
Cloning of murine transformed cell lines in suspension culture with efficiencies near 100
8
Printed in Sweden Copyright @ 1978 by Academic Press. Inc. All rights of reproduction in any form reserved 0014-4827/78/l Il2-0309$02.00/O Experimental Cell Research I1 1 (1978) 309-3 16 CLONING OF MURINE TRANSFORMED CELL LINES IN SUSPENSION CULTURE WITH EFFICIENCIES NEAR 100% WALDEMAR LERNHARDT,’ JAN ANDERSSONP ANTONIO COUTINHO’ and FRITZ MELCHERS’ ‘Base1Institute for Immunology, CH4055 Base15, Switzerland, and 2Biomedicum, University of Uppsala, S-75123 Uppsala, Sweden SUMMARY Addition of 3 x lo6 thymus cells from either syngeneic, allogeneic or xenogeneic animals increases the cloning efftciencies of murine thymomas (EL-4, WC-2), B-lymphomas (McPC 1748, 38C-13), Abelson-virus transformed cell lines (F and K), mastocytomas (PSlS), myelomas (AbPC 22, X63-AC& 5563, MOPC 104 E, RPC 5, W 3469) and hybrids of myelomas and normal B-lympho- cytes (Sp-1), all adapted to tissue culture, to near 100 %. Thymus cells also increase the efficiencies of growth initiation in primary in vitro cultures of myeloma tumor cells (S117) transplanted in vivo, and of cells fused between the azaguanine-resistant X63-AG8 myeloma cell line and nor- mal, LPS-stimulated B-lymphocyte blasts. Normal and malignant T (thymus-derived)- and B (bone marrow-derived, bursa- equivalent)-lymphocytes can be cloned in vitro in semi-solid culture systems [l-8] or, by limiting dilutions, in suspension cul- tures. The cloning efficiency, however, is never near lOO%, but varies between 0.1 and 2 % for different established lymphoma and myeloma cell lines. Normal T- and B- lymphocytes, stimulated to growth by mito- gens [9] show similar problems in the effi- ciencies of initiation to clonal growth. They cease to grow below cell concentrations of 3~ 104/ml, either in semisolid or suspen- sion culture [8, 10, 111. Recently it has be- come possible to increase the efftciency of clonal growth of mitogen-stimulated normal T- and B-lymphocytes by factors near lo4 by adding to the cultures mitogen-unreac- tive “filler” cells from a lymphoid organ, from the thymus in the case of mitogen- stimulated B-lymphocytes [8, 1l-141. It appeared that every mitogen-reactive B-lymphocyte now could initiate clonal growth [15]. From these results it could be hoped that addition of “filler” thymus cells to malignant T- and B-lymphocytes in sus- pension culture would increase their cloning efficiencies. Addition of “tiller” thymus cells, how- ever, makes it mandatory that single clones of growing lymphocytes can be detected amongst the large excess of “filler” cells. For myeloma cells secreting immunoglobu- lin (Ig)-molecules a plaque assay is available which uses protein A-coated sheep red cells (SRC) and anti Ig-antibodies to detect every secreting cell [16]. Within a population of Exp Cell Res I I I (1978)
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cloning of murine transformed cell lines in suspension culture with efficiencies near 100

Printed in Sweden Copyright @ 1978 by Academic Press. Inc.
All rights of reproduction in any form reserved 0014-4827/78/l Il2-0309$02.00/O
Experimental Cell Research I1 1 (1978) 309-3 16
CLONING OF MURINE TRANSFORMED CELL LINES
IN SUSPENSION CULTURE WITH EFFICIENCIES
NEAR 100%
WALDEMAR LERNHARDT,’ JAN ANDERSSONP ANTONIO COUTINHO’ and FRITZ MELCHERS’
‘Base1 Institute for Immunology, CH4055 Base1 5, Switzerland, and 2Biomedicum, University of Uppsala, S-751 23 Uppsala, Sweden
SUMMARY Addition of 3 x lo6 thymus cells from either syngeneic, allogeneic or xenogeneic animals increases the cloning efftciencies of murine thymomas (EL-4, WC-2), B-lymphomas (McPC 1748, 38C-13), Abelson-virus transformed cell lines (F and K), mastocytomas (PSlS), myelomas (AbPC 22, X63-AC& 5563, MOPC 104 E, RPC 5, W 3469) and hybrids of myelomas and normal B-lympho- cytes (Sp-1), all adapted to tissue culture, to near 100 %. Thymus cells also increase the efficiencies of growth initiation in primary in vitro cultures of myeloma tumor cells (S117) transplanted in vivo, and of cells fused between the azaguanine-resistant X63-AG8 myeloma cell line and nor- mal, LPS-stimulated B-lymphocyte blasts.
Normal and malignant T (thymus-derived)- and B (bone marrow-derived, bursa- equivalent)-lymphocytes can be cloned in vitro in semi-solid culture systems [l-8] or, by limiting dilutions, in suspension cul- tures. The cloning efficiency, however, is never near lOO%, but varies between 0.1 and 2 % for different established lymphoma and myeloma cell lines. Normal T- and B- lymphocytes, stimulated to growth by mito- gens [9] show similar problems in the effi- ciencies of initiation to clonal growth. They cease to grow below cell concentrations of 3~ 104/ml, either in semisolid or suspen- sion culture [8, 10, 111. Recently it has be- come possible to increase the efftciency of clonal growth of mitogen-stimulated normal T- and B-lymphocytes by factors near lo4 by adding to the cultures mitogen-unreac-
tive “filler” cells from a lymphoid organ, from the thymus in the case of mitogen- stimulated B-lymphocytes [8, 1 l-141. It appeared that every mitogen-reactive B-lymphocyte now could initiate clonal growth [15]. From these results it could be hoped that addition of “filler” thymus cells to malignant T- and B-lymphocytes in sus- pension culture would increase their cloning efficiencies.
Addition of “tiller” thymus cells, how- ever, makes it mandatory that single clones of growing lymphocytes can be detected amongst the large excess of “filler” cells. For myeloma cells secreting immunoglobu- lin (Ig)-molecules a plaque assay is available which uses protein A-coated sheep red cells (SRC) and anti Ig-antibodies to detect every secreting cell [16]. Within a population of
Exp Cell Res I I I (1978)

310 Lernhardt et al.
Ig-secreting myeloma cells, between 1.5 and 75 % of all cells will produce a plaque in this assay. Furthermore, lymphoma and myeloma cells are easily detectable under the microscope in a suspension culture with thymus cells because of their large size. With these assays for clonal growth we de- termined in this paper cloning efficiencies of a series of T- and B-lymphomas and myelomas in suspension cultures in the presence of “filler” cells either from syn- geneic, allogeneic or xenogeneic thymus or from xenogeneic peripheral lymph. The ad- dition of “filler” cells increases the cloning effkiencies of all tested cell lines to near 100 %. Similarly it increases the frequencies of clones growing in selective medium after fusion of azaguanine-resistant myeloma cells with normal splenic lymphocytes [17, 181 and supports the establishment of new cell lines from in vivo murine tumours not yet adapted to tissue culture.
MATERIALS AND METHODS
Animals
C57Bl/6J/Fiil mice and A/J/Fiii mice, 4 weeks of age, BALB/c/Ftil mice, 6-8 weeks of age, and Lewis strain rats, 4-8 weeks of age, were obtained from the Institut fur biologisch-medizinische Forschung A.G.. Fiilhnsdorf, Switzerland.
Cell lines and myelomas
From the murine AKR thymoma F22, isolated by Dr P. Krammer [19] and adapted to tissue culture by Dr M. Schreier, in our Institute, a number of clones were derived. The cloned cell line used here. caged WC-2. secretes protein with binding activity for complexed rabbit Ig (A. Coutinho et al., in prenaration). The T- cell leukemia EL-4 of C57Bi/6 origin and the DBA/Z mastocytoma P815 were given to us by Dr G. Kiihler of our Institute. The chemically induced B-lymphoma 38C-13 (producing IgM) originally isolated and de- scribed by Haran-Ghera & Peled [20], was adapted to tissue culture by Dr Y. Bergman and given to us by Dr Haimovich (both from the Weizmann Institute. Rehovot). The myeloma AbPC22 (secreting IgM) and RPCS (secreting IgG) maintained as transplantable tllmnurs in BALB/c mice, were kindly given to us by
Dr M. Potter, NCI, NIH, Bethesda, Md, and adapted to tissue culture in our laboratory. The BALB/c B lymphomas McPC 1748 and W3469 (both producing IaM) were originally obtained from Dr J. Buxbaum, New York University, Department of Medicine, New York, who kindly donated the lines.
The S117 myeloma, secreting IgA, was given to us bv Dr M. Potter and was kept as a transplantable tumor in BALB/c mice.
The tissue culture line of myeloma 5563, secreting IgG2 (raised originally in C3H-mice), was kindly given to us by Dr B. A. Askonas, National Institute for Medi- cal Research, Mill Hill, London. The azaguanine- resistant cell line X63-AGS of myeloma MOPC21, secreting IgGl, [ 171 (raised originally in BALB/c mice bv Dr M. Potter). was kindly given to us by Dr C. Mil- stein, MRC Postgraduate School of Molecular Biology, Cambridge, UK. The hybrid cell line Sp-1 producing IgM specific for SRC was obtained after fusion be- tween X63-A68 and BALB/c spleen cells [21] and was kindly given to us by Dr G. Kiihler of our Institute.
Abelson murine leukemia transformed tumors [22] were induced in vivo by intraperitoneal injections of virus preparations (kindly provided by Dr Philip, Microbiological Associates, Bethesda, Md) into neo- natal BALB/c mice. Tumors were raised by Dr B. Wei- mann and adapted to continuous suspension cultures by Dr M. Schreier and Dr B. Weimann, both of our Institute. The cloned cell lines F and K secrete Abel- son murine leukemia virus into the culture medium.
Tissue culture
RPM1 1640 medium (Microbiological Associates, Bethesda, Md), supplemented with glutamine (2 mM); penicillin and streptomycin (5 OOfl III/ml each), Hepes buffer (10 mM) pH 7.3,2-mercaptoethanol(5 x 10V5 M), was used throughout all experiments. Cell lines X63- AG8, 5563, MOPC 104E, 38C-13, McPC 1748 were grown in the presence of 10% horse serum (batch L267301, Gibco Bio-Cult, Irvine), all other lines and the adapted lines of S117-myeloma and of the fused cells were grown in 20% fetal calf serum (batch K 255701 D, Gibco, Bio-Cult). Cells were grown in Linbro Dispo trays (model FB16-27, TC; Linbro Scientific, New Haven, Conn.) holding 1 ml medium. Mouse and rat thymus cells were prepared as de- scribed previously [15]. Rabbit thymus cells (from 3- month-old New Zealand strain rabbits) were prepared accordingly. Sheep peripheral lymphocytes, obtained by lymph drainage, were kindly provided by Dr R. Cahill of our Institute.
Mitogenic activation of B-cells and fusion with X63-AC8 myeloma cells
BALB/c spleen (3x 10S/ml) were activated with LPS (50 pg/ml, batch EDTEN 18735, a gift from Drs C. Galanos and 0. Ltideritz, Max-Planck-Institut tir Im- munbiologie, Freiburg), as described previously [23, 101. For hybridization experiments [18] 108 LPS- activated spleen cells were taken at day 5 after activa-
Exp Cell RPS I I I (1978)

Cloning of lymphomas and myelomas 3 11
Table 1. Growth of X63-AG8 myeloma cells in the presence and absence of syngeneic mouse thymus cells
0 a2 a4 0.6 06 1.0 1.2 1.4 1.6 100
37F-
‘hi,
1 I” 1.36
Fig. 1. Abscissa: no. of X63-AG8 cells/culture; ordi- nate: % non-responding cultures.
Titration of growing X63-AG8 myeloma cells in sus- pension cultures in the CO, presence; O-O, or absence of 3~10~ syngeneic thymus cells/ml. Ab- scissa indicates the number of input myeloma cells per culture. Each point in the figure is based on 40 assays. The assays for IgGl-secreting, plaque-forming myeloma cells were done at day 5 of culture.
tion and spun together with 10’ X63-AG8 azaguanine- resistant myeloma cells. One ml of 45 % polyethylene- glycol (mol. wt 4000, Lot No. 2871-125, Polysciences Inc., Warrington, Pa), 15 % dimethylsulphoxide (Merck, Darmstadt) in medium at pH 7.8 was added under slow stirring within 1 min, then 20 ml of medium were added within the next 10 min. All operations were carried out at 37°C. The fused cells were washed twice, then plated in culture medium with or without thymus “filler” cells. Twenty-four hours later the cul- ture medium was changed for hypoxanthine, amino- pterin, thymidine, glycine containing medium [24].
Plaque assay for Ig secreting cells A modified hemolytic plaque assay using protein A- coated SRC and mouse Ig specific antisera as develop- ing antibodies in the presence of complement was employed to detect and enumerate Ig-secreting cells [ 161. Antisera were raised in rabbits by repeated injec- tions of myeloma proteins. Anti-mouse IgM antibodies were raised against purified MOPC 104E IgM (A, p) anti-mouse IgGl antibodies against purified MOPC 21 IgGl (K; n), anti-mouse IgG2 antibodies against puri- fied Adj-PC-5 IgG2 (K; -y.& and anti-mouse IgA anti- bodies against purified MOPC 315 IgA (X, a) myeloma protein. Protein A used in the coating of sheep red cells was obtained from Dr H. Wigzell, Biomedicum, University of Uppsala, UppsaIa. Complement, appro- priately diluted with BSS, was from Bio-Cult.
IgGl-PFC/culture”
Number of syngeneic BALB/c thymus X63-AG8 cells/mP myeloma cells in culturec 0 3x105 3x106 3x10’
600 2000 2000 2000 2000 200 1800 2000 2000 60 650d 1800
El 2000
20 loo” 9006 2000 2000 6 5 80” 2 000 2 000
(i.6) 5 5 5 5 850d 15oe 780d 180”
a X63-AG8 myeloma cells secrete IgG 1 and can, there- fore, be detected in a plaque assay with protein A- SRC and anti IgGl-antibodies [ 161. Assay was done at day 5. The values given are the average of 10 deter- minations. b No nlaaues were detected with either 3 x 105. 3 X lo6 or 3 x iOr thymus cells alone in control cultures: c 1 ml cultures. d 80 % or more of all cultures positive. e Between 20 and 50% of all cultures positive.
RESULTS
Cloning efJiciency of a myeloma cell line, X63-AG8, in suspension cultures in the presence and absence of syngeneic thymus cells
The effect of syngeneic BALB/c thymus cells on the growth of IgGl-secreting X63- AG8 myeloma cells was tested in suspen- sion cultures containing either no thymus cells, or 3x 105, 3x IO6 or 3x 10’ thymus cells/ml. Myeloma cells from an exponenti- ally growing culture were diluted from 2 000 cells to an average of 0.6 cells/culture in medium containing these graded numbers of thymus cells or no thymus cells. Growth was scored by determining the number of IgGl-secreting, plaque forming X63-AG8 cells in the protein A-SRC plaque assay with anti IgGl antibodies as developing antibodies.
Exp CellRes III (1978)
3 12 Lernhardt et al.
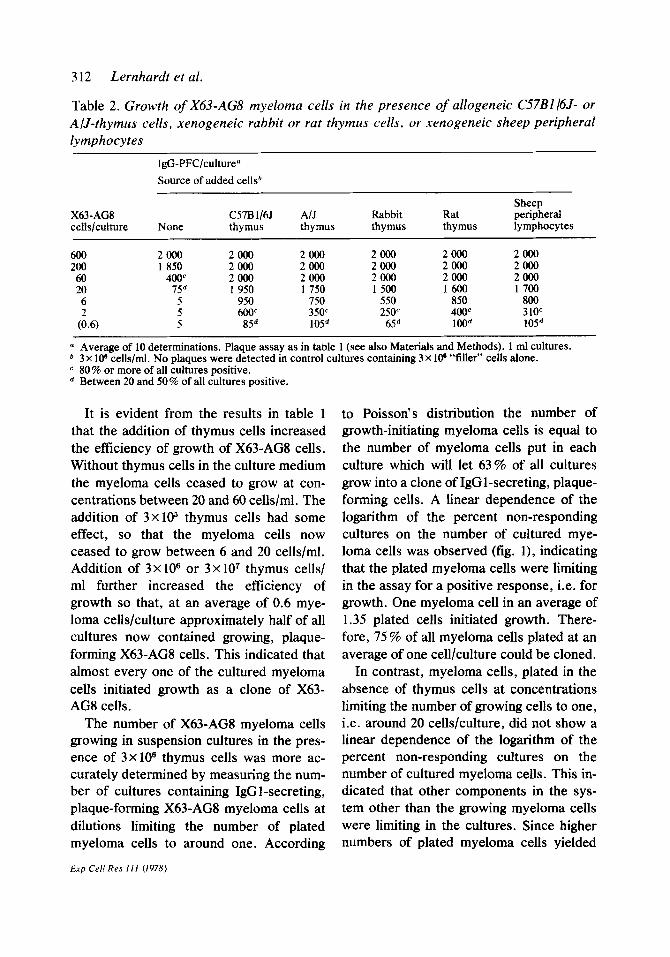
Table 2. Growth of X63-AG% myeloma cells in the presence of allogeneic C57B1 I6J- or A/J-thymus cells, xenogeneic rabbit or rat thymus cells, or xenogeneic sheep peripheral lymphocytes
IgG-PFC/culture” Source of added cells”
Sheep X63-AG8 c57B l/65 A/J Rabbit Rat peripheral cells/culture None thymus thymus thymus thymus lymphocytes
600 2000 2000 2000 2000 2000 2000 200 1850 2000 2000 2000 2000 2000 60 400c 2000 2000 2000 2000 2000 20 156 1950 1 750 1500 1600 1700 6 5 950 750 550 850 800
(i.6) 5 600’ 35oc 250" 400' 310' 5 85d 105d 6Y' 1OOd 10Sd
o Average of 10 determinations. Plaque assay as in table 1 (see also Materials and Methods). 1 ml cultures. b 3x 10” cells/ml. No nlaaues were detected in control cultures containing 3 x 106 “tiller” cells alone. c 80 % or more of all cultures positive. d Between 20 and 50% of all cultures positive.
It is evident from the results in table 1 that the addition of thymus cells increased the efficiency of growth of X63-AG8 cells. Without thymus cells in the culture medium the myeloma cells ceased to grow at con- centrations between 20 and 60 cellslml. The addition of 3x IO5 thymus cells had some effect, so that the myeloma cells now ceased to grow between 6 and 20 cells/ml. Addition of 3x IO6 or 3 x 10’ thymus cells/ ml further increased the efficiency of growth so that, at an average of 0.6 mye- loma cells/culture approximately half of all cultures now contained growing, plaque- forming X63-AG8 cells. This indicated that almost every one of the cultured myeloma cells initiated growth as a clone of X63- AGS cells.
The number of X63-AG8 myeloma cells growing in suspension cultures in the pres- ence of 3X lo6 thymus cells was more ac- curately determined by measuring the num- ber of cultures containing IgGI-secreting, plaque-forming X63-AG8 myeloma cells at dilutions limiting the number of plated myeloma cells to around one. According
Exp Cell Res Ill (1978)
to Poisson’s distribution the number of growth-initiating myeloma cells is equal to the number of myeloma cells put in each culture which will let 63 % of all cultures grow into a clone of IgGl-secreting, plaque- forming cells. A linear dependence of the logarithm of the percent non-responding cultures on the number of cultured mye- loma cells was observed (fig. l), indicating that the plated myeloma cells were limiting in the assay for a positive response, i.e. for growth. One myeloma cell in an average of 1.35 plated cells initiated growth. There- fore, 75 % of all myeloma cells plated at an average of one cell/culture could be cloned.
In contrast, myeloma cells, plated in the absence of thymus cells at concentrations limiting the number of growing cells to one, i.e. around 20 cells/culture, did not show a linear dependence of the logarithm of the percent non-responding cultures on the number of cultured myeloma cells. This in- dicated that other components in the sys- tem other than the growing myeloma cells were limiting in the cultures. Since higher numbers of plated myeloma cells yielded

Cloning of lymphomas and myelomas 313
Fig. 2. A hybrid cell clone after 8 days of culture in selective medium in the presence of rat thymus filler cells. At this time of culture the thymus cells appear
disproportionately higher numbers of re- sponding, i.e. growing cultures, it may be that the myeloma cells in cultures helped each other to grow.
Other sources of lymphoid cells as “fillet’ cells
Growth-enhancing effects could also be ob- served with lymphoid cells of allogeneic or xenogeneic origin. Thymus cells from allo- geneic mice (CS7B1/6J, A/J) and of xeno- geneic origin, such as rabbit or rat thymus cells and sheep peripheral lymphocytes, when added to the suspension cultures at 3 X lo6 cells/ml all increased the efficiency of cloning of X63-AG8 myeloma cells to near 100% (table 2). It shows that the growth-enhancing effect is not species- specific and can be supplied by thymus cells as well as by peripheral lymphocytes.
as dCbris surrounding the growing hybrid cells. The hybrid was formed after fusion between the X63-AG8 myeloma cell line and normal BALB/c spleen cells.
Sheep red cells, in concentrations be- tween 3 x 105 and 3x 10’ cells/ml, on the other hand, did not enhance growth of X63-AG8 myeloma cells. This indicates that the addition of thymus cells or pe- ripheral lymphocytes is not due to a mere crowding eff:ct in which the myeloma cells have to be in contact with any other cell surface.
Cloning of different T- and B-lymphomas and of other myeloma lines in suspension cultures in the presence of rat thymus cells
Cells of established culture lines of two T-lymphomas, three B-lymphomas, one mastocytoma, two Abelson virus-trans- formed cell lines, four myelomas and one hybrid myeloma cell line were diluted in suspension cultures with or without rat
Exp Cc// Res 111 (1978)

314 Lernhardt et al.
Table 3. Growth of thymoma, B-lymphoma, Abelson virus-transformed, mastocytoma and myeloma tissue culture lines in the ab- sence and presence of thymus ‘filler” cells
Name and type of line
No. of cells/culture where approx.” 37 % of all cultures did not yield positive cultures containing grow- ing tumour cells
Absent Present*
EL-4 thymoma WC-2 thymoma W 3469 - McPC 1748 B-lymphoma 38C-13 P 815 mastocytoma Abelson virus trans- formed cell line F-tine K-line
AbPC 22 myeloma MOPC 104E myeloma 5563 myeloma Sp-1 hybridC myeloma RPCS myeloma
2x102 1 6x 10’ 1 n.d. 1 6x 10’ 1 nd. 1 2x 10’ 1
n.d. 1 n.d. 1 n.d. 1 2x 103 1 n.d. 1 n.d. 1 n.d. 1
u According to Poisson’s distribution, an average of one percursor of a clone of growing cells is present in each culture when 37% of all cultures do not yield growing cells. In our experiments between 20% and 60% of all cultures did not yield growing cells; there- fore, we have only determined approximate frequen- cies. b Rat thymus tiller cells added at a rate of 3X lo6 cells/ml. n.d., not done. c Fused cell line, secreting SRC-specific IgM [17].
thymus cells as “fillers” to determine clon- ing efficiencies. T- and B-lymphoma growth was screened under the microscope (fig. 2), while myeloma growth was recorded in ad- dition by the protein A-SRC plaque assay with the appropriate class-specific anti- bodies as developing antibodies. The dif- ferent cell lines.showed different cloning ef- ficiencies in cultures without “filler” thy- mus cells, ranging from 1 in 2X lo3 to 1 in 20 cells (table 3). Addition of 3 x lo6 rat thymus cells per millilitre to the medium in- creased the frequency of growing cells for all cell lines. The data in table 3 indicate
that cloning efficiencies near 100% were obtained for all cell lines. Establishment and cloning of myelomas grown in vivo and offused cell lines The addition of “filler” cells appeared to increase the frequency of growing cells for every established murine lymphoid cell line and for a mastocytoma cell line to 1. We therefore tried to use this growth-support- ing effect of thymus cells to establish new cell lines in tissue culture. Cells, taken either from a myeloma transplanted in vivo, or from fusions between normal, LPS (bac- terial lipopolysaccharides)-stimulated B- lymphocytes [23] and an azaguanine-resist- ant myeloma [17] (Melchers, F, in prepara- tion), were plated at different cell concen- trations in the presence or absence of rat thymus cells. Growth was monitored un- der the microscope and by the develop- ment of Ig-secreting plaque-forming cells. (S117 myeloma cells secrete IgA; X63-AG8 cells secrete IgGl ; 90 % of all LPS-blasts at day 5 secrete IgM [ 111. Fused cells were, therefore, monitored for their capacity to secrete IgM.) Results in table 4 show that the addition of thymus cells to the primary cultures increased the frequencies of grow- ing clones of either S117-myeloma cells, or of fused cells between normal, LPS-stimu- lated cells and an azaguanine-resistant mye- loma cell line growing in tissue culture, for at least as long as 2 months after initiation of the primary cultures.
DISCUSSION Addition of thymus cells as growth-sup- porting “filler” or “feeder” cells increases the cloning efficiencies of a series of thymomas, B-lymphomas, Abelson virus- transformed cell lines, as well as myeloma cell lines adapted to tissue culture near to 100%. It also increases the efficiencies of growth initiation of primary cultures from
Exp Cd Res III (1978)

Table 4. Establishment of primary in vitro cell lines from in vivo growing SlI7-mye- loma cells and from cells fused between normal, L.PS-stimulated B-cells and X63- AG8 myeloma cells”
No. of positive, growing cultures* per total number of cultures plated
No. of Sl17 mveloma cells Fused cells’ plated cells/ culture Absent Presentd Absent Presentd
3x106 20120 20120 7110 loll0 3x105 l/20 20120 o/10 loll0 3x104 O/20 20120 o/10 2110 3x103 0120 20120 0110 0110 3x102 O/20 16120 - - 3x 10’ O/20 3120 - - 3x100 O/20 O/20 - -
a Splenic B-cells of normal, BALBlc-mice were stimu- lated for 5 days with LPS in culture under conditions described elsewhere [lo]. These LPS-blasts (10 * cells1 ml) were then fused with azaguanine-resistant X63- AGS-myeloma cells (Kiihler & Milstein [ 17
3 (10’ cells/
ml) in 1 ml 45 % polyethyleneglycol [18 . 15% di- methylsulphoxide in RPM1 1640 medium (Melchers, F, in preparation). * Scoredunder the microscope (at day 14 of culture) and by plaque assay (for S 117 myeloma as IgA-secret- ing cells (at day 17 of culture)). c The number of plated cells/culture corresponds to the original number of normal, LPS-stimulated B- blast cells used in the fusion reaction. d Rat thymus cells added at 3 x 106 cells/ml.
Cloning of lymphomas and myelomas 3 15
appears, again, of prime practical impor- tance. Cell lines obtained after fusion be- tween the azaguanine-resistant myeloma and normal, LPS-stimulated B-cells are ex- pected to contain a spectrum of B-cell lines, each of them secreting one type of Ig with one antigen-binding specificity within the total repertoire of specificities con- tained within the population of B-cells stim- ulated polyclonally by LPS [23]. Addition of thymus filler cells to recently formed hybrids will allow a larger fraction of fused cells to grow out into clones of Ig-pro- ducing hybrid cell lines. Subsequent cloning of hybrid cell lines producing a certain wanted antibody specificity is then again eased by the aid of thymus tiller cells. This appears of importance whenever B-cells with a wanted antibody specificity are rare within the total population of fused B-cells, i.e. as in the case of antibodies against histocompatibility antigens [ 181.
myelomas growing in vivo, and of cells fused between a myeloma cell line and nor- mal splenic B-cells. These findings are of considerable practical use. Cloning effici- encies of near 100% for established cell lines will enable us to determine, in such cells, mutation rates for selected genetic markers, as for instance for the resistance to azaguanine [25, 261 (W. Lernhardt & F. Melchers, in preparation). It will ease the selection from these established lines of new lines with a given genetic marker, for which selective media or a selective assay exists.
We have developed an assay for killer T-lymphocytes in which the target cells are plaque-forming myeloma cells [8]. The as- say makes use of our finding presented in this paper, that every myeloma cell initiates growth in cultures with thymus cells. Therefore, when, on the average, two to five myeloma ‘cells are plated, every cul- ture, after 5 days of growth, contains be- tween 20&500 PFC. If, however, killer T- lymphocytes, specific for the myeloma target and activated by either concanavalin A or in the mixed lymphocyte reaction, are added to these 2-5 myeloma target cells at the beginning of the culture, no PFC ap- pear. The assay appears to detect single killer T-lymphocytes and is, therefore, or- ders of magnitude more sensitive than cur- rently available assays for killer cell ac- tivities.
The establishment of new in vitro lines We know nothing of the molecular nature from in vivo tumors or from fused cells of the “filler” or “feeder” effect of thymus
Exp Cell Xes 111 (1978)

3 16 Lernhardt et al.
cells. We have not yet investigated whether other cell sources could substitute for thy- mus cells. We know that thymus cells from other species will support growth of mouse cells. It is not clear yet whether thymus cells from mouse or rat will support the growth of malignant or normal cells from other species.
Normal [ 11, 12, 151 and transformed murine B-cells increase their cloning efft- ciencies in the presence of thymus cells. Conversely, normal T-cells-stimulated by mitogen such as concanavalin A or in the mixed lymphocyte reaction-increase their growth efficiencies in the presence of “fill- er” cells devoid of T-cells, such as spleen cells from genetically athymic, nude (nu/ nu) mice [8, 13, 141. The hope remains that molecules can be extracted and purified from lymphocytes which will substitute for whole “filler” cells in cultures of normal or transformed T- or B-lymphocytes. Such “filler” cell-free cultures would make it pos- sible to assay T-cell responses in a quanti- tative fashion on a clonal level.
The study of genetically or environmen- tally determined factors influencing growth and differentiation of eukaryotic cells re- quires the quantitative analysis of the num- ber of cells involved in a given response and their fate after the response. This can best be done in vitro where every cell can be separated from all others-at least in principle. However, it requires that every cell has an equal, maximal chance of survi- val, response and growth in culture. At least for lymphocytes, transformed (this paper) or normal mitogen-reactive B-cells or not yet reactive precursors of B-cells [27], this requirement for maximal effici- ency of growth initiation appears fulfilled through the addition of “filler” thymus cells. It remains to be investigated whether thymus “filler” cells will also increase the
Exp Cdl Res III (1978)
efticiency of growth initiation for other eukaryotic cells in suspension culture. The able technical assistance of MS Helen Campbell and MS Joy Monckton is gratefully acknowledged.
Jan Andersson was supported by the Swedish Can- cer Society.
1. 2. 3.
4.
5.
6.
7.
8.
9. 10.
11.
12. 13.
14.
15.
16.
17. 18.
19.
20.
21.
22.
REFERENCES Metcalf, D, J cell physiol8 1 (1973) 397. - J immunol 113 (1974) 235. Metcalf, D, Warner, N L, Nossal, G J V, Miller, J F A P, Shortman, K & Rabellino, E, Nature 255 (1975a) 630. Metcalf, D, Nossal, G J V, Warner, N L, Miller, J F A P. Mandel. T E. Lavton. J E & Gutman. GA. J exp med 142 (1975b) 1534. Fibach. E. Gerassi. E & Sachs. L. Nature 259 (1976) i27.’
,
Riou, N, Biozard, G, Alcalay, D, Goube de Laforest, P & Tanzer, J, Ann immunol (Inst Pasteur) 127C (1976) 83. Wilson, J D & Dalton, G, Austral j exp biol med sci 54 (1976) 27. Watanabe, T, Fathman, C G & Coutinho, A, Transnlant rev 35 ( 1977) 3. Molle;, G, Transplant rev 11 (ed) (1972). Melchers. F. Coutinho. A. Heinrich. G & Anders- son, J, Stand j immundl4’(1975) 853. Andersson, J, Coutinho, A, Melchers, F & Watanabe, T, Cold Spring Harbor symp quant biol 41 (1977) 227. Stocker, J W, Immunology 30 (1976) 181. Schilling, R M, Phillips, R A & Miller, R G, .I exp med 144 (1976) 241. Teh, H S, Harley, E, Phillips, R A & Miller, R G, J immunol 118 (1977) 1049. Andersson, J, Coutinho, A, Lemhardt, W & Melchers, F, Cell 10 (1977) 27. Gronowicz, E, Coutinho, A & Melchers, F, Eur j immunol6 (1976) 588. Kiihler, G & Milstein. C. Nature 256 (1975) 495. Galfre, G, Howe, S d, Milstein, C, Butcher, G W & Howard, J C, Nature 266 (1977) 550. Krammer, P H, Citronbaum, R, Read, S E, Fomi, L & Lang, R, Cell immunol21 (1976) 97. Haran Ghera, N & Peled, A, Nature 241 (1973) 396. Kohler, G & Milstein, C, Eur j immunol 6 (1976) 511. Abelson, H T & Rabstein, L S, Cancer res 30 (1970) 2208.
23. ‘Andersson, J, Melchers, F, Galanos, C & Lii- deritz, 0, J exp med 137 (1973) 943.
24. Littlefield. J W. Proc natl acad sci US 50 (1963) 568.
. I
25. Anderson, E P & Law, L W, Ann rev biochem 29 (1%9) 577.
26. Chu, E J Y, Brimer, P & Mailing, H V, Genetics 61 (1%9) 10.
27. Melchers, F, Eur j immunol(l977). In press.
Received June 30, 1977 Accepted September 1, 1977




















