
Cloning, overexpression, purification, and matrix-assisted refolding of DevS (Rv 3132c) histidine...
6
Protein Expression and Purification 25, 203-208 (2002) doi: 10.1006/prep.2002.1628 ® Cloning, Overexpression, Purification, and Matrix-Assisted Refolding of DevS (Rv 3132c) Histidine Protein Kinase of Mycobacterium tuberculosis Deepak Kumar Saini,* Neha Pant,* Taposh K. Das,1- and Jaya Sivaswami Tyagi *,1 :kDepartment of Biotechnology and "(Department of Anatomy, All India Institute of Medical Sciences, New Delhi 110029, India Received October 30, 2001, and in revised form January 16, 2002 The devR-devS (Rv 3133c-Rv 3132c) two-component system of Mycobacterium tuberculosis was identified in our laboratory by RNA subtractive hybridization. This genetic system was predicted to encode a response regulator and histidine protein kinase, respectively. The putative histidine kinase protein DevS was over- expressed to high levels in Escherichia coli as a fusion protein with a hexahistidine tag, His(6)-DevS2ol, in the form of inclusion bodies. Here we report a "redox- based" method of matrix-bound renaturation of DevS protein. The refolded protein was biochemically active in an autophosphorylation reaction characteristic of histidine kinases and was suitable for the generation of polyclonal antibodies and as an antigen in ELISA. ~ 2002 Elsevier Science (USA) Key Words: Mycobacterium tuberculosis; two-compo- nent system; sensor kinase; poly-His tag; inclusion bod- ies; protein renaturation. The devR-devS two-component system of Mycobacte- rium tuberculosis (Rv 3133c-Rv 3132c) was identified by RNA subtractive hybridization among several genes differentially expressed in the virulent H37Rv strain of M. tuberculosis cultured in vitro (1). This genetic system is conserved in organisms belonging to the M. tuberculosis complex. The genes are cotranscribed to- gether with an upstream gene, Rv 3134c, encoding a putative alanine-valine-rich protein. The devR gene is also expressed in vivo within human monocytes (2). A To whom correspondence should be addressed at the Department of Biotechnology, All India Institute of Medical Sciences, New Delhi 110029, India. Fax: + +91-11-6862663. E-mail: [email protected]. 1046-5928/02 $35.00 © 2002 Elsevier Science (USA) All rights reserved. devR mutant strain of M. tuberculosis is attenuated in virulence in guinea pigs (Kapur et al., submitted for publication), implicating the devR-devS system in the virulence phenotype of M. tuberculosis. The devS gene is predicted to encode a 578-amino-acid-long protein that shows homology to typical histidine kinase (HK)2 proteins in its C-terminal region. The C-terminal region of HKs comprises the kinase catalytic core, -350 amino acids in length, and contains five conserved amino acid motifs, namely H, N, D/G1, and G2 boxes. In most HKs, the H box is part of the dimerization domain and the N, D/G1, and G2 boxes form the ATP-binding site (3, 4). HKs undergo an ATP-dependent autophospho- rylation at a conserved His residue in the H box. In DevS of M. tuberculosis, the conserved His residue at position 395 is predicted to be the site of phosphoryla- tion. The kinase catalytic core of several HKs has been phosphorylated in vitro at the conserved His residue using ATP (3, 4). Several laboratories have expressed HKs in soluble form in Escherichia coli and purified them by conventional procedures (4-6). Here we report the overexpression and refolding of the 201-amino-acid- long kinase C-terminal catalytic core of DevS (desig- nated DevS2ol). Overexpression in E. coli always re- sulted in the localization of His(6)-DevS2ol in inclusion bodies. The insoluble protein was not amenable to solubilization and refolding by the conventional strate- gies of dialysis or dilution-based refolding (7-9). Alter- natively, the protein was folded on solid matrix as re- ported earlier (10). These protocols have largely led to solubilization of proteins whose biochemical status was 2 Abbreviations used: HK, histidine protein kinase; IPTG, isopro- pyl- 1-thio-fl-D-galactopyranoside; Gm- HC1, guanidinium hydrochlo- ride; GSSG, oxidized glutathione; GSH, reduced glutathione. 203
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cloning, overexpression, purification, and matrix-assisted refolding of DevS (Rv 3132c) histidine...

Protein Expression and Purification 25, 203-208 (2002)
doi: 10.1006/prep.2002.1628 ®
Cloning, Overexpression, Purification, and Matrix-Assisted Refolding of DevS (Rv 3132c) Histidine Protein Kinase of Mycobacterium tuberculosis
Deepak K u m a r Saini,* Neha Pant,* Taposh K. Das,1- and J a y a Sivaswami Tyagi *,1 :kDepartment of Biotechnology and "(Department of Anatomy, All India Institute of Medical Sciences, New Delhi 110029, India
Received October 30, 2001, and in revised form January 16, 2002
The devR-devS (Rv 3133c-Rv 3132c) t w o - c o m p o n e n t s y s t e m of Mycobacterium tuberculosis w a s ident i f i ed in our l a b o r a t o r y by R N A s u b t r a c t i v e h y b r i d i z a t i o n . This g e n e t i c s y s t e m w a s p r e d i c t e d to e n c o d e a r e s p o n s e r e g u l a t o r and h i s t i d i n e p r o t e i n k inase , respect ive ly . The p u t a t i v e h i s t i d i n e k i n a s e p r o t e i n D e v S w a s over- e x p r e s s e d to h igh l eve l s in Escherichia coli a s a f u s i o n p r o t e i n w i t h a h e x a h i s t i d i n e tag, His(6)-DevS2ol, in the f o r m o f i n c l u s i o n bodies . Here w e repor t a "redox- based" m e t h o d of m a t r i x - b o u n d r e n a t u r a t i o n of DevS prote in . The re fo lded p r o t e i n w a s b i o c h e m i c a l l y ac t ive in an a u t o p h o s p h o r y l a t i o n r e a c t i o n c h a r a c t e r i s t i c of h i s t i d i n e k i n a s e s and w a s su i tab le for the g e n e r a t i o n of p o l y c l o n a l a n t i b o d i e s and as an a n t i g e n in ELISA. ~ 2002 E l sev ier S c i e n c e (USA)
Key Words: Mycobacterium tuberculosis; t w o - c o m p o - n e n t system; s e n s o r k inase; po ly-His tag; i n c l u s i o n bod- ies; p r o t e i n r e n a t u r a t i o n .
The devR-devS two-component system of Mycobacte- rium tuberculosis (Rv 3133c-Rv 3132c) was identified by RNA subtractive hybridization among several genes differentially expressed in the virulent H37Rv strain of M. tuberculosis cultured in vitro (1). This genetic system is conserved in organisms belonging to the M. tuberculosis complex. The genes are cotranscribed to- gether with an upstream gene, Rv 3134c, encoding a putative alanine-valine-rich protein. The devR gene is also expressed in vivo within human monocytes (2). A
To whom correspondence should be addressed at the Department of Biotechnology, All India Institute of Medical Sciences, New Delhi 110029, India. Fax: + +91-11-6862663. E-mail: [email protected].
1046-5928/02 $35.00 © 2002 Elsevier Science (USA) All rights reserved.
devR mutant strain of M. tuberculosis is attenuated in virulence in guinea pigs (Kapur et al., submitted for publication), implicating the devR-devS system in the virulence phenotype of M. tuberculosis. The devS gene is predicted to encode a 578-amino-acid-long protein that shows homology to typical histidine kinase (HK) 2 proteins in its C-terminal region. The C-terminal region of HKs comprises the kinase catalytic core, -350 amino acids in length, and contains five conserved amino acid motifs, namely H, N, D/G1, and G2 boxes. In most HKs, the H box is part of the dimerization domain and the N, D/G1, and G2 boxes form the ATP-binding site (3, 4). HKs undergo an ATP-dependent autophospho- rylation at a conserved His residue in the H box. In DevS of M. tuberculosis, the conserved His residue at position 395 is predicted to be the site of phosphoryla- tion. The kinase catalytic core of several HKs has been phosphorylated in vitro at the conserved His residue using ATP (3, 4). Several laboratories have expressed HKs in soluble form in Escherichia coli and purified them by conventional procedures (4-6). Here we report the overexpression and refolding of the 201-amino-acid- long kinase C-terminal catalytic core of DevS (desig- nated DevS2ol). Overexpression in E. coli always re- sulted in the localization of His(6)-DevS2ol in inclusion bodies. The insoluble protein was not amenable to solubilization and refolding by the conventional strate- gies of dialysis or dilution-based refolding (7-9). Alter- natively, the protein was folded on solid matrix as re- ported earlier (10). These protocols have largely led to solubilization of proteins whose biochemical status was
2 Abbreviations used: HK, histidine protein kinase; IPTG, isopro- pyl- 1-thio-fl-D-galactopyranoside; Gm- HC1, guanidinium hydrochlo- ride; GSSG, oxidized glutathione; GSH, reduced glutathione.
203

2 0 4 SAINI ET AL.
not assessed. We report the renaturation of His(6)- DevS2ol of M. tuberculosis by matrix-assisted refolding to yield biochemically active protein that is also suitable for antibody generation and ELISA and represents to the best of our knowledge the first example of on-column refolding of a histidine kinase.
MATERIALS AND METHODS
Cloning of DevS2ol. A 603-bp DNA fragment that maps within the DevS coding region and encodes the putative kinase core (amino acid residues 378 to 578) was selected for overexpression in E. coli. Its transla- tion product, DevS2o~, is predicted to contain the charac- teristic kinase core, but to lack the putative transmem- brane regions, TM1 to TM3 (Fig. 1). DNA was amplified using Pfu polymerase (Promega, Madison, WI) from M. tuberculosis H37Rv DNA using primers BamF and BamR with a BamHI restriction site engineered in them (Fig. 1). The amplicon was digested with BamHI and ligated into BamHI-digested pPROEx-HTc expression vector (Invitrogen, Inc., San Diego, CA). This expression system is inducible with isopropyl-l-thio-fl-D-galacto- pyranoside (IPTG) and foreign proteins are expressed with a covalently attached N-terminal His(6) tag.
Overexpression of DevS2ol. An overnight culture of E. coli BL21(DE3) transformed with recombinant pPROEx-HTc-devS2ol was grown at 37°C in 250 ml of 2×YT containing ampicillin (100/~g/ml) to an OD6oo of 0.4-0.6. Recombinant protein expression was induced by the addition of 0.6 mM IPTG (final concentration) and the culture was incubated at 37°C with vigorous shaking for an additional 5 h. The cells were centrifuged at 8000g for 15 min at 4°C and checked for expression of the fusion protein by 12.5% SDS-polyacrylamide gel electrophoresis (PAGE) under reducing conditions and stored at -20°C.
Solubilization of inclusion bodies. The cell pellet was resuspended in 10 ml of solubilization]denatur- ation buffer (20 mM Tris • HC1, pH 8.0, 500 mM NaC1,
10% glycerol containing 8 M urea) and incubated at 37°C for 2 h with shaking, followed by centrifugation at 12,000g for 15 min to remove the cell debris and other insoluble material. The supernatant fraction con- taining the solubilized inclusion bodies was processed for protein purification. Gm • HC1 at 6 M could also be used in place of urea with identical results.
DevS2ol purification and matrix-assisted refolding. The purification and refolding scheme employed for DevS2ol is outlined in Fig. 2. The denatured proteins were loaded onto a 2-ml column of preequilibrated Ni+2-NTA agarose (Qiagen, GmbH, Germany) and in- cubated for 2 h at room temperature. The column was shifted to 16°C and the unbound protein was collected as flowthrough. The column was washed with 3 bed volumes of denaturation buffer containing 20 mM imid- azole and then washed stepwise with decreasing con- centrations of urea (6 to 1 M) in refolding buffer (20 mM Tris • HC1, pH 8.0, 500 mM NaC1, 10% glycerol, 20 mM imidazole, 0.5 mM oxidized glutathione (GSSG) and 5 mM reduced glutathione (GSH)). The column was then washed with 2 bed volumes of refolding buffer without urea to remove contaminating E. coli proteins. The bound protein was eluted with 5 ml refolding buffer containing 250 mM imidazole in fractions of 1 ml. The fractions were checked by SDS-PAGE and the peak fractions (generally fractions 2-5) were pooled, dialyze d against storage buffer (50 mM Tris • HC1, pH 8.0, 50 mM NaC1, 50% glycerol, 0.1 mM DTT), and stored at -20°C. The protein was quanti tated by Bradford's method of protein estimation using BSA as the stand- ard (12). The use of GSSG and GSH provided a good indication of the redox milieu of the column. As the GSSG/GSH entered the column, the oxidation state of Ni +2 changed and the matrix color also changed gradu- ally from bluish white to white and finally pinkish white. During elution with buffer containing 250 mM imidazole, the oxidation state and the color returned to blue, serving as a useful indicator of elution of refolded protein. The non-redox-refolded DevS201 was renatured
Sensor domain
TM1 TM2
203 295
TM3
342
Kinase catalytic core I
H395 [L~ N DIG1 G2
~- 578 DevS
4 - BamR
H395 . , ~ N DIG1 G2 378 ~ []- 578 DevS~..,.
FIG. 1. Salient features of the DevS protein. The three N-terminal transmembrane domains (TM1, TM2, TM3) of DevS were predicted by TMpred (www.expasy.ch) and are shown as black boxes. The H box containing the conserved His-395 residue and the N, D/G1, and G2 boxes are shown in gray. The 201-amino-acid-long cytosolic fragment located in the C-terminal region of DevS is referred to as DevS2o~.

DEVS HISTIDINE KINASE OF Mycobacterium tuberculosis
I IPTG - induced cell culture
l I Solubilized protein I
[ (His)6DevS2o, -immobilized on Ni .2 - NTA agarose l
Column washed with stepped urea I gradient in refolding buffer (8 - 0 M urea) I
l I Column washed with refolding buffe~
Refolded (His)sDevS2ol eluted with imidazole buffer
RESULTS FIG. 2. Flow chart of matrix-assisted purification and redox-based refolding of His(6)-DevS~ol.
exactly as outlined in Fig. 2 except for the exclusion of GSSG/GSH.
Autophosphorylation of DevS2o~. Purified and re- folded DevS2o~ (15 #M) was incubated with 5 ;tCi of [~-32P]ATP (5000 Ci/mmol; BRIT, India) in 10 ~1 reac- tion buffer (50 mM Tris • HC1, pH 8.0, 50 mM KC1, 10 mM MgC12, 50 ttM ATP) at 25°C. The reaction was terminated at various time points as indicated with 5 #1 3× sample buffer (250 mM Tris • HC1, pH 6.8, 10% glycerol, 1% SDS, 280 mM fl-mercaptoethanol, 0.01% bromophenol blue). Since 98% of the maximum radioac- tivity was incorporated into DevS2o~ P in a 60-min pe- riod, the reactions were terminated at this point (D.K.S., unpublished results). The samples were ana- lyzed by 15% SDS-PAGE. The gel was rinsed in water and subjected to autoradiography.
Production of a rabbit antiserum against Dev- $2Ol. Purified and refolded DevS2oi was used to raise polyclonal antisera in two locally bred rabbits. Weekly injections of 100 #g purified protein emulsified with incomplete Freund's adjuvant in a volume of 500 ttl were given intramuscularly in the thigh muscle. After three injections, a test bleed was taken, serum was collected, and its t i ter was estimated by ELISA. Subse- quently boosters of 75 ttg protein were given fortnightly with bleeding being done 9 days after each injection. The serum was collected and stored in aliquots at -20°C.
205
Western blot analysis. A 200-/A culture equivalent of induced and uninduced E. coli BL21(DE3) with the pPROEx-HTc-devS2ol construct and I t~g of purified and refolded protein were resolved on 12.5% SDS-PAGE gel. Proteins were transferred onto a nitrocellulose membrane at 0.8 mA/cm 2 for 1 h. The membrane was blocked with 3% BSA for 2 h, followed by incubation with primary anti-DevS2oi serum collected after the second dose of antigen at a dilution of 1:1000 in PBS for 1 h. The blot was washed thoroughly with 0.05% Tween 20-PBS, followed by incubation with donkey anti-rabbit horseradish peroxidase conjugate (Amers- ham-Pharmacia, UK) at a dilution of 1:1000 in PBS for i h. The blot was washed and developed with diamino- benzidine. All the incubations and washes were per- formed at room temperature.
Immunoelectron microscopy. Immunoelectron mi- croscopy was essentially performed as described (2). A 25-ml culture of induced E. coli BL21(DE3) carrying pPROEx-HTc-devS2ol was probed with anti-DevS2ol an- tibody at 1:2000 dilution.
Cloning and overexpression of DevS2ol. The DevS HK protein (578 amino acids) is predicted to contain three t ransmembrane segments between residues 187 and 203, 274 and 295, and 325 and 342, respectively (TM1, TM2, and TM3, Fig. 1), that probably anchor it to the cell membrane. Overexpression of full-length sensor kinase proteins with membrane-spanning seg- ments has been unsuccessful due to toxicity to host cells (5, 13). Therefore, the putative cytoplasmic kinase core of DevS of M. tuberculosis was overexpressed as a pro- tein of 26.5 kDa inclusive of the 3-kDa His(6) tag.
Cells expressing recombinant protein were lysed by sonication or by use of B-Per Bacterial Protein Extrac- tion Reagent II (Pierce Biologicals, Rockford, IL). Anal- ysis of the soluble and insoluble protein fractions from induced E. coli cell pellets by SDS-PAGE indicated that the DevS2oi protein was expressed at high levels but was exclusively present in the insoluble fraction (Fig. 3A). The expression of DevS2ol protein occurred at a level of 50 to 60% of the total cellular protein. A variety of parameters reported to enhance the expression of soluble protein (14, 15) were tested to drive DevS2ol protein expression into the soluble form. These included varying the temperature of induction, growth media, host strains, and use of additives like 3% ethanol, 0.4 M NaC1 during induction, to name a few. However, these measures failed to produce the protein in a soluble form (not shown). DevS2ol was localized in large inclusion bodies by immunoelectron microscopy using rabbit anti- DevS2ol antibody (Fig. 3B). Hence, refolding of DevS2ol became a necessity prior to biochemical characteriza- tion of the putative HK.
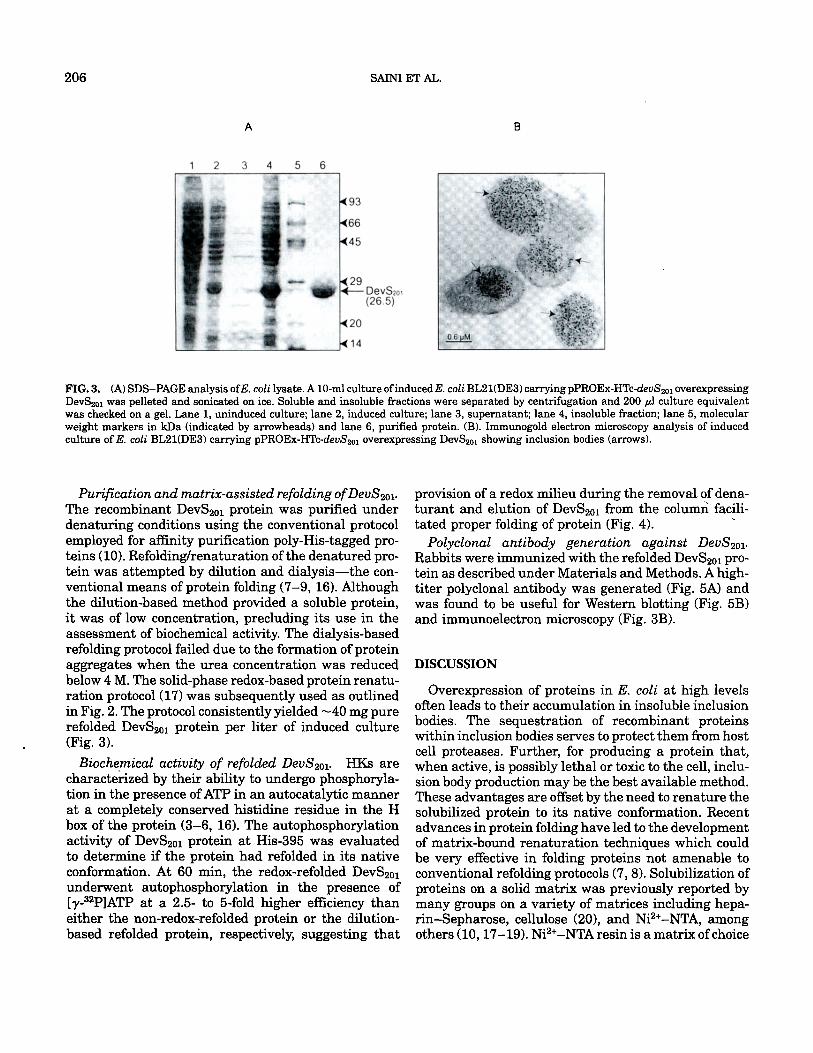
206 SAINI ET AL.
A
1 2 3 4 5 6
FIG. 3. (A)SDS-PAGEanalysisofE.coli~ysate.A~m~cu~tureofinducedE.c~liBL2~(DE3)carryingpPROEx-HTc-devS2~1overexpressing DevS201 was pelleted and sonicated on ice. Soluble and insoluble fractions were separated by centrifugation and 200/A culture equivalent was checked on a gel. Lane 1, uninduced culture; lane 2, induced culture; lane 3, supernatant; lane 4, insoluble fraction; lane 5, molecular weight markers in kDa (indicated by arrowheads) and lane 6, purified protein. (B). Immunogold electron microscopy analysis of induced culture ofE. coli BL21(DE3) carrying pPROEx-HTc-devS2ol overexpressing DevS2ol showing inclusion bodies (arrows).
Purification and matrix-assisted refolding of DevS2ol. The recombinant DevS2ol protein was purified under denaturing conditions using the conventional protocol employed for affinity purification poly-His-tagged pro- teins (10). Refolding/renaturation of the denatured pro- tein was attempted by dilution and dialysis--the con- ventional means of protein folding (7-9, 16). Although the dilution-based method provided a soluble protein, it was of low concentration, precluding its use in the assessment of biochemical activity. The dialysis-based refolding protocol failed due to the formation of protein aggregates when the urea concentration was reduced below 4 M. The solid-phase redox-based protein renatu- ration protocol (17) was subsequently used as outlined in Fig. 2. The protocol consistently yielded ~40 mg pure refolded DevS201 protein per liter of induced culture (Fig. 3).
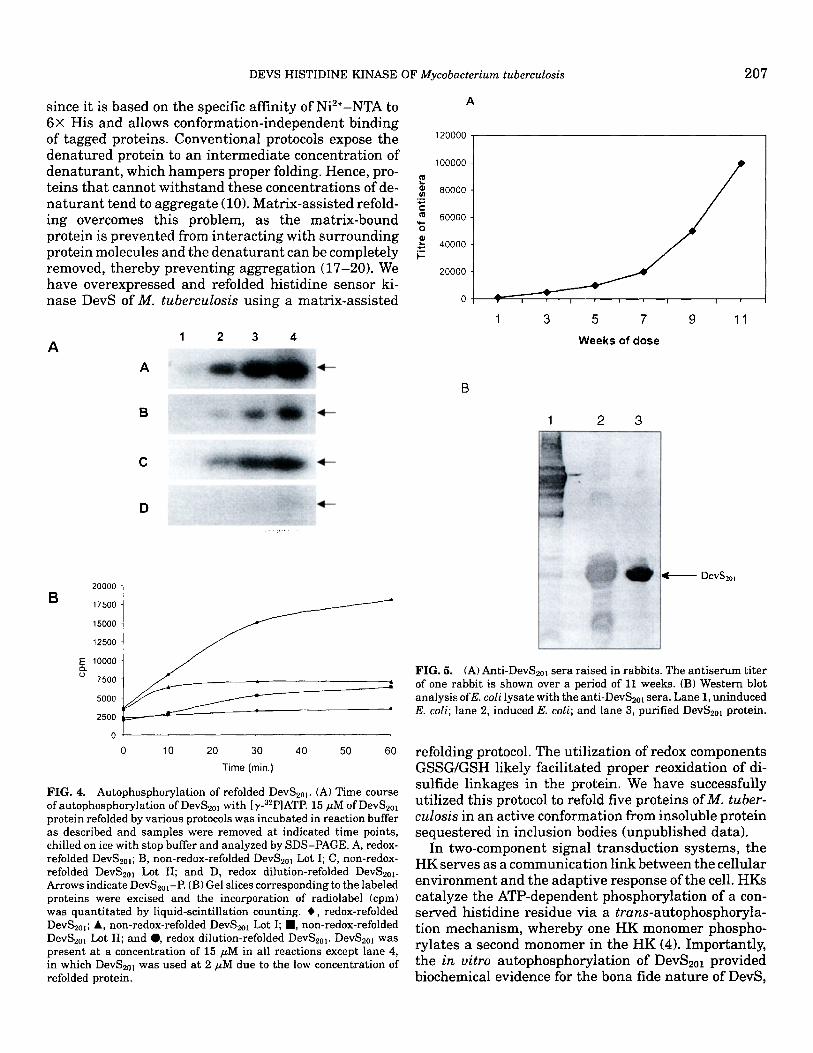
Biochemical activity of refolded DevS2ol. HKs are characterized by their ability to undergo phosphoryla- tion in the presence of ATP in an autocatalytic manner at a completely conserved histidine residue in the H box of the protein (3-6, 16). The autophosphorylation activity of DevS2ol protein at His-395 was evaluated to determine if the protein had refolded in its native conformation. At 60 rain, the redox-refolded DevS2ot underwent autophosphorylation in the presence of [7-s2p]ATP at a 2.5- to 5-fold higher efficiency than either the non-redox-refolded protein or the dilution- based refolded protein, respectively, suggesting that
provision of a redox milieu during the removal of dena- turant and elution of DevS2ol from the column facili- tated proper folding of protein (Fig. 4). "
Polyclonal antibody generation against DevS2ol. Rabbits were immunized with the refolded DevS2ol pro- tein as described under Materials and Methods. A high- titer polyclonal antibody was generated (Fig. 5A) and was found to be useful for Western blotting (Fig. 5B) and immunoelectron microscopy (Fig. 3B).
DISCUSSION
Overexpression of proteins in E. coli at high levels often leads to their accumulation in insoluble inclusion bodies. The sequestration of recombinant proteins within inclusion bodies serves to protect them from host cell proteases. Further, for producing a protein that, when active, is possibly lethal or toxic to the cell, inclu- sion body production may be the best available method. These advantages are offset by the need to renature the solubilized protein to its native conformation. Recent advances in protein folding have led to the development of matrix-bound renaturation techniques which could be very effective in folding proteins not amenable to conventional refolding protocols (7, 8). Solubilization of proteins on a solid matrix was previously reported by many groups on a variety of matrices including hepa- rin-Sepharose, cellulose (20), and Ni2+-NTA, among others (10, 17-19). Ni2+-NTA resin is a matrix of choice
DEVS HISTIDINE KINASE OF Mycobacterium tuberculosis 207
since it is based on the specific affinity of Ni2+-NTA to 6× His and allows conformat ion- independent binding of tagged proteins. Conventional protocols expose the 12oooo dena tured protein to an in termedia te concentration of
1 ooooo dena turan t , which hampers proper folding. Hence, pro- teins tha t cannot wi ths tand these concentrat ions of de- ~ 8oooo n a t u r a n t tend to aggregate (10). Matrix-assisted refold- c
m 60000 ing overcomes this problem, as the matr ix-bound protein is prevented from interact ing with sur rounding ~ 4oooo protein molecules and the dena tu ran t can be completely ~= removed, thereby prevent ing aggregation (17-20). We 20000 have overexpressed and refolded hist idine sensor ki- nase DevS of M. tuberculosis using a matrix-assisted o
A 1 2 3 4
A
B -~1--
A
1 3 5 7 9 11
W e e k s o f d o s e
1 2 3
C
D , : . -~ . ~
20000
B 17500
15000
12500
E 10000 Q. o
7500
5000
2500
~ f
o
o lO 20 30
Time(min.)
40 50 60
FIG. 4. Autophosphorylation of refolded DevS2ot. (A) Time course ofautophosphorylation of DevS2ol with [~-32p]ATP. 15 #M of DevS2ol protein refolded by various protocols was incubated in reaction buffer as described and samples were removed at indicated time points, chilled on ice with stop buffer and analyzed by SDS-PAGE. A, redox- refolded DevS2ol; B, non-redox-refolded DevS2ol Lot I; C, non-redox- refolded DevS2ol Lot II; and D, redox dilution-refolded DevS2ol. Arrows indicate DevS2orP. (B) Gel slices corresponding to the labeled proteins were excised and the incorporation of radiolabel (cpm) was quantitated by liquid-scintillation counting. O, redox-refolded DevS2ol; A, non-redox-refolded DevS2ol Lot I; I , non-redox-refolded DevS2ol Lot II; and 0, redox dilution-refolded DevS2ol. DevS2ol was present at a concentration of 15 tiM in all reactions except lane 4, in which DevS2ol was used at 2 ttM due to the low concentration of refolded protein.
• ; L
B
DcvS2o,
FIG. 5. (A) Anti-DevS2o~ sera raised in rabbits. The antiserum titer of one rabbit is shown over a period of 11 weeks. (B) Western blot analysis ofE. coli lysate with the anti-DevS2ot sera. Lane 1, uninduced E. coil; lane 2, induced E. coli; and lane 3, purified DevS2ol protein.
refolding protocol. The utilization of redox components GSSG/GSH likely facilitated proper reoxidation of di- sulfide l inkages in the protein. We have successfully utilized this protocol to refold five proteins of M. tuber- culosis in an active conformation from insoluble protein sequestered in inclusion bodies (unpublished data).
In two-component signal t ransduct ion systems, the HK serves as a communicat ion link between the cellular envi ronment and the adaptive response of the cell. HKs catalyze the ATP-dependent phosphorylat ion of a con- served hist idine residue via a trans-autophosphoryla- tion mechanism, whereby one HK monomer phospho- rylates a second monomer in the HK (4). Importantly, the in vitro autophosphorylat ion of DevS2ol provided biochemical evidence for the bona fide na ture of DevS,

208 SAINI ET AL.
which was predicted to be a sensor histidine kinase on the basis of homology with bacterial HKs (3, 4). The overexpression and purification of sufficient quantities of functionally active DevS2ol and generation of anti- body therefrom have paved the way for further struc- tural, biochemical, and physiological characterization of the DevR-DevS two-component system of M. tuber- culosis.
ACKNOWLEDGMENTS
D.K.S. is thankful to the University Grants Commission for a Senior Research Fellowship and Neha Pant to the Department of Biotechnol- ogy for a M.Sc. fellowship. The authors acknowledge S. Chakravorty for pPROEx-HTc-devS2ol. Financial assistance provided to J.S.T. by the Council for Scientific and Industrial Research, Government of India, is acknowledged.
REFERENCES
1. Kinger, A. K., and Tyagi, J. S. (1993) Identification and cloning of genes differentially expressed in the virulent strain of Myco- bacterium tuberculosis. Gene 131, 113-117.
2. Dasgupta, N., Kapur, V., Singh, K. K., Das, T. K., Sachdeva, S., Jyothisri, K., and Tyagi, J. S. (2000) Characterization of a two- component system, devR-devS, of Myeobacterium tuberculosis. Tuberc. Lung Dis. 80, 141-159.
3. Parkinson, J. S., and Kofoid, E. C. (1992) Communication mod- ules in bacterial signaling proteins.Annu. Rev. Genet. 26, 71-112.
4. Stock, J. B., Surette, M. G., Levit, M., and Park, P. (1995) Two- component signal transduction systems: Structure-function re- lationships and mechanisms of catalysis, in "Two-Component Signal Transduction" (Hoch, J. A., and Silhavy, T. J., Eds.), pp. 25-51, ASM Press, Washington, DC.
5. Haydel, S. E., Dunlap, N. E., and Benjamin, W. H., Jr. (1999) In vitro phosphorylation between the M. tuberculosis TrcRfIYcS proteins. Microb. Pathog. 26, 195-206.
6. Mukai, K., Kawata, M., and Tanaka, T. (1990) Isolation and phosphorylation of the Bacillus subtilis degS and degU gene products. J. Biol. Chem. 265, 20000-20006.
7. De Bernardez Clark, E. (1998) Refolding of recombinant proteins. Curr. Opin. Biotechnol. 9, 157-163.
8. Rudolph, R., B6hm, G., Lilie, H., and Jaenicke, R. (1997) Folding proteins, in "Protein Function: A Practical Approach" (Crieghton, T. E., Ed.), pp. 57-99, 2nd ed., IRL Press, Oxford.
9. Lilie, H., Schwarz, E., and Rudolph, R. (1998) Advances in refold- ing proteins produced in E. coll. Curr. Opin. Biotechnol. 9, 497-501.
10. Holzinger, A., Phillips, K. S., and Weaver, T. E. (1996) Single- step purification/solubilization of recombinant proteins: Applica- tion to surfactant protein B. Biotechniques 20, 804-807.
11. Cole, S. T., Brosch, R., Parkhill, J., Garnier, T., Churcher, C., Harris, D., Gordon, S. V., Eiglmeier, K., Gas, S., Barry, C. E., III, Tekaia, F., Badcock, K., Bashyam, D., Brown, D., Chillingworth, T., Connor, R., Davies, R., Devlin, K., Feltwell, T., Gentles, S., Hamlin, N., Holroyd, S., Hornsby, T., Jagels, K., Krough, A., McLean, J., Moule, S., Murphy, L., Oliver, K., Osborne, J., Quail, M. A., Rajandream, M. A., Rogers, J., Rutter, S., Seeger, K., Skelton, J., Squares, R., Squares, S., Sulston, J. E., Taylor, K., Whirehead, S., and Barrell, B. G. (1998) Deciphering the biology of Mycobacterium tuberculosis from the complete genome se- quence. Nature 393, 537-544.
12. Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the prin- ciple of protein-dye binding. Anal. Biochem. 72, 248-254.
13. Gilles-Gonzalez, M. A., Ditta, G. S., and Helsinki, D. R. (1994) A haemoprotein with kinase activity encoded by the oxygen sen- sor of Rhizobium meliloti. Nature 350, 170-172.
14. Baneyx, F. (1999) Recombinant protein production in E. coli. Curr. Opin. Biotechnol. 10, 411-421.
15. Georgiou, G., and Valax, P. (1996) Expression of correctly folded proteins in E. coli. Curl:. Opin. Biotechnol. 7, 190-197.
16. Himpens, S., Locht, C., and Supply, P. (2000) Molecular character- ization of the mycobacterial SenX3-RegX3 two-component sys- tem: Evidence for autoregulation. Microbiology 146, 3091-3098.
17. Zouhar, J., Nanak, E., and Brzobohaty, B. (1999) Expression, single-step purification and matrix-assisted refolding of a maize cytokinin glucoside-specific fl-glucosidase. Protein Expression Purif. 17, 153-162.
18. Longstaff, M., Powell, K. S., Gatehouse, J. A., Raemaekers, J. A., Newell, C. A., and Hamilton, W. D. O. (1998) Production and purification of active snowdrop lectin in E. coll. Eur. J. Biochem. 252, 59-65.
19. Zahn, R., yon Schroetter, C., and Wtithrich, K. (19971 Human prion proteins expressed in E. coli and purified by high affinity column refolding. FEBS Lett. 417, 400-404.
20. Berdichevsky, Y., Lamed, R., Frenkel, D., Gophna, U., Bayer, E. A., Yaron, S., Shoham, Y., and Benhar, I. (1999) Matrix-assisted refolding of single-chain Fv-cellulose binding domain fusion pro- teins. Protein Expression Purif 17, 249-259.












![% :.I COPY AVAILA']RV lj£gIE ~l - DTIC](https://static.fdokumen.com/doc/165x107/631e931825add517740b1bd2/-i-copy-availarv-ljgie-l-dtic.jpg)







