exchange(p. 45 A, B).
In addition, astrocytes play a decisive part inmaintaining the interior environment of theCNS, particularly the ion balance. Potassiumions released upon excitation of groups ofneurons are removed from the extracellularspace via the network of astrocyteprocesses. Astrocytes probably also take upCO2 released by nerve cells and thus keepthe interstitial pH at a constant value of 7.3.Astrocyte processes enclose the synapsesand seal off the synaptic cleft. They also takeup neurotransmitters (uptake and release ofGABA via astrocytes have been demon-strated).
Oligodendrocytes have a smaller, darkercell nucleus and only a few, sparselybranched processes. In the graymatter, theyaccompany neurons (satellite cells) (B). In thewhite matter, they lie in rows between thenerve fibers (intrafascicular glia). They pro-duce and maintain the myelin sheath (p. 39,B and C). In the peripheral nervous systemthemyelin sheath is formed by Schwann cells.
Microglial cells have an oval or rodlike cellnucleus and short, branched processes.They exhibit ameboid mobility and can mi-grate within the brain tissue. In response totissue destruction, they phagocytose mate-rial (scavenger cells) and round up intospheres (gitter cell).
The frequently expressed view that micro-glia were not derived from the ectoderm butfrom the mesoderm (mesoglia) is not sup-ported by evidence.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

43
BasicElem
ents
Neuroglia
A Equivalent images of neuroglia: Nissl staining (top row), silver impregnation (bottom row)
Fibrous astrocyte Protoplasmic astrocyte Oligodendroglia Microglia
B Oligodendrocytes assatellites of a nerve cell
C Astrocyte in tissue culture
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

44Ba
sicElem
ents
Basic Elements of the Nervous System
Blood Vessels
Cerebral blood vessels are of mesodermalorigin. They grow during development fromthe mesodermal coverings into the braintissue. In histological preparations, they aremostly surrounded by a narrow empty cleft(Virchow–Robin space, perivascular space),an artifact caused by tissue shrinkageduring histological preparation. Arteriesand arterioles are of the elastic type, that is,their muscles are poorly developed andtheir contractility is limited. The capillariesexhibit a nonfenestrated closed endotheliumand a closed basal membrane. There are nolymph vessels in the CNS.
Astrocyte processes extend to the capillariesand widen into perivascular glial feet(AB1). In electron micrographs, capillariesare completely covered by perivascular feet.The capillary wall consists of endothelial cells(BE2) which overlap at their margins likeroof tiles and are joint together by zonulaeoccludentes (tight junctions). The capillaryis enclosed by the basal lamina (BE3) and theastrocyte covering (BE4). The latter can becompared to the glial limiting membrane(p. 42); both structures separate the ec-todermal tissue of the CNS from the adja-cent mesodermal tissue.
The sealing of the brain tissue from the restof the body manifests itself in the blood–brain barrier, a selective barrier for numer-ous substances that are prevented frompenetrating from the bloodstream throughthe capillary wall into the brain tissue.
This barrierwas first demonstrated by Gold-mann’s experiments using trypan blue. If thedye is injected intravenously into experi-mental animals (Goldmann’s first experiment)(C), almost all organs stain blue, but thebrain and spinal cord remain unstained.Minor blue staining is only found in the graytubercle (C5), the postremal area, and thespinal ganglia. The choroid plexus (C6) andthe dura (C7) show a distinct blue staining.The same pattern is observed in cases ofjaundice in humans; the bile pigment stainsall organs yellow, only the CNS remains un-stained. If the dye is injected into the space
of the cerebrospinal fluid (Goldmann’s secondexperiment) (D), brain and spinal cord arediffusely stained on the surface, while therest of the body remains unstained. Thus,there exists a barrier between the CSF andthe blood but not between the CSF and theCNS. We therefore distinguish between ablood–brain barrier and a blood–cere-brospinal fluid barrier. The two barriersbehave in different ways.
The site of the blood–brain barrier is thecapillary endothelium (E) (see also vol. 2);in the brain, it forms a closed wall withoutfenestration. By contrast, the capillary wallsof many other organs (liver, kidney [E8]) ex-hibit prominent fenestration which allowsfor extensive metabolite exchange. The bar-rier effect has been demonstrated fornumerous substances in studies using iso-topes. The barrier may result in a completeblockade or in a delay of penetration.Whether or not drugs can penetrate thisbarrier has major practical implications.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

45
BasicElem
ents
5
6
1
5 7
7
24
3
1
2 83
4
Perivascular Glial Feet, Blood–Brain Barrier, Blood–CSF Barrier
A Blood vessel surrounded by astrocytes,silver impregnation
B Blood vessel with perivascular glial feet(diagram according to Wolff)
C Goldmann’s first experiment
D Goldmann’s second experiment
C–D Blood-brain barrier in the rab-bit (according to Spatz)
E Brain capillary (left) and kidney capillary(right), diagram based on electron-microscopic findings
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

46Ba
sicElem
ents
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Spinal Cord and SpinalNerves
Overview 48The Spinal Cord 50Peripheral Nerves 70
Cervical Plexus 72Posterior Branches 72Brachial Plexus 74Nerves of the Trunk 84Lumbosacral Plexus 86Sacral Plexus 90
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

48Sp
inal
Cord
Spinal Cord and Spinal Nerves
Overview
The spinal cord, or spinal medulla, is lo-cated in the channel of the spinal column,the vertebral canal, and is surrounded by thecerebrospinal fluid. It has two spindle-shaped swellings: one in the neck region,the cervical enlargement (C1), and one in thelumbar region, the lumbar enlargement (C2).At the lower end, the spinal cord tapers intothe medullary cone (BC3) and ends as a thinthread, the terminal filament (C4). The ante-rior median fissure at the ventral side and theposterior median sulcus (BC5) at the dorsalside mark the boundaries between the twosymmetrical halves of the spinal cord. Nervefibers enter dorsolaterally and emerge ven-trolaterally at both sides of the spinal cordand unite to form the dorsal roots, posteriorroots, and the ventral roots, anterior roots. Theroots join to form short nerve trunks of 1 cmin length, the spinal nerves. Intercalatedinto the posterior roots are spinal ganglia(B6) containing sensory nerve cells. Only theposterior roots of the first cervical spinalnerves do not have a spinal ganglion, or onlya rudimentary one.
In humans, there are 31 pairs of spinal nerveswhich emerge through the intervertebralforamina from the vertebral canal. Each spi-nal nerve pair supplies one body segment.The spinal cord itself is unsegmented. Theimpression of segmentation is created bythe bundling of nerve fibers emerging fromthe foramina (p. 66).
The spinal nerves are subdivided into cervi-cal nerves, thoracic nerves, lumbar nerves, sacralnerves, and coccygeal nerves (A). There are
! 8 pairs of cervical nerves (C1–C8) (thefirst pair emerges between occipital boneand atlas)
! 12 pairs of thoracic nerves (T1–T12) (thefirst pair emerges between the first andsecond thoracic vertebrae
! 5 pairs of lumbar nerves (L1–L5) (the firstpair emerges between the first and sec-ond lumbar vertebrae)
! 5 pairs of sacral nerves (S1–S5) (the firstpair emerges through the upper sacralforamina)
! one pair of coccygeal nerves (emerging be-tween the first and second coccygealvertebrae)
Spinal cord and vertebral canal are initiallyof the same length so that each spinal nerveemerges from the foramen lying at its ownlevel. During development, however, thevertebral column increases much more inlength than does the spinal cord. As a result,the lower end of the spinal cord movesfurther up in relation to the surroundingvertebrae. In the newborn, the lower end ofthe spinal cord lies at the level of the thirdlumbar vertebra, and in the adult, at thelevel of the first lumbar or twelfth thoracicvertebra. Thus, the spinal nerves no longeremerge at their levels of origin; instead,their roots run down a certain distancewithin the vertebral canal to their foramenwhere they emerge. The more caudally theroots originate from the spinal cord, thelonger their run within the vertebral canal.The levels where the spinal nerves emergeare therefore no longer identical with thecorresponding levels of the spinal cord.
From the medullary cone (BC3) onward, thevertebral canal contains only a dense massof descending spinal roots, known as thecauda equina (tail of a horse) (B7).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

49
Spinal
Cord
1
2
3
4
5
6
78
1
2
3
4
5
6
7
8
9
10
11
12
1
2
3
4
512345
1
5
2
4
7
3
5
6
Spinal Cord and Spinal Nerves
A Lateral view of the spinal nerves
Cerv
ical
nerv
esTh
orac
icne
rves
Lum
barn
erve
s
Sacralnerves
Coccygeal nerves
B Dorsal view of the spinal ganglia
Coccyx
C Spinal cord
C 1
C 7C 8T 1
T 12
L 1
L 5
S 1
C 2
T 1
L 1
S 1
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

50Sp
inal
Cord
Spinal Cord and Spinal Nerves
The Spinal Cord
Structure (A, B)
The gray matter, substantia grisea (nervecells), appears in transverse section of thespinal cord as a butterfly configuration sur-rounded by the white matter, substantia alba(fiber tracts).We distinguish on either side adorsal horn (posterior horn) (AB1) and a ven-tral horn (anterior horn) (AB2). Both formcolumns in the longitudinal dimension ofthe spinal cord, the anterior column and theposterior column. Between them lies the cen-tral intermediate substance (A3) with theobliterated central canal (A4). In thethoracic spinal cord, the lateral horn (AB5) isinterposed between the anterior and poste-rior horns. The lateral posterior sulcus (A6) isthe site where the posterior root fibers(AB7) enter. The anterior root fibers (AB8)leave the anterior side of the spinal cord asfine bundles.
The posterior horn is derived from the alarplate (origin of sensory neurons) and con-tains neurons of the afferent system (B). Theanterior horn is derived from the basal plate(origin of motor neurons) and contains theanterior horn cells, the efferent fibers ofwhich run to the muscles. The lateral horncontains autonomic nerve cells of the sym-pathetic nervous system (p. 292).
The white matter is subdivided into thedorsal column, or posterior funiculus (A9),which reaches from the posterior septum(A10) to the posterior horn, the lateralcolumn, or lateral funiculus (A11), whichreaches from the posterior horn to the ante-rior root, and the ventral column, or anteriorfuniculus (A12), which reaches from theanterior root to the anterior fissure (A13).The latter two form the anterolateral column.The white commissure (A14) connects thetwo halves of the spinal cord.
Reflex Arcs (C–G)
The afferent fibers of the posterior root,which originate from the nerve cells of thespinal ganglion, transmit sensory signals tothe posterior horn cells of the spinal cord,
and these pass them on to the brain (C). Therelay may also take place in the medulla ob-longata. However, the afferent fibers mayalso run to the anterior horn cells and trans-mit the signal directly to these cells. The re-sulting muscle reaction is called reflex, theunderlying neuronal circuit is called reflexarc (D). In general, the afferent fibers do notrun directly to the motor neuron (mono-synaptic reflex arc) but via interneurons thatare interposed (multisynaptic reflex arc) (E).
The monosynaptic intrinsic reflex (stretchreflex) and themultisynaptic extrinsic reflex(withdrawal reflex) are of clinical impor-tance. In the stretch reflex (F), the muscle isbriefly stretched by a tap on its tendon.Stimulation of the muscle receptors (p. 314)results in a momentary contraction of themuscle as a counter reaction. The reflex in-volves only a few neurons at any level of thespinal cord. In the withdrawal reflex (G), skinreceptors are stimulated (pain); the with-drawal movement is brought about by thecoordinated action of several musclegroups. The signal spreads through severallevels of the spinal cord and involves manyinterneurons.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

51
Spinal
Cord
13
109
11 5
1
7
2
812
813
4
6 7
514
2
Gray and White Matter, Reflex Arcs
A Cross section of spinal cord B Longitudinal zones of spinal cord
C Afferent fibers (ascending tracts) D Monosynaptic reflex arc
E Multisynaptic reflex arc
F Stretch reflex G Withdrawal reflex
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

52Sp
inal
Cord
Spinal Cord and Spinal Nerves: Spinal Cord
Gray Substance and IntrinsicSystem (A–E)
The posterior horn is formed by the nucleusproprius (A1), the major portion of the pos-terior horn from which the dorsal nucleus(Clarke’s nucleus) (A2) is set apart. The gelat-inous substance (Rolando’s substance) (A3)borders dorsally on the nucleus proprius.On it sits like a cap the end of the posteriorhorn, the marginal zone (nucleus postero-marginalis) (A4). The posterior horn is sepa-rated from the surface of the spinal cordthrough the posterolateral tract (Lissauer’stract) (A5). Between posterior horn andanterior horn lies the intermediate graymat-ter (A6) and lateral to it the lateral horn (A7).The border to the white matter betweenposterior horn and lateral horn is diffuse(reticular formation) (A8).
In the anterior horn, the motor neurons arearranged in groups of nuclei.
Medial group of nuclei
! Anteromedial nucleus (A9)! Posteromedial nucleus (A10)
Lateral group of nuclei
! Anterolateral nucleus (A11)! Posterolateral nucleus (A12)! Retroposterolateral nucleus (A13)
Central group of nuclei in the cervical spi-nal cord
! Phrenic nucleus! Accessory nucleus
For example, in the cervical spinal cord (B),the anterior horn is subdivided somatotopi-cally so that the neurons of the medialgroup of nuclei supply neck and backmuscles, intercostal, and abdominalmuscles (B14). The neurons of the antero-lateral nucleus supply the muscles ofshoulder girdle and upper arm (B15), andthe neurons of the posterolateral nucleussupply the muscles of lower arm and hand(B16). Finally, the retroposterolateral nu-cleus contains particularly large motor neu-rons that supply the small finger muscles(B17).
The neurons for the extensor muscles (B18)lie in the anterior field of the anterior horn,and those for the flexor muscles (B20) lieposterior to them. The somatotopic subdivi-sions do not occupy a single plane in theanterior horn but are spread over a certainheight in such a way that the neurons forthe shoulder girdle lie at a higher level,below them those for the upper arm, andstill deeper those for the lower arm andhand. Diagram (C) illustrates the nervesupply to all body muscles.
To bring about an orderly movement duringcontraction of a muscle group, there mustbe simultaneous relaxation of the corre-sponding antagonists. This is achievedthrough inhibition of the correspondinganterior horn cells (D). For example, if animpulse is passed on by a neuron of the ex-tensor muscles (D18), it is simultaneouslytransmitted by an axon collateral to inhibi-tory interneurons, the Renshaw cells (D19),which then inhibit the neurons of the flexormuscles (D20).
Intrinsic system of the spinal cord (E).Other interneurons mediate the spread ofimpulses over several levels, either on thesame side or on the opposite side. Their as-cending and descending fibers run in basicbundles, fasciculi proprii (E21), which borderdirectly on the gray matter. In general, theascending and descending fibers reach onlyone or two root levels. However, thefasciculi proprii also contain long fibersconnecting the cervical spinal cord and thelumbar spinal cord (as shown in cats andmonkeys). These fibers transmit excitatoryand inhibitory impulses to anterior hornmotor cells, a fact that is thought to be im-portant for coordinated movement of theanterior and posterior extremities duringlocomotion. About half of the posterolateraltract (Lissauer’s tract) (E5) consists of fibersof the intrinsic system.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
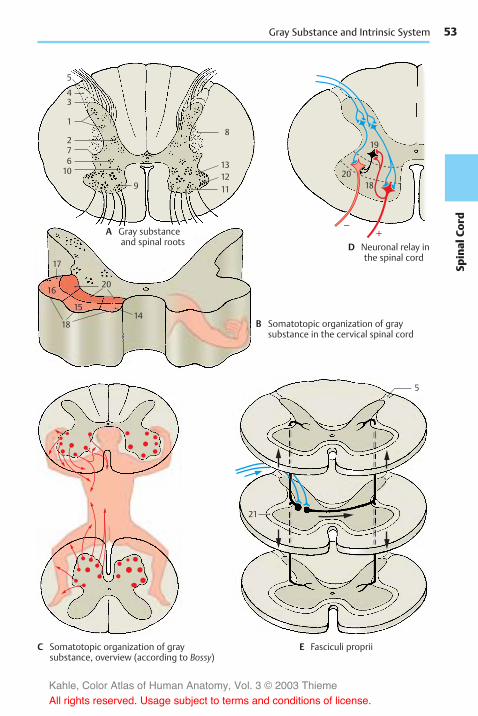
53
Spinal
Cord–
+
5
43
1
276
9 111213
819
2018
10
21
15
17
20
1418
16
5
Gray Substance and Intrinsic System
A Gray substanceand spinal roots
B Somatotopic organization of graysubstance in the cervical spinal cord
C Somatotopic organization of graysubstance, overview (according to Bossy)
D Neuronal relay inthe spinal cord
E Fasciculi proprii
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

54Sp
inal
Cord
Spinal Cord and Spinal Nerves: Spinal Cord
Cross Sections of the Spinal Cord(A–D)
Cross sections at different levels (left, my-elin stain; right, cellular stain) vary con-siderably. In the regions of cervical enlarge-ment and lumbar enlargement, the cross-sectional area is larger than in the rest of thespinal cord; it is largest at the C4–C5 andL4–L5 levels. In both swellings, the numer-ous nerves that supply the extremitiescause an increase in gray matter.
Thewhitematter is most extensive in the cer-vical region and diminishes gradually incaudal direction; the ascending sensorytracts increase in number from the sacral tothe cervical region as more fibers are added,while the descending motor tracts decreasefrom the cervical to the sacral regions asfibers terminate at various levels.
The butterfly configuration of the graymatterchanges in shape at the various levels, andso does the posterolateral tract (Lissauer’stract) (A–D1).
The posterior horn is narrow in the cervicalspinal cord; its tip ends in the cap-shapedmarginal zone (nucleus posteromarginalis)(A2). The lateral angle between the poste-rior and anterior horn is occupied by the re-ticular formation (AD3). The gelatinous sub-stance (Rolando’s substance) (A–D4) con-tains small, mostly peptidergic neuronswhere posterior root fibers of various cali-bers terminate; it also contains descendingfibers from the brain stem (raphe nuclei,p. 108, B28; reticular formation, p. 146). Un-myelinated processes of neurons ascend ordescend for one to four root levels withinthe posterolateral tract (Lissauer’s tract) andthen reenter into the gelatinous substance.Some of the processes run within the lateralspinothalamic tract to the thalamus(p. 324). The fibers of proprioceptive sensi-bility in the muscles (muscle spindles) ter-minate in the posterior thoracic nucleus (dor-sal nucleus of Clarke) (AB5) where the tractsto the cerebellum begin. The reduced graymatter of the thoracic spinal cord has aslender posterior horn with a prominentdorsal nucleus. In the plump posterior horn
of the lumbar and sacral spinal cords, thegelatinous substance (CD4) is much en-larged and borders dorsally on the narrowband of the marginal zone (CD2).
The lateral horn forms in the thoracic spi-nal cord the lateral intermediate substance(B6). It contains sympathetic nerve fibersmainly for the vasomotor system, the effer-ent fibers of which emerge via the anteriorroot. Sympathetic neurons also lie mediallyin the intermediomedial nucleus (B7). In thesacral spinal cord, parasympathetic neuronsform the intermediolateral nucleus und in-termediomedial nucleus (D8).
The anterior horn expands in the cervicalspinal cord and contains several nuclei withlargemotor neurons, all of which are cholin-ergic.
Medial group of nuclei! Anteromedial nucleus (A9)! Posteromedial nucleus (A10)
Lateral group of nuclei! Anterolateral nucleus (A11)! Posterolateral nucleus (A12)! Retroposterolateral nucleus (A13)
In the region supplying the upper limbs, theanterior horn is far more differentiated thanin the thoracic spinal cord where only a fewcell groups can be identified. The expanded,plump anterior horn of the lumbar andsacral spinal cords, which supplies thelower limbs, again contains several groupsof nuclei.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

55
Spinal
Cord
124
35
1312
10119
4
5
76
11
9
124
3
119
124
81210
11
9
1210
21
Transverse Sections of the Spinal Cord
A Cervical spinal cord
B Thoracic spinal cord
C Lumbar spinal cord
D Sacral spinal cord
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

56Sp
inal
Cord
Spinal Cord and Spinal Nerves: Spinal Cord
Ascending Pathways (A–D)
Tracts of the Anterolateral Funiculus (A)
Lateral spinothalamic tract (A1). The affer-ent, poorly myelinated posterior root fibers(A2) (first neuron of sensory pathway) bi-furcate in the posterolateral tract (Lissauer’stract) and terminate at the cells of the gelat-inous substance of the posterior horn. Thefibers of the tract originate here, cross in thewhite commissure to the opposite side, andascend in the lateral funiculus to thethalamus (second neuron). The pathwaytransmits pain and temperature sensation,exteroceptive and proprioceptive impulses. Itis somatotopically subdivided; sacral (S)and lumbar (L) fibers are located dorsolater-ally, while thoracic (T) and cervical (C) fibersare located ventromedially. Fibers for painsensation probably lie superficially, whilethose for temperature sensation lie moredeeply.
Anterior spinothalamic tract (A3). The af-ferent fibers (A4) (first neuron) bifurcateinto ascending and descending branchesand terminate at posterior horn cells, thefibers of which cross to the opposite sideand ascend in the anterior funiculus to thethalamus (second neuron). They transmitcrude touch and pressure sensations. To-gether with the lateral tract, they form thepathway of protopathic sensibility (p. 324).
The spinotectal tract (A5) carries painfibers to the roof of the midbrain (contrac-tion of pupils when in pain).
Pathways of the Posterior Funiculus (C, D)
Fasciculus gracilis (of Goll) (C6) andfasciculus cuneatus (of Burdach) (C7). Thethick heavily myelinated fibers ascendwithout relay in the ipsilateral posteriorfuniculi. They belong to the first neuron ofthe sensory pathway and terminate at thenerve cells of the posterior funiculus nuclei(second neuron) (p. 140, B5, B6). They trans-mit exteroceptive and proprioceptive im-pulses of the epicritic sensibility (exterocep-tive, information on localization and qualityof tactile sensation; proprioceptive, infor-
mation on limb position and body posture).The posterior funiculi are somatotopicallysubdivided; the sacral fibers lie medially,followed laterally by the lumbar andthoracic fibers (fasciculus gracilis). Thefibers from T3 to C2 lie laterally and formthe fasciculus cuneatus.
Short ascending collaterals (C8) branchfrom the ascending fibers. They terminate atthe posterior horn cells and form compactbundles, namely, the comma tract ofSchultze (D9) in the cervical spinal cord,Flechsig’s oval field (D10) in the thoracic spi-nal cord, and the Phillippe–Gombault tri-angle (D11) in the sacral spinal cord.
Cerebellar Pathways of the LateralFuniculus (B)
Posterior spinocerebellar tract (Flechsig’stract) (B12). The afferent posterior hornfibers (first neuron) terminate at the cells ofthe dorsal nucleus of Clarke (B13) fromwhere the tract (second neuron) originates.It runs along the margin of the ipsilaterallateral funiculus to the cerebellum andtransmits mainly proprioceptive impulses(from joints, tendons, muscle spindles).
Anterior spinocerebellar tract (Gowers’tract) (B14). The cells of origin lie in the pos-terior horn. Their fibers (second neuron) as-cend ipsilaterally as well as contralaterallyalong the anterolateral margin of the spinalcord to the cerebellum, to which they trans-mit exteroceptive and proprioceptive im-pulses. Both cerebellar pathways are soma-totopically subdivided; the sacral fibers liedorsally, the lumbar and thoracic fibers ven-trally.
The spino-olivary tract (B15) and vesti-bulospinal tract (B16) arise from the poste-rior horn cells of the cervical spinal cord;they transmit mainly proprioceptive im-pulses to the inferior olive of the oppositeside and to the vestibular nuclei.
A–C17 Neurons in the spinal ganglion (firstneuron) (p. 71, A7).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

57
Spinal
Cord
142
1412 17
12
14
17
67
7
6
17
15
16
13
4 4
2
1
1
35
17
9
10
8
11
8
Ascending Pathways
A Lateral and anteriorspinothalamic tracts
B Anterior and posteriorspinocerebellar tracts
C Fasciculus gracilis,fasciculus cuneatus
D Descending fibers of theposterior funiculi
S
L
T
S
L
T
SL
TC
C 1C 8 T L S
C
L
S
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

58Sp
inal
Cord
Spinal Cord and Spinal Nerves: Spinal Cord
Descending Pathways (A–C)
Corticospinal Tract, Pyramidal Tract (A)
The fibers of the pyramidal pathway origi-nate mostly from the precentral gyrus andthe cortex in front of it (areas 4 and 6)(p. 308, A1, A2). Furthermore, some of thefibers are thought to be derived from thecortical regions of the parietal lobe. Eightypercent of all fibers cross in the lowermedulla oblongata to the collateral side(pyramidal decussation) (A1) and run as lateralcorticospinal tract (A2) in the lateralfuniculus. The remaining fibers run un-crossed as anterior corticospinal tract (A3) inthe anterior funiculus and cross only at thelevel of their termination. More than half ofthe pyramidal tract fibers terminate in thecervical spinal cord to supply the upperlimb, and one-fourth terminate in the lum-bar spinal cord to supply the lower limb. Thelateral funiculus is subdivided somatotopi-cally, with the fibers for the lower limb lyingat the periphery and those for the trunk andupper limb lying further inside. Most of thefibers terminate on interneurons that trans-mit impulses for the voluntary motor sys-tem to the anterior horn cells. However, thefibers not only conduct excitation to theanterior horn cells but also mediate corticalinhibition via interneurons (p. 308, p. 316).
Extrapyramidal Tracts (B)
The extrapyramidal tracts comprise de-scending systems from the brain stem thatinfluence the motor system (p. 310):
! Vestibulospinal tract (B4) (balance, muscletone)
! Anterior and lateral reticulospinal tract (B5)from the pons
! Lateral reticulospinal tract (B6) from themedulla oblongata
! Tegmentospinal tract (B7) from the mid-brain
The rubrospinal tract (B8) (in humans largelyreplaced by the tegmentospinal tract) andthe tectospinal tract (B9) terminate in the cer-vical spinal cord and influence only thedifferentiated motor system of the head andupper limb. The medial longitudinal fascicle
(B10) contains various fiber systems of thebrain stem (p. 142).
Autonomic Pathways (C)
The autonomic pathways consist of poorlymyelinated or unmyelinated fibers andrarely form compact bundles. The par-ependymal tract (C11) runs along both sidesof the central canal. Its ascending and de-scending fibers can be traced back up to thediencephalon (hypothalamus) and arethought to transmit impulses for genitalfunction, micturition, and defecation. Ante-rior to the lateral pyramidal tract runs thedescending pathway for vasoconstrictionand sweating (Foerster) (C12) with a soma-totopic subdivision corresponding to that ofthe lateral pyramidal tract.
Visualization of Pathways (D–E)
The various pathway systems cannot beidentified on normal transverse sections ofthe spinal cord. Only under special circum-stances (in experimental transection, in spi-nal cord injury, or during developmentwhentracts become myelinated at differenttimes), can they be distinguished from eachsother, such as the late myelinating pyra-midal tract (D2). In case of injuries, distalfibers separated from the perikaryondegenerate so that their area in the spinalcord becomes visible, such as the fasciculusgracilis (E13).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

59
Spinal
Cord
9
6
10
7
8
45
1112
1
2
2
13
3
Descending Pathways, Visualization of Pathways
A Anterior and lateral corticospinaltract (pyramidal tract)
B Descending pathways
C Autonomic pathways
D Unmyelinated pyramidal tractin the newborn
E Degeneration of the fasciculus gracilisafter injury to the spinal cord
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

60Sp
inal
Cord
Spinal Cord and Spinal Nerves: Spinal Cord
Blood Vessels of the Spinal Cord(A–E)
The spinal cord is supplied with blood fromtwo sources, the vertebral arteries and thesegmental arteries (intercostal arteries andlumbar arteries).
Vertebral arteries (A1). Before they unite,they give off two thin posterior spinal arter-ies that form a network of small arteriesalong the posterior surface of the spinalcord. At the level of the pyramidal decussa-tion, two additional branches of the verte-bral arteries join to form the anterior spinalartery (AD2) which runs along the anteriorsurface of the spinal cord at the entrance tothe anterior sulcus.
Segmental arteries (C3). Their posteriorbranches (C4) and the vertebral arteries giveoff spinal branches (C5) which enter throughthe intervertebral foramina and divide atthe spinal roots into dorsal and ventralbranches to supply the spinal roots and thespinal meninges. Of the 31 spinal arteries,only 8 to 10 extend to the spinal cord andcontribute to its blood supply. The levels atwhich the radicular arteries approach thespinal cord vary, and so do the sizes of thevessels. The largest vessel approaches thespinal cord at the level of the lumbar en-largement between T12 and L3 (large radicu-lar artery) (A6).
The anterior spinal artery is widest at the levelof the cervical and lumbar enlargements. Itsdiameter is much reduced in the mid-thoracic region of the spinal cord. As this re-gion is also the border area between twosupplying radicular arteries, this segment ofthe spinal cord is especially at risk in case ofcirculatory problems (A, arrow). Dependingon the variation of the radicular arteries,this may also apply to other segments of thespinal cord.
The anterior spinal artery gives off numer-ous small arteries into the anterior sulcus,the sulcocommissural arteries (D7). In the cer-vical and thoracic spinal cords, they turn al-ternately to the left and right halves of thespinal cord; in the lumbar and sacral spinalcords, they divide into two branches. In ad-
dition, anastomoses arise between the ante-rior and posterior spinal arteries, so that thespinal cord is surrounded by a vascular ring(vasocorona) (D8) fromwhere vessels radiateinto the white matter. Injection of tracersrevealed that the gray matter is much morevascularized than the white matter (D).
Areas of blood supply (E). The anterior spi-nal artery supplies the anterior horns, thebases of the posterior horns, and the largestpart of the anterior lateral funiculi (E9). Theposterior funiculi and the remaining partsof the posterior horns are supplied by theposterior spinal arteries (E10). The marginalzone of the anterior lateral funiculus is sup-plied by the plexus of the vasocorona (E11).
The spinal veins (B) form a network inwhich one anterior spinal vein and two pos-terior spinal veins stand out. The efferentveins run along the spinal roots and openinto the epidural venous plexus (see vol. 2).The spinal veins lack valves prior to theirpenetration through the dura.
C12 Aorta.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

61
Spinal
Cord
6
1
2 5
3
4
12
8
7
2
11 9
10
Spinal Blood Vessels
A B
Arteries and veinsof the spinal cord
C Afferent blood vessels
D Vascularization of the spinal cord
E Areas supplied by the spinal cordarteries (according to Gillilan)
C 1
C 3
C 5
T 1
T 3
T 5
T 8
T10
L 2
L 5
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

62Sp
inal
Cord
Spinal Cord and Spinal Nerves: Spinal Cord
Spinal Ganglion and Posterior Root(A–H)
The posterior spinal root contains a spindle-shaped bulge, the spinal ganglion (A), anaccumulation of cell bodies of sensory neu-rons; their bifurcated processes send onebranch to the periphery and the otherbranch to the spinal cord (p. 70, A7). They lieas cell clusters or as cell rows between thebundles of nerve fibers.
Development of the ganglia (C). The cellsoriginate from the lateral zone of the neuralplate (C1); however, they do not participatein the formation of the neural tube but re-main at both sides as the neural crest (C2).Hence, the spinal ganglia can be regarded asgray matter of the spinal cord that becametranslocated to the periphery. Other deriva-tives of the neural crest are the cells of theautonomic ganglia, the paraganglia, and theadrenal medulla.
From the capsule (A3) of the spinal gan-glion,whichmerges into the perineuriumofthe spinal nerve, connective tissue extendsto the interior and forms a sheath aroundeach neuron (endoganglionic connectivetissue) (B4). The innermost sheath, however,is formed by ectodermal satellite cells (BE5)and is surrounded by a basal membranecomparable to that around the Schwanncells of the peripheral nerve. The large nervecells (B6, E) with their myelinated processconglomerated into a glomerulus representonly one-third of the ganglion. They trans-mit impulses of epicritic sensibility (p. 322).The remainder consists of medium-sizedand small ganglion cells with poorly myeli-nated or unmyelinated nerve fibers whichare thought to conduct pain signals and sen-sations from the intestine. There are alsosome multipolar nerve cells.
Development of the ganglion cells (D). Thespinal ganglion cells are initially bipolarcells. During development, however, thetwo processes fuse to form a single trunkwhich then bifurcates in a T-shaped man-ner. The cells are therefore called pseudo-unipolar nerve cells.
The posterior root is thicker than the ante-rior root. It contains fibers of various cali-bers, two-thirds of them being poorly my-elinated or unmyelinated fibers. The thinpoorly myelinated and unmyelinated fibers,which transmit impulses of the protopathicsensibility (p. 324), enter through the lateralpart of the root into the spinal cord (F7). Thethick myelinated fibers transmit impulsesof the epicritic sensibility and enter throughthe median part of the root into the spinalcord (F8).
At the entrance into the spinal cord, there isa narrow zonewhere themyelin sheaths arevery thin so that the fibers appear unmyeli-nated. This zone is regarded as the boundarybetween the central and the peripheralnervous systems (Redlich–Obersteiner zone)(G). In the electron-microscopic image (H),however, this boundary does not exactlycoincide with the Redlich–Obersteinerzone. For each axon, the boundary ismarkedby the last node of Ranvier prior to the en-trance into the spinal cord. Up to this point,the peripheral myelin sheath is surroundedby a basal membrane (blue in H). The nextinternode no longer has a basal membrane.For unmyelinated fibers, the boundary isalso marked by the basal membrane of theenveloping Schwann cell. Thus, the basalmembranes around the spinal cord form aboundary that is only penetrated by theaxons.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

63
Spinal
Cord
5
5
6 4
3
1
2 2
7
8
Spinal Ganglion and Posterior Root
A Spinal ganglion B Detail of A
C Development of the spinal ganglion
D Development of the pseudounipolar ganglion cell
E Spinal ganglion cell andsatellite cells
F Posterior root
G Redlich–Obersteiner zone H Posterior root, electron-microscopic diagram (according to Andres)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

64Sp
inal
Cord
Spinal Meninges (A–E)
The spinal cord in the vertebral canal is sur-rounded by the following connective tissuemembranes: the tough spinal meninx(pachymeninx), or spinal dura mater (A1), andthe soft spinal meninx (leptomeninx) con-sisting of the spinal arachnoidea (A2) and thespinal pia mater (A3).
The spinal dura mater forms the outermostsheath which is separated from the perios-teum-like lining of the vertebral canal, theendorhachis (A4), by the epidural space (A5).The space is filled with adipose tissue andcontains an extensive venous plexus, the in-ternal vertebral venous plexus (see vol. 2).The dura mater forms caudally the dural sac(B6), enveloping the cauda equina (B7), andfinally extends together with the terminalfilament as a thin cord up to the periosteumof the coccyx (dural terminal filament) (B8).Only at the oral end at the foramen magnum(occipital bone) is the dural sac attached tobone. The epidural space forms a resilientcushion for the dural sac, which moves to-gether with the vertebral column and thehead. Bending the head pulls the dural sacupward, causing mechanical stress on thespinal cord; when bending the head for-ward, roots and blood vessels are stretched(D9), when bending the head backward,they are compressed (D10).
The arachnoidea borders closely onto theinner surface of the dura mater. It forms theboundary of the subarachnoidal space (AC11),which is filled with cerebrospinal fluid (CSF).Between the inner surface of the dura andthe arachnoidea lies a capillary cleft, thesubdural space, which widens into a realspace only under pathological conditions(subdural bleeding). Dura and arachnoideaaccompany the spinal roots (AC12), passwith them through the intervertebralforamina, and also envelope the spinal gan-glia (AC13). The funnel-like root sleevescontain CSF in their proximal portions. Thedura then turns into the epineurium (A14),and the arachnoidea into the perineurium(A15) of the spinal nerves. The part of theroot leaving the vertebral canal, the radicu-
lar nerve (A16), runs obliquely downward inthe cervical and lumbosacral regions andobliquely upward in the midthoracic region(C).
The spinal pia mater borders directly ontothe marginal glial layer of the spinal cord.This represents the boundary between me-sodermal envelopes and ectodermal nervetissue. The pia mater contains numeroussmall blood vessels that penetrate from thesurface into the spinal cord. A connectivetissue plate, the denticulate ligament (A17),extends on both sides of the spinal cordfrom the pia to the dura and is attached tothe latter by individual pointed processes.The ligament extends from the cervical spi-nal cord to the midlumbar spinal cord, thuskeeping the spinal cord, which floats in theCSF, in position.
Clinical Note: Under sterile conditions, cere-brospinal fluid may be safely withdrawn for ex-amination from the lower segment of the duralsac that contains only the fibers of the caudaequina. For this purpose, with the patient bendingover, a needle is deeply inserted between theprocesses of the second to fifth lumbar vertebraeuntil CSF begins to drop (lumbar puncture) (E).
Spinal Cord and Spinal Nerves: Spinal Cord
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

65
Spinal
Cord
1 23
11
45
1712
16
1314
15
12
1316
11
9 10
13
6
7
8
1 23
11
45
1712
16
1314
15
12
1316
11
9 10
13
6
7
8
Spinal Meninges
A Position of the spinal cord in thevertebral canal, cross section(according to Rauber-Kopsch)
B Cauda equina
C Dorsal view of root, radicularnerve, and spinal ganglion
D Cervical spinal cord with neckflexed and extended (accordingto Breig)
E Lumbar puncture
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

66Sp
inal
Cord
Spinal Cord and Spinal Nerves: Spinal Cord
Segmental Innervation (A–C)
The vertebrate body, with the exception ofthe head, is originally subdivided into seg-ments ormetameres. The vertebrae, ribs, andintercostal muscles can be regarded as rem-nants of such a segmentation in humans.Metamerism concerns only tissues of themesoderm (myotomes, sclerotomes) but notderivatives of the ectoderm. Thus, there areno spinal cord segments, only the levels atwhich the individual spinal roots enter andemerge. However, the spinal fibers join toform the spinal nerves as they emergethrough the metameric intervertebralforamina, thus creating an apparent second-ary segmentation. The sensory fibers of thespinal nerves supply stripe-shaped zones ofthe skin, called dermatomes in analogy tomyotomes and sclerotomes. This, too, is asecondary segmentation and reflects the in-nervation of each dermatome by a singleposterior root (segmental innervation).
Clinical Note: The dermatomes play an im-portant role in the diagnosis and localization ofspinal cord injuries. Loss of sensibility in certaindermatomes indicates a specific level of injury inthe spinal cord. Simplified reference points arethe line through the nipples, regarded as theboundary between T4 and T5, and the groin, re-garded as the boundary between L1 and L2. Thefirst cervical spinal nerve has no sensory repre-sentation on the body surface, for the spinal gan-glion of its posterior root is absent or rudimen-tary.
There are slightly different segmentalboundaries for various modalities, such astouch and pain, and for sweating andpiloerection. The diagram (A) was designedaccording to the decrease in sensibility (hy-poesthesia) resulting from disk prolapse; itshows how the dermatomes extendingaround the trunk become elongated in thelimbs. They may even lose their continuitywith the midline (C7, L5). They becometranslocated to the distal limb areas duringembryonic development when the limbsare budding (C).
The dermatomes overlap like roof tiles, as il-lustrated by the shift in boundaries thathave been determined according to the ex-
panded areas in case of posterior root pain(hypersensitivity to pain, hyperalgesia) (B).The loss of a single posterior root cannot bedemonstrated for touch sensation, since thecorresponding dermatome is also suppliedby the neighboring posterior roots. The der-matomes for pain and temperature sensa-tion are narrower, and the loss of a posteriorroot can still be demonstrated when thesemodalities are tested.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

67
Spinal
Cord
4 43
56
78
123
5678
23
345
78
61
23
3
5
7
9
11
4
6
8
10
12
Dermatomes
A Dermatomes(according toKeegan and Garrett)
B Overlap of dermatomes (accord-ing to Förster)
C Development of dermatomes inthe upper limb (according to Bolk)
C 3C 4C 5C 6 T 1
T 3
T 6
T 9
C 5C 6
C 7 C 8
L 1
L 2
L 3
T 12
S 2S 3
L 4
L 5
S 1C 5
C 3 C 4
T 1T 2
L 1
L 2L 3
C 2C 3C 4C 5
C 6C 7C 8
T 1T 6
T 12
L 1L 3L 5
C 6C 7
C 8S 1
S 2
S 3
L 5
L 4
S 1
S 2
C 3
T 1
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

68Sp
inal
Cord
Spinal Cord and Spinal Nerves: Spinal Cord
Spinal Cord Syndromes (A–C)
The anatomy of the spinal cord causes veryspecific patterns of functional deficienciesafter injury; depending on the site of lesion,different pathways and therefore differentfunctions are lost.
Complete transection (A) cuts off all de-scending motor pathways, causing completeparalysis below the injured level. At thesame time, it interrupts all ascending path-ways, causing a complete loss of all sensa-tions. If the lesion is above the sacral spinalcord, it results in the loss of voluntary con-trol over urination and defecation. If the le-sion lies above the lumbar enlargement,both lower limbs are paralyzed (paraplegia),and if it lies above the cervical enlargement,both upper limbs are also paralyzed (tet-raplegia).
Hemisection of the spinal cord (B) results inthe Brown–Séquard’s syndrome. For example,hemisection on the left interrupts thelateral and anterior corticospinal tracts (B1)and results in left-sided paralysis. Transec-tion of the vasomotor pathway causes ipsi-lateral vasomotor paralysis. Transection ofthe posterior funiculi (B2) and the cerebel-lar lateral funiculi (B3) leads to severely im-paired deep sensibility (posture sensation).On the same side as the lesion, there is alsohyperesthesia (touch is perceived as pain).This is thought to be caused by a loss ofepicritic sensibility (posterior funiculi) withretention of the protopathic sensibility(crossing pathways of the anterior corti-cospinal tract ascend contralaterally) (B4).Finally, there is dissociated anesthesia on theintact right side from the lesion downward;while touch sensation is hardly impaired,pain and temperature sensations are lost(ipsilateral interruption of the crossingpathway of the anterior corticospinal tract)(B5). The anesthetic zone (B6) above thetransection on the affected side is attributedto destruction of the posterior root entrancezone at the level of the spinal cord lesion.
Central injury (C) to the gray substance ofthe spinal cord also causes dissociated anes-thesia at the corresponding levels. The
epicritic sensibility transmitted via the ipsi-lateral posterior funiculi (C2) is retained.However, pain and temperature sensationsare lost (analgesia and thermoanesthesia), be-cause their fibers, which cross through thewhite commissure, are interrupted (C5).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

69
Spinal
Cord
3
1
52
4
2
5
6
Spinal Cord Syndromes
A Complete transection of thespinal cord
B Brown–Séquard’s syndrome inhemisection of the spinal cord
C Dissociated anesthesia in case ofinjury to the central spinal cord
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

70Sp
inal
Cord
Spinal Cord and Spinal Nerves
Peripheral Nerves
The peripheral nerves may contain fourdifferent types of fibers:
! Somatomotor (efferent) fibers (A1) forstriated muscles
! Somatosensory (afferent) fibers (A2) forskin sensibility
! Visceromotor fibers (A3) for smoothmuscles
! Viscerosensory fibers (A4) for inner organs
The spinal nerves usually contain severaltypes of fibers; they are mixed nerves.
The different fibers have the following path-ways. The somatomotor fibers pass from theanterior horn cells (A5) through the anteriorroot (A6); the somatosensory andviscerosensory fibers originate from thenerve cells of the spinal ganglia (A7); andthe visceromotor fibers of the lateral horncells (A8) pass mostly through the anteriorroot. Anterior and posterior roots (A9) jointo form the spinal nerve (A10), which con-tains all types of fibers. This short nervetrunk then divides into four branches:
! The meningeal branch (A11), a recurrentsensory branch extending to the spinalmeninges
! The posterior branch (A12)! The anterior branch (A13)! The communicating branch (A14)
The posterior branch supplies motor fibers tothe deep (autochthonous) muscles of theback and sensory fibers to the skin areas onboth sides of the vertebral column (p. 84).The anterior branch supplies motor fibers tothe muscles of the anterior and lateral wallsof the trunk and to themuscles of the limbs;it also supplies sensory fibers to the corre-sponding skin areas. The communicatingbranch connects with the sympathetic chainganglion (A15) (autonomic nervous system,p. 292). It usually forms two independentcommunicating branches, the white com-municating branch (A16) (myelinated) andthe gray communicating branch (A17) (un-myelinated). The visceromotor fibers passvia the white branch to the sympatheticchain ganglion, where they are relayed toneurons, the axons of which partly reenter
the spinal nerve as postganglionic fibers(p. 297 A5) via the gray branch.
Nerve Plexusus (B)
At the level of the limbs, the anteriorbranches of the spinal nerves form networks(plexusus) in which fibers are exchanged.The resulting nerve trunks, which then ex-tend to the periphery, possess a newly or-ganized supply of fibers derived from differ-ent spinal nerves.
Cervical plexus (p. 72). The plexus of theneck is formed by the anterior branch of thefirst four spinal nerves. The followingnerves originate here: the lesser occipitalnerve (B18), the greater auricular nerve(B19), the transverse nerve of the neck(B20), the supraclavicular nerves (B21), thephrenic nerve (B22), and also the roots ofthe deep cervical ansa (B23).
Brachial plexus (p. 74). The plexus of thearm is formed by the anterior branches ofspinal nerves C5 to C8 and by a part of the T1nerve. We distinguish between a sectionlying above the clavicle, the supraclavicularpart, and a section lying below the clavicle,the infraclavicular part. The anterior branchespass through the scalene gap into the poste-rior cervical triangle, where they form threeprimary trunks above the clavicle:
! The superior trunk (B24) (C5, C6)! The medial trunk (B25) (C7)! The inferior trunk (B26) (C8, T1)
The nerves that originate here form the su-praclavicular part (p. 74). Below the clavicle,three secondary cords form; they are namedaccording to their position relative to theaxillary artery (B27):
! The lateral cord (B28) (p. 74) (from theanterior branches of the superior andmedial trunks)
! The medial cord (B29) (p. 78) (from theanterior branch of the inferior trunk)
! The posterior cord (B30) (p. 80) (from thedorsal branches of the three trunks)
The lateral cord gives rise to themusculocu-taneous nerve (B31). The remaining fiberstogether with fibers of the medial cord form
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

71
Spinal
Cord
97
11
13
12
13
210
16
14
17
15
4
6
8
5
19
24
25
26
18
23
2120
22
30
28
27
29
37
32
33
31
38 34 3635
97
11
13
12
13
210
16
14
17
15
4
6
8
5
19
24
25
26
18
23
2120
22
30
28
27
29
37
32
33
31
38 34 3635
Composition of Peripheral Nerves, Cervical and Brachial Plexus
A Composition of the peripheral nerves
B Cervical plexus and brachial plexus(preparation by Professor Platzer)
Cervical plexusBrachial plexusParts of the lateral cordParts of the medial cordParts of the posterior cord
C 1
C 2
C 3
C 4
C 5
C 6
C 7
C 8
T 1
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

72Sp
inal
Cord
the median loop (B32) (p. 76, AC1) and uniteto form the median nerve (B33). The medialcord gives rise to the ulnar nerve (B34), themedial cutaneous nerve of the forearm (B35),and the medial cutaneous nerve of the arm(B36 ). The posterior cord gives off the axil-lary nerve (B37) and continues as the radialnerve (B38).
Spinal Cord and Spinal Nerves: Peripheral Nerves
Cervical Plexus (C1–C4) (A–D)
Innervation of the muscles (A). Shortnerves run from the anterior branchesdirectly to the deep neck muscles, namely,the anterior (A1) and lateral (A2) rectuscapitismuscles, the longmuscle of the head,and the long muscle of the neck (A3). Fromthe anterior branch of C4, nerves run to theupper part of the anterior scalene muscle(A4) and to the medial scalene muscle (A5).
The anterior branches of C1–C3 form thedeep cervical ansa (C6): fibers from C1 andC2 temporarily appose the hypoglossalnerve (AC7) and then leave it as the superiorroot (anterior) (AC8); the fibers for the thy-rohyoid muscle (A9) and the geniohyoidmuscle then continue with the hypoglossalnerve. The superior root combines with theinferior root (posterior) (AC10) (C2, C3) toform the cervical ansa, from wherebranches run to supply the infrahyoidmuscles, namely, the omohyoid muscle(A11), the sternothyroid muscle (A12), andsternohyoid muscle (A13).
Innervation of the skin (B, C). The sensorynerves of the plexus pass behind the sterno-cleidomastoid muscle through the fascia,where they form the punctum nervosum(B14). From here they spread over head,neck, and shoulder; the lesser occipital nerve(BC15) (C2, C3) extends to the occiput, thegreater auricular nerve (BC16) (C3) into thearea surrounding the ear (auricula, mastoidprocess, region of the mandibular angle).The transverse nerve of the neck (BC17) (C3)supplies the upper neck region up to thechin, while the supraclavicular nerves (BC18)(C3, C4) supply the subclavicular fossa andthe shoulder region.
Area innervated by the phrenic nerve (C,D). The phrenic nerve (CD19) (C3, C4) con-tains fibers of the fourth, and often also of
the third, spinal nerve. It crosses the ante-rior scalene muscle and enters into the su-perior thoracic aperture in front of the sub-clavian artery. It extends through the medi-astinum to the diaphragm and, on its way,gives off fine branches for sensory supply tothe pericardium, the pericardiac branches(D20). At the surface of the diaphragm, itbranches and supplies all muscles of the di-aphragm (D21). Fine branches provide thesensory fibers for themembranes borderingon the diaphragm, that is, cranially thepleura and caudally the peritoneum of thediaphragm and the peritoneal covering ofthe upper intestinal organs.
Clinical Note: Injury to the cervical spinalcord or its roots at the C3 – C5 levels results in pa-ralysis of the diaphragm and in reduced respira-tion. In case of paralysis of the thoracic muscles,on the other hand, respiration can still be main-tained by the cervical spinal cord via the phrenicnerve.
Posterior Branches (C1–C8)
The dorsal branches of the cervical nerves, orposterior branches, supply motor fibers toneck muscles belonging to the autoch-thonous muscles of the back and sensoryfibers to the skin of the neck.
The posterior branch of the first cervicalnerve is exclusively motor and runs as sub-occipital nerve to the small muscles in theregion of occiput, atlas, and axis.
The greater occipital nerve runs from thesecond cervical nerve to the occiput andsupplies its skin up to the vertex (p. 84 D4).
The posterior branch of the third cervicalspinal nerve, the third occipital nerve, sup-plies sensory fibers to the neck region.
The remaining posterior branches of thecervical spinal nerves supply sensory fibersto the skin area bordering caudally andmotor fibers to the autochthonous backmuscles of this region.
Innervation of the skin (B). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

73
Spinal
Cord
12
37
85
7
815
16
17
10
18 19
64
10
9
13
12
11
19
20
15
16
141817
2121
Cervical Plexus
A Muscles supplied bythe cervical plexus
B Skin area supplied by thecervical plexus(according to Lanz-Wachsmuth)
C Cervical plexus
D Area supplied by the phrenic nerve
C 1C 2
C 3
C 4
C 1
C 2
C 3
C 4
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

74Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Brachial Plexus (C5–T1)
Peripheral sensory innervation. The in-nervation of the skin by peripheral nervesoriginating from the plexus differs from thesegmental innervation (p. 66). The regionssupplied by individual nerves overlap attheir margins. The region innervated by asingle nerve is called the autonomic zone(dark blue), and the total area supplied bythe nerve including the area cosupplied byadjacent nerves is called themaximum zone(light blue).
Clinical Note: Interruption of a nerve causescomplete insensibility (anesthesia) in the auto-nomic zone but only a decreased sensibility (hy-poesthesia) in the maximum zones.
Supraclavicular Part (A–C)
The supraclavicular part gives rise to motornerves that innervate the muscles of theshoulder girdle.
The following nerves run to the posteriorand lateral surfaces of the thorax: the dorsalscapular nerve (A1) (C5) to the scapularmuscle (C2) and to the lesser (C3) andgreater (C4) rhomboid muscles; the longthoracic nerve (A5) (C5 – C7), the branches ofwhich terminate at the lateral thoracic wallin the peaks of the anterior serratus muscle(B6); and the thoracodorsal nerve (A7) (C7,C8), which supplies the latissimus dorsimuscle (C8). The muscles of the shoulderblade are innervated at the posterior surfaceof the shoulder blade (supraspinous muscle[C9] and infraspinous muscle [C10]) by thesuprascapular nerve (A11) (C5, C6), and at theanterior surface by the subscapular nerve(A12) (C5–C7), which extend to the sub-scapular muscle and the greater teresmuscle (C13).
The following nerves reach the anterior sur-face of the thorax: the subclavius nerve (A14)(C4–C6) (to the subclavius muscle [B15),the lateral pectoral nerve (A16) (C5–C7) andthe medial pectoral nerve (A17) (C7–T1),which supply the greater (B18) and lesser(B19) pectoral muscles.
Clinical Note: Injury to the supraclavicularpart leads to paralysis of the muscles of theshoulder girdle and makes it impossible to raisethe arm. This type of upper brachial plexus paraly-sis (Erb’s palsy) may be caused by dislocation ofthe shoulder joint during birth, or through im-proper positioning of the arm during anesthesia.Injury to the infraclavicular part of the brachialplexus results in lower brachial plexus paralysis(Klumpke ’s palsy), which predominantly involvesthe small muscles of the hand and possibly alsothe flexor muscles of the forearm.
Infraclavicular Part (D–F)
Three main trunks of the anterior branches,the superior, middle, and inferior trunks ofthe brachial plexus, give rise to three cords,the lateral, middle, and posterior fascicles;they are named according to their positionrelative to the axillary artery.
Lateral Fascicle
The lateral fascicle gives rise to themusculocutaneous nerve and the mediannerve.
Musculocutaneous nerve (C5–C7) (D–F).The nerve passes through the coraco-brachial muscle and runs between the bi-ceps muscle and the brachial muscle downto the elbow. It gives off branches (E20) tothe flexor muscles of the upper arm,namely, to the coracobrachial muscle (D21),to the short head (D22) and long head (D23)of the biceps muscle of the arm, and to thebrachial muscle (D24).
The sensory fibers of the nerves come to thesurface through the fascia at the elbow andsupply the skin in the lateral region of theforearm as lateral cutaneous nerve of the fore-arm (D–F25). Injury to this nerve causesloss of sensibility in a small zone of theelbow; diminished sensibility extends tothe middle of the forearm.
Innervation of the skin (F). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

75
Spinal
Cord
15
19
18
1
11
14
7 12 5
2
3
4
9
10
13
8
567
1617
6
21
22
24 25
20
25
23
25
Brachial Plexus
A Supraclavicular part of thebrachial plexus
B Muscles supplied by the supra-clavicular part, frontal view
C Muscles supplied by the supra-clavicular part, dorsal view
D Muscles supplied by themusculocutaneous nerve(according to Lanz-Wachsmuth)
E Sequence of branches
F Innervation of skin
C 4
C 5
C 6
C 7
C 8
T 1
C 5
C 6C 7
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

76Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Infraclavicular Part (continued)
Lateral Fascicle (continued) (A–D)
Median nerve (C6–T1). Parts of the lateraland medial fascicles form the median loop(AC1) at the anterior surface of the axillaryartery and join to form the median nerve.
The nerve extends in the medial bicipitalsulcus along the surface of the brachialartery to the elbow, where it passes be-tween the two heads of the round pronatormuscle to the forearm. It runs between thesuperficial flexor muscle of the fingers andthe deep flexor muscle of the fingers to thewrist. Prior to its passage through the carpaltunnel, it lies superficially between the ten-dons of the radial flexor muscle of the wristand the long palmar muscle. In the carpaltunnel, it ramifies into its terminalbranches.
The muscular branches (C2) of the nervesupply the pronator muscles and most of theflexor muscles of the forearm, namely, theround pronator muscle (A3), the radialflexor muscle of the wrist (A4), the long pal-mar muscle (A5), and the superficial flexormuscle of the fingers with radial head (A6)and humeroulnar head (A7). In the elbow,the anterior interosseous nerve of the forearm(AC8) branches off and runs along the inter-osseous membrane to the quadrate prona-tor muscle (A9). It gives off branches to thelong flexor muscle of the thumb (A10) andto the radial part of the deep flexor muscleof the fingers.
In the lower third of the forearm, thesensory palmar branch of the median nerve(A–C11) branches off to the skin of the ballof the thumb (thenar eminence), to theradial side of the wrist, and to the palm.
After passing through the carpal tunnel, themedian nerve divides into three branches:the common palmar digital nerves I– III(A–C12), each of which bifurcates at thelevel of the metacarpophalangeal joints intotwo proper palmar digital nerves (A–C13).From the first common palmar digital nerve,a branch extends to the thenar eminence(short abductor muscle of thumb [A14], su-
perficial head of the short flexor muscle ofthumb [A15 ], and opposing muscle ofthumb [A16]). The common palmar digitalnerves supply the lumbrical muscles I– III(A17). They run to the interdigits and bifur-cate in such a way that each pair of properpalmar digital nerves provides sensoryfibers to the lateral aspects of an interdigit.Thus, the first pair of nerves supplies theulnar side of the thumb and the radial sideof the index finger, the second pair suppliesthe ulnar side of the index finger and theradial side of the middle finger, and thethird pair supplies the ulnar side of themiddle finger and the radial side of the ringfinger. The area innervated by the properpalmar digital nerves on the posterior sideincludes the end phalanx of the thumb aswell as the distal and middle phalanges ofthe fingers (B).
The median nerve gives off branches to theperiosteum, elbow joint, radiocarpal joint,and mediocarpal joint. At the level of thewrist, there is normally an anastomosiswith the ulnar nerve.
Clinical Note: After injury to the nerve, pro-nation of the forearm is no longer possible andflexion is severely restricted. As to the hand, thethumb, index finger and middle finger can nolonger be flexed at the end and middle phalanges,resulting in a characteristic feature of median pa-ralysis, the so-called hand of oath (D). On passingthe carpal tunnel, the nerve can be injured bypressure in older persons (carpal tunnel syn-drome).
Innervation of the skin (B). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

77
Spinal
Cord
1
9
10
7
6
11
13
1611
1415 12
17
13
458
3
1
2
8
1112
13
A Muscles supplied by the median nerve(according to Lanz-Wachsmuth)
B Skin supplied by the median nerve
C Sequence of branches
D Paralysis of the median nerve(according to Lanz-Wachsmuth)
C 6C 7C 8T 1
Brachial Plexus
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

78Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Infraclavicular Part (continued)
Medial Fascicle (A–D)
Ulnar nerve (C8–T1). Initially, the ulnarnerve runs in the upper arm in the medialbicipital sulcus without giving off anybranches.
On the ulnar side of the upper arm, thenerve runs down behind the medial inter-muscular septum, being covered by the me-dial head of the triceps muscle. It crossesthe elbow joint on the extensor side in abony groove, the sulcus for the ulnar nerve,at the medial epicondyle of the humerus.Here, the nerve can be palpated, and thepressure causes an electrifying pain radiat-ing into the ulnar side of the hand. Thenerve then passes between the two heads ofthe ulnar flexor muscle of the wrist to theflexor side of the forearm and runs beneaththis muscle down to the wrist. It does notpass through the carpal tunnel but extendsover the flexor retinaculum to the palm ofthe hand, where it divides into a superficialbranch and a deep branch.
In the forearm, the nerve gives off branches(C1) to the ulnar flexor muscle of the wrist(A2) and to the ulnar half of the deep flexormuscle of the fingers (A3). A sensorybranch, the dorsal branch of the ulnar nerve(BC4), branches off in themiddle of the fore-arm and runs to the ulnar side of the back ofthe hand where it supplies the skin. As forthe rest of the back of the hand, its area ofinnervation overlaps with that of the radialnerve. Another sensory branch, the palmarbranch of the ulnar nerve (BC5), branches offin the distal third of the forearm. It extendsto the palm and supplies the skin of the hy-pothenar eminence.
The superficial branch runs as common palmardigital nerve IV (BC6) toward the interdigitalspace between ring finger and little fingerand divides into the proper palmar digitalnerves (BC7), which supply sensory fibers tothe volar aspects of the little finger and theulnar side of the ring finger and reach to thedistal phalanges on the extensor side ofboth fingers. There is a connection to abranch of the median nerve, called the com-
municating branch of the median nervewith the ulnar nerve (C8).
The deepbranch (AC9) sinks into the depth ofthe palm and curves toward the thenar emi-nence. It gives off branches for allmuscles ofthe hypothenar eminence (C10) (abductormuscle of fifth finger [A11], short flexormuscle of fifth finger [A12], opposingmuscle of fifth finger [A13]), for all dorsaland palmar interosseous muscles (A14), forthe lumbrical muscles III and IV (A15), andfinally, at the thenar eminence, for the ab-ductor muscle of thumb (A16) and the deephead of the short flexor muscle of thumb(A17).
Clinical Note: Injury to the ulnar nerve causesthe formation of a so-called clawhand (D), wherethe fingers are extended in the metacarpo-phalangeal joints but flexed in the proximal anddistal interphalangeal joints. This characteristicposture of the fingers is caused by paralysis of theinterosseous muscles and lumbrical muscles,which flex the phalanges in the metacarpo-phalangeal joints but extend them in the proxi-mal and distal interphalangeal joints. Failure ofthe flexor muscles causes the fingers to remain inthis posture due to the now predominant exten-sor muscles. Since the little finger and the adduc-tors of the thumb are paralyzed, thumb and littlefinger can no longer touch each other.
Innervation of the skin (B). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

79
Spinal
Cord
23
11
121315
1414
16179
54
6
7
106
7
8
5 4
1
9
Brachial Plexus
A Muscles supplied by the ulnar nerve(according to Lanz-Wachsmuth)
B Skin supplied by the ulnar nerve(according to Lanz-Wachsmuth)
C Sequence of branches
D Paralysis of the ulnar nerve(according to Lanz-Wachsmuth)
C 8
T 1
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

80Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Infraclavicular Part (continued)
Medial Fascicle (continued) (A–C)
In addition to the ulnar nerve, the medialfascicle gives rise to the medial cutaneousnerve of the arm and the medial cutaneousnerve of the forearm; both are exclusivelysensory nerves supplying the skin on themedian side of the arm.
Medial cutaneous nerve of the arm (C8 –T1) (A, B). The nerve approaches the ante-rior surface of the upper arm below the axil-lary fossa. Here it ramifies and supplies theskin of themedial aspect between axilla andelbow joint. It reaches to the flexor sidewithits anterior branches and to the extensorside of the upper arm with its posteriorbranches. Frequently, there are anasto-moses to the intercostobrachial nerve.
Medial cutaneous nerve of the forearm(C8 – T1) (A, C). The nerve runs below thefascia on the ulnar side of the forearm andpasses in the lower third through the fasciawith two branches, the anterior branch (AC1)and the ulnar branch (AC2). The anteriorbranch supplies themedial flexor side of theforearm almost up to the midline, and theulnar branch supplies the upper region ofthe medial extensor side almost up to themidline. The area innervated by the medialcutaneous nerve of the forearm extendsslightly to the upper arm and to the hand.
Posterior Fascicle (D, F)
The posterior fascicle gives rise to the axil-lary nerve and the radial nerve.
Axillary nerve (C5 – C6). This runs deep in-side the axilla and across the capsule of theshoulder joint around the surgical neck onthe back of the humerus. It passes throughthe lateral axillary gap and extends beneaththe deltoid muscle to the anterior margin ofthe latter.
Before the nerve trunk passes through thelateral axillary gap, it gives off a motorbranch (DF3) to the lesser teresmuscle (D4),which also passes through the lateral axil-lary gap. At the same level, the superiorlateral cutaneous nerve of the arm (D–F5)
branches off and reaches the skin at the pos-teriormargin of the deltoidmuscle, where itsupplies the skin of the lateral aspects ofshoulder and upper arm. From the nervetrunk extending beneath the deltoid muscleto the front, numerous branches (D6) to thedeltoid muscle (D7) branch off and supplyits various parts.
Clinical Note: As a result of its location on thecapsule of the shoulder joint, the nerve can be in-jured by dislocation of the humerus or byhumeral neck fracture. This causes anesthesia inthe skin area over the deltoid muscle.
Innervation of the skin (B, C, E). Auto-nomic zone (dark blue) and maximum zone(light blue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

81
Spinal
Cord
2
1
1
2
754
36
6 3
5 5
Brachial Plexus
A Sequence of branches of the medialcutaneous nerve of the arm and of themedial cutaneous nerve of the forearm
B Skin supplied by the medialcutaneous nerve of the arm(according to Lanz-Wachsmuth)
D Muscles supplied by the axillarynerve
E Skin supplied by the axillary nerve(according to Lanz-Wachsmuth)
F Sequence of branches
C 8T 1
C 5
C 6
C Skin supplied by the medialcutaneous nerve of theforearm (according toLanz-Wachsmuth)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

82Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Infraclavicular Part (continued)
Posterior Fascicle (continued) (A–D)
Radial nerve (C5–C8) (A–C). The mainnerve of the posterior cord supplies the ex-tensor muscles of upper arm and forearm.
The nerve trunk extends from the axilla intothe proximal third of the medial bicipitalsulcus and then spirals around the dorsalsurface of the humerus, to which it isdirectly apposed in the sulcus of the radialnerve. In the distal third of the upper arm, itpasses to the flexor side between brachialmuscle and brachioradial muscle. In the sul-cus of the radial nerve, the nerve can easilybe injured by pressure or by bone fracturesbecause of its proximity to the bone. Thenerve crosses the elbow joint on the flexorside and divides at the level of the head ofradius into two terminal branches, the su-perficial branch and the deep branch. The su-perficial branch continues in the forearm onthe medial surface of the brachioradialmuscle and then runs in the lower third be-tween brachioradial muscle and radius tothe extensor side in order to reach the backof the hand. The deep branch obliquelypenetrates the supinator muscle, gives offnumerous muscular branches, and extendsas the thin posterior interosseous nerve of theforearm to the wrist.
For the upper arm, the radial nerve gives offthe posterior cutaneous nerve of the arm(A–C1), which supplies a skin area on theextensor side of the upper armwith sensoryfibers, and the inferior lateral cutaneous nerveof the arm (A–C2). In the middle third of theupper arm, it gives off muscular branches(AC3) for the long head, the lateral head, andthe medial head of the triceps muscle (A4).The branch for themedial head gives off alsothe branch for the anconeus muscle (A5).
The posterior cutaneous nerve of the forearm(A–C6) branches off in the region of theupper arm; it supplies a strip of skin on theradial extensor side of the forearm. At thelevel of the lateral epicondyle, muscularbranches (C7) extend to the brachioradialmuscle (A8) and to the long radial extensormuscle of the wrist (A9). The nerve trunk
then ramifies into its two major branches inthe forearm.
At the back of the hand, the superficial branch(A–C10) gives off the dorsal digital nerves(A–C11); they supply sensory fibers to theradial back of the hand, the extensor side ofthe thumb, the proximal phalanges of indexandmiddle fingers, and the radial half of theextensor side of the ring finger. The ulnarcommunicating branch of the radial nerveconnects with the ulnar nerve (C12).
The deep branch (AC13) gives off muscularbranches to the short radial extensor muscleof the wrist (A14) and to the supinatormuscle, while passing through the supina-tor muscle. Thereafter, it gives off motorbranches to the hand extensor muscles,namely, to the common extensor muscle ofthe fingers (A15), the extensormuscle of thelittle finger (A16), the ulnar extensormuscleof the wrist (A17), the long abductor muscleof the thumb (A18), and the short extensormuscle of the thumb (A19). Finally, the ter-minal branch of the deep branch, the poste-rior interosseous nerve, gives off branches tothe long extensor muscle of the thumb(A20) and to the extensor muscle of theindex finger (A21).
The nerve sends sensory branches to theshoulder joint and wrist.
Clinical Note: Injury to the main nerve trunkin the area of the upper arm results in paralysis ofthe extensor muscles. This mainly affects thehand, leading to the so-called wristdrop (D)characteristic for radial paralysis: extension ispossible neither in the wrist nor in the fingers,thus making the hand drop down limply.
Innervation of the skin (B). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

83
Spinal
Cord
1
2
4
6
8
12
3
7
9
15
13
14
18
19
11
10
5
3
17
20
21
16
1
6
2
2
10
11
13
10
11
12
6
Brachial Plexus
A Muscles supplied by the radial nerve(according to Lanz-Wachsmuth)
B Skin supplied by the radial nerve(according to Lanz-Wachsmuth)
C Sequence of branches
D Paralysis of the radialnerve (according toLanz-Wachsmuth)
C 5
C 8T 1
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

84Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Nerves of the Trunk
In the trunk region, the original metamer-ism of the body can still be recognizedthrough the arrangement of the ribs andtheir intercostal muscles. The thoracicnerves, too, fit in well with this segmentalorganization.
Each of the twelve thoracic spinal nervesdivides into a posterior branch (A1) and ananterior branch (A2).
Posterior Branches (A, D)
Each posterior branch divides into a medialand a lateral branch. Both supply motorfibers to the deep autochthonous backmuscles. Sensory innervation of the backcomes mainly from the lateral branches ofthe posterior branches (AD3). The area sup-plied by the posterior branches of cervicalspinal nerves expands widely and includesthe occiput (greater occipital nerve) (D4). Inthe lumbar region, sensory innervation ofthe back comes from the posterior branchesof the lumbar spinal nerves L1–L3 and thesacral spinal nerves S1–S3 (superior clunealnerves [D5] and medial cluneal nerves [D6]).
Anterior Branches (A–D)
The anterior branches of the spinal thoracicnerves run as intercostal nerves between theribs, initially on the inner surface of thethorax and later within the internal inter-costal muscles. We distinguish between anupper group and a lower group of inter-costal nerves.
Thenerves of the upper group (T1–T6) runup to the sternum and supply the intercostalmuscles (C7), the superior and inferior pos-terior serrate muscles, and the transversethoracic muscle. They give off sensorybranches to the skin of the thorax, namely,the lateral cutaneous branches (AD8) at theanterior margin of the anterior serratemuscle, which further divide into anteriorand posterior branches, and the anterior cu-taneous branches (AD9) close to the sternum,which also divide into anterior and poste-rior branches. The lateral and medial cu-
taneous branches of anterior branches 4 – 6,which extend to the area of the mammarygland, are referred to as lateral and medialmammary branches.
The nerves of the lower group (T7–T12),the intercostal segments of which no longerend at the sternum, extend across the costalcartilages up to the white line. They take anincreasingly oblique downward path andsupply the muscles of the abdominal wall(abdominal transverse muscle [C10], exter-nal [C11] and internal [C12] abdominal ob-lique muscles, rectus abdominis muscle[C13] and pyramidal muscle).
Special features. Intercostal nerve 1 partici-pates in forming the brachial plexus andsends only a thin branch to the intercostalspace. Intercostal nerve 2 (and often 3 aswell) gives off its lateral cutaneous branch tothe upper arm (intercostobrachial nerve)(B14), where it connects with themedial cu-taneous nerve of the arm. The last intercostalnerve running beneath the twelfth rib is re-ferred to as the subcostal nerve; it runs ob-liquely downward across the iliac crest.
The inguinal region and hip region receivetheir sensory innervation from the upper-most branches of the lumbar plexus,namely, from the iliohypogastric nerve (D15)(lateral branch and anterior branch), fromthe ilioinguinal nerve (D16), and from thegenitofemoral nerve (genital branch [D17,femoral branch [D18]).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

85
Spinal
Cord
2
14
2
13
8
9
13
3
4
88
9
15
18
17
16
1556
11
10
12
7
Nerves of the Trunk
A Course of a thoracic nerve
B Intercostobrachial nerve
C Muscles supplied by the intercostal nerves D Innervation of the skin of the trunk
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

86Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Lumbosacral Plexus (A)
The lumbosacral plexus is formed by theanterior branches of the lumbar and sacralspinal nerves. Its branches provide sensoryand motor innervation to the lower limb.The branches of L1–L3 and part of L4 formthe lumbar plexus, the roots of which liewithin the psoas muscle. The obturator nerve(A1) and the femoral nerve (A2) originatefrom here, in addition to several shortmuscular branches. The remainder of thefourth lumbar nerve and the L5 nerve join toform the lumbosacral trunk (A3), which thenunites in the small pelvis with sacralbranches 1 – 3 to form the sacral plexus.The sacral branches emerge from the ante-rior sacral foramina of the sacrum and formtogether with the lumbosacral trunk thesacral plexus; the main nerves originatingfrom here are the sciatic nerve (A4) (commonperoneal nerve [A5] and tibial nerve [A6]).
Lumbar Plexus
The lumbar plexus gives off direct shortmuscular branches to the hip muscles,namely, to the greater and lesser psoasmuscles (L1–L5), the lumbar quadratemuscle (T12–L3), and the lumbar inter-costal muscles. The upper nerves of theplexus are still roughly organized in thesame way as the intercostal nerves. To-gether with the subcostal nerve (A7), theyrepresent transitional nerves between theintercostal nerves and the lumbar nerves.
Iliohypogastric Nerve (T12, L1)
The iliohypogastric nerve (A8) initially runson the inside of the lumbar quadrate musclealong the dorsal aspect of the kidney andthen between the abdominal transversemuscle and the internal oblique muscle ofthe abdomen. It participates in the innerva-tion of the broad abdominal muscles. Itgives off two main branches, namely, thelateral cutaneous branch which supplies thelateral hip region, and the anterior cu-taneous branch which penetrates theaponeurosis of the external oblique muscleof the abdomen cranially to the outer ingui-
nal ring and supplies the skin of this regionas well as the pubic region (p. 84, D15; p. 96,C16).
Ilioinguinal Nerve (L1)
The ilioinguinal nerve (A9) runs along the in-guinal ligament and inguinal canal with thespermatic cord up to the scrotum, or withthe round ligament of the uterus up to thegreater lips in the female, respectively. Itparticipates in the innervation of the broadabdominal muscles and supplies sensoryfibers to the skin of the mons pubis and theupper part of the scrotum, or labia majora,respectively (p. 84, D16).
Genitofemoral Nerve (L1, L2)
The genitofemoral nerve (A10) divides al-ready in, or on, the psoas muscle into twobranches, the genital branch and the femoralbranch. The genital branch runs in theabdominal wall along the inguinal ligamentthrough the inguinal canal and reaches thescrotum with the spermatic cord or, in thefemale, the labia majora with the roundligament of the uterus. It innervates thecremaster muscle and supplies sensoryfibers to the skin of the scrotum, or the labiamajora, respectively, and the adjacent skinarea of the thigh (p. 84, D17; p. 96, C15). Thefemoral branch continues to below the in-guinal ligament and becomes subcutaneousin the saphenous hiatus. It supplies the skinof the thigh lateral to the region of the geni-tal branch (p. 84, D18).
A11 Lateral cutaneous nerve of femur(p. 88, A).
A12 Posterior cutaneous nerve of femur(p. 90, D).
A13 Pudendal nerve (p. 96, AB1).A14 Superior gluteal nerve (p. 90, E).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
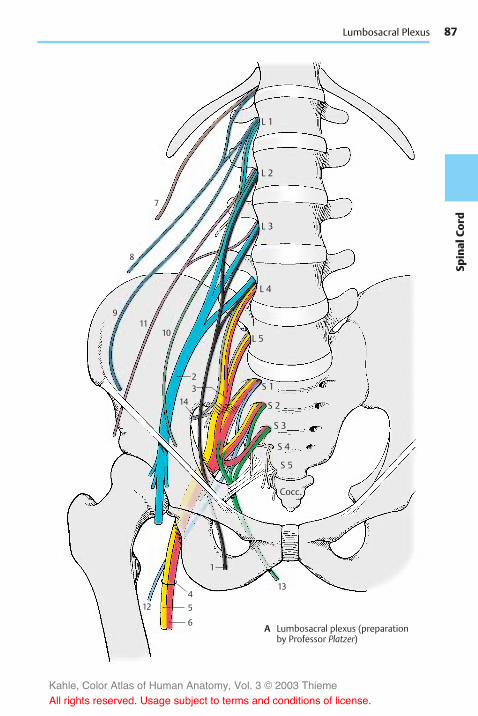
87
Spinal
Cord
7
8
911
10
3
12 5
4
1
6
13
2
14
Lumbosacral Plexus
A Lumbosacral plexus (preparationby Professor Platzer)
L 1
L 2
L 3
L 4
L 5
S 1
S 2
S 3
S 4
S 5
Cocc.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

88Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Lumbar Plexus (continued)
Lateral Cutaneous Nerve of Thigh (L2–L3)(A)
The nerve runs over the iliac muscle tobelow the superior anterior iliac spine. Itthen extends underneath the inguinal liga-ment through the lateral part of the muscu-lar lacuna to the outer aspect of the thighand passes through the fascia lata to theskin. The nerve is exclusively sensory andsupplies the skin of the lateral aspect of thethigh down to the level of the knee.
Femoral Nerve (L1–L4) (B–D)
The nerve runs along the margin of thegreater psoasmuscle up to the inguinal liga-ment and underneath it through themuscu-lar lacuna to the front of the thigh. The nervetrunk divides below the inguinal ligamentinto several branches, namely, a mostlysensory group, the anterior cutaneousbranches (B–D1), a lateral and medial groupof motor branches for the extensor musclesof the thigh, and the saphenous nerve(B–D2). The saphenous nerve extends tothe adductor canal and enters into it. Itpenetrates the vastoadductor membraneand runs along the medial side of the kneejoint and the lower leg together with thegreat saphenous vein down to the medialankle.
In the small pelvis, the femoral nerve givesoff fine branches (D3) to the greater psoasmuscle (B4) and to the iliac muscle (B5).Below the inguinal ligament, a branch (D6)extends to the pectineal muscle (B7). Theanterior cutaneous branches (B–D1) origi-nate slightly more distally, with thestrongest one continuing along the middleof the thigh down to the knee. They supplysensory fibers to the skin of the anterior andmedial aspects of the thigh.
The lateral group of branches (D8) consistsof muscular branches for the sartorius muscle(B9), the rectus femoris muscle (B10), thelateral vastus muscle (B11), and the inter-mediate vastus muscle (B12). The muscularbranch (D13) for the medial vastus muscle(B14) runs along the medial margin of the
sartorius muscle. The muscular branches al-ways ramify into several branches for theproximal and distal portions of the muscles.The muscular branches also give off finesensory branches to the capsule of the kneejoint and the periosteum of the tibia. Fibersfrom the branch for the medial vastusmuscle extend to the femoral artery andfemoral vein.
The saphenous nerve (CD2) is exclusivelysensory. Below the knee joint, it gives off theinfrapatellar branch (B–D15) which sup-plies the skin below the patella. The remain-ing branches, the medial crural cutaneousbranches, supply the skin of the anterior andmedial aspects of the lower leg. The sup-plied area extends on the anterior side overthe edge of the tibia and may reach to thegreat toe along themedial aspect of the foot.
Clinical Note: Injury to the femoral nervemakes it impossible to extend the leg in the kneejoint. Flexion in the hip joint is reduced, and thepatellar tendon reflex is absent.
Innervation of the skin (A, C). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

89
Spinal
Cord
1
2
15
36
131
2
8
15
12
5
1
4
11
14
10
79
2
15
Lumbar Plexus
A Skin supplied by the lateralcutaneous nerve of thigh(according to Lanz-Wachsmuth)
B Muscles supplied by the femoralnerve (according to Lanz-Wachsmuth)
C Skin supplied by the femoralnerve (according to Lanz-Wachsmuth)
D Sequence of branches
L 1
L 2
L 3
L 4
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

90Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Lumbar Plexus (continued) (A–C)
Obturator Nerve (L2–L4)
The nerve providesmotor innervation to theadductor muscles of the thigh. Medial to thegreater psoas muscle, it extends along thelateral wall of the small pelvis down to theobturator canal through which it passes toreach the thigh. It gives off a muscularbranch to the external obturator muscle(AB1) and then divides into a superficialbranch and a deep branch. The superficialbranch (AB2) runs between the long adduc-tor muscle (A3) and short adductor muscle(A4) and innervates both. The nerve alsogives off branches to the pectineal muscleand the gracilis muscle (A5) and finally ter-minates in a cutaneous branch (A–C6) to thedistal region of the medial aspect of thethigh. The deep branch (AB7) runs along theexternal obturatormuscle and then down tothe great adductor muscle (A8).
Clinical Note: Paralysis of the obturator nerve(for example, as a result of pelvic fracture) causesloss of adductor muscle function. This restrictsstanding and walking, and the affected leg can nolonger be crossed over the other leg.
Sacral Plexus (D–F)
The lumbosacral trunk (parts of L4 and L5)and the anterior branches of S1 – S3 join onthe anterior surface of the piriform muscleto form the sacral plexus. Direct branchesextend from the plexus to themuscles of thepelvic region, namely, to the piriformmuscle, the gemellus muscles (F9), the in-ternal obturator muscle, and the quadratemuscle of thigh (F10).
Superior Gluteus Nerve (L4–S1) (E)
The nerve extends across the upper marginof the piriform muscle in dorsal directionthrough the suprapiriform foramen to thegluteus medius (E11) and minimus (E12)muscles and supplies both with motorfibers. The nerve continues between thetwomuscles to the tensor muscle of the fas-cia lata (E13).
Clinical Note: Paralysis of the nerve weakensabduction of the leg. Standing on the affected legand lifting the healthy leg makes the pelvis of theother side drop (Trendelenburg’s symptom).
Inferior Gluteus Nerve (L5–S2) (F)
The nerve leaves the pelvis through the in-frapiriform foramen and gives off severalbranches to the gluteus maximus muscle(F14).
Clinical Note: Paralysis of the nerve weakensextension of the hip joint (for example, whenstanding up or climbing stairs).
Posterior Cutaneous Nerve of Thigh(S1–S3) (D)
The nerve leaves the pelvis together withsciatic nerve and inferior gluteus nervethrough the infrapiriform foramen andreaches below the gluteus maximus muscleto the posterior aspect of the thigh. Locateddirectly beneath the fascia lata, it extendsalong the middle of the thigh into the pop-liteal fossa. This exclusively sensory nervegives off branches to the lower part of thebuttock, the inferior cluneal nerves, and to theperineal region, the perineal branches. It pro-vides sensory innervation to the posterioraspect of the thigh from the lower buttockregion into the popliteal fossa and reachesto the proximal aspect of the lower leg.
Innervation of the skin (C, D). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

91
Spinal
Cord
1
65
8
7
61
72
6
2
4
3
12 9
10
14
11
13
Lumbar Plexus, Sacral Plexus
A Muscles supplied bythe obturator nerve(according to Lanz-Wachsmuth)
B Sequence of branches
C Skin supplied by theobturator nerve (accordingto Lanz-Wachsmuth)
D Skin supplied by theposterior cutaneousnerve of thigh(according toLanz-Wachsmuth)
E Muscles supplied by the superiorgluteal nerve (according to Lanz-Wachsmuth)
F Muscles supplied bythe inferior glutealnerve (according toLanz-Wachsmuth)
L 1
L 2L 3
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

92Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Sacral Plexus (continued)
Sciatic Nerve (L4–S3) (A–C)
The nerve has two components, the commonperoneal nerve (common fibular nerve) andthe tibial nerve; they appear as a uniformnerve trunk (AC1) because they are sur-rounded by a common connective-tissuesheath in the small pelvis and in the thigh.The sciatic nerve leaves the pelvis throughthe infrapiriform foramen and extendsbeneath the gluteus maximus muscle andbicepsmuscle along the posterior aspects ofthe internal obturator muscle, the quadratemuscle of femur, and the great adductormuscle in the direction of the knee joint.Peroneal nerve and tibial nerve separateabove the knee joint. In the pelvis within theconnective tissue sheath, the peronealnerve is on the top and the tibial nervebelow. In the thigh, the peroneal nerve lieslaterally and the tibial nerve medially.However, both may run completely sepa-rately, in which case only the tibial nervepasses through the infrapiriform foramen,while the peroneal nerve penetrates the pir-iform muscle.
Common peroneal nerve (common fibu-lar nerve) (L4 – S2). In the thigh, the per-oneal part (AC2) of the sciatic nerve gives offa muscular branch to the short head of thebiceps muscle of the thigh (A3).
After division of the sciatic nerve, the com-mon peroneal nerve extends along the bi-ceps muscle at the lateral edge of the pop-liteal fossa to the head of the fibula. It thenwinds around the neck of the fibula to theanterior aspect of the lower leg and entersinto the long peroneal (fibular) muscle. Thecommon peroneal nerve divides within thismuscle into the superficial peroneal nerve(AC4) and the deep peroneal nerve (AC5).The superficial peroneal nerve is predomi-nantly sensory and runs between the longperoneal muscle and the fibula to the backof the foot. The deep peroneal nerve is pre-dominantly a motor nerve; it turns towardthe front to the extensor muscles of thelower leg and extends on the lateral surfaceof the anterior tibial muscle to the back ofthe foot.
At the lateral margin of the popliteal fossa,the common peroneal nerve gives off twomain branches for the skin, the lateral suralcutaneous nerve (A–C6), which supplies theskin at the lateral aspect of the lower leg,and the fibular communicating branch (C7),which joins themedial sural cutaneous nerveto form the sural nerve.
The superficial peroneal nerve gives offmuscu-lar branches (AC8) to the long (A9) and short(A10) peroneal muscles. The rest of thenerve is exclusively sensory; it ramifies intoterminal branches, the medial dorsal cu-taneous nerve (BC11) and the intermediatedorsal cutaneous nerve (BC12), which supplythe skin of the back of the foot except for theinterdigital space between great toe andsecond toe.
The deep peroneal nerve gives off severalmuscular branches (AC13) to the extensormuscles of the lower leg and the foot,namely, to the anterior tibial muscle (A14),the long (A15) and short (A16) extensormuscles of toes, and the long (A17) andshort (A18) extensor muscles of the greattoe. The terminal branch is sensory and sup-plies the apposing skin surfaces of the inter-digital space between great toe and secondtoe (B19).
Clinical Note: Injury to the nerve affects theextensor muscles of the foot. The foot can nolonger be lifted in the ankle joint. When walking,the foot hangs down and the toes drag along thefloor. The leg must be lifted higher than normal,resulting in the so-called steppage gait.
Innervation of the skin (B). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

93
Spinal
Cord
14
1315
11
11
12
19
12
6
4
7
6
2
1
5
8
17
1612
13
13
11
18
10
10
9
6
4
3
2
1
5
89
6
Sacral Plexus
A Muscles supplied by the common peronealnerve (according to Lanz-Wachsmuth)
B Skin supplied by thecommon peroneal nerve(according toLanz-Wachsmuth)
C Sequence of branches
L 4L 5S 1S 2
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

94Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Sacral Plexus (continued)
Sciatic Nerve (continued) (A–D)
Tibial nerve (L4–S3). Several motorbranches (AC1) originate from the tibial por-tion of the sciatic nerve, namely, those forthe proximal and distal parts of the semi-tendinous muscle (A2), for the long head ofthe biceps muscle (A3), and a branch divid-ing further for the semimembraneousmuscle (A4) and themedial part of the greatadductor muscle (A5).
After the division of the sciatic nerve, thetibial nerve descends vertically through themiddle of the popliteal fossa and under-neath the gastrocnemius muscle. It then liesunder the tendinous arch of the soleusmuscle and, further distal, between the longflexor muscle of the great toe and the longflexor muscle of toes. It extends betweenthe tendons of both muscles to the back ofthe medial ankle and winds around it.Below the ankle, it divides into two terminalbranches, the medial plantar nerve and thelateral plantar nerve.
The medial sural cutaneous nerve (C6)branches off in the popliteal fossa; it de-scends between the two heads of thegastrocnemius muscle and joins the com-municating branch of the peroneal nerve toform the sural nerve (BC7). The latter extendslaterally from the Achilles tendon behindthe lateral ankle and around it to the lateralaspect of the foot. It gives off the lateral cal-caneal branches (BC8) to the skin of thelateral side of the heel and the lateral dorsalcutaneous nerve (BC9) to the lateral aspect ofthe foot.
Also in the popliteal fossa, motor branches(AC10) go off to the flexor muscles of thelower leg, namely, to the two heads of thegastrocnemius muscle (A11), to the soleusmuscle (A12), to the plantar muscle and thepopliteal muscle (A13). The popliteal branchgives rise to the interosseous nerve of the leg(C14), which runs along the dorsal surface ofthe interosseous membrane and providessensory innervation to the periosteum ofthe tibia, the upper ankle joint, and the tib-iofibular joint. The tibial nerve gives off
muscular branches (C15) to the posteriortibial muscle (A16), the long flexor muscleof toes (A17), and the long flexor muscle ofthe great toe (A18). Before the nerve trunkramifies into terminal branches, it sends offthemedial calcaneal branches (B19) to theme-dial skin area of the heel.
The medial of the two terminal branches,the medial plantar nerve (CD20), innervatesthe abductor muscle of the great toe (D21),the short flexor muscle of toes (D22), andthe short flexor muscle of the great toe(D23). Finally, it divides into the three com-mon plantar digital nerves (BC24), whichsupply lumbrical muscles 1 and 2 (D25) anddivide further into the proper plantar digitalnerves (BC26 ) for the skin of the interdigitalspaces from the great toe up to the fourthtoe.
The second terminal branch, lateral plantarnerve (CD27), divides into a superficial branch(C28) with the common plantar digitalnerves (C29) and proper plantar digitalnerves (BC30) for the skin of the little toearea and into a deep branch (CD31) with themuscular branches for the interosseousmuscles (D32), the adductor muscle of greattoe (D33 ) and the three lateral lumbricalmuscles. D34, short flexor muscle of thelittle toe.
Clinical Note: Injury of the tibial nerve leadsto paralysis of the flexor muscles of toes and foot.The foot can no longer be moved in plantar direc-tion: tiptoeing becomes impossible.
Innervation of the skin (B). Autonomiczone (dark blue) and maximum zone (lightblue).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

95
Spinal
Cord
7
26
24
1919
89
30
32
34
33
2728
22
31
20
21
25
23
5
3
2
4
1
13
17
3
10
14
15
6
1
7
9
8
2720
3124
3026
19
16
12
18
11
12
1110
28
29
Sacral Plexus
A Muscles supplied by the tibial nerve(according to Lanz-Wachsmuth)
B Skin supplied by the tibial nerve(according to Lanz-Wachsmuth)
C Sequence of branches
D Foot muscles supplied by the ti-bial nerve (according to Lanz-Wachsmuth)
L 4L 5S 1S 2S 3
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

96Sp
inal
Cord
Spinal Cord and Spinal Nerves: Peripheral Nerves
Sacral Plexus (continued)
Pudendal Nerve (S2–S4) (A, B)
The pudendal nerve (AB1) leaves the pelvisthrough the infrapiriform foramen (AB2),extends dorsally around the sciatic spine(AB3) and passes through the lesser sciaticforamen (AB4) into the ischioanal fossa. Itthen runs along the lateral wall of the fossawithin the pudendal canal (Alcock’s canal) tobelow the symphysis, sending its terminalbranch to the dorsal side of the penis or cli-toris, respectively.
Numerous branches are given off in the pu-dendal canal; the inferior rectal nerves(A–C5), which may also originate directlyfrom the second to fourth sacral nerves,penetrate through the wall of the canal tothe perineum and supplymotor fibers to theexternal sphinctermuscle of anus (AB6) andsensory fibers to the skin around the anus aswell as the lower two-thirds of the analcanal.
The perineal nerves (AB7) subdivide intodeep and superficial branches. The deepbranches participate in the innervation ofthe external sphincter muscle of the anus.More superficially, they supply the bulbo-cavernous muscle, the ischiocavernousmuscle, and the superficial transverse per-ineal muscle. The superficial branchessupply sensory fibers to the posterior part ofthe scrotum (posterior scrotal nerves) (AC8)in males and to the labia majora (posteriorlabial nerves) (BC9) in females. They alsosupply the mucosa of the urethra and thebulb of penis in males, and the externalurethral opening and the vestibule of vaginain females.
The terminal branch, the dorsal nerve of thepenis (A10) or dorsal nerve of the clitoris (B11),respectively, sends motor branches to thedeep transverse perineal muscle, the deepsphincter muscle, and the sphincter muscleof urethra (B12). After passing through theurogenital diaphragm (AB13), it gives off abranch to the cavernous body of the penis inmales, and to the cavernous body of the cli-toris in females. In males, the nerve runsalong the dorsum of the penis and gives off
sensory branches to the skin of the penisand the glans. In females, it supplies sensoryfibers to the clitoris including the glans.
Muscular Branches (S3, S4)
The levator ani muscle and the coccygealmuscle are supplied directly by nervebranches from the sacral plexus.
Coccygeal Plexus (S4 – Co) (A–C)
The anterior branches of the fourth and fifthsacral nerves form a fine plexus on thecoccygeal muscle, the coccygeal plexus(AB14). The anococcygeal nerves originatefrom here; they supply sensory fibers to theskin over the coccyx and between coccyxand anus (C14).
Sensory Innervation of Pelvis andPerineum (C)
In addition to the sacral and coccygealnerves, the following nerves participate: theilioinguinal nerve and the genitofemoralnerve (C15), the iliohypogastric nerve (C16),the obturator nerve (C17), the posterior cu-taneous nerve of the thigh (C18), the inferiorcluneal nerves (C19), and the medial clunealnerves (C20).
External openings of genitals, bladder, andrectum are border areas between involun-tary smooth intestinal muscles and volun-tary striated muscles. Accordingly, auto-nomic and somatomotor fibers are here in-tertwined. The pudendal nerve contains,apart from sensory, somatomotor, and sym-pathetic fibers, also parasympathetic fibersfrom the sacral spinal cord. Sympatheticfibers also originate as pelvic splanchnicnerves from the second to fourth sacralnerves.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

97
Spinal
Cord
2
4
5
31 14
10
8
137
6 15 15
9 17
18
1620
1914
15
8
2
4
5
31 14
7 4 6
11
913
12
2
4
5
31 14
10
8
137
6 15 15
9 17
18
1620
1914
15
8
2
4
5
31 14
7 4 6
11
913
12
Sacral Plexus
A Pudendal nerve in the male
B Pudendal nerve in the female
C Sensory innervation of the per-ineum (according to Haymakerand Woodhall)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

98Sp
inal
Cord
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Brain Stem andCranial Nerves
Overview 100Cranial Nerve Nuclei 106Medulla Oblongata 108Pons 110Cranial Nerves (V, VII – XII) 112Parasympathetic Ganglia 128Midbrain 132Eye–Muscle Nerves (Cranial Nerves III,
IV, VI) 138Long Pathways 140Reticular Formation 146Histochemistry of the Brain
Stem 148
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

100Br
ainStem
Brain Stem and Cranial Nerves
Overview
The brain stem, or encephalic trunk, is sub-divided into three sections: the medulla ob-longata (elongated spinal cord) (C1), thepons (bridge) (C2), and the mesencephalon(midbrain) (C3).
It is this part of the brain that is underlain bythe chorda dorsalis (notochord) duringembryonic development and from whereten pairs of genuine peripheral nerves(cranial nerves III – XII) emerge. The cerebel-lum, which in ontogenetic terms alsobelongs to it, will be discussed separatelybecause of its special structure (p. 152).
The medulla oblongata between the pyra-midal decussation and the lower border ofthe pons represents the transition from thespinal cord to the brain. The anterior medianfissure, which is interrupted by the pyra-midal decussation (A4), and the antero-lateral sulcus (AD5) on each side extend upto the pons. The anterior funiculi thickenbelow the pons to form the pyramids (A6).Lateral to them on each side bulge the olives(AD7).
The pons forms a broad arching bulge withprominent transverse fibers. Here, de-scending pathways from the brain are re-layed to neurons extending to the cerebel-lum.
The posterior surface of the brain stem iscovered by the cerebellum (C8). Upon itsremoval, the cerebellar peduncles are cutthrough on both sides, namely, the inferiorcerebellar peduncle (or restiform body)(BD9), the middle cerebellar peduncle (orbrachium pontis) (BD10), and the superiorcerebellar peduncle (or brachium conjunc-tivum) (BD11). Removal of the cerebellumopens the fourth ventricle (C12), the tent-shaped roof of which is formed by the supe-rior medullary velum (C13) and the inferiormedullary velum (C14). The floor of thefourth ventricle, the rhomboid fossa, thusbecomes exposed (B). Medulla oblongataand pons together form the hindbrain, alsoknown as rhombencephalon, named after thisfossa. The posterior funiculi (see p. 56)
thicken on both sides to form the tubercle ofthe cuneate nucleus (B15) and the tubercle ofthe gracile nucleus (B16); they are borderedby the posterior median sulcus (B17) and bythe posterolateral sulcus on each side (B18).
The fourth ventricle forms on each side thelateral recess (B19) which opens to the sub-arachnoid space by the lateral aperture(foramen of Luschka) (B20). An unpairedopening lies below the inferior medullaryvelum, the median aperture (foramen ofMagendie) (p. 282, D14). The floor of therhomboid fossa shows bulges near the me-dian sulcus (B21); they are caused by cranialnerve nuclei, namely, the medial eminence(B22), the facial colliculus (B23), the trigonof the hypoglossal nerve (B24), the trigon ofthe vagus nerve (B25), and the vestibulararea (B26). The rhomboid fossa is crossed bymyelinated nerve fibers, the medullarystriae (B27). The pigmented nerve cells ofthe locus ceruleus (B28) shine blueishthrough the floor of the rhomboid fossa.They are mostly noradrenergic and projectinto the hypothalamus, the limbic system,and the neocortex. The locus ceruleus alsocontains peptidergic neurons (enkephalin,neurotensin).
The anterior surface of the midbrain, ormesencephalon, is formed by the cerebralpeduncles (A29) (descending cerebral path-ways). Between them lies the interpeduncu-lar fossa (A30); its floor is perforated bynumerous vessels and is known as the pos-terior perforated substance. At the posteriorsurface of the midbrain lies the tectal plate(or quadrigeminal plate) (BD31) with twoupper elevations, superior colliculi (D32),the relay station of the optic system, andtwo lower elevations, the inferior colliculi(D33), the relay station of the acoustic sys-tem.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

101
BrainStem
29
30
6
7
5
4
32
3331
29
11
910
7
5
3
2
1
13
812
14
31
22 1110
28
21
20 19 15
1617
18
26
24
25
23 9
27
Anatomy of Brain Stem
A Basal view of the brain stem B Dorsal view of the brain stem,rhomboid fossa
C Subdivisions of the brain stem
D Lateral view of the brain stem
III
V
VI
VIIVIII
IXX
XIIXI
C1
IV
III
VIIVIII
VI
IX
XII
C1
IV
V
X
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

102Br
ainStem
Brain Stem and Cranial Nerves: Overview
Longitudinal Organization (A)
The longitudinal organization of the neuraltube (A1) can still be recognized in the brainstem. However, it is modified through thewidening of the central canal into the fourthventricle (A2, A3).
The dorsoventral arrangement of motorbasal plate (A4), visceromotor (A5) andviscerosensory (A6) regions, and sensoryalar plate (A7) changes to amediolateral ar-rangement on the floor of the rhomboidfossa when the neural tube unfolds (A2).The somatomotor zone lies medially, nextcomes the visceromotor zone; theviscerosensory and somatosensory zones aretransposed laterally. The cranial nerve nu-clei in the medulla oblongata are arrangedaccording to this blueprint (A3) (p. 106,p. 108).
Cranial Nerves (B)
According to classical anatomical nomen-clature, there are 12 pairs of cranial nerves,although the first two pairs are not reallyperipheral nerves. The olfactory nerve (I)consists of the olfactory fibers, the bundledprocesses of sensory cells in the olfactoryepithelium which enter the olfactory bulb(B8) (p. 228, A). The optical nerve (II) is acerebral pathway; the origin of the opticalfibers, the retina, together with the pig-mented epithelium of the eyeball repre-sents an evagination of the diencephalon(p. 342, A). Optic chiasm (B9), optical tract(B10).
The eye-muscle nerves (p. 138) are somato-motor nerves. The oculomotor nerve (III)leaves the brain on the floor of the inter-peduncular fossa (B11); the trochlear nerve(IV) emerges at the dorsal surface of themidbrain and extends around the cerebralpeduncles to the basal surface (p. 101, BDIV); the abducens nerve (VI) emerges fromthe lower border of the pons.
Five nerves have developed from thebranchial arch nerves of lower vertebrates:the trigeminal nerve (V) (p. 124), the facialnerve (VII) (p. 122), the glossopharyngeal
nerve (IX) (p. 118), the vagus nerve (X)(p. 114), and the accessory nerve (XI)(p. 112, CD). The muscles supplied by thesenerves are derived from the branchial archmuscles of the foregut. Hence, these nerveshave originally been visceromotor nerves. Inmammals, the branchial arch muscles havechanged into the striated muscles ofpharynx, oral cavity, and face. Unlikegenuine striated muscles, they are notcompletely voluntary (e.g., facial expressionin response to emotion).
The vestibulocochlear nerve (VIII) (p. 120)with its vestibular part represents a phylo-genetically old connection to the organ ofbalance already present in lower vertebrates.
The trigeminal nerve (V) emerges from thelateral part of the pons. Its sensory root ex-tends to the trigeminal ganglion (semilunarganglion, Gasser’s ganglion) (B12); its motorroot (B13) bypasses the ganglion. The facialnerve (VII) and the vestibulocochlear nerve(VIII) leave the medulla oblongata at thecerebellopontine angle. The taste fibers ofthe facial nerve emerge as an independentnerve, the intermediate nerve (B14). Theglossopharyngeal nerve (IX) and the vagusnerve (X) emerge dorsal to the olive. Supe-rior ganglion of the vagus nerve (B15). Thecervical roots of the accessory nerve (XI)unite to form the spinal root (B16). Theupper fibers originating from the medullaoblongata form the cranial root; they run ashort course in the nerve and change over tothe vagus nerve as internal branch (B17).
The hypoglossal nerve (XII) (p. 112, AB) is asomatomotor nerve; in ontogenetic terms,it represents the remnants of several cervi-cal nerves that have become included in thebrain region secondarily and now have onlyrudimentary sensory roots.
B18Olfactory tract.B19 Lateral olfactory stria.B20 Anterior perforated substance.B21Hypophyseal stalk.B22 Choroid plexus (flower spray of Boch-
dalek) (p. 282, D15).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

103
BrainStem
8
18
199
21
20
10
11
17
22
16
15
14
13
12
1 2
4
56
7
3
Longitudinal Organization, Cranial Nerves
A Longitudinal organization of the medulla oblongata (according to Herrick)
B Cranial nerves, base of the brain
I
II
III
IV
V
VI VIIVIII
IX
X
XI
XII
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

104Br
ainStem
Brain Stem and Cranial Nerves: Overview
Base of the Skull (A)
The base of the skull holds the brain. Threebony depressions on each side correspondto the basal aspects of the brain; the basalaspect of the frontal lobe lies in the anteriorcranial fossa (A1), that of the temporal lobe inthe middle cranial fossa (A2), and the basalaspect of the cerebellum in the posteriorcranial fossa (A3). (For the participation ofbones in and the boundaries of the cranialfossae, see vol. 1.) The cranial cavity is linedby a hard meninx, the dura mater (p. 288);its two layers form both a cover for the brainand the periosteum. Embedded betweenthese two layers are large venous sinuses(p. 288). Nerves and blood vessels passthrough numerous foramina in the base ofthe skull (see vol. 1).
On the floor of the anterior cranial fossaclose to the midline, the olfactory nervespass through the openings of the thinlamina cribrosa to the olfactory bulb (A4).The sella turcica rises between the twomiddle cranial fossae; its depression con-tains the hypophysis (A5), which is attachedto the floor of the diencephalon. Lateral tothe sella turcica, the internal carotid artery(A6) passes through the carotid canal intothe cranial cavity. Its S-shaped course by-passes the cavernous sinus (A7). The opticnerve (A8) enters the cranial cavity throughthe optic canal in the medial area of thefossa, while the eye-muscle nerves leave thecavity through the superior orbital fissure(see vol. 1). The paths of the abducens nerve(A9) and the trochlear nerve (A10) are charac-terized by their intradural position. The ab-ducens nerve enters the dura at the middlelevel of the clivus, and the trochlear nerveenters at the edge of the clivus at the attach-ment of the tentorium. The oculomotor nerve(A11) and the trochlear nerve run throughthe lateral wall of the cavernous sinus, andthe abducens nerve through the laterobasalsinus of the internal carotid artery (see vol.2). The trigeminal nerve (A12) reaches belowa dural bridge into the middle cranial fossawhere the trigeminal ganglion (A13) lies in apocket formed by the two dural layers, thetrigeminal cavity. The three trigeminal
branches leave the cranial cavity throughdifferent openings; after passing throughthe wall of the cavernous sinus, the ophthal-mic nerve (A14) extends with its branchesthrough the orbital fissure, the maxillarynerve (A15) through the round foramen, andthe mandibular nerve (A16) through the ovalforamen.
The two posterior cranial fossae surroundthe foramen magnum (A17) to which theclivus (A18) descends steeply from the sellaturcica. The brain stem rests on the clivus,and the cerebellar hemispheres fit into thetwo basal fossae. From the confluence ofsinuses (A19), the transverse sinus (A20) em-braces the posterior cranial fossa and opensinto the internal jugular vein (A21). The facialnerve (A22) and the vestibulocochlear nerve(A23) enter the auditory canal, the internalacoustic meatus. Basal to the meatus, theglossopharyngeal nerve (A24), vagus nerve(A25), and accessory nerve (A26) passthrough the anterior part of the jugularfossa. The fiber bundles of the hypoglossalnerve (A27) pass as a single nerve throughthe hypoglossal canal.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

105
BrainStem
4
810
75
6
14
15
1316 10
11
2425
26
17
9
13
9122223
27
19
20
21
1
18
2
3
Base of the Skull
A Base of the skull, viewedfrom above (preparation byProfessor Platzer)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

106Br
ainStem
Brain Stem and Cranial Nerves
Cranial Nerve Nuclei
As in the spinal cord, where the anteriorhorn represents the area of origin of motorfibers and the posterior horn the area of ter-mination of sensory fibers, the medulla ob-longata contains the nuclei of origin (withthe cell bodies of efferent fibers) and thenuclei of termination (for the axon termi-nals of afferent fibers), the pseudounipolarcells of which lie in sensory ganglia outsidethe brain stem.
The somatomotor nuclei lie close to themidline:
! The nucleus of the hypoglossal nerve (AB1)(tongue muscles)
! The nucleus of the abducens nerve (AB2)! The nucleus of the trochlear nerve (AB3)! The nucleus of the oculomotor nerve (AB4)
(eye muscles)
The visceromotor nuclei follow laterally,namely, the genuine visceromotor nucleibelonging to the parasympathetic nervoussystem and the originally visceromotor nu-clei of the transformed branchial archmuscles. The parasympathetic nuclei in-clude:
! The dorsal nucleus of vagus nerve (AB5)(viscera)
! The inferior salivatory nucleus (AB6) (pre-ganglionic fibers for the parotid gland)
! The superior salivatory nucleus (AB7) (pre-ganglionic fibers for the submandibularand sublingual glands)
! The Edinger–Westphal nucleus (accessorynucleus of oculomotor nerve) (AB8) (pre-ganglionic fibers for the sphincter muscleof pupil and the ciliary muscle)
The series ofmotor nuclei of the branchialarch nerves begins caudally with the spinalnucleus of the accessory nerve (AB9) (shouldermuscles), which extends into the cervicalspinal cord. The series continues craniallywith the ambiguous nucleus (AB10), which isthe motor nucleus of the vagus nerve andthe glossopharyngeal nerve (muscles ofpharynx and larynx), and the nucleus of thefacial nerve (AB11) (facial muscles). The facialnucleus lies deep, as do all motor nuclei of
the branchial arch nerves. Its fibers run in acurve directed dorsally, extend on the floorof the rhomboid fossa (facial colliculus)around the abducens nucleus (internal genuof facial nerve) (A12), and then descendagain to the lower border of the pons wherethey emerge from the medulla oblongata.The most cranial nucleus of the branchialarch nerves is the motor nucleus of thetrigeminal nerve (AB13) (masticatorymuscles).
The sensory nuclei are located laterally;most medially lies the viscerosensory soli-tary nucleus (AB14), in which the sensoryfibers of the vagus nerve and the glos-sopharyngeal nerve, as well as all taste fibers,terminate. Further laterally extends the nu-clear area of the trigeminal nerve, which hasthe largest expanse of all cranial nerves andconsists of:
! The pontine nucleus of the trigeminal nerve(principal sensory nucleus) (AB15)
! The mesencephalic nucleus of the trigeminalnerve (AB16)
! The spinal nucleus of the trigeminal nerve(AB17)
All fibers of the exteroceptive sensibility offace,mouth, and maxillary sinuses terminatein this area.
Finally, most laterally lies the area of the ves-tibular nucleus (B18) and the cochlear nucleus(B19), in which the fibers of the vestibularroot (organ of balance) and the cochlear root(organ of hearing) of the vestibulocochlearnerve terminate.
A20 Red nucleus.A21Olive.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

107
BrainStem
820
4
3
16
13
2
1215111476
5211
10
17
9
8
4
3
11
7
6
10
5
1
9
17
14
18
19
15
16
2
13
Cranial Nerve Nuclei
A Medial view of a median sectionthrough the medulla oblongatashowing the nuclei of cranialnerves (according to Braus andElze)
B Dorsal view of the rhomboid fossashowing the cranial nerve nuclei
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

108Br
ainStem
Brain Stem and Cranial Nerves
Medulla Oblongata
The semi-schematic cross sections show thecellular stain (Nissl) on the left and thecorresponding fiber stain (myelin) on theright.
Cross Section at the Level of theHypoglossal Nerve (A)
The dorsal part, the tegmentum, shows thecranial nerve nuclei, and the ventral partshows the olive (AB1) and the pyramidaltract (AB2).
In the tegmentum, the magnocellular nu-cleus of the hypoglossal nerve (AB3) lies me-dially, and dorsally to it lie the posterior nu-cleus of the vagus nerve (AB4) and the soli-tary nucleus (AB5); the latter nucleus con-tains a large number of peptidergic neurons.The posterior funiculi of the spinal cord ter-minate dorsolaterally in the gracilis nucleus(A6) and in the cuneate nucleus (AB7) wherethe secondary sensory pathway, the mediallemniscus, originates. Ventrally to thecuneate nucleus lies the spinal nucleus of thetrigeminal nerve (AB8). The large cells of theambiguous nucleus (AB9) stand out in thecenter of the field; they lie in the region ofthe reticular formation, of which only theslightly denser lateral reticular nucleus(AB10) can be delimited. The olive (AB1),the fibers of which extend to the cerebellum(p. 164, A11), is accompanied by two nuclei,the posterior accessory olivary nucleus(AB11) and the medial accessory olivary nu-cleus (AB12). Along the ventral aspect of thepyramid stretches the arcuate nucleus(AB13) where collaterals of the pyramidaltract synapse (arcuatocerebellar tract)(p. 164, C18).
The fibers of the hypoglossal nerve (A14)cross the medulla oblongata to reach theirexit point between pyramid and olive. Theposterior longitudinal fasciculus (Schütz’sbundle) (AB15) (p. 144, B) lies dorsally to thehypoglossal nucleus; laterally lies the soli-tary tract (AB16) (p. 114, B12; p. 118, B10)and ventrally the medial longitudinalfasciculus (AB17) (p. 142, A). From the nuclei
of the posterior funiculus, the internal ar-cuate fibers (AB18) radiate broadly into themedial lemniscus (AB19) (p. 140, B). The spi-nal tract of the trigeminal nerve (AB20)(p. 124, B5) runs laterally, and the centraltegmental tract (A21) (extrapyramidalmotor tract, p. 144, A) descends dorsally tothe main olivary nucleus. The fibers of theolivocerebellar tract (AB22) run through thehilum of the olive, while the superficial ar-cuate fibers (AB23) (arcuate nucleus, cere-bellum) run along the lateral part of theolive. The ventral area is occupied by thepyramidal tract (AB2) (p. 140, A).
Cross Section at the Level of theVagus Nerve (B)
The fourth ventricle has become wider. Thesame nuclear columns as in A are found inits floor. Ventrally to the hypoglossal nucleus(AB3) appears Roller’s nucleus (B24) anddorsally the intercalate nucleus (Staderini’snucleus) (B25); the fiber connections ofthese two nuclei are not known. In thelateral field, the nuclei of the posteriorfuniculus disappear and make room for thevestibular nuclei (medial vestibular nucleus)(B26). In the middle of the medulla oblon-gata, the decussating fibers form a seam, theraphe (B27). On both sides of the raphe liesmall groups of cells, the raphe nuclei (B28);their serotoninergic neurons project to thehypothalamus, the olfactory epithelium,and the limbic system. Along the lateralaspect, spinal fibers running to the cerebel-lum aggregate in the inferior cerebellarpeduncle (restiform body) (B29). Afferentand efferent fibers of the vagus nerve (B30)cross the medulla oblongata. Ventrally tothem, the spinothalamic tract (B31) (p. 140,B8) and the spinocerebellar tract (B32)(p. 164, A1; p. 166, B14) ascend along thelateral aspect. The olivocerebellar fibers(B33) (p. 144, A12), which run to the inferiorcerebellar peduncle, aggregate dorsally tothe olive.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

109
BrainStem
6
7
4
17
1516
20
21
1
23
222
18
19
14
3
8
9
1011
1
5
1216
1545
637
8
10
1
8
7
26 25
11
109
8
7
5 4 24
17
1629
1
20
18
19
3
10
1
911
12
13
2
19
18
22
23
26 5 25 24 16
15
29
3020
3132
33
1
2
13
23
213
27
12
28911
223231
34
17
28
27
12
14
17 20
21
13
Medulla Oblongata, Cross Sections
Planes of sections
A Cross section through the medulla oblongataat the level of the hypoglossal nerve (XII)
B Cross section though the medulla oblongataat the level of the vagus nerve (X)
XII
X
BA
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

110Br
ainStem
Brain Stem and Cranial Nerves
Pons
The semi-schematic cross sections show thecellular stain (Nissl) on the left and thecorresponding fiber stain (myelin) on theright.
Cross Section at the Level of theGenu of the Facial Nerve (A)
Beneath the floor of the rhomboid fossa liesthe magnocellular nucleus of the abducensnerve (A1) and, ventrolaterally to it, thenucleus of the facial nerve (A2). Thevisceroefferent superior salivatory nucleus(A3) is seen between the abducens and fa-cial nuclei. The lateral field is occupied bythe sensory terminal nuclei of the vestibularnerve and the trigeminal nerve, namely, themedial vestibular nucleus (Schwalbe’s nu-cleus) (A4), the lateral vestibular nucleus(Deiters’ nucleus) (A5), and the spinal nu-cleus of the trigeminal nerve (A6).
The fibers of the facial nerve bend aroundthe abducens nucleus (A1) and form the fa-cial colliculus (A7). We distinguish an as-cending limb (A8) and, cranial to the il-lustrated section, a descending limb. Theapex is the internal genu of the facial nerve(A9). The fibers of the abducens nerve (A10)descend through the medial field of the teg-mentum. The medial longitudinal fasciculus(AB11) is seen medially and the posteriorlongitudinal nucleus (Schütz’s bundle)(AB12) dorsally to the abducens nucleus.Deep in the pontine tegmentum run thecentral tegmental tract (AB13) and thespinothalamic tract (A14). Secondary fibersof the auditory pathway collect from theanterior cochlear nucleus as a wide fiberbundle, the trapezoid body (AB15); theycross ventral to the medial lemniscus (A16)to the opposite side where they ascend inthe lateral lemniscus (B17). They synapse inpart in the adjacent nuclei of the trapezoidbody, namely, the anterior nucleus of thetrapezoid body (A18) and the posterior nu-cleus of the trapezoid body (superior olive)(AB19). The spinal tract of the trigeminalnerve (A20) lies in the lateral field.
The pontine bulb is formed by the transversepontine fibers (A21). They are corticopon-tine fibers, which synapse in the pontine nu-clei (A22), and pontocerebellar fibers, whichare postsynaptic and extend to the cerebel-lum in the medial cerebellar peduncle(brachium pontis) (A23). Embedded in themiddle of the longitudinally cut fiberbundles is the transversely cut fiber bundleof the pyramidal tract (AB24).
Cross Section at the Level of theTrigeminal Nerve (B)
The medial field of the pontine tegmentumis occupied by the nuclei of the tegmentum.These nuclei, of which only the inferior cen-tral tegmental nucleus (B25) is well defined,belong to the reticular formation. In thelateral field, the trigeminal complex hasreached its widest expansion; lateral is thepontine nucleus of the trigeminal nerve(principal nucleus) (B26), medial to it themotor nucleus of the trigeminal nerve (B27),and dorsal the nucleus of themesencephalictrigeminal root (B28). Afferent and efferentfibers unite to form the strong trunk emerg-ing at the anterior aspect of the pons.
The lateral lemniscus (AB17), the trapezoidbody (AB15), and the adjacent posterior nu-cleus of the trapezoid body (AB19) lie ven-trally to the trigeminal nuclei. The followingascending and descending pathways can berecognized: the posterior longitudinalfasciculus (AB12), the medial longitudinalfasciculus (AB11), and the central tegmentaltract (AB13).
AB29 Pontine tegmentum.AB30 Pontine bulb.
Planes of sections
A
B
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

111
BrainStem
11 10 9 7 12 8
13
16
20
14
15
23
21
24
541326
191822
345
29
30
6 2
191615
111 9
82010
18
21
2926
28
27
19 25
15
11
12
13
17
30
27
25
26
28 11 12
1517
13
19
24
24
Pons, Cross Sections
A Cross section through the pons atthe level of the genu of the facialnerve (VII)
B Cross section through the pons atthe level of the trigeminal nerve(V)
V
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

112Br
ainStem
Brain Stem and Cranial Nerves
Cranial Nerves (V, VII – XII)
Hypoglossal Nerve (A, B)
The twelfth cranial nerve is an exclusivelysomatomotor nerve for the tongue muscles.Its nucleus, the nucleus of the hypoglossalnerve (B1), forms a column of large multi-polar neurons in the floor of the rhomboidfossa (trigon of hypoglossal nerve). It consistsof a number of cell groups, each of which in-nervates a particular muscle of the tongue.The nerve fibers emerge between pyramidand olive and form two bundles that thencombine into a nerve trunk.
The nerve leaves the skull through the canalof the hypoglossal nerve (B2) and descendslaterally to the vagus nerve and the internalcarotid artery. It forms a loop, the arch of thehypoglossal nerve (A3), and reaches the rootof the tongue slightly above the hyoid bonebetween the hypoglossal muscle and themy-lohyoid muscle, where it ramifies into termi-nal branches.
Fiber bundles of the first and second cervi-cal nerves adhere to the hypoglossal nerve.They form the deep cervical ansa (branchesfor the lower hyoid bone muscles) bybranching off again as superior root (A4) andcombining with the inferior root (A5) (sec-ond and third cervical nerve). The cervicalfibers for the geniohyoid muscle (A6) and thethyrohyoid muscle (A7) continue to run inthe hypoglossal nerve. The hypoglossalnerve gives off the lingual branches to thehypoglossal muscle (A8), the genioglossalmuscle (A9), the styloglossal muscle (A10),and to the intrinsic muscles of the body ofthe tongue (A11). Innervation of the tonguemuscles is strictly ipsilateral.
Clinical Note: Injury to the hypoglossal nervecauses hemilateral shrinkage of the tongue (hemi-atrophy). When the tongue is stuck out, it turns tothe affected side because the genioglossal muscle,which moves the tongue to the front, dominateson the healthy side.
Accessory Nerve (C, D)
The eleventh cranial nerve is exclusively amotor nerve; its external branch supplies thesternocleidomastoid muscle (D12) and thetrapeziusmuscle (D13). Its nucleus, the spinalnucleus of the accessory nerve (C14),forms a narrow cell column from C1 to C5 orC6. The large multipolar neurons lie at thelateral aspect of the anterior horn. The cellsof the caudal section supply the trapeziusmuscle, and those of the cranial sectionsupply the sternocleidomastoid muscle. Thenerve fibers emerge from the lateral aspectof the cervical spinal cord between poste-rior root and anterior root and combine toform a bundle that enters the skull as thespinal root (C15) alongside the spinal cordthrough the foramen magnum. Fiberbundles from the caudal part of the ambigu-ous nucleus join the nerve here as cranialroots (C16). Both components pass throughthe jugular foramen (C17). Immediatelyafter passing, the fibers change from theambiguous nucleus as internal branches (C18)over to the vagus nerve (C19). The fibersfrom the cervical spinal cord form the exter-nal branch (C20), which supplies as accessorynerve the sternocleidomastoid muscle andthe trapezius muscle. It passes through thesternocleidomastoid muscle and reachesthe trapezius muscle with its terminalbranches.
Clinical Note: Injury to the accessory nervecauses the head to tilt (plagiocephaly). The armcan no longer be lifted above the horizontal.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

113
BrainStem
1
1
2
9 11
10
3
76
8 4 5
12
13
17
19
18
20
16
15
14
Eleventh and Twelfth Cranial Nerves
A Muscles suppliedby the hypoglossalnerve
B Nuclear region and exitof the hypoglossal nerve
C Nuclear region and exit of theaccessory nerve
D Muscles supplied bythe accessory nerve
C1
C2
C3
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

114Br
ainStem
Brain Stem and Cranial Nerves: Cranial Nerves (V, VII – XII)
Vagus Nerve (A–F)
The tenth cranial nerve not only suppliesareas in the head region like the othercranial nerves, it also descends into thoraxand abdomen where it ramifies in theviscera like a plexus. It is the strongest para-sympathetic nerve of the autonomic nervoussystem and, hence, the most important antag-onist of the sympathetic nervous system(p. 292).
It has the following components:
! Motor fibers (branchial arch muscles)! Exteroceptive sensory fibers! Visceromotor fibers! Viscerosensory fibers! Taste fibers
The fibers emerge directly from behind theolive, unite to form the nerve trunk, andleave the skull through the jugular foramen(B1). In the foramen, the nerve forms the su-perior ganglion of the vagus nerve (jugular gan-glion) (B2) and, after passing through it, themuch larger inferior ganglion of the vagusnerve (nodose ganglion) (B3).
The motor fibers for the branchial archmuscles (AB4) originate from large multi-polar neurons in the ambiguous nucleus(AB5).
The visceromotor fibers (AB6) originate in theparvocellular posterior nucleus of thevagus nerve (AB7), which lies laterally tothe nucleus of the hypoglossal nerve in thefloor of the rhomboid fossa.
The exteroceptive sensory fibers (AB8) origi-nate from neurons in the superior ganglion.They descend with the terminal trigeminalroot (B9) and terminate in the spinal nu-cleus of the trigeminal nerve (AB10).
The cells of the viscerosensory fibers (AB11)lie in the inferior ganglion (nodose ganglion).The fibers run as part of the solitary tract(B12) in caudal direction and terminate atvarious levels of the solitary nucleus(AB13). This nucleus is rich in peptidergicneurons (VIP, corticoliberin, dynorphin).
The taste fibers (AB14) also originate fromcells of the inferior ganglion and terminate
in the cranial part of the solitary nucleus(p. 328, B7).
Head Region (B–D)
In addition to a meningeal branch (sensorysupply to the dura mater in the posteriorcranial fossa), the vagus nerve gives off theauricular branch (B15). The latter branchesoff at the superior ganglion, passes throughthemastoid canaliculus, and reaches the ex-ternal meatus through the tympanomastoidfissure. It supplies the skin of the meatus inthe dorsal and caudal region (D) and a smallarea of the auricula (C) (exteroceptivesensory component of the nerve).
Cervical Region (B, E, F)
Inside a common connective-tissue sheath,the nerve descends in the neck togetherwith the internal carotid artery, the com-mon carotid artery, and the internal jugularvein; it emerges with them through theupper thoracic aperture.
It gives off four branches:
1 The pharyngeal branches (B16) at thelevel of the inferior ganglion. They com-bine in the pharynx with fibers of theglossopharyngeal nerve and the sympa-thetic nervous system to form the pharyn-geal plexus. At the outer surface of themuscles and in the submucosa of thepharynx, the latter forms a meshwork offine fibers with groups of neurons. Thevagal fibers provide sensory innervationto the muscosa of the trachea and mu-cosa of the esophagus including theepiglottis (E, F). The taste buds (E) on theepiglottis are also supplied by the vagusnerve. (Continued p. 116.)
B17 Superior laryngeal nerve (p. 116, A2).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
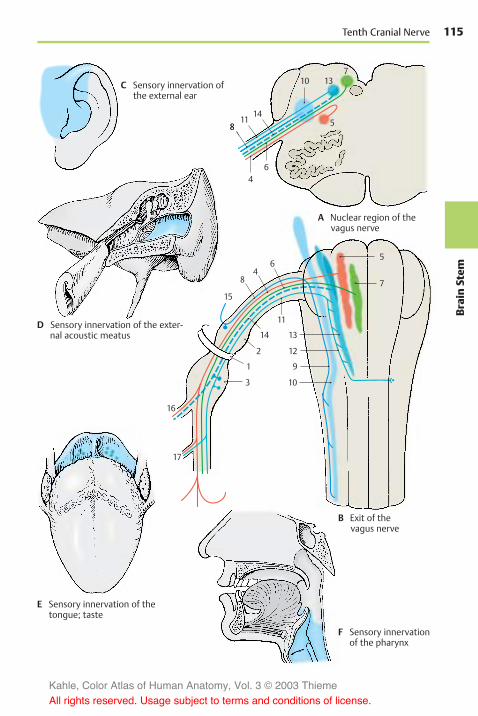
115
BrainStem
15
48
6
11
14
2
1
3
16
17
5
7
13
12
9
10
6
811
14
1310
5
7
4
15
48
6
11
14
2
1
3
16
17
5
7
13
12
9
10
6
811
14
1310
5
7
4
Tenth Cranial Nerve
A Nuclear region of thevagus nerve
B Exit of thevagus nerve
C Sensory innervation ofthe external ear
D Sensory innervation of the exter-nal acoustic meatus
E Sensory innervation of thetongue; taste
F Sensory innervationof the pharynx
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

116Br
ainStem
Brain Stem and Cranial Nerves: Cranial Nerves (V, VII – XII)
Vagus Nerve (continued)
Cervical Region (continued) (A–CEF)
1 The pharyngeal branches (continued).The motor fibers of the vagus nerve in-nervate muscles of the soft palate and thepharynx; these are the muscles of thetonsillar sinus, the levator muscle of thevelum palatinum, and the constrictormuscles of the pharynx (B1).
2 The superior laryngeal nerve (A2) origi-nates below the inferior ganglion (nodoseganglion) and divides at the level of thehyoid bone into an external branch(motor branch for the cricothyroidmuscle) and an internal branch (sensorybranch for the mucosa of the larynx as fardown as the vocal cords).
3 The recurrent laryngeal nerve (A3)branches off in the thorax after the vagusnerve has extended on the left over thearch of the aorta (A4) and on the rightacross the subclavian artery (A5). Itpasses on the left around the aorta andthe ligamentum arteriosum and on theright around the subclavian artery andthen ascends behind the artery. Betweentrachea and esophagus, to which it givesoff the tracheal branches (A6) and theesophageal branches, it extends up to thelarynx. Its terminal branch, the inferiorlaryngeal nerve (A7), supplies motor fibersto all laryngeal muscles except for the cri-cothyroid muscle, and sensory fibers tothe laryngeal mucosa below the vocalcords.
Themotor fibers originate from the ambigu-ous nucleus, the cell groups of which showa somatotopic organization: The fibers ofthe glossopharyngeal nerve arise in thecranial part, those of the superior laryngealnerve further below, and those of the inferiorlaryngeal nerve caudally, whereby the neu-rons for abduction and adduction are ar-ranged one below the other (C).
4 The cervical cardiac branches (pregan-glionic parasympathetic fibers). The supe-rior branches (A8) depart at various levelsand run with the large vessels to theheart, where they terminate in the para-
sympathetic ganglia of the cardiac plexus.One of the branches carries viscero-sensory fibers that transmit informationabout aortic wall tension. Stimulation ofthese fibers causes a fall in blood pres-sure (depressor nerve). The inferior cervi-cal cardiac branches (A9) depart from therecurrent laryngeal nerve or from themain trunk and terminate in the gangliaof the cardiac plexus.
Clinical Note: Injury to the vagus nerve leadsto deficiencies in both pharynx and larynx (F)(see vol. 2). In the case of unilateral paralysis ofthe levator muscle of the velum palatinum (F18),the soft palate and uvula are pulled toward thehealthy side. The vocal cord of the affected side(F19) remains immobilized in the cadaver positionowing to paralysis of the internal laryngealmuscles (recurrent laryngeal paralysis). E showsthe normal positions.
Thoracic and Abdominal Part (A, D)
The vagus nerve loses its identity as a singlenerve; as a visceral nerve, it spreads out likea network. It forms the pulmonary plexus(A10) at the hilum of the lung, which itcrosses dorsally, and the esophageal plexus(A11), from which the anterior vagal trunk(A12) and the posterior vagal trunk (A13) ex-tend to the anterior and posterior aspects ofthe stomach, forming the anterior (A14) andposterior gastric branches. The hepaticbranches (A15) run to the hepatic plexus, theceliac branches (A16) to the celiac plexus,and the renal branches (A17) to the renalplexus.
The preganglionic visceromotor (parasym-pathetic) fibers originate from the posteriornucleus of the vagus nerve, in which a so-matotopic organization of the visceralsupply can be recognized (D).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

117
BrainStem
1
7
2
8
6
53
9
10
43
11
12
16
13
15
17
14
18
19
Tenth Cranial Nerve
A Thoracic and abdominalparts of the vagus nerve(according to Feneis)
B Muscles innervated bythe vagus nerve
C Somatotopic organization ofthe ambiguous nucleus(according to Crosby,Humphrey, and Lauer)
Superior laryn-
geal nucleus
(crico-
thyroid muscle)
Adduction
D Somatotopic organization ofthe posterior nucleus of thevagus nerve(according to Getz and Sienes)
Lung
Abdomen
Heart
Trachea andesophagus
E Velum palatinum andvocal cords (normal)
F Velum palatinum andvocal cords in caseof paralysis of theleft vagus nerve
Inferior laryn-
geal nucleus
Abduction
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

118Br
ainStem
Brain Stem and Cranial Nerves: Cranial Nerves (V, VII – XII)
Glossopharyngeal Nerve (A–E)
The ninth cranial nerve supplies sensoryfibers for themiddle ear, areas of the tongueand of the pharynx, and motor fibers for themuscles of the pharynx. It contains motor,visceromotor (parasympathetic), viscero-sensory, and taste fibers. It emerges fromthe medulla oblongata behind the oliveright above the vagus nerve and leaves theskull together with the vagus nerve throughthe jugular foramen (B1). In the foramen, itforms the superior ganglion (B2) and, afterpassing through it, the larger inferior gan-glion (petrosal ganglion) (B3). Laterally tothe internal carotid artery and the pharynx,it forms an arch to the root of the tonguewhere it ramifies into several terminalbranches.
The motor fibers (AB4) originate from thecranial part of the ambiguous nucleus(AB5), while the visceroefferent fibers(secretory fibers) (AB6) originate from theinferior salivatory nucleus (AB7). The cellsof the viscerosensory fibers (AB8) and the tastefibers (AB9) lie in the inferior ganglion anddescend in the solitary tract (B10) to termi-nate at specific levels of the solitary nu-cleus (AB11).
The first branch, the tympanic nerve (B12),originates from the inferior ganglion withviscerosensory and preganglionic secretoryfibers in the petrosal fossula. It runs throughthe tympanic canaliculus into the tympaniccavity, where it receives fibers from theplexus of the internal carotid artery via thecaroticotympanic nerve and forms the tym-panic plexus. It supplies sensory fibers to themucosa of tympanic cavity and auditory(eustachian) tube (C). The secretory fibersrun as lesser petrosal nerve to the otic gan-glion (p. 130).
Apart from connections with the vagusnerve, facial nerve, and sympatheticnervous system, the inferior ganglion givesoff the (viscerosensory) branch of the carotidsinus (B13), which descends to the bifurca-tion of the common carotid artery and ter-minates in the wall of the carotid sinus (B14)and in the carotid glomus (B15) (see vol. 2).
The nerve transmits impulses of themechanoreceptors of the sinus and thechemoreceptors of the glomus to themedulla oblongata and via collaterals to theposterior nucleus of the vagus nerve (affer-ent limb of the sinus reflex). Preganglionicfibers run from the vagal nucleus to groupsof neurons in the cardiac atria, the axons ofwhich (postganglionic parasympatheticfibers) terminate at the sinoatrial node andthe atrioventricular node (efferent limb ofthe sinus reflex). This system registers andregulates the blood pressure and heart rate.
The nerve also gives off the pharyngealbranches (B16); together with the part of thevagus nerve, they form the pharyngealplexus and participate in the sensory (E)and motor supply of the pharynx. A motorbranch, the stylopharyngeal branch (B17), in-nervates the stylopharyngeal muscle, whilesome sensory tonsillar branches (D18) extendto the tonsils and the soft palate. The nervedivides below the tonsils into the lingualbranches (D19), which supply the posteriorthird of the tongue including the vallatepapillae with sensory as well as taste fibers(D20).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

119
BrainStem
119
8
46
7
5
7
6
9
10
118
1
4
2
312
5
13
151617
19
20
14
18
Ninth Cranial Nerve
A Nuclear region of the glossopharyngeal nerve
B Exit of the glossopharyngeal nerve
C Sensory innervation of the middle ear
D Sensory innervationof the tongue; taste
E Sensory innervation ofthe pharynx
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

120Br
ainStem
Brain Stem and Cranial Nerves: Cranial Nerves (V, VII – XII)
Vestibulocochlear Nerve
The eighth cranial nerve is an afferentnerve consisting of two components, thecochlear root for the organ of hearing and thevestibular root for the organ of balance.
Cochlear Root (A)
The nerve fibers originate from the bipolarneurons of the spiral ganglion (A1), a band ofcells following the spiral course of thecochlea. The peripheral processes of thecells terminate at the hair cells of Corti’sorgan; the central processes form smallbundles that organize into the foraminousspiral tract (A2) and combine in the floor ofthe inner auditory canal, the internal acous-tic meatus, to form the cochlear root (A3). Thelatter extends, together with the vestibularroot (B) inside a common connective-tissuesheath, through the internal acoustic meatusinto the cranial cavity. At the entrance of theeighth cranial nerve into the medulla ob-longata at the cerebellopontine angle, thecochlear component lies dorsally and thevestibular component ventrally.
The cochlear fibers terminate in the ante-rior cochlear nucleus (A4) and in the pos-terior cochlear nucleus (A5). From theanterior nucleus, the fibers cross over to theopposite side (trapezoid body) (A6) (p. 110,AB15); after partly synapsing in the trape-zoid nuclei (B7), they ascend as lateral lem-niscus (A8) (central auditory tract, p. 378).The fibers originating from the posteriorcochlear nucleus cross partly as medullarystriae (posterior acoustic striae) just belowthe rhomboid fossa; they ascend in thelateral lemniscus as well.
Vestibular Root (B)
The nerve fibers originate from bipolar neu-rons of the vestibular ganglion (B9) which liesin the internal acoustic meatus. The periph-eral processes of these cells terminate at thesensory epithelia of the semicircular ducts(B10), the saccule (B11), and the utricle (B12)(p. 377, D). Their central processes unite toform the vestibular root (B13) and terminate,after bifurcation into the ascending and de-
scending branches, in the vestibular nucleiof the medulla oblongata. Only a small por-tion reaches directly into the cerebellum viathe inferior cerebellar peduncle (restiformbody).
The vestibular nuclei lie in the floor of therhomboid fossa below the lateral recess: thesuperior nucleus (Bechterew’s nucleus)(B14), the medial nucleus (Schwalbe’s nu-cleus) (B15), the lateral nucleus (Deiters’nucleus) (B16), and the inferior nucleus(B17). The primary vestibular fibers termi-nate mostly in the medial nucleus. Second-ary fibers run from the vestibular nuclei tothe cerebellum and into the spinal cord (ves-tibulospinal tract) (B18).
The function of the vestibular apparatus playsan important role for balance and uprightposture. The tracts to the cerebellum and thespinal cord serve this purpose. The vesti-bulospinal tract has an effect on musculartension in various parts of the body. The ves-tibular apparatus controls especially move-ments of the head and fixation of visionduring movement (tracts to the eye-musclenuclei) (p. 383, C).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

121
BrainStem
8
6 7
1413
9
1112
1016 15
17
18
14 16 15
54
5
3
2
1
4
Eighth Cranial Nerve
A Vestibulocochlear nerve, nuclear region,and entrance of the cochlear root
B Vestibulocochlear nerve, nuclearregion, and entrance of the vesti-bular root
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

122Br
ainStem
Brain Stem and Cranial Nerves: Cranial Nerves (V, VII – XII)
Facial Nerve (A–F)
The seventh cranial nerve supplies motorfibers to the muscles of facial expression; ina nerve bundle emerging separately fromthe brain stem, called the intermediatenerve, it carries taste fibers and viscero-efferent secretory (parasympathetic) fibers.The motor fibers (AB1) originate from thelarge, multipolar neurons in the nucleus ofthe facial nerve (AB2). They arch aroundthe abducens nucleus (AB3) (internal genu ofthe facial nerve) and emerge on the lateralaspect of the medulla oblongata from thelower border of the pons. The cells of thepreganglionic secretory fibers (AB4) form thesuperior salivatory nucleus (AB5). Thetaste fibers (AB6) originate from the pseudo-unipolar cells in the geniculate ganglion (BC7)and terminate in the cranial section of thesolitary nucleus (AB8). The visceroefferentand taste fibers do not arch around the ab-ducens nucleus but join the ascending limbof the nerve and emerge as intermediatenerve (B9) between the facial nerve and thevestibulocochlear nerve.
Both parts of the nerve pass through theinner auditory canal, the internal acousticmeatus (petrous part of temporal bone, in-ternal acoustic pore, see vol. 1), and enterthe facial canal as a nerve trunk. At the bendof the nerve in the petrous bone (externalgenu of the facial nerve) lies the geniculateganglion (BC7). The canal continues abovethe tympanic cavity (p. 367, A10) and turnscaudally toward the stylomastoid foramen(BC10), through which the nerve leaves theskull. The nerve ramifies into terminalbranches (parotid plexus) (E11) in the parotidgland.
The greater petrosal nerve (BC12), thestapedius nerve (BC13), and the chordatympani (BC14) branch off inside the facialcanal. The greater petrosal nerve (pregan-glionic secretory fibers for the lacrimalgland, nasal glands, and palatal glands)originates from the geniculate ganglion, ex-tends through the hiatus for the lesserpetrosal nerve into the cranial cavity andover the anterior aspect of the petrous bonethrough the foramen lacerum and finally
through the pterygoid canal to the pterygo-palatine ganglion (C15). The stapedius nervesupplies the stapedius muscle in the middleear. The chorda tympani (BC14) branches offabove the stylomastoid foramen, runsbeneath the mucosa through the tympaniccavity (p. 365, A22) and further to thepetrotympanic fissure, and finally joins thelingual nerve (C16). It contains taste fibersfor the anterior two-thirds of the tongue (D)and preganglionic fibers for the subman-dibular and sublingual glands as well asvarious lingual glands.
Before it enters the parotid gland, the facialnerve gives off the posterior auricular nerve(E17) as well as branches to the posteriorbelly of the digastric muscle (CE18) and tothe stylohyoid muscle (C19). The parotidplexus gives off the temporal branches (E20),the zygomatic branches (E21), the buccalbranches (E22), the marginal mandibularbranch (E23), and the cervical branch (E24)for the platysma (see vol. 1). The branchesprovide innervation to all the muscles of fa-cial expression.
Ramifications of the cervical branch lyingbeneath the platysma form the superficialcervical ansa by anastomosing withbranches of the sensory transverse cervicalnerve (p. 72, BC17). The small branchesdeparting from the ansa are mixed sen-sorimotor nerves. The terminal ramifica-tions of temporal branches, buccalbranches, and marginal mandibular branchform similar plexuses with branches of thetrigeminal nerve.
Clinical Note: Injury to the nerve results inatony of all muscles of the affected half of theface. The mouth region drops, and the eye can nolonger close (F). There is increased sensitivity tosound, hyperacusis (p. 366).
C25 Trigeminal ganglion.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

123
BrainStem
7 12 25
1410
18
19
15
16
13
14
6 25
3
8
62 3
5
8
41
12 9
13
1410
7
20
21
22
23
24
18
1117
Seventh Cranial Nerve
A Nuclear region of the facial nerve
B Exit of the facial nerve
C Facial nerve,course withinthe petrous bone
D Tongue area suppliedby taste fibers
E Muscles innervatedby the facial nerve
F Paralysis of theleft facial nerve
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

124Br
ainStem
Brain Stem and Cranial Nerves: Cranial Nerves (V, VII – XII)
Trigeminal Nerve (A-F)
The fifth cranial nerve carries sensoryfibers for the facial skin and mucosa andmotor fibers for the masticatory muscles,for the mylohyoid muscle, the anterior bellyof the digastric muscle, and probably also forthe tensor muscle of the velum palatinumand the tensor muscle of the tympanic mem-brane. It emerges from the pons with a thicksensory root (greater portion) and a thinnermotor root (lesser portion) and then passes tothe front over the petrous bone. Thetrigeminal ganglion (semilunar ganglion,Gasser’s ganglion) lies in a dural pocket, thetrigeminal cave, and gives off three mainbranches, namely, the ophthalmic nerve, themaxillary nerve, and the mandibular nerve(see p. 104, A14 –A16).
The sensory fibers (B1) originate from thepseudounipolar cells in the trigeminal gan-glion (semilunar ganglion, Gasser’s gan-glion) (BE2); the central processes of thesecells terminate in the sensory trigeminalnuclei. Most of the fibers of the epicritic sen-sibility (p. 322) terminate in the pontine nu-cleus of the trigeminal nerve (principal nu-cleus) (AB3), while those of the protopathicsensibility (p. 324) terminate in the spinalnucleus of the trigeminal nerve (BC4).Fibers descend as spinal tract (B5) down tothe upper cervical spinal cord and terminatein a somatotopic arrangement (C): fibers forthe perioral region terminate cranially,those for the adjacent skin areas morecaudally. The fibers for the outermost semi-circle terminate furthest caudally (onionskin arrangement of central sensory inner-vation). The mesencephalic tract (B6) carriesproprioceptive impulses from the mastica-tory muscles.
The mesencephalic nucleus of the trigeminalnerve (AB7) consists of pseudounipolar neu-rons, the processes of which run throughthe trigeminal ganglion without interrup-tion. These are the only sensory fibers forwhich the cells of origin do not lie in a gan-glion outside the CNS but in a nucleus of thebrain stem, so to speak representing asensory ganglion located inside the brain.
The motor fibers originate from large, multi-polar neurons in the motor nucleus of thetrigeminal nerve (AB8).
Innervation of mucosa (D)
The ophthalmic nerve supplies the frontaland sphenoid sinuses as well as the nasalseptum (D9); the maxillary nerve suppliesthe maxillary sinuses, nasal conchae, palate,and gingiva (D10); and the mandibularnerve supplies the lower region of the oralcavity (D11) and the cheeks.
Ophthalmic Nerve (E)
The ophthalmic nerve (E12) gives off a re-current tentorial branch and divides into thelacrimal nerve (E13), the frontal nerve (E14),and the nasociliary nerve (E15). Thesebranches pass through the superior orbitalfissure into the orbit; the nasociliary nerveenters through the medial section of the fis-sure, the two other branches enter throughthe lateral section.
The lacrimal nerve runs to the lacrimal gland(E16) and innervates the skin of the lateralcorner of the eye. Via a communicatingbranch, it receives postganglionic secretory(parasympathetic) fibers from the zygo-matic nerve for innervation of the lacrimalgland.
The frontal nerve divides into the supra-trochlear nerve (E17) (medial corner of theeye) and the supraorbital nerve (E18), whichpasses through the supraorbital notch (con-junctiva, upper eyelid, and the skin of theforehead).
The nasociliary nerve runs to the medialcorner of the eye, which it supplies with itsterminal branch, the infratrochlear nerve(E19). The nasociliary nerve gives off the fol-lowing branches: a communicating branchto the ciliary ganglion (E20), the long ciliarynerves (E21) to the eyeball, the posterior eth-moidal nerve (E22) to the sphenoidal andethmoidal sinuses, and the anterior eth-moidal nerve (E23); the latter runs throughthe anterior ethmoidal foramen to the eth-moidal plate and through the plate into thenasal cavity. Its terminal branch, the exter-nal nasal branch, supplies the skin of thedorsum and the tip of the nose.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

125
BrainStem
73
86
7
2
4
1
3
8
4
5
9 10 9
11
122
23
21 20
17
19
18
16
13 14 22 15
Fifth Cranial Nerve
A Nuclear region of the trigeminal nerve
B Exit of the trigeminal nerve
C Somatotopic organization ofthe spinal nucleus of thetrigeminal nerve (accordingto Dejerine)
D Sensory innervation ofmucosa by the threebranches of thetrigeminal nerve
E Ophthalmic nerve(according to Feneis)
F Skin innervated bythe ophthalmic nerve
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

126Br
ainStem
Brain Stem and Cranial Nerves: Cranial Nerves (V, VII – XII)
Trigeminal Nerve (continued)
Maxillary Nerve (A, B)
The maxillary nerve (A1) gives off a menin-geal branch and then passes through theround foramen (A2) into the pterygopalatinefossa, where it divides into the zygomaticnerve, the ganglionic branches (pterygo-palatine nerves), and the infraorbital nerve.
The zygomatic nerve (A3) reaches through theinferior orbital fissure to the lateral wall ofthe orbit. It gives off a communicatingbranch, which contains postganglionicsecretory (parasympathetic) fibers from thepterygopalatine ganglion for the lacrimalgland, to the lacrimal nerve and divides intothe zygomaticotemporal branch (A4) (temple)and the zygomaticofacial branch (A5) (skinover the zygomatic arch).
The ganglionic branches (A6) are two or threefine filaments running to the pterygo-palatine ganglion (p. 128, A10). The fibersprovide sensory innervation to the upperpharynx, nasal cavity, and hard and soft pal-ates.
The infraorbital nerve (A7) reaches throughthe inferior orbital fissure into the orbit andthrough the infraorbital canal (A8) to thecheek, where it supplies the skin betweenlower eyelid and upper lip (B). It gives offthe posterior superior alveolar nerves (A9)(molar teeth), the middle superior alveolarnerve (A10) (premolar teeth), and the ante-rior superior alveolar nerves (A11) (incisors).The nerves form the superior dental plexusabove the alveoli.
Mandibular Nerve (C–F)
After passing through the oval foramen andgiving off ameningeal branch (C12) in the in-fratemporal fossa, the nerve divides into theauriculotemporal nerve, the lingual nerve,the inferior alveolar nerve, the buccal nerve,and the pure motor branches.
The pure motor branches leave the mandibu-lar nerve shortly after its passage throughthe foramen: the masseteric nerve (C13) forthe masseter muscle (F14), the deep temporalnerves (C15) for the temporal muscle (F16),
and the pterygoid nerves (C17) for the ptery-goid muscles (F18). Motor fibers for the ten-sor tympani muscle and for the tensormuscleof the velum palatinum run to the otic gan-glion (p. 131, AB1) and emerge from it asnerve of the tensor tympanimuscle and as nerveof the tensor veli palatini muscle.
The auriculotemporal nerve (C19) (temporalskin, external acoustic meatus, and tym-panic membrane) usually originates withtwo roots that embrace the middle menin-geal artery and then unite to form the nerve(p. 131, A15). The lingual nerve (C20) de-scends in an arch to the base of the tongueand supplies sensory fibers to the anteriortwo-thirds of the tongue (D). It receives itstaste fibers from the chorda tympani (facialnerve). The inferior alveolar nerve (C21) con-tains motor fibers for the mylohyoid muscleand the anterior belly of the digastricmuscle; furthermore, it contains sensoryfibers, which enter the mandibular canaland give off numerous inferior dentalbranches (C22) for the teeth of the lower jaw.The main branch of the nerve, the mentalnerve (C23), passes through themental fora-men and supplies sensory fibers to the chin,the lower lip, and the skin over the body ofthe mandible (E). The buccal nerve (C24)passes through the buccinator muscle (C25)and supplies the mucosa of the cheek.
BC26 trigeminal ganglion.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

127
BrainStem
8 3 4
12
20
21
13
16
17
25
24
23
22
19
15
7
26
2 1
26
69
1011
5
1814
Fifth Cranial Nerve
A Maxillary nerve(according to Feneis)
B Skin suppliedby the maxillarynerve
C Mandibular nerve(according to Feneis)
D Sensory innervationof the tongue
E Skin supplied by themandibular nerve
F Innervation of muscle
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

128Br
ainStem
Brain Stem and Cranial Nerves
Parasympathetic Ganglia
The fibers of the visceroefferent nuclei(visceromotor and secretory) synapse inparasympathetic ganglia to form postgan-glionic fibers. Apart from the parasympa-thetic root (preganglionic fibers), each gan-glion has a sympathetic root (synapsing inthe ganglia of the sympathetic chain, p. 296)and a sensory root, the fibers of which passthe ganglionwithout interruption. Thus, thebranches leaving the ganglion contain sym-pathetic, parasympathetic, and sensoryfibers.
Ciliary Ganglion (A, B)
The ciliary ganglion (AB1) is a small, flatbody lying laterally to the optical nerve inthe orbit. Its parasympathetic fibers fromthe Edinger–Westphal nucleus run in theoculomotor nerve (AB2) and cross over to theganglion as oculomotor root (AB3) (para-sympathetic root). The preganglionic sym-pathetic fibers originate from the lateralhorn of the spinal cord C8–T2 (ciliospinalcenter) (B4) and synapse in the superior cer-vical ganglion (B5). The postganglionicfibers ascend in the carotid plexus (B6) assympathetic root (B7) to the ciliary ganglion.Sensory fibers originate from the nasociliarynerve (nasociliary root) (AB8).
The short ciliary nerves (AB9) extend from theganglion to the eyeball and penetrate thesclera to enter the interior of the eyeball.Their parasympathetic fibers innervate theciliary muscle (accommodation) and thesphincter pupillae muscle; the sympatheticfibers innervate the dilator pupillae muscle(p. 358).
Clinical Note: The pupil is antagonistically in-nervated by parasympathetic fibers (constrictionof pupil) and sympathetic fibers (dilatation ofpupil). Injury to the ciliospinal center or the spi-nal roots C8, T1 (paralysis of the lower brachialplexus, p. 74) results in ipsilateral constriction ofthe pupil.
Pterygopalatine Ganglion (A, B)
The pterygopalatine ganglion (AB10) lies atthe anterior wall of the pterygopalatinefossa below the maxillary nerve (AB11),which gives off ganglionic branches (pterygo-palatine nerves) (AB12) to the ganglion(sensory root). The parasympatheticsecretory fibers from the superior salivatorynucleus extend in the facial nerve (interme-diate nerve) (AB13) up to the genu of the fa-cial nerve where they branch off as thegreater petrosal nerve (AB14). The nervepasses through the foramen lacerum to thebase of the skull and through the pterygoidcanal to the ganglion (parasympatheticroot). Sympathetic fibers from the carotidplexus form the deep petrosal nerve (AB15)(sympathetic root) and join the greaterpetrosal nerve to form the nerve of the ptery-goid canal (AB16).
The branches leaving the ganglion carrysecretory fibers for the lacrimal gland andfor the glands of the nasal cavity. The para-sympathetic fibers (B17) for the lacrimalgland (AB18) synapse in the ganglion. Thepostganglionic fibers run in the ganglionicbranches (AB12) to the maxillary nerve(AB11) and reach the lacrimal gland via thezygomatic nerve (AB19) and its anastomosis(A20) with the lacrimal nerve (A21).
The remaining parasympathetic secretoryfibers run in the orbital branches (B22) to theposterior ethmoidal cells, in the lateral poste-rior nasal branches (B23) to the nasal con-chae, in the nasopalatine nerve across thenasal septum and through the incisive canal,and in the palatine nerve (AB24) to the hardand soft palates.
The taste fibers (B25) for the soft palate runin the palatine nerves and in the greaterpetrosal nerve.
A26 Trigeminal ganglion.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

129
BrainStem
2
9 1 3
8
17
14
1219
16 25 15
13
6
4
5
18
22
11
7
21 9
1 3 8 2
15
1426
26
1318
20
19 12
102416
11
23 24
10
Ciliary Ganglion, Pterygopalatine Ganglion
A Topographical arrangement ofthe ciliary ganglion and the ptery-gopalatine ganglion
B Pathways of the ciliary ganglionand the pterygopalatine ganglion
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

130Br
ainStem
Brain Stem and Cranial Nerves: Parasympathetic Ganglia
Otic Ganglion (A, B)
The otic ganglion (AB1) is a flat body lyingbelow the oval foramen on the medial sideof the mandibular nerve (A2), from wheresensory and motor fibers (sensorimotorroots) (AB3) enter the ganglion and passthrough without synapsing. The pregan-glionic parasympathetic fibers originatefrom the inferior salivatory nucleus. Theyrun in the glossopharyngeal nerve andbranch off, together with the tympanicnerve, from the inferior ganglion of the glos-sopharyngeal nerve in the petrous fossula tothe tympanic cavity. The fibers leave thetympanic cavity through the hiatus for thelesser petrosal nerve as a fine branch, thelesser petrosal nerve (AB4) (parasympatheticroot). The nerve runs beneath the duramater along the surface of the petrous boneand reaches the otic ganglion after passingthrough the foramen lacerum. The fibers ofthe sympathetic root (AB5) originate fromthe plexus of the middle meningeal artery.
The motor fibers from the motor root of thetrigeminal nerve pass through the ganglionand leave it in the nerve to tensor veli palatini(B6) (soft palate) and in the nerve to tensortympani (B7) (for the muscle that tightensthe tympanic membrane). Motor fibers (B8)for the levator veli palatini from the facialnerve (VII) are thought to run in the chordatympani (AB9) and cross over into the gan-glion via the communicating branch withchorda tympani (AB10). They pass throughwithout synapsing and enter via a com-municating branch (A11) the greaterpetrosal nerve (A12), in which they reachthe pterygopalatine ganglion (A13). Theypass to the palate in the palatine nerves(A14).
The postganglionic secretory (parasympa-thetic) fibers together with sympatheticfibers enter the auriculotemporal nerve(AB15) via a communicating branch andfrom here into the facial nerve (AB16) viaanother anastomosis. The fibers then ramifyin the parotid gland (AB17) together withbranches of the facial nerve. Apart from theparotid gland, they supply the buccal andlabial glands via the buccal nerve and the in-ferior alveolar nerve.
Submandibular Ganglion (A, B)
The submandibular ganglion (AB18) lies to-gether with several small secondary gangliain the floor of the mouth above the subman-dibular gland (AB19) and below the lingualnerve (AB20), to which it connects viaseveral ganglionic branches. Its pregan-glionic parasympathetic fibers (B21) origi-nate from the superior salivatory nucleus,run in the facial nerve (intermediate nerve),and leave the nerve together with the tastefibers (B22) in the chorda tympani (AB9). Inthe latter, the fibers reach the lingual nerve(AB20) and extend in it to the floor of themouth where they cross over into the gan-glion (AB18). Postganglionic sympatheticfibers from the plexus of the external carotidartery reach the ganglion via the sympa-thetic branch (B23) given off by the plexus ofthe facial artery; they pass through the gan-glion without synapsing.
The postganglionic parasympathetic andsympathetic fibers pass partly in the glan-dular branches to the submandibular gland,partly in the lingual nerve to the sublingualgland (AB24) and to the glands in the distaltwo-thirds of the tongue.
A25 Ciliary ganglion.A26 Trigeminal ganglion.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

131
BrainStem
79
515
16
23
17
1924
18
4
3
18
6
21
20
1022
154
12
2625
13
14
113
2
9
10
16
1720
24
18
19
15
Otic Ganglion, Submandibular Ganglion
A Topographical arrangement ofthe otic ganglion and the sub-mandibular ganglion
B Pathways of the otic ganglion andthe submandibular ganglion
VII
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

132Br
ainStem
Brain Stem and Cranial Nerves
Midbrain
Structure (A–C)
Apart from certain modifications in themedulla oblongata (A1), pons (A2), andmesencephalon (A3), the brain stem has auniform structure. The phylogenetically oldpart of the brain stem, which is common toall three parts and contains the cranialnerve nuclei, is the tegmentum (A4). At thelevel of themedulla oblongata and the pons,it is overlain by the cerebellum and in themesencephalon by the tectum (quadrigemi-nal plate) (A5). The ventral part of the brainstem mainly contains the large tracts de-scending from the telencephalon; they formthe pyramids (A6) in the medulla oblongata,the pontine bulb (A7) in the pons, and thecerebral peduncles (A8) in the mesen-cephalon.
The ventricular system undergoes consider-able narrowing in the midbrain, the aque-duct of the mesencephalon (cerebral aqueduct,aqueduct of Sylvius) (A–D9). Duringdevelopment, the lumen of the neural tubebecomes increasingly narrowed as the teg-mentum of the midbrain increases involume (B), while the blueprint of theneural tube survives. The motor derivativesof the basal plate lie ventrally: the nucleus ofthe oculomotor nerve (BC10), the trochlearnucleus (eye muscles), the red nucleus (C11),and the substantia nigra (C12) (consisting ofan outer reticular part and an inner compactpart). The sensory derivatives of the alarplate lie dorsally: the tectum of the mesen-cephalon (quadrigeminal plate) (C13) (syn-aptic relay station for auditory and visualpathways).
Cross Section Through the InferiorColliculi of the Midbrain (D)
The inferior colliculus with its nucleus (nu-cleus of inferior colliculus) (D14) (synapticrelay station of the central auditory path-way) is seen dorsally. The transitional re-gion between pons and cerebral pedunclesand the most caudal cell groups of the sub-stantia nigra (D15) lie ventrally. The magno-cellular nucleus of the trochlear nerve (D16)
is clearly visible in the center of the teg-mentum below the aqueduct, and thelateroposterior tegmental nucleus (D17) issituated dorsal to it. Further lateral are thecells of the locus ceruleus (D18) (the pontinerespiratory center reaching into the mid-brain; it contains noradrenergic neurons,p. 100, B28). The relatively large, scatteredcells dorsal to the locus ceruleus form themesencephalic nucleus of the trigeminalnerve (D19). The lateral field is occupied bythe pedunculopontine tegmental nucleus(D20). At the ventral margin of the tegmen-tum lies the interpeduncular nucleus (D21),which is rich in peptidergic neurons (pre-dominantly enkephalin). The habenulo in-terpeduncular tract (fasciculus retroflexus,Meynert’s bundle) (p. 176, A11), which de-scends from the habenular nucleus, termi-nates here.
The lateral lemniscus (D22) radiates ven-trally into the nucleus of the inferior col-liculus (D14) (p. 378, A5). The fibers of thepeduncle of the inferior colliculus (D23)aggregate at the lateral aspect and run to themedial geniculate body (central auditorypathway, p. 378). In the medial field lie themedial longitudinal fasciculus (D24) (p. 142)and the decussation of the superior cerebellarpeduncle (D25) (p. 166, B5). The fiber plateof the medial lemniscus (D26) (p. 140, B) lieslaterally. The fibers of the cerebral peduncle(D27) are cut transversely and are inter-spersed with a few pontine fibers runningacross.
D28 Periaqueductal gray substance, centralgray.
C29 Edinger–Westphal nucleus (accessorynucleus of the oculomotor nerve).
Plane of section
D
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

133
BrainStem
4
16 7 2 3
4 4
5
139
29
8
9
9 11
12
10
10
14
28
19 9
18
201716
15
19
20
18
16
21
17
9
24
22
25 26
27
26
22
23
24 25
21
14
4
16 7 2 3
4 4
5
139
29
8
9
9 11
12
10
10
14
28
19 9
18
201716
15
19
20
18
16
21
17
9
24
22
25 26
27
26
22
23
24 25
21
14
Structure of the Midbrain, Inferior Colliculi
A Structure of medulla oblongata, pons, and midbrain
B Development of the midbrain C Organization of the midbraininto basal and alar plates
D Cross section through the midbrain atthe level of the inferior colliculi, cellularstaining (Nissl) and fiber staining (myelin)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

134Br
ainStem
Brain Stem and Cranial Nerves: Midbrain
Cross Section Through the SuperiorColliculi of the Midbrain (A)
The two superior colliculi (A1) are seen dor-sally. In lower vertebrates, they representthe most important visual center and con-sist of several layers of cells and fibers. Inhumans, they are only a relay station for re-flex movements of the eyes and pupillaryreflexes, and their stratification is rudimen-tary. In the superficial gray layer (A2) termi-nate the fibers from the occipital fields ofthe cortex (corticotectal tract) (A3). Theoptic layer (A4), which in lower vertebratesconsists of fibers of the optical tract, isformed in humans by fibers from the lateralgenicular body. The deeper layers of cellsand fibers are collectively known as stratumlemnisci (A5). Here terminate the spinotec-tal tract (p. 56, A5), fibers of the medial andlateral lemnisci, and fiber bundles of the in-ferior colliculi.
The aqueduct is surrounded by the peri-aqueductal gray, or central gray (AB6). Itcontains a large number of peptidergic neu-rons (VIP, enkephalin, cholecystokinin, andothers). The mesencephalic nucleus of thetrigeminal nerve (A7) lies laterally to it, andventrally to it lie the nucleus of the oculomotornerve (A8) and the Edinger-Westphal nucleus(accessory oculomotor nucleus) (A9)(p. 138, AD19). Dorsally to both nuclei runsthe posterior longitudinal fasciculus(Schütz’s bundle) (p. 144, B) and ventrally tothem the medial longitudinal fasciculus(A10) (p. 142). The main nucleus of the teg-mentum is the red nucleus (AB11) (p. 136;p. 148, A2); it is delimited by a capsule con-sisting of afferent and efferent fibers(among others, the dentatorubral fasciculus)(A12). At its medial margin descend fiberbundles of the oculomotor nerve (A13) inventral direction. Tectospinal fibers (pupil-lary reflex) and tectorubral fibers cross themidline in the superior tegmental decussa-tion (Meynert’s decussation) (A14) and teg-mentospinal fibers in the inferior decussa-tion (Forel’s decussation) (A15). The lateralfield is occupied by the medial lemniscus(AB16) (p. 140, B).
Ventrally to the tegmentum border the sub-stantia nigra (pars compacta [A17] and parsreticulata [A18], p.136; p. 148, A1). The ven-tral aspect on both sides is formed by thecorticofugal fiber masses of the cerebralpeduncles (AB19). The dorsal aspect isformed by themedial genicular body (AB20).
Cross Section Through the PretectalRegion of the Midbrain (B)
The pretectal region (B21) situated orally tothe superior colliculi represents the tran-sition from the midbrain to the dien-cephalon. Hence, the cross section alreadycontains structures of the diencephalon:dorsally on each side lies the pulvinar (B22),in the middle the epithalamic commissure(B23), and ventrally the mamillary bodies(B24). The pretectal region extends dor-solaterally with the principal pretectal nucleus(B25). The latter is an important relay sta-tion for the pupillary reflex (p. 358, A2). Thefibers of the optical tract and the fibers ofthe occipital cortical fields terminate here.An efferent pathway of the nucleus extendsacross the epithalamic commissure to theEdinger–Westphal nucleus (accessoryoculomotor nucleus). Ventral to the aque-duct are the Darkshevich’s nucleus (B26) andthe interstitial nucleus (of Cajal) (B27), therelay stations in the system of the mediallongitudinal fascicle (p. 142, A8, A9).
Animal experiments have shown that theinterstitial nucleus of Cajal and the prestitialnucleus situated further orally are impor-tant relay stations for automatic motions(p. 192, B) within the extrapyramidal motorsystem (p. 310). The essential synapses forthe rotation of the body around its longi-tudinal axis lie in the interstitial nucleus,those for raising the head and upper body inthe prestitial nucleus.
B28 Supramamillary commissure.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

135
BrainStem
6 9 10
43 16
25
78
20
17
18
24
5
20
1718
11
7 89 16
4 3
12
19
14
11
1513
12 19
13
1415
1
22
25
2627
2025
2726
28
16
23
11
21
1924
1623
6
6 9 10
43 16
25
78
20
17
18
24
5
20
1718
11
7 89 16
4 3
12
19
14
11
1513
12 19
13
1415
1
22
25
2627
2025
2726
28
16
23
11
21
1924
1623
6
6 9 10
43 16
25
78
20
17
18
24
5
20
1718
11
7 89 16
4 3
12
19
14
11
1513
12 19
13
1415
1
22
25
2627
2025
2726
28
16
23
11
21
1924
1623
6
Superior Colliculi, Pretectal Region
Planes of sections
A Cross section through the mid-brain at the level of the superiorcolliculi
B Cross section though the mid-brain at the level of the pretectalregion
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

136Br
ainStem
Brain Stem and Cranial Nerves: Midbrain
Red Nucleus and Substantia Nigra
Lateral View of the Brain Stem (A)
The two large nuclei extend far toward thediencephalon. The substantia nigra (AB1)reaches from the oral part of the pons (A2)to the pallidum (AB3) in the diencephalon.Both nuclei are important relay stations ofthe extrapyramidal system (p. 310).
Red Nucleus (A, B)
The nucleus (AB4) appears reddish in afresh brain section (high iron content,p. 148, A). It consists of the parvocellularneorubrum and the magnocellular paleoru-brum situated ventrocaudally.
Afferent connections
! The dentatorubral fasciculus (B5) of thedentate nucleus (B6) of the cerebellumruns in the superior cerebellar peduncleand terminates in the contralateral rednucleus.
! The tectorubral tract (B7) of the superiorcolliculus terminates in the ipsilateraland contralateral paleorubrum.
! The pallidorubral tract (B8) consists ofpallidotegmental bundles from the innersegment of the pallidum.
! The corticorubral tract (B9) from the fron-tal and precentral cortex terminates inthe ipsilateral red nucleus.
Efferent connections
! The rubroreticular and rubro-olivaryfibers (B10) run in the central tegmentaltract (p. 144, A) and terminate primarilyin the olive (neuronal circuit: dentate nu-cleus – red nucleus – olive – cerebellum).
! The rubrospinal tract (B11) (poorlydeveloped in humans) crosses in Forel’stegmental decussation and terminates inthe cervical spinal cord.
Functional significance. The red nucleus isa relay and control station for cerebellar,pallidal, and corticomotor impulses that areimportant formuscle tone, posture, and loco-motion. Injury to this nucleus causes passivetremor (shaking), changes in muscle tone,and choreic-athetoid hyperactivity.
Substantia Nigra (A–C)
This consists of the dark compact part (nervecells with black melanin pigment) (C) andthe reticular part (of reddish color and rich iniron). The tracts of the substantia nigra formonly loose pathways of fine fibers ratherthan compact bundles.
Afferent connections terminating in theanterior part
! Fibers of the caudate nucleus, strionigralfasciculus (B12)
! Fibers of the frontal cortex (areas 9 to 12),corticonigral fibers (B13)
Afferent connections terminating in thecaudal part
! Fibers of the putamen (B14)! Fibers of the precentral cortex (areas 4
and 6) (B15)
Efferent connections
! Nigrostriatal fibers (B16), running from thecompact part to the striatum
! Fibers of the reticular part, running to thethalamus
The majority of efferent fibers ascend to thestriatum, to which the substantia nigra isclosely connected functionally by the nigro-striatal system. In the axons of the dopaminer-gic nigral neurons (compact part), dopamineis transported to the striatum, where it is re-leased by the axon terminals. There is atopographical relationship between thesubstantia nigra and the striatum (caudatenucleus and putamen); cranial and caudalsegments of the substantia nigra are con-nected with the corresponding segments ofcaudate nucleus and putamen. Caudate nu-cleus and putamen are under the control ofmassive input from totally different neocor-tical zones (B17).
Functional significance. The substantianigra is of special importance for the controlof involuntary coordinated movement andthe rapid onset of movement (starter func-tion). Injury causes muscle stiffness, passivetremor, and loss of coordinated movementand facial expression (masklike expression ofthe face).
B18 Posterior thalamus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

137
BrainStem
34
2
1517
13
12
14
9
18
8
7
4
5 6
16
1
10
11
3
1
34
2
1517
13
12
14
9
18
8
7
4
5 6
16
1
10
11
3
1
Red Nucleus, Substantia Nigra
A Positions of red nucleus andsubstantia nigra in the brainstem, lateral view
B Connecting tracts of red nucleusand substantia nigra
C Melanin-containing pigment cellsof the substantia nigra
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

138Br
ainStem
Brain Stem and Cranial Nerves
Eye-Muscle Nerves (CranialNerves III, IV, and VI)
Abducens Nerve (C, E)
The sixth cranial nerve (C1) is an exclu-sively somatomotor nerve, which innervatesthe lateral rectusmuscle (E2) of the extra-ocu-lar muscles. Its fibers originate from thelarge, multipolar neurons of the nucleus ofthe abducens nerve (C3), which lies in thepons in the floor of the rhomboid fossa(p. 110, A1). The fibers exit at the basal mar-gin of the pons above the pyramid. Aftertaking a long intradural course, the nervepasses through the cavernous sinus andleaves the cranial cavity through the supe-rior orbital fissure.
Trochlear Nerve (B, C, E)
The fourth cranial nerve (BC4) is an exclu-sively somatomotor nerve and innervatesthe superior oblique muscle (E5) of the extra-ocular muscles. Its fibers originate from thelarge, multipolar neurons of the nucleus ofthe trochlear nerve (BC6) (p. 132, D16), whichlies in the midbrain below the aqueduct atthe level of the inferior colliculi. The fibersascend dorsally in an arch, cross above theaqueduct, and leave the midbrain at thelower margin of the inferior colliculi. Thenerve is the only cranial nerve leaving thebrain stem at its dorsal aspect. It descends inthe subarachnoid space (p. 289, A13) to thebase of the skull, where it enters the duramater at the margin of the tentorium andcontinues through the lateral wall of thecavernous sinus. It enters the orbit throughthe superior orbital fissure.
Oculomotor Nerve (A, C–F)
The third cranial nerve (AC7) contains so-matomotor and visceromotor (parasympa-thetic) (A8) fibers. It innervates the remain-ing outer eye muscles (E) and, with itsvisceromotor portion, the intra-ocularmuscles. The fibers exit from the floor of theinterpeduncular fossa at the medial marginof the cerebral peduncle in the oculomotor
sulcus. Laterally to the sella turcica, theypenetrate the dura mater, run through theroof and then through the lateral wall of thecavernous sinus, and enter the orbitthrough the superior orbital fissure. Here,the nerve divides into a superior branch,which supplies the levator muscle of theupper eyelid and the superior rector muscle(E9), and into an inferior branch, which sup-plies the inferior rector muscle (E10), theme-dial rector muscle (E11), and the inferior ob-lique muscle (E12).
The somatomotor fibers originate fromlargemultipolar neurons of the nucleus of theoculomotor nerve (AC13) (p. 134, A8), whichlies in the midbrain below the aqueduct atthe level of the superior colliculi.
The longitudinally arranged cell groups in-nervate specific muscles. The neurons forthe inferior rector muscle (D14) lie dor-solaterally, those for the superior rectormuscle (D15) dorsomedially; below themlie the neurons for the inferior obliquemuscle (D16), those for the medial rectormuscle (D17) ventrally, and those for the le-vator muscle of the upper eyelids (caudalcentral oculomotor nucleus) (D18) dorso-caudally. In the middle third between thetwo paired main nuclei there usually lies anunpaired cell group, Perlia’s nucleus, whichis thought to be associated with ocular con-vergence (p. 358, C).
The preganglionic visceromotor (parasym-pathetic) fibers originate from the parvo-cellular Edinger–Westphal nucleus, the acces-sory oculomotor nucleus (ACD19). They runfrom the oculomotor nucleus to the ciliaryganglion where they synapse. The postgan-glionic fibers enter through the sclera intothe eyeball and innervate the ciliary muscle(F20) and the sphincter pupillae muscle(F21) (p. 358, A, B).
(For extra-ocular muscles, see p. 340.)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

139
BrainStem
1913
78
4
6
7
136
19
4
1 3
19
18
16
17
15
15 19 14
1716
14
14
15
18
20
21
9
2
10
5
11
12
Third, Fourth, and Sixth Cranial Nerves
A–C Nuclear regions and exits of abducens nerve,trochlear nerve, and oculomotor nerve
D Somatotopic arrangement of neurons inthe oculomotor nucleus (accordingto Warwick)
E Extra-ocular muscles
F Intra-ocular muscles
A
B C
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

140Br
ainStem
Brain Stem and Cranial Nerves
Long Pathways
Corticospinal Tract andCorticonuclear Fibers (A)
The pyramidal tract, or corticospinal tract(p. 58, A; p. 308), runs through the basal partof the brain stem and forms the pyramids inthe medulla oblongata (p. 100, A6).
Some of the pyramidal tract fibers termi-nate in the motor nuclei of cranial nerves(corticonuclear fibers):
! Bilaterally in the oculomotor nucleus (III),in the motor nucleus of the trigeminalnerve (V), in the caudal part of the facialnucleus (VII) (forehead muscles), and inthe ambiguous nucleus (X)
! After crossing to the contralateral nu-cleus: in the abducens nucleus (VI), in therostral part of the facial nucleus (VII) (fa-cial muscles with the exception of theforehead muscles), and in the hypoglos-sal nucleus (XII)
! Uncrossed in the ipsilateral trochlear nu-cleus (IV)
Clinical Note: In central facial paralysis, paral-ysis of facial muscles is caused by injury to thecorticobulbar fibers, yet the mobility of the bi-laterally innervated forehead muscles is retained.
Aberrant fibers (Déjérine) (A1). At variouslevels of the midbrain and the pons, finefiber bundles branch off from the corticonu-clear fibers and unite to form the mesen-cephalic aberrant tract and the pontine aber-rant tract. Both descend in the medial lem-niscus (A2) and terminate in the con-tralateral abducens nucleus (VI) and hypo-glossal nucleus (XII), in the two ambiguousnuclei (X), and in the spinal accessory nu-cleus (XI).
Medial Lemniscus (B)
This fiber system includes the most impor-tant ascending pathways of the exteroceptivesensibility from the spinal cord and the brainstem. It is subdivided into the spinal lemnis-cus and the trigeminal lemniscus. The spinallemniscus contains the sensory pathways
for trunk and limbs (bulbothalamic tract,spinothalamic tract, spinotectal tract),while the trigeminal lemniscus contains thesensory pathways for the face (anteriortegmental fasciculus).
1. Bulbothalamic tract (B3). The fibers rep-resent the extension of the posteriorfuniculi of the spinal cord (B4) (epicritic sen-sibility). They originate in the gracile nu-cleus (B5) and the cuneate nucleus (B6),cross as arcuate fibers (decussation of lem-nisci) (B7), and form the medial lemniscusin the narrower sense (p. 108, A19). Thecuneate fibers initially lie dorsally to thegracile fibers, while they lie medially tothem in the pons and midbrain. They termi-nate in the thalamus.
2. Spinothalamic tract (lateral and ante-rior) (B8). The fibers (protopathic sensibility,pain, temperature, coarse tactile sensation)have already crossed to the contralateralside at various levels of the spinal cord andform slightly spread, loose bundles (spinallemniscus) in the medulla oblongata. Theyjoin the medial lemniscus (p. 133, D26;p. 135, A16) in the midbrain.
3. Spinotectal tract (B9). The fibers runtogether with those of the lateralspinothalamic tract. They form the lateraltip of the lemniscus in themidbrain and ter-minate in the superior colliculi (pupillary re-flex on sensation of pain).
4. Anterior tegmental fasciculus (Spitzer)(B10). The fibers (protopathic and epicriticsensibilities of the face) cross in smallbundles from the spinal nucleus of thetrigeminal nerve (principle nucleus) to thecontralateral side (trigeminal lemniscus)and join themedial lemniscus at the level ofthe pons. They terminate in the thalamus.
5. Secondary taste fibers (B11). These origi-nate from the rostral part of the solitary nu-cleus (B12), probably cross to the con-tralateral side, and occupy the medial mar-gin of the lemniscus. They terminate in thethalamus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

141
BrainStem
2
11
10
12
4
5
6
7
3
8
10 9
3
11
8
1
Pyramidal Tract System, Medial Lemniscus
B Ascending pathwaysof the medial lemniscus
A Pyramidal system: corticospinaltract and corticonuclear fibers
III III
V
IV IV
V
VI
VII VII
X X
XII
XI XI
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

142Br
ainStem
Brain Stem and Cranial Nerves: Long Pathways
Medial Longitudinal Fasciculus (A)
Themedial longitudinal fasciculus is not auniform fiber tract but contains differentfiber systems that enter and exit at variouslevels. It reaches from the rostral midbraininto the spinal cord and interconnectsnumerous nuclei of the brain stem. On crosssections through the brain stem, it is foundin the middle of the tegmentum, ventrallyfrom the central gray (p. 109, AB17; p. 111,A11; p. 133, D24).
Vestibular part. Crossed and uncrossedfibers run in the longitudinal fasciculusfrom the lateral (A1), medial (A2), and infe-rior (A3) vestibular nuclei to the abducensnucleus (A4) and to the motor cells of theanterior horn of the cervical spinal cord.From the superior vestibular nucleus (A5),fibers ascend to the ipsilateral trochlear nu-cleus (A6) and oculomotor nucleus (A7).The vestibular fibers finally terminate in theipsilateral or contralateral interstitial nu-cleus of Cajal (A8) and in Darkshevich’s nu-cleus (A9) (decussation of the epithalamiccommissure [A10]). The longitudinalfasciculus connects the vestibular apparatuswith the eye and neck muscles and with theextrapyramidal system (p. 382).
Extrapyramidal part. The interstitial nu-cleus of Cajal and Darkshevich’s nucleus areintercalated in the course of the longitudi-nal fasciculus. They receive fibers from thestriatum and pallidum and crossed fibersfrom the cerebellum. They send a fiber tract,the interstitiospinal fasciculus (A11), in thelongitudinal fasciculus to the caudal brainstem and into the spinal cord.
Internuclear part. This consists of connect-ing fibers between motor nuclei of cranialnerves, namely, between abducens nucleus(A4) and oculomotor nucleus (A7), facialnucleus (A12) and oculomotor nucleus, fa-cial nucleus and motor nucleus of thetrigeminal nerve (A13), hypoglossal nucleus(A14) and ambiguous nucleus (A15).
The interconnections of motor nuclei ofcranial nerves allow certain muscle groupsto interact functionally, for example, duringthe coordination of eye muscles with the
movements of the eyeball, coordination ofeyelid muscles during opening and closingof the eyelids, and coordination of mastica-tory muscles and muscles of tongue andpharynx during swallowing and speaking.
Internuclear Connections of theTrigeminal Nuclei
Only a few secondary trigeminal fibers en-ter the medial longitudinal fasciculus. Themajority of fibers run primarily uncrossedin the dorsolateral region of the tegmentumto the motor nuclei of cranial nerves; theyform the basis of numerous important re-flexes. Crossed and uncrossed fibers run tothe facial nucleus as the basis of the cornealreflex (eyelids close upon touching the cor-nea). There are connections to the superiorsalivatory nucleus for the lacrimal reflex.Fibers to the hypoglossal nucleus, to theambiguous nucleus, and to the anteriorhorn cells of the cervical spinal cord (cells oforigin of the phrenic nerve) are the basis ofthe sneezing reflex. The pharyngeal reflex isbased on fiber connections to the ambigu-ous nucleus, the posterior vagus nucleus,and the motor nucleus of the trigeminalnerve. Connections with the posterior vagusnucleus are the basis of the oculocardial re-flex (slow heart rate upon pressure on theeyeballs).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

143
BrainStem
8
7
6
13
12
4
5
3
2
1
1514
11
10
9
Medial Longitudinal Fasciculus
A Medial longitudinal fasciculus (ac-cording to Crosby, Humphrey, andLauer)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

144Br
ainStem
Brain Stem and Cranial Nerves: Long Pathways
Central Tegmental Tract (A)
The central tegmental tract is themost im-portant efferent pathway of the extrapy-ramidal motor system (p. 310). It runs fromthe midbrain to the lower portion of theolive (A1) where the majority of its fibersterminate. The remaining fibers are thoughtto continue into the spinal cord via shortneurons that synapse in series (reticuloretic-ular fibers) (A2). In the caudal midbrain, thetract lies dorsolaterally from the decussa-tion of the superior cerebellar peduncles; itforms a large, not clearly demarcated fiberplate (p. 111, AB13) in the pons.
The tract consists of three components:
! The pallido-olivary fibers (A3) from the stri-atum (A4) and the pallidum (A5), whichextend in the pallidotegmental bundle(A6) to the capsule of the red nucleus(A7) and further to the olive. Fibers fromthe zona incerta (A8) join them.
! The rubro-olivary fibers (A9) from theparvocellular part (neorubrum) of the rednucleus. In humans, they form a strongfiber tract, the rubro-olivary fasciculus,representing the most important de-scending pathway of the red nucleus.
! The reticulo-olivary fibers (A10) join thetegmental tract from various levels,namely, from the red nucleus, the centralgray of the aqueduct (A11), and the retic-ular formation of pons and medulla ob-longata.
Impulses received by the olive from the ex-trapyramidal motor centers, and probablyalso from the motor cortex, are relayed tothe cerebellar cortex via the olivocerebellarfibers (A12).
Posterior Longitudinal Fasciculus(B)
The posterior longitudinal fasciculus(Schütz’s bundle) (p. 197, B11) contains as-cending and descending fiber systems thatconnect the hypothalamus with various nu-clei of the brain stem and provide connec-tions between the visceroefferent parasym-pathetic nuclei. A large portion of the fibers
are peptidergic (somatostatin, amongothers). They originate or terminate, respec-tively, in the septum, the oral hypothalamus,the gray tubercle (B13), and the mamillarybodies (B14). They aggregate in the mid-brain below the ependyma (p. 284) of theaqueduct and form the longitudinalfasciculus, which runs beneath theependyma on the floor of the fourth ven-tricle to the lower portion of the medullaoblongata (p. 107, AB15; p. 111, AB12).
Fibers branch off to the superior colliculi(B15) and to the parasympathetic nuclei,namely, the Edinger–Westphal nucleus (ac-cessory nucleus of oculomotor nerve) (B16),the superior (B17) and inferior (B18) sali-vatory nuclei, and the posterior vagus nu-cleus (B19). Other fibers terminate in thecranial nerve nuclei, namely, in the motornucleus of the trigeminal nerve (B20), the fa-cial nucleus (B21), and the hypoglossal nu-cleus (B22). Fibers are also exchanged withthe nuclei of the reticular formation.
The posterior longitudinal fascicle receivesolfactory impulses via the lateroposteriortegmental nucleus (habenular nucleus – in-terpeduncular nucleus – lateroposteriortegmental nucleus).
Long ascending pathways. Fibers, probablytaste fibers, ascend from the solitary nucleus(B23) to the hypothalamus. The fibers ofserotoninergic neurons can be traced byfluorescence microscopy from the posteriorraphe nucleus (B24) into the region of theseptum.
The posterior longitudinal fasciculus re-ceives hypothalamic, olfactory, and gusta-tory impulses that are relayed to the motorand sensory nuclei of the brain stem (reflexmovement of the tongue, secretion ofsaliva).
A25 Subthalamic nucleus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

145
BrainStem
4
8 5
6
79
10
3 2511
12
1
2
13
14
15
16
24
17
20
21
22
18
1923
Central Tegmental Tract, Posterior Longitudinal Fasciculus
A Central tegmental tract (accord-ing to Spatz)
B Posterior longitudinal fasciculus
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

146Br
ainStem
Brain Stem and Cranial Nerves
Reticular Formation
The scattered neurons of the tegmentumand their network of processes form the re-ticular formation. This occupies the centralarea of the tegmentum and expands fromthe medulla oblongata into the rostral mid-brain. Several areas of different structurecan be distinguished (A). In the medial partare magnocellular nuclei from where longascending and descending fiber tracts originate.The parvocellular lateral part is regarded asan association area.
Many of the neurons have long ascending ordescending axons, or axons bifurcating intoan ascending and a descending branch. Asshown by Golgi impregnation, such a neu-ron (B1) can simultaneously reach caudalcranial nerve nuclei (B2) and diencephalicnuclei (B3). The reticular formation containsa large number of peptidergic neurons(enkephalin, neurotensin, and others).
Afferent connections. The reticular forma-tion is reached by impulses of all sensorymodalities. Sensory spinoreticular fibersterminate in the medial field of medulla ob-longata and pons, and so do secondaryfibers of the trigeminal and vestibular nu-clei. Collaterals of the lateral lemniscusbring in acoustic impulses, while fibers ofthe tectoreticular fasciculus bring in opticimpulses. Experimental studies on stimula-tion have shown that reticular neurons areexcitedmore by sensory (pain), acoustic andvestibular stimuli than by optic stimuli.Other afferent fibers originate from thecerebral cortex, the cerebellum, the red nu-cleus, and the pallidum.
Efferent connections. The reticulospinaltract (p. 58, B5, B6) runs from the medialfield of medulla oblongata and pons into thespinal cord. Bundles of the reticulothalamicfasciculus ascend to the intralaminar nucleiof the thalamus (truncothalamus) (p. 180,B). Fiber bundles from the midbrain termi-nate in the oral hypothalamus and in theseptum.
Respiratory and cardiovascular controlcenters. Groups of neurons regulate respi-
ration (C), heart rate, and blood pressure(changes upon physical activity or emotion).The neurons for inspiration are localized inthe central field of the lower portion of themedulla oblongata (C4), those for expirationare further dorsal and lateral (C5). Thehigher relay stations for inhibition andstimulation of respiration lie in the pons(locus ceruleus). The autonomic nuclei of theglossopharyngeal nerve and the vagusnerve are involved in regulating heart rateand blood pressure (D). Electrical stimula-tion in the caudal central field of themedulla oblongata causes a drop in bloodpressure (depressor center) (D6), while elec-trical stimulation of the remaining reticularformation in the medulla oblongata (D7)leads to an increase in blood pressure.
Effect on the motor system. The reticularformation has a differential effect on thespinal motor system. In the medial field ofthemedulla oblongata lies an inhibition cen-ter; upon stimulation, the muscle tonedrops, reflexes fail, and the electric stimula-tion of the motor cortex no longer triggers areaction. By contrast, the reticular forma-tion in pons and midbrain has an enhancingeffect on the motor system.
Ascending activation systems. The reticu-lar formation has an effect on consciousnessvia connections to the intralaminar nuclei ofthe thalamus. When strongly stimulated bysensory or cortical input, the organism sud-denly becomes fully alert, a prerequisite forattention and perception. Upon electricalstimulation of the reticular formation, thiswake-up function can be objectively assessedby electroencephalography (EEG).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
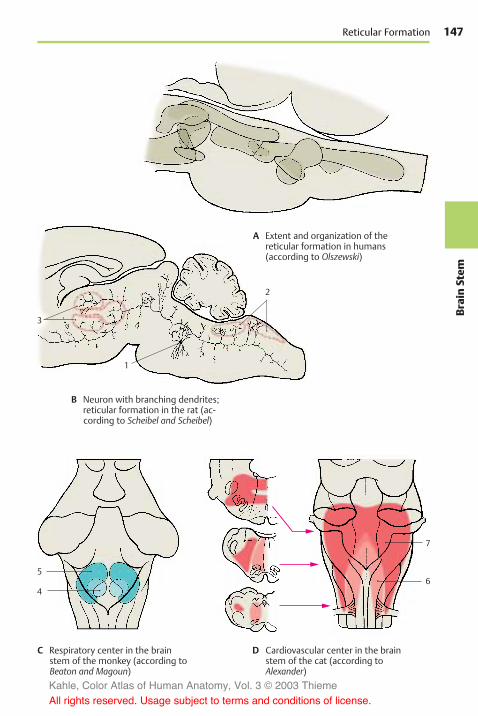
147
BrainStem
2
1
7
6
3
5
4
Reticular Formation
A Extent and organization of thereticular formation in humans(according to Olszewski)
B Neuron with branching dendrites;reticular formation in the rat (ac-cording to Scheibel and Scheibel)
C Respiratory center in the brainstem of the monkey (according toBeaton and Magoun)
D Cardiovascular center in the brainstem of the cat (according toAlexander)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

148Br
ainStem
Brain Stem and Cranial Nerves
Histochemistry of the BrainStem
Different regions of the brain stem arecharacterized by different contents inchemical substances. The delimitation ofareas according to their chemical composi-tion is called chemoarchitectonics. Substancescan be demonstrated by quantitativechemical analysis after homogenization ofthe brain tissue, or by treating histologicalsections with certain chemicals that make itpossible to show the exact localization of asubstance in the tissue. The methods com-plement each other.
Iron was one of the first substances forwhich different distributions were demon-strated. By means of the Berlin blue reac-tion, a high iron content can be demon-strated in the substantia nigra (A1) and inthe pallidum, while a lower iron content isfound in the red nucleus (A2), in the dentatenucleus of the cerebellum, and in the stri-atum. The iron is contained in neurons andglial cells in the form of small particles. Thishigh iron content is a characteristic of thenuclei that make up the extrapyramidal sys-tem (p. 310).
Neurotransmitter substances and theenzymes required for their synthesis anddegradation show marked regional varia-tions. While catecholaminergic and sero-toninergic neurons form specific nuclei inthe tegmentum (p. 33), the motor nuclei ofcranial nerves are characterized by a highcontent in acetylcholine and acetylcholineesterase. Quantitative chemical analysis ofbrain tissue yields a relatively high contentof norepinephrine in the tegmentum of themidbrain (B3), but a considerably lowercontent in the tectum (B4) and in the teg-mentum of the medulla oblongata (B5). Thecontent of dopamine is particularly high inthe substantia nigra (B1) and very low in therest of the brain stem.
Metabolic enzymes (C) also show regionalvariations in their distribution. Activity ofoxidative enzymes is generally higher in graymatter than in white matter. In the brain
stem, activity is particularly high in thecranial nerve nuclei, the lower portion ofthe olive, and the pontine nuclei. The differ-ences refer not only to the individual areasbut also to the localization of enzyme activ-ity within the cell bodies (somatic type) or inthe neuropil (dendritic type).
Neuropil. The substance between the cellbodies, which appears amorphous in Nissl-stained material, is called the neuropil. Itconsists mainly of dendrites and also ofaxons and glial processes. Themajority of allsynaptic contacts are found in the neuropil.
The distribution in themedulla oblongata ofsuccinate dehydrogenase (an enzyme of thecitric acid cycle) serves as an example fordifferent localizations of an oxidative meta-bolic enzymewithin the tissue: in the oculo-motor nucleus (C6), its activity in the peri-karya and in the neuropil is high, while it islow at both locations in the solitary nucleus(C7). In the posterior nucleus of the vagusnerve (C8), the cell bodies contrast with theneuropil owing to their high activity. Bycomparison, the highly active neuropil inthe gracile nucleus (C9) lets the poorly re-acting perikarya appear as light spots. Fibertracts (for example, the solitary tract) (C10)show very low activity. The distribution ofenzymes is characteristic for each nucleararea and is referred to as the enzyme pattern.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

149
BrainStem
10
9
7
8
6
2
1
4
1
53
Histochemistry of the Brain Stem
A Iron content of red nucleus andsubstantia nigra in humans(according to Spatz)
B Distribution of norepine-phrine and dopamine in thehuman brain stem
C Activity of succinate dehydro-genase in the medullaoblongata of a rabbit(according to Friede)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Cerebellum
Structure 152Functional Organization 162Pathways 164
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

152Ce
rebe
llum
Cerebellum
Structure
Subdivision (A–D)
The cerebellum is the integrative organ forthe coordination and fine-tuning of move-ment and for the regulation of muscle tone. Itdevelops from the alar plate of the brainstem and forms the roof of the fourth ven-tricle. The superior surface (C) is covered bythe cerebrum. Embedded into the inferiorsurface (D) is the medulla oblongata (seep. 101, C). There is an unpaired central part,the vermis of the cerebellum (ACD1, B), andthe two cerebellar hemispheres. This triparti-tion is only visible at the inferior surface,where the vermis forms the floor of a fossa,the vallecula of the cerebellum (D2). The sur-face of the cerebellum exhibits numerousnarrow, almost parallel convolutions, thefolia of the cerebellum.
Phylogenetic studies indicate that the cere-bellum consists of old portions (developedearly in evolution, present in all vertebrates)and new portions (developed late, presentonly in mammals). Accordingly, the cerebel-lum is subdivided into two parts, the floc-culonodular lobe and the cerebellar body (A3).The two are separated by the posterolateralfissure (A4). The cerebellar body is furthersubdivided by the primary fossa (AC5) intoanterior lobe and posterior lobe.
Flocculonodular Lobe (A6)
Together with the lingula (AB7), this is theoldest portion (archicerebellum). Function-ally, it is connected to the vestibular nucleithrough its fiber tracts (vestibulocerebellum)(p. 164, B).
Anterior Lobe of the Cerebellar Body (A8)
This is a relatively old component; togetherwith its central sections, which belong tothe vermis (central lobule [A–C9], culmen[A–C10]) and other sections of the vermis(uvula [ABD11], pyramid [ABD12]), it formsthe paleocerebellum. It receives thespinocerebellar tracts for proprioceptivesensibility from the muscles (spinocerebel-lum) (p. 164, A).
Posterior Lobe of the Cerebellar Body(A13)
This is the new portion (neocerebellum); itsenormous enlargement in primates con-tributes significantly to the formation of thecerebellar hemispheres. It receives the largecorticocerebellar tracts from the cerebralcortex via the pontine nuclei (pontocerebel-lum) and represents the apparatus for fine-tuning of voluntary movements.
Traditional Nomenclature
The individual sections of the cerebellumhave traditional names unrelated to theirdevelopment or function. According to thisclassification, most sections of the vermisare associated with a pair of hemisphericlobes: the central lobule (A–C9) with thewing of the central lobule (A14) on each side,the culmen (A–C10) with the quadrangularlobule (AC15), the declive (A–C16) with thesimple lobule (AC17), the folium (A–C18)with the superior semilunar lobule (ACD19),the tuber (ABD20) with the inferior semi-lunar lobule (AD21) and part of the gracilelobule (AD22), the pyramid (ABD12) withpart of the gracile lobule and the biventrallobule (AD23), the uvula (AB11) with thetonsilla (A24) and the paraflocculus (A25),and the nodulus (AB26) with the flocculus(AD27). Only the lingula (AB7) is not as-sociated with any lateral lobe.
The pale red arrow A in diagram B refers tothe direction of viewing the anterior surfaceof the cerebellum as illustrated on p. 155, A.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

153
Cerebe
llum
19
1715
10
9 5
16
18
2
20
12
1
21
22
23
19
27
11
1
8 9
10
1618 19
21
22
1016 18
20
12
11
9
7
26
2324
25
427
20 1
12
11
26
15
14
5
17
13
3
6
7
Subdivision of the Cerebellum
A Subdivision of cerebellum
B Median sectionthrough thevermis
C View from above
D View from below
A
C
D
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

154Ce
rebe
llum
Cerebellum: Structure
Cerebellar Peduncles and Nuclei(A–C)
Anterior Surface (A)
On both sides, the cerebellum is connectedto the brain stem by the cerebellar peduncles(A1). All afferent and efferent pathwayspass through them. The anterior surface ofthe cerebellum becomes fully visible onlyafter cutting through the peduncles and re-moving the pons and medulla oblongata.Between the cerebellar peduncles lies theroof of the fourth ventricle with the superiormedullary velum (A2) and the inferiormedullary velum (A3). The anterior parts ofthe vermis are exposed, namely, the lingula(A4), the central lobule (A5), the nodulus(A6), the uvula (A7), and also the flocculus(A8). The vallecula of the cerebellum (A9) issurrounded on both sides by the tonsillae(A10).
The following parts are also visible: biven-tral lobule (A11), superior semilunar lobule(A12), inferior semilunar lobule (A13),simple lobule (A14), quadrangular lobule(A15), and wing of the central lobule (A16).
Nuclei (B)
The cross section shows cortex and nuclei ofthe cerebellum. The sulci are heavilybranched, resulting in a leaflike configura-tion of the cross sectioned folia. The sagittalsection thus shows a tree-like image, thearbor vitae (tree of life) (C17).
Deep in the white matter are the cerebellarnuclei. The fastigial nucleus (B18) lies close tothe median line in the white matter of thevermis. It receives fibers from the cortex ofthe vermis, the vestibular nuclei, and theolive. It sends fibers to the vestibular nucleiand other nuclei of the medulla oblongata.The globose nucleus (B19), too, is thought toreceive fibers from the cortex of the vermisand to send fibers to the nuclei of themedulla oblongata. Fibers of the cerebellarcortex from the region between vermis andhemisphere (intermediate part) are thoughtto terminate at the hilum of the dentate nu-cleus in the emboliform nucleus (B20). Thefibers of the latter nucleus run through the
superior cerebellar peduncle to thethalamus. The dentate nucleus (B21) appearsas a heavily folded band with the medialpart remaining open (hilum of dentate nu-cleus). The cortical fibers of the hemisphereterminate in the dentate nucleus, and fibersextend from here as superior cerebellarpeduncle to the red nucleus (p. 137, B) andto the thalamus (p. 185, A).
Cerebellar Peduncles (A, C)
The efferent and afferent pathways of thecerebellum run through three cerebellarpeduncles:
! The inferior cerebellar peduncle (restiformbody) (AC22), which ascends from thelower medulla oblongata; it contains thespinocerebellar tracts and the connec-tions to the vestibular nuclei
! The medial cerebellar peduncle (brachiumpontis) (AC23) with the fiber massesfrom the pons, which originate from thepontine nuclei and represent the con-tinuation of the corticopontine tracts
! The superior cerebellar peduncle (brachiumconjunctivum) (AC24), which containsthe efferent fiber systems extending tothe red nucleus and the thalamus
C25 Tectal plate.C26Medial lemniscus.C27 Lateral lemniscus.C28 Trigeminal nerve.C29 Facial nerve.C30 Vestibulocochlear nerve.C32Olive.C32 Central tegmental tract.C33 Anterior cerebellar tract.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

155
Cerebe
llum
2
673
1
5
24 2322
8
11
4 1516
14
12
13
18
19 21
20
25
17
2433
28
26
27
2930
3231
2322
9
10
Cerebellar Peduncles and Nuclei
A Anterior view
B Cerebellar nuclei
C Cerebellar peduncles(according to Büttner)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

156Ce
rebe
llum
Cerebellum: Structure
Cerebellar Cortex (A–D)
Overview (A)
The cortex lies immediately below the sur-face and follows the course of the sulci andfolia. Projection of the convoluted relief ofthe human cerebellum onto a plane resultsin an expanse of 1m in length in the oro-caudal dimension (from the lingula to thenodulus). The cortex is regularly structuredthroughout all regions of the cerebellum. Itconsists of three layers:
! The molecular layer! The Purkinje cell layer! The granular layer
The molecular layer (A1) lies beneath thesurface; it contains few cells and consistsmainly of unmyelinated fibers. Among itsneurons we can distinguish the outer stel-late cells (lying close to the surface) and theinner basket cells. The narrow Purkinje celllayer (ganglionic layer) (A2) is formed bythe large neurons of the cerebellum, thePurkinje cells. Then follows the granularlayer (A3). It is very rich in cells, consistingof densely packed, small neurons, thegranule cells. There are also scattered largercells, the Golgi cells.
Purkinje Cells (B–D)
The Purkinje cell represents the largest andmost characteristic cell of the cerebellum.The Nissl stain shows the pear-shaped cellbody (B4) filled with coarse Nissl bodies.Also visible are the basal portions of two orthree dendrites (B5) at the upper pole of thecell. However, the cell’s entire expanse withall its processes can only be visualized byGolgi impregnation or intracellular staining.The primary stems of the dendrites ramifyinto further branches, and these again intofine arborizations that form the dendritic tree(B6). The dendrites spread in a two-dimen-sional plane like the branches of an espaliertree. The Purkinje cells are arranged in astrictly geometric fashion; spaced at rela-tively regular intervals, they form a row be-tween granular and molecular layers andsend their dendritic trees into themolecularlayer toward the surface of the folium.
Without exception, the flattened dendritictrees extend in a plane perpendicular to thelongitudinal axis of the cerebellar folium(D).
The initial branches of the dendritic tree(primary and secondary dendrites) have asmooth surface (C7) and are covered withsynapses. The fine terminal branches aredotted with short spines (C8). Each Purkinjecell carries approximately 60000 spinoussynapses. Different fiber systems terminateat the smooth and spiny sections of the cell:the climbing fibers end at the smooth sec-tion and the parallel fibers at the spiny sec-tion (p. 160).
The axon (B9) departs from the base of thePurkinje cell and extends through thegranular layer into the white matter. Theaxons of Purkinje cells terminate at neuronsof the cerebellar nuclei (p. 160, D). They giveoff recurrent collaterals. Purkinje cells useGABA as neurotransmitter.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
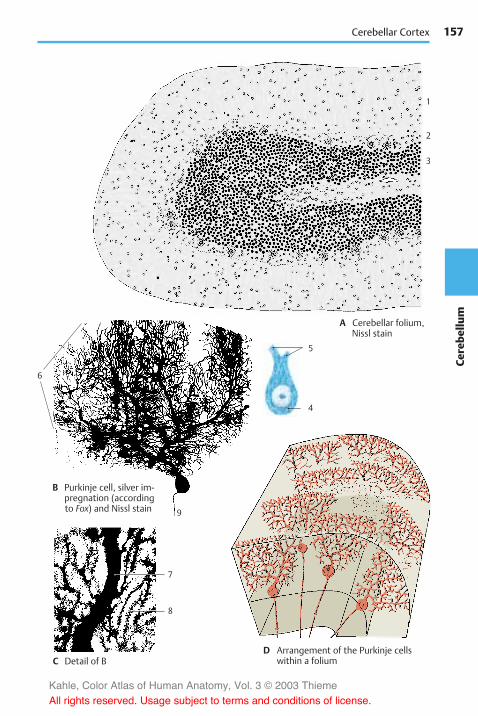
157
Cerebe
llum
6
5
4
9
7
8
1
2
3
Cerebellar Cortex
A Cerebellar folium,Nissl stain
B Purkinje cell, silver im-pregnation (accordingto Fox) and Nissl stain
D Arrangement of the Purkinje cellswithin a foliumC Detail of B
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

158Ce
rebe
llum
Cerebellum: Structure
Cerebellar Cortex (continued)
Stellate Cells and Basket Cells (A, B)
In the upper half of the molecular layer liethe stellate cells. The dendrites of these smallneurons run in all directions and reach ap-proximately 12 Purkinje dendritic trees.Their axons terminate either at the Purkinjecell bodies or run horizontally below thesurface of the folium.
In the lower third of the molecular layer liethe slightly larger basket cells (A1). Theirlong axons run horizontally above thePurkinje cell bodies and give off collaterals,the terminal branches of which form net-works (baskets) around the Purkinje cellbodies. The electron-microscopic imageshows that the basket cell fibers formnumerous synaptic contacts (B2) with thePurkinje cell, namely, at the base of the cellbody (axon hillock) and at the initial seg-ment of the axon up to where the myelinsheath begins. The rest of the Purkinje cellbody is enveloped by Bergmann’s glial cells(B3). The positioning of the synapses at theaxon hillock indicates the inhibitorycharacter of the basket cells.
Granule Cells (C)
These small, densely packed neurons formthe granular layer. At high magnification,the Golgi impregnation shows three to fiveshort dendrites which carry clawlike thick-enings at their terminal branches. The thinaxon (C4) of the granule cell ascends verti-cally through the Purkinje cell layer into themolecular layer, where it bifurcates at rightangles into two parallel fibers (p. 161, C5).
Cerebellar Glomeruli. The granular layercontains small, cell-free islets (glomeruli),in which the clawlike dendritic endings ofthe granule cells form synaptic contactswith the axon terminals of afferent nervefibers (mossy fibers, p. 161, B3). In addition,the short axons of the Golgi cells terminatehere. The electron-microscopic imageshows complex, large synapses (glomeru-lus-like synaptic complexes), which are en-veloped by glial processes.
Golgi Cells (E)
These aremuch larger than the granule cellsand are scattered throughout the granularlayer, usually slightly below the Purkinjecells (p. 161, C9). Their dendritic trees,which ramify predominantly in themolecu-lar layer and extend toward the surface ofthe folium, are not flattened like those of thePurkinje cells but are spread in all direc-tions. The cells have short axons, whicheither terminate in a glomerulus or ramifyinto a fine, dense fiber network. The Golgicells belong to the inhibitory interneurons.
Glia (D)
Apart from the regular glial cell types, suchas the oligodendrocytes (D5) and proto-plasmic astrocytes (D6) commonly found inthe granular layer, there are also glial cellsthat are characteristic for the cerebellum:Bergmann’s glia and the penniform glia ofFañanás.
The cell bodies of the Bergmann’s cells (D7) liebetween the Purkinje cells and send longsupporting fibers vertically toward the sur-face, where their small end-feet form a lim-iting glial membrane against the meninges.The supporting fibers carry leaflikeprocesses and form a dense scaffold. Berg-mann’s glia begins to proliferate wherePurkinje cells go dead. The Fañanás cells (D8)have several short processes with a charac-teristic penniform structure.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

159
Cerebe
llum
4
13
2
2
8
7
6
5
4
13
2
2
8
7
6
5
Cerebellar Cortex
A Basket cell (according to Jacob)
B Purkinje cell with basket cell synapses,electron-microscopic diagram(according to Hámori Szentágothai)
C Granule cell
D Glial cells of the cerebellum E Golgi cell (according to Jacob)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

160Ce
rebe
llum
Cerebellum: Structure
Neuronal Circuits
Afferent Fibers (A, B)
The afferent fiber systems terminate in thecerebellar cortex and give off axon collater-als to the cerebellar nuclei. There are twodifferent types of terminals: climbing fibersand mossy fibers.
The climbing fibers (AC1) terminate at thePurkinje cells by splitting up and attachinglike tendrils to the ramifications of the den-dritic tree. Each climbing fiber terminates ata single Purkinje cell and via axon collateralsalso at some stellate and basket cells. Theclimbing fibers originate from neurons ofthe olive and its accessory nuclei.
The mossy fibers (BC2) divide into widelydivergent branches and finally give offnumerous lateral branches with smallrosettes of spheroid terminals. These fit intothe clawlike terminals of the granule celldendrites and form synaptic complexeswith them (B3). Mossy fibers terminate thespinocerebellar and pontocerebellar tractsand also fibers from nuclei of the medulla.
Cortex (C)
The structure of the cerebellar cortex is de-termined by the transverse orientation ofthe flattened dendritic trees of the Purkinjecells (ACD4) and the longitudinally extend-ing parallel fibers of the granule cells(B–D5), which form synapses with thePurkinje cell dendrites. The Purkinje cellsrepresent the efferent elements of the cor-tex. They receive excitatory input throughdirect contact with climbing fibers (C1) andindirectly through mossy fibers (C2) via in-terposed granule cells (CD6). The axons ofgranule cells bifurcate in the molecularlayer into two parallel fibers each, whichmeasure approximately 3mm in totallength and travel through approximately350 dendritic trees. About 200000 parallelfibers are thought to pass through each den-dritic tree.
Stellate cells (C7), basket cells (C8), andGolgi cells (C9) are inhibitory inter-neurons which inhibit Purkinje cells. They
are coexcited by every incoming impulse,either via synapses in the glomeruli, viasynapses with parallel fibers, via synapsesof Golgi cells with mossy fibers, or via axoncollaterals of afferent fibers. In this way, theexcitation of one row of Purkinje cells inhib-its all neighboring Purkinje cells; this sharp-ens the boundaries between active Purkinjecell clusters (contrast formation).
Functional Principle of the Cerebellum (D)
The axons of the Purkinje cells (D4) termi-nate at the neurons of the subcortical nuclei(D10) (cerebellar nuclei and vestibular nu-clei). Purkinje cells are inhibitory neuronswith a high GABA content. They have astrong inhibitory effect on the neurons ofthe cerebellar nuclei, which continuouslyreceive excitatory input exclusively via axoncollaterals (D11) of the afferent fibers(climbing fibers and mossy fibers) (D12).However, these impulses cannot be passedon further, because the nuclei are under theinhibitory control of the Purkinje cells. Onlywhen Purkinje cells are inhibited by inhibi-tory interneurons (D13) is their inhibitingeffect lifted so that excitation is passed on tothe corresponding segments of the nuclei.
Hence, the cerebellar nuclei are independentsynaptic centers, which receive and transmitimpulses and in which there is continuousexcitation. Transmission is regulated by thecerebellar cortex by means of fine-tuned in-hibition and disinhibition.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

161
Cerebe
llum
–
–
6
5
4
13
10
11
12
5
7
11
12
4
1
4
1
2
2
1
5
8
9
5
2
3
66
6
Neuronal Circuits
A Terminal of a climbing fiber
B Terminal of a mossy fiber
C Neuronal connections in thecerebellar cortex (diagram)
D Neuronal connections between cere-bellar cortex and nuclei (accordingto Eccles, Ito, and Szentágothai)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

162Ce
rebe
llum
Cerebellum
Functional Organization
The afferent fiber systems terminate in cir-cumscribed areas of the cortex; in turn, cor-ticofugal fibers of different cortical areas ex-tend to specific portions of the cerebellarnuclei. The functional organization of thecerebellum is based on these fiber projec-tions. Studies in experimental animals (rab-bits, cats, monkeys) have cast light on theserelationships.
Fiber Projection (A, B)
The way in which the afferent fiber sys-tems terminate reveals a functional triparti-tion of the cerebellum into vermis, hemi-sphere, and the intermediate zone lying be-tween them. The spinocerebellar fibers,namely, posterior spinocerebellar tract,cuneocerebellar tract (p. 164, A1 and A2),and anterior spinocerebellar tract (p. 166,B14), terminate as mossy fibers in the ver-mis of the anterior lobe, in the pyramid anduvula, and in the intermediate zone lyinglaterally to them (A1). The corticopontocere-bellar fibers, which enter through the middlecerebellar peduncle, terminate as mossyfibers in the cerebellar hemisphere (A2).The vestibulocerebellar fibers terminate in theflocculonodular lobe and in the uvula (A3).Afferent pathways that synapse in the oliveand its accessory nuclei and in the lateral re-ticular nucleus terminate according to theirorigin: the spinal ones in the vermis, thecortical ones in the hemisphere.
The tripartition is also obvious from the pro-jection of corticofugal axons to the cere-bellar nuclei. In the vestibular nuclei (B4)terminate fibers from the vermis (anteriorlobe, pyramid, uvula, and nodulus) and theflocculus (B5). In the fastigial nucleus (B6)terminate fibers from the entire vermis(B7). The emboliform nucleus and globosenucleus (a complex in experimental ani-mals) (B8) receive the fibers of the interme-diate zone (B9) and, finally, in the dentatenucleus (B10) terminate the fiber masses ofthe hemisphere (B11).
Results of ExperimentalStimulation (C, D)
The pathways terminate following a set pat-tern in which lower limb, trunk, upper limb,and head area are arranged one after theother. This somatotopic organization hasbeen confirmed by stimulation experi-ments. Electrical stimulation of the cerebel-lar cortex in a decerebrate animal resultedin contractions and tonic changes in the ex-tensor and flexor muscles of the limbs, inconjugate eye movements, and in contrac-tions of the facial and cervical muscles (C)(the figure shows upper and lower surfacesof the cerebellum projected onto the sameplane).
Similar results were obtained by tactilestimulation of various body parts andsimultaneous electrical recording of the re-sulting potentials in the cerebellar cortex(evoked potentials) (D). In addition, localiza-tion of the potentials demonstrated the ipsi-lateral representation of the body half in theanterior lobe and simple lobule (D12) andthe bilateral representation in the parame-dian lobule (D13).
As this somatotopic organization has beendemonstrated in rabbits, cats, dogs, andmonkeys, we may assume that the same or-ganization is present in all mammals. Thesomatotopic organization hypothesized forthe human brain is illustrated in E.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

163
Cerebe
llum
119
7
56 8
4
10
13
12
2
1
3
Fiber Projections, Results of Experimental Stimulation
A Terminals of the vestibulocerebel-lar tract and the spinocerebellartract (according to Brodal)
B Projection of the cerebellar cortexto the cerebellar nuclei and thevestibular nucleus (according toJansen and Brodal)
C Somatotopic organization of thecerebellar cortex, motor effects inthe cat (according to Hampson,Harrison, and Woolsey)
D Somatotopic organization of the cerebellarcortex, evoked potentials duringsensory stimulation (according to Snider)
E Hypothetical somatotopic organi-zation of the human cerebellarcortex (according to Hampson,Harrison, and Woolsey)
Dorsal
Leg
Arm
Face
Eye and ear
Face
Arm
LegVentral
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

164Ce
rebe
llum
Cerebellum
Pathways
Inferior Cerebellar Peduncle(Restiform Body) (A–C)
The inferior cerebellar peduncle containsthe following fiber systems:
Posterior spinocerebellar tract andcuneocerebellar tract (A). The fibers of theposterior spinocerebellar tract (Flechsig’stract) (A1) originate from cells of the poste-rior thoracic nucleus (Clarke’s column) (A3),in which the afferent fibers of the proprio-ceptive sensibility terminate (tendon or-gans, muscle spindles, pp. 312, 314). The re-gion supplied by this tract is restricted tothe lower extremity and the lower trunk.The fibers of the posterior spinocerebellartract terminate as mossy fibers in the ver-mis and intermediate zone of the anteriorlobe and in the pyramid. The correspondingfibers for the upper extremity and the upperpart of the trunk collect in the lateralcuneate nucleus (Monakow’s nucleus) (A4)and extend as cuneocerebellar tract (A2) tothe same areas. The anterior spinocerebellartract reaches the cerebellum via the supe-rior cerebellar peduncle (brachium con-junctivum) (p. 166, B).
Vestibulocerebellar tract (B). The cerebel-lum receives primary and secondary vesti-bular fibers. The primary fibers (B5) origi-nate from the vestibular ganglion (B6) (pre-dominantly from the semicircular ducts)and run to the cerebellum without synaps-ing. The secondary fibers (B7) synapse inthe vestibular nuclei (B8). Nearly all fibersterminate in the nodulus, flocculus (B9),and fastigial nucleus (B10) but some termi-nate in the uvula. The connection with thevestibular nuclei also contains cerebellofu-gal fibers (cerebellovestibular tract), whichoriginate from the terminal areas just men-tioned and from the vermis of the anteriorlobe. Some of them synapse in the lateralvestibular nucleus and extend in the vesti-bulospinal tract (p. 382, A9) to the spinalcord.
Olivocerebellar tract (A). The olive (A11),which may be regarded as a cerebellar nu-cleus transposed ventrally, sends all itsfibers to the cerebellum. The olive and itsaccessory nuclei receive ascending fibersfrom the spinal cord (spino-olivary tract)(A12), fibers from the cerebral cortex andfrom extrapyramidal nuclei (centraltegmental tract, p. 144, A). The fibers syn-apse in specific segments of the olive toform the olivocerebellar tract (A13), whichcrosses to the opposite side and extends tothe contralateral half of the cerebellum. Thefibers of the olivary complex terminate asclimbing fibers in the cerebellar cortex: thefibers of the accessory nuclei (terminationzone of the spino-olivary tract) run to thecortex of vermis and intermediate zone ofthe anterior lobe, while the fibers of themain nucleus (termination zone of corticalfibers and tegmental tract) run to the cere-bellar hemispheres.
Reticulocerebellar tract, nucleocerebellartract, and arcuatocerebellar tract (C). Thelateral reticular nucleus (C14) receives ex-teroceptive sensory fibers which ascend to-gether with the spinothalamic tracts. Thepostsynaptic fibers run as reticulocerebellartract (C15) through the ipsilateral cerebellarpeduncle to the vermis and the hemisphere.The nucleocerebellar tract (C16) transmitstactile impulses of the facial area primarilyfrom the trigeminal nuclei (C17) to the cere-bellum. The fibers of the arcuatocerebellartract (C18) originate in the arcuate nucleus(C19) and run to the floor of the fourth ven-tricle, where they form themedullary striae.They run crossed and uncrossed and arethought to terminate in the flocculus.
The uncinate fasciculus of cerebellum, a cere-bellospinal tract originating in the con-tralateral fastigial nucleus, has not been un-equivocally demonstrated in the humanbrain.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

165
Cerebe
llum
15 18
12 1
3
11
4
213
6
578
10 9
16
17
14
19
Inferior Cerebellar Peduncle
A Posterior spinocerebellar tract,olivocerebellar tract
B Vestibulocerebellartract
C Reticulocerebellar tract,nucleocerebellar tract,arcuatocerebellar tract
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

166Ce
rebe
llum
Cerebellum: Pathways
Middle Cerebellar Peduncle(Brachium Pontis) (A)
Pathways from the cerebral cortex of thefrontal and temporal lobes extend to thepons. Together with the pyramidal tractthey form the cerebral peduncles (A1) inwhich they occupy the lateral and medialsegments: laterally lies the temporopontinetract (Türck’s bundle) (A2) and medially thefrontopontine tract (Arnold’s bundle) (A3).The fibers of the first neuron terminate inthe pontine nuclei (A4). The fibers of thesecond neuron cross from the pontine nu-clei to the opposite side and form as thepontocerebellar tract the middle cerebellarpeduncle. The fibers terminate as mossyfibers mostly in the contralateral cerebellarhemisphere but some also bilaterally in themiddle segment of the vermis.
Superior Cerebellar Peduncle(Brachium conjunctivum) (B)
Most of the efferent cerebellar pathwaysrun through the superior cerebellarpeduncle. The only afferent bundle en-tering it is the anterior spinocerebellar tract.The efferent fibers of the superior cerebellarpeduncle enter the midbrain tegmentum atthe level of the inferior colliculi and cross inthe decussation of the superior cerebellarpeduncle (B5) to the opposite side, wherethey divide into a descending (B6) and anascending limb (B7). The descending fiberbundles originate from the fastigial nucleus(B8) and the globose nucleus (B9). They ter-minate in the medial nuclei of the reticularformation (B10) in pons and medulla ob-longata, where they synapse to form the ret-iculospinal tract. Thus, cerebellar impulsesare transmitted to the spinal cord via twopathways, namely, the reticulospinal tractand the vestibulospinal tract. From bothtracts, interneurons affect the anterior horncells.
The fibers of the stronger ascending limboriginate predominantly from the dentatenucleus (B11), but partly also from the em-boliform nucleus. They terminate in twoareas: (1) in the red nucleus (B12), its sur-
roundings, and various nuclei of the mid-brain tegmentum (Edinger–Westphal nu-cleus, accessory oculomotor nucleus, Dark-shevich’s nucleus, etc.) that connect thecerebellum to the extrapyramidal system,and (2) in the dorsal thalamus (B13) fromwhere the impulses are passed on to thecerebral cortex, mainly to the motor cortex.
These connecting tracts create a large neu-ronal circuit; cerebellar impulses affect thecerebral cortex via brachium conjunctivumand thalamus. The cerebral cortex, in turn,affects the cerebellum via the corticopon-tocerebellar and cortico-olivocerebellarsystems. Thus, motor cortex and cerebellumare under mutual control.
Anterior spinocerebellar tract (Gowers’tract) (B14). The fibers originate in the pos-terior horn where fibers primarily from thetendon organs synapse. The postsynapticbundles run crossed or uncrossed; however,they do not enter the inferior cerebellarpeduncle but extend as far as the uppermargin of the pons, where they turn andenter through the superior cerebellarpeduncle (C). They terminate as mossyfibers in vermis and intermediate zone ofthe anterior lobe and in the uvula.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

167
Cerebe
llum
13
1 2
4
3
12
7
65
14
89
11
10
13
1 2
4
3
12
7
65
14
89
11
10
Middle and Superior Cerebellar Peduncles
A Corticopontine tract, pontocere-bellar tract
B Cerebelloreticular tract,cerebellorubral tract,cerebellothalamic tract,anterior spinocerebellar tract
C Anterior spinocerebellar tract
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Diencephalon
Development of theProsencephalon 170
Structure 172Epithalamus 176Dorsal Thalamus 178Subthalamus 192Hypothalamus 194Hypothalamus and Hypophysis 200
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

170Dienc
epha
lon
Diencephalon
Development of theProsencephalon
Brain and spinal cord develop from theneural tube which forms several brain ves-icles at its anterior segment, namely, therhombencephalon (A1),mesencephalon (A2),diencephalon (A3), and telencephalon (A4).The lateral walls of the vesicles thicken tobecome the brain substance proper inwhich the nerve cells and their processesdifferentiate. The developmental processbegins in the rhombencephalon andspreads to the mesencephalon and dien-cephalon. Development of the telen-cephalon is greatly delayed. A thin-walledvesicle forms on each side so that the telen-cephalon becomes subdivided into threeparts, namely, the two symmetrical hemi-spheres (A5) and the unpaired median por-tion (A6), which forms the anterior wall ofthe third ventricle (terminal lamina).
The vesicles of the telencephalon begin tocover the diencephalon. As they expand,particularly in caudal direction, thetelodiencephalic boundary becomes dis-placed. Initially, it represents the frontalborder line (A7) but then runs more andmore obliquely (A8) until it finally becomesthe lateral border (A9) of the diencephalon.The diencephalon thus comes to lie be-tween the two hemispheres, having almostno outer surface. The three brain sectionsthat are initially arranged one behind theother, namely, mesencephalon, dien-cephalon (red), and telencephalon (yellow),become largely positioned one within theother in the mature brain.
Telodiencephalic Boundary (B–E)
Only the floor of the diencephalon is visibleat the surface of the brain; it forms the opticchiasm, the tuber cinereum, and the mamil-lary bodies at the base of the brain (p. 12, A).The roof of the diencephalon becomes vis-ible only after cutting horizontally to re-move the corpus callosum (B). This exposesthe roof of the third ventricle and the twothalami. The entire region is covered by a
vascularized connective tissue plate, thetela choroidea (D10), the removal of whichopens the third ventricle (C). Over the thirdventricle and in themedial wall of the hemi-sphere, the brain tissue is extremely thinand becomes invaginated into the ventricu-lar cavity by protruding vascular loops(p. 282, A). The vascular convolutions lyingin the ventricle form the choroid plexus(D11) (production of cerebrospinal fluid).Upon removal of tela choroidea and choroidplexus, the thinned wall of the cerebralhemisphere is torn away and only the sepa-ration line remains as the choroid line (C12).The surface of the thalamus (C13) becomesexposed up to this separation line, while it isstill covered laterally by the thinned wall ofthe hemisphere. The segment of thethinned wall of the hemisphere betweenthe attachment of the plexus and thethalamostriate vein (C–E14) is called thelamina affixa (CD15). It adheres to the dorsalsurface of the thalamus and fuses with it inthe mature brain (E16). The thalamostriatevein (C–E14), which runs betweenthalamus and caudate nucleus (C–E17),marks the boundary between diencephalonand telencephalon when viewed fromabove.
BCE18 Fornix.BC19 Epiphysis.C20 Quadrigeminal plate.C21 Habenula.D22 Telodiencephalic fissure.BCE23 Corpus callosum.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
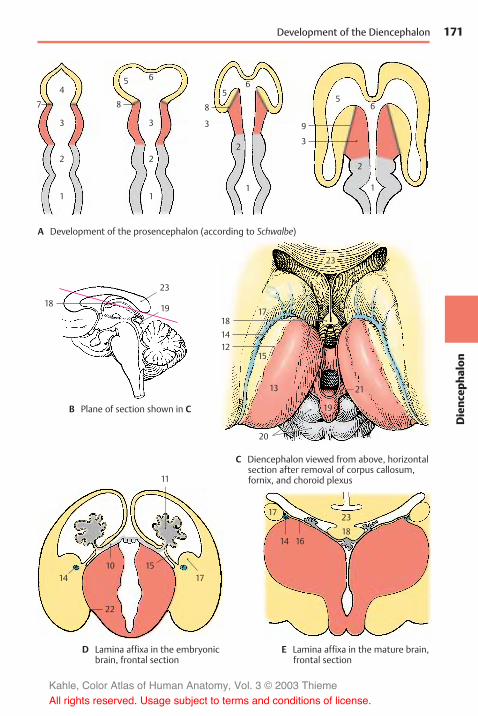
171
Dienc
epha
lon
56
1
9
6
1
2
8
3
5
65
8
4
3
7
2
1
3
2
1
3
2
19
19
23
23
23
18
18
20
13 21
15
17
1214
18
11
14 1710 15
22
17
14 16
Development of the Diencephalon
A Development of the prosencephalon (according to Schwalbe)
B Plane of section shown in C
C Diencephalon viewed from above, horizontalsection after removal of corpus callosum,fornix, and choroid plexus
D Lamina affixa in the embryonicbrain, frontal section
E Lamina affixa in the mature brain,frontal section
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

172Dienc
epha
lon
Diencephalon
Structure
Subdivision (A–C)
The diencephalon is subdivided into fourlayers lying on top of each other:
! The epithalamus (A–C1)! The dorsal thalamus (A–C2)! The subthalamus (A–C3)! The hypothalamus (A–C4)
The simple arrangement of these layers isstill clearly visible in the embryonic brain.However, it changes considerably duringdevelopment owing to differences in re-gional growth. In particular, the extraordi-nary increase inmass of the dorsal thalamusand the expansion of the hypothalamus inthe region of the tuber cinereum determinethe structure of the diencephalon.
The epithalamus (p. 176) consists of thehabenulae, a relay station for pathways be-tween the olfactory centers and the brainstem, and of the pineal gland (epiphysiscerebri). Owing to the increasing size of thethalamus, the dorsally situated epithalamus(B1) becomes medially transposed and ap-pears only as an appendage of the dorsalthalamus (C1).
The dorsal thalamus (p. 178) is the terminalstation of sensory pathways (cutaneoussensibility; taste; visual, acoustic, and vesti-bular pathways). It is connected to the cere-bral cortex by efferent and afferent fibersystems.
The subthalamus (p. 192) is the continua-tion of the midbrain tegmentum. It containsnuclei of the extrapyramidal motor system(zona incerta, subthalamic nucleus, globuspallidus) and may be regarded as the motorzone of the diencephalon.
The globus pallidus, or pallidum (CD5), is aderivative of the diencephalon. It becomesseparated from the other gray regions of thediencephalon as a result of the ingrowingfiber masses of the internal capsule (CD6)during development and finally becomesdisplaced into the telencephalon. Only asmall medial rest of the pallidum remainswithin the unit of the diencephalon; this is
the entopeduncular nucleus. As a constitu-ent of the extrapyramidal system, theglobus pallidus should logically be regardedas part of the subthalamus.
The hypothalamus (p. 194) is derived fromthe lowest layer and forms the floor of thediencephalon from which the neurohy-pophysis (A7) protrudes. It is the highestregulatory center of the autonomousnervous system.
Frontal Section at the Level of theOptic Chasm (D)
A section through the anterior wall of thethird ventricle shows parts of the dien-cephalon and telencephalon. Ventrally liesthe fiber plate of the decussation of theoptic nerve, the optic chasm (D8). A rostralexcavation of the third ventricle, the preop-tic recess (D9), is seen above it. The globuspallidus (CD5) appears laterally to the inter-nal capsule. All other structures belong tothe telencephalon: the two lateral ven-tricles (D10) and the septum pellucidum(D11) enclosing the cave of the septum pel-lucidum (D12), the caudate nucleus (D13),the putamen (CD14), and at the base, the ol-factory area (D15) (anterior perforated sub-stance). The corpus callosum (D16) and theanterior commissure (D17) connect the twohemispheres. Other fiber systems shown inthe section are the fornix (D18) and thelateral olfactory stria (D19).
Plane of section
D
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

173
Dienc
epha
lon
123
1
1
2
3
4
2
77
4
2
2
1
3
4 4
3
614
5
1
34
6
14
5
19
13
16
10
1112
18
9
8
15
17
Subdivision of the Diencephalon, Frontal Section (Optic Chiasm)
A Development of the layers of thediencephalon
B Structure of the diencephalon inthe embryonic brain
C Structure of the diencephalon inthe adult brain
D Frontal section through the rostral wall of the third ventricle (according to Villiger and Ludwig)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

174Dienc
epha
lon
Diencephalon: Structure
Frontal Section Through the TuberCinereum (A)
The plane of section lies just behind the in-terventricular foramen (foramen of Monro).Lateral ventricle and third ventricle are sep-arated by the thin base of the choroid plexus(A1). From here to the thalamostriate vein(AB2) extends the lamina affixa (AB3). Itcovers the dorsal surface of the thalamus(A4), of which only the anterior nuclei arevisible. Ventrolaterally to it and separatedby the internal capsule (AB5) lies the globuspallidus (AB6), which is divided into twoparts, the internal (A7) and the external seg-ment of the pallidum (A8). It stands outagainst the adjacent putamen (AB9) becauseof its higher myelin content. At the basalmargin and at the tip of the pallidum thereexit the lenticular fasciculus (Forel’s fieldH2) and the lenticular ansa (A10). The latterforms an arch in dorsal direction around themedial tip of the pallidum. The ventral partof the diencephalon is occupied by the hy-pothalamus (tuber cinereum [A11] and in-fundibulum [A12]), which appears markedlypoor in myelin, in contrast to the heavilymyelinated optic tract (AB13).
The diencephalon is enclosed on both sidesby the telencephalon, but without a clearlyvisible boundary. The closest nuclei of thetelencephalon are the putamen (AB9) andthe caudate nucleus (AB14). Anterior to theglobus pallidus lies a nucleus belonging tothe telencephalon, the basal nucleus (Mey-nert’s nucleus) (A15). It receives fibers fromthemidbrain tegmentum. Its large choliner-gic neurons project diffusely into the entireneocortex. The fornix (AB16) is seen twicebecause of its arched course (p. 233, C).
Frontal Section at the Level of theMamillary Bodies (B)
The section shows both thalami; their in-crease in volume has lead to secondary fu-sion in the median line, resulting in the in-terthalamic adhesion (B17). Myelinated fiberlamellae, the medullary layers of thethalamus, subdivide the thalamus intoseveral large complexes of nuclei. Dorsally
lies the anterior nuclear group (B18), ven-trally to it the medial nuclear group (B19),which borders medially on several smallparaventricular nuclei (B20) and is laterallyseparated by the internal medullary layer(B21) from the lateral nuclear group (B22).Subdivision of the latter into a dorsal and aventral area is less distinct. The entire com-plex is enclosed by a narrow, shell-shapednucleus, the reticular nucleus of thethalamus (B23), which is separated from thelateral nuclear group by the external medul-lary layer (B24).
Ventrally to the thalamus lies the sub-thalamuswith the zona incerta (B25) and thesubthalamic nucleus (Luys’ body) (B26). Thezona incerta is delimited by two myelinatedfiber plates, dorsally by Forel’s field H1(thalamic fasciculus) (B27) and ventrally byForel’s field H2 (lenticular fasciculus) (B28).Below the subthalamic nucleus appears therostral pole of the substantia nigra (B29).The floor of the diencephalon is formed bythe two mamillary bodies (B30). The mamil-lothalamic fasciculus (Vicq d'Azyr’s bundle)(B31) ascends from the mamillary body tothe thalamus.
AB32 Corpus callosum.A33 Amygdaloid body (amygdala).B34 Hippocampus.A35 Anterior commissure.B36 Medullary stria (p. 176, A2).
Planes of sections
A B
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

175
Dienc
epha
lon
1618
14
22
32 2423
215
920
17
31
27
26
29
34
13
28 2530
32
14 23
14
8
710
151113
33
1656
9
3516
12
3619
23
6
Frontal Sections (Tuber Cinereum, Mamillary Bodies)
A Frontal section through the diencephalon at the level of the tuber cinereum(according to Villiger and Ludwig)
B Frontal section through the diencephalon at the level of the mamillary bodies(according to Villiger and Ludwig)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

176Dienc
epha
lon
Diencephalon
Epithalamus
The epithalamus includes the habenula(with habenular nuclei, habenular commis-sure, and medullary stria), the pineal gland,and the epithalamic commissure (posteriorcommissure).
Habenula (A)
The habenula (A1) (p. 171, C21) with its af-ferent and efferent pathways forms a relaysystem in which olfactory impulses aretransmitted to efferent (salivatory andmotor) nuclei of the brain stem. In this way,olfactory sensation is thought to affect foodintake. The habenular nucleus containsnumerous peptidergic neurons.
The afferent pathways reach the habenularnuclei via the medullary stria of the thalamus(A2). It contains fibers from the septal nuclei(A3), the anterior perforated substance (ol-factory area) (A4), and the preoptic region(A5). Furthermore, it receives fibers fromthe amygdaloid body (amygdala) (A6) cross-ing over from the terminal stria (A7).
The efferent pathways extend into the mid-brain. The habenulotectal tract (A8) transmitsolfactory impulses to the superior colliculi.The habenulotegmental tract (A9) terminatesin the dorsal tegmental nucleus (A10), fromwhere there is a link to the posterior longi-tudinal fasciculus (p. 144, B) with connec-tions to the salivatory and motor nuclei ofthe masticatory and deglutitory muscles(olfactory stimuli leading to secretion ofsaliva and gastric juice). The habenulo-inter-peduncular tract, Meynert’s bundle (A11), ter-minates in the interpeduncular nucleus(A12) (p. 132, D21) which is connected tovarious nuclei of the reticular formation.
Pineal Gland (B–D)
The pineal gland (pineal body, epiphysiscerebri) (A13, B), is a small peg-shaped bodyat the posterior wall of the third ventricleabove the quadrigeminal plate (p. 170BC19). Its cells, the pinealocytes, are groupedinto lobules by connective tissue septa. In
silver-impregnated sections, they showlong processes with club-shaped terminalswellings (C), which terminate predomi-nantly at blood vessels (D). In adults, thepineal gland contains large foci of calcifica-tion (B14), which are visible on radiographs.
In lower vertebrates, the pineal gland is a photo-sensitive organ; it registers changes from light todark either by a special parietal eye or just by thelight penetrating through the thin roof of theskull. By doing so, it influences the day and nightrhythm of the organism. For example, it regulatesthe color change in amphibians (dark pigmenta-tion during the day, pale pigmentation at night)and the corresponding change in the animal’s be-havior. The pineal gland also registers the trans-ition from bright summertime to dark wintertimeand thus brings about seasonal changes in thegonads.
In higher vertebrates, the light does not penetratethe thick roof of the skull. The rhythm of day andnight is transmitted to the pineal gland throughthe following route: via retinal fibers to the su-prachiasmatic nucleus in the hypothalamus, thenvia efferent hypothalamic fibers to the interme-diolateral nucleus. and finally via postganglionicfibers of the cervical sympathetic chain to thepineal gland.
In humans, the pineal gland is thought to inhibitmaturation of the genitals until puberty. As in an-imals, it is supposed to have an antigonadotropicaction. Hypergonadism has been observed insome cases of pineal gland destruction inchildren.
Epithalamic commissure (posterior com-missure) (B). Not all fiber systems that passthrough the epithalamic commissure (B15)are known. Habenulotectal fibers cross in it.From the various pretectal nuclei that sendfibers through the commissure, the intersti-tial nucleus of Cajal and Darkshevich’s nu-cleus are the most important ones. Vestibu-lar fibers are also thought to cross in thiscommissure (p. 135, B23).
A16 Olfactory bulb.A17 Chiasm.A18 Hypophysis.B19 Pineal recess.B20 Habenular commissure.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

177
Dienc
epha
lon
72
13
89
11
126
1817
16
2014
31
4 5
10
15
19
Habenula and Epiphysis
A Fiber connections ofthe habenula
B Pineal gland
C Pinealocytes, silver impregnation(according to Hortega)
D Histological appearance of the pineal gland,silver impregnation (according to Hortega)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

178Dienc
epha
lon
Diencephalon
Dorsal Thalamus
The dorsal thalami are two large, ovoid nu-clear complexes. Their medial surfaces formthe wall of the third ventricle, while theirlateral surfaces border on the internal cap-sule. They extend from the interventricularforamen (foramen of Monro) to the quad-rigeminal plate of the midbrain.
The two thalami are the relay stations formost sensory pathways, almost all of whichterminate in the contralateral thalamus.Fiber bundles connect the thalami also withthe cerebellum, globus pallidus, striatum,and hypothalamus.
Thalamic Radiation (A)
The thalamus (A1) is connected to the cere-bral cortex by the corona radiata, orthalamic radiation (A2–4). The fibers runobliquely through the internal capsuletoward the cerebral cortex. The more prom-inent bundles are the anterior thalamic radia-tion (A2) (to the frontal lobe), the superiorthalamic radiation (A3) (to the parietal lobe),the posterior thalamic radiation (A4) (to theoccipital lobe), and the inferior thalamicradiation (to the temporal lobe).
The great variety of fiber connections indi-cates the central function of the thalamus,which is directly or indirectly integratedinto most systems. Accordingly, it is not auniform structure but a highly organizedcomplex consisting of diversely structurednuclear groups.
Based on their fiber connections, two typesof thalamic nuclei are distinguished. Nucleiwith fiber connections to the cerebral cor-tex are collectively called specific thalamicnuclei; nuclei without any connection to thecortex but with connections to the brainstem are the nonspecific thalamic nuclei.
Clinical Note: Knowledge of the basic struc-ture of the thalamus is of practical importance be-cause motor disturbances and conditions of paincan be treated by stereotactic surgery at thethalamus.
Specific Thalamic Nuclei (B–D)
The cortex-dependent specific thalamic nu-clei are subdivided into the following nu-clear groups (or complexes):
! The anterior nuclear group, anteriorthalamic nuclei (green) (B–D5).
! The medial nuclear group, medial thalamicnuclei (red) (BD6).
! The lateral nuclear group, ventrolateralthalamic nuclei (blue) (CD7); this group isfurther divided into a lateral tier, thelateral nuclei, and a ventral tier, the ven-tral nuclei
! The lateral geniculate nucleus (BC8).! The medial geniculate nucleus (BC9).! The pulvinar (BC10).! The reticular nucleus of the thalamus (D11).
The nuclear groups are separated by layersof fibers: the internal medullary lamina (D12)(between the medial nuclear group and thelateral and anterior nuclear groups) and theexternal medullary lamina (D13) (between thelateral nuclear group and the reticular nu-cleus which encloses the lateral surface ofthe thalamus).
The reticular nucleus and nonspecific nu-clei, with the exception of the centromediannucleus (B14), have been omitted from thereconstruction of nuclear groups (B, C). Themost anterior nuclear groups are the ante-rior nuclei (B5) to which the medial nuclei(B6) border caudally. In the lateral complex,we distinguish between a nuclear group lo-cated dorsally (the lateral nuclei, lateral dor-sal nucleus [C15] and lateral posterior nu-cleus [C16]), and a group located ventrally(the ventral nuclei, ventral anterior nucleus[C17], ventral lateral nucleus [C18], and ven-tral posterior nucleus [C19]).
BC20 Superficial dorsal nucleus.B21 Interventricular foramen
(foramen of Monro).C22 Anterior commissure.C23 Optic chiasm.C24 Mamillary body.C25 Optic tract.A26 Cut surface of corpus callosum.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

179
Dienc
epha
lon
6
5
20
10
14
9
8
21
15
17
5
16
20
19
1810
95
67
13
11
128
25
7
2423
22
26
4
31
2 21
6
5
20
10
14
9
8
21
15
17
5
16
20
19
1810
95
67
13
11
128
25
7
2423
22
26
4
31
2 21
Thalamic Radiation, Specific Thalamic Nuclei
A Thalamic radiation, fiber preparation(according to Ludwig and Klingler)
B, C Diagrammatic reconstructionof the thalamic complexes
B Median view
C Lateral view
D Frontal sectionshowing nuclearorganization
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

180Dienc
epha
lon
Diencephalon: Dorsal Thalamus
Specific Thalamic Nuclei(continued) (A)
Each nuclear group is connected to aspecific region (field of projection) in thecerebral cortex; hence the term specificthalamic nuclei. Within this system, thenuclei project to their cortical fields and, inturn, the cortical fields project to the re-spective thalamic nuclei. Thus, there exists aneuronal circuit with a thalamocorticallimb and a corticothalamic limb. The neu-rons of the specific thalamic nuclei transmitimpulses to the cerebral cortex and are, inturn, influenced by the respective corticalfields. Hence, the function of a cortical fieldcannot be examined without the thalamicnucleus belonging to it; likewise, the func-tion of a thalamic nucleus cannot be ex-amined without the cortical field belongingto it.
When neurons of the specific thalamic nu-clei become separated from their axon ter-minals, they respond by retrogradedegeneration. Therefore, destruction of cir-cumscribed cortical fields results in neu-ronal death in the respective thalamic nu-clei. The thalamic projection fields of thecerebral cortex can be delineated in thisway. The anterior nuclei (A1) are connectedwith the cortex of the cingulate gyrus (A2)and the medial nuclei (A3) with the cortex ofthe frontal lobe (A4). The lateral nuclei (A5)project to the dorsal and medial cortex ofthe parietal lobe (A6), with the lateral dorsalnucleus partly supplying the retrosplenialpart of the cingulate gyrus. Among the ven-tral nuclei, the ventral anterior nucleus (A7) isconnected with the premotor cortex (A8),the ventral lateral nucleus (A9) with themotorprecentral area (A10), and the ventral poste-rior nucleus (A11) with the sensory postcen-tral area (A12). The pulvinar (A13) projects tothe cortical parts of the parietal and tem-poral lobes (A14) and to the cuneus (A15).The lateral geniculate nucleus (A16) is con-nected through the visual pathway with thevisual cortex (striate area) (A17), the medialgeniculate nucleus (A18) through the auditorypathway with the auditory cortex (trans-verse temporal gyri, Heschl’s convolutions)(A19).
Nonspecific Thalamic Nuclei (B)
These nuclei have fiber connections to thebrain stem, to the diencephalic nuclei, and tothe corpus striatum, but no direct connec-tion to the cerebral cortex has been demon-strated anatomically. Their neurons are notinjured by removal of the entire cerebralcortex; they are cortex-independent. Twogroups of nuclei are distinguished:
! The median nuclei (nuclei of the centralthalamic gray matter) (B20), which aresmall cell clusters located along the wallof the third ventricle
! The intralaminar nuclei (B21), which areembedded into the internal medullarylamina; the largest of them is the cen-tromedian nucleus (B22)
Electrical stimulation of these nuclei doesnot lead to excitation of individual corticalareas but to changes in the electrical activityof the entire cerebral cortex. Hence, they arecalled nonspecific nuclei. The pathwaysthroughwhich cortical activity is influencedare unknown. The ascending pathways ofthe reticular formation (ascending activat-ing system, p. 146) terminate in the in-tralaminar nuclei.
Alternative Nuclear Subdivision (C)
The subdivision of the thalamus according toHassler differs from the traditional arrange-mentmainly with respect to the subdivisionof the lateral nuclear complex. The nucleuslocated most orally is called the lateropolarnucleus (C23). Then follows the division intodorsal, ventral, and central zones. Thesethree zones are further divided into oral, in-termediate, and caudal segments. This re-sults in the following nuclei: dorsally lie thedorso-oral nucleus (C24), the dorsointerme-diate nucleus (C25), and the dorsocaudal nu-cleus (C26), and ventrally lie the ventro-oralnucleus (C27), the ventrointermediate nu-cleus (C28), and the ventrocaudal nucleus(C29).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

181
Dienc
epha
lon
4
2
31
5
1197
19
4
12108
6
14
23
24
2526
29
22
21 20
21 20
2827
16
18
13
12108
6
15
17
Specific Thalamic Nuclei, Nonspecific Thalamic Nuclei
A Cortical projections of thethalamus (according to Walker)
B Nonspecific thalamic nucleiC Alternative nuclear subdivision(according to Hassler)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

182Dienc
epha
lon
Diencephalon: Dorsal Thalamus
Anterior Nuclear Group (A)
The anterior nuclear group (anterior com-plex) (A1) consists of a principal nucleusand several smaller nuclei. All these nucleihave bidirectional connections to the cingu-late gyrus (A2), which lies at the medialaspect of the hemisphere directly above thecorpus callosum. Predominantly afferentfibers from the mamillary body (A3) reachthe anterior nucleus as a thick, myelinatedbundle, the mamillothalamic tract (Vicqd'Azyr’s bundle) (A4). Fibers of the fornix arealso thought to terminate in the anteriornucleus. The nucleus is thought to representa relay station in the limbic system (p. 332).However, its functional significance is notyet precisely known. Electrical stimulationleads to autonomic reactions (changes inblood pressure and respiration rate) due toconnections with the hypothalamus.
Medial Nuclear Group (B)
The medial nuclear group (medial complex)(B5) consists of a medial magnocellular nu-cleus, a lateral parvocellular nucleus, and acaudal nucleus. All nuclei project to thefrontal lobe, namely, to the premotor cortex,the polar cortex, and the orbital cortex (B6).Afferent fiber bundles run via the inferiorthalamic radiation from the globus pallidus(B7) and from the basal nucleus of Meynert(substantia innominata) (p. 174, A15) to themedial nuclear group. The medial magno-cellular nucleus has fiber connections to thehypothalamus (B8) (preoptic area and tubercinereum) and to the amygdaloid body(amygdala). The lateral parvocellular nu-cleus receives fibers from the adjacent ven-tral nuclei of the thalamus.
The medial nuclear complex is thought toreceive visceral and somatic impulses viapathways from the hypothalamus and ven-tral nuclei; the impulses are integrated hereand then transmitted via the anteriorthalamic radiation to the frontal cortex. Thebasic affective mood, which is essentially de-termined by unconscious stimuli from thevisceral and somatic spheres, is thought toenter consciousness in this way.
Clinical Note: It has been observed in patientswith severe agitation that incision of thethalamocortical tracts (prefrontal lobotomy) hada calming effect but was also associated with in-difference and a regression of personality. Thestereotactic destruction of the medial nuclearcomplex has similar effects.
Centromedian Nucleus (C)
The centromedian nucleus (C9) is thelargest of the nonspecific thalamic nuclei andbelongs to the intralaminar nuclei sur-rounding the medial nuclear complex. It isdivided into a ventrocaudal parvocellularpart and a dorso-oral magnocellular part.The fibers of the superior cerebellarpeduncle, which terminate in this nucleus,originate from the emboliform nucleus of thecerebellum (C10). Apart from these crossedfibers, the nucleus also receives ipsilateralfibers from the reticular formation (C11).Fibers from the internal segment of the globuspallidus (C12) branch off from the lenticularfasciculus (Forel’s field H2) and radiate intothe nucleus. Fibers from the precentral cor-tex (area 4) are thought to terminate here aswell. Efferent fiber bundles run from themagnocellular region to the caudate nucleus(C13) and from the parvocellular region tothe putamen (C14). These tracts provide aconnection between cerebellum and stri-atum.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

183
Dienc
epha
lon
2
1
4
3
6
8
9
13
10
12
14
11
5
7
2
1
4
3
6
8
9
13
10
12
14
11
5
7
Anterior and Medial Nuclear Groups, Centromedian Nucleus
A Fiber connections ofthe anterior complex(anterior thalamic nuclei)
B Fiber connections of the medialcomplex (medial thalamic nuclei)
C Fiber connections of thecentromedian nucleus
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

184Dienc
epha
lon
Diencephalon: Dorsal Thalamus
Lateral Nuclear Group (A)
The lateral nuclei form the dorsal tier of thelateroventral nuclear complex. Neither ofthe two lateral nuclei, the lateral dorsal nu-cleus (A1) and the lateral posterior nu-cleus (A2), receives any extrathalamicinput; they are connected only with otherthalamic nuclei and are therefore viewed asintegration nuclei. They send their efferentfibers to the parietal lobe (A3).
Ventral Nuclear Group (A, B)
Ventral anterior nucleus (VA) (A4). It re-ceives afferent fibers mainly from the inter-nal segment of the globus pallidus (A5)(they probably originate in the caudate nu-cleus [A6]) and from the nonspecificthalamic nuclei. Additional fibers arethought to stem from the substantia nigra,the interstitial nucleus of Cajal, and the re-ticular formation. The nucleus projects tothe premotor cortex (A7) but depends onlypartially on the cortex because only half ofits neurons die after injury to the cortical re-gion. The ventral anterior nucleus is inte-grated into the ascending activation system;stimulation within the nuclear zone causesa change in electrical activity of the cortex.
Ventral lateral nucleus (VL) (AB8). Themost important afferent system of the nu-cleus are the fibers of the crossed superiorcerebellar peduncle (A9). In the anteriorsegment of the ventral lateral nucleus thereterminate fibers from the globus pallidus(thalamic fasciculus) (A10). The efferentfibers (A11) extend to the cortex of the pre-central gyrus (A12). The somatotopic or-ganization of the ventral lateral nucleus isobvious in this system (AB8); the lateralpart of the nucleus is connected to the legregion of the precentral cortex, the adjacentparts to the trunk and arm regions, and themedial part to the head region. Thus,thalamic nucleus and cortical area show acorresponding topical subdivision. Informa-tion from the cerebellum (body posture,coordination, muscle tone) reaches themotor cortex via the ventral lateral nucleus;the cerebellum influences voluntary move-
ment in this way. A narrow caudal region ofthe nucleus is distinguished as ventral inter-mediate nucleus (B13). Forel’s dorsolateraltegmental fasciculus from the ipsilateralvestibular nuclei terminates here (head andgaze turn to the same side).
Ventral posterior nucleus (VP) (A14). Thenucleus is the terminus for the crossed, sec-ondary sensory pathways (A15). Originatingfrom the nuclei of the posterior column, themedial lemniscus (p. 140, B) radiates intothe lateral segment of the nucleus, the ven-tral posterolateral nucleus (VPL) (B16). Thefibers of the gracile nucleus lie laterally,those of the cuneate nucleus medially. Theresulting somatotopic organization of thenucleus can also be demonstrated electro-physiologically; the lower limb is repre-sented laterally, trunk and upper limb me-dially. The secondary trigeminal fibers(lemniscus trigeminal) terminate in theme-dial segment of the nucleus, the ventral post-eromedial nucleus (VPM) (B17). They transmitsensory information from the head and oralcavity, thus completing the homunculus ofthe contralateral half of the body. The ter-minus of the secondary gustatory pathway(p. 328) lies most medially. The pathways ofthe protopathic sensibility (see p. 324),namely, the spinothalamic tract and thetrigeminal pain fibers, are thought to termi-nate bilaterally in the basal areas of the nu-cleus. The efferent fibers of the nucleus(A18) extend to the sensory postcentral area(p. 250), the somatotopic organization ofwhich results from the topical subdivisionof the ventral posterior nucleus and its pro-jection to the cerebral cortex.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

185
Dienc
epha
lon
7
3
184
8
105
15
14
9
4
8
13
1617
1
2
1112
6
Lateral and Ventral Nuclear Groups
A Fiber connections of the lateral complex(lateroventral thalamic nuclei)
B Somatotopic organization of theventral nuclei, horizontal section(according to Hassler)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

186Dienc
epha
lon
Diencephalon: Dorsal Thalamus
Ventral Nuclear Group (continued)
Functional Topography of the VentralNuclei (A, B)
Knowledge of the functional organizationmakes it possible to stop severe painthrough stereotactic destruction within theventral posterior nucleus without affectingtactile sensation. By destroying areas withinthe ventral lateral nucleus, motor distur-bances (hyperkinesis) can be eliminatedwithout producing concomitant paralysis.The controlled stimulation required forthese procedures yields clues regarding therepresentation of various body regions. Asshown in the stimulation diagram (red sym-bols) (A), the representation of the bodyruns obliquely from dorsolateral (leg region,| ; arm region, – ) to mediobasal (head re-gion, (!)). Controlled stimulation of theventral lateral nucleus (orange symbols) (B)results also in involuntary sound produc-tion (") or eruptively uttered sentences (!),thereby revealing which of the two thalamiis the dominant one, depending on thedominant hemisphere (p. 262) (in right-handed persons, this is the left thalamus).
Lateral Geniculate Body (C)
This nucleus (C1) lies somewhat isolated atthe ventrocaudal aspect of the thalamus andis a relatively independent structure. Itshows stratification into six cell layerswhich are separated by the afferent fiberbundles of the optic tract. Crossed and un-crossed optic fibers terminate in a regulararrangement (p. 256, A) in each of the twogeniculate nuclei. In the left lateral genicu-late body, the temporal half of the retina ofthe left eye and the nasal half of the retina ofthe right eye are represented; in the rightlateral geniculate body, the temporal half ofthe retina of the right eye and the nasal halfof the retina of the left eye are represented(p. 355B). The fibers from themacula, whichis the region of greatest visual acuity, termi-nate in a central wedge-shaped area, whichextends through all cell layers (p. 190, A9).The neurons of the lateral geniculate nu-cleus send their axons to the visual cortex,
the striate area (C2) at the medialhemispheric surface of the occipital lobe(central optic radiation or occipitothalamicradiation).
Medial Geniculate Body (D)
This nucleus (D3) is the diencephalic relaystation of the auditory pathway. It appears asan externally visible eminence medially tothe lateral geniculate body. Its afferentfibers form the brachium of the inferior col-liculus from the ipsilateral inferior colliculus(D4). Some fiber bundles of the auditorypathway come from the nucleus of the trape-zoid body and the ipsilateral cochlear nuclei;however, most fibers originate from thecontralateral cochlear nuclei. The efferentfibers of the medial geniculate nucleus ex-tend to the auditory cortex (D5), which liesin the transverse temporal gyri, or Heschl’s con-volutions (p. 253, C1) of the temporal lobe.
Pulvinar
The pulvinar (p. 179, BC10) occupies thecaudal third of the thalamus and is dividedinto several nuclei. Its functional signifi-cance is not understood. Since it does not re-ceive any extrathalamic input, it must beviewed as an integration nucleus. Afferentfibers from the lateral geniculate nucleus(collaterals of the optic fibers) and probablyalso fibers from the medial geniculate nu-cleus enter the pulvinar.
There are reciprocal fiber connections be-tween the pulvinar and the cortex of theparietal lobe and the dorsal temporal lobe.Hence, the pulvinar is not only integratedinto the optic and acoustic systems but isalso connected with the cortical areas im-portant for language and symbolic thinking(p. 250).
Clinical Note: Injury (or electrical stimula-tion) of the pulvinar causes speech disorder inhumans.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

187
Dienc
epha
lon
2
1
3
4
5
Ventral Nuclei, Lateral and Medial Geniculate Bodies
A Motor responses following stimu-lation of the ventral lateral nu-cleus (according to Schaltenbrand,Spuler, Wahren and Rümler)
B Responses of speech and soundfollowing stimulation of the ven-tral lateral nucleus (according toSchaltenbrand, Spuler, Wahren andRümler)
C Fiber connections ofthe lateral geniculate body
D Fiber connections of the medialgeniculate body
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

188Dienc
epha
lon
Diencephalon: Dorsal Thalamus
Frontal Section Through the RostralThalamus (A–C)
In the myelin staining, the anterior and me-dial nuclear groups are clearly distinguisha-ble from the lateral nuclear group by theirpoor and delicate myelination. The dorsallylocated anterior nuclear group (green) (A–C1)bulges against the interventricular foramen(foramen of Monro) (AB2) and forms thethalamic eminence. The medial nuclear group(red) is enveloped by the internal medullarylamina (B3) and the intralaminar nuclei (C4)which separate it from the lateral portion.Within the medial nuclear group, a medialmagnocellular portion (AC5) and a lateralparvocellular portion (AC6) surroundingthe medial one can be distinguished.
The largest part of the thalamus is formedby the lateroventral nuclear group (blue),which surrounds the medial portion like abroad shell. It contains considerably moremyelin, and a difference between its dorsaland ventral regions can be recognized in themyelin stained section (B). As compared tothe dorsal region (lateral dorsal nucleus)(A–C7), the ventral nuclear region hasmoreprominent and coarser myelin fibers. Its di-vision into a medial and a lateral segmentcan be easily recognized in the overview.The section shows the ventral lateral nu-cleus. In its medial segment (A–C8) termi-nate fibers from the midbrain tegmentum.Lateral to it is seen the rostral part of the nu-cleus (A–C9) where the fiber bundles of thesuperior cerebellar peduncle terminate; itsprojection to the precentral area (area 4) re-veals a somatotopic organization.
The lateral surface of the thalamus isformed by the reticular nucleus of the thalamus(A–C10). As a narrow layer of cells, thisnucleus laterally surrounds the entirethalamus like a shell and extends from therostral pole, where it is widest, to the pulvi-nar and to the lateral geniculate nucleus. Itis separated from the lateral nuclear com-plex by a lamella of myelin fibers, the exter-nal medullary lamina (B11). The relation-ships between cerebral cortex and reticularnucleus vary for different nuclear segments:the frontal cortex is connected with the ros-
tral portion of the nucleus, the temporalcortex with the middle portion, and theoccipital cortex with the caudal portion. Thefunctional significance of this nucleus is un-known. Its neurons send many collaterals tothe other thalamic nuclei.
Fiber relationships between the thalamicnuclei and certain cortical areas have beenestablished by experimentally destroyingthe cortical segments or by severing thefibers. The neurons of the respective nucleiundergo retrograde degeneration once theiraxons have been severed. Neurons of the re-ticular nucleus, however, are thought to un-dergo transneuronal degeneration ratherthan retrograde degeneration, that is, theynot degenerate because their axons havebeen severed but because they have lost af-ferent fibers terminating on them. Thiswould mean that the cortex projects to thereticular nucleus, while the latter does notproject to the cortex.
Plane of section
A–C
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

189
Dienc
epha
lon
10
9
8
6
7
5
2
1
3
7
8
104 10
7
8
9
65
1
11
9
1
2
Rostral Thalamus, Frontal Section
A–C Frontal section through the rostral thalamusA Cellular staining (Nissl)
B Fiber staining (myelin) C Nuclear organization(according to Hassler)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

190Dienc
epha
lon
Diencephalon: Dorsal Thalamus
Frontal Section Through the CaudalThalamus (A–C)
At this level, the section again shows themedial nuclear group (red) (A–C1) and thelateroventral nuclear group (blue) (A–C2). Itincludes themost caudal parts of themedialnucleus. Dorsally separated from the super-ficial dorsal nucleus (A–C3) by a narrowmy-elin layer, they are otherwise surrounded bythe internal medullary lamina and the in-tralaminar nuclei. The nonspecific thalamicparts here reach special expansion throughthe centromedian nucleus (A–C4).
The most rostral nuclear portions of thepulvinar (A–C5) lie dorsally between themedial and the lateral nuclear groups. Thisrostral part of the pulvinar projects to theupper convolutions of the temporal lobeand is thought to receive fibers from thelateral lemniscus; it is therefore assumed tobe an integration nucleus of the acousticsystem.
The ventral posterior nucleus (A–C6) is seenin the lateroventral area. Themedial lemnis-cus, the spinothalamic pathways, and thesecondary trigeminal fibers terminate here.The outer portion, which receives the fibersfor the limbs and the trunk, is rich in myeli-nated fibers and has fewer cells than theinner portion, which receives the fibers forthe head region. The inner portion is rich incells and has thinlymyelinated fibers. It sur-rounds the centromedian nucleus ventrallyand laterally; it appears as a crescent-shaped figure in the myelin-stained sectionand, hence, is called the semilunar nucleus(B7).
The lateral geniculate body (A–C8) liesslightly apart from the complex of thethalamus at the ventral surface of the dien-cephalon. It is indented at the base and pro-trudes laterally (lateral geniculum). It ischaracterized by prominent stratificationinto six layers of cells and five intercalatinglayers of fibers. The latter are formed by thefibers of the optic tract, which disperse ac-cording to a set pattern and terminate atneurons of different cell layers (p. 257, A).The upper four of these layers are parvo-
cellular, the lower two layers are magno-cellular. In the second, third, and fifth layersterminate the fibers from the retina of theipsilateral eye (uncrossed optic nervefibers), while those from the contralateraleye (crossed optic nerve fibers) terminate inthe first, fourth, and sixth layers. Fibers fromthe site of visual acuity, the macula, termi-nate in the central area (A9). When themac-ula is destroyed, the geniculate cells of thisarea undergo transneuronal degeneration.The lateral geniculate body is surrounded bya dense capsule of myelinated fibers. Theseare the dorsally and laterally emergingfibers of the optic radiation (geniculocal-carine tract) (p. 258, C).
Medially to the lateral geniculate body, thesection shows the caudal portion of the me-dial geniculate body (A–C10). The reticularnucleus (AC11) forms the lateral capsule. Itwidens ventrally and also encloses thelateral geniculate body.
Plane of section
A–C
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

191
Dienc
epha
lon
3
1
4
7
6
9
8
10
2
11
5
1
3 5
2
67
4
10
8
3
1
1
10
4
5
6
7 2
11
8
1
3
1
4
7
6
9
8
10
2
11
5
1
3 5
2
67
4
10
8
3
1
1
10
4
5
6
7 2
11
8
1
Caudal Thalamus, Frontal Section
A–C Frontal section throughthe caudal thalamus
A Cellular staining (Nissl)
B Fiber staining (myelin) C Nuclear organization(according to Hassler)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

192Dienc
epha
lon
Diencephalon
Subthalamus
Subdivision (A)
The zona incerta (A1) (p. 174, B25) betweenForel’s field H1 (A2) and Forel’s field H2 (A3)(p. 174, B27, B28) is supposed to be a relaystation for descending fibers of the globuspallidus.
The subthalamic nucleus (Luys’ body) (A4)(p. 174, B26) between Forel’s field H2 andthe internal capsule (A5) has close connec-tions to the pallidum (A6), namely, afferentfibers from the outer segment of the globuspallidus and efferent fibers to the inner seg-ment of the globus pallidus. Bidirectionaltracts run to the tegmentum and to the con-tralateral subthalamic nucleus and globuspallidus (supramamillary commissure).
Clinical Note: In humans, injury to the sub-thalamic nucleus leads to hyperkinesis, whichmay develop into paroxysmal violent flingingmovements of the contralateral arm or even theentire body half (hemiballismus, Luys’ body syn-drome). In monkeys, the same symptom complexcan be induced by destroying the subthalamic nu-cleus.
The globus pallidus (Pallidium) (A6)(p. 174, AB6) is divided by amyelinated fiberlamella into an outer (lateral) and an inner(medial) segment. Both segments are con-nected by fibers to one another and to theputamen (A7) and the caudate nucleus (A8).Bidirectional connections exist to the sub-thalamic nucleus (A4), with the sub-thalamopallidal fibers (A9) terminating inthe inner segment and the pallidosub-thalamic fibers (A10) originating in the outersegment. Nigropallidal fibers (A11) run fromthe substantia nigra (A12) to the inner seg-ment of the pallidum.
The lenticular fasciculus (A13) emerges at thedorsal margin of the inner segment of thepallidum and forms Forel’s field H2 ventrallyto the zona incerta. The lenticular ansa (A14)(p. 174, A10) emerges from the ventral partof the inner segment and extends in an archthrough the inner capsule. Lenticularfasciculus and ansa unite to form thethalamic fasciculus (A15), which forms Forel’s
field H1 and radiates into the thalamus (ven-tral anterior nucleus, ventral lateral nucleus,medial nucleus). Fibers from the inner seg-ment of the pallidum run as pallidotegmen-tal tract (A16) into the tegmentum of themidbrain.
Clinical Note: Contrary to the old view thatParkinson’s disease (paralysis agitans) would re-sult from injury to the pallidum, destruction ofthe pallidum does not lead to motor disorders.Unilateral elimination of the pallidum in patientswith Parkinson’s disease removes muscle stiff-ness on the contralateral side and reduces tremor.Bilateral elimination causes mental disorders(postconcussional syndrome: irritability, fatiga-bility, difficulty in concentrating).
Responses to Stimulation of theSubthalamus (B)
Electrical stimulation results in an increasein muscle tone, increased reflex excitabilityand promotion of cortically induced move-ments.
Automatic motions can be induced in certainareas. As passing fiber bundles are also af-fected, the stimulatory effects cannot pro-vide information on the function of in-dividual nuclei. Stimulation at the level ofthe posterior commissure (fiber region ofthe interstitial nucleus) results in loweringthe head ( ); medial stimulation in theprerubral field results in lifting the head( ). The field for turning and rolling mo-tions ( ) corresponds to the fiber area ofthe superior cerebellar peduncle. The fieldfor ipsilateral turning motions ( ) corre-sponds to the dorsolateral tegmentalfasciculus (vestibulothalamic tract). Thearea for contralateral turning motions ( )corresponds to the zona incerta.
B17 Fornix.B18Mamillothalamic tract.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

193
Dienc
epha
lon
7
8
5
9 1
4 3
2
6
11 16
14
10
1315
12
17
18
Subdivision of the Subthalamus, Stimulation Results
A Fiber connections ofthe subthalamus
B Motor responses following stimulation of subthalamus and tegmentum;horizontal section through the diencephalon of the cat (according to Hess)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

194Dienc
epha
lon
Diencephalon
Hypothalamus
The hypothalamus forms the lowest layerand the floor of the diencephalon, con-sisting of the optic chiasm, the tuber cin-ereum tapering into the funnel-shaped in-fundibulum (hypophysial stalk), and themamillary bodies. The hypothalamus is thecentral region for the control of autonomicfunctions; it influences not only the auto-nomic nervous system but also the en-docrine system via its connection to the hy-pophysis, and it coordinates the two. Thehypothalamus is divided into two parts, thepoorly myelinated and the richly myelinatedhypothalamus.
Poorly Myelinated Hypothalamus(A–C)
This includes the preoptic region in front ofthe optic chiasm, the tuber cinereum, thelateral field (A1) lying dorsolaterally to thetuber cinereum, and the dorsocaudal area(B2) above themamillary bodies. The poorlymyelinated hypothalamus is the part of thebrain that is richest in peptidergic neurons.The different neuropeptides can be demon-strated in diffusely scattered cells (luliberin,cholecystokinin, thyoliberin), in per-iventricular cell aggregations (soma-tostatin), and in the nuclear areas (neu-ropeptides in various compositions).Numerous fiber connections within the hy-pothalamus and some of the long projectiontracts are peptidergic.
The preoptic region (C3) extends from theanterior commissure (C4) to the optic chiasm(C5) as a parvocellular field surrounding themost rostral excavation of the third ven-tricle, the preoptic recess. The region con-tains a large number of peptidergic neurons(containing primarily enkephalin). Twoprominent magnocellular nuclei lie adja-cent to it, namely, the supraoptic nucleus(AC6) and the paraventricular nucleus (AC7).The supraoptic nucleus borders on the optictract (A8). Its neurons contain cholecys-tokinin and dynorphin. The paraventricularnucleus lies close to the wall of the thirdventricle, separated from the ependyma
(the simple cell layer lining the walls of theventricular system, p. 284) only by a layer ofglial fibers, and extends as a narrow bandobliquely upward into the region of thezona incerta. Its peptidergic neurons havebeen shown to contain corticoliberin, neu-rotensin, cholecystokinin, as well as otherneuropeptides.
The principal nucleus of the tuber cinereum(A9) is the ventromedial nucleus (AC10), around body occupying most of the tubercinereum. It contains medium-sized neu-rons, among them many peptidergic (con-taining primarily neurotensin), and is sur-rounded by a delicate fiber capsule formedby the pallidohypothalamic fasciculus. Thedorsomedial nucleus (AC11) is less distinctand contains small neurons. At the base ofthe infundibulum lies the parvocellular in-fundibular nucleus (AC12). Its cells encirclethe infundibular recess and reach directlyinto the ependyma. The neurons of this nu-cleus contain primarily endorphin andACTH (adrenocorticotropic hormone).
Richly Myelinated Hypothalamus(B–C)
The mamillary body (BC13) forms the caudalsegment of the hypothalamus. It appears asa round bulge, a myelinated region that issurrounded by a prominent capsule. Thelatter is formed by afferent and efferentfiber tracts, namely, medially by fibers ofthemamillothalamic fasciculus (Vicq d'Azyr’sbundle) and themamillotegmental fasciculus(Gudden's tract), and laterally by fibers ofthe fornix.
The mamillary body is divided into a medialand a lateral nucleus. The large medial nu-cleus (B14) contains small neurons. It isspheroid and makes up the round body thatis visible at the base of the brain. The lateralnucleus (B15) is dorsolaterally attached tothe medial nucleus like a small cap. Themamillary body is surrounded by nuclearregions of the poorly myelinated hypo-thalamus, namely, the premamillary nucleus(C16) and the tuberomamillary nucleus(B17).
AC18 Fornix.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

195
Dienc
epha
lon
18
11
7
1
8
6 10
12 9
17
15
14
4
3
106
512
16
13
18
11
13
2
7
Poorly Myelinated and Richly Myelinated Hypothalamus
A Frontal section through the tuber cinereum(poorly myelinated hypothalamus)
B Frontal section through themamillary body (richly myelinatedhypothalamus)
C Lateral view of the hypothalamus(according to Diepen)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

196Dienc
epha
lon
Diencephalon: Hypothalamus
Vascular Supply (A)
The close association of the nervous systemwith the endocrine system is revealed bythe extraordinarily rich vascular supply ofindividual hypothalamic nuclei. The su-praoptic nucleus (A1) and the paraventricu-lar nucleus (A2) are about six times morevascularized than the rest of the gray mat-ter. Their neurons have close contact withthe capillaries, some of which are even en-closed by the neurons (endocellular capil-laries).
Fiber Connections of the PoorlyMyelinated Hypothalamus (B)
Numerous connections transmit olfactory,gustatory, viscerosensory, and soma-tosensory impulses to the hypothalamus.These are usually loose and divergent sys-tems, and almost all of them are bidirec-tional. The following are themost importantones.
The medial forebrain bundle (medial telen-cephalic fasciculus), containing the olfacto-hypothalamotegmental fibers (B3), con-nects almost all hypothalamic nuclei withthe olfactory centers and with the reticularformation of the midbrain. The bundle con-tains a large number of peptidergic fibers(VIP [vasoactive intestinal peptide],enkephalin, somatostatin).
The terminal stria (B4) runs in an arch aroundthe caudate nucleus and connects the amyg-daloid body (B5) (pp. 226, 228) with the pre-optic region (B6) and the ventromedial nu-cleus (B7). It is rich in peptidergic fibers.
The fibers of the fornix (BC8) (p. 232, B15, C)originate from the pyramidal cells of thehippocampus (BC9) and the subiculum(p. 232, A12) and terminate in the mamil-lary body (BC10). A large portion of the for-nix fibers are peptidergic. At the level of theanterior commissure, fibers branch off fromthe fornix to the preoptic region and to thetuberal nuclei (B7).
The dorsal longitudinal fasciculus, or Schütz’sbundle (p. 144, B) (B11), is the most impor-tant component of an extensive periventric-
ular fiber system. At the transition to themidbrain, fibers collect to form a compactbundle that connects the hypothalamuswith the nuclei of the brain stem.
Connections with thalamus and pallidum: Thehypothalamic nuclei are connected by peri-ventricular fibers with the medial nuclei ofthalamus (B12), the fibers of which projectto the frontal cortex. In this way, an indirectconnection is established between hypo-thalamus and frontal cortex. The pallidohy-pothalamic fasciculus runs from the globuspallidus to the tuberal nuclei (ventromedialnucleus).
Commissures: The commissures in the regionof the hypothalamus contain almost nofibers from hypothalamic nuclei; fibersfrom the midbrain and pons cross in thedorsal supraoptic commissure (Ganser’scommissure) and in the ventral supraopticcommissure (Gudden’s commissure), whilefibers from subthalamic nuclei cross in thesupramamillary commissure.
Fiber Connections of the RichlyMyelinated Hypothalamus (C)
Primarily afferent pathways of the mamil-lary body (BC10) are the fornix (BC8), thefibers of which terminate primarily in themamillary body, and the peduncle of themamillary body (C13) from the tegmental nu-clei of the midbrain, which is thought tocontain also gustatory fibers, vestibularfibers, and fibers from the medial lemnis-cus.
Primarily efferent pathways are repre-sented by the mamillothalamic fasciculus(Vicq d'Azyr’s bundle) (C14) (p. 174, B31)which ascends to the anterior nucleus of thethalamus (C15). The projection of the ante-rior nucleus to the cingulate gyrus estab-lishes a connection between the hypo-thalamus and the limbic association cortex.The mamillotegmental fasciculus (C16) termi-nates in the tegmental nuclei of the mid-brain. All pathways contain a high propor-tion of peptidergic fibers.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

197
Dienc
epha
lon
2
1
6
8
12
4
5
9
9
7
3
11
10
815
14
13
10 16
2
1
6
8
12
4
5
9
9
7
3
11
10
815
14
13
10 16
Vascular Supply and Fiber Connections
A Vascular supply of the hypothalamus in the rhesus monkey(according to Engelhardt)
B Fiber connections of the poorlymyelinated hypothalamus
C Fiber connections of the richlymyelinated hypothalamus
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

198Dienc
epha
lon
Diencephalon: Hypothalamus
Functional Topography of theHypothalamus
The centers of the hypothalamus influenceall processes that are important for the in-ternal environment of the body; they regu-late the performance of organs according tothe actual physical load, that is, they controlthe balance of temperature, water, and elec-trolytes; the activity of the heart; circula-tion and respiration; metabolism; and therhythm of sleeping and waking.
Vital functions (such as food intake,gastrointestinal activity and defecation,fluid intake and urination) are controlledfromhere, and so are the processes essentialfor the preservation of the species (procrea-tion and sexuality). These vital activities aretriggered by physical needs that are per-ceived as hunger, thirst, or sexual drive. In-stinctive activities serving the preservationof the organism are usually accompanied bya strong affective component, such as desireor aversion, joy, anxiety, or anger. Hypo-thalamic excitation plays an important rolein the creation of these emotions.
Dynamogenic and Trophotropic Zones(A–C)
Electric stimulation of the hypothalamusevokes autonomic reactions that can bedivided into two groups (A): those as-sociated with regeneration and metabolicprocesses (blue symbols: contraction of thepupils ; slowing down of respiration ;drop in blood pressure ; urination ; defe-cation ), and those associated with in-creased performance in response to the en-vironment (red symbols: dilation of thepupils ; accelerated respiration ; in-crease in blood pressure ). Stimulation re-sponses from specific regions have estab-lished a dorsocaudal and lateral region forthe dynamogenic mechanisms, the dynamo-genic zone (B1), and a ventro-oral region forthemechanisms promoting regeneration, thetrophotropic zone (B2). The two regionscorrespond to the subdivision of the periph-eral autonomic nervous system into a sym-pathetic component (dynamogenic) and aparasympathetic component (trophotropic)
(p. 292). In humans, controlled stimulationof the caudal hypothalamus (C) yielded sim-ilar results, namely, dilation of pupils (C3),increase in blood pressure (C4), and accel-erated respiration (C5).
Stimulation and Lesion Experiments(D–F)
Experiments of these types reveal the sig-nificance of defined regions for the regula-tion of certain processes; destruction of thetuber cinereum in the region of the infun-dibular nucleus (D6) in juvenile animalsleads to atrophy of the gonads. On the otherhand, lesions between optic chiasm andtuber cinereum in infantile rats result inpremature sexual development. Estrous cycleand sexual behavior are affected by lesionsto the hypothalamus. Lesions to the caudalhypothalamus between the tuber cinereumand the premamillary nucleus (E7) lead toadipsia (cessation of spontaneous drinking).More dorsal lesions result in aphagia (re-fusal to eat), while dorsal stimulation (F8)results in hyperphagia (compulsive eating).In the anterior hypothalamus at the level ofthe optic chiasm lies a region responsiblefor the control of body temperature. Stimula-tion in the vicinity of the fornix triggers fitsof rage and aggressive behavior (perifornicalzone of rage).
Clinical Note: In humans, pathologicalprocesses in the hypothalamus cause similarchanges (premature sexual development, bulimia,anorexia nervosa).
A9 Hypophysial stalk.B10Hypophysis.B11Mamillary body.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

199
Dienc
epha
lon
2
1
10
11
6 7
5 43
8
9
Functional Topography of the Hypothalamus
A Trophotropic and dynamogenic zones in thediencephalon of the cat, sagittal section,stimulation pattern (according to Hess)
B Trophotropic and dynamogeniczones in the cat, sagittal section
C Responses following stimulation in the caudalhypothalamus in humans, frontal section(according to Sano, Yoshioka, Ogishiwa, et al.)
D Gonadotropic region in cats andrabbits (according to Spatz,Bustamante, and Weisschedel)
E Adipsia, experimental lesionin the rat(according to Stevenson)
F Hyperphagia, stimulationexperiment in the cat(according to Brügge)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

200Dienc
epha
lon
Diencephalon
Hypothalamus andHypophysis
Development and Subdivision ofthe Hypophysis (A, B)
The hypophysis, or pituitary gland (see vol. 2),consists of two parts, namely, the adeno-hypophysis (anterior lobe) (AB1), whichdevelops from an evagination of the roof ofthe primitive foregut (Rathke’s pouch), andthe neurohypophysis (posterior lobe)(AB2), which represents an evagination ofthe floor of the diencephalon. The adenohy-pophysis is an endocrine gland, while theneurohypophysis is a part of the brain com-posed of nerve fibers, a capillary bed, and aunique type of glia, the pituicytes. The twoparts of the hypophysis adjoin each other,bringing the nervous system into close asso-ciation with the endocrine system.
Infundibulum (A–D)
The tuber cinereum tapers at its base toform the infundibulum (hypophysial stalk)(A3) and the infundibular recess (A4). Thefunnel-shaped descent of the infundibulum,where the contact between nervous and en-docrine systems is established, is also calledthemedian eminence of the tuber. A thin tissuelayer of the adenohypophysis reaches to thetuber cinereum and covers the anterior sideof the infundibulum (infundibular part of theadenohypophysis) (A5); several islets of tissuecover also its back. Thus, we can distinguisha proximal part of the hypophysis borderingon its tuber cinereum (infundibulum andinfundibular part of the adenohypophysis)and a distal part lying in the sella turcica(adenohypophysis with the intermediate part[A6, hypophysial cavity [A7], and neuro-hypophysis). The proximal contact surface isof special significance for the interconnec-tion of the nervous and endocrine systems.The outer glial fiber layer (A8), which sealsoff the rest of the brain surface, is absenthere, and portal capillaries (C) enter fromthe adenohypophysis into the infun-dibulum. These are vascular loops with anafferent and an efferent limb, which take
a convoluted and sometimes spiral course(D).
Blood Vessels of the Hypophysis(C–E)
A prominent vascular supply ensures thecoupling of the nervous and endocrine partsof the hypophysis. The afferent vessels, thesuperior hypophysial artery (E9) and the infe-rior hypophysial artery (E10), branch off fromthe internal carotid artery. The two superiorhypophysial arteries form an arterial ringaround the proximal part of the infun-dibulum, from where small arteries extendthrough the adenohypophysial cover intothe infundibulum and disperse into portalcapillaries (E11). The recurrent limbs of thelatter collect in the portal veins (E12), whichtransport the blood to the capillary bed ofthe adenohypophysis. The trabecular arteries(E13) extend to the adenohypophysis, as-cend caudally, and supply the distal seg-ment of the infundibulum. The blood thenruns from the capillary bed of the adenohy-pophysis into the veins.
The two inferior hypophysial arteriessupply the neurohypophysis; with severalbranches in the area of the intermediatepart, they also form special vessels (E14);from here, the blood also runs via short por-tal vessels into the capillary bed of the ade-nohypophysis.
Thus, the adenohypophysis receives nodirect arterial supply. The latter goes intothe infundibulum and into the neurohy-pophysis, from where the blood flows intothe adenohypophysis via the portal vesselsand, only then, drains into the venous side(E15) of the circulation.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

201
Dienc
epha
lon
8
57 3
621
4
21
35
5
3 9
11
11
12
15
1015
1 2
14
13
11
Development and Blood Vessels of the Hypophysis
A Hypophysis and infundibulum(according to Christ) B Development
C Specialized vessels of the infun-dibulum (according to Christ)
D Specialized vessel(according to Sloper)
E Hypophysial blood vessels(according to Xuereb, Prichard,and Daniel)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

202Dienc
epha
lon
Diencephalon: Hypothalamus and Hypophysis
Neuroendocrine System (A–D)
The hypophysis is under the control of thehyopothalamic centers. Unmyelinated fiberbundles from the hypothalamus run in thehypophysial stalk and in the hypophysialposterior lobe. Severing the hypophysialstalk leads to retrograde cellular changes,namely, to extensive loss of neurons in thenuclei of the tuber cinereum when transec-tion is carried out at a high level.
Neurons in the hypothalamus produce sub-stances that migrate within the axons intothe hypophysis and enter there into thebloodstream. This endocrine function ofneurons is called neurosecretion. The sub-stances are produced in the perikarya andappear there as small secretory droplets(B1). The cells, which are genuine neuronswith dendrites and axons, represent a tran-sitional stage between neurons andsecretory cells. Both these cell types are ofectodermal origin and are closely relatedwith respect to physiology and metabolism.Both of them produce a specific substancethat they secrete in response to a nervous orhumoral stimulus: the neurons (A2) releasetransmitter substances (neurotransmitters)and the secretory cells (A3) release theirsecretion. Transitional forms between thetwo cell types are the neurosecretory cells(A4) and the endocrine cells A5), both ofwhich release their secretion into the blood-stream.
Hypothalamohypophysial fiber tracts.Corresponding to the structure of the hy-pophysis with an anterior lobe (adenohy-pophysis) and a posterior lobe (neurohy-pophysis), there are two different fiber sys-tems extending from the hypothalamus tothe hypophysis, namely, the tuberoinfun-dibular tract and the hypothalamohypophy-sial tract. In both, the coupling of the neuralsystem with the endocrine system isachieved by the sequential arrangement ofnerve fibers and capillaries (neurovascularchain).
Tuberoinfundibular System (C, D)
The tuberoinfundibular tract (D) consistsof thin nerve fibers that originate in thetuberal nuclei, namely, the ventromedial nu-cleus (D6), the dorsomedial nucleus (D7), andthe infundibular nucleus (D8), and extendinto the hypophysial stalk. The substancesproduced in the perikarya enter from theaxon terminals into the portal capillaries(D9) and pass through the portal veins(D10) into the capillary bed of the adenohy-pophysis. They are stimulating substances,releasing factors, which cause the release ofglandotropic hormones (messengers thataffect other endocrine glands) by the ade-nohypophysis (see vol. 2).
The production of specific releasing factorscannot be attributed to individual hy-pothalamic nuclei. The regions from whichan increased secretion can be induced byelectrical stimulation (C) do not correspondto the tuberal nuclei. Stimulation of the pre-optic region (C11) results in an increasedsecretion of luteotropic hormone. Stimula-tion caudally to the optic chiasm (C12) leadsto the release of thyrotropic hormone, andstimulation of the ventral hypothalamus(tuber cinereum to mamillary recess) (C13)leads to the release of gonadotropic hor-mone. In addition to releasing factors, inhib-iting factors, inhibiting factors have beenidentified that block the release of hor-mones in the adenohypophysis.
CD14 Optic chiasm.CD15 Mamillary body.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

203
Dienc
epha
lon
2
4
5
3
1
117
6
15
14
7
6
8
15
10 9
12
13
14
Neurosecretion, Tuberoinfundibular System
A Nerve cells and secretory cells B Neurosecretory cells in humans(according to Gaupp and Scharrer)
C Regions where stimulation trig-gers the secretion of hypophysialhormones (according to Harris)
D Tuberoinfundibular tract
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

204Dienc
epha
lon
Diencephalon: Hypothalamus and Hypophysis
Neuroendocrine System(continued)
Hypothalamohypophysial System (A–D)
The hypothalamohypophysial tract (D) consistsof the supraopticohypophysical tract and theparaventriculohypophysial tractwhich origi-nate in the supraoptic nucleus (D1) and inthe paraventricular nucleus (D2), respec-tively. The fibers run through the hypophy-sial stalk into the hypophysial posterior lobewhere they terminate at the capillaries. Thehormones produced by the neurons of bothhypothalamic nuclei migrate along thispathway to the axon terminals and enterfrom here into the bloodstream. Electricalstimulation of the supraoptic nucleus (C3)leads to an increased secretion of vasopres-sin (antidiuretic hormone), while stimula-tion of the paraventricular nucleus (C4)leads to an increased secretion of oxytocin.In this system, the neurons do not releasestimulating substances that affect the secre-tion of a hormone by an endocrine gland(such as the glandotropic hormones or re-leasing factors of the tuberoinfundibularsystem), but they themselves produce hor-mones that have a direct effect on the targetorgans (effector hormones). The carrier sub-stances to which the hormones are boundduring their migration in the axons can bedemonstrated histologically. These Gomori-positive substances often cause swellings ofthe axons (Herring bodies) (B5).
The neurosecretory substances in axons andswellings appear in the electron-micro-scopic image as granules that are muchlarger than synaptic vesicles. At the capillar-ies of the neurohypophysis, the axons formclub-shaped endings (AD6) containingsmall, clear synaptic vesicles in addition tothe large granules. At the sites of contactwith axon terminals, the capillary walls lackthe glial covering layer that, in the centralnervous system, forms the boundary be-tween ectodermal and mesodermal tissuesand envelops all vessels (p. 44). It is herethat the neurosecretory product enters thebloodstream. At the terminal bulbs of theneurosecretory cells, there are also syn-apses (A7) of unknown origin, which
nevertheless certainly influence the releaseof the hormones.
Presumably the regulation of neurosecretion isachieved not only via synaptic contacts butalso via the bloodstream. The exceptionallyrich vascularization of hypothalamic nucleiand the existence of endocellular capillariessupport this hypothesis. This arrangementprovides a pathway for humoral feedbackand forms a regulatory circuit for control-ling the production and secretion of hor-mones, consisting of a neural limb (su-praopticohypophysial tract) and a humorallimb (circulation).
CD8 Optic chiasm.CD9 Mamillary body.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

205
Dienc
epha
lon
67
54
38
9
2
9
1
8
6
Hypothalamohypophysial System
A Supraopticohypophysial tract,electron-microscopic diagram(according to Bargmann)
C Regions where stimulation trig-gers the secretion of hypophysialhormones (according to Harris)
B Herring bodies(according to Hild)
D Hypothalamohypophysialtract
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Telencephalon
Overview 208Sections Through the
Telencephalon 214Paleocortex and Amygdaloid
Body 224Archicortex 230Neostriatum 236Insula 238Neocortex 240Imaging Procedures 264
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

208Te
lenc
epha
lon
Telencephalon
Overview
Subdivision of the Hemisphere(A, B)
The embryonic hemispheric vesicle (A)clearly shows the subdivision of the telen-cephalon into four parts, some of whichdevelop early (phylogenetically old por-tions), while others develop late (phylo-genetically new portions). The four parts arethe paleopallium, the striatum, the neopal-lium, and the archipallium.
The hemispheric wall is called the pallium,or brainmantle, because it covers the dien-cephalon and brain stem and envelops themlike a mantle.
The paleopallium (blue) (AB1) is the oldestportion of the hemisphere. It forms the floorof the hemisphere and corresponds, withthe olfactory bulb (A2) and adjacent paleo-cortex (p. 224ff), to the olfactory brain, orrhinencephalon, in the narrower sense. Theneostriatum (deep yellow) (AB3) (p. 236)develops above the paleopallium; it, too, ispart of the hemispheric wall, although itdoes not appear on the outer aspect of thehemisphere.
The largest area is made up by the neopal-lium (light yellow). Its outer aspect, the neo-cortex (p. 240ff) (AB4), develops very lateand encircles ventrally a transitional area tothe paleocortex that lies over the striatum;this is the insula (p. 238) (B14).
The medial hemispheric wall is formed bythe archipallium (red) (AB5), an old por-tion of the brain; its cortical band, the archi-cortex (p. 230ff), later curls up to form thehippocampus (Ammon’s horn).
The relationships in the mature brain aredetermined by themassive expansion of theneocortex, which pushes the paleocortexand the transitional cortex of the insula intothe deeper parts of the brain. The archicor-tex becomes displaced caudally and appearson the surface of the corpus callosum onlyas a thin layer (B5, F10).
Rotation of the Hemisphere (C–F)
The hemispheric vesicle does not expandevenly in all directions during its develop-ment but widens primarily in caudal andbasal directions. The temporal lobe isformed in this way, and it finally turns ros-trally in a circular movement (C); to a lesserdegree, such a rotation can also be observedwith the frontal lobe. The axis aroundwhichthe hemispheric vesicle rotates is the insu-lar region; like the putamen (E6) lyingbeneath it, the insula does not participate inthe movement. Other structures of thehemisphere, however, follow the rotationand end up having an arched shape in themature brain. The lateral ventricle (D7)forms such an arch with its anterior and in-ferior horns. The lateral portion of the stri-atum, the caudate nucleus (E8), participatesin the rotation as well and follows preciselythe arched shape of the lateral ventricle. Themain part of the archipallium, the hippo-campus (F9), moves from its original dorsalposition in basal direction and comes to liein the temporal lobe. The remnants of thearchipallium on the dorsal aspect of the cor-pus callosum, the indusium griseum (F10),and the fornix (F11) reflect the arched ex-pansion of the archipallium. The corpus cal-losum (F12) also expands in caudal directionbut follows the rotation only partially as itdevelops only late toward the end of thisprocess.
D13 Third ventricle.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

209
Telenc
epha
lon
3
4
1
14
2
5
1
5
13
7
8
6
12
9
10 11
3
4
1
14
2
5
1
5
13
7
8
6
12
9
10 11
Subdivision and Rotation of the Hemisphere
A, B Subdivision of the hemispheres
A Embryonic brain B Adult brain
C Rotation of the hemisphere(according to Jacob and Spatz)
D Ventricles
E Caudate nucleus and putamen
F Hippocampus (archipallium)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

210Te
lenc
epha
lon
Telencephalon: Overview
Evolution (A–D)
During primate evolution, the telen-cephalon has undergone changes similar tothose taking place during human embry-onic development; it developed late andthen overgrew the other parts of the brain.Thus, the cerebellum (A1) is still completelyexposed in the brain of primitive mammals(hedgehog), while it becomes more andmore covered by the hemispheres of the tel-encephalon during primate evolution.
The paleopallium (rhinencephalon) (blue)(A–C2) with olfactory bulb (A–C3) and pir-iform lobe (A–C4) forms the largest part ofthe hemisphere in the primitive mam-malian brain (A), and the archipallium (red)(A–D5) still has its original dorsal positionabove the diencephalon. These two oldcomponents of the hemisphere then be-come overgrown by the neopallium (yellow)(A–D6) during the course of evolution. Thepaleopallium of prosimians (C) is still ofconsiderable size. In humans (D), however,it becomes displaced deep into the base ofthe brain and no longer appears in thelateral view of the brain. The archipallium(hippocampus), which lies above the dien-cephalon in the hedgehog (A5), appears as apart of the temporal lobe at the base of thebrain in humans (D5). Only a narrow rem-nant remains above the corpus callosum(indusium griseum).
The positional changes largely correspondto the rotation of the hemisphere duringembryonic development; they also lead tothe formation of the temporal lobe (B–D7).While still absent from the brain of thehedgehog (A), the temporal lobe is alreadyrecognized as a ventrally directed projec-tion in the brain of the tree shrew (Tupaia),the most primitive of primates (B). In theprosimian brain (C), a caudally directedtemporal lobe has developed that finallyturns rostrally in the human brain (D). In ad-dition, sulci and gyri develop in the regionof the neopallium. Whereas the neopalliumof primitive mammals is smooth (lissen-cephalic brains), a relief of convolutionsdevelops only in higher mammals (gyren-cephalic brains). The development of sulci
and gyri considerably enlarges the surfaceof the cerebral cortex. In humans, only one-third of the cortical surface lies at the sur-face of the hemispheres, two-thirds lie deepin the sulci.
Two types of cortical areas can be distin-guished on the neocortex: the primary areasof origin (light red) and termination areas(green) of long pathways, and betweenthem the secondary association areas (yel-low).
The area of origin of motor pathways, themotor cortex (A–D8), constitutes the entirefrontal lobe in the hedgehog. An associationarea (B–D9) appears for the first time inprimitive primates (Tupaia) and achievesextraordinary expansion in the humanbrain. The termination area of sensory path-ways, the sensory cortex (A–D10), borderscaudally on the motor cortex. Owing to theenlargement of the adjacent associationarea, most of the termination area of thevisual pathway, the visual cortex (A–D11),becomes displaced to the medialhemispheric surface in humans. The termi-nation area of the acoustic pathway, theauditory cortex (CD12), becomes displaceddeep into the lateral sulcus (fissure of Syl-vius) by the expansion of the temporal asso-ciation areas. Thus, the association areas ex-pand much more during evolution than theprimary areas; they represent the largestpart of the neocortex in humans.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

211
Telenc
epha
lon
10 11 6 583
6
1
1
1
1
6
6
6
7 12
27425
5
5
611108
93
4 22
12 11
11
610
7
8
9
3
108
9
242
6
Evolution
A Hedgehog
B Tupaia
C Lemur
D Homo sapiens
A–D Evolution of the telencephalon (modified after Edinger, Elliot Smith, and Le Gros-Clark)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

212Te
lenc
epha
lon
Telencephalon: Overview
Cerebral Lobes (A–C)
The hemisphere is divided into four cere-bral lobes:
! The frontal lobe (red) (p. 246)! The parietal lobe (light blue) (p. 250)! The temporal lobe (dark blue) (p. 252)! The occipital lobe (purple) (p. 254)
The hemispheric surface consists of grooves,or sulci, and convolutions, or gyri. We distin-guish primary, secondary, and tertiary sulci.The primary sulci appear first and areequally well developed in all human brains(central sulcus, calcarine sulcus). The sec-ondary sulci are variable. The tertiary sulciappear last, being irregular and different ineach brain. Thus, each brain has its own sur-face relief as an expression of individuality,like the features of the face.
The frontal lobe extends from the frontalpole (AC1) to the central sulcus (AB2), whichtogether with the precentral sulcus (A3) de-fines the precentral gyrus (A4). The latter isgrouped with the postcentral gyrus (A5) toform the central region, which spreads be-yond the edge of the hemisphere (AB6) to theparacentral gyrus (B7). Furthermore, thefrontal lobe exhibits three major convolu-tions: the superior frontal gyrus (A8), themiddle frontal gyrus (A9), and the inferiorfrontal gyrus (A10 ); they are separated bythe superior frontal sulcus (A11) and the in-ferior frontal sulcus (A12). Three parts aredistinguished at the inferior frontal gyrusthat define the lateral sulcus (sulcus of Syl-vius) (AC13): the opercular part (A14), thetriangular part (A15), and the orbital part(A16).
The parietal lobe adjoins the frontal lobewith the postcentral gyrus (A5) which is de-fined caudally by the postcentral sulcus(A17). This is followed by the superiorparietal lobule (A18) and the inferior parietallobule (A19), which are separated by the in-traparietal sulcus (A20). The end of thelateral sulcus is surrounded by the supra-marginal gyrus (A21); the angular gyrus(A22) lies ventrally to it. The medial surfaceof the parietal lobe is formed by the pre-cuneus (B23).
The temporal lobe includes the temporalpole (AC24) and three major convolutions:the superior temporal gyrus (A25), themiddle temporal gyrus (A26), and the inferiortemporal gyrus (AC27), which are separatedby the superior temporal sulcus (A28) andthe inferior temporal sulcus (A29). The trans-verse temporal gyri (Heschl ’s convolutions)of the dorsal aspect of the temporal lobe liein the depth of the lateral sulcus (p. 252, C).On the medial surface is the parahippocam-pal gyrus (BC30) whichmerges rostrally intothe uncus (BC31) and caudally into the lin-gual gyrus (BC32). It is separated by the col-lateral sulcus (BC33) from the middle occipi-totemporal gyrus (BC34). Ventrally lies thelateral occipitotemporal gyrus (BC35),delimited by the occipitotemporal sulcus(BC36).
The occipital lobe includes the occipitalpole (A–C37) and is crossed by the trans-verse occipital sulcus (A38) and the deep cal-carine sulcus (B39). Together with theparieto-occipital sulcus (B40), the latter de-fines the cuneus (B41).
The cingulate gyrus (limbic gyrus) (green)(B42) extends around the corpus callosum(B43). Caudally, it is separated by the hippo-campal sulcus (B44) from the dentate gyrus(dentate band) (B45) and tapers rostrallyinto the paraterminal gyrus (B46) and intothe subcallosal area (parolfactory area)(B47). Isthmus of cingulate gyrus (B48).
Base of the brain. The basal aspect of thefrontal lobe is covered by the orbital gyri(C49). Along the edge of the hemisphereruns the gyrus rectus (C50), laterally definedby the olfactory sulcus (C51) into which theolfactory bulb (C52) and the olfactory tractare embedded. The olfactory tract splits intothe two olfactory striae which embrace theanterior perforated substance (olfactoryarea) (C53).
C54Hippocampal sulcus.C55 Longitudinal cerebral fissure.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

213
Telenc
epha
lon
172311
89
45
2118
20
19
22
25
1326
27
15
14
16
24373828
29
10
2
6
47 46
43
427
23
40
41
3239
37
48
33
1327
35
34
30
49
51
1
52
24
53
50
54
31 37
55
32
36
3336
45
304431
34
35
12
6
1
Cerebral Lobes
A Lateral view of the hemisphere
B Median view of the hemisphere
C Basal view of the two hemispheres
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

214Te
lenc
epha
lon
Telencephalon
Sections Through theTelencephalon
Frontal Sections
The posterior cut surface is shown for eachbrain section.
Section at the Level of the Exit of theOlfactory Tract (A)
The cut surface shows the two hemispheresseparated by the cerebral longitudinal fissure(AB1); the gray matter (cortex and nuclei) iseasily distinguished from the white matter(myelinated fiber masses). The corpus callo-sum (AB2) connects the two hemispheres.The section shows the cingulate gyrus (AB3)above the corpus callosum.
The lateral field of the section shows thedeep lateral sulcus (AB4). Dorsally to it liesthe frontal lobe with the superior frontalgyrus (AB5), the middle frontal gyrus (AB6),and the inferior frontal gyrus (AB7). They areseparated by the superior frontal sulcus(AB8) and the inferior frontal sulcus (AB9).Ventrally to the lateral sulcus lies the tem-poral lobe with the superior temporal gyrus(AB10), the middle temporal gyrus (AB11),and the inferior temporal gyrus (AB12). Thetemporal gyri are separated by the superiortemporal sulcus (AB13) and inferior temporalsulcus (AB14). The lateral sulcus expandsdeep into the lateral fossa (fossa of Sylvius)(AB15), on the inner surface of which is theinsula. The insular cortex extends basally al-most to the exit of the olfactory tract (A16).It represents a transitional area betweenpaleocortex and neocortex.
In the depth of the hemisphere lies the neo-striatum which is divided by the internalcapsule (AB17) into the caudate nucleus(AB18) and the putamen (AB19). The sectionshows the anterior horn (AB20) of the lateralventricle. The lateral wall of the ventricle isformed by the caudate nucleus, while itsmedial wall is formed by the septum pel-lucidum (AB21) containing the cavity of theseptum pellucidum (AB22). At the lateralaspect of the putamen lies a narrow, cup-shaped layer of gray matter, the claustrum(AB23). It is separated from the putamen bythe external capsule (AB24) and from the in-sular cortex by the extreme capsule (AB25).
Section at the Level of the AnteriorCommissure (B)
At this level, the section shows the centralregions of the frontal lobe and the temporallobe. The lateral fossa is closed, and the in-sula is covered by the frontal operculum(AB26) and the temporal operculum (AB27).The ventral regions of both hemispheres areconnected by the anterior commissure (B28)where fibers of the paleocortex and thetemporal neocortex cross. Above the com-missure appears the globus pallidus (B29)(part of the diencephalon), and close to themidline lies the septum pellucidum (AB21),or more specifically, its wide ventral seg-ment containing the septal nuclei (alsoknown as peduncle of the septum pel-lucidum). The mediobasal aspect of thehemisphere is covered by the paleocortex,the olfactory cortex (B30).
Claustrum. In the past, the claustrum(AB23) was either grouped together withthe striatum to form the so-called basal gan-glia or was assigned to the insular cortex asan additional cortical layer. Developmentalstudies and comparative anatomical inves-tigations, however, suggest that it consistsof cell clusters of the paleocortexwhich havebecome displaced during development. Theclaustrum merges with its wide base intopaleocortical regions (namely, the prepiri-form cortex and the lateral nucleus of theamygdaloid body). Unmyelinated fibersfrom the cortices of parietal, temporal, andoccipital lobes are thought to terminate inthe claustrum in a topical arrangement. Thefunction of the claustrum is largely un-known.
B31Optic chiasm.
Planes of sectionsA BKahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

215
Telenc
epha
lon
1
8
5
6
9
717
19
18
15
16
1
5
8
96
7
26
427
25
2423
31
3028
29
2119
18
15
12
14
11
1310
1722 202
3
1
252423 27
15
4
26
22 221 20
3
10
13
11
1412
Frontal Sections
A Frontal section at the exit of the olfactory tract
B Frontal section at the level of the anterior commissure
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

216Te
lenc
epha
lon
Telencephalon: Sections
Frontal Sections (continued)
Section at the Level of the AmygdaloidBody (A)
At this level the central sulcus (AB1), whichruns obliquely from dorsocaudal to ven-trorostal, has been cut in the more rostralpart; the frontal lobe, which is dorsal to it,therefore occupies a far larger part of thesection than the parietal lobe, which is ven-tral to it. The convolution above the centralsulcus is the precentral gyrus (AB2); the con-volution below it is the postcentral gyrus(AB3). Deep in the temporal lobe appearsthe amygdaloid body (amygdala) (A4). Itreaches the surface at the medial aspect ofthe temporal lobe and might therefore beregarded partly as cortex, partly as nucleus,or rather as a transition between the twostructures. Since not only the surroundingperiamygdalar cortex but also its cortico-medial half belong to the primary olfactorycenters, the amygdaloid body can be as-signed to the paleocortex, despite its nu-clear features. The claustrum (AB5) endsabove this region with a wide base.
Between the hemispheres lies the dien-cephalon with thalamus (AB6), globus pal-lidus (AB7), and hypothalamus (A8). Later-ally to the diencephalic nuclei border theneostriatum with putamen (AB9) and cau-date nucleus (AB10). Below the corpus callo-sum (AB11) lies a strong fiber bundle, thefornix (AB12). Also seen are the longitudinalcerebral fissure (AB13), the lateral cerebralsulcus (AB14), the lateral fossa (AB15), theoptic tract (A16), and the infundibulum(A17).
Section at the Level of the Hippocampus(B)
Once the more caudally cut sections nolonger show the amygdaloid body, the hip-pocampus (B18) appears in the medial areaof the temporal lobe. This most importantportion of the archicortex is a cortical for-mation that has curled up and projectsagainst the inferior horn of the lateral ven-tricle (B23). The section also shows thecaudal part of the lateral fossa (B15). The
inner surface of the temporal operculum ex-hibits prominent convolutions; these arethe obliquely cut transverse temporal gyri(B19), or Heschl’s convolutions, repre-senting the auditory cortex. In the ventralregion of the diencephalon lie the sub-thalamic body (B20), the mamillary body(B21), and the substantia nigra (B22), whichis a part of the midbrain.
Basal Gaglia. The gray nuclear complexesdeep in the hemisphere are collectivelyknown as basal ganglia. Some authors usethe term only for the striatum and the pal-lidum, while others include the amygdaloidbody and the claustrum, some even thethalamus. As this term is vague and ill-de-fined, it is not used in the present descrip-tion. Earlier anatomists viewed the pal-lidum and the putamen as parts of the len-tiform nucleus (a concept still surviving aslenticular ansa and lenticular fasciculus), aterm that is no longer used.
Planes of sectionsA B
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

217
Telenc
epha
lon
2 1
3
14
15
16
7
178
13
10
21
3
9
6
7
5
4
1211
13
10
9
5
20
18
21
22
76
1211
23
14
19
15
1
2
3
Frontal Sections
A Frontal section at the level of the amygdaloid body
B Frontal section at the level of the hippocampus
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

218Te
lenc
epha
lon
Telencephalon: Sections
Frontal Sections (continued)
Section at the Level of Midbrain and Pons(A)
The caudal portion of the lateral fossa (A1) isopen to the lateral aspect of the hemi-sphere. Dorsally to the lateral sulcus (A2)lies the parietal lobe, ventrally the temporallobe. The dorsal convolutions of the latter,which lie deep in the lateral sulcus and rep-resent the transverse temporal gyri (A3)(p. 252, C1), are cut obliquely. At the bottomof the lateral fossa lies the insular cortex,which rests on the caudal extensions ofclaustrum (A4) and putamen (A5). The cau-date nucleus (A6) appears at the lateral wallof the lateral ventricle (A7). At the medialaspect of the temporal lobe, concealed bythe parahippocampal gyrus (A8), the cortexcurls up to form the hippocampus (Ammon’shorn) (A9). Corpus callosum (A10) and for-nix (A11) are seen above the choroid plexus.
The field between the hemispheres repre-sents the transition between diencephalonand midbrain. The section shows the caudalnuclear regions of the thalamus (A12). Sepa-rated from the main complex lies the lateralgeniculate body (A13), and medially to theventricular wall lies the habenular nucleus(A14). The plane of section has beenoriented according to Forel’s axis (p. 4, B),thus showing telencephalon and dien-cephalon in frontal section, while the struc-tures of midbrain and pons (Meynert’s axis;p. 4, B) have been cut obliquely. Ventral tothe aqueduct (A15) lies the decussation ofthe superior cerebellar peduncle (A16). A nar-row strip of dark cells, the substantia nigra(A17), extends ventrally on both sides. Thecerebral peduncles (A18) are seen laterally toit; the course of their fiber masses can betraced from the internal capsule to the pons(A19).
Section at the Level of the Splenium ofthe Corpus Callosum (B)
In this section, the dorsal part of the hemi-sphere belongs to the parietal lobe and theventral part to the temporal lobe; at thisplane of section, the latter is merging intothe occipital lobe. The boundary between
parietal lobe and temporal lobe lies in theregion of the angulate gyrus (B20). Thelateral sulcus and the lateral fossa are nolonger present in the section. The cut sur-face of the corpus callosum is particularlywide at the level of the splenium (B21)(p. 220, A6; p. 260, E14). Dorsally and ven-trally to it lies the cingulate gyrus (B22),which encircles the splenium in an arch. Theparahippocampal gyrus (B23) adjoins ven-trally. Neither the hippocampus nor the cal-carine sulcus are present in the section;hence, the section lies behind the hippo-campus but in front of the calcarine sulcus.The two lateral ventricles are remarkablywide, each representing the most anteriorpart of the posterior horn at the transitioninto inferior horn and central part (seep. 281, BC7 – 9).
The lower aspects of the hemispheresborder on the cerebellum. The medulla ob-longata appears in the middle, the obliquesection shows the fourth ventricle (B24),the olives (B25), and the pyramids (B26).
Planes of sections A B
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
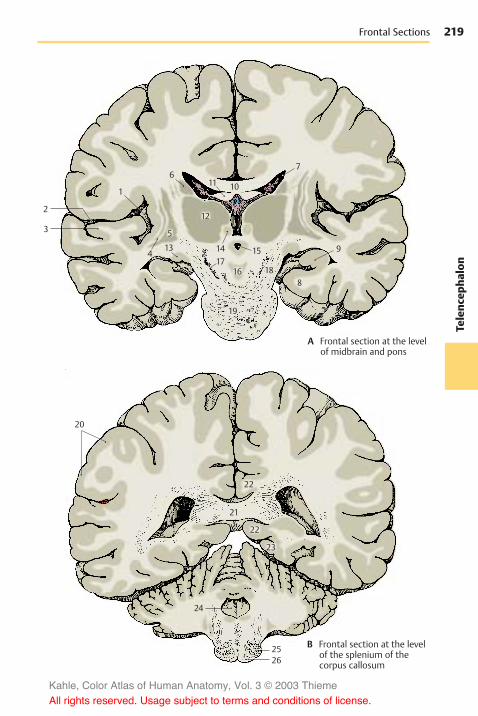
219
Telenc
epha
lon
61011
12
1417
16
15
18
19
20
22
22
21
23
24
2526
8
9
7
5
134
1
2
3
Frontal Sections
A Frontal section at the levelof midbrain and pons
B Frontal section at the levelof the splenium of thecorpus callosum
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

220Te
lenc
epha
lon
Telencephalon: Sections
Horizontal Sections
Superior Aspect of Corpus Callosum andLateral Ventricles (A)
The horizontal section through the brainhas been cut above the corpus callosum, andthe superior aspect of the corpus callosumand the lateral ventricles have been exposedby removal of deeper portions of whitemat-ter. The section shows the frontal lobes (A1)at the top, the temporal lobes (A2) on bothsides, and the occipital lobes (A3) at thebottom. The superior surface of the corpuscallosum (A4) belongs to the free brain sur-face lined by the pia mater and arachnoidea.Lying deep in the brain, it is covered by theconvolutions of the medial walls of thehemispheres. Rostrally, the superior surfaceof the corpus callosum turns in ventraldirection and forms the genu of the corpuscallosum (A5) (p. 260, E11); caudally, itforms the splenium of the corpus callosum(A6) (p. 260, E14). On the superior aspect ofthe corpus callosum extend four myelinatedfiber ridges: one lateral longitudinal stria(A7) and one medial longitudinal stria ofLancisi (A8) run along each half of the cor-pus callosum (see p. 230). Their fiber tractsextend from the hippocampus to the sub-callosal area. Between the two longitudinalstriae lies a thin layer of gray matter con-sisting of a narrow layer of neurons, the in-dusium griseum. This is a cortical portion ofthe archicortex that regressed as a result ofthe extensive development of the corpuscallosum (p. 7, E) and subsequent displace-ment of the archicortex into the inferiorhorn of the lateral ventricle (see p. 209, F).
The anterior horns (A9) of the lateral ven-tricles (p. 280, A1) are opened in the area ofthe frontal lobes, and the posterior horns(A10) in the area of the occipital lobes. Theprotruding hippocampus (A11) forms thefloor of the inferior horn. The central partand the inferior horn of the lateral ventriclecontain the choroid plexus (A12) (p. 282).
Exposure of the Roof of the Diencephalon(B)
This is an oblique horizontal section belowthe corpus callosum, which has beencompletely removed. Upon opening the twolateral ventricles, the dorsal aspect of thecaudate nucleus (B13) and, bordering medi-ally, the dorsal aspect of the thalamus (B14)become visible. Parts of the diencephalonbecome exposed as well, namely, the pinealgland (B15) and both habenulae (B16) whichare connected to it. The two fornices (B17)between the heads of the two caudate nu-clei have been cut in their rostral part(columns of fornix). The septum pellucidum(B18) extends from there to the corpus cal-losum.
The lateral wall of the hemisphere containsa particularly wide medullary layer be-tween the cortex and the ventricle, thesemioval center (B19). The central sulcus(B20) cuts into it and separates the frontallobe (at the top of the figure) from theparietal lobe (bottom). Starting from thecentral sulcus, the precentral gyrus (B21)and the postcentral gyrus (B22) can be lo-cated.
Caudally in the longitudinal cerebral fissure(AB23), the cerebellum (B24) is visible. Thecaudal portion of the hemisphere is formedby the occipital lobe. The striate area (B25),the visual cortex, lies in this region and oc-cupies primarily the calcarine sulcus (B26) atthe medial aspect of the occipital lobe,while extending only a short distance ontothe occipital pole. It can be distinguishedeven by the naked eye from the rest of thecortex through a white streak, the line ofGennari (B27), which divides the cortex intotwo gray bands. Gennari’s line is a wideband of myelinated nerve fibers corre-sponding to the slightly narrower externalband of Baillarger in the other areas of theneocortex (see p. 240, A16; p. 254).
B28Mesencephalic tectum.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

221
Telenc
epha
lon
10
12
6
2
11
9
1
4
78
5
23
3
18
23
1713
2120
22
25
1516
19
14
28
24
26
27
10
12
6
2
11
9
1
4
78
5
23
3
18
23
1713
2120
22
25
1516
19
14
28
24
26
27
Horizontal Sections
A Horizontal section withsuperior surface of thecorpus callosum
B Horizontal section exposingthe roof of the diencephalon
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

222Te
lenc
epha
lon
Telencephalon: Sections
Horizontal Sections (continued)
Horizontal Section through theNeostriatum (A)
At this level, the lateral cerebral fossa (AB1)is exposed in its longitudinal expansion. Thelateral sulcus (A2) is found more rostrally,with the frontal operculum (AB3) in front ofit and the elongated temporal operculum(AB4) caudally to it. The longitudinal expan-sion is also apparent in the deep structuresof the telencephalon, the claustrum (AB5)and the putamen (AB6). The arched struc-tures have been cut twice; the corpus callo-sum (A7) appears rostrally with its anteriorpart, the genu of the corpus callosum, andcaudally with its end, the splenium. The cau-date nucleus has been cut twice as well; thehead of the caudate nucleus (AB8) is seenrostrally and the tail of the caudate nucleus(AB9) caudolaterally to the thalamus(AB10). The thalamus is separated from theglobus pallidus (AB11) by the internal cap-sule which, in horizontal sections, exhibitsthe shape of a hook made up of the anteriorlimb (AB12) and the posterior limb (AB13).Also the lateral ventricle has been exposedtwice. Its anterior horn (A14) has been cut inthe area of the frontal lobe and, caudally, inthe transition to the posterior horn (A15).The two anterior horns are separated by theseptum pellucidum (A16), which spans be-tween corpus callosum and fornix (A17).
The section also shows the frontal lobes(AB18), the temporal lobes (AB19), theoccipital lobes (A20), the longitudinal cere-bral fissure (AB21), and the striate area(visual cortex) (A22).
Horizontal Section at the Level of theAnterior Commissure (B)
While the section still shows the entirefrontal lobe and temporal lobe, the occipitallobe has only been cut in its anterior part atthe transition to the temporal lobe. Betweenthe two hemispheres appears the cone-shaped dorsal aspect of the cerebellum(B23). The anterior horn of the lateral ven-tricle and the corpus callosum are no longerseen in this section. Instead there is the
anterior commissure (B24) connecting thetwo hemispheres. The two columns of thefornix (B25), lying close together in the pre-vious section, are separated at the level ofthe anterior commissure. While the poste-rior limb of the internal capsule (AB13) re-tains its usual width, the anterior limb(AB12) is only indicated by some fiberbundles. As a result, the head of the caudatenucleus (AB8) is no longer separated fromthe putamen (AB6), and the striatum is seenas uniform nuclear complex. In the area ofthe temporal lobe, the curled-up corticalband of the hippocampus (Ammon’s horn)(B26) is almost covered by the parahippo-campal gyrus (B27).
B28Mesencephalic tectum.
Planes of sections
AB
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

223
Telenc
epha
lon
5
9
1919
26
10
11
6
3
1
25
13
28
27
4
18
1224 8
23
21
21
21 22
3
21
4
5
16 17
10
611
8
7
7
14
18
12
1319
9
15
20
Horizontal Sections
A Horizontal section at thelevel of the neostriatum
B Horizontal section at thelevel of the anterior commissure
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

224Te
lenc
epha
lon
Telencephalon
Paleocortex and AmygdaloidBody
Paleocortex
Subdivision (A, B)
The paleocortex (blue) is the oldest corticalarea of the telencephalon. Together with theolfactory bulb and the olfactory tract itforms the olfactory brain, or rhinencephalon.In primitive mammals (hedgehog) (A), thisis the largest part of the telencephalon. Thelarge, compact olfactory bulb (A1) lies ros-trally and, adjacent to it, the olfactorytubercle (A2), or olfactory cortex. The rest ofthe base of the brain is occupied by the pir-iform lobe (A3) with the uncus (A4). The pir-iform lobe contains various cortical areas,namely, laterally the prepiriform area (A5),medially the diagonal band of Broca (ban-deletta diagonalis) (A6), and caudally theperiamygdalar area (A7). The caudal part ofthe piriform lobe is occupied by the entorhi-nal area (A8), a transitional area (orange)between archicortex (red) and neocortex.Medially appears a portion of the hippo-campal formation, the uncus with the su-perficial dentate gyrus (dentate band) (A9).
The enormous expansion of the neocortexin humans (B) has displaced the paleocortexinto the depth where it represents only asmall part of the base of the brain. Theslender olfactory bulb (B10) is connected bythe olfactory tract (B11) with the olfactorycortex. The fibers of the tract divide at theolfactory trigonum (B12) into two (but ofteninto three or more) bundles: the medial ol-factory stria (B13) and the lateral olfactorystria (B14). They enclose the olfactorytuberclewhich, in humans, has sunk into thedepth as anterior perforated substance (B15).It is delimited caudally by the diagonal bandof Broca (B16) which contains afferent fibersfor the olfactory bulb.
The rotation of the hemisphere in humanshas displaced the other parts of the piriformlobemainly to the medial aspect of the tem-poral lobe, where they form the ambientgyrus (B17) and the semilunar gyrus (B18).The ambient gyrus is occupied by the pre-
piriform cortex (B19), and the semilunargyrus by the periamygdalar cortex (B20).Ventrocaudally to it the uncus (B21) bulgeswith the superficial end of the dentate gyrusknown as Giacomini’s band. It merges intothe parahippocampal gyrus (B22) which iscovered by the entorhinal cortex (B23).
Olfactory Bulb (C)
The olfactory bulb has regressed in humans,who belong to the microsmatic mammals.Mammals with a highly developed sense ofsmell (macrosmatic mammals) possess alarge olfactory bulb of complex structure(p. 211, AB3). In the human olfactory bulbwe distinguish a glomerular layer (C24), amitral layer (C25), and a granular layer(C26). The mitral cells of the glomerularlayer form synaptic contacts with the termi-nals of the olfactory nerves (p. 228, A). Theaxons of the mitral cells run through the ol-factory tract to the primary olfactory cen-ters. The olfactory tract contains a discon-tinuous aggregation of medium-sized neu-rons along its entire length, the anterior ol-factory nucleus. Their axons join the fibers ofthe olfactory tract and partly cross to thecontralateral olfactory bulb.
Anterior Perforated Substance (D)
The anterior perforated substance, which ischaracterized by numerous vascular per-forations (D27), is covered externally by anirregular layer of small pyramidal cells, thepyramidal layer (D28), and internally by theloose multiform layer (D29) with individualclusters of dark cells, the islands of Calleja(D30). Olfactory bulb, olfactory tract, andanterior perforated substance contain largenumbers of peptidergic neurons (corti-coliberin, enkephalin, and other peptides).
D31Nucleus of the diagonal band.D32 Longitudinal cerebral fissure.D33 Lateral ventricle.D34 Paraterminal gyrus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

225
Telenc
epha
lon
1
43
25
69
47
8
14
18
17
21
22
15
16
12 13
10
11
19
20
21
23
2625
24
3033
31
34
32
2928
30
27
1
43
25
69
47
8
14
18
17
21
22
15
16
12 13
10
11
19
20
21
23
2625
24
3033
31
34
32
2928
30
27
Paleocortex
A Base of the brainin the hedgehog(according to Stephan)
B Base of the human brain
C Olfactory bulb,frontal section
D Anterior perforated substance, olfactorycortex (according to Crosby and Humphrey)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

226Te
lenc
epha
lon
Telencephalon: Paleocortex and Amygdaloid Body
Amygdaloid Body
The amygdaloid body (amygdala) lies at themedial aspect of the temporal lobe (B). Itconsists of a cortical part, the cortical nu-cleus, and a nuclear part lying in the depth;hence, it must be viewed as a transition be-tween cortex and nucleus. The nuclear com-plex is covered by the periamygdalar cortex(A1).
Subnuclei (A–D)
The complex is divided into several subnu-clei, namely, the superficial cortical nucleus(ACD2), the central nucleus (ACD3), thebasal nucleus (CD4) consisting of a parvo-cellular medial part (A5) and a magnocellu-lar lateral part (A6), and the lateral nucleus(ACD7). The assignment of the medial nu-cleus (A8) to the amygdala complex isquestionable. The amygdaloid body is richin peptidergic neurons. Primarilyenkephalin and corticoliberin can be dem-onstrated in the central nucleus and VIP inthe lateral nucleus.
The subnuclei form two groups: the phylo-genetically old corticomedial group (corticalnucleus, central nucleus) and the phylo-genetically younger basolateral group (basalnucleus, lateral nucleus). The corticomedialgroup receives fibers of the olfactory bulband is the area of origin of the stria termi-nalis. The basolateral group has fiber con-nections with the prepiriform area and theentorhinal area. Electrophysiological re-cordings have demonstrated that only thecorticomedial group receives olfactory im-pulses, while the basolateral group receivesoptic and acoustic impulses.
Functional Organization (C–E)
Electrical stimulation of the amygdala andits surroundings induces autonomic andemotional responses. Anger (!) or flight re-action (") with the corresponding auto-nomic phenomena (dilatation of pupils, risein blood pressure, increase in cardiac andrespiratory rates) can be triggered by stimu-lation of the collecting area of the stria ter-minalis fibers (C). Other sites produce reac-
tions of alertness associated with turningthe head. Stimulation may induce chewing(#), licking ($), or salivation (%) (D). It mayalso result in food uptake, secretion of gas-tric juice, and increased intestinal motilityor bulimia. Hypersexuality may occur as aresult of stimulation but may also be pro-duced by lesions to the basolateral group ofnuclei. Urination (&) or defecation may beinduced as well.
The stimulation responses are difficult toarrange topically; many fibers run throughthe nuclear complex, and the stimulationresponses may originate not only from thesite of stimulation but also from affectedfiber bundles of other nuclei. The medialpart of the basal nucleus has been assignedto the corticomedial group of nuclei, and anattempt has been made to correlate the twonuclear groups with the different re-sponses; the corticomedial group (E9) isthought to promote aggressive behavior,sexual drive, and appetite, while the lateralgroup (E10) has an inhibitory effect.
Clinical Note: Stimulation of the amygdaloidbody in humans (a diagnostic measure in thetreatment of severe epilepsy) may trigger angeror anxiety, but also a feeling of tranquillity and re-laxation. The patients may feel “transformed” or“in a different world”. The response will essen-tially be influenced by the emotional state at theonset of the stimulation.
A–E11 Optic tract.A12 Hypothalamus.A13 Claustrum.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

227
Telenc
epha
lon
1111
9
9 9
10
10
3
2
2
11
4
7
3
4
7
313 8
11
12
2
1
5
6
7
Amygdala
A Subdivision of the amygdaloid body, frontal section, semi-diagram
B Location of section in A C Fight-or-flight reaction, stimulation experimentin the cat (according to de Molina and Hunsperger)
D Autonomic reactions, stimulation experimentsin the cat (according to Ursin and Kaada) E Functional organization
(according to Koikegami)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

228Te
lenc
epha
lon
Telencephalon: Paleocortex
Fiber Connections
Olfactory Bulb (A)
The bundled axons of the olfactory cells(A1) (p. 331, C) pass as olfactory nerves (1stneuron) through the openings of the cri-briform lamina (A2) into the olfactory bulb(A3). Here they terminate on the dendritesof the mitral cells (A4) with which they formglomeruli (A5). In this glomerular system,one mitral cell is in contact with numeroussensory cells. Other cell types, such asgranule cells, periglomerular cells, and tuftedcells, belong to the integration center of theolfactory bulb. The axons of the mitral cells(2nd neuron) pass through the olfactory tract(A6) to the primary olfactory centers. Me-dium-sized neurons are scattered along theolfactory tract; they constitute the anteriorolfactory nucleus (AC7). The axons, or theircollaterals, of the mitral cells terminatehere. The neuronal processes partly crossthrough the anterior commissure to thecontralateral olfactory bulb, where theyform themedial olfactory stria (B8).
Lateral Olfactory Stria (B)
All fibers of the mitral cells extend in thelateral olfactory stria to the primary ol-factory centers, namely, the anterior per-forated substance (olfactory area) (BC9), theprepiriform area (B10), and the periamyg-dalar area (B11) including the cortical nu-cleus of the amygdaloid body. The prepir-iform area and the periamygdalar area arethought to be the olfactory cortex proper forthe conscious perception of olfactorystimuli. The medial olfactory stria isthought to receive exclusively fibers run-ning from the olfactory cortex to the ol-factory bulb.
Fiber systems extend from the olfactory cor-tex (olfactory impulses for the search forfood, food uptake, and sexual behavior) tothe entorhinal area (B12), to the basolateralnuclear group of the amygdaloid body(BC13), to the anterior and lateral portionsof the hypothalamus (B14), and to the mag-nocellular nucleus of the medial thalamicnuclei (B15). A connection to the centers of
the brain stem is established through fibersrunning to the habenular nuclei (B16)(p. 176, A). These association pathways donot directly belong to the olfactory system.
Amygdaloid Body (B)
The basolateral nuclear group receivesfibers from the premotor, prefrontal, andtemporal cortices; from the magnocellularnucleus of the medial thalamic nuclei; andfrom nonspecific thalamic nuclei. The mostimportant efferent fiber system of theamygdaloid body is the stria terminalis(BC17). It arches in the sulcus between cau-date nucleus and thalamus and runs belowthe thalamostriate vein (p. 171, C14; p. 175,AB2) as far as the anterior commissure. Itsfibers terminate in the septal nuclei (B18), inthe preoptic area (B19), and in the nuclei ofthe hypothalamus. Fiber bundles cross fromthe stria terminalis into the medullary stria(B20) and extend to the habenular nuclei.Other efferent bundles from the basolateralportion of the amygdaloid body extend asventral amygdalofugal fibers (B21) to the en-torhinal area, to the hypothalamus, and tothe medial thalamic nuclei, from where ad-ditional connections lead to the frontal lobe.The stria terminalis is rich in peptidergicfibers.
Anterior Commissure (C)
In the anterior part of the anterior commis-sure, fibers of the olfactory tract (anteriorolfactory nucleus) (AC7) and fibers of the ol-factory cortex (BC9) cross to the con-tralateral side. The anterior part is poorlydeveloped in humans. The main part isformed by the posterior part, where fibers ofthe temporal cortex (C22) cross; they areprimarily from the cortex of the medialtemporal gyrus. Furthermore, the posteriorpart contains crossing fibers from the amyg-daloid bodies (BC13) and the striae (termi-nales) (BC17).
B23Optic chiasm.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

229
Telenc
epha
lon
9
1322
17
7
435
6 7
2
1
18
19
15
20 17
16
8
911
14
13
1221
10
23
Fiber Connections
A Olfactory bulb
B Lateral olfactory stria
C Anterior commissure
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

230Te
lenc
epha
lon
Telencephalon
Archicortex
Subdivision and FunctionalSignificance (A–D)
The hippocampus (A–D1) is the main part ofthe archicortex. It lies at the medial aspectof the temporal lobe in the depth and islargely covered by the parahippocampalgyrus. The left hemisphere has been re-moved in the preparation, showing the cutsurface of the corpus callosum (A2) withonly the left hippocampus being left intact.The latter looks like a paw with claws, thedigitations. The temporal lobe of the righthemisphere in the background illustratesthe position of the hippocampus in the tem-poral lobe. The hippocampus extends to thecaudal end of the corpus callosum. Here, itbecomes reduced to a thin layer of graymatter, the indusium griseum (A3), which ex-tends along the superior surface of the cor-pus callosum to its rostral end in the regionof the anterior commissure (A4). Two nar-row fiber bundles, the lateral and the mediallongitudinal striae of Lancisi (p. 220, A7 andA8) also run here bilaterally. On the dorsalsurface of the hippocampus lies a thick fiberband, the fimbria of hippocampus (A–D5),which separates from the hippocampusbeneath the corpus callosum and continuesas fornix (A6), arching down to to mamillarybody (A7).
In a horizontal section through the temporallobe, the inferior horn (BC8) and the poste-rior horn (B9) of the lateral ventricle are ex-posed, and the protrusion of the hippocam-pus into the ventricle is visible. Medially, al-ready at the outer aspect of the temporallobe, lies the fimbria and, beneath it, thedentate gyrus (fascia dentata) (B–D10), sepa-rated from the parahippocampal gyrus (en-torhinal area) (B–D11) by the hippocampalsulcus (BC12).
In a frontal section the hippocampal cortexforms a curled band, Ammon’s horn, whichprotrudes against the ventricle and iscovered by a layer of fibers, the alveus (C13).Ammon’s horn shows considerable varia-tions at different planes of section (D).
In the past, the hippocampus has been as-signed to the rhinencephalon, but it has nodirect relationship with the olfactory sense.In reptiles, which do not have a neocortex,the telencephalon is the highest integrationorgan. Electrical recordings from the hippo-campus of mammals show that it receivesoptic, acoustic, tactile, and visceral input,but only a few olfactory impulses. It is anintegration organ influencing endocrine,visceral, and emotional processes via its con-nections to the hypothalamus, septal nuclei,and cingulate gyrus. Furthermore, the hip-pocampus plays a major role in processes oflearning and memory.
Clinical Note: Bilateral removal of the hippo-campus in humans (treatment of severe epilepticseizures) leads to a loss in memory. While oldmemories are retained, new information can beremembered only for a few seconds. Such a short-term memory may persist for years. The hippo-campal neurons possess a very low absolutethreshold for convulsive discharges. Thus, thehippocampus is of special importance for theorigin of epileptic seizures and memory deficits.
C14Optic tract.C15 Choroid plexus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

231
Telenc
epha
lon
2
47
1
6
5
3
10 10
11
11
105
11
105
11
14
1513
1
8
1
1
12510
1
128
1 5
9
11
Subdivision of the Archicortex
A Hippocampus after removal of the rest of theleft hemisphere (according to Ludwig and Klingler)
B Hippocampus viewed from above(according to Sobotta)
C Frontal section through hippocampus,Ammon’s horn
D Ammon’s horn at differentplanes of sections
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

232Te
lenc
epha
lon
Telencephalon: Archicortex
Ammon’s Horn (A)
The hippocampus is subdivided into fourparts according to width, cell size, and celldensity:
! Field CA1 (A1) contains small pyramidalcells.
! Field CA2 (A2) is characterized by a nar-row, dense band of large pyramidal cells.
! Field CA3 (A3) is characterized by a wideloose band of large pyramidal cells.
! Field CA4 (A4) forms the loosely struc-tured inner zone. Recently, it has beencalled into question whether a separateCA4 region can be delimited from theCA3 region.
The narrow band of densely packed granulecells of the dentate gyrus (fascia dentata)(A5) surrounds the ending band of pyra-midal cells. The dentate gyrus is fused withthe surface of the curled-up Ammon’s hornand appears only partially at the surface ofthe brain. It is separated by the hippocampalsulcus (A6) from the parahippocampal gyrus(A7) and by the fimbriodentate sulcus (A8)from the fimbria of the hippocampus (A9).The inner layer bordering on the ventricle isthe alveus of the hippocampus (A10), inwhich the efferent fibers collect before leav-ing the hippocampus via the fimbria. Thetransitional area between Ammon’s hornand the bordering entorhinal cortex (A11) iscalled the subiculum (A12).
Fiber Connections (B, C)
Afferent Pathways (B)
The fiber bundles from the entorhinal area(B13) are thought to be the most importantafferent system, where the pathways fromthe primary olfactory centers (prepiriformarea), from the amygdaloid body, and fromvarious regions of the neocortex terminate.Direct connections between olfactory bulband hippocampus have not been demon-strated.
The fibers from the cingulate gyrus collectin the cingulum (B14) and extend primarilyto the subiculum.
The fornix (B15) contains bundles from theseptal nuclei (B16) but above all fibers fromthe hippocampus and the entorhinal area ofthe contralateral hemisphere (via the com-missure of the fornix).
Efferent Pathways (B)
Apart from a few fibers leaving the hippo-campus via the longitudinal stria (B17), thefornix contains all other efferent pathways.It is divided into a precommissural part anda postcommissural part. The fibers of theprecommissural fornix (B18) terminate in theseptum, in the preoptic area (B19), and inthe hypothalamus (B20). The fibers of thepostcommissural fornix (B21) terminate in themamillary body (B22) (predominantly in themedial nucleus of the mamillary body), in theanterior thalamic nucleus (B23), and in thehypothalamus. Some fibers of the fornix ex-tend to the central gray matter of the mid-brain.
A large neuronal circuit can be recognized inthis system of pathways. Hippocampal im-pulses are conducted via the fornix to theanterior thalamic nucleus. The latter is con-nected with the cingulate gyrus, fromwhere there is feedback via the cingulum tothe hippocampus (Papez circuit) (p. 332, C).
Fornix (C)
At the inferior surface of the corpus callo-sum, the two limbs of the fornix (C24) uniteto form the commissure of the fornix (psal-terium) (C25) and the body of the fornix(C26), which then divides again into the twocolumns of the fornix (C27) above the fora-men of Monro.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

233
Telenc
epha
lon
9
8
3
5
4
12
111
2
10
67
16
1823 17
14
2022
2726 24
25
13
19 21
15
9
8
3
5
4
12
111
2
10
67
16
1823 17
14
2022
2726 24
25
13
19 21
15
Ammon’s Horn, Fiber Connections
A Ammon’s horn, frontal section through the hippocampus
B Fiber connections of thehippocampus
C Hippocampus and fornix(according to Feneis)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

234Te
lenc
epha
lon
Telencephalon: Archicortex
Hippocampal Cortex (A, B)
The structure of the archicortex is simplerthan that of the neocortex, and its neuronalcircuits are therefore easier to elucidate. Thehippocampal cortex belongs to those brainregions where inhibitory and excitatoryneurons have been identified both histo-logically and electrophysiologically.
Fields CA1 (A1), CA2 (A2), and CA3 (A3) showdifferences with respect to organization andfiber connections. The majority of afferentfibers enter Ammon’s horn via the perforantpath (A4), and only a few do so via the alveusof hippocampus. They terminate on thedendrites of the pyramidal cells (AB5). Manyof the fibers (AB6) extend to the granule cells(AB22) of the dentate gyrus (fascia dentata);their axons, mossy fibers (AB7), too, havesynaptic contacts with the dendrites of pyr-amidal cells. However, mossy fibers runonly to field CA3; they are absent from fieldsCA1 and CA2.
The pyramidal cells are the efferent el-ements; their axons collect in the alveus(AB8) and leave the cortex through the fim-bria (A9). The axons of the CA3 pyramidalcells give off recurrent collaterals (Schaffercollaterals) (AB10) that form synapses withdendrites of the CA1 pyramidal cells. Theefferent fibers running to the septum origi-nate in CA3, the fibers for the mamillarybody and the anterior thalamic nucleusoriginate in CA1. Many of the efferent fibersof the hippocampus, however, run to thesubiculum.
Organization of layers. Ammon’s horn con-sists of the following layers: the alveus(AB8) with the efferent fibers lies inside andis followed by the stratum oriens (B11 ) withthe basket cells (B12), the axons of whichsplit up and fill the pyramidal layer with adense fiber network (B13). The fibers en-velope the pyramidal cell bodies and formsynaptic contacts (axosomatic synapses)with them. Basket cells are inhibitory neu-rons that are excited by the axon collateralsof the pyramidal cells and cause pyramidalcell inhibition following pyramidal cell dis-charge. The pyramidal cells form the
stratum pyramidale (B14). Their apices areoriented toward the subsequent stratumradiatum (B15), their bases toward thestratum oriens. They send dense dendritictrees in both directions. The long apical den-drite reaches with its branches into thestratum lacunosum-moleculare (B16). In theCA3 region, one can also distinguish astratum lucidum (B20) where the mossyfibers run.
The afferent fibers originating from differentregions run in different layers. Many of thecommissural fibers from the contralateralhippocampus extend into the stratum oriens(B11) and the stratum radiatum (B15). Thefibers of the entorhinal area (B5) extendinto the stratum lacunosum-moleculare(B16) and form contacts with the outermostbranches of the apical dendrites (B17).Schaffer collaterals (B10) have contact withdistal segments of the apical dendrites ofthe CA1 pyramidal cells, while the mossyfibers (B7) have contact with proximal seg-ments of the CA3 pyramidal cells. The den-drites of granule cells of the dentate gyrusare contacted in a similar way; entorhinalfibers terminate on distal dendritic seg-ments, while commissural fibers terminateon proximal segments of the dendrites. Inaddition to the principal cells—pyramidalcells and granule cells—the afferent fibers ofthe hippocampus also form synaptic con-tacts with inhibitory GABAergic inter-neurons (feed-forward inhibition of princi-pal neurons, p. 35, C). Apart from the basketcells mentioned above (B12), which formaxosomatic synapses, GABAergic cells havebeen found in recent years that form synap-tic contacts on the initial segment of theaxon (axo-axonal cells or chandelier cells)(B18) or on the dendrites (B19) of the princi-pal cells. From the course of the fibers andfrom electrophysiological studies, the fol-lowing impulse flow emerges in the hippo-campus: glutamatergic, entorhinal afferentfibers activate granule cells which, in turn,activate CA3 pyramidal cells via mossyfibers. These then activate CA1 pyramidalcells via Schaffer collaterals (trisynaptic ex-citatory pathway of the hippocampus).
B21Hilus of dentate gyrus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

235
Telenc
epha
lon
2
1
3
16
15
14
11
8
8
76
4
5
10
9
10
13
19 12
7
20 21
22
18
17 5 6
22
Hippocampal Cortex
A Organization of the hippocampus (according to Cajal)
CA1 CA3 Dentate gyrus
B Neuronal circuits in the hippocampus
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

236Te
lenc
epha
lon
Telencephalon
Neostriatum
The neostriatum (or striatum) is the highestintegration site of the extrapyramidal motorsystem (p. 310). It is a large, gray complex inthe depth of the cerebral hemisphere and isdivided into two parts by the internal cap-sule (ABD1), namely, the caudate nucleus(ABD2) and the putamen (ABD3) (p. 214,AB18 and AB19; p. 216, AB9 and AB10). Thecaudate nucleus consists of the large head ofthe caudate nucleus (A4), the body of the cau-date nucleus (A5), and the tail of the caudatenucleus (A6). Immunohistochemical assaysfor neurotransmitter substances yield aspotty, mosaic-like structure created by theterminals of various fiber tracts. The spotsform a system of interconnected fields(striosomes) that stand out from the rest ofthe tissue because of their content of aspecific neurotransmitter.
Afferent Pathways (B–D)
Corticostriate fibers (B8). Fibers extendfrom all areas of the neocortex to the neo-striatum. They are the axons of medium-sized and small pyramidal cells of the fifthlayer (see p. 240). However, there are nofiber connections extending from the stri-atum to the cortex. The corticostriate pro-jection reveals a topical organization (C):the frontal lobe projects to the head of thecaudate nucleus (red) and is followed by theparietal lobe (light blue), the occipital lobe(purple), and the temporal lobe (dark blue)(see p. 213). The projection of the precentralmotor area in the putamen reveals a soma-totopic organization (D): head (red), arm(light red), and leg (hatched area). A soma-totopic projection of the postcentralsensory area to the dorsolateral region ofthe caudate nucleus has been demon-strated. The fibers from areas adjoining thecentral sulcus are the only ones that partlycross via the corpus callosum to the con-tralateral neostriatum (B9).
Centrostriate fibers (B10). These fiberbundles extend from the centromedianthalamic nucleus to the neostriatum; thosefor the caudate nucleus originate in the dor-sal part, those for the putamen in the ven-
tral part of the nucleus. Impulses from thecerebellum and from the reticular forma-tion of the midbrain reach the neostriatumvia these fibers.
Nigrostriate fibers (B11). Fibers extendingfrom the substantia nigra to the neostri-atum can be traced by fluorescence micros-copy. They are the axons of dopaminergicneurons, and they cross the inner capsule ingroups. They run without interruptionthrough the globus pallidus to the neostri-atum (p. 136, B16).
Serotoninergic fiber bundles from theraphe nuclei.
Efferent Pathways (B)
The efferent fibers extend to the globus pal-lidus. The fibers of the caudate nucleus ter-minate in the dorsal parts of the two seg-ments of the pallidum (B12), while thefibers of the putamen terminate in the ven-tral parts (B13). Here, they synapse with thepallidofugal system, namely, with the pal-lidosubthalamic fibers, the lenticular ansa,the lenticular fasciculus, and the pal-lidotegmental fibers (p. 192, A16).
Strionigral fibers (B14). Fibers of the cau-date nucleus terminate in the rostral partand fibers of the putamen in the caudal partof the substantia nigra (p. 136, B12, B14).
Functional Significance
Both the topical organization of the cor-ticostriate fiber systems and its mosaic-likestructure show that the neostriatum isdivided into many functionally differentsectors. It receives stimuli from the frontalcortex, from the optic, acoustic, and tactilecortical fields and their association areas.These areas are thought to have an effect onthe motor system via the stratum (sensorymotor integration, cognitive function of theneostriatum). The neostriatum has no directcontrol over elementary motor processes(its destruction does not lead to an appreci-able loss of motor functions). Rather, it isviewed as a higher integration system thatinfluences the behavior of an individual.
A7 Amygdaloid body.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

237
Telenc
epha
lon
2
5
4 3
61
7
2
1
2
2
3
3
3
88
8
9
10
1
131411
8
2
3
12
2
5
4 3
61
7
2
1
2
2
3
3
3
88
8
9
10
1
131411
8
2
3
12
Neostriatum
A Neostriatum follo-wing removal ofadjacent brain structures(according to Ludwig and Klingler)
B Fiber connections ofthe neostriatum
C Projection of the cortex to thecaudate nucleus in the monkey(according to Kemp and Powell)
D Projection of the precentral areaonto the putamen in the monkey(according Künzle)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

238Te
lenc
epha
lon
Insula
The insula is the region at the lateral aspectof the hemisphere that lags behind duringdevelopment and becomes covered by themore rapidly growing adjacent regions ofthe hemisphere. The parts of the hemi-sphere overlapping the insula are calledopercula. They are named according to thecerebral lobe they belong to: the frontaloperculum (A1), the parietal operculum (A2),and the temporal operculum (A3). In diagramA, the opercula have been moved apart toexpose the insula. They normally leave onlya cleft, the lateral cerebral sulcus (fissure ofSylvius, p. 10, A4), which widens over the in-sula into the lateral fossa (p. 216, AB15). Theinsula has roughly the shape of a triangleand is bordered at its three sides by thecircular sulcus of the insula (A4). The centralsulcus of the insula (A5) divides the insulainto a rostral and a caudal part. At its lowerpole, the limen of insula (A6), the insular re-gion merges into the olfactory area, thepaleocortex.
The insular cortex represents a transitionalregion between paleocortex and neocortex.The lower pole of the insula is occupied bythe prepiriform area (B7) (blue) whichbelongs to the paleocortex. The upper partof the insula is covered by the isocortex (neo-cortex; see p. 244) (B8) (yellow) with the fa-miliar six layers (p. 240). Between bothparts lies a transitional region, the mesocor-tex (proisocortex, see p. 244) (B9) (hatchedarea). Unlike the paleocortex, it has six lay-ers; however, these are only poorlydeveloped as compared to the neocortex.The fifth layer (C10) is characteristic for themesocortex by standing out as a distinctnarrow, dark stripe in the cortical band. Itcontains small pyramidal cells that aredensely packed like palisades, a featureotherwise found only in the cortex of thecingulate gyrus.
Stimulation responses (D). Stimulation ofthe insular cortex is difficult because of thehidden position of the region; it has beencarried out in humans during surgical treat-ment of some specific forms of epilepsy. It
caused an increase (+) or decrease (–) in theperistaltic movement of the stomach.Nausea and vomiting (!) were induced atsome stimulation sites, while sensations inthe upper abdomen or stomach region (!)or in the lower abdomen (") were producedat other sites. At several stimulation sites,taste sensations were induced (#). Al-though the stimulation chart does not showa topical organization of these effects, theresults do indicate viscerosensory andvisceromotor functions of the insular cortex.Experiments with monkeys yielded notonly salivation but also motor responses inthe muscles of the face and the limbs. Inhumans, surgical removal of the insular re-gion does not lead to any functional losses.
Telencephalon
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

239
Telenc
epha
lon
2
4
5 31
6
10
8
9
7
Insula
A Insula with the opercula movedapart (according to Retzius)
B Cortical areas of the insula(according to Brockhaus)
C Mesocortex D Stimulation map of the human insularcortex (according to Penfield and Faulk)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

240Te
lenc
epha
lon
Telencephalon
Neocortex
Cortical Layers (A–C)
The neocortex (isocortex) exhibits a stratifi-cation into six layers running parallel to thesurface of the hemisphere. The stratificationcan be demonstrated by silver impregnation(A1), cellular staining according to Nissl(A2), myelin staining (A3), and pigmentstaining (B). The layers are distinguished ac-cording to the different shapes, sizes andnumbers of their neurons and by the differ-ent densities of myelinated nerve fibers.
Cellular staining (A2) reveals the followingfeatures:
! The outermost layer, themolecular layer(layer I) (A4), contains few cells.
! The external granular layer (layer II)(A5) is densely packed with small granulecells.
! The external pyramidal layer (layer III)(A6) contains predominantly medium-sized pyramidal cells.
! The internal granular layer (layer IV)(A7) consists of densely packed smallgranule cells.
! The internal pyramidal layer (ganglioniclayer) (layer V) (A8) contains large pyra-midal cells.
! The multiform layer (layer VI) (A9)completes the stratification with a loosemixture of different cell types.
Silver impregnation (A1), which shows theneuron with all its processes (p. 18), makesit possible to identify the granule cells oflayer II as small pyramidal cells and stellatecells, and the granule cells of layer IV pre-dominantly as stellate cells. The pyramidalcell (C) is the typical neuron of the neocor-tex. Its axon (C10) takes off from the base ofthe cell, where the basal dendrites (C11)branch off at the margins. One long, thickdendrite, the apical dendrite (C12), ascends tothe surface of the cortex. The dendrites havethousands of spines at which other neuronssynapse.
Myelin staining (A3) of the nerve fibers re-veals the following layers based on thedifferent densities of tangential fibers:
! The tangential layer (A13).! The dysfibrous layer (A14).! The suprastriate layer (A15).! The external (A16) and internal (A17)
Baillarger’s bands of high fiber density,the external band being created bybranches of afferent fibers, the internalband by axon collaterals of pyramidalcells.
! The substriate layer (A18) completes thestratification.
! In addition, there are the vertical bundlesof radial fibers (A19).
Pigment staining (B). The various neuronsdiffer in their degree of pigmentation. Thedifferent pigment contents cause thecharacteristic stratification of the cortex,usually with two unpigmented bands corre-sponding to the two Baillarger’s bands.
Vertical Columns (D)
The basic functional units of the neocortexare vertical cell columns that reach throughall layers and have a diameter of 200 –300µm. Electrophysiological studies haveshown that, in the cortical projection areas,each cell column is connected to a definedperipheral group of sensory cells. Stimula-tion of the peripheral field always yields aresponse from the entire column.
Fiber tracts connect the cortical columnswith each other (D): the fibers of a column(D20) run either to columns of the ipsi-lateral hemisphere (association fibers, seep. 260) or via the corpus callosum to mostlysymmetrically localized columns of thecontralateral hemisphere (commissuralfibers, see p. 260). Branches of individualfibers terminate in different columns (D21).It is estimated that the neocortex ismade upof 4 million columns.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

241
Telenc
epha
lon
4 13
14
15
16
17
19 18
11
12
10
20
21
21
5
6
7
8
9
1 2 3
I
II
III
IV
V
VI
Neocortex, Cortical Layers, and Vertical Columns
200–30
0 µm
200–300 µm
A Layers of the neocortex: 1, silverimpregnation; 2, cellular staining;3, myelin staining (according toBrodmann)
B Pigment staining C Pyramidal cell andapical dendrite(according to Cajal)
D Connection of vertical columns in the neocortex(Szentágothai according to Goldman and Nauta)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

242Te
lenc
epha
lon
Telencephalon: Neocortex
Cell Types of the Neocortex (A)
In principle, we distinguish between projec-tion neuronswith long axons (excitatory gluta-matergic pyramidal cells) and interneuronswith short axons (inhibitory GABAergic inter-neurons).
The pyramidal cell (A1) is characterized byone apical dendrite (A2), which ascends tothe molecular layer and branches there, andnumerous basal dendrites (A3). Its de-scending axon gives off numerous recurrentcollaterals (A4). The cell-deficient molecu-lar layer (layer I) contains Cajal–Retzius cells(A5) with tangentially running axons. Thedifferent types of granule cells or stellatecells are predominantly interneurons andare found in all layers at various densities.They include Martinotti’s cells (A6), the ver-tically ascending axons of which ramify invarious cortical layers and reach as far as themolecular layer. The cellules à doublebouquet dendritique of Cajal, cells with twovertically oriented dendritic trees (A7) (pri-marily in layers II, III, and IV), possess longascending or descending axons. The axon ofsome stellate cell types arborizes after ashort course (A8), or it bifurcates and termi-nates with basketlike networks (basketcells) (A9) on adjacent pyramidal cells. Axonbifurcations may run horizontally and ter-minate on distant pyramidal cells (A10).Their inhibitory function has been con-firmed by detection of GABA in the synapsesof basket cells.
The Module Concept (B)
The results of histological and electrophysi-ological studies have made it possible to de-sign models in which the described celltypes are organized in a functional group.The vertical column is conceived as a mod-ule, that is, as a group of elements forming afunctional unit.
The efferent elements of the column arethe pyramidal cells (B11). Their axons eitherrun to other cortical columns, where theirterminal ramifications end at the spines ofother pyramidal cells, or they run to subcor-tical groups of neurons. The numerous axon
collaterals (A4) terminate at the pyramidalcells of nearby columns.
There are two kinds of afferent fibers: theassociation fibers from other columns(p. 240, D) and the specific sensory fibersfrom peripheral sensory areas. In everylayer the association fibers (B12) give offbranches that terminate at the spines ofpyramidal cells. They ascend to the molecu-lar layer, where they branch into horizon-tally running fibers. The latter have synapticcontacts with apical dendrites within aradius of 3mm. The excitation transmittedby them reaches far beyond the column;however, it remains weak because the num-ber of synaptic contacts is limited. Thespecific fibers (B13) terminate in layer IV oninterneurons (B14), primarily on the cellswith two dendritic trees (B15). The axons ofthe latter ascend vertically along the apicaldendrites of pyramidal cells and form syn-apses with their spines (B16). These seriesof synapses result in powerful transmission.The basket cells (B17), which are inhibitoryinterneurons, send their axons to the pyra-midal cells of adjacent columns and inhibitthem, thereby restricting the excitation. Thebasket cells themselves are activated by re-current collaterals of the excitatory pyra-midal cells. The axons of Martinotti’s cells(B18) ascend to the molecular layer wherethey form branches.
The number of neurons per column is esti-mated to be 2500, approximately 100 ofwhich are pyramidal cells. It should be con-sidered, however, that a vertical column isnot a clearly defined histological entity.Possibly, it does not represent a permanentmorphological unit but rather a functionalunit, which forms and disintegrates accord-ing to the level of excitation.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

243
Telenc
epha
lon
16
5
2 2
1
15 10
8
13
39
7
6 4
15
11
14 14
16
11
18
17
1312
Cell Types, Module Concept
A Cellular elementsfired of theneocortex (accordingto Colonnier)
B Simplified model of a column(according to Szentágothai)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

244Te
lenc
epha
lon
Telencephalon: Neocortex
Cortical Areas (A, B)
All regions of the neocortex develop in asimilar way. First, a wide cell layer, the corti-cal plate, forms on the surface of the hemi-sphere which then divides into six layers.Because of this similar development, theneocortex is also known as isogenetic cortex,or briefly, isocortex, or as homogenetic cor-tex.
Nevertheless, the adult neocortex exhibitsconsiderable regional variations, and wedistinguish several regions of differentstructure, the cortical areas. The individuallayers may vary considerably in these areas:wide or narrow, with densely or looselypacked cells. The cells may vary in size, or aspecific cell type may predominate. Thedefinition of individual areas according tosuch criteria is called architectonics. Depend-ing on the staining method employed(p. 240A), the following terms are used: cy-toarchitectonics, myeloarchitectonics, or pig-ment architectonics. A map of cortical areascan be reconstructed on the surface of thehemisphere, similar to a geographic map.The cytoarchtectonic map of cortical areasestablished by Korbinian Brodmann (A, B)has been confirmed repeatedly and is gen-erally accepted.
Types of cortices. A special feature of pro-jection areas (terminals of ascending sensorypathways) is the prominent development oftheir granular layers. In the sensory cortex(area 3) as well as in the auditory cortex(areas 41 and 42, temporal transverse gyri),the granular layers (layers II and IV) arewide and rich in cells, while the pyramidalcell layers are less well developed. This typeof cortex is referred to as koniocortex, orgranular cortex. The visual cortex (area 17,striate area) even exhibits a duplication oflayer IV (p. 255, A). These sensory corticalareas, which are the terminals of afferentprojection fibers, are involved in associativeprocesses, where relay neurons with shortaxons play an important role. On the otherhand, the granular layers in the motor cortex(areas 4 and 6) are largely reduced in favorof the pyramidal layers (agranular cortex).The pyramidal cells are projection neurons
with long axons; for example, in the motorcortex, their axons form the corticospinaltract (pyramidal pathway).
Border zones. Wherever the isocortexborders on the archicortex or paleocortex,its structure becomes simpler. The transi-tional formation with the simpler structureis referred to as proisocortex. The proisocor-tex includes the cortex of the cingulate gyrus,the retrosplenial cortex (lying around theposterior end of the corpus callosum), andparts of the insular cortex. The proisocortexis phylogenetically older than the neocor-tex.
Paleocortex and archicortex are also sur-rounded by a border zone, the structure ofwhich approximates that of the neocortex.The border zones are known as periarchicor-tex and peripaleocortex. The periarchicortexincludes, for example, the entorhinal areabordering on the hippocampus.
Allocortex. The allocortex is often con-trasted with the isocortex. The term refersto the paleocortex and the archicortex. Asboth are completely different parts of thetelencephalon, genetically as well as struc-turally and functionally, this collective termis not justified. The only thing they have incommon is that they are different from theisocortex. Their simpler structure (for ex-ample, no stratification into six layers as it ischaracteristic for the neocortex) does notjustify to label them as primitive regions.Rather, they are phylogenetically old, highlyspecialized structures.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
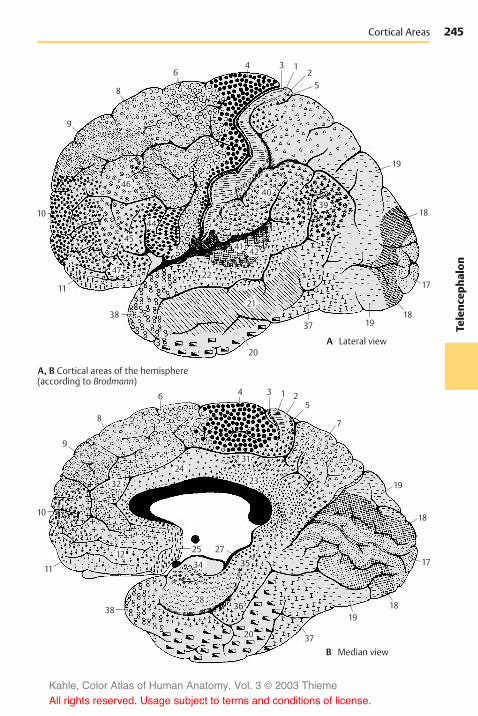
245
Telenc
epha
lon
46
9
9
8
6 4 3 1 25
7
19
18
1819
37
38
25 27
11
10
17
8
64 3 1
52
19
18
17
181937
20
20
38
11
10
4039
4241
22
21
2431
2332
1234 35
2836
47
45
44
Cortical Areas
A, B Cortical areas of the hemisphere(according to Brodmann)
A Lateral view
B Median view
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

246Te
lenc
epha
lon
Telencephalon: Neocortex
Frontal Lobe (A–C)
We distinguish the precentral area (themotor cortex proper) and the premotor, theprefrontal, and the orbitofrontal corticalareas.
Agranular Cortex
The cortex of the precentral area (red), con-sisting of primary motor cortex (area 4) andpremotor cortex (area 6) (C), is characterizedby the reduction or loss of granular layersand a general increase in pyramidal cells.Also typical are the exceptional thickness ofthe cortex and its gradual transition into thewhite matter. These features are especiallyprominent in the cortex of area 4 (A), wherecertain regions of layer V contain giant pyra-midal cells (Betz’s cells) (A1). The latterpossess the thickest and longest axons inthe nervous system, reaching as far as thesacral spinal cord.
The prefrontal cortex (area 9, light red) isshown for comparison (B). It is not only nar-rower and better delimited against thewhite matter by a distinct layer VI, but alsopossesses well-developed granular layers (IIand IV).
The agranular cortex (areas 4 and 6) is theprincipal site of origin of the pyramidal tractand is regarded as prototype of the motorcortex. Nevertheless, it also receives afferentfibers: following the stimulation of skin atthe extensor and flexor sides of the limbs,electrical potentials can be recorded at theprecentral area. They are probably afferentsystems for controlling and fine-tuning themotor system. On the other hand, motor re-sponses can be induced by enhanced stimu-lation at some points of the postcentral areaof the parietal lobe (somatosensory area,blue) (C) and the premotor area of the fron-tal lobe. Accordingly, pyramidal cells withlong axons are found in layer V of theseareas (!). Physiologists therefore speak of amotosensory area, Ms I (predominantlymotor) and of a sensorimotor area, Sm I (pre-dominantly sensory). However, these find-ings do not affect the basic fact that the pre-central area represents the motor cortex,
while the postcentral area represents thesomatosensory (tactosensory) cortex.
Granular Cortex
The prefrontal and orbitofrontal corticesexhibit well-developed granular layers (B).
Clinical Note: Injury to the granular frontalcortex results in severe changes in personality.This affects the formal intellectual capacity lessthan it does initiative, ambition, concentration,and judgment. The patients show a silly self-satisfied euphoria, are only interested in everydaytrivia, and are unable to plan ahead.Similar changes have been observed in patientswith prefrontal lobotomy. This is a complete surgi-cal severing of the frontal fiber connections,which was performed as a form of treatment ofraving, mentally deranged patients and inpatients with the most severe conditions of pain(this treatment has now become obsolete thanksto psychopharmaceuticals). The operationachieved permanent calming and indifference ofthe patients. A characteristic change was ob-served in the emotional sphere: the patents stillfelt the pain, but they did not perceive it as trou-blesome; previously intolerable pain had becomeimmaterial. Radical changes in character alsooccur following injury of the orbitofrontal area(basal neocortex). Previously cultured and edu-cated people exhibit a disintegration of decency,tact, and sense of shame, which may lead to seri-ous social offenses.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

247
Telenc
epha
lon
1
89
6
4
4445
1
89
6
4
4445
Frontal Lobe
I
B Area 9
C Precentral cortical areas, areas 4and 6 (according to Bonin)
II
III
IV
V
VI
I
II
III
IV
V
VI
A Area 4, motor cortex
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

248Te
lenc
epha
lon
Telencephalon: Neocortex
Frontal Lobe (continued)
Somatotopic Organization of thePrecentral Area (A, B)
Electrical stimulation of individual corticalregions of the precentral area (A, B1) causesmuscle contraction in specific body regions.The area exhibits a somatotopic organiza-tion. The head region lies above the lateralsulcus, with the lowest parts representingthe throat (A2), the tongue (A3), and thelips. The areas for hand, arm, trunk, and legfollow dorsally, with the leg area reachingbeyond the edge of the pallium to the me-dian surface of the hemisphere. This createsan invertedmotor homunculus. The areas forindividual body parts differ in size; thoseparts of the body where muscles have toperform differentiated movements are rep-resented by particularly large areas. The fin-gers and the hand occupy the largest areas,and the trunk the smallest.
Each body half is represented on the con-tralateral hemisphere, that is, the left bodyhalf on the right hemisphere and the rightbody half on the left hemisphere. Unilateralstimulation yields bilateral responses ofmasticatory, laryngeal, and palatal muscles,and partly also of the trunk muscles. Themuscles of face and limbs yield strictly con-tralateral responses. Representation of thelimbs is organized in such a way that areasfor distal parts of the limbs lie deep in thecentral sulcus, while those for the proximalparts lie more rostrally on the precentralgyrus (B1).
Supplementary Motor Areas (B)
Apart from the precentral area (Ms I), thereare two additional motor areas. Their soma-totopic organization has been demon-strated in monkeys but not in humans. Thesecond motosensory area, Ms II (B4), lies onthe medial surface of the hemisphere abovethe cingulate gyrus adjacent to areas 4 and6. The second sensorimotor area, Sm II (B5),which is predominantly a tactosensory arearather than a motor area, lies above thelateral sulcus and roughly corresponds toarea 40. The functional significance of these
areas within the entire motor system hasnot yet been elucidated.
Frontal Eye Field (C)
Conjugated eye movements can be inducedby electrical stimulation of the precentralarea and, above all, of area 8. This is the fron-tal eye field for voluntary eye movements.In general, stimulation causes the gaze toturn to the opposite side, sometimes withsimultaneous turning of the head. The fibersof area 8 do not terminate directly at the nu-clei of the eye-muscle nerves. Their im-pulses are probably relayed in the intersti-tial nucleus of Cajal.
Broca’s Motor Speech Area (D)
Injury to the region in the lower frontalgyrus (areas 44 and 45) of the dominanthemisphere (p. 262) causes motor aphasia.The patients are no longer able to form andarticulate words, even though the speechmuscles (lips, tongue, larynx) are not para-lyzed. Speech comprehension is not af-fected.
It is certainly impossible to localize the fac-ulty of speech to a defined cortical area(“speech center”). Speech is one of thehighest cerebral functions and involveslarge regions of the cortex. However, Broca’smotor speech area is without doubt a cru-cial relay station in the complex neuronalfoundation of speech (sensory aphasia,p. 262).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
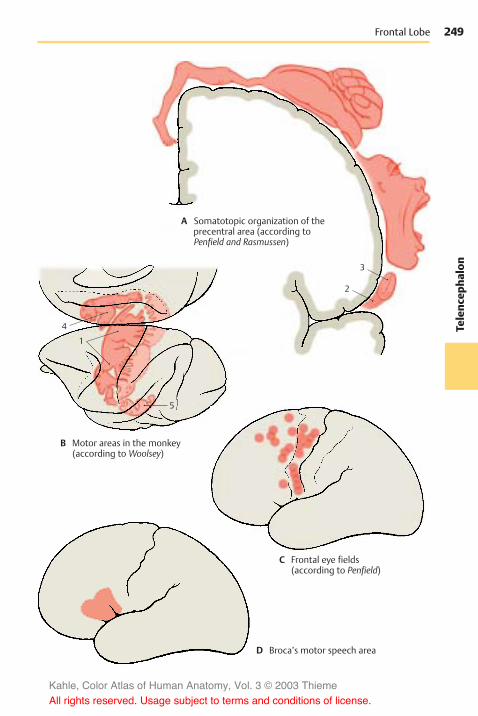
249
Telenc
epha
lon
4
1
5
2
3
Frontal Lobe
A Somatotopic organization of theprecentral area (according toPenfield and Rasmussen)
B Motor areas in the monkey(according to Woolsey)
C Frontal eye fields(according to Penfield)
D Broca’s motor speech area
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

250Te
lenc
epha
lon
Telencephalon: Neocortex
Parietal Lobe (A–C)
Postcentral Area
The terminal station of the sensory path-ways, the somatosensory cortex, lies on themost anterior convolution of the parietallobe, the postcentral gyrus. It contains areas3, 1, and 2. Area 3 lies on the frontal surfaceof the gyrus and deep in the central sulcus;area 1 covers the top of the gyrus as a nar-row band; and area 2 covers the posteriorsurface (p. 245, A).
In contrast to themotor cortex, the cortex ofarea 3 (A) is extremely narrow and clearlyseparated from the white matter. The pyra-midal layers (III and V) are narrower andcontain few cells, while the granular layers(II and IV) aremuchwider. Hence, the cortexof area 3 is part of the koniocortex, or granu-lar cortex (p. 244). The cortex of area 40 isshown for comparison (B); it covers the su-pramarginal gyrus and may be regarded asthe prototype of the parietal cortex. Bothgranular and pyramidal layers are welldeveloped, and the radial striation runningthrough all layers is clearly visible.
The somatosensory cortex receives its affer-ent fibers from the ventral posteriorthalamic nucleus in a somatotopic organiza-tion that results in representation of thecontralateral body parts in specific corticalareas. The areas for throat and oral cavity lieabove the lateral sulcus (C1), and the areasfor face, arm, trunk, and leg are superior toit. The leg area reaches beyond the edge ofthe pallium to the median surface, wherethe representations of bladder, colon, andgenitals (C2) complete the sequence. Thiscreates a sensory homunculus. The cu-taneous regions of highly differentiatedsensibilities, such as hand and face, are rep-resented by particularly large cortical areas.The areas for distal parts of the limbs areusually larger than those for proximal parts.
According to clinical and electrophysiologi-cal studies, the superficial skin sensibility isrepresented by area 3, and the deep sensi-bility by area 2 (predominantly impulsesfrom joint receptors). In area 2, position andmovement of the limbs are constantly regis-tered.
Functional Significance of the ParietalCortex
The function of this area has become knownthrough mental deficiencies following in-jury of the parietal lobe.
Clinical Note: Various types of agnosia mayoccur. Although sensory impressions are per-ceived, the importance and characteristics of ob-jects are not recognized. Such disturbances mayalso affect tactile, optic, or acoustic sensations.There may be disturbances in symbolic thinkingwhen the parietal lobe (angular gyrus) of thedominant hemisphere (p. 262, A) is affected: theloss of understanding of letters or numbersmakes reading and writing, counting and calcu-lating impossible.Furthermore, disturbances of the body schememay be observed. They may involve the inabilityto distinguish between left and right. Also, one’sown paralyzed or nonparalyzed limbs may besensed as foreign limbs, for example, one’s ownarmmay be perceived as a heavy iron bar lying onthe chest. The disturbance may affect an entirebody half, which is then perceived as a differentperson, “my brother” (hemidepersonalization).The parietal cortex, which lies between the tactileand the optic cortices and is closely linked to bothby fiber connections, is thought to have a specialimportance for bringing about three-dimensionalspace perception. Injury to the parietal lobe mayresult in the destruction of this sense.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

251
Telenc
epha
lon
2
1
Parietal Lobe
I
B Area 40
C Somatotopic organization of thepostcentral area (according toPenfield and Rasmussen)
II
III
IV
V
VI
I
II
III
IV
V
VI
A Area 3, sensory cortex
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

252Te
lenc
epha
lon
Telencephalon: Neocortex
Temporal Lobe (A–C)
Auditory Cortex
The main convolutions on the lateral sur-face of the temporal lobe run mostly longi-tudinally. However, two convolutions on thesuperior surface, the transverse temporal gyri,or Heschl’s convolutions (C1), run trans-versely. They lie deep in the lateral sulcusand become visible upon removal of theoverlying parietal operculum. The cortex ofthe anterior transverse convolution is theterminal station of the acoustic radiation(p. 380) originating from the medial genicu-late body. The cortical areas of the twotransverse convolutions correspond to area41 (A) and area 42; they represent the audi-tory cortex. Like all receptive cortical areas,they are part of the koniocortex, or granularcortex (p. 244). The external granular layer(II) and, even more so, the internal granularlayer (IV) are rich in cells and very wide. Thepyramidal layers (III and V), on the otherhand, are narrow and contain only smallpyramidal cells. The cortex of area 21 (B) isshown for comparison; it covers the medialtemporal gyrus and represents a typicaltemporal cortex with prominent granularlayers, wide pyramidal layers, and distinctradial striation.
Electrical stimulation of area 22 close to thetransverse convolutions induces acousticsensations, such as humming, buzzing, andringing. The auditory cortex is organized ac-cording to tone frequencies (tonotopic or-ganization, p. 381C). In the human auditorycortex, the highest frequencies are assumedto be registered medially and the lowestfrequencies laterally.
Functional Significance of the TemporalCortex
Electrical stimulation of the remaining partsof the temporal lobe (performed during sur-gical treatment of temporal lobe epilepsy)induces hallucinations involving fragmentsof past experiences. The patients hear thevoices of people familiar to them in theiryouth. They relive momentary episodes oftheir own past. These are mainly acoustichallucinations and less often visual ones.
During stimulation of the temporal lobe,however, misinterpretation of the currentsituation may occur as well. A new situationmay thus appear as an old experience (déjàvu). Surrounding objects may move away orcome closer. The entire surroundings maytake on an uncanny or threateningcharacter.
Such phenomena occur only upon stimula-tion of the temporal lobe and cannot be elic-ited from any other part of the cortex. It istherefore assumed that the temporal cortexplays a special role in the conscious and un-conscious availability of one ’s own past andof things experienced in the past. Only if weare continuously aware of past experiencesare we able to judge and interpret new situa-tions correctly. Without this ability, wewould not find our way around in our sur-roundings. Therefore, the temporal cortexhas also been called the interpretative cor-tex.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

253
Telenc
epha
lon
1
Temporal Lobe
I
B Area 21
C Transverse temporal gyri,Heschl’s convolutions
II
III
IV
V
VI
II
III
I
IV
V
VI
A Area 41, auditory cortex
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

254Te
lenc
epha
lon
Telencephalon: Neocortex
Occipital Lobe (A–C)
The medial surface of the occipital lobe iscrossed by the horizontally running cal-carine sulcus (BC1); its deepest point corre-sponds to an eminence on the ventricularwall, the calcar avis (B2). Frontal sectionsthrough the occipital lobe clearly show afiber plate in the white matter, the tapetum(B3). It contains commissural fibers of thecorpus callosum that run through thesplenium and radiate in an arch into theoccipital lobe (forceps major, see p. 260,F16).
Visual Cortex (Striate Area)
Area 17 (AC4) is the terminal station of theoptic radiation and represents the visual cor-tex. It lies on themedial surface of the occip-ital lobe and spreads slightly to the convexpart at the pole. The cortex lines the cal-carine sulcus and extends further to its dor-sal and ventral lips. Area 17 is surrounded byarea 18 (A5) (left side of figure, the arrowshows the boundary between areas 17 and18) and area 19, which represent the opticintegration areas.
Like all receptive cortical areas, the cortex ofarea 17 is characterized by reduced pyra-midal layers and well-developed granularlayers. The cortex is very thin and set apartfrom thewhitematter by a cell-rich layer VI.The internal granule layer (IV) is divided bya zone deficient in cells. In myelin-stainedsections, this zone corresponds to Gennari’sline (B6). Because of the striate appearance,the visual cortex is therefore also calledstriate cortex. In the cell-deficient zone (IVb)are remarkably large cells, the giant stellatecells or Meynert’s cells. The two cellular lay-ers of the internal granular layer (IVa andIVc) contain very small granule cells. Theseare the layers with the highest cell densitywithin the entire cerebral cortex. Area 18exhibits a uniform granular layer made upof large granule cells. Area 19 forms a trans-ition to the parietal and temporal cortices.
Functional Organization of the VisualCortex
Electrophysiological studies on the visualcortex of experimental animals have shownthat there are two main types of neurons inthe striate area: simple cells and complexcells. A simple cell receives impulses from acell group of the retina. Its response isstrongest to narrow lines of light, to darklines against a light background, or tostraight lines of light/dark boundaries.Orientation of the lines is crucial: some cellsrespond only to horizontal lines of light,others only to vertical or oblique lines.
Complex cells also respond to lines of light ofa specific orientation. However, whereas asimple cell becomes excited only by its re-ceptive field, a complex cell responds tomo-bile lines of light that move over the retina:each complex cell is stimulated by a largenumber of simple cells. It is assumed thatthe axons of numerous simple cells termi-nate on one complex cell. The internalgranular layers consist almost entirely ofsimple cells, while the complex cells aggre-gate in the external granular layer. Morethan one half of all neurons in areas 18 and19 are complex or hypercomplex cells. It is as-sumed that they play a special role in shaperecognition.
Electric stimulation of the visual cortex(area 17) causes the sensation of light sparksor flashes. Stimulation of the parastriate andperistriate areas (areas 18 and 19) is thoughtto produce figures and shapes; it also causesthe gaze to turn (occipital eye field). Eyemovements induced by the occipital lobeare purely reflex, in contrast to the volun-tary movements directed by the frontal eyefield (p. 248, C).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

255
Telenc
epha
lon
1
4
2
6
3
1
5
4
Occipital Lobe
I
B Occipital lobe, frontal section C Median view of hemisphere,showing area 17
II
III
IVa
IVb
IVc
V
VI
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

256Te
lenc
epha
lon
Telencephalon: Neocortex
Occipital Lobe (continued)
Functional Organization of the VisualCortex (continued) (A, B)
Subdivision of the visual cortex intocolumns. In addition to the structural sub-division into cell layers, the visual cortexshows a functional subdivision intocolumns; these run vertically to the cell lay-ers and through the entire width of the cor-tex; they are 0.3 – 0.5mm in diameter. Eachcolumn is connected to a defined field of theretina.When the sensory cells of such a fieldare stimulated, all neurons of the respectivecolumn respond. Each column is connectedto a peripheral field of only one of the tworetinas. In the visual cortex (A1), columnsfor the right retina alternate with columnsfor the left retina (ocular dominancecolumns). Hence, the impulses from eachretina are segregated along the entire visualpathway.
The nerve fibers from the two correspond-ing halves of the retina terminate in thelateral geniculate body: the fibers from the leftretinal halves of both eyes (right halves ofthe visual fields) terminate in the leftgeniculate body (A2), and the fibers fromthe right retinal halves (left halves of thevisual fields) terminate in the right genicu-late body. The fibers from correspondingfields of both retinal halves terminate indifferent cell layers of the geniculate body:the uncrossed fibers (A3) from the ipsi-lateral retina extend to the second, third,and fifth layers, while the crossed fibers(A4) from the contralateral retina run to thefirst, fourth, and sixth layers. The neuronsreceiving the optic fibers from correspond-ing points of the two retinas lie along a linethat runs through all cell layers (projectioncolumn). Their axons project via the opticradiation (A5) to the visual cortex. Eachgeniculate fiber is thought to branch exten-sively and to terminate on several thou-sands of stellate cells of cortical layer IV.Fibers conducting the excitation of the ipsi-lateral retina extend to other columns thanthe fibers for excitation of the contralateralretina.
The organization of the visual cortex into verticalcolumns has been visualized by administer-ing 14C-deoxyglucose to experimental ani-mals and determining the varying distribu-tion of the substance by autoradiography.Excited neurons possess an increased me-tabolism and rapidly take up 14C-deoxyglu-cose, whereas resting cells do not.
The visual cortex of an experimental animalwith both eyes open (rhesus monkey)showed a bandlike distribution of theradioactive label, corresponding to the fa-miliar stratification of cells (B6). The dis-tribution of the label indicated that layers I,II, III, and V have a low glucose content,while layer VI has a higher content and layerIV the highest. When both eyes of the testanimal were closed, there were no signifi-cant differences between individual layers;instead, low-grade labeling of relatively uni-form distribution throughout the cortex(B7) was found. If one eye was open whilethe other one was closed, this methodyielded a series of columns running perpen-dicularly to the cell layers and showing in-tensely dark columns alternating with paleones (B8). The pale columns, where theneurons did not take up the label, repre-sented the retina of the closed eye. The darkcolumns contained newly incorporated 14C-deoxyglucose because they received inputfrom the retina of the open eye. Again, layerIV was most intensely labeled. The columnswere absent in a small area (B9) repre-senting the monocular zones of the retina,namely, the outermost margin of the retinaand the blind spot.
In addition to the ocular dominancecolumns, periodically arranged cell popula-tions have been demonstrated that respondin a characteristic fashion to the orientationof lines in the visual field (orientationcolumns). By demonstrating the enzyme cy-tochromoxidase in sections cut parallel tothe cerebral surface, periodically arrangedspots have been found that represent neu-rons responding to color stimuli mediatedby one eye (color columns).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

257
Telenc
epha
lon
6
7
9
8
4
2
5
1
3
Occipital Lobe
I
B Functional columns of the visual cortex(according to Kennedy, Des Rosiers, Sakurada,Shinohara, Reivich, Jehle, and Sokoloff)
II
IIIIV
VVI
I– IIIIV
V
VI
A Fiber connections between thegeniculate body and the visualcortex (according to Spatz)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

258Te
lenc
epha
lon
Telencephalon: Neocortex
Fiber Tracts (A–C)
A broad layer of white matter lies betweenthe cerebral cortex and the gray nuclei inthe depth. It consists of fiber bundles origi-nating from the neurons of the cortex orthose extending to the cortex and terminat-ing on the cortical neurons. There are threedifferent types of fiber systems:
! Projection fibers! Association fibers! Commissural fibers
Projection fibers provide connections be-tween the cerebral cortex and the subcorti-cal centers, either as ascending systems ter-minating in the cortex or as descending sys-tems extending from the cortex to thedeeper-lying centers. Association fibers pro-vide connections between different corticalareas of the same hemisphere. Commissuralfibers connect the cortices of both hemi-spheres; they are really nothing else but in-terhemispheric association fibers. However,commissural fibers often connect identicalregions of the two hemispheres.
Projection Fibers
The pathways descending from differentcortical areas merge and form a fanlikestructure known as the internal capsule.The ascending fibers pass through the inter-nal capsule and then radiate outward like afan. In this way, ascending and descendingfibers form a radiating crown of fibersbeneath the cortex, the corona radiata(A1).
The internal capsule (A2, B) appears in hor-izontal sections as an angle consisting of ananterior limb (B3), bordered by the head ofthe caudate nucleus (B4), globus pallidus(B5) and putamen (B6), and a posterior limb(B7), bordered by the thalamus (B8), globuspallidus and putamen (see p. 222, A). Be-tween both limbs lies the genu of the internalcapsule (B9). The different fiber tracts passthrough specific parts of the internal cap-sule. The frontopontine tract (B10) (redlines) and the anterior thalamic radiation(B11) (blue lines) pass through the anteriorlimb. The corticonuclear fibers supplying the
cranial nerve nuclei run through the genu ofthe internal capsule. The fibers of the corti-cospinal tract (red dots) pass through theposterior limb in a somatotopic organiza-tion: upper extremity, trunk, and lower ex-tremity. The thalamocortical fibers leadingto area 4 and the corticorubral and corti-cotegmental fibers coming from area 6 passthrough the same region. The caudal part ofthe posterior limb is occupied by the fibersof the central thalamic radiation (blue dots)(B12) which extend to the postcentral area.The fibers of the posterior thalamic radiation(B13) (light blue dots) and of the tem-poropontine tract (light red dots) (B14) passobliquely through the caudal part.
The most important projection pathwaysinclude the acoustic radiation and the opticradiation. The fibers of the acoustic radia-tion originate in the medial geniculatebody, extend across the lateral geniculatebody, and cross the internal capsule at theinferiormargin of the putamen. In thewhitematter of the temporal lobe, they ascend al-most vertically to the anterior transversegyrus (Heschl’s convolution) (pp. 252, 380).The optic radiation originates in the lateralgeniculate body. The fibers fan out into awide medullary lamina (A15) and run to thetemporal lobe, where they form the tem-poral genu (C16) of the visual pathway. Theythen pass in an arch around the inferiorhorn of the lateral ventricle and through thewhite matter of the occipital lobe to the cal-carine sulcus (C17).
A18 Corpus callosum.A19 Cerebral peduncle.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

259
Telenc
epha
lon
17
16
1
18
2
19
15
12
8
7
5
9
411
3
10
6
14
13
Projection Fibers
A Projection fibers, fiber prepara-tion (according to Ludwig andKlingler)
B Somatotopic organization of theinternal capsule
C Optic radiation during myelinmaturation (according to Flechsig)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

260Te
lenc
epha
lon
Telencephalon: Neocortex
Fiber Tracts (continued)
Association Fibers (A–D)
The connections between different corticalareas are very different in length. For thepurpose of simplification, we distinguishshort and long association fibers.
The short association fibers, or arcuatefibers (B), provide connections within acerebral lobe (B1) or from one convolutionto the next (B2). The shortest fibers connectadjacent cortical parts. They reenter thecortex after running just a short distancethrough the white matter. The layer of ar-cuate fibers lies immediately beneath thecortex.
The long association fibers connect differ-ent cerebral lobes and form compactbundles that can be recognized with thenaked eye. The cingulum (D3) is a strong sys-tem of shorter and longer fibers lying un-derneath the cingulate gyrus; it follows theentire course of the cingulate gyrus. Thelong fibers extend from the parolfactoryarea and the rostrum of the corpus callosumto the entorhinal area. The subcallosalfasciculus (superior occipitofrontalfasciculus) (CD4) lies dorsolaterally to thecaudate nucleus below the radiation of thecorpus callosum. Its fibers connect the fron-tal lobe with the temporal lobe and theoccipital lobe. Some of the fibers run to theinsula, others connect the frontal lobe withthe caudate nucleus. The superior longitudi-nal fasciculus (ACD5) lying dorsolaterally tothe putamen is a strong association bundlebetween frontal lobe and occipital lobewithfibers branching to the parietal lobe andtemporal lobe. The inferior occipitofrontalfasciculus (ACD6) passes through the ventralpart of the extreme capsule from the frontallobe to the occipital lobe. The inferior longi-tudinal fasciculus (C7) extends betweenoccipital lobe and temporal lobe. The unci-nate fasciculus (AC8) connects the temporalcortex with the frontal cortex. Its ventralpart provides a connection between the en-torhinal area and the orbital area of thefrontal lobe. Other fiber bundles are the ver-tical occipital fasciculus (AC9) and the orbi-tofrontal fasciculus (C10).
Commissural Fibers (E, F)
The interhemispheric association fibers passthrough the corpus callosum, the anteriorcommissure (p. 228, C), and the commissureof the fornix (p. 232, C25) to the con-tralateral hemisphere. The most importantcommissure of the neocortex is the corpuscallosum (E). Its curved rostral part is thegenu of the corpus callosum (E11) with thepointed rostrum (E12). It is followed by themiddle part, the trunk of the corpus callosum(E13), and the thickened end, the spleniumof the corpus callosum (E14). The fibers ofthe corpus callosum spread through thewhite matter of both hemispheres and formthe radiation of the corpus callosum. Thearched fibers passing through the genu ofthe corpus callosum and connecting bothfrontal lobes are called minor forceps (F15),those passing through the splenium andconnecting both occipital lobes are calledmajor forceps (F16).
We distinguish between homotopic andheterotopic interhemispheric fibers. Homo-topic fibers connect the same cortical areasin both hemispheres, while heterotopic fibersconnect different areas. The vast majority offibers of the corpus callosum is homotopic.Not all areas are connected to the same ex-tent with their counterpart in the otherhemisphere. The hand and foot parts of bothsomatosensory areas, for example, possessno interhemispheric fiber connections; norare the two visual cortices connected toeach other. However, very strong fiber con-nections exist between both areas 18, whichare regarded as optic integration areas.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

261
Telenc
epha
lon
5
9
68
15
13
12
11 14
16
6
5
3
4
1 2
5
4
10
8
6
7
9
Association Fibers and Commissural Fibers
A Long association fibers, fiberpreparation (according toLudwig and Klingler)
B Short association fibers
F Radiation of thecorpus callosum
C D
C, D Long association fibers, lateral view and frontal section
E Commissural fibers, corpus callosum
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

262Te
lenc
epha
lon
Telencephalon: Neocortex
Hemispheric Asymmetry (A, B)
Consciousness is dependent on the cerebralcortex. Only those sensory stimuli reachconsciousness that are transmitted to thecerebral cortex.
The faculty of speech is unique to humans.Internal speech is the prerequisite forthought, just as the spokenword is the basisfor communication and writing is the infor-mation transmitted over thousands ofyears. In the individual person, speech de-pends on the integrity of specific corticalareas that usually lie only in one hemi-sphere. This hemisphere is called the domi-nant hemisphere and is normally the left onein right-handed persons. In left-handedpersons, it may be the right or the left hemi-sphere, or the faculty may be represented inboth hemispheres. Thus, handedness is not areliable indication for dominance of thecontralateral hemisphere.
In the posterior region of the superior tem-poral gyrus of the dominant hemisphere liesWernicke’s speech center (A1). Injury to thisarea results in disturbed word comprehen-sion (receptive aphasia, or sensory aphasia).It is an integration area that is indispensablefor the continuous availability of learnedword patterns and for the interpretation ofheard or spoken words. Patients withsensory aphasia utter a senseless wordsalad (schizophasia), and the speech of otherpersons sounds to them like an incompre-hensible foreign language. Injury to theangular gyrus (A2), which borders on the su-pramarginal gyrus (A3) (see p. 212, A21), re-sults in the loss of the abilities to write(agraphia) and to read (alexia). Stimulationof adjacent areas (A5), especially of themiddle temporal gyrus, causes disturbancein spontaneous speech or writing. Broca ’sarea for motor speech coordination (A4) liesin the inferior frontal gyrus (p. 248, D).
Injury to the right (nondominant) hemi-sphere may cause disturbance of the visualand spatial orientation or the appreciationof music (amusia). Although speech is pre-served, the melody of the language and theemotional timbre of speech are affected.
Different ways of thinking have often beenassigned to one of the two hemispheres: theleft, dominant hemisphere is thought towork in a logical, rational, and analyticalway, while the right hemisphere is sup-posed to be integrative, synthetic, and intui-tive. These generalizations are largelyspeculative.
Transection of the corpus callosum (splitbrain). There are no changes in personalityor intelligence following the transection ofthe corpus callosum. The patients arecompletely normal in everyday life. Onlyspecial tests of the tactile and visual systemswill reveal any shortcomings (B).
Touch sensation of the left hand is regis-tered in the right hemisphere, that of theright hand in the left hemisphere (in right-handed persons, the dominant hemispherecontrols the ability to speak). Visual stimuliaffecting the left halves of each retina aretransmitted to the left hemisphere, whilethe stimuli for the right halves are trans-mitted to the right hemisphere (p. 256). Anexamination shows that right-handed per-sons with a severed corpus callosum canonly read with the left halves of the retinas.They cannot name objects perceived withthe right halves of the retina. However, theycan illustrate the use of these objectsthrough movements with their hands. Thesame phenomenon occurs when such per-sons have their eyes covered and receive anobject in their left hand: they are unable todescribe it verbally but can indicate its usethrough gestures. Objects perceived withthe right hand, or with the left halves of theretina which are connected to the “speak-ing” hemisphere, are immediately named.
Movements performed with one limb can-not be repeated with the contralateral limb,because the one hemisphere has not beeninformed about the impulses been sent outby the other hemisphere.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

263
Telenc
epha
lon
4
3
1
2
5
Hemispheric Asymmetry, Split Brain
A Areas for speech and writing in a right-handed person
B Split brain experiment (adaptedfrom Sperry and Gazzaniga)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

264Te
lenc
epha
lon
Telencephalon
Imaging Procedures
The development of imaging methods forclinical diagnostics has been extremelyrapid during the last 20 years. The currentlymost important methods for visualizing thecentral nervous system are:
! Contrast medium-assisted projection radi-ography for the visualization of bloodvessels and the ventricular system
! Computed tomography (CT)! Magnetic resonance imaging (MRI)
(p. 266)
Conventional radiography is mainly usedfor the visualization of bony structures. Ul-trasound is not suitable for the centralnervous system because sonic waves do notpenetrate the cranial bones in adults.However, examination by ultrasound doesplay a major role in diagnostics during earlychildhood. In addition to purely anatomicalimaging methods, there is also the possi-bility of visualizing functional parametersfollowing the injection of a suitable radioac-tive nuclide. The methods of nuclear medi-cine—single-photon emission computed to-mography (SPECT) and positron emission to-mography (PET) (p. 266)—can be used forstudying cerebral blood flow and metabolicactivity as well as for mapping specific re-ceptor systems. In the following section,some areas of application are briefly de-scribed for the most important procedures.
Contrast Radiography (A)
Radiation emitted by an X-ray source pene-trates the body. Absorbing structures(mainly the bones) appear as silhouettes onthe recording screen. The screen consistseither of a light-sensitive film or, to an in-creasing extent, of digital receiver arrange-ments. For the visualization of blood vessels,a contrast medium is injected into thepatient’s vessels between two recordings.By subtracting the images obtained prior toand after the injection of contrast medium,the (unchanged) bony structures are elimi-nated. The resulting subtraction imageselectively shows the blood vessels filled
with contrast medium. The contrast me-dium is usually administered to a selectedvessel by introducing a catheter into thevascular system of interest. The result is aselective high-resolution image of thecorresponding vascular segments (A).However, intra-arterial digital subtractionangiography (DSA) subjects the patient toconsiderable stress due to the introductionof the arterial catheter. Intravenous DSA,where contrast medium is injected into abrachial vein, produces images of lowercontrast because the contrast medium is di-luted by the blood volume; furthermore, itresults in nonspecific coloration of the ves-sels. Hence, it has not yet gained acceptancein the practice.
The same recording principle is used formyelography, in which contrast medium isadministered directly into the cerebrospinalfluid following puncture of the lumbarspine.
Computed Tomography (B)
Computed tomography (CT), or computedaxial tomography (CAT), also produces im-ages by means of X-rays. However, the im-aging principle is not based on the creationof silhouettes as in the projection method.Rather, individual transmissions through athin layer of tissue (“slice”) defined by thegeometry of the X-ray are recorded. Afterobtaining the absorption profiles at differ-ent projection angles (in Figure B shown asred, green, and blue areas), a two-dimen-sional image of the slice penetrated by theX-rays is computed by means of the filteredback projection. Only digital photocells ofvery high sensitivity are used as detectors.These allow the detection of even soft tissuestructures with great sensitivity and preci-sion, in addition to bony structures.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

265
Telenc
epha
lon
Subtraction Angiography, Computed Tomography
A Digital subtraction angiography (DSA): Selective visualization of the internal carotid artery,lateral view (Courtesy of M. Orszagh, Department of Radiodiagnostics, University of Freiburg,Germany)
B Principle of computed tomography (CT): After recording the absorption profiles (red, green,and blue areas) of fan-shaped X-rays, the image is reconstructed by the computer and dis-played on the monitor (Courtesy of J. Lautenberger, Department of Radiodiagnostics, Univer-sity of Freiburg, Germany)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

266Te
lenc
epha
lon
Telencephalon: Imaging Procedures
Computed Tomography (continued)
The main area of CT application for the CNSis above all the examination of patients withhead trauma: apart from high-precision im-aging of bony displacements, CT is also verysensitive for the visualization of hemor-rhages. Furthermore, all types of space-oc-cupying processes can be examined bymeans of CT because of the highly detailedprecision of the anatomical imaging.However, these areas of application are nowshifting increasingly to magnetic resonanceimaging owing to its higher sensitivity.
Magnetic Resonance Imaging (A, B)
Magnetic resonance imaging (MRI) is basedon the magnetic properties of the atomicnuclei in the body (mainly hydrogen nuclei)and uses radiofrequency resonance tech-nique to measure the magnetic moment in-duced by an external magnetic field. Varia-tions imposed on the magnetic field allowspatial visualization of the T1 and T2 relaxa-tion times, which are nuclear magneticproperties governed by the chemical formof the hydrogen atoms containing the nucleiin the tissues. These relaxation times, whichmainly reflect the ratio of free to boundwatermolecules, produce a contrastmecha-nism that delineates not only different tis-sues but also pathological tissue changes.The images in A and B display the same ana-tomic section and illustrate the exquisitecontrast achievable by MRI, which makes itan excellent method for the identification ofpathological processes. The static magneticand electromagnetic fields used with MRIhave not been found to be hazardous topatients. Furthermore, MRI has the advan-tage over CT that the images can be recon-structed in any plane. The spatial resolutionof MRI is in the range of 0.7 – 1.0mm for asection thickness of 5mm. It takes severalminutes to acquire and process the data forthe axial, coronal, and sagittal sections con-stituting a standard cranial MRI.
PET and SPECT (C, D)
Both of these imaging methods of nuclearmedicine are based on detecting the radia-tion of radioactive nuclides administeredprior to the investigation. Single-photonemission computed tomography (SPECT)uses gamma-emitters. Detection of thegenerated radiation is carried out by meansof photocells in a similar way as in CT. Thegeometry of the detected radiation,however, is not predetermined by the ar-rangement of transmitter and receiver; in-stead, both intensity and direction of the re-ceived gamma rays must be determined.This yields a spatial resolution of about1 cm, clearly less favorable than when usingcomputed tomography. Hence, high-resolu-tion anatomical images cannot be obtained;nevertheless, SPECT is suitable for the deter-mination of cerebral blood flow by means oftime-resolved kinetics of the detected sig-nals.
Positron emission tomography (PET) usesemitters of beta particles (positrons). Thepositrons almost instantly combine withelectrons in the immediate surrounding,with both particles undergoing mutualannihilation and giving rise to two gammarays of 511keV energy emitted in oppositedirections (C). The geometry of thesegamma rays can be much better definedthan in the case of SPECT, namely, by rec-ording the coincidence of the two gammaphotons received in the detector ring.Therefore, the spatial resolution of PET liesat 2 – 4mm, though this is clearly still not asgood as in the case of CT and MRI. Throughincorporation of suitable radioisotopes intobiologically active molecules, PET also al-lows us to carry out metabolic studies in ad-dition to measurements of blood flow (D), orto selectively visualize receptor systems (forexample, dopamine receptors).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

267
Telenc
epha
lon
MRI, PET
A, B Magnetic resonance imaging(MRI): Two different scans ofthe head at the level above theventricles
A Spin–lattice relaxation, T1 B Spin–spin relaxation, T2
C Principle of positron emission tomography (PET):Detection of gamma rays emitted simultaneously in opposite directionsduring the decay of a radioisotope (Courtesy of F. Jüngling, Departmentof Radiodiagnostics, University of Freiburg, Germany)
D PET scans of four parallel slices (intensity of glucose utilization in the brain)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Cerebrovascular andVentricular Systems
Cerebrovascular SystemArteries 270Veins 276
Cerebrospinal Fluid Spaces 280Meninges 288
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

270Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems
Cerebrovascular System
Arteries
The brain is supplied by four large arteries:the two internal carotid arteries and the twovertebral arteries (see vol. 2).
The internal carotid artery (A1) (p. 272)passes through the dura mater medially tothe anterior clinoid process of the sphenoidbone. Between the subarachnoid and the piamater, it gives off the superior hypophysialartery (p. 200, E9), the ophthalmic artery,the posterior communicating artery (A16),and the anterior choroidal artery (A2). Itthen divides into two large terminalbranches, the anterior cerebral artery (A4)and the middle cerebral artery (A7).
The anterior choroidal artery (A2) runs alongthe optic tract to the choroid plexus (A3) inthe inferior horn of the lateral ventricle. Itgives off fine branches that supply the optictract, the temporal genu of the optic radia-tion, the hippocampus, the tail of the cau-date nucleus, and the amygdaloid body.
The anterior cerebral artery (A4) runs on themedial surface of the hemisphere across thecorpus callosum. The two anterior cerebralarteries are interconnected by the anteriorcommunicating artery (A5). The long centralartery (recurrent artery of Heubner) (A6)branches off shortly after the communicat-ing artery. It passes through the anteriorperforated substance into the brain andsupplies the anterior limb of the internalcapsule, the adjacent region of the head ofthe caudate nucleus, and the putamen.
The middle cerebral artery (A7) runs laterallytoward the lateral sulcus; above the per-forated substance it gives off 8 – 10 striatebranches that enter the brain. At the en-trance of the lateral fossa, it divides intoseveral large branches that spread over thelateral surface of the hemisphere.
The two vertebral arteries (A8) arise fromthe two subclavian arteries and enter thecranial cavity through the foramen mag-num; they unite at the upper margin of themedulla oblongata to form the unpaired
basilar artery (A9). The latter ascends alongthe ventral surface of the pons and bifur-cates at the upper margin of the pons intothe two posterior cerebral arteries (A10). Thevertebral artery gives off the posterior infe-rior cerebellar artery (A11), which suppliesthe lower surface of the cerebellum and thechoroid plexus of the fourth ventricle. Thebasilar artery gives off the anterior inferiorcerebellar artery (A12), which also suppliesthe lower surface of the cerebellum as wellas the lateral parts of medulla and pons. Thelabyrinthine artery (A13) runs as a finebranch together with the facial nerve andthe vestibulocochlear nerve through the in-ternal acoustic meatus into the inner ear. Itmay arise from the basilar artery or from theanterior inferior cerebellar artery. Numer-ous small branches reach as pontine arteries(A14) directly into the pons. The superiorcerebellar artery (A15) runs along the uppermargin of the pons and extends deep intothe cisterna ambiens around the cerebralpeduncles to the dorsal surface of the cere-bellum.
The circle ofWillis. The posterior communi-cating arteries (A16) connect on both sidesthe posterior cerebral arteries with the in-ternal carotid arteries so that the blood flowof the vertebral arteries can communicatewith the carotid circulation. The anteriorcerebral arteries are interconnectedthrough the anterior communicating artery.This way, a closed cerebral arterial circle iscreated at the base of the brain. However,the anastomoses are often so thin that theydo not allow for a significant exchange ofblood. Under conditions of normal in-tracranial pressure, each hemisphere is fedby the ipsilateral internal carotid artery andthe ipsilateral posterior cerebral artery.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

271
Cerebr
ovas
cularan
dVen
tricular
System
s
65
1
7
4
16
10 2
3
9
8
11
15
14
12
10
13
Arteries of the Base of the Brain
A Arteries of the base of the brain
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

272Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems: Arteries
Internal Carotid Artery (A–C)
The internal carotid artery (C1) can be sub-divided into a cervical part (between thecarotid bifurcation and the base of theskull); a petrosal part (in the carotid canal ofthe petrous bone); a cavernous part (withinthe cavernous sinus, p. 104, A7); and a cere-bral part. The cavernous and cerebral seg-ments of the artery form an S-shaped curve(carotid siphon) (C2). The inferior hypophy-sial artery (p. 200, E10) comes off in thecavernous part, followed by small branchesto the dura and to cranial nerves IV and V.After giving off the superior hypophysialartery (p. 200, E9), the ophthalmic artery,and the anterior choroidal artery in the cere-bral part, the internal carotid artery dividesinto two large terminal branches, the ante-rior cerebral artery and the middle cerebralartery.
The anterior cerebral artery (BC3) turnstoward the longitudinal cerebral fissureafter giving off the anterior communicatingartery. The postcommunical part of theartery (pericallosal artery) (BC4) runs on themedial hemispheric wall around the ros-trum and the genu of the corpus callosum(B5) and along the superior surface of thecorpus callosum toward the parieto-occipi-tal sulcus. It gives off branches to the basalsurface of the frontal lobe (medial fron-tobasal artery) (B6). The other branchesspread over the medial surface of the hemi-sphere; they are the frontal branches (BC7),the callosomarginal artery (BC8), and theparacentral artery (B9) which supplies themotor area for the leg.
The middle cerebral artery (AC10) extendslaterally to the bottom of the lateral fossa,where it divides into several groups ofbranches. The artery is divided into threesegments. The sphenoidal part gives off thecentral arteries (fine branches for the stri-atum, thalamus, and internal capsule), andthe insular part gives rise to the short insulararteries (C11) for the insular cortex, thelateral frontobasal artery (A12), and the tem-poral arteries (A13) for the cortex of thetemporal lobe. The last segment, the termi-nal part, is formed by the long branches for
the cortex of the central region and theparietal lobe (AC14). There are considerablevariations regarding the bifurcation andcourse of individual arteries.
The posterior cerebral artery (BC15) developsas a branch of the internal carotid artery.In adults, however, it lies relatively farcaudally. It is only connected with the inter-nal carotid artery through the thin posteriorcommunicating artery. Because most of itsblood comes from the vertebral arteries, it isregarded as part of their supply area; thelatter consists of the subtentorial parts ofthe brain (brain stem and cerebellum) andthe supratentorially located occipital lobe,the basal part of the temporal lobe, and thecaudal parts of the striatum and thalamus(tentorium of the cerebellum, p. 288, B5). Allparts of the frontal brain lying in front of itreceive their blood from the internal carotidartery.
The posterior cerebral artery ramifies on themedial surface of the occipital lobe and onthe basal surface of the temporal lobe. Itgives off the posterior choroidal artery forthe choroid plexus of the third ventricle aswell as fine branches for the striatum andthalamus. The ophthalmic artery (C16) runsfrom the internal carotid artery to the orbit.
Carotid angiogram (arterial phase). A dia-gram of a carotid angiogram is shown in C.For diagnostic purposes, a contrast mediumis injected into the internal carotid artery.Within a few seconds, the contrast mediumspreads through the supply area of theartery. A radiograph taken immediately il-lustrates the arterial vascular tree. Whenviewing arteriographic images, it must betaken into consideration that all blood ves-sels are seen in a single plane (see contrastradiography, p. 264, A).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

273
Cerebr
ovas
cularan
dVen
tricular
System
s
14 14
14
948
7
5
63
15
14
1310
12
8
7
3
11
4
10 1516 2
1
Internal Carotid Artery
A Arteries of the brain,lateral view
B Arteries of the brain, medialsurface of the hemisphere
C Cerebral angiogram, arterial phase(according to Krayenbühl and Richter)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

274Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems: Arteries
Areas of Blood Supply
Anterior Cerebral Artery (A, B)
The short branches of the anterior cerebralartery (AB1) extend to the optic chiasm, tothe septum pellucidum, to the rostrum andgenu of the corpus callosum, while the longrecurrent artery of Heubner (long centralartery) runs to themedial part of the head ofthe caudate nucleus and to the anterior limbof the internal capsule. The corticalbranches supply the medial parts of thebase of the frontal brain and the olfactorylobe, and also the frontal and parietal cortexon the medial surface of the hemisphereand the corpus callosum as far as thesplenium. The supply area of the artery ex-tends beyond the edge of the pallium to thedorsal convolutions of the convexity.
Middle Cerebral Artery (A, B)
The striate branches of the middle cerebralartery (AB2) terminate in the globus pal-lidus, in parts of the thalamus, in the genuand parts of the anterior limb of the internalcapsule. The branches given off by the insu-lar arteries ramify in the insular cortex andin the claustrum and reach into the externalcapsule. The area supplied by the corticalbranches includes the lateral surface offrontal, parietal, and temporal lobes andalso a large part of the central region and thetemporal pole. The branches not onlysupply the cortex but also the white matteras far as the lateral ventricle, including thecentral part of the optic radiation.
Posterior Cerebral Artery (A, B)
The posterior cerebral artery (AB3) gives offfine short branches that supply the cerebralpeduncles, pulvinar, geniculate bodies,quadrigeminal plate, and splenium of thecorpus callosum. The cortical supply areaoccupies the basal part of the temporal lobeand the occipital lobe with the visual cortex(striate area); however, the latter is alsoreached in the region of the occipital pole bythe most inferior branches of the middlecerebral artery.
Blood Supply for the Nuclei ofDiencephalon and Telencephalon (C, D)
The recurrent artery of Heubner and thestriate branches (D4) of the middle cerebralartery (D5) supply the head of the caudatenucleus, putamen, and internal capsule. Therole of the anterior choroidal artery (C6) inthe supply of deep structures varies; itsbranches extend not only to the hippocam-pus and amygdaloid body but also to partsof the pallidum and thalamus. The rostralpart of the thalamus receives a branch fromthe posterior communicating artery (C7),the thalamic branch (C8). The middle andcaudal parts of the thalamus are supplied bythe basilar artery (C9), from where directbranches (C10) may run to the thalamus.Other fine thalamic branches are given offby the posterior choroidal artery (C11) andthe posterior cerebral artery (C12).
Vascularization. The large cerebral vesselslie without exception on the surface of thebrain. They give off small arteries and arteri-oles that penetrate vertically into the brainand ramify. The capillary network is verydense in the gray matter but far less so inthe white matter.
Clinical Note: Sudden obstruction of an arteryby a thrombus, air bubble, or fat droplet in thebloodstream (embolism) causes the death of braintissue in the supply area of the affected artery.The anastomoses between the vascular areas arenot adequate to supply the affected regionthrough nearby areas in case of a sudden obstruc-tion. Especially affected are the middle cerebralartery and its branches.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

275
Cerebr
ovas
cularan
dVen
tricular
System
s
2
2
3
1
2
3
1
1
1
8
6
12
11 4
57
109
Areas of Blood Supply
A Regions supplied by the arteries,lateral view
B Regions supplied by thearteries, medial view
C Arterial supply of the thalamus (accordingto Van den Bergh and Van der Eeken)
D Arterial supply to the neostratum
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

276Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems
Veins (A–C)
The major veins lie on the surface of thebrain in the subarachnoid space; some deepveins run beneath the ependyma. The cere-bral veins do not possess valves. They ex-hibit considerable variations with respect tocourse and drainage. Quite often there areseveral small vessels instead of a singlemajor vein. The cerebral veins are dividedinto two groups: the superficial cerebral veinswhich drain the blood into the sinuses of thedura mater (see vol. 2), and the deep cerebralveins which drain the blood into the greatcerebral vein (great vein of Galen).
Superficial Cerebral Veins
We distinguish between the group of supe-rior cerebral veins and the group of inferiorcerebral veins.
The superior cerebral veins (AC1), totalingabout 10 – 15 veins, collect the blood fromthe frontal and parietal lobes and carry itinto the superior sagittal sinus (BC2). Theyrun within the subarachnoid space andempty into the lateral lacunae (BC3),pouchlike cavities of the superior sagittalsinus. For a short distance, they passthrough the subdural space. Here, the thin-walled veins can easily rupture during headinjury and bleed into the subdural space(subdural hematoma). Strangely, the veinsempty into the superior sagittal sinus at anoblique angle against the dominating bloodflow in the sinus.
The inferior cerebral veins receive theblood from the temporal lobe and from thebasal regions of the occipital lobe; theyempty into the transverse sinus and the su-perior petrosal sinus. The largest and mostconsistent of these veins is the superficialmiddle cerebral vein (AC4) located in thelateral sulcus; it often consists of severalvenous trunks. It drains the blood frommost of the lateral aspect of the hemisphereinto the cavernous sinus (p. 104, A7).
The superior and inferior cerebral veins areinterconnected only by a few anastomoses.The most important one is the superior
anastomotic vein (Trolard’s vein) (AC5); itempties into the superior sagittal sinus andis connected with the superficial middlecerebral vein. The central vein (C6) locatedin the central sulcus may also form anasto-moses with the middle cerebral vein. Theinferior anastomotic vein (Labbé’s vein,Browning ’s vein) (AC7) connects the super-ficial middle cerebral vein with the trans-verse sinus.
Carotid angiogram (venous phase). A dia-gram of the venous phase of a carotid angio-gram is shown in C (for arterial phase, seep. 272). A radiograph taken only secondsafter injection of the contrast mediumshows its drainage via the venous vasculartree. Superficial and deep veins are seen in asingle plane.
Deep cerebral veins (S. 278):
BC8 Great cerebral vein(great vein of Galen).
C9 Internal cerebral vein.C10 Thalamostriate vein
(terminal vein).C11 Vein of the septum pellucidumC12 Interventricular foramen
(foramen of Monro).C13 Basal vein (Rosenthal’s vein).BC14 Straight sinus.BC15 Inferior sagittal sinus.BC16 Confluence of sinuses
(p. 104, A19).
(For the large venous channels, the sinusesof dura mater, see vol. 2.)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

277
Cerebr
ovas
cularan
dVen
tricular
System
s
5
4
3
7
2
1
1
15
8
14
16
9
3
1 5 6
1
89
15
2
714
16
13
4
10
1112
Superficial Veins
A Veins of the brain, lateral view
B Veins of the brain, medialsurface of the hemisphere
C Cerebral angiogram, venous phase(according to Krayenbühl and Richter)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

278Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems: Veins
Deep Cerebral Veins (A, B)
The deep cerebral veins collect the blood fromthe diencephalon, the deep structures of thehemispheres, and the deep white matter. Inaddition, there are thin transcerebral veinsrunning along the fibers of the corona radi-ata from the outer white matter and fromthe cortex. They connect the superficialdrainage areas with the deep ones. The deepcranial veins empty their blood into thegreat cerebral vein (great vein of Galen). Thedrainage system of the deep veins is there-fore also known as the system of the greatcerebral vein.
The great cerebral vein (AB1) is a shortvascular trunk formed by the confluence offour veins, namely, the two internal cerebralveins and the two basal veins. It curvesaround the splenium of the corpus callosumand empties into the straight sinus. Veinsfrom the surface of the cerebellum and fromthe occipital lobe (B2) may drain into it.
The basal vein (Rosenthal’s vein) (AB3)arises at the anterior perforated substance(A4) by junction of the anterior cerebral veinand the deep middle cerebral vein.
The anterior cerebral vein (A5) receives itsblood from the anterior two-thirds of thecorpus callosum and the adjacent convolu-tions. It extends around the genu of the cor-pus callosum to the base of the frontal lobe.The deep middle cerebral vein (A6) arises inthe insular area and receives veins from thebasal parts of putamen and globus pallidus.
The basal vein crosses the optic tract and as-cends in the cisterna ambiens around thecerebral peduncle (A7) to below thesplenium, where it empties into the greatcerebral vein. Along its course it receivesnumerous venous tributaries, namely, veinsfrom the optic chiasm and the hypo-thalamus, the interpeduncular vein (A8), theinferior choroid vein (A9) from the choroidplexus (A10) of the inferior horn, and veinsfrom the internal segment of the globus pal-lidus and from the basal parts of thethalamus.
The internal cerebral vein (AB11) arises atthe interventricular foramen (foramen ofMonro) by junction of the vein of the septumpellucidum, the thalamostriate vein, and thesuperior choroid vein.
The thalamostriate vein (terminal vein)(B12) runs in the terminal sulcus betweenthalamus (B13) and caudate nucleus (B14)in rostral direction to the interventricularforamen. It receives venous tributaries fromthe caudate nucleus, from the adjacentwhite matter, and from the lateral corner ofthe lateral ventricle. The vein of the septumpellucidum (B15) receives venous branchesfrom the septum pellucidum (B16) and fromthe deep frontal white matter. The choroidvein (B17) runs with the choroid plexus tothe inferior horn. In addition to the vesselsof the plexus, it receives the veins from thehippocampus and from the deep temporalwhite matter.
The internal cerebral vein extends from theinterventricular foramen across the medialsurface of the thalamus at the margin of theroof of the diencephalon to the region of thepineal gland, where it unites with the con-tralateral internal cerebral vein and thebasal veins to form the great cerebral vein.Along its course it receives tributaries fromthe fornix (B18), from the dorsal parts of thethalamus, from the pineal gland (epiphysis)(B19) and, variably, from the deep whitematter of the occipital lobe.
In summary, the dorsal parts of thalamus,pallidum, and striatum drain into the inter-nal cerebral vein, while the ventral partsdrain into the basal vein.
Clinical Note: Obstruction of a cerebral veincauses congestion and hemorrhage in the affectedregion. In case of birth trauma, rupture of thethalamostriate vein in newborns may lead tohemorrhage into the ventricles.
A20 Superficial middle cerebral vein.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

279
Cerebr
ovas
cularan
dVen
tricular
System
s
1
15
14
1618
12 13
3
2
17
11
19
20
4
87
11
5
6
3
1
10
9
1
15
14
1618
12 13
3
2
17
11
19
20
4
87
11
5
6
3
1
10
9
Deep Veins
A Basal veinsof the brain
B Deep veins of the brain,view from above
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

280Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems
Cerebrospinal Fluid Spaces
Overview
The central nervous system (CNS) iscompletely surrounded by the cerebrospinalfluid (CSF), which also fills the inner cavitiesof the brain, the ventricles. We thereforedistinguish between internal and externalcerebrospinal fluid spaces. They communi-cate with each other in the region of thefourth ventricle.
Internal Cerebrospinal Fluid Spaces (A–C)
The ventricular system consists of four ven-tricles, namely, the two lateral ventricles(A1) of the telencephalon, the third ventricle(A–C2) of the diencephalon, and the fourthventricle (A–C3) of the rhombencephalon(pons and medulla oblongata). The twolateral ventricles are connected with thethird ventricle by the interventricular foramen(foramen of Monro) (AC4) located on eachside in front of the thalamus. The third ven-tricle, in turn, communicates with thefourth ventricle through a narrow channel,the cerebral aqueduct (aqueduct of Sylvius)(A–C5).
Owing to the rotation of the hemispheres(p. 208) the lateral ventricle has a semi-circular configuration with a caudallydirected spur. We distinguish the followingparts:
! The anterior horn, frontal horn (BC6), inthe frontal lobe is bordered laterally bythe head of the caudate nucleus, mediallyby the septum pellucidum, and dorsallyby the corpus callosum
! The narrow central part (BC7) above thethalamus
! The inferior horn, temporal horn (BC8), inthe temporal lobe
! The posterior horn, occipital horn (BC9), inthe occipital lobe
The lateral wall of the third ventricle isformed by the thalamus with the inter-thalamic adhesion (C10) (p. 10, C18) and thehypothalamus. The optic recess (C11) andthe infundibular recess (C12) project ros-trally, and the suprapineal recess (C13) andthe pineal recess (C14) do so caudally.
The fourth ventricle creates a tent-shapedspace above the rhomboid fossa betweencerebellum and medulla oblongata; on bothsides, it sends out a long lateral recess(BC15). At the end of each lateral recess isthe lateral aperture of the fourth ventricle(foramen of Luschka, foramen of Key andRetzius). At the attachment of the inferiormedullary velum lies the median aperture(foramen of Magendie) (p. 282, D14).
External Cerebrospinal Fluid Spaces (A)
The external cerebrospinal fluid space liesbetween the two layers of the leptome-ninges. It is delimited on the inside by thepia mater and on the outside by thearachnoidea mater (subarachnoid space orcavity; p. 288, A13). The space is narrowover the convexity of the hemispheres andwidens only in several areas at the base ofthe brain to form cisterns. Whereas the piamater adheres closely to the surface of theCNS, the arachnoidea spans across sulci andfossae; in regions of deep indentations thisarrangement creates larger spaces filledwith CSF, the subarachnoid cisterns. Thelargest space is the cerebellomedullary cis-tern (A16) between cerebellum andmedullaoblongata. In the corner formed by dien-cephalic floor, cerebral peduncles and ponslies the interpeduncular cistern (A17), andthe chiasmatic cistern (A18) lies in front of itnear the optic chiasm. The cerebellar sur-face, quadrigeminal plate, and pineal glanddelimit the cisterna ambiens (A19), which istraversed by a loose network of connectivetissue.
Circulation of Cerebrospinal Fluid (A)
The cerebrospinal fluid is produced by thechoroid plexus (p. 282). It flows from thelateral ventricles into the third ventricle,and from there through the aqueduct intothe fourth ventricle. Here it passes throughthe median and lateral apertures into theexternal cerebrospinal fluid space. Drainageof CSF into the venous circulation takesplace partly in the arachnoid granulations(p. 288, A15) that protrude into the venoussinuses or lateral lacunae, and partly at theexits of the spinal nerves where the fluid en-ters into the dense venous plexuses and intothe nerve sheaths (drainage into the lym-phatic circulation).Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

281
Cerebr
ovas
cularan
dVen
tricular
System
s
2
1
3
4
5
19
186
7
8
2
3
9
9
7
64
10
2 1314
5
8
3
15
1112
5
17
16
15
2
1
3
4
5
19
186
7
8
2
3
9
9
7
64
10
2 1314
5
8
3
15
1112
5
17
16
15
2
1
3
4
5
19
186
7
8
2
3
9
9
7
64
10
2 1314
5
8
3
15
1112
5
17
16
15
Cerebrospinal Fluid Spaces and Circulation
A Cerebrospinal fluid spaces
B Ventricular system,view from above
C Ventricular system,lateral view
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

282Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems: Cerebrospinal Fluid Spaces
Choroid Plexus
Lateral Ventricles (A, C)
The choroid plexus consists of convolutionsof blood vessels protruding into the ven-tricles from specific parts of the wall. Thearea of the wall adhering to the medial sur-face of the hemisphere (choroid lamina)(A1) becomes thinner during embryonicdevelopment and is pushed into theventricular lumen by the vascular loops ofthe pia mater (A2). At the beginning ofdevelopment, all vascular convolutions arecovered by a thin layer of hemispheric wall.The latter finally turns into a layer ofepithelial cells, the plexus epithelium. Thus,the adult choroid plexus consists of twocomponents, namely, the vascularized con-nective tissue of the pia mater and theplexus epithelium (transformed hemi-spheric wall). Once the choroid plexus hasinvaginated into the ventricular cavity, it re-mains connected with the external piamater only through a narrow channel, thechoroid fissure (A3). When the choroidplexus is removed, the thinned parts of thehemispheric wall tear away at this location.The line of attachment is called taenia(band). One such line is attached to the for-nix (p. 230, A6) and to the fimbria of the hip-pocampus (p. 230, ACD5); it is known as thetaenia fornicis (taenia of the fornix) (C4).The other line runs along the lamina affixa(p. 170, D15, E16) and is known as the taeniachoroidea (choroid line) (C5).
Owing to the rotation of the hemispheres(p. 208), the choroid plexus forms a semi-circle along the medial wall of the ventricle,reaching from the interventricular foramen(foramen of Monro) via the central part (C6)to the inferior horn (C7). The anterior horn(C8) and the posterior horn (C9) do not con-tain a plexus.
Tela choroidea (A–C)
When the hemispheres overgrow the dien-cephalon, the leptomeninges of both partsof the brain come to lie on top of each otherto form a duplication (A10), the tela choroidea(B), a connective-tissue plate spanning be-
tween the telencephalic hemispheres andthe diencephalon. At its lateral margins, thepia mater forms the vascular convolutionfor the choroid plexus of the lateral ven-tricles. In themiddle, the tela covers the roofof the third ventricle (tela choroidea of thethird ventricle) (BC11). In this region, tworows of convoluted blood vessels protrudeinto the lumen of the third ventricle andform the choroid plexus of the third ventricle.When removing the ventricular roof, thetaenia thalami (C12) remains as line of at-tachment running along the medullary striaacross the thalamus.
Fourth Ventricle (D, E)
Above the fourth ventricle, the tela choroideaof the fourth ventricle is also formed as a du-plication of the pia mater by apposition ofthe lower surface of the cerebellum to thesurface of the rhombencephalon (E). Theroof of the rhombencephalon becomes re-duced to a thin epithelial layer and ispushed into the ventricle by the vascularloops originating from the tela. The telachoroidea of the fourth ventricle consist ofpia mater only, as the arachnoidea does notadhere to the surface of the cerebellum butspans the cerebellomedullary cistern. At theobex (D13), the attachment of the tela abovea narrow medullary fold, lies the medianaperture (foramen of Magendie) (D14). Onboth sides open the lateral apertures(foramina of Luschka), through which thelateral end of the choroid plexus protrudes(Bochdalek’s flower basket) (D15).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
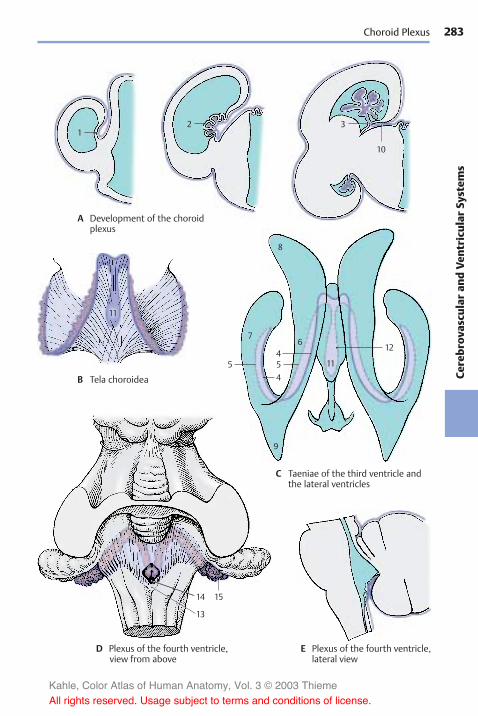
283
Cerebr
ovas
cularan
dVen
tricular
System
s
1
11
2 3
10
8
7
5
1514
13
9
454
6
11
12
Choroid Plexus
A Development of the choroidplexus
B Tela choroidea
C Taeniae of the third ventricle andthe lateral ventricles
D Plexus of the fourth ventricle,view from above
E Plexus of the fourth ventricle,lateral view
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

284Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems: Cerebrospinal Fluid Spaces
Choroid Plexus (continued) (A, B)
The treelike ramification of the choroidplexus (A) creates a very large surface. Eachbranch contains one or more vessels, suchas arteries, capillaries, and thin-walledvenous caverns. The vessels are surroundedby a loose meshwork of collagen fibers (B1)which, in turn, is covered by the plexusepithelium (B). The plexus epithelium con-sists of a single layer of cuboidal cells thatcarry a fine brush border on their apical sur-face. Their cytoplasm contains vacuoles andcoarse granules as well as inclusions of lipidand glycogen.
The choroid plexus is the site of cerebrospi-nal fluid production. Fluid is transferredfrom the vascular system of the choroidplexus through the epithelium into the ven-tricles. Whether this occurs through secre-tion by the plexus epithelium or throughdialysis (a form of filtration) has yet to beestablished.
Like the pia-arachnoid and dura, the choroidplexus is richly innervated (the meningesare innervated by the trigeminal nerve,vagus nerve, and autonomic fibers). Thechoroid plexus and meninges are thereforesensitive to pain, whereas the brain tissueitself is largely insensitive.
Ependyma (C, D)
Thewalls of the ventricular system are linedby a single cell layer, the ependyma (C).Each ependymal cell has a basal process, theependymal fiber, which extends into thebrain. The cell surface facing the ventricularlumen often carries several cilia, with thebasal bodies, or kinetosomes (C2), lined upbeneath the cell surface.
In the electron-microscopic image, theventricular surface of the ependymal cellsexhibits numerous vesicle-containing pro-trusions (D3). The cilia (D4) contain micro-tubules in the characteristic 9 + 2 arrange-ment: two single microtubules in the center(D5) and ninemicrotubule doublets (D6) ar-ranged around them. The basal body of eachcilia is surrounded by a dense zone (D7) into
which numerous short rootlets (D8) radiate.A basal foot (D9) is located on one side of thebasal body; it may play a role in directingthe beat of the cilia. The ependymal cells areinterconnected along their lateral surfacesby zonulae adherentes (adherent junctions)(D10) and by zonulae occludentes (tightjunctions) (D11); the latter seal the cere-brospinal fluid space against the brain. Neu-ronal processes (D12) run between theependymal cells. The layer underneath theependyma consists of radially or horizon-tally running glial fibers (C13) and containsonly few cells. Below it lies the subependy-mal cell layer (C14). It contains undifferen-tiated cells in addition to astrocytes. Ac-cording to recent studies, not only glial cellsbut also neurons are generated herethroughout life. Intensive investigations areunder way to test whether neuronal stemcells of the subependymal zone can be usedfor neuronal replacement in various formsof neuronal degeneration.
The structure of the ventricular wall varieswidely in different regions. The ependymalcover or the subependymal layer of glialfibers may be completely absent in certainareas. The subependymal glial cell layer ismost prominent above the head of the cau-date nucleus and at the base of the anteriorhorn but is absent above the hippocampus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

285
Cerebr
ovas
cularan
dVen
tricular
System
s
1
3
56
712
8
9
2
13
10
11
14
4
Choroid Plexus, Ependyma
A Choroid plexus
B Plexus epithelium
C Ependyma and subependymal zone D Ependymal cell, electron-microscopicdiagram (according to Brightman and Palay)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

286Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems
Circumventricular Organs (A–D)
In lower vertebrates, the ependyma hassecretory functions and probably also re-ceptor functions. This has resulted in thedevelopment of special structures that canbe demonstrated also in mammals. Suchcircumventricular organs include:
! The vascular organ of the terminal lamina! The subfornical organ! The area postrema! The paraphysis! The subcommissural organ
These organs are regressed in humans, andsome of them (paraphysis and subcommis-sural organ) appear only temporarily duringembryonic development. Their functionsare unknown. It is assumed that they play arole in the regulation of CSF pressure andcomposition and that they are somehow re-lated to the neuroendocrine system of thehypothalamus.What is striking is their loca-tion at narrow passages of the ventricularsystem, their high vascularization, and thepresence of fluid-filled spaces.
Vascular Organ of the Terminal Lamina(A, D)
This organ (A, D1) lies in the terminallamina which spans as rostral end of thethird ventricle between the anterior com-missure and the optic chiasm. We distin-guish between a highly vascularized outerzone beneath the pia mater and an innerzone consisting mainly of glia. The vesselsform a dense plexus with sinuslike dilata-tions. In the inner zone run nerve fibersfrom the supraoptic nucleus that containGomori-positive material (Herring bodies,p. 204, B5). It also receives peptidergic fibersfrom the hypothalamus.
Subfornical Organ (B, D)
The subfornical organ (B, D2) lies as a small,pinhead-sized nodule between the two in-terventricular foramina in the roof of thethird ventricle at the rostral end of the telachoroidea. In addition to glial cells and iso-lated neurons, it contains large roundparenchymal cells, the neuronal characterof which is disputed.
Electron microscopy has demonstratedependymal canaliculi penetrating from thesurface and interconnecting with wide in-tercellular clefts. Vascular loops penetratefrom the tela choroidea into the inside of thesubfornical organ. Peptidergic nerve fibers(somatostatin, luliberin) terminate at capil-laries and in the area of the ependymalcanaliculi.
Area postrema (C, D)
The area postrema (C, D3) consists of twosymmetrical narrow structures at the floorof the rhomboid fossa; they lie at the fun-nel-shaped entrance of the central canal.The loose tissue of this region containsnumerous small cavities. It consists of gliaand parenchymal cells, which are generallyregarded as neuronal cells. The tissue con-tains numerous convoluted capillaries thatappear highly fenestrated in the electron-microscopic image. The area postrema istherefore one of the few brain regionswhere the blood–brain barrier is permeable(p. 44, C–E).
Paraphysis and Subcommissural Organ(D)
Both structures appear in humans only tem-porarily during embryonic development(transitory structures). The paraphysis is asmall saclike evagination of the roof of thethird ventricle caudally to the interventricu-lar foramina. The subcommissural organ (D4)consists of a complex of cylindrical ependy-mal cells underneath the epithalamic com-missure. These cells produce a secretionthat does not dissolve in the CSF but con-denses to form a long, thin filament,Reissner’s fiber. In animals inwhich the cen-tral canal is not obliterated, Reissner’s fiberextends into the lower spinal cord.
D5 Choroid plexus.D6 Interventricular foramen
(foramen of Monro).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

287
Cerebr
ovas
cularan
dVen
tricular
System
s
26
4
5
1
3
Circumventricular Organs
A Vascular organ of the terminal lamina,horizontal view (according to Kuhlenbeck)
B Subfornical organ
C Area postrema
D Locations of the organs
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

288Ce
rebr
ovas
cularan
dVen
tricular
System
sCerebrovascular and Ventricular Systems
Meninges
The brain is surrounded by mesodermalcoverings, the meninges. The outer layer isthe tough pachymeninx, or dura mater (A1).The inner layer is the soft leptomeninxwhich consists of two sheets, the arachnoidmater (A2) and the pia mater (A3).
Dura Mater (B)
The duramater lines the inner surface of theskull and also forms the periosteum. Sturdysepta extend from it deep into the cranialcavity. A sickle-shaped fold of the dura, thefalx of the cerebrum (B4), suspends verti-cally between the two cerebral hemi-spheres. It is attached rostrally to the cristagalli and extends over the frontal crest tothe internal occipital protuberance, where itturns into the tentorium of the cerebellum(B5) spanning both sides. The falx dividesthe superior part of the cranial cavity insuch a way that each hemisphere is sup-ported in its own space. The tentorium ofthe cerebellum stretches like a tent acrossthe cerebellum, lying in the posteriorcranial fossa. It is attached along the trans-verse sulcus of the occipital bone and theupper margin of the petrous bone, leavingrostrally a wide opening for the passage ofthe brain stem (B6). At the lower surface ofthe tentorium and along the occipital crest,the falx of the cerebellum projects into theposterior cranial fossa.
The large venous channels, the sinuses of thedura mater, are embedded between the twosheets of the dura mater (see vol. 2). The di-agram shows cross sections of the superiorsagittal sinus (B8) and the transverse sinus(B9).
Certain structures are encapsulated by duralpockets and thus separated from the rest ofthe inner cavity. The sellar diaphragm (B10)spans the sella turcica and contains an aper-ture for passage of the hypophysial stalk, thediaphragmatic hiatus (B11). The trigeminalganglion (semilunar ganglion, Gasser’s gan-glion) on the anterior surface of the petrousbone is enclosed by a dural pocket, thetrigeminal cave (Meckel’s space).
Arachnoid Mater (A)
The arachnoidmater (A2) adjoins closely theinner surface of the dura mater and is sepa-rated from it only by a capillary cleft, thesubdural space (A12). It encloses the sub-arachnoid space (A13), which containscerebrospinal fluid and is connected withthe pia mater by trabeculae (A14) andseptae that form a dense meshwork andcreate a system of communicating cham-bers.
Peduncledmushroomlike vegetations of thearachnoid protrude into the large sinuses,the arachnoid granulations (meningealgranules, or pacchionian bodies) (A15). Theyconsist of an arachnoid meshwork and arecovered by mesothelium. The dura mater,which encloses them, is reduced to a mem-brane. These arachnoid villi are most abun-dant in the vicinity of the superior sagittalsinus (A16) and at the lateral lacunae (A17),and less frequent at the exits of spinalnerves. The CSF is absorbed into the venousblood in the area of the granulations. Inolder people, the granulations may alsopenetrate into the bone (formation ofgranular foveolae) (A18) and invaginate intothe diploic veins.
Pia Mater (A)
The pia mater (A3) is the meningeal cover-ing that contains the blood vessels. It bordersdirectly on the brain and forms the me-sodermal side of the pia–glia barrier. Fromhere vessels enter the brain, and they aresurrounded by the pia mater for some dis-tance (pial funnel).
A19 Scalp.A20 Skull.A21Diploë (see vol. 1).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

289
Cerebr
ovas
cularan
dVen
tricular
System
s
19
20
12
3
1413 12
4
11
10 6 5
8
9
18
17 15 16 21
Meninges
A Meninges and subarachnoid space
B Pachymeninx (dura mater)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Autonomic NervousSystem
Overview 292Sympathetic Trunk 296Autonomic Periphery 300
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

292Aut
onom
icNervo
usSy
stem
Autonomic Nervous System
Overview
The vegetative or autonomic nervous sys-tem supplies the internal organs and theircoverings. Almost all tissues of the body arepermeated by a plexus of very delicatenerve fibers. We distinguish between affer-ent (viscerosensory) and efferent (visceromo-tor and secretory) fibers. The neurons withsensory fibers lie in the spinal ganglia. Theneurons giving rise to efferent fibers formcell clusters scattered throughout the body;these clusters are surrounded by a connec-tive-tissue sheath and are known as auto-nomic ganglia.
The main function of the autonomicnervous system is to stabilize the internalenvironment of the organism and to regulatethe function of the organs in accordancewiththe changing requirements of the surround-ings. This regulation is achieved by interac-tion of two antagonistic parts of the auto-nomic system, the sympathetic nervous system(A1) (yellow) and the parasympatheticnervous system (A2) (green). The sympatheticnervous system is stimulated by increasedphysical activity, resulting in elevated bloodpressure, accelerated heart rate and respira-tory rate, dilated pupils, raised hair, and in-creased perspiration. At the same time, theperistaltic activity of the gastrointestinaltract is suppressed and secretion by intesti-nal glands is reduced. When the parasym-pathetic system predominates, it increasesperistaltic activity and intestinal secretion,stimulates defecation and urination, and re-duces the heart rate and respiratory rate,while the pupils constrict. The sympatheticnervous system is responsible for increasedperformance under stress and in states ofemergency, while the parasympatheticnervous system promotes metabolism, re-generation, and the buildup of body re-serves.
The division of the autonomic nervous sys-tem into a sympathetic part and a parasym-pathetic part refers to visceromotor andsecretory fibers. Such a distinction is notpossible in case of viscerosensory fibers.
Central Autonomic System
We distinguish between a peripheral and acentral autonomic nervous system. The cen-tral cell groups of the sympathetic and para-sympathetic nervous systems lie in differ-ent regions. Parasympathetic neurons formnuclei in the brain stem (see p. 106):
! The Edinger-Westphal nucleus (A3)! The salivatory nuclei (A4)! The dorsal nucleus of the vagus nerve (A5)
The sacral spinal cord also contains para-sympathetic neurons (A6). The sympatheticneurons occupy the lateral horn in thethoracic and upper lumbar segments of thespinal cord (A7) (p. 297, A1). Localization ofthe parasympathetic nuclei is thereforecraniosacral, while that of the sympatheticnuclei is thoracolumbar.
The highest integration organ of the auto-nomic nervous system is the hypothalamus. Italso regulates the endocrine glands throughits connection to the hypophysis, and itcoordinates the autonomic nervous systemand the endocrine system. Cell groups in thereticular formation of the brain stem alsoparticipate in the central regulation of organfunctions (heart rate, respiratory rate, bloodpressure, p. 146).
A8 Sympathetic trunk.A9 Superior cervical ganglion.A10 Stellate ganglion
(cervicothoracic ganglion).A11 Celiac ganglion.A12 Superior mesenteric ganglion.A13 Inferior mesenteric ganglion.A14Hypogastric plexus.A15Greater splanchnic nerve.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

293
Aut
onom
icNervo
usSy
stem
3
4
5
29
8
10
11
12
13
14
6
15
7
1
Sympathetic and Parasympathetic Nervous Systems
A Sympathetic and parasympathetic nervous systems(adapted from Villiger and Ludwig)
Eyes
Lacrimal glands,salivary glands
Blood vesselsof the head
Heart
Lungs
Stomach
Liver
Pancreas
Kidneys
Intestine
Rectum
Urinary bladder
Genitals
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

294Aut
onom
icNervo
usSy
stem
Autonomic Nervous System: Overview
Peripheral Autonomic System (A)
Parasympathetic Nervous System
The fibers of the central parasympatheticneurons runwithin various cranial nerves tothe parasympathetic ganglia in the head re-gion (pp. 128, 130) where they synapse; thepostganglionic fibers extend to the effectororgans. The vagus nerve (A1), which is theprincipal nerve of the parasympatheticnervous system, descends together with thelarge cerebral vessels (neurovascular trunkof the neck); after passing through the supe-rior thoracic aperture, it divides into plex-uses in the regions of the thoracic andabdominal viscera (p. 116).
The cells lying in the intermediolateral nu-cleus and intermediomedial nucleus of thesacral spinal cord send their axons throughthe third and fourth sacral root (A2) to thepudendal nerve; from here the fibers pass aspelvic nerves into the inferior hypogastricplexus and to the pelvic organs (urinarybladder [A3], rectum, and genitals). Syn-apses with postganglionic neurons areformed in the inferior hypogastric plexus orin small ganglia of the various organ plex-uses. As is the case with the sympatheticnervous system, the peripheral supply isprovided by two neurons: the first neuron(preganglionic neuron) in the spinal cord,and the second neuron (postganglionic neu-ron) in the ganglia (p. 297, B, C).
Sympathetic Nervous System
The sympathetic neurons in the thoracicand lumbar lateral horn send their axons viathe communicating branches (A4) to thesympathetic trunk (A5) (p. 296, D). The lat-ter consists of a chain of sympathetic gan-glia which lie on each side of the vertebralcolumn, in front of the transverse processesof each vertebra, and extend from the baseof the skull to the coccyx. They are intercon-nected by interganglionic branches (A6).There are three ganglia in the cervical seg-ment, namely, the superior cervical ganglion,the variable middle cervical ganglion (A7),and the stellate ganglion (cervicothoracicganglion) (A8). The thoracic segment contains
10 – 11 ganglia, the lumbar segment usuallyfour, and the sacral segment also four ganglia.The chain is completed by the small un-paired ganglion (A9) which lies in themiddle in front of the coccyx. The sacralganglia receive their preganglionic fibers viainterganglionic branches from spinal cordlevels T12 – L2.
From the thoracic and lumbar sympathetictrunk ganglia, nerves extend to ganglia thatlie within dense nervous plexuses on bothsides of the abdominal aorta. The uppergroup of ganglia are the celiac ganglia (A10)to which the greater splanchnic nerve (A11)extends from the fifth to the ninth sympa-thetic trunk ganglia. Below it lies the supe-rior mesenteric ganglion (A12) and the infe-rior mesenteric ganglion (A13). The superiorhypogastric plexus (A14) and the inferior hy-pogastric plexus (A15) expand in the pelvis.
Adrenergic and CholinergicSystems
The transmission of impulses is mediated inthe sympathetic nervous system bynorepinephrine, and in the parasympa-thetic nervous system by acetylcholine. Thesympathetic nervous system is therefore alsoknown as the adrenergic system and the para-sympathetic nervous system as the choliner-gic system. All preganglionic fibers of thesympathetic nervous system are choliner-gic, and only the postganglionic fibers arenoradrenergic (p. 297, C). The postgan-glionic sympathetic fibers innervating thesweat glands of the skin are also cholinergic.
The antagonism between sympathetic andparasympathetic nervous systems is clearlyapparent for some organs (heart, lungs).Other organs are regulated by the increasedor decreased tone of just one system. Thus,the adrenal glands and the uterus are sup-plied only by sympathetic fibers (theadrenal gland as paraganglion is supplied bypreganglionic fibers, see vol. 2). The func-tion of the urinary bladder is regulated byparasympathetic fibers; the role of sympa-thetic fibers in this control is disputed.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

295
Aut
onom
icNervo
usSy
stem
8
1
4
6
11
10
5
15
2
9
7
12
13
14
3
Sympathetic and Parasympathetic Nervous Systems
A Autonomic nervoussystem (accordingto Hirschfeldand Léveillé)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

296Aut
onom
icNervo
usSy
stem
Autonomic Nervous System: Overview/Sympathetic Trunk
Neuronal Circuit (A–C)
The sympathetic neurons in the interme-diomedial nucleus and in the intermediolateralnucleus (lateral horn) (A1) of the thoracicspinal cord send their axons through theanterior root (A2) into the spinal nerve.They run through the white communicatingbranch (A3) to reach the sympathetic trunkganglion (A4) as preganglionic fibers. Here,some of them terminate at neurons fromwhere postganglionic fibers return into thespinal nerve via the gray communicatingbranch (A5). The preganglionic fibers en-tering the ganglion are myelinated so thatthe connecting branch appears white(white communicating branch). The post-ganglionic fibers exiting the ganglion areunmyelinated so that the connecting branchappears gray (gray communicating branch).
Other postganglionic fibers (A6) extendfrom the sympathetic trunk via autonomicnerves to the organs. Some preganglionicfibers (A7) pass through the ganglionwithout synapsing and terminate in the pre-vertebral ganglia (A8) which lie on both sidesof the aorta. Numerous small and very smallterminal ganglia (A9) lie in the regions of in-ternal organs. They are part of the neu-roplexuses spreading through each organ;they are found in the sheaths (extramuralganglia) as well as inside the organs (in-tramural ganglia). Whereas preganglionicand postganglionic fibers of the parasympa-thetic nervous system are both cholinergic(B), the cholinergic preganglionic fibers ofthe sympathetic nervous system synapse inthe ganglia with noradrenergic neurons (C).
According to their localization, we distin-guish three different types of ganglia inwhich preganglionic fibers synapse withpostganglionic neurons:
! The sympathetic trunk ganglia! The prevertebral ganglia! The terminal ganglia
The sympathetic trunk ganglia and the pre-vertebral ganglia are sympathetic ganglia,whereas the terminal ganglia are mostly,but not exclusively, parasympathetic gan-glia.
Sympathetic Trunk
Cervical and Upper ThoracicSegments (D)
The cervical ganglia are reduced to three;the uppermost, the superior cervical ganglion(D10), lies below the base of the skull nearthe nodose ganglion (inferior ganglion ofthe vagus nerve). It receives fibers from theupper thoracic segment via the intergan-glionic branches. Its postganglionic fibersform plexuses around the internal carotidartery and external carotid artery. Branchesextend from the internal carotid plexus tothe meninges, to the eyes, and to the glandsof the head region. The superior tarsalmuscle of the upper eyelid and the ophthal-mic muscles at the posterior wall of theorbit are innervated by sympathetic fibers.Injury to the superior cervical gangliontherefore leads to drooping of the uppereyelid (ptosis) and to a backward displace-ment of the eyeball (enophthalmos).
The middle cervical ganglion (D11) may beabsent, and the inferior cervical ganglion hasin most cases fused with the first thoracicganglion to form the stellate ganglion (D12).Its postganglionic fibers form plexusesaround the subclavian artery and aroundthe vertebral artery. Fiber bundles connect-ing the stellate ganglion with the middlecervical ganglion extend across the sub-clavian artery and form the subclavian ansa(D13). Nerves from the cervical ganglia(D14) and nerves from the upper thoracicganglia (D15) extend to the heart and to thehila of the lungs, where they participate to-gether with the parasympathetic fibers ofthe vagus nerve in the formation of the car-diac plexus (D16) and the pulmonary plexus(D17). The branches of the fifth to ninthsympathetic trunk ganglia join to form thegreater splanchnic nerve (D18) which ex-tends to the celiac ganglia.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

297
Aut
onom
icNervo
usSy
stem
2
1
4
8
11
10
14
12
17
16
13
15
76
9
5
3
18
Neuronal Circuit/Sympathetic Trunk: Cervical and Upper Thoracic Segments
A Neuronal circuit
B Cholinergic system(parasympatheticnervous system)
C Synaptic switch from acholinergic to a norad-renergic neuron (sympa-thetic nervous system)
D Sympathetic trunk, cervical andupper thoracic segments
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

298Aut
onom
icNervo
usSy
stem
Autonomic Nervous System: Sympathetic Trunk
Lower Thoracic and AbdominalSegments (A)
Branches originating from the thoracic andupper lumbar sympathetic trunk gangliaextend to the prevertebral ganglia of theabdominal aortic plexus. There are severalgroups of ganglia. At the exit of the celiactrunk lie the celiac ganglia (A1) where thegreater splanchnic nerve (A2) (T5–T9) andthe lesser splanchnic nerve (A3) (T9–T11)terminate. Their postganglionic fibers ex-tend with the branches of the aorta to stom-ach, duodenum, liver, pancreas, spleen, andadrenal gland (gastric plexuses, hepaticplexus, splenic plexus, pancreatic plexus,suprarenal plexus). Preganglionic fibers runto the adrenal medulla (see vol. 2).
The postganglionic fibers from the superiormesenteric ganglion (A4), together with thebranches of the celiac ganglion, supply thesmall intestine, the ascending colon, and thetransverse colon. The fibers from the inferiormesenteric ganglion (A5) supply the de-scending colon, the sigmoid colon, and therectum. The preganglionic fibers (lumbarsplanchnic nerves) of both ganglia originatefrom levels T11–L2. Some branches extendto the renal plexus, which also containsfibers from the celiac ganglia and the supe-rior hypogastric plexus.
Parasympathetic fibers also participate inthe formation of the visceral plexuses.Stimulation of parasympathetic fibers in thedigestive tract leads to increased peristalsisand secretion as well as relaxation of thesphincter muscles, while stimulation ofsympathetic fibers causes reduced peristal-sic and secretion as well as contraction ofthe sphincter muscles.
The pelvic organs are supplied by the supe-rior hypogastric plexus (A6) and the inferiorhypogastric plexus. Both plexuses receivepreganglionic sympathetic fibers from thelower thoracic and upper lumbar spinalcord, and parasympathetic fibers from thesacral spinal cord.
The urinary bladder is predominantly inner-vated by the parasympathetic fibers of thevisceral plexus that supply the muscles for
bladder contraction (detrusor muscle). Thesympathetic fibers terminate at the smoothmuscles of the orifice of the urethra andboth ureteral orifices. Regulation of bladdertone and urination takes place via spinal re-flexes which, in turn, are controlled by thehypothalamus and by cortical areas.
The genitals are supplied by the prostaticplexus in the male and by the uterovaginalplexus in the female. Stimulation of para-sympathetic fibers dilates the vessels of thecavernous bodies and thus triggers erectionin the male (nervi erigentes). Stimulation ofsympathetic fibers leads to vasoconstrictionand to ejaculation. The uterine muscles areinnervated by sympathetic and parasympa-thetic fibers. Their functional significance isnot clear because even a denervated uterusis fully functional during pregnancy andparturition.
Innervation of the Skin (B)
The sympathetic fibers returning from thesympathetic trunk ganglia into the spinalnerves (p. 297, A5) run in the peripheralnerves to the skin, where they innervatevessels, sweat glands, and erector pilimuscles (vasomotor, sudomotor, pilomotorfunctions) in the corresponding derma-tomes (p. 66). Segmental loss of these func-tions is of diagnostic significance in injuriesto the spinal cord.
Clinical Note: In specific skin regions, Head’szones (zones of hyperalgesia) (B), disease of anorgan may give rise to pain or hypersensitivity,with each organ being represented in a specificzone: diaphragm (B7) (C4), heart (B8) (T3/4),esophagus (B9) (T4/5), stomach (B10) (T8), liverand gallbladder (B11) (T8 – 11), small intestine(B12) (T10), large intestine (B13) (T11), urinarybladder (B14) (T11–L1), kidneys and testes (B15)(T10–L1). The Head’s zones are of practical im-portance in the diagnosis.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

299
Aut
onom
icNervo
usSy
stem
2
1
3
1
1
4
5
6
9 8
10
11
12
13
1415
7
Lower Thoracic and Abdominal Segments, Head’s Zones
A Sympathetic trunk, lower thoracicand abdominal segments
B Head’s zones
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

300Aut
onom
icNervo
usSy
stem
Autonomic Nervous System
Autonomic Periphery
Efferent Fibers
The preganglionic fibers are myelinated,while the postganglionic fibers are unmyeli-nated. The unmyelinated fibers are sur-rounded by the cytoplasm of Schwann cells,and a single Schwann cell envelopes severalaxons (p. 38, A8).
Afferent Fibers
The viscerosensory fibers are myelinated.They are regarded as being neither sympa-thetic nor parasympathetic in nature. Ingeneral, they accompany the sympatheticnerves and enter the spinal cord throughthe posterior roots. The fibers from theheart run through the upper thoracic roots,those from the stomach, liver, and gallblad-der run through the middle thoracic roots,and those from the colon and appendix runthrough the lower thoracic roots. The re-spective dermatomes of these roots corre-spond roughly to the various Head’s zones.
Intramural Plexus (A–E)
The autonomic nerves enter the internal or-gans together with the vessels and form afine network of noradrenergic (A) or cholin-ergic (B) fibers (enteric plexus). The fibersterminate on smooth-muscle cells and onglands. Vascular muscles influence thefunction of many organs (regulation ofblood flow by contraction or dilatation ofvessels). It is not known whether parenchy-mal organs such as liver or kidneys containsecretory fibers.
The digestive tract is supplied by two differ-ent plexuses, namely, the submucous plexus(Meissner’s plexus) and the myenteric plexus(Auerbach’s plexus). The submucous plexus(C1, D) forms a three-dimensional networkthroughout the entire submucosa. It is an ir-regular meshwork of bundles containingmedium-sized to very fine nerve fibers thatbecome finer and narrower toward the mu-cosa. Aggregations of neurons form smallintramural ganglia at the intersection offibers.
The intramural ganglia (E) contain primarilymultipolar, rarely unipolar, neurons withgranular Nissl substance. The neurons aresurrounded by flat sheath cells, and theirnumerous long dendrites are thin and oftenindistinguishable from axons. The axons areextremely thin, unmyelinated or poorly my-elinated, and often arise from a dendriterather than the perikaryon. Between theneurons lies a dense network of fibers inwhich it is difficult to differentiate betweendendrites, terminating axons, and fiberspassing through the ganglion. Sympatheticand parasympathetic neurons have thesame shape; they can only be distinguishedhistochemically.
The myenteric plexus (C2) is embedded ina narrow space between the transverse andlongitudinal muscles of the intestine. It con-sists of a relatively regular network ofcoarse and fine fiber bundles. Apart fromthe intramural ganglia at the intersections,there are numerous neurons that are oftenarranged in rows along the fiber bundles.
The enormous number of neurons scatteredthroughout the tissue almost amounts to anindependent nervous organ. This explainsthe local autonomy of the gastrointestinaltract, which remains functional even afterdenervation.
As derivatives of the neural crest (p. 62, C2),the paraganglia and the adrenal medulla areregarded as part of the autonomic nervoussystem (see vol. 2).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

301
Aut
onom
icNervo
usSy
stem
1
2
Intramural Plexus
A Sympathetic fibers, fluorescencemicroscopy
C Intestinal wall, diagram
D Submucous plexus E Intramural ganglion
A, B Vas deferens in the guinea pig (after J. Winkler)
B Parasympathetic fibers, acetylcholineesterase histochemistry
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

302Aut
onom
icNervo
usSy
stem
Autonomic Nervous System: Autonomic Periphery
Autonomic Neurons
Structure (A, B, D)
The autonomic nervous system consists ofnumerous individual elements, the auto-nomic neurons (A) (neuron doctrine). Thetheory of neuronal continuity, which hadlong been postulated for the terminal rami-fications of the autonomic nervous system,has been disproved by electron-microscopicstudies. According to the continuity theory,the terminal ramifications of an intramuralplexus were thought to form a continuousnetwork (terminal reticulum), in which theprocesses of different neurons would fusewith each other and with the innervatedmuscle cells and glandular cells. The net-work was thought to represent a syncytiumwhere nerve fibers share a common cyto-plasm. The electron-microscopic imagedoes not show such continuity.
However, the postganglionic neurons dohave specific features. The bundles of nervefibers exhibit numerous axo-axonal syn-apses (D1), not only among sympathetic andparasympathetic fibers themselves but alsobetween sympathetic and parasympatheticfibers. In the area of terminal ramification,specific structures corresponding to themotor end plates (p. 313, B and C) at striatedmuscles are absent. The only remarkablefeatures are varicose swellings (varicosities)(A–D2) along the terminal branches of theaxons.
The axon swellings may lead to indenta-tions on the smooth-muscle cells or mayeven invaginate the cells. In general,however, they lie between the muscle cellswithout direct membrane contact (as theywould in a synapse, see p. 27, D). The swell-ings contain clear and granular vesicles (C3)similar to those in presynaptic terminalboutons (p. 27, C). The granular vesicles havebeen shown to contain norepinephrine, theneurotransmitter of the sympatheticnervous system. The sheath of Schwann cells(B4) surrounding the terminal branches isabsent in the area of the swelling, and theadjacent wall segment of the smooth-muscle cell (BC5) lacks a basement mem-
brane (B6). This is the site of signal trans-mission from the autonomic nerve fiber tothe smooth-muscle cell.
Signal Transmission (C)
The vesicles contained in the axon swellingsrelease their content into the intercellularspace (C7). The neurotransmitter moleculesdiffuse into the intercellular space, therebytransmitting the signal to a large number ofsmooth-muscle cells. Signal transmissionalso spreads via membrane contacts be-tween the muscle cells. The smooth-musclecells are interconnected by gap junctions(B8) which function as electrical synapses(see p. 26). There is no basement membraneat gap junctions.
Efferent autonomic nerve fibers innervatethe smooth muscles and glandular cells(secretory fibers). Innervation of glandularcells (blue cells in C) is essentially the sameas that of muscle cells. The neurotransmit-ters released from the vesicles in the axonswellings (green dots in C) activate G pro-tein-coupled receptors (C9) on the surfaceof the innervated cell. G protein mediatesthe opening of ion channels (C10), thus trig-gering an intracellular cascade of signals.This results in the contraction of smooth-muscle cells or in the synthesis and releaseof glandular secretion by the glandular cells.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

303
Aut
onom
icNervo
usSy
stem
6
6
2
5
5
5
9
6
4
28
8
2
10
2
7
3
2
2
2
1
9
Autonomic Neurons
A Autonomic neuron B Autonomic nerve fiber, electron-microscopic diagram
C Signal transmission
D Innervation of smooth-muscle cells
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Functional Systems
Brain Function 306Motor Systems 308Sensory Systems 318Limbic System 332
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

306Fu
nction
alSy
stem
sFunctional Systems
Brain Function
The central nervous system enables the or-ganism to adapt to the environment and tosurvive. It receives stimuli from outside andinside the body through the sensory organs;it then filters the stimuli and processesthem into information. In accordance withthis information, it sends impulses to theperiphery of the body so that the organismcan react in a meaningful way to the con-stantly changing conditions (p. 2, EF). Thefunctional systems described in this chapterare by no means independent, isolated sys-tems. The highly simplified and schematicdescription is supposed to provide an ap-proximate illustration of the extremelycomplex interactions among billions ofnerve cells.
For centuries there has been the simplemechanistic idea that sensations reach thebrain and the brain then triggers motor re-actions. According to Descartes, optic stimuliare transmitted from the eyes to the pinealgland (epiphysis), which then sends im-pulses that travel to the muscles (A). Heviewed the pineal gland as the seat of thesoul. Franz Gall was the first to postulate theimportance of the cerebral convolutionsand the cerebral cortex for brain function.He localized the “organs of the soul” on thesurface of the hemispheres, and he believedthat mental faculties could be determinedto various degrees by the topographical fea-tures on the surface of the skull (phrenology)(B). He used Roman numerals for localizingthese features: I, reproductive instinct; II,love of offspring; III, friendship; IV, bravery;V, instinct to eat meat; VI, intelligence; VII,greed and cleptomania; VIII, pride and arro-gance; IX, vanity and ambition; and so on.
On the basis of brain injuries, Kleist arrivedat localizations of higher cerebral functions(C). He assumed that positive capacities, or“functions,” would correspond to deficits inrecognition and thinking, motivation andaction, among others, which were ex-pressed as negative pathological findings.However, this is not so. While it is possible
to localize symptoms of deficit, this cannotbe done for capacities (von Monakow). Crit-ics of this theory of localization and centersspoke of “brain mythology.” Ultimately,specific capacities cannot be attributed tospecific brain regions because immensenumbers of other neuronal groups are al-ways participating in the stimulation, inhi-bition, or modulation of a capacity. The so-called “centers” can be regarded, at best, asimportant relay stations for a particularcapacity. Nor is the central nervous system arigid apparatus; rather it exhibits a con-siderable degree of plasticity. Especially inthe infant brain, other centers can take overand perform functions in place of injuredparts of the brain. Plasticity of our principalorgan is also a prerequisite for the capacityto learn (language, writing, physical skills).
Information processing in the telen-cephalon is known as integration. It refersto the combination and interconnection ofsensations, including stored experience, toform a higher and complex functional unit.In this way, the functions of the organismare guided by means of the meticulousmutual coordination between groups ofneurons. During human evolution, the inte-grative processes of regulating and coordi-nating elementary biological tasks havedeveloped into conscious recognition,thought, and action. Cybernetics and com-puter technology have provided us withmodels for the function of the brain (cogni-tive science). According to these models,various “functions” are based on the con-tinuously changing stimuli within intercon-nected neuronal circuits.
Despite our highly developed instrumenttechnology, there is ultimately only one in-strument available for brain research: thebrain itself. In other words, a human organis involved in an attempt to investigate itsown structure and function.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

307
Func
tion
alSy
stem
s
Brain Function
A Transmission of visual stimulifrom the eyes to the pineal gland(according to René Descartes1662)
B Localization of centers on theskull (according to Gall 1810)
C Localization of cerebral functions(according to Kleist 1934)
Turning move-ment oftrunk
Motivation
Course ofaction
Activethou
ght
Mentality
Percep
tion
of stren
gthAction
Course ofaction
Memoryof place
Arithmetic
Reading
Senseof
direction
Color re-cognition
Acoustic attention
XXIIXXIII
XXVII
X
VIIIIX
III
II
I
V
VIVII
IV
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

308Fu
nction
alSy
stem
sFunctional Systems
Motor Systems
Corticospinal Tract (A, B)
The corticospinal tract (pyramidal tract)and the corticonuclear fibers (p. 58, A; p. 140,A) are regarded as pathways of voluntarymovements. It is through them that the cor-tex controls the subcortical motor centers.The cortex can have a reducing and inhib-iting effect, but it also produces a continu-ous tonic stimulus that promotes quick,sudden movements. The mechanical andstereotyped motions controlled by the sub-cortical motor centers must be modified bythe influence of pyramidal impulses inorder to produce specific, fine-tuned move-ments.
The fibers of the corticospinal tract origi-nate in the precentral areas 4 (A1) and 6(A2), in regions of the parietal lobe (areas 3,1, and 2), and in the second sensorimotorarea (area 40) (see pp. 246, 248, 250).Roughly two-thirds stem from the precen-tral area, and one-third from the parietallobe. Only about 60% of the fibers are myeli-nated; the other 40% are unmyelinated. Thethick fibers of Betz’s giant pyramidal cells(p. 246, A) in area 4 account for only 2 – 3%of the myelinated fibers. All other fibersstem from smaller pyramidal cells.
The fibers of the pyramidal tract passthrough the internal capsule (p. 258, A2, B).At the transition to the midbrain, they ap-proach the base of the brain and togetherwith the corticopontine tracts form thecerebral peduncles. The corticospinal tractfibers occupy the central part, and the fibersfrom the parietal cortex occupy the mostlateral position (B3). They are followed bythe corticospinal tracts for the lower limb (L,S), trunk (T), and upper limb (C), and finallythe corticonuclear fibers for the facial re-gion (B4). When passing through the pons,the fiber tracts rotate so that the corticonu-clear fibers now lie dorsally, followed by thebundles terminating in the cervical,thoracic, lumbar, and sacral regions, respec-tively. In the medulla oblongata, the corti-conuclear fibers terminate on the cranialnerve nuclei (p. 140, A). Most of the fibers
(70 – 90%) cross to the opposite side in thepyramidal decussation (AB5) (p. 59, A1) andform the lateral corticospinal tract (AB6). Thefibers for the upper limb cross dorsally tothe fibers for the lower limb. Within thelateral corticospinal tract, the fibers for theupper limb lie medially, while the longfibers for the lower limb lie laterally(p. 58A). The uncrossed fibers continue inthe anterior corticospinal tract (AB7) and crossto the opposite side only at the level of theirtermination above the white commissure(p. 50, A14). The anterior tract shows varia-ble degrees of development; it may beasymmetric or even completely absent. Itreaches only into the cervical or thoracicspinal cord.
The majority of corticospinal tract fibersterminate on interneurons in the interme-diate zone between anterior and posteriorhorns (p. 52, A6). Only a small portionreaches the motor neurons of the anteriorhorn, predominantly those supplying thedistal segments of the limbs, which areunder the special control of the corticospi-nal tract. Impulses from the corticospinaltract activate neurons that innervate theflexor muscles but inhibit neurons that in-nervate the extensor muscles.
Fibers originating from the parietal lobe ter-minate in the dorsal column nuclei (gracilenucleus and cuneate nucleus) and in the sub-stantia gelatinosa of the posterior horn.They regulate the input of sensory impulses.Hence, the corticospinal tract is not a uni-form motor pathway but contains de-scending systems of different functions.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

309
Func
tion
alSy
stem
s
2 1
3
4
5
6
7
6
6
7
6
6
5
Corticospinal Tract
A Corticospinal tract (pyramidal tract)
B Position of the corticospinal tractin brain stem and spinal cord
CT
L, S
CT L
S
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

310Fu
nction
alSy
stem
sFunctional Systems: Motor Systems
Extrapyramidal Motor System (A)
Apart from the precentral area and corti-cospinal (pyramidal) tract, numerous othercortical areas and pathways control motoractivity. They are collectively known as theextrapyramidal motor system. It is phylo-genetically older than the corticospinaltract and, unlike it, consists of chains ofmultisynaptic neurons. Originally, the termreferred to a group of nuclei characterizedby its high iron content: neostriatum (puta-men [A1] and caudate nucleus [A2]), pal-lidum (A3), subthalamic nucleus (A4), rednucleus (A5), and substantia nigra (A6) (ex-trapyramidal motor system in the narrowersense). This group of nuclei is connected toother centers that are important for motoractivity; however, these are integration cen-ters rather than motor nuclei: cerebellum(A7), thalamic nuclei (A8), reticular forma-tion, vestibular nuclei (A9), and some corti-cal areas. They are collectively known as theextrapyramidal system in the broader sense.
Function
When one limb is moved voluntarily,muscle groups of the other limbs and of thetrunk are simultaneously activated so thatbalance and posture are maintained underthe changed static conditions and themove-ment can be completed smoothly. The ac-companying muscle activities, which areoften nothing more than the increased ten-sion or relaxation of certain muscle groups,are carried out involuntarily and are notexperienced consciously. Without them,however, coordinated movement would beimpossible. Such unconscious motions in-clude associated movements (synkinesis)(such as arm movements while walking) aswell as many movements that have beenpracticed for a long time and thus occur au-tomatically. They are all under the control ofthe extrapyramidal system; this may becompared to a servomechanism that sup-ports all voluntary movements in an auto-nomic way and without reaching the levelof consciousness.
Pathways
Afferent pathways reach the system throughthe cerebellum. Cerebellar tracts terminatein the red nucleus (cerebellorubral tract)(A10) and in the centromedian nucleus ofthalamus (A11) from which fibers extend tothe striatum. Cortical fibers run to the stri-atum (A12), to the red nucleus (A13), and tothe substantia nigra (A14). Vestibular fibersterminate in the interstitial nucleus of Cajal(A15).
The efferent pathway of the system is the cen-tral tegmental tract (A16) (p. 144, A). Otherdescending pathways are:
! The reticulospinal tract (A17)! The rubrospinal tract! The vestibulospinal tract (A18)! The interstitiospinal tract (A19)
The extrapyramidal centers are intercon-nected by numerous neuronal circuits to en-sure mutual control and adjustment. Thereare two-way connections between pallidumand subthalamic nucleus, and between stri-atum and substantia nigra (A20). A largeneuronal circuit runs from the cerebellumthrough the centromedian nucleus ofthalamus to the striatum and from there viapallidum, red nucleus, and olive (A21) backto the cerebellum. Other functional circuitsare formed by cortical fibers to the striatum,with a recurrent circuit passing throughpallidum, ventral anterior nucleus and ven-tral lateral nucleus of thalamus, and back tothe cortex.
The frontal and occipital visual fields(p. 248, C; p. 254) together with the regionsof parietal and temporal lobes, from wherecomplex massive movements can be elic-ited by a strong electrical current, areknown as extrapyramidal cortical fields.However, the inclusion of cortical fields inthe extrapyramidal system is controversial,even though numerous corticostriatal con-nections have been demonstrated (A12).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

311
Func
tion
alSy
stem
s
1213 14
11 8
3
2
1
4
20
155
20
6
10
7
21
18
9
19
16
17
Extrapyramidal Motor System
A Extrapyramidalmotor system
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

312Fu
nction
alSy
stem
sFunctional Systems: Motor Systems
Motor End Plate (A–C)
The axons of the motor neurons (A1) ar-borize in the muscles so that each musclefiber (AB2) is reached by an axonal branch(A3). The number of muscle fibers suppliedby one axon varies considerably. While asingle axon may innervate two to threemuscle fibers in themuscles of eyes and fin-gers, it may supply 50 – 60 muscle fibers inother muscles. The anterior horn cell and itsaxon (!-motoneuron) together with thegroup of muscle fibers it supplies is called amotor unit. When the neuron is stimulated,the muscle fibers contract in unison. Theterminal branches of the axon lose their my-elin sheaths before terminating and formtangled ramifications. In the terminal re-gion, the surface of the muscle fiber forms aflat eminence (hence the term end plate)(A4).
The area of axonal arborization (A5) con-tains a number of cell nuclei. The nucleilying on top of the axonal ramificationsbelong to Schwann cells that envelop theaxon terminals (teloglia) (B6). The nucleilying beneath the ramifications (B7) aremuscle fiber nuclei in the region of the endplate. At the junction between axoplasmand sarcoplasm, the axon terminals are sur-rounded by a palisade layer (B8) which con-sists of infoldings of the sarcolemma, asshown by electron microscopy.
The axons terminate with boutonlike swell-ings (B9) that dip into the surface of the endplate. These grooves are lined by the mem-brane of the sarcoplasm (sarcolemma) and abasement membrane. The heavily foldedsarcolemma of the grooves (subneural clefts)(C10) greatly enlarges the surface area of themuscle fiber.
The motor end plate is a specialized syn-apse. Its presynaptic membrane is the axo-lemma (C11), and its postsynaptic mem-brane is the folded sarcolemma (C12). Thesubstance transmitting nerve impulses tothe muscle fiber is acetylcholine. It is con-tained in clear synaptic vesicles (BC13).Upon stimulation of the axon, the neu-rotransmitter is released into the synaptic
cleft, resulting in receptor-mediated (ni-cotinic acetylcholine receptors) depolariza-tion of the membrane of the muscle fiber.
Tendon Organ (D–F)
The Golgi tendon organ lies at the junctionbetween tendon and muscle. It consists of agroup of collagen fibers (D14), which is sur-rounded by a thin sheath of connectivetissue and is innervated by a myelinatednerve fiber (D15). The nerve fiber loses itsmyelin sheath after passing through theconnective tissue capsule and divides into anumber of branches that coil around thecollagen fibers. The loosely arranged col-lagen fibers (E) are thought to tighten upontension (F) and thus exert pressure onto thenerve endings. The resulting impulse istransmitted by the nerve fiber via the poste-rior root to the spinal cord. Here it has an in-hibitory effect on the motor neurons, thuspreventing the muscle from excessivestretching or contraction.
B16 Basement membrane of the axon ter-minal.
B17 Basement membrane of the musclefiber.
C18 Synaptic cleft with common basementmembrane of axon terminal andmuscle fiber.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

313
Func
tion
alSy
stem
s
1
4
3
4
2
5
2
1811
13
12
10
17 16 9
13
2
6
15
14
7
8
Motor End Plate, Tendon Organ
A Motor end plates,overview
B End plate, electron-microscopicdiagram (according to Couteaux)
C Magnified detail of B (accordingto Robertson)
D Tendon organ (according toBridgeman)
E Tendon organ with muscle relaxed(according to Bridgeman)
F Tendon organ with muscle con-tracted (according to Bridgeman)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

314Fu
nction
alSy
stem
sFunctional Systems: Motor Systems
Muscle Spindle (A–F)
A muscle spindle, or neuromuscularspindle, consists of 5 – 10 thin striatedmuscle fibers (intrafusal muscle fibers)(A1) which are surrounded by a fluid-filledconnective-tissue capsule (A2). The fibers ofthe up to 10mm long spindles are arrangedin parallel with the other fibers of themuscle (extrafusal muscle fibers) and attacheither to the tendons of the muscle or to theconnective-tissue poles of the capsule. Asthe intrafusal fibers lie in the same longi-tudinal orientation as the extrafusal fibers,stretching and shortening of the muscle af-fects them in the same way. The number ofspindles within individual muscles is quitevariable. Muscles participating in delicateand precise movements (finger muscles)possess a large number of spindles, whereasmuscles for simple movements (trunkmuscles) contain far fewer spindles.
The central equatorial part (A3) of an intra-fusal fiber contains several cell nuclei but nomyofibrils; this part of the spindle is notcontractile. Only the two segments (A4) thatcontain striated myofibrils are contractile. Athick sensory nerve fiber (A5) terminates atthe central part; its terminal branches windaround the muscle fibers like spirals andform the annulospiral endings (AC6; B). Adelicate sensory fiber (A7) attaches in anumbelliform fashion (flower-spray ending)(A8, D) at one side, or at both sides, of theannulospiral ending.
Both contractile polar segments are inner-vated by thin fusimotor fibers (!-fibers)(A9). Their small motor end plates have onlypoorly developed subneural clefts; like theextrafusal muscle fibers, they are epilem-mal. The sensory annulospiral endings liebelow the basement membrane of themuscle fiber (C10) and, hence, are hypolem-mal. The !-fibers stem from small motorneurons in the anterior horn (!-mo-toneurons); impulses from these neuronscause contraction of the polar segments ofthe intrafusal fiber. This results in stretchingof the equatorial segment and does not onlystimulate the annulospiral ending but altersthe sensitivity of the spindle as well.
The muscle spindle is a stretch receptor,which is stimulated when the muscle isstretched but becomes inactive when themuscle is contracted. Upon stretching themuscle the impulse frequency increaseswith the change in muscle length. This way,the spindles transmit information on theprevailing length of the muscle. The im-pulses are transmitted not only via thespinocerebellar tracts to the cerebellum butalso via reflex collaterals directly to thelarge anterior horn cells ("-motoneurons).Stimulation of the latter neurons duringsudden stretching results in immediatemuscle contraction (stretch reflex, p. 50, F).
The muscle spindle contains two differenttypes of intrafusal fibers: the nuclear chainfibers (EF11) and the nuclear bag fibers (EF12).Both types of fiber are innervated by an-nulospiral endings. Flower-spray endingsare predominantly found at nuclear chainfibers. The thicker nuclear bag fibers re-spond to the ongoing stretching of themuscle, whereas the continuous state ofmuscle stretching is registered by the thin-ner nuclear chain fibers. Thus, musclespindles transmit to the cerebellum notonly information on the length of themuscle but also on the speed of musclestretching.
Apart from tendon organs and musclespindles, there are sensory end organs injoint capsules and ligaments (tension recep-tors) that constantly send information onmovement and posture of trunk and limbsto the cerebellum (anterior and posteriorspinocerebellar tracts).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

315
Func
tion
alSy
stem
s
4
3
5
6
10
12
12
12
11
11
11
6
7
8
1
24
9
Muscle Spindle
A Muscle spindle
B Annulospiral ending
C Detail of annulospiral ending
D Flower-spray ending
E Nuclear chain fibersand nuclear bag fibers
F Cross section througha muscle spindle
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

316Fu
nction
alSy
stem
sFunctional Systems: Motor Systems
Common Terminal Motor Pathway(A)
The common terminal pathway of all cen-ters involved in motor activity is the largeanterior horn cell (A1) and its axon (!-mo-toneuron), which innervates the voluntaryskeletal muscles. Most of the tracts runningto the anterior horn do not end directly onthe anterior horn cells but terminate on in-terneurons. The latter influence the motorneurons either directly or act by inhibitingor activating the reflexes between musclereceptors and motor neurons. The anteriorhorn is therefore not simply a relay stationas described earlier (p. 50) but a complex in-tegration apparatus that regulates motoractivity.
The central regions that influence motoractivity via descending pathways are inter-connected in many ways. The most impor-tant afferent pathways stem from the cere-bellum, which receives the impulses ofmuscle receptors via the spinocerebellartracts (A2) and the stimuli of the cortex viathe corticopontine tracts (A3). The cerebellarimpulses are transmitted via the parvo-cellular part of the dentate nucleus (A4) andthe ventral lateral nucleus of thalamus (A5)to the precentral cortex (area 4) (A6). Thecorticospinal (pyramidal) tract (A7) de-scends from area 4 to the anterior horn andgives off collaterals in the pons (A8) that re-turn to the cerebellum. Additional cerebel-lar impulses are transmitted via the em-boliform nucleus (A9) and the centromediannucleus of the thalamus (A10) to the striatum(A11) and via the magnocellular part of thedentate nucleus (A12) to the red nucleus(A13). From here fibers run in the centraltegmental tract (A14) via the olive (A15) backto the cerebellum and in the rubroreticu-lospinal tract (A16) to the anterior horn.Fibers from the globose nucleus (A17) run tothe interstitial nucleus of Cajal (A18) andfrom there in the interstitiospinal fasciculus(A19) to the anterior horn. Finally, cerebello-fugal fibers are relayed in the vestibular nu-clei (A20) and in the reticular formation(A21) to the vestibulospinal tract (A22) andthe reticulospinal tract (A23), respectively.
The descending pathways can be dividedinto two groups according to their effect onthemuscles: one group stimulates the flexormuscles, and another group stimulates theextensor muscles. The corticospinal tract andthe rubroreticulospinal tract activate mainlythe neurons of the flexor muscles and in-hibit the neurons of the extensor muscles.This corresponds to the functional impor-tance of the corticospinal tract for delicateand precise movements, especially those ofhand and finger muscles where flexormuscles play an important role. In contrast,the fibers of the vestibulospinal tract and thefibers from the pontine reticular formation in-hibit the flexors and activate the extensors.They belong to a phylogenetically old motorsystem that is directed against the effect ofgravity and, thus, is of special importancefor body posture and balance.
The peripheral fibers that run through theposterior root into the anterior horn origi-nate from themuscle receptors. The afferentfibers of the annulospiral endings (A24) ter-minate with their collaterals directly on the!-motoneurons, while the fibers of the ten-don organs (A25) terminate on inter-neurons. Many descending pathways in-fluence the !-neurons via the spinal reflexapparatus. They terminate on the large !-neurons and on the small "-neurons (A26).Since the "-neurons have a lower thresholdof stimulation, they are stimulated first,which results in the activation of musclespindles. The latter send their impulses tothe !-neurons. Thus, the "-neurons andmuscle spindles have a starter function forvoluntary movements.
A27 Accessory olive.A28 Skeletal muscles.A29Muscle spindle.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

317
Func
tion
alSy
stem
s
11
6
5
412
9
17
21
20
18
13
3 7
827
15
1619 22 23
2
25 24
26 1 1 26
2928
14
10
Common Terminal Motor Pathway
A Common terminal motor path-way (according to Hassler)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

318Fu
nction
alSy
stem
sFunctional Systems
Sensory Systems
Cutaneous Sensory Organs
The skin is endowed with a large number ofend organs that differ in their structures andtheir sensitivities to specific stimuli. Sincethe assignment of different sensory quali-ties to specific end organs is amatter of con-troversy, the nerve endings are classified ac-cording to morphological aspects:
! Free nerve endings! Encapsulated nerve endings! Transitional forms between these two
types
Free Nerve Endings (A–C, F)
Free nerve endings are found in almostevery tissue of the body. In the skin, theyreach into the lower layers of the epidermis(stratum germinativum). At the end of theirterminal branches, the axons send nodularor fingerlike evaginations through gaps inthe Schwann cell sheath. These evagina-tions are covered only by the basementmembrane and represent the receptor seg-ments of free nerve endings to which thesensations of pain and cold are attributed.
Delicate nerve fibers encircle the hair fol-licles, and their terminal segments ascend ordescend parallel to the hair shaft (A). The re-ceptor terminals lose their myelin sheathand are enclosed between two Schwanncells (B1) (sandwich arrangement), whichleave a cleft along the entire terminal seg-ment (B2). Through this cleft the axon ter-minal reaches the surface where it iscovered only by the basement membrane.The nerve endings (C3) are radially arrangedaround the hair follicle (C4) in such a waythat the sensory clefts face the follicle. Everymovement of the hair causes mechanicalstimulation of the nerve endings (red ar-rows in C), which is perceived as touch.
Also associated with hair follicles areMerkel’s touch cells (F5). These are large,clear epithelial cells that lie between thebasal cells (F6) of the outer root sheath andsend out fingerlike processes (F7) into theirsurroundings. Deformation of these cells
through movement of the hair results instimulation of the associated nerve fiber(F8). The nerve fiber loses its myelin sheathas it penetrates the basement membrane(F9) and sends branches to several tactilecells. The terminal segment widens into atactile meniscus (F10) and forms synapselikemembrane contacts (F11) with the Merkelcell.
Encapsulated Nerve Endings (D, E)
Meissner’s tactile corpuscles (D, E) lie inthe papillae of the dermis. They are mostdensely located in the palmar and plantarsurfaces of hands and feet. Their densities atthe distal ends, or digital pulps (especially atthe digital pulp of the index finger), are farhigher than at the rest of the surface. Thispattern of distribution is an indication ofhow important these corpuscles are fordelicate touch sensation.
The tactile corpuscles (E) are ovoid struc-tures that consist of stacks of flattened cells(probably Schwann cells) and are sur-rounded by a thin capsule. One or moreaxons (E12) enter the corpuscle from below,lose their myelin sheath, and run in convo-luted spirals between the stacked cells. Theclub-shaped swellings (E13) of the axonsrepresent the receptor terminals. Collagenfiber bundles (E14) radiate into the capsuleof the tactile corpuscle. They are continuouswith the tonofibrils in the epidermal cellsand transmit any mechanical deformationof the skin surface to the receptor terminals.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
319
Func
tion
alSy
stem
s
3
4
11 10 56 7
9
7 5 8
14
13
12
2
1
Cutaneous Sensory Organs
B Free nerve ending, electron-microscopicdiagram (according to Andres)
C Arrangement of nerve endings around a hairfollicle (according to Andres and von Düring)
D Meissner’s corpuscles in the skin
E Meissner’s tactile corpuscleF Merkel’s touch cells, electron-microscopic
diagram (according to Andres and von Düring)
A–C Free nerve endings associated with hair follicles
A Overview
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

320Fu
nction
alSy
stem
sFunctional Systems: Sensory Systems
Cutaneous Sensory Organs(continued)
Encapsulated Nerve Endings (continued)(A–C)
Pacini’s corpuscles, or lamellar corpuscles(A–C), are relatively large, up to 4mm longbodies located below the skin in the subcu-taneous tissue. They are also found in theperiosteum, in the vicinity of joints, and atthe surface of tendons and fasciae. Theyconsist of a large number of concentriclamellae which are arranged in three layers:
! The capsule! The lamellar outer core! The inner core
The capsule (A1) is formed by a few denselypacked lamellae that are reinforced by con-nective-tissue fibers. The lamellar outer core(AC2) consists of closed annular proto-plasmic lamellae (B3) that are separatedfrom each other by noncommunicating,fluid-filled spaces. Capsule and lamellarouter core are considered to be differentia-tion products of the perineurium. Thetightly packed layers of the fluid-free innercore (A–C4) are formed by Schwann cells.The inner core consists of two symmetricalstacks of semiannular lamellae (B5) that areseparated by a radial cleft. The protoplasmiclamellae are arranged alternately in such away that two juxtaposed lamellae stemfrom two different cells. The nerve fiber(AC6) enters the lamellar corpuscle at thelower pole and runs in the center of theinner core to the end of the core. Upon en-tering the inner core, the axon loses its my-elin sheath (A7), and it terminates with afew club-shaped swellings. The unmyeli-nated segment of the axon (A–C8) in theinner core represents the receptor portion.Perineurium (A9) (see p. 41, CD9).
Pacini’s corpuscles are not only pressure re-ceptors but, above all, vibration receptors.Electrical recordings from isolated cor-puscles have demonstrated that they arestimulated by both deformation anddecompression, but not by continuous pres-sure. In experiments, these highly sensitivevibration receptors registered, for example,
the vibration of the floor caused by some-one passing by.
Transitional Forms (D, E)
There are numerous different transitionalforms between free and encapsulated nerveendings. In all of them the nerve fibersramify into terminal complexes in whichthe delicate axonal branches form glomerulior treelike ramifications and terminate withclub-shaped swellings. These formationsare enclosed in a more or less prominentconnective tissue capsule. They include theovoid bulboid corpuscles (Krause’s end bulbs)in the papillae of the dermis, the roundGolgi–Mazzoni corpuscles, and the elon-gated Ruffini’s corpuscles (D) in the subcu-taneous tissue. Such structures are foundnot only in the skin but also in the mucosa,in joint capsules, in the coverings of innerorgans, and in the adventitial coat of largearteries. The most diverse forms are foundin the genitals, especially in glans (E) andclitoris.
The assignment of the different sensoryqualities to specific end organs is amatter ofcontroversy. It is, however, recognized thatthe corpuscular end organs are importantmechanoreceptors and that free nerveendings are pain receptors. How the sensa-tion of heat and cold is brought about hasnot yet been elucidated. However, thedifferent structures of the receptors suggestthat there is selectivity with respect to thetype of stimulus processed.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

321
Func
tion
alSy
stem
s
48
12796
4
3
85
8
4
2
6
Cutaneous Sensory Organs
A Pacini’s corpuscle, longitudinalsection and cross section
B Centre of a Pacini’s corpuscle,electron-microscopic diagram(according to Quilliam)
C Pacini’s corpuscle(according to Munger)
D Ruffini’s corpuscle(according to Ruffini)
E End organ from the skin of theglans penis (according to Dogiel)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

322Fu
nction
alSy
stem
sFunctional Systems: Sensory Systems
Pathway of the Epicritic Sensibility(A–C)
The nerve fibers transmitting impulses forthe senses of touch, vibration, and joints origi-nate from neurons of the spinal ganglia(A1), while the fibers for face and sinusesstem from neurons of the trigeminal gan-glion (Gasser’s ganglion, semilunar gan-glion) (A2) (1st neuron). Touch stimuli aretransmitted by two types of fibers; thick,well-myelinated nerve fibers terminate atthe corpuscular end organs, while thin nervefibers terminate at hair follicles. The cen-tripetal axons of the neurons enter the spi-nal cord via the posterior root, with thethick myelinated fibers running through themedial portion of the root (p. 63, F8). Theymerge with the posterior funiculi (AB3) insuch a way that the newly entering fibersborder laterally; as a result, the sacral andlumbar fibers lie medially and the thoracicand cervical fibers laterally. The sacral, lum-bar, and thoracic bundles form the gracilefasciculus (Goll’s tract) (B4), while the cervi-cal fibers form the cuneate fasciculus (Bur-dach’s tract) (B5).
The primary fibers (gracile funiculus andcuneate funiculus) terminate in a corre-sponding arrangement on the neurons ofthe dorsal column nuclei (A6) (2nd neuron),gracile nucleus (B7), and cuneate nucleus(B8), which therefore exhibit the same so-matotopic arrangement as the posteriorfuniculi. Each neuron in the dorsal columnnuclei receives its impulses from a specifictype of receptor. The cutaneous supply areaof a neuron is small in the distal segments ofthe limbs (hand, finger) but larger in theproximal segments. As electrophysiologicalstudies have demonstrated, the neurons re-ceiving impulses from specific receptorsshow a somatotopic arrangement as well;close to the surface of the nuclei lie the neu-rons for the hair follicle receptors, in themiddle those for the touch organs, and stilldeeper those for the vibration receptors.
Corticofugal fibers from the central region(precentral gyrus and postcentral gyrus)run via the corticospinal (pyramidal) tractto the dorsal column nuclei; fibers from the
lower limb area of the central region termi-nate in the gracile nucleus, while fibersfrom the upper limb area terminate in thecuneate nucleus. The corticofugal fibershave a postsynaptic or presynaptic inhibi-tory effect on neurons of the dorsal columnnuclei and therefore attenuate the incomingafferent impulses. Thus, the cortex is able toregulate in these relay nuclei the input ofimpulses coming from the periphery.
The secondary fibers ascending from thedorsal column nuclei (2nd neuron) form themedial lemniscus (B9). In the decussation ofthe medial lemnisci (B10) the fibers cross tothe opposite sides, with the fibers from thegracile nucleus lying ventrally and thosefrom the cuneate nucleus lying dorsally.Later, the gracile fibers take a lateral posi-tion (B11) and the cuneate fibers take a me-dial position (B12). The secondary fibersfrom the trigeminal nuclei (B13), trigeminallemniscus, join the medial lemniscus at thelevel of the pons and become located dor-somedially to it in the midbrain (B14).
The medial lemniscus extends to the lateralpart of the ventral posterior nucleus ofthalamus (AC15); the fibers of the gracilenucleus terminate laterally, while those ofthe cuneate nucleus terminate medially.The trigeminal fibers (C16) terminate in themedial part of the ventral posterior nucleus.This results in a somatotopic organization ofthe nucleus. The arrangement of fibers ispreserved in the projections of thethalamocortical fibers (3rd neuron) to thecortex of the postcentral gyrus (A17) andforms the basis for the somatotopic organi-zation of the postcentral area (p. 250).
Hence, the pathway of the epicritic sensi-bility consists of three neurons relayed intandem, with a demonstrated somatotopicorganization in each relay station and at theterminal station.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
323
Func
tion
alSy
stem
s
15 16
17
15
13
6
2
3
1
12 11
14
13
8
10
7
4
5
4
5
3
3
4 3
9 12
11
Epicritic Sensibility
C Terminal somatotopic arrangement inthe thalamus (according to Hassler)
A Pathway ofthe sensation of touch
B Position of the posterior funiculusand the medial lemniscus
SL TC
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

324Fu
nction
alSy
stem
sFunctional Systems: Sensory Systems
Pathway of the ProtopathicSensibility (A–C)
The thin, poorly myelinated or unmyeli-nated nerve fibers for the senses of pain andtemperature originate from the small neu-rons of the spinal ganglia (A1) (1st neuron).Their centripetal axons enter the spinal cordthrough the lateral part of the posterior root(p. 63, F7). They bifurcate in Lissauer’s tractand terminate in the dorsal border region ofthe substantia gelatinosa and in the poste-rior horn (p. 57, A2). The secondary fiberscross to the opposite side and ascend inthe anterolateral funiculus as lateralspinothalamic tract (B2) (2nd neuron). Thetract does not form a discrete fiber bundlebut consists of loosely arranged fibers thatare mixed with fibers of other systems. Thefibers entering at various root levels joinventromedially. Thus, the sacral fibers lie atthe surface, and the cervical fibers thatjoined last lie in the inner part of the an-terolateral funiculus (p. 57, A1; p. 140, B8).
The input of impulses is controlled by de-scending fibers that originate in the centralregion, in the anterior lobe of the cerebel-lum, and in the reticular formation. Thesefibers terminate in the substantia gelati-nosa, a relay station in which the peripheralimpulses aremodulated by the excitatory orinhibitory influences of higher centers.Numerous axo-axonal synapses, which aretypical for presynaptic inhibition, have beendemonstrated in the substantia gelatinosa.
In the medulla oblongata, the lateralspinothalamic tract (spinal lemniscus) is lo-cated at its lateral margin above the oliveand gives off numerous collaterals to the re-ticular formation. Here, too, a considerableportion of the fibers (spinoreticular tract)terminate. The reticular formation is part ofthe ascending activation system (p. 146), thestimulation of which puts the organism intoa state of alertness. Hence, the impulsestransmitted via the pain pathway not onlycause a conscious sensation but also in-crease the attention via the reticular forma-tion. By contrast, the pathway of theepicritic sensibility runs through the brainstem without giving off any collaterals.
The spinothalamic fibers join the mediallemniscus in the midbrain and take a dor-solateral position. A large portion of themterminate on the cells of the ventral poste-rior nucleus of thalamus (AC3) (3rd neuron)in somatotopic organization, predomi-nantly in a ventral parvocellular region. Ter-tiary fibers extend from here to the postcen-tral region (A4). Other spinothalamic fibersterminate in other thalamic nuclei, for ex-ample, in the intralaminar nuclei.
The anterior spinothalamic tract (B5)transmits crude senses of touch and pressure.Its fibers cross from the posterior horn (2ndneuron) to the contralateral anteriorfuniculus (p. 57, A3). The position of thetract in the medulla oblongata is a matter ofcontroversy. It is though to lie either medi-ally to themedial lemniscus (B6) or laterallyto the olive (B7). In the pons and midbrain,the fibers join the medial lemniscus (B8)and terminate on the cells of the ventral pos-terior nucleus of thalamus (3rd neuron).
Pain and temperature fibers for the face andsinuses originate from the neurons of thetrigeminal ganglion (A9), the centripetalaxons of which terminate in the spinal nu-cleus of the trigeminal nerve (AB10) (p. 125,BC4). In the spinal tract of the trigeminalnerve, the pain-transmitting fibers arethought to lie laterally and those transmit-ting temperature further medially. The sec-ondary trigeminal fibers (B11) (trigeminallemniscus ) join the medial lemniscus.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

325
Func
tion
alSy
stem
s
11
10
72
2
2
2
6
5
5
5
3
28
3
9 10
1
4
Protopathic Sensibility
C Termination of the spinothalamicfibers in the thalamus
A Pathways of the sensation of pain
B Position of the spinothalamictract and the medial lemniscus
C T L
S
CT
LS
CT
LS
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

326Fu
nction
alSy
stem
sFunctional Systems: Sensory Systems
Gustatory Organ (A–E)
Taste Buds
Different taste sensations are registered bythe taste buds which, together with the ol-factory epithelium, constitute the chemore-ceptors. Taste buds, or gustatory receptors(B–D1; E), are found in large numbers inthe lateral walls of the vallate papillae (AB2),and in moderate numbers in the fungiformpapillae and in the foliate papillae. In addi-tion, there are isolated taste buds in the softpalate, in the posterior pharyngeal wall, andin the epiglottis.
A taste bud consists of up to 50 spindle-shaped, modified epithelial cells. Each bar-rel-shaped taste bud has a small opening atthe epithelial surface, the taste pore (C3),into which the sensory cells send short cy-toplasmic processes. New sensory cells arecontinuously generated from epithelial cellsat the base of the taste bud. The conversionof epithelial cells into sensory cells takesplace at the periphery, while the fullydeveloped sensory cells lie in the center.Taste buds can multiply by fission, and twinbuds or multiple buds are found occa-sionally (C4).
The base of the taste bud is innervated bythin, myelinated nerve fibers (D5) thatbranch and supply the adjacent epitheliumas well. We therefore distinguish betweenextragemmal and intragemmal nerve fibers.Upon entering the taste bud, the intragem-mal nerve fibers (usually 2 – 3) lose theirSchwann cell sheath and arborize. Accord-ing to electronmicroscopic studies, the deli-cate nerve endings lie between the sensorycells and invaginate these so deeply thatthey appear to be intracellular. When a tastenerve is severed, the taste buds degenerate.Regeneration of the nerve induces the for-mation of new taste buds in the epitheliumof the tongue.
Electron micrographs show the long neck(E6) of the sensory cell extending into thetaste pore and ending with a dense borderof microvilli (E7). This is the receptor por-tion of the cell. The microvilli considerablyenlarge the cell surface coming into contact
with the substances to be tasted. The tastebud contains three different cell types thatrepresent different developmental stages ofthe sensory cells: the dark supporting cells(E8), which contain numerous secretorygranules, the clear taste cells (E9) withoutgranules, and the small basal cells (E10). Thefloor of the taste pore as well as the cleftsbetween the receptor portions of the cellscontain amucous substance that is probablysecreted by the taste cells.
There are different theories as to how a gus-tatory stimulus is produced. Molecules thattrigger a specific taste sensation bind tospecific receptor sites on the sensory cellmembrane. These sites recognize the con-figuration of the molecule and cause achange in membrane potential, which fi-nally leads to excitation of the associatedaxon terminal.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

327
Func
tion
alSy
stem
s
2
2
1 1
3
4
1
4
5
1
9
10
7
6
8
Taste Buds
A Tongue with papillae
B Section through a vallate papilla
C–E Taste buds
C Overview, detail of B
D Nerve endings
E Electron-microscopic diagram(according to Popoff)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

328Fu
nction
alSy
stem
sFunctional Systems: Sensory Systems
Gustatory Organ (continued)
Taste Buds (continued) (A)
The four basic taste sensations (sweet, sour,salty, and bitter) are not equally well per-ceived by all taste buds. Some buds reactonly to sweet or only to sour, while othersregister two or three qualities without ex-hibiting morphological differences. Theperception of individual qualities on thesurface of the tongue is differentially dis-tributed. Sour is perceived especially at thelateral margins of the tongue (A1), salty atthe margins and the tip (A2), bitter at thebase of the tongue (A3), and sweet at the tipof the tongue (A4).
Taste Fibers (B, C)
The taste fibers are assigned to three cranialnerves, namely, the facial nerve (intermediatenerve) (B5), the glossopharyngeal nerve (B6),and the vagus nerve (B7). The fibers originatefrom pseudounipolar neurons in the cranialnerve ganglia, namely, the geniculate gan-glion (B8), the petrosal ganglion (inferiorganglion of the glossopharyngeal nerve)(B9), and the nodose ganglion (inferior gan-glion of the vagus nerve) (B10). Taste fibersof the facial nerve run in the chorda tympanito the lingual nerve (B11) (p. 131, AB9) andsupply the receptors of the fungiformpapillae on the anterior two-thirds of thetongue. Taste fibers of the glossopharyngealnerve run in the lingual branches of theglossopharyngeal nerve to the posteriorthird of the tongue and supply the receptorsof the vallate papillae. In the tonsillarbranches of the glossopharyngeal nerve,taste fibers run to the soft palate. Tastefibers of the vagus nerve reach the epiglottisand the epipharynx through the pharyngealbranches of the vagus nerve.
The centripetal axons of the neurons enterthemedulla oblongata and form the solitarytract. The primary taste fibers terminate inthe solitary nuclear complex (BC12) approxi-mately at the level of the nerve entry. Thesolitary nuclear complex widens in this re-gion and contains a cell column also re-ferred to as the gustatory nucleus.
The secondary taste fibers originate from thesolitary nuclear complex. Their exact coursein the brain stem is not known. It is assumedthat the majority of fibers cross to the op-posite side as arcuate fibers and join theme-dial lemniscus (C13). They are thought to oc-cupy the most medially located part in thelemniscus. The secondary taste fibers termi-nate in the medial part of the ventral poste-rior nucleus of thalamus (C14). From here, ter-tiary taste fibers project to a cortical area onthe ventral aspect of the parietal operculum(C15), below the postcentral region. The ter-minals in thalamus and cerebral cortex havebeen confirmed by experiments inmonkeys. In humans, a loss of taste percep-tion on the contralateral half of the tonguehas been observed when these regions aredestroyed by disease.
Some of the secondary taste fibers run tothe hypothalamus. They are thought tobranch off from the medial lemniscus in themidbrain and to run through the mamillarypeduncle to the mamillary body. Otherfibers are relayed in the ventral tegmentalnucleus and are thought to reach the hy-pothalamus through the posterior longi-tudinal fasciculus (Schütz’s bundle)(p. 144B).
Collaterals extend from the neurons of thesolitary nuclear complex to the parasympa-thetic salivatory nuclei. This way, taste sen-sation can cause reflex secretion of saliva.Collaterals running to the posterior nucleusof the vagus nerve form the connection bywhich taste stimuli produce reflex secretionof gastric juice.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

329
Func
tion
alSy
stem
s
14
13
13
11
6
9
7
8 5
10 12
12
15
1 2
3 4
Taste Pathways
A Localization of different tastequalities on the tongue: 1, sour;2, salty; 3, bitter; 4, sweet
B Peripheral taste fibers C Position of the taste pathway
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

330Fu
nction
alSy
stem
sFunctional Systems: Sensory Systems
Olfactory Organ (A–D)
In humans, the olfactory epithelium occu-pies a small region in both nasal cavities (ol-factory region) (A1) at the upper margin ofthe superior nasal concha and on the op-posite surface of the nasal septum. Themul-tilayered sensory epithelium is composed ofsupporting cells (C2) and receptor cells (C3)that are characterized by pale, deep-lyingcell nuclei. The olfactory region also con-tains small mucous glands, the olfactoryglands (Bowman’s glands). Their secretioncovers the olfactory mucosa as a thin film.
The apical part of the sensory cell tapersinto a thin shaft that slightly extends be-yond the surface of the epithelium. Thisknoblike olfactory vesicle (C4) is occupied bya number of olfactory hairs. At the basal end,the ovoid cell body forms a fine process that,together with several other processes, is en-veloped by Schwann cells. The bundledprocesses (fila olfactoria) represent the ol-factory nerves (AC5) and extend through theopenings of the cribriform plate to the ol-factory bulb (A6) (p. 228, A). In the olfactorybulb, the processes terminate in the ol-factory glomeruli, where they form synapticcontacts with the dendrites of mitral cells.The epithelial sensory cells are bipolar neu-rons; the short dendrite represents the re-ceptor part and the long axon runs as cen-tripetal fiber to the olfactory bulb.
Electron micrographs of olfactory cells (inthe cat) show that the apical shaft of the cell(D7) terminates with a knob (D8) fromwhich numerous long olfactory cilia (D9) ex-tend. The terminal parts of the sensory cilialie in the mucous layer (D10) that seals theentire surface of the olfactory epitheliumagainst the air space. Shaft and knob containmicrotubules, numerous mitochondria(D11), and some lysosomes (D12). The knobextends above the surface of the supportingcells, which exhibit a dense border of micro-villi (D13).
How the sensory cells receive the differentodors is currently the topic of intensive re-search. The odoriferous substances must bewater-soluble in order to dissolve in the
superficial mucous layer and reach thesensory cilia where they bind to specificmembrane receptors. In sufficiently highconcentrations, they induce depolarizationof the membrane, which is conducted as anaction potential in the cell’s axon. As in thecase of taste, it is assumed that there are afew basic qualities of odor and that onesensory cell registers only one particular ba-sic quality via specific receptors. Since sub-stances belonging to a group of odors haveroughly the same molecular size, it seemspossible that the membrane of an olfactorycilium reacts to only one particular molecu-lar size. Indeed, recent studies suggest thateach sensory cell expresses only one recep-tor type. In the mouse, the sensory cells ofone receptor type project to only two of the1800 glomeruli in the olfactory bulb (centralolfactory pathway, p. 228, AB).
Apart from the olfactory nerves, two otherpaired nerves run from the nasal cavity tothe brain, namely, the terminal nerve andthe vomeronasal nerve. The terminal nerve(B14) consists of a bundle of delicate nervefibers that extends from the nasal septumthrough the cribriform plate to the terminallamina and enters below the anterior com-missure into the brain. The bundle containsnumerous neurons and is regarded as an au-tonomic nerve. The vomeronasal nerve (B15),which runs from the vomeronasal organ (Ja-cobson’s organ) to the accessory olfactorybulb, is well developed in lower vertebratesbut can be demonstrated in humans onlyduring embryonic development. In reptiles,the vomeronasal organ is a sensoryepithelium in a pocket of the mucosa of thenasal septum; it is thought to play an impor-tant role in tracking down prey.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

331
Func
tion
alSy
stem
s
12
7
11
8
13
10
9
6
51
14
15
5
3
2
4
Olfactory Organ
A Position of the olfactoryepithelium
B Terminal nerve andvomeronasal nerve
C Olfactory epithelium
D An olfactory cell between two supporting cells, electron-microscopic diagram (according to Andres)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

332Fu
nction
alSy
stem
sFunctional Systems
Limbic System
Overview
The phylogenetically old parts of the telen-cephalon, together with its border zonesand its connections to subcortical centers,are collectively known as the limbic system.It is not a circumscribed, topically organizedsystem of pathways but rather a collectionof functionally closely related nuclei andcortical areas. The system has also beencalled the visceral or emotional brain, acatchphrase highlighting its functional sig-nificance. As the concept of the limbic sys-tem is based on functional connections, itsbasic anatomical structures are onlyvaguely defined.
Subdivision (A)
The cortical regions belonging to the limbicsystem form a C-shaped complex on themedial aspect of the hemisphere that con-sists of the parahippocampal gyrus (A1), thecingulate gyrus (A2), and the subcallosalarea (parolfactory area) (A3). The cingulategyrus, which is also called limbic gyrus, gavethe system its name. An inner and an outerarch can be distinguished at the medialaspect of the hemisphere. The outer arch(parahippocampal gyrus, cingulate gyrus,and parolfactory area) is formed by theborder zones of the archicortex (periarchi-cortex) and by the indusium griseum of thecorpus callosum (rudimentary archicortex).The inner arch consists of archicortical andpaleocortical regions, namely, the hippo-campal formation (A4), the fornix (A–C5),the septal area (A6), the diagonal band ofBroca, and the paraterminal gyrus (A7). Animportant component is also the amyg-daloid body. Some subcortical nuclei withclose fiber connections to the limbic cortexare included in the system, such as themamillary body, the anterior nuclei ofthalamus, the habenular nucleus, and addi-tionally, in the midbrain the posteriortegmental nucleus, the anterior tegmentalnucleus, and the interpeduncular nucleus.
Pathways (B, C)
The limbic system is connected with the ol-factory centers through several fiberbundles. The fibers of the lateral olfactorystria terminate in the cortical parts of theamygdaloid body (p. 228, B).
The limbic system influences the hypo-thalamus via three pathways:
! The fornix, the precommissural fibers ofwhich terminate in the preoptic area (B8)and in the nuclei of the tuber cinereum(B9)
! The terminal stria (B10), which runs fromthe amygdaloid body (B11) to the tuberalnuclei
! The ventral amygdalofugal fibers (B12)
The connection to the tegmental nuclei ofthe midbrain is established through thedescending bundle of the habenular nu-cleus (habenulotegmental tract andhabenulopeduncular tract) and through thepathways of the mamillary body (mamillarypeduncle and mamillotegmental fasciculus).The efferent mamillotegmental fasciculus(C13) and the afferent peduncle of themamillary body (C14) form a neuronal cir-cuit.
Within the limbic system runs a multiplepathway, the neuronal circuit of Papez. Theefferent fibers of the hippocampus reachthe mamillary body (C15) through the for-nix (C5). Here the impulses are relayed tothe bundle of Vicq d'Azyr (C16), which ex-tends to the anterior nuclei of thalamus(C17). The latter project to the cortex of thecingulate gyrus (C18), from where the cin-gulate fiber bundles (C19) return to the hip-pocampus.
Connections of the neocortex to the limbicsystem exist especially via the entorhinalarea in the parahippocampal gyrus (A1),which projects to the hippocampus (per-forant pathway, see p. 234, A4).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

333
Func
tion
alSy
stem
s
3 7
1
65
4
2
5
10
98
1819
17 516
15
13
14
12
11
3 7
1
65
4
2
5
10
98
1819
17 516
15
13
14
12
11
Limbic System: Subdivision and Pathways
A Cortical areas of the limbic sys-tem (according to Stephan)
B Connections of the limbic systemwith the hypothalamus (accordingto Akert and Hummel)
C Neuronal circuit of Papez(according to Akertand Hummel)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

334Fu
nction
alSy
stem
sFunctional Systems: Limbic System
Cingulate Gyrus
The cingulate gyrus is connected with theolfactory cortex, the hypothalamus, thefrontal cortex, the caudal portion of theorbital cortex, and the rostral portion of theinsular cortex. Electrical stimulation of itsrostral region in humans leads to changes inblood pressure, pulse rate, and respirationrate. Changes in temperature, erection ofthe hair, dilatation of pupils, increasedsalivation, and altered gastric motility havebeen observed in stimulation or lesion ex-periments in monkeys.
The cortex of the cingulate gyrus influencesthe hypothalamus and the autonomicnervous system. The limbic system ob-viously plays an important role in the regu-lation of basic vital processes, such as foodintake, digestion, and reproduction (see alsohypothalamus, p. 198, and amygdaloidbody, p. 226). These are the primary vitalfunctions that serve self-preservation aswell as species preservation and are alwaysaccompanied by pleasurable sensations orreluctance. Hence, emotional states havebeen attributed to the limbic system.
Septal Area (A–C)
There are strong connections to the hippo-campus, the central structure of the limbicsystem. Cholinergic and GABAergic neuronsof the medial septal nucleus project to thehippocampus and the dentate gyrus; col-laterals of the CA3 pyramidal cells (p. 234)project back to the lateral septal nucleus.
As with stimulation of the amygdaloid body,electrical stimulation of the septal area(BC1) triggers oral reactions (licking, chew-ing, retching), excretory reactions (defeca-tion, urination), and sexual reactions (erec-tion). The septal area, especially the diago-nal band of Broca, is also the preferred local-ization for self-stimulation experiments inthe rat (A, B). Rats carrying an implantedelectrode that facilitates stimulation of thisarea continuously stimulate themselves bypressing a button. The urge for stimulationis so strong that the animals prefer stimula-tion to feeding, even after periods of hunger
or thirst. Stimulation near the anterior com-missure (C2) produces a euphoric reactionand the general feeling of well-being inhumans.
Klüver–Bucy syndrome (D). This syn-drome was observed following bilateral re-moval of the temporal lobe (D3) in themonkey. The lobectomy affected the neo-cortex, hippocampus, and amygdaloid body,and resulted in a polysymptomatic be-havioral syndrome. The animals becametame and trusting; they lost their naturalbehavior and shyness even toward danger-ous objects or animals (such as snakes). Inaddition, they completely lost any sexual in-hibition (hypersexuality). Familiar objectswere no longer recognized visually (visualagnosia); instead, they were put in themouth (oral tendency) and examined re-peatedly this way as if theywere completelyunknown each time theywere encountered.The disturbed memory indicates a specialrole of the hippocampus in processes oflearning and memory. Information from theneocortex, such as visual impressions, istransmitted to the hippocampus via the en-torhinal area and examined for its noveltyvalue.
There is no doubt that the limbic system in-cludes very divergent and complex func-tions. It is likely that increases in knowledgewill soon lead us to abandon the summaryand vague concept of one limbic system.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

335
Func
tion
alSy
stem
s
1
1
2
3
Stimulation Experiments
A, B Self-stimulation experiment in the rat (according to Olds)
A Experimental design
B Area of stimulation
C Foci of stimulation for euphoricreactions in humans (according toSchaltenbrand, Spuler, Wahren, andWilhelm)
D Removal of the temporal lobe,Klüver–Bucy syndrome (accord-ing to Klüver and Bucy)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Sensory Organs The Eye
Structure 338Visual Pathway and Ocular
Reflexes 354
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

338Th
eEy
eThe Eye
Structure
Eyelids, Lacrimal Apparatus, andOrbital Cavity
Eyelids (A–C)
The eyeball is embedded in the orbit and iscovered by the eyelids. The upper eyelid (A1)and the lower eyelid (A2) demarcate thepalpebral fissure. The latter ends in themedialangle of the eye (A3) with a recess enclosingthe lacrimal caruncle (A4).
In Oriental People, the upper eyelid con-tinues medially onto the side of the nose asa vertical fold of skin, the palpebronasal fold.The fold can also be observed as a transitoryformation in infants and is known as epican-thus.
The eyelids are reinforced by firm plates ofconnective tissue consisting of collagenousfibers, tarsus superior (B5) and tarsus infe-rior (B6); the tarsal plates, or palpebral car-tilages, are attached to the margin of theorbit by the lateral palpebral ligament (B7)and the medial palpebral ligament. The tar-sal plates contain elongated tarsal glands,the meibomian glands (C8), which spreadover the entire height of the eyelids. Theirsecretion prevents the tears from flowingover the edge of the eyelids. They open atthe posterior margin of the eyelid. Severalrows of eyelashes, the cilia (C10), emergefrom the anterior margin (AC9). The innerwall of the eyelids is lined by the conjunctiva(C11), which extends to the anterior aspectof the eyeball at the conjunctival fornix(C12). The smooth superior tarsal muscle(C13) and inferior tarsal muscle (C14) (inner-vated by the sympathetic nervous system),which control the size of the palpebral fis-sure, attach at the tarsus. The eyelids areclosed by the orbicular muscle of the eye(C15) (facial nerve, p. 122). The upper lid islifted by the levator muscle of the upper eye-lid (BC16) (oculomotor nerve), which at-taches at the upper margin of the opticcanal. Its superficial tendinous membrane(C17) penetrates into the subcutaneous con-nective tissue of the upper lid, while thedeep tendinousmembrane (C18) attaches atthe upper margin of the tarsus.
Lacrimal Apparatus (B)
The lacrimal gland (B19) lies above the lateralangle of the eye; it is divided by the tendonof the levator muscle of the upper eyelidinto an orbital part (B20) and a palpebralpart (B21). Its excretory ducts at the con-junctival fornix secrete the lacrimal fluid(tears), which keeps the anterior aspect ofthe eyeball continuously moist and collectsin the lacrimal lake of the medial angle ofthe eye. Here at the inner surface of eacheyelid is a small opening, the lacrimal point(B22), which leads to the lacrimalcanaliculus (B23). The canaliculi ascend anddescend, respectively, and then turn at aright angle to join and open into the lacrimalsac (tear sac) (B24), from where the na-solacrimal duct (tear duct) (B25) leads to theinferior meatus of the nose. Blinking notonly results in uniform moistening of theeyeball surface but also has a sucking effecton the flow of tears by expanding and con-stricting the nasolacrimal duct.
Orbit (C)
The orbital cavity (eye socket) is lined by theperiosteum (periorbita) (C26) and is filledwith fatty tissue, the orbital fat body (C27),in which the eyeball (C28), the optic nerve(C29), and the eye muscles (C30) areembedded. At the anterior margin of theorbit, the fatty tissue is demarcated by theorbital septum (BC31). The fatty tissue isseparated from the eyeball by a connectivetissue capsule, the bulbar sheath (C32),which encloses the sclera (C33).
C34 Choroidea.C35Osseous wall of the orbit.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

339
TheEy
e
20
19
17
153112
13
11
8
928
32343330
29
30
10
1431
15
18 35
2616 27
7
31
6
5
1623
24
25
22
21
4
3
1
29
20
19
17
153112
13
11
8
928
32343330
29
30
10
1431
15
18 35
2616 27
7
31
6
5
1623
24
25
22
21
4
3
1
29
Eyelids, Lacrimal Apparatus, Orbital Cavity
A The eye
B Lacrimal apparatus
C Longitudinal section throughthe orbit
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

340Th
eEy
eThe Eye: Structure
Muscles of the Eyeball (A–E)
The eyeball is attached bymembranes to thecapsule of the orbital fat body, and it canmove in all directions. Movements areachieved by six extra-ocular muscles,namely four rectusmuscles and two obliquemuscles. The tendons of origin of the rectusmuscles form a funnel-shaped ring aroundthe optic canal, the common annular tendon(AB1). The superior rectus muscle (A–C2)(oculomotor nerve) runs above the eyeballin a slightly oblique, outward direction. Theinferior rectus muscle (A–C3) (oculomo-tor nerve) runs beneath the eyeball in thesame direction. At the nasal aspect of theeyeball lies themedial rectus muscle (AC4)(oculomotor nerve), and at the temporalaspect lies the lateral rectus muscle(A–C5) (abducens nerve). At a distance of0.5–1 cm from themargin of the cornea, theflat tendons of the rectus muscles attach tothe sclera of the eyeball. The superior ob-lique muscle (AC6) (trochlear nerve) origi-nates medially at the lesser wing of thesphenoid bone and extends almost to themargin of the orbit. Here, its tendon passesthrough the trochlea (A7), a wide loop con-sisting of fibrous cartilage and lined with asynovial sheath. The tendon then turns at anacute angle in posterolateral direction andattaches underneath the superior rectusmuscle on the temporal side of the uppereyeball. The inferior oblique muscle (BC8)(oculomotor nerve) originates medially atthe infraorbital margin and runs to the tem-poral side of the eyeball. Levator muscle ofupper eyelid (B9).
Movements of the eyeball:
! Rotation around the vertical axis towardthe nose (adduction) and toward thetemple (abduction)
! Rotation around the horizontal axis up-ward (elevation) and downward (depres-sion)
! Rotation around the sagittal axis with roll-ing of the upper half of the eyeball towardthe nose (inward rotation, or intorsion)and toward the temple (outward rotation,or extorsion)
The medial rectus muscle (C4) causes adduc-tion; the lateral rectus muscle (C5) abduction.
The superior rectus muscle (C2) raises theeyeball and causes also a slight adductionand intorsion; the inferior rectus muscle (C3)lowers the eyeball and causes also a slightadduction and extorsion.
The superior oblique muscle (C6) rotates theupper pole of the eyeball inward slightlydepresses and abducts the eyeball; the infe-rior oblique muscle (C8) rotates the upperpole of the eyeball outward and slightly ele-vate and abducts the eyeball.
This functional description applies only whenlooking straight ahead (medial gaze) and whenboth eyeballs have parallel axes of vision. Duringocular movements and simultaneous reactions ofconvergence (p. 358, C) and divergence, the func-tions of individual muscles changes, e.g., the twomedial rectus muscles are synergists during con-vergence and antagonists during lateral gaze. Thechange in function is determined by the deviationof the visual axis from the anatomical axis of theorbit. When the two axes overlap during abduc-tion of the eyeball by 23!, the superior rectusmuscle (D10, looking straight ahead) and the infe-rior rectus muscle lose their accessory functions;the first turns into a true levator of the eyeball(D11) and the latter into a true depressor of theeyeball. During maximal adduction of the eyeballup to 50!, the superior oblique muscle (E12, look-ing straight ahead) turns into a true depressor ofthe eyeball (E13) and the inferior oblique muscleinto a true levator of the eyeball. All extra-ocularmuscles are involved with tension and relaxationduring every eye movement, and the position ofthe eyeball determines the function of eachmuscle.
Precision and speed of muscle function rely onstructural characteristics. Apart from the intra-fusal fibers of muscle spindles, numerous extra-fusal fibers are supplied with sensory anulospiralnerve endings. The motor units are extremelysmall; about six ocular muscle fibers are suppliedby one nerve fiber. For comparison, one nervefiber innervates 100–300 muscle fibers in themuscle of the fingers, in other muscles often morethan 1500 fibers.
Clinical Note: Paralysis of individual eyemuscles causes double vision, when objects in thesurroundings are perceived twice. The relativeposition of the two images—side by side or shiftedobliquely above or obliquely below one another—indicates which muscle is paralyzed.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

341
TheEy
e
7
4
62
5
1
2
9
5
8
3
1
3
4
35
10 11 12 13
2
6
8
7
4
62
5
1
2
9
5
8
3
1
3
4
35
10 11 12 13
2
6
8
Muscles of the Eyeball
A Extraocular muscles, view fromabove (right eye)
B Extraocular muscles, lateral view
C Functions of the extraocular muscles of the right eye
D Superior rectus muscle of the right eye, gazestraight ahead (left) and abducted by 23! (right)
E Superior oblique muscle of the right eye, gazestraight ahead (left) and adducted by 50! (right)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

342Th
eEy
eThe Eye: Structure
The Eyeball
Development (A)
The light-sensitive part of the eye is aderivative of the diencephalon. At end of thefirst month of embryonic development, thetwo optic vesicles (A1) are formed as evagi-nations of the prosencephalon (A2). Theoptic vesicles then induce thickenings in theectoderm of the head, the lens placodes (A3),which later invaginate as lens vesicles (A4).The epithelial cells of the posterior vesiclewall elongate into lens fibers (A5), whichlater form the main part of the lens. Thecells of the anterior vesicle wall persist aslens epithelium. The anterior and posteriorwalls of the optic vesicle approximate eachother to form the optic cup (A6). The lumenof the vesicle, originally a part of theventricular system, the optic ventricle (A7),becomes a narrow cavity. The optic cup con-sists of an inner layer, the neural layer (A8),and an outer layer, the pigmented layer (A9);both are layers of the retina. The hyaloidartery (A10) first extends to the lens butlater regresses.
Structure (B)
The anterior aspect of the eyeball consists ofthe transparent cornea (B11). Behind it liesthe lens of the eye (B12), overlaid by the iris(B13) with its central opening, the pupil. Theoptic nerve (B14) exits at the posteriorwall ofthe eyeball, slightly medial to the optic axis.There are three cavities in the eye:
! The anterior chamber (B15), which isbordered by cornea, iris, and lens
! The posterior chamber, which forms a ringaround the lens (B16)
! The interior of the eye, which containsthe vitreous body (B17)
The vitreous body is a transparent gel con-sisting mostly of water. The two eye cham-bers contain a clear fluid, the aqueoushumor.
The wall of the eyeball consists of three lay-ers:
! The fibrous tunic of the eyeball, or sclera! The vascular tunic of the eyeball, or uvea
! The internal (sensory) tunic of the eye-ball, or retina
The sclera (B18) is a thick, stretch-resistantconnective-tissue capsule that consistsmainly of collagenous fibers and some elas-tic fibers; in conjunction with the intraocu-lar pressure, it maintains the shape of theeyeball.
The vascular uvea forms the iris and the cili-ary body (B19) in the anterior part of the eye-ball, and the choroidea (B20) in the posteriorpart.
The posterior part of the retina (optic part)(B21) contains the light-sensitive receptorcells as well as pigmented epithelium, while
the anterior part (blind part) (B22) containsonly pigmented epithelium. The border be-tween the two retinal parts is known as theora serrata (B23).
The eyeball has an anterior pole (B24) and aposterior pole (B25); the equator of the eye-ball (B26) runs between them. Some bloodvessels and muscles follow themeridians ofthe eyeball (B27) which run from pole topole.
The eyeball can be divided into an anteriorpart and a posterior part, which fulfill differ-ent functions. The anterior part contains theimage-forming apparatus, the refractinglens system. The posterior part contains thephotoreceptive surface, the retina. Hence, theeye can be compared to a camera thatpossesses a lens system with an aperture inthe front—the iris of the eye—and a light-sensitive film at the back—the retina.
B28 Subarachnoidal cavity (p. 351, D9).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

343
TheEy
e
2
11
4
7
3
8
9
6
6
7
789
5
10
24
151319
11
16
27
12
18
20
14
28
25
21
26
17
2223
Development, Structure of the Eyeball
A Development of the eye
B Horizontal section through theeyeball
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

344Th
eEy
eThe Eye: Structure
Anterior Part of the Eye
Cornea (A, B)
The cornea (A1, B) is positioned like awatch-glass on the eyeball. Because of its markedcurvature, it has the effect of a focusing lens.Its anterior surface is formed by a multi-layered, nonkeratinizing squamous epi-thelium (B2) that rests upon a thick base-mentmembrane, called the anterior limitinglamina (Bowman’s membrane) (B3). Belowthis lies the stroma of the cornea (substantiapropria) (B4); its straight collagen fibersform lamellae that lie parallel to the surfaceof the cornea. At the posterior surface liesanother basement membrane, the posteriorlimiting lamina (Descemet’s membrane)(B5) and a single-layered endothelium (B6).The cornea contains unmyelinated nervefibers but no blood vessels. Its transparencyis due to a specific fluid content and thestate of swelling of its components. Anychange in the turgor causes turbidity of thecornea.
Anterior Chamber of the Eye (A)
The anterior chamber of the eye (A7) con-tains the aqueous humor generated by theblood vessels of the iris. The wall of theiridocorneal angle (A8) consists of looseconnective-tissue strands (reticulumtrabeculare, or pectinate ligament) (A9); theaqueous humor filters through the spacesbetween the fibers into the venous sinus ofthe sclera, or Schlemm’s canal (A10), andpasses into the bloodstream.
Iris (A, C)
The iris (A11) forms an aperture in front ofthe lens. It is attached to the ciliary body atthe root of the iris (A12) and extends to thepupillary margin (A13). The iris consists oftwo layers, namely, the mesodermal stroma(AC14) and the ectodermal posterior aspectof the iris, known as the iridial part of theretina (AC15). The stroma is made up of con-nective-tissue strands and is pigmented. Ahigh pigment content results in brown eyes,while a low pigment content gives the eyesa green or blue appearance. Numerous
blood vessels branch radially off the greaterarterial circle of the iris (AC16). The ectoder-mal part of the iris, a derivative of the opticcup (p. 342, A6), gives rise to two smoothmuscles, namely, the sphincter muscle of thepupil (AC17) and the thin dilator muscle ofthe pupil (AC18).
Ciliary Body (A, D)
The lens is suspended at the circular ciliarybody (A19, D). The muscles of the ciliarybody control the curvature of the lens(p. 358, B) and, hence, the visual acuity forclose and distant vision. It consists of aradially folded surface, the ciliary disk(D20), from where approximately 80 ciliaryprocesses (D21) protrude (ciliary crown). Theanterior part of the disk is occupied by theciliary muscle; its muscle fibers, the merid-ional fibers (A22), span between the ora ser-rata and the reticulum trabeculare as well asDescemet’s membrane. From here, radialmuscle fibers extend inward and bend totake a circular course (circular fibers) (A23).The posterior surface of the ciliary body iscovered by the ciliary part of the retina(A24). Delicate fibers, the zonular fibers(AD25), extend from the ciliary body to thelens and form the ciliary zone (D26). Manyfibers originate from the area of the ora ser-rata and extend to the anterior surface ofthe lens. They cross the shorter fibers aris-ing from the ciliary processes and ending atthe posterior surface of the lens.
Lens of the Eye (A, D, E)
The biconvex lens (AD27) is made up byelongated epithelial cells, the lens fibers,which show a lamellar arrangement. Theends of the fibers push against each other atthe anterior and posterior surfaces of thelens; in the newborn, they form a three-pointed star (E) at their border. The lensfibers are continuously being formedthroughout life.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

345
TheEy
e
20
2125
26
27
23
4
5
6
13 17
7
1
11
18 10
9
15
8
1927
25 12 16 23 22
24
14
14
17
18
16
15
Anterior Part of the Eye
A Anterior part of the eye
B Cornea
C Iris
D Ciliary body and lens, posterior viewE Fibers in the lens of a newborn
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

346Th
eEy
eThe Eye: Structure
Vascular Supply (A)
The eye has two different vascular systems:the ciliary arteries and the central retinalartery. All these vessels arise from the oph-thalmic artery (p. 273, C16). The posteriorciliary arteries are the branches supplyingthe vascular tunic of the eyeball, or uvea,which forms the iris (A1), the ciliary body(A2), and the choroidea proper of the poste-rior wall of the eyeball (A3). This vascularsystem not only supplies the blood, it is alsoessential for maintaining the intraocularpressure and the tension of the eyeball.
Long posterior ciliary arteries (A4). Thesearteries penetrate the sclera near the exit ofthe optic nerve. One of them runs in thetemporal, the other one in the nasal wall ofthe eyeball to the ciliary body and to the iris.At the root of the iris they form the greaterarterial circle of the iris (A5), from wherevessels radiate to the lesser arterial circle ofthe iris (A6) near the pupil.
Short posterior ciliary arteries (A7). Theseform the vascular plexus of the choroidea,which extends from the posterior wall ofthe eyeball to the ora serrata (A8). The innerchoroid layer consists of especially widecapillaries, the choriocapillary layer, andborders on the pigmented epithelium of theretina. Whereas the choriocapillary laminais firmly attached to the pigmentedepithelium, the outer aspect of thechoroidea is separated from the sclera bythe perichoroidal space and, thus, can be dis-placed.
Anterior ciliary arteries (A9). These runfrom the rectus muscles to the sclera wherethey branch in the episcleral tissue and inthe conjunctiva. In the conjunctiva theyform the marginal loops (A10) around themargin of the cornea.
The veins unite to form the four posterior cil-iary veins, or vorticose veins (A11), whichobliquely penetrate the sclera at the poste-rior wall of the eyeball.
Central retinal artery (A12). This artery en-ters the optic nerve approximately 1 cm be-hind the eyeball and extends in the middle
of the nerve to the papilla of the optic nerve(see below). It then divides into brancheswhich run along the inner surface of the ret-ina within the layer of nerve fibers. The reti-nal vessels are end arteries. Their capillariesreach as far as the inner nuclear layer (seep. 349, A12). The venules unite to form thecentral vein of the retina (A13), which takes acourse similar to that of the central artery.
The visual cells are nourished from bothsides of the retina: from the outside by thecapillary system of the short posterior cili-ary arteries, and from the inside by the cen-tral arteries.
Fundus of the Eye (B)
The posterior pole of the eyeball, the fun-dus, can be examined through the pupilwith an ophthalmoscope. It is reddish incolor. In the nasal half lies the papilla of theoptic nerve (blind spot) (B14), where all nervefibers of the retina combine to leave the eyeas the optic nerve. The papilla is a whitishdisk with a central shallow depression, theexcavation of the optic disk (AB15) (p. 351,D). In the papilla, the central artery dividesinto several branches, and the veins unite toform the central vein. The arteries are rela-tively light in color and are thin, while theveins are darker and slightly thicker. Thevessels run radially in the nasal direction,while they arch in the temporal direction.Numerous vessels run to the macula (yellowspot) (B16), the area of highest visual acuity.Its traversely oval, slightly yellowish surfacecontains a small depression in the center,the central fovea (AB17).
A18Optical part of the retina.A19 Ciliary part of the retina.A20 Iridial part of the retina.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

347
TheEy
e
16
17
1514
9
10
1
6
20
2
19
8 18 3
11
4
7
13125
Vascular Supply, Fundus of the Eye
A Vascular supply of the eye
B Fundus of the eye
15
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

348Th
eEy
eThe Eye: Structure
Retina (A, B)
The retina consists of two layers, namely, theouter pigmented layer and the inner neurallayer (see p. 342, A8, A9). The two layersfirmly adhere to each other only in the re-gions of papilla and ora serrata.
Neural Layer
The neural layer contains three layers ofcells. Adjacent to the pigmented epithelium(AB1) lies the neuroepithelial layer of the ret-ina, the layer of photoreceptors. Next fol-lows the ganglionic layer of the retina, a layerof bipolar neurons. Last comes the gan-glionic layer of the optic nerve, a layer ofmul-tipolar neurons; their axons form the opticnerve. Thus, the sensory cells of the retinado not face the incoming light with their re-ceptor parts; rather, they turn away fromthe light and are covered by neurons andnerve fibers. This reverse order is known asinversion of the retina. The inner surface ofthe retina is separated from the vitreousbody by a basement membrane, the internallimiting membrane (B2). A glial membrane,the external limiting membrane (B3), sepa-rates the receptor parts of the sensory cellsfrom the rest of the neural epithelium. Be-tween both membranes expand elongatedsupporting glial cells of Müller (B4) withtheir leaf-like processes.
Neuroepithelial layer (AB5). The neuralepithelium contains two different types ofphotoreceptor cells, the retinal rods (B6) andcones (B7). The rod cells are for light–darkperception in dim light (night vision), whilethe cone cells are responsible for color per-ception in bright light (color vision) andvisual acuity (duplicity theory). The cell nu-clei of the photoreceptors form the externalnuclear layer (A8). The rods and cones reston the pigmented epithelium and do notfunction without contacting with the pig-mented epithelium. The photoreceptorsrepresent the 1st neuron of the visual path-way.
In lower vertebrates, the pigmentedepithelium has a brush border of microvilli(B9) that reach between the rods and cones.
When exposed to light, the pigment movesinto the microvilli and surrounds the recep-tor parts of the sensory cells. The pigmentreturns to the cell body in darkness. In somespecies, the sensory cells can elongate andpress against the pigmented epitheliumwhen exposed to light. Such retinomotoricmovement has not been clearly demon-strated in mammals.
Ganglionic layer of the retina (AB10). Thislayer consists of bipolar neurons (B11) (2ndneuron). Their dendrites extend to thesensory cells and contact the synaptic ter-minals of the photoreceptors. Their axonshave contact with the large neurons of theganglionic layer of the optic nerve. The cellnuclei of the bipolar neurons form the innernuclear layer (AB12). Their synapses lie inthe outer plexiform layer (AB13) and innerplexiform layer (AB14).
Ganglionic layer of the optic nerve (AB15).The uppermost cell layer consists of a row oflarge multipolar neurons (B16) (3rd neuron);their short dendrites form synapses in theinner plexiform layer with the axons of thebipolar neurons. Their axons extend as un-myelinated fibers in the layer of nerve fibers(AB17) to the papilla of the optic nerve(blind spot). They form the optic nerve andterminate in the lateral geniculate body (p.256).
Association cells. There are transverse con-nections in the individual layers. In theouter plexiform layer they are formed bythe axons of the horizontal cells (B18) and inthe inner plexiform layer by the dendrites ofthe amacrine cells (B19).
A20 Choriocapillary layer.A21Direction of incoming light.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
349
TheEy
e
15
10
5
15
2
17
21
14
12
13
8
10
5
1
17
14
13
120
8
12
19
16
18
11
4
7
9
6
3
Retina
A Retina, cellular staining
B Structure of the retina, diagram(modified from Schaffer)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

350Th
eEy
eThe Eye: Structure
Retina (continued) (A–C)
Regional Structures of the Retina
The retina is divided into three parts:
! The optic part of the retina (p. 347, A18)covers the fundus of the eye and containsthe light-sensitive sensory cells (photore-ceptors).
! The ciliary part of the retina (p. 344, A24;p. 347, A19) lies below the ciliary bodyand is separated from the optic part bythe ora serrata (p. 343, B23).
! The iridial part of the retina (p. 344, A15;p. 347, A20) covers the posterior surfaceof the iris.
Neither the ciliary part nor the iridial partcontain sensory cells; they consist only of adouble-layered pigmented epithelium.They are therefore collectively known as theblind part of retina.
The human retina contains approximately120 million rod cells and 6 million conecells. Their distributions vary regionally. Therod-to-cone ratio in the vicinity of the cen-tral fovea is 2:1. The portion of conesdecreases toward the periphery, and thelateral region of the retina is dominated byrods.
Macula and central fovea. The macula luteadoes not contain any rods but only cones,and the unusually long and narrow shape ofits cones (AB1) differs from the usual struc-ture of cones. The ganglionic layer of the ret-ina and the ganglionic layer of the opticnerve are relatively thick in the macula, butthey diminish in the region of the centralfovea, leaving only a thin tissue layer on topof the cone cells. Thus, the incoming lighthas immediate access to the photoreceptorsin the fovea. Macula and fovea are the areasof highest visual acuity. This is not only dueto the absence of the upper retinal layersbut also to special neuronal circuits.
Each cone in the central fovea is connectedwith a single bipolar neuron (B2). Eachbipolar neuron in the region around thefovea (perifovea) synapses with six conecells. Such convergence (p. 34) is even morepronounced at the periphery of the retina
(C). Here a single optic nerve neuron mayreceive input from over 500 receptors. Atotal of only onemillion of optic nerve fiberscorrespond to 130 million photoreceptors.
A circumscribed fiber bundle, the papil-lomacular bundle, extends from the neuronsof the macula to the papilla.
Neuronal Circuits (B, C)
The retina is an extremely complex relaysystem. The eye is not just a sensory organtransmitting light impulses. Rather, theseimpulses already undergo processing in theretina. Electrophysiological studies haveshown that groups of sensory cells are com-bined to receptive fields and react asfunctional units. A receptive field is formedby the dendritic tree of a large optic nerveganglion cell and the dendrites of theamacrine interneurons in the inner plexi-form layer. Stimulation of a receptive fieldinhibits neurons in the vicinity. This inhibi-tion is made possible by the transverse con-nections of the horizontal interneurons inthe outer plexiform layer.
Optic Nerve (D)
The nerve fibers of the retina extend inbundles to the papilla of the optic nerve (D3)where they unite to form the optic nervebefore exiting the eyeball. Sclera andchoroidea are very thin at the site of pene-tration, and the sclera is perforated (laminacribrosa) (D4). Once the extremely delicatenerve fibers have passed the sclera, they be-come enveloped by myelin sheaths. Theoptic nerve is actually a fiber tract of theCNS and contains astrocytes and oligoden-drocytes; hence, its nerve fibers do not haveSchwann cell sheaths. As part of the brain,the optic nerve is surrounded by mengines.The dural sheath (D5) and the arachnoidsheath (D6) merge with the sclera (D7). Be-tween arachnoid sheath and pial sheath (D8)lies a CSF-filled space (D9) which makes ashift between nerve and sheath possible. Anumber of septae from the pia mater extendbetween the nerve bundles.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

351
TheEy
e
98
3
4
765
1
1
2
10
Retina, Optic Nerve
A Central fovea
B, C Neuronal circuitsin the retina(according to Polyak)
B Cone system ofthe central fovea
C Mixed rod andcone system
D Papilla of theoptic nerve
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

352Th
eEy
eThe Eye: Structure
Photoreceptors (A–D)
The light-sensitive sensory cells have thesame structural design in all vertebrates.Next to the pigmented epithelium lies theouter segment of the photoreceptor cell; it ispartly buried in a pigmented epithelial cell.The outer segment of rod cells is a cylinder(ABC1) containing several hundreds ofstacked, disk-shaped membrane pouches ofuniform size. The outer segment of conecells (B2, D) is of a conical shape, and theproximal membrane folds are larger thanthe distal ones. A thin, eccentric cyto-plasmic bridge, the connecting cilium(ACD3), links the outer segment to the innersegment (AB4). The bridge contains a mod-ified ciliumwith 9 pairs of microtubules butwithout the central pair characteristic ofother cilia (see p. 285, D5, D6). The connect-ing cilium is relatively long in some species,leaving a distinct space between the twosegments (A). In humans, however, it is soshort that both segments touch each otherwithout leaving a visible space betweenthem (B). The inner segment containsnumerous Golgi stacks, ribosomes, andlongitudinally arranged mitochondria. Thecell body then tapers to form an axonlikeprocess (AB5) containing neurofilamentsand microtubules. The cell nucleus (AB6)lies either at the transition from the innersegment to the axon or within the axon. Thecell terminates with an end bulb, the synap-tic terminal (A7). In addition to the usualsynapses, the terminal develops invaginatedsynapses (A8) in which the presynapticmembrane becomes invaginated and sur-rounds the postsynaptic complex on allsides.
The outer segment is the actual receptorpart of the cell where the light is absorbed.The stacked membranes are formed by in-foldings of the plasma membrane (C9) inthe proximal part of the outer segment. Inthe rod cells, they detach from the outermembrane and form isolated disks in thedistal part of the segment (C10). The visualpigment of the rod cells, rhodopsin, is boundto the membrane of the disks. The forma-tion of visual pigment in the inner segment
and its migration through the connectingbridge into the outer segment can be fol-lowed using autoradiography by labelingthe protein component of rhodopsin with aradioactive amino acid. Once the labeledsubstance has passed the bridge, it forms aband that migrates to the outer end andthen disappears (in rats within 10 days). Themigrating band represents a membranedisk that has incorporated labeled rhodop-sin. Thus, new disks are being continuouslyformed in the rod cells, migrate to the distalend, and are shed there. Fragments of theshed disks have been found in the pig-mented epithelial cells. There is no new for-mation of disks in the outer segments ofcone cells (D). The infoldings of the mem-brane are permanent and, in contrast to rodcells, there is no detachment of membraneinvaginations from the plasma membrane.
Only rod cells contain rhodopsin. The ab-sorption of light changes the molecularstructure of rhodopsin, causing it to breakdown into its protein and pigment com-ponents. From these components, rhodop-sin is continuously resynthesized in therods (rhodopsin–retinin cycle). It absorbslight of all wavelengths and, thus, is not in-volved in color vision. Rod cells are light–dark receptors. The three different types ofcone cells each contain a different pigmentthat absorbs only light of a specificwavelength. Cone cells are color receptors.
There are some animal species in which theretina contains only cones, while it containsonly rods in other species (cat, cattle). Ani-mals with a retina containing only rods can-not distinguish between colors. The bull,which is said to react to red, is actually colorblind.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

353
TheEy
e
8
9
6
5
4
3
1
7
4
6
6
5
12
10
3 3
Photoreceptors
A Rod cell, electron-microscopic diagram
B Rod cell (left)and cone cell(right) of thehuman retina
C Outer segmentof a rod cell
D Outer segmentof a cone cell
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

354Th
eEy
eThe Eye
Visual Pathway and OcularReflexes
Visual Pathway (A, B)
The visual pathway consists of four neuronsconnected in tandem:
! 1st neuron, the photoreceptors! 2nd neuron, the bipolar neurons of the ret-
ina, which transmit the impulses fromthe rods and cones to the large ganglioncells of the retina
! 3rd neuron, the large ganglion cells, theaxons of which combine to form the opticnerve and extend to the primary visualcenters (lateral geniculate nucleus)
! 4th neuron, the geniculate cells, the axonsof which project as the optic radiation tothe visual cortex (striate area)
The optic nerve (A1) enters the cranial cavitythrough the optic canal. At the base of thediencephalon, together with the con-tralateral optic nerve, it forms the optic chi-asm (A2). The fiber bundle starting from thechiasm is known as the optic tract (A3). Thetwo tracts run around the cerebralpeduncles to the two lateral geniculate bod-ies (A4). Before reaching these, each tractdivides into a lateral root (A5) and a medialroot (A6). Whereas most of the fibers runthrough the lateral root to the lateralgeniculate body, the medial fibers continuebelow the medial geniculate body (A7) tothe superior colliculi. They contain visualreflex pathways (p. 358, A16). The opticnerve fibers are thought to give off collater-als to the pulvinar of the thalamus (A8)prior to terminating in the lateral geniculatebody. The optic radiation (radiation of Gra-tiolet) (B9) begins at the lateral geniculatebody and extends as a broad fiber plate tothe calcarine sulcus at the medial aspect ofthe occipital lobe and, while doing so, formsthe outward-arching temporal genu (B10)(p. 258, C16). Numerous fibers bend ros-trally (occipital genu) (B11) in the occipitallobe to reach the anterior areas of the visualcortex.
The optic nerve fibers originating from thenasal halves (B12) of the retina cross in the
optic chiasm. The fibers from the temporalhalves (B13) do not cross but continue onthe ipsilateral side. Hence, the right tractcontains the fibers from the temporal half ofthe right eye and from the nasal half of theleft eye. The left tract contains fibers fromthe temporal half of the left eye and fromthe nasal half of the right eye. In a cross sec-tion of the tract, the crossed fibers liemostlyventrolaterally, and the uncrossed fibersdorsomedially; in between, the fibers aremixed.
The crossed and uncrossed fibers of the optic tractextend to different cell layers of the lateralgeniculate body (p. 256, B). The number ofgeniculate cells, approximately one million,corresponds to the number of optic nerve fibers.However, the optic nerve fibers usually end onfive to six cells located in different cell layers. Cor-ticofugal fibers of the occipital cortex also end inthe lateral geniculate body. They probably controlthe input of impulses, as suggested by the pres-ence of axo-axonal synapses characteristic forpresynaptic inhibition.
The axons of the geniculate cells form the opticradiation. Its fibers are arranged according to thedifferent regions of the retina (p. 356). The fibersfor the lower half of the retina, especially thosefor the periphery of the retina, arch most rostrallyin the temporal genu. The fibers for the upper halfof the retina and for the central region of the mac-ula arch only slightly in the temporal lobe.
In the striate area (B14) of the right hemi-sphere terminate the fibers for the righthalves of the retinae; hence, it receivessensory input from the left halves of thevisual fields. In the striate area of the lefthemisphere there terminate the fibers ofthe left halves of the retinaewith input fromthe right halves of the visual fields. The righthand and the right visual field are thereforeboth represented in the left hemisphere,which dominates in right-handed persons(p. 262).
B15 Visual fields.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

355
TheEy
e
1
2
3
5
4
7
13
10
6 8
9
11
14
12
15
13
Visual Pathway
A Optical nerve and optical tract
B Arrangement of optic nerve fibers(according to Polyak)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

356Th
eEy
eThe Eye: Visual Pathway and Ocular Reflexes
Topographic Organization of theVisual Pathway (A)
The fibers from individual regions of the ret-ina occupy specific positions in the differentparts of the optic system. To illustrate this ina simple way, the retina is subdivided intofour quadrants that all share the center,namely, the macula with the central fovea(area of highest visual acuity). The fibers ofthe fovea show a regular point-to-pointconnection between fovea, lateral genicu-late body, and striate area.
The halves of the visual fields of each eye(visual hemifields) (A1) are projected ontothe respective contralateral halves of the re-spective retina (hemiretinas) (A2). Immedi-ately after the exit of the optic nerve from theeyeball (A3), the macular fibers lie on thelateral side of the nerve, with the fibers fromthe nasal half of themacula lying in the cen-ter, surrounded by fibers of the temporalhalf of the macula. Further along the opticnerve, the macular bundle comes to occupythe center (A4).
The fibers of the nasal hemiretinas (con-tinuous lines) cross to the opposite side inthe optic chiasm (A5). While doing so theytake a strange course. The medial fiberscross, then run a short distance into the con-tralateral optic nerve, and finally turn in aright angle into the contralateral optic tract.The lateral fibers run a short distance intothe ipsilateral optic tract and then turnabruptly into the contralateral tract. Thefibers of the temporal hemiretinas (brokenlines) do not cross but continue in the ipsi-lateral tract.
The optic tract (A6) thus contains the fibersof the corresponding halves of both retinas:the left tract contains the fibers of the lefthemiretinas, the right tract contains thefibers of the right hemiretinas. The fibers ofthe two superior retinal quadrants lie ven-tromedially, those of the two inferior quad-rants lie dorsolaterally, while the fibers ofthe macula take a central position. Prior toradiating into the lateral geniculate body(A7), the fibers rearrange so that the macu-lar fibers form a central wedge, the fibers
from the upper retinal quadrants come to liemedially, and the fibers from the lower reti-nal quadrants lie laterally.
The fibers in the lateral geniculate body (A8)terminate in the same arrangement. Thecentral wedge of the terminal macularfibers makes up almost half of the genicu-late body. The fibers from the periphery ofthe retina terminate in the most anteriorand ventral regions of the lateral geniculatebody. The terminals of the ipsilateral andcontralateral fibers in the geniculate layersare shown schematically in light gray anddark gray (A9) (see also p. 257, A). Thegeniculate cells of the central wedge projectto the posterior region of the striate area(A10). The area of highest acuity, which inthe human retina measures slightly morethan 2mm in diameter, is represented bythe largest portion of the visual cortex. Ros-trally to it lie the much smaller areas for theremaining parts of the retina. The upperquadrants of each retina are represented inthe upper lip of the calcarine sulcus, and thelower quadrants in the lower lip.
Clinical Note: Corresponding to the arrange-ment of fibers, injury to the visual pathway inspecific segments results in various patterns ofloss of vision. It should be taken into considera-tion that the lower halves of each retina registerthe input from the upper halves of the visualfields, while the upper halves of each retina regis-ter the input from the lower halves of the visualfields. The same is true for the left and righthalves of each retina. Injury to the optic tract,lateral geniculate body, or visual cortex on the leftside affects the left halves of each retina and theright halves of each visual field. The result is ho-monymous hemianopia on the right side. In caseof bitemporal heteronymous hemianopia, injuryto the crossing nasal fibers of both retinae (e.g., incase of tumors of the hypophysis near the opticchiasm) results in bilateral loss of the temporalhalves of the visual fields. Damage of both visualcortices causes visual agnosia.
A11 Blind spot (papilla of the optic nerve).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

357
TheEy
e
1
2
11
3
4
5
6
7
8 9
3
4
5
6
7
810
4
3
2
1
Topographic Organization of the Visual Pathway
A Position of retinal quadrants in the visualpathway (modified from Polyak)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

358Th
eEy
eThe Eye: Visual Pathway and Ocular Reflexes
Ocular Reflexes (A–C)
During the visual process the eye must con-tinuously compensate for changes fromlight to dark and from near to far. Both theaperture and lens system must continu-ously adapt to the prevailing conditions.Whereas light–dark adaptation is achieved bydilatation or contraction of the pupil, near–far adaptation requires a change in the curva-ture of the lens (accommodation), a changein the lines of sight (convergence), and achange in pupillary width. During negativeaccommodation (adjustment for long dis-tances), the surface of the lens is onlyslightly curved, the lines of sight run paral-lel, and the pupils are dilated. During posi-tive accommodation (adjustment for shortdistances), the surface of the lens is dis-tinctly curved, the lines of sight cross at adistance that corresponds to the fixated ob-ject, and the pupils are constricted.
Pupillary Light Reflex (A)
When light falls onto the retina, the pupilconstricts. The afferent loop of this reflexarch is formed by optic nerve fibers (A1) ex-tending to the pretectal nucleus (A2). Thelatter is connected with the rostral part ofthe Edinger–Westphal nucleus (accessoryoculomotor nucleus) (A3), the fibers ofwhich (A4) extend as the efferent limb of thereflex arch to the ciliary ganglion (A5)(p. 128). The postganglionic fibers (A6) in-nervate the sphincter muscle of the pupil(A7). Both pretectal nuclei are connected viathe epithalamic commissure (A8). In addi-tion, the optic nerve fibers of each side ter-minate in the two pretectal nuclei. This ex-plains the bilaterality of the light reflex;when light falls only onto one eye, the pupilof the other eye constricts as well (consen-sual pupillary response). Ciliospinal center(A9); sympathetic fibers (A10) for the di-lator of the pupil (A11).
Accommodation (B)
The accommodation apparatus consists ofthe lens, its suspension mechanism (ciliaryzone), the ciliary body, and the choroidea.These parts form a tense, elastic system
which spans the entire eyeball and main-tains the flat, slightly curved form of thelens (B12) (negative accommodation). Duringpositive accommodation the circular ciliarymuscle (B13) contracts. The meridionalmuscle fibers pull the origins of the longzonular fibers forward, and the circularfibers move the ciliary processes closer tothe margin of the lens. This relaxes both thezonular fibers (B14), and the lens capsule,thus causing the lens to round off (B15).
The fiber tracts of the accommodation reflexare less well known. As fixation of an objectis the prerequisite of accommodation, theoptic nerve is the afferent loop. The reflexarch runs propably via the visual cortex(striate area) to the pretectal nuclei,possibly also via the superior colliculi (A16).The efferent loop begins in the caudal part ofthe Edinger–Westphal nucleus. Its fiberssynapse in the ciliary ganglion with post-ganglionic fibers that innervate the ciliarymuscle.
Convergence (A, C)
When an object approaching from a far dis-tance is fixated with both eyes, the medialrectus muscles (C17) increasingly adductboth eyeballs, and the lines of sight whichinitially run parallel (broken arrows) beginto cross each other. The fixated object re-mains in the intersection of the lines of sightwhile being projected onto each macula.
The visual fixation reflex probably runs via thevisual pathway to the occipital cortex andvia the corticofugal fibers (A18) to the supe-rior colliculi, to the pretectal region, and tothe nuclei of the ocular muscles (A19). Theoccipital cortex is therefore regarded as areflex center (occipital eye fields).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

359
TheEy
e
17
18
8
162
19
1915
14
12
4
5
610
3
11 7
9
1
13
17
18
8
162
19
1915
14
12
4
5
610
3
11 7
9
1
13
Ocular Reflexes
A Pupillary light reflex
B Accommodation(modified from Rohen)
C Convergence
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

The Ear
Structure 362Auditory Pathway and Vestibular
Pathways 378
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

362Th
eEa
rThe Ear
Structure
Overview (A, D)
The ear contains two sensory organs withdifferent functions; morphologically theyform a single complex, the inner ear, or ves-tibulocochlear organ. One part of it, thecochlea, is the organ of hearing, or spiralorgan (organ of Corti). The other part con-sists of saccule, utricle, and semicircularducts; it registers changes in body position,especially those of the head, and representsthe organ of balance, or vestibular appara-tus. The ear is divided into three parts: theexternal ear, the middle ear (p. 364f), andthe internal ear (p. 368ff).
The external ear includes the auricle (A, D1)and the external acoustic meatus (D2).
The middle ear consists of the tympanic cav-ity (D3), the mastoid cells (air cells) (p. 364,A6), and the auditory tube (eustachian tube)(D4). The tympanic cavity with the auditoryossicles is a narrow space filled with air. Itlies behind the eardrum and extends as theepitympanic recess (D5) above the externalacoustic meatus. The tympanic cavitymerges anteriorly into the auditory tube(tympanic opening) (D6). The tube extendsobliquely downward and forward and opensin front of the posterior pharyngeal wallinto the pharyngeal cavity (pharyngealopening) (D7). The auditory tube is linedwith a ciliated epithelium and consists of anosseous and a cartilaginous section that joinat the isthmus of the tube (D8). The tubal car-tilage (D24) leaves open a cleft that is linedby connective tissue (membranous lamina).The tendon of the tensor tympani muscle(D9) attaches to the base of the manubriumof the malleus (p. 365, A25). The tympaniccavity communicates with the pharyngealcavity through the auditory tube, thus per-mitting exchange of air and the equilibra-tion of pressure in themiddle ear. The open-ing of the auditory tube, however, is nor-mally closed and opens only when thepharyngeal muscles contract (swallowing).
The internal ear consists of the bony laby-rinth (D10), which contains the mem-branous labyrinth and the internal acousticmeatus.
Outer Ear (A–C)
The auricle, or pinna (A, D1), with the ex-ception of the lobule, contains a scaffold ofelastic cartilage. The shapes of the auricularprojections and depressions are different ineach person and are genetically determined.The shapes of the following parts are in-herited: helix (A11), antihelix (A12), scapha(A13), concha (A14), tragus (A15), antitragus(A16), and triangular fossa (A17). In the past,the features of the auricle were of specialimportance for establishing paternity.
The entrance of the outer ear canal, or ex-ternal acoustic meatus (D2), is formed by agroovelike continuation of the auricular car-tilage (D25) and is completed with connec-tive tissue to form a uniform passage. Thepassage is lined with epidermis, and largeceruminous glands lie beneath the epider-mis.
The outer ear canal ends with the eardrum,or tympanic membrane (B, D18), which isobliquely placed in the meatus. Whenviewed from the outside, the mallear stria(B19) can be recognized; it is caused by theattachment of the manubrium of the mal-leus, which reaches to the umbo of the tym-panic membrane (B20), the innermost pointof the funnel-shaped eardrum. Above theupper end of the mallear stria (mallearprominence) lies a lax, thin part of the ear-drum, the reddish pars flaccida (B21), whichis separated from the firm, gray and shinypars tensa (B22) by two mallear folds. Theeardrum is covered externally by skin andinternally by a mucosa. Between these liesthe lamina propria of the pars tensa; it con-tains radial and nonradial fibers (C). The lat-ter are circular, parabolic, and transversal.The fibrocartilaginous annulus (C23) formsthe anchoring tissue of the eardrum.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

363
TheEa
r
22
21
1113
17
1514
12
16
23
19
20
1
5 3
10
92
25
18 6 8 4
7
24
Overview, Outer Ear
A Auricula (external ear)
B Right eardrum(preparation by Platzer)
C Course of fibers in the lamina propria ofthe eardrum (according to Kirikae)
D View of the middle ear and internal ear
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

364Th
eEa
rThe Ear: Structure
Middle Ear
Tympanic Cavity (A, B, D)
The tympanic cavity is a narrow, verticalspace with the eardrum (AD1) obliquelyplaced in its lateral wall. Its medial wall hastwo openings leading to the internal ear,namely, the oval window, or vestibular win-dow (D2), and the round window, or cochlearwindow (D3). The roof of the tympanic cav-ity, the tegmental wall, is relatively thin andborders on the surface of the petrous pyra-mid. The floor of the tympanic cavity isformed by a thin layer of bone; beneath itruns the jugular vein.
The tympanic cavity continues in anteriordirection as the auditory tube (A4) (p. 363,D4). Its superior part opens posteriorly intothe mastoid antrum (A5), a round space intowhich numerous small cavities open, themastoid air cells (A6). These air-containingcavities are lined with mucosa and form asystem of chambers that penetrates the en-tire mastoid bone; they may even extendinto the petrous bone.
Auditory Ossicles (A, C, D)
The three auditory ossicles, or ear bones,form together with the eardrum the sound-conducting apparatus. They are called thehammer, or malleus (CD7), the anvil, or incus(CD8), and the stirrup, or stapes (CD9). Themanubrium of the malleus (ACD10) is firmlyattached to the eardrum and connected viaits neck (C11) to the head of the malleus(C12). The malleus has a saddle-shaped ar-ticular surface that contacts the body of theincus (C13). Its lenticular process (AC14),which is attached to the long limb of theincus and projects at a right angle, carriesthe articular surface for the head of thestapes (C15). The footplate of the stapescloses the vestibular window; it is attachedat themargin by the annular ligament of thestapes (D16). Several ligaments (A17) con-nected to the wall of the tympanic cavitykeep the ossicles in place.
The auditory ossicles transmit to the innerear the vibration of the eardrum caused bysound waves. For this purpose, malleus and
incus act like an angular lever, and thestapes performs a tilting movement. Thefootplate of the stapes transmits the vibra-tion to the fluid of the inner ear. The move-ment of the fluid through the cochlea issimplified in diagram (D); in reality, it takesa spiral course inside the cochlea (p. 369, A;p. 371, C). Tension in the system is controlledby two muscles with antagonistic effects,namely, the tensor tympani muscle (A18)(p. 363, D9) and the stapedius muscle (A19)(p. 367, C22).
The mucosa lining the tympanic cavity andcovering the auditory ossicles forms variousfolds, such as the anterior (A20) and poste-rior mallear folds (A21) which envelope thechorda tympani (A22). The folds formseveral mucosal pouches. The superiorrecess of the tympanic membrane, Prussak’spouch (D23), lies between the pars flaccidaof the eardrum and the neck of the malleus;it plays an important role in ear infections.
A24 Facial nerve.A25 Tendon of the tensor tympani muscle.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

365
TheEa
r
12
13
11
14
15 9
8
7
10
10
1
16
2
238
9
3
7
17
22
5 6
17
21
20
25
1
4
18
14
24
1019
Middle Ear
A Tympanic cavity, view of theinner aspect of the eardrum(preparation by Platzer)
B Position of tympanic cavity andauditory tube in the skull
C Auditory ossicles
D Function of theauditory ossicles
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

366Th
eEa
rThe Ear: Structure
Middle Ear (continued)
Medial Wall of the Tympanic Cavity (A–C)
The medial wall, or labyrinthic wall, sepa-rates the tympanic cavity from the internalear. The prominence in its middle region,the promontory of the tympanic cavity (A1), iscaused by the basal convolution of thecochlea. In a bifurcated groove, the sulcus ofthe promontory (A2), lies the tympanicplexus (C3); it is formed by the tympanicnerve (C4) of the glossopharyngeal nerveand by the sympathetic fibers of the carotidplexus of the internal carotid artery. Thepromontory is anteriorly delimited by thetympanic cells (A5). In the medial wall, theoval window, vestibular window (A6 ), and theround window, cochlear window (A7), openinto the inner ear. The stapes (C8) rests inthe vestibular window and closes it with itsfoot plate. The cochlear window is closed bythe secondary tympanic membrane. In theposterior wall opening to the mastoid an-trum (A9) run two arched canals, the facialcanal (A10) and the lateral semicircularcanal (A11); both canals cause protrusionson the wall of the tympanic cavity, namely,the prominence of the facial canal and theprominence of the lateral semicircular canal.A bony protrusion, the pyramidal eminence(A12), contains an opening at its tip throughwhich the tendon of the stapedius muscle(C13) enters. In anterior direction, the tym-panic cavity leads into the semicanal of theauditory tube (A14). Above it lies the semi-canal of the tensor tympani muscle (A15).The two semicanals are incompletely sepa-rated by a bony septum and together formthe musculotubal canal. The medial wall atthe level of the opening of the tympanictube (carotid wall) separates the tympaniccavity from the carotid canal (A16); the bonyfloor (jugular wall) separates it from thejugular fossa (A17). Also shown are the jugu-lar vein (B18 ) and the internal carotid artery(B19).
Clinical Note: The osseous roof and floor ofthe tympanic cavity may be very thin. In case ofpurulent otitis media, the infection can penetrateeither through the roof and progress to meningesand brain (meningitis, cerebral abscess in the
temporal lobe), or through the floor and progressto the internal jugular vein (jugular thrombosis).
Muscles of the Tympanic Cavity (C)
The tensor tympani muscle (C20) origi-nates from the cartilaginous wall of thetympanic tube and from the osseous wall ofthe canal. Its narrow tendon bends awayfrom the cochleariform process (C21) and in-serts on the manubrium of malleus. Themuscle is innervated by the tensor tympaninerve of the mandibular nerve. The stape-dius muscle (C22) originates in a smallbony canal which usually communicateswith the facial canal. Its small tendon passesthrough the opening of the pyramidal emi-nence and inserts at the head of the stapes.The muscle is innervated by the stapediusnerve of the facial nerve (C23).
The two muscles control the tension of thesound-conducting apparatus. The tensor tym-pani muscle pulls the eardrum inward andpushes the footplate of the stapes into thevestibular window, thus increasing the sen-sitivity of the transmission. The stapediusmuscle levers the footplate of the stapes outof the vestibular window and thus dampensthe transmission. Hence, the two musclesare antagonists.
Clinical Note: Paralysis of the facial nervecauses loss of function in the stapedius muscleand deficient dampening of sounds; the patientssuffer from hyperacusis, an increased sensitivityto sound.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
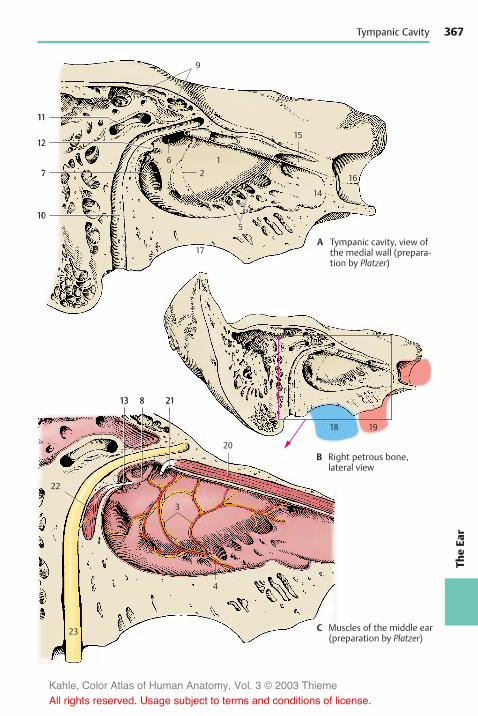
367
TheEa
r
11
12
7
10
9
6 12
17
18 19
5
15
16
14
22
13 8 21
3
20
4
23
11
12
7
10
9
6 12
17
18 19
5
15
16
14
22
13 8 21
3
20
4
23
Tympanic Cavity
A Tympanic cavity, view ofthe medial wall (prepara-tion by Platzer)
B Right petrous bone,lateral view
C Muscles of the middle ear(preparation by Platzer)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

368Th
eEa
rThe Ear: Structure
Inner Ear (A–D)
The membranous labyrinth is a system ofvesicles and canals that is surrounded on allsides by a hard, bony capsule. The cavities inthe bone have the same shapes as the mem-branous structures, and their cast (C) pro-vides a crude representation of the mem-branous labyrinth. We therefore distinguishbetween an osseous (bony) labyrinth and amembranous labyrinth. The osseous laby-rinth contains a clear, aqueous fluid, theperilymph (light greenish-blue), in which themembranous labyrinth is suspended. Theperilymphatic space communicates withthe subarachnoid space via the perilym-phatic duct (A1) at the posterior edge of thepetrous bone. The membranous labyrinthcontains the endolymph (dark greenish-blue) which is a viscous fluid.
The vestibular window (AC2) is closed by thestapes and leads into the middle part of theosseous labyrinth, the vestibule of the ear(AC3). The vestibule communicates anteri-orly with the bony cochlea (C4) and at itsposterior wall with the bony semicircularcanals (C5).
The vestibule contains two membranousparts, the saccule (AB6) and the utricle (AB7).Both structures contain sensory epitheliumin a circumscribed part of the wall (blue),themacula of the saccule (AB8) and themac-ula of the utricle (AB9), and are intercon-nected by the utriculosaccular duct (AB10).The latter gives off the slender endolym-phatic duct (A11) which runs to the poste-rior surface of the petrous bone and endsbeneath the dura mater as a flattened ves-icle, the endolymphatic sac (A12). The unit-ing duct (AB13) forms a connection betweenthe saccule and the membranous cochlearduct.
The osseous cochlea (C4) has about twoand a half turns. The spiral canal of thecochlea (C14) contains the membranouscochlear duct (AB15) which starts with ablind end, the vestibular cecum (B16), andends in the tip of the cochlea, or cupula(C17), with the cupular cecum (B18). Theperilymphatic spaces are above and below
the cochlear duct, or scala media; the scalavestibuli (AB19) lies above it and opens intothe vestibule, and the scala tympani (AB20)lies beneath it and is closed by the cochlearwindow (A–C21).
The three bony semicircular canals (C5)emanating from the vestibule contain themembranous semicircular ducts (A22),which are connected to the utricle. They aresurrounded by perilymph and attached tothe walls of the perilymphatic space by con-nective-tissue fibers. The three semicircularducts are arranged perpendicularly to eachother. The convexity of the anterior semi-circular duct (B23) is oriented toward thesurface of the petrous pyramid, the posteriorsemicircular duct (B24) runs parallel to theposterior surface of the petrous bone, andthe lateral semicircular duct (B25) runs hori-zontally.
Each semicircular duct has a dilatation at itstransition to the utricle, the membranousampulla (B26), which corresponds to an os-seous ampulla in the bony canal. The ante-rior and the posterior semicircular ductsjoin to form the common membranous crus(AB27). Each ampulla contains sensoryepithelium, the ampullary crest.
The courses taken by the semicircular ductsdo not correspond to the axes of the body.The anterior and posterior semicircularducts diverge from the median and frontalplanes by 45!; the lateral semicircular ductis tilted in posterocaudal direction by 30!towards the horizontal plane.
C28 Eardrum.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

369
TheEa
r
14
2821
2
5
174
3
22
27
11
9102 3
1
1321
20
19
7
68
15
12
23
25
27
7
6
26
8
9
13 18
19
26
10
202116
15
24
Inner Ear
A Internal ear, schematic view
B Membranous labyrinth(according to Krieg)
C Osseous labyrinth(according to Platzer)
D Position of the internal earin the skull
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

370Th
eEa
rThe Ear: Structure
Inner Ear (continued)
Cochlea (A–C)
The cochlea spirals around a conical bonyaxis, the central pillar of cochlea, or modiolus(AC1), which contains the neurons of thespiral ganglion (AB2) (p. 377, D9), the nervefibers originating from them (AB3), and theradix cochlearis (A4) (p. 377, D11) in the cen-ter. A double plate of bone, the osseous spirallamina (A–C5), protrudes from the modio-lus far into the cochlear duct (A6, B). Itforms a spiral like the cochlea, but it doesnot reach into the end of the uppermostconvolution and terminates in a free, hook-like process, the hamulus of spiral lamina(C7). The bony spiral lamina is mostly hol-low and contains nerve fibers extending tothe organ of Corti. Opposite to it at thelateral wall, in the lower half of the basalconvolution, lies the secondary spirallamina.
The spiral canal of cochlea contains themembranous cochlear duct (scala media)(A–C8) which is filled with endolymph.Above the duct lies the scala vestibuli (A–C9)and below it the scala tympani (A–C10); boththese spaces contain perilymph. The lowerwall of the cochlear duct is formed by thebasilar membrane (B11) which carries thesensory receptor for hearing, the organ ofCorti (B12). The width of the membrane var-ies in individual convolutions. The finefibers of the membrane radiate like a fanprior to attaching to the lateral wall of thecochlear canal, forming the spiral ligamentof cochlea (B13), which looks like a sickle incross section. Its part above the basilarmembrane forms the lateral wall of thecochlear duct; it is known as vascular stria(B14) because it is rich in endolymph-pro-ducing capillaries. The upper wall of thecochlear duct is a thinmembrane of double-layered epithelium, Reissner’s membrane orvestibular wall of the cochlear duct (B15).
The scala vestibuli communicates with theperilymphatic space of the vestibule andturns into the scala tympani at the heli-cotrema (AC16). The scala tympani runstoward the cochlear window (p. 365, D)
which is closed by the secondary tympanicmembrane. The connection between thetwo ducts is made possible by separation ofthe spiral lamina from the modiolus andformation of the hamulus. In this way, thehelicotrema is created medially. Only thescala vestibuli and the cochlear duct ascendto the uppermost tip of the cochlea, thecupula (A17). In contrast to the rest of thecochlea, the cupula thus contains only twomembranous spaces.
Frequency analysis in the cochlea. Theoscillations of sound waves are transmittedto the perilymph through the vestibularwindow via eardrum and auditory ossicles.The resulting movements of the fluid as-cend in the scala vestibuli and descend inthe scala tympani to the cochlear window,where thewaves ofmovement are absorbed(C). Movements of the fluid lead to oscilla-tions of the basilar membrane (travelingwaves). The site ofmaximal displacement ofthe basilar membrane (and hence stimula-tion of the receptor cells in the organ ofCorti) depends on the frequency of thetraveling wave or stimulating sound. Hightonal frequencies cause maximal displace-ment of the basilar membrane in the basalconvolutions (where the basilar membraneis narrow), middle frequencies in themiddle of the cochlea, and low frequenciesin the uppermost convolutions (where thebasilar membrane is wide). Hence, differentfrequencies are registered in different partsof the cochlea, namely, frequencies of20000Hz in the basal convolutions andfrequencies of 20Hz in the uppermost con-volutions. This local arrangement providesthe basis of the tonotopic organization ofthe acoustic system (p. 381, C).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
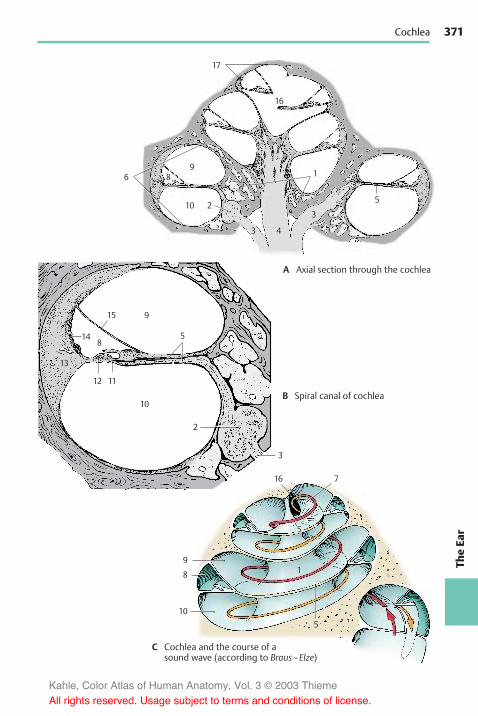
371
TheEa
r
17
16
9
915
14
13
58
12 11
10
2
3
16 7
5
1
510
1
5
86
10 2
3 4
3
9
8
Cochlea
A Axial section through the cochlea
B Spiral canal of cochlea
C Cochlea and the course of asound wave (according to Braus–Elze)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

372Th
eEa
rThe Ear: Structure
Inner Ear (continued)
Organ of Corti (A–C)
The basilar membrane (AB1) is covered on itslower surface by connective tissue cells thatline the scala tympani (AB2); on its uppersurface rests the organ of Corti, or spiralorgan (A3, B). Lateral to the spiral organ, theepithelium continues as the vascular stria(A4), which contains numerous intra-epithelial capillaries. Medial to the spiralorgan, at the margin of the bony spirallamina, lies a thickened tissue layer derivedfrom the inner periosteum, the limbus of thespiral lamina (A5); it is covered by epithelialcells. The limbus tapers off into two lips, thetympanic lip of the limbus (A6) and the vesti-bular lip of the limbus (A7), which enclosethe internal spiral sulcus (A8).
The organ of Corti extends in a spiral fromthe basal convolution to the cupula of thecochlea. The figures (A, B) show cross sec-tions through the organ; it consists ofsensory cells and a variety of supportingcells. The center is occupied by the innertunnel (B9), which contains a perilymph-like fluid, the cortilymph. Its medial wall isformed by the inner pillar cells (B10) and itslateral wall by the obliquely arranged outerpillar cells (B11). The pillar cells have a widebase (B12) containing the cell nucleus, anarrow middle part, and a head. They con-tain long bundles of supporting tonofila-ments. The inner pillar cell forms the headplate (B13), while the outer pillar cell formsa round head part (B14), which approachesthe head plate from below, as well as a flatprocess, the phalangeal process (B15). Later-ally follows the group of Deiters’ supportingcells (outer phalangeal cells) (B16); thesecarry the sensory cells (B17, C) on a protru-sion of their lower part. Their tonofilamentbundles branch below the sensory cells toform a support basket (B18). From eachDeiters’ cell, a narrow process ascends be-tween the sensory cells and ends with a flatphalangeal process (C19). The phalangealprocesses together form a perforated super-ficial membrane, the reticular membrane,and the apical ends of the sensory cells arefirmly secured in the openings of the mem-
brane. Between the outer pillar cells and theDeiters’ cells lies the inner tunnel, or Corti’sspace (B20), and laterally to the Deiters’cells lies the small outer tunnel, or Nuel’sspace (B21). This is followed by a group ofsimple, elongated supporting cells thatmerge into the epithelium of the vascularstria to form the medial wall of the externalspiral sulcus (A22). The inner phalangealcells border onto the inner pillar cells.
The sensory cells (C) include the inner haircells (C23), which form only one row, andthe outer hair cells (C24); the latter formthree rows in the basal convolution of thecochlea, four rows in the middle convolu-tion, and five rows in the upper convolution.All hair cells have below their apical surfacea dense terminal web of microfilaments(C25) through which the sensory hairs(C26) are firmly secured. The sensory hairsare stiff, specialized microvilli (stereovilli)which are arranged in a semicircle, usuallyin three rows of different lengths. Nervefibers with synapse–like contacts (C27) ter-minate at the base of the hair cells.
A nonoscillating gelatinous mass, the tec-torial membrane (AB28), lies on top of thehair cells; it covers the spiral limbus and ex-tends beyond its vestibular lip. It is not clearwhether or not the stereovilli of the sensorycells are attached to the tectorial membraneand their tangential deflection during oscil-lation of the basilar membrane is broughtabout by a shift against the tectorial mem-brane. It is also possible that the stereovillido not touch the tectorial membrane at alland are merely moved by the flow of en-dolymph.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

373
TheEa
r
4
2232887
6
17
18
13 14 15 28
2021
1 12 16 2
26
25
26 19
24
27
23
27
119
10
1 2
5
Organ of Corti
A Cochlear duct
B Organ of Corti
C Inner hair cell and outer hair cellof the organ of Corti (accordingto Wersäll and Lundquist)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

374Th
eEa
rThe Ear: Structure
Inner Ear (continued)
Vestibular Apperatus (A–D)
Saccule, utricle, and the three semicircularducts emanating from the utricle form theorgan of balance, the vestibular apparatus.It contains several sensory fields, namely,the two acoustic maculae, macula of sacculeand macula of utricle, and the three ampullarycrests. They all register acceleration andpositional changes and, therefore, servespatial orientation. The maculae react tolinear acceleration in different directions,the crests react to rotational acceleration.The maculae occupy specific positions inspace (p. 369, B8, B9); the macula of theutricle lies roughly horizontally on the floorof the utricle, and the macula of the sacculelies vertically on the anterior wall of the sac-cule. Thus, both are arranged at right anglesto each other. (For positions of the semi-circular ducts, see p. 369.)
Acoustic maculae (A). The epithelium lin-ing the endolymphatic space increases inheight in the oval areas of the maculae anddifferentiates into supporting cells andsensory cells. The supporting cells (A1) carryand surround the sensory cells (A2). Eachsensory cell is shaped like a flask or am-poule and bears 70–80 stereovilli on its api-cal surface (A3). The sensory epithelium issurmounted by a gelatinous membrane, thestatoconic (otolithic) membrane (A4), whichcarries crystalline particles of calcium car-bonate, the ear crystals, or statocones(otoliths) (A5). The stereovilli of the sensorycells do not directly project into the stato-conic membrane but are surrounded by anarrow space containing endolymph.
Function of the maculae. The properstimulus for the stereovilli is a shearingforce affecting the macula; with increasingacceleration there is a tangential shift be-tween sensory epithelium and statolithicmembrane. The resulting deflection of thestereovilli leads to stimulation of thesensory cell and to induction of a nerve im-pulse.
Ampullary crest (B, C). The crest (BC6) isformed by a protrusion in the ampulla and isoriented transversely to the course of thesemicircular duct (C). Its surface is coveredby supporting cells (B7) and sensory cells(B8). Each sensory cell bears approximately50 stereovilli (B9) that are considerablylonger than those of the macular cells. Theampullary crest occupies about one third ofthe height of the ampulla. It is surmountedby a gelatinous cap, the ampullary cupula(B–D10), which reaches to the roof of theampulla. The cupula is traversed by longchannels into which the hair bundles of thesensory cells protrude. The bases of thesensory cells are innervated by nerveendings (A–C11).
Function of the semicircular canals (D).The semicircular canals respond to ro-tational acceleration which sets the en-dolymph inmotion. The resulting deflectionof the ampullary cupula bends thestereovilli of the sensory cells and acts asthe triggering stimulus. For example, if thehead is turned to the right (red arrows), theendolymph of the lateral semicircular ductinitially remains in place due to its inertia;this results in a relative movement in theopposite direction (hydrodynamic inertia,black arrows) so that both cupulae are de-flected toward the left (D12). The en-dolymph then slowly follows the rotation ofthe head. However, once the rotation hasstopped (broken, arrested arrows), it con-tinues to flow for a certain distance in thesame direction so that the cupulae are de-flected to the right (D13). The function ofthe semicircular ducts serves primarily thereflex eye movements. Rapid eye move-ments caused by rotation of the head (ro-tatory nystagmus) depend on cupular de-flection. The slow component of nystagmusalways follows the direction of the cupulardeflection.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

375
TheEa
r
4
5
3
2
11
1
10
10
10
116
12
13
10
9
8
6
7
11
Vestibular Apparatus
A Acoustic macula
B Ampullary crest
C Ampulla with crest
D Function of the semicircularcanals (according to Trincker)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

376Th
eEa
rThe Ear: Structure
Inner Ear (continued)
Vestibular Sensory Cells (A–C)
The hair cells of the acoustic maculae andampullary crests share the same structuralprinciple. They are mechanoreceptors re-sponding to the tangential deflection oftheir sensory hairs. There are two types ofvestibular hair cells: type I cells have theshape of a flask, while type II cells have theshape of a cylinder.
Type I hair cells (A1) have a round cell bodyand a narrow neck; below their apical sur-face lies a dense terminal web (A2). The api-cal cell surface is differentiated into ap-proximately 60 stereovilli (A3) of graduatedlengths and a single, very long motilekinocilium (A4) with a basal body at itsorigin. Each type I hair cell is surrounded onits lateral and basal surfaces by a nerve calyx(A5), which is formed by a thick nervefiber. The upper part of the calyx containsvesicles and closely adjoins the hair cell;this region is therefore regarded as the syn-aptic part of the hair cell. The nerve calyx, inturn, is contacted by heavily granulatednerve endings (A6), possibly representingthe terminals of efferent nerve fibers.
Type II hair cells (A7) are equipped with anidentical set of sensory hairs. At the base ofthe cell lie more (A8) or less heavily granu-lated terminals of nerve fibers.
All hair cells of a sensory field show a uni-form orientation of their kinocilia (B). Elec-trophysiological studies have shown thatbending of the stereovilli toward thekinocilium results in stimulation (greenarrow), while bending in the opposite direc-tion leads to inhibition (red arrow) (C).Movements in intermediate directionscause stimulation or inhibition below thethreshold of sensation.
In this way, the vestibular apparatus is ableto register each movement precisely. Thesemicircular ducts (kinetic labyrinth) controlprimarily the eye movements, whereas theacoustic maculae (tonic labyrinth) directly af-fect muscular tone, in particular, the tensionof extensor muscles and neck muscles(p. 383 C).
Spiral Ganglion and Vestibular Ganglion(D)
The spiral ganglion (D9) consists of a chainof nerve cell clusters lying in the modiolusat the margin of the osseous spiral lamina.Together these clusters form a spiral. Theycontain bipolar neurons, the peripheralprocesses (dendrites) of which extend to thehair cells of the organ of Corti. Their centralprocesses (axons) run as foraminous spiraltract (D10) to the axis of the modiolus,where they unite to form the radixcochlearis (root of the cochlear nerve) (D11).
The vestibular ganglion (D12) lies at thefloor of the internal acoustic meatus. It con-sists of a superior part and an inferior part.The bipolar neurons of the superior part(D13) send their peripheral processes to theampullary crests of the anterior semicircu-lar duct (D14) (anterior ampullary nerve)and the lateral semicircular duct (D15)(lateral ampullary nerve), to the macula ofthe utricle (D16) (utricular nerve) and topart of the macula of the saccule (D17). Theneurons of the inferior part (D18) supply theampullary crest of the posterior semicircu-lar duct (D19) (posterior ampullary nerve)and part of themacula of the saccule (saccu-lar nerve). The central processes form theradix vestibularis (root of the vestibularnerve) (D20), which together with the radixcochlearis extends in a common nervesheath through the internal acoustic meatusinto the middle cranial fossa.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

377
TheEa
r
32
1
6
7
8
5
4
14
15
17
16
19
9
20 11
1012
13
18
Vestibular Sensory Cells, Spiral Ganglion, and Vestibular Ganglion
A
Vestibular sensory cells:type I and type II haircells, electron-microscopicdiagram (accordingto Wersäll)
B Arrangement of sensory cells(according to Flock and Wersäll)
C Function of the sensory hairs(stereovilli plus kinocilium)
D Nerve supply of the internal ear(according to Krieg)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

378Th
eEa
rThe Ear
Auditory Pathway andVestibular Pathways
Auditory Pathway
Cochlear Nuclei (A, B)
The fibers of the radix cochlearis (A1) enterthe medulla oblongata at the level of theanterior cochlear nucleus (AB2) and bifurcate.The ascending branches extend to the poste-rior cochlear nucleus (AB3), and the de-scending branches to the anterior cochlearnucleus. The projection of the cochlea ontothe nuclear complex is highly organized;fibers from the basal cochlear convolutionsterminate in the dorsomedial parts of thenuclei, while fibers from the uppermostconvolutions terminate in the ventrolateralparts. This regular distribution of the affer-ent fibers is the basis for the subdivision ofthe cochlear nuclei according to tonalfrequencies (see p. 370, frequency analysisin the cochlea).
Such a tonotopic organization of thecochlear complex can be demonstrated byelectrical recordings in an experimental an-imal (cat) (B). Recordings from individualneurons and simultaneous sonication withdifferent sounds can detect the frequency atwhich an individual cell responds best. Thefrontal section through the oral region ofthe cochlear nuclei shows how the elec-trode, which scans from top to bottom, reg-isters in the posterior nucleus (B3) therecorded points in a precise sequence fromhigh to low frequencies. Here lie the neu-rons for specific tonal frequencies in a regu-lar sequence. When the electrode enters theanterior nucleus (B2), this sequence sud-denly stops and frequencies oscillate only ina specific range.
The secondary fibers of the auditory path-way originate from the neurons of thecochlear nuclei. Fiber bundles from theanterior cochlear nucleus cross to the op-posite side as a broad fiber plate mixed withnerve cells, the trapezoid body (A4) (p. 111,AB15), and then ascend as the lateral lem-niscus (A5) (p. 133, D22) to the inferior col-liculus (A6). Fibers originating from the pos-
terior cochlear nucleus cross obliquely asposterior acoustic striae (A7). A large portionof the lemniscus fibers run from thecochlear nuclei directly to the inferior col-liculus. A considerable number of fibers,however, are relayed to tertiary fibers in theintermediate nuclei of the auditory path-way, namely, in the posterior nucleus of thetrapezoid body (superior olive) (A8), in theanterior nucleus of the trapezoid body (A9), andin the nuclei of the lateral lemniscus. A tono-topic organization has been demonstratedin the nucleus of the posterior trapezoidbody. The accessory nucleus lying mediallyto it, the medial nucleus of the superior olive(A10), receives fibers from the cochlear nu-clei of both sides and is interposed into afiber system serving directional hearing. Thefiber connections from the posterior nu-cleus of the trapezoid body to the abducensnucleus (A11) (reflex eye movements result-ing from the sensation of sound) are still dis-puted. The fibers are thought to extend be-yond the abducens nucleus to the con-tralateral cochlear nucleus and terminate asefferent fibers on the hair cells of the organof Corti. They probably control the inflow ofstimuli. The lateral nuclei of the lemniscusare scattered clusters of nerve cells inter-posed in the course of the lateral lemniscus.From the posterior nucleus of the lateral lem-niscus (A12), fibers cross to the con-tralateral lemniscus (Probst ’s commissure)(A13).
Inferior Colliculus (A)
The predominating portion of the laterallemniscus terminates in the principal nu-cleus of the inferior colliculus in a topicalpattern. Electrophysiological studies havedemonstrated a tonotopic organization ofthis nucleus. It is a relay station for acousticreflexes, from where acoustico-optic fibersextend to the superior colliculi and tec-tocerebellar fibers to the cerebellum. Theinferior colliculi are interconnected by thecommissure of the inferior colliculi (A14).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

379
TheEa
r
25.021.820.918.316.515.014.0
12.512.020.524.329.0
28.529.028.524.821.8
3
2
6 14
13
12
3
2
8
10
9
5 11 7
4
1
Auditory Pathway
A Auditory pathway
B Tonotopic organization of thecochlear nuclei (according to Roseand Mountcastle)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

380Th
eEa
rThe Ear: Auditory Pathway and Vestibular Pathways
Auditory Pathway (continued)
Medial Geniculate Body (A, B)
The next section of the auditory pathway isthe peduncle of the inferior colliculi (A1),which extends as a strong fiber tract fromthe inferior colliculus to themedial geniculatebody (AB2), from where the acoustic radia-tion originates. The medial geniculate bodyis thought also to contain somatosensoryfibers from the spinal cord as well as cere-bellar fibers. Obviously, it is not solely arelay station of the acoustic system but isalso involved in other systems. Some fiberbundles of the peduncle of the inferior col-liculi are derived from the trapezoid nucleiand reach the medial geniculate bodywithout synapsing in the inferior colliculus.The two medial geniculate bodies arethought to be interconnected by crossingfibers running through the inferior supra-optic commissure (Gudden’s commissure)(A3). It is uncertain whether such connec-tions exist in humans. However, the pres-ence of fibers descending from the auditorycortex and terminating in the geniculatebody has been confirmed.
Acoustic Radiation (A, B)
The fibers of the acoustic radiation (AB4) runfrom the medial geniculate body trans-versely through the inferior posterior partof the internal capsule and ascend verticallyin the temporal lobe to the auditory cortex.The fibers show a topical organization withindividual sections of the medial geniculatebody projecting to specific regions of theauditory cortex. Within the acoustic radia-tion, the fibers undergo a spiral rotation sothat the rostral parts of the geniculate bodyproject to the caudal cortical areas and thecaudal parts of the geniculate body projectto rostral cortical areas (B). This rotation hasbeen demonstrated experimentally inmonkeys and during maturation of the my-elin sheath in humans.
Auditory Cortex (A–C)
Electrical recordings from the cortex ofvarious experimental animals (cat, monkey)
under simultaneous exposure to sounds ofdifferent frequencies have uncovered thetonotopic organization of the auditory cor-tex (AB5, C), where the uncoiled cochlea isrepresented from the basal convolution tothe cupula. Three regions of hearing havebeen demonstrated: the primary auditoryarea (AI) (C6), the secondary auditory area (AII)(C7), and the area of the posterior ectosylviangyrus (Ep) (C8). In the primary auditory area,the neurons responding best to highfrequencies (large blue dots) lie rostrally;the neurons responding best to lowfrequencies (small blue dots) lie caudally. Inthe secondary auditory area, the frequencysensitivity is arranged in the reverse order.Auditory area AI is the primary terminal ofthe acoustic radiation, while auditory areasAII and Ep are regarded as secondary audi-tory areas. The relationships may be com-pared to those of the visual cortex, wherearea 17 is the terminal of the optic radiationwhile areas 18 and 19 are secondary inte-gration areas. Auditory area AI correspondsin humans to area 41, which covers Heschl’sconvolutions (transverse temporal gyri) andis the terminal of the acoustic radiation(p. 252, C1). Areas 42 and 22 are secondaryauditory areas and include Wernicke’sspeech center for the comprehension of lan-guages (p. 262, A1). Hence, the auditory cor-tex must be regarded as a region muchlarger than Heschl’s convolutions.
The auditory pathway has several commis-sural systems along its course that providefiber exchanges at various levels. Some fiberbundles also ascend to the ipsilateral audi-tory cortex. The latter therefore receives im-pulses from both organs of Corti, and this isespecially important for directional hearing.
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

381
TheEa
r
5
4
2
1
4
3
5
2
6
87
Auditory Pathway
A Auditory pathway
B Fiber relationship between themedial geniculate body and theauditory cortex (according toWalker)
C Tonotopic organization of the auditory cortexof the cat (according to Woolsey)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

382Th
eEa
rThe Ear: Auditory Pathway and Vestibular Pathways
Vestibular Pathways
Vestibular Nuclei (A, B)
The fibers of the radix vestibularis (A1) enterthe medulla oblongata at the level of thelateral vestibular nucleus (Deiters’ nucleus)(AB2) and bifurcate into ascending and de-scending branches that terminate in the su-perior vestibular nucleus (Bechterew’s nu-cleus) (AB3), in the medial vestibular nucleus(Schwalbe’s nucleus) (AB4), and in the infe-rior vestibular nucleus (AB5) (p. 120, B). Thenerve fibers for different parts of the laby-rinth extend to specific regions of the nu-clear complex. Fiber bundles for the maculaof the saccule (B6) terminate in the lateralpart of the inferior nucleus, while fibers forthemacula of the utricle (B7) end in the me-dial part of the inferior nucleus and in thelateral part of the medial nucleus. Fibers forthe ampullary crests (B8) terminate pri-marily in the superior nucleus and in theupper part of the medial nucleus.
Certain groups of neurons respond to linearacceleration and others to rotational accel-eration. Some neurons respond to ipsilateralrotation, others to contralateral rotation.The vestibular complexes of both sides areinterconnected by commissural fibers,through which some nerve cell clusters arestimulated by the labyrinth of the con-tralateral side. In addition to labyrinthinefibers, cerebellar fibers from the vermis andthe fastigial nuclei (p. 164, B) as well as spi-nal fibers transmitting impulses from thejoint receptors terminate in the nuclearcomplex. Efferent fibers for central controlrun from the vestibular nuclei back to thesensory epithelia.
Secondary Vestibular Pathways (A, C)
These are connections to the spinal cord, tothe reticular formation, to the cerebellum,and to the oculomotor nuclei. The vesti-bulospinal tract (A9) originates from the neu-rons of Deiters’ nucleus (lateral vestibularnucleus) and reaches into the sacral spinalcord. Its fibers terminate at the spinal inter-neurons and activate the !- and "-mo-toneurons of the extensor muscles.
The numerous fibers extending to the retic-ular formation stem from all vestibular nu-clei. To the cerebellum run direct fibersfrom the vestibular ganglion as well as fiberbundles from the medial and inferiorvestibular nuclei. They terminate in thenodulus and flocculus (A10) and in parts ofthe uvula (vestibulocerebellum, p. 152, A6;p. 164, B). The fibers ascending to the oculo-motor nuclei (AC11) stem primarily fromthe medial and superior vestibular nuclei;they form part of the medial longitudinalfasciculus (A12). There is also a vesti-bulocortical connection through thethalamus (ventral intermediate nucleus,p. 184, B13). Electrophysiological studies re-sulted in a projection of vestibular impulsesto a small region in the anterior postcentralarea near the facial region of the sensoryhomunculus (p. 251, C).
Interaction of eye muscles, neck muscles,and organ of balance (C). The connectionbetween vestibular complex and oculomo-tor nuclei brings circumscribed groups ofneurons into contact with each other. Nervecell clusters receiving impulses from aspecific semicircular duct are probably con-nectedwith nerve cell clusters innervating aspecific eye muscle. This would explain theexceptionally precise interaction of vestib-ular apparatus, eye muscles, and neckmuscles, thus permitting fixation of an ob-ject even during movements of the head.We always perceive a stationary, verticalimage of our surroundings despite our headmovements. To guarantee such a constantvisual impression, each head movement iscompensated for by rotation of the eyeballs.The finely tuned interaction of neckmusclesand ocular muscles is controlled by the ves-tibular apparatus via "-motoneurons (C13).
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
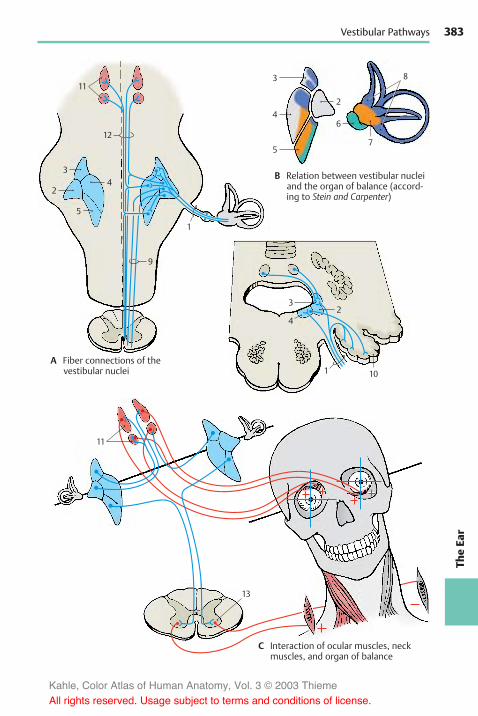
383
TheEa
r
4
3
57
8
2
32
10
11
1
4
6
3
11
4
1
9
12
2
5
13
Vestibular Pathways
A Fiber connections of thevestibular nuclei
B Relation between vestibular nucleiand the organ of balance (accord-ing to Stein and Carpenter)
C Interaction of ocular muscles, neckmuscles, and organ of balance
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

384
Further Reading
Textbooks, generalAriëns Kappers, C. U., G. C. Huber, E. C. Crosby: TheComparative Anatomy of the Nervous System ofVertebrats, Including Man. Hafner, New York1936, Reprint 1960
Brodal, A.: Neurological Anatomy. Oxford UniversityPress, Oxford 1981
Carpenter, M. B.: Core Text of Neuroanatomy. Wil-liams & Wilkins, Baltimore 1978
Clara, M.: Das Nervensystem des Menschen. Barth,Leipzig 1959
Creutzfeld, O. D.: Cortex Cerebri. Springer, Berlin,Heidelberg, New York, Tokyo 1983
Curtis, B. A., S. Jakobson, E. M. Marcus: An Introduc-tion to the Neurosciences. Saunders, Philadelphia1972
Dejerine, J.: Anatomie des centres nerveux. Rueff,Paris 1895–1901
Eccles, J. C.: Das Gehirn des Menschen. Piper,München 1973
Friede, R. L.: Topographic Brain Chemistry. Aca-demic Press, New York 1966
Greger, R., U. Windhorst: Comprehensive HumanPhysiology. From Cellular Mechanisms to Integra-tion. Vol. I und II. Springer, Berlin, Heidelberg,New York 1996
Kandel, E. R., J. H. Schwartz, T. M. Jessell: Principlesof Neural Science. 3 rd Edition. Appleton & Lange1991
Ludwig, E., J. Klingler: Atlas cerebri humani. Karger,Basel 1956
Nieuwenhuys, R., J. Voogd, Chr. van Huizen: TheHuman Nervous System: A Synopsis and Atlas,3rd ed. Springer, Berlin 1988
Retzius, G.: Das Menschenhirn. Norstedt, Stockholm1896
Schaltenbrand, G., P. Bailey: Einführung in diestereotaktischen Operationenmit einemAtlas desmenschlichen Gehirns. Thieme, Stuttgart 1959
Schmidt, R. F., G. Thews (Hrsg.): Physiologie desMenschen, 26. Aufl. Springer, Berlin 1995
Sidman, R. L., M. Sidmann: Neuroanatomie pro-grammiert. Springer, Berlin 1971
Thompson, R. F.: Das Gehirn. Von der Nervenzellezur Verhaltenssteuerung. 2. Auflage. Spektrum,Heidelberg, Berlin, Oxford 1994
Villiger, E., E. Ludwig: Gehirn und Rückenmark.Schwabe, Basel 1946
Zigmond, M. J., F. E. Bloom, S. C. Landis, J. L. Roberts,L. R. Squire: Fundamental Neuroscience. Aca-demic Press, San Diego, London, Boston 1999
IntroductionBullock, Th. H.: Introduction to Nervous System. San
Francisco 1977Bullock, Th. H., G. A. Horridge: Structure and Func-
tion in the Nervous System of Invertebrates. Uni-versity Chicago Press, Chicago 1955
Drescher, U., A. Faissner, R. Klein, F. G. Rathjen, C.Stürmer (eds.): Molecular bases of axonal growthand pathfinding. Cell Tissue Res. Special Issue 290(1997) 187–470
Eccles, J. C.: How the Self Controls the Brain.Springer, Berlin 1994
Edelman, G. M., W. E. Gall, W. M. Cowan (eds.):Molecular Bases of Neural Development. JohnWiley & Sons, New York, Chichester, Brisbane1985
Goodman, C. S., C. J. Shatz: Developmental mecha-nisms that generate precise patterns of neuronalconnectivity. Cell 72/Neuron 10 (Suppl.) (1993)77–98
Hamburger, V.: The Heritage of ExperimentalEmbryology. Hans Spemann and the Organizer.Oxford University Press, New York 1988
Herrick, J. C.: Brains of Rats andMen. University Chi-cago Press, Chicago 1926
Herrick, J. C.: The Evolution of Human Nature. Uni-versity Texas Press, Austin 1956
Kahle,W.: Die Entwicklung dermenschlichen Groß-hirnhemisphäre. Springer, Berlin, Heidelberg1969
Kölliker, A.: Entwicklungsgeschichte des Menschenund der höheren Tiere. 2. Ausgabe.W. Engelmann,Leipzig 1879
Le Gros Clark, W. E.: Fossil Evidence for Human Evo-lution. University Chicago Press, Chicago 1955
Le Gros Clark, W. E.: The Antecedents of Man. Edin-burgh University Press. Edinburgh 1959
Sherrington, Sir Charles: Körper und Geist – DerMensch über seine Natur. Schünemann, Bremen1964
Sperry, R.W.: Chemoaffinity in the orderly growthofnerve fiber patterns and connections. Proc. Natl.Acad. Sci. USA 50 (1963) 703–710
Tessier-Lavigne, M., C. S. Goodman: The molecularbiology of axon guidance. Science 274 (1996)1123–1133
Tobias, P. V.: The Brain inHominid Evolution. Colum-bia University Press, New York 1971
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

385
Basic Elements ofthe Nervous SystemAkert, K., P. G. Waser: Mechanisms of Synaptic
Transmission. Elsevier, Amsterdam 1969Babel, J., A. Bischoff, H. Spoendlin: Ultrastructure of
the Peripheral Nervous System and Sense Organs.Thieme, Stuttgart 1970
Barker, J. L.: The Role of Peptides in Neuronal Func-tion. Dekker, Basel 1980
de Belleroche, J., G. J. Dockray: Cholecystokinin inthe Nervous System. Verlag Chemie, Weinheim1984
Björklund, A., T. Hökfeld: Classical Neurotransmit-ters in the CNS. Handbook of Chemical Neu-roanatomy. Elsevier, Amsterdam 1984
Bloom, F. E.: The functional significance of neu-rotransmitter diversity. Am. J. Physiol. 246 (1984)C184–C194
Cajal, S. R.: Histologie du système nerveux del'homme et des vertébrés. Maloine, Paris 1909–1911
Causey, G.: The Cell of Schwann. Livingstone, Edin-burgh 1960
Cold Spring Harbour Symposia 40: The Synapse.Cold Spring Harbour Laboratory, New York 1976
Cottrell, G. A., P. N. R. Usherwood: Synapses. Blackie,Glasgow 1977
Cowan, W. M., M. Cuenod: The Use of Axonal Trans-port for Studies of Neuronal Connectivity. El-sevier, Amsterdam 1975
De Robertis, E. D. P., R. Carrea: Biology of Neuroglia.Elsevier, Amsterdam 1965
Eccles, J. C.: The physiology of synapses. Springer,Berlin, Göttingen, Heidelberg, New York 1964
Emson, P. C.: Chemical Neuroanatomy. Raven, NewYork 1984
Eränkö, O.: Histochemistry of Nervous Transmis-sion. Elsevier, Amsterdam 1969
Fedoroff, S., A. Vernadakis: Astrocytes. AcademicPress, London 1986
Frotscher, M., U. Misgeld (eds.): Central CholinergicSynaptic Transmission. Birkhäuser, Basel, Boston,Berlin 1989
Fuxe, K., M. Goldstein, B. Hökfeld, T. Hökfeld: CentralAdrenalin Neurons. Pergamon, Oxford 1980
Gray, E. G.: Axo-somatic and axo-dendritic synapsesof the cerebral cortex: an electron microscopestudy. J. Anat. 93 (1959) 420–433
Heimer, L., L. Záborszky (eds.): NeuroanatomicalTract-TracingMethods 2. Recent Progress. PlenumPress, New York, London 1989
Heuser, J. E., T. S. Reese : Structure of the synapse. In:Handbook of Physiology, Section 1: The NervousSystem. Vol. I: Cellular Biology of Neurons, Part 1(E. R. Kandel, Ed.), American Physiological Society,Bethesda, Md., 261–294, 1977
Jonas, P., H. Monyer (eds.): Ionotropic Glutamate Re-ceptors in the CNS. Handbook of Experimental
Pharmacology. Vol. 141. Springer, Berlin, Heidel-berg, New York 1999
Jones, D. G.: Synapses and Synaptosomes. Chap-mann & Hall, London 1975
Landon, D. N.: The Peripheral Nerve. Chapmann &Hall, London 1976
Loewi, O.: Über humorale Übertragbarkeit der Herz-nervenwirkung. Pflügers Arch. Gesamte Physiol.189 (1921) 239–242
Nakai, J.: Morphology of Neuroglia. Igaku-Shoin,Osaka 1963
Neher, E.: Ion channels for communication betweenand within cells. Neuron 8 (1992) 606–612
Pappas, G. D., D. P. Purpura: Structure and Functionof Synpases. Raven, New York 1972
Penfield, W.: Cytology and Cellular Pathology of theNervous System. Hoeber, New York 1932
Peters, A., S. L. Palay, H. F. Webster: The Fine Struc-ture of the Nervous System. Oxford University.Press, New York 1991
Pfenninger, K. H.: Synaptic morphology and cyto-chemistry. Prog. Histochem. Cytochem. 5 (1973)1–86
Ransom, B., H. Kettenmann (eds.): Neuroglia. OxfordUniversity Press, Oxford 1995
Rapoport, St. J.: Blood-Brain Barrier in Physiologyand Medicine. Raven, New York 1976
Roberts, E., T. N. Chase, D. B. Tower: GABA in NervousSystem Function. Raven, New York 1976
Sakmann, B.: Elementary steps in synaptic trans-mission revealed by currents through single ionchannels. Neuron 8 (1992) 613–629
Sakmann, B., E. Neher: Single-channel recording.Plenum, New York 1983
Schoffeniels, E., G. Franck, L. Hertz: Dynamic Proper-ties of Glia Cells. Pergamon, Oxford 1978
Stjaerne, L.: Chemical Neurotransmission. AcademicPress, London 1981
Szentágothai, J.: Neuron Concept Today. AdakémiaiKiadó, Budapest 1977
Uchizono, K.: Excitation and inhibition. Elsevier,Amsterdam 1975
Unwin, N.: Neurotransmitter action: opening ofligand-gated ion channels. Cell 72/Neuron 10(Suppl.) (1993) 31–41
Usdin, E., W. E. Burney, J. M. Davis: Neuroreceptors.Chichester, New York 1979
Watson,W. E.: Cell Biology of Brain. Chapman&Hall,London 1976
Windle,W. F.: Biology of Neuroglia. Thomas, Spring-field 1958
Weiss, D. G.: Axoplasmic Transport. Springer, Berlin1982
Yamamura, H. J., S. J. Enna, M. J. Kuhar: Neu-rotransmitter Receptor Binding. Raven, New York1985
Zimmermann, H.: Synaptic transmission : Cellularand Molecular Basis. Thieme, Stuttgart 1993
Further Reading
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

386
Spinal Cord and Spinal NervesBrown, A. G.: Organization of the Spinal Cord.
Springer, Berlin 1981Dyck, P. J., P. K. Thomas, E. H. Lambert: Peripheral
Neuropathy, Vol. I. Biology of the Peripheral Sys-tem. Saunders, Philadelphia 1975
Foerster, O.: Spezielle Anatomie und Physiologie derperipheren Nerven. In: Handbuch der Neurologie,Bd. II/I, hrsg. von O. Bumke, O. Foerster. Springer,Berlin 1928
Foerster, O.: Symptomatologie der Erkrankungendes Rückenmarks und seiner Wurzeln. In: Hand-buch der Neurologie, Bd. V, hrsg. von O. Bumke, O.Foerster. Springer, Berlin 1936
Hubbard, J. I.: The Peripheral Nervous System.Plenum, New York 1974
Kadyi, H.: Über die Blutgefäße des menschlichenRückenmarkes. Gubrynowicz & Schmidt, Lemberg1886
Keegan, J. J., F. D. Garrett: The segmental distributionof the cutaneous nerves in the limbs of man. Anat.Rec. 102 (1948) 409–437
v. Lanz, T., W. Wachsmuth: Praktische Anatomie,Bd. I/2.–4. Springer, Berlin 1955–1972
Murnenthaler, M., H. Schliack: Läsionen periphererNerven. 5. Aufl. Thieme, Stuttgart 1987
Noback, Ch. N., J. K. Harting: Spinal cord. In: Prima-tologia, Bd. II/1, hrsg. von H. Hofer, A. H. Schultz, D.Starck. Karger, Basel 1971
Villiger, E.: Die periphere Innervation. Schwabe,Basel 1964
Willis, W. D., R. E. Coggeshall: Sensory mechanismsof the spinal cord. 2nd ed. Plenum, New York 1991
Brain Stem and Cranial NervesBrodal, A.: The Cranial Nerves. Blackwell, Oxford1954
Brodal, A.: The Reticular Formation of the BrainStem. Oliver & Boyd, Edinburgh 1957
Clemente, C. D., H. W. Magoun: Der bulbäre Hirn-stamm. In: Einführung in die stereotaktischenOperationen mit einem Atlas des menschlichenGehirns, hrsg. von G. Schaltenbrand, P. Bailey,Thieme, Stuttgart 1959
Crosby, E. C., E. W. Lauer: Anatomie des Mittelhirns.In: Einführung in die stereotaktischen Opera-tionen mit einem Atlas des menschlichen Ge-hirns, hrsg. von G. Schaltenbrand, P. Bailey.Thieme, Stuttgart 1959
Delafresnaye, J. F.: Brain-Mechanisms and Consci-ousness. Blackwell, Oxford 1954
Duvernoy, H. M.: Human Brain Stem Vessels.Springer, Berlin 1978
Jasper, H., L. D. Proctor, R. S. Knighton, W. C. Noshay,R. T. Costello: Reticular Formation of the Brain.Churchill, Oxford 1958
Mingazzini, G.: Medulla oblongata und Brücke. In:Handbuch der mikroskopischen Anatomie, Bd. IV.Springer, Berlin 1928
Olszewski, J., D. Baxter: Cytoarchitecture of theHuman Brain Stem. Karger, Basel 1954
Pollak, E.: Anatomie des Rückenmarks, der Medullaoblongata und der Brücke. In: Handbuch der Neu-rologie, Bd. I, hrsg. von O. Bumke, O. Foerster.Springer, Berlin 1935
Riley, H. A.: An Atlas of the Basal Ganglia, Brain Stemand Spinal Cord. Williams & Wilkins, Baltimore1943
Spatz, H.: Anatomie des Mittelhirns. In: Handbuchder Neurologie, Bd. I, hrsg. von O. Bumke, O. Foer-ster, Springer, Berlin 1935
CerebellumAngevine, jr. J. B., E. L. Mancall, P. I. Yakovlev: TheHuman Cerebellum. Little, Brown, Boston 1961
Chain-Palay, V.: Cerebellar Dentate Nucleus.Springer, Berlin 1977
Dichgans, J., J. Bloedel, W. Precht: Cerebellar Func-tions. Springer, Berlin 1984
Dow, R. S., G. Moruzzi: The Physiology and Pathol-ogy of the Cerebellum. University MinnesotaPress, Minneapolis 1958
Eccles, J. C., M. Ito, J. Szentágothai: The Cerebellumas a Neuronal Machine. Springer, Berlin 1967
Fields, W. S., W. D. Willis: The Cerebellum in Healthand Disease. Green, St. Louis 1970
Ito, M.: The Cerebellum and Neural Control. RavenPress, New York 1984
Jakob, A.: Das Kleinhirn. In: Handbuch der mik-roskopischen Anatomie, Bd. IV, hrsg. von W. v.Moellendorff. Springer, Berlin 1928
Jansen, J., A. Brodal: Das Kleinhirn. In: Handbuch dermikroskopischen Anatomie, Bd. IV, hrsg. von W.Bargmann, Erg. zu Bd. IV/I. Springer, Berlin 1958
Larsell, O., J. Jansen: The Comparative Anatomy andHistology of the Cerebellum. University of Minne-sota Press. Minneapolis 1972
Llinás, R.: Neurobiology of Cerebellar Evolution andDevelopment. American Medical Association,Chicago 1969
Palay, S. L.: Cerebellar Cortex. Springer, Berlin 1974
Further Reading
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

387
DiencephalonAkert, K.: Die Physiologie und Pathophysiologie desHypothalamus. In: Einführung in die stereo-taktischen Operationen mit einem Atlas desmenschlichen Gehirns, hrsg. von G. Schalten-brand, P. Bailey. Thieme, Stuttgart 1959
Ariëns Kappers, J., J. P. Schadé: Structure and Func-tion of the Epiphysis Cerebri. Elsevier, Amsterdam1965
Bargmann, W., J. P. Schadé: Lectures on the Dien-cephalon. Elsevier, Amsterdam 1964
De Wulf, A.: Anatomy of the Normal HumanThalamus. Elsevier, Amsterdam 1971
Diepen, R.: Der Hypothalamus. In: Handbuch dermikroskopischen Anatomie, Bd. IV/7, hrsg. von W.Bargmann. Springer, Berlin 1962
Emmers, R., R. R. Tasker: The Human SomestheticThalamus. Raven, New York 1975
Frigyesi, T. L., E. Rinvik, M. D. Yahr: Thalamus. Raven,New York 1972
Harris, G. W., B. T. Donovan: The Pituitary Gland,Bd. III. Pars Intermedia and Neurohypophysis.Butterworths, London 1966
Hassler, R.: Anatomie des Thalamus. In: Einführungin die stereotaktischen Operationen mit einemAtlas des menschlichen Gehirns, hrsg. von G.Schaltenbrand, P. Bailey. Thieme, Stuttgart 1959
Haymaker, W., E. Anderson, J. H. Nauta: Hy-pothalamus. Thomas, Springfield/III. 1969
Kuhlenbeck, H.: The human diencephalon. Confin,neurol. (Basel), Suppl. 14 (1954)
Macchi, G., A. Rustioni, R. Speafico: SomatosensoryIntegration in the Thalamus. Elsevier, Amsterdam1983
Morgane, P. J.: Handbook of the Hypothalamus (3Vol.) Dekker, Basel 1979–81
Nir, I., R. J. Reiter, R. J. Wurtman: The Pineal Gland.Springer, Wien 1977
Pallas, J. E.: La journée du thalamus, Marseille 1969Purpura, D. P.: The Thalamus, Columbia University
Press, New York 1966Wahren, W.: Anatomie des Hypothalamus. In:Einführung in die stereotaktischen Operationenmit einem Atlas des menschlichen Gehirns, hrsg.von G. Schaltenbrand, P. Bailey. Thieme, Stuttgart1959
Walker, A. E.: The Primate Thalamus. University ofChicago Press, Chicago 1938
Walker, A. E.: Normale und pathologische Physio-logie des Thalamus. In: Einführung in diestereotaktischen Operationenmit einemAtlas desmenschlichen Gehirns, hrsg. von G. Schalten-brand, P. Bailey. Thieme, Stuttgart 1959
Wolstenholme, G. E. W., J. Knight: The Pineal Gland.Ciba Foundation Symposium Churchill-Livings-tone, London 1971
Wurtman, R. J., J. A. Axelrod, D. E. Kelly: The Pineal.Academic Press, New York 1968
TelencephalonAlajouanine, P. Th.: Les grandes activitées du lobetemporal. Masson, Paris 1955
v. Bonin, G.: Die Basalganglien. In: Einführung in diestereotaktischen Operationenmit einemAtlas desmenschlichen Gehirns, hrsg. von G. Schalten-brand, P. Bailey. Thieme, Stuttgart 1959
Braak, H.: Architectonics of the Human Telen-cephalic Cortex. Springer, Berlin 1980
Brazier, M. A. B., H. Petsche: Architectonics of theCerebral Cortex. Raven Press, New York 1978
Brodmann, K.: Vergleichende Lokalisationslehre derGroßhirnrinde. Barth, Leipzig 1925
Bucy, P. C.: The Precentral Motor Cortex. Universityof Illinois Press, Urbana 1949
Chan-Palay, V., C. Köhler (eds.): The Hippocampus -New Vistas. Neurology and Neurobiology. Vol.52. Alan R. Liss, Inc., New York 1989
Ciba Foundation Symposium 58: Functions of theSepto-Hippocampal System. Elsevier, Amsterdam1977
Creuzfeldt, O. D.: Cortex Cerebri. Springer, Berlin1983
Critchley, M.: The Parietal Lobes. Arnold, London1953
Denny-Brown, D.: The Basal Ganglia. Oxford Univer-sity Press, London 1962
Descarries, L., T. R. Reader, H. H. Jasper: MonoaminInnervation of the Cerebral Cortex. Alan R. Riss,New York 1984
Dimond, St.: The Double Brain. Churchill-Livings-tone, Edinburgh 1972
Divac, I. R., G. E. Öberg: The Neostriatum. Pergamon,Oxford 1979
Eccles, J. C.: Brain and Conscious Experience.Springer, Berlin 1966
v. Economo, C., G. N. Koskinas: Die Cytoarchitek-tonik der Hirnrinde des erwachsenen Menschen.Springer, Berlin 1925
Eleftheriou, B. E.: The Neurobiology of the Amyg-dala. Plenum, New York 1972
Feremutsch, K.: Basalganglien. In: Primatologia,Bd. II/2, hrsg. von H. Hofer, A. H. Schultz, D. Starck.Karger, Basel 1961
Freund, T. F., G. Buzsáki (eds.): Interneurons of theHippocampus. Hippocampus 6 (1996) 347–473
Frotscher, M., P. Kugler, U. Misgeld, K. Zilles: Neu-rotransmission in the Hippocampus. Advances inAnatomy, Embryology and Cell Biology, Vol. 111,Springer, Berlin, Heidelberg 1988
Fuster, J. M.: The Prefrontal Cortex. Raven, New York1980
Gainotti, G., C. Caltagirone: Emotions and the DualBrain. Springer, Berlin 1989
Gastaud, H., H. J. Lammers: Anatomie du rhinen-céphale. Masson, Paris 1961
Goldman, P. S., W. J. Nauta: Columnar distribution ofcortico-cortical fibers in the frontal association,
Further Reading
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

388
limbic and motor cortex of the developing rhesusmonkey. Brain Res. 122, 393–413 (1977)
Goodwin, A. W., J. Darian-Smith: Handfunction andthe Neocortex. Springer, Berlin 1985
Hubel, D. H., T. N. Wiesel: Anatomical demonstra-tion of columns in the monkey striate cortex. Na-ture (Lond.) 221 (1969) 747–750
Isaacson, R. L., K. H. Pribram: The Hippocampus.Plenum, New York 1975
Jones, E. G., A. Peters: Cerebral Cortex. Vol.1–6. Plenum, New York 1984–1987
Kahle,W.: Die Entwicklung dermenschlichen Groß-hirnhemisphäre. Springer, Berlin 1969
Kennedy, C., M. H. Des Rosiers, O. Sakurada, M.Shinohara, M. Reivich, J. W. Jehle, L. Sokoloff: Met-abolic mapping of the primary visual system ofthe monkey by means of the autoradiographicC14deoxyglucose technique. Proc. nat. Acad. Sci.(Wash.) 73 (1976), 4230–4234
Kinsbourne, M., W. L. Smith: Hemispheric Discon-nection and Cerebral Function. Thomas, Spring-field 1974
Passouant, P.: Physiologie de le hippocampe. Editiondu centre national de la recherche scientifique.Paris 1962
Penfield, W., H. Jasper: Epilepsy and the FunctionalAnatomy of the Human Brain. Little, Brown,Boston 1954
Penfield, W., T. Rasmussen: The Cerebral Cortex ofMan. Macmillan, New York 1950
Penfield, W., L. Roberts: Speech and Brain Mecha-nisms. Princeton University Press. Princeton 1959
Ploog, D.: Die Sprache der Affen. In: Neue Anthropo-logie, hrsg. von H. G. Gadamer, P. Vogler. Thieme,Stuttgart 1972
Rose, M.: Cytoarchitektonik und Myeloarchitek-tonik der Großhirnrinde. In: Handbuch der Neu-rologie, Bd. I, hrsg. von O. Bumke, O. Foerster.Springer, Berlin 1935
Sanides, F.: Die Architektonik des menschlichenStirnhirns. Springer, Berlin 1962
Schneider, J. S., T. J. Lidsky: Basal Ganglia and Be-havior. Huber, Bern 1987
Schwerdtfeger, W. K.: Structure and Fiber Connec-tions of the Hippocampus. Springer, Berlin 1984
Seifert, W.: Neurobiology of the Hippocampus. Aca-demic Press, London 1983
Squire, L. R.: Memory and Brain. Oxford UniversityPress, New York 1987
Stephan, H.: Allocortex. In: Handbuch der mik-roskopischen Anatomie, Bd. IV/9, hrsg. von W.Bargmann. Springer, Berlin 1975
Valverde, F.: Studies on the Piriform Lobe. HarvardUniversity Press. Cambridge/Mass. 1965
Weinstein, E. A., R. P. Friedland: Hemiinattentionand Hemisphere Spezialization. Raven, New York1977
Cerebrospinal Fluid SystemHofer, H.: Circumventrikuläre Organe des Zwischen-
hirns. In: Primatologia, Bd. II/2, hrsg. von H. Hofer,A. H. Schultz, D. Starck. Karger, Basel 1965
Lajtha, A., D. H. Ford: Brain Barrier System. Elsevier,Amsterdam 1968
Millen, J. W. M., D. H. M. Woollam: The Anatomy ofthe Cerebrospinal Fluid. Oxford University Press,London 1962
Schaltenbrand, G.: Plexus und Meningen. In: Hand-buch der mikroskopischen Anatomie, Bd IV/2,hrsg. von W. Bargmann. Springer, Berlin 1955
Sterba, G.: Zirkumventrikuläre Organe und Liquor.VEB Fischer, Jena 1969
Cerebrovascular SystemDommisee, G. F.: The Arteries and Veins of the
Human Spinal Cord from Birth. Churchill-Livings-tone, Edinburgh 1975
Hiller, F.: Die Zirkulationsstörungen des Rücken-marks und Gehirns. In: Handbuch der Neurologie,Bd. III/11, hrsg. von O. Bumke, H. Foerster.Springer, Berlin 1936
Kaplan, H. A., D. H. Ford: The Brain Vascular System.Elsevier, Amsterdam 1966
Krayenbühl, H., M. G. Yasargil: Die zerebrale Angiog-raphie,2. Aufl.Thieme,Stuttgart1965;3. Aufl.1979
Luyendijk; W.: Cerebral Circulation. Elsevier,Amsterdam 1968
Szilka, G., G. Bouvier, T. Hovi, V. Petrov: Angiographyof the Human Brain Cortex. Springer, Berlin 1977
Autonomic Nervous SystemBurnstock, G., M. Costa: Adrenergic Neurons, Chap-
man & Hall, London 1975Csillik, B., S. Ariens Kappers: Neurovegetative Trans-mission Mechanisms. Springer, Berlin 1974
Furness, J. B., M. Costa: The enteric nervous system.Churchill Livingstone, Edinburgh 1987
Gabella, G.: Structure of the Autonomous NervousSystem. Chapman & Hall, London 1976
Kuntz, A.: The Autonomic Nervous System. Lea & Fe-biger, Philadelphia 1947
Langley, J. N.: The autonomic nervous system, part1.Heffer, Cambridge 1921
Mitchell, G. A. G.: Anatomy of the AutonomicNervous System. Livingstone, Edinburgh 1953
Newman, P. P.: Visceral Afferent Functions of theNervous System. Arnold, London 1974
Pick, J.: The Autonomic Nervous System. Lippincott,Philadelphia 1970
Thews, G., G. Vaupel.: Vegetative Physiologie.Springer, Berlin 1990
White, J. C., R. H. Smithwick: The AutonomicNervous System. Macmillan, New York 1948
Further Reading
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

389
Functional SystemsAdey, W. R., T. Tokizane: Structure and Function ofthe Limbic System. Elsevier, Amsterdam 1967
Andres, K. H., M. v. Dühring: Morphology of cuta-neous receptors. In: Handbook of SensoryPhysiology, Bd. II, hrsg. von H. Autrum, R. Jung, W.R. Loewenstein, D. M. MacKay, H. L. Teuber.Springer, Berlin 1973
Barker, D.: The morphology of muscle receptors. In:Handbook of Sensory Physiology. Bd. III/2, hrsg.von H. Autrum, R. Jung, W. R. Loewenstein, D. M.MacKay, H. L. Teuber. Springer, Berlin 1974
Campbell, H. J.: The Pleasure Areas. Eyre Methuen,London 1973
Couteaux, R.: Motor endplate structure. In: TheStructure and Function of Muscle, hrsg. von G. H.Bourne. Academic Press, New York 1973
Douek, E.: The Sense of Smell and Its Abnormalities.Churchill-Livingstone, London 1974
Gardner, H.: Dem Denken auf der Spur, der Weg derKognitionswissenschaft. Klett-Cotta, Stuttgart 1989
Halata, Z.: The Mechanoreceptors of the Mamma-lian Skin, Advances in Anatomy, Embryology andCell Biology, Bd. 50/5. Springer, Berlin 1975
Isaacson, R. L.: The Limbic System. Plenum Press,New York 1974
Janzen, R., W. D. Keidel, G. Herz, C. Steichele:Schmerz. Thieme, Stuttgart 1972
Jung, R., R. Hassler: The extrapyramidal motor sys-tem. In: Handbook of Physiology, Section 1, Bd. 2,hrsg. von J. Field, H. W. Magoun, V. E. Hall. Ameri-can Physiological Society Washington 1960
Knight, J.: Mechanisms of Taste and Smell in Verte-brates. Ciba Foundation Symposium. Churchill,London 1970
Lassek, A. M.: The Pyramidal Tract. Thomas, Spring-field/Ill. 1954
Monnier, M.: Functions of the Nervous System,Bd. 3: Sensory Functions and Perception. Elsevier,Amsterdam 1975
Munger, B. L.: Patterns of organization of peripheralsensory receptors. In: Handbook of Sensory Physi-ology, Bd. I/1, hrsg. von H. Autrum, R. Jung, W. R.Loewenstein, D. M. MacKay, H. L. Teuber. Springer,Berlin 1971
de Reuk, A. V., S. J. Knight: Touch, Heat and Pain. CibaFoundation Symposium. Churchill, London 1966
Sezntágothai, J., J. Hámori, M. Palkovits: RegulatoryFunctions of the CNS: Motion and OrganizationPrinciples. Pergamon, Oxford 1981
Wiesendanger, M.: The pyramidal tract. In: Ergeb-nisse der Physiologie, Bd. 61. Springer, Berlin 1969
Zacks, S. J.: The Motor Endplate. Saunders, Philadel-phia 1964
Zotterman, Y.: Olfaction and Taste. Pergamon, Ox-ford 1963
Zotterman, Y.: Sensory Mechanisms. Elsevier,Amsterdam 1966
Zotterman, Y.: Sensory Functions of the Skin. Perga-mon, Oxford 1977
The EyeCarpenter, R. H. S.: Movements of the Eyes. Pion,London 1977
Fine, B. S.,M. Yanoff: Ocular histology. Harper & Row,New York 1972
Hollyfield, J. G.: The Structure of the Eye. Elsevier,Amsterdam 1982
Hubel, D. H., T. N. Wiesel: Die Verarbeitung visuellerInformationen. Spektrum der Wissenschaft,November 1979
Livingston, M., D. Hubel: Segregation of form, color,movement, and depth: Anatomy, physiology andperception. Science 240 (1988) 740–749
Masland, R. H.: Die funktionelle Architektur derNetzhaut. Spektrum der Wissenschaft, 66–75,Februar 1989
Marr, D.: Vision. Freeman, San Francisco 1982Polyak, St.: The Vertebrate Visual System. University
Chicago Press, Chicago 1957Rodieck, R. W.: Vertebrate Retina. Freeman, San
Francisco 1973Straatsma, B. R., M. O. Hall, R. A. Allen, F. Crescitelli:
The Retina Morphology, Function and ClinicalCharacteristics. University California Press, Berke-ley 1969
Wässle, H., B. B. Boycott: Functional architecture ofthe mammalian retina. Physiol. Rev. 71 (1991)447–480
Walsh, F. W., W. F. Hoyt: Clinical Neuroophthalmol-ogy. Williams & Wilkins, Baltimore 1969
Warwick, R.: Wolff’s Anatomy of the Eye and Orbit.Lewis, London 1976
Zeki, S.: AVisionof theBrain. Blackwell, London1993
Organ of Hearing and BalanceAdes, H. W., H. Engström: Anatomy of the inner ear.In: Handbook of Sensory Physiology, Bd. V/1, hrsg.von H. Autrum, R. Jung, W. R. Loewenstein, D. M.MacKay, H. L. Teuber. Springer, Berlin 1974
Brodal, A., O. Pompeiano, F. Walberg: The VestibularNuclei and Their Connexions. Oliver & Boyd, Edin-burgh 1962
Gualtierotti, T.: The Vestibular System. Springer,Berlin 1981
Kolmer, W.: Gehörorgan. In: Handbuch der mik-roskopischen Anatomie, Bd. III/1, hrsg. von W. v.Moellendorff. Springer, Berlin 1927
Precht, W.: Neuronal Operations in the VestibularSystems. Springer, Berlin 1978
Rasmussen, G. L., W. F. Windle: Neural Mechanismsof the Auditory and Vestibular Systems. Thomas,Springfield/Ill. 1960
deReuck, A. V. S., J. Knight: Hearing Mechanisms inVertebrates. Ciba Foundation Symposium,Churchill, London 1968
Whitfield, I. C.: The Auditory Pathway. Arnold, Lon-don 1960
Further Reading
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

390
Index
Page numbers in bold indicate extensive coverage of the subject
AAccommodation, 358, 359negative, 358positive, 358
Acetylcholine (ACh), 26, 148,294, 312
nicotinic receptor, 30, 31Acetylcholinesterase, 28, 148Adaptationlight–dark, 358near–far, 358
Adenohypophysis, 200Adhesion, interthalamic, 10,
174Adrenergic system, 294Agnosia, 250
visual, 334Agraphia, 262Alexia, 262Allocortex, 244Alveus, 230, 232, 234Alzheimer’s disease, 32Ampullae, membranous, 368Amusia, 262Amygdala, 174, 176, 216,
226–229subnuclei, 226
Analgesia, 68Anesthesia, 74dissociated, 68
Angleiridocorneal, 344medial, of the eye, 338
Annulus, fibrocartilaginous,362
Ansacervical, 72
deep, 70, 72, 112superficial, 122
lenticular, 174, 192subclavian, 296
Antrum, mastoid, 364, 366Aortic arch, 116Aperturelateral, 100, 282median, 100, 282
Aphasiamotor, 248sensory, 248, 262
Apparatuslacrimal, 338, 339
vestibular, 362, 374Aqueduct, 218cerebral (of Sylvius), 10, 132,
134, 280Arachnoidea mater, 288spinal, 64
Arbor vitae, 10, 154Archicerebellum, 152Archicortex, 208, 230–235,
332rudimentary, 332
Archipallium, 208–210Architectonics, 244Area(s)association, 210auditory
primary, 380secondary, 380
Broca’s motor speech, 248,249
cortical, 244, 245dorsocaudal, 194entorhinal, 224, 228, 232,
244motor, 180, 248, 249motosensory, 246, 248of origin, 210olfactory, 172, 176, 212, 228optic integration, 254orbitofrontal, 246parolfactory, 332periamygdalar, 224, 228postcentral, 250, 251postremal, 44, 286, 287precentral, 246, 247, 248,
249motor, 180
prefrontal, 246premotor, 246preoptic, 228prepiriform, 224, 228, 238projection, 244sensorimotor, 246, 248sensory postcentral, 180septal, 332, 334, 335striate, 220, 222, 254, 354,
356subcallosal, 212termination, 210vestibular, 100
Artery(ies), 44, 270–275basilar, 270, 274
callosomarginal, 272carotid, internal, 104, 270,
272, 273, 366central, 272cerebellar
inferioranterior, 270posterior, 270
superior, 270cerebral
anterior, 270, 272, 274middle, 270, 272, 274posterior, 270, 272, 274
choroidalanterior, 270, 272, 274posterior, 274
ciliary, 346anterior, 346posterior
long, 346short, 346
communicatinganterior, 270posterior, 270, 274
frontobasallateral, 272medial, 272
hyaloid, 342hypophysial
inferior, 200, 272superior, 200, 270, 272
insular, 272intercostal, 60labyrinthine, 270lumbar, 60ophthalmic, 270, 272, 346paracentral, 272pericallosal, 272pontine, 270radicular, 60
large, 60recurrent artery of Heubner,
270, 274retinal, central, 346segmental, 60spinal
anterior, 60posterior, 60
subclavian, 116, 270sulcocommissural, 60temporal, 272trabecular, 200
A Accommodation
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

391
vertebral, 60, 270spinal branches, 60
Astrocytes, 42, 43, 44, 158fibrous, 42, 43protoplasmic, 42, 43
Astroglia, 42Auricle, 362, 363Autonomic nervous system, 2,
292–303central, 292peripheral, 294, 300–303
Axolemma, 36Axon(s), 18, 36collaterals, 18corticofugal, 162hillock, 18terminals, 18, 24transport, 28
Axoplasm, flow of, 28, 29
BBalance, 362, 382
organ of, 374, 375see also Ear
BandBaillarge’s, 240diagonal, of Broca, 224, 332,
334Giacomini’s, 224Vicq d’Azyr’s, 174, 182, 194,
196Barr body, 18Basal membrane, 62Bladder, urinary, 298Blind spot, 346, 356Blood vessels, 44, 45, 270–279,
288eye, 346, 347hypophysis, 200, 201spinal cord, 60, 61visualization, 264
Blood–brain barrier, 44, 286Blood–cerebrospinal fluid
barrier, 44Bochdalek’s flower basket, 282Body(ies)
amygdaloid, 174–176, 196,216, 217, 226–229,236, 332
subnuclei, 226basal, 284cavernous, 96cerebellar, 152
anterior lobe, 152posterior lobe, 152
ciliary, 342, 344, 345geniculatelateral, 178–180, 186, 187,
190, 218, 256, 354–356
medial, 132–134, 178–180, 186, 187, 190,380, 381
Herring, 204, 205Luys’, 174, 192mamillary, 10–12, 134, 144,
170, 174, 175, 178, 194,198, 202–204, 216,230–232, 332
peduncle, 196Nissl, 18, 22, 156orbital fat, 338pacchionian, 288pineal, 176restiform, 100, 108, 120, 154,
164subthalamic, 216tigroid, 18trapezoid, 110, 120, 378
anterior nucleus, 378posterior nucleus, 378
vitreous, 342Boutons, 18, 19, 24
see also SynapsesBrachium conjunctivum, 100,
154, 166Brachium pontis, 100, 110, 154,
166Brain, 4, 6–15
age-related atrophy, 8axes, 4, 5
Forel’s, 4Meinert’s, 4
development, 6, 7, 14divisions, 4, 6emotional, 332evolution, 14, 15function, 306, 307gyrencephalic, 210lissencephalic, 210mantle, 208olfactory, 224visceral, 332weight, 8
Brain stem, 4–12, 32, 100, 101,132, 136, 180
divisions, 4histochemistry, 148–149
Brown–Séquard’s syndrome,68, 69
BulbKrause’s end, 320
olfactory, 6, 12, 32, 102–104,176, 208–212, 224,225, 228, 229, 330
pontine, 110, 132Bundle
Arnold’s, 166forebrain, medial, 196Meynert’s, 132, 176papillomacular, 350Schütz’s, 108, 110, 134, 144,
196, 328Türck’s, 166
CCalcar avis, 254Calyx, nerve, 376Canal(s)
Alcock’s, 96carotid, 104, 366central, 8facial, 122, 366hypoglossal, 104, 112mandibular, 126musculotubal, 366outer ear, 362pterygoid, 122, 128pudendal, 96Schlemm’s, 344semicircular, 368
lateral, 366spiral, of the cochlea, 368vertebral, 48
Canaliculus(i)ependymal, 286lacrimal, 338mastoid, 114tympanic, 118
Capillaries, 44, 45endothelium, 44
Capsuleexternal, 214extreme, 214internal, 174, 192, 214, 222,
258, 259Carpal tunnel syndrome, 76Cartilage, tubular, 362Caruncle, lacrimal, 338Catecholamines, 26, 148Cauda equina, 48, 64, 65Cave, trigeminal, 288Cavity
subarachnoidal, 342trigeminal, 104tympanic, 118, 362, 364–367
promontory, 366
Cavity C
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

392
Cecumcupular, 368vestibular, 368
Cell(s)amacrine, 348anterior horn, 316axo-axonal, 32basal, 326basket, 156, 158, 159, 160,
234, 242GABAergic, 32
Bergmann’s glial, 158Betz’s, 246Cajal–Retzius, 242complex, 254cone cells, retina, 348–352endothelial, 40, 44Fa[ntilde]anás, 158ganglion, 354geniculate, 354glitter, 42Golgi, 156, 158, 159, 160granule, 158, 159, 160, 228,
234, 240hair
inner, 372, 373outer, 372, 373vestibular, 376, 377
horizontal, 348hypercomplex, 254Martinotti’s, 242mastoid (air), 362, 364Merkel’s touch, 318, 319Meynert’s, 254mitral, 228, 330nucleus, 18, 22periglomerular, 228phalangeal
inner, 372outer, 372
pillarinner, 372outer, 372
Purkinje, 156, 157, 158–160pyramidal, 232–234, 240–
242, 246giant, 246
receptor, 330Renshaw, 52rod cells, retina, 348–352satellite, 42, 62scavenger, 42Schwann, 36–38, 40, 302simple, 254stellate, 156, 158, 160, 240,
254supporting, 326, 330, 374
Deiters’, 372glial, of Müller, 348, 350
taste, 326tufted, 228tympanic, 366
Centercardiovascular control, 146,
147ciliospinal, 128depressor, 146inhibition, 146respiratory control, 146, 147semioval, 220speech (Wernicke’s), 248,
262Central gray, 132, 134Central nervous system, 2, 3Cerebellum, 4–12, 32, 100,
152–167, 210, 220–222, 310
cortex, 156–159tentorium, 288vermis, 12, 152, 153, 162
Cerebrospinal fluid (CSF), 4, 8,48, 264, 280, 284
circulation, 280Cerebrum, 4
falx, 288Chemoarchitectonics, 148Chemoreceptors, 118, 326Chiasm, 176
optic, 10–12, 102, 170–172,178, 194, 202–204,214, 228, 354, 355
Choline acetyltransferase, 28Cholinergic system, 294Chorda dorsalis, 4, 100Chorda tympani, 122, 130, 364
communicating branch, 130Choroidea, 338, 342, 346, 350Cilium(a)
connecting, 352olfactory, 330
Cingulum, 232, 260Circle
arterial, of the irisgreater, 344, 346lesser, 346
of Willis, 270Cisterna ambiens, 280Cistern(s), 280
cerebellomedullary, 280chiasmatic, 280interpeduncular, 280marginal, 22subarachnoid, 280
Claustrum, 214–218, 222, 226
Clawhand, 78Cleft(s)
subneural, 312synaptic, 24
Clivus, 104Cochlea, 362, 368, 370, 371
frequency analysis, 370osseous, 368
Collaterals, Schaffer, 234Colliculus(i)
facial, 100, 110inferior, 100, 132, 378, 379,
380brachium, 186
superior, 100, 134–136, 144Column(s)
anterior, 50anterolateral, 50Burdach’s, 322Clarke’s, 164color, 256dorsal, 50Goll’s, 322lateral, 50ocular dominance, 256orientation, 256posterior, 50projection, 256ventral, 50vertical, 240, 241
Commissure, 196anterior, 10, 172–174, 178,
194, 214, 215, 222,223, 228, 229, 230
epithalamic, 134, 176Gudden’s, 380habenular, 176of the fornix, 232posterior, 176Probst’s, 378supraoptic, inferior, 380supramamillary, 134white, 50
Computed tomography (CT),264–266
Conductionsaltatory, 40velocity, 40
Cone(s)medullary, 48retinal, 348–352
Conjunctiva, 338Connective tissue, 64
endoganglionic, 62Contrast radiography, 264, 265Convergence, 358, 359Convolutions, 212
C Cecum
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

393
Heschl’s, 180, 186, 212, 216,252, 253
Cord(s)lateral, 70, 71, 74–76medial, 70–74, 78–80posterior, 70–74, 80–82vocal, 117see also Spinal cord
Cornea, 342, 344, 345Corona radiata, 178, 258Corpus callosum, 6, 10, 170–
174, 178, 208, 214–218, 220, 221, 222,258–260
development, 7rostrum, 260splenium, 218–222, 260transection, 262
Corpus striatum, 180Corpuscles
bulboid, 320Golgi–Mazzoni, 320lamellar, 320Meissner’s tactile, 318, 319Ruffini’s, 320, 321Vater–Pacini, 320, 321
Cortexagranular, 244, 246auditory, 180, 210, 252, 253,
380, 381cerebellar, 156–159, 160,
161granular layer, 158molecular layer, 156Purkinje cells, 156, 157
cerebral, 10, 32entorhinal, 224frontal, 136granular, 244, 246, 250,
252hippocampal, 234, 235homogenetic, 244insular, 238, 239, 244interpretative, 252isogenetic, 244motor, 210, 244–247orbital, 182orbitofrontal, 246periamygdalar, 224, 226polar, 182precentral, 136, 246, 247,
248, 249, 316prefrontal, 246premotor, 180, 182, 246prepiriform, 224retrosplenial, 244sensory, 210
somatosensory, 250striate, 254visual, 180, 210, 222, 244,
254–257see also Area(s)
Crestampullary, 368, 374, 375, 382neural, 62
Crown, ciliary, 344Culmen, 152Cuneus, 180, 212Cup, optic, 342Cupula, 368, 374Cytoarchitectonics, 244Cytoplasm, 22, 36
DDeclive, 152Decussation
Forel’s, 134inferior, 134medial lemnisci, 322Meynert’s, 134pyramidal, 58, 100, 308superior cerebellar peduncle,
132, 166, 218superior tegmental, 134
Dendrite(s), 18apical, 240, 241
Dendritic tree, 156, 160Dermatomes, 66, 67Diaphragm
paralysis of, 72sellar, 288urogenital, 96
Dictyosomes, 22Diencephalon, 4–12, 170–205,
220, 221Digital subtraction angiogra-
phy (DSA), 264, 265Disinhibition, 34Disk
ciliary, 344optic, excavation of, 346prolapse, 66
Dopamine (DA), 26, 148, 149Duct(s)
cochlear, 368, 370, 373endolymphatic, 368nasolacrimal (tear), 338perilymphatic, 368semicircular, 120, 362, 368,
374, 376anterior, 368lateral, 368
posterior, 368uniting, 368utriculosaccular, 368
Dura mater, 104, 288, 289spinal, 64
Dynein, 28
EEar, 362–383
external, 115, 362, 363inner, 362, 363, 368–377middle, 118, 119, 362, 363,
364–367vestibule, 368
Eardrum, 362–368Embolism, 274Eminence
hypothenar, 78mamillary, 6medial, 100pyramidal, 366thalamic, 188thenar, 76, 78
Encephalon, 4Endolymph, 368Endoneural sheath, 40Endoneurium, 40Endoplasmic reticulum
rough, 18–22smooth, 22
Endorhachis, 64Enlargement
cervical, 48, 54lumbar, 48, 54
Enophthalmos, 296Enzyme pattern, 148Ependyma, 284, 285, 286Epicanthus, 338Epineurium, 40, 64Epiphysis, 10, 170, 176
cerebri, 176Epithalamus, 172, 176, 177Epithelium
neural, 348olfactory, 102, 330, 331plexus, 282, 284, 285
Erb’s palsy, 74Evoked potentials, 162Extrapyramidal motor system,
310, 311Eye, 338–359
chambers, 342anterior, 342, 344, 345posterior, 342
equator of, 342
Eye (-e) E
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

394
medial angle of, 338muscles, 102, 106, 340–341
nerves, 138, 139oblique
inferior, 340superior, 340, 341
rectusinferior, 340lateral, 340medial, 340superior, 340, 341
Eyeball, 338, 342, 343movements, 340
Eyelids, 338anterior margin, 338lower, 338posterior margin, 338upper, 338
FFascia
dentata, 230, 232lata, 90
Fasciclelateral, 74medial, 74
longitudinal, 58posterior, 74
Fasciculus(i)cuneatus, 56, 57, 322dentatorubral, 134, 136gracilis, 56, 57, 322degeneration, 58, 59
interstitiospinal, 142, 310,316
lenticular, 174, 192longitudinal, 144
dorsal, 196inferior, 260medial, 108–110, 132–
134, 142, 143posterior, 108–110, 134,
144, 145, 328superior, 260
mamillotegmental, 194, 196,332
mamillothalamic, 174, 194,196
occipital, vertical, 260occipitofrontal
inferior, 260superior, 260
of Burdach, 56of Goll, 56of Spitzer, 140
orbitofrontal, 260pallidohypothalamic, 196proprii, 52, 53reticulothalamic, 146retroflexus, 132rubro-olivary, 144strionigral, 136subcallosal, 260tegmental, anterior, 140telencephalic, medial, 196thalamic, 174, 184, 192uncinate, 164, 260
Fiber(s), 36, 37amygdalofugal, 228
ventral, 332arcuate, 260
internal, 108superficial, 108
association, 240, 242, 258,260, 261
centrostriate, 236cerebellofugal, 316circular, 344climbing, 156, 160, 161commissural, 240, 258, 260,
261cortical, 310corticonigral, 136corticonuclear, 140, 141, 308corticopontocerebellar, 162corticostriate, 236cuneate, 140, 322Déjérine, 140exteroceptive, 114extragemmal, 326!-fibers, 314gracile, 322heterotopic, 260homotopic, 260intragemmal, 326lens, 342meridional, 344mossy, 158–161, 234motor, 114–118, 122, 130muscle
extrafusal, 314intrafusal, 314
nigropallidal, 192nigrostriate, 136, 236nuclear bag, 314, 315nuclear chain, 314, 315olivocerebellar, 108, 144pallido-olivary, 144pallidosubthalamic, 192parallel, 156–160pontine, transverse, 110projection, 258, 259
radial, 240Reissner’s, 286reticulo-olivary, 144reticuloreticular, 144rubro-olivary, 136, 144rubroreticular, 136secretory, 292somatomotor, 70somatosensory, 70specific, 242spinocerebellar, 162strionigral, 236subthalamopallidal, 192taste, 114, 118, 122, 128, 328,
329primary, 328secondary, 140, 328tertiary, 328
unmyelinated, 38vestibular, 310vestibulocerebellar, 162visceroefferent, 118, 122visceromotor, 70, 114, 116,
292viscerosensory, 70, 114, 118,
292, 300zonular, 344see also Axon(s)
Fieldcortical, extrapyramidal, 310eye
frontal, 248, 249, 254occipital, 254
Flechsig’s oval, 56Forel’s, 174, 192lateral, 194of projection, 180receptive, 350
Fila olfactoria, 330Filament, terminal, 48, 64
dural, 64Fimbria, 230, 232, 234Fissure
anterior median, 48, 100cerebral, longitudinal,
10–12, 212–216, 220–224
choroid, 282lateral, 8of Sylvius, 8, 238orbital, 104
superior, 138palpebral, 338posterolateral, 152telodiencephalic, 170tympanomastoid, 114see also Sulcus
E Eye (-m)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

395
Flocculus, 152, 154Fold
mallearanterior, 364posterior, 364
palpebronasal (mongolian),338
Folium(a), 152, 157Foramen, 4
ethmoidal, anterior, 124infrapiriform, 96interventricular, 8–10, 178,
276, 280, 286intervertebral, 4, 48jugular, 112, 114, 118lacerum, 122, 128magnum, 64, 104mental, 126of Key and Retzius, 280of Luschka, 100, 280, 282of Magendie, 100, 280, 282of Monro, 8–10, 178, 276,
280, 286oval, 104, 126round, 104, 126sciatic, lesser, 96stylomastoid, 122
Forcepsmajor, 254, 260minor, 260
Forebrain, 4, 32Formation
hippocampal, 332reticular, 32, 52, 108, 146,
147, 182, 292, 310, 316pontine, 316
Fornix, 10, 32, 170–174, 192–196, 208, 216–222,230, 232, 332
conjunctival, 338postcommissural, 232precommissural, 232
Fossacranial
anterior, 104middle, 104posterior, 104
infratemporal, 126interpeduncular, 100, 102ischioanal, 96jugular, 104, 366lateral, 8–10, 214–218, 222,
238of Sylvius, 214popliteal, 94primary, 152pterygopalatine, 126
rhomboid, 100, 101, 107, 120Fovea, central, 346, 350, 351,
356Fundus, 346, 347Funiculus(i)
anterior, 50, 100tracts, 56, 58
cuneate, 322gracile, 322lateral, 50, 58
tracts, 56, 58posterior, 50, 100, 108, 323
pathways, 56, 57Funnel, pial, 288
GG proteins, 30Gamma-aminobutyric acid
(GABA), 26, 160receptors, 30
Ganglion(a), 4autonomic, 292basal, 216celiac, 292, 294, 298cervical
inferior, 296middle, 294, 296superior, 128, 292–296
cervicothoracic, 292, 294ciliary, 124, 128, 129, 130,
358extramural, 296Gasser’s, 102, 124, 288geniculate, 122, 328glossopharyngeal nerveinferior, 118superior, 118
intramural, 296–301jugular, 114mesenteric
inferior, 292, 294, 298superior, 292, 294, 298
nodose, 114, 328otic, 118, 130, 131parasympathetic, 128–131petrosal, 118, 328prevertebral, 296pterygopalatine, 122, 128,
129, 130semilunar, 102, 124, 288spinal, 44, 48, 49, 62, 63, 292
development, 62, 63spiral, 120, 370, 376stellate, 292–296submandibular, 130, 131
sympathetic trunk, 296terminal, 296trigeminal, 102–104, 122–
130, 288vagus nerveinferior, 114superior, 102, 114
vestibular, 120, 164, 376Gap junctions, 26, 302Geniculum, lateral, 190Genitals, 298Genu
corpus callosum, 220, 222,260
facial nerveexternal, 122internal, 106, 110, 122
internal capsule, 258occipital, 354temporal, 258, 354
Gland(s)Bowman’s, 330ceruminous, 362lacrimal, 124, 128, 338meibomian, 338olfactory, 330parotid, 106, 122, 130pineal, 10, 172, 176, 177, 220pituitary, 200
see also Hypophysissublingual, 106, 130submandibular, 106, 130tarsal, 338
Glia, 18, 42, 43, 158, 159Bergmann’s glial cells, 158intrafascicular, 42of Fa[ntilde]anás, 158
Glial feet, perivascular, 44, 45Globus pallidus, 172–174, 182–
184, 192, 214–216,222
see also PallidumGlomerulus(i), 62, 228
cerebellar, 158olfactory, 330
Glomus, carotid, 118Glutamate, 26, 32
receptors, 30Glycine, 26Goldmann’s experiments, 44,
45Golgi complex, 22Granulations, arachnoid, 280,
288Granules
chromatin, 22meningeal, 288
Granules G
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

396
Gray matter, 12, 13intermediate, 52spinal cord, 50–54
Gyrus(i), 10, 212ambient, 224angular, 212, 218, 250, 262cingulate, 32, 180–182, 212–
214, 218, 332, 334isthmus, 212
dentate, 212, 224, 230–234ectosylvian, posterior, 380frontal
inferior, 212, 214middle, 212, 214superior, 212, 214
limbic, 212, 332lingual, 212occipitotemporal
lateral, 212middle, 212
orbital, 212paracentral, 212parahippocampal, 212, 218,
222–224, 230–232,332
paraterminal, 212, 224, 332postcentral, 10, 212, 216,
220, 250precentral, 10, 184, 212, 216,
220rectus, 212semilunar, 224supramarginal, 212, 250temporal
inferior, 212, 214middle, 212, 214, 252superior, 212, 214transverse, 180, 186, 212,
216–218, 252, 253
HHabenula(e), 170–172, 176,
177, 220Hair follicles, roots, 318Hairs
olfactory, 330organ of Corti, 372
Hallucinations, 252Hand of oath, 76Helicotrema, 370Hematoma, subdural, 276Hemiatrophy, 112Hemiballismus, 192Hemidepersonalization, 250Hemispheres
cerebellar, 12, 152, 162cerebral, 4–10, 170, 208, 212,
213asymmetry, 262, 263dominant, 262rotation, 208, 209
Hiatus, diaphragmatic, 288Hindbrain see Rhomben-
cephalonHippocampus, 32, 174, 208–
210, 216–222, 230Histological techniques, 20Homunculus
motor, 248sensory, 250
Hormonegonadotropic, 202luteotropic, 202thyrotropic, 202
HornAmmon’s, 208, 218, 222, 230,
231, 232, 233, 234anterior, 50–54, 214, 222,
280, 316dorsal, 50frontal, 280inferior, 230, 280lateral, 52, 54occipital, 280posterior, 50–54, 222, 230,
280temporal, 280ventral, 50
Humor, aqueous, 342, 344Hyperacusis, 122Hyperalgesia, 66Hypoesthesia, 66, 68, 74Hypophysial stalk, 102, 198–
202Hypophysis, 10–12, 104, 176,
198, 200, 201, 202blood vessels, 200, 201
Hypothalamohypophysial sys-tem, 204, 205
Hypothalamus, 32, 172–174,194–201, 202, 216,226–228, 232, 292
functional topography, 198,199
oral, 144
IImmunocytochemistry, 20Incus, 364Indusium griseum, 208, 220,
230
Infundibulum, 174, 194, 200,201, 216
Inhibitionfeed-forward, 34postsynaptic, 34recurrent, 34
Insula, 6, 8, 208, 238, 239stimulation responses, 238,
239Interneurons, 18, 32, 242Iris, 342, 344Iron, 148, 149Isocortex, 238, 240, 244
KKaryoplasm, 22Kinesin, 28, 29Kinetosomes, 284Kinocilia, 376Klumpe’s palsy, 74Klüver–Bucy syndrome, 334Koniocortex, 244, 250, 252
LLabyrinth, 362, 368
kinetic, 376membraneous, 368, 369osseous, 368, 369tonic, 376
Lacunae, lateral, 276, 288Lamina
affixa, 170, 171, 174basal, 40, 44choroid, 282cribosa, 104, 350limiting
anterior, 344posterior, 344
medullaryexternal, 178, 188internal, 178, 188
membranous, 362spiral
bony, 370hamulus of, 370limbus of, 372secondary, 370
tectal, 10terminal, 6, 170
vascular organ, 286, 287Larynx, muscles, 116Lemniscus
lateral, 110, 120, 132, 154,190, 378
G Graymatter
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

397
nuclei, 378medial, 108–110, 132–134,
140, 141, 154, 190,322–324, 328
spinal, 140, 324trigeminal, 140, 184, 322–
324Lens, 342, 344, 345
accommodation, 358Leptomeninx, 64, 288, 289Ligand-gated ion channels, 30Ligament(s)
denticulate, 64inguinal, 88palpebral
lateral, 338medial, 338
pectinate, 344spiral, of the cochlea, 370
Limbic system, 332–335Line(s)
choroid, 170, 282Gennari’s, 220, 254period, 36, 38
Lingula, 152, 154Lobe
floculonodular, 152frontal, 8–12, 180–182, 212,
214–216, 220–222,246–249
occipital, 8–10, 212, 220–222, 254–257
olfactory, 12parietal, 8–10, 180, 212, 218,
250, 251piriform, 210, 224temporal, 8–10, 180, 210,
212, 214, 218–222,230, 252, 253
Lobotomy, prefrontal, 246Lobule
biventral, 152, 154central, 152, 154
wing, 152, 154gracile, 152parietal
inferior, 212superior, 212
quadrangular, 152, 154semilunar
inferior, 152, 154superior, 152, 154
simple, 152, 154Locus caeruleus, 32, 100, 132,
146Loop, median, 72, 76Lumbar puncture, 64, 65
Lysosomes, 22
MMacroglia, 42Macula(e), 186, 346, 350, 356,
374, 375, 376adherentes, 24saccule, 368, 374, 382utricle, 368, 374, 382
Magnetic resonance imaging(MRI), 266, 267
Malleus, 364Margin, pupillary, 344Meatus, acoustic
external, 362internal, 104, 120, 122, 376
Mechanoreceptors, 118Medulla oblongata, 6–12, 100,
103, 106, 107, 108,109, 132, 133, 218
Medulla spinalis, 4see also Spinal cord
Membranebasilar, 370, 372Bowman’s, 344Descemet’s, 344limiting
external, 348glial, 42, 44internal, 348
postsynaptic, 24presynaptic, 24Reissner’s, 370reticular, 372statoconic (otolithic), 374tectorial, 372tympanic, 362secondary, 366, 370
Meninges, 288, 289spinal, 64
Meniscus, tactile, 318Mesaxon, 38Mesencephalon, 6, 8, 100, 132,
170see also Midbrain
Mesocortex, 238, 239Metameres, 66Microglia, 42, 43Midbrain, 132–137, 218, 219
see also MesencephalonMitochondria, 22Modiolus, 370Motor end plate, 312, 313Motor system
ecotropic somatomotor, 2
idiotropic visceromotor, 2Motor unit, 312Muscle spindle, 314, 315, 316Muscle(s)
abdominal, 86oblique
external, 84, 86internal, 84
transverse, 84adductor, 90
great, 94long, 90short, 90
anconeus, 82anus, external sphincter, 96autochthonous, 72, 84biceps
armlong head, 74short head, 74
thighlong head, 94short head, 92
brachial, 74brachioradial, 82branchial arch, 102, 114bulbocavernous, 96ciliary, 106, 128, 138coccygeal, 96coracobrachial, 74cremaster, 86cricothyroid, 116deltoid, 80detrusor, 298digastric, 122, 124, 126dilator muscle of the pupil,
128, 344eye-muscles see Eyefibular, 92fingers
abductor, 78extensor, 82
common, 82flexor
deep, 76, 78short, 78superficial, 76
opposing, 78forearm, flexor, 76gastrocnemius, 94gemellus, 90genioglossal, 112geniohyoid, 72, 112gluteus
greatest, 90least, 90middle, 90
Muscle(s) (-g) M
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

398
gracilis, 90head, long, 72hypoglossal, 112iliac, 88infrahyoid, 72infraspinous, 74intercostal, 84
lumbar, 86interosseous, 78, 94ischiocavernous, 96laryngeal, 116
paralysis, 116latissimus dorsi, 74levator, of the upper eyelid,
338, 340levator ani, 96lumbrical, 76, 78, 94
lateral, 94masseter, 126masticatory, 124mylohyoid, 112, 124, 126neck
deep, 72long, 72
obliqueinferior, 138superior, 138
obturatorexternal, 90internal, 90
omohyoid, 72orbicular, of the eye, 338palmar, long, 76pectineal, 88, 90pectoral
greater, 74lesser, 74
perineal, transversedeep, 96superficial, 96
peroneallong, 92short, 92
pharynx, 116constrictor muscles, 116
piriform, 90plantar, 94popliteal, 94pronator, 76
quadrate, 76round, 76
psoas, 86greater, 86, 88lesser, 86
pterygoid, 126pyramidal, 84quadrate
lumbar, 86thigh, 90
rectorinferior, 138medial, 138superior, 138
rectusabdominis, 84capitis
anterior, 72lateral, 72
femoris, 88lateral, 138
rhomboidgreater, 74lesser, 74
sartorius, 88scalene
anterior, 72medial, 72
scapular, 74semimembraneous, 94semitendinous, 94serratus
anterior, 74, 84posterior
inferior, 84superior, 84
shoulder girdle, 74soleus, 94sphincter
deep, 96external, 96pupillary, 106, 128, 138,
344stapedius, 364, 366sternocleidomastoid, 112sternohyoid, 72sternothyroid, 72styloglossal, 112stylohyoid, 122stylopharyngeal, 118subclavius, 74subscapular, 74supinator, 82supraspinous, 74tarsal
inferior, 338superior, 338
temporal, 126tensor tympani, 124–126,
130, 362–366tensor veli palatini, 124, 126,
130teres
greater, 74lesser, 80
thoracic, transverse, 84thumb
abductor, 78long, 82short, 76
extensorlong, 82short, 82
flexorlong, 76short, 76, 78
opposing, 76thyrohyoid, 72, 112tibial
anterior, 92posterior, 94
toesabductor, 94adductor, 94extensor
long, 92short, 92
flexorlong, 94short, 94
tongue, 106, 112trapezius, 112triceps, 82vastusintermediate, 88lateral, 88medial, 88
wristextensor
long, 82short, 82ulnar, 82
flexorradial, 76ulnar, 78
Myelin sheath, 18, 36–38, 62CNS nerves, 38, 39development, 38, 39period lines, 36, 38peripheral nerves, 40ultrastructure, 36, 37
Myeloarchitectonics, 244Myelography, 264Myotomes, 66
NNeocerebellum, 152Neocortex, 32, 208, 236, 240–
263Neopallium, 208, 210
M Muscle(s) (-g)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

399
Neorubrum, 136Neostriatum, 236, 237, 310Nerve cell see Neuron(s)Nerve fibers see Fiber(s)Nerve(s)
abducens, 102–104, 110,138, 139
accessory, 102–104, 112, 113external branch, 112
afferent, 2alveolar
inferior, 126, 130superior
anterior, 126middle, 126posterior, 126
ampullaranterior, 376lateral, 376posterior, 376
anococcygeal, 96arm, cutaneous
lateralinferior, 82superior, 80
medial, 76, 80, 81, 84posterior, 82
auriculargreater, 70, 72posterior, 122
auriculotemporal, 126, 130axillary, 72, 80, 81branchial arch, 102
motor nuclei, 106buccal, 126, 130caroticotympanic, 118cervical, 48, 49, 72ciliary
long, 124short, 128
clitoris, dorsal nerve, 96cluneal
inferior, 90, 96medial, 84, 96superior, 84
coccygeal, 48, 49cranial, 4, 12, 100, 102, 103,
112–127nuclei, 106, 107
cutaneousdorsal
intermediate, 92lateral, 94medial, 92
surallateral, 92medial, 92, 94
depressor, 116digital
dorsal, 82palmar
common, 76, 78proper, 76, 78
plantarcommon, 94proper, 94
efferent, 2ethmoidal
anterior, 124posterior, 124
facial, 102–104, 110, 122,123, 130, 154, 328
buccal branches, 122cervical branch, 122marginal mandibular
branch, 122paralysis, 122, 123, 366temporal branches, 122zygomatic branches, 122
femoral, 86, 88, 89anterior cutaneous
branches, 88muscular branches, 88
femur, cutaneouslateral, 86, 88, 89posterior, 86, 90
fibularcommon, 92communicating branch, 92
forearmcutaneous
lateral, 74medial, 72, 80, 81posterior, 82
interosseousanterior, 76posterior, 82
frontal, 124genitofemoral, 84, 86, 96femoral branch, 86genital branch, 86
glossopharyngeal, 102–104,114–116, 118, 119,328
lingual branches, 118pharyngeal branches, 118stylopharyngeal branch,
118tonsillar branches, 118
glutealinferior, 90, 91superior, 86, 90, 91
hypoglossal, 72, 102–104,108, 112, 113
arch, 112lingual branches, 112trigon, 100, 112
iliohypogastric, 84, 86, 96ilioinguinal, 84, 86, 96infraorbital, 126infratrochlear, 124intercostal, 84, 85intercostobrachial, 84, 85intermediate, 102, 122, 328labial, posterior, 96lacrimal, 124laryngeal
inferior, 116recurrent, 116
esophageal branches,116
tracheal branches, 116superior, 114, 116
external branch, 116internal branch, 116
leg, interosseous, 94lingual, 122, 126, 130, 328lumbar, 48, 49mandibular, 104, 124, 126,
127, 130meningeal branch, 126motor branches, 126
masseteric, 126maxillary, 104, 124, 126, 127
ganglionic branches, 126,128
meningeal branch, 126median, 72, 74, 76, 77
communicating branch, 78muscular branches, 76palmar branch, 76paralysis, 76, 77
mental, 126motor, 2musculocutaneous, 70, 74,
75nasociliary, 124, 128nasopalatine, 128neck, transverse, 70, 72obturator, 86, 90, 91, 96
cutaneous branch, 90deep branch, 90superficial branch, 90
occipitalgreater, 72, 84lesser, 70, 72third, 72
oculomotor, 102–104, 128,134, 138, 139
olfactory, 102–104, 228–330
Nerve(s) (-o) N
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

400
ophthalmic, 104, 124, 125tentorial branch, 124
optic, 12, 102–104, 342, 350,351, 354–356
palatine, 128, 130pectoral
lateral, 74medial, 74
pelvic, 294penis, dorsal nerve, 96perineal, 96peripheral, 4, 36, 40, 41,
70–72, 100peroneal
common, 86, 92, 93deep, 92superficial, 92
petrosaldeep, 128greater, 122, 128, 130lesser, 118, 130
hiatus, 122phrenic, 70–73
pericardiac branches, 72plantar
lateral, 94medial, 94
pterygoid, 126pterygopalatine, 126, 128pudendal, 86, 96, 97
muscular branches, 96radial, 72, 82, 83
deep branch, 82muscular branches, 82paralysis, 82, 83superficial branch, 72ulnar communicating
branch, 82radicular, 64, 65rectal, inferior, 96saccular, 376sacral, 48, 49, 96saphenous, 88
cutaneous branches, 88infrapatellar branch, 88
scapular, dorsal, 74sciatic, 86, 92–94scrotal, posterior, 96sensory, 2spinal, 4, 48, 49, 70
anterior branches, 70, 84communicating branch, 70cutaneous branches
anterior, 84lateral, 84
lumbar, 84, 86mammary branches
lateral, 84medial, 84
meningeal branch, 70posterior branches, 70, 84sacral, 84, 86thoracic, 84, 85
splanchnicgreater, 292–298lesser, 298
stapedius, 122, 366subclavius, 74subcostal, 84, 86suboccipital, 72subscapular, 74supraclavicular, 70, 72supraorbital, 124suprascapular, 74supratrochlear, 124sural, 92, 94
calcaneal brancheslateral, 94medial, 94
muscular branches, 94temporal, deep, 126tensor tympani, 126, 130,
366tensor veli palatini muscle,
126, 130terminal, 330, 331thigh, cutaneous
lateral, 86, 88, 89posterior, 86, 90, 91, 96
thoracic, 48, 49long, 74
thoracodorsal, 74tibial, 86, 92, 94, 95
motor branches, 94paralysis, 94popliteal branch, 94
trigeminal, 102–104, 110,124–127, 154
spinal tract, 108, 110trochlear, 102, 104, 138, 139tympanic, 118, 366ulnar, 72, 78, 79
deep branch, 78dorsal branch, 78palmar branch, 78paralysis, 78, 79superficial branch, 78
utricular, 376vagus, 102–108, 112, 114–
117, 294, 328auricular branch, 114celiac branches, 116cervical cardiac branches,
116
gastric branches, 116hepatic branches, 116meningeal branch, 114paralysis, 116, 117pharyngeal branches, 114–
116renal branches, 116trigon, 100
vestibulocochlear, 102–106,120, 121, 154
vomeronasal, 330, 331zygomatic, 124, 126see also Ganglion(a);
Nucleus(i)Nervous system
autonomic, 2central, 2, 3development, 2, 3functional circuits, 2, 3parasympathetic, 2, 114peripheral, 2position of, 4, 5somatic, 2sympathetic, 2, 50, 114
Neural crest, 62Neural groove, 2, 6Neural network, 2, 3, 35Neural plate, 2, 62Neural tube, 2, 6, 102
longitudinal zones, 12primary cavity, 8
Neurite, 18Neuroendocrine system, 202–
205Neurofibrils, 18, 19, 22Neurofilaments, 22Neuroglia see GliaNeurohypophysis, 200Neurokeratin, 36Neuronal circuit(s), 34, 35, 296,
297, 310cerebellum, 160, 161of Papez, 332, 333retina, 350, 351
Neuron(s), 18–23autonomic, 302, 303bipolar, 18, 348–350, 354,
376catecholaminergic, 28, 29,
148cholinergic, 28, 32, 33dopaminergic, 32, 136GABAergic, 32, 34glutamatergic, 32multipolar, 18noradrenergic, 32peptidergic, 32, 146
N Nerve(s) (-o)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

401
projection, 18, 242serotoninergic, 28, 32, 148ultrastructure, 22unipolar, 18
Neuropeptides, 26Neuropil, 148Neuropore, oral, 6Neurosecretion, 202, 204Neurotransmitter receptors,
30, 31ligand-gated ion channels,
30receptors coupled to G pro-
tein, 30Neurotransmitters, 26–28,
148, 302pharmaceutical effects, 32
Neurotubules, 22, 28, 29Nigrostriatal system, 136Nodes of Ranvier, 36–41Nodulus, 152, 154Norepinephrine (NE), 26, 148,
149, 294Notch, supraorbital, 124Notochord, 100Nucleolus, 18, 22Nucleus(i)
abducens, 106, 110, 122, 138,378
accessory, 52spinal, 106, 112
ambiguous, 106, 108, 112–118
anterolateral, 52, 54anteromedial, 52, 54arcuate, 108basal, 32, 174, 182, 226Bechterew’s, 120, 380Cajal’s, 134, 176, 316caudate, 136, 172–174, 182,
192, 208, 209, 214–222, 236, 310
cell, 18, 22central, 226centromedian, 178cerebellar, 154, 155, 160Clarke’s, 52, 54, 56cochlear, 106, 378, 379
anterior, 120, 378posterior, 120, 378
cranial nerves, 106, 107caudal, 146
cuneate, 108, 308, 322lateral, 164tubercle, 100
Darkshevich’s, 134, 176Deiter’s, 110, 120, 382
dentate, 136, 154, 162, 316diencephalic, 146, 180dorsal, 52, 54, 56dorsomedial, 194, 202Edinger–Westphal, 106, 132–
134, 138, 144, 292emboliform, 154, 162, 182,
316entopeduncular, 172facial nerve, 106, 110, 122,
144fastigial, 154, 162geniculate see Body, genicu-
lateglobose, 154, 162, 316glossopharyngeal nerve, 106gracilis, 108, 308, 322
tubercle, 100gustatory, 328habenular, 32, 144, 176, 218,
332hypoglossal, 106, 108, 112,
144inferior colliculus, 132infundibular, 194, 202integration, 184, 186intercalate, 108intermediolateral, 54, 296intermediomedial, 54, 296interpeduncular, 32, 132,
144, 176, 332interstitial, 134, 176, 316lateral, 194, 226lentiform, 216magnocellular, 146medial, 194, 232Meynert’s, 174, 182Monakow’s, 164oculomotor nerve, 106, 132–
134, 138, 148accessory, 106, 132–134,
138, 144of origin, 106of termination, 106olfactory, anterior, 224, 228olivary, accessory
medial, 108posterior, 108
parasympathetic, 106paraventricular, 194, 196,
204Perlia’s, 138phrenic, 52pontine, 110posterolateral, 52, 54posteromarginalis, 54posteromedial, 52, 54
premamillary, 194prestitial, 134pretectal, 134proprius, 52raphe, 32, 108
posterior, 32, 144red, 106, 132–134, 136, 137,
310, 316reticular, lateral, 108, 164retroposterolateral, 52, 54Roller’s, 108salivatory, 292
inferior, 106, 118, 130, 144superior, 106, 110, 122,
130, 144Schwalbe’s, 110, 120, 382sensory, 106septal, 32, 176, 228, 232solitary, 32, 106–108, 114,
118, 122, 144, 148, 328somatomotor, 106Staderini’s, 108subthalamic, 144, 174, 192,
310supraoptic, 194, 196, 204tegmental
anterior, 332central, inferior, 110dorsal, 176pedunculopontine, 132posterior, 332
thalamic, 174, 178–187, 228,310
anterior, 178–180, 182,183, 188, 196, 232, 332
centromedian, 180, 182,183, 190, 316
dorso-oral, 180dorsocaudal, 180dorsointermediate, 180intralaminar, 180, 182, 188lateral, 178–180, 184, 185,
188–190dorsal, 178, 180, 184posterior, 178, 184
lateropolar, 180medial, 178–180, 182, 183,
188–190, 196, 228nonspecific, 178, 180–187reticular, 174, 178, 188, 190semilunar, 190specific, 178–180ventral, 178, 180, 184–187,
188, 190anterior, 178, 180intermediate, 184lateral, 178–180, 184–
188, 316
Nucleus(i) (-t) N
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

402
posterior, 178–180, 184–186, 190, 322–324,328
posterolateral, 184posteromedial, 184
ventro-oral, 180ventrocaudal, 180ventrointermediate, 180ventrolateral, 178
thoracic, posterior, 54, 164trapezoid, 120, 186
anterior, 110posterior, 110
trigeminal nerve, 110internuclear connections,
142mesencephalic, 106, 124,
132–134motor, 106, 110, 124, 144pontine, 106, 110, 124spinal, 106–110, 114, 124,
125, 324trochlear nerve, 106, 132,
138tuberal, 196tuberomamillary, 194vagus nervedorsal, 106, 292motor, 106posterior, 108, 114–117,
144, 148ventromedial, 194, 196, 202vestibular, 106–110, 160–
164, 310, 316, 382,383
inferior, 120, 382lateral, 110, 120, 382medial, 108–110, 120, 382superior, 120, 382
visceromotor, 106
OOligodendrocytes, 36–39, 43,
158Oligodendroglia, 42, 43Olive, 100, 106–108, 136, 144,
154, 164, 218, 316accessory, 316superior, 110, 378
Openingpharyngeal, 362tympanic, 362
Operculum(a), 8, 238frontal, 238parietal, 238
temporal, 222, 238Ora serrata, 342, 346, 350Orbit, 338, 339Organ(s)
acoustic, 362see also Ear
circumventricular, 286, 287gustatory, 326–329Jacobson’s, 330of balance, 374, 375of Corti, 120, 370, 372, 373olfactory, 330, 331sensory, cutaneous, 318–321spiral, 372subcommissural, 286subfornical, 286, 287tendon, 312, 313vascular, of the terminal
lamina, 286, 287vomeronasal, 330
Ossicles, auditory, 362, 364,365
Otoliths, 374Oxytocin, 204
PPachymeninx, 64, 288Paleocerebellum, 152Paleocortex, 208, 214, 224,
225Paleopallium, 208, 210Paleorubrum, 136Pallidum, 136, 144, 172, 310Pallium, 208Papilla(e)
foliate, 326fungiform, 326optic nerve, 346, 350, 351,
356vallate, 326, 327
Paraflocculus, 152Paraganglia, 300Paralysis, 68
brachial plexuslower (Klumpke’s palsy),
74upper (Erb’s palsy), 74
diaphragm, 72facial, 140facial nerve, 122, 123, 366gluteus nerveinferior, 90superior, 90
levator muscle of the velumpalatinum, 116
median nerve, 76, 77obturator nerve, 90radial nerve, 82, 83recurrent laryngeal, 116tibial nerve, 94ulnar nerve, 78, 79vagus nerve, 116, 117
Paraphysis, 286Paraplegia, 68Parasympathetic nervous sys-
tem, 2, 114, 292, 294Parkinson’s disease, 192Pathway(s)
auditory, 378–381common terminal motor,
316, 317vestibular, 382, 383visual, 354–357see also Tract(s)
Peduncle(s)cerebellar, 100, 154, 155
inferior, 100, 108, 120, 154,164
medial, 100, 110, 154, 166superior, 100, 136, 154,
166, 182–184, 218cerebral, 100, 132–134, 166,
218, 258inferior colliculi, 380mamillary body, 196, 332septum pellucidum, 214thalamic
anterior, 258dorsal, 258posterior, 258
Periarchicortex, 244, 332Perifovea, 350Perikaryon, 18Perilymph, 368Perineurium, 40, 64, 320Periorbita, 338Periosteum, 338Peripaleocortex, 244Peripheral nervous system, 2Pharynx, 114, 115, 118, 119
constrictor muscles, 116Phillippe–Gombault triangle,
56Photoreceptors, 352, 353,
354Pia mater, 288
spinal, 64Pigment architectonics, 244Pinealocytes, 176, 177Pinna, 362Placodes, lens, 342Plagiocephaly, 112
N Nucleus(i) (-t)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

403
Plasticity, 306Plate
alar, 4, 12, 50, 102, 132basal, 4, 12, 50, 102, 132commissural, 6cortical, 244motor end plate, 312, 313neural, 2, 62quadrigeminal, 10, 100, 132,
170tarsal, 338tectal, 100, 154
Platysma, 122Plexus, 70
aortic, abdominal, 298Auerbach’s, 300brachial, 4, 70, 71, 74–83
infraclavicular part, 70,74–83
paralysis, 74supraclavicular part, 70,
74, 75cardiac, 116, 296carotid, internal, 296celiac, 116cervical, 70, 71, 72, 73choroid, 10, 44, 102, 170, 174,
220, 230, 280, 282–285, 286
coccygeal, 96dental, superior, 126enteric, 300esophageal, 116hepatic, 116hypogastric, 292
inferior, 294, 298superior, 294, 298
intramural, 300, 301lumbar, 84, 86–90lumbosacral, 4, 86, 87Meissner’s, 300myenteric, 300parotid, 122pharyngeal, 114, 118prostatic, 298pulmonary, 116, 296renal, 116sacral, 86, 90–96submucous, 300, 301tympanic, 118, 366uterovaginal, 298venousepidural, 60internal vertebral, 64
visceral, 298Point, lacrimal, 338Pole
frontal, 212
occipital, 212temporal, 212
Pons, 6–12, 100, 110, 111, 132,133, 136, 218, 219, 316
Pontocerebellum, 152Positron emission tomography
(PET), 264, 266, 267Pouch, Prussak’s, 364Precuneus, 212Process(es)
ciliary, 344cochleariform, 366neuronal, 12, 284phalangeal, 372
Proisocortex, 244Prosencephalon, 4, 6, 170Psalterium, 232Ptosis, 296Pulvinar, 134, 178–180, 186,
190Punctum nervosum, 72Pupil, 128, 342Putamen, 136, 172–174, 182,
192, 208, 209,214–218, 222, 236,310
Pyramids, 100, 132, 140, 152,218
RRadiation
acoustic, 252, 258, 380corpus callosum, 260, 261occipitothalamic, 186optic (of Gratiolet), 254, 258,
259, 354central, 186, 190
thalamic, 179anterior, 178inferior, 178posterior, 178superior, 178
Radiography, 264, 265Radix
cochlearis, 370, 376, 378vestibularis, 376, 382
Raphe, 108Recess
epitympanic, 362infundibular, 200, 280lateral, 100, 280optic, 280pineal, 176, 280preoptic, 172superior, of the tympanic
membrane, 364
suprapineal, 280Redlich–Obersteiner zone, 62,
63Reflex, 50
accommodation, 358corneal, 142fixation, 358lacrimal, 142ocular, 358, 359oculocardial, 142pharyngeal, 142pupillary, 140, 358, 359sneezing, 142stretch, 50, 51, 314withdrawal, 50, 51
Reflex arcs, 50, 51monosynaptic, 50, 51multisynaptic, 50, 51
Regioncentral, 212, 316olfactory, 330paranodal, 38preoptic, 176, 194, 196pretectal, 134, 135
Reticulum trabeculare, 344Retina, 342, 346, 348–351, 356
cerebral layer, 348ciliary part, 344, 350ganglionic layer, 348iridial part, 344, 350neuroepithelial layer, 348optic part, 350
Rhinencephalon, 208, 210, 224Rhodopsin, 352Rhombencephalon, 100, 170Ribosomes, 22Rods, retinal, 348–352Root(s)
anterior, 48, 70cochlear, 106, 120, 121cranial, 102, 112dorsal, 48inferior, 72, 112lateral, 354medial, 354motor, 102, 124parasympathetic, 128, 130posterior, 48, 62, 63, 70
pain, 66sensorimotor, 130sensory, 124, 128spinal, 102, 112superior, 72, 112sympathetic, 128, 130trigeminal, terminal, 114ventral, 48vestibular, 106, 120, 121, 376
Roots R
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

404
SSac
dural, 64endolymphatic, 368lacrimal (tear), 338
Saccule, 120, 362, 368, 374Scala
media, 368, 370tympani, 368, 370vestibuli, 368, 370
Schizophasia, 262Schmidt–Lanterman incisures,
36, 37, 40Sclera, 338, 342, 350Sclerotomes, 66Segment
interannular, 36internodal, 36
Segmentation, 66Sella turcica, 104Sensation
loss of, 68pain, 56, 66, 324, 325pressure, 56, 324taste, 326, 327temperature, 56, 66, 324touch, 56, 318, 322–324
Sensibilitydisk prolapse and, 66epicritic, 56, 62, 68, 124, 140,
322, 323exteroceptive, 2, 56, 106,
140proprioceptive, 2, 56protopathic, 56, 62, 68, 124,
140, 324, 325Sensory systems, 318–331Septum(a), 144
posterior, 50Septum pellucidum, 10, 172,
214, 220cave of, 172
Serotonin (5-HT), 26, 148receptor, 30
Sheatharachnoid, 350bulbar, 338dural, 350pial, 350see also Myelin sheath
Signal transmission, 302Single-photon emission com-
puted tomography(SPECT), 264, 266, 267
Sinus(es)carotid, 118
viscerosensory branch, 118cavernous, 104confluence of, 104, 276dura mater, 288petrosal, superior, 276sagittal
inferior, 276superior, 276, 288
straight, 276transverse, 104, 276, 288venous, of the sclera, 344
Skin, 318encapsulated end organs,
318–321free nerve endings, 318, 319innervation, 72–95, 124–
127, 298, 299Skull base, 104, 105Somatic nervous system, 2Somatotopic organization, 162,
163Space(s)
cerebrospinal fluid, 280–287external, 280internal, 280
epidural, 64Meckel’s, 288Nuel’s, 372perichoroidal, 346perinuclear, 22perivascular, 44subarachnoid, 64, 288, 289subdural, 64, 288
Spinal cord, 4, 8, 12, 48, 50–69blood vessels, 60, 61cross sections, 54, 55intrinsic system, 52structure, 50syndromes, 68
central injury, 68, 69hemisection, 68, 69transection, 68, 69
Spinocerebellum, 152Stain, 18
Golgi’s method, 18, 19, 20Nissl’s method, 18, 19, 20
Stapes, 364, 366Statocones, 374Steppage gait, 92Stereocilia, 376Stimuli
exteroceptive, 2proprioceptive, 2
Stratumcerebral, 342lacunosum-moleculare, 234lemnisci, 134
oriens, 234pigmented, 342pyramidale, 234radiatum, 234
Stria(e)acoustic, posterior, 120, 378longitudinal, 232
lateral, 220, 230medial (of Lancisi), 220,
230mallear, 362medullary, 100, 120, 164,
174, 176olfactory, 12, 102, 212
lateral, 172, 224, 228, 229medial, 224, 228
terminal, 176, 196, 228, 332vascular, 372
Striatum, 32, 136, 144, 208,214–216, 222, 223,316
blood supply, 275Stroma, 344Subiculum, 232Substance
gelatinous, 52, 54, 308intermediate
central, 50lateral, 54
Nissl, 18perforated
anterior, 12, 102, 172, 176,212, 224, 225, 228
posterior, 100Rolando’s, 52, 54
Substantiaalba, 12, 50grisea, 12, 52, 310innominata, 174, 182nigra, 32, 50, 132–134, 136,
137, 174, 192, 216–218
compact part, 136reticular part, 136
spongiosa, 52Subthalamus, 172, 174, 192, 193Succinate dehydrogenase, 148,
149Sulcus(i), 10, 212
anterolateral, 100calcarine, 212, 220, 254central, 212, 216, 220
insula, 238cerebral, lateral, 238circular, 238collateral, 212fimbriodentate, 232
S Sac
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

405
frontalinferior, 212, 214superior, 212, 214
hippocampal, 212, 230, 232intraparietal, 212lateral, 8–10, 212–218, 222
posterior, 50median, 100occipital, transverse, 212occipitotemporal, 212of Sylvius, 10, 212olfactory, 212parieto-occipital, 212postcentral, 212posterior median, 48, 100posterolateral, 100precentral, 212spiral
external, 372internal, 372
telodiencephalic, 6temporal
inferior, 212, 214superior, 212, 214
see also FissureSympathetic nervous system,
2, 50, 114, 292, 294Synapse(s), 24–27
asymmetric, 24axo-axonal, 24, 302axodendritic, 24axosomatic, 24axospinous, 24chemical, 26complex, 26, 27electrical, 26excitatory, 24glomerulus-like complexes,
26inhibitory, 24invaginated, 352parallel contact, 26, 27spinous, 26, 27, 156symmetric, 24
Synaptic transmission, 30, 31
TTaenia, 282, 283choroidea, 282fornicis, 282thalami, 282
Tapetum, 254Tarsusinferior, 338superior, 338
Taste buds, 326–329Tectum, 132, 220, 222Tegmentum, 108, 110, 132, 146Tela choroidea, 170, 282, 283Telencephalization, 14Telencephalon, 4, 6, 12, 170,
208–267evolution, 14, 210, 211
Telodiencephalic boundary,170, 171
Teloglia, 312Tendon, common annular, 340Tetraplegia, 68Thalamus, 32, 170, 174, 178,
216–222blood supply, 274, 275caudal, 190, 191dorsal, 172, 178, 179medullary layers, 174posterior, 136rostral, 188, 189
Thermoanesthesia, 68Tissue culture, 20Tongue, 112, 115, 118, 119, 123,
126, 127, 327–329Tonotopic organization, 378–
381Tonsilla(e), 152, 154Tracers, 20Tract(s)aberrant
mesencephalic, 140pontine, 140
arcuatocerebellar, 108, 164,165
ascending, 56, 57bulbothalamic, 140cerebellar, anterior, 154cerebelloreticular, 167cerebellorubral, 167, 310cerebellothalamic, 167cerebellovestibular, 164comma tract of Schultze, 56corticopontine, 167, 316corticorubral, 136corticospinal, 58, 59, 140,
141, 244, 258, 308,309, 316
anterior, 58, 59, 308lateral, 58, 59, 308
corticotectal, 134cuneocerebellar, 162, 164descending, 58, 59extrapyramidal, 58Flechsig’s, 56, 164frontopontine, 166, 258geniculocalcarine, 190
Gowers’, 56, 166habenulointerpeduncular,
132, 176habenulopeduncular, 332habenulotectal, 176habenulotegmental, 176, 332hypothalamohypophysial,
202interstitiospinal, 310Lissauer’s, 52, 54, 56, 324mamillothalamic, 182, 192mesencephalic, 124nucleocerebellar, 164, 165olfactory, 12, 102, 212–215,
224, 228olivocerebellar, 108, 164, 165optic, 102, 174, 178, 186, 190,
216, 226, 230, 354–357
pallidorubral, 136paraventriculohypophysial,
204parependymal, 58pontocerebellar, 166, 167posterolateral, 52–56pyramidal, 58, 59, 108–110,
140, 246, 308, 316reticulocerebellar, 164, 165reticulospinal, 146, 166, 310,
316anterior, 58lateral, 58
rubroreticulospinal, 316rubrospinal, 58, 136, 310solitary, 108, 114, 118spinal, 124spino-olivary, 56, 164spinocerebellar, 108, 163,
316anterior, 56, 57, 162–167posterior, 56, 57, 162, 164,
165spinoreticular, 324spinotectal, 56, 140spinothalamic, 108–110,
140, 325anterior, 56, 57, 140, 324lateral, 56, 57, 140, 324
spiral, foraminous, 120, 376supraopticohypophysial,
204, 205tectorubral, 136tectospinal, 58tegmental, central, 108–110,
136, 144, 145, 154,164, 310, 316
tegmentospinal, 58
Tract(s) (-t) T
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

406
temporopontine, 166, 258tuberoinfundibular, 202vegetative, 58vestibulocerebellar, 163, 164,
165vestibulospinal, 56–58, 120,
164–166, 310, 316,382
visualization of, 58see also Pathway(s)
Transportanterograde, 20, 21, 28axonal, 28, 29retrograde, 20, 21, 28
Trendelenburg’s symptom, 90Trigone, olfactory, 12, 224Trochlea, 340Trunkencephalic, 4, 6, 100inferior, 70, 74lumbosacral, 86, 90medial, 70, 74superior, 70, 74sympathetic, 292, 294, 296–
299vagalanterior, 116posterior, 116
Tubeauditory (eustachian), 118,
362, 364neural, 2, 6, 102
longitudinal zones, 12primary cavity, 8
Tubercinereum, 170, 174, 175, 194,
195median eminence, 200
Tuberclegray, 44, 144olfactory, 224
Tuberoinfundibular system,202, 203
UTunnelinner, 372outer, 372
Ultrasound, 264Uncus, 212, 224Utricle, 120, 362, 368, 374Uvea, 342Uvula, 152, 154
VVallecula, cerebellum, 152, 154Varicosities, 302Vasocorona, 60Vasopressin, 204Vein(s), 276–279anastomotic
inferior, 276superior, 276
basal, 276, 278, 279Browning’s, 276central, 276
of the retina, 346cerebral
anterior, 278deep, 276, 278, 279great (of Galen), 276, 278inferior, 276internal, 276, 278middle, 276, 278superficial, 276, 278superior, 276
choroid, 278inferior, 278superior, 278
ciliary, posterior, 346interpendicular, 278jugular, 366
internal, 104Labbé’s, 276Rosenthal’s, 276septum pellucidum, 276, 278spinal, 60
anterior, 60posterior, 60
thalamostriate (terminal),170, 174, 276, 278
transcerebral, 278Trolard’s, 276vorticose, 346
Velummedullary
inferior, 100, 154superior, 100, 154
palatinum, 117levator muscle, 116, 130
paralysis, 116tensor muscle, 124, 126,
130Ventricle(s), 209, 280, 281fourth, 8–10, 100, 108, 218,
280, 282, 283lateral, 8, 172, 208, 214, 218,
220, 221, 222–224,280, 282, 283
optic, 342
third, 8, 172, 208, 280Vermis, cerebellum, 12, 152,
153, 162Vesicle(s)Golgi, 22granular, 26lens, 342olfactory, 330optic, 6, 342synaptic, 24–27, 312telencephalic, 6
Vestibular apparatus, 120Vestibulocerebellum, 152Virchow-Robin space, 44
WWall, labyrinthic, 366White matter, 12, 13, 258spinal cord, 50, 54
Windowoval (vestibular), 364–368round (cochlear), 364–368
Wristdrop, 82
YYellow spot, 346
ZZona incerta, 144, 174, 192Zone(s)
autonomic, 74border, 244ciliary, 344dynamogenic (ergotropic),
198Head’s, 298intermediate, 162longitudinal, 12maximum, 74neurohemal, 200of apposition, 24Redlich–Obersteiner, 62, 63somatomotor, 102somatosensory, 102trophotropic, 198visceromotor, 102viscerosensory, 102
Zonulae, 24adherentes, 284occludentes, 40, 44, 284
T Tract(s) (-t)
Kahle, Color Atlas of Human Anatomy, Vol. 3 © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.