
Codend selection in the deep-water crustacean trawl fishery in Portuguese southern waters
12
Fisheries Research 85 (2007) 49–60 Codend selection in the deep-water crustacean trawl fishery in Portuguese southern waters Paulo Fonseca a,∗ , Aida Campos a , Russell B. Millar b a INIAP/IPIMAR-Portuguese Institute for Agriculture and Fisheries Research, Avenida de Bras´ ılia, 1449-006 Lisboa, Portugal b Department of Statistics, University of Auckland, Private Bag 92019, New Zealand Received 8 August 2006; received in revised form 14 November 2006; accepted 28 November 2006 Abstract In this paper, the potential improvement of size-selectivity by increasing codend mesh size and changing twine type is addressed for the main commercial species exploited by crustacean trawlers in waters off the Portuguese south coast. Three cruises were conducted between autumn 1998 and summer 1999 onboard the F/V “Porto Bravo”, resulting in a total of 93 valid hauls using 3 different sizes of diamond mesh in the codend (stretched nominal sizes of 55, 70 and 80 mm) and 2 types of twine, polyamide and polyethylene. A non-linear mixed-effects model was employed to include the effect of covariates and between-haul variation in the estimation of codend selectivity parameters for the main target species in this fishery, Norway lobster, Nephrops norvegicus (L.), rose shrimp, Parapenaeus longirostris (Lucas 1846), and ‘blue and red’ shrimp, Aristeus antennatus (Risso, 1816). The chosen selectivity model found a significant (p < 0.001) linear effect of mesh size for all three target species, and a significant effect of cruise for Norway lobster (p < 0.05) and rose shrimp (p < 0.001). The cruise effect was due to a difference between the autumn 1998 cruise and the two latter cruises. This effect coincides with a change from polyamide twine in the autumn cruise to the use of polyethylene twine in the latter cruises. Other covariates, such as depth, codend-catch weight and species catch weight did not show any systematic effect on selectivity and thus were not included in the final models. The retention of a high number of undersized (or immature) individuals, while using the legal minimum mesh sizes, suggests the need for a change in the status quo fishing pattern. © 2006 Elsevier B.V. All rights reserved. Keywords: Diamond mesh size; Codend selection; Crustacean trawling; Non-linear mixed effects; Parapenaeus longirostris; Nephrops norvegicus; Aristeus antennatus; Portuguese coast 1. Introduction Bottom trawling for deep-water crustaceans is one of the most important m´ etiers of the Portuguese fisheries, due to the extremely high market value attained by these species (1176 tonnes, D 15,372 × 10 3 in 2003). 1 A total of about 30 vessels are engaged in this fishery, constituting a well-defined fleet component (Campos et al., 2007), with length overall, gross tonnage and engine power ranging between 23 and 32 m, 96–241 GT and 363–589kW, respectively. 2 Fishing is carried out off the south and southwest coasts (ICES Functional Units 28 and 29) at depths ranging from about 150–700 m. Species ∗ Corresponding author. Tel.: +351 213027163; fax: +351 213015948. E-mail addresses: [email protected] (P. Fonseca), [email protected] (A. Campos), [email protected] (R.B. Millar). 1 Data from the National Institute for Statistics (INE). 2 Data from the Directorate-General for Fisheries and Aquaculture (DGPA) for 2003. caught include the rose shrimp, Parapenaeus longirostris (Lucas 1846), the Norway lobster, Nephrops norvegicus (L.), and deep- water shrimps such as the ‘blue and red’ shrimp, Aristeus antennatus (Risso, 1816), the giant red shrimp, Aristaeomor- pha foliacea (Risso, 1827), and the scarlet shrimp, Aristaeopsis edwardsiana (Johnson, 1867). Along with crustaceans, a high number of commercial and non-commercial fish species are caught as by-catch. The hake, Merluccius merluccius (L.), and the monkfishes (Lophius spp.) are among the most valuable fish by-catch species, while the blue whiting, Micromesistius poutassou (Risso, 1826) is captured in high quantity but often discarded. The Norway lobster and the rose shrimp constitute the bulk of the catches (both in weight and in value) and, according to their relative abundance, the fleet may re-direct the effort prefer- entially to one of them, as happened from 1994 to 2003 when the decline of Norway lobster was compensated by shrimp catches. Deep-water shrimps (mainly the ‘blue and red’ shrimp) repre- sent a much smaller fraction of the total catches of crustaceans, 0165-7836/$ – see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.fishres.2006.11.036
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Codend selection in the deep-water crustacean trawl fishery in Portuguese southern waters

A
ca(ttas1tsl©
Ka
1
mt(vflg9o2
(
f
0d
Fisheries Research 85 (2007) 49–60
Codend selection in the deep-water crustacean trawl fisheryin Portuguese southern waters
Paulo Fonseca a,∗, Aida Campos a, Russell B. Millar b
a INIAP/IPIMAR-Portuguese Institute for Agriculture and Fisheries Research, Avenida de Brasılia, 1449-006 Lisboa, Portugalb Department of Statistics, University of Auckland, Private Bag 92019, New Zealand
Received 8 August 2006; received in revised form 14 November 2006; accepted 28 November 2006
bstract
In this paper, the potential improvement of size-selectivity by increasing codend mesh size and changing twine type is addressed for the mainommercial species exploited by crustacean trawlers in waters off the Portuguese south coast. Three cruises were conducted between autumn 1998nd summer 1999 onboard the F/V “Porto Bravo”, resulting in a total of 93 valid hauls using 3 different sizes of diamond mesh in the codendstretched nominal sizes of 55, 70 and 80 mm) and 2 types of twine, polyamide and polyethylene. A non-linear mixed-effects model was employedo include the effect of covariates and between-haul variation in the estimation of codend selectivity parameters for the main target species inhis fishery, Norway lobster, Nephrops norvegicus (L.), rose shrimp, Parapenaeus longirostris (Lucas 1846), and ‘blue and red’ shrimp, Aristeusntennatus (Risso, 1816). The chosen selectivity model found a significant (p < 0.001) linear effect of mesh size for all three target species, and aignificant effect of cruise for Norway lobster (p < 0.05) and rose shrimp (p < 0.001). The cruise effect was due to a difference between the autumn998 cruise and the two latter cruises. This effect coincides with a change from polyamide twine in the autumn cruise to the use of polyethylene
wine in the latter cruises. Other covariates, such as depth, codend-catch weight and species catch weight did not show any systematic effect onelectivity and thus were not included in the final models. The retention of a high number of undersized (or immature) individuals, while using theegal minimum mesh sizes, suggests the need for a change in the status quo fishing pattern.2006 Elsevier B.V. All rights reserved.
n-lin
c1wapenct
eywords: Diamond mesh size; Codend selection; Crustacean trawling; Nontennatus; Portuguese coast
. Introduction
Bottom trawling for deep-water crustaceans is one of theost important metiers of the Portuguese fisheries, due to
he extremely high market value attained by these species1176 tonnes, D 15,372 × 103 in 2003).1 A total of about 30essels are engaged in this fishery, constituting a well-definedeet component (Campos et al., 2007), with length overall,ross tonnage and engine power ranging between 23 and 32 m,
6–241 GT and 363–589 kW, respectively.2 Fishing is carriedut off the south and southwest coasts (ICES Functional Units8 and 29) at depths ranging from about 150–700 m. Species∗ Corresponding author. Tel.: +351 213027163; fax: +351 213015948.E-mail addresses: [email protected] (P. Fonseca), [email protected]
A. Campos), [email protected] (R.B. Millar).1 Data from the National Institute for Statistics (INE).2 Data from the Directorate-General for Fisheries and Aquaculture (DGPA)
or 2003.
fipd
otedDs
165-7836/$ – see front matter © 2006 Elsevier B.V. All rights reserved.oi:10.1016/j.fishres.2006.11.036
ear mixed effects; Parapenaeus longirostris; Nephrops norvegicus; Aristeus
aught include the rose shrimp, Parapenaeus longirostris (Lucas846), the Norway lobster, Nephrops norvegicus (L.), and deep-ater shrimps such as the ‘blue and red’ shrimp, Aristeusntennatus (Risso, 1816), the giant red shrimp, Aristaeomor-ha foliacea (Risso, 1827), and the scarlet shrimp, Aristaeopsisdwardsiana (Johnson, 1867). Along with crustaceans, a highumber of commercial and non-commercial fish species areaught as by-catch. The hake, Merluccius merluccius (L.), andhe monkfishes (Lophius spp.) are among the most valuablesh by-catch species, while the blue whiting, Micromesistiusoutassou (Risso, 1826) is captured in high quantity but ofteniscarded.
The Norway lobster and the rose shrimp constitute the bulkf the catches (both in weight and in value) and, according toheir relative abundance, the fleet may re-direct the effort prefer-
ntially to one of them, as happened from 1994 to 2003 when theecline of Norway lobster was compensated by shrimp catches.eep-water shrimps (mainly the ‘blue and red’ shrimp) repre-ent a much smaller fraction of the total catches of crustaceans,

5 ies Re
bsw
oaCCa(7lcsMtgmo
cl1omoda2Frtto1stl
Seswctnau(rb
q
v
ttls
2
2
oomd5oodewfirgh
2
sswaruoa
The codends were constructed of diamond mesh of threenominal stretched sizes: 55 mm (the MMS for shrimps), 70 mm(the MMS for the Nephrops target fishery) and 80 mm. Theircharacteristics and construction followed that of the commercial
0 P. Fonseca et al. / Fisher
ut the high price attained in the first market sale justifies thatome fishing units can preferentially exploit the fishing groundshere they occur.The Norway lobster is the only species, with its wide area
f distribution (from Iceland to the southern coast of Portugalnd the Mediterranean), present across European Union waters.onsequently, it is managed within the scope of the Internationalouncil for the Exploration of the Sea (ICES), where total allow-ble catches (TAC’s) are defined yearly for each functional unitor group of units) and minimum stretched mesh size (MMS,0 mm) and minimum landing sizes (MLS, 20 mm carapaceength, CL) are established. On the other hand, the Portugueseoast lies at the northernmost distribution of the above shrimppecies within the northeast Atlantic. There are no TAC’s, and
LS’s have been set for only the rose shrimp (24 mm, CL) andhe ‘blue and red’ shrimp (29 mm, CL), both of which can be tar-eted with a MMS of 55 mm. Additional regulations consist ofinimum percentages of target species in the catch and temporal
r spatial fishing closures.The ICES Working Group on Nephrops (WGNEPH) noted a
onsistent decrease in catch-per-unit-effort (CPUE) of Norwayobster in Portuguese waters from about 120 kg/vessel-day in989 to about 50 kg in the period 1995–2003, remaining morer less stable thereafter (ICES, 2004). Stock biomass and recruit-ent also show a similar declining trend. Assessments carried
ut in 2001 (ICES, 2001) indicate this to be ‘. . . a severelyeclining stock.’ resulting from over-exploitation. Subsequentppraisals led the WGNEPH to propose zero TAC’s for 2003,004 and 2005 to rebuild the stock (ICES, 2002, 2003, 2004).or rose shrimp, a tentative assessment was tried in 1991 forose shrimp (Pestana and Ribeiro-Cascalho, 1991), suggestinghe maintenance, or even the reduction, of fishing effort as a cau-ionary measure. Nonetheless, the catch increased until a peakf 1696 tonnes in 1999, followed by a marked drop to about07 tonnes in 2004. The overall trend for the ‘blue and red’hrimp3 is similar; the average landings of about 220 tonnes inhe 1998–2000 period were followed by a strong decline, withandings of 19 tonnes in 2004 (DGPA, 1998–2004).
A group of experts (SGMOS, 2003) convened under thecientific, Technical and Economical Committee for Fish-ries (STECF), proposed the implementation of a managementcheme for Norway lobster and hake stocks in Southern EUaters, based on the limitation of effort, complemented by area
losures. In spite of the current situation additional gear restric-ions will be difficult to implement given the mixed-speciesature of the fisheries, although previous studies (Campos etl., 2002, 2003) have evidenced a mismatch between currentlysed MMS and the MLS (for shrimps) or size of first maturation4
for Norway lobster). Therefore, in spite of the urgent need toeduce fishing mortality, which will be more readily achievedy effort restrictions, there is still the need for adjustment in
3 Under this name there are other shrimp species landed, although in smalluantities, such as Aristaeomorpha foliacea and Aristaeopsis edwardsiana.4 First maturation size (Lmat) refers to the length for which 50% of the indi-iduals are mature.
F((
search 85 (2007) 49–60
he fishing pattern. To determine appropriate mesh sizes for thearget species, the present study analyses experimental data col-ected during commercial surveys using a range of codend meshizes, over three different seasons.
. Materials and methods
.1. Haul location
The selectivity experiments took place on three differentccasions, in the autumn of 1998, and the spring and summerf 1999. In each of the experiments, three different sizes of dia-ond mesh were tested in the codend. Each cruise lasted for 10
ays, onboard the F/V ‘Porto Bravo’, a 31.5 m overall length,89 kW, commercial crustacean trawler. Fishing was carried outn the grounds commonly exploited by the crustacean trawl fleetff the western region of the Portuguese south coast (Fig. 1), atepths from 200 to 570 m, during daylight. Minimum interfer-nce with commercial fishing practice was adopted; the skipperas encouraged to follow the usual fishing tactics, choice ofshing grounds and trawling speed (3.1–3.2 knots). The onlyestriction was the haul duration (i.e., time of the gear on theround), which was set to 3 h, to facilitate comparability amongauls.
.2. Gear and test codends
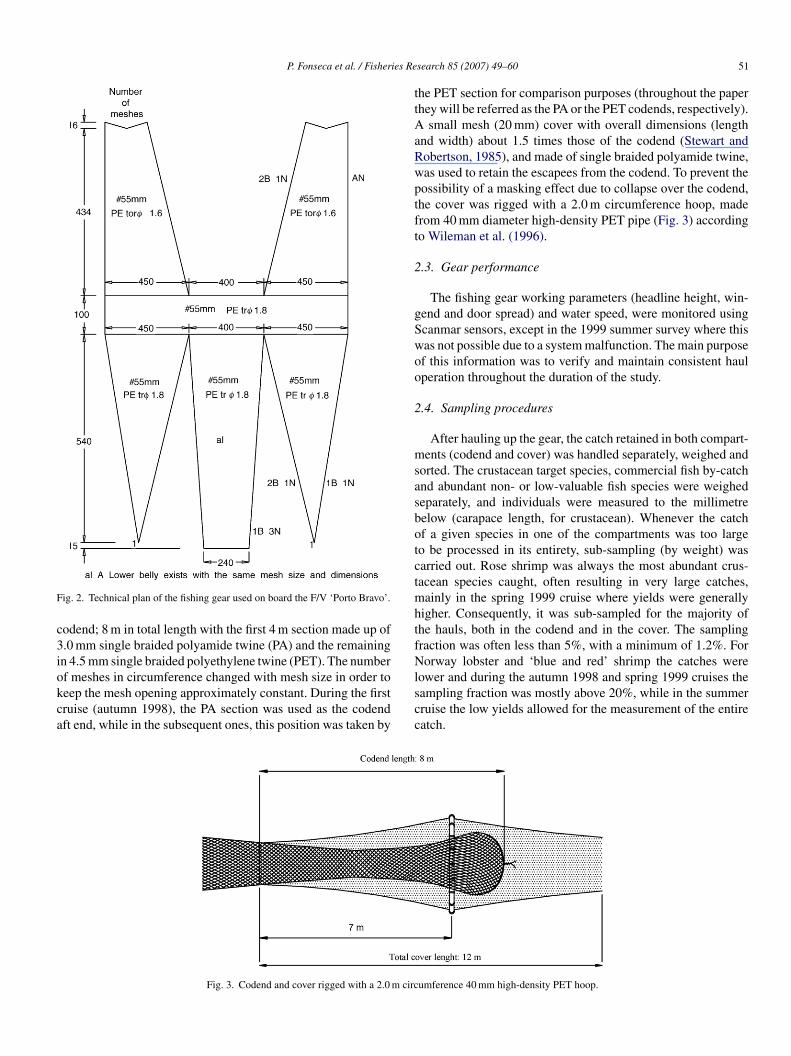
A standard commercial trawl net was used, and it was con-tructed and rigged by the crew according to the skipper’specifications (Fig. 2). In particular, the upper and lower wingsere made in a single piece with 1700 meshes in circumference
t the footrope level and about 480 meshes at the codend joining-ow. Mesh size (stretched mesh) of 55 and 1.8 mm diameter wassed throughout the wings and body of the net. ‘Sounete’ semi-val otterboards of Portuguese design, weighing 390 kg and withn area of about 3.2 m2, were used.
ig. 1. Location of the experimental hauls. Open circles—autumn 1998 haulsn = 26); filled circles—spring 1999 hauls (n = 23); crosses—summer 1999 haulsn = 28).
P. Fonseca et al. / Fisheries Re
F
c3iokca
ttAaRwptft
2
gSwoo
2
msasbotctmhtfN
ig. 2. Technical plan of the fishing gear used on board the F/V ‘Porto Bravo’.
odend; 8 m in total length with the first 4 m section made up of.0 mm single braided polyamide twine (PA) and the remainingn 4.5 mm single braided polyethylene twine (PET). The number
f meshes in circumference changed with mesh size in order toeep the mesh opening approximately constant. During the firstruise (autumn 1998), the PA section was used as the codendft end, while in the subsequent ones, this position was taken bylscc
Fig. 3. Codend and cover rigged with a 2.0 m circ
search 85 (2007) 49–60 51
he PET section for comparison purposes (throughout the paperhey will be referred as the PA or the PET codends, respectively).
small mesh (20 mm) cover with overall dimensions (lengthnd width) about 1.5 times those of the codend (Stewart andobertson, 1985), and made of single braided polyamide twine,as used to retain the escapees from the codend. To prevent theossibility of a masking effect due to collapse over the codend,he cover was rigged with a 2.0 m circumference hoop, maderom 40 mm diameter high-density PET pipe (Fig. 3) accordingo Wileman et al. (1996).
.3. Gear performance
The fishing gear working parameters (headline height, win-end and door spread) and water speed, were monitored usingcanmar sensors, except in the 1999 summer survey where thisas not possible due to a system malfunction. The main purposef this information was to verify and maintain consistent haulperation throughout the duration of the study.
.4. Sampling procedures
After hauling up the gear, the catch retained in both compart-ents (codend and cover) was handled separately, weighed and
orted. The crustacean target species, commercial fish by-catchnd abundant non- or low-valuable fish species were weighedeparately, and individuals were measured to the millimetreelow (carapace length, for crustacean). Whenever the catchf a given species in one of the compartments was too largeo be processed in its entirety, sub-sampling (by weight) wasarried out. Rose shrimp was always the most abundant crus-acean species caught, often resulting in very large catches,
ainly in the spring 1999 cruise where yields were generallyigher. Consequently, it was sub-sampled for the majority ofhe hauls, both in the codend and in the cover. The samplingraction was often less than 5%, with a minimum of 1.2%. Fororway lobster and ‘blue and red’ shrimp the catches were
ower and during the autumn 1998 and spring 1999 cruises theampling fraction was mostly above 20%, while in the summerruise the low yields allowed for the measurement of the entireatch.
umference 40 mm high-density PET hoop.

5 ies Research 85 (2007) 49–60
2
vctsyaToctu
Tab2hrrtr52r
L
wcartT
L
w(ta
S
wN
3
3
gssbs
Ft
sccsav
2 P. Fonseca et al. / Fisher
.5. Selectivity analysis
For each species, the length frequency data from every indi-idual deployment (over the three codend mesh sizes and threeruises) were appended into a single file. This file also containedhe relevant covariates (i.e., explanatory variables), including theampling fraction if the catch was sub-sampled. Statistical anal-sis of selectivity used the mixed-effects approach of Millar etl. (2004), implemented using the SAS procedure NLMIXED.his approach allows the selectivity model to include the effectsf covariates and between-haul variability. Moreover, in the pro-ess of fitting the selection curves, the model explicitly includeshe effect of sub-sampling, thereby avoiding the need to manip-late the observed length frequency data (Millar, 1994).
The main covariates of interest were mesh size and cruise.otal codend-catch weight and species total weight, as wells depth, have been found to be responsible for uncontrolledetween-haul variability in selectivity (Campos et al., 2002,003), and therefore were also included in the analysis. Between-aul variability was included by allowing length of 50%etention (L50) and selection range (SR: L75 – L25) to varyandomly between each deployment, according to a normal dis-ribution (Millar et al., 2004). Specifically, the L50 value for theth replicate deployment of gear j (j = 1, 2, 3, corresponding to5, 70 and 80 mm mesh codends, respectively) on cruise i (i = 1,, 3, corresponding to the autumn, spring and summer cruises,espectively) was modelled as (Model 1):
50 = αi + βj + δwi,j,r + γdi,j,r + εi,j,r (1)
here wi,j,r denotes the catch weight (either total weight inodend or species weight), and di,j,r denotes depth. The εi,j,r
re independent and identically distributed Normal(0,σ2) andepresent the random between-haul variation. A reduced form ofhis model, with a linear effect of mesh size, was also considered.hat is (Model 2),
50 = αi + βmj + δwi,j,r + γdi,j,r + εi,j,r (2)
here mj denotes mesh size, and β is interpreted as the increasemm) in L50 arising from a one mm increase in mesh size. Selec-ion range was modelled as a function of between-haul variationnd gear.
R = μSRj + ξi,j,r (3)
here ξi,j,r are independent and identically distributedormal(0,τ2).
. Results
.1. Species catch
The seasonal variability of catches (in weight) of the tar-et species and main commercial and non-commercial by-catch
pecies, including the ‘trash’ fraction (discards constituted bymall organisms, either fish or crustacean and other inverte-rates) is displayed in Fig. 4. The most productive season was thepring, with the catches of commercial crustaceans (mostly rosefatw
ig. 4. Seasonal variability of catches in weight. The ‘other’ fraction includesrash and discarded species.
hrimp) attaining 44% of the total. For Norway lobster the per-entages always remained under 4%. Concurrently, there werehanges in the proportion of by-catch species. Blue whiting con-tituted about 50% of the total catches in the summer and theutumn trials, but only 16% in the spring, while hake, the mostaluable by-catch species, was scarcely captured. The ‘trash’
raction always constituted a large component of the catch,ttaining about 37% in the spring. The latter figures (along withhe fact that when the trials were carried out the blue whitingas not a valuable by-catch species) points to a high level of dis-
P. Fonseca et al. / Fisheries Research 85 (2007) 49–60 53
Fs(
ca
3
ad
m2I
Fs
abawmsncrbso
ig. 5. Seasonal length frequency distribution (sex combined) for the threepecies in study. (a) Rose shrimp; (b) ‘blue and red’ shrimp; (c) Norway lobster.—) Autumn; (· · ·) spring; (– – –) summer.
ards in this fishery constituting at least 50% of the total catchnd reaching as much as 75%.
.2. Length frequency distributions
The length frequency distributions (LFD) by cruise (season)nd by depth strata for the commercial crustacean species areisplayed in Figs. 5 and 6, respectively.
The rose shrimp length distributions for the spring and sum-er cruises were almost coincident, both in range (from about
0–40 mm, carapace length) and in modal values (26–27 mm).n the autumn two modes could be perceived, a principal mode
ta
d
ig. 6. Length frequency by depth strata for the three species in study. (a) Rosehrimp; (b) ‘blue and red’ shrimp; (c) Norway lobster.
t 23 mm and a secondary one at 28 mm. Norway lobsteretween about 18 and 59 mm carapace length were caught inll three surveys, although individuals below the 20 mm MLSere extremely scarce. The LFD’s from the spring and sum-er were very similar, while the autumn one displayed a clear
hift to the left, due to the simultaneous occurrence of a largeumber of smaller individuals and the scarcity of larger lengthlasses. Otherwise, for the ‘blue and red’ shrimp the lengthanges were almost coincident for the three cruises, varyingetween 15 and 68 mm. The summer and autumn cruises pre-ented similar LFD’s with the main mode locate in the rangef 30–32 mm, while in the spring where the shrimps belowhe MLS (29 mm) were much scarcer the mode was located
t 37 mm.Fig. 6 evidences the marked influence of depth on the abun-ance and size distribution of these crustacean species. The

5 ies Research 85 (2007) 49–60
riwTdiarhn‘wtoffi
3
trmuea
M(cs
Uiabateswawmsbsmnrc
tStss
Table 1Final results of the modelling process: predictive models
Parameter Estimate S.E. p-Value Hauls (total/used)
Rose shrimpμSR 11.32 0.32 <0.0001β 0.41 0.01 <0.0001 73/73α2&3 −3.03 0.85 0.001σ2 11.72 2.17 <0.0001
SR fixedRed shrimp
μSR 11.07 0.24 <0.0001β 0.44 0.01 <0.0001 57/50α2&3
a – – –σ2 13.65 3.19 <0.0001
Norway lobsterμSR 7.38 0.35 <0.0001β 0.41 0.02 <0.0001 65/28α2&3
a −3.75 1.41 0.0131σ2 8.96 3.01 0.0061
a
atwt
3
−mTasT
c(aeosacidOnt(c
4 P. Fonseca et al. / Fisher
ose shrimp was captured from 200 to about 600 m, display-ng a clear trend for the increase in mean carapace length sizeith depth (24 mm until 400 m and 27 mm in deeper waters).he catch rates (numbers per hour) decrease markedly withepth until about 500 m, but then showed a very pronouncedncrease in the deeper stratum. Both the ‘blue and red’ shrimpnd the Norway lobster displayed a considerably narrower depthange distribution, starting to occur at about 300 m but with theighest yields only taking place in waters deeper than 400 m,amely at the 500 m stratum. Similarly to the rose shrimp, theblue and red’ shrimp displayed a depth-related size distributionith a mean carapace length of 34.3 mm in the shallower stra-
um (until 400 m) and above 39 mm in deeper waters. On thether hand, the Norway lobster did not display a marked dif-erence among the mean carapace sizes within the depth rangeshed.
.3. Size-selectivity
In general, PROC NLMIXED was frequently unable to ini-iate the model fit when both L50 and SR were modelled withandom between-haul variation, indicating that attempting toodel between-haul variability in both of these parameters was
nrealistically complex for the available data. Thus, the mod-lling of random between-haul variability was restricted to L50lone.
Using Akaike’s information criterion (AIC) it was found thatodel 2 (Eq. (2)) was preferred to the unrestricted Model 1
Eq. (1)). The covariates depth, codend-catch weight and speciesatch weight were all non-significant (p > 0.05) for all threepecies.
Model 2 assumes a linear effect of mesh size on L50 (Eq. (2)).nder this model it makes sense to not only test for a difference
n the cruise effect parameters (αi, i = 1, 2, 3, in Eq. (2)), butlso to test whether these three parameters can all be set to zeroecause the hypothesis αi = 0 corresponds to modelling the L50s being geometrically similar, in the sense that L50 is propor-ional to mesh size. For the ‘blue and red’ shrimp, none of thestimated cruise effect parameters (αi, i = 1, 2, 3, in Eq. (2)) wereignificantly different from zero (p > 0.05) and hence these termsere excluded from the final model (Table 1). For rose shrimp
nd Norway lobster, a significant difference between cruisesas detected (p < 0.05). Pairwise comparisons between the esti-ated cruise effects were implemented using the CONTRAST
tatement within PROC NLMIXED. These revealed that, foroth species, the estimated autumn cruise parameter α1 was notignificantly different from zero. However, the spring and sum-er cruise estimates were significantly different from zero, but
ot from each other. The final models (Table 1) fitted to theose shrimp and Norway lobster data set α1 = 0 and estimate aommon value, α2&3 for the common effect of cruises 2 and 3.
Mesh size did not have a significant effect (p > 0.05) on selec-ion range. That is, for each of the three target species, a constant
R value, μSR, was assumed for that species. To check whetherhis was a sensible conclusion for these data, separate analy-es were also conducted for each mesh size. These analyseshowed no systematic pattern in SR attributable to mesh size for
3
f
a This parameter should be interpreted as a codend twine effect, PA for autumnnd PET for spring/summer (see Section 4 for details).
ny of the species. Indeed, using Bonferroni correction for thehree pairwise comparisons between mesh sizes, the SR valuesere not significantly different (corrected p > 0.05) for any of
he species.
.4. Rose shrimp
The cruise parameter for the final model was estimated as3.03, implying that the selectivity (L50) of the spring and sum-er experiments is about 3 mm smaller compared to the autumn.he values of the selection parameters, by mesh size for theutumn and the combined spring and summer cruises, and theelection curves, based on the model of Table 1 are displayed inable 2 and Fig. 7, respectively.
Table 3 demonstrates the influence of mesh size on theodend retention proportion of individuals below the MLSunwanted catch) and the proportion retained by the coverbove this size (lost commercial catch). The main featuresmerging from the data concern the fast reduction in the catchf undersized shrimp with increasing mesh size, and a corre-ponding increase in the loss of commercial sized individuals,nd the differences between the autumn and spring/summerruises. In particular, the proportion of retention below MLSn the 55 and 70 mm codends was much higher (0.64 and 0.31)uring the spring/summer than in the autumn (0.43 and 0.14).n the other hand, the proportion of escapees above MLS didot show a similar overall pattern; for the 55 mm mesh codendhe proportion in the autumn was higher than in spring/summer0.32 versus 0.20) but the opposite happened for the 70 mmodend (0.49 versus 0.54).
.5. Red shrimp
As noted above, the final model (Table 1) contains no dif-erence in selectivity among the three cruises; i.e., only mesh

P. Fonseca et al. / Fisheries Research 85 (2007) 49–60 55
Table 2Selectivity parameters estimated for the three species in study, from the models presented in Table 1
Mesh size Parameter Estimate 95%CI S.E. Yield (kg h−1)
Species Total
Rose shrimpAutumn
55 mm μL50 22.38 21.23–23.51 0.57 13.8 10370 mm μL50 28.48 27.03–29.93 0.73 9.8 8180 mm μL50 32.55 30.89–34.20 0.83 18.5 138Common μSR 11.32 10.69–11.95 0.32Common SF 0.41 – –
Spring + summer55 mm μL50 19.34 18.32–20.37 0.52 16.4 10870 mm μL50 25.44 24.44–26.45 0.50 23.6 15280 mm μL50 29.51 28.48–30.55 0.52 20.7 102Common μSR 11.32 10.69–11.95 0.32
SF 0.36a – –
Red shrimpb
All cruises55 mm μL50 24.24 23.33–25.15 0.45 3.8 5270 mm μL50 30.85 29.69–32.01 0.58 5.6 6180 mm μL50 35.26 33.94–36.59 0.66 3.7 70Common μSR 11.07 10.59–11.55 0.24Common SF 0.44 – –
Norway lobsterAutumn
55 mm μL50 22.70 20.74–24.65 0.95 1.7 5070 mm μL50 28.89 28.40–31.37 1.21 1.0 4780 mm μL50 33.01 30.17–35.85 1.38 6.2 91Common μSR 7.38 6.66–8.11 0.35Common SF 0.41 – –
Spring + summer55 mm μL50 18.94 17.21–20.68 0.84 2.7 5970 mm μL50 25.13 23.45–26.82 0.82 2.2 8480 mm μL50 29.26 27.51–31.01 0.85 6.3 73Common μSR 7.38 6.66–8.11 0.35
SF 0.36a – –
SF represents the selection factor: L50/mesh size. Yields (in kg h−1) per mesh size, for both species and total catch are also presented.a These are average SFs since here L50 is not proportional to mesh size due to the cruise factor.
siastst0ciri
3
c
tml(c7
tsthSa
b Data corresponding to three cruises.
ize emerged as statistically significant. Therefore, the selectiv-ty parameters (L50 and SR) by mesh size presented in Table 2,long with their 95% confidence interval and complementarytatistics, and the selection curves plotted in Fig. 7, correspondo the data from the three cruises. The proportion of undersizedhrimps retained by the 55 mm codend was very high, 0.49, buthe use of a higher mesh size of 70 mm decreased that figure to.18, although at the costs of an increase in the loss of commer-ially sized individuals from 0.07 to 0.25 (Table 3). A furtherncrease in mesh size to 80 mm provided a further reduction inetention of undersized shrimp (0.12), but resulted in a greatlyncreased loss of those above the MLS (0.39).
.6. Norway lobster
Similarly to the rose shrimp, the modelling of selectivity overruises found a significant cruise effect (i.e., the autumn versus
a3ar
he spring and summer) (Table 1). The selection parameters byesh size can be found in Table 2, where the systematically
ower L50’s during the autumn trials (ca. 3.8 mm) are apparentFig. 7). In particular, the autumn 80 mm codend has a selectionurve that is almost coincident with that of the spring/summer0 mm codend.
Undersized individuals were seldom present (Fig. 5) in eitherhe codend or cover. Regarding the catches of commerciallyized lobsters, two main conclusions may be drawn. First, in allhree cruises, the increase of codend mesh size from 55 to 70 mmas an almost negligible impact (only 2–4% increase) in losses.econdly, there are major differences in retention between theutumn and spring/summer cruises, with the former displaying
consistently lower (about 20%) proportion. Using a MLS of0 mm, the L50 for the first maturation, reveals a situation whereconsiderable fraction of immature lobsters would always be
etained.

56 P. Fonseca et al. / Fisheries Research 85 (2007) 49–60
Table 3Proportions in codend (retained) and cover (escapees), below and above thecurrent MLS
Codend Cover
Rose shrimpAutumn
55 mm<MLS 0.43 0.57≥MLS 0.68 0.32
70 mm<MLS 0.14 0.86≥MLS 0.51 0.49
80 mm<MLS 0.12 0.88≥MLS 0.34 0.66
Spring + summer55 mm
<MLS 0.64 0.36≥MLS 0.80 0.20
70 mm<MLS 0.31 0.69≥MLS 0.46 0.54
80 mm<MLS 0.17 0.83≥MLS 0.33 0.67
Norway lobsterAutumn
55 mm<MLS 0.00 (0.51) 0.00 (0.49)≥MLS 0.75 (0.82) 0.25 (0.18)
70 mm<MLS 0.00 (0.38) 1.00 (0.62)≥MLS 0.73 (0.84) 0.27 (0.16)
80 mm<MLS 0.00 (0.33) 0.00 (0.67)≥MLS 0.48 (0.56) 0.52 (0.44)
Spring + summer55 mm
<MLS 0.00 (0.84) 1.00 (0.16)≥MLS 0.97 (0.99) 0.03 (0.01)
70 mm<MLS 1.00 (0.65) 0.00 (0.35)≥MLS 0.93 (0.95) 0.07 (0.05)
80 mm<MLS 0.00 (0.22) 1.00 (0.78)≥MLS 0.77 (0.85) 0.23 (0.15)
Red shrimpa
All cruises55 mm
<MLS 0.49 0.51≥MLS 0.93 0.07
70 mm<MLS 0.18 0.82≥MLS 0.75 0.25
80 mm<MLS 0.12 0.88≥MLS 0.61 0.39
For Nephrops, proportions in brackets correspond to the same estimates takingas a reference point the first maturation size (30 mm) instead of the current MLS(20 mm). Highlight in bold: proportions of undersized individuals retained andof commercial size lost.
a Data corresponding to three cruises.
Fig. 7. (a–c) Mean selection curves by mesh size for the three species in studyaccording to the estimates in Table 2. (—) 55 mm; (– – –) 70 mm; (· · ·) 80 mm.Far
4
4
scTswwpta
or rose shrimp and Norway lobster, black and grey lines correspond to theutumn and spring–summer estimates, respectively. The vertical dotted linesepresent the MLS and the length of first maturation (Lmat).
. Discussion
.1. Nature of the among-cruise differences
For two out of the three crustacean species under study (rosehrimp and Norway lobster) the modelling found a statisti-ally significant reduction (p < 0.001 and <0.05, respectively,able 1) in L50 between the autumn 1998 and the combinedpring and summer 1999 cruises. A similar reduction in L50as also observed for the ‘blue and red’ shrimp, however it
as not statistically significant (p = 0.11). This split correspondsrecisely to the use of polyamide twine in the codend mesh ofhe autumn cruise, and use of polyethylene twine in the springnd summer cruises. Thus, the effects of cruise (i.e., season)
ies Re
adstc
lHHaaiOpw2veCfiNare
imgistlsdacAas
ckssccPaLai0Nrddu
e1
4
hacJaccwmbi5ocascc
csoHt(fvsusa(utt
itv3ashture of stronger swimmers (larger individuals) which otherwisewould be able to escape (Wardle, 1983, 1986), but this potentialeffect is species-dependent. However, it is unlikely that poor-
P. Fonseca et al. / Fisher
re confounded with twine and therefore the explanation of theifference requires careful consideration of twine versus sea-onal effects based on existing knowledge on the life cycle ofhese species, and corresponding biochemical and morphologi-al alterations.
For fish species, body girth is considered the main morpho-ogical trait determining the retention by a given mesh size (e.g.,amley, 1975; Wileman et al., 1996; Reis and Pawson, 1992;ovgard and Lassen, 2000). On the other hand, periodic alter-
tion in external factors such as water temperature and foodvailability may also induce changes in selectivity by alter-ng fish behaviour and/or swimming capacity (Ozbilgin, 1998;¨ zbilgin et al., 2006). The latter studies suggest that swimmingower associated to fish ‘condition’ and water temperature mayell be more important than girth (see also, Ozbilgin and Wardle,002). No similar studies, or ad hoc evidences, of such seasonalariability in selectivity, based on either internal (biological) orxternal (environmental) factors are published for crustaceans.ontrary to fishes which exhibit a more or less active reaction toshing gears (e.g., Wardle, 1983, 1986, 1987) crustaceans, bothorway lobster and shrimps, have a much reduced swimming
bility and mainly react upon direct physical stimuli – ‘passive-eactive’ behaviour (see, Newland and Chapman, 1989; Hannaht al., 2003).
It would be plausible to reason that pre-spawning and spawn-ng females (including Norway lobster egg-carrying females)
ight have their swimming ability and behaviour towards fishingears partially impaired, thus affecting negatively the selectiv-ty in the spring and summer cruises, but no direct evidenceeems to be documented in the published literature. Moreover,he patterns of variation of the species biochemistry along theife cycle do not support the hypothesis of environmental (sea-onal) induced changes in their behaviour able to explain theifferences in selectivity found in this study. In fact, springnd summer seasons correspond to an overall better nutritionalondition (Rosa and Nunes, 2002a,b, 2003a,b, 2004, 2005).s such, the contrasting codend selection between the autumn
nd spring/summer surveys found for Norway lobster and rosehrimp is most likely attributable to codend twine material.
The higher selective properties of polyamide codends whenompared to those made up in polyethylene have long beennown (see Holden, 1971). Polyamide twine is generally lesstiff, has a lower roughness, and higher elasticity and exten-ibility (Klust, 1982) thus seemingly facilitating enmeshedrustacean to force themselves through meshes. Very few directomparative studies have been carried out (Isaksen et al., 1990;olet and Redant, 1994; Tokac et al., 2004; Deval et al., 2006),nd only the second and the fourth address crustaceans (Norwayobster and rose shrimp, respectively). These authors found that67 mm single braided PA codend displayed a higher selectiv-
ty than a 79 mm double braided PET codend (SF: 0.47 and..37, respectively). Here, the L50 values for rose shrimp andorway lobster were estimated to decrease by 3.0 and 3.8 mm,
espectively, when using the PET codend, and these estimatedecreases were statistically significant (p < 0.05). Red shrimpid not show a statistically significant decrease (p = 0.12) whensing the PET codend, but the estimated effect was in the
s
a
search 85 (2007) 49–60 57
xpected direction, and was an estimated decrease in L50 of.7 mm.
.2. Comparison with former studies
On account of their economic importance, these three speciesave been the subject of intensive research both on their biologynd on fisheries. In the early 1980s, a series of ad hoc studies onodend selectivity were carried out on Norway lobster (Mariaose Figueiredo, unpublished5; Figueiredo and Castro, 1983)nd shrimps (Ribeiro-Cascalho, 1988) off the Portugueseoast, on board INIAP/IPIMAR research vessels using non-ommercial gears. While the experiments on Norway lobsterere performed over a multitude of gears, codend mesh sizes,aterials, and haul durations, rendering difficult the compara-
ility with recent studies, those on shrimps were carried outn more controlled conditions, being concentrated mostly on5 mm PET codends. More recently, in the early 1990s, a seriesf experiments were carried out on a research vessel in controlledonditions and with a gear design closer to the commercial ones,iming at the characterization of the selectivity of Norway lob-ter and shrimps Campos et al. (2002, 2003). These studiesompared 55, 60 and 70 mm diamond, and 55 mm square meshodends.
The earlier experiments by Ribeiro-Cascalho (1988) werearried out on a small (27 m LOA and 463 HP) research ves-el, while those by Campos et al. (2002, 2003) were carriedut on the R/V Noruega, a much larger vessel (47.5 m, 1500P). The former study reports considerably higher selection fac-
ors (L50/mesh size) for diamond meshes than the latter study0.41–0.45 versus 0.39–0.40 and 0.46–0.49 versus 0.41–0.43or rose shrimp and ‘blue and red’ shrimp, respectively). Con-ersely, the results obtained in the current paper for the threepecies are closer to those in Campos et al. (2002, 2003) whosed Fryer’s method for incorporating between-haul variation inelectivity estimation (Fryer, 1991). However, for rose shrimpnd Norway lobster, similarity applies only to the autumn cruiseboth SF’s equal to 0.41) while in the spring/summer the SF val-es were much lower (0.36 and 0.34). It would be expected thathe selectivity of PET codends would be in the range of those ofhe former studies and that of the PA codends would be higher.
Explaining the reasons behind the similarities and differencess not straightforward, when data were obtained under con-rasting conditions: different codend materials, different sizedessels and tow duration (1 h onboard the research vessel versush under commercial conditions). Vessel size is one of the vari-bles thought to have a potential influence on selectivity but haseldom been investigated (Tschernij and Holst, 1999). Longerauls are likely to affect the selectivity by promoting the cap-
wimming species such as crustaceans would be affected by
5 Available as INIP (currently INIAP/IPIMAR) internal reports no. 24 (1984)nd nos. 48 and 49 (1985).

5 ies Re
ccamcpcbyecirafmgpvosofciC
ctAoroisa4tds1P1dtrr(ecv
ctCto
Ift2oaccioof0air0S(6r
sdHswcmlfsarioo
4
mvrlteacaio
8 P. Fonseca et al. / Fisher
hanging haul duration. It is conceivable that longer commer-ial hauls may have resulted in an overall decrease of selectivity,cross mesh sizes and codend twine materials, due to codendesh saturation, but this would imply that catches during the
ommercial hauls had been much higher. However, a com-arative catch analysis showed that, besides the differences inrustacean yields (in kg h−1), attributable to cyclic alternationetween rose shrimp and Norway lobster abundance, overallields were considerably higher (maximum about 4:1) in thearly study due to the catch of large quantities of both commer-ial and non-commercial fish by-catch. This is a well-knownnfluencing factor on selectivity, although with contradictoryesults (positive or negative correlation with L50 and/or SR)ccording to the different authors. The between-studies dif-erences discussed herein seem to be in accordance with theechanical hypothesis by O’Neill and Kynoch (1996), who sug-
est that for relatively low catches (<500 kg) this relationship isositive, becoming negative for higher catches. In fact, a pre-ious work by Charreau (1992) based on experiments carriedut for Norway lobster between 1975 and 1980 had already pre-ented an expression relating the increase in SF with the weightf by-catch. Consequently, the overall lower selectivity resultsound in recent experiments may be explained based on the lowatch rates which, in the case of the Autumn PA codend exper-ments, brought the SF’s to the same level of PET codends inampos et al. (2002).
Similar studies for other stocks of the same species have beenarried out from the northeastern Atlantic (for Norway lobster)o the Mediterranean sea (for all three species). While for thetlantic fisheries the codend mesh sizes tested are in the rangef those under analysis, in the Gulf of Cadiz and the Mediter-anean, where commercial mesh sizes are much smaller (40 mmr less), meshes over 50 mm were seldom used during selectiv-ty trials. In a review of Norway lobster diamond mesh codendelectivity trials carried out from 1965 to 1984, Briggs (1986)nalysed data from Iceland to the Gulf of Cadiz (mesh sizes from4 to 80 mm; SF between 0.31 and 0.78) and concluded thathe highly contrasting selection results were intrinsically depen-ent ‘upon the gear used and prevailing conditions’. Subsequenttudies were carried out both in the Atlantic (Robertson et al.,986; Kirkgaard et al., 1989; Larsvik and Ulmestrand, 1992;olet and Redant, 1994) and in the Mediterranean (Sarda et al.,993; Stergiou et al., 1997; Mytilineou et al., 1998). Althoughisplaying a narrower range of variation for SF (0.31–0.57) forhe same mesh size range as those prior to 1986, their resultseinforce the importance of codend characteristics (twine mate-ial/twine thickness) (Polet and Redant, 1994) and mesh shapeLarsvik and Ulmestrand, 1992; Robertson et al., 1986; Stergiout al., 1997). The specific conditions of the fisheries, such as theatch rates, emerge as the likely explanation for the remainingariability in the results.
Rose shrimp has its northern East-Atlantic limit at the northoast of Spain, being the subject of important fisheries from
he southwest and south coasts of Portugal through the Gulf ofadiz, and the Mediterranean Sea. The ‘blue and red’ shrimp dis-ribution is more restricted, it is fished in the southwest and southf Portugal but its main fisheries are in central Mediterranean.
cte(
search 85 (2007) 49–60
n recent years, codend selection studies have been carried outor rose shrimp in the Gulf of Cadiz (Sobrino et al., 2000) andhe Mediterranean (Rinelli et al., 2005; Guijarro and Massuti,006; Ragonese and Bianchini, 2006; Deval et al., 2006), andnly in the Mediterranean for ‘blue and red’ shrimp (Carlucci etl., 2006; Guijarro and Massuti, 2006). For rose shrimp, there isonsiderable variability in the SF values, although apparently theodend material was always the same (PA), with the value foundn our study for the polyamide codend (0.41) being in the rangef those presented by those authors. Sobrino et al. (2000) carriedut experiments both in commercial and research vessels, andound that SF results were higher in the latter (0.39–0.49 versus.32–0.37). On the other hand, Rinelli et al. (2005), Ragonesend Bianchini (2006) and Guijarro and Massuti (2006) in exper-ments carried out on board commercial vessels and within aestrict mesh size range (31–40 mm) obtained SF values between.39 and 0.44, most of them closer to the upper limit. Red shrimpF data from Carlucci et al. (2006) and Guijarro and Massuti2006) studies were 0.39 (40 mm mesh size) and 0.43 (50 and0 mm), respectively, the latter being very close to our ownesults of 0.44.
The restricted number of studies carried out so far on shrimpelectivity limits the interpretations of the underlying factorsetermining the observed variability in the selection of shrimps.owever, a very contrasting trend is observed between the
hrimp selection ranges (L75–L25) on the Portuguese coast,hich are much higher, and those in the Mediterranean, espe-
ially for codend mesh sizes less than 60 mm. Although lessarked, the same overall trend is observable for the Norway
obster. In their study for the latter species, Campos et al. (2002)ound a statistically significant positive influence of both meshize and codend catch on the selection range. Although thoseuthors did not identify the same factors as determinants for theose shrimp selectivity, from the comparison of the various stud-es it is noticeable that different ranges of mesh size exhibit SR’sf very different magnitude, and even the potential influence ofverall catch cannot be ruled out.
.3. Potential impact of altering mesh size
The simultaneous exploitation of species with very differentorphologic characteristics and life cycles is a complex conser-
ation problem that calls for very strict and specific managementules. However, the current legislation at both EU and nationalevels does not take into account the different characteristics ofhe shrimp species caught in this fishery, especially their differ-nt size ranges. Both the rose and the ‘blue and red’ shrimps,nd also the giant red shrimp may be fished with trawls usingodend mesh sizes from 55 to 59 mm, provided they constitute,t least, 30% of the catch, while for the Norway lobster the min-mum mesh size (MMS) for a targeted fishery should be equalr higher than 70 mm.
From Table 3 it becomes obvious that the use of 55 mm
odend mesh sizes is inadequate for both shrimp species givenhe extremely high number of undersized individuals retained,specially for the rose shrimp during the spring/summerpolyethylene codends) trials (64%). However, given the size
ies Re
rf7iacot‘li
v5folhppstiwclti
5
tcbsspssaiciasalecdif
t
os
tcaeasstatoaobm
A
‘KlFokrpT
R
B
C
C
C
C
C
P. Fonseca et al. / Fisher
anges of both species no single mesh size would be adequateor improving the fishing pattern. While the use of a MMS of0 mm would result in a large decrease in the catch of undersizedndividuals for both species (from about 50–67%), the losses ofround 43% of commercial sized rose shrimp would be unac-eptable. In any case, an increase in mesh size to at least 60 mmr, in a first phase, the adoption of polyamide netting would con-ribute to lower the catches under MLS, and since shrimps arerelatively’ short-lived,6 high-fecundity, fast-growing species,osses would be partially compensated in the short term by thencrease of the available biomass.
The case of the Norway lobster is apparently different; indi-iduals below the current MLS are rare in the catches, even when5 mm codends are used. However, the 50% maturation lengthor this species is about 30 mm, meaning that a high percentagef immature lobsters are being retained (65% for the polyethy-ene 70 mm codend). This status quo certainly contributes to aigh extent, along with overfishing, for the current state of theopulation units which led to the implementation of a recoverylan (E.U., 2005). Furthermore, the fact that both Norway lob-ter and ‘blue and red’ shrimp are not separately targeted sinceheir depth distribution partially overlaps, adds to the complex-ty of their management. Since they may be legally capturedith different mesh sizes, there is no guarantee that all skippers
omply with the mandatory mesh size when targeting Norwayobster. Nevertheless, the fact that by 2004 more than 80% ofhe crustacean trawlers were licensed for using both mesh sizess somewhat encouraging.
. Conclusions
The overall results obtained in the current study establishedhe importance of codend characteristics in the selection ofrustacean species, which dominates any potential influence ofehavioural changes along their life cycle. Actually, although theelection of both shrimp and Norway lobster has been demon-trated not to be a purely mechanical question, they display aassive behaviour during most part of the catch process. Asuch, it is likely that other factors (in addition to mesh size)uch as changing mesh opening and twine stiffness will haven impact on the retention of the different length classes. Theres little doubt that the use of polyethylene twine was the mainause of the smaller estimated L50’s in the spring–summer exper-ments. This reduction in L50 was observed for all three species,nd was statistically significant for Norway lobster and rosehrimp (Table 1). Former studies carried out both in the Atlanticnd the Mediterranean, especially those concerning the Norwayobster partially support that conclusion. Moreover, the differ-nces in selectivity (mainly in L50) compared to former studies
arried out in Portuguese waters can be attributed with a greategree of certainty to the differences in by-catch of fish speciesn both periods. This confirms what has long been referencedor the Norway lobster and strongly suggests that the influence6 The life span of the rose shrimp is about 2–3 years, while for the red shrimphe majority of the studies point to 4–5 years.
D
E
search 85 (2007) 49–60 59
f total catch weight on mesh opening is equally important forhrimps.
The adoption of management measures to prevent the reten-ion of undersized and immature individuals based on codendharacteristics, should therefore take into consideration thebove conclusions, to minimize as much as possible the influ-nce of external factors on the selection. Amongst the potentiallternatives to a simple increase in mesh size, the use of square-haped mesh codends was shown to be a practical and effectiveolution by Campos et al. (2002, 2003). That mesh configura-ion displayed a much higher selectivity, both for Norway lobsternd shrimp species. Simultaneously, it proved to be a very effec-ive way of releasing low-commercial fish by-catch with smallerr null losses of highly valuable species. A further alternative,lready successfully tested by Fonseca et al. (2005), consistsf the use of a rigid selective grid placed in an extension pieceefore the codend, allowing for the release of fish by-catch withinor losses of the crustacean target species.
cknowledgements
The collaboration of Mestre ‘Zeca’, skipper of the F/VPorto Bravo’, and his crew is gratefully acknowledged. Prof.arim Erzini’s (University of Algarve) keen comments on the
ast version of the manuscript were much appreciated. Dr.rancesc Sarda (ICM, Barcelona) provided helpful informationn shrimps and Norway lobster biology. The suggestions andnowledgeable comments by the referees facilitated much theevision process. This work was partially financed by the Euro-ean Commission under the Study contract 96/61 “TRASEL –rawl Selectivity in Region 3”.
eferences
riggs, R.P., 1986. A general review of mesh selection for Nephrops norvegicus(L.). Fish. Res. 4, 59–73.
ampos, A., Fonseca, P., Erzini, K., 2002. Size selectivity of diamond and squaremesh cod ends for rose shrimp (Parapenaeus longirostris) and Norway lob-ster (Nephrops norvegicus) off the Portuguese south coast. Fish. Res. 58,281–301.
ampos, A., Fonseca, P., Erzini, K., 2003. Size selectivity of diamond and squaremesh cod ends for four by-catch species in the crustacean fishery off thePortuguese south coast. Fish. Res. 60, 79–97.
ampos, A., Fonseca, P., Fonseca, T., Parente, J., 2007. Definition of fleetcomponents in the Portuguese bottom trawl fishery 3. Fish. Res. 83, 185–191.
arlucci, R., D’Onghia, G., Sion, L., Maiorano, P., Tursi, A., 2006. Selectivityparameters and size at first maturity in deep-water shrimps, Aristaeomorphafoliacea (Risso, 1827) and Aristeus antennatus (Risso, 1816), from the North-Western Ionian Sea (Mediterranean Sea). Hydrobiologia 557, 145–154.
harreau, A., 1992. Preliminary investigations of the use of a flume tank forthe study of Nephrops selectivity. In: EC Workshop on Cod-End Selectivity.Marine Laboratory Aberdeen, Scotland.
eval, M.C., Bok, T., Ates, C., Ozbilgin, H., 2006. Selectivity of PE and PAmaterial codends for rose shrimp (Parapenaus longirostris) in Turkish twinrigged beam trawl fishery. Fish. Res. 81, 72–79.
.U., 2005. Council Regulation (EC) No. 2166/2005 of 20 December 2005 estab-lishing measures for the recovery of the Southern hake and Norway lobsterstocks in the Cantabrian Sea and Western Iberian peninsula and amendingRegulation (EC) No. 850/98 for the conservation of fishery resources throughtechnical measures for the protection of juveniles of marine organisms.

6 ies Re
F
F
F
G
H
H
H
H
I
I
I
I
I
K
K
L
M
M
M
N
O
O
O
O
P
P
R
R
R
R
R
R
R
R
R
R
R
S
S
S
S
T
T
W
W
W
0 P. Fonseca et al. / Fisher
igueiredo, M.J., Castro, M., 1983. Studies on the Selectivity of Nephrops offyhr Portuguese Coast. ICES CM 1983/K:27.
onseca, P., Campos, A., Larsen, R., Borges, T.C., Erzini, K., 2005. Using amodified Nordmøre grid for by-catch reduction in the Portuguese crustaceantrawl fishery. Fish. Res. 71, 223–239.
ryer, R.J., 1991. A model of between-haul variation in selectivity. ICES J. Mar.Sci. 48, 281–290.
uijarro, B., Massuti, E., 2006. Selectivity of diamond- and square-meshcodends in the deepwater crustacean trawl fishery off Balearic Islands (west-ern Mediterranean). ICES J. Mar. Res. 63, 52–67.
amley, J.M., 1975. Review of Gill-net selectivity. J. Fish. Res. Board Can. 32,1943–1969.
annah, R.W., Jones S.A., Matteson, K.M., 2003. Observations of fish andshrimp behaviour in ocean shrimp (Pandalus jordani) Trawls. OregonDepartment of Fish Wild., Information Rept. Ser., Fish. No. 2003-03, 28pp.
olden, M.J. (Ed.), 1971. Report of the ICES/ICNAF Working Groups onSelectivity Analysis. Coop. Res. Rep. ICES, Ser. A, No. 25, 144 pp.
ovgard, H., Lassen, H., 2000. Manual on estimation of selectivity for gillnetand longline gears in abundance estimation. FAO Fish. Tech. Paper 397, 84pp.
CES, 2001. Report of the Working Group on Nephrops Stocks. ICES CM2001/ACFM:16.
CES, 2002. Report of the Working Group on Nephrops Stocks. ICES CM2002/ACFM:15.
CES, 2003. Report of the Working Group on Nephrops Stocks. ICES CM2003/ACFM:18.
CES, 2004. Report of the Working Group on Nephrops Stocks. ICES CM2004/ACFM:19.
saksen, B., Lisovsky, S., Sakhno, V.A., 1990. A Comparison of the Selectivityin Codends Used by the Soviet and Norwegian Trawler Fleet in the BarentsSea. ICES CM 1990/B:51.
irkgaard, E., Nielsen, N.A., Bagge, O., 1989. Mesh Selection of Nephrops in60 and 70 mm Nephrops Trawl. ICES CM 1989/B:32, 22 pp.
lust, G., 1982. Netting Materials for Fishing Gear, second ed. Fishing NewsBooks Ltd., England, p. 175.
arsvik, M., Ulmestrand, L., 1992. Square and Diamond Mesh Trawl CodendSelection on Nephrops norvegicus (L.), Analysed with the Curve-Fit MethodIsotonic Regression. ICES CM 1992/B:36.
illar, R.B., 1994. Sampling from trawl gears used in size selectivity experi-ments. ICES J. Mar. Sci. 51, 293–298.
illar, R.B., Broadhurst, M.K., Macbeth, W.G., 2004. Modelling between-haulvariability in the size selectivity of trawls. Fish. Res. 67, 171–181.
ytilineou, C., Politou, C.-Y., Fourtouni, A., 1998. Trawl selectivity studies inNephrops norvegicus (L.) in the eastern Mediterranean Sea. Sci. Mar. 62(Suppl. 1), 107–116.
ewland, P.L., Chapman, C.J., 1989. The swimming and orientation behaviourof the Norway lobster, Nephrops norvegicus (L.), in relation to trawling.Fish. Res. 8, 63–80.
’Neill, F.G., Kynoch, R.J., 1996. The effect of cover mesh size and cod-endcatch size on cod-end selectivity. Fish. Res. 28, 291–303.
¨ zbilgin, H., 1998. The seasonal variation of trawl cod-end selectivity and therole of learning in mesh penetration behaviour of fish. Thesis Presentedfor the Degree of Doctor of Philosophy at the University of Aberdeen. 206pp. + Annexes.
¨ zbilgin, H., Wardle, C.S., 2002. Effect of seasonal temperature changes on theescape behaviour of haddock, Melanogrammus aeglefinus, from the codend.Fish. Res. 58, 323–331.
¨ zbilgin, H., Ferro, R.S.T., Robertson, J.H.B., Holtrop, G., Kynoch, R.J., 2006.Seasonal variation in trawl codend selection of northern North Sea haddock.
ICES J. Mar. Sci. 63, 737–748.estana, G., Ribeiro-Cascalho, A., 1991. Effects of changing trawl mesh size andfishing effort on deep water Rose shrimp (Parapenaeus longirostris) fromthe Southern Portugal (ICES Division IXa). ICES, Shellfish Committee. CM1991/K.45, 18 pp.
W
search 85 (2007) 49–60
olet, H., Redant, F., 1994. Selectivity experiments in the Belgian Norwaylobster (Nephrops norvegicus) fishery. ICES CM 1994/B:39, 10 pp.
agonese, R., Bianchini, M.L., 2006. Trawl selectivity trials on the deep-waterrose shrimp (Parapenaeus longirostris) in Sicilian waters. Hydrobiologia557, 113–119.
eis, E.G., Pawson, M.G., 1992. Determination of gill-net selectivity for bass(Dicentrarchus labrax L.) using commercial catch data. Fish Res. 13,173–187.
inelli, P., Giordano, D., Perdichizzi, F., Greco, S., Ragonese, S., 2005. Trawlgear selectivity on the deep-water rose shrimp (Parapenaeus longirostris,Lucas, 1846) in the Southern Tyrrhenian Sea (central Mediterranean). Cah.Biol. Mar. 46, 1–7.
ibeiro-Cascalho, A., 1988. Biology, Ecology and Fishery of Deep-WaterShrimps, Parapenaeus longirostris and Aristeus antennatus in PortugueseWaters. Instituto Nacional de Investigacao das Pescas. Dezembro de 1998,169 pp. (in Portuguese).
obertson, J.H.B., Emslie, D.C., Ballantyne, K.A., Chapman, C.J., 1986. Squareand Diamond Mesh Trawl Cod-End Selection Trials on Nephrops norvegicus(L.). ICES CM 1986/B:12, 5 pp.
osa, R., Nunes, M.L., 2002a. Biological changes during the reproductive cycleof the deep-sea decapod Nephrops norvegicus on the south coast of Portugal.Mar. Biol. 141, 1001–1009.
osa, R., Nunes, M.L., 2002b. Influence of the reproductive cycle on the bio-chemical composition of deep-sea decapod Parapenaeus longirostris (Lucas,1846) in the Portuguese south coast. J. Shellfish Res. 21, 843–849.
osa, R., Nunes, M.L., 2003a. Tissue biochemical composition in relation tothe reproductive cycle of deep-sea decapod Aristeus antennatus in the Por-tuguese south coast. J. Mar. Biol. Assoc. U.K. 83, 963–970.
osa, R., Nunes, M.L., 2003b. Seasonal changes in nucleic acids, amino acidsand protein content in juvenile Norway lobster (Nephrops norvegicus). Mar.Biol. 143, 565–572.
osa, R., Nunes, M.L., 2004. RNA, DNA and protein concentrations andamino acid profiles of deep-sea decapod Aristeus antennatus: an indicationfor seasonal variations of nutrition and growth. Aquat. Living Resour. 17,25–30.
osa, R., Nunes, M.L., 2005. Seasonal patterns of nucleic acid concentrationsand amino acid profiles of Parapenaeus longirostris (Crustacea, Decapoda):relation to growth and nutritional condition. Hydrobiologia 537, 207–216.
arda, F., Conan, G.Y., Fuste, X., 1993. Selectivity of Norway lobster Nephropsnorvegicus (L.) in the northwestern Mediterranean. Sci. Mar. 57, 167–174.
obrino, I., Garcıa, T., Baro, J., 2000. Trawl gear selectivity and the effect ofmesh size on the deep-water rose shrimp (Parapenaeus longirostris, Lucas,1846). Fish. Res. 44, 235–245.
tergiou, K.I., Petrakis, G., Politou, C.-Y., 1997. Size selectivity of diamondand square mesh cod-ends for Nephrops norvegicus in the Aegean Sea. Fish.Res. 29, 203–209.
tewart, P.A.M., Robertson, J.H.B., 1985. Small mesh codend covers. ScottishFish. Res. Rep. No. 32, 11 pp.
schernij, V., Holst, R., 1999. Evidence of Factors at Vessel-Level AffectingCodend Selectivity in Baltic Demersal Fishery. ICES CM 1999/R:02.
okac, A., Ozbilgin, H., Tosunoglu, Z., 2004. Effect of PA and PE material oncodend selectivity in Turkish bottom trawl. Fish. Res. 67, 317–327.
ardle, C.S., 1983. Fish reactions to fishing gears. In: MacDonald, A.G., Priede,I.G. (Eds.), Experimental Biology at Sea. Academic Press, London, pp.168–195.
ardle, C.S., 1986. Fish behaviour and fishing gears. In: Pitcher, T.J. (Ed.),The Behaviour of Teleost Fishes. Croom Helm, London and Sydney, pp.463–495.
ardle, C.S., 1987. Investigating the behaviour of fish during capture. In: Bailey,
R.S., Parrish, B.B. (Eds.), Developments in Fisheries Research in Scotland.Fishing News Books, London, pp. 139–155.ileman, D., Ferro, R.S.T., Fonteyne, R., Millar, R.B., 1996. Manual of Methodsof Measuring the Selectivity of Towed Fishing Gear. ICES Coop. Res. Rep.215, 126 pp.




















