
Clewell-R.A. 2010 Toxicol.-Vitro
8
Assessing the relevance of in vitro measures of phthalate inhibition of steroidogenesis for in vivo response Rebecca A. Clewell a,b, * , Jerry L. Campbell b , Susan M. Ross b , Kevin W. Gaido b , Harvey J. Clewell III b , Melvin E. Andersen b a University of North Carolina, Chapel Hill, NC 27599, USA b The Hamner Institutes for Health Sciences, RTP, NC 27709, USA article info Article history: Received 16 May 2009 Accepted 13 August 2009 Available online 20 August 2009 Keywords: Phthalates Developmental toxicity Testosterone inhibition Fetal rat Testes abstract Human phthalate exposure occurs as mixtures of diesters with varying activity towards testosterone- dependent development. Dibutyl (DBP), diethylhexyl (DEHP) and butylbenzyl (BBP) phthalate disrupt sexual development in the fetal rat. Dimethyl (DMP) and diethyl (DEP) phthalate do not. These differ- ences in potency may result from differential delivery of the monophthalates to the testes or from var- iation in the abilities of the compounds to alter steroidogenesis. We tested five phthalates in pregnant rats (500 mg/kg-d, GD12–19) and analyzed the fetal testes for corresponding monoesters (MMP, MEP, MBP, MEHP, MBeP). Testes MMP and MEP levels were 2–40-fold higher than the active monoesters, MBP and MEHP. BBP exposure led to low concentrations of MBeP, but similar MBP levels to DBP. An in vitro MA-10 cell assay measured the direct effect of monophthalates on testosterone production. MEHP inhibited LH-stimulated testosterone production at 1 lM. RT-PCR confirmed down-regulation of genes associated with cholesterol transport and steroid synthesis and metabolism by MEHP. Five additional phthalates were tested for testosterone inhibition. MBP and mono-n-octyl phthalate were similar to MEHP; MMP, MEP and MBeP were poor inhibitors of testosterone production. Based on these results, dif- ferences in the phthalates’ ability to interfere with sexual development in vivo appears to be more asso- ciated with differential potency for testosterone inhibition than differences in tissue doses. Ó 2009 Elsevier Ltd. All rights reserved. 1. Introduction Exposure to phthalate esters occurs through drinking water, food and personal care products. Some of these esters are also used as a coating for medications (Heudorf et al., 2007). Metabolites of several phthalates are found in the blood and urine from biomon- itoring studies in the United States and Europe at all age ranges from infants to adults, and in milk from lactating women (Brock et al., 2002; Högberg et al., 2008; Kohn et al., 2000; Silva et al., 2004). A recent study in Europeans found that food sources ac- counted for 40–90% of the overall exposure to di-n-butyl phthalate (DBP) and di-2,4-ethylhexyl phthalate (DEHP); inhalation and der- mal exposures are dominant sources of diethyl (DEP) and dimethyl phthalate (DMP) (Wormuth et al., 2006). Some phthalates, such as DEHP, DBP and butylbenzyl phthalate (BBP), disrupt male sexual development in the rat. Administration of DBP, DEHP, or BBP to pregnant rats causes reduced anogenital distance (AGD), nipple retention, hypospadias, delayed testes des- cent, and vaginal pouch development in male offspring (Gray et al., 2000; Mylchreest et al., 2000). Several other phthalate esters, how- ever, including diethyl phthalate (DEP), dimethyl phthalate (DMP), and dioctyl terephthalate (Gray et al., 2000) do not affect sexual development of the male rat. The monoester metabolites are the biologically active forms of the phthalate diesters. DBP and DEHP and their monoesters, MBP and MEHP, have been studied extensively in vivo in an effort to elu- cidate their mechanism of action. While the molecular targets have not been identified, the monoester metabolites of the endocrine active phthalates have been shown act directly on the testes, and more specifically the Leydig cell, reducing the steroidogenic re- sponse to luteinizing hormone (LH) (Jones et al., 1993; Parks et al., 2000). Gene array studies performed on fetal rat testes fol- lowing gestational phthalate exposure (Lehmann et al., 2004; Liu et al., 2005; Shultz et al., 2001) showed consistent down-regulation of genes associated with steroidogenesis, including genes encoding proteins at multiple points in the testosterone biosynthetic path- way. Some of the key genes identified in these studies include scav- enger receptor class B family 1 (SR-B1), steroid acute regulatory protein (StAR), cytochrome P450 17a1 (Cyp17a1), and P450 side chain 0887-2333/$ - see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.tiv.2009.08.003 * Corresponding author. Address: The Hamner Institutes for Health Sciences, 6 Davis Drive, Research Triangle Park, NC 27709, USA. Tel.: +1 919 558 1307; fax: +1 919 558 1300. E-mail address: [email protected] (R.A. Clewell). Toxicology in Vitro 24 (2010) 327–334 Contents lists available at ScienceDirect Toxicology in Vitro journal homepage: www.elsevier.com/locate/toxinvit
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Clewell-R.A. 2010 Toxicol.-Vitro

Toxicology in Vitro 24 (2010) 327–334
Contents lists available at ScienceDirect
Toxicology in Vitro
journal homepage: www.elsevier .com/locate / toxinvi t
Assessing the relevance of in vitro measures of phthalate inhibitionof steroidogenesis for in vivo response
Rebecca A. Clewell a,b,*, Jerry L. Campbell b, Susan M. Ross b, Kevin W. Gaido b,Harvey J. Clewell III b, Melvin E. Andersen b
a University of North Carolina, Chapel Hill, NC 27599, USAb The Hamner Institutes for Health Sciences, RTP, NC 27709, USA
a r t i c l e i n f o
Article history:Received 16 May 2009Accepted 13 August 2009Available online 20 August 2009
Keywords:PhthalatesDevelopmental toxicityTestosterone inhibitionFetal ratTestes
0887-2333/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.tiv.2009.08.003
* Corresponding author. Address: The Hamner InsDavis Drive, Research Triangle Park, NC 27709, USA. T919 558 1300.
E-mail address: [email protected] (R.A. Clew
a b s t r a c t
Human phthalate exposure occurs as mixtures of diesters with varying activity towards testosterone-dependent development. Dibutyl (DBP), diethylhexyl (DEHP) and butylbenzyl (BBP) phthalate disruptsexual development in the fetal rat. Dimethyl (DMP) and diethyl (DEP) phthalate do not. These differ-ences in potency may result from differential delivery of the monophthalates to the testes or from var-iation in the abilities of the compounds to alter steroidogenesis. We tested five phthalates in pregnantrats (500 mg/kg-d, GD12–19) and analyzed the fetal testes for corresponding monoesters (MMP, MEP,MBP, MEHP, MBeP). Testes MMP and MEP levels were 2–40-fold higher than the active monoesters,MBP and MEHP. BBP exposure led to low concentrations of MBeP, but similar MBP levels to DBP. Anin vitro MA-10 cell assay measured the direct effect of monophthalates on testosterone production. MEHPinhibited LH-stimulated testosterone production at 1 lM. RT-PCR confirmed down-regulation of genesassociated with cholesterol transport and steroid synthesis and metabolism by MEHP. Five additionalphthalates were tested for testosterone inhibition. MBP and mono-n-octyl phthalate were similar toMEHP; MMP, MEP and MBeP were poor inhibitors of testosterone production. Based on these results, dif-ferences in the phthalates’ ability to interfere with sexual development in vivo appears to be more asso-ciated with differential potency for testosterone inhibition than differences in tissue doses.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Exposure to phthalate esters occurs through drinking water,food and personal care products. Some of these esters are also usedas a coating for medications (Heudorf et al., 2007). Metabolites ofseveral phthalates are found in the blood and urine from biomon-itoring studies in the United States and Europe at all age rangesfrom infants to adults, and in milk from lactating women (Brocket al., 2002; Högberg et al., 2008; Kohn et al., 2000; Silva et al.,2004). A recent study in Europeans found that food sources ac-counted for 40–90% of the overall exposure to di-n-butyl phthalate(DBP) and di-2,4-ethylhexyl phthalate (DEHP); inhalation and der-mal exposures are dominant sources of diethyl (DEP) and dimethylphthalate (DMP) (Wormuth et al., 2006).
Some phthalates, such as DEHP, DBP and butylbenzyl phthalate(BBP), disrupt male sexual development in the rat. Administrationof DBP, DEHP, or BBP to pregnant rats causes reduced anogenital
ll rights reserved.
titutes for Health Sciences, 6el.: +1 919 558 1307; fax: +1
ell).
distance (AGD), nipple retention, hypospadias, delayed testes des-cent, and vaginal pouch development in male offspring (Gray et al.,2000; Mylchreest et al., 2000). Several other phthalate esters, how-ever, including diethyl phthalate (DEP), dimethyl phthalate (DMP),and dioctyl terephthalate (Gray et al., 2000) do not affect sexualdevelopment of the male rat.
The monoester metabolites are the biologically active forms ofthe phthalate diesters. DBP and DEHP and their monoesters, MBPand MEHP, have been studied extensively in vivo in an effort to elu-cidate their mechanism of action. While the molecular targets havenot been identified, the monoester metabolites of the endocrineactive phthalates have been shown act directly on the testes, andmore specifically the Leydig cell, reducing the steroidogenic re-sponse to luteinizing hormone (LH) (Jones et al., 1993; Parkset al., 2000). Gene array studies performed on fetal rat testes fol-lowing gestational phthalate exposure (Lehmann et al., 2004; Liuet al., 2005; Shultz et al., 2001) showed consistent down-regulationof genes associated with steroidogenesis, including genes encodingproteins at multiple points in the testosterone biosynthetic path-way. Some of the key genes identified in these studies include scav-enger receptor class B family 1 (SR-B1), steroid acute regulatoryprotein (StAR), cytochrome P450 17a1 (Cyp17a1), and P450 side chain

Fig. 1. Genes encoding proteins in the Leydig cell steroidogenic pathway. Brokenlines represent transport proteins, solid arrows represent enzymatic proteins.
328 R.A. Clewell et al. / Toxicology in Vitro 24 (2010) 327–334
cleavage (P450scc) (Fig. 1). SR-B1 and StAR encode proteins respon-sible for the transport of cholesterol into the cell and across themitochondrial membrane, respectively. P450scc is required forthe conversion of cholesterol to pregnenolone, which is eventuallymetabolized by Cyp17a1 to form androstenedione, the immediateprecursor to testosterone.
Inactive phthalates, such as DEP and DMP, do not reduce testos-terone or expression of steroidogenic genes the testes of fetal rats.It is not clear why some phthalates disrupt testosterone-dependentdevelopment and others do not. The difference in potency may re-sult from the length and extent of branching in the alkyl chains(Gray et al., 2000). The structural differences in the monoalkylest-ers could affect metabolite kinetics, as well as their ability to inter-fere with steroidogenesis. The parent diester and monoestermetabolites for some of the more common phthalates are shownin Fig. 2.
Fig. 2. Chemical structure of c
Because human exposure occurs as a mixture of active and inac-tive phthalates, one of the more important considerations in acomprehensive risk assessment is accounting for the differencesin potency among phthalates. Ideally, the cumulative risk assess-ment for a family of chemicals such as the phthalates would at-tempt to account for the combined exposure to only thosechemicals with the same molecular targets. The studies describedin this paper were designed to address these issues specifically.First, an in vivo study was performed to determine the target tissue(fetal testes) concentrations associated with phthalate develop-mental effects. Second, an in vitro assay was developed to testwhether the measured phthalate concentrations would be suffi-cient to inhibit steroidogenesis directly in the steroidogenic cellof the testes, the Leydig cell. The described assay can provide usefulestimates of phthalate potency in vivo.
2. Methods
Six different phthalates were chosen to represent compoundswith different potencies for steroidogenic effects: butylbenzylphthalate (BBP), DBP, DEP, DEHP, DMP, and di-n-octyl phthalate(DnOP). Two of the tested phthalates are known inhibitors ofmale rat sexual development: DEHP and DBP. Two of the testedphthalates do not disrupt reproductive tract developmentin vivo: DEP and DMP. Butylbenzyl phthalate (BBP) treatmentcauses similar developmental effects and gene expression changesto DEHP and DBP (Gray et al., 2000; Liu et al., 2005). However, itis not known which metabolite is responsible for the anti-andro-genic effects, as BBP may be metabolized to MBP or MBeP prior toabsorption. The in vivo activity of DnOP has not been tested in thefetal rat.
2.1. Determination of fetal testes phthalate concentrations associatedwith developmental effects
2.1.1. AnimalsPregnant Sprague–Dawley (Crl:CD(SD)) rats (sperm plug posi-
tive GD 0) were obtained from Charles River Laboratories (Raleigh,NC) and housed in the Animal Care Facility of the The Hamner
ommon phthalate esters.

R.A. Clewell et al. / Toxicology in Vitro 24 (2010) 327–334 329
Institutes for Health Sciences, which is accredited by the Associa-tion for Assessment and Accreditation of Laboratory Animal CareInternational. Rats were acclimated in a temperature- and humid-ity-controlled, HEPA-filtered environment on a 12 h light–dark cy-cle. NIH rodent diet (NIH-07, Zeigler Bros., Gardner, PA) andreverse-osmosis water were provided ad libitum. This study wasapproved by The Hamner Institute’s Animal Care and UseCommittee.
2.1.2. TreatmentAnimal exposures, sacrifice and tissue collection has been de-
scribed previously (Liu et al., 2005). Briefly, dosing solutions(500 mg/mL) were prepared by mixing DEHP, DBP, BBP, DEP orDMP and corn oil (Sigma–Aldrich, St. Louis, MO). Pregnant ratswere dosed via oral gavage with corn oil (1.0 mL/kg/day) or oneof the phthalates (500 mg/kg/day) dissolved in corn oil (1.0 mL/kg) from GD 12 through 19. The dams were euthanized by CO2
asphyxiation and exsanguination 2 h after the final dose. Fetuseswere delivered by caesarean section and sex was determined byinternal inspection of gonads. Testes from each male fetus werecollected, snap frozen in liquid nitrogen, and stored individuallyat �80 �C. Testes samples that were not used for gene expressiondata published by Liu et al. (2005) were generously provided tous for measurement of phthalate concentrations.
2.1.3. Phthalate analysisThe monoester phthalates (MEHP, MBP, MBeP, MEP, and
MMP) were extracted from testes prior to analysis using a previ-ously described method for MBP (Clewell et al., 2008b). Each tes-tes pair was homogenized in 1% PBS (500 lL) with a Mixer Mill300 for 5 min at 30 Hz/s at room temperature. The homogenatewas transferred to a 2.0 mL microcentrifuge tube and 200 lL of1% formic acid/water and 25 lL of [13C]-MBP (6 mg/mL) wereadded to the tube. The sample was vortexed for 5 s and thencentrifuged at 1900g for 6 min at 4 �C. The supernatant wasloaded onto an LC-18 SPE column (Honeywell Burdick and Jack-son, Morristown, NJ) that had been pre-activated with an aque-ous solution of 2 mL of 0.1 formic acid, added in two 1 mLincrements. After passing 2 mL of a 0.1% formic acid in water/0.1 formic acid in methanol (75%/25%) in 1 mL increments, themonoester was eluted with a methanol solution of 0.1% formicacid (2 mL) in 1 mL increments. The samples were evaporatedto dryness under nitrogen and reconstituted in 200 lL of 0.1%formic acid in ACN/H2O (90%/10%) for analysis by LC-MS/MS,performed according to previously published methods (Fennellet al., 2004).
2.2. Development of an in vitro assay for phthalate inhibition ofsteroidogenesis
2.2.1. Cell cultureMouse Leydig tumor (MA-10) cells were obtained from Dr. Mar-
io Ascoli (Department of Pharmacology, University of Iowa, Collegeof Medicine, Iowa City). Stock cells were grown in 150 mm tissue-culture dishes as described by Ascoli (1981) in complete medium(Waymouth’s MB 752/1 supplemented with 15% horse serum, 2%L-glutamine, 20 mM HEPES, and 50 lg/mL Gentamicin). For all cellculture experiments, dishes were coated with a 0.1% solution ofgelatin for 30 min before plating cells. All incubations were per-formed at 37 �C in a humidity-controlled atmosphere of 5% CO2-air.
2.2.2. Progesterone and testosterone inhibitionCells were plated in 24-well plates at a density of 1 � 105 cells/
well in 0.5 mL complete medium and allowed to incubate for 24 h.The medium was then removed and replaced with serum-freemedium (SFM) containing 0 (0.1% DMSO), 1, 3, 10, 30, 100, 300,
1000 lM MEHP. After 24 h, the medium was replaced with SFMcontaining the same MEHP concentration supplemented with a100 ng/mL LH. The medium was collected after 6 h and stored at�20 �C. Progesterone (PG) and testosterone in the medium weremeasured using ELISA assays (Neogen Corporation, Lexington,KY) according to the manufacturer’s directions.
2.2.3. Protein content and viability analysisCells from each well were treated with 100 lL of lysis buffer
(0.1 M Tris–HCl (pH 8.0), 0.05 M EDTA, 0.1 M NaCl, 1% sodiumdodecyl sulfate, 1% sarcosyl) supplemented with HALT ProteaseInhibitor Cocktail (10 lL/1 mL buffer) (Thermo Fisher ScientificInc., Rockford, IL) and dislodged using a cell scraper. The resultinglysate was homogenized and stored at �80 �C prior to analysis. To-tal protein content was measured using a bicinchoninic acid (BCA)assay (Sigma–Aldrich) according to the manufacturer’s directions.Cell viability was also tested in three separate assays (threewells/assay) using the ATPlite luminescence assay (Perkin Elmer,Boston, MA) according to manufacturer’s instructions.
2.2.4. Quantitative RT-PCRMA-10 cells were plated at a density of 3 � 106 cells/well in
100 mm cell culture dishes in 20 mL complete medium. About24 h after incubation at 37 �C, medium was replaced with 20 mLSFM containing 0 (DMSO) or 100 lM MEHP. Cells were incubatedfor 24 h and medium was replaced with SFM containing the sameinhibitor concentration supplemented with PBS (no treatmentcontrol) or 100 ng/mL LH. After 6 h, medium was aspirated fromthe plate, cells were washed with Ca2+/Mg2+-free PBS and removedfrom the plate using a disposable cell scraper. Total RNA was iso-lated from the cells of each cell culture dish using an RNeasy MiniKit (Qiagen, Valencia, CA). cDNA was prepared for each sampleusing the High Capacity cDNA kit (Applied Biosystems, Foster City,CA) according to the manufacturer’s protocol. Reverse transcrip-tion (RT) reactions were performed on the Taqman 7900HT sys-tem (Applied Biosystems, Foster City, CA). RNA and DNA wereanalyzed spectrophotometrically. Taqman gene expression assaysfor mouse Cyp17A1, GAPDH, P450scc, SR-B1, and StAR were pur-chased from Applied Biosystems (Foster City, CA). Samples(n = 6/treatment group) were run in triplicate and the change inexpression of target genes were normalized to GAPDH. Foldchange in GAPDH between LH controls (1.0 + 0.2) and MEHP trea-ted cells (0.9 + 0.2) was calculated using the 2�DCt method.Expression of GAPDH was not significantly different between con-trol and MEHP treated cells, indicating that it is a reasonable con-trol for this assay. The fold change for each gene normalized toGAPDH and relative to expression in no treatment control (NTC)samples was determined by the 2�DDCt method (Livak and Sch-mittgen, 2001).
2.3. Comparison of Phthalate Potency for Inhibition of Steroidogenesis
The ability of five additional phthalates to inhibit testosteronesynthesis in the MA-10 cell was examined using the method de-scribed for MEHP above. MBP (TCI America), MEP (Accustandard),and MnOP, MBeP, and MMP (Sigma–Aldrich) were dissolved sepa-rately in DMSO to obtain stock solutions (1 M) for each of thephthalates. Cells were pre-treated with 0, 1, 3, 10, 30, or 100 lMof an individual phthalate for 24 h in SFM. Cells were then stimu-lated with 100 ng/mL LH in the presence of the monophthalate (orDMSO) and medium was collected for hormone analysis. Cell pro-tein content was assayed for all tested compounds at each dose.Cell viability assays were also run for each treatment in three sep-arate assays (three wells/assay) using the ATPlite luminescence as-say (Perkin Elmer, Boston, MA).

330 R.A. Clewell et al. / Toxicology in Vitro 24 (2010) 327–334
3. Results
3.1. Determination of fetal testes phthalate concentrations associatedwith developmental effects
Despite the number of in vivo effect studies, little information isavailable regarding phthalate disposition during gestation. In fact,active metabolite (MBP) concentrations in the target tissue (fetaltestes) have only been published for DBP (Clewell et al., 2008b).In order to interpret the effect studies and determine the appropri-ate phthalate concentrations for in vitro assays, it is necessary tohave some estimation of active metabolite concentrations in thefetal testes. Thus, testes were collected from fetuses of damsadministered 500 mg/kg/day DMP, DEP, BBP, DBP, or DEHP fromgestation day (GD) 12–19, and their free monoester metabolites,MMP, MEP, MBP or MBeP, MBP and MEHP, respectively, were mea-sured using HPLC/MS-MS (Fig. 3).
All of the monoesters were found in the fetal testes after mater-nal exposure to the dialkyl esters. MMP and MEP, the monoestersderived from the two inactive phthalates were present at the high-est concentrations, with mean concentrations of 396 and 409 lM,respectively. Mean testes concentrations of MBP (189 lM) indicatethat it is also efficiently transported into the fetus. MEHP, on theother hand, was present at much lower concentrations (12 lM)than MBP (189 lM), despite the fact that both DBP and DEHP arepotent inhibitors of fetal development at this administered dose(500 mg/kg/day). In rats exposed to BBP, MBP was the majormetabolite found in the testes: the mean concentration (124 lM)was 5-fold greater than MBeP (21 lM).
3.2. Validation of an in vitro assay for phthalate inhibition ofsteroidogenesis
The PD effect of interest for the phthalates is delayed sexualdevelopment of the fetal rat (Gray et al., 2000; Liu et al., 2005).Inhibition of testosterone production in the Leydig cell of the fetaltestes has been shown to precede the more overt physiological ef-fects, such as nipple retention and hypospadias (Borch et al.,2004; Lehmann et al., 2004; Parks et al., 2000) and may be a use-ful early marker of sexual development disruption. To testwhether the phthalates have different potencies for steroid inhibi-tion, an assay was needed that allows direct exposure of theLeydig cell and has a measurable biomarker. In the absence of aLH-responsive rat cell line, an assay in the murine Leydig tumorMA-10 cell line was developed. MA-10 cells are known to produceprogesterone (PG; a testosterone precursor) in response to stimu-lation by luteinizing hormone and human chorionic gonadotropin.The MA-10 cell has been widely used as a model for studying thesteroidogenic pathway in the rodent Leydig cell, measuring alter-ations in gene expression and PG production under a variety ofconditions (Wang et al., 1999). The MA-10 cells have also been
Fig. 3. Measured monoester concentrations in the fetal testes from dams given daily orafter the final dose. Bars represent mean + SD from 3 fetuses of 2–3 dams.
shown to be susceptible to phthalates; Dees et al. (2001) reportedinhibition of PG production after stimulation with human chori-onic gonadotropin.
As PG was previously understood to be the terminal point inMA-10 steroid synthesis, initial assay development focused onthe ability of MEHP, a known inhibitor of steroid production, toinhibit LH-stimulated PG production. Treatment of MA-10 cellswas found to inhibit PG production significantly at 30 lM(Fig. 4). Cell viability assessed via intracellular ATP and total pro-tein content were >90% of control values at concentrations up to100 lM. At 300 and 1000 lM, significant cell loss was observed(Fig. 5).
Expression of the steroidogenic genes, SR-B1, StAR, Cyp17a1 andP450scc, were evaluated in MA-10 cells by quantitative RT-PCR. Allfour genes were significantly up-regulated within 6 h of LH-stimu-lation compared to controls (Fig. 6A). When pre-treated with100 lM MEHP, Cyp17a1, P450scc, SR-B1 and StAR were decreasedrelative to LH-stimulated cells (Fig. 6B).
Cyp17a1 changes were particularly interesting due to the factthat the enzyme is responsible for metabolism of PG, indicatingthat the enzymes necessary for synthesis of the subsequent ste-roids, androstenedione and testosterone, are present in the MA-10 cell. In response to these observations, medium from controland LH-stimulated MA-10 cells was analyzed for testosterone byELISA assay. The ELISA assay has less than 1% cross reactivity withthe precursor steroids, including progesterone, pregnenolone, 17-hydroxyprogesterone, and androstenedione. The MA-10 cells werefound to produce testosterone in response to LH-stimulation. At100 ng/mL LH, testosterone concentration in the medium was0.27 + 0.3 ng/mL (vs. <0.01 ng/mL in no treatment controls). Whenthe cells were pre-treated with MEHP, testosterone production wasreduced in a dose-dependent manner. MEHP concentrations as lowas 1 lM resulted in statistically significant (p < 0.05) testosteroneinhibition (Fig. 7A). The IC50 calculated from the one-site inhibitioncurve was 4.1 lM (Fig. 7B).
3.3. Comparison of phthalate potency for inhibition of steroidogenesis
MEHP induced inhibition of testosterone and steroidogenicgene expression in the MA-10 cell is consistent with in vivo phthal-ate studies, indicating that this assay may provide a reasonablemodel for the action of phthalate monoesters on Leydig cell func-tion. Because the in vitro assay eliminates pharmacokinetic (PK)differences inherent in in vivo studies, the MA-10 in vitro assaywas used to compare the PD behavior of different phthalates(Fig. 8). The dose–response curve for MBP, another endocrine ac-tive phthalate, was remarkably similar to that of MEHP, with sig-nificant inhibition at 3 lM and a calculated IC50 of 3.3 lM. Incontrast, neither MMP nor MEP inhibited testosterone synthesissignificantly at concentrations up to 100 lM. Although the 10 lMMMP treatment group was significantly different from controls,
al doses of DMP, DEP, DBP, DEHP, or BBP from GD 12–19. Testes were collected 2 h
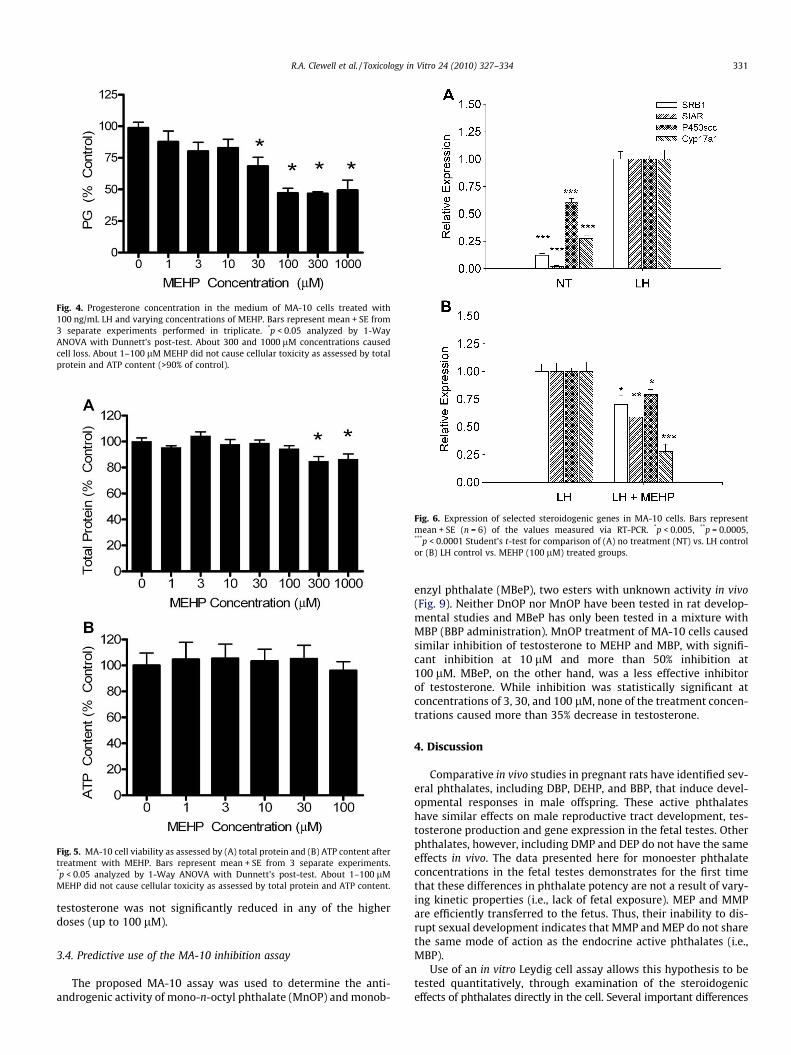
Fig. 5. MA-10 cell viability as assessed by (A) total protein and (B) ATP content aftertreatment with MEHP. Bars represent mean + SE from 3 separate experiments.*p < 0.05 analyzed by 1-Way ANOVA with Dunnett’s post-test. About 1–100 lMMEHP did not cause cellular toxicity as assessed by total protein and ATP content.
Fig. 6. Expression of selected steroidogenic genes in MA-10 cells. Bars representmean + SE (n = 6) of the values measured via RT-PCR. *p < 0.005, **p = 0.0005,***p < 0.0001 Student’s t-test for comparison of (A) no treatment (NT) vs. LH controlor (B) LH control vs. MEHP (100 lM) treated groups.
Fig. 4. Progesterone concentration in the medium of MA-10 cells treated with100 ng/mL LH and varying concentrations of MEHP. Bars represent mean + SE from3 separate experiments performed in triplicate. *p < 0.05 analyzed by 1-WayANOVA with Dunnett’s post-test. About 300 and 1000 lM concentrations causedcell loss. About 1–100 lM MEHP did not cause cellular toxicity as assessed by totalprotein and ATP content (>90% of control).
R.A. Clewell et al. / Toxicology in Vitro 24 (2010) 327–334 331
testosterone was not significantly reduced in any of the higherdoses (up to 100 lM).
3.4. Predictive use of the MA-10 inhibition assay
The proposed MA-10 assay was used to determine the anti-androgenic activity of mono-n-octyl phthalate (MnOP) and monob-
enzyl phthalate (MBeP), two esters with unknown activity in vivo(Fig. 9). Neither DnOP nor MnOP have been tested in rat develop-mental studies and MBeP has only been tested in a mixture withMBP (BBP administration). MnOP treatment of MA-10 cells causedsimilar inhibition of testosterone to MEHP and MBP, with signifi-cant inhibition at 10 lM and more than 50% inhibition at100 lM. MBeP, on the other hand, was a less effective inhibitorof testosterone. While inhibition was statistically significant atconcentrations of 3, 30, and 100 lM, none of the treatment concen-trations caused more than 35% decrease in testosterone.
4. Discussion
Comparative in vivo studies in pregnant rats have identified sev-eral phthalates, including DBP, DEHP, and BBP, that induce devel-opmental responses in male offspring. These active phthalateshave similar effects on male reproductive tract development, tes-tosterone production and gene expression in the fetal testes. Otherphthalates, however, including DMP and DEP do not have the sameeffects in vivo. The data presented here for monoester phthalateconcentrations in the fetal testes demonstrates for the first timethat these differences in phthalate potency are not a result of vary-ing kinetic properties (i.e., lack of fetal exposure). MEP and MMPare efficiently transferred to the fetus. Thus, their inability to dis-rupt sexual development indicates that MMP and MEP do not sharethe same mode of action as the endocrine active phthalates (i.e.,MBP).
Use of an in vitro Leydig cell assay allows this hypothesis to betested quantitatively, through examination of the steroidogeniceffects of phthalates directly in the cell. Several important differences
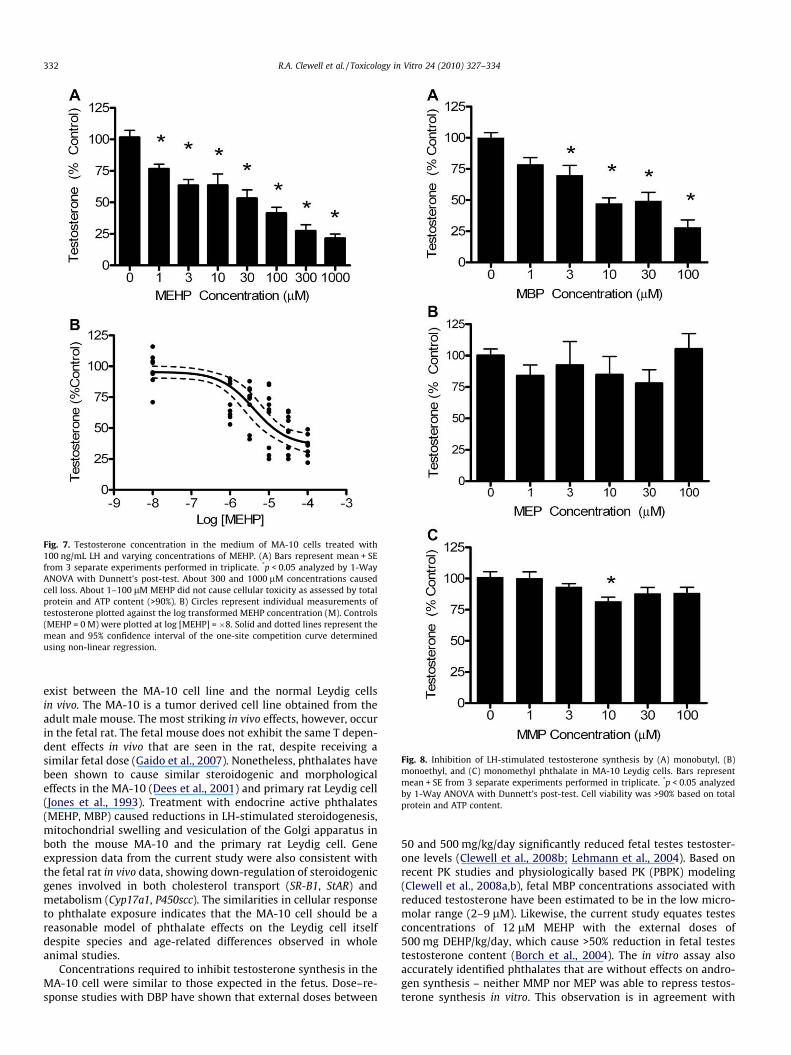
Fig. 7. Testosterone concentration in the medium of MA-10 cells treated with100 ng/mL LH and varying concentrations of MEHP. (A) Bars represent mean + SEfrom 3 separate experiments performed in triplicate. *p < 0.05 analyzed by 1-WayANOVA with Dunnett’s post-test. About 300 and 1000 lM concentrations causedcell loss. About 1–100 lM MEHP did not cause cellular toxicity as assessed by totalprotein and ATP content (>90%). B) Circles represent individual measurements oftestosterone plotted against the log transformed MEHP concentration (M). Controls(MEHP = 0 M) were plotted at log [MEHP] = �8. Solid and dotted lines represent themean and 95% confidence interval of the one-site competition curve determinedusing non-linear regression.
Fig. 8. Inhibition of LH-stimulated testosterone synthesis by (A) monobutyl, (B)monoethyl, and (C) monomethyl phthalate in MA-10 Leydig cells. Bars representmean + SE from 3 separate experiments performed in triplicate. *p < 0.05 analyzedby 1-Way ANOVA with Dunnett’s post-test. Cell viability was >90% based on totalprotein and ATP content.
332 R.A. Clewell et al. / Toxicology in Vitro 24 (2010) 327–334
exist between the MA-10 cell line and the normal Leydig cellsin vivo. The MA-10 is a tumor derived cell line obtained from theadult male mouse. The most striking in vivo effects, however, occurin the fetal rat. The fetal mouse does not exhibit the same T depen-dent effects in vivo that are seen in the rat, despite receiving asimilar fetal dose (Gaido et al., 2007). Nonetheless, phthalates havebeen shown to cause similar steroidogenic and morphologicaleffects in the MA-10 (Dees et al., 2001) and primary rat Leydig cell(Jones et al., 1993). Treatment with endocrine active phthalates(MEHP, MBP) caused reductions in LH-stimulated steroidogenesis,mitochondrial swelling and vesiculation of the Golgi apparatus inboth the mouse MA-10 and the primary rat Leydig cell. Geneexpression data from the current study were also consistent withthe fetal rat in vivo data, showing down-regulation of steroidogenicgenes involved in both cholesterol transport (SR-B1, StAR) andmetabolism (Cyp17a1, P450scc). The similarities in cellular responseto phthalate exposure indicates that the MA-10 cell should be areasonable model of phthalate effects on the Leydig cell itselfdespite species and age-related differences observed in wholeanimal studies.
Concentrations required to inhibit testosterone synthesis in theMA-10 cell were similar to those expected in the fetus. Dose–re-sponse studies with DBP have shown that external doses between
50 and 500 mg/kg/day significantly reduced fetal testes testoster-one levels (Clewell et al., 2008b; Lehmann et al., 2004). Based onrecent PK studies and physiologically based PK (PBPK) modeling(Clewell et al., 2008a,b), fetal MBP concentrations associated withreduced testosterone have been estimated to be in the low micro-molar range (2–9 lM). Likewise, the current study equates testesconcentrations of 12 lM MEHP with the external doses of500 mg DEHP/kg/day, which cause >50% reduction in fetal testestestosterone content (Borch et al., 2004). The in vitro assay alsoaccurately identified phthalates that are without effects on andro-gen synthesis – neither MMP nor MEP was able to repress testos-terone synthesis in vitro. This observation is in agreement with

Fig. 9. Inhibition of LH-stimulated testosterone synthesis by (A) mono-n-octyl and(B) monobenzyl phthalate in MA-10 Leydig cells. Bars represent mean + SE from 3separate experiments performed in triplicate. *p < 0.05 analyzed by 1-Way ANOVAwith Dunnett’s post-test. Cell viability was >90% based on total protein and ATPcontent.
R.A. Clewell et al. / Toxicology in Vitro 24 (2010) 327–334 333
data from isolated rat Leydig cells (Jones et al., 1993) and in vivostudies where DEP and DMP did not cause the reproductive (Grayet al., 2000) or gene expression changes (Liu et al., 2005), despitesignificant exposure of the fetus to the monoester metabolites ver-ified in this study.
Application of this in vitro assay to other phthalates enables theclassification of various phthalate metabolites according to theirability to inhibit steroid production, supporting a more informedassessment of human risk. MnOP has not been tested in the fetalrat. It reduced testosterone synthesis in the MA-10 cell, indicatingthat it likely shares the same mechanism of action as MBP andMEHP. This conclusion is consistent with studies in the adult rat,showing similar effects of DEHP and DnOP administration in theLeydig cells of adult rats in vivo, as well as reduced testosteroneproduction in cultured rat Leydig cells (Jones et al., 1993). Interest-ingly, MBeP does not appear to be an effective inhibitor of steroido-genesis in vitro (when compared to MEHP or MBP), despite the factthat BBP has been shown to be a potent anti-androgen in fetal rats(Gray et al., 2000; Lehmann et al., 2004). The low levels of testos-terone inhibition by MBeP in vitro, together with the testes metab-olite data showing minimal transfer of MBeP to the fetus, indicatesthat the observed fetal effects after administration of BBP are likelya result of the preferential hydrolysis of BBP to MBP in the rat(Eigenberg et al., 1986; Nativelle et al., 1999).
The use of this MA-10 assay to evaluate mixed exposures wouldbe particularly useful in the effort to predict cumulative humanrisk, since human exposure occurs as a mixture of both endocrineactive and inactive compounds. In vivo efforts to evaluate mixed
exposures are limited by the expense incurred for the many setsof exposures required and the challenges in accounting for differ-ences in pharmacokinetics when rats are dosed with multiple com-pounds at high doses that produce dose-dependent changes inkinetics (Clewell et al., 2008a). A reliable in vitro test would also al-low examination of relative potencies and the combined effect ofoxidative metabolites in addition to the intact monoesters. MEHP,for example, is only a minor metabolite of DEHP in the blood of therat (Albro and Lavenhar, 1989). In the human, blood concentrationsof potentially endocrine active oxidative metabolites such asmono(2-ethyl-5-oxohexyl) phthalate and mono(2-ethyl-5-hydrox-yhexyl) phthalate are similar to MEHP (Koch et al., 2005). A betterunderstanding of the relative potency of the monoester derivativesand their combined effect on the Leydig cell would be valuable inpredicting human risk and in determining the most informativemeasure of human exposure for biomonitoring data collection.Nonetheless, the current studies provide information on the phar-macokinetic and pharmacodynamic factors governing the differingpotency of several prominent phthalate esters.
From these studies, it is evident that pharmacokinetic factorsare not responsible for the differences in phthalate potencyin vivo. The ability of the phthalate esters to cause anti-androgeniceffects is determined by their different potency for steroid inhibi-tion within the Leydig cell itself. The MA-10 assay described hereprovides a means for estimating relative anti-androgenic potentialin vitro and may be used to inform cumulative risk assessment ef-forts. For example, because MBP and MEHP are commonly found inhuman urine, and have anti-androgenic activity in the fetal rat andin vitro, both would need to be included in a cumulative phthalaterisk assessment. Despite the lack of information on DnOP fetaldevelopment, it would also be advisable to account for its potentialexposure in a cumulative risk assessment, as its primary metabo-lite reduces Leydig cell steroid production. Limited studies indicatethat tissue and plasma kinetics of MnOP are similar to MEHP(Oishi, 1990). MEP and MMP, on the other hand, should be ex-cluded from calculations of total phthalate exposure in relationto fetal testes effects as they do not share the same mode of action.BBP is an interesting case, in that despite the adverse effects seenin rat developmental studies, it might still be excluded from acumulative risk assessment in the human. The observed effectsin the rat are clearly driven by MBP concentrations. The human,however, preferentially hydrolyzes BBP to MBeP (Anderson et al.,2001), which exhibits limited anti-androgenic activity in vitro. Inthis case, species PK differences indicate that the human wouldbe less susceptible than the rat to developmental toxicity fromBBP exposure.
5. Conflict of interest statement
The authors declare no conflict of interest.
Acknowledgements
The authors express their gratitude to Dr. Mark Sochaski andTrista Kohen for assistance with analytical chemistry. This studywas funded by the Long-range Research Initiative of the AmericanChemistry Council (ACC-LRI) and US EPA – Science to Achieve Re-sults (STAR) Program (Grant # RD83386701). Financial sponsorsdid not participate in the design or implementation of the studies,or in manuscript preparation.
References
Albro, P.W., Lavenhar, S.R., 1989. Metabolism of di(2-ethylhexyl)phthalate. DrugMetabolism Reviews 21 (1), 13–34.

334 R.A. Clewell et al. / Toxicology in Vitro 24 (2010) 327–334
Anderson, W.A., Castle, L., Scotter, M.J., Massey, R.C., Springall, C., 2001. A biomarkerapproach to measuring human dietary exposure to certain phthalate diesters.Food Additives and Contaminants 18 (12), 1068–1074.
Ascoli, M., 1981. Characterization of several clonal lines of cultured Leydig tumorcells: gonadotropin receptors and steroidogenic responses. Endocrinology 108(1), 88–95.
Borch, J., Ladefoged, O., Hass, U., Vinggaard, A.M., 2004. Steroidogenesis in fetal malerats is reduced by DEHP and DINP, but endocrine effects of DEHP are notmodulated by DEHA in fetal, prepubertal and adult male rats. ReproductiveToxicology 18 (1), 53–61.
Brock, J.W., Caudill, S.P., Silva, M.J., Needham, L.L., Hilborn, E.D., 2002. Phthalatemonoesters levels in the urine of young children. Bulletin of EnvironmentalContamination and Toxicology 68 (3), 309–314.
Clewell, R.A., Kremer, J.J., Williams, C.C., Campbell Jr., J.L., Andersen, M.E., Borghoff,S.J., 2008a. Tissue exposures to free and glucuronidated monobutylyphthalatein the pregnant and fetal rat following exposure to di-n-butylphthalate:Evaluation with a PBPK model. Toxicological Sciences 103 (2), 241–259.
Clewell, R.A., Kremer, J.J., Williams, C.C., Campbell, J.L., Sochaski, M.A., Andersen,M.E., Borghoff, S.J., 2008b. Kinetics of selected di-n-butyl phthalate metabolitesand fetal testosterone following repeated and single administration in pregnantrats. Toxicology 255 (1–2), 80–90.
Dees, J.H., Gazouli, M., Papadopoulos, V., 2001. Effect of mono-ethylhexylphthalate on MA-10 Leydig tumor cells. Reproductive Toxicology 15 (2), 171–187.
Eigenberg, D.A., Bozigian, H.P., Carter, D.E., Sipes, I.G., 1986. Distribution, excretion,and metabolism of butylbenzyl phthalate in the rat. Journal of Toxicology andEnvironmental Health 17 (4), 445–456.
Fennell, T.R., Krol, W.L., Sumner, S.C., Snyder, R.W., 2004. Pharmacokinetics ofdibutylphthalate in pregnant rats. Toxicological Sciences 82 (2), 407–418.
Gaido, K.W., Hensley, J.B., Liu, D., Wallace, D.G., Borghoff, S., Johnson, K.J., Hall, S.J.,Boekelheide, K., 2007. Fetal mouse phthalate exposure shows that gonocytemultinucleation is not associated with decreased testicular testosterone.Toxicological Sciences 97 (2), 491–503.
Gray, L.E.Jr., Ostby, J., Furr, J., Price, M., Veeramachaneni, D.N., Parks, L., 2000.Perinatal exposure to the phthalates DEHP, BBP, and DINP, but not DEP, DMP, orDOTP, alters sexual differentiation of the male rat. Toxicological Sciences 58 (2),350–365.
Heudorf, U., Mersch-Sundermann, V., Angerer, J., 2007. Phthalates: Toxicology andexposure. International Journal of Hygiene and Environmental Health 210 (5),623–634.
Högberg, J., Hanberg, A., Berglund, M., Skerfving, S., Remberger, M., Calafat, A.M.,Filipsson, A.F., Jansson, B., Johansson, N., Appelgren, M., Håkansson, H., 2008.Phthalate diesters and their metabolites in human breast milk, blood or serum,and urine as biomarkers of exposure in vulnerable populations. EnvironmentalHealth Perspectives 116 (3), 334–339.
Jones, H.B., Garside, D.A., Liu, R., Roberts, J.C., 1993. The influence of phthalate esterson Leydig cell structure and function in vitro and in vivo. Experimental andMolecular Pathology 58 (3), 179–193.
Koch, H.M., Bolt, H.M., Preuss, R., Angerer, J., 2005. New metabolites of di(2-ethylhexyl)phthalate (DEHP) in human urine and serum after single oral dosesof deuterium-labelled DEHP. Archives of Toxicology 79 (7), 367–376.
Kohn, M.C., Parham, F., Masten, S.A., Portier, C.J., Shelby, M.D., Brock, J.W., Needham,L.L., 2000. Human exposure estimates for phthalates. Environmental HealthPerspectives 108 (10), A440–A442.
Lehmann, K.P., Phillips, S., Sar, M., Foster, P.M., Gaido, K.W., 2004. Dose-dependentalterations in gene expression and testosterone synthesis in the fetal testes ofmale rats exposed to di (n-butyl) phthalate. Toxicological Sciences 81 (1), 60–68.
Liu, K., Lehmann, K.P., Sar, M., Young, S.S., Gaido, K.W., 2005. Gene expressionprofiling following in utero exposure to phthalate esters reveals new genetargets in the etiology of testicular dysgenesis. Biology of Reproduction 73 (1),180–192.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression using real-time quantitative PCR and the 2-DDCt method. Methods 25, 402–408.
Mylchreest, E., Wallace, D.G., Cattley, R.C., Foster, P.M., 2000. Dose-dependentalterations in androgen-regulated male reproductive development in ratsexposed to Di(n-butyl) phthalate during late gestation. Toxicological Sciences55 (1), 143–151.
Nativelle, C., Picard, K., Valentin, I., Lhuguenot, J.C., Chagnon, M.C., 1999. Metabolismof n-butyl benzyl phthalate in the female Wistar rat. Identification of newmetabolites 1. Food and Chemical Toxicology 37 (8), 905–917.
Oishi, S., 1990. Effects of phthalic acid esters on mitochondrial functions in the rat.Archives of Toxicology 63, 143–147.
Parks, L.G., Ostby, J.S., Lambright, C.R., Abbott, B.D., Klinefelter, G.R., Barlow, N.J.,Gray, L.E.Jr., 2000. The plasticizer diethylhexyl phthalate induces malformationsby decreasing fetal testosterone synthesis during sexual differentiation in themale rat. Toxicological Sciences 58 (2), 339–349.
Shultz, V.D., Phillips, S., Sar, M., Foster, P.M., Gaido, K.W., 2001. Altered gene profilesin fetal rat testes after in utero exposure to di(n-butyl) phthalate. ToxicologicalSciences 64 (2), 233–242.
Silva, M.J., Barr, D.B., Reidy, J.A., Malek, N.A., Hodge, C.C., Caudill, S.P., Brock, J.W.,Needham, L.L., Calafat, A.M., 2004. Urinary levels of seven phthalate metabolitesin the US population from the National Health and Nutrition ExaminationSurvey (NHANES) 1999–2000. Environmental Health Perspectives 112 (3), 331–338.
Wang, X., Walsh, L.P., Stocco, D.M., 1999. The role of arachidonic acid on LH-stimulated steroidogenesis and steroidogenic acute regulatory proteinaccumulation in MA-10 mouse Leydig tumor cells. Endocrine 10 (1), 7–12.
Wormuth, M., Scheringer, M., Vollenweider, M., Hungerbühler, K., 2006. What arethe sources of exposure to eight frequently used phthalic acid esters inEuropeans? Risk Analysis 26 (3), 803–824.




















