
Classification and status of wild populations of parrots
310
Transcript of Classification and status of wild populations of parrots


Manual of Parrot Behavior

Manual of Parrot Behavior
Andrew U. Luescher, Editor

Andrew U. Luescher, DVM, PhD, is Director of theAnimal Behavior Clinic at Purdue University. Heestablished the Animal Behavior Clinic in 1997 and iscertified as an applied animal behaviorist by theAnimal Behavior Society and is a diplomate of theAmerican College of Veterinary Behaviorists. He hasbeen treating animals with behavioral problems formore than 20 years.
© 2006 Blackwell PublishingAll rights reserved
Blackwell Publishing Professional2121 State Avenue, Ames, Iowa 50014, USA
Orders: 1-800-862-6657Office: 1-515-292-0140Fax: 1-515-292-3348Web site: www.blackwellprofessional.com
Blackwell Publishing Ltd9600 Garsington Road, Oxford OX4 2DQ, UKTel.: +44 (0)1865 776868
Blackwell Publishing Asia550 Swanston Street, Carlton, Victoria 3053, AustraliaTel.: +61 (0)3 8359 1011
Authorization to photocopy items for internal or per-sonal use, or the internal or personal use of specificclients, is granted by Blackwell Publishing, providedthat the base fee of $.10 per copy is paid directly to theCopyright Clearance Center, 222 Rosewood Drive,Danvers, MA 01923. For those organizations that havebeen granted a photocopy license by CCC, a separatesystem of payments has been arranged. The fee codefor users of the Transactional Reporting Service is ISBN-13: 978-0-8138-2749-0; ISBN-10: 0-8138-2749-3/2006 $.10.
Printed on acid-free paper in the United States ofAmerica
First edition, 2006
Library of Congress Cataloging-in-Publication Data
Manual of parrot behavior / edited by Andrew U.Luescher.— 1st ed.
p. cm.Includes bibliographical references and index.ISBN-13: 978-0-8138-2749-0 (alk. paper)ISBN-10: 0-8138-2749-3 (alk. paper)1. Parrots—Behavior. I. Luescher, Andrew U.
SF473.P3.M27 2006636.6�865—dc22
2005028547The last digit is the print number: 9 8 7 6 5 4 3 2 1

Contents
Contributors vii
Preface ix
01 Classification and the Status of Wild Populations of Parrots 3Dominique G. Homberger
02 Behavior of Wild Amazona and Rhynchopsitta Parrots, with Comparative Insights 13from Other PsittacinesErnesto C. Enkerlin-Hoeflich, Noel F.R. Snyder, and James W. Wiley
03 Parrot Conservation, Trade, and Reintroduction 27Charles A. Munn
04 Sensory Capacities of Parrots 33Jennifer Graham, Timothy F. Wright, Robert J. Dooling, and Ruediger Korbel
05 Social Behavior of Psittacine Birds 43Lynne M. Seibert
06 Captive Parrot Nutrition: Interactions with Anatomy, Physiology, and Behavior 49Kevin David Matson and Elizabeth A. Koutsos
07 Comfort Behavior and Sleep 59Laurie Bergman and Ulrike S. Reinisch
08 Parrot Reproductive Behavior, or Who Associates, Who Mates, and Who Cares? 63Tracey R. Spoon
09 Nest Box Preferences 79Scott George Martin and April Romagnano
10 Hand-Rearing: Behavioral Impacts and Implications for Captive Parrot Welfare 83Rebecca Fox
11 Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 93Phoebe Greene Linden with Andrew U. Luescher
12 Handler Attitude and Chick Development 113Brenda Cramton
v

13 Grey Parrot Cognition and Communication 133Irene M. Pepperberg
14 Behavior Analysis and Parrot Learning 147S.G. Friedman, Steve Martin, and Bobbi Brinker
15 Behavior Classes in the Veterinary Hospital: Preventing Problems Before They Start 165Kenneth R. Welle
16 Clinical Evaluation of Psittacine Behavioral Disorders 175Kenneth R. Welle and Liz Wilson
17 Diagnostic Workup of Suspected Behavioral Problems 195Susan E. Orosz
18 Aggressive Behavior in Pet Birds 211Kenneth R. Welle and Andrew U. Luescher
19 Parrot Vocalization 219Laurie Bergman and Ulrike S. Reinisch
20 Parrots and Fear 225Liz Wilson and Andrew U. Luescher
21 Problem Sexual Behaviors of Companion Parrots 233Fern Van Sant
Color Plates
22 Mate Trauma 247April Romagnano
23 Feather-Picking Disorder in Pet Birds 255Lynne M. Seibert
24 Psittacine Behavioral Pharmacotherapy 267Kenneth M. Martin
25 Behavior of Captive Psittacids in the Breeding Aviary 281G. Heather Wilson
26 Housing and Management Considerations for Problem Prevention 291Andrew U. Luescher and Liz Wilson
27 Captive Parrot Welfare 301Cheryl Meehan and Joy Mench
Index 319
vi Contents

Contributors
vii
Numbers in brackets refer to chapters
Laurie Bergman, VMD, DACVB [7,19]Co-ordinator Clinical Animal Behavior ServiceUniversity of California Veterinary Medical
Center, San Diego
Bobbi Brinker [14]Parrottalk.com
Brenda Cramton, MS, JD [12]Arroyo Veterinary HospitalDenair, CA
Robert J. Dooling, PhD [4]Department of PsychologyUniversity of Maryland, College Park
Ernesto C. Enkerlin-Hoeflich [2]www.conanp.gob.mx/
Rebecca Fox [10]Department of Animal ScienceUniversity of California, Davis
S.G. Friedman, PhD [14]Department of PsychologyUtah State University, Logan
Jennifer Graham, DVM, Diplomate, ABVP,Avian [4]
Affiliate Assistant Professor, Department ofComparative Medicine,School of Medicine,University of Washington
Veterinary Specialty Center of Seattle,Lynnwood, WA
www.seattleveterinaryspecialists.com
Dominique G. Homberger [1]Professor of Zoology, Dept. of Biological
SciencesLouisiana State University, Baton Rouge
Ruediger Korbel, Prof. Dr. med. vet., Dr. med.vet. habil. [4]
Director Institute of Avian Diseases, Ludwig-Maximilians-University Munich
Cert. Spec. Avian Medicine, Cert. Spec. Vet.Ophthalmol., Dipl. ECAMS. Univ.–Institutfuer Gefluegelkrankheiten
Elizabeth A. Koutsos [6]Department of Animal ScienceUniversity of Missouri, Saint Louis
Phoebe Greene Linden, MA [11]Santa Barbara Bird FarmSanta Barbara, CA
Andrew U. Luescher, DVM, PhD, DACVB[Editor, 11,18,20,26]
Director of the Animal Behavior ClinicPurdue University, West Lafayette, IN
Kenneth M. Martin, DVM [24]Veterinary Behavior ConsultationsNew Orleans, LA
Scott George Martin, MS, DVM [9]Animal Health ClinicJupiter, FL
Steve Martin [14]Natural Encounters, Inc.Winter Haven, FL
Kevin David Matson [6]Department of BiologyUniversity of Missouri, St. Louis
Cheryl Meehan, PhD [27]Associate Director, Center for Animal WelfareUniversity of California, Davis

Joy Mench [27]Professor and Director of the Center for Animal
Welfare, Department of Animal ScienceUniversity of California, Davis
Charles A. Munn, PhD [3]Chairman of the Board, Tropical NatureArlington, VAwww.tropicalnature.org
Susan E. Orosz, PhD, DVM, Diplomate,ABVP, Avian, and Diplomate, ECAMS [17]
Perrysburg Animal Care, Perrysburg, OhioAdjunct Professor, The University of Tennessee,
College of Veterinary Medicine, KnoxvilleConsultant, Lafeber Company, Cornell, Illinois
Irene M. Pepperberg [13]Research Associate Professor, Dept. of
Psychology, Brandeis UniversityResearch Scientist, MIT School of Architecture
and Planning
Ulrike S. Reinisch, DVM [7,19]Resident–Clinical Animal Behavior ServiceUniversity of California, Davis
April Romagnano, PhD, DVM, ABVP (AvianPractice) [9,22]
Animal Health ClinicJupiter, FL
Lynne M. Seibert, DVM, MS, PhD, DiplACVB [5,23]
Veterinary Specialty Center of SeattleLynnwood, WA
Noel F.R. Snyder [2]Portal, AZ
Tracey R. Spoon [8]Department of BiologyUniversity of Massachusetts, Boston
Fern Van Sant, DVM [21]For the BirdsSan Jose, CA
Kenneth R. Welle, DVM, Diplomate, ABVP,Avian [15, 16, 18]
All Creatures Animal HospitalAdjunct Assistant Professor, University of
Illinois College of Veterinary MedicineUrbana, IL
James W. Wiley [2]USGS-BRDMaryland Cooperative Fish and Wildlife
Research Unit, Princess Anne, MD
G. Heather Wilson, DVM, Diplomate, ABVP,Avian [25]
Assistant Professor Exotic Animal, Wildlife, andZoological Medicine, Department of SmallAnimal Medicine, College of VeterinaryMedicine
University of Georgia, Athens
Liz Wilson, CVT [16,20,26]Parrot Behavior ConsultantLevittown, PA
Timothy F. Wright, PhD [4]Genetics Lab, National Museum of Natural
History, Smithsonian Institution, SmithsonianNational Zoo
Washington, DC
viii Contributors

Many have forgotten this truth, but you must not forget it. You remain responsible, forever, forwhat you have tamed.
—Antoine de Saint-Exupery
Preface
ix
The untamed beauty of parrots has fascinatedhumans for centuries and keeps us in its spell tothe present time. Parrots are beautiful, they canfly, they are different from us, they are intelligent,and they remain mysterious. However, our rela-tionship with parrots has changed greatly overtime. Once considered a plentiful natural resourceworth exploiting, we now make great efforts toprotect their dwindling natural populations. At thesame time we have come a long way in how wekeep parrots in our homes. They no longer are butbrilliant exhibition pieces chained to a T-stand buthave become members of our families whose sen-sitivities, cognitive abilities, and emotions werespect and try to understand.
Yet parrot-keeping is a challenging endeavor.We admire their wildness, yet we bring them intoa very unnatural captive environment for whichthey have not evolved. We admire their flight, yetin most cases where we keep parrots as pets weneed to clip their wings. We like them becausethey are social creatures, yet we frequently keepthem as solitary birds so they will redirect theiraffection toward us, and in most cases we leavethem alone for extended periods of time. We rec-ognize their intelligence, yet maintain them in avery restricted and confining environment.
No wonder behavior problems in parrots areplentiful and the numbers of abandoned parrotsending up in sanctuaries is increasing. Stories ofparrots relegated to small cages in the basement,
neglected, covered with a towel to keep themquiet, are much too common. Even normal parrotbehavior such as vocalization, chewing, and beingmessy does not fit well with people’s lifestylesand can result in a broken human-animal bond.This book is written by authors who understandand love parrots in order to help foster a mutual-ly beneficial and enjoyable relationship betweenparrots and their humans. We hope it can set upnew parrot-human relationships for success andrekindle the joy that should be inherent in suchrelationships in cases where it has been lost. Weintend to promote a deepened understanding andresponsible attitude toward parrots in the wild aswell as in captivity. We hope this will contributeto the welfare of parrots and help develop arespect for and appreciation of these fascinatingbeings.
Although scientific interest in parrot behavioris growing, knowledge in this area is still limited.This is especially true for behavior problems ofpet birds and their treatment. The information inthis book is based on scientific principles andavailable publications but, where specific andproven information is not available, may reflectthe opinion and the personal experience of theauthors. Therefore, there may be some degree ofcontradiction or difference in interpretationbetween chapters. This inconsistency was inten-tionally maintained to offer the reader differentperspectives.

Manual of Parrot Behavior

THE ORDER PSITTACIFORMES AND ITSRELATIONSHIPS WITHIN THE CLASSAVES
The roughly 350 species in about 74 genera ofparrots and cockatoos (Forshaw 1989; Collar1997; Rowley 1997; Juniper & Parr 1998) aregrouped within the Psittaciformes, one of themost distinctive and largest of the 28 avian orders(Brooke & Birkhead 1991). Parrot and cockatoospecies are usually easily recognized as psittaci-form (or “psittacine”) birds because of theircurved beaks, in which the tip of the maxilla proj-ects beyond the shorter mandible, and their zygo-dactylous feet, in which the second and third toespoint forward and oppose the first and fourthtoes, which point caudally. Other characteristicsinclude a usually colorful plumage; a very largebrain; curiosity, lifelong capacity for learning,and adaptability to changing environmental con-ditions; distinctive vocalizations; a feeding ecolo-gy as seed predators; versatile feeding mecha-nisms; a complex social behavior; lifelong pairbonding; nesting in cavities; white eggshells; andnidicolous young.
In the past, there have been some attempts atidentifying the avian orders that are most closelyrelated to the Psittaciformes by looking for com-mon features, but it has become clear that anysuch commonalities reflect traits that haveevolved in adaptation to similar environmentalconditions and not traits that have been retainedfrom a common ancestor. Furthermore, most ofthe common features at the ordinal level resembleone another only superficially and are easily rec-ognized as having evolved independently in vari-
ous avian orders. For example, in the curved beakof owls (Strigiformes) and raptors (Falconi-formes), the mandible points straight forward, andthe hooked maxilla serves to get a grip whengrabbing or tearing apart prey. In the zygodactylfeet of woodpeckers (Piciformes) and cuckoos(Cuculiformes), the limb musculature differsfrom that of Psittaciformes, and the scaly skin dif-fers in the shape and number of the scales. Thesedifferences indicate that the zygodactyl feetreflect an adaptation to an arboricole lifestyle,which has evolved separately in the ancestors ofeach order, rather than one that has evolved in acommon ancestor of all three orders.
THE EVOLUTIONARY ORIGIN OFPSITTACIFORMES
The evolutionary origin of the Psittaciformes canbe reconstructed from a combination of function-al morphological, ecological, phylogenetic, bio-geographical, geological, and paleoecologicaldata (Cracraft 2001; Homberger 1991, 2003). Thezygodactylous feet that are especially adept atclimbing tree trunks and the predominant nestingin tree cavities suggest that the Psittaciformesoriginated as forest birds. The white color of theeggshells indicates that the ancestral species incu-bated their eggs in cavities (probably of trees),where they would not need camouflaging colorpattern to escape the attention of predators.
The functional morphology of their feedingapparatus provides additional support for apsittaciform origin from an ancestor that wasadapted to living in forests (Homberger 2003).The quadratomandibular, or jaw, joint is uniquely
3
1Classification and Status of Wild Populations of Parrots
Dominique G. Homberger

shaped to allow lateral movements of the lowermandible relative to the upper maxilla. However,parrots and most cockatoos, such as the Whiteand Pink Cockatoos (Cacatua spp., Eolophusroseicapillus, Lophochroa leadbeateri, Plictolo-phus spp.), the Cockatiel (Nymphicus hollandi-cus), the Yellow-tailed and White-tailed BlackCockatoos (Calyptorhynchus [Zanda]), and thePalm Cockatoo (Probosciger aterrimus), use thiscapacity only during bouts of bill honing and forminor adjustments when positioning food itemsbetween their mandible to bite into them. It isunlikely, therefore, that the psittaciform jaw artic-ulation was evolved in conjunction with the billmovements observed in these species. It has longbeen suspected that it was a feature that originat-ed in a psittaciform ancestor as part of a feedingbehavior that differed from that which is commonamong extant parrots (Homberger 1981).
In contrast, the lateral deflection of the lowermandible is an integral part of the feeding mech-anism in most Red-tailed Black Cockatoos(Calyptorhynchus banksii subspecies), the GlossyBlack Cockatoo (C. lathami), and the Gang-gangCockatoo (Callocephalon fimbriatum) (Homber-ger 2001, 2003). They align one of the paired,projecting corners of their V-shaped lower bill tipwith their upper bill tip. They do this in order touse their beak as pincer-like pliers to tear apartwoody branches to extricate wood-boring orgallicole insect larvae or to break apart woody-fibrous capsules to extract seeds (Homberger2001, 2003). These species also possess a bonysuborbital arch that juts out on the sides of theirskull and is firmly buttressed against the postor-bital and zygomatic processes of the cranium. Thejaw muscles that attach to this suborbital archassume an orientation that emphasizes transverse-ly directed force components, which are instru-mental for the lateral deflections of the mandibleduring feeding in these species. In this “calyp-torhynchid” feeding apparatus, the shapes of thejaw joint, skull, and bill are structurally and func-tionally integrated with the feeding mechanism totear apart food sources that are made of fibrouswood, which are prevalent in a wooded or forestedenvironment (Homberger 2003). The tight func-tional integration of the features of the calyp-torhynchid feeding apparatus also indicates thatthey are part of an ancestral condition for Psittaci-formes. The calyptorhynchid feeding apparatus
may have originated in a psittaciform ancestorfirst to extract wood-boring or gallicole insect lar-vae and subsequently been applied with few, ifany, modifications, to extract seeds from fibrous-woody fruits.
In the “psittacid” feeding apparatus of parrotsand most cockatoos (except the Red-tailed BlackCockatoos, Calyptorhynchus lathami and mostC. banksii subspecies, and the Gang-gangCockatoo, Callocephalon fimbriatum), in con-trast, the structure and function of the jaw jointdoes not fit the bill shape and feeding behavior.The psittacid feeding apparatus relies on special-ized surface structures, such as the transversestep and filing ridges on the inside of the upperbill tip, to provide grip for seeds that are cut openwith the cutting edge of the lower mandible(Homberger 1980a, 1980b, 2003). Psittaciformswith a psittacid bill (except the Pesquet’s Parrot,Psittrichas fulgidus) remove the shells of allseeds before swallowing them, and they do sowith a stereotypical seed-shelling mechanismthat does not require lateral movement of themandible. During this seed-shelling procedure,the tip of the tongue places and holds a seedagainst the corrugated upper bill tip and its trans-verse step, while the cutting edge of the mandiblecuts open the seed-shell. The bony suborbitalarch is generally absent so that the transversecomponent of the jaw muscles is much reducedin favor of the longitudinal and vertical forcecomponents. If a suborbital arch is present, as inmany South American species, it is less massiveand fused only with the postorbital process of thecranium (see Smith 1975). This functional disso-ciation of the various structural and functionalfeatures indicates that the shapes of the jaw joint,skull, and bill of parrots and cockatoos with apsittacid feeding apparatus have changed underthe influence of a variety of selective regimesarising from environmental conditions that differfrom those to which the psittaciform ancestorwas adapted.
The most significant selective advantage of thepsittacid feeding apparatus over the calyp-torhynchid feeding apparatus is that the formercan use both sides of the jaw musculature simul-taneously to maximize the bite force of the man-dible. This selective advantage, however, can beutilized only in environments in which plants withseeds enclosed in fruits that are not woody-
4 Manual of Parrot Behavior

fibrous predominate (Homberger 2003). Most ofthese fruits have a sclerotic endocarp (i.e.,“stone”) that can be split, or cracked open, byapplying a focused pressure, such as by the cut-ting edge of the mandible, onto their preformedweak points or sutures that facilitate the germina-tion of the seeds. The selective advantage of apsittacid feeding apparatus appears to be consid-erable because it has evolved multiple times inseparate lineages of parrots and cockatoos,including among them some of the populationsand subspecies of Red-tailed Black Cockatoos(Calyptorhynchus banksii). This convergent evo-lution of the psittacid feeding apparatus is madeevident by the great variability of the individualcomponents and features, such as the pattern andconfiguration of the filing ridges and corneouspalate, the shape and expression of the transversestep, the shape of the cutting edge of themandible, and the configuration and degree of thereduction of the suborbital arch (Homberger1980a, 1980b, 2003).
The large brain of the Psittaciformes earnedthem the epithet “avian primates.” As in primates,it is correlated with curiosity and exploratorybehaviors and a lifelong capacity for learning(e.g., Mettke-Hofmann et al. 2002; Pepperberg2002). This high degree of encephalization sup-ports the hypothesis that the Psittaciformes origi-nated from ancestors that were feeding on station-ary food items that were hidden from sight (i.e.,wood-boring or gallicole insect larvae and seedswithin fruits) and, therefore, need to be locatedthrough indirect evidence and learning fromexperienced individuals. These arboreal fooditems further support the hypothesis that Psit-taciformes originated in a forested environment.
Psittaciformess are concentrated in the conti-nents and islands of the Southern Hemispherewith only limited expansions into the adjacentnorthern regions. Contrary to general impres-sions, Psittaciformes are not restricted to tropicalregions, as several species occur in the colderregions of China, New Zealand, New Guinea,Tasmania, and South America. Such a distribu-tion pattern can be understood only on the basisof past geological events. Biogeography has beensuggestive of a psittaciform origin in theSouthern Hemisphere (Boetticher 1959; Forshaw1989) even before geological data could demon-strate that the southern continents were formed
through the disintegration of the Mesozoic south-ern continent called Gondwana and their subse-quent migration northward toward the equator(Frakes & Vickers-Rich 1991; Schodde &Tidemann 1986; Stevens 1991).
Gondwana’s climate in the Cretaceous wasgenerally temperate to subtropical, and Gond-wana itself was covered mostly with evergreenmesic forest and rain forest (White 1990). As thecontinents moved northwards, they tended tobecome more arid with the rising temperatures(Frakes & Vickers-Rich 1991; Stevens 1991;White 1994). The original plant communities thatincluded southern gymnosperms (e.g., Arau-caria), Casuarinas, Proteaceae (e.g., Banksia,Protea, Grevillea), Myrtaceae (e.g., ancestors ofEucalyptus), Podocarpaceae, Nothofagaceae(e.g., Southern beeches—Nothofagus), and soforth, adapted to the changing conditions, werereplaced by other plant communities, or retreatedto refugia in which the original Gondwanan con-ditions were retained or changed but little. SuchGondwanan refugia are found today in Australiaalong its eastern coast, the southeastern andsouthwestern corners, and in Tasmania; in NewZealand, New Caledonia, and Fiji; in the centralhighlands of New Guinea; in the Drakensbergs ofeastern South Africa; and in the Valdivian andPatagonian rain forests along the eastern coast ofsouthern South America and the cooler Atlanticrain forests in Southern Brazil.
In Australia, several of these seed plants (e.g.,Casuarinas, Proteaceae, Myrtaceae) occur pre-dominantly in the Gondwanan refugia (Schodde& Tidemann 1986) and bear complex inflores-cences that mature into multi-seeded, fibrous-woody infructescences, called cones, cobs, orcapsules. Several species have also becomeserotinous (i.e., they retain their mature fibrous-woody fruits for several years in their canopyinstead of shedding their mature seeds), presum-ably in adaptation to their fire-prone environment(Homberger 2003). That the psittaciform speciesthat possess a calyptorhynchid feeding apparatus(most Red-tailed Black Cockatoos, Calyptorhyn-chus banksii subspecies; the Glossy BlackCockatoo, C. lathami; and the Gang-gang Cock-atoo, Callocephalon fimbriatum) not only occurin these refugia but also have a feeding apparatusthat is specifically adapted to exploiting theseplants supports the hypothesis that the calyp-
1 / Classification and Status of Wild Populations of Parrots 5

torhynchid feeding apparatus is the ancestral con-dition for Psittaciformes.
In the other southern continents and islands,the Gondwanan refugia are dominated byGondwanan plants whose seeds are enclosed inthinner seed-shells (e.g., Araucariaceae, Notho-fagaceae, some Podocarpaceae) or sclerotic endo-carps with preformed weak points and sutures(e.g., some Podocarpaceae). The psittaciformspecies that feed on these seeds and are restrictedto Gondwanan refugia can be surmised to haveevolved their psittacid feeding apparatus alreadyin adaptation to these plants before the breakup ofGondwana and were able to retain it because theirenvironment changed little, if at all. This is prob-ably the situation, for example, of the Austral andSlender-billed Conures (Enicognathus ferrug-ineus and E. leptorhynchus) in southern SouthAmerica; the Vinaceous Amazon (Amazonavinacea) in southern Brazil; the Cape Parrot(Poicephalus r. robustus) in southeastern Africa;and the non-cacatuid psittaciforms with a psit-tacid feeding apparatus in the Australo-Pacificregion.
The greatest diversity of Psittaciformes at thefamilial and subfamilial levels is found in theAustralo-Pacific region (see Figure 1.1). Thisindicates that this part of Gondwana may havecontained the greatest psittaciform diversity evenbefore its separation from the remainder ofGondwana and further breakup into what isknown today as Australia, New Guinea, NewZealand, New Caledonia, and Fiji.
THE SUBDIVISION ANDCLASSIFICATION OF THEPSITTACIFORMES
The very ease with which psittaciforms can beidentified as such is compensated by the difficul-ties that are encountered trying to subdivide thislarge order into smaller, hierarchically arrangedtaxonomic units that are united by common char-acteristics (i.e., families, subfamilies, tribes, gen-era). Such a classification creates order within themultitude of species, which is needed for scientif-ic research (e.g., systematics, comparative mor-phology, evolutionary biology) and applied biolo-gy (e.g., evaluation of susceptibility to certaindiseases, choice of foster parents for the manage-ment of endangered species). However, it must bekept in mind that every classification is only a
hypothesis that needs to be tested continuously asnew data emerge and earlier interpretations arere-evaluated in light of new observations.Changes in the nomenclature of taxa and in thehierarchical levels of taxonomic subdivisions are,hence, reflective of intense scientific activity butare not an end in themselves.
Numerous classifications have been proposedover the last 200 years, but all have faced consid-erable difficulties. One of the underlying reasonsfor this situation is that the Psittaciformes repre-sent a very old group that had to adapt to numer-ous environmental changes in the course of itslong history dating back to the early Tertiary (ca.60 million years ago). Because similar environ-mental changes (e.g., aridification, tropicaliza-tion, colonization of volcanic islands, etc.) haveoccurred in different regions, many derived fea-tures have been acquired independently and con-vergently by different psittaciform lineages inadaptation to these new environments. This preva-lence of convergent (i.e., homoplastic) featuresamong the Psittaciformes as a group has ham-pered earlier efforts in classifying this avianorder, mainly because many convergent and othernon-homologous features have been misidenti-fied as homologous ones that would indicate evo-lutionary relationships (for discussions, seeHomberger 1980a, 1991; Güntert 1981).
The distinction between homologous and con-vergent features is one of the most challengingtasks for evolutionary biologists, because the firststep in this procedure requires the analysis of boththe structure and function of the features, as wellas their biological role in the natural environment.Two examples will illustrate the basic approach.The first example will use the bony suborbitalarch to demonstrate the possible pitfalls in ana-lyzing features in isolation. A recent functional-anatomical analysis of the bony suborbital arch incockatoos revealed that it is a component of thefeeding apparatus and as such cannot be used as afeature in isolation. It also revealed that its mostcomplete configuration is intimately connectedwith lateral mandibular movements during feed-ing in Black Cockatoos that possess a calyptor-hynchid feeding apparatus. Various configu-rations of less complete suborbital arches indifferent psittaciform lineages that possess a psit-tacid feeding apparatus can, therefore, be inter-preted as derived remnants of the ancestral condi-
6 Manual of Parrot Behavior

7
Figure 1.1. Phylogram of psit-taciform genera based conserva-tively on established criteria.

tion that is still present in psittaciforms with acalyptorhynchid feeding apparatus. This reinter-pretation of the evolutionary history of the bonysuborbital arch is contrary to the original inter-pretation by Hofer (1950, 1953) and Zusi (1993),both of whom did not have access to observationsof psittaciforms in their natural environment.
The second example will use the oral plate ofthe upper rhamphotheca (i.e., corneous sheath ofthe maxilla) to demonstrate that a particular struc-ture may be composed of several features thatprovide different insights for the reconstruction ofthe evolutionary history of the Psittaciformes.The oral plate of the upper rhamphotheca consistsof three parts: The inside of the upper bill tip, thetransverse step, and the corneous palate. Theinside of the upper bill tip of parrots that possessa psittacid feeding apparatus is corrugated by fil-ing ridges. These filing ridges, however, arearranged in patterns and are formed by the under-lying soft tissues in a manner that is highly vari-able among, but generally characteristic of,species. The inside of the upper bill tip of psittaci-forms that possess a calyptorhynchid feeding ap-paratus is smooth and lacks any surface structure(Homberger 2003). The evolutionary transitionfrom the ancestral to the derived condition of theinside of the upper bill tip is modeled by the var-ious populations and subspecies of the Red-tailedBlack Cockatoo (Calyptorhynchus banksii) and isclearly correlated with the derived seed-shellingbehavior of psittaciforms with a psittacid feedingapparatus (Homberger 2003). In contrast, the sur-face structure of the corneous palate, which is thefeature with the greatest diagnostic value for theidentification of genera in psittaciforms, does nothave any functional significance (Homberger1980a).
Mosaic evolution, that is, the presence of prim-itive and derived characters in a single species asa result of asynchronous evolutionary changes,has been another source of difficulties for theclassification of the Psittaciformes. Because of it,a phylogeny that is based on a particular set offeatures, such as the feeding apparatus, may notsimply correspond to another phylogeny that isbased on a different set of features. As a conse-quence, the evolutionary history of each lineageand species needs to be reconstructed by careful-ly analyzing, weighing, and integrating a varietyof data and observations. A simplified example
taken from the Cacatuidae may illustrate such acase. Among birds in general, a large body size isa derived character, because flight has a muchgreater safety margin in small birds than in largerbirds and, therefore, has probably originated insmall avian ancestors whose flight apparatus maynot have been perfected yet (Homberger & deSilva 2000; Homberger 2003). According to thiscriterion, the Cockatiel (Nymphicus hollandicus)could be considered the most ancestral cockatoo.This interpretation could be supported by its darkplumage color and pattern, which are similar tothose of the Black Cockatoos (Calyptorhynchusspp.) and clearly more ancestral than the plumagecolors and patterns of the White and PinkCockatoos. But the Cockatiel’s psittacid feedingapparatus and its ecology in Australia’s more aridwoodlands indicate that it has also acquiredderived characters in adaptation to the aridifica-tion of Australia. In contrast, the Red-tailed BlackCockatoos and the Gang-gang Cockatoo are char-acterized by ancestral plumage colors and pat-terns and by the ancestral calyptorhynchid feed-ing apparatus. At the same time, the Red-tailedBlack Cockatoos are among the larger cockatoos,whose body size may have evolved in conjunctionwith their more massive bills to handle their dietof large fibrous-woody fruits (Homberger 2003).
At this point in time, the best classification ofthe Psittaciformes may be one that is based on alarge number of features, whose biological andevolutionary significance has been analyzed andis well understood. Unfortunately, we are still farfrom this goal. The proposed classification (seeFigure 1.1) is presented as a pragmatic proposalthat combines simplicity and familiarity andavoids some of the errors of earlier classifications.
THE STATUS OF WILD POPULATIONSOF PARROTS
Over the millions of years since their origin in theearly Tertiary, many psittaciform species havesurvived and continued to adapt successfully tochanging environmental conditions, as we canconclude from their present geographical distri-bution and the number of existing species andindividuals. Other species have not been able todo so and have become extinct, as we know fromhistorical records or from fossils in regions, suchas Europe, in which psittaciforms have beenabsent in historical times. Rates of extinction are
8 Manual of Parrot Behavior

difficult to estimate from the fossil record,because fossilization is a rare event in any caseand especially so for organisms, such as the an-cestral and many other Psittaciformes, that arerelatively small and live in microorganism-richforest environments with their characteristicallyrapid degradation of organic materials. Neverthe-less, the large number of species that are knownto have existed at least until the more recent rashof extinctions testifies to the success and proba-bly net increase in number of species and individ-uals of the Psittaciformes over the course of theirevolutionary history.
Although extinctions of species are a normalpart of biological evolution, extinction must becounterbalanced by speciation, that is, the appear-ance of new species, if a taxon, or group ofspecies, as a whole is to survive. The appearanceof new species, however, is presently not occur-ring any longer, at least not naturally. This processnormally starts when a certain portion of a popu-lation becomes separated from the rest of the pop-ulation by the appearance of a geographical barri-er, such as a river having changed direction, anarea having been divided by the uplifting of amountain or the formation of a desert, or a num-ber of individuals having migrated permanentlyto an island. This separation, or isolation, pre-vents the exchange of genetic materials betweenthe separate populations and provides the condi-tions for the two populations to accumulate dis-tinctive mutations, undergo distinctive selectiveprocesses, and, thereby, acquire distinctive traitssimply by themselves or in adaptation to distinctenvironmental conditions.
The main reason for natural speciation not tobe initiated any longer is the accelerating andwell-documented shrinking of the natural habi-tats, so that psittaciform populations cannotexpand and subsequently be subdivided into non-interbreeding populations. However, the recentsuccessful establishment of self-sustaining parrotpopulations from aviary and transport escapees invarious urban and suburban places in regions thathad been devoid of natural populations ofpsittaciforms may be considered an experiment inhuman-induced speciation. As we can extrapolatefrom earlier such experiments in the late 18th andearly 19th centuries, during which Europeansongbirds (e.g., House Sparrows, Starlings, Chaf-finches, Blackbirds, European Goldfinches) were
transported to colonies in North America,Australia, and New Zealand by homesick Euro-pean emigrants, the successful psittaciform expa-triate populations in Germany, England, andNorth America may eventually become genetical-ly distinct from their source populations. Butmodifications of external features will take manygenerations to become noticeable, as they did inthe various domesticated psittaciforms, and thesechanges may reflect adaptations to the new envi-ronments or the lack of specific selection pres-sures (e.g., in cases of variable plumage colors),unless these mostly urban populations will berepeatedly swamped by new escapees and acci-dental releases. The possibility of such artificialspeciation events may be a consolation, but hard-ly a compensation for the current progressive lossof the amazing diversity of psittaciform species intheir natural environment.
There is no denying that the single-most threatto natural populations is the capture of individu-als for aviculture and the pet market. Captivebreeding of parrots by private individuals for con-servation purposes should be recognized as thesmoke screen that it is (Beissinger et al. 1991;Beissinger 2001; Snyder et al. 1997; Wright et al.2001). Only a single psittaciform species, thePuerto Rican Amazon (Amazona vittata) (Wilsonet al. 1994, Wunderle et al. 2003) has beenbrought back from the brink of extinction, whichwas made possible only through the lavish invest-ment of governmental funding. The success ofother governmental rescue programs for the Kaka(Nestor meridionalis) and Kakapo (Strigopshabroptilus) in New Zealand (Beggs & Wilson1991; Lloyd & Powlesland 1994) and the Orange-bellied Parrot (Neophema chrysogaster) inAustralia (Drechsler 1998) is still uncertain. Suchmassive financial investments for the rescue ofsingle species are beyond the possibilities of evenvery wealthy persons. Furthermore, althoughthere have been successful reintroductions of cap-tive individuals into the wild provided that thesecould be integrated with natural populations ofthe same species (Brightsmith et al. 2003), simplereleases of captive-bred psittaciforms into naturalenvironments, whose resources are characteristi-cally seasonal and unpredictable, have not beensuccessful (Snyder et al. 1994). The reason forthese difficulties may well be based in the evolu-tionary origin of the Psittaciformes with their spe-
1 / Classification and Status of Wild Populations of Parrots 9

cialized diet of wood-boring and gallicole insectlarvae, which could be detected only throughindirect evidence and through learning fromexperienced individuals.
ACKNOWLEDGMENTS
I thank Andrew Luescher for his kind invitation tocontribute to this volume and his excellent editor-ship. I also thank David Ray for designing thephylogram.
REFERENCES
Beggs, J.R., and P.R. Wilson. 1991. The kaka Nestormeridionalis, a New Zealand parrot endangered byintroduced wasps and mammals. Biol Conserv 56(1):23–38.
Beissinger, S.R. 2001. “Trade of live wild birds: Poten-tials, principles and practices of sustainable use.” InConservation of exploited species, ed. J.D.Reynolds, G.M. Mace, K.H. Redford, and J.G.Robinson, pp. 182–202. Cambridge: CambridgeUniversity Press.
Beissinger, S.R., N.F.R. Snyder, S.R. Derrickson, F.C.James, F.C., and S.M. Lanyon, S.M. 1991. Interna-tional trade in live exotic birds creates a vast move-ment that must be halted. Auk 108 (4):982–984.
Boetticher, H. von. 1959. Papageien. WittenbergLutherstadt, Germany: A. Ziemsen Verlag.
Brightsmith, D., J. Hilburn, A. del Campo, J. Boyd, M.Frisius, R. Frisius, F. Guille, and D. Janik. 2003.“Survival and reproduction of hand-raised scarletmacaws (Ara macao) in the wild.” In Abstracts of VIINeotropical Ornithological Congress, Termas dePuyehue, Chile: Program and book of abstracts, p.106. Chile: Neotropical Ornithological Society.
Brooke, M., and T. Birkhead. 1991. The Cambridgeencyclopedia of ornithology. Cambridge: Cam-bridge University Press.
Collar, N.J. 1997. “Family Psittacidae (parrots).” InHandbook of the birds of the world. Volume 4:Sandgrouse to cuckoos, ed. J. del Hoyo, A. Elliott,and J. Sargatal, pp. 280–477. Barcelona: LynxEdicions.
Cracraft, J. 2001. Avian evolution, Gondwana biogeog-raphy and the Cretaceous-Tertiary mass extinctionevent. Proc R Soc Lond B 268 (1466):459–469.
Drechsler, M. 1998. Spatial conservation managementof the orange-bellied parrot Neophema chryso-gaster. Biol Conserv 84 (3):283–292.
Forshaw, J.M. 1989. Parrots of the world, 3rd ed. Mel-bourne: Lansdowne Editions.
Frakes, L.A., and P. Vickers-Rich. 1991. “Palaeocli-matic setting and palaeogeographic links of Austra-lia in the Phanerozoic.” In Vertebrate palaeontology
of Australasia, ed. P. Vickers-Rich, J.M. Monghan,R.F. Baird, and T.H. Tich, pp. 111–146. Melbourne:Monash University Publications Committee.
Güntert, M. 1981. Morphologische Untersuchungenzur adaptiven Radiation des Verdauungstraktes beiPapageien (Psittaci). Zool Jahrb Anat 106:471–526.
Hofer, H. 1950. Zur Morphologie der Kiefermusku-latur der Vögel. Zool Jahrb Anat 70 (4):427–556.
Hofer, H. 1953. Die Kiefermuskulatur der Papageienals Evolutionsproblem. Biol Zentralbl 62 (5/6):225–232.
Homberger, D.G. 1980a. Funktionell morphologischeUntersuchungen zur Radiation der Ernährungs undTrinkmethoden der Papageien (Psittacidae). [Func-tional morphological studies on the radiation of thefeeding and drinking methods of the parrots]. BonnerZoologische Monographien, No. 13, pp. 192.
Homberger, D.G. 1980b. “Functional morphology andevolution of the feeding apparatus in parrots, withspecial reference to the Pesquet’s Parrot, Psittrichasfulgidus (lesson).” In Conservation of new worldparrots, ed. R.F. Pasquier, pp. 471–485. Interna-tional Council for Bird Preservation Technical PaperNo. 1. Washington, DC: Smithsonian InstitutionPress.
Homberger, D.G. 1981. Morphological foundations ofthe bill honing behavior in parrots (Psittacidae).Amer Zool 21 (4):1039.
Homberger, D.G. 1991. “The evolutionary history ofparrots and cockatoos: A model for evolution in theAustralasian avifauna.” Acta XX Congr Int Ornithol,pp. 398–403.
Homberger, D.G. 2001. “The case of the cockatoo bill,horse hoof, rhinoceros horn, whale baleen, andturkey beard: The integument as a model system to explore the concepts of homology and non-homology.” In Vertebrate functional morphology:Horizon of research in the 21st century, ed. H.M.Dutta and J.S. Datta Munshi, pp. 317–343. Enfield,NH: Science Publishers Inc.
Homberger, D.G. 2003. “The comparative biomechan-ics of a prey-predator relationship: The adaptivemorphologies of the feeding apparatus of Australianblack cockatoos and their foods as a basis for thereconstruction of the evolutionary history of thePsittaciformes.” In Vertebrate biomechanics andevolution, ed. V.L. Bels, J.-P. Gasc, and A. Casinos,pp. 203–228. Oxford: BIOS Scientific Publishers.
Homberger, D.G., and K.N. de Silva. 2000. Functionalmicroanatomy of the feather-bearing avian integu-ment: Implications for the evolution of birds andavian flight. Amer Zool 40 (4):553–574.
Juniper, T., and M. Parr. 1998. Parrots: A guide to par-rots of the world. New Haven: Yale University Press.
Lloyd, B.D., and R.G. Powlesland. 1994. The decline of
10 Manual of Parrot Behavior

the kakapo Strigops habroptilus and attempts at con-servation by translocation. Biol Conserv 69:75–85.
Mettke-Hofmann, C., H. Winkler, and B. Leisler. 2002.The significance of ecological factors for explo-ration and neophobia in parrots. Ethology 108(3):249–272.
Pepperberg, I.M. 2002. Alex studies: Cognitive andcommunicative abilities of grey parrots. Cambridge,MA: Harvard University Press.
Rowley, I. 1997. “Family Cacatuidae (cockatoos).” InHandbook of the birds of the world. Volume 4: Sand-grouse to cuckoos, ed. J. del Hoyo, A. Elliott, and J.Sargatal, pp. 246–279. Barcelona: Lynx Edicions.
Schodde, R., and S.C. Tidemann. 1986. Reader’sDigest complete book of Australian birds. Sydney:Reader’s Digest.
Smith, G.A. 1975. Systematics of parrots. Ibis117:18–68.
Snyder, N.F.R., S.R. Derrickson, S.R. Beissinger, J.W.Wiley, T.B. Smith, W.D. Toone, and B. Miller. 1997.Limitations of captive breeding: Reply to Gippolitiand Carpaneto. Conserv Biol 11 (3):808–810.
Snyder, N.F.R., S.E. Koenig, J. Koschmann, H.A. Sny-der, and T.B. Johnson. 1994. Thick-billed parrotreleases in Arizona. Condor 96:845–862.
Stevens, G.R. 1991. “Geological evolution and bioticlinks in the Mesozoic and Cenozoic of the southwestPacific.” Acta XX Congr Int Ornithol, pp. 361–382.
White, M.E. 1990. The flowering of Gondwana; The400 million year story of Australia’s plants. Prince-ton: Princeton University Press.
White, M.E. 1994. After the greening: The browning ofAustralia. Kenthurst, NSW: Kangaroo Press.
Wilson, M.H., C.B. Kepler, N.F.R. Snyder, S.R.Derrickson, F.J. Dein, J.W. Wiley, J.M. Wunderle,A.E. Lugo, D.L. Graham, and W.D. Toone. 1994.Puerto Rican parrots and potential limitations of themetapopulation approach to species conservation.Conserv Biol 8 (1):114–123.
Wright, T.F., C.A. Toft, E. Enkerlin-Hoeflich, J.Gonzalez-Elizondo, M. Albornoz, A. Rodríguez-Ferraro, F. Rojas-Suárez, V. Sanz, A. Trujillo, S.R.Beissinger, V. Berovides A., X. Gálvez A., A.T.Brice, K. Joyner, J. Eberhard, J. Gilardi, S.E. Koenig,S. Stoleson, P. Martuscelli, J.M. Meyers, K. Renton,A.M. Rodríguez, A.C. Sosa-Asanza, F. Vilella, andJ.W. Wiley. 2001. Nest poaching in Neotropical par-rots. Conserv Biol 15:710–720.
Wunderle, J.M., Jr., N.F. Snyder, S.R. Beissinger, J.M.Meyers, and J.W. Wiley. 2003. “Struggling out of thebottleneck: Puerto Rican parrot recovery from 1973to 2000.” In VII Neotropical Ornithological Con-gress, Termas de Puyehue—Chile, October 5–11,2003: Program and book of abstracts, p. 155. Chile:Neotropical Ornithological Society.
Zusi, R.L. 1993. “Pattern of diversity in the avianskull.” In The skull. Volume 2: Patterns of structuraland systematic diversity, ed. J. Hanken and B.K.Hall, pp. 391–437. Chicago: University of ChicagoPress.
1 / Classification and Status of Wild Populations of Parrots 11

Research on the behavioral characteristics ofselected wild psittacines may be important in es-tablishing management and conservation guide-lines for these species, both in the wild and incaptivity. An understanding of such behavioralcharacteristics may also have wider significancein aiding the interpretations of behavioral andnatural history parameters in other psittacinebirds. Prior to the 1970s, intensive biologicalstudies of wild Neotropical parrots were nearlynonexistent. This gap in ornithological knowl-edge is now being rapidly remedied with numer-ous species under investigation throughout Cen-tral and South America and the West Indies. Yetto date there have been few attempts to integratethe information from various studies into coher-ent frameworks of biological understanding. Inthis offering, we provide a number of preliminaryhypotheses about parrot behavior, based largelyon studies of a variety of species, mainly in thegenera Amazona and Rhynchopsitta. These hy-potheses appear to have wide explanatory power,yet need to be tested in additional genera andspecies before their validity can be consideredfirm.
Our basic approach is the comparative one,looking at features of behavior that vary amongspecies and attempting to correlate these differ-ences with underlying ecological imperativesfaced by the species under consideration. As rawmaterials for these comparisons, we rely heavily
on intensive studies of (1) the Puerto Rican Parrot(Amazona vittata) by Snyder et al. (1987); (2)various subspecies of the Cuban Parrot (Amazonaleucocephala) by Gnam (1991), Wiley (unpub-lished), and others; (3) the Hispaniolan Parrot(Amazona ventralis) by Wiley (unpublished); (4)the Jamaican Black-billed and Yellow-billedParrots (Amazona agilis and collaria) by Koenig(1999); (5) the St. Lucia Parrot (Amazona versi-color), the Imperial Parrot (Amazona imperialis),the Red-necked Parrots (Amazona arausiaca) ofthe Lesser Antilles by Snyder, Koenig, and manyothers (unpublished); and (6) three species ofamazons in northeastern Mexico by Enkerlin-Hoeflich (1995, unpublished)—the Red-crownedParrot (Amazona viridigenalis), the Yellow-headed Parrot (Amazona oratrix), and the Red-lored Parrot (Amazona autumnalis). Studies ofRhynchopsitta pachyrhyncha and terrisi havebeen primarily carried out by Enkerlin-Hoeflich(unpublished), Cruz-Nieto et al. (1998), Snyder etal. (1999), Lanning and Shiflett (1981, 1983), andLawson and Lanning (1982).
The particular behavioral features we considerhere are (1) the values of intraspecific sociality invarious species, (2) timing of nesting seasons, (3)site and pair fidelity, and nest reuse, (4) feedingbehavior and rates, (5) relationships of speciesconspicuousness and nest accessibility to exploi-tation in the pet trade, and (6) deficits in breedingeffort.
13
2Behavior of Wild Amazona and
Rhynchopsitta Parrots, withComparative Insights from
Other Psittacines
Ernesto C. Enkerlin-Hoeflich, Noel F.R. Snyder, and James W. Wiley

BACKGROUND
As generally understood, the genus Amazonaincludes 31 extant species limited largely to trop-ical areas of the Western Hemisphere. The variousspecies occur from Argentina and Chile norththrough virtually all of mainland South Americaand Central America to northern Mexico (Juniper& Parr 1998). Feral populations of several speciesalso exist in a number of southern cities of theUnited States, Mexico, and Puerto Rico(Enkerlin-Hoeflich & Hogan 1997; Mabb 1997).The West Indies have a particularly goodAmazona fauna with nine extant species aboutevenly divided between the Greater Antilles andLesser Antilles. Many Amazona species are nowendangered, and most all are declining, threat-ened mainly by bird trade and habitat changes,but also in some cases by hunting, introducedexotic species, and other factors (Snyder et al.2000). The various species of Amazona are farfrom monolithic in their behavior and ecology.Species vary in clutch and brood sizes, diets,feeding rates, degrees of sociality, reproductiveeffort, and reproductive success, to name just afew facets of interest.
The genus Rhynchopsitta has only two livingspecies, both distributed in the highlands ofnorthern Mexico in more or less “island-like” for-est habitats. The Thick-billed Parrot (R. pachy-rhyncha) is a tree-cavity nester inhabiting theSierra Madre Occidental in western Chihuahuaand eastern Sonora south through the mountainsof Durango, Sinaloa, Nayarit, and Jalisco toColima and Michoacan. It was also formerly aregular inhabitant of extreme southeastern Ari-zona and probably bred there until the early 20th
century, although no historical records of nestsexist for this region. The Maroon-fronted Parrot(R. terrisi) is a cliff-cavity nester that occurs inthe Sierra Madre Oriental of northeastern Mexicoin southeastern Coahuila, central western NuevoLeón, and southwestern Tamaulipas (Juniper &Parr 1998). Like many of the Amazona, bothextant species of Rhynchopsitta are globallythreatened. In addition, on the basis of fossils,Rea (1997) recently described a third species ofRhynchopsitta (R. phillipsi), now extinct, that wasapparently sympatric with both pachyrhyncha andterrisi in the Sierra Madre Oriental in the latePleistocene. The former sympatry of pachyrhyn-cha and terrisi makes it quite clear that these two
parrots are distinct species and not simply racesof a single species, as they were considered byForshaw (1990).
VALUES OF SOCIALITY AMONGAMAZONA AND RHYNCHOPSITTAPARROTS
In general, Amazona parrots tend to be highlysocial in foraging and roosting habits and some-what social in nesting habits, but some conspicu-ous differences exist among species in thesetendencies. Four species that stand out in their dis-inclination to travel in groups larger than familygroups are the large amazons of the LesserAntilles—the St. Lucia Parrot, the St. VincentParrot (A. guildingii), the Imperial Parrot, and theRed-necked Parrot. Although these species some-times assemble in groups larger than family unitsat roosts and at rich food sources, they normallynest in relatively dispersed arrays and travel onlyas singles, pairs, or small family groups in movingfrom nests or roosts to foraging areas. In contrast,other amazons, including most of those in theGreater Antilles and Mexico, show clear tenden-cies toward clumped nesting, often travel in muchlarger groups, and typically feed in large aggrega-tions. Low sociality in the Lesser Antillean speciesis also reflected in the fact that male and femaleadults often separate in foraging activities in thebreeding season and often feed their young inde-pendently. Such independent provisioning of nestsis virtually unknown in amazons of the GreaterAntilles and the mainland Neotropics.
Are there any obvious ecological correlates toexplain the relatively low sociality of the largeamazons of the Lesser Antilles? One promisingpossibility is the fact that these species, essential-ly alone among species in the genus, live in habi-tats that are effectively free of predation threatsfrom large raptors. Whereas most Greater Antil-lean amazons and essentially all mainlandAmazona have to deal with threats from large rap-tors such as Red-tailed Hawks (Buteo jamaicen-sis), Peregrine Falcons (Falco peregrinus), andvarious Accipiter species, no large accipiters orbuteos occur in the Lesser Antilles, and the Pere-grine Falcons of these islands are largely winter-ing birds utilizing coastal areas separate from therain forest habitats occupied by the LesserAntillean parrot species.
The largest raptor in parrot habitats of most of
14 Manual of Parrot Behavior

the Lesser Antilles is the Broad-winged Hawk,which is too small to represent a credible threat tothe parrots and for which there are no records ofparrot predation. The Red-tailed Hawk, on theother hand, is not a species to be underestimatedin its capacities to take Amazona parrots. Recordsexist of it successfully dispatching a variety ofAmazona and Rhynchopsitta species in theGreater Antilles and on the mainland.
Thus, to the extent that conspicuous flockingbehavior has often been suggested as primarily anadaptation to reduce risks of avian predation, theLesser Antillean amazons might be expected togain little by flocking behavior and the tendencymay never have evolved or may have disappearedin the evolutionary history of these speciesbecause of very low predation threats. Flockingbehavior has often been envisioned as primarily ameans to reduce predation via the increased vigi-lance possible when the combined sensory capac-ities of multiple individuals are available andwhen specific individuals can serve as sentinelsfor groups (see discussions in Snyder et al. 1987and Yamashita 1987).
Only one of the Greater Antillean amazonsshows social behavior similar to that of the LesserAntillean species, the race of the Cuban Parrot onCayman Brac (Amazona leucocephala hesterna).Like the Lesser Antillean species, the CaymanBrac Parrot rarely travels in groups larger thanfamily groups, and its male and female adultsoften feed their young independently (Wiley,unpublished). And like the Lesser Antilleanspecies, and as one might predict from the pre-ceding discussion, this parrot lives in an environ-ment free of significant avian predators. No Red-tailed Hawks or large accipiters occur on CaymanBrac, and the Peregrine Falcons that are seenthere occasionally are mostly on the coast, posingno significant risks to the parrots.
The associations of low sociality with low pre-dation risks and high sociality with high preda-tion risks strongly suggest a causal connection ofthese features. Further reinforcing this conclusionis the fact that the race of the Cuban Parrot onGrand Cayman (Amazona leucocephala cayma-nensis) shows the typical Amazona tendencytoward large flocks and apparently feeds its youngas pairs. Significantly, there are Red-tailed Hawkson Grand Cayman, unlike nearby Cayman Brac.Thus the parrots of the various Cayman Islands in
themselves give strong evidence for the impor-tance of avian predators in producing social ten-dencies among Amazona parrots.
We also call attention to the especially well-developed sociality of the Thick-billed andMaroon-fronted Parrots of Mexico. These Rhyn-chopsitta species are similar to the amazons insize and face predation risks from the same sortsof avian predators. In particular, these speciesface significant predation threats from both Red-tailed Hawks and Peregrine Falcons, and in thecase of the Thick-billed Parrot, also from ApacheGoshawks (Accipiter gentilis apache). In ourexperience, sociality in the Rhynchopsitta speciesis even more highly developed than in anyAmazona species for which we have data. In fact,pairs of the Thick-billed Parrot often nest veryclose together, sometimes with more than onepair in the same tree, while Maroon-frontedParrots typically nest in dense colonies in cliffs.Moreover, observations indicate that breedingmales of the Thick-billed Parrot typically associ-ate in combined flocks for foraging, often waitingfor one another to leave the nesting areas as agroup. Such coordinated male behavior has notbeen regularly recorded for any Amazona species.
Other explanations for the flocking and social-ity of amazon parrots—for example, traditionalarguments for advantages in food finding in birds(see Krebs 1974)—have difficulty in accountingfor the variations in sociality seen in variousAmazona species. That there might be any basicdifferences in food availability for the CaymanBrac Parrot and the Lesser Antillean species thatcould explain their low sociality is undocumentedand does not seem intuitively likely. Available evi-dence suggests that they feed on much the samefoods that are taken by other more social speciesin the genus.
Regardless of what factors are truly mostimportant in producing the relatively high degreeof sociality found in most Amazona and Rhyn-chopsitta, this characteristic is generally consid-ered to be adaptive in the lives of these species.Modern circumstances, however, can produce sit-uations where this is clearly not true. We callattention to a recent instance of mass drowning ofRhynchopsitta terrisi in an artificial water catch-ment where the species suffered a major popula-tion stress precisely because of its high degree ofsociality. In this instance, in 1994, at least 52 R.
2 / Behavior of Wild Amazona and Rhynchopsitta Parrots 15

terrisi perished when they were unable to exitfrom an artificial cement water tank that they hadapparently entered for drinking and/or bathingpurposes. When one considers that the total pop-ulation of this species is only about 3,000 birdsand the annual recruitment of young is only about200 individuals, this event was nothing short ofcatastrophic (Macías-Caballero et al. 2001).
As an aside, Rhynchopsitta parrots, like manyspecies of Australian parrots, but unlike mostNeotropical parrots inhabiting humid environ-ments and consuming foods high in water con-tent, come to water sources, such as waterfalls, todrink on a daily basis. This behavior has beendocumented in both species of Rhynchopsitta(Snyder et al. 1999; Macías-Caballero et al.2001), and, like the parrot assemblages at claylicks of the Amazon basin, constitutes a mar-velous spectacle. Unfortunately, man-inducedchanges in the environment have both reduced theavailability of springs and waterfalls in the land-scape and increased the presence of artificialwater catchments that can pose inadvertent risksof mortality to the species.
Another species for which high sociality mayhave led to major population stress from humansources is the extinct Carolina Parakeet (Conu-ropsis carolinensis). Flocks of this species wereexceedingly vulnerable to shooting, and the ten-dency of the species to roost together in largegroups in hollow trees made it susceptible toheavy harvest for the pet trade, both of which fac-tors were of presumed importance in the species’decline (Snyder & Russell 2002). The high social-ity of this species may also have rendered it high-ly susceptible to the spread of exotic diseases.
Finally, as another aside, we note that theabsence of any strong tendency for flocking in theLesser Antillean and Cayman Brac Amazona is afactor that makes censusing of these species espe-cially difficult. Although counts of large flocksentering and leaving roosts have proven an effec-tive way to census many other Amazona species—for example, the Puerto Rican Parrot and theBahama Parrot (A. leucocephala bahamensis)—itis not a practical option for species with lowflocking tendencies.
TIMING OF NESTING SEASONS
In the West Indies, most amazon parrots beginegg laying in the late winter and early spring, with
March usually the peak month. This timing is ingeneral correlated with the dry season, and couldbe related primarily to minimizing risks of nestloss to flooding, although it could alternatively bekeyed to seasonal aspects of food availability.Strongly suggesting the latter is the abnormallylate egg-laying period seen in the Bahama Parrotsof Abaco, which do not normally lay until lateMay and early June, just before onset of the rainyseason in that region (Gnam 1991). Here, layingappears to be timed to take advantage of the abun-dance of poisonwood (Metopium toxiferum)fruits, wild guava (Tetrazygia bicolor) fruits andappropriate-aged pine (Pinus caribea) seeds inmidsummer, the most important known foods forthe species in provisioning young. The BahamaParrots on Inagua Island apparently lay at a moretypical time in the early spring, in line with otheramazons of the West Indies (Snyder et al. 1982).Pine is absent from Inagua, and poisonwood isnot nearly as conspicuous an element of the floraon this island as on Abaco.
Breeding seasons of mainland amazons havebeen especially closely studied in northeasternMexico (Enkerlin-Hoeflich 1995) and are similarto most West Indian amazons, with peak laying in late March and early April (Table 2-1). Thesympatric Red-crowned Parrot, Yellow-headedParrot, and Red-lored Parrot have similar egg-laying dates. Food is abundant during spring andsummer for Mexican Amazona. There is no clear-cut dry season, although spring and summer usu-ally show peaks in rain and winter is normallydry. Their breeding season is earlier as one movessouth and would indicate that it is more related tophotoperiod or temperature than to food avail-ability (Enkerlin-Hoeflich 1995, unpublisheddata).
Breeding seasons of the Rhynchopsitta speciesare extremely delayed relative to the Amazonaspecies, and this delay is almost surely keyed totheir specialized diets, primarily of variousconifer seeds, which do not normally becomeabundantly available until midsummer, with earlyJune being the low point in seasonal availabilityof seeds for the conifer species in the ranges ofthe species. The mean egg-laying date of theThick-billed Parrot in Chihuahua has been mid-July, with most chicks fledged by the first or sec-ond week of October (Snyder et al. 1999). TheMaroon-fronted Parrot starts somewhat later with
16 Manual of Parrot Behavior

most egg-laying in late July to early August andchicks fledging at the end of October through thefirst week of November.
Thus, the evidence for importance of diet indetermining the timing of breeding is highly sug-gestive both in the Bahama Parrot and theRhynchopsitta parrots, and diet may be the mostimportant factor with the other Amazona as well,although this is less clear from available data.Future studies focused on crop sampling of nest-lings of a variety of species to rigorously deter-mine dietary relationships (Enkerlin-Hoeflich etal. 1999), combined with studies of seasonalavailability of primary foods, may help solidifyknowledge of the most important factors deter-mining the timing of breeding.
NEST SITE AND PAIR FIDELITY, ANDCAVITY REUSE
In general, nest site and pair fidelity tend to behigh for psittacine birds (Snyder et al. 1987;Rowley & Chapman 1991), although there arevariations to be seen among species. High pairfidelity, for example, has been found in twospecies of Amazona in northeastern Mexico—the Red-crowned and Yellow-headed Parrots(Enkerlin-Hoeflich 1995), and as with nest fideli-ty, may often be associated with improved pro-ductivity as the years of experience accumulate.In many studies, cases of divorce have been large-ly limited to instances of reproductive incompe-tence of one of the pair members (Snyder et al.1987; Rowley & Chapman 1991).
2 / Behavior of Wild Amazona and Rhynchopsitta Parrots 17
Table 2.1. Clutch initiation, incubation periods, feeding visits, and fledging age forAmazona parrotsa
A. autumnalis A. oratrix A. viridigenalisDescriptive statistics (n = 24) (n = 6) (n = 26)
Week (number of nests)1 = 19–24 March 3 1 12 = 25–31 March 4 3 73 = 1–7 April 10 1 104 = 8–14 April 5 1 25 = 15–21 April 1 0 56+ = after 22 April 1 0 1
“Mean” (week of initiation)b 3.00 2.33 3.23Range (week of initiation) 1 to 6 1 to 4 1 to 6Standard deviation 1.22 1.03 1.27Coefficient of variation 0.41 0.44 0.39Average initiation of clutch (date)c 2 April 31 March 5 AprilMean duration of incubation (days) 28d 28d 27 (n = 7)Mean daily feeding visits to the nestse 2.09 2.18 2.08Range of daily visits to the neste 0–3 0–3 0–4Mean age at fledging (days) 55 (n = 4) 57 (n = 2) 53 (n = 9)
aBased on nests inspected with a burrow probe in the 1993 and 1994 breeding seasons. For A. oratrix,two additional nests from the 1992 season were included to increase sample size; although no burrowprobe was available in 1992, these two nests were shallow enough to be inspected directly.
bAn index calculated from six categories (weeks) assigned based on day of initiation. A test usingKruskal-Wallis on this index showed no difference among species (KW = 2.5, df = 2, p < 0.281).
cCalculated from actual date of initiation for each nest.dAs reported in the popular captive breeding literature.eEstimated by multiplying the average number of visits per observation session by two as justified in
Enkerlin-Hoeflich 1995. The range of visits also refers to the observation sessions only.

Maroon-fronted Parrots nest in colonies rang-ing from one or two to more than 100 pairs. Pairsseem to have strong site fidelity, at least to thesame colony, if not the same nest hole, as demon-strated by returns of birds carrying radio trans-mitters over periods of several years. Similarly,established pairs of most Amazona exhibit amarked degree of philopatry. For example, innortheastern Mexico in 1993, four pairs of visual-ly distinctive Amazona that switched nest sitesmoved to new nests within a 50 m radius of theirprevious nests. In 1994, five pairs had new nestswithin a 50 m radius of their previous nests andtwo pairs moved within a radius of only 100 m.The attachment to specific nesting areas can besomething that occurs rapidly: a female Red-crowned Parrot released with a radio collar estab-lished her nest sites in two successive nestingperiods in trees within 200 m of the release cage(Enkerlin-Hoeflich 1995).
At least six pairs of Red-crowned Parrots andfive pairs of Yellow-headed Parrots individuallyrecognizable by feather characteristics showedmate fidelity between successive nesting periods,and at least three of each species exhibited fideli-ty for three nesting periods. Such high matefidelity has also been documented in the PuertoRican Parrot by Snyder et al. (1987) and may begenerally true in the genus Amazona.
Fidelity to specific nest sites, however, is morevariable. Enkerlin-Hoeflich’s (1995) studies ofRed-crowned and Yellow-headed Parrots in 1993and 1994 revealed that fidelity to specific siteswas low compared to that reported in otherAmazona (Snyder et al. 1987; Gnam 1991; Rojas-Suárez 1994). In large measure, this differencemay reflect species differences in cavity availabil-ity, with suitable cavities being considerably moreabundant for the Red-crowned and Yellow-headedParrots than for other amazons, although addi-tional factors may well have been involved aswell. Nest switching is standard in many cavity-nesting birds (e.g., Boreal Owls, Aegoliusfunereus, and California Condors, Gymnogypscalifornianus—see Hayward & Hayward 1993and Snyder & Schmitt 2002), and may offer gen-eral advantages, such as reductions in parasiteinfestations, that need to be balanced againstadvantages that may result from maintaining sitefidelity, especially in cavity-poor environments.
In many species, there is a tendency for pairs to
switch nest sites after failures to fledge young anda tendency to stay with nest sites after success infledging young (Saunders 1982). One pair ofPuerto Rican Parrots studied over many years fol-lowed this pattern religiously, while other pairsexhibited strong nest-site fidelity regardless ofsuccess or failure in the sites over the years (Sny-der et al. l987). As an aside, until it was learnedthat the latter pattern was the more typical one forthis species, efforts to multiple-clutch wild pairswere held in abeyance because of concerns thatsuch efforts would drive pairs into using new nestsites for replacement clutches that might be vul-nerable to predation by Pearly-eyed Thrashers(Margarops fuscatus). But once the strong ten-dency of pairs to stick with nest sites, despitefailure in the sites, was established, multiple-clutching efforts were initiated with considerablesuccess and without causing pairs to abandon sites.
Even with relatively low levels of nest reuti-lization, pairs of Red-crowned and Yellow-headedParrots have exhibited greater tendencies to reusesites in which they have succeeded than sites inwhich they have failed. Similarly, studies ofMaroon-fronted Parrot nesting colonies indicatethat cavities producing fledglings are generallythe cavities most frequently reused over several-year periods.
Thus there are reasons to suspect that poachingof entire broods from nests of many species maynot only remove immediate reproduction but mayalso affect future reproduction by stimulatingpairs to move to new and untested nest sites, bothbecause poachers frequently destroy nest sites inharvesting them and because they often stimulatethe birds to move even if they do not harm thenest sites. If instead parrot trappers were to allowat least one young to fledge per nest and were notto harm nest sites in harvesting young, both par-rots and trappers might ultimately benefit fromgreater overall parrot populations and nest suc-cess in the populations. Instituting such relativelyprudent harvesting procedures, unfortunately, isunlikely in areas subject to unregulated harvest,because maximization of short-term benefitstends strongly to overbalance maximization oflong-term benefits.
FEEDING BEHAVIOR AND RATES
Amazon parrots of the mainland, such as Red-crowned Parrots of northeastern Mexico, almost
18 Manual of Parrot Behavior

invariably feed their nestlings only twice a day,regardless of the age of the nestlings—once inearly to mid-morning and once in late afternoon(Enkerlin-Hoeflich 1995). This pattern alsoapplies to Lilac-crowned (A. finschi), Yellow-headed, Yellow-naped (A. auropalliata) , White-fronted (A. albifrons), and Red-lored Parrots ofMexico and Central America (personal observa-tions; Renton & Salinas-Melgoza 1999). In con-trast, amazon parrots of the West Indies, both inthe Greater and Lesser Antilles, typically feedtheir young four to five times per day, a major dif-ference. The Puerto Rican Parrot, for example,averages about 4.6 feeding trips per day duringthe nestling period (Snyder et al. 1987). Similarly,the Hispaniolan Parrot (A. ventralis) averages 4.3feeding trips per day and the Cuban Parrot (A.leucocephala) averages 4.5 feeding trips per day(Wiley, unpublished). The differences amongspecies in provisioning rates have potentiallyimportant consequences, for example withrespect to vulnerabilities of adults and nests topredation, and it is of considerable interest to seekpotential causes of the differences.
As studied by Enkerlin-Hoeflich (1995) inRed-crowned, Yellow-headed, and Red-loredParrots, the overall commitment of mainlandamazons to a regimen of two feedings per day atnests is extreme. The first feeding visit to nestsusually takes place about one hour after sunrise,presumably after a foraging bout. The secondvisit occurs about one and a half hours beforesunset, shortly before adults assemble in roosts.Other mainland Amazons exhibit similar patterns(e.g., Renton & Salinas-Melgoza 1999). In thesestudies, provisioning trips were highly stereo-typed in timing even in cases where feedings wereomitted at the expected time in the morning orwere interrupted in the morning due to distur-bance (e.g., presence of a predator) or otherunknown factors. The birds did not attempt com-pensatory feedings during the middle of the dayto make up for the missed feedings, and insteadwaited until the next normal evening feeding peri-od to resume feeding the young. On a few occa-sions, we observed that as many as two consecu-tive feeding sessions were omitted, resulting in upto 36 hours of fasting for the chicks. Never-theless, the chicks involved eventually fledgedsuccessfully. The pattern of visiting the nest onlyonce in the morning and once in the evening
seems so highly ingrained in these species thatthey seem almost incapable of provisioning atother times of day.
In contrast, amazons of the West Indies charac-teristically make four to five feeding trips to theirnests per day, including multiple trips during themidday hours, and this is true of both the largeLesser Antillean species and the much smallerGreater Antillean species. Somewhat intermedi-ate is the Black-billed Parrot of Jamaica, with anaverage of 3.8 feeding trips per day (Koenig1999).
What factors could explain the differences inprovisioning rates between the mainland andisland amazons? Two possible explanations comeimmediately to mind—differences in the nutri-tional quality of diets and different daily regimesof temperature stress. The mainland populationsstudied to date have essentially all been close tosea level in regions with high midday tempera-tures potentially offering stress for adults in for-aging at that time of day, whereas the island ama-zon populations studied have mostly been rela-tively high-elevation rain forest populations thatare spared comparable midday heat stress. But inaddition, there appears to be a major difference inquality of foods offered to nestlings, with cropsampling of nestlings indicating very high pro-portions of seeds in the diet of mainland species(Enkerlin-Hoeflich 1995), and with most foodsdocumented for the island species being variouskinds of soft fruits (e.g., Snyder et al. 1987). Asseeds are a much more concentrated form ofnutrition than fruits, the differences in provision-ing rates at nests could be largely a consequenceof a necessity for the fruit eaters to process largevolumes of food to compensate for the low nutri-tional quality of fruit foods.
These hypotheses are not mutually exclusiveand surely are not the only explanations that couldbe offered for the feeding rate differences, but itis worth examining available data to see if boththese hypotheses are consistent with existinginformation. Here, we caution that comprehen-sive determinations of diet fed to nestlings aredifficult to achieve by observations of adult feed-ing behavior, as studies of Enkerlin-Hoeflich etal. (1999) have shown that crop sampling ofnestlings often yields rather different evaluationsof diet than observations of adults foraging.Nevertheless, where crop sampling has been
2 / Behavior of Wild Amazona and Rhynchopsitta Parrots 19

employed most comprehensively—for mainlandamazons—it appears that seeds are indeed theoverwhelming food types given to nestlings.Much less crop sampling of nestlings has beenperformed with the island amazons, and althoughthese data are generally consistent with a predom-inance of fruit feeding, the database is not nearlyas good as for the mainland amazons.
With respect to temperature stress explana-tions, a precise quantification of environmentalconditions faced by various species has not beenachieved, and could also be related to timing ofnesting as was discussed earlier. Nevertheless, wenote that the Bahama Parrots of Abaco Islandexist in a near-sea-level environment with highmidday temperatures, yet also exhibit a high rateof provisioning at nests (averaging about fivetrips per day). Although these parrots have notbeen studied with comprehensive crop sampling,they do appear to take many fruits as breeding-season food, especially poisonwood and wildguava, although they also take substantial quanti-ties of pine seeds. Thus the diet of this species onAbaco seems relatively similar to that of otherGreater Antillean species and the high feedingrates of the Abaco parrots provide some apparentsupport for dietary explanations of provisioningrates and lack of support for major temperatureeffects. Similarly, studies of sea-level populationsof the Cuban Parrot in Cuba have also yieldedhigh provisioning rates (4.5 trips per day) in spiteof potential temperature stresses. The latter alsoshow overall diets with a substantial proportion offruit (Gálvez-Aguilera et al. 1998).
Somewhat different conclusions apply to theBlack-billed Parrot of Jamaica, with an interme-diate feeding rate of about 3.8 trips per day to thenest. One might hypothesize that the Black-billedParrots might be feeding relatively higher por-tions of seeds to their nestlings, but this has yet tobe shown conclusively. The Jamaican Black-billed Parrots have been studied at mid-elevations, in relation to some other amazons of theWest Indies, so they would presumably fall some-where in the middle in temperature relationships,although this has not been carefully documented.
The Rhynchopsitta parrots also offer somepotential for distinguishing food quality versustemperature effects on feeding rates. Crop sam-pling of Thick-billed Parrot nestlings indicates ahigh proportion of seeds in the diet, primarily
pine seeds and, to a lesser extent, acorns, and yetthe species occupies a relatively cool and temper-ate high-elevation range during the breeding sea-son (Snyder et al. 1999). On the basis of a seeddiet, one might anticipate only two feedings perday in Thick-bills, by comparison with the ama-zon species, but on the basis of temperature rela-tionships, one would anticipate an absence ofmajor midday temperature stress, and thereforethe potential for more feeding trips per day. Datafor the Thick-billed Parrot (Snyder et al. 1999)indicate a usual provisioning rate of three to fourtrips per day, an intermediate result apparentlymost consistent with temperature explanationsbut not entirely inconsistent with nutritionalexplanations, as the cool high-elevation habitatsoccupied by these species may increase food orenergy needs relative to those of lowland seed-eating amazons. This is reinforced by the pres-ence of brood reduction due to starvation ofyounger chicks in Thick-billed Parrots, rarelyseen in Amazona.
Clearly, progress in resolving alternativehypotheses could be achieved by intensive crop-sampling efforts with nestlings of a variety ofamazon species, showing a variety of feeding ratepatterns, and by quantification of environmentalconditions in nesting habitats of the variousspecies.
RELATIONSHIPS OF SPECIESCONSPICUOUSNESS AND NESTACCESSIBILITY TO EXPLOITATION INTHE PET TRADE
The three species of Amazona studied byEnkerlin-Hoeflich (1995) in northeasternMexico—the Red-crowned, the Yellow-headed,and the Red-lored Parrots—offer an especiallyinstructive look at the influence of behavior onthe comparative vulnerability of parrots toexploitation by the pet trade, especially throughnest robbing. For the three species, nests of theRed-crowned Parrot are by far the easiest to find,and this species has been so heavily exploited bypoachers that it has been totally eliminated frommany areas, especially in riparian habitats that areeasily accessible (Iñigo-Elias & Ramos 1997;Clinton-Eitniear 1986). The Yellow-headed Parrothas also been very popular in trade because of itscapacities to talk, and has also suffered greatlyfrom poaching, although its nests are less easy to
20 Manual of Parrot Behavior

find. The most inconspicuous species, the Red-lored Parrot, is in contrast more abundant andwidely distributed, and may even be increasing innumbers in the same region inhabited by the othertwo species. Even without the legal protectionenjoyed by the Red-crowned Parrot and theYellow-headed Parrot, this species has been cap-tured and traded within Mexico to a considerablylesser extent. Its nest trees are as easy to climb asthose of the Red-crowned Parrot and Yellow-headed Parrot, and it is abundant in some second-growth and agricultural landscapes, so the lesserdegree of human impacts on the species deservesanalysis.
Perhaps the most crucial factor in the successof the Red-lored Parrot is its overall wariness andinconspicuousness around nests, leading to majordifficulties for humans in finding nests. Pairs ofRed-lored Parrots are loud in the general vicinityof nests but secretive in their movements close totheir nest trees. Approaches to, landings at, andtakeoffs from nests are all done silently. Further,Red-lored Parrot pairs often wait for long periodsin a tree distant from the nest tree before movingto a nest. Pairs are usually very loud during inter-or intraspecific interactions, but this is usuallydistant from the nest and rarely helpful in de-termining a nest location. The routines for eachpair are difficult to establish and on several occa-sions have led to mistakes in identifying their nestlocations.
In contrast, the nests of the Red-crownedParrot are not difficult to find, because the ap-proach of an adult (whether a lone male ap-proaching a nest to feed a female or both birdsarriving together) is very deliberate and isannounced by a series of characteristic calls.Often members of a pair do not land directly onthe nest tree but close to it, with subsequent flightto the nesting tree. If the female is inside the nest,the male usually calls and the female promptlycomes out. Both birds usually fly together withcharacteristic takeoff squawks and land nearby.This series of events is stereotyped and clearlyadvertises the presence and location of the nest. Ifthe birds are disturbed by a human coming closeto the nest location, they take off with loud rau-cous calls but land nearby and soon return to thenest (within 14–79 minutes, n = 13), if the humanobserver has concealed him- or herself.
The nests of the Yellow-headed Parrot are inter-
mediate in difficulty of detection. Pairs of thisspecies tend to vocalize for long periods near thenest tree, but they rarely approach the actual nesttree and do not move to the nest tree if they per-ceive the presence of an observer. In response tothe arrival of the male, the female sometimescomes out of the nest promptly, but more oftenshe takes many minutes to come out. In contrastto the Red-crowned Parrot, Yellow-headed Parrotsdo not give any characteristic calls when depart-ing from the nest. Together, these characteristicsmake it relatively difficult to find nests of Yellow-headed Parrots.
In keeping with the relative difficulties in find-ing nests of the three species, the first new nestslocated each breeding season in the study ofEnkerlin-Hoeflich (1995) were predominantlythose of Red-crowned Parrots (13 of 21), where-as nests found in the second half of the breedingseason were predominantly those of Red-loredParrots (16 of 21). Although the timing of egglaying was the same in both these species, thesedifferences reflect the much longer times it takesto find the nests of Red-lored Parrots.
Thus, the easy detectability of nests has evi-dently made the Red-crowned Parrot exceedinglyvulnerable to illegal harvest of nestlings, andalthough nests of the Yellow-headed Parrot areconsiderably more difficult to find, the high mar-ket value of this species has likely greatlyincreased the motivation of poachers to overcomethe problems involved in finding its nests. In con-trast, the extreme difficulty in finding nests of theRed-lored Parrot and its lesser market valueappear to have enabled it to survive and eventhrive in spite of unregulated harvest. One mighthypothesize that species such as the Red-crownedParrot and the Yellow-headed Parrot are likelyunder strong selective pressures toward becomingless conspicuous around nests, but as yet any suchpotential changes in behavior have apparentlybeen insufficient to prevent continuous popula-tion declines.
The amazon species of the West Indies varygreatly in their conspicuousness around nests.Many are relatively conspicuous, advertising nestlocations by loud vocalizations, but two species—the Imperial Parrot and the Cayman BracParrot—are so extremely inconspicuous aroundnests that their nests are infrequently found andharvested. This has been especially true for the
2 / Behavior of Wild Amazona and Rhynchopsitta Parrots 21

Imperial Parrot. On Cayman Brac, the harvest hasbeen more frequent due to the concerted search-ing efforts of poachers and absence of many nesttrees that are difficult to climb. As in Mexico, har-vest of nestlings of the West Indian species hasbeen especially severe for the species that aremost conspicuous, and of the island amazons per-haps only those in the Lesser Antilles havereceived any substantial protection from poachersby relatively frequent inaccessibility of their nestsin enormous and difficult-to-climb canopy trees.However, despite such difficulties, poachers onSt. Vincent have traditionally managed a substan-tial harvest, having become especially adeptclimbers.
Both species of Rhynchopsitta in Mexico areconspicuously noisy around nests and nests arenot difficult to find, yet there has been relativelylittle harvest of nestlings of either species. For R.terrisi, the main factor preventing nestling har-vest has surely been the awesome inaccessibilityof essentially all nests. Nests are all in deep solu-tion holes high in precipitous, towering cliffs, andeven if climbers might occasionally get to espe-cially low nest entrances, they can rarely get tothe chicks far within. For R. pachyrhyncha, therehas been some harvest of nestlings, but a substan-tial fraction of nests have been in huge dead treesthat are difficult and unsafe to climb and too largeto be cut down easily.
Thus both inconspicuousness and inaccessibili-ty of nests appear to be important factors decreas-ing poaching rates of Amazona and Rhynchopsittaparrot nests, and although some species have ap-parently benefited by possessing one or the otheror both of these traits, a great many others havebeen stressed, apparently to the point of endanger-ment in some cases, by lacking either trait.
DEFICITS IN BREEDING EFFORT
The Yellow-headed Parrot of Mexico and thePuerto Rican Parrot have shown a chronic tenden-cy for the existence of many pairs that adopt nestsites but do not lay eggs (Snyder et al. 1987;Enkerlin-Hoeflich 1995). Roughly half the pairsof the Puerto Rican Parrot studied over the yearshave been non-egg-laying, and statistics havebeen worse with the Yellow-headed Parrot.Similarly, various macaws in the Amazon and inMexico have a large proportion of non-breeders(Munn 1992; Marineros S. 1993; Iñigo-Elias
1996). As many of these species are consideredendangered, it would be highly advantageous todiscover the causes of the apparent reluctance tobreed in many pairs, as the causes may be suscep-tible to reversal by conservation actions.
In both Amazona just mentioned, the causes oflow breeding effort do not appear to lie in ascarcity of good nest sites, but research to datehas not yielded a clear understanding of whatother causes may be involved. Among the varioushypotheses are potential dietary limitations, prob-lems with sex ratios and homosexual pairs, andproblems with abnormal age distributions inpopulations.
The comparative approach has not yet yielded aclear resolution among alternatives, although itmay do so in the future. In most amazons understudy, the pairs adopting and defending nest siteshave almost always been egg-laying pairs, but it ispossible in these other species that pairs with noreal potential for egg laying may generally fail toform associations with nest sites, so they are sim-ply missed by standard survey techniques. But,assuming that all species may have similar tenden-cies to form associations with nest holes, we havebeen unable to find other obvious features thatcould explain the apparent differences in propor-tions of breeding pairs in any conclusive fashion.
Nevertheless, with the Puerto Rican Parrot, afortuitous event in 1989 has provided some intri-guing and suggestive clues. This was the directstrike of the parrot range by Hurricane Hugo.Although this storm caused tremendous damageto vegetation and a loss of approximately half theparrot population, it was remarkably followed bya tremendous increase in the numbers of breedingpairs, which lasted for several years and then sub-sided back to a more usual level. Clearly a sub-stantial number of birds that had been chronicnon-breeders before the storm became breedersafter the storm, but the effect did not last for morethan a few years. These facts strongly suggest thatthe basic causes of non-breeding were not to befound in sex ratio or age distribution problems.
Instead, we suspect that the causes are muchmore likely to be found in some subtle dietaryrelationships. Many trees and shrubs of Carib-bean rain forests show greatly enhanced fruitingfollowing hurricanes, and there are also well-documented insect blooms following hurricanesthat apparently underlie enhanced breeding in
22 Manual of Parrot Behavior

other avian species following hurricanes (Arendt1992; Wunderle 1995; Wiley & Wunderle 1993).Detailed dietary studies of species such as thePuerto Rican Parrot and the Yellow-headed Parrotin comparison with other species, especiallyusing crop-sampling techniques carried out overperiods of many years, might shed light on thepossibility that the species showing generally lowbreeding effort might suffer from nutritional lim-itations and might respond to dietary modifica-tions that could be introduced as managementtechniques.
As an aside, we point out that despite the obvi-ous devastating effects of Caribbean hurricaneson diverse amazon parrots, these storms may havean important positive role to play in the biology ofthese species, not just from a dietary standpointbut also from the standpoint of creation of nestsites through breakage and subsequent rotting oftree limbs. Breeding in the Puerto Rican Parrotwas evidently sufficiently enhanced by HurricaneHugo such that the population had recovered tonearly 90% of its former size only four years afterthe storm. Further, it is conceivable that some fre-quency of hurricanes is actually beneficial andeven necessary to the ecology of this species. Theabsence of major hurricanes between the 1930sand 1989 could have been an important factor inthe progressive decline of the species.
SUMMARY
Comparative studies of the behavior and ecologyof Amazona and Rhynchopsitta parrots allow theevaluation of a variety of hypotheses concerningthe determinants of important characteristics ofthe species. While many of the conclusions pre-sented here are tentative and demand furtherresearch for confirmation, they offer potentialsfor enhancing the conservation of the species in anumber of respects. We particularly emphasizethe potential benefits of careful quantitativedietary studies of many species utilizing crop-sampling techniques, as dietary relationships mayunderlie much of the behavior of these species.
ACKNOWLEDGMENTS
Much of the information reported has resultedfrom a long-term program focused on parrot ecol-ogy and conservation initiated by the first authorin 1990 as part of doctoral studies at Texas A&MUniversity and continued at Monterrey Institute
of Technology (ITESM), Mexico, since 1994. The“parrot team” that provided direct assistance inmany years of field work was led at differenttimes by Claudia Macías-Caballero, TiberioMonterrubio-Rico, Miguel Angel Cruz-Nieto, S.Gabriela Ortiz-Maciel and included a large num-ber of volunteers, students, ranch hands, indige-nous and local communities, and so forth. Thisproject would not have been possible withoutassistance and cooperation from a wide range ofpeople and institutions. I particularly appreciatethe help and advice of Jane Packard, who chairedmy doctoral advisory committee. Also RandyBrue, Wylie Barrow, Michael Schindlinger, TilaPérez, and Robert B. Hamilton. Among these Iwant to thank for participation in different stagesof the field research Jim Shiflett, Martjan Lam-mertink, Steve Scheid, Javier Cruz, Diana Vene-gas, Roger Otto, Emilio Rojas, Ali Taylor, theTutuaca Ejido, and the community of Vallecillo.
The project could never have succeeded with-out the financial and administrative support ofCentro de Calidad Ambiental at ITESM.
Financial support was generously provided bymany agencies and organizations. Wildlife Trustprovided consistent economic support duringthese studies. CONABIO (Mexican Commissionfor Biodiversity Research), FMCN (MexicanFund for Nature Conservation), CONACYT(Consejo Nacional de Ciencia y Tecnologia), theAmerican Zoo and Aquarium Association, theSacramento Zoo, the National Fish and WildlifeFoundation, the Arizona Game and Fish Depart-ment, and the USFWS/SEMARNAT (U.S. Fishand Wildlife Service/Secretaria de MedioAmbiente y Recursos Naturales) program in Bio-diversity Conservation all were important finan-cial contributors. Additional support has beenprovided by Louisiana State University and anumber of private individual donors.
REFERENCESArendt, W.J. 1992. Impact of Hurricane Hugo on pearly-
eyed thrasher reproduction in the Luquillo rain for-est, Puerto Rico. Pitirre 5(3):13–14.
Clinton-Eitniear, J. 1986. Status of the green-cheekedamazon in Northeastern Mexico. Watchbird Dec/Jan:22–24.
Cruz-Nieto, M.A., E. Enkerlin-Hoeflich, and N.F.R.Snyder. 1998. “Estatus y ecología de anidación de laCotorra Serrana Occidental (Rhynchopsitta pachy-rhyncha) en México: efectos de las prácticas del uso
2 / Behavior of Wild Amazona and Rhynchopsitta Parrots 23

del suelo y perspectivas futuras de manejo.” InCongreso de Investigación y Extensión del SistemaITESM. Generación, transferencia y aplicación delconocimiento, ed. ITESM, pp. 73–81. Monterrey,N.L. Mexico: Tomo I.
Enkerlin-Hoeflich, E.C. 1995. Comparative ecologyand reproductive biology of three species ofAmazona parrots in northeastern Mexico. PhD diss.,Texas A&M University, College Station.
Enkerlin-Hoeflich, E.C., and K.M. Hogan. 1997. “Red-crowned Parrot, Amazona viridigenalis.” In Thebirds of North America, ed. F.B. Gill and A. Poole, p.20. Philadelphia, PA: American Ornithologists’Union and Academy of Natural Sciences ofPhiladelphia.
Enkerlin-Hoeflich, E.C., J.M. Packard, and J.J.González-Elizondo. 1999. Safe field techniques fornest inspections and nestling crop sampling of par-rots. J Field Ornithology 70:8–17.
Forshaw, J.M. 1990. Parrots of the world. Ontario,Canada: Silvio Matachione & Co.
Gálvez-Aguilera, X., J. Rivera R., F. Quiala G., andJ.W. Wiley. 1998. Breeding season diet of the Cubanparrot Amazona leucocephala in Los Indios Ecolo-gical Reserve, Isla de Juventud, Cuba. Papageien-kunde 2:325–334.
Gnam, R.S. 1991. Breeding biology of the Bahamaparrot (Amazona leucocephala bahamensis). PhDthesis, the City University of New York, New York.
Hayward, G.D., and P.H. Hayward. 1993. “Boreal owl(Aegolius funereus).” In The birds of North America,No. 63, ed. A. Poole and F. Gill. Philadelphia:Academy of Natural Sciences; Washington, DC:American Ornithologists’ Union.
Iñigo-Elias, E.E. 1996. Ecology and breeding biologyof the Scarlet Macaw (Ara macao) in theUsumacinta drainage basin of Mexico and Guate-mala. PhD diss., University of Florida, Gainesville.
Iñigo-Elias, E.E., and M.A. Ramos. 1997. “El comer-cio de los psitácidos en México.” In Uso y conser-vación de la vida silvestre neotropical, ed. J.G.Robinson, K.H. Redford, and J.E. Rabinovich, pp.445–458.. México: Fondo de Cultura Económica.
Juniper, T., and M. Parr. 1998. Parrots: A guide to par-rots of the world. New Haven and London: Yale Uni-versity Press.
Koenig, S.E. 1999. The reproductive biology ofJamaica’s black-billed parrot (Amazona agilis) andconservation implications. PhD diss., Yale Uni-versity, New Haven, CT.
Krebs, J.R. 1974. Colonial nesting and social feeding asstrategies for exploiting food resources in the greatblue heron (Ardea herodias). Behaviour 51:99–134.
Lanning, D.V., and J.T. Shiflett. 1981. “Status and nest-ing ecology of the thick-billed parrot (Rhynchopsitta
pachyrhyncha).” In Conservation of new world par-rots, ed. R.F. Pasquier, pp. 393–401. St. Lucia:Smithsonian Institution Press.
Lanning, D.V., and J.T. Shiflett. 1983. Nesting ecologyof thick-billed parrots. Condor 85:66–73.
Lawson, P.W., and D.V. Lanning. 1982. “Nesting andstatus of the maroon-fronted parrot (Rhynchopsittaterrisi).” In Conservation of new world parrots, ed.R.F. Pasquier, pp. 385–392. St. Lucia: SmithsonianInstitution Press.
Mabb, K.T. 1997. Roosting behavior of naturalizedparrots in the San Gabriel Valley, California.Western Birds 28:202–208.
Macías-Caballero, C.M., E. Enkerlin-Hoeflich, G.Ortíz-Maciel, A. Madero-Farias, J.J. Manzano-Loza,and M.-L. J.R. 2001. Estudio de la cotorra serranaoriental (Rhynchopsitta terrisi) en México: progra-ma de conservación y manejo de ecosistemas.Reporte final presentado al FMCN. Proyecto A-1-98/93 Centro de Calidad Ambiental, ITESM,Monterrey, N.L. Mexico.
Marineros S., M. 1993. La Lapa Roja (Psittacidae: Aramacao): ecología, turismo y pautas para su manejoen la reserva biológica Carara, Costa Rica. Maestríaen Manejo de Vida Silvestre. Universidad Nacional,Heredia, Costa Rica.
Munn, C.A. 1992. Macaw biology and ecotourism, or“When a bird in the bush is worth two in the hand.”In New world parrots in crisis: Solutions from con-servation biology, ed. S.R. Beissinger and N.F.R.Snyder, pp. 47–72. Washington, DC: SmithsonianInstitution Press.
Rea, A.M. 1997. “The indeterminate parrot of NuevoLeon.” In The era of Alan R. Phillips: A festschrift,pp. 167–176. Albuquerque, NM: Horizon.
Renton, K., and A. Salinas-Melgoza. 1999. Nestingbehavior of the lilac-crowned parrot. Wilson Bull111:488–493.
Rojas-Suárez, F. 1994. “Biología reproductiva de lacotorra Amazona barbadensis (Aves: Psittaci-formes) en la península de Macanao, estado NuevaEsparta.” In Biología y conservación de los psitáci-dos en Venezuela, Caracas, ed. G. Morales, I. Novo,D. Bigio, A. Luy, and F. Rojas-Suárez, pp. 73–87.Caracas, Venezuela: PROVITA.
Rowley, I., and G. Chapman. 1991. The breeding biol-ogy, food, social organisation, demography and con-servation of the Major Mitchell or pink cockatoo,(Cacatua leadbeateri), on the margin of the WesternAustralian wheatbelt. Australian Journal of Zoology39:211–261.
Saunders, D.A. 1982. The breeding behaviour andbiology of the short-billed form of the white-tailedblack cockatoo Calyptorhynchus funereus. Ibis 124:422–455.
24 Manual of Parrot Behavior

Snyder, N.F.R., E.C. Enkerlin-Hoeflich, and M.A.Cruz-Nieto. 1999. “Thick-billed parrot, Rhyncho-psitta pachyrhyncha.” In The birds of NorthAmerica, ed. F.B. Gill and A. Poole. Philadelphia:American Ornithologists’ Union and Academy ofNatural Sciences of Philadelphia.
Snyder, N.F.R., W.B. King, and C.B. Kepler. 1982.Biology and conservation of the Bahama parrot.Living Bird 19:91–114.
Snyder, N., P. McGowan, J. Gilardi, and A. Grajal, eds.2000. Parrots: Status survey and conservationaction plan 2000–2004. Gland, Switzerland andCambridge, U.K.: IUCN World Conservation Union.
Snyder, N.F.R., and K. Russell. 2002. “Carolina para-keet (Conuropsis carolinensis).” In The birds ofNorth America, No. 667, ed. A. Poole and F. Gill.Philadelphia: Birds of North America, Inc.
Snyder, N.F.R., and N.J. Schmitt. 2002. “Californiacondor (Gymnogyps californianus).” In The birds of
North America, No. 610, ed. A. Poole and F. Gill.Philadelphia: Birds of North America, Inc.
Snyder, N.F.R., J.W. Wiley, and C.B. Kepler. 1987. Theparrots of Luquillo: Natural history and conserva-tion of the Puerto Rican parrot. Los Angeles:Western Foundation of Vertebrate Zoology.
Wiley, J.W., and J.M. Wunderle, Jr. 1993. The effects ofhurricanes on birds, with special reference to Carib-bean islands. Bird Conservation International 3 (4):319–349.
Wunderle, J.M., Jr. 1995. Pre- and post-hurricane fruit-ing phenologies: Potential implications for PuertoRican parrots. Pitirre 10 (1):38.
Yamashita, C. 1987. Field observation and commentson the indigo macaw (Anodorhynchus leari), a high-ly endangered species from northeastern Brazil.Wilson Bull 99:280–282.
2 / Behavior of Wild Amazona and Rhynchopsitta Parrots 25

CONSERVATION AND TRADE
Of the 345 or so species of parrots, about a thirdof them are threatened or declining, about threetimes the rate of threat found in average birds, ofwhich about 10% are threatened. Of these threat-ened species of parrots, about half are endangeredprimarily by trade, and the other half by conver-sion of their habitats to agricultural lands. Thespecies endangered by trade are invariably color-ful, large, or good talkers. It is no accident that thelarger wild parrots in private collections (asopposed to the ubiquitous, captive-bred Budgeri-gars and Cockatiels) match closely the colorful,large, talking species that are endangered bytrade. For nearly all of these wild parrots kept aspets, destruction of their wild habitat is a minor orvery much secondary conservation issue for thesespecies, while regional, national, and internation-al demand for the pet trade is the main threat totheir survival in the wild. In not a single case sofar in the history of the world has the keeping ofcaptive parrots by private citizens been of signif-icant use in a successful recovery effort for athreatened population of wild parrots.
Conversely, none of the parrots threatened byhabitat destruction are popular in captivity, eventhough they are exactly the species that couldpotentially benefit from a backup population incase they disappear in the wild because of neartotal destruction of their wild habitat. Of course,private parrot owners feel appropriately guiltyabout keeping wild parrots in cages while tryingto justify it by saying it is helpful to the species.They justify this keeping and breeding of color-ful, large, talking wild parrots by saying, in part,
that they are “saving wild Polly from having hishabitat rug pulled out from under him,” or wordsto that effect. In fact, the reason that the large,colorful, talkative species of wild parrots are introuble is not because of shortage of habitat butbecause so many humans like to keep them aspets. Loving them to death, it would seem!
When a number of years ago I suggested to 200parrot owners, breeders, and dealers that theyshould house backup populations of the primarilydrab, small, non-talking species of parrots that arein danger of extinction because of habitat destruc-tion, the idea was received with stony silence. Noone showed interest in donating to establishingbackup populations of species that have “no com-mercial value.”
Trade in all wild parrots clearly should be ille-gal (as it has been for ten years now in the UnitedStates) unless the source country can prove toserious, independent “conservation inspectors”that there is no harm to wild populations and thatthe trade is helping add value to wild habitat andproduce rural jobs related to parrot conservation.Theoretically, at least, a system of wild ranchingof parrots could be devised, but so far no one hasdone so. Any such systems would have to beimmune to the obvious and less obvious forms ofcheating, including banding or chipping unsus-tainably trapped wild birds to launder them forsale as sustainably ranched. By outlawing allinternational trade in wild parrots that cannot beshown to be sustainably produced, the world mayeventually see the development of independentlyverifiable methods of sustainable, humane ranch-ing of wild parrots.
27
3Parrot Conservation,
Trade, and Reintroduction
Charles A. Munn

While talking about humane methods of ranch-ing wild parrots, it could be argued that the onlyhumane, conservative, and responsible way toranch wild parrots would be by harvesting the lasthatched babies from some or all nests. But beforeany harvest of wild babies, it would be importantto raise the wild populations to carrying capacityand beyond by supplemental feeding of the wildpopulation and by erecting nest boxes or other-wise improving natural nest cavities and wildhabitat and ensuring that all hatched young aretreated for parasites and fledge in good condition.One might be able to raise the carrying capacityof the wild habitat by adding balanced food sup-plementation and nest sites. Such measures mightpermit a controlled harvest of last-hatched youngor even entire first clutches while providing sup-plemental food to the laying hens, who would inmany cases re-lay. In other words, use the besttechniques from traditional aviculture and, in gen-eral, livestock ranching to increase the output ofthe population, while not worrying about psycho-logical or plucking problems in the wild birds, aswild birds never pluck and don’t need behavior-ists and psychologists.
All the simple quota-based harvest systemsthat are used still in some tropical countries innortheastern South America and West and CentralAfrica are particularly crude and inhumane, asthey break up families and even catch breedingadults that leave babies to starve in wild nests.Thus, I am unalterably opposed to simple captureor export quotas of parrots. The only humane andsustainable method for parrot ranching for the pettrade would have to be the harvest of the last,starvation-bound babies in wild populations keptat carrying capacity. If these wild populations aretotally protected from hunting or trapping whilealso given supplemental feeding and an overabun-dance of superb nest cavities, then there is everyreason to think that their wild populations mightincrease to a new, higher carrying capacity. Inother words, parrot producers should treat wildparrots as a rancher would treat his or her mostpampered cattle. Wild parrots brought to pam-pered carrying capacity in a wilderness settingwill add more value to the habitat than any otheruse of the forest—by being major tourist attrac-tions in the wild at the same time that they are theraw material that produce valuable babies reliablyevery year.
So far, every example of parrot trapping in thetropics around the world BUT ONE is a classiccase of the tragedy of the commons, in which rov-ing bands of greedy trappers catch all wild birds,adults and babies, while cutting down or hackingopen all nest trees and breaking up breeding fam-ilies. Each trapper figures that he might as welltrap and sell the last adults and babies before thenext guy gets them.
The one example that I know of a trapper (aformer trapper in this case) being a rational parrotrancher (to the extent that he could be given theconstraints of acting outside the legal system) isone of the most skilled Hyacinth trappers of alltime—Lourival Lima of the state of Piaui, Brazil.Lima and his band of followers effectively had aunique bird harvesting concession in an extensivearea of roughly 1,000 square miles of wildernessin the dry forest region of northeastern Brazil.Lima never allowed any of his band of eightexpert trappers to catch or hurt the adult popula-tion of breeding birds. Rather, as over decades hisfather (who himself was master macaw trapper)and later he himself had established a territorialsystem to keep other wholesalers out of his part ofBrazil, he managed the Hyacinth Macaw resourcerationally, like a responsible rancher. That is, heonly took babies from his professional providers,and he knew that by not hurting adults, he wasguaranteed a sustainable, never-ending harvest ofhealthy, happy wild babies. He also knew that hiscliff-climbing trappers could not reach more thanperhaps 50% of all wild nests of the species, andexperience showed that each year, the same pairsof Hyacinths would lay eggs and hatch young inthe same cliff cavities, despite the depredationsby his band of pro climbers. This system ensureda steady, never-ending supply of high-quality,parent-hatched, parent-fed Hyacinth nestlings,which his team would harvest in their third monthof life—when they were large and strong andwould survive the harvesting very well.
Lima reports that using these methods, histeam never lost a single baby Hyacinth, and hisfather and he sold perhaps 1,000 large Hyacinthbabies between 1976 and 1994, when he stoppedtrading and started protecting all wildlife in hisregion as a paid warden, paid by funds suppliedby me and by the Bird Clubs of Virginia. In this18-year period, the Hyacinth population nevershowed any sign of decline.
28 Manual of Parrot Behavior

I wish I could say that there are many examplesof such informal but effective “harvest conces-sions” that produced incentives for rational man-agement such as practiced by Lima. But to this day,I have never heard of or seen another rational, sus-tainable example of macaw or parrot harvest any-where else on Earth. And I have heard of more than100 or 200 examples of unsustainable, “tragedy ofthe commons” examples of the annihilation ofparrot populations in Indonesia to Central Africa,Mexico, Central America, and South America.
If we could find ways to experiment withLima’s system in different parts of the globe, itmight be possible to get it right, as Lima got itright. During the entire time that he and his fatherwere trapping and trading thousands of macawsfrom that part of Brazil, the practice was illegal.Nevertheless, no one ever caught them or evenmade a serious attempt to catch them, and theymanaged their Hyacinth populations sustainably.Weird but true—an example of a successful expe-riment in sustainability.
The only reason that Lima started working forme as a parrot protector instead of trader is that bythe mid-90s, law enforcement in Brazil was final-ly catching up with parrot trappers, and Limarealized that he could not continue for more thana few more years without having a nasty run-inwith the law. His major buyer had gone to prisonfor wildlife dealing, and he realized that the jigwas up and he had to look for other options.Fortunately, in my work for the Wildlife Conser-vation Society I long had been interested in con-verting Lima to the “good side of the force,” as itwere. I had met him in 1987 while investigatingtraffic in Hyacinths, and he was “the Man.” By1994, however, with law enforcement breathingdown his neck, I was able to convert him. He hasnever gone back.
Despite the fact that he has renounced birdtrapping, there is no question in my mind that hewas the first and perhaps only sustainable har-vester of Hyacinths in the world, if not the world’sfirst and only documented case of a sustainableharvester of any large, commercially valuableparrot. He has a lot to teach field conservationistsand writers of environmental laws. He consideredthat brutalizing or catching adult breeding birdswas a cretinous, destructive practice, yet mostarmchair theorists on the topic of parrot quotasand trade still espouse standard catch and export
quotas, regardless of the resulting “tragedy of thecommons” race to remove from the forest ALLwild parrots of any age.
Parrot trade normally is tragic and destructiveto wild populations of commercially valuablewild parrots. The only way to change this andmake wild parrots a force for habitat conservationand rural development is to develop systems thatgrant unique, enforceable concessions of parrothabitat to individuals, clans, or organized com-munities so that they can ranch and pamper theirown wild populations of valuable parrots. Such asystem would have to include independent moni-toring, as few tropical countries currently haveadequate abilities to police remote parrot produc-tion areas. The details do not have to be workedout here, as they are beyond the scope of thischapter. Suffice it to say that parrot trade current-ly is out of control virtually everywhere on theplanet except where it is either too difficult to getthe birds out to markets (e.g., remote parts of theAmazon or the interior of New Guinea) or wherethe local cultures value wild birds more in thewild than as trade goods to be sold as pets. Theonly places that seem to cherish wild parrot pop-ulations in the wild and prohibit trapping areAustralia, New Zealand, Gabon, and parts ofCosta Rica, Peru, Ecuador, and Brazil. In the firstthree countries, cultural norms prohibit parrottrapping, while in the latter four cases, parrot eco-tourism has become significant business and sohas created a local conservation culture thatdiverges from the national norm.
Unfortunately, the areas that protect wild par-rots and prevent all trade in large parrots areunlikely to amount to more than 10% of theremaining major tropical forest parrot habitataround the globe, but we would like to protectALL remaining tropical forests. Therefore, weshould experiment with methods for making theparrots in the rest of the world’s tropical forestsworth more alive and free flying in the wild thantrapped to extinction decades before their foresthome is in any significant danger. Sustainablyranched wild parrots might provide an option formaintaining forest cover and rural livelihoods in20–30% of the remaining tropical forests of theworld. The sooner we start systematic experi-ments with sustainable ranching of wild parrots,the sooner we can stop all the current wasteful,unsustainable, and inhumane quote-based trade.
3 / Parrot Conservation, Trade, and Reintroduction 29

The World Parrot Trust’s proposal to ban parrotimport into the European Union is a very usefulmove, as would be experiments with methodssuch as those pioneered by Lourival Lima in the1970s and 1980s in Brazil.
REINTRODUCTION
It is obvious to anyone who travels to Palm Beachor Miami, Florida, to parts of San Francisco orother parts of coastal California, to Lima, Peru, orhundreds or thousands of other towns and citiesaround the tropics, subtropics, and milder parts ofthe temperate latitudes that many species of par-rots have become reintroduced and are flourish-ing. These birds all were inadvertently “reintro-duced” by release or escape of cage birds. Sothere is no question that parrots can be reintro-duced. This truth, however, does not contradictthe fact that not all of the hundreds of species ofparrots will be equally easy to reintroduce. Infact, reintroduction should rarely, if ever, be amethod that one should have to rely on to savewild populations of the very rarest parrots.
The reasons for this are mostly economic.Unless a wild species is down to a population ofless than 100 wild birds and less than five or tenwild nests, the option of reintroducing captive-bred individuals normally will prove to be a muchmore expensive or riskier way of increasing wildparrot populations than simply increasing the out-put from existing wild nests or translocating wildbirds from other locations.
A good example of a parrot species thatdeserves a major effort in reintroduction is theSpix’s Macaw. The species went extinct in thewild in 2000 when the last wild bird perished, yetthere are more than 70 birds in captivity, most ofwhich are very inbred and for whom the studbookdata have not been made available by the privateowners. If the private owners of Spix’s Macawcannot organize themselves to reintroduce theSpix’s Macaw to the wild, then what hope is therethat any other private owners of rare parrots willever contribute captive birds to a future reintro-duction effort for a similarly endangered parrotspecies?
Possibly the Blue-throated Macaw qualifies fora reintroduction effort, but so far, only nonprofit,public-interest conservation organizations andtheir board members have shown interest indeveloping a technically valid reintroduction
project for this species. Private owners of thespecies have not come forward to help.
The most successful parrot recovery effort inhistory is that of the Echo Parakeet, which hasrecovered from less than 20 birds 15 years ago tomore than 200 birds in the wild now, all thanks tothe generous work of a number of nonprofit con-servation groups such as the Jersey WildlifePreservation Trust and the World Parrot Trust.But none of the birds used in this successful rein-troduction effort came from private bird ownersor collectors. Rather, they were bred at the non-profit Wildlife Preservation Trust or, more typi-cally, produced by assisting the fledging of wild-laid eggs in wild nests. In fact, work with theEcho Parakeet in Mauritius, and with Scarlet andGreen-winged Macaws in Manu and Tambopata,Peru, and in Costa Rica, has shown that the bestway to increase the local parrot population in awild setting is to help all wild-laid eggs survive tohatch and fledge healthy young. The methods fordoing this may involve assisting eggs or babies inwild nests, or temporarily removing wild eggs orwild babies to field-based labs where they can beassisted to fledge into adjacent, appropriate habi-tat as soon as possible, thus minimizing their tem-porary captivity.
Another notably successful parrot reintroduc-tion project was that of the Margarita Amazon inVenezuela, a project that reintroduced hundredsof birds confiscated from trappers and traders.With very few quarantine precautions and with aminimum of fuss and muss, it appears that mostof these reintroduced birds flourished and becameactively reproductive, thus increasing the wildpopulation by severalfold in just a few years.Persons who are wary of disease risks caution thatthis case may have been handled less cautiouslythan is appropriate according to the best practicesof modern avian veterinarians, but then it also ispossible that many diseases seen in captive par-rots may be a manifestation of captivity itselfrather than a problem for wild birds. In otherwords, perhaps wild birds are largely free of clin-ical signs of many diseases that are present in thewild, while captive birds become ill and often diefrom the same diseases, with captivity itself beingthe culprit rather than the pathogen, per se. It isimportant to examine the assumptions surround-ing pathogens in captivity and in the wild beforeplanning a major release effort for captive birds.
30 Manual of Parrot Behavior

It also is notable that, without taking any specialprecautions whatsoever, wild parrots have reintro-duced themselves successfully in so many thou-sands of tropical, subtropical, and temperate loca-tions around the globe. It would be a fertileresearch field for avian veterinarians to investi-gate which diseases are caused by or are exacer-bated by captivity.
The best example of a reintroduction projectgone awry may have been the attempt to reintro-duce naive, lab-raised Thick-billed Parrots insouthern Arizona. In this case, the project suf-fered from a number of problems, namely the factthat the Thick-billed Parrot is among the most dif-ficult of parrots to reintroduce and the reintroduc-tion area was plagued with a high density of hun-gry Red-tailed Hawks happy to eat reintroducedbirds. The Thick-bill is among the hardest of allparrots to reintroduce because it has a very spe-cialized feeding ecology and normally lives intight flocks that fly each day at high speed forenormous distances. Thus, in order to blend into awild flock and so not be singled out and attackedby hungry hawks, released Thick-bills need to beable to extract pinecone seeds at a phenomenallyfast rate and then blend perfectly into a flock as ittwists and turns in high-speed, marathon dailyflights. The only reintroduced Thick-bills to havenotable survival success after release were wild
birds that were captured in forests in Mexico andquickly released in the target forest in southernArizona. These wild birds knew how to eat effi-ciently and fly fast and precisely, and so were ableto survive in the face of the major hawk popula-tions in that part of the United States.
A notable advantage of using a variety of tech-niques to increase the productivity of wild nestsand wild populations is that you will normally notneed to take very elaborate, expensive precau-tions to prevent disease transmission, as thesetechniques involve either no or very little time incaptivity, and even when in captivity, the birds arekept in single-species, dedicated facilities close toor in the wild habitat.
Thus, normally, reintroduction of captive-bredparrots should be avoided unless there are few orno wild birds left to work with. It will be muchless expensive, much easier, and much more suc-cessful to assist wild populations to increase asfast as possible (as was the case with the EchoParakeet). Working to help wild populationsincrease their reproductive success avoids all thecomplications and expense of elaborate quaran-tines for captive-raised birds. Additionally,captive-bred birds often will be naive and at highrisk in the wild when compared with savvy, wild-born or wild-trained birds.
3 / Parrot Conservation, Trade, and Reintroduction 31

INTRODUCTION
Parrots are gregarious and vocal creatures thatcommunicate in ways we have yet to understand.How do parrots perceive the world? By under-standing some of the unique adaptations of aviananatomy, we may better understand parrot behav-ior. This chapter will discuss the sensory capaci-ties of parrots including vision, hearing, taste,smell, and touch perceptions.
VISION
The majority of birds rely heavily on visual abili-ties in their daily activities. Visual acuity isenhanced in avian species, approximately two toeight times higher than in mammals, as the avianeye is large in relation to the size of the head,allowing a large image to be projected on the reti-na.[1–5] Visual acuity is also enhanced becausethe retina of diurnal birds has a large number ofcones compared to humans; for example, thehawk fovea contains around 300,000 cones/mm2,while the human fovea contains around 147,000cones/mm2.[1] In addition, nearly every cone inthe avian eye is represented by an individual axontraveling to the brain, while the eye of humanscontains six to seven million cones but only onemillion axons in the entire optic nerve.[1]
While eyes come in different shapes dependingon the species of bird, parrots have a “flat” eye.The flat eyeball is characterized by a short axisthat projects a relatively smaller image on the reti-na, decreasing visual acuity compared to otherspecies such as birds of prey.[1] The eyeball ofbirds is asymmetric, favoring binocular vision.[1]
The sclera of the eye is strengthened by ten to 18small bones called scleral ossicles.[1–5]
Because the eyeball almost completely fills theorbit, the eye movements of the bird are generallyfewer than those of mammals.[1] However, birdscan move their heads and necks extensively, andthis compensates for the small eye movements.[1]Movement of the orbits is independent betweenboth eyes in parrots, in contrast to mammals.[1]
A feature of the avian eye is that the sphincterand dilator muscles of the pupil contain mainlystriated fibers, compared to the mammalian coun-terpart that contains only smooth muscle.[1–5]Because of this anatomic feature, the pupillaryopening is under voluntary control in parrots.Rapid dilation and constriction of the pupillaryopening is often observed in aggressive or excit-ed parrots.[6] While pupillary light reflexes dooccur in birds, complete decussation of the opticnerve axons prevents true consensual pupillarylight reflex.[2, 3] The iris is the colored part of theeye that contains chromatophores that can createvarying iris colors based on age, gender, andspecies of the parrot.[1–3]
Unlike the mammalian counterpart, the avianretina is devoid of blood vessels, which decreasesscattering of light and shadows.[5] The pecten isa unique vascular structure found only in theavian eye in association with the retina. The func-tion of the pecten is likely to provide nutrition tothe eye, as retinal vessels are lacking.[1, 5, 7]
Parrots often turn their head or body sidewayswhen presented with a new toy or object. Be-havioral studies in many birds have shown that
33
4Sensory Capacities of Parrots
Jennifer Graham, Timothy F. Wright, Robert J. Dooling, and Ruediger Korbel

they prefer the use of a lateral and monocularfield to observe distant objects.[5, 8–12] Basedon monocular data in pigeons, visual resolution ishigher in the lateral field than frontal field, thusexplaining this preference.[5, 13]
The lens of the avian eye is softer than that ofmammalian species.[1] Unlike the yellow-tintedmammalian lens that filters out wavelengths oflight below 400 nm, the clear avian lens transmitswavelengths below 400 nm.[1] Colored oildroplets on the ends of the cones provide protec-tion against the effects of ultraviolet (UV)light.[5, 14] Birds are able to see UV light below400 nm due to the combined effects of cone oildroplets and visual pigments.[4, 5, 15] Whiletrichromatic color vision in humans is based onthree colors (blue, green, and red), the tetrachro-matic, or pentachromatic in some avian species,system of birds includes UV, fluorescent, blue,green, and red.[4, 16–24] UV perception of par-rots likely plays an important role in behavior.Many parrots’ feathers reflect UV and studieshave shown that UV reflection of feathers affectsmate choice (see Plates 1 and 2 in color sec-tion).[4, 16, 25–29] While some parrots are notvisibly sexually dimorphic to the human eye, UVreflection from plumage and skin varies betweensexes of some birds.[4] Some types of fruits andberries, such as kaki, green grapes, and figs,reflect UV light and ripeness of the food may bedetermined by this characteristic.[4, 30] Certainflower patterns, insects, and urine and feces ofrodents also reflect UV light that can be detectedby birds.[4, 30–32] Additionally, highly UV-reflective areas within the oral cavity play animportant role in triggering reflexes to feed youngbirds that demonstrate their oral cavity to theirparents. Birds may use UV receptors in combina-tion with color receptors for navigation by detect-ing sun-based color gradients.[33–35] Fluores-cence, which occurs when short wavelength lightis absorbed and re-emitted at a longer wave-length, occurs on parrot feathers and may be animportant avian signaller.[25, 36]
Birds are able to detect a spatial frequency ofaround 160 frames/second or hertz (Hz), com-pared to 50–60 Hz in humans.[37–39] Becausemost artificial lights produce noncontinuous lightat a frequency of around 100–120 Hz, a strobo-scopic effect not detectable to humans results andmay be detrimental to birds.[4, 39–41] In addi-
tion, artificial lights and sunlight passing throughwindows do not provide full-spectrum light.While studies are currently under way to examinethe effects of artificial lights on birds, current rec-ommendations have been made to provide full-spectrum light and high frequency sources thatemit continuous light.[37] Suggestions have alsobeen made to consider light source and presenceof full-spectrum light when performing ethologi-cal studies.[17, 37, 42, 43] Because video or com-puter monitors have refresh rates of around 50–95Hz, welfare issues may arise when performingvideo playback experiments in birds.[39, 44–46]
Familiarity with the unique anatomic and phys-iologic variations of the avian eye compared tothat of mammals is important when assessingbehavior alterations in parrots. Behavior changes,such as reluctance to fly or step onto an extendedhand, abnormal head posture, inappetence, andothers, can certainly result from ocular abnormal-ities. In addition, permanent ocular problems,such as blindness resulting from cataract forma-tion, are a common occurrence in parrots and canbe managed in such a way as to maintain qualityof life. Provision of full-spectrum lighting, nor-mal light cycles, and continuous-emitting lightsources should be considered when addressingbehavioral problems in birds.
HEARING
Birds rely on their hearing ability for detectingpredators and prey, orienting in the environment,and communicating with conspecifics. The songsand calls produced by birds are among the mostcomplex auditory signals known,[47] and thiscomplexity has generated much interest in howbirds hear sounds.[48] In the case of parrots,many of these vocalizations appear to be learnedthrough experience,[49] which has led to furtherinterest in the connections between perception,learning, and vocal production.
The anatomy of the avian ear presents somemarked contrasts to the more familiar mammalianear. These differences include the absence of anexternal ear; a single middle ear bone, the col-umella, in place of the three bones found in mam-mals; and the much shorter sensory auditoryepithelium in the inner ear. In Budgerigars, forinstance, the sensory surface of the inner ear, thebasilar papilla, is about 3–4 mm in length (com-pared to around 30 mm in humans). The columel-
34 Manual of Parrot Behavior

lar middle ear and the short auditory sensoryepithelium in birds probably both exert limita-tions on the range of hearing in birds compared tomammals.[50] Another interesting differencebetween birds and mammals is in the organizationof sensory hair cells on the auditory epithelium.Mammals typically have one row of inner haircells and three rows of outer hair cells across thewidth of the auditory epithelium, while birdsshow more rows of hair cells and considerablevariation in the structure and orientation of thesehair cells.[51] The functional consequence ofthese differences remains obscure. But, in strikingcontrast to mammalian hair cells, avian hair cellsare known to be capable of regenerating afterdamage caused by exposure to excessive noise orototoxic drugs.[52, 53] Here the functional conse-quences are enormous. Birds regain their hearingwhen their hair cells regenerate. Many forms ofhuman deafness are related to defects in or loss of hair cell function,[54] and thus the discovery ofhair cell regeneration in birds has spurred arenewed interest in avian ear anatomy.
The anatomical complexity of the bird ear isnot fully understood and has led to much interestin how well birds are able to detect, discriminate,and learn complex sounds. We are fortunate toknow a great deal about the hearing in one parrotspecies, the Budgerigar, because of its small sizeand tractability in the laboratory for behavioralstudies of hearing. Less is known of the hearingabilities in other parrot species, but what isknown suggests that many abilities of theBudgerigar are shared across species. The behav-ioral methods used for studying hearing involveoperant conditioning or training the bird torespond to a sound—or the change in a sound—by pecking a switch in order to obtain food.[55]These methods have been highly successful andhave been used in a wide range of studies exam-ining how parrots and other birds detect sounds,discriminate among similar sounds, and classifysounds into perceptual categories.
One of the most basic measures of hearingabilities is the audiogram. The audiogram is a plotof the least detectable amount of sound energy abird can hear in the quiet at different frequenciesover its range of hearing. Figure 4.1 shows audio-grams for three parrots—the Budgerigar, theCockatiel, and the Orange-fronted Conure. Theseaudiograms show that these parrots, like many
bird species, hear best at frequencies betweenabout 1 and 5 kHz and less well at frequenciesbelow about 500 Hz and above 10 kHz. The low-est threshold approaches 0 dB in the Budgerigar,5 dB in the Cockatiel, and 20 dB in the Orange-fronted Conure.[56, 57]
In all three species these lowest thresholds inthe quiet occur at frequencies between 2 and 4kHz. This is also the frequency range in whichmost of the acoustic energy is found in their mostcommon type of vocalizations, the contactcall.[57, 58] Contact calls are probably designedfor distance communication under more noisyconditions than found in the laboratory. Interest-ingly, when hearing thresholds are measured inthe presence of a masking noise, Budgerigars,Cockatiels, and Orange-fronted Conures alsoshow the best signal-to-noise ratios in this samefrequency region. These signal-to-noise ratios(called critical ratios) are shown in Figure 4.2.These critical ratio functions show the level (indecibels) above the background noise that asound must be in order to be heard. Most birdsshow a pattern like the Cockatiel; that is, criticalratios increase monotonically at roughly 3 dB forevery octave increase in frequency. The Budgeri-gar and Orange-fronted Conure, by contrast,show a 5–10 dB increase in sensitivity between 2and 4 kHz relative to the typical avian critical
4 / Sensory Capacities of Parrots 35
Figure 4.1. Hearing thresholds under quietconditions for three species of parrot. Figureredrawn from Wright et al. (2003).

ratio function.[56, 57] While the function of thisincreased sensitivity in these species is uncertain,it is intriguing to note that it corresponds well tothe frequency range of maximum energy in theircontact calls and may help in discriminatingamong different calls within large noisy flocks.
Birds in general, with the exception of noctur-nal predators such as the Barn Owl, are not verygood at localizing sound. Because of their smallheads and closely spaced ears, the time differenceor intensity difference between sounds arriving atthe two ears of a bird is negligible. One parrot, theBudgerigar, has been tested in the laboratory andminimum audible angles are in the range of22–52 degrees for pure tones and 24 degrees forbroadband sounds such as noises and vocaliza-tions.[59]
Hearing is much more than the detection orlocalization of sounds. In order to communicate,an animal also must be able to discriminateamong different sounds with potentially very dif-ferent meanings. The complex temporal and fre-quency structure of many bird vocalizations haslong prompted suspicions that birds may haveparticularly good abilities to detect small differ-ences in frequency, amplitude, and temporal char-acteristics of sound. In some cases this prediction
is borne out, while in other areas the abilities ofbirds are very similar to those of mammals andother terrestrial animals. For example, studies offrequency discrimination in the Budgerigar andthe Orange-fronted Conure have shown that, likemost birds, they are able to discriminate amongtones that differ by about 1% of their frequen-cy.[57, 60] This threshold is roughly in the rangeof humans and other animals that have been test-ed. In contrast, these parrots are worse thanhumans at discriminating differences in the inten-sity of two tones; humans can discriminate a 1 dBdifference in the intensity of pure tones, whilebirds, including Budgerigars and Orange-frontedConures, typically require a difference of 2–5dB.[57, 61] One can imagine that in discriminat-ing vocalizations in the real world, frequency cuesmight be far more reliable than intensity cues dueto degradation of signals during transmissionthrough the environment, so perhaps this is onereason that intensity discrimination abilities areless well developed.
The detection abilities measured using puretone stimuli may not be perfect predictors of theability of parrots to distinguish among complexspecies-specific calls. Several studies have exam-ined the ability of parrots to discriminate amongand classify their contact calls. One study com-pared the abilities of Budgerigars and ZebraFinches to detect the presence of contact calls ina noisy background, and compared these thresh-old levels to those found when birds were askedto discriminate among the same calls in the pres-ence of noise.[62] Thresholds were 2–5 dB lowerfor detection than for discrimination among thesame calls, suggesting that discrimination is amore difficult task requiring more of the informa-tion in the calls to be clearly perceived.
A second study compared the ability of Bud-gerigars, Zebra Finches, and Canaries to discrim-inate among a set of stimuli including four con-tact calls from each species.[63] All three specieshad more difficulty discriminating between callsfrom the same species than between calls fromdifferent species. Furthermore, all three speciescould discriminate more easily between callsfrom their own species than between two callsfrom a different species. These results suggestthat discrimination is more difficult when callsare acoustically more similar (i.e., from the samespecies) but that different species may have spe-
36 Manual of Parrot Behavior
Figure 4.2. Hearing thresholds under noisyconditions for three species of parrot. Thresholdsare given as the critical ratio between signal leveland the masking noise at the threshold of detec-tion. Figure redrawn from Wright et al. (2003).

cial hearing abilities that aid in the perception oftheir own calls. Such specializations may ariseeither through innate differences in auditorycapabilities or through learned preferences devel-oped through selective exposure to conspecificsounds as nestlings or fledglings.
A third study examined the ability of theOrange-fronted Conure to form perceptual cate-gories for different individuals based on theacoustic properties of their calls (T. Wright, K.Cortopassi, J. Bradbury, and R. Dooling, unpub-lished data). Subjects listened to a repeating back-ground of ten calls from a single individual inter-spersed with calls from different individuals.Subjects quickly learned to avoid responding tothe differences between different renditions of thecontact call by a single individual and to respondto the differences among calls of different indi-viduals. Their ability to learn this distinction rap-idly suggests that they are able to form perceptu-al categories for the calls of different individualsthat allow them to focus on those acoustic fea-tures that reliably differ between different individ-uals. Such perceptual abilities may be critical foracoustic recognition of a variety of social levels inparrots, including individuals, pairs, flocks,roosts, and geographic regions.
TASTE AND SMELL
Taste buds lie on the tongue base in most of theavian species studied such as the chicken, pigeon,swift, raptor, and songbird.[1, 64, 65] In parrots,taste buds are found along the choanal opening onthe roof of the oropharynx in association withsalivary glands.[1, 65] Compared to mammals,birds have a poor sense of taste; while humanshave around 9,000 taste buds, parrots are estimat-ed to have 300–400.[66, 67] Parrots have a high-er number of taste buds than most other avianspecies, such as the chicken with 250–350 and thepigeon with only 37–75.[64, 66, 67] Despite thelow number of taste buds found in birds, manystudies have shown that flavors can affect foodchoice and quantity consumed.[66, 68–76] Whileit has been stated that most birds easily detectsalts and acids but sweet substances are not effec-tive stimuli, the response to different flavorsvaries widely among birds.[1, 76] Some parrotsand Budgerigars, as well as other birds, have beenshown to prefer sugar solutions over water.[67]Studies in captive Cockatiels examined threshold
and preference for water, sodium chloride, potas-sium chloride, sucrose, glucose, fructose, sodiumphosphate buffer, and citric acid buffer solu-tions.[66, 76] In the Cockatiel studies, all testedcompounds added to the water resulted in de-creased consumption of the test solution and in-creased consumption of pure water. No test com-pound was preferred by the Cockatiels.[66, 76]While future study is needed to determine the sig-nificance of taste preference in parrots, there isno question that taste plays a role in food accept-ance and avoidance.
The receptors of the nasal cavity that detectodor are generally located on the caudal nasalconchae.[1] Receptor nerve fibers run from theconchae olfactory epithelium to an area withinthe brain called the olfactory bulb, which is rela-tively small in the parrot compared to other avianspecies.[1] Interestingly, the avian orders with rel-atively small olfactory bulbs have high olfactorythresholds.[77, 78] Compared to mammals suchas man, dogs, and rats, birds have proven to havecomparable olfactory capacities in conditioningstudies.[77, 79–83] Although research intopsittacine olfactory abilities is scarce, variousavian species use olfactory cues for food location,orientation and navigation, returning to nest sites,reproduction and parenting, and selection of nestmaterial.[81, 84–89]
TOUCH
There are many types of sensory receptors,including those for touch, heat, and pain, locatedwithin the parrot beak and skin that give the birdmore information about its environment. The dif-ferent types of touch receptors, or mechanorecep-tors, in birds are Herbst corpuscles, Merkel cellreceptors, Grandry corpuscles, and Ruffini end-ings.[90] Herbst corpuscles, which are vibration-sensitive, are the most numerous skin receptorsand are found in the beak, leg, and skin.[90, 91]Because they lie in close association with featherfollicles and muscles associated with the follicles,Herbst corpuscles relay feather position in rela-tion to the body. Merkel cells are found mainly inthe beak of non-aquatic birds, while Grandry cor-puscles are present in aquatic birds; both arenumerous in the bill tip organ that is important forfood prehension.[90, 91] While Ruffini’s cor-puscles can be found in joint capsules of birds,Ruffini endings have only been identified in
4 / Sensory Capacities of Parrots 37

the bill of geese and the beak of the Japanesequail.[90, 92–94] Mechanoreceptors are involvedwith behavioral responses, including the initiationof a feeding response in baby birds upon beakmanipulation and the ability of parrots to manip-ulate food with their beak and feet.[90] The sen-sitive mechanoreceptors in the feet of parrots mayallow them to feel earthquakes that are unde-tectable to owners.[90, 95] Disorders of theplumage may be detected by mechanoreceptorsand stimulate preening behavior.[90, 96] Flightcontrol and patterns may be regulated bymechanoreceptors detecting vibrations caused byair stream turbulence.[90, 97, 98]
Avian thermoreceptors may be free nerve end-ings and are present in the skin, especially thebeak and tongue.[90, 99] Thermoreceptors in theskin assist with body thermoregulation and thosein the beak may be used for regulating incubationtemperatures in some birds.[100] Pain receptors,or nociceptors, respond to mechanical and ther-mal stimuli and are present in the beak andskin.[90, 101, 102] Research indicates that birdsrespond to pain by either a reflex/escape responseor by immobility; these responses may be mediat-ed by different types of pain receptors.[103] Inaddition, beak amputation studies show that birdsmay experience chronic pain.[103]
CONCLUSION
Parrots experience the world in ways both similarand different to mammals. It is apparent thatvision, hearing and vocalization, taste, olfaction,and touch perception play vital roles in the dailylife of the parrot. Many of the normal and abnor-mal behaviors of parrots can be better understoodby examining how birds perceive the environmentaround them. Further research in the area of sen-sory perception of parrots will expand our knowl-edge and likely enable us to improve the lives ofthese magnificent creatures.
REFERENCES
1. King, A., and J. McLelland, 1984. “Special senseorgans.” In Birds, their structure and function, ed.A. King and J. McLelland, pp. 284–314. Phila-delphia: Balliere-Tindall.
2. Williams, D. 1994. “Ophthalmology.” In Avianmedicine: Principles and application, ed. B.W.Ritchie, G.J. Harrison, and L.R. Harrison, pp.673–694. Lake Worth, FL: Wingers Publishing.
3. Kern, T.J. 1997. “Disorders of the special senses.”In Avian medicine and surgery, ed. R.B. Altman,S.L. Clubb, G.M. Dorrestein, and K. Quesen-berry, pp. 563–589. Philadelphia: WB Saunders.
4. Korbel, R.T. 2000. “Avian ophthalmology: A clin-ically oriented approach.” Proceedings of theAssociation of Avian Veterinarians, pp. 439–456.
5. Gunturkun, O. 2000. “Sensory physiology:Vision.” In Sturkie’s avian physiology, ed. G.C.Whittow, pp. 1–19. San Diego: Academic Press.
6. Blanchard, S. 1999. The companion parrot hand-book. Oakland: PBIC, Inc., p. 162.
7. Korbel, R.T., B. Nell, P.T. Redig, I. Walde, and S.Reese. 2000. “Video fluorescein angiography inthe eyes of various raptors and mammals.” Pro-ceedings of the Association of Avian Veterina-rians, pp. 89–95.
8. Blough, P.M. 1971. The visual accuity of thepigeon for distant targets. Journal of the Experi-mental Analysis of Behavior 15:57–67.
9. Bischof, H.J. 1988. The visual field and visuallyguided behavior in the zebra finch (Taeniopygiaguttata). Journal of Comparative Physiology163:329–337.
10. Fox, R., Lehmkuhle S.W., and D.H. Westendorf.1976. Falcon visual acuity. Science 192:263–265.
11. Friedman, M.B. 1975. “How birds use their eyes.”In Neural and endocrine aspects of behaviour inbirds, ed. P. Wright, P. Cryl, and D.M. Volwes, pp.182–204. Amsterdam: Elsevier.
12. Reymond, L. 1985. Spatial visual accuity of theeagle Aquila audax: A behavioral, optical, andanatomic investigation. Vision Res 25:1477–1491.
13. Gunturkun, O., and U. Hahmann. 1994. Cerebralasymetries and visual acuity in pigeons. BehavBrain Res 60171–175.
14. Emmerton, J. 1983. “Vision.” In Physiology andbehavior of the pigeon, ed. M. Abs, pp. 245–266.London: Academic Press.
15. Chen, D.M., and T.H. Goldsmith. 1986. Fourspectral classes of cone in the retinas of birds. JComp Physiol 159:473–479.
16. Burkhardt, D. 1989. UV vision: a bird’s eye viewof feathers. Journal of Comparative PhysiologyA164:787–196.
17. Cuthill, I., J.C. Partridge, and A.T. Bennett. 2000.“Avian UV vision and sexual selection.” In Ani-mal signals. Signalling and signal design in ani-mal communication, ed. Y. Epsmark, T. Amund-sen, and G. Rosenqvist, pp. 87–106. Trondheim,Norway: Royal Norwegian Society of Sciencesand Letters, The Foundation Tapir Publishers.
18. Cuthill, I., J.C. Partridge, A.T. Bennett, S.C.Church, and N.S. Hart. 2000. Ultraviolet vision inbirds. Adv Study Behav 29:159–214.
38 Manual of Parrot Behavior

19. Bennett, A.T., I. Cuthill, and K. Norris. 1994.Sexual selection and the mismeasure of color. AmNat 144:848–860.
20. Osorio, D., M. Vorobyev, and C.D. Jones. 1999.Colour vision of domestic chicks. J Exp Biol202:2951–2959.
21. Osorio, D., C.D. Jones, and M. Vorobyev. 1999.Accurate memory for colour but not pattern con-trast in chicks. Curr Biol 9:199–202.
22. Vorobyev, M., D. Osorio, A.T. Bennett, N.J.Marshall, and I. Cuthill. 1998. Tetrachromacy, oildroplets and bird plumage colors. J Comp PhysiolA183:621–633.
23. Bowmaker, J.K., L.A. Heath, S.E. Wilkie, andD.M. Hunt. 1997. Visual pigments and oil drop-lets from six classes of photoreceptors in the reti-nas of birds. Vis Res 37:2183–2194.
24. Wilkie, S.E., P.M. Vissers, D. Das, W.J. DeGrip,J.K. Bowmaker, and D.M. Hunt. 1998. Themolecular basis for UV vision in birds: Specialcharacteristics, cDNA sequence and retinal local-isation of the UV-sensitive pigment of thebudgerigar (Melopsittacus undulatus). Biochem J330:541–547.
25. Pearn, S.M., T.D. Andrew, A.T. Bennett, and I.Cuthill. 2001. Ultraviolet selection, fluorescenceand mate choice in a parrot, the budgerigarMelopsittacus undulatus. Proc R Soc Lond268:2273–2279.
26. Burkhardt, D., and E. Finger. 1991. Black, white,and UV: How birds see birds. Naturwissen-schaften 78:279–280.
27. Cuthill, I., A.T. Bennett, J.C. Partridge, and E.Maier. 1999. Plumage reflectance and the objec-tive assessment of avian sexual dichromatism. AmNat 160:183–200.
28. Finger, E., and D. Burkhardt. 1994. Biologicalaspects of bird colouration and avian colourvision including ultraviolet range. Vis Res34:1509–1514.
29. Finger, E., D. Burkhardt, and J. Dyck. 1992.Avian plumage colors: Origin of UV reflection in a black parrot. Naturwissenschaften 79:187–188.
30. Burkhardt, D. 1982. Birds, berries, and UV. Anote on some consequences of UV vision inbirds. Naturwissenschaften 69(4):153–157.
31. Viitala, J., E. Korplmakl, P. Palokangas, and M.Kolvula. 1994. Attraction of kestrels to vole scentmarks visible in ultraviolet light. Nature373:425–427.
32. Chittka, L., A. Shmida, N. Troje, and R. Menzel.1994. Ultraviolet as a component of flower reflec-tions, and the color perception of Hymenoptera.Vis Res 34(11):1489–1508.
33. Bradbury, J.W., and S.L. Vehrencamp. 1998.“Light signal reception.” In Principles of animalcommunication, ed. A.D. Sinauer, pp. 239–278.Sunderland, MA: Sinauer Associates, Inc.
34. Bennett, A.T., and I. Cuthill. 1994. Ultravioletvision in birds: What is its function? Vis Res34:1471–1478.
35. Coemans, M.A., J.J. vos Hzn, and J.F.W. Nuboer.1994. The relation between celestial colour gradi-ents and the position of the sun, with regard to thesun compass. Vis Res 34:1461–1470.
36. Arnold, K.E., I.P. Owens, and N.J. Marshall. 2002.Fluorescent signaling in parrots. Science 295:92.
37. Korbel, R.T., and U. Gropp. “Ultraviolet percep-tion in birds.” Proceedings of the Association ofAvian Veterinarians, pp. 77–81.
38. Brundett, G.W. 1974. Human sensitivity to flick-er. Lighting Res Technol 6:127–143.
39. Maddocks, S.A., A.R. Goldsmith, and I. Cuthill.2001. The influence of flicker rate on plasmacorticosterone levels of European starlings,Sturnus vulgaris. Gen Comp Endocrinol 124(3):315–320.
40. Boshouwers, F.M.G., and E. Nicaise. 1992.Artificial light sources and their influence onphysical activity and energy expenditure of layinghens. Brit Poultry Sci 34:11–19.
41. Nuboer, J.F.W. 1993. “Visual ecology in poultryhouses.” Proceedings of the Fourth Symposiumof Poultry Welfare, Edinburgh, Scotland, pp.39–44.
42. D’Eath, R.B., and M.S. Dawkins. 1996. Layinghens do not discriminate between video images ofconspecifics. Anim Behav 52:903–912.
43. Hunt, S., I. Cuthill, J.P. Swaddle, and A.T.Bennett. 1997. Ultraviolet vision and band-colour preferences in female zebra finches,Taeniopygia guttata. Anim Behav 54:1383–1392.
44. D’Eath, R.B. 1998. Can video images imitate realstimuli in animal behaviour experiments? BiolRev 73:267–292.
45. Fleishman, L.J., and J.A. Endler. 2000. Somecomments on visual perception and the use ofvideo playback in animal behaviour studies. ActaEthol 3:15–27.
46. Cuthill, I., N.S. Hart, J.C. Partridge, A.T. Bennett,S. Hunt, and S.C. Church. 2000. Avian colourvision and avian video playback experiments.Acta Ethol 3:29–37.
47. Catchpole, C.K., and P.J.B. Slater. 1995. Birdsong: Biological themes and variations. Cam-bridge: Cambridge Univ. Press.
48. Dooling, R.J., B. Lohr, and M.L. Dent. 2000.“Hearing in birds and reptiles.” In Comparativehearing in birds and reptiles, ed. R.J. Dooling,
4 / Sensory Capacities of Parrots 39

R.R. Fay, and A.N. Popper, pp. 308–359. NewYork: Springer-Verlag.
49. Farabaugh, S.M., and R.J. Dooling. 1996.“Acoustic communication in parrots: Laboratoryand field studies of Budgerigars, Melopsittacusundulatus.” In Ecology and evolution of acousticcommunication in birds, ed. D.E. Kroodsma andE.H. Miller, pp. 97–117. Ithaca, NY: Cornell Uni-versity Press.
50. Saunders, J.C., R.K. Duncan, D.E. Doan, and Y.L.Werner. 2000. “The middle ear of reptiles andbirds.” In Comparative earing: Birds and reptiles,ed. R.J. Dooling, A.N. Popper, and R.R. Fay, pp.13–69. New York: Springer-Verlag.
51. Gleich, O., and G.A. Manley. 2000. “The hearingorgan of birds and Crocodilia.” In Comparativehearing: Birds and reptiles, ed. R.J. Dooling,A.N. Popper, and R.R. Fay, pp. 70–138. NewYork: Springer-Verlag.
52. Ryals, B.M., and E.W. Rubel. 1988. Hair cellregeneration after acoustic trauma in adultCoturnix quail. Science 240:1774–1776.
53. Corwin, J.T., and D.A. Cotanche. 1988. Regen-eration of sensory hair cells after acoustic trauma.Science 240:1772–1774.
54. Steel, K.P., and C.J. Kros. 2001. A geneticapproach to understanding auditory function.Nature Genet 22:143–149.
55. Park, T.J., K. Okanoya, and R.J. Dooling. 1985.Operant conditioning of small birds for acousticdiscrimination. J Ethol 3:5–9.
56. Okanoya, K., and R.J. Dooling. 1987. Hearing inpasserine and psittacine birds: A comparativestudy of absolute and masked thresholds. J CompPsychol 101 (1):7–15.
57. Wright, T.F., K.A. Cortopassi, J.W. Bradbury, andR.J. Dooling. In press. Hearing and vocalizationsin the orange-fronted conure, Aratinga canicu-laris. J Comp Psychol.
58. Farabaugh, S.M., M.L. Dent, and R.J. Dooling.1998. Hearing and vocalizations of wild-caughtAustralian budgerigars (Melopsittacus undula-tus). J Comp Psychol 112:74–81.
59. Park, T.J., and R.J. Dooling. 1991. Sound local-ization in small birds: Absolute localization inazimuth. J Comp Psychol 105:125–133.
60. Dent, M.L., R.J. Dooling, and A.S. Pierce. 2000.Frequency discrimination in budgerigars (Melop-sittacus undulatus): Effects of tone duration and tonal context. J Acoust Soc Am 107(5):2657–2664.
61. Dooling, R.J., and J.C. Saunders. 1975. Auditoryintensity discrimination in the parakeet (Melop-sittacus undulatus). J Acoust Soc Am 58:1308–1310.
62. Lohr, B., T.F. Wright, and R.J. Dooling. In press.Detection and discrimination of natural calls inmasking noise by birds: Estimating the activespace of a signal. Anim Behav.
63. Dooling, R.J., S.D. Brown, G.M. Klump, and K.Okanoya. 1992. Auditory perception of conspe-cific and heterospecific vocalizations in birds:Evidence for special processes. J Comp Psychol106:20–28.
64. El Boushy, A.R., A.F.B. Van der Poel, J.C.J.Verhaart, and D.A. Kennedy. 1989. Sensoryinvolvement control feed intake in poultry.Feedstuffs 61 (25):16–41.
65. Nalavade, M.N., and A.T. Varute. 1977.Histochemical studies on the mucins of the verte-brate tongues. XI. Histochemical analysis ofmucosubstances in the lingual glands and tastebuds of some birds. Acta Histochem 60 (1):18–31.
66. Matson, K.D., J.R. Millam, and K.C. Klasing.2000. Taste threshold determination and side-preference in captive cockatiels (Nymphicus hol-landicus). 2000. Applied Animal BehaviourScience 69:313–326.
67. Kare, M.R., and J.R. Mason. 1986. “The chemicalsenses in birds.” In Avian physiology, ed. P.D.Sturkie, pp. 59–73. New York: Springer-Verlag.
68. Bartholomew, G.A., and T.J. Cade. 1958. Effectsof sodium chloride on the water consumption ofhouse sparrows. Physiol Zool 31:304–310.
69. Cummings, J.L., J.R. Mason, D.L. Otis, J.E.Davis, and T.J. Ohashi. 1994. Evaluation ofmethiocarb, ziram, and methyl anthranilate asbird repellants applied to dendrobium orchids.Wildl Soc Bull 22:633–638.
70. Hainsworth, F.R., and L.L. Wolf. 1976. Nectarcharacteristics and food selection by humming-birds. Oecologia 25:101–113.
71. Harriman, A.E., and M.R. Kare. 1966. Aversionto saline solutions in starlings, purple grackles,and herring gulls. Physiol Zool 39:123–126.
72. Harriman, A.E., and J.S. Milner. 1969. Preferencefor sucrose solutions by Japanese quail (Coturnixcoturnix japonica) in two-bottle drinking tests.Am Midland Natural 81:575–578.
73. Harriman, A.E., and E.G. Fry. 1990. Solutionacceptance by common ravens (Corvus corax)given two-bottle preference tests. Psychol Rep67:19–26.
74. Kare, M.R., and H.L. Pick. 1960. The influenceof the sense of taste on feed and fluid consump-tion. Poult Sci 39:697–706.
75. Kare, M.R., R. Black, and E.G. Allison. 1957.The sense of taste in the fowl. Poult Sci 36:129–138.
40 Manual of Parrot Behavior

76. Matson, K.D., J.R. Millam, and K.C. Klasing.2001. Thresholds for sweet, salt, and sour tastestimuli in cockatiels (Nymphicus hollandicus).Zoo Biology 20:1–13.
77. Mason, J.R., and L. Clark. 2000. “The chemicalsenses in birds.” In Sturkie’s avian physiology, ed.G.C. Whittow, pp. 39–56. San Diego: AcademicPress.
78. Clark, L., and P.S. Shah. 1993. Chemical birdrepellants: Possible use in cyanide ponds. J WildlManage 57:657–664.
79. Davis, R.G. 1973. Olfactory psychophysicalparameters in man, rat, dog, and pigeon. J CompPhysiol Psychol 85:221–232.
80. Clark, L., and J.R. Mason. 1987. Olfactory dis-crimination of plant volatiles by the Europeanstarling. Anim Behav 35:227–235.
81. Clark, L., and C.A. Smeraski. 1990. Seasonalshifts in odor acuity by starlings. J Exp Zool177:673–680.
82. Clark, L. 1991. Odor detection thresholds in treeswallows and cedar waxwings. Auk 108:177–180.
83. Clark, L., K.V. Avilova, and N.J. Bean. 1993.Odor thresholds in passerines. Comp BiochemPhysiol 104:305–312.
84. Houston, D.C. 1987. Scavenging efficiency ofturkey vultures in tropical forests. Condor88:318–323.
85. Clark, L., and P.S. Shah. 1992. “Information con-tent of prey odor fumes: What do foragingLeach’s storm petrels know?” In Chemical sig-nals in vertebrates, ed. R.L. Doty and D. Muller-Schwarze, pp. 421–428. New York: Plenum Press.
86. Wallraff, H.G., J. Kiepenheuer, and A. Streng.1993. Further experiments on olfactory naviga-tion and non-olfactory pilotage by homingpigeons. Behav Ecol Sociobiol 32:387–390.
87. Harriman, A.E., and R.H. Berger. 1986. Olfactoryacuity in the common raven (Corvus corax).Physiol Behav 36:257–262.
88. Cohen, J. 1981. Olfaction and parental behaviorin Ring Doves. Biochem Sys Ecol 9:351–354.
89. Clark, L., and J.R. Mason. 1988. Effect of biolog-ically active plants used as nest material and thederived benefit to starling nestlings. Oecologia77:174–180.
90. Necker, R. 2000. “The somatosensory system.” InSturkie’s avian physiology, ed. G.C. Whittow, pp.57–69. San Diego: Academic Press.
91. Gottschaldt, K.M. 1985. “Structure and functionof avian somatosensory receptors.” In Form andfunction in birds, ed. A. King and J. McLelland,pp. 375–461. London: Academic Press.
92. Halata, Z., and B.L. Munger. 1980. The ultra-structure of Ruffini and Herbst corpuscles in thearticular capsule of domestic pigeon. Anat Rec198:681–692.
93. Halata, Z., and M. Grim. 1993. Sensory nerveendings in the beak skin of Japanese quail. AnatEmbryol 187:131–138.
94. Gottschaldt, K.M., H. Fruhstorfer, W. Schmidt,and I. Kraft. 1982. Thermosensitivity and its pos-sible fine-structural basis in mechanoreceptors inthe beak skin of geese. J Comp Neurol 205:219–245.
95. Shen, J.X., and Z.M. Xu. 1994. Response charac-teristics of Herbst corpuscles in the interosseousregion of the pigeon’s hind limb. J Comp Physiol175:667–674.
96. Delius, J.D. 1988. Preening and associated com-fort behaviour in birds. Ann NY Acad Sci 525:40–55.
97. Gewecke, M., and M. Woike. 1978. Breast feath-ers as an air-current sense organ for the control offlight behaviour in a songbird (Carduelis spinus).Z Tierpsychol 47:293–298.
98. Bilo, D., and A. Bilo. 1978. Wind stimuli controlvestibular and optokinetic reflexes in the pigeon.Naturwissenschaften 65:161–162.
99. Hensel, H. 1973. “Cutaneous thermoreceptors.”In Handbook of sensory physiology: Somato-sensory system, ed. A. Iggo, pp. 79–110. Berlin/Heidelberg/New York: Springer-Verlag.
100. Frith, H.J. 1959. Incubator birds. Sci Am 201:52–58.
101. Necker, R., and B. Reiner. 1980. Temperature-sensitive mechanoreceptors, thermoreceptors,and heat nociceptors in the feathered skin ofpigeons. J Comp Physiol 135:201–207.
102. Gentle, M.J. 1989. Cutaneous sensory afferentsrecorded from the nervus intramandibularis ofGallus gallus var. domesticus. J Comp Physiol164:763–774.
103. Gentle, M.J. 1992. Pain in birds. Anim Welfare,1:235–247.
4 / Sensory Capacities of Parrots 41

Due to the popularity of keeping psittacine birdsas pets, a better understanding of their social be-havior, in both natural and captive environments,is crucial in order to provide for their social andphysical needs. Many psittacine species do nothave an extensive history of domestication.Understanding and addressing their social needswill improve the welfare of captive psittacinebirds, behavior problems may be more effectivelymanaged, and captive breeding programs can ben-efit when social behaviors are better understood.Social behavior varies among the differentpsittacine species. Solitary behavior is the excep-tion (Kakapo, Strigops habroptilus), with mostspecies showing complex social organization.
FLOCK FORMATION
Flock formation is important for predator detec-tion and avoidance, access to mates, defense ofterritories, and foraging efficiency (Wilson1975). Psittacine birds often form flocks, a behav-ior that is promoted by unstable food resources(irregularly distributed sources that are unpre-dictable through time) and indefensible areas.Feeding together in organized flocks may be ad-vantageous to the individual, who is able to bene-fit from the collective knowledge of the group.By following the flock, an individual has a betterchance of locating adequate amounts of foodwhen resources are unpredictable. Small foraginggroups are better able than individual birds to ex-clude competitors from feeding sites (Wilson1975). There is some evidence that smaller birdswith more limited fasting ability are more likelyto flock than larger birds (Gill 1995).
Indefensible areas also promote flocking be-
havior in birds. There is increased security in alarge group, with individuals nearest the center ofthe flock having the least chance of becoming thevictim of a predator. Flocking improves the effi-ciency of predator detection, allowing the individ-ual more time for other activities. Alarm calling iscommon among flocks and serves to alert othermembers of the group to possible danger. Birdsmay also participate in cooperative mobbing be-havior against intruders (Gill 1995).
Flocks range in size and species composition,with mixed-species flocks observed in nativehabitats. Nuclear species form the main elementsof the organization, while additional species thatjoin the flock opportunistically are referred to as“followers.” The formation of multi-speciesflocks appears to provide additional advantagesfor group members. There may be less conspe-cific competition for similar food and nestingsites. One theory that explains the variety of dis-tinctive plumage coloration in psittacine birdsproposes that the coloration promotes recognitionof conspecifics for breeding purposes when dif-ferent species are living in close proximity(Butcher & Rohwer 1989).
DOMINANCE RELATIONSHIPS
The importance and meaning of dominance inter-actions in psittacine social groups was reviewedby Seibert (2003). Stability of social groups re-quires both mutual recognition of members and asystem for allocation of limited group resources.A dominance relationship exists when predictabledominance-subordinance responses occur be-tween members of a stable social group, based onthe outcome of prior interactions between the in-
43
5Social Behavior of
Psittacine Birds
Lynne M. Seibert

dividuals. Once relationships are established,there is consistency in social interactions, result-ing in fewer, or less intense, aggressive assertionsof dominance (Bernstein 1981). Dominance rela-tionships function to reduce the occurrence ofcompetitive conflicts between members of a so-cial group.
Agonistic behaviors consist of both aggressiveand submissive actions within the context of a so-cial interaction (Wilson 1975). Agonistic encoun-ters are observed more frequently when relation-ships are unclear, such as the introduction of newindividuals. Dominance relationships also appearto require periodic reinforcement, even in the ab-sence of incentive, to prevent extinction (Bern-stein 1981).
Subordinate individuals respond to aggressivebehaviors performed by higher-ranking individu-als with appeasement or submissive signals.Submissive postures allow avoidance of combat.Patterns of communication that function to termi-nate aggression are labeled submissive (Bernstein1981).
BENEFITS OF RANK
The advantages of occupying positions of higherstatus in the flock have not been determined formost psittacine species. Higher-ranking individu-als may have greater access to feeding or roostingsites, lower visibility to predators, or more matingopportunities. Aggressive encounters in a group ofOrange-fronted Parakeets were most frequent dur-ing feeding, followed by bathing or seeking roost-ing places (Hardy 1965). However, no aggressionoccurred in the context of foraging in a captiveflock of Cockatiels, but higher-ranking males didappear to have greater access to mates and pre-ferred nest boxes (Seibert & Crowell-Davis 2001).Female domestic gallinaceous hens were found toselect mates with larger than average combs, orhigher-ranking males if information about socialdominance was available (Graves et al. 1985).Female Speckled Parrotlets also appeared to pur-sue higher-ranking partners, but males showed nopreference for higher-ranking females (Garnetzke-Stollmann & Franck 1991).
GENDER EFFECTS ON AGGRESSIVENESS
In most avian species studied, males show higherfrequencies of aggressive behaviors than females(Jackson 1991; Nol et al. 1996; Seibert &
Crowell-Davis 2001; Wilson 1992; Wingfield etal. 1987; Woolfenden & Fitzpatrick 1977). How-ever, Sandell and Smith (1997) found that femaleEuropean Starlings became more aggressive thanmales during the breeding season. Tarvin andWoolfenden (1997) reported similar findings infemale Blue Jays during the breeding season.Further studies are needed to explore the causesof gender differences in aggressiveness.
Seibert and Crowell-Davis (2001), studying acaptive flock of Cockatiels, found that femaleswere significantly more likely to direct aggres-sion against other females than against males inthe flock. There was not a significant differencefor the male Cockatiels in the gender of their op-ponents. Female competition for access to mateshas been suggested as an explanation for thesegender differences in female aggressiveness (San-dell & Smith 1997; Tarvin & Woolfenden 1997).
INDICATORS OF DOMINANCE
Reliable indicators of dominance status have notbeen determined for most psittacine species.Some postulated indicators of dominance rela-tionships are the frequency of threats and attacksand access to resources. Rushen (1984) proposedthat social dominance within established flocks ofdomestic chickens could be determined using ob-servations of agonistic encounters within the en-tire flock, rather than paired contests. Seibert andCrowell-Davis (2001) measured dominance rela-tionships by recording the outcomes of all agonis-tic encounters during focal sampling of eachflock member. Power (1966) recorded displace-ment at feeding and roosting sites to determinerelative social status in a breeding flock ofOrange-chinned Parakeets.
Studies of other bird populations have shownthat social status is directly related to size, age,and gender (Gill 1995). However, Hardy (1965)found no correlation between dominance rankand physical attributes in the group of parrots hestudied. Instead, he noted a direct correlation be-tween pair bonding and dominance rank. Otherresearchers have found that pair bonding in-creases the social status of psittacine birds withina group (Levinson 1980).
MEASURING DOMINANCE
The agonistic display behaviors of White-frontedAmazon Parrots were classified as low, medium,
44 Manual of Parrot Behavior

or high intensity (Levinson 1980). Threat behav-iors (aggressive components of agonistic behav-ior) are composed of one or more componentsthat differ in valence (intimidatory effectiveness)(Hardy 1965). The displays are partially stereo-typed, such that components appear in a charac-teristic order, but the display may be terminated atany point in the series when intimidation has beenaccomplished.
In species for which dominance interactionshave been recorded, the following behaviors wererecorded as aggressive (Garnetzke-Stollmann &Franck 1991; Hardy 1965; Levinson 1980; Power1966; Seibert & Crowell-Davis 2001).
Turn threat: the aggressing bird abruptly turnedtoward opponent with head and neck extended
Beak gape: aggressing bird directed open beak to-ward opponent
Peck threat: aggressor pecked at opponent but didnot make contact
Beak spar: short bouts in which birds’ beaks madecontact
Peck: aggressor’s beak closed on some part of re-cipient
Wing flapping: perched aggressor flapped wingswhile facing opponent
Sidle approach: perched aggressor approachedwith side of the body directed toward opponent
Slow advance: perched aggressor walked directlytoward opponent
Rushing: perched aggressor ran at opponentFlight approach: aggressor flew directly toward
opponent
Hardy (1965) and Power (1966) also reportedstationary threat behaviors including plumage ap-pression, or sleeking of the body feathers, andmalar fluffing, fluffing of feathers in the malar re-gion, which causes the bird’s head to appearlarger and draws attention to the beak.
Submissive behaviors, also referred to as ap-peasement behaviors, appear to be less ritualized.Submissive behaviors performed in response toaggressive displays consist of crouching, fluffingfeathers, head wagging, foot lifting, or avoidance(Hardy 1965).
Variations in threat display complexity of dif-ferent psittacine species may be explained by thegames-theory approach. Games theory predictsthat as the risk of physical injury increases,
species evolve less dangerous strategies for re-solving disputes, such as more complex ritualizedpostural displays. Serpell (1982) studied nine dif-ferent taxa of Trichoglossus parrots that differedin their beak size and the complexity of threat dis-plays. The findings of this study suggest that aninterspecific difference in beak length, which isdirectly related to the risk of injury from the bitesof conspecifics, influences the nature of threatdisplays. With increasing beak size, displays be-came more complex, and there was a reduced in-clination to attack a mirror-image opponent(Serpell 1982).
GENDER EFFECTS ON DOMINANCERELATIONSHIPS
Several studies have found that male birds tend tooccupy higher social positions than female birds(Seibert & Crowell-Davis 2001; Tarvin & Wool-fenden 1997; Weinhold 1998; Woolfenden &Fitzpatrick 1977).
Aggressiveness, or the tendency to initiate ag-onistic interactions, may or may not be correlatedwith dominance status. Cloutier et al. (1995)found that subordinate hens were less likely to en-gage in aggressive behaviors than dominant hens.Graves et al. (1985) also found that aggressive-ness was correlated with dominance rank inWhite Leghorn cocks. Higher-ranking Cockatielshad significantly higher rates of aggression thanlower-ranking flock members (Seibert & Crowell-Davis 2001).
AFFILIATIVE RELATIONSHIPS
Affiliative behaviors in birds consist of allopreen-ing, allofeeding, maintenance of close proximity,pair bonding, and reproductive behaviors.Garnetzke-Stollmann and Franck (1991) de-scribed affiliative interactions in a group of cap-tive parrots (Forpus conspicillatus) that includedperching in close contact, allopreening, and solic-itation of allopreening.
Spatial organization of flock members is not ran-dom. Members maintain relationships with otherflock members that can be measured based on spa-tial patterns and proximity. Sparks (1964) foundthat the members of a flock of Red Avadavats(Amandava amandava) were not randomly dis-persed. Grigor et al. (1995) found that spatial asso-ciations in domestic chickens were influenced bythe social relationships within the flock. Seibert
5 / Social Behavior of Psittacine Birds 45

and Crowell-Davis (2001) also found the spacingin a flock of Cockatiels to be non-random and in-dicative of preferred associations. Preferred spa-tial associations coincided with mating groups,but in addition, males and females in the flocksometimes had same-gender preferred associates.Other researchers have found that mated pairs ofbirds maintain close spatial associations (Silcox &Evans 1982; Trillmich 1976; Wechsler 1989).
ALLOPREENING
Allopreening, which occurs when an individualuses its beak to groom another bird, is cited as themost important mechanism for maintenance ofthe pair bond (Gill 1995). In a review of the allo-preening behavior of different psittacine species,Harrison (1994) reported that allopreening wasconfined to the head and neck region in amazonparrots, lovebirds, and the genus Melopsittacus.Allopreening involved the head, wings, and tail inAratinga, Brotogeris, Ara, and Cacatua species.
Since allopreening behavior has been associ-ated with the formation of pair bonds, a predilec-tion for cross-gender allopreening has been sup-ported in various avian species (Gaston 1977;Harrison 1965; Spruijt et al. 1992). Seibert andCrowell-Davis (2001) found that males allo-preened females significantly more than theyallopreened other males. However, isosexual allo-preening does occur and should be viewed as ev-idence of a social bond. Garnetzke-Stollmann andFranck (1991) observed that preferred associa-tions, allopreening, and support in agonistic inter-actions were significantly more common amongsiblings than among unrelated birds in a flock ofSpeckled Parrotlets.
ALLOFEEDING
Allofeeding is closely associated with copulationin birds (Skeate 1984). The female solicits feed-ing by crouching, lowering her head, ruffling herfeathers, and vocalizing. The male displays headbobbing, grasps the female’s beak at a right angle,and regurgitates food to the female. Allofeedingoccurs year-round in some amazon parrots,conures, lovebirds, and Grey-cheeked Parakeets(Harrison 1994).
PAIR BONDING
Pair bonding has been defined as a mutually ben-eficial relationship between sexually mature fe-
male and male birds, serving primarily for the co-operative rearing of young (Doane & Qual-kinbush 1994; Wilson 1975). Pairs are character-ized by allofeeding, pair participation in agonisticencounters, and close spatial associations(Garnetzke-Stollmann & Franck 1991; Levinson1980; Trillmich 1976). Many psittacine speciesare thought to maintain pair bonds throughout theyear.
Advent of the breeding season can alter the so-cial hierarchy. Power (1966) found that singlebirds were more successful in an aggressive en-counter if their mate was nearby, even if the matedid not appear to be actively participating.Levinson (1980) reported pair participation in ag-onistic encounters in White-fronted AmazonParrots (Amazona albifrons), a species that main-tained pair bonds throughout the year.
According to Butterfield (1970), perching inclose proximity can be interpreted as evidence ofpair bond formation. Arrowood (1988) observedclose spatial associations among bonded pairs ofCanary-winged Parakeets (Brotogeris v. versi-colorus). Mates maintained very close proximity,usually touching. In addition, once pairing wasachieved, individual mates no longer displayed af-filiative behaviors toward any other flock mem-bers, as long as the mate was present in the flock,and agonistic displays did not occur between pair-bonded individuals.
MATING PAIRS AND GROUPS
The social behavior of most psittacine species hasnot been studied in natural habitats, and captivepopulations have commonly been housed in pairsfor breeding. The assumption that psittacine birdsmaintain exclusive pair bonds is not accurate forall species. Extra-pair matings have been ob-served in Speckled Parrotlets, Budgerigars, andCockatiels (Baltz & Clark 1997; Garnetzke-Stollmann & Franck 1991; Seibert & Crowell-Davis 2001). Allopreening occurs less frequentlybetween the male and the secondary female.Extra-pair courtship activities tend to occur whilethe primary female partner is incubating eggs andunable to observe the activity (Baltz & Clark1997).
While some psittacine males will actuallyassist in incubating the eggs (Cockatiels,macaws, conures, and some cockatoo species),all male psittacine birds appear to assist in the
46 Manual of Parrot Behavior

rearing of young. Male birds feed the hen as sheincubates, guard the nest entrance, and feed thehatchlings. Many psittacine offspring have rela-tively long infancies, with weaning taking up toa year in some species, increasing the require-ment for parental care (Doane & Qualkinbush1994).
Successful nesting behavior has been describedfor endangered Puerto Rican Parrots (Amazonavittata) in a field setting. Successful mating pairs,or those producing fledged chicks, followed dis-tinct patterns of nesting activities (Wilson et al.1995). Females increased nest attendance duringegg laying and incubation, while males rarelyentered the nest at this stage. Allofeeding of the hen occurred close to the nest. During earlychick rearing, male attentiveness increased, whilefemales began to spend more time away from the nest. Regular allofeeding of the young by the male was essential for proper growth.Knowledge of the typical patterns of nest atten-dance can be used to detect problems in captivebreeding programs.
CONCLUSION
An understanding of the social behaviors ofpsittacine birds has implications for the preven-tion and treatment of various undesirable behav-iors of pet birds. Problems may occur in birds notprovided with appropriate socialization opportu-nities, or as the birds reach sexual maturity, withreports of behaviors that seem to indicate bond-ing with a human caregiver. Undesirable behav-iors include attempts to preen, allofeed (regurgi-tate), and copulate with the person; masturbation;aggressive attempts to drive away other membersof the family; and defense of the cage as a nest-ing site (Harrison & Davis 1986). In addition tosexual behaviors, abnormal behaviors indicativeof stress or anxiety can occur including featherpicking, barbering, and self-mutilation; scream-ing, aggression and biting; and phobias. The im-portance of flock social interactions to variousspecies and the effects of isolation or pair hous-ing on welfare are pertinent issues. Spatial andsocial relationships have important implicationsfor the management of psittacine species, includ-ing providing the appropriate amount of space forthe flock, optimizing feeding locations, deter-mining stocking density, and identifying matingpairs.
REFERENCES
Arrowood, P.C. 1988. Duetting, pair bonding, and ago-nistic display in parakeet pairs. Behaviour 106:129–157.
Baltz, A.P., and A.B. Clark. 1997. Extra-pair courtshipbehaviour of male budgerigars and the effect of anaudience. Animal Behavior 53:1017–1024.
Bernstein, I.S. 1981. Dominance: The baby and thebath water. Behavioral and Brain Sciences4:419–457.
Butcher, G.S., and S. Rohwer. 1989. The evolution ofconspicuous and distinctive coloration for commu-nication in birds. Current Ornithology 6:51–108.
Butterfield, P.A. 1970. “The pair bond of the zebrafinch.” In Social behavior in birds and mam-mals: Essays on the social ethology of animals, ed.J.H. Crook, pp. 249–278. New York: AcademicPress.
Cloutier, S., J.P. Beaugrand, and P.C. Laguë. 1995. Theeffect of prior victory or defeat in the same site asthat of subsequent encounter on the determination ofdyadic dominance in the domestic hen. Behav Proc34:293–298.
Doane, B.M., and T. Qualkinbush. 1994. My parrot, myfriend: An owner’s guide to parrot behavior. NewYork: Macmillan.
Garnetzke-Stollmann, K., and D. Franck. 1991.Socialisation tactics of the speckled parrotlet(Forpus conspicillatus). Behaviour 119:1–29.
Gaston, A.J. 1977. Social behaviour within groups ofjungle babblers (Turdoides striatus). AnimalBehavior 25:828–848.
Gill, F.B. 1995. Ornithology, 2nd ed. New York: WHFreeman and Company.
Graves, H.B., C.P. Hable, and T.H. Jenkins. 1985.Sexual selection in Gallus: Effects of morphologyand dominance on female spatial behavior. BehavProc 11:189–197.
Grigor, P.N., B.O. Hughes, and M.C. Appleby. 1995.Social inhibition of movement in domestic hens.Animal Behavior 49:1381–1388.
Hardy, J.W. 1965. Flock social behavior of the orange-fronted parakeet. Condor 67:140–156.
Harrison, C.J.O. 1965. Allopreening as agonistic be-haviour. Behaviour 24:161–209.
Harrison, G.J. 1994. “Perspective on parrot behavior.”In Avian medicine: Principles and application,ed.B.W. Ritchie, G.J. Harrison, and L.R. Harrison,pp. 96–108. Lake Worth, FL: Wingers Publishing.
Harrison, G.J., and C. Davis. 1986. “Captive behaviorand its modification.” In Clinical avian medicineand surgery, ed. G.J. Harrison, pp. 20–28. Phila-delphia: WB Saunders Company.
Jackson, W.M. 1991. Why do winners keep winning?Behav Ecol Sociobiol 28:271–276.
5 / Social Behavior of Psittacine Birds 47

Levinson, S.T. 1980. The social behavior of the white-fronted Amazon (Amazona albifrons). In Conser-vation of new world parrots: Proceedings of theICBP Parrot Working Group Meeting, ed. R.F.Pasquier, pp. 403–417. Washington, DC: Smithson-ian Institution Press.
Nol, E., K. Cheng, and C. Nichols. 1996. Heritabilityand phenotypic correlations of behaviour and domi-nance rank of Japanese quail. Anim Behav52:813–820.
Power, D.M. 1966. Agonistic behavior and vocaliza-tions of orange-chinned parakeets in captivity. Con-dor 68:562–581.
Rushen, J. 1984. How peck orders in chickens aremeasured: A critical review. Appl Anim Ethol11:255–264.
Sandell, M.I., and Smith, H.G. 1997. Female aggres-sion in the European starling during the breedingseason. Anim Behav 53:13–23.
Seibert L.M. 2003. “Social dominance: The peck orderrevealed.” Proc Assoc Avian Vet, Pittsburgh, PA, pp.187–188.
Seibert, L.M., and Crowell-Davis, S.L. 2001. Gendereffects on aggression, dominance rank, and affilia-tive behaviors in a flock of captive adult cockatiels(Nymphicus hollandicus). Appl Anim Behav Sci 71(2):155–170.
Serpell, J.A. 1982. Factors influencing fighting andthreat in the parrot genus Trichoglossus. AnimalBehaviour 30:1244–1251.
Silcox, A.P., and Evans, S.M. 1982. Factors affectingthe formation and maintenance of pair bonds in thezebra finch, Taeniopygia guttata. Anim Behav30:1237–1243.
Skeate, S.T. 1984. Courtship and reproductive behav-iour of captive white-fronted Amazon parrots (Ama-zona albifrons). Bird Behaviour 5:103–109.
Sparks, J.H. 1964. Flock structure of the red avadavatwith particular reference to clumping and allopreen-ing. Anim Behav 12:125–136.
Spruijt, B.M., VanHooff, J.A., and Gispen, W.H. 1992.Ethology and neurobiology of grooming behavior.Physiol Rev 72:825–852.
Tarvin, K.A., and Woolfenden, G.E. 1997. Patterns ofdominance and aggressive behavior in blue jays at afeeder. Condor 99:434–444.
Trillmich, F. 1976. Spatial proximity and mate-specificbehaviour in a flock of budgerigars. Z Tierpsychol41:307–331.
Wechsler, B. 1989. Measuring pair relationships injackdaws. Ethology 80:307–317.
Weinhold, J. 1998. Analysis of the social behavior of acommunity of blue-fronted Amazons (Amazonaaestiva) kept in an aviary. Amazona Quarterly14:11–13.
Wilson, E.O. 1975. Sociobiology. Cambridge: BelknapPress.
Wilson, J.D. 1992. Correlates of agonistic display bygreat tit Parus major. Behaviour 121:168–214.
Wilson, K.A., Field, R., and Wilson, M.H. 1995.Successful nesting behavior of Puerto Rican parrots.Wilson Bulletin 107 (3):518–529.
Wingfield, J.C., Ball, G.F., Dufty, A.M., Hegner, R.E.,and Ramenofsky, M. 1987. Testosterone and aggres-sion in birds. Amer Scientist 75:602–608.
Woolfenden, G.E., and Fitzpatrick, J.W. 1977. Domi-nance in the Florida scrub jay. Condor 79:1–12.
48 Manual of Parrot Behavior

OVERVIEW OF PSITTACINE NUTRITION
The selection or formulation of appropriate dietsthat meet the nutrient requirements of psittacinebirds is based upon several factors. First, the wild-type foraging habits and behaviors of a particularparrot species provide important information re-garding the animal’s evolutionary adaptations tofeeds and feeding. Second, an understanding of ananimal’s digestive anatomy and physiology, whichoften reflects its wild-type feeding habits, assistsnutritionists in determining the nutrient and foodrequirements of that species. Third, optimal dietformulation is enabled by experimental data con-cerning the specific nutrient requirements of aparticular species, based on data collected in ei-ther that species or in a closely related species.This factor is especially important, since knowl-edge of an animal’s wild-type foraging strategiesand its digestive anatomy and physiology onlyprovide estimates of nutritional needs, while ex-perimental data tests hypotheses and firmly estab-lishes nutritional needs for a particular animal.This chapter explores topics in parrot nutrition in-cluding wild-type feeding strategies, digestiveanatomy, and physiology with a special focus onavian gustation, calculated and experimentally de-termined nutrient requirements, and behavioraland immunological impacts of an improper diet.
WILD-TYPE DIETS
In the wild, the majority of parrots and othermembers of the order Psittaciformes consumeplant-based (folivorous) diets. Folivorous psit-
tacines range from nectarivorous (nectar-/pollen-eating) to granivorous (seed-eating), whileconsumption of several plant-based feedstuffs(omnivory) is most common. A condensed listingof wild-type feeding strategy and common dietingredients can be found in Table 6-1, and moredetailed information concerning a wider range ofspecies is available (Koutsos et al. 2001a).
DIGESTIVE ANATOMY ANDPHYSIOLOGY
A bird’s digestive anatomy, including the beakand oral cavity, esophagus, crop, proventriculus,gizzard, small and large intestine, ceca (generallyabsent in parrots), and cloaca, determines its abil-ity to acquire, digest, and absorb nutrients, andoften reflects its wild-type diet. For example,beak shape and size is generally correlated towild-type diets (Klasing 1998). Granivorous birdsoften have ridged beaks that enhance seed-cracking abilities (Homberger & Brush 1986),and large granivores (particularly parrots) gener-ally have a cartilaginous connection between theskull and beak to absorb the shock of crackinglarge seeds. Compared to granivores, frugivorousbirds tend to have wider beaks and oral cavities.The remainder of the gastrointestinal tract, in-cluding the crop, proventriculus, gizzard, and in-testines, is generally similar in psittacine birds, al-though the gizzards of frugivorous birds tend tohave reduced musculature as compared to grani-vores or omnivores, reflecting the reduced needfor particle grinding (Klasing 1998). Finally,
49
6Captive Parrot Nutrition:
Interactions with Anatomy,Physiology, and Behavior
Kevin David Matson and Elizabeth A. Koutsos

larger birds generally have longer GI tracts, whichresults in increased time of retention of diet ingre-dients as compared to that of smaller birds.
GUSTATION IN PSITTACINES
Gustation, or the act of tasting food particles, oc-curs in the structures of the oral cavity. In the caseof birds, these structures include the salivary
glands, the tongue and its associated taste buds,and the beak. One important function of the struc-tures associated with the oral cavity is the percep-tion of the chemical qualities of potential food.Because these qualities transmit informationabout the suitability of potential food, it is impor-tant to understand how parrots sense their chemi-cal environment.
50 Manual of Parrot Behavior
Table 6.1. Feeding strategies and common diet ingredients of some wild Psittaciformes
Species name Strategy Common diet ingredients References
Blue and Gold macaw Florivore Seeds, fruits, nuts (Abramson et al. 1995)(A. ararauna)
Red-faced parrot Florivore Flowers, berries, shoots, (Toyne & Flanagan (H. pyrrhops) seeds, seed pods 1997)
Scaly-headed parrot Florivore Seeds (70%), flowers (20%), (Galetti 1993)(P. maximiliani) grain (8%), fruit pulp (2%)
Blue-throated macaw Frugivore Palm fruit, nuts, milk (Abramson et al. 1995)(A. glaucogularis)
Buffon’s macaw Frugivore Fruits, flowers (Abramson et al. 1995)(A. ambigua)
Green-winged macaw Frugivore Fruits (Hymenaea), palm nuts, (Abramson et al. 1995)(A. chloroptera) seeds
Orange-winged amazon Frugivore Fruit (85% from palm fruit) (Bonadie & Bacon (A. amazonica) 2000)
Red-bellied macaw Frugivore Fruit (96% from palm fruit), (Bonadie & Bacon (A. manilata) flowers, seed pods 2000)
Vulturine parrot Frugivore One or two of the 38 extant (Mack & Wright 1998)(P. fulgidus) species of figs (Ficus spp.)
Red-fronted macaw Frugivore- Fruits, seeds (Pitter & Christiansen (A. rubrogenys) Granivore 1995)
Regents parrot Frugivore- Fruits, seeds (Long & Mawson (P. anthopeplus) Granivore 1994)
Scarlet macaw Frugivore- Fruits, nuts, bark, leaves and (Abramson et al. 1995)(A. macao) Granivore shoots
Budgerigar (M. undulatus) Granivore Seeds (Wyndham 1980)Cockatiel (N. hollandicus) Granivore Seeds (prefers soft, young over (Jones 1987)
mature, hard seeds)Ground parrot (P. wallicus) Granivore Seeds, some insect larvae (Mcfarland 1991)Hyacinth macaw Granivore Palm nuts (50% lipid content) (Abramson et al. 1995)
(A. hyacinthinus) Lear’s macaw (A. leari) Granivore 1° palm nuts, fruit (Abramson et al. 1995)Red-fronted macaw Granivore Nuts, seeds, fruit (Abramson et al. 1995)
(A. rubrogenys)Spix’s macaw (C. spixii) Granivore Palm nuts (Abramson et al. 1995)Hooded parrot Omnivore 1° seeds (1° sesame), flowers, (Garnett & Crowley
(P. dissimilis) invertebrates, 1995)Red-tailed amazon Omnivore Seeds, fruits, flowers, leaves, (Martuscelli 1995)
(A. brasiliensis) nectar and insects

Chemosensory Perception
The importance of chemosensory perception (i.e.,gustation and olfaction) in birds has typicallybeen downplayed with the emphasis, instead,being placed on sight and hearing. Nonetheless,olfaction and gustation are employed by birds in avariety of nutritive pursuits. For example, theProcellariiform “tubenose” seabirds use scent tolocate food sources when navigating vast pelagicenvironments (Malakoff 1999; Nevitt 1999b;Nevitt 1999a), while Red-winged Blackbirds(Agelaius phoeniceus) reject dilute solutions ofarthropod defensive secretions on the basis oftaste (Yang & Kare 1968). In addition to toxinavoidance, foodstuffs are also evaluated by tastein order to detect nutrient levels (Herness &Gilbertson 1999). This phenomenon has been ex-tensively researched in nectarivores such as hum-mingbirds and sunbirds (Martinez del Rio 1990;Martinez del Rio et al. 1992; Downs & Perrin1996). Further, plasticity in the transduction path-ways of gustatory chemoreceptors may result in aconnection between the nutritional status of birdsand their taste responses, thereby allowing for“specific appetites” for certain minerals (Herness& Gilbertson 1999).
While the avian sense of taste has received lit-tle attention, is has been demonstrated that tastebuds are present in the oral cavity of birds on thefloor of the pharynx and the base of the tongue(El Boushy et al. 1989). The number of avian tastebuds appears to be extremely low compared tomammals. Humans (with about 9,000) havearound 25 times more taste buds than parrots,which are reported to have 300–400 (Kare &Mason 1986). Fibers stemming from the glos-sopharyngeal nerve innervate the taste buds (ElBoushy et al. 1989). Most of the additional stud-ies of the microanatomy and physiology of tastebuds have been limited to mammals, and little isknown about what generalities about these struc-tures can be made across taxa.
Testing Taste in Birds
The gustatory abilities of birds have been testedin several species by a variety of methods. Themethods typically involve comparisons of con-sumption variables (e.g., number of sips or pecksor volume consumed) that are measured simulta-neously for pure water compared to an aqueous
solution of the test chemical, offered in separatebottles. Most commonly, two-choice taste-preference tests are conducted. However, concernsabout arbitrary side preferences or biases have re-sulted in some investigators varying the numberof bottles (from two to six), the relative positionof each bottle (usually changing on an hourly ordaily basis), and the location of bottles withineach cage (Bartholomew & Cade 1958; Harriman& Fry 1990; Jackson et al. 1998). In addition tonumerous studies testing the sugar preferences ofnectarivores and frugivores, many wild and do-mestic species have been tested to determinewhether consumption patterns change followingthe addition of taste stimuli: domestic chickens(Gallus domesticus [Kare et al. 1957; Kare &Pick 1960; Fuerst & Kare 1962; Gentle 1972]),Rock Doves (Columba livia [Duncan 1960;Crocker et al. 1993]), common crows (Corvuscorax [Harriman & Fry 1990]), Japanese Quail(Coturnix coturnix japonica [Brindley 1965;Harriman & Milner 1969]), Bobwhite Quail(Colinus virginianus [Brindley 1965; Brindley &Prior 1968]), Laughing Gulls (Larus atricilla[Harriman 1967]), Herring Gulls (Larus argenta-tus smithsonianus), European Starlings (Sturnusvulgaris), Common Grackles (Quiscalus quiscula[Harriman & Kare 1966]), and House Finches(Carpodacus mexicanus [Bartholomew & Cade1958]). This group of studies suggests that birdsrespond differently to a range of taste stimuli andthe response depends on the species, the com-pound, and the concentrations being tested.
Taste Thresholds of Cockatiels
To better understand the role of taste in foodchoice of parrots, a series of tests were conductedusing captive Cockatiels, Nymphicus hollandicus,as a model (Matson et al. 2000; Matson et al.2001). Two-choice taste-preference tests were de-signed to determine thresholds of taste for varioustaste stimuli in the sweet, salt, bitter, and sour cat-egories. Taste threshold was defined as the lowestconcentration at which the consumed volume of asolution of test compound was significantly dif-ferent from the consumed volume of pure water.
Results of this work are summarized in Table 6-2. In general, these results show that the birds’gustatory acuity differs according to taste cate-gory (i.e., salt, sweet, bitter, or sour). A 4,000-folddifference in concentration was found between
6 / Captive Parrot Nutrition: Interactions with Anatomy, Physiology, and Behavior 51

the highest (sweet, using fructose) and lowest(bitter, using quinine) thresholds. Further, for alltest substances, the Cockatiels either significantlyincreased intake of pure water or significantly de-creased intake of the test solution—no solutionswere preferable to water.
Cockatiels demonstrated the greatest gustatoryacuity for bitter flavors. Thresholds for these com-pounds ranged from 0.1–10 mmol*L�1, with thelowest threshold for quinine (0.1 mmol*L�1)—acommonly used bitter tasting alkaloid (K. Matson,unpublished data). The quinine threshold ofCockatiels is similar to the threshold of most hu-mans (0.09 mmol*L�1 in distilled water [Schall1990]), but lower than the thresholds of mam-malian florivores (e.g., browser threshold, 3.0mmol*L�1; grazer threshold, 0.67 mmol*L�1
[Glendinning 1994]).Both chloride salts that were tested were de-
tected at the same threshold (160.0 mmol*L�1)(Matson et al. 2001). As the normal range for cir-culating sodium concentrations in captive parrotsis 130.0–157.0 mmol*L�1, the threshold levelsrepresent a marginally hypertonic concentration(Lane 1996; Polo et al. 1998). Rejection of hy-pertonic salt solutions is common in birds. For ex-ample, Laughing Gulls, Herring Gulls, and Euro-pean Starlings show an aversion to solutions ofsodium chloride in the range of 150.0–200.0mmol*L�1 (Harriman & Kare 1966; Harriman1967). One possible explanation for salt rejectionis that hormones responsible for regulating waterand salt balance also regulate the amiloride-
sensitive sodium channel, a sodium gustatorytransduction mechanism (Herness & Gilbertson1999).
Some reports indicate that parrots prefer solu-tions of sugar to pure water, while others hypoth-esize that granivores, such as Cockatiels, shouldreact neutrally or negatively to sugar consumption(El Boushy et al. 1989; Kare & Mason 1986, re-spectively). No significant preferences werefound when two monosaccharides (glucose andfructose) and one disaccharide (sucrose) weretested independently (Matson et al. 2000; Matsonet al. 2001). Threshold concentrations were, how-ever, higher on average than those found for bitterand salt compounds.
Sour taste is mostly based upon acidity or freeprotons (H+). Therefore, two buffer systems wereused so that the concentration of free protonscould be manipulated—a 0.05 mol*L�1 buffer ofsodium citrate and citric acid and a 0.05 mol*L�1
buffer of mono- and dibasic sodium phosphate.For these stimuli, the pH (rather than simple con-centration) was varied and the pH was used as wellfor reporting taste thresholds. Despite the abilityof protons to trigger many classes of ion channelsthat serve as gustatory transduction mechanisms,only citrate buffer resulted in the determination ofa threshold (Matson et al. 2000; Matson et al.2001). Thus, taste trials in Cockatiels, as well as inchickens, correspond to neurological studiesdemonstrating greater responses to organic acids(i.e., citrate buffer) than inorganic acids (i.e.,phosphate buffer [Fuerst & Kare 1962]).
52 Manual of Parrot Behavior
Table 6.2. Taste thresholds of Cockatiels, Nymphicus hollandicus
Taste category Chemical stimulus Test range Taste threshold
Salt Sodium chloride 20.0–180.0 mmol*L�1 160.0 mmol*L�1
Potassium chloride 100.0–250.0 160.0Sweet Fructose 160.0–490.0 mmol*L�1 400.0 mmol*L�1
Glucose 160.0–200.0 160.0Sucrose 80.0–560.0 360.0
Bitter Quinine 0.001–0.5 mmol*L�1 0.1 mmol*L�1
Gramine 0.001–1.0 1.0Tannic acid 0.001–1.0 0.5Watte tannins 0.001–10.0 10.0
Sour Citrate buffer (0.05 mol*L�1) pH 5.0–5.5 pH 5.5Phosphate buffer (0.05 mol*L�1) pH 4.9–7.7 None determined
Source: Matson et al. 2000; Matson et al. 2001; Matson, unpublished data.

Implications
Given the wide range of Cockatiel taste thresh-olds, it is important to consider the effects of gus-tation on diet formulation and dietary changes incaptive psittacines. Food palatability is the prod-uct of the food’s chemical qualities and the bird’staste abilities. Despite the fact that the gustatoryacuity of Cockatiels and other birds is generallyequal to or less than humans, certain chemicals—particularly bitter ones—are rejected by birds onthe basis of taste and these should be avoidedwhen formulating diets. Various levels of bitter-tasting tannins are present in many grains cropsand if these grains are used in seed mixes or for-mulated diets, food intake may be reduced. Othertastes, such as salt and sweet, are associated withnutrients required by all birds, and thus, the ac-ceptance or rejection response to these taste stim-uli may vary with nutritional status. Finally, tastesand specific appetites may sometimes drive con-sumption. Therefore, it is possible that becausethe Cockatiels used in the taste preference trialsjust described were all offered a salt and energysufficient diet ad libitum (0.18% Na, 0.27% Cl,crude protein minimum 11%; five primary ingre-dients: ground corn, ground wheat, peanut meal,soy oil, and soy meal; Maintenance Crumbles,Roudybush, Inc., Cameron Park, California), thethresholds as measured may have been affected. Ifthe birds had been fed a deficient diet, prefer-ences for solutions containing the deficient nutri-ent might be expected.
THEORETICAL AND CALCULATEDNUTRIENT REQUIREMENTS
Knowledge of an animal’s wild-type diet, gas-trointestinal anatomy and physiology, and gusta-tory capacity in combination with experimentaldata obtained in that species or related speciescan be used to determine specific nutrient re-quirements. It is important to keep in mind, how-ever, that nutrient requirements are generallybased upon meeting the nutrient needs of the ma-jority of a population. Inevitably, there are ani-mals whose individual nutrient needs are differentfrom the majority of the population. Therefore,even when feeding individual animals a diet be-lieved to be nutritionally adequate, it is importantto be observant for signs of deficiency or toxicity,in addition to behavioral changes indicating nutri-tional problems.
Water
Water is often overlooked as a nutrient, but mustbe supplied to maintain cellular homeostasis,food digestion processes, waste excretion, andnumerous metabolic reactions. Water require-ments will vary with size of the animal (in gen-eral, smaller animals need less water) and envi-ronmental temperature (warmer temperaturestend to increase water requirements). Underthermo-neutral conditions, the daily water re-quirement of adult parrots is calculated to be~2.4% of body weight (MacMillen & Baudinette1993).
Energy
Energy can be supplied in the diet by lipid, pro-tein, or carbohydrate, and functions to supportbasal metabolism, thermoregulation, and activity.A bird’s energy requirement will change withenvironmental temperature (colder temperaturesincrease energy needs), activity level (higher ac-tivity levels also increase energy needs), andphysiological state (maintenance animals requireless energy than breeding or growing animals).Estimates of the daily metabolizable energy re-quirements for adult psittacine birds range from154.6 kcal/kg0.75 (indoor cage) to 226.1 kcal/kg0.75 (outdoor aviary, cold weather) (see Koutsoset al. 2001a).
Protein/Amino Acids
Birds require 12 essential amino acids: phenylala-nine, valine, tryptophan, methionine, arginine,threonine, hisitidine, isoleucine, lysine, leucine,glycine, and proline. Additionally, a source of ni-trogen (e.g., protein) must also be present in thediet. Like energy requirements, the physiologicalstate of an animal has a dramatic effect on itsprotein and amino acid requirements. In general,requirements are highest in growing chicks and fe-males laying large clutches of eggs, while require-ments are generally lowest for adult animals atmaintenance. For example, the African GreyParrot (Psittacus erithacus) requires 10–15% pro-tein for maintenance (Kamphues et al. 1997), buthigher requirements would be predicted for grow-ing chicks and egg-laying females. Similarly, 11%crude protein is sufficient to support Cockatiels(Nymphicus hollandicus) at maintenance (Koutsoset al. 2001b), but growing Cockatiel chicks require20% crude protein (Roudybush & Grau 1986).
6 / Captive Parrot Nutrition: Interactions with Anatomy, Physiology, and Behavior 53

In addition to physiological state, dietary feed-ing strategy impacts the protein requirement ofanimals. For example, frugivorous species ofbirds have lower rates of obligatory nitrogenlosses as compared to granivorous species (Pryor1999). Therefore, these specialist feeders are ex-pected to have lower protein requirements thangranivores or omnivores, and this hypothesis hasbeen confirmed in the Rainbow Lorikeet (Tricho-glossus haemotodus), which required less than3% crude protein when provided a high-qualityprotein source (egg white) (Frankel & Avram2001).
Lipids
Birds, like other animals, require essential fattyacids to provide membrane integrity, intracellularsignaling molecules, and hormones. However, lit-tle research has been conducted to quantify thefatty acid requirements of psittacines. Therefore,nutritionists generally use poultry guidelines as areference (NRC 1994); domesticated poultry re-quire approximately 1% linoleic acid and 4–5%total lipid in poultry diets is not uncommon.
Vitamins
Presumably, parrots and other psittacines requirethe same vitamins as do other birds. However, lit-tle research has been conducted to quantify theserequirements. In general, research has focused onthe fat-soluble vitamins (A, D, E, and K); due totheir chemical nature, these vitamins can be diffi-cult to excrete, resulting in enhanced susceptibil-ity to toxicity. Research in Cockatiels has demon-strated that diets containing 2,000–10,000 IUvitamin A/kg were sufficient to support animalsat maintenance, and 4,000 IU vitamin A/kg dietwas sufficient to support chick growth to fledging(Koutsos & Klasing 2002). Interestingly, in thesame trial, adult birds fed 100,000 IU vitaminA/kg diet developed vitamin A toxicity within 3months, while birds fed 0 IU vitamin A/kg dietdid not develop a vitamin A deficiency during thetwo-year experimental period. However, 0 IU vi-tamin A/kg diet did not support normal growthand development of Cockatiel chicks. These re-sults demonstrate that adult Cockatiels are moresusceptible to vitamin A toxicity than to vitaminA deficiency, and that growing chicks require adietary vitamin A source. Additionally, Cockatielchicks fed 2.4 mg ß-carotene/kg diet and no vita-
min A grew normally and exhibited no signs ofvitamin A deficiency. This observation supportsthe use of pro-vitamin A carotenoids in psittacinediets, perhaps as a means to avoid vitamin A tox-icity. However, the exact rate of ß-carotene con-version to vitamin A in psittacines has not yetbeen determined.
Minerals
Because of the requirements for eggshell forma-tion and skeletal calcification, as well as variablelevels of calcium in different diet ingredients, theprimary mineral of concern to avian nutritionistsis calcium. The calcium requirement for psit-tacines has not been experimentally determined,so the established chicken requirement of 0.1% ofthe diet (NRC 1994) is commonly used. This is ofparticular importance when diets composed en-tirely of seed are fed, since seeds generally con-tain less than 0.1% calcium. Additionally, highercalcium requirements are expected during periodsof eggshell formation and laying; egg-layingchickens are commonly fed 3–4% calcium.African Grey Parrots are commonly diagnosedwith hypocalcemia (Rosskopf et al. 1985), al-though whether the calcium requirements or defi-ciency pathologies of this species are similaracross parrot species remains to be determined. Aswith calcium, the requirements of psittacines forother essential minerals have not been examined,and poultry guidelines are often the only dataavailable concerning avian mineral requirements.
EFFECTS OF NUTRIENT DEFICIENCIES/EXCESSES
Nutrient deficiency and excess can induce numer-ous pathologies, which can include defects in re-production, embryogenesis, and the growth anddevelopment of chicks; increased susceptibility todisease; undesired behavioral changes; poorhealth; and ultimately, death. A deficiency or tox-icity of an individual nutrient will likely result ina unique combination of signs and symptoms, al-though the underlying etiology of these patholo-gies is often very similar.
Immune Function Effects
The immune response can be dramatically alteredby the nutritional status of an animal (see reviewby Koutsos & Klasing 2001). These effects aremost pronounced when deficiencies or toxicities
54 Manual of Parrot Behavior

occur during development, and a chronic defi-ciency of virtually any required nutrient during theperiod of immune system development (primarilyduring in ovo development, but also in post-hatchchicks) negatively impacts immunocompetence.In general, those required nutrients that functionin regulating cell differentiation (e.g., vitamins Aand D) are particularly detrimental to the develop-ment of immunocompetence. Similarly, post-hatch nutritional status can affect all facets of theimmune system, and nutrient deficiency or excessmay lead to increased disease susceptibility andmay enhance the virulence or pathogenicity ofcertain organisms. Finally, many nutrients, specif-ically fatty acids and antioxidants, may modulatethe immune response depending on their rate ofdietary inclusion. For example, vitamin E hasbeen shown to have anti-inflammatory propertiesat moderate dietary levels, while high dietary lev-els are associated with dampened immune respon-siveness (e.g., broiler chickens [Leshchinsky &Klasing 2001]). However, the optimal levels ofthese nutrients for psittacine birds have not beendetermined, therefore excessive supplementationshould be avoided until further research has beencompleted.
Behavioral Effects
Nutritional deficiency and excess can impact an-imal behavior in a variety of ways. First, a severenutrient deficiency can change the behavior of ananimal in terms of activity level. In rats, for exam-ple, deficiencies of protein, vitamin D, vitamin A,thiamin, riboflavin, or magnesium cause reduc-tions in activity (Hughes & Wood-Gush 1973).Calcium-deficient chicks have increased move-ment and pecking behaviors, while sodium-deficient chickens have increased pecking fre-quency (Hughes & Wood-Gush 1973). Second,some nutrient deficiencies result in a “specific ap-petite” for that particular nutrient. This term refersto the ability of an animal to identify the propor-tion of a particular nutrient in a feedstuff, and toadjust the consumption of that feedstuff relative toits nutrient needs (Murphy 1994). For example, aspecific appetite for calcium has been demon-strated in egg-laying female pheasants, in whichincreased consumption of a calcium-containingsupplement (limestone) occurred when birdswere fed calcium-deficient diets. While less re-search has been completed in birds as compared
to mammals, specific appetites for calcium(Hughes & Wood-Gush 1971a; Ranft & Hennig1993), sodium (Hughes & Wood-Gush 1971b),amino acids (Newman & Sands 1983; Noble et al.1993), protein (Ranft & Hennig 1993), energy(Ranft & Hennig 1993), and thiamin (Hughes &Wood-Gush 1971b) have been identified in birds.Third, undernourishment during post-hatch devel-opment may have severe consequences on the de-velopment of behavior and motor coordination,since cerebellum development (which plays amajor role in the development of motor coordina-tion in birds and mammals) is more affected byundernutrition than other parts of the brain. Inmammals, undernourished infants have reducedlocomotor activity, which is not reversed by re-feeding (Altman et al. 1971). Finally, novel be-havior effects resulting from nutritional defi-ciency or excess have recently been reported.Cockatiels fed deficient (0 IU vitamin A/kg diet),excessive (10,000 IU vitamin A/kg diet), or toxic(100,000 IU vitamin A/kg diet) levels of vitaminA had altered vocalization patterns compared toanimals maintained on adequate vitamin A diets(2,000 IU/kg diet) (Koutsos et al. 2001c). Speci-fically, the total number of vocalizations (Figure6.1), average length of vocalizations (reduced by
6 / Captive Parrot Nutrition: Interactions with Anatomy, Physiology, and Behavior 55
Figure 6.1. Effect of dietary vitamin A level onthe number of vocalizations over a two-minuteperiod made by adult female Cockatiels(Nymphicus hollandicus) at maintenance. Birdshad consumed assigned diets for three monthsprior to vocalization analysis. Data from Koutsoset al. 2001c.

high or low vitamin A), peak frequency (Hz) ofvocalizations (reduced by high or low vitamin A),and peak amplitude (dB) and total power (dB)(reduced by deficient diets) were significantly af-fected by dietary vitamin A level.
In addition to the effects of nutrient deficiencyand excess, other types of behavioral responses todiet and nutrition have been demonstrated. First,specific preferences for food types have beendemonstrated in cockatoos (Rowley et al. 1989).In general, when offered a choice of seed size,smaller birds preferred smaller seeds and largerbirds preferred larger seeds, although individualpreference was quite variable. Additionally, thenovelty of a food item may play a role in the ani-mal’s response, since animals tend to ingestsmaller amounts of a novel food as compared toconsumption of a familiar food (Forbes 1998).Second, “nutritional enrichment” provides amechanism for altered behavior patterns in re-sponse to diet choice and/or presentation. For ex-ample, providing sources of foraging enrichmentaltered parrot behavior (Coulton et al. 1997).Specifically, birds spent more time with the en-richment tool, altered the time spent on otherperches, and increased allopreening events. Inaddition, birds chose to work for food by partici-pating in the foraging enrichment apparatus, al-though identical food was freely available. A will-ingness to “work” for food has been observed ina variety of species. When starlings (Sturnus vul-garis) were offered a choice between freely avail-able mealworms and hidden mealworms (re-quiring searching and foraging behaviors), theypreferentially foraged for and ate hidden meal-worms (Inglis & Ferguson 1986). Only whenbirds were nutritionally deprived for long periodsof time did they choose freely available meal-worms. Similar results have been seen in pigeons(Neuringer 1969) and in domestic fowl (Duncan& Hughes 1972). Finally, aversion to a novel foodwill occur if the food contains excessive toxins,has a major nutrient imbalance (e.g. amino acidimbalance), and/or causes nausea or discomfort(Forbes 1998).
CONCLUSIONS
Determining the nutrient requirements of an ani-mal requires knowledge of its wild-type feedingstrategy, gastrointestinal anatomy and physiology,
and calculated and theoretical nutrient require-ments. Once nutrient requirements have been es-tablished, diets may be selected to meet those nu-trient needs, although several considerationsshould be made at this time. First, the bird’s evo-lutionary adaptations may direct the choice offeedstuffs (e.g., incorporating large seeds in thecase of a large parrot or liquid diet in the case ofa nectarivore). Second, knowledge of specific ap-petites can allow for effective supplementation ofdiets (e.g., providing a supplemental calciumsource to egg-laying birds). Third, knowledge ofgustatory preferences and aversions can facilitatediet formulation by maximizing palatability andfeed acceptance. Fourth, providing enrichmentthrough nutrition (e.g., providing foraging oppor-tunities) may enhance the animal’s feeding expe-rience and reduce stereotypic behaviors. Finally,it is critical to observe an animal subjected to di-etary changes in order to evaluate its response tothe new diet and, if necessary, to respond appro-priately to changes in the animal’s condition,health, and behavior.
REFERENCES
Abramson, J, B.L. Speer, and J.B. Thomson. 1995. Thelarge macaws. Fort Bragg, CA: Raintree.
Altman, J., K. Sudarshan, G.D. Das, N. McCormick,and D. Barnes. 1971. The influence of nutrition onneural and behavioral development. 3. Developmentof some motor, particularly locomotor patterns dur-ing infancy. Dev Psychobiol 4 (2):97–114.
Bartholomew, G.A., and T.J. Cade. 1958. Effects ofsodium chloride on the water consumption of housefinches. Phys Zool 31:304–310.
Bonadie, W.A., and P.R. Bacon. 2000. Year-round utili-sation of fragmented palm swamp forest by red-bellied macaws (Ara manilata) and orange-wingedparrots (Amazona amazonica) in the Nariva Swamp(Trinidad). Biol Cons 95:1–5.
Brindley, L.D. 1965. Taste discrimination in bobwhiteand Japanese quail. Anim Behav 16:304–307.
Brindley, L.D., and S. Prior. 1968. Effects of age ontaste discrimination in the bobwhite quail. AnimBehav 16 (2):304–307.
Coulton, L.E., N.K. Waran, and R.J. Young. 1997.Effects of foraging enrichment on the behaviour ofparrots. Anim Welfare 6 (4):357–363.
Crocker, D.R., S.M. Perry, M. Wilson, J.D. Bishop, andC.B. Scanlon. 1993. Repellency of cinnamic acidderivatives to captive rock doves. J Wildl Manag 57(1):113–122.
56 Manual of Parrot Behavior

Downs, C.T., and M.R. Perrin. 1996. Sugar preferencesof some southern African nectarivorous birds. Ibis138 (3):455–459.
Duncan, C.J. 1960. Preference tests and the sense oftaste in the feral pigeon (Columba livia Var Gmelin).Anim Behav 8:54–60.
Duncan, I.J.H., and B.O. Hughes. 1972. Free and oper-ant feeding in domestic fowls. Anim Behav20:775–777.
El Boushy, A.R., A.F.B. Van der Poel, J.C.J. Verhaart,and D.A. Kennedy. 1989. Sensory involvement con-trols feed intake in poultry. Feedstuffs 61 (25):16–41.
Forbes, J.M. 1998. Dietary awareness. Appl AnimBehav Sci 57 (3–4):287–297.
Frankel, T.L., and D.S. Avram. 2001. Protein require-ments of rainbow lorikeets, Trichoglossus haemato-dus. Aust J Zool 49:435–443.
Fuerst, W.F., and M.R. Kare. 1962. The influence of pHon fluid tolerance and preferences. Poult Sci 41:71–77.
Galetti, M. 1993. Diet of the scaly-headed parrot(Pionus maximiliani) in a semideciduous forest inSoutheastern Brazil. Biotropica 25:419–425.
Garnett, S., and G. Crowley. 1995. Feeding ecology ofhooded parrots Psephotus dissimilis during the earlywet season. Emu 95:54–61.
Gentle, M.J. 1972. Taste preference in the chicken(Gallus domesticus L.). Br Poult Sci 13 (2):141–155.
Glendinning, J.I. 1994. Is the bitter rejection responsealways adaptive? Physiol Behav 56 (6):1217–1227.
Harriman, A.E. 1967. Laughing gulls offered saline inpreference and survival tests. Physiol Zool40:273–279.
Harriman, A.E., and E.G. Fry. 1990. Solution accept-ance by common ravens (Corvus corax) given two-bottle preference tests. Psychol Rep 67 (1):19–26.
Harriman, A.E., and M.R. Kare. 1966. Aversion tosaline solutions in starlings, purple grackles, andherring gulls. Physiol Zool 39:123–126.
Harriman, A.E., and J.S. Milner. 1969. Preference forsucrose solutions by Japanese quail (Coturnix co-turnix japonica) in two-bottle drinking tests. AmMidland Natur 81:575–578.
Herness, M.S., and T.A. Gilbertson. 1999. Cellularmechanisms of taste transduction. Annu Rev Physiol61:873–900.
Homberger, D.G., and A.H. Brush. 1986. Functional-morphological and biochemical correlations of thekeratinized structures in the African gray parrot,Psittacus erithacus (Aves). Zoomorphology (Berl)106:103–114.
Hughes, B.O., and D.G.M. Wood-Gush. 1971a. A spe-cific appetite for calcium in domestic chickens.Anim Behav 19:490–499.
Hughes, B.O., and D.G.M. Wood-Gush. 1971b.Investigations into specific appetites for sodium andthiamine in domestic fowls. Physiol Behav 6(4):331–339.
Hughes, B.O., and D.G.M. Wood-Gush. 1973. An in-crease in activity of domestic fowls produced by nu-tritional deficiency. Anim Behav 21:10–17.
Inglis, I.R., and N.J.K. Ferguson. 1986. Starlings searchfor food rather than eat freely-available identicalfood. Anim Behav 34 (2):614–617.
Jackson, S., S.W. Nicolson, and C.N. Lotz. 1998. Sugarpreferences and “side bias” in cape sugarbirds andlesser double-collared sunbirds. Auk 115 (1):156–165.
Jones, D. 1987. Feeding ecology of the cockatiel,Nymphicus hollandicus, in a grain-growing area.Aust Wild Res 14:105–115.
Kamphues, J., W. Otte, and P. Wolf. 1997. “Effects ofincreasing protein intake on various parameters ofnitrogen metabolism in grey parrots (Psittacuserithacus erithacus).” In First International Sympo-sium on Pet Bird Nutrition, ed. J. Kamphues, P. Wolf,and N. Rabehl, p. 118. Hannover, Germany: Institutfur Tierernahrung.
Kare, M.R., R. Black, and E.G. Allison. 1957. Thesense of taste in the fowl. Poult Sci 36:129–138.
Kare, M.R., and J.R. Mason. 1986. “The chemicalsenses in birds.” In Avian physiology, ed. P.D.Sturkie and C.A. Benzo, pp. 59–73. New York:Springer-Verlag.
Kare, M.R., and H.L. Pick. 1960. The influence of thesense of taste on feed and fluid consumption. PoultSci 39:697–706.
Klasing, K.C. (1998). Comparative avian nutrition.New York: CAB International.
Koutsos, E.A., and K.C. Klasing. 2001. “Interactionsbetween the immune system, nutrition, and produc-tivity of animals.” In Recent advances in animal nu-trition, ed. P.C. Garnsworthy & J. Wiseman.Nottingham: Nottingham University Press.
Koutsos, E.A., and K.C. Klasing. 2002. “Vitamin A nu-trition of cockatiels.” In Proc. Comparative Nutri-tion Society, pp. 210–211. Silver Spring, MD:Comparative Nutrition Society.
Koutsos, E.A., K.D. Matson, and K.C. Klasing. 2001a.Nutrition of birds in the order Psittaciformes: A re-view. J Avian Med Surg 15:237–275.
Koutsos, E.A., H.N. Pham, J.R. Millam, and K.C. Klas-ing. 2001b. “Vocalizations of cockatiels (Nymphicushollandicus) are affected by dietary vitamin A con-centration.” 35th International Congress of theISAE, Davis, CA.
Koutsos, E.A., J. Smith, L. Woods, and K.C. Klasing.2001c. Adult cockatiels (Nymphicus hollandicus)undergo metabolic adaptation to high protein diets. JNutr 131:2014–2020.
6 / Captive Parrot Nutrition: Interactions with Anatomy, Physiology, and Behavior 57

Lane, R.A. 1996. “Avian hematology: Basic cell identi-fication, white blood cell count determinations, andclinical pathology.” In Diseases of cage and aviarybirds, ed. W.J. Rosskopf and R.W. Woerpel, pp.739–772. Baltimore: Williams and Wilkins.
Leshchinsky, T.V., and K.C. Klasing. 2001. Rela-tionship between the level of dietary vitamin E andthe immune response of broiler chickens. Poult Sci80 (11):1590–1599.
Long, J.L., and P.R. Mawson. 1994. Diet of regent par-rots (Polytelis anthopeplus) in the south-west ofWestern Australia. Western Austr Natur 19:293–299.
Mack, A.L., and D.D. Wright. 1998. The vulturine par-rot, Psittrichas fulgidus, a threatened New Guineaendemic: Notes on its biology and conservation.Bird Cons Int 8:185–194.
MacMillen, R.E., and R.V. Baudinette. 1993. Watereconomy of granivorous birds: Australian parrots.Functional Ecol 7 (6):704–712.
Malakoff, D. 1999. Following the scent of avian olfac-tion. Science 286 (5440):704–705.
Martinez del Rio, C. 1990. Sugar preferences in hum-mingbirds: The influence of subtle chemical differ-ences on food choice. Condor 92 (4):1022–1030.
Martinez del Rio, C., H.G. Baker, and I. Baker. 1992.Ecological and evolutionary implications of diges-tive processes: Bird preferences and the sugar con-stituents of floral nectar and fruit pulp. Experientia48 (6):544–551.
Martuscelli, P. 1995. Ecology and conservation of thered-tailed amazon Amazona brasiliensis in south-eastern Brazil. Bird Cons Int 5:405–420.
Matson, K.D., J.R. Millam, and K.C. Klasing. 2000.Taste threshold determination and side-preference incaptive cockatiels (Nymphicus hollandicus). ApplAnim Behav Sci 69 (4):313–326.
Matson, K.D., J.R. Millam, and K.C. Klasing. 2001.Thresholds for sweet, salt, and sour taste stimuli incockatiels (Nymphicus hollandicus). Zoo Biol 20(1):1–13.
Mcfarland, D.C. 1991. The biology of the ground par-rot, Pezoporus wallicus, in Queensland [Australia]:I. Microhabitat use, activity cycle and diet. WildlifeRes 18:168–184.
Murphy, M.E. 1994. Dietary complementation by wildbirds: Considerations for field studies. J Biosci(Bangalore) 19 (4):355–368.
Neuringer, A.J. 1969. Animals respond for food in thepresence of free food. Science 166 (903):399–401.
Nevitt, G. 1999a. Foraging by seabirds on an olfactorylandscape. American Scientist 87 (1):46–53.
Nevitt, G. 1999b. Olfactory foraging in Antarcticseabirds: A species-specific attraction to krill odors.Marine Ecology Progress Series 177:235–241.
Newman, R.K., and D.C. Sands. 1983. Dietary selec-tion for lysine by the chicks. Physiology andBehaviour 31:13–20.
Noble, D.O., M.L. Picard, E.A. Dunnington, G. Uzu,A.S. Larsen, and P.B. Siegel. 1993. Food intake ad-justments of chicks: Short term reactions of geneticstocks to deficiencies in lysine, methionine or tryp-tophan. Br Poult Sci 34 (4):725–735.
NRC, ed. 1994. Nutrient requirements of poultry.Washington, D.C.: National Academy Press.
Pitter, E., and M.B. Christiansen. 1995. Ecology, statusand conservation of the red-fronted macaw Ararubrogenys. Bird Cons Int 5:61–78.
Polo, F.J., V.I. Peinado, G. Viscor, and J. Palomeque.1998. Hematologic and plasma chemistry values incaptive psittacine birds. Avian Dis 42 (3):523–535.
Pryor, G.S. 1999. Comparative protein requirementsand digestive strategies of three species of parrotswith distinct dietary specializations. Am Zool 39(5):93A.
Ranft, U., and A. Hennig (1993). “New experimentalresults on nutrient selection of laying hens.” InEndocrine and nutritional control of basic biologi-cal functions, ed. H. Lehnert, pp. 167–172. Seattle:Hogrefe & Huber.
Rosskopf, W.J., R.W. Woerpel, and R.A. Lane. 1985.“The hypocalcemia syndrome in African greys: Anupdated clinical viewpoint with current recommen-dations for treatment.” Proc Assoc Avian Vet:129–131.
Roudybush, T.E., and C.R. Grau. 1986. Food and waterinterrelations and the protein requirement for growthof an altricial bird, the cockatiel (Nymphicus hol-landicus). J Nutr 116:552–559.
Rowley, I., E. Russell, and M. Palmer. 1989. The foodpreference of cockatoos: An aviary experiment.Austr Wildl Res 16 (1):19–32.
Schall, J.J. 1990. Aversion of whiptail lizardsCnemidophorus to a model alkaloid. Herpetologica46 (1):34–38.
Toyne, E.P., and J.N.M. Flanagan. 1997. Observationson the breeding, diet and behaviour of the red-facedparrot Hapalopsittaca pyrrhops in southernEcuador. Bull Br Ornithologists’ Club 117:257–263.
Wyndham, E. 1980. Environment and food of thebudgerigar Melopsittacus undulatus. Aust J Ecol5:47–61.
Yang, R.S.H., and M.R. Kare. 1968. Taste response ofa bird to constituents of arthropod defense secre-tions. Ann Entomol Soc America 61:781–782.
58 Manual of Parrot Behavior

Although many people are attracted to parrots aspets by their playful antics and vocal mimicry,when looking at the time budgets of most parrotspecies in the wild one sees that these birds spendthe most time each day engaged in relatively quietand sedate activities. Sleep and rest occupy themajor part of a 24-hour day. The activities thattake up the greatest portions of their waking dayare foraging and grooming. Grooming and othercomfort behaviors are major behavioral activitiesfor parrots of all species.
There are two basic ways to look at sleep:studying the patterns of brain waves during thedifferent phases of sleep and looking at sleep aspart of an animal’s daily activity pattern.Electroencephalographic (EEG) studies, record-ing brain activity, electromyographic (EMG)studies, recording muscular activity, and elec-trooculographic studies, recording eye move-ments, have been performed in several species ofbirds, including two psittacine species, Half-moon Conures (Aratinga canicularis) and Bud-gerigars (Melopsittacus undulatus) (Ayala-Guerrero et al. 1987, 1989). As in mammals,birds have been found to have two phases ofsleep. The first, slow wave sleep (SWS) is charac-terized by slow, high-voltage brain waves. Thistype of sleep appears to be the most importantsleep stage, the one that may have restorativefunctions. The second, called paradoxical sleep(PS) or REM (rapid eye movement) sleep, showslow-voltage brain waves, similar to those seenduring wakefulness. These low-voltage brainwaves are often accompanied by movement of theeyes. In people, dreaming occurs during PS. PSmay be involved in brain development and learn-ing (Carlson 2001).
In the psittacines studied, PS was always pre-
ceded by SWS. These birds were found to havespent shorter overall percentages of their totalsleep time in PS than mammals or most otheravian species. However, this may representchanges in the birds’ sleep patterns due to beingkept in constant light for the studies, not an actualdifference in psittacine PS as compared to otherbirds (Ayala-Guerrero et al. 1987, 1989).
Missing from the sleep of all avian speciesstudied are sleep spindles, bursts of activity thatare thought to represent a mechanism that de-creases the individual’s sensitivity to sensoryinput. Sleep spindles arise from the neocortex, astructure that is not well developed in the avianbrain (Ookawa 1972; Ayala-Guerrero et al. 1987).
Unihemispheric sleep was observed in theHalf-moon Conures studied. Unihemisphericsleep is a means of maintaining predator detec-tion where only one hemisphere of the brain isasleep at a time. EEGs of the conures showedtimes of low-voltage fast waves in one cerebralhemisphere and large amplitude slow waves in theother hemisphere (Ayala-Guerrero et al. 1987). Itis believed that while flying, especially on migra-tory flights over waters, birds engage in unihemi-spheric sleep. Unihemispheric sleep has been ob-served in most orders of birds and in marinemammals (Rattenburg et al. 2000).
Sleep is the single behavior that occupies thegreatest proportion of a parrot’s day (Rowley1990; Snyder 1987; Wirminghaus et al. 2001).Even when kept under constant illumination, theHalf-moon Conure was found to spend almost57% of a 24-hour period in some state of sleep(drowsiness, SWS, or PS). Budgerigars in similarconditions slept for an average of 38% of a 24-hour day. These studies also showed sustained pe-riods of slow wave sleep and an increase in para-
59
7Comfort Behavior and Sleep
Laurie Bergman and Ulrike S. Reinisch

doxical sleep between 7:00 P.M. and 7:00 A.M.This implies that this sleep pattern is due to en-dogenous circadian rhythms, not simply due toentrainment by external stimuli (Ayala-Guerreroet al. 1987, 1989).
As a social species, a parrot flock usuallysleeps and rests as a group. In general, a flock’swaking day begins with sunrise or before, withthe first light of the day, and ends with sunset.Parrot flocks usually sleep in a roosting area thatis distinct from the feeding area where they havespent the day. In the case of breeding pairs with anest the roosting area is near or at the nest site.Non-breeding birds may roost in trees that areconvenient to the area where they have been feed-ing. As dusk approaches birds will return to theirroosting area or a nearby convenient area. Duringthe middle of the day parrots will also have a pe-riod of quiet time during which they may sleep ordrowse (Rowley 1990; Snyder 1987; Wirming-haus et al. 2001). During this rest period the birdsmay experience short periods of SWS and PS(Ayala-Guerrero et al. 1987).
A parrot typically sleeps while perching up-right or lowered in a somewhat horizontal posi-tion with its body touching the perch. As the par-rot becomes drowsy it will fluff its feathers andblinking and eye movements will decrease. Oncethe bird is asleep its eyes are closed. In REMsleep the eyes remain closed and the head maydroop as neck muscle tone decreases. At the endof a period of PS the birds often suddenly raisetheir heads but do not awaken, returning to SWS.In prolonged sleep parrots turn their heads 180degrees and tuck their heads under their scapularfeathers. Parrots may sleep with both feet on theperch or with one foot raised (Ayala-Guerrero etal. 1987; Rowley 1990).
Although sleep is recognized as an essentialbehavior in both humans and animals and is thesubject of much research, little is actually knownabout the functions of sleep. It is believed thatSWS provides a restorative period for the brainand PS is involved in memory and learning.Laboratory animals subjected to prolonged sleepdeprivation eventually died (Carlson 2001). It hasbeen estimated that as much as 50% of the adulthuman population of America is sleep deprived(Maas 1999). This may also be true for ourpsittacine pets and may contribute to some behav-ior problems. As most species of parrots are from
tropical or semi-tropical regions, their days nor-mally have roughly 12 hours of light and 12 hoursof dark (Forshaw 1989). Despite the laboratoryfindings that parakeets kept in constant light con-tinued to do the bulk of their sleeping between7:00 P.M. and 7:00 A.M. (Ayala-Guerrero et al.1987, 1989), it appears that many pet parrots arenot given the opportunity to sleep as they wouldin the wild. Pet parrots are often kept in cages lo-cated in the main living areas of homes, wherethey are exposed not only to durations of “day-light” that extend beyond 12 hours/day but also tonoises and visual stimulation from televisions, ra-dios, and people moving about the house. Petparrots should be provided with a quiet, darksleeping area, ideally separate from their daytimeliving area (e.g., a separate small sleep cage), andclose to 12 hours of “nighttime” to sleep.
In addition to sleep, parrots spend a great dealof time engaged in grooming: preening and otherbody maintenance behaviors. Collectively thesebehaviors are often called comfort behaviors. Notonly do these behaviors normally occur when thebird is comfortable and at ease but the behaviorsthemselves appear to be comforting and soothingto the birds. It is probably due to this aspect ofthese behaviors that they often appear as displace-ment behaviors when parrots are under stress orbehaviorally conflicted. This can lead to these be-haviors becoming liberated from their normalcontexts and appearing as “problem behaviors”ranging from minor annoyances to owners to self-injurious behaviors such as feather picking andself-mutilation.
Grooming occupies the largest amount of awild parrot’s waking hours after foraging (Rowley1990; Snyder 1987). Preening serves severalfunctions, starting with maintenance of feathers.Maintaining feathers in good condition is not onlycrucial for flight but also for thermoregulation,waterproofing, camouflage, and communication(Cech et al. 2001). Other grooming behaviors thathave been noted in parrots include scratchingwith the feet, cleaning feet and legs with the beak,stretching, yawning, and beak rubbing and grind-ing (Lefebvre 1982; Rowley 1990; Wirminghauset al. 2000).
Despite their widespread geographic distribu-tion, parrots of different species tend to have sim-ilar grooming behaviors (Lefebvre 1982; Rowley1990; Wirminghaus et al. 2000). Based on a study
60 Manual of Parrot Behavior

of grooming behavior in Budgerigars, groomingtends to proceed from head-to-toe as has alsobeen found to be the case in several species ofmammals. Also in common with other speciesstudied, Budgerigars moved from grooming oneregion of the body to the next based mainly on ad-jacency and clustered their grooming in distinctanatomical regions (e.g., head, wings and trunk,lower region, and preen gland) (Lefebvre 1982).Individual feathers are preened by grasping thefeather in the beak and pulling it between theupper and lower beak while nibbling at thefeather. This serves to reattach disconnected bar-bules (Rowley 1990).
Long and short grooming bouts were noted inthe Budgerigars studied. Bouts were defined as asequence of grooming acts that was uninterruptedby non-grooming behavior for less that 30 sec-onds. Short bouts contained seven or feweracts/bout, whereas the number of acts/bout in along bout could be over 100. Short bouts seemedto occur mainly in response to an irritating stimu-lus. Short bouts of grooming often includedbehaviors such as shaking, stretching, andscratching, whereas preening took place almostexclusively in long bouts (Lefebvre 1982).
The other types of grooming behaviors thathave been described in parrots are also well pre-served between the species. Scratching with feetis an important grooming behavior. Because ofthese birds’ flexibility they are able to reach mostareas on the head that are inaccessible to preeningwith their feet. Stretching has also been describedin parrots, often occurring after a period of restbefore beginning another activity, including as theopening sequences of a grooming bout. Yawning,also called jaw or beak stretching, commonly oc-curs. There are also a variety of stretches that in-volve the wings, including arching the wings overthe back and stretching a wing and ipsilateral legdownward at the same time. This stretch is ac-companied by tail fanning. This procedure is thenrepeated with the other wing and leg. This oftenoccurs before flight. Other grooming behaviorsinclude chewing at inedible objects and beakgrinding, both of which serve to keep the beak ingood condition by wearing down any overgrowthand sharpening the cutting edge of the mandible.Beak grinding produces a soft, audible grindingsound (Lefebvre 1982; Rowley 1990).
In addition to self-directed grooming, when
given the opportunity, parrots also engage in al-logrooming. Allogrooming serves a variety offunctions. It allows for grooming of areas, specif-ically the head, which are inaccessible by the in-dividual being groomed. It also is an importantsocial behavior. Allopreening is often reciprocaland usually, but not always, takes place between abonded pair or parents and offspring. However,allopreening does occur between non-related,non-bonded individuals in a flock. Allopreeningis usually solicited by one bird lowering its headto present its neck to be preened. The preeningbird then moves on to groom the neck, head, andface of the other bird. The birds will often switchroles after a bout of allopreening. In Budgerigars,the proportion of allopreening given versus re-ceived was almost the same (Lefebvre 1982;Rowley 1990). The importance of allopreeningcan be seen in the amount of time devoted to it.Galahs have been reported to engage in allopreen-ing sessions that last as long as five minutes(Rowley 1990). In Budgerigars allopreening oc-cupied almost the same proportion of longgrooming bouts as preening of the wings(Lefebvre 1982).
Parrot flocks will groom for a brief period inthe morning before flying from their nighttimeroosting area to forage for the day. After feedingand filling their crops the flock will rest in a con-venient tree (near the foraging area or, if they arefeeding young, in the roosting/nesting area). Thebirds will groom while digesting their morningmeal. This pattern is repeated with grooming afteran afternoon meal and upon arriving at the roost-ing area at dusk (Rowley 1990; Snyder 1987).
As was noted earlier, comfort behaviors canbecome problematic, either of their own right oras signs of underlying stress. Shortly after one ofthe authors adopted a rescued parrot that had beenhoused in a dark basement, she noticed smallpiles of black powder in the bird’s cage in themorning. These piles were the result of the birdcontinually grinding his beak through the night.In this case, the bird did not do any permanentdamage to himself and, as he became more re-laxed and less fearful in his new home, the beakgrinding stopped. Pet parrots may substitute pet-ting from their owners for allogrooming, whichmay contribute to problems with aggression, vo-calizations, and misplaced sexual behavior, due to“pair bonding” with the owner. These and other
7 / Comfort Behavior and Sleep 61

behavior problems related to comfort behaviors,such as feather picking and self-mutilation, arediscussed in chapters 18, 21, and 23.
REFERENCES
Ayala-Guerrero, F., M.C. Perez, and A. Calderon. 1987.Sleep patterns in the bird Aratinga canicularis.Physiology and Behavior 43 (5):585–589.
Ayala-Guerrero, F. 1989. Sleep patterns in the parakeetMelopsittacus undulates. Physiology and Behavior46 (5):787–791.
Carlson, N.R. 2001. Physiology of behavior. Boston:Allyn and Bacon.
Cech, R., J.B. Dunning Jr., and C. Elphick. 2001.“Behavior.” In The Sibley guide to bird life and be-havior, ed. Chris Elphick, John B. Dunning, Jr., andDavid Allen Sibley, pp. 51–79. New York; Alfred A.Knopf.
Forshaw, J.M. 1989. Parrots of the world, 3rd (rev.) ed.London: Blandford.
Lefebvre, L. 1982. The organization of grooming inbudgerigars. Behavioural Processes 7:93–106.
Maas, J. 1999. Power sleep: The revolutionary programthat prepares your mind for peak performance. NewYork: Harper Collins.
Ookawa, T. 1972. Avian wakefulness and sleep on thebasis of recent electroencephalographic observa-tions. Poultry Science 51:1565–1574.
Rattenburg, W.C. et al. 2000. Behavioral, neurophysio-logical and evolutionary perspectives on unihemi-spheric sleep. Neuroscience and BiobehavioralReviews 24 (8):817–842.
Rowley, I. 1990. Behavioral ecology of the galah,Eolophus roseicapillus: In the wheatbelt of WesternAustralia. Chipping Norton, NSW: Surrey Beatty.
Snyder, N.F.R. 1987. The parrots of Luquillo: Naturalhistory and conservation of the Puerto Rican parrot.Los Angeles: Western Foundation of VertebrateZoology.
Wirminghaus, J.O., C.T. Downs, C.T. Symes, E.Dempster, and M.R. Perrin. 2000. Vocalizations andbehaviours of the cape parrot Poicephalus robustus(Psittaciformes:Psittacidae). Durban MuseumNovitates 25:12–17.
Wirminghaus, J.O., C.T. Downs, M.R. Perrin, C.T.Symes. 2001. Abundance and activity patterns of thecape parrot (Poicephalus robustus) in two afromon-tane forests in South Africa. African Zoology 36(1):71–77.
62 Manual of Parrot Behavior

INTRODUCTION
Thorough knowledge of parrot breeding biologyrequires information about the composition ofbreeding groups, the process by which individu-als form breeding relationships, nest-site estab-lishment, parental care patterns, and factors thatinfluence breeding success. A comprehensive un-derstanding also requires an evaluation and ap-preciation of the variation both between andwithin species regarding these aspects ofpsittacine reproductive behavior. On the surface,parrots appear to represent a fairly uniform groupwith respect to reproductive biology, but recentevidence suggests more variability and complex-ity than initially believed. Researching parrot re-productive biology not only has a basic scientificimportance but has practical implications as well.Given the poor conservation status of a signifi-cant number of parrot species (Collar & Juniper1992; Bennett & Owens 1997), an in-depth un-derstanding of the reproductive systems ofpsittacines becomes crucial to protecting andrestoring declining populations. Managementpractices both in the wild and in captivity wouldbenefit from additional rigorous scientific studiesof the reproductive behavior of psittacines (e.g.,Monterrubio et al. 2002; Manning et al. 2004).For example, recent studies indicate that supple-mental feeding of breeding female Kakapos, anextremely endangered psittacine with a rare lek-type mating system (see later), appears to result inan undesirable male bias in offspring sex ratios(Clout et al. 2002; Sutherland 2002).
With these issues in the forefront, I will beginthis review of psittacine reproductive behaviorwith a few caveats about methodology. Next, thischapter will review the mating systems of parrotswith a particular eye toward discerning variationwithin species and discrepancies between the so-cial and genetic aspects of mating patterns.Finally, I will turn to behaviors used by parrotsduring mate assessment, courtship, and breedingand follow with a consideration of psittacine par-ental care. I give preference to studies conductedon wild populations or semi-natural captivegroups that allow individuals to express variationin behavior. Compared to captive breeding situa-tions with individuals housed in pairs, large cap-tive populations of mixed sex and age likely offera more complete picture of the process by whichindividuals form breeding relationships and thedegree of variation that exists in the species. Suchstudies likely provide the most valid and reliablescientific information. On a practical note, man-agement practices in the wild and in captivity thatpreserve the behavioral variation in a species andfulfill the behavioral needs of individuals willlikely have the most success in breeding produc-tivity and long-term conservation goals (Derrick-son & Snyder 1992).
The following discussion of psittacine matingpatterns and reproductive behavior requires a fewmethodological qualifications. First, assessmentsof psittacine mating systems are often based onobservations of the number of parrots attending anest or the social associations between individu-
63
8Parrot Reproductive Behavior,
or Who Associates,Who Mates, and Who Cares?
Tracey R. Spoon

als within a larger flock. For example, based onobservations of two individuals attending a nestor duos of birds associating within a larger flock,researchers usually assert that a particular speciesexhibits monogamy, a mating system in whicheach breeding unit consists of one male and onefemale. However, in order to definitively statethat a species truly exhibits a particular matingsystem, the members of breeding units must beindividually identifiable either through naturallyoccurring physical differences or marks placed byobservers—a formidable task for many parrotspecies. Due to the inherent difficulty of trackingbirds beyond the immediate vicinity of the nestsite, a member of an assumed monogamous pairmay attend more than one nest, or more than onemale or female may attend a single nest unbe-knownst to an observer who cannot identify indi-viduals. For instance, in a study of Glossy BlackCockatoos (Calyptorhynchus lathami), a speciesidentified as strongly monogamous, occasionallya male approached and fed an incubating femaleonly to be chased off by a different male who alsofed the female and then accompanied her back tothe nest (Garnett et al. 1999). Determining the so-cial and mating relationships in such cases re-quires long-term studies on known individuals.
A second difficulty in studying the reproduc-tive behavior of parrots rests on the fact that manyspecies display sexual monomorphism (Forshaw1981, 1989; Alderton 1991); in other words,males and females cannot be readily distin-guished based on physical characteristics. Inmany cases, individuals are assigned a sex basedon behavioral differences in nest attendance andsexual behavior. Because sex identification thenrelies on behavioral differences, this methodologymay obscure rather than reveal important behav-ioral variability within the sexes both betweenand within species.
MATING SYSTEMS
Although a moderate degree of variation existsamong parrot species, most psittacines appear toexhibit social monogamy, meaning that the pre-dominant breeding unit consists of one male andone female (Forshaw 1981, 1989; Higgins 1999).Social monogamy differs from genetic mono-gamy in that socially monogamous pairmates mayassociate primarily with each other and mayjointly establish a nest but may copulate and cre-
ate offspring with an individual other than theirsocial mate (Dunn & Lifjeld 1994). Because ofdifficulties observing parrots in the wild and test-ing genetic parentage, most studies have focusedon the social aspects of mating systems. For ex-ample, detailed studies on identifiable individualsin the wild have revealed that White-tailed BlackCockatoos (Calyptorhynchus funereus, Saunders1982), Puerto Rican Amazons (Amazona vittata,Snyder et al. 1987), Galahs (Eolophus roseicapil-lus, Rowley 1990), Major Mitchell’s Cockatoos(Cacatua leadbeateri, Rowley & Chapman1991), Green-rumped Parrotlets (Forpus passeri-nus, Waltman & Beissinger 1992), Monk Para-keets (Myiopsitta monachus, Eberhard 1998), andBurrowing Parrots (Cyanoliseus patagonus,Masello et al. 2002) predominantly or exclusivelyexhibit social monogamy. Additional studies inthe wild on Crimson Rosellas (Platycercus ele-gans, Krebs 1998), Glossy Black Cockatoos(Garnett et al. 1999), and several species of ama-zon parrots (Yellow-headed, Amazona oratrix;Red-crowned, A. viridigenalis; Red-lored, A. au-tumnalis [Enkerlin-Hoeflich 1995]; Lilac-crowned, A. finschi [Renton & Salinas-Melgoza1999]; Black-billed, A. agilis, and Yellow-billed,A. collaria [Koenig 2001]) strongly suggest a so-cially monogamous mating system, although theidentification of individuals and sex in these stud-ies typically or occasionally relied on observa-tions of nest attendance, association patterns, andbehavioral patterns rather than an independent as-sessment of identification and sex. Likewise, inseveral studies on captive groups that allowed in-dividuals to express variability in mating patterns,several species of lovebirds (Agapornis species,Dilger 1960; Stamm 1962), Orange-frontedConures (Aratinga canicularis, Hardy 1963),Budgerigars (Melopsittacus undulatus, Brockway1964; Trillmich 1976), White-fronted Amazons(Amazona albifrons, Levinson 1980), Rainbowand Scaly-breasted Lorikeets (Trichoglossus hae-matodus and T. chlorolepidotus, Serpell 1981),Canary-winged Parakeets (Brotogeris versicol-orus, Arrowood 1987), Spectacled Parrotlets(Forpus conspicillatus, Garnetzke-Stollman &Franck 1991), and Cockatiels (Nymphicus hol-landicus, Spoon et al. 2004) predominatelyformed socially monogamous breeding groups.These monogamous pair relationships seem par-ticularly strong as evidenced by the close associ-
64 Manual of Parrot Behavior

ation between mates who perform most of theirdaily activities together throughout the year (e.g.,Rainbow and Scaly-breasted Lorikeets [Serpell1981]; Canary-winged Parakeets [Arrowood1987]; Galahs [Rowley 1990]; SpectacledParrotlets [Garnetzke-Stollman & Franck 1991,Wanker et al. 1996 cited in Wanker et al. 1998];Cockatiels [Spoon et al. 2004]; Brown-headedParrots, Poicephalus cryptoxanthus [Taylor &Perrin 2004]). Galah mates, for example, return totheir nest site together every evening to roost(Rowley 1990).
Despite this tendency toward monogamy, sev-eral typically monogamous species exhibit varia-tion in breeding group composition. For example,polygamous groups have been observed inMasked Lovebirds (Agapornis personata, Stamm1962), Spectacled Parrotlets (Garnetzke-Stollman& Franck 1991), Yellow-headed Amazons(Enkerlin-Hoeflich 1995), Monk Parakeets(Eberhard 1998), and Cockatiels (Seibert &Crowell-Davis 2001). The role of these “extra”birds in breeding remains little understood.Normally the accessory bird in trios of Yellow-headed Amazons only flew with the breedingpair, but in one trio the accessory bird partici-pated minimally in nesting and was once ob-served feeding the female (Enkerlin-Hoeflich1995). In a captive group of Cockatiels, bothmales of a polygamous trio cared for the female’syoung although she was only observed to copu-late with one of them, and a male who associatedand copulated with two females subsequentlycared for the young of both in two separate nestboxes (Seibert & Crowell-Davis 2001). As notedpreviously, instances of two birds feeding onenesting female have been recorded in GlossyBlack Cockatoos (Garnett et al. 1999). In twoMonk Parakeet trios consisting of one female andtwo males, both males participated in nest build-ing and feeding the female. Interestingly, a com-plete copulation bout between the two males wasobserved near the nest in which the female wasincubating (Eberhard 1998). In a Monk Parakeettrio of two females and one male, all three partic-ipated in nest building but only one female incu-bated the eggs (Eberhard 1998). Not only dopolygamous trios occasionally occur in MonkParakeets, but the species may exhibit some de-gree of cooperative or communal breeding amongpairs with incipient helping-at-the-nest as well
(Eberhard 1998). Evidence also suggests thatGolden Conures (Aratinga guarouba) may pos-sess a communal breeding system in whichgroups of several males and females or possiblygroups of pairs utilize the same nest cavity withmultiple females contributing eggs and multiplemale and female attendants subsequently caringfor the young; single pairs may also breed alone(Oren & Novaes 1986; Forshaw 1989). This coop-erative or communal pattern is quite differentfrom the more general pattern in which monoga-mous pairs aggregate with conspecifics duringfeeding and roosting and then defend the nest sitefrom other pairs during breeding (Rowley &Chapman 1991; Wanker et al. 1998).
In addition to variation in the composition ofbreeding units, the degree of permanency of pairrelationships also varies between individuals andspecies. Long-term studies on White-tailed BlackCockatoos (Saunders 1982), Puerto RicanAmazons (Snyder et al. 1987), Galahs (Rowley1990), and Major Mitchell’s Cockatoos (Rowley& Chapman 1991) indicate that mates often re-main together until the death of one member. Inthese studies, very few of the former mates of in-dividuals known to have changed mates were ob-served again, suggesting that the mate change oc-curred because the former mate died and left theremaining member without a mate. However,these results also indicate that occasionally indi-viduals of these species switch mates despite thefact that their former mate remains alive. In sev-eral additional studies, observations indicate thatindividuals usually retain their mates betweenbreeding seasons (e.g., Green-rumped Parrotlets,Waltman & Beissinger 1992; Yellow-headed andRed-crowned Amazons, Enkerlin-Hoeflich 1995;Monk Parakeets, Eberhard 1998; BurrowingParrots, Masello et al. 2002). In contrast, captiveCockatiels (Spoon 2002) and Canary-wingedParakeets (Arrowood 1987) were not as devotedto their mates; when several previously confinedpairs were housed together or additional birdswere added to the flock, several paired Cockatielsand Canary-winged Parakeets, respectively,switched mates. Similarly, in one study of captiveMasked Lovebirds, mate change frequently oc-curred (Stamm 1962), yet Dilger (1960) claimedthat lovebird mates usually remain together untildeath, based on his study of several lovebirdspecies. Spectacled Parrotlet pairs remained sta-
8 / Parrot Reproductive Behavior, or Who Associates, Who Mates, and Who Cares? 65

ble even after changes in group composition(Garnetzke-Stollman & Franck 1991). Based onthese studies it is difficult to tell whether captiv-ity increases the frequency of mate change ormerely renders it more easily observed.
As mentioned previously, social monogamydoes not necessarily mean genetic monogamy.Galahs (Rowley 1990) and Budgerigars (Brock-way 1964; Baltz & Clark 1997), for example, willengage in sexual behavior and copulate outsidethe pair relationship, but often restrict these extra-pair interactions to times when their mates are inthe nest cavity or otherwise unable to observe theinfidelity. Paired Cockatiels appear to use extra-pair sexual behavior and copulation to assess po-tential mates and form new pair relationships,apparently in response to a low degree of compat-ibility with their current mate (Spoon 2002).Among captive Spectacled Parrotlets, Garnetzke-Stollman and Franck (1991) observed extra-pair copulations only under one very specificcircumstance—the secondary female of a polyga-mous trio copulated repeatedly with the male of aneighboring pair while his female incubated aclutch. Interestingly, this extra-pair relationshipinvolved only sexual behavior with no affiliativebehavior or courtship feeding (Garnetzke-Stollman & Franck 1991). Very few studies haveactually assessed the genetic mating systems ofparrots. In a captive group of Golden Conures,the eggs of one female were fertilized by twomales, but no information was available on thesocial relationships between the female and malesor on which individuals cared for the eggs oryoung (Oren & Novaes 1986). In contrast, a de-tailed study of wild Burrowing Parrots discoveredno instances of extra-pair paternity, suggestingthat these parrots exhibit genetic as well as socialmonogamy (although the study did identify onecase of apparent intraspecific brood parasitism)(Masello et al. 2002).
Despite the apparent predominance of socialmonogamy among parrots, some species exhibitmating systems vastly different from socialmonogamy. The Kakapo (Strigops habroptilus)and Vasa Parrot (Coracopsis vasa), for instance,display mating systems that differ dramaticallyfrom the socially monogamous systems just de-scribed. The highly endangered Kakapo of NewZealand appears to possess a lek-type breedingsystem in which males establish courts called
track-and-bowl systems used to attract femalesfor copulation (Merton et al. 1984; Powlesland etal. 1992). During mating periods, which occuronly once every two to five years, females travelto the male’s court, which appears to be usedsolely for mating and not for feeding or nestingby the female. Otherwise, Kakapos live solitarily,and females alone assume all parental responsi-bilities (Powlesland et al. 1992). The Greater VasaParrot exhibits a strongly polygynandrous matingsystem in which both males and females copulatewith several individuals (Ekstrom 2002). Maleand female Vasa Parrots do not form pair relation-ships or pair bonds typical of most parrots, andfemales generally lay clutches sired by multiplemales.
PAIR RELATIONSHIPS
Although the parrot family exhibits greater diver-sity regarding mating systems than previously be-lieved, the most complete information on repro-ductive behavior exists primarily for sociallymonogamous species. For these species, the rela-tionship between pairmates not only forms thebasis of the breeding unit, but because these rela-tionships often last year-round for multiple years,they also form the basis of parrot social organiza-tion. In some species, establishing a pair relation-ship may prove vital to social rank within theflock. For instance, Spectacled Parrotlet pair-mates hold the same rank within the flock, andmonogamous pairs hold the highest ranks(Garnetzke-Stollman & Franck 1991). Similarly,Orange-fronted Conure pairmates appear to helpeach other establish a higher rank, and when apair member is removed, the remaining memberoften loses status (Hardy 1963). Major Mitchell’sCockatoos offer further evidence of the impor-tance of pair relationships in parrot reproductiveand social behavior—reproductively immaturebirds may form pair bonds up to a year prior toactually breeding (Rowley & Chapman 1991).Thus, understanding these pair relationships is crucial to understanding parrot reproductivebehavior.
Affiliative interactions such as close proxim-ity, allopreening, and reduced aggression typi-cally characterize associations between psitta-cine pairmates. The predominant pattern appearsto be for mates to follow each other closely andmaintain significantly closer proximity to each
66 Manual of Parrot Behavior

other than to non-mates. As a result, matesremain together almost constantly and performmany of their daily activities together (e.g.,Budgerigars [Brockway 1964; Trillmich 1976];Rainbow and Scaly-breasted Lorikeets [Serpell1981]; Canary-winged Parakeets [Arrowood1987]; Galahs [Rowley 1990]; Spectacled Par-rotlets [Garnetzke-Stollman & Franck 1991];Major Mitchell’s Cockatoos [Rowley & Chap-man 1991]; Cockatiels [Spoon et al. 2004]).Between mates of several lovebird species, main-tenance behaviors hold a strong mimetic valuesuch that a bird often joins its mate in perform-ing particular behaviors such as preening (Dilger1960). Similarly, Cockatiel pairmates not onlymaintain significantly closer proximity to theirmates but also exhibit greater behavioral syn-chrony, meaning that mates often perform thesame behaviors simultaneously (Spoon et al.2004). Indeed, Cockatiel pairs that exhibitgreater affiliative behavior and synchrony andless aggression enjoy greater reproductive suc-cess and display less extra-pair sexual behavior(Spoon 2002; Spoon et al., in press). This strongpreference to associate with a mate extends to theauditory realm as well; in controlled choice stud-ies, Spectacled Parrotlets prefer the contact callsof their mates over non-mates (Garnetzke-Stollman & Franck 1991). While in close prox-imity, members of a pair often engage in bouts ofallopreening in which they preen as well as so-licit preening from their partners. In most so-cially monogamous parrot species, allopreeningoccurs primarily with a mate rather than non-mate (e.g., lovebirds, Dilger 1960; Orange-fronted Conures, Hardy 1963; Rainbow andScaly-breasted Lorikeets, Serpell 1981; Galahs,Rowley 1990; Green-rumped Parrotlets, Walt-man & Beissinger 1992) and in some studies hasbeen observed only between established mates(e.g., White-tailed Black Cockatoos, Saunders1974; Budgerigars, Trillmich 1976; Canary-winged Parakeets, Arrowood 1987; SpectacledParrotlets, Garnetzke-Stollman & Franck 1991;Cockatiels, Spoon et al. 2004). Bill touching, an-other affiliative behavior of Budgerigars, also oc-curs predominantly between mates (Trillmich1976). In addition to increased affiliative behav-iors, Budgerigars (Trillmich 1976) and Cock-atiels (Spoon et al. 2004) display reduced aggres-sion toward mates compared to non-mates.
Moreover in many parrot species, mates displaythese various affiliative behaviors during non-breeding periods, indicating the year-round na-ture of these relationships (e.g., Orange-frontedConures [Hardy 1963]; Cockatiels [Zann 1965;Spoon et al. 2004]; Puerto Rican Amazons[Snyder et al. 1987]; Galahs [Rowley 1990]). Asan additional indication of the specificity ofthese pair relationships, paired SpectacledParrotlets (Garnetzke-Stollman & Franck 1991)and Budgerigars (Trillmich 1976), for example,tend not to display affiliative behaviors towardtheir extra-pair sexual partners.
Many psittacine species also display coopera-tive or coordinated pair behaviors especially dur-ing agonistic interactions with non-mate con-specifics. For example, Orange-chinned Parakeet(Brotogeris jugularis, Power 1966), Canary-winged Parakeet (Arrowood 1987), Puerto RicanAmazon (Snyder et al. 1987), and Yellow-napedAmazon (Amazona auropalliata, Wright & Dorin2001) mates perform coordinated duets duringagonistic or territorial encounters with non-mates. Likewise, in several different Trichoglos-sus (lorikeet) species, Serpell (1981) recordedeight distinct types of cooperative displays withboth vocal and visual elements performed bymates, again often during agonistic interactionswith conspecifics. Hardy (1963) also describedhighly coordinated vocalizations given byOrange-fronted Conure mates while approachingtheir nest site.
Although pair relationships form a critical partof most parrot social groups and many affiliativeand sexual behaviors characterize those relation-ships, the process of pair formation appears sub-tle and remains poorly understood. For example,Arrowood (1987) and Hardy (1963) could iden-tify no specific or obvious behavioral methodsused to attract a mate or form pairs in Canary-winged Parakeets and Orange-fronted Conures,respectively. In Spectacled Parrotlets, young firstestablish close sibling relationships and thenoften form a series of non-exclusive relationshipswith potential mates until exclusive pair forma-tion occurs (Garnetzke-Stollman & Franck 1991;Wanker et al. 1996). Similarly, in Cockatiels, amale often approaches and sings to one or moreselected females until the female permits sexualor affiliative behavior (Zann 1965; T. Spoon, per-sonal observation). Interestingly, in the one case
8 / Parrot Reproductive Behavior, or Who Associates, Who Mates, and Who Cares? 67

of pair formation in Cockatiels that Zann (1965)observed, pairing appeared complete once the fe-male allowed the male to allopreen her; in con-trast, in the several instances of pair formationthat I observed in Cockatiels, copulation often oc-curred prior to allopreening and seemed to con-sistently indicate the occurrence of pair formation(Spoon 2002). Furthermore, pair formation oftenoccurs outside of an immediate breeding context(e.g., Budgerigars, Trillmich 1976; Cockatiels,Spoon et al. 2004). In some cases, specific behav-iors do appear to be associated with pair forma-tion; for example, head shaking by maleBudgerigars (Brockway 1964) and head bowingby male Puerto Rican Amazons (Snyder et al.1987) seem to be associated primarily with pairformation. An intriguing series of studies onBudgerigars revealed that males learn to imitatethe contact calls of females with whom they arepaired (Hile et al. 2000) and that the ability to doso plays an important role in mate choice and pairformation (Hile 2002; Striedter et al. 2003). Afew studies have contributed vital information tounderstanding pair formation in parrots by exam-ining the characteristics that individuals use to se-lect a mate. Some degree of mate choice inBudgerigars depends on ultraviolet fluorescentproperties of the plumage; both males and fe-males prefer partners in which the fluorescentcharacteristics of the feathers remain visible(Pearn et al. 2001; Arnold et al. 2002). Deter-mining whether Budgerigars use natural variationin fluorescence to select among potential mateswould offer important insight into the process ofmate choice in parrots. Likewise, male and fe-male Burrowing Parrots may prefer mates withlarger abdominal red feather patches, which ap-pear to signal individual quality (Masello &Quillfeldt 2003). In a free-choice test, female butnot male Spectacled Parrotlets preferred potentialmates of high rank (Garnetzke-Stollman &Franck 1991). In this species, pairmates hold thesame rank within the flock and selecting a high-ranking mate may confer significant advantages.Cockatiels allowed to self-select a mate experi-ence a reproductive advantage over pairs formedrandomly by investigators (Yamamoto et al.1989); several non-psittacine species display asimilar pattern (Klint & Enquist 1981; Bluhm1985; Lupo et al. 1990; Bottoni et al. 1993; Ryan& Altmann 2001).
COURTSHIP AND COPULATION
In addition to affiliative and cooperative behav-iors, courtship and pre-copulatory displays repre-sent an important component of parrot reproduc-tive behavior. The documented courtship displaysof many socially monogamous psittacines containseveral common elements. For example, male dis-plays of several species in the Cacatuinae sub-family involve at least partially raised crests,switch sidling or strutting back and forth along aperch, head bowing, and flaring the wings orholding the carpal joints slightly away from thebody (e.g., Cockatiels [Zann 1965; Spoon 2002];White-tailed Black Cockatoos [Saunders 1974;Forshaw 1981]; Red-tailed Black Cockatoos[Calyptorhynchus banksii, Forshaw 1981]).Forms of repetitive strutting are also observed insuch diverse species as Orange-fronted Conures(Hardy 1963), Budgerigars (Brockway 1964),Black-billed Amazons (Cruz & Gruber 1980),Monk Parakeets (Eberhard 1998), and severallovebird species (Dilger 1960). Likewise, malesof various species such as Orange-frontedConures (Hardy 1963), Budgerigars (Brockway1964), Thick-billed Parrots (Rhynchopsitta pachy-rhyncha, Lanning & Shiflett 1983), White-frontedAmazons (Skeate 1984), Puerto Rican Amazons(Snyder et al. 1987), Green-rumped Parrotlets(Waltman & Beissinger 1992), and several love-bird species (Dilger 1960) perform some type ofhead bobbing and/or head bowing display. PuertoRican Amazon (Snyder et al. 1987) and Green-rumped Parrotlet males (Waltman & Beissinger1992) also perform rapid wing flicking as part oftheir courtship displays. As in many bird species,vocalizations also comprise an important aspectof parrot courtship behavior (e.g., Orange-frontedConures [Hardy 1963]; Budgerigars [Brockway1964]; Cockatiels [Zann 1965; Spoon 2002];White-tailed Black Cockatoos [Saunders 1974];King Parrots, Alisterus scapularis [Forshaw1981]). However, unlike passerines, most parrotsdo not produce vocalizations regarded as song.Males of a few parrot species produce courtshipvocalizations that may be considered song in thatthe vocalizations consist of melodious, recogniz-able syllables (e.g., warbling in Budgerigars[Brockway 1964]; courtship song in Orange-fronted Conures [Hardy 1963] and Cockatiels[Zann 1965; Spoon 2002]). Warbling by maleBudgerigars not only correlates with sperm pro-
68 Manual of Parrot Behavior

duction but also stimulates female ovarian activ-ity and thus represents an integral component ofreproductive behavior in this species (Brockway1965). Interestingly, female Budgerigars occa-sionally warble but at a substantially lower rateand complexity than males (Farabaugh et al.1992). Administering testosterone to adult maleand female Budgerigars appears to lower thethreshold for male-typical courtship behaviors in-cluding vocalizations (Brockway 1968; Nespor etal. 1996). Jackson (1963) also describes a yodel-ing song produced by female Kakas (Nestormeridionalis), and Ekstrom (2002) reportssinging by female Greater Vasa Parrots in the con-text of allofeeding (see later).
Because the courtship activities of the lek-breeding Kakapo illustrate the diversity that ex-ists within psittacines, they deserve special no-tice. During breeding periods, males activelymaintain their courts known as track-and-bowlsystems by excavating up to several shallow de-pressions or bowls and cleaning the trails thatconnect them (Merton et al. 1984). From theircourts, males of this flightless, nocturnal speciesproduce courtship vocalizations including boom-ing, which can be heard up to several kilometersaway. Indeed, males often locate their bowls at thebase of natural sound reflectors. As mentionedpreviously, the track-and-bowl systems appear tobe used exclusively for courtship and mating andnot feeding or nesting. In fact, a male’s track-and-bowl system usually occurs several kilometersfrom his home range (Merton et al. 1984).
Observers typically have recorded only malesperforming courtship behaviors in most parrotspecies. However, in a few species, observershave noted females displaying as well. For exam-ple, female Cockatiels often join their male part-ners in switch sidling, especially in instances thatlead to female copulation solicitation (Zann 1965;T. Spoon, personal observation). In King Parrots,both males and females erect their head feathersbut sleek the remaining body feathers, contracttheir pupils, and call (Forshaw 1981), and inPuerto Rican Amazons males and females engagein a mutual bowing display during pair formation(Snyder et al. 1987). Either member of Green-rumped Parrotlet pairs may initiate courtship be-havior, which often occurs in conjunction withnest prospecting and normally consists of headbowing combined with tail fanning alternating
between mates (Waltman & Beissinger 1992).Female Budgerigars usually remain motionless(or may leave or respond aggressively) to malepre-copulatory courtship but occasionally re-spond with nudging or pumping, behavioral com-ponents typical of male courtship, prior to solicit-ing copulation (Brockway 1964).
Courtship feeding or allofeeding occurs be-tween pairmates in numerous parrot species. Inthe following discussion, I use the terms“allofeeding” and “courtship feeding” inter-changeably to refer to regurgitation of food byone individual for another excluding the provi-sioning of a mate during incubation or broodingand the feeding of chicks. Yet, courtship feedingand provisioning an incubating female appear in-terrelated—courtship feeding occurs primarily inspecies in which the male provides food for thefemale while she alone incubates. For example,species such as White-tailed Black Cockatoos(Saunders 1982), Puerto Rican Amazons (Snyderet al. 1987), Green-rumped Parrotlets (Waltman& Beissinger 1992), Monk Parakeets (Eberhard1998), Orange-fronted Conures (Hardy 1963),Rainbow and Scaly-breasted Lorikeets (Serpell1981; Forshaw & Cooper 1981), and Budgerigars(Brockway 1964) exhibit courtship feeding and aparental care system in which the female alone in-cubates while the male provisions her; in contrast,species such as Gang-gang Cockatoos (Callo-cephalon fimbriatum, Forshaw 1981), Galahs(Rowley 1990), and Cockatiels (Spoon 2002)rarely if ever engage in allofeeding and exhibit aparental care system in which both members ofthe pair share incubation duties and the male doesnot provision the female.
Most authors prefer the term “courtship feed-ing” because the behavior often occurs in associ-ation with courtship displays and copulation.Moreover, some courtship displays such as headbobbing and bill clasping appear to be ritualizedforms of courtship feeding (Dilger 1960;Brockway 1964; Waltman & Beissinger 1992).Allofeeding or an apparently ritualized formoccurs in a courtship context in Budgerigars(Brockway 1964), King Parrots (Forshaw 1981),Green-rumped Parrotlets (Waltman & Beissinger1992), and several lovebird species (Dilger 1960).In species such as Spectacled Parrotlets(Garnetzke-Stollman & Franck 1991), Green-rumped Parrotlets (Waltman & Beissinger 1992),
8 / Parrot Reproductive Behavior, or Who Associates, Who Mates, and Who Cares? 69

Puerto Rican Amazons (Snyder et al. 1987),White-tailed Black Cockatoos (Saunders 1982),and Monk Parakeets (Eberhard 1998), allofeedingonly occurs during, or at least increases in fre-quency at the beginning of, the breeding cycle. InPuerto Rican Amazons, for example, courtshipfeeding occurs in close association with copula-tion, although the success rate of copulation re-mains similar whether or not courtship feedingoccurs (Snyder et al. 1987). However, in somespecies allofeeding occurs throughout the year oreven if restricted to the breeding season may notbe directly associated with copulation. Allofeed-ing occurs year-round in Orange-fronted Conures(Hardy 1963), White-fronted Amazons (Skeate1984), and several lovebird species (Dilger 1960;Stamm 1962) although in lovebirds the frequencyincreases with the onset of breeding (Dilger1960). In most psittacines, only the male allo-feeds the female; however, female SpectacledParrotlets (Garnetzke-Stollman & Franck 1991)and Puerto Rican Amazons (Snyder et al. 1987)rarely but occasionally feed their mates, andMadagascar (Agapornis cana) and AbyssinianLovebird (A. taranta) females frequently feedtheir mates (Dilger 1960).
Some form of courtship behavior typically pre-cedes copulation, another important componentin the reproductive lives of parrots. Copulation inparrots tends to be a lengthy affair compared toother birds whose copulations generally last onlya few seconds (Birkhead & Møller 1992). Thefollowing examples illustrate the extended dura-tion of copulation in a variety of socially monog-amous psittacines: mean of 30 seconds but up to1.5 minutes in Budgerigars (Brockway 1964), ap-proximately one minute in Black-billed Amazons(Koenig 2001) and White-tailed Black Cockatoos(Saunders 1974), approximately 1.5 minutes inOrange-fronted Conures (Hardy 1963) andGalahs (Rowley 1990), 1.5–2 minutes in Cock-atiels (Zann 1965; T. Spoon, personal observa-tion), approximately 1–3.5 minutes in Green-rumped Parrotlets (Waltman & Beissinger 1992),a few seconds to more than six minutes in love-birds (Dilger 1960; Stamm 1962), and a mean ofslightly more than 4.5 minutes with multiplemounts in Monk Parakeets (Eberhard 1998). Themost remarkable copulation sequence belongs tothe polygynandrous Greater Vasa Parrot, in whichcopulation may exceed 1.5 hours and begins with
the male mounting the female but ends with thebirds perched side by side while their cloacae re-main locked together (Wilkinson & Birkhead1995). During the breeding season, the cloacae ofVasa Parrot males and, to a lesser extent, femalesbecome greatly enlarged, and during copulation,the male’s cloacal protrusion appears to com-pletely enter the female’s opening. While the tworemain interlocked, the male makes vigorous cop-ulatory movements and may allofeed the femaleseveral times (Wilkinson & Birkhead 1995). Inaddition to the duration of copulation, anotherdifference between parrots and most birds is thatmale parrots mount by stepping onto rather thanflying onto the back of the female (Dilger 1960;Hardy 1963; Brockway 1964; Eberhard 1998; T.Spoon, personal observation). Male Thick-billed(Lanning & Shiflett 1983) and amazon parrots(Amazona species, Snyder et al. 1987) usually donot even fully mount the female during copula-tion but place one foot on the female’s back whilethe other foot grasps the perch. An interestingtwist occurs in Peach-faced Lovebirds and thewhite eye-ringed group of lovebirds in whichpseudofemale copulation solicitation by malessometimes results in female mounting; the sexu-ally dichromatic lovebird species, on the otherhand, do not appear to display this behavioral re-versal (Dilger 1960). The degree to which parrotslimit copulation to the breeding season variesamong species. Green-rumped Parrotlets (Walt-man & Beissinger 1992), Budgerigars (Trillmich1976), Galahs (Rowley 1990), and Vasa Parrots(Wilkinson & Birkhead 1995) tend to limit copu-lation primarily or entirely to around the time ofnest prospecting and egg laying. In contrast,Cockatiel mates copulate throughout year (Zann1965) and often multiple times a day, although thefrequency tends to be highest during pair forma-tion and breeding (Spoon et al. 2004).
NESTING
Although the majority of parrot species nest intree cavities, psittacines exhibit a moderate de-gree of variability in nest sites. Most psittacinesare secondary cavity nesters, meaning that theyutilize existing cavities in trees as nest sites(Forshaw 1981, 1989). Some species modify thecavity by adding layers of nesting material; forexample, Palm Cockatoos (Probosciger ater-rimus) may create a layer of twigs two to three
70 Manual of Parrot Behavior

meters deep in the bottom of a cavity (Forshaw1981), and Galahs line their nests with greenleaves, the only cockatoo species known to usegreen material (Rowley 1990). Members of thegenus Agapornis display a great deal of diversityin nest construction inside their nesting hollows,ranging from a simple pad in Madagascar andAbyssinian Lovebirds to a well-formed cup inPeach-faced Lovebirds to a roofed nest chamberwith tunnel in Masked Lovebirds (Dilger 1960).Additionally, many parrot species, while cavitynesters, use chambers in termite or ant mounds(Red-Faced Lovebirds, Agapornis pullaria[Dilger 1960]; Orange-fronted Conures [Hardy1963]; Golden-shouldered Parrots, Psephotuschrysopterygius [Forshaw 1981]; Canary-wingedParakeets [Paranhos & Marcondes-Machado2000]) or cliff faces rather than trees (Maroon-fronted Parrots, Rhynchopsitta pachyrhynchusterrisi [Lawson & Lanning 1980]; BurrowingParrots [Masello et al. 2001]). Rock Parakeets(Neophema petrophila) nest in crevices in rocksor under rock overhangs (Forshaw 1981). The useof alternative nesting niches such as chambers intermitaria, ant mounds, and cliffs may haveevolved in response to increased predation pres-sures corresponding to the evolutionary radiationof mammalian predators (Brightsmith 2005).Several species exhibit the flexibility to nest in avariety of cavities; Paradise Parakeets (Psephotuspulcherrimus) may use chambers in termitaria orin banks along watercourses (Forshaw 1981),Bahama Amazons (Amazona leucocephala ba-hamensis) typically nest in tree cavities but willuse limestone karst cavities in the ground on is-lands lacking suitable cavity-containing trees(Snyder et al. 1987), Puerto Rican and His-paniolan Amazons (A. ventralis) may nest in ei-ther tree or cliff cavities (Snyder et al. 1987), andGalahs nest primarily in tree cavities but will nestin concrete pipes and crevices in rock and clifffaces (Rowley 1990). On the other hand, GroundParrots (Pezoporus wallicus), Night Parrots(Geopsittacus occidentalis), and Kakapos do notnest in formal cavities but use shallow excava-tions or hollows located under overhanging vege-tation or near the base of a shrub or tree (Forshaw1981; Powlesland et al. 1992). Similarly, Keas(Nestor notabilis) nest in burrows normally exca-vated under boulders (Diamond & Bond 1999).Monk Parakeets are unusual among parrots in
that each breeding pair uses sticks to construct anenclosed nest chamber, which is often integratedwith other such chambers into a large compoundnest (Navarro et al. 1992; Eberhard 1998).Interestingly, both breeder and non-breeder MonkParakeets participate in building the large, com-pound structure that they actively maintain year-round and use for roosting during non-breedingperiods as well as nesting during the breedingseason (Navarro et al. 1992; Eberhard 1998).However, twig theft commonly occurs betweenpairs, suggesting a lack of true cooperation in nestconstruction (Eberhard 1998).
Spacing between conspecific nests also variesgreatly between species, ranging from colonialsituations with little distance between nests towidely dispersed distributions with kilometersbetween nests. Species such as Maroon-frontedParrots (Lawson & Lanning 1980; Snyder et al.1987), Monk Parakeets (Eberhard 1998), Burrow-ing Parrots (Masello et al. 2002), and Peach-facedand the white-eye ringed group of lovebirds(Dilger 1960) nest colonially with little distanceand often minimal aggression between breedingpairs. In contrast, Major Mitchell’s Cockatoosmaintain an average distance of almost three kilo-meters between nests (Rowley & Chapman 1991)due to intense aggression by breeding pairs to-ward conspecifics (Saunders et al. 1985). Highlevels of intraspecific aggression around nestingtrees by Blue-and-yellow Macaws (Ara ararauna)may prevent the use of many apparently suitablenesting cavities and limit the density of breedingpairs (Renton 2004). In between these extremes,many species appear to exhibit an intermediatepattern in which nests appear somewhat clumpedbut not as much so as colonially breeding species(e.g., Thick-billed Parrots, Lanning & Shiflett1983; Galahs, Rowley 1990; several Amazonaspecies, reviewed in Snyder et al. 1987 andEnkerlin-Hoeflich 1995). The degree of clumpingand aggression between breeding pairs may re-late, in part, to the availability of suitable nestingsites. Bahama Parrots nesting in a region with su-perabundant nest sites in the form of solutionholes in the ground exhibit little territorial ag-gression and close nest spacing compared toheightened territorial aggression exhibited by thesame species in an area with few potential nestsites (Snyder et al. 1987). Some species such asGalahs maintain an interest in their nest sites
8 / Parrot Reproductive Behavior, or Who Associates, Who Mates, and Who Cares? 71

throughout the year, and pairs return to them toroost every evening (Rowley 1990). During thenon-breeding season, Puerto Rican Amazon pairsroost near their nests only intermittently and de-fend the territory somewhat but much less vigor-ously than during the breeding season (Snyder etal. 1987). Defense of nest sites by Imperial(Amazona imperialis) and Hispaniolan Amazonpairs has been observed during non-breedingtimes (Snyder et al. 1987). In contrast, White-tailed Black Cockatoos, Red-tailed Black Cocka-toos, and Corellas (Cacatua tenuirostris) inhabittheir nesting areas only during breeding (Saun-ders 1982; Saunders et al. 1982).
Following nest-site selection and preparation,egg laying, incubation, and chick rearing com-mence. “Parrot encyclopedias” such as Forshaw(1981, 1989), Alderton (1991), Collar (1997), andHiggins (1999) provide species information,when available, on age at first breeding, breedingseason, clutch size, incubation, and fledging.Masello and Quillfeldt (2002) also provide a use-ful summary of various breeding parameters forpsittacines reported in the literature. Due to spaceconstraints, I will discuss general trends and no-table variability in these reproductive parametersand refer readers to these sources for species-specific information. In general, parrots exhibitdelayed sexual maturity; many of the large-bodiedspecies of cockatoos, for example, do not reachsexual maturity until approximately four to fiveyears of age (Forshaw 1981), while Budgerigarsmay pair and breed in captivity as young as fivemonths of age (Kavanau 1987). Natural popula-tions of parrots exhibit a remarkable range inclutch size from one in some large cockatoos(Red-tailed Black [Smith & Saunders 1986];Glossy [Garnett et al. 1999]; Palm [Murphy et al.2003]) to an average of six (range 1–11) in MonkParakeets (Navarro et al. 1992) and seven (range5–10) in Green-rumped Parrotlets (Waltman &Beissinger 1992). In species with variable clutchsize, the female’s nutritional status may affectclutch size; Green-rumped Parrotlet females fedmore often by their mates during egg laying sub-sequently produced larger clutches (Waltman &Beissinger 1992). Among Australian psittacines,average clutch size decreases, egg weight in-creases, and incubation duration increases asspecies body weight increases (Saunders et al.1984). A similar trend regarding clutch size and
body weight occurs among Amazona species(Enkerlin-Hoeflich 1995). This trend does notapply universally across all parrot taxa, however;Double-eyed Fig Parrots (Opopsitta dioph-thalma), for example, have both a small bodyweight and small clutch size (average two eggs)(Forshaw 1981). Thus, in species of large-bodiedparrots such as Palm Cockatoos, the convergenceof late sexual maturity, small clutch size, failureto breed every year, extremely low nesting suc-cess (81% of active nests failed to produce fledg-lings in one population of Palm Cockatoos), andstrong competition for limited nesting sites resultin populations highly sensitive to environmentalperturbations (Murphy et al. 2003).
PARENTAL CARE
In most parrot species, the female incubates alonewhile the male provides regurgitated food to her(Forshaw 1981, 1989). In these species, the fe-male usually leaves the nest only to receive food.For example, incubating female Glossy BlackCockatoos normally leave the nest only in theevening when the male returns from foraging tofeed her (Garnett et al. 1999). Similarly, femaleWhite-tailed Black Cockatoos remain in the nestduring incubation except to receive food from themale in the morning and evening (Saunders1982). Female Green-rumped Parrotlets spend onaverage about 85% of their time during the day inthe nest, and males feed the incubating femalesalmost once per hour during this time (Waltman& Beissinger 1992). An interesting provisioningpattern emerges among amazon parrots—in gen-eral, males of mainland species feed their matesonly twice per day, whereas males of islandspecies feed their mates multiple times each day(Enkerlin-Hoeflich 1995; Renton & Salinas-Melgoza 1999). This pattern may relate to differ-ential predation pressure, but again, exceptionsexist to this general trend. Yellow-billed Amazonson the island of Jamaica display the twice-dailyfeeding pattern (Koenig 2001). Even the polygy-nandrous Vasa Parrot conforms to the pattern offemale-only incubation coupled with male provi-sioning but with an interesting twist—femalestypically receive food from several males andeach male provisions several females at widelyseparated nests (Ekstrom 2002). Furthermore, inkeeping with the association between food andsex, female Vasa Parrots generally perform ritual-
72 Manual of Parrot Behavior

ized or false copulation (in the sense that it doesnot involve cloacal contact or the prolonged join-ing described previously) with each of the provi-sioning males.
However, the parrot family contains several no-table exceptions to this pattern of female incuba-tion and male provisioning. The Black Cockatoosof the genera Probosciger and Calyptorhynchusexhibit the aforementioned pattern of female-onlyincubation and male provisioning, whereas thecockatoo genera of Eolophus, Cacatua, Callo-cephalon, and Nymphicus exhibit biparental incu-bation without males provisioning females(Rowley 1990). Among these latter species, matesmust coordinate incubation shifts to ensure thattheir eggs receive adequate coverage and protec-tion. Cockatiel mates that demonstrate greater co-ordination of incubation subsequently experiencegreater hatching success of fertile eggs, and thosemates that exhibit greater affiliation and behav-ioral synchrony and less aggression subsequentlydisplay greater incubation coordination and thushatching success (Spoon et al., in press). Kakaposalso exhibit a somewhat different incubation pat-tern in that although the female alone broods,males do not provision incubating females orchicks (Merton et al. 1984; Powlesland et al.1992). Additional notable exceptions includeGolden Conures, in which multiple females maylay eggs in one nest and several males and fe-males may participate in nest attendance (Oren &Novaes 1986), and Monk Parakeets, in whichsome form of incipient helping at the nest by in-dividuals other than the pair members may occur(Eberhard 1998).
After hatching, parrot chicks experience a rela-tively lengthy period of dependence upon theirparents. Compared to raptors, owls, and pigeonsof equivalent body weight, parrots exhibit a rela-tively extended nestling period (Saunders et al.1984). With the only known exception of theKakapo, in which the female alone cares for herchicks, male and female psittacine parents jointlycare for their young. In general, in species withfemale-only incubation and male provisioning,the male initially continues to provision the fe-male who in turn feeds the newly hatched chicks;as the nestlings age, the female spends progres-sively more time away from the nest, returning tofeed the young with the male who may also beginto directly feed the chicks (several lovebird
species, Dilger 1960; White-tailed Black Cock-atoos, Saunders 1982; Thick-billed Parrots,Lanning & Shiflett 1983; Budgerigars, Stamps etal. 1985; Green-rumped Parrotlets, Waltman &Beissinger 1992; Monk Parakeets, Eberhard1998; Glossy Black Cockatoos, Garnett et al.1999; Lilac-crowned Amazons, Renton &Salinas-Melgoza 1999; Black-billed Amazons,Koenig 2001). Keas appear to take this pattern tothe extreme in that initially the male feeds the fe-male, who in turn feeds the young chicks, but asthe chicks age the male begins to feed them di-rectly, and by fledging the male alone feeds themand continues to do so alone for several weeksafter fledging (reviewed in Diamond & Bond1999). Although the Vasa Parrot displays a simi-lar chick-rearing pattern in that females feed thenestlings with regurgitated food received frommales, this species once again provides an inter-esting twist on this general pattern—only femaleVasa Parrots sing, and they appear to do so prima-rily during chick-rearing to attract provisioningmales. Females that sing more frequently or morecomplex songs receive more provisions frommales and produce larger clutches and broods(Ekstrom 2002). In contrast to the widespreadpattern of female-only incubation and male provi-sioning, species such as Galahs (Rowley 1990)and Cockatiels (Spoon 2002) exhibit a parentalcare system in which both parents incubate andfeed the chicks directly.
Another important aspect of parrot reproduc-tion is hatching asynchrony (Waltman &Beissinger 1992). Because many parrots begin in-cubation with the first egg but often continue tolay eggs for several days, a clutch may experiencestrong hatching asynchrony, resulting in chicks ofvastly discrepant sizes and competitive abilities.Because of this, last-hatched chicks may die ofstarvation. In Green-rumped Parrotlets, all chicksin small broods experienced uniformly high sur-vival, whereas the last and penultimately hatchedchicks in large clutches experienced reduced sur-vival, apparently due to starvation (Waltman &Beissinger 1992). A similar pattern occurs inBurrowing Parrots (Masello & Quillfeldt 2002).White-tailed Black Cockatoos offer an even moredramatic example with two eggs laid approxi-mately eight days apart. If the first chick survivesuntil the second egg hatches, the second-hatchedchick typically dies of starvation within a couple
8 / Parrot Reproductive Behavior, or Who Associates, Who Mates, and Who Cares? 73

of days (Saunders 1982). This pattern may repre-sent insurance against infertile eggs or, at least inthe case of the Green-rumped Parrotlet, mayallow pairs to raise additional young in exception-ally good years.
Parrot parents may either overcome or facili-tate the discrepant competitive abilities of chicksby taking an active role in the distribution of foodto chicks. Studies on Crimson Rosellas (Krebs &Magrath 2000; Krebs 2001) and Budgerigars(Stamps et al. 1985) demonstrate that parents canpreferentially allocate food to certain chicks andthat males and females may show different feed-ing preferences based on age and hunger level ofchicks. After fledging occurs, parents may con-tinue to feed the chicks for several weeks ormonths and may continue to associate with themfor several months (White-fronted Amazons,Skeate 1984; Corellas, Beeton 1985; Galahs,Rowley 1990; Major Mitchell’s Cockatoo,Rowley & Chapman 1991; Green-rumpedParrotlets, Waltman & Beissinger 1992; Keas,Diamond & Bond 1999; Burrowing Parrots,Masello et al. 2002; Cockatiels, T. Spoon, per-sonal observation) and in some cases until theparents enter their next reproductive cycle the fol-lowing year (e.g., White-tailed Black Cockatoo,Saunders 1982).
SUMMARY
Parrots are often regarded as a rather uniformgroup of species, and in many respects they con-form to this generalization. Most parrot speciesexhibit social monogamy and biparental care ofyoung. In these species, pair relationships com-monly endure for several breeding attempts andpersist throughout the year. In addition, many af-filiative and courtship behaviors characterizethese relationships. However, two known and in-triguing exceptions occur—lek-breeding Kakaposand highly polygynandrous Vasa Parrots, neitherspecies of which exhibits the close pair bonds ob-served in socially monogamous species. In addi-tion, Kakapo females alone care for their eggs andyoung, and Vasa females receive regurgitated foodprovisions from several males while caring for off-spring. Two species, Monk Parakeets and GoldenConures, may also engage in some degree of co-operative or communal breeding. Yet, even in so-cially monogamous species, individuals may not
display absolute sexual monogamy, and in someinstances polygamous breeding groups occur.Among Australian psittacines, larger-bodiedspecies produce smaller clutches of larger eggswith longer incubation and nestling periods.Parrots nest in cavities or chambers or at least inhollows under overhanging vegetation or rocks.Most parrots also exhibit a prolonged associationwith their parents and delayed sexual maturation.Another important characteristic of psittacine re-production involves a pronounced hatching asyn-chrony in many species that often results in thedeath of the youngest chicks.
Although researchers have made substantialprogress in the study of parrot reproductive be-havior, successful conservation and managementof these species both in the wild and in captivitydepend upon a more thorough understanding ofthe social and reproductive requirements of par-rots. At the most basic level, this requires infor-mation on the composition of breeding groups,the processes by which breeding relationshipsform, nest-site selection, and parental care pat-terns. A more in-depth but perhaps equally criti-cal understanding includes an appreciation of thenatural variability both within and betweenspecies. Unrecognized variability may thwartconservation efforts on multiple levels. Lack ofsuch knowledge may cause difficulties in provid-ing the social and physical environments that bestpromote breeding success. Similarly, that whichgoes unrecognized or unappreciated may be lost,fundamentally altering the behavioral and per-haps genetic composition of a species.
REFERENCES
Alderton, D. 1991. The atlas of parrots of the world.Neptune City, NJ: T. F. H. Publications.
Arnold, K.E., I.P.F. Owens, and N.J. Marshall. 2002.Fluorescent signaling in parrots. Science 295:92.
Arrowood, P.C. 1987. Duetting, pair bonding and ago-nistic display in parakeet pairs. Behaviour 106:129–157.
Baltz, A.P., and A.B. Clark. 1997. Extra-pair courtshipbehaviour of male budgerigars and the effect of anaudience. Animal Behaviour 53:1017–1024.
Beeton, R.J.S. 1985. The little corella: A seasonallyadapted species. Proceedings of the EcologicalSociety of Australia 13:53–63.
Bennett, P.M., and I.P. Owens. 1997. Variation in ex-tinction risk among birds: Chance or evolutionary
74 Manual of Parrot Behavior

predisposition? Proceedings of the Royal Society ofLondon, Series B 264:401–408.
Birkhead, T.M., and A.P. Møller. 1992. Sperm competi-tion in birds: Evolutionary causes and conse-quences. London: Academic Press.
Bluhm, C.K. 1985. Mate preferences and mating pat-terns in canvasback ducks (Aythya valisineria). InAvian monogamy, ed. P.A. Gowaty and D.W. Mock,pp. 45–56. Washington, DC: American Ornitho-logists’ Union.
Bottoni, L., R. Massa, R.W. Lea, and P.J. Sharp. 1993.Mate choice and reproductive success in the red-legged partridge (Alectoris rufa). Hormones andBehavior 27: 308–317.
Brightsmith, D.J. 2005. Competition, predation andnest niche shifts among tropical cavity nesters:Phylogeny and natural history evolution of parrots(Psittaciformes) and trogons (Trogoniformes).Journal of Avian Biology 36:64–73.
Brockway, B.F. 1964. Ethological studies of thebudgerigar: Reproductive behavior. Behaviour23:294–324.
Brockway, B.F. 1965. Stimulation of ovarian develop-ment and egg laying by male courtship vocalizationin budgerigars (Melopsittacus undulatus). AnimalBehaviour 13:575–578.
Brockway, B.F. 1968. Influences of sex hormones onthe loud and soft warbles of male budgerigars.Animal Behaviour 16:5–12.
Clout, M.N., G.P. Elliott, and B.C. Robertson. 2002.Effects of supplementary feeding on the offspringsex ratio of kakapo: A dilemma for the conservationof a polygynous parrot. Biological Conservation107:13–18.
Collar, N.J. 1997. “Family Psittacidae (parrots).” InHandbook of the birds of the world. Volume 4: Sand-grouse to cuckoos, ed. J. del Hoyo, A. Elliot, and J.Sargatal, pp. 280–477. Barcelona: Lynx Edicions.
Collar, N.J., and A.T. Juniper. 1992. “Dimensions andcauses of the parrot conservation crisis: Solutionsfrom conservation biology.” In New world parrots incrisis, ed. S.R. Beissinger and N.F.R. Snyder, pp.1–24. Washington, DC: Smithsonian InstitutionPress.
Cruz, A., and S. Gruber. 1980. “The distribution, ecol-ogy, and breeding biology of Jamaican amazon par-rots.” In Conservation of new world parrots:Proceedings of the ICBP Parrot Working GroupMeeting, ed. R.F. Pasquier, pp. 403–417. Wash-ington, DC: Smithsonian Institution Press.
Derrickson, S.R., and N.F.R. Snyder. 1992. “Potentialsand limits of captive breeding in parrot conserva-tion.” In New world parrots in crisis, ed. S. R.Beissinger and N. F. R. Snyder, pp. 133–163.Washington, DC: Smithsonian Institution Press.
Diamond, J., and A.B. Bond. 1999. Kea, bird of para-dox: The Evolution and Behavior of a New ZealandParrot. Berkeley: University of California Press.
Dilger, W.C. 1960. The comparative ethology of theAfrican parrot genus Agapornis. Zeitschrift furTierpsychologie 17:649–685.
Dunn, P.O., and J.T. Lifjeld. 1994. Can extra-pair pater-nity be used to predict extra-pair paternity in birds?Animal Behaviour 47:983–985.
Eberhard, J.R. 1998. Breeding biology of the monkparakeet. Wilson Bulletin 110:463–473.
Ekstrom, J. 2002. Singing for your dinner but not foryour mates: Female song and polygynandry in thegreater vasa parrot. Abstract from V. Kirindy Sym-posium, 23 August at University of Sheffield, UK.
Enkerlin-Hoeflich, E.C. 1995. Comparative ecologyand reproductive biology of three species ofAmazona parrots in northeastern Mexico. PhD diss.,Texas A&M University, College Station.
Farabaugh, S.M., E.D. Brown, and R.J. Dooling. 1992.Analysis of the warble song of the budgerigar(Melopsittacus undulatus). Bioacoustics 4:111–130.
Forshaw, J.M. 1981. Australian parrots, 2nd ed.Melbourne, Lansdowne Editions.
Forshaw, J.M. 1989. Parrots of the world, 3rd ed.London: Blanford Press.
Forshaw, J.M., & W.T. Cooper. 1981. Australian par-rots. Melbourne: Lansdowne Press.
Garnett, S.T., L.P. Pedler, and G.M. Crowley. 1999. Thebreeding biology of the glossy black cockatooCalyptorhynchus lathami on Kangaroo Island, SouthAustralia. Emu 99:262–279.
Garnetzke-Stollman, K., and D. Franck. 1991.Socialisation tactics of the spectacled parrotlet(Forpus conspicilllatus). Behaviour 119:1–29.
Hardy, J.W. 1963. Epigamic and reproductive behaviorof the orange-fronted parakeet. Condor 65:169–199.
Higgins, P.J., ed. 1999. Handbook of Australian, NewZealand, and Antarctic birds. Volume 4: Parrots todollarbirds. Victoria: Oxford University Press.
Hile, A.G. 2002. Effects of male vocal learning on fe-male behavior in the budgerigar. Brain, Behaviorand Evolution Abstracts 60:62–63.
Hile, A.G., T.K. Plummer, and G.F. Striedter. 2000.Male vocal imitation produces call convergence dur-ing pair bonding in budgerigars, Melopsittacus un-dulatus. Animal Behaviour 59:1209–1218.
Jackson, J.R. 1963. Studies at a kaka’s nest. Notornis10:168–175.
Kavanau, J.L. 1987. Lovebirds, cockatiels, budgeri-gars: Behavior and evolution. Los Angeles: ScienceSoftware Systems, Inc.
Klint, T., and M. Enquist. 1981. Pair formation and re-productive output in domestic pigeons. BehaviouralProcesses 6:57–62.
8 / Parrot Reproductive Behavior, or Who Associates, Who Mates, and Who Cares? 75

Koenig, S.E. 2001. The breeding biology of black-billed parrot Amazona agilis and yellow-billed par-rot Amazona collaria in Cockpit Country, Jamaica.Bird Conservation International 11:205–225.
Krebs, E.A. 1998. Breeding biology of crimson rosel-las Platycercus elegans in Black Mountain,Australian Capital Territory. Australian Journal ofZoology 46:119–136.
Krebs, E.A. 2001. Begging and food distribution incrimson rosella (Platycercus elegans) broods: Whydon’t hungry chicks beg more? Behavioral Ecologyand Sociobiology 50:20–30.
Krebs, E.A., and R.D. Magrath. 2000. Food allocationin crimson rosella broods: Parents differ in their re-sponses to chick hunger. Animal Behaviour59:739–751.
Lanning, D.V., and J.T. Shiflett. 1983. Nesting ecologyof thick-billed parrots. Condor 85:66–73.
Lawson, P.W., and D.V. Lanning. 1980. “Nesting andstatus of the maroon-fronted parrot (Rhynchopsittaterrisi).” In Conservation of new world parrots:Proceedings of the ICBP Parrot Working GroupMeeting, ed. R.F. Pasquier, pp. 385–392. Washing-ton, DC: Smithsonian Institution Press.
Levinson, S.T. 1980. “The social behavior of the white-fronted amazon (Amazona albifrons).” In Conser-vation of new world parrots: Proceedings of theICBP Parrot Working Group Meeting, ed. R.F.Pasquier, pp. 403–417. Washington, DC: Smithson-ian Institution Press.
Lupo, C., L. Beani, L., R. Cervo, R., L. Lodi, L., and F.Dessi-Fulgheri. 1990. Steroid hormones and repro-ductive history of the grey partridge (Perdix perdix).Bollettino di Zoologia 57:247–252.
Manning, A.D., D.B. Lindenmayer, and S.C. Barry.2004. The conservation implications of bird repro-duction in the agricultural “matrix”: A case study ofthe vulnerable superb parrot of south-easternAustralia. Biological Conservation 120:367–378.
Masello, J.F., G.A. Pagnossin, G.E. Palleiro, and P.Quillfeldt. 2001. Use of miniature security camerasto record behaviour of burrow-nesting birds.Vogelwarte 4:150–154.
Masello, J.F., and P. Quillfeldt. 2002. Chick growth andbreeding success of the burrowing parrot. Condor104:574–586.
Masello, J.F., and P. Quillfeldt. 2003. Body size, bodycondition, and ornamental feathers of burrowingparrots: Variation between years and sexes, assorta-tive mating and influences on breeding success. Emu103:149–161.
Masello, J.F., A. Stramkova, P. Quillfelt, J.T. Epplen,and T. Lubjuhn, T. 2002. Genetic monogamy in bur-rowing parrots Cyanoliseus patagonus. Journal ofAvian Biology 33:99–103.
Merton, D.V., R.B. Morris, and I.A.E. Atkinson. 1984.Lek behaviour in a parrot: The kakapo Strigopshabroptilus of New Zealand. Ibis 126:277–283.
Monterrubio, T., E. Enkerlin-Hoeflich, and R.B.Hamilton. 2002. Productivity and nesting success ofthick-billed parrots. Condor 104:788–794.
Murphy, S., S. Legge, and R. Heinsohn. 2003. Thebreeding biology of palm cockatoos (Proboscigeraterrimus): A case of a slow life history. Journal ofZoology 261:327–339.
Navarro, J.L., M.B. Martella, and E.H. Bucher. 1992.Breeding season and productivity of monk parakeetsin Cordoba, Argentina. Wilson Bulletin 104:413–424.
Nespor, A.A., M.J. Lukaszewicz, R.J. Dooling, andG.F. Ball. 1996. Testosterone induction of male-likevocalizations in female budgerigars (Melopsittacusundulatus). Hormones and Behavior 30:162–169.
Oren, D.C., and F. Novaes. 1986. Observations on thegolden parakeet Aratinga guarouba in northernBrazil. Biological Conservation 36:329–337.
Paranhos, S.J., and L.O. Marcondes-Machado. 2000.Breeding behavior of Brotogeris versicolorus chiriri(Aves, Psittacidae) in Sao Paulo, Brazil. IheringiaSerie Zoologia 88:61–66.
Pearn, S.M., A.T.D. Bennett, and I.C. Cuthill. 2001.Ultraviolet vision, fluorescence and mate choice in aparrot, the budgerigar Melopsittacus undulatus. Pro-ceedings of the Royal Society Biological SciencesSeries B 268:2273–2279.
Power, D.M. 1966. Antiphonal duetting and evidencefor auditory reaction time in the orange-chinnedparakeet. Auk 83:314–319.
Powlesland, R.G., B.D. Lloyd, H.A. Best, and D.V.Merton. 1992. Breeding biology of the kakapo Stri-gops habroptilus on Stewart Island, New Zealand.Ibis 134:361–373.
Renton, K. 2004. Agonistic interactions of nesting andnonbreeding macaws. Condor 106: 354–362.
Renton, K., and A. Salinas-Melgoza. 1999. Nesting be-havior of the lilac-crowned parrot. Wilson Bulletin111:488–493.
Rowley, I. 1990. Behavioural ecology of the galah,Eolophus roseicapillus. New South Wales, Aus-tralia: Surrey Beatty & Sons, Chipping Norton.
Rowley, I., and G. Chapman. 1991. The breeding biol-ogy, food, social organization, demography and con-servation of the Major Mitchell or pink cockatoo,Cacatua leadbeateri, on the margin of the WesternAustralia wheatbelt. Australian Journal of Zoology39:211–261.
Ryan, K.K., and J. Altmann. 2001. Selection for malechoice based primarily on mate compatibility in theoldfield mouse, Peromyscus polionotus rhoadsi.Behavioral Ecology and Sociobiology 50:436–440.
76 Manual of Parrot Behavior

Saunders, D.A. 1974. The function of displays in thebreeding of the white-tailed black cockatoo. Emu74:43–46.
Saunders, D.A. 1982. The breeding behavior of theshort-billed form of the white-tailed black cockatoo(Calyptorhynchus funereus). Ibis 124:422–455.
Saunders, D.A., I. Rowley, and G.T. Smith. 1985. “Theeffects of clearing for agriculture on the distributionof cockatoos in the southwest of Western Australia.”In Birds of the eucalypt forests and woodlands:Ecology, conservations, management, ed. A. Keast,H. F. Recher, H. Ford, and D. Saunders, pp. 309–321.Sidney Australia: Royal Australasian OrnithologistsUnion and Surrey Beatty and Sons.
Saunders, D.A., G.T. Smith, and Campbell, N. A. 1984.The relationship between body weight, egg weight,incubation period, nestling period and nest site in thePsittaciformes, Falconiformes, Strigiformes andColumbiformes. Australian Journal of Zoology32:57–65.
Saunders, D.A., G.T. Smith, and I. Rowley. 1982. Theavailability and dimensions of tree hollows that pro-vide nest sites for cockatoo (Psittaciformes) inWestern Australia. Australian Wildlife Research9:541–546.
Seibert, L.M., and S.L. Crowell-Davis. 2001. Gendereffects on aggression, dominance rank, and affilia-tive behaviors in a flock of captive adult cockatiels(Nymphicus hollandicus). Applied Animal Be-haviour Science 71:155–170.
Serpell, J.A. 1981. Duets, greetings, and triumph cere-monies: Analogous displays in the parrot genusTrichoglossus. Zeitschrift fur Tierpsychologie55:268–283.
Skeate, S.T. 1984. Courtship and reproductive behaviorof captive white-fronted amazon parrots Amazonaalbifrons. Bird Behaviour 5:103–109.
Smith, G.T., and D.A. Saunders. 1986. Clutch size andproductivity in three sympatric species of cockatoo(Psittaciformes) in the south-west of WesternAustralia. Australian Wildlife Research 13:275–285.
Snyder, N.F.R., J.W. Wiley, and C.B. Kepler. 1987. Theparrots of Luquillo: Natural history and conserva-tion of the Puerto Rican parrot. Los Angeles:Western Foundation of Vertebrate Zoology.
Spoon, T.R. 2002. Reproductive success, parenting,and fidelity in a socially monogamous parrot (cock-atiels, Nymphicus hollandicus): The influence of so-cial relationships between mates. PhD diss.,University of California, Davis.
Spoon, T.R., J.R. Millam, and D.H. Owings. 2004.Variation in the stability of cockatiel (Nymphicushollandicus) pair relationships: The roles of males,
females, and mate compatibility. Behaviour 141:1211–1234.
Spoon, T.R., J.R. Millam, and D.H. Owings. In press.The importance of mate behavioural compatibilityin parenting and reproductive success by cockatiels(Nymphicus hollandicus). Animal Behaviour.
Stamm, R.A. 1962. Aspekte des Paarverhaltens vonAgapornis personata (Aves, Psittacidae, Loriini).Behaviour 19:1–56.
Stamps, J., A. Clark, P. Arrowood, and B. Kus. 1985.Parent offspring conflict in budgerigars. Behaviour94:1–40.
Striedter, G.F., L. Freibott, A.G. Hile, and N.T. Burley.2003. For whom the male calls: An effect of audi-ence on contact call rate and repertoire in budgeri-gars, Melopsittacus undulatus. Animal Behaviour65:875–882.
Sutherland, W.J. 2002. Science, sex and the kakapo.Nature 419:265–266.
Taylor, S., and M.R. Perrin. 2004. Intraspecific associ-ations of individual brown-headed parrots (Poiceph-alus cryptoxanthus). African Zoology 39:263–271.
Trillmich, F. 1976. Spatial proximity and mate specificbehaviour in a flock of budgerigars (Melopsittacusundulatus, Aves, Psittacidae). Zeitschrift furTierpsychologie 41:307–331.
Waltman, J.R., and S.R. Beissinger. 1992. Breeding be-havior of the green-rumped parrotlet. Wilson Bulle-tin 104:65–84.
Wanker, R., J. Apein, B. Jennerjahn, and B. Waibel.1998. Discrimination of different social companionsin spectacled parrotlets (Forpus conspicillatus):Evidence for individual vocal recognition. Be-havioral Ecology and Sociobiology 43:197–202.
Wanker, R., B.L. Cruz, and D. Franck. 1996. So-cialization of spectacled parrotlets Forpus conspicil-latus: The role of parents, crèches and sibling groupsin nature. Journal of Ornithology 137:447–461.
Wilkinson, R., and T.R. Birkhead. 1995. Copulation be-haviour in the Vasa parrots Coracopsis vasa and C.nigra. Ibis 137:117–119.
Wright, T.F., and M. Dorin. 2001. Pair duets in theyellow-naped amazon (Psittaciformes: Amazona au-ropalliata): Responses to playbacks of different di-alects. Ethology 107:111–124.
Yamamato, J.T., K.M. Shields, J.R. Millam, T.E.Roudybush, and C.R. Grau. 1989. Reproductive ac-tivity of force-paired cockatiels (Nymphicus hol-landicus). Auk 106:86–93.
Zann, R. 1965. Behavioural studies of the quarrion(Nymphicus hollandicus). B.S. thesis, University ofNew England, Armidale, New South Wales.
8 / Parrot Reproductive Behavior, or Who Associates, Who Mates, and Who Cares? 77

SUMMARY
Traditionally, cavity-type nest boxes are preferredover all other options for captive breeding of mostpsittacine species. These species indeed havegreater reproductive success when they are givencavity-type boxes. The basis for this informationis largely field studies. The idea of the cavity-typenest boxes is an extrapolation from the wild,where the majority of psittacine birds nest in liv-ing trees. However, one scientific study, con-ducted by the first author, exists in the literatureand corroborates this choice. This study alsogives alternatives to improve reproductive successfor the naive (first-time layers), the recently non-breeding, and the floor-laying psittacine birds, in-stead of re-pairing or even culling these poten-tially reproductively successful birds. However,other studies have shown that the efficacy of nestboxes may depend upon early experience withthem since Cockatiel chicks parent-reared with-out nest boxes, or hand-reared by humans, do notreadily use cavity-type nest boxes as adults. Thisproblem will certainly be exacerbated with in-creasing numbers of hand-reared psittacine birdsgoing into captive breeding situations.
REVIEW OF THE NEST BOX SELECTIONSTUDY
The demand for captively bred parrots for the petbird market is ever increasing due to the tradebans imposed in response to the threatened or en-dangered status of the majority of psittacinespecies, and due to the increasing popularity ofbirds as pets.[3] Birds are now the third mostcommon pet in the United States and their popu-larity is growing. Further, since nearly all psitta-cine birds are cavity nesters in the wild, it is espe-cially important to understand problems relating
to nest box use in captivity to increase reproduc-tive efficiency.[3].
For many temperate-zone birds, variation in thephotoperiod is known to provide the most impor-tant initial cue for control of the reproductivecycle.[3, 4, 5] However, photoperiod is often sup-plemented by other proximate factors to initiatereproduction.[3] This is especially true for cavity-nesting parrots, for which provision of a nest boxis perhaps the most important environmentalstimulus available to induce reproductive devel-opment. Further, it has been shown that maxi-mum levels of serum-luteinizing hormone infemale Cockatiels is only achieved when a com-bination of photostimulation, nest box access, andfull mate access is available.[3–5] Other studieshave shown that the efficacy of nest boxes maydepend upon early experience with them sinceCockatiel chicks reared without nest boxes didnot readily use them as adults.[4] Further,Cockatiel chicks reared in nest boxes by parentsare far more likely to use nest boxes as adults,when compared to chicks hand-reared by humansand lacking early exposure to nest boxes.[3, 4]With the increase in domestically hand-rearedbirds that subsequently go on to be breeders, ini-tiating successful reproduction and use of thecavity-type nest boxes are likely to become pro-gressively problematic.
Characteristics of nest boxes such as size,shape, construction material, location, and inte-rior light intensity all influence nest box selectionin different species of psittacine birds (Figure 9.1and Table 9-1).[1–3, 6, 7] In communal groups,Cockatiels prefer higher nest boxes, and thosewith smaller nest entrances, over lower nest boxesand those with larger entrances.[3] Preferentialnest-site selection involving a variety of nest-site
79
9Nest Box Preferences
Scott George Martin and April Romagnano

characteristics has been observed in severalspecies.[1–3, 6, 7] However, occasionally individ-ual psittacine birds may choose nesting sites thatresemble neither natural nor artificially providedones. Floor laying is an example of such an aber-rant nest-site selection behavior. This behaviordecreases hatchability through either egg surfaceor interior contamination secondary to egg soilingor minor cracks or breakage. In captive parrotsfloor laying is most common in the smallerspecies. This behavior is costly and consideredmaladaptive.
The etiology of floor laying is unknown, butthis behavior can affect up to 25% of Cockatielpairs in captive breeding colonies, despite thepresence of conventional cavity-type nest boxesand proper long day photoperiod stimulation.[3]It is possible that such birds may have floor laideven if the two previously mentioned stimuli, cav-ity-type nest boxes and photostimulation, wereabsent. Since the etiology of this aberrant layingbehavior has not been elucidated, and since it canbe devastating from both a financial and a conser-vational standpoint, the first author conducted astudy to address the nature of nest box selectionin captivity. This study was composed of two ex-periments in which nest boxes of two differentnest entrance types were offered as choices toCockatiels with or without a history of floor lay-ing, as well as to birds that had not laid in a pre-vious breeding trial, and finally to reproductivelynaive birds. The nest box entrance types chosen
80 Manual of Parrot Behavior
Figure 9.1. Some basic nest box types. Upperleft is a vertical box (VB), as is the one below;these VBs come in various sizes depending onthe species. The upper right-hand box is a hori-zontal macaw (HM) and is always 16� x 16� x48�. The lower right-hand box is known as an L-shaped or boot box. WF stands for wire frontand WL stands for wire ladder.
Table 9-1 Nest box types for different species
Species Nest box type
Budgerigars, Cockatiels, Quaker Parrots, Lovebirds, Will happily breed in VB boxes of various sizesParrotlets, Grey- and Green-cheeked Parakeets, and numerous other smaller psittacine birds, Eclectus, Cockatoos of various sizes (except for Palm Cockatoos, who prefer a vertical open-top box 2� x 2� x 6� [VO]), smaller Macaws, Conures, Amazons, Pionus, Caiques, hawk heads, thick-billed parrots and the lesser vasas
Large and medium-sized macaws and Greater Will happily breed in HM boxes of various sizesVasas
African Greys Will happily breed in L-shaped or boot boxes that give a lot of privacy to the female and heryoung

were a small hole for the classic cavity-type nestboxes and a shelf-type entrance as the alternative.
NEST BOX SELECTION
In the nest box selection study, the nest box en-trance preferences of Cockatiels were examinedand recorded.[3] Some of the birds were histori-cally floor layers, some were cavity-type layers,some had not recently laid, and the rest were re-productively naive (first-time layers).
The nest box entrances of the cavity-type nestboxes were circular and relatively small, while theentrances of shelf-type boxes were rectangularand relatively large (Figure 9.2).
The study found that birds with histories of lay-ing in cavity-type boxes tended to choose cavity-type boxes again and repeated this preference,while those birds that were previously floor layersor had not recently laid tended to choose shelf-type boxes for their first clutches but laterswitched to cavity-type boxes for subsequentclutches. Hence, with reproductive experience af-forded by the shelf-type nest box, these birdslater, when they were more experienced layers,preferred the conventional cavity-type nest box.Reproductively naive Cockatiels were divided intheir nest box preferences. Approximately one-third of the reproductively naive Cockatiels choseshelf-type boxes for their first clutches but in-creasingly used cavity-type boxes for subsequentclutches.
Further, it was noted that Cockatiels that laid incavity-type boxes never switched to shelf-typeboxes. Fertility, hatchability, and fledging suc-cess, defined as survival to three weeks of age,were all greater in cavity-type boxes, resulting ingreater chick production.
The question that was asked, and that wasclearly answered by this study, was whether thereis a nest box selection preference among Cock-atiels.[3] Cavity-type nest box selection was pre-ferred by Cockatiels that had historically bred andused cavity nests. Further, naive, non-laying, andfloor-laying Cockatiels also all eventually movedtoward cavity-type nest boxes as their preferrednesting sites. The presence of the shelf-type nestbox, however, initiated successful laying in theseproblem birds.
It has previously been noted that given the rightmaterials, most all psittacine birds seek out orbuild cavity-type nests. This clear and universal
pattern leads one to believe that cavity-type nest-ing must be the closest to natural parrot nesting inwild populations.
WILD NESTS
According to Thompson, parrots in the wild nestin cavities that are primarily found in living trees,or they nest in cavities within dead trees.[6] It isclear that parrots prefer to lay their eggs in spacesthat are cavity- or hole-like, so as to afford theeggs more protection from both predators and theenvironment.[6] New world birds use abandonedwoodpecker nests, which are great for many of thesmaller parrots. Larger parrots have a harder timefinding appropriate-sized cavity nests that areabandoned.[6] Nesting cavities must accommo-date the nesting female, her mate, and their poten-tial young. Most psittacine young will grow to thesize of their parents by weaning/fledging.[6]
Thompson also notes that a subgroup com-posed of various parrot and parrot-like specieschooses nesting sites in cavities other thantrees.[6] Some of these sites include tunnels in
9 / Nest Box Preferences 81
Figure 9.2. The shelf-type and the cavity-typenest box setup, used in the Cockatiel nest boxselection study. They were installed side by sideas depicted.

cliff sides, active and inactive termite mounds,and chambers or hollows found within the rockson the ground. However, any parrot species thatnests on the ground is more vulnerable and thusvery endangered. Examples of ground-nestingpsittacine birds include the Bahama Parrot andthe Kakapo.[6]
CONCLUSION
The authors believe that given the results of thefirst author’s nest box selection study and explor-ing the general nesting habits of different speciesof parrots in the wild, one can, and should, at-tempt to give each psittacine species a nest boxchoice, if the classic captive cavity-type nestboxes are unacceptable, that is, if the birds do notbreed. Of course, this is assuming that the psitta-cine pair in question is a true pair, consisting ofboth a female and a male of the same species, andthat they are sufficiently healthy for breeding.Hence, one should always attempt to match thenatural nesting needs of captive psittacine birdsfirst, and if this does not work, employ the shelf-type nest box choice to reintroduce or introducethe cavity-type nest box nesters, the naive, thenon-layers, and the floor-laying birds to a suitablenest before thinking of re-pairing or culling birds.It is likely that most psittacine birds would bene-fit from a shelf-type nesting opportunity and thenif also given the choice, as in the study, wouldlikely eventually move toward cavity-type nestboxes as their preferred nesting site.
Since the etiology of rejecting the cavity-typenest box is unknown in captive, otherwise healthytrue psittacine pairs, one would have to assumethat experience plays a significant and growingrole. It is well known that the fostering of naturalbehaviors, such as parental rearing in a cavity nestbox if just for a brief period of time, is an ap-proach that has been demonstrated to improve re-productive success and the general well-being ofbreeding psittacine birds.[7] Styles states thatbirds properly socialized to their own species areindeed superior in their reproductive abilities,while improperly socialized birds are much lesslikely to become successful breeders.[7]
The findings discussed in this chapter agreewith other Cockatiel studies where the efficacy ofcavity-type nest boxes appeared to depend uponearly experience with them, in that parent-raisedCockatiel chicks reared without nest boxes andCockatiel chicks hand-reared by humans and thusalso lacking early exposure to nest boxes do notreadily use them as adults. Hence, with the in-crease in domestically hand-reared birds that willsubsequently go on to become domestic breeders,the likelihood of problem layers can be expectedto increase.
REFERENCES
1. Clubb, S.L. 1997. “Avicultural medicine and flockhealth management.” In Avicultural medicine andsurgery, ed. R.B. Altman, S.L. Clubb, G.M. Dorre-stein, and K. Quesenberry, pp. 101–116. Philadel-phia: WB Saunders Co.
2. Johnson, T., and K. Clubb. 1992. “Aviary design andconstruction.” In Psittacine aviculture, perspec-tives, techniques and research, ed. R.M. Schubot,K.J. Clubb, and S.L. Clubb S.L., pp. 4.1–4.12.Loxahatchee, FL: Avicultural Breeding and Res-earch Center..
3. Martin, S.G., and J.R. Millam. 1995. Nest box se-lection by floor laying and reproductively naive cap-tive cockatiels (Nymphicus hollandicus). Appl AnimBehav Sci 43:95–109.
4. Meyers, S.A., J.A. Millam, T.E. Roudybush, andC.R. Grau. 1988. Reproductive success of hand-reared vs parent reared cockatiels (Nymphicus hol-landicus). Auk 105:536–542.
5. Millam, J.R., B. Zhang, and M.E. el Halawani.1996. Egg production of cockatiels (Nymphicushollandicus) is influenced by number of eggs in nestafter incubation begins. General and ComparativeEndocrinology 101 (2):205–210.
6. Thompson, D.R. 1998. “The importance of nest-boxes.” International Aviculturists Society Con-vention.
7. Styles D. 2001. “Captive psittacine behavioral re-productive husbandry and management: Sociali-zation, aggression control, and pairing techniques.”Proceedings of the Association of Avian Veterina-rians (Specialty Advanced Program), pp. 3–14.
82 Manual of Parrot Behavior

INTRODUCTION
Since the Wild Bird Conservation Act of 1992banned the importation of most wild-capturedparrots into the United States, hand-rearing of do-mestically bred parrot chicks has become thedominant method of producing parrots for the pettrade. That hand-reared parrots are popular aspets should not be surprising: young hand-rearedparrots are tame, apparently enjoy contact withhumans, and seem much better adapted to life asa pet than do most parent-reared parrots (Millam2000). Hand-reared parrots may be more likely totalk than parent-reared birds (see the “SocialBehavior” section of this chapter). However,hand-rearing is time-intensive and technically de-manding, and both the scientific and lay literaturesuggest that hand-reared birds may exhibit a num-ber of behavioral abnormalities.
Hand-rearing requires that parrot chicks beseparated from their parents for long periods oftime (several weeks to several months, dependingon the species). It has long been appreciated thateven short-term separations are stressful (cf. Vaz-quez 1997 for examples in rats), and that disrup-tions in parental care can disrupt normal behav-ioral and physiological development (cf. Levine2001; Capitanio 1986). In rat pups, maternal be-havior during the first two to three weeks of lifeis critical to both the regulation of the pups’ phys-iological state and to normal brain development(Meaney 2001, Levine 2001, Vazquez 1997).Even brief periods of maternal separation (greaterthan three hours) during the first two to three
weeks of life are sufficient to produce long-lasting changes in responsiveness to stress and re-activity to novelty in rats and other mammals in-cluding mice and guinea pigs (Meaney 2001;Levine 2001; Albers et al. 2000; Vazquez 1997).These changes are primarily mediated by changesin the hypothalamic-pituitary-adrenal (HPA) axisinduced by maternal separation (Francis et al.2002; Meaney 2001; Levine 2001; Albers et al.2000; Vazquez 1997; Biagini et al. 1998).Maternally-separated animals also experiencedelays in tissue growth and altered circadianrhythms, effects that are probably also mediatedby increased HPA axis responsivity to stressors(high levels of glucocorticoids, which are pro-duced when the HPA axis is activated by stress,have catabolic effects) (Biagini et al. 1998). Fur-thermore, maternal separation affects not only theseparated animals themselves but their offspringas well. Meaney (2001) demonstrated that mater-nal behavior influences how rat pups will behavetoward their own offspring.
Although maternal separation effects have notbeen directly demonstrated in parrots, some evi-dence suggests that these effects do occur. Astudy of growth rate differences in hand-rearedand parent-reared chicks at Loro Parque suggeststhat parrots may respond to maternal separationwith slower growth rates. In at least two psittacinespecies (Ara rubrogenys, Pyhurra p. perlata),hand-reared chicks show a growth-rate deficitsimilar to that of maternally-separated rat pups(Navarro & Castanon 2001). However, this deficit
83
10Hand-Rearing: Behavioral
Impacts and Implications forCaptive Parrot Welfare
Rebecca Fox

may be alleviated by leaving the chicks with theparents during the first week of life and beginninghand-rearing thereafter, suggesting some aspectof parental care during the first two weeks of lifemay be responsible for maintaining normalgrowth rates in parrots (Navarro & Castanon2001).
Hand-reared parrots typically experience farlonger periods of maternal separation than thosethat cause striking behavioral changes in mam-mals such as rats (separations of weeks to monthsfor hand-reared parrots versus separations ofhours in maternally-separated mammals in manyexperimental paradigms). Nursery-reared mon-keys, which experience a comparable level of ma-ternal separation to hand-reared parrots, fre-quently develop abnormal behaviors includingself-mutilation (possibly analogous to featherpicking in companion birds), increased responsiv-ity to stress, and altered social behavior (Capi-tanio 1985).
Furthermore, hand-rearing deprives parrots ofparental contact that may be required to establishnormal social and sexual preferences. A numberof studies (cf. Immelmann 1972, 1975 for re-views) have shown that the presence of parents isoften a critical component of the development ofnormal species identity and sexual imprinting.Sexual imprinting refers to the process by whichanimals learn the characteristics of appropriatemates by learning the characteristics of their par-ents or siblings. Among parrots, Galahs (Cacatuaroseicapilla) naturally cross-fostered to the sym-patric Leadbeater’s Cockatoo (C. leadbeateri)apparently imprint on their foster parents and as-sociate solely with C. leadbeateri individualsduring adulthood (Rowley & Chapman 1986).Cross-fostered Galahs also choose to mate withC. leadbeateri, suggesting that sexual imprintingdoes occur in these species and may be disruptedby changing the neonatal environment (Rowley &Chapman 1986). Anecdotal evidence suggeststhat sexual imprinting may occur in a number ofparrot species, including Budgerigars (Melopsit-tacus undulatus) and Senegal Parrots (Poicepha-lus senegalus senegalus) (Klinghammer 1967).Thus, hand-rearing may also have negative repro-ductive consequences.
In this chapter, I will discuss two studies of thereproductive and behavioral consequences ofhand-rearing in Cockatiels (Nymphicus hollandi-
cus) and Orange-winged Amazon Parrots (Ama-zona amazonica). The first study, conducted byMyers et al. (1988), examined differences in nest-ing behavior and reproductive success in hand-reared and parent-reared Cockatiels. The secondstudy is part of my own research, and examineddifferences in social preferences, vocal behavior,and neophobia (fear of novel objects) in 3-month-old to 12-month-old hand-reared and parent-reared parrots. I will also discuss the possiblewelfare consequences of hand-rearing for captiveparrots.
BEHAVIORAL DIFFERENCES IN HAND-REARED VERSUS PARENT-REARED PARROTS
Reproductive Behavior
Rowley and Chapman (1986) suggest thatparental identity strongly influences the learningof species identity and sexual imprinting in par-rots. Sexual imprinting refers to a process bywhich exposure to certain characteristics early inlife increases an animal’s preferences for thosecharacteristics in a mate. Furthermore, habitatimprinting may be important for parrots to learnthe characteristics of appropriate nest sites.Habitat imprinting refers to a process in whichexposure to a particular habitat feature early indevelopment increases preference for that featurelater in life. Hand-reared parrots, which are gen-erally reared in brooders and fed by humans, aredeprived of the opportunity to learn the character-istics of appropriate nest sites and mates and canshow inappropriate reproductive behavior (Myerset al. 1988). In Cockatiels (Nymphicus hollandi-cus), hand-rearing decreases reproductive successby altering both nesting behavior and normal sex-ual behavior (see Table 10-1), probably becausematernal separation disrupts normal sexual andhabitat imprinting (Myers et al. 1988).
Maternal separation (i.e., hand-rearing) dis-rupts habitat imprinting in both male and femaleCockatiels. Some effects of hand-rearing on nest-ing behavior were sex-specific: hand-reared (H)males were less likely to inspect nest boxes thanparent-reared (P) males, but the female’s rearingcondition had no effect on nest inspection (Myerset al. 1988). However, both the male’s and the fe-male’s rearing condition influences where a pairchooses to lay their eggs. The mates of H-males
84 Manual of Parrot Behavior

were about three times more likely to lay eggs onthe cage floor rather than in the nest box, regard-less of rearing condition. Early experience in fe-males also influences where they choose to laytheir eggs—floor laying was more common inpairs containing H-females than in pairs contain-ing P-females (Myers et al. 1988).
Hand-rearing apparently influences sexual im-printing in males more strongly than in females.H-male pairs have significantly lower egg fertilityand reproductive success when compared to P-male pairs, presumably because males exhibit ab-normal courtship behavior or copulation (Myerset al. 1988). However, normal sexual imprintingis apparently less critical for female receptivity, asH-females did not exhibit significantly impairedreproductive success relative to P-females (Myerset al. 1988). Because the Cockatiels used in thisstudy were force-paired, hand-rearing may haveeffects on mate choice in females that were notdetected (Myers et al 1988).
Hand-rearing/maternal separation causes sex-specific disruptions in normal sexual behavior inparrots. In males, nest box recognition and use areimpaired, and normal copulatory behavior isprobably impaired as well (Myers et al. 1988).Certainly, hand-rearing has a significant negativeeffect on a male’s reproductive success. The be-havioral disruptions in females seem to be less se-rious. Although hand-reared females exhibithigher levels of floor laying, the reproductive suc-cess of pairs containing H-females is not im-paired relative to P-female pairs.
Social Behavior
SOCIAL PREFERENCES
Abnormal sexual behavior may be broadly re-lated to disruptions in normal social preferencesinduced by hand-rearing. My recent work, which
compared the behavior of hand-reared and par-ent-reared Orange-winged Amazon Parrots(Amazona amazonica) suggests that hand-rearingalters social preferences and normal vocaliza-tions in juvenile (3–12 months of age) Orange-winged Amazons.
Hand-reared parrots exhibit a strong preferencefor social contact with humans and reduced pref-erences for contact with conspecifics. When ahand-reared Amazon is placed in a choice cagewith a human handler to one side and two parrotsin their home cage to the other, the hand-rearedbird will perch on the side of the cage nearest thehandler significantly more frequently than pre-dicted by chance. Conversely, non-tame parent-reared birds perch on the side of the cage nearestconspecifics significantly more often than pre-dicted by chance. Parent-reared birds that havebeen tamed by occasional neonatal handling (e.g.,Aengus & Millam 1999) exhibit approximatelyequal preference for human and conspecific com-panionship, suggesting that while tameness influ-ences social preferences in parrots, hand-rearingdisrupts social preferences to a much greater ex-tent (see Figure 10.1).
VOCAL BEHAVIOR
Parent-reared and hand-reared birds also differedconsiderably in the extent and the speed withwhich they acquired human vocalizations, sug-gesting that hand-rearing may also disrupt normalvocal development in parrots. Three of sixOrange-winged Amazons hand-reared in 2001began mimicking human vocalizations while stillin the nursery (i.e., before three months of age),and the remaining three birds began mimickinghuman speech in the presence of experimentersby four months of age. Of the parent-reared birds,only two of the five birds that had been tamed by
10 / Hand-Rearing: Behavioral Impacts and Implications for Captive Parrot Welfare 85
Table 10-1 Comparison of reproductive behavior and success in hand-reared (H) andparent-reared (P) cockatiels (modified from Myers et al. 1988)
Nest inspection Floor laying by male by female Egg fertility Overall RS*
H-male Low High Lower Lower P-male High Low Higher HigherH-female No difference High No difference No differenceP-female No difference Low No difference No difference
*RS: Reproductive success, defined by the number of offspring surviving to fledging.

neonatal handling ever mimicked human speechin the presence of humans, and they did not do sountil six to nine months of age. The two parent-reared birds that did develop human vocalizationswere also qualitatively the tamest of the parent-reared group. Hand-reared birds also developedfar more extensive repertoires than did parent-reared birds (see Table 10-2).
The high levels of human mimicry exhibited by the hand-reared birds are probably related toseveral factors, of which extensive exposure tohuman speech and the perception of human con-tact as rewarding are probably the most pertinent.Early removal from the nest also seems to preventthe learning of species-typical vocalizations(hand-reared birds do not develop the loud high-pitched call typical of Orange-winged Amazonsuntil they have been housed with normally vocal-izing birds for at least a week), and human soundsmay thus occupy a larger-than-normal part ofhand-reared birds’ repertoires.
Research by Pepperberg et al. (1998) and Westand King (1990) suggests that the learning ofhuman vocalizations by mimetic species such asparrots and starlings is heavily dependent on so-cial interaction with humans. Hand-reared birds,which are fed as many as five times/day or more
during the first few months of life, have muchmore extensive contact with humans than doparent-reared birds: even those birds that have beenreared under neonatal handling paradigms are only handled five times/week for approximately20 minutes/session. Because the hand-rearedbirds have more extensive contact with humans,they also have more opportunities to learn humanvocalizations. It also seems reasonable that thosebirds that find human attention especially reward-ing will be more likely to use human vocaliza-tions in the presence of people (e.g., Pepperberg1999). The hand-reared Orange-winged Amazonsin this study appeared to use human vocalizationsto elicit human attention. They typically vocalizedwhen an experimenter or handler entered theroom, and they often used human vocalizationsduring handling sessions. These vocalizationsgenerally elicited increased attention, or at least avocal response, from the human handlers. Theparrots’ human vocalizations were typically ac-
86 Manual of Parrot Behavior
Figure 10.1. Numbers of instantaneous sam-ples (mean ± S.E.) in which hand-reared (H),parent-reared with human handling (PH), andparent-reared (P) birds were observed perchingnear either a familiar handler or near a cagecontaining two conspecifics during a preferencetest. Each subject’s location in the test cage wasrecorded every 30 seconds for 15 minutes.
Table 10-2 Human vocalizations used by3- to 12-month-old hand-reared and par-ent-reared Orange-winged Amazons
Bird Vocalizations
Valentine “step up,” “hello,” “hi,” “good bird,” “squeak,” “hi, Val,”kissing noises, various whistles
Squeeker “step up,” “hello,” “good bird,” “good boy,” “good Squeeker,”“squeak,” “hi, Val,” “Squeeker,”kissing noises, various whistles
Julian “step up,” “hello,” “hi,” “good boy,” “good girl,” various whis-tles, kissing noises, "brrrring!"
Andy “step up,” “hello,” “hi,” “good,” “good boy,” “good girl,” “goodboygirl,” “good bird,” “I loveyou,” “Andy,” “what doing,”various whistles, kissing noises,“brrrring!”
Kelly “step up,” “hi,” “good girl,” “good bird,” kissing noises, variouswhistles, “brring!”
Taylor “step up,” “hi,” “good boy”Mackenzie* “Step up,” “hi,” “brrrr”Robin* “Step up,” “hi,” kissing noise
*Parent-reared bird.

companied by excited displays such as rapid pupildilation and contraction (“pinpointing”), headand neck feather erection, and tail fanning. Parrot“speech” is likely related to extensive exposure tohuman speech and reinforced by operant condi-tioning in which human attention serves as the re-ward. Hand-reared birds are, in general, morelikely to talk and less likely to exhibit normal vo-calizations than are parent-reared birds, eventhose that have been tamed by neonatal handling.
Neophobia
Maternal separation and hand-rearing affect notonly social preferences and learned vocalizationsbut may also change the developmental trajectoryfor certain behaviors. Neophobia refers to theavoidance of novel objects and/or foods by ani-mals and is related to animals’ reactivity to nov-elty (cf. Meaney 2001). Less neophobic animalswill approach a novel object more readily thanmore neophobic animals and will exhibit higherlevels of exploratory behavior in a novel environ-ment. Neophobia probably represents an adaptiveresponse for avoiding potentially poisonous fooditems and predators, and, at least in ravens, seemsto develop after juveniles are weaned and theirfood preferences have solidified (Heinrich 1998).
In order to distinguish neophobia from a lackof interest in novel items, I tested hand-reared andparent-reared birds by presenting them with adish of peanuts (a highly favored food) overwhich a novel object had been hung. Novel ob-jects included a green plastic mug, a blue toydump truck, a large black cooking spoon, and atoy elephant. The birds’ latency to approach thedish/novel object combination was measured andused as an index of neophobia. More neophobicbirds would presumably take longer to approachthe dish /novel object combination than wouldless neophobic birds.
After a two-week habituation period in whichthe birds were habituated to the dish of peanutsalone, five consecutive tests in which the birdswere presented with the dish and a novel objectwere administered approximately ten days apartwhen the chicks were approximately 135–180days old. Striking differences in neophobia wereapparent in parent-reared and hand-reared birdsthroughout most of the testing period (Figure10.2). Parent-reared birds were already somewhatneophobic by 135 days and showed latencies of a
several minutes to approach the dish + objectcombination. Conversely, hand-reared birds ap-proached the dish + object combination within afew seconds. Neophobia (latency to approach thedish + novel object combination) peaked at 160days in parent-reared birds and then began to de-cline, but neophobia remained low in hand-rearedbirds until 180 days, when hand-reared birdsabruptly began reacting fearfully to novel objects.However, the behavior of hand-reared birds at oneyear of age suggests that hand-rearing only delaysthe onset of neophobic behavior in hand-rearedbirds but does not permanently alter the level ofneophobia; 12-month-old hand-reared and parent-reared birds behave very similarly in response tonovelty. When presented with a novel toy, 12-month-old hand-reared and parent-reared birdsdisplayed comparable levels of fearful behaviorwhen their behavior was scored on a scale of 0(approach toy) to 5 (frantic attempts to escape)(Figure 10.3).
The differences in neophobia may be due eitherto delayed maturation in the hand-reared birds orto a more generalized habituation to novelty.Research in rats has shown that maternal separa-
10 / Hand-Rearing: Behavioral Impacts and Implications for Captive Parrot Welfare 87
Figure 10.2. Latencies (mean ± S.E.) toapproach and feed from a dish of favored foodin the presence of a novel object for parent-reared and hand-reared birds in a series of fivetests administered between the ages of 135 and180 days. Data were log-transformed to homog-enize variance. Repeated-measures ANOVAshowed that latencies differed significantlybetween parent-reared and hand-reared birds(F1,17 = 9.25, P = 0.007).

tion early in life acts to disrupt or delay the matu-ration of certain regions of the amygdala that re-late to stress reactivity and responsivity to novelty(Vazquez 1997, Levine 2001). In birds, the medialarchistriatum controls fearfulness and reactivity tonovelty, and lesions of the medial archistriatumgreatly reduce fearfulness in wild birds (Butler &Hodos 1996). Similar effects are seen in rhesusmacaques with neonatal lesions to the central nu-cleus of the amygdala, who exhibit very low levelsof behavioral inhibition in response to stimuli thatinduce fearful behavior in normal macaques(Prather et al. 2001). It is possible that maternalseparation affects the maturation of the medialarchistriatum in parrots, leading to greatly reducedlevels of neophobia in parrots. However, hand-
reared birds also generally experience higher lev-els of novelty (different caregivers, more extremechanges in light level, exposure to a greater vari-ety of objects, etc.) during the nestling period thando parent-reared birds, which remain in a darknest box until fledging. The reduced neophobiadisplayed by young hand-reared birds couldsimply reflect an adaptation to novelty.
After weaning, the parent-reared and hand-reared birds were housed under identical condi-tions, so the disappearance of behavioral differ-ences between the two groups may be related to aphenomenon described by Francis et al. (2002), inwhich environmental conditions (in this case, en-richment) can reverse the effects of maternal sep-aration in prepubertal rats. Alternatively, althoughthe birds were provided with toys, the toys werechanged relatively infrequently (every fewmonths), and the level of novelty experienced bythe hand-reared birds may not have been suffi-cient to maintain low levels of neophobia thatthey exhibited at the beginning of the study.
Summary
From a behavioral perspective, hand-rearing reli-ably produces tame, human-habituated birds thattend to talk more than do parent-reared birds. Forthis reason, young hand-reared birds are quiteappealing as companion animals. However, hand-rearing is also a potent disrupter of normal behav-ioral development in Cockatiels and Orange-winged Amazons (see Table 10-3). Hand-rearingcan cause abnormalities in sexual behavior andnesting, especially in male Cockatiels. These ab-normalities are probably related to abnormal sex-ual and habitat imprinting (Myers et al. 1988).Anecdotal evidence suggests that hand-rearedparrots frequently imprint sexually on humansand may even prefer to court their owners overconspecifics. Certainly, hand-rearing alters socialpreferences to a much greater degree than does
88 Manual of Parrot Behavior
Figure 10.3. Neophobia scores (mean ± S.E.)for parent-reared (P), parent-reared with humanhandling (PH), and hand-reared (H) birds whenpresented with a novel object at 12 months of age. A score of 0 represents immediate ap-proach to the toy, while a score of 5 representsfrantic attempts to avoid the toy. Neophobiascores did not differ significantly between thethree rearing conditions.
Table 10-3 Summary of behavioral differences between hand-reared and parent-rearedCockatiels and Orange-winged Amazon parrots
Nest Social Humanrecognition preference vocalizations Onset of neophobia
Hand-reared Poor Humans Common 6 months of ageParent-reared Normal Con-specifics Rare On or before 4.5 months
of age

taming by neonatal handling, with hand-rearedbirds showing a much stronger preference forhuman companionship than neonatally-handledbirds. Hand-reared Orange-winged Amazons alsoshow developmental differences in neophobia,with hand-reared birds developing a fear of novelobjects much later than parent-reared birds.However, when parent-reared and hand-rearedbirds are housed under identical conditions, dif-ferences in neophobic behavior disappear by thetime the birds are 12 months of age.
IMPLICATIONS FOR PARROTS INCAPTIVITY
Breeding Programs
Because of restrictions on the importation of wildparrots imposed by the Wild Bird ConservationAct, the success of breeding programs in theUnited States (both for conservation purposes andfor the purpose of producing parrots for the pettrade) rests on the continued availability of repro-ductively competent domestically raised parrots.Because hand-reared Cockatiels show a numberof abnormalities in reproductive behavior and thereproductive success of hand-reared Cockatielspaired with hand-reared Cockatiels is extremelylow (Myers et al. 1988), it is likely shortsighted toemphasize the production of hand-reared birdsover more reproductively normal parent-rearedbirds. The loss of wild-captured breeding stock toage or disease could be disastrous, especially forthose species that are rare or difficult to breed incaptivity, unless healthy, reproductively normalparent-reared birds are available to replace them.
However, Myers et al. (1988) also showed thatthe reproductive success of hand-reared birds (atleast Cockatiels) can be improved by pairing themwith a parent-reared, reproductively normal part-ner. This is particularly true for female parrots(Myers et al. 1988). Careful pairing of hand-reared birds with more experienced parent-rearedmates may help to mitigate the effects of hand-rearing on reproductive behavior. Unfortunately,hand-reared birds may also be more difficult topair than parent-reared birds, since they exhibitgreatly reduced preference for conspecific com-panionship.
Companion Parrots
Young hand-reared parrots seem like the perfectpet: tame, talking parrots that have little fear of
new things and are quite willing to interact withhumans. However, hand-rearing may also be atthe root of a number of behavior problems incompanion parrots.
Hand-rearing alters Orange-winged Amazons’social preferences and sexual behavior and maylead to hand-reared parrots inappropriately di-recting sexual behavior (both courtship behaviorand aggression) toward humans. Anecdotally,sexual aggression directed at humans is commonin a number of parrot species, especially in maleamazon parrots and male cockatoos. Similarly,parrot owners often report that their parrots re-gurgitate for them or attempt to masturbateagainst their owners’ hands or bodies. Although astrong social preference for humans may be en-dearing in a parrot chick, sexual aggression andinappropriate sexual behavior can be problematicin older birds.
Hand-rearing may also be related to the “pho-bic” behavior that has been recently described inpet parrots. In the lay literature, phobic behavioris defined as a behavior pattern in which “a pre-viously tame and affectionate parrot ‘suddenly’seems afraid of almost everything and everyone”(Blanchard 2001). This sudden, apparently inex-plicable, fearfulness usually appears in hand-reared birds around the age at which they wouldtypically become independent of their parents inthe wild (Blanchard 2001). “Phobic” or anxiousparrots often injure themselves trying to avoidaversive stimuli, often breaking blood feathersand injuring their keels in repeated falls (Clark2001). Obviously, the sudden appearance of such extreme and apparently unexplainable fear-ful behavior in a previously fearless bird can bedisconcerting and would certainly appear patho-logical.
This pattern seems analogous to the pattern ob-served in the Orange-winged Amazons, in whichhand-reared birds exhibited markedly low levelsof neophobia for at least two months longer thandid parent-reared birds. Furthermore, the onset ofneophobia in the hand-reared birds was quite sud-den. In the week between the fourth and fifthnovel object tests, hand-reared birds’ latencies toapproach the novel object/dish combinationshowed a striking increase (Figure 10.2). Althoughthis sudden appearance of avoidance behavior mayseem startling, the fact that hand-reared andparent-reared birds react almost identically to a
10 / Hand-Rearing: Behavioral Impacts and Implications for Captive Parrot Welfare 89

novel toy at 12 months of age suggests that this“sudden” fearfulness may actually represent nor-mal, if delayed, maturation. Although “phobic”behavior may not actually be pathological, the layliterature (cf. Blanchard 2001; Clark 2001) sug-gests that pet owners find it extremely disturbing,and the increased chance of injury during this“phobic” period is a very real risk.
CONCLUSION
Hand-reared parrot chicks are endearing, oftentalkative, and fearless. However, this period ofoutgoing fearlessness is only temporary. By oneyear of age, hand-reared and parent-rearedOrange-winged Amazons behave identically inresponse to a novel object, suggesting that theperiod in which hand-reared birds appear partic-ularly well adapted to captivity may only be tem-porary. Furthermore, hand-reared Cockatielsshow serious deficits in reproductive behaviorthat are probably related to abnormal sexualand/or habitat imprinting (Myers et al. 1988).Abnormal sexual imprinting and a strong socialpreference for humans may cause behavior prob-lems in pet parrots, which are probably morelikely to inappropriately direct sexual behavior attheir owners. Hand-reared birds may exhibitother behavior problems as well, most notablyso-called “phobic” behavior. Hand-rearing is alsotime intensive (hand-reared parrots often must befed several times a day for several months), tech-nically demanding, and risky (aspiration of hand-rearing formula can kill or sicken chicks veryquickly).
However, research has shown that occasional(four to five times/week for 30 minutes) handlingof parent-reared Orange-winged Amazon chicksbetween two weeks of age and fledging also pro-duces tame chicks that are less responsive tostress than birds that have been parent-rearedwithout handling (cf. Aengus & Millam 1999).Because neonatally handled birds that remain inthe nest have much more contact with parents andsiblings than they do with humans, it is also likelythat they will show normal reproductive behaviorat maturity (Aengus & Millam 1999). Neonatalhandling also requires far less time, effort, andexpense (in terms of hand-rearing supplies andequipment) than does hand-rearing and may rep-resent a viable alternative for producing parrotsfor the pet trade.
REFERENCES
Aengus, W.L., and J.R. Millam. 1999. Taming parent-reared orange-winged amazon parrots by neonatalhandling. Zoo Biology 18: 177–187.
Albers, P.C.H., P.J.A. Timmermans, and J.M.H. Vossen.2000. Effects of frequency and length of separationbouts between mother and offspring on later explo-rative behaviour of young guinea-pigs (Caviaaperea f. porcellus). Behaviour 137:1487–1502.
Biagini, G., E.M. Pich, P. Carani, P. Marrama, and L.F.Agnati. 1998. Postnatal maternal separation duringthe stress hyporesponsive period enhances theadrenocortical response to novelty in adult rats byaffecting feedback regulation in the ca1 hippocam-pal field. International Journal of DevelopmentalNeuroscience 16:187–197.
Blanchard, S. 2001. Working with phobic parrots.Companion Parrot Quarterly 54:43.
Butler, A.B., and W. Hodos. 1996. Comparative verte-brate neuroanatomy. New York: Wiley-Liss.
Capitanio, J.P. 1986. “Behavioral pathology.” In Com-parative primate biology. Volume 2A: Conservationand ecology, ed. G. Mitchell and J. Erwin, pp.411–454. Alan R. Liss, Inc.
Clark, P. 2001. A vicious cycle: Helping the anxiousparrot. Companion Parrot Quarterly 54:70–79.
Francis, D.D., J. Diorio, D. Liu, and M.J. Meaney. 1999.Nongenomic transmission across generations of ma-ternal behavior and stress responses in the rat.Science 286:1155–1158.
Francis, D.D., J. Diorio, P.M. Plotsky, and M.J. Meaney.2002. Environmental enrichment reverses the effectsof maternal separation on stress reactivity. Journalof Neuroscience 22:7840–7843.
Heinrich, B. 1998. Mind of the raven: Adventures andinvestigations with wolf-birds. New York: CliffStreet Books.
Immelmann, K. 1972. Sexual and other long-term as-pects of imprinting in birds and other species. Ad-vances in the Study of Behavior 4:147–174.
Immelmann, K. 1975. Ecological significance of im-printing and early learning. Annual Review of Eco-logical Systems 6:15–37.
Klinghammer, E. 1967. “Factors influencing choice ofmate in altricial birds.” In Early behavior: Coopera-tive and developmental approaches, ed. H.W. Ste-venson, pp. 5–42. New York: Wiley.
Levine, S. 2001. Primary social relationships influencethe development of the hypothalamic-pituitary-adrenal axis in the rat. Physiology and Behavior73:255–260.
Meaney, M.J. 2001. Maternal care, gene expression,and the transmission of individual differences instress reactivity across generations. Annual Reviewof Neuroscience 24: 1161–1192.
90 Manual of Parrot Behavior

Millam, J.R. 2000. Neonatal handling, behaviour, andreproduction in orange-winged amazons and cock-atiels Amazona amazonica and Nymphicus hol-landicus at the Department of Animal Science,University of California. International Zoo Yearbook37:220–231.
Myers, S.A., J.R. Millam, T.E. Roudybush, and C.R.Grau. 1988. Reproductive success of hand-reared vs.parent-reared cockatiels (Nymphicus hollandicus).Auk 105:536–542.
Navarro, A., and I. Castanon. 2001. Comparative studyof the growth rates of hand-raised and parent-raisedpsittacids in Loro Parque Fundacion. Cyanopsitta60:12–16.
Pepperberg, I.M. 1999. The Alex studies. Boston: Har-vard University Press.
Pepperberg, I.M, J.R. Baughton, and P.A. Banta. 1998.Allospecific vocal learning by grey parrots (Psit-
tacus erithacus): A failure of videotaped instructionunder certain conditions. Journal of ComparativePsychology 109:182–195.
Prather, M.D., M.L. Lavenex, W. Mauldin-Jourdain, A.Mason, J.P. Capitanio, S.P. Mendoza, and D.G.Amaral. 2001. Increased social fear and decreasedfear of objects in monkeys with neonatal amygdalalesions. Neuroscience 106:653–658.
Rowley, I., and G. Chapman. 1986. Cross-fostering,imprinting, and learning in two sympatric species ofcockatoo. Behaviour 96:1–16.
Vazquez, D.M. 1997. Stress and the developing limbic-hypothalamic-pituitary-adrenal axis. Psychoneuro-endocrinology 23:663–700.
West, M.J., and A.P. King. 1990. Mozart’s starling.American Scientist 78:106–114.
10 / Hand-Rearing: Behavioral Impacts and Implications for Captive Parrot Welfare 91

INTRODUCTION
Psittacine development progresses along a pre-dictable path wherein certain behaviors becomeapparent in the young birds as parallel physicalattributes mature. Simply put, when their eyesopen, they begin to see, so eye-opening time is theideal time for human caretakers to provide envi-ronmental enhancements that encourage visualacuity. Likewise, when we see pre-fledglings flaptheir fully feathered wings and hover over a perch,we provide an environment suitable for safefledging. The fledging process itself contributesin many ways to long-term success for psittacinecompanions because during fledging, early be-haviors culminate in a fully active animal totallyengaged in and interactive with its environment,including human caretakers.
The environment is thought to have a strongimpact on young animals, and experiences duringa relatively short period early in the animal’s lifemay have lasting effects on how the animal willreact later in life (Levine 1962; Escorihuela et al.1995). This also holds true for parrots (Coulton etal. 1997; Nicol & Pope 1993; Sheehan 2001).
No matter the stage—from egg to aviary—theenvironment provided to domestically situatedpsittacine birds is largely controlled by humans,with the notable exceptions of weather and nature.
We offer reinforcement suitable for us, not them,when we enrich our home-based situations toencourage desirable and ignore undesirable be-haviors. In the ideal environment, changes areexpected, welcomed, and result in an increase inpositive behavior. Ridley suggests that genes and be-havior are interdependent in that genes are “activeduring life; [genes] switch one another on and off;they respond to the environment” (Ridley 2003).
I intend to describe the initial behaviors ofpsittacine hatchlings, or neonates, who kick them-selves free of their eggs and quickly grow intofuzzy babies. Next, their eyes open and they beginto look at the world around them. Soon their legsare capable of carrying their body weight; theylearn to climb onto the edge of a box to lookaround, flap their growing wings, and vocalizewith their neighbors. By the time they fledge, theyactively participate in their surroundings, and it’sthis environment that becomes the testing andproving grounds of companion behavior. Invari-ably, the most “thoughtfully arranged environ-ments” (Friedman 2002) where parrots live showtelltale signs of birds. Play gyms in the familyroom, organic pellets in the cupboard, a perch inthe shower; parrots have a way of taking over ourhomes with the same ease that they capture our at-tention with their showy plumage and antics.
93
11Behavioral Development of
Psittacine Companions: Neonates,Neophytes, and Fledglings
Phoebe Greene Linden with Andrew U. Luescher
Once you have flown,/ you will walk the Earth with your eyes turned skyward;/ for there youhave been,/ and there you long to return.
—Leonardo DaVinci

THOUGHTFUL ARRANGEMENT OF THEENVIRONMENT
From hatchling to fledgling, young psittacinesmethodically use a thoughtfully arranged envi-ronment to set in motion a fascinating series ofidentifiable developmental progressions that re-spond to experience through constant reorganiza-tion and rebuilding (Ridley 2003; Figure 11.1).Under the right conditions, tiny neonates whoseem helpless grow into competent fledglings. Asthey grow, the skills of psittacine birds aredemonstrated through learned behaviors. We willexamine only the most observable in this text.
Thoughtful caregivers keep the environmentsuitable for keen interaction with the birds as they change and develop. For example, we putaway baby cuddle toys and boxes to make roomfor fledgling landing spots when fledging is im-minent. We offer diets for a variety of age-appropriate circumstances; we’d no more encour-age a 15-month-old cockatoo to take food from asyringe than we’d expect a newly hatched baby tochew through a guava branch. Friedman suggeststhat the developmental process should be one richwith opportunities to “select and strengthen den-drites for companion behavior and let others forfree-ranging behavior fall off the brain-tree”(S.G. Friedman, personal communication, 2003).As a caregiver of young psittacine birds, I thinkoften about which behaviors seen in young birdsenhance, and which detract from, lifelong com-
panionship with humans. Once we find out whatis reinforcing to our birds—depending upon theirage and circumstance—and provide those itemsand interactions, we can deliberately select be-haviors that increase a bird’s success in the envi-ronment of the human home.
Although this chapter concerns itself withyoungsters, it is also pertinent to adult compan-ions. The rigorous environmental, nutritional, andinteractive demands that neonates make are mim-icked by the equally unapologetic demands madeby highly functioning psittacine adults. Babies re-quire warmth, humidity, food, and care in care-fully calibrated amounts without which a panoplyof complications can result. Psittacine caregiversare wise to avail themselves of species-specificneonatal protocols when rearing babies. As weobserve our neonates, we want to encourage be-haviors identified as desirable in companions anddiscourage those that undermine companionship.In these ways, I suggest that appropriate experi-ences in early developmental periods can help ac-climate birds toward growth into successful com-panions.
This approach fosters more bird-like attributesthan those historically accepted in many homesbecause this formula encourages birds to fly, for-age for food, make predictable but not persever-ing noises, be reasonably social to strangers andcontinue to make decisions regarding their dailyactions and reactions. Avian companions I haveraised fly to people of their own volition, becausethey desire interaction. This methodology doesnot result in cookie-cutter companions but the re-verse. When raised in environments tailored to-ward their growth, psittacine companions developindividual personalities, preferences, and pro-pensities.
A Case Study of One
I describe the 240+ psittacine species currentlyidentified by scientists as one group with specificreferences made to individual birds as experienceand space allows. Behavioral scientist SusanFriedman, PhD, advises that each student of be-havior examine his or her subjects best when thesubjects are considered “a case study of one”(Friedman 2002). When we account for individu-ality in this manner, we include considerations ofthe specific set of genetics and environmental in-fluences in order to see each animal distinct from
94 Manual of Parrot Behavior
Figure 11.1. Neophyte parrots such as thisyoung Amazon and Eclectus look out of theirtowel-covered box. (Photo by Layne DavidDicker, courtesy of Santa Barbara Bird Farm.)

others. This is not to say, however, that knowledgeaccumulated over years from many subjects is su-perfluous. Indeed, experience both widens andhones our abilities so that careful observers con-sider more influences on individual behavior, notfewer.
Designed to Be Wild; Bred to Be Captive
For many years I actively cared for psittacineneonates, some only minutes from the egg. Othersbenefited from early parental care. What followsare my observations of these companions who aredesigned to be wild and fly but who are bred to betamed and clipped.
One large contrast between wild and domesticparrots, for example, is that wild psittacines even-tually achieve total independence from their par-ents but captive psittacines do not gain such free-dom from their caregivers. Parrots in domesticsituations remain dependent upon their care-givers, who may sometimes supply and some-times withhold rewards. Often, reinforcementsoffered to captive parrots are commensurate withhuman values, not psittacine sensibilities.Sometimes captivity and companionship mingleuneasily for parrots and people, especially forpersons who are unaware of the lifelong depend-ence inherent in caring for such a potentiallylong-lived animal companion. This enforced de-pendence motivates us to scrutinize behavior atits elemental level—the observable—in order toimprove the fate of tamed captive psittacine com-panions.
NEONATES
Because they hatch blind and mainly naked,psittacine neonates are designated altricial birds.They differ from precocial birds that see, walk,and forage within hours of hatching by being last-ingly reliant upon care and teaching instead ofquickly independent.
Their seeming helplessness, however, does notindicate that psittacine neonates are inactive—onthe contrary. The hatching process is vigorous anddemands that before hatching, psittacine chicksmaneuver, rotate, pip the eggshell, push againstthe egg, and even vocalize (Abramson et al.1995). The hatching process itself can take an ex-traordinarily long time, “up to 79 hours from pip-ping to fully hatched,” reports Abramson for oneHyacinth Macaw (Anodorhynchus hyacinthinus)
(Abramson et al. 1995). Some pre-nestlings napduring the long hatching process. Although mosthatch sooner than Hyacinths, all who emerge inoptimal health have absorbed the yolky nutrientsthat provide the first fuel needed for growth andvigor. I have held and vocalized with hatchingeggs and have watched twisting, pushing babiesemerge from their egg. They often move to aplace away from the cut edge of the broken shellsand immediately fall asleep to recover fromhatching exertions.
The first dropping excreted by neonates I notebecause it signals that the digestive system func-tions. Once the babies wake, they are ready to eatan appropriately thinned-down formulated diet.
Because they sleep so much, newly hatchedpsittacine birds might again seem inactive.However, even this early in development, theirnascent readiness to act in response to the in-creasingly complex environments they will en-counter as they grow becomes apparent. Whenthey are awake, chicks even two days old willstand straight up on their haunches and repeti-tively vocalize when hungry. They push theirsightless heads toward something to touch, lifttheir rumps off the towels upon which they sit,and open their beaks wide. When the flanges(sides) of their beaks are touched, the babiespump up and down to indicate their reflexivereadiness to eat. Though I hand-fed from day 1less than 25 incubator-hatched birds, all were re-sponsive to early stimuli. Their tightly closedeyes, undeveloped muscles, and inability tothermo-regulate indicate developmentally normalphysical limitations. Early sensitivity to vocaliza-tions, food, and gentle touch massage all indicatethat, as Friedman writes, “the feedback loop be-tween behavior and the environment begins”(S.G. Friedman, personal communication, 2003).
When the proper types of comfort are providedand when the babies are healthy, some behaviorsof psittacine neonates are contextually relatedsuch as snuggling/sleeping, pumping/eating, andwiggling/pooping. Snuggling precedes sleeping,pumping precedes eating, and wiggling precedespooping. Backing up, a few quick wiggles offeatherless rumps and tails, a suspenseful pause,the emergence of a well-formed broken string offecal matter surrounded by white urates and clearurine—these are endearing and notable actions.These motions are generally accompanied by a
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 95

sound that ranges from a soft sigh to a tiny grunt.Hatchlings that remain healthy and behave in themanners just described are considered well ontheir way toward further development.
In contrast, chicks who are dehydrated, ill, toocold, or too warm behave differently. If fed an in-appropriate or inadequate diet, if housing is sub-optimal, or if health is questionable, neonates willregurgitate, shiver or pant, cry, stay awake afterfeeding, and act either listless or hyper. Drop-pings may be viscous, scant, or smelly. Babieswho demonstrate these behaviors and types ofelimination need immediate medical evaluation.
Overall, biological parent birds are the optimalcaregivers for psittacine neonates. When they re-spond competently to neonates’ demands, adultpairs make the best psittacine early caregivers. Acockatoo hen, for instance, who wakes at mid-night in order to deliver just one bite of perfectlymixed food at its most palatable temperature toher drowsy baby is the creature most suited to thatjob. Aviculturists are wise to consider ways to in-crease parenting behaviors in captive pairs ratherthan resorting to labor-intensive and sometimesdetrimental artificial incubation and raising ofhatchlings.
Adept parent birds handle hatchling neonateswith ease. In contrast, aviculturists who artifi-cially incubate have alarms bleeping them awakein the dead of night so they can carefully preparethe correct food, check its temperature, wakesleeping chicks, feed them, clean the substrate,return the chicks to the brooder, and hope sleepreturns. I found hand-feeding from day 1 deleteri-ous to my health. In addition, we generally knowthat parent-raised babies are fitter, fatter, andcalmer than birds human-raised from day 1.Christine Sellers, DVM, considers all psittacinebabies fed by humans from day 1 to be “at leastsomewhat compromised” (C. Sellers, personalcommunication, 1998) when compared to parent-fed chicks.
Because we don’t want to allow yelling forfood to become reinforced by the provision offood, we don’t feed neonates when they yell forfood. We feed upon early demand, when chickswake and are hungry, before they cry. A babymonitor alerts us during the night and we respondefficiently (if groggily) in order to avoid teachingour birds that, if they scream loud and long, theirmost basic needs are met.
NEOPHYTES
Once babies’ eyes open, I call them “neophytes”because many of their behaviors are new (Linden1999b; Figure 11.2). As their bodies grow, theirbehaviors both increase and expand daily. This isan ideal time for practitioners to quite deliberatelypromote certain behaviors and deter others.
We are active participants in and enrichers of thegrowing parrot’s environment. The primary (un-learned) reinforcers—finding comfort, gettingfood, taking rest—are in place and now the psit-tacine caregiver begins to control his or her contin-gency on the young parrot’s behavior in order toencourage companionable characteristics and todiminish others. By preventing their success, wecan help deter behaviors that do not promote healthand happiness in captivity while we actively pro-vide reinforcement for behaviors that do.Simultaneously, we promote captivity-related ben-eficial behaviors, dissuade non-companionable be-haviors, and arrange the environment for success.
Just as we keep a careful eye on humidity, tem-perature, and substrate for the neonate, we do thesame for the neophyte, with appropriate amend-ments. Neophytes develop properly in environ-ments large enough for flapping, climbing, hid-ing, and tumbling; where chances for socialinteraction with other birds and with humansabound; where diets conducive to foraging are in-troduced; where opportunities to touch and betouched are prevalent; and where the important
96 Manual of Parrot Behavior
Figure 11.2. Healthy neophytes, such as theseAlexandrine chicks, have bright eyes and appearinquisitive during their busyness. See also colorsection. (Photo by Alice J. Patterson, courtesy ofSanta Barbara Bird Farm.)

lifelong skills of preening and bathing are intro-duced and demonstrated. Enrichment at this stageis important to prevent abnormal behavior such asfeather picking (Meehan & Mench 2002;Meehan, Millam, & Mench 2003). In the follow-ing paragraphs I describe the types of environ-ments that promote these behaviors.
Healthy neophytes grow larger and more inter-active by the moment. At five weeks old, for ex-ample, our African Greys (Psittacus erithacus)are downy-covered lumps that dreamily raise theirheads for feeding. By six weeks, these same ba-bies begin a cautious walk toward light; theytouch colored fabrics, foods, and toys with theirbeaks; and they even begin to preen themselvesand clutchmates. By seven weeks old, they pickup foods in their beaks, vocalize in response tocues, and stare steadily at the sights within theirview. They also quickly scamper back to a dark-ened corner of the box if they are startled in anyway. For this reason, we (aviculturists Abramson,Cravens, Linden, and Speed, among others) raisebabies in containers that provide both privacy andplaces for peeking out. Sheltered areas for restmay be important to reduce skeletal problems inyoung growing birds (Harcourt-Brown 2004). Wewant to encourage birds to look out at the worldwhen they feel safe to do so and to seek and read-ily find comfort when they need it. We give themenvironments where they can escape from feel-
ings of helplessness, fear, and lasting discomfortand experience quick recoveries from upset andtrauma (Figure 11.3).
Along with visual development, neophytes in-crease their physical development. Birds whoonly a week ago mainly slept, ate, and poopednow start to look at, touch, and move toward stim-ulation (Figure 11.4). We watch them as theywatch their environment and its inhabitants—us.Their ears are open too—we want them to hearpleasant sounds because we know that psittacineadult voices can ratchet up volume, tempo, andpitch. Therefore, we keep the neophytes’ habitatas free as possible of screeching and/or meaning-less droning repetitive noises. I once saw a videoof young parrots being fed in a large institutionalenvironment where feedings were dispensed ac-cording to a rigid schedule. Long before “feedingtime,” some of the birds started screaming and theunheeded screams escalated into piercing noisesthat quickly woke the other babies, all of whomscreamed until the ear-plugged hand feeder en-tered the room at the appointed time, wordlesslyfed the birds, then left. This early experience al-lows screaming to continue unabated too long be-cause thereby, “function is established betweenbehavior and consequence” (S.G. Friedman, per-sonal communication, 2003). Screaming forfood—or for any need—is largely avoidable andnot beneficial for birds who, as adults, routinelylose their homes because of loud endless noise
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 97
Figure 11.4. The comfort and security of thecardboard box allow opportunities for this youngGrey Parrot to either peek out, hide, nap, or play.(Photo by Liz Wilson, courtesy of Santa BarbaraBird Farm.)
Figure 11.3. Before fledging, this youngMoluccan Cockatoo tests her feet and balancewhen she climbs onto the edge of the box sheshares with other psittacids. (Photo by LayneDavid Dicker, courtesy of Santa Barbara BirdFarm.)

making. Instead, we want to reward hungry birdswith their food when they make those first “ac-ceptable” noises. Development incorporates notonly food and feedings, however.
Slowly, steadily, over the next few weeks,psittacine birds increase the amount of time theyspend looking out of their containers and de-crease the amount of time they spend hiding andsleeping. However, they still need to both sleepand hide so dual-function containers that affordboth dark and peeking-out sections continue to beused. Inside the box, we add toys and bowls ofcolored foods for touching and visual interest be-cause we want to promote curiosity and explo-ration as we lessen the numbing effects of blankenvironments (Meehan & Mench 2002). We alsotouch the babies to replicate parental contact andto acclimate the birds for future handling. Avicul-turist Katy McElroy’s videotape of MoluccanCockatoo (Cacatua moluccensis) parent birds asthey care for a hatchling is surprising because itshows lavish amounts of attention directed to onelone chick (McElroy 1998).
Neophytes’ observable behaviors include look-ing, touching, and moving but now these behav-iors are more looking around, more touching ofobjects with the beak, and pronounced increasesin physical activities like eating, preening, andtheir first unstudied attempts at exercise.
Neophytes Exercise
At the beginning of the neophyte stage, psittacinewings are just beginning to produce long viableflight feathers, and neophytes regularly flap thesestubby wings (Harcourt-Brown 2004). EclectusParrots (Eclectus roratus) are not the only ones toput their little heads down in the substrate, firmlyplant their feet, and flap their wings repeatedly.African Greys, Psittacula, Cockatoos, Amazons,and Macaws do the same. Their bottom-heavyphysiques serve as ready ballast during these flap-ping exercises.
In preparation for flight, psittacine birds startto hop, jump, and strut. Amazons are especiallygood at strutting. They push their chests out, holdheads high and wings akimbo—then pounce ontoys or roll on their backs for tummy tickles. Allpre-fledged psittacine birds should be givenample opportunity to exercise their legs and feetfor the sheer joy of it, and as good preparation forlandings after flight.
Pre-fledged birds also need to practice climb-ing. Up the sides of their containers they climband perch on the edge. Soon, they are climbingand flapping, then flapping while climbing. Atfirst, they grip tightly onto the perch. Later, theycan be seen flapping vigorously while hovering abare inch from the perch, as they test the forma-tive abilities of their untried wings.
Neophytes Interact
Neophytes act independently. They also minglewith each other. Increasingly, they actively partic-ipate in their environments and interact with theircaregivers. Environmental enrichment early in lifewill increase the motivation for exploration andreduce neophobia (Nicol & Pope 1993; Holsen1986). Early environmental enrichment also fos-ters species-typical behavior and reduces abnor-mal and detrimental behavior (Clayton & Krebs1994; van Hoek & King 1997; Coulton et al.1997). Enrichment also positively influences braindevelopment (e.g., Turner et al. 2002; Rosenzweig& Bennett 1996; Rosenzweig 1984) and learningability (Winocur & Greenwood 1999; Cooper &Zubek 1958; Renner & Rosenzweig 1987).Housing young birds in pairs has been shown to bebeneficial compared to single-bird housing. Pair-housed birds were more interactive with the envi-ronment, less fearful, and engaged more in adap-tive behavior and performed less detrimental orabnormal behavior (Meehan, Garner, & Mench2003). Gentle early handling imposes a mild stresson the young bird but is beneficial because it re-duces detrimental reaction to non-avoidablechronic stress while intensifying potentially life-saving reactions to acute stress (Levine 1960/1973). The more early stimulation is provided tothe young animal, the greater the decrease in emo-tional reactivity (Denenberg 1969).
Some of the behaviors we see in neophytes in-clude these:
Independently, they look at, approach, and finallytouch objects housed in their boxes. Later, inpreparation for fledging, they run, hop, andpractice flapping.
They snuggle, eat, and sleep with each other.Sometimes they pump on each other’s wingtips or other body parts in simulated feedingmotions. If one wakes, is hungry, and beginsmoving, the others quickly wake and homoge-
98 Manual of Parrot Behavior

nous squirming ensues, soon accompanied byfood begging sounds.
They vocalize in response to their caregivers andoften assume the feeding posture: flat bottomon the substrate, neck extended, beak openwide. They also will sit still and even nap whilebeing held and softly touched.
Neophytes Eat
Neophytes remain on formulated diets with appro-priate amounts administered according to the bestspecies-specific schedules. The method of deliv-ery of foods matters little as long as the caliber ofthe feeding experience is positive, unrushed, andas infused with interaction as it is with nutrition.Syringe, paper cup, spoon, or by hand—the imple-ments that deliver food are not nearly as importantas is the undivided attention of the deliverer. Wewant to instill the ideas that eating with a group isfun, that eating a variety of foods is even more fun,and that eating it, shredding it, pulverizing it, andwasting it are the best fun ever.
Here’s how a feeding of neophytes typically oc-curs at my house:
A box inhabitant stirs and so I scrub my hands,then start to warm the food. As neophyteswake, one flaps and falls over, another justflaps. Most stretch upon waking and somepractice making a step or two, but not alwayswith forward motion. The oldest birds advancetoward the opening first and it is usually a pin-feathered head that greets me.
I peel back the towel in response to their posturesand body language. I then establish vocal andphysical contact with the neophytes. For exam-ple, if one still sleeps, I take care to be quietuntil the baby wakes; then vocalizations remainsoft and actions deliberate, predictable, andmodified to soothe, not alarm. As the feedingbegins, one baby invariably stands front andcenter. This is most likely the oldest or most-developed chick. In a mixed clutch box, for ex-ample, an older macaw might be less devel-oped than the younger Grey. The Grey, in thisinstance, would be first to the feeding.
Round-robin type feeding (where each bird gets apredetermined number of morsels or strictmeasure of cc’s in a set rotation) is anathema tome. I feed based on individual assessment ofeach baby in my care. If one ate a small break-
fast, I depend upon that chick to be extra recep-tive to the mid-morning meal. As already notedwith feeding utensils, hand feeders develop, re-fine, and practice the hand-feeding techniquethat works best for them and the babies theynurture. Some colleagues feed exactly meas-ured amounts, others use a syringe or spoon,and still others feed in regimented order, butthose aspects of hand-feeding are of limitedconsequence except if they expedite carefulrecord-keeping. The individual identity of eachbaby under human care is of primary impor-tance, as is that individual’s progress, health,and comfort and the hand feeder’s ability totrack and encourage these important measuresof wellness.
Neophytes Touch
I feed by hand because that’s what I like to do. Ihold real pieces of softened formulated diet be-tween my fingers, which touch the babies approx-imately where and how parent birds use theirbeaks to touch babies’ beaks while feeding. Itwould seem like each feeding would becomemore automatic than the next, but the reverse istrue for me. With each feeding, neophyteschange, albeit slightly. Over a week, cumulativesmall adjustments make a huge difference in thefeeding technique. One day the baby macaw swal-lows the food whole, the next day she mouthseach piece. Some pump on fingers or utensils,some seem to grab food from the air if it’s tossedin their general vicinity. My job includes watch-ing for these subtle shifts as babies use their beaksand tongues to eat, preen, and touch more objects.
An important skill begins at this stage when weteach psittacine neophytes how to interact withand touch human hands. I enjoy reinforcing themwhen they touch me in appropriately gentle ways,just as they provide me with valued reinforcementby being happy to see me, eager for the food, andsolicitous of my touch. I learn about each of themthrough tactile connection, and they learn thatwhen they repeatedly reach for a human hand, re-wards they value result.
Neophytes Preen
The variety of preening-like skills displayed inthis early stage of development is surprising. Thecurrent epidemic of feather-picking in companionparrots (Jenkins 2000) is a bleak situation that has
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 99

so far resisted solution. Because all possibilitiesfor improvement are welcome, hand feeders arewell advised to encourage early beak/tongue co-ordination. For example, the ability to pull amany-segmented item through the beak is a goodone that neophytes can practice with carrot tops,Italian parsley, and other leafy green food items.These skills practiced with greens benefit youngbirds who lack parental or sibling preening andmust solo preen.
Preening and eating skills begin to overlap.Neophytes offered fresh wet greens touch, lick,preen, and shred them. At first, they don’t ingestpalpable quantities of these offerings but inges-tion is not the goal here, practice is: we want themto generalize preening of greens into preening offeathers. We note how they use their tiny tonguesand already surprisingly dexterous beaks to ex-plore the fissures and intricacies of carrots tops,dandelion greens, or chard. Substantive pieces offresh vegetables, especially greens, provide visualas well as tactile stimulation to the increasinglyactive neophytes. Six-week-old Greys (P. eritha-cus) avidly lick wet green leaves while Eclectus(E. roratus) drink the droplets as they fall. Cock-atoo neophytes (Cacatua) will preen carrot topsand baby macaws (Ara) will boldly shred ro-maine. Little tongues and beaks move quickly inrepetitive measured motions. Fine motor skills aresolicited, practiced, and enhanced when we pro-vide appropriate enrichments such as greens andtoys (Figure 11.5).
A plethora of enrichment may ameliorate somefeather-destructive behaviors, especially those tiedto lack of preening practice, motivation, or materi-als (Meehan, Millam, & Mench 2003). We encour-age neophytes to preen inanimate objects when wecan first influence those early attempts to preen.We provide them with enrichments so they canpractice these activities with diligence. Furtherstudy may indicate whether or not the mastery ofearly tongue-beak-preen skills correlates with laterlack of feather-destructive behaviors.
Early preening actions allow neophytes to prac-tice physical skills. Twist and balance of supplespines is challenged as the neophytes bend over,lean down, and nibble at the sheaths of their earlyfeathers. Surprisingly nimble, neophytes ruffletheir budding feathers with deliberation, althoughsometimes they tumble over when they do so.When offered enrichments such as greens, they
stretch their necks long in order to grab a hangingleaf as they jostle together for access to the cho-sen particle. Often, they perform the same mo-tions on the feathers as they do on the greens: lit-tle repetitive nibbles.
At this stage, we want to encourage potentialyoung companions to eat healthy foods, to touchand accept touching from human hands, and topreen themselves and each other. Therefore, weadd reinforcement appropriate to their increasingproficiencies. In so doing, we encourage observ-able behaviors with an eye toward long-term suc-cess. Because the neophyte stage passes quickly,we find ourselves changing reinforcers often,even daily. We encourage exercise, interaction,good eating skills, nice touching behaviors, andsolid preening actions when we arrange the envi-ronment so that the neophytes are positively rein-forced by participating in these activities.
Once psittacine birds reach the stage wherethey are ready, many of the behaviors already ob-served begin to shift unmistakably toward the ul-timate psittacine activity—flying. Precursors tofledging including exercise, interaction, and diet,and all culminate in a series of events that lead toself-propelled airborne flight complete with take-offs, landings, mid-air turns, and recall. I have ex-perienced great joy while fledging psittacinebirds.
100 Manual of Parrot Behavior
Figure 11.5. The young Eclectus male discov-ers that fledging means not only flying throughthe air but also landing in interesting spots.Once in the food tub, he selects a freshly cutguava leaf as his morsel of the moment. (Photoby Liz Wilson, courtesy of Santa Barbara BirdFarm.)

FLEDGLINGS
Fledging principles are best guided by neitherhard right-wing nor hard left-wing practices.Hard right-wing thinking assumes that birds whodo not fledge don’t miss the experience. Hardleft-wing thinking—that all birds must fly all thetime, preferably “freely” outside—is equally un-supportable for all but a minority of psittacinecaregivers. Like the obsolete one wing only clip,either hard direction results in a crash. We canachieve a balanced approach to fledging, onewhere the birds get a chance to experience theirpersonal kinesiology and are adequately preparedfor life and are then later suitably confined bywell-measured standards of domesticity.
A balanced fledging program includes assess-ment of the physical characteristics of all whowill participate in the young psittacine’s domesticflight. Caregivers, children, dogs, cats, and visi-tors all participate in the equation.
A program that balances the bird’s capabilitiesin a thoughtfully arranged environment can beachieved, but it takes total “flock” cooperation aswell as knowledge about the domestication proc-ess. Domestication is in its earliest stages withparrots, most of whom are only one generation outof the jungle. Price writes, “Domestication is aprocess of adaptation to the captive environmentthat includes both genetic changes occurring overgenerations and environmentally induced develop-mental events (such as taming) that occur withinthe lifetime of an individual” (Price 1984).Further, the “capacity to perform the behaviorsseen in the repertoire of wild counterparts re-mains, although the threshold for performancemay be altered” (Price 1999). It is this behavioralthreshold that caregivers must consider for giv-ing parrots the richest experience possible as ourcompanions.
Before I describe my limited domesticallybased fledging experiences, I must give credit totwo major flight-related influences, one human,one psittacine. The former is Eb Cravens, an avi-culturist in Hawaii whose articles published in thelate 1980s recounted experiences with birds inflight both indoors and outdoors (AFA Watchbird,“Birdkeeping Naturally,” 1988–1990). Cravens’swritings continue to influence many. The psitta-cine influence on my fledging program, describednext, comes from a flock of feral amazons.
OBSERVATIONS OF WILD FLEDGLINGS
A Brief History of a Feral Flock of AmazonParrots
A flock of amazon parrots, currently nine in num-ber, visits our property and inspires countlesshours of observation. Our aviary inhabitants greettheir free-flying counterparts with recognizableyells while my spouse, Harry, and I quickly fillfeed bowls to further reinforce their visits.Harry’s interest in this flock goes back nearly 25years to when he first watched a Double Yellow-Headed Amazon (Amazona ochrocephala; Figure11.6) and three Lilac-crowned Amazons (Ama-zona finschi) forage in a tree located close to thefreeway, seven miles from our house. This obso-lete viewing area is now covered in concrete, thetree long replaced by urban blankness, and theflock has moved its foraging areas further up themountains. Fortuitously, they include our prop-erty in the hills in their chosen territory. Theymake brief stops at the feeding stations we supplyfor them but it’s clear to us that their territory isvast because sightings extend for at least 35 milesin every land-based direction from our propertyor from their nesting sites, which we believe areclose by. We have yet to find their nesting sites,although we look with diligence. We havewatched wild pairs visit two different sites. As the
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 101
Figure 11.6. The naturalized Double Yellow-headed Amazon, a proven male, keeps lookoutfor his fledglings amid the trees at the SantaBarbara Bird Farm, where the wild flock’s dailyactivities are noted. See also color section.(Photo by Harry A. Linden, courtesy of SantaBarbara Bird Farm.)

frequency of their visits increase, so does our an-ticipation. We have watched them gnaw at and en-large openings, sit inside good nesting areas, andeven mate close by. However, even after weeks offixing up, they have left these observable sites forplaces still undiscovered by us. Clearly, theseobservations—made in our backyard and the sur-rounding rural areas—do not constitute a scien-tific study. The flock’s almost daily visits give uslimited information on their many activities butwe avidly watch them anyway.
Initially, this flock was formed of probable es-capees from local aviaries or homes. The DoubleYellow-headed Amazon (Amazona ochrocephalatresmariae), who we later discovered was a male,and his chosen Lilac-crowned Amazon (Amazonafinschi; Figure 11.7) hen were the first to repro-duce. Although they both disappeared during thewinter of 2001, they produced many offspring,three of whom still visit. There are two otherLilac-crowneds initially referred to as “the sen-try” and “the babysitter.” We assumed they werethe same sex because we never saw interactionbetween them but they eventually proved them-selves a viable pair. They’ve fledged five babies,one of whom still frequents our property. Overthe years, the A. O. tresmariae and A. finschi pairproduced eight hybrid chicks that we watchedfledge, three of whom still visit. Undoubtedly,both pairs have produced babies who never madeit to the fledging grounds we observe in ourbackyard and surrounding mountains and val-leys, and so these are not included in this countnor meant as a commentary on fertility. Thesenumbers do, however, reflect survivability aftersuccessful hatching and fledging for this groupof birds.
How We Determine Unseen Nesting Schedules
Gravity bound, we humans are locked into limitedobservations from which we make suppositionsthat sometimes prove out, sometimes do not.Since these birds have never—so far—nestedwhere we can see them, we have to calculate theirseasonal schedules.
For example, we assume that these wild birdsgo to nest when the hens begin to miss the twice-daily feedings. When the hens stop coming, wemark this date on a calendar kept for this pur-pose. At the same time that the hens cease theirvisits, we consistently make two interesting ob-
servations. First, the male birds come more fre-quently than the normal dawn and sunset times.Second, at the same time the hens stop coming,the males’ beaks become encrusted with food,presumably from feeding the roosting hens, un-doubtedly a messy process, at least for thesepairs of birds.
Now, the males make their way in solitude andgo from place to place among the feeding stationsas they peer into each food cup. If we do not pro-vide extra food quickly enough (our schedulesmust change to accommodate theirs), they call orpace while looking closely at the door from whichtheir food servers emerge. One feeding malecomes right to our windows and peers inside,yells loudly, then waits, several times a day. Othertimes the males sit camouflaged by a tree in ab-solute silence, depending on the hawk status andtheir hunger.
Approximately four weeks after the hens stopcoming, males increase their visits and the inten-sity of their solicitations aimed at us. We note thatthe males have even messier beaks. At this time,the feeding males have very little interaction withother flock members; they eat quickly and flyaway, often only to return an hour or so later,seemingly impatient and important. As I stand inmy carefully disinfected kitchen, I watch themales with their food-coated beaks gobbling foodto feed to undoubtedly equally untidy hens. Wemark this date on our calendar with the note“probable hatch.”
102 Manual of Parrot Behavior
Figure 11.7. This Lilac-crowned Amazon fledg-ling received flying lessons from her parents andolder siblings. (Photo by Harry A. Linden, cour-tesy of Santa Barbara Bird Farm.)

About eight weeks after the probable hatch, the mother birds show back up at our feeders. Thehens’ beaks are also noticeably soiled, but themother birds are decidedly less active than the males. If other parrots are present, the feedinghens hang around the food stations and preenthemselves while the males fly away, then back,then away again.
Day of Celebration: A Wild Fledgling Appears
With amazons, we expect fledging at approxi-mately 10.5 weeks after our probable hatch date.Ten days to three weeks after that day, if fortuneis with us, we see a wild fledgling, new to theproperty. Actually, we hear about new arrivalswell before we see them because all of our aviaryand pet birds comment—loudly, as if heralding agreat event—on the first sighting of a new parrot.These enthusiastic greetings continue for severaldays and are soon enough answered by the fledg-lings’ own contributions to flock communication,food begging, and calling.
These wild-hatched fledglings make few foodbegging noises, but they do make some, espe-cially early in the fledging process. Mostly theyare closely attended and thus excessive noises areunnecessary. In particular, the father birds areconsistently attentive. Rarely do we hear pro-longed food begging noises that, with amazons,consist of distinctly resonant honking noises,identifiable nearly a mile from the source.
We watch fledglings in the fruit trees as theywatch their parents and older siblings. The moreexperienced birds march through the foliage, se-lect an orange, for instance, take a single bite ofit, drop it, and continue on their way. The largedark eyes of the fledglings are riveted on thesefeeding activities and we watch them similarlymesmerized. Field glasses allow us to see the de-tails of their faces, marked by dark eyes, unmis-takably untidy beaks, and individualized features.
After several observational sessions, we see thefledglings bounce along flexible tree branches andmake their initial attempts at independent feeding.During the next weeks, parent birds continue tofeed the wild fledglings, but with decreasing fre-quency. The fledglings increase their independentforaging and eating activities (Figure 11.8).
The begging and honking noises also diminishand are replaced by tremendous amounts of air-borne activities. Often, we worry that the fledg-
lings aren’t eating enough, because they fly and flyand spend measurably less time actually at thefeeding stations than do older flock members. Theyalleviate our fears when their flight proficiency in-creases day by day. Therefore, foraging, eating, andflying sessions increase in frequency and duration.
After the morning feeding, fledglings will waiton a branch and are eventually met by one flockmember who takes them through the sky, acrossthe valley, and back to landing perches. Thesedaily flying lessons occur for about two weeks.During this time some of the successful fledg-lings follow their flight instructor, coordinatethemselves, and keep up with physical demandsof flight. Unfortunately, others do not.
The Wild Fledgling Who Didn’t Come WhenCalled
In 1998, we observed two hybrid fledglings andnoted them as healthy looking. One stayed closeby the parent birds but the other was late to arriveand missed several gatherings altogether. One dayI heard both parent birds as they sat on a high ex-posed tree branch and called long and loudly.After ten minutes, the fledgling arrived and theyall eventually ate. Two days later, the parent birdsresumed their loud calling and the tardy fledglingeventually arrived. The next day, the parent birdscalled but the fledgling didn’t come. The next day,the parent birds did not call and the fledgling wasnever again seen.
Whenever there is excess vocalization, trouble
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 103
Figure 11.8. Another hybrid chick from aproven disparate and now long-missing pair.(Photo by Harry A. Linden, courtesy of SantaBarbara Bird Farm.)

is often brewing. I am usually heartened by howlittle noise the wild fledglings make. Mainly, theirneeds are met, quietly. When they play in trees,however, they play loudly, usually with each otheror with older siblings. Games such as “get yourtummy,” “hang from one toe while yelling,” and“swing on a branch” seem to be flock favorites.One such fledgling, however, didn’t play suchgames and frankly confounded us when he or shedid not stop making food begging noises.
The Wild Fledgling Who Cried
Well after the other fledglings had stopped mak-ing food-solicitation noises, a particular fledgingbird repetitively honked morning and evening.Whole hours went by punctuated by the noise yetwe never saw this fledgling come to the feedingstations. Field glasses afforded me views of thisyoung amazon (A. finschi) who usually had anolder sibling in a nearby branch and who beggedand begged for food. According to our calcula-tions, this bird was nearly nine months old beforehe or she was seen no more and the noises ceased.
HOME-BASED EXPERIENCES WITHFLEDGING
Many years ago when I first raised psittacinebirds, it was not unusual to clip their wings on theday of their first flight. After all, it is highly in-convenient, usually messy, and sometimes dan-gerous to have young birds taking off and landingwith abandon. However, because many of our petcompanions are in full flight, it seems unfair toclip the babies who repeatedly indicated theirreadiness to fly (Figure 11.9). While the olderpets capable of flight stay on their perches andpreen, chew toys, or just hang out, fledglingsquiver, flap, look for landing spots, and seem rar-ing to go. After several seasons of clipping birds,it became apparent that early wing clipping isanathema to their genetic impulses. The chal-lenges became clear: to promote flight so that thebenefits associated with fledging are experienced(Linden 1998) while simultaneously increasingthe characteristics that contribute to successfulcaptivity. Concurrently, we want to overlap be-haviors that encourage daily explorations of vasthabitats and far-ranging food foraging excursionswith skills that include shorter-distance con-trolled flight and foraging in a cage, aviary, orother designated habitat. Because these not-yet-
domesticated animals surely share their wildcounterparts’ “behavioral capacities and responsethresholds” (Meehan, Millam, & Mench 2003),they can be expected to be highly motivated toperform behavior associated with food procure-ment in the wild even though captive feedingmethods may meet their nutritional needs. Forparrots, foraging is its own reward.
Gradual clipping remains a viable alternativefor many companion birds and leaves them some-where between fully flighted and fully clipped.Birds should be left in full flight for as long aspossible but after a time, in order to achieve bal-ance between wild and domestic success, manyneed to be trimmed at least somewhat. In a grad-ual clip, only the first four to six feathers aretrimmed so that some flight is possible and en-couraged. In this clip, clients are advised thattheir birds can fly and indeed birds so trimmedoften fly in a straight line from a play gym to awaiting hand, for instance. A well-tended homepoliced by vigilant responsible clients must becoupled with well-behaved birds in order to allowfor successful partial or full flight (Linden 1998).
The Fledgling Aviary
The boon to any home-based fledging program isa well-equipped and commodious (Cravens 1990)
104 Manual of Parrot Behavior
Figure 11.9. Fledgling aviaries should be largeenough for the parrots to fully extend their wingsin many directions and then fly and land, notmerely hop and leap. The aviary enjoyed bythese Slender-billed Cockatoos is 15’ x 15’ andsits amid loquat, orange, and nectarine trees.(Photo by Layne David Dicker, courtesy of SantaBarbara Bird Farm.)

fledgling aviary (Figure 11.10). We include in-ducements to exercise such as rope and naturalperches, multitudes of bowls, colorful toys, forag-ing opportunities, and showers in the aviary. Ofcourse, the human attendant is usually the mostfavored enrichment. We resist the temptation tostuff the aviary so full of toys that the birds do nothave room to spread their wings and fly. Here,they continue to practice the lessons they firstlearned in the house: fling food around, find a sib-ling to tickle, fly to a waiting hand, follow me toanother side of the flight cage, land in lots ofplaces. When they miss a landing spot, the birdslearn to right themselves up, shake off the fall andresume action newly informed. Their learningcurve is amazingly fast as long as the environ-ment supplies thoughtfully arranged enticementsand continual reinforcement.
How Fledglings Exercise: Physical Benefits ofFledging
Most obviously, we see that psittacine birds whosuccessfully complete a fledging process attain arobust physique. Birds who fly develop pectoralmuscles that are firm when palpated. They standtall, walk and hop across moving branches, foragefor foods, and eat just the right amounts. Duringfledging, athletic abilities are strengthened—healthy respiration, strong musculature, smooth
coordination (Linden 1998) and life-enhancingeating habits result.
Strength, endurance, climbing skills, and land-ing abilities blossom during fledging and posi-tively influence confidence. Without confidence,wild psittacids would become overdependent bur-dens on flock mates; those lacking confidencewould surely not survive. Coordination comesfrom using personal kinesiology; coordinationleads to confidence, a distinct psychological hall-mark of well-actuated beings (Linden 1999a).Domestic parrots also benefit from confidencewhen they amuse themselves with independentplay, napping, preening, and foraging. Abilities toself-occupy are highly desirable in adult psitta-cine companions and are the result of these form-ative experiences.
How Fledglings Interact: Social Benefits ofFledging
Early experiences with flight introduce youngbirds to the necessity of social skills becausehealthy fledging includes flock activities wheremany participate. Aunts, uncles, cousins, grand-parents, older siblings—surely all influence theyoung fledglings. In addition to the social experi-ences inherent with hand-rearing, birds learn crit-ical social lessons when fledging. The importanceof following instructions and careful observationof others and the environment are reinforced.Surely well-traveled psittacine fledglings run intomany new acquaintances where they experiencedeferment and well-modulated assertion. Theylearn to leave unsavory situations, propelled bytheir physical strength. During this time, fledg-lings learn to come, follow, and obey flock sig-nals. They also balance dependence—being withthe flock—with independence when they maketheir own decisions, try a new path, or stop at theberry patch for an unscheduled snack. Duringfledging, a healthy mix of dependence and inde-pendence is tried and amended according to expe-rience.
How Fledglings Learn: Mental Benefits ofFledging
If we want to replicate a full fledging process inthe domestic psittacine—one that includes impor-tant lessons learned by wild psittacines—thechallenges are many. Wild birds definitely have amuch richer fledging experience than birds con-
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 105
Figure 11.10. The author’s one-year-old Blue-fronted Amazon, Bucket, meets his prospectivemate, Bonnet, for the first time as she sits on aperch with her clutch mates. Fifteen years later,they are still together. See also color section.(Photo by Margaret Ames, courtesy of SantaBarbara Bird Farm.)

fined in homes, but these rich experiences are notwithout dangers, as illustrated by the death rate ofnaturalized parrots. Still, in the ideal world, oncethey land on the branch of a tree, wild psittacinefledglings look around their immediate habitat—they take in the lay of the land, so to speak. Theywatch the sky and ground for predators, calculatethe distance to the feeding stations, gauge anglesof descent and ascent, and watch a bug crawlalong the vein of a leaf. Once airborne, these ob-servations and calculations continue as they fly influid formation with the flock and learn the times,places, and circumstances under which to land,keep going, or flee with alacrity. Certain calls per-colate during flight—these are not eating noises,nor are they standard calling noises. The callsmade during flight use another vocabulary andvolume altogether.
In contrast, domestically situated parrotsoften cope with fully clipped wings (enforceddisablement) and are confined in a cage for con-secutive hours where they endure hours of quietsedentary solitude. We combat these domesti-cally inflicted negatives—as best as possible—through home-based fledging and full-timehuman attendants.
How Fledglings Eat: The Benefits of Foraging
Right before they fly, fledglings’ appetites de-crease in concert with an increase in the time theyspend exercising and exploring. At this time,fledglings’ eating skills center around foragingexcursions.
I used to chase after fledglings and beg them tostop flying in order to eat. Now I encourage themto come find the food I’ve placed in a variety oflocations (Figure 11.11). In this way, we inducehours of busy time while we check unnaturalsedentary behaviors.
Psittacine fledglings remain unweaned forvarying amounts of time. From the time they flyto the time they wean is specific to the variousspecies. Moreover, it’s frequently reported thatbabies wean on individual schedules as well.“Weaning is the process by which birds learn toeat on their own without the assistance of theirparents. Once a bird reaches its peak weight, itbegins to eat on its own, perch, fly, and finallywean” (Abramson et al. 1995). Therefore, fledg-ing and weaning are both processes that birds un-dergo somewhat concurrently.
How Fledglings Touch: The Benefits ofBusyness
Fledglings use their tongues to explore foot toys,fissures, and food. Their beak strength also in-creases and they repeatedly try to bite, gnaw, andchew on whatever is handy. Usually, human handsare most handy. This is the ideal time to discour-age the propensity young birds have for chewingwhat’s attached to nerve endings and to encouragechewing, instead, appropriate inanimate objects.We supply the environment with many items ap-propriate for chewing: soft wood, fresh treebranches, untreated wicker baskets, species-appropriate toys. We stock our pockets with sur-prises sure to please inquisitive beaks.
Whenever young birds are intent upon playingwith a toy, we let their attention remain riveted onthat object (Figure 11.12). We do not interrupt abusy bird. Instead, we wait for the play session tobe over, then we praise the results of the bird’s ac-tivity. Hopefully, this is a pile of pulverized or-ganic materials.
Many fledglings enjoy cuddling sessions andthese are certainly part of many long-termhuman/psittacine relationships. However, it’s easyfor trimmed birds, especially cockatoos andmacaws, to want, to like, then to vocally demand,an amount of physical contact incompatible withhuman time allowances. Psittacines who scream
106 Manual of Parrot Behavior
Figure 11.11. Rosie, a one-year-old Green-winged Macaw, occasionally needs encourage-ment to eat foods selected by her caregiver. Ahandheld bowl and lots of verbal interactionassist her in making good food choices. Seealso color section. (Photo by Layne David Dicker, courtesy of Santa Barbara Bird Farm.)

for attention do so because they have been rein-forced for it in the past, and this noise usually hap-pens on a schedule that conflicts with human-based reality. Therefore, it’s best to keep touching,cuddling, and holding sessions on a fluid schedule.However, most well-trained humans do recognizeand at least intermittently (and happily) reinforcecompanion birds who appropriately request neckrubs. Neck, head, and face scratching are favoredareas for physical interaction and most sociallycomfortable birds will accept assistance in preen-ing these areas. Additionally, we want to regularlytouch birds’ toes, feet, and legs in order to accli-mate them to nail trims and blood draws. We wantto familiarize psittacine birds with handling bytouching them and by allowing them to touch us.
How Fledglings Preen: The Benefits of GoodHygiene
Fledglings need lots of space in order to properlypreen. Preening in a cage is akin to a person doingjumping jacks in a broom closet. When givenshowers and plenty of space, fledglings learn topreen their own tails, wings, and backs (Figures11.13 and 11.14). The results are breathtaking forcaregivers and may provide psittacine compan-ions with lifelong preening skills as an additionalline of defense against boredom.
Height Inconvenience: A Lesson of Fledging
Now that our psittacine fledglings exercise, inter-act, learn, eat, touch, and preen appropriately, it’s
time for them to practice some manners that helpthem integrate into a human flock. I teach two les-sons that are important to me: (1) How to stepdown from tall heights, and (2) how to land onplaces that are not my head. As an adjunct to Dr.Friedman’s discussion about height dominance inchapter 14, I’d like to add the results of my ownobservations on this topic.
Previously, I went along with the construct“height dominance” because I observed that thewild parent birds would not feed fledglings whostood above them. Obviously, it’s inconvenient forparent birds to regurgitate uphill. They ignore thesolicitation of babies who are overhead and in-stead feed those on the downward slope closest to
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 107
Figure 11.12. This young African Grey Parrottries her beak at unwrapping a birthday gift.(Photo by Phoebe Greene Linden, courtesy ofSanta Barbara Bird Farm.)
Figure 11.13. The art of preening occupiesmany moments in parrots’ daily lives. Humancaretakers can promote preening with praise andby giving parrots the baths and space necessaryfor expansive preening sessions. See also colorsection. (Photo by Kelly Flynn, courtesy of SantaBarbara Bird Farm.)

their beaks. The parents favor this position for itsease and the fledglings favor it for results: theylearn to sit slightly below or right next to parentsand get fed; sit above and go hungry. Begging ba-bies who sit overhead are not trying to dominatetheir parents, they just demonstrate their positionon the learning curve. Undoubtedly, arboreal ani-mals such as parrots enjoy sitting in trees andsometimes the view is best from the highestbranches. That doesn’t mean, however, that hang-ing out in the middle branches is submissive anymore than foraging in the middle dense foliage isuseless.
For me, the inconvenience of reaching overheadto grasp parrots that fly to the tops of curtain rodsor ceiling beams is a deterrent. In the past, I wasconcerned that parrots—even young fledglings—
would try to dominate me if they got above myhead, so I anxiously removed them from all thesepreferred (to them) places. In 2003, however, Itested a new theory explained by Dr. Luescherwho thought that birds were reluctant to step onfingers that came from below because this handposition was unfamiliar to them (A.U. Luescher,personal communication, 2003). This made greatsense to me—a hand that is offered palm up orwith the flat ridge presented at chest level cer-tainly looks very different from waggling fingersproffered from below.
I decided to train my fledglings and my maturecompanions to see if they would accept bunchedfingertips that come from below as a suitablestep surface. Once I stopped grabbing for theirtoes, it took, on average, three tries before allwould step onto this new finger configuration. Irealized that previously they weren’t trying todominate me, they were just unsure of such astrange stepping place and needed training inorder to accept it. I learned that having birdsperched overhead doesn’t make them dominantany more than it makes me submissive, but itdoes remain inconvenient.
Still, in order to supply them with a fledgingexperience commensurate with their needs, notmine, I modified places that were previously “offlimits” into new landing spaces. The top of the re-frigerator, tops of cages, and even the top of mygrandmother’s antique secretary (covered withwashable padding) became takeoff and landingspots for the 2003 fledglings: four Greys, twoMacaws, two Cockatoos, two Amazons, and oneDerbyan (Psittacula derbiana) Gidget. Withoutexception, these birds learned to step up as read-ily from these overhead places as they did fromchest-level places when they received rewardsthey valued for doing so. Although they now read-ily step from overhead places, it’s still inconven-ient for me to stand on tiptoe with my armstretched as far as it will go in order to get thebirds to come to me. Instead, I decided to teachthem to fly down from their lofty perches(Cravens 2001).
It proved simple to train the fledglings to flydown. Keeping them from landing on top of myhead was more difficult. Apparently, the combi-nation of slippery hair and a dome-shaped land-ing surface necessitates the use of eight sharpnails to secure good touchdown. Rather quickly I
108 Manual of Parrot Behavior
Figure 11.14. Young Blue-and-gold Macawtwists and reaches as she preens her tail featherwhile outdoors. See also color section. (Photo byKelly Flynn, courtesy of Santa Barbara BirdFarm.)

knew that I would need to enrich the non-head en-vironment in order to save my scalp. Alternatelanding spots were created: baskets weighted inthe bottoms were placed all over and the birdswere densely reinforced for landing on them.Similarly, I became adept at shooting my hand upin the air right behind my head to intercept theparrot landing gear. Birds who landed on my handor on basket handles were reinforced with ticklesand praise; those who landed on my head weresilently set down elsewhere. In four days, the headlanding incidents disappeared while the overalllandings increased.
Psittacine Companions Who Never Fledgedand Who Do Not Fly
Sadly, the majority of psittacids raised for thecompanion market will not experience a truefledging process and may never actually fly be-cause their environments are not provisioned forsuch development. Space, time, and commitmentlimitations abound, and some aviculturists, evi-dence to the contrary, contend that fledging is un-necessary or extravagant. The question remains:Can a suitably developed psittacine companionwho never flies remain a viable lifelong pet? Theanswer to that question depends, of course, onwhat environments shape the experiences duringthe time of development normally occupied byflight and after.
If caregivers can turn on the impulses for exer-cise, interaction, eating, foraging, touching, andpreening that normally activate during flight, thenyes, the result can be a well-developed compan-ion. Persons who decide to not fledge birds obli-gate themselves to construct experiences com-mensurate with fledging wherein parrots exercise,interact, learn, and forage in equivalent ways.When deprived of flight, many birds use repeti-tious behaviors like yelling and overpreening assubstitute actions and therefore require competentredirection. Clipped companion birds can be rein-forced for athletic exercises other than flight suchas climbing, swinging, and flapping. However,flapping stubby clipped wings is not nearly as sat-isfying as flapping full luxuriant wings, and soclipped birds must be appropriately and extrava-gantly enticed and reinforced to keep up the exer-cise needed for cardiovascular, respiratory, andmuscular health.
Even clipped birds can learn to interact socially
so long as they are taught that they need not bitein order to drive away unwanted attentions.Flighted birds flee from unsolicited interest andso find biting largely unnecessary, but clippedbirds, lacking escape, often bite to drive away per-ceived intruders and other annoyances. The socialinteractions of clipped birds often land on the sideof “overdependent” because, lacking their ownresources for exploration, they depend uponhuman caregivers for entertainment, transporta-tion, and to save them from less-favored persons.Therefore, conscientious caregivers of clippedcompanion parrots provide rope walkways and avariety of play gyms and daily activities to ensurewell-adjusted lifelong companions who can bothavoid and seek out social interaction, dependingon what is reinforcing for the birds, not for thehumans.
Of course clipped psittacids can develophealthy eating habits, but their environments alsoneed to be thoughtfully arranged to stimulate for-aging, shredding, and mulching behaviors. Com-panions who get only one neatly prepared bowl offood must be given other enrichments that en-courage movement and action.
Many clipped companion parrots toleratetouching and learn to touch their primary care-givers with gentle ease, but lacking diversity inexperience, they can become more limited asyears go by. Because they are stuck on perches,clipped birds are often the unwitting recipients ofunwanted touching by strangers and may learn toeschew touching as a result. Similarly, flightedbirds also avoid unneeded, unwanted, and unap-preciated touches, the value of which seems to bedistinctly human, not avian. With few exceptions,humans like to touch birds more than most birdslike to be touched by humans. Many psittacids—clipped or unclipped—learn to tolerate touching,but few enjoy it into adulthood. Nevertheless,both clipped and flighted birds can and do touchhumans in very positive ways by stepping up onthe offered hand, by taking treats nicely, and bysitting on laps, chests, or shoulders during relax-ation times (Figure 11.15).
Clipped birds can develop very good preeningskills, especially when they are supplied withlarge outdoor play gyms that they occupy underclose supervision. Nevertheless, the immediatescrutiny needed to guard defenseless clippedbirds from danger often diminishes the freedom
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 109

of large outdoor perches. Such supervision neednot diminish good preening skills. Just like weleave alone birds intent upon toy destruction inorder to pursue their goal, we also leave birds whoare preening to their own devices.
It’s difficult for clipped birds to develop a senseof independence when they must be constantlymonitored. However, flighted birds require com-mensurate supervision since it’s not feasible tohave fully flighted animals zooming heedlesslyaround one’s home. Therefore, we must concludethat all birds in captivity need good supervision inthoughtfully arranged environments.
SUMMARY
Psittacine neonates, neophytes, and fledglings allbenefit from thoughtfully arranged environmentsthat reinforce desirable companion characteristicsand skills. As development progresses, we seepsittacines respond to their environment throughconstant reorganization of observable behaviors.Domestically situated psittacines, whetherflighted or clipped, remain dependent upon theircaregivers to supply their environment with en-richment and rewards that are valuable to them.
During fledging, I see many of the characteris-tics I value in companion birds come to fruition.Exercise, interaction, foraging for foods, touch-ing, and good preening skills obliterate (or at leastgreatly reduce) screaming, picking, biting, and
other cage-bound type behaviors. Surely thegreatest challenges are still ahead of psittacinecaregivers as we reinforce our psittacid compan-ions for adapting to our environment while simul-taneously enriching that environment in order toaccommodate greater portions of their wildness.
REFERENCES
Abramson, J., B.L. Speer, and J.B. Thomsen. 1995. Thelarge macaws, pp. 177, 204. Fort Bragg, CA:Raintree Publications.
Clayton, N.S., and J.R. Krebs. 1994. Hippocampalgrowth and attrition in birds affected by experience.Proceedings of the National Academy of ScienceUSA 19:7410–7414.
Cooper, R.M., and J.P. Zubek. 1958. Effects of enrichedand restricted early environments on the learningability of bright and dull rats. Canadian Journal ofPsychology 12:159–164.
Coulton, L.E., N.K. Waran, and R.J. Young. 1997.Effect of foraging enrichment on the behavior ofparrots. Animal Welfare 6:357–363.
Cravens, E. 1988–present. Various columns and articlesas published in AFA Watchbird, Parrots, OriginalFlying Machine, and others.
Cravens, E. 2002. The backyard aviculturist. OriginalFlying Machine 2:58–59.
Cravens, E. 2002. The links between mental alertnessand flight. Parrots 55:12–13.
Denenberg, V.H. 1969. Open field behavior in the rat:What does it mean? Annals of the New York Academyof Sciences 159:852–859.
Escorihuela, R.M., A. Fernandez-Teruel, A. Tobena,N.M. Vivas, F. Marmol, A. Badia, A., and M. Diers-sen. 1995. Early environmental stimulation produceslong-lasting changes on ß-adrenoceptor transductionsystems. Neurobiology of Learning and Memory64:49–57.
Friedman, S.G. 2002. “Living and learning with par-rots.” Online class at www.parrottalk.com.
Harcourt-Brown, N. 2004. Development of the skele-ton and feathers of dusky parrots (Pionus fuscus) inrelation to their behavior. Veterinary Record 154:42–48.
Holsen, R.R. 1986. Feeding neophobia: A possible ex-planation for the differential maze performance ofrats reared in enriched or isolated environments.Physiology and Behavior 38:191–201.
Jenkins, T. 2000. “Feather picking in companion par-rots.” CPQ Conference paper.
Levine, S. 1960/1973. “Stimulation in infancy.” InReadings from Scientific American: The nature andnurture of behavior—Developmental psychology,pp. 55–61. Scientific American Ltd.
110 Manual of Parrot Behavior
Figure 11.15. Pauly, a mature Yellow-napedAmazon male, sits protectively on his humancaregiver’s arm. The healthy glow of good nutri-tion and hygiene are evident in his colors andfeather condition. (Photo by Layne David Dicker,courtesy of Santa Barbara Bird Farm.)

Levine, S. 1962. “Psychophysiological effects of infan-tile stimulation.” In Roots of behavior: Genetics, in-stinct and socialization in animal behavior, ed. E.L.Bliss, pp. 246–253. New York: Harper and Brothers.
Linden, P.G. 1999a. “Fledging and flight for aviancompanions.” Proceedings of the Association ofAvian Veterinarians, New Orleans, LA, pp. 61–76.
Linden, P.G. 1999b. Teaching psittacine birds to learn.Seminars in Avian and Exotic Pet Medicine 8(4):154–164.
Linden, P.G. 1998. “Behavioral development of thecompanion psittacine bird.” Proceedings of the As-sociation of Avian Veterinarians, St Paul, MN, pp.138–143.
McElroy, K. 1998. “Parent-rearing moluccan cockatoosand breeding cockatoos in outdoor aviaries in north-ern climates.” Proceedings of the Midwest AvianResearch Exposition, Toledo, OH, pp. 95–118.
Meehan, C.L., J.P. Garner, and J.A. Mench. 2003.Isosexual pair housing improves the welfare ofyoung Amazon parrots. Applied Animal BehaviorScience 81:73–88.
Meehan, C. L., and J.A. Mench. 2002. Environmentalenrichment affects the fear and exploratory re-sponses to novelty of young Amazon parrots.Applied Animal Behavior Science 79:75–88.
Meehan, C. L., J.R. Millam, and J.A. Mench. 2003.Foraging opportunity and increased physical com-plexity both prevent and reduce psychogenic featherpicking by young amazon parrots. Applied AnimalBehavior Science 80:71–85.
Nicol, C.J., & S.J. Pope. 1993. A comparison of the be-havior of solitary and group-housed budgerigars.Animal Welfare 2:269–277.
Price, E.O. 1984. Behavioral aspects of animal domes-tication. Q Rev Biol 59:1–32.
Price, E.O. 1999. Behavioral development in animalsundergoing domestication. Applied Animal BehaviorScience 65:245–271.
Renner, M.J., and M.R. Rosenzweig, M.R. 1987.Enriched and impoverished environments: Effectson brain and behavior. New York: Springer-Verlag.
Ridley, M. 2003. Nature via nurture: Genes, experi-ence, and what makes us human. New York: HarperCollins.
Rosenzweig, M.R. 1984. Experience, memory and thebrain. American Psychologist 39:365–376.
Rosenzweig, M.R., and E.L. Bennett. 1996. Psycho-biology of plasticity; effects of training and expe-rience on brain and behavior. Behavioral BrainResearch 78:57–65.
Sheehan, K.L. 2001. The effects of environmental en-richment and post-natal handling on the develop-ment, emotional reactivity and learning ability ofjuvenile nanday conures (Nandayus nenday). MSthesis. Purdue University.
Turner, C.A., M.C. Yang, and M.H. Lewis. 2002. Envi-ronmental enrichment: Effects on stereotyped be-havior and regional neuronal metabolic activity.Brain Research 938:15–21.
van Hoek, C.S., and C.E. King. 1997. Causation and in-fluence of environmental enrichment on featherpicking of the crimson-bellied conure (Pyrrhuraperlata perlata). Zoo Biology 16:161–172.
Winocur, G., and C.E. Greenwood. 1999. The effects ofhigh fat diets and environmental influences on cog-nitive performance in rats. Behavioral BrainResearch 101:153–161.
11 / Behavioral Development of Psittacine Companions: Neonates, Neophytes, and Fledglings 111

INTRODUCTION
Avian Companions and the Human-AnimalBond
Human relationships with companion animalshave existed for thousands of years (Lorenz1953). Artifacts from ancient civilizations attestto our long-standing relationships with birds(Vriends 1984). Our relationships with birds con-tinue to flourish. Today, there are millions of petbirds in the United States (Harris 1989).
Empirical studies have demonstrated conclu-sively that relationships with companion animalsprovide humans with valuable physical and psy-chosocial benefits (see, e.g., Mugford &M’Comisky, 1975; Beck & Katcher, 1989;Loughlin & Dowrick, 1993). Likewise, humanshave been shown to influence the behavior ofbirds. Imprinting studies conducted early in thiscentury by Lorenz are probably the most widelyknown experiments on the reactions of domesticavian species to interaction with human beings(Duncan 1992). Imprinting has been defined as apreprogrammed learned behavior (Alcock 1993).It allows the rapid establishment of a behavioralbond between an offspring and its parent. Lorenz(1952) found that baby Mallard ducks andGreylag goslings that he reared from hatchingformed an immediate attachment to him either bysight (Greylags) or vocal expression (Mallards)and would maintain close proximity to him ratherthan their mother, another adult female of thesame species, or another human.
Handling
More recently, studies have begun to focus on thelong-term effects of other types of early experi-
ence on the human-animal bond. It has beenshown that an animal’s fear of humans can be re-duced by habituation. One habituation techniqueis called “handling” (Duncan 1992). Although itis difficult to modify relationships between adultanimals and humans (Murphy & Duncan 1978),young animals are responsive to learning experi-ences that include the formation of social attach-ments during a sensitive or critical period (Jones& Waddington 1993). Neonatal, or postnatal,handling has been shown to produce psychophys-iological effects such as decreased fearfulness,decreased emotionality in open field tests (Den-enberg & Zarrow 1971), and decreased novelty-induced fear (Bodnoff et al. 1987), as well as in-creased resistance to stress (Levine 1957, 1962).Many studies on mammals have shown that han-dled animals, such as puppies (Scott & Fuller1965) and kittens (Karsh 1983), become closelyattached to humans. Likewise, several studieshave found similar effects in domestic poultryspecies (see, e.g., Jones & Faure 1981; Jones &Waddington 1993; Gross & Siegel 1979; Nicol1992). Most neonatal handling studies have beenconducted using mammals, primarily rats andmice. Levine initiated the classical infant-handling studies using rats to examine the effectsof early experience on neural development andfunction under stress conditions (Smythe et al.1994). He hypothesized from his results that“handling constitutes a stressful situation for theinfant organism and that early experience withstress results in a greater ability of the organismto adapt to psychological and physiological stressin adulthood” (1957, p. 405). Meaney and his col-leagues undertook studies to identify the mecha-
113
12Handler Attitude and Chick Development
Brenda Cramton

nism that enables this adaptation to occur. Theirfindings suggest that the hypothalamic-pituitary-adrenal (HPA) axis is altered by early experiencessuch as postnatal handling.
The HPA Axis
The HPA axis is highly responsive to stress (Selye1950). Any type of stress stimulates the neuronsin the paraventricularis nucleus of the hypothala-mus to secrete corticotropin-releasing hormone(CRH) into the portal system, which drains intothe anterior lobe of the pituitary. CRH results inan increase in the release of adrenocorticotropin(ACTH) from the pituitary. The elevated level ofACTH stimulates an increase in the output of glu-cocorticoids from the adrenals. Elevated concen-trations of glucocorticoids inhibit subsequent se-cretion of CRH from the hypothalamus andACTH from the pituitary (Meaney et al. 1988).This negative feedback system involves the inter-action of the hormones ACTH and CRH and a cy-tosolic glucocorticoid receptor in neural tissue(Meaney et al. 1985b).
Although glucocorticoids assist the organismunder stressful conditions by increasing the avail-ability of energy substrates, continued exposureto glucocorticoids may be detrimental to the or-ganism after the termination of the stressor.Elevated glucocorticoid levels may lead to sup-pression of anabolic processes, muscle atrophy,hypertension, hyperlipidemia, arterial disease,impairment of growth and tissue repair, and im-munosuppression (Meaney et al. 1996). There-fore, the capacity to effectively cope with thesestimuli is adaptive. Gross and Siegel (1979) foundthat male Shaver Starcross chickens that wereadapted to their handler produced more antibodyto sheep, horse, or human erythrocytes; had moreblood protein; gained more weight; and weremore resistant to a Mycoplasma gallisepticumchallenge than unadapted birds that were allowedminimal human contact.
Effect of Early Experience on the Maturationof the HPA Axis
In the rat, the pituitary-adrenal stress responseand the stress-related, negative feedback systemdo not mature until after birth; therefore, the de-velopment of these systems is under way duringthe period when an animal is exposed to early en-vironmental stimulation (Meaney et al. 1985b).
Meaney and his colleagues (1985a) suggest thatthe mechanism by which handling influences thedevelopment of the stress response involves theregulation of glucocorticoid receptor concentra-tions in the hippocampus and, perhaps, trans-cortin receptors in the pituitary. They found thathandled rats showed an increase in hippocampalglucocorticoid receptor concentrations and a de-crease in pituitary transcortin binding comparedto non-handled animals. Since transcortin recep-tors bind circulating corticosterone, it is not ableto inhibit the release of ACTH; therefore, highertranscortin levels in non-handled animals wouldlead to greater adrenocortical activity. The re-searchers hypothesize that early handling in-creases hippocampal glucocorticoid receptor con-centration in one of two ways: (1) by increasingthe number of receptor sites per cell, or (2) bystimulating postnatal neurogenesis in the hip-pocampus, which leads to an overall higher con-centration of receptor sites. Their work also sug-gests that thyroid hormones have some role in themediation of the development of glucocorticoidreceptor concentrations in the hippocampus be-cause their levels are known to be low from birthuntil about day 4, when they begin to increase,and peak at adult levels at the end of the secondweek of life; this pattern is identical to the devel-opmental increase in glucocorticoid receptor con-centrations in some brain regions (Meaney et al.1987). Also, it is known that thyroid hormonesare released during hypothermia, and handling isknown to result in a transient period of mild hy-pothermia (Mitchell et al. 1990).
Increases in serotonin activity may also medi-ate the developmental changes in hippocampalglucocorticoid receptors and influence the effectsof environmental events such as neonatal han-dling (Mitchell et al. 1990). Hippocampal con-centrations of serotonin increase over the first twoweeks of life and peak on day 14, which is simi-lar to the developmental changes in hippocampalglucocorticoid receptor binding (Mitchell et al.1990). Mitchell and his colleagues (1990) foundthat adult animals treated with a serotonin neuro-toxin (5,7-dihydroxytryptamine) in the first fewdays of life had reduced hippocampal glucocorti-coid receptor binding. In contrast, treatments thatincreased receptor binding capacity, such asneonatal handling or exogenous thyroid hormonetreatment, increased hippocampal serotonin turn-
114 Manual of Parrot Behavior

over. Furthermore, the effects of handling wereblocked when the serotonin receptor antagonist,ketanserin, was administered concurrently (seealso Smythe et al. 1994).
Postnatal Handling in Parrots
Since the interest in parrots as pets has grown inrecent years, many species have become threat-ened due to capture for the pet trade. Habitat de-struction and hunting for food have also con-tributed to the reduction in wild populations (Toft1993). Every effort, therefore, must be made toalleviate collection from wild populations.Captive breeding is an alternative that can be em-ployed to meet the demand for parrots for the pettrade market.
While many of the postnatal handling studieswith rats and mice involve minimal amounts ofhandling (such as simple removal from the cageand placement in a container), the traditional avi-culturalist’s approach to taming chicks is at theopposite end of the handling spectrum: newlyhatched chicks are permanently separated fromparent birds and hand-fed by humans until wean-ing. This form of hand-raising may, however, in-troduce several sources of error. Inappropriatediets and thermal environments as well as the as-piration of food into the lungs may increase mor-tality rates of hand-raised chicks (Davis &Millam 1997). Furthermore, hand-raised chicksmay lose their capacity to reproduce if they im-print on the humans that hand-rear them. Theseissues prompted Davis and Millam (1997) to askwhether a combination of hand-rearing andparent-rearing might alleviate some of these prob-lems while at the same time producing tame par-rots. A shared method of rearing tamed chickswould not only decrease the risk of chick mortal-ity but also significantly decrease the amount ofhuman labor necessary to produce tame birds.Davis and Millam demonstrated that tamed par-rots could be produced by short, regular periodsof handling by humans while parent birds pro-vided the primary care for the chicks. In theirstudy, chicks were handled daily for 15 to 30 min-utes from day 12 until fledging. The researchersfound that handled chicks scored high on a seriesof tameness tests. A follow-up study (Davis &Millam 1997) revealed that chicks that were han-dled later in life (handling began at 35 days ofage) were more tame than the early handled birds
and the experimenters judged the birds to be astame as hand-raised birds.
Collette et al. (2000) confirmed and extendedDavis and Millam’s findings. They found thatpostnatally handled chicks were more tame on allbehavioral tests than non-handled chicks. Theyalso examined the influence of taming on immunestatus. After fledging, birds were physically re-strained for a period of ten minutes. For handledbirds, the restraint consisted only of perching ona human hand; however, non-handled birds wouldnot perch on a human hand so they were insteadwrapped in a towel. After the period of restraint,immune status was assessed by responses to (1)humoral response to a killed Newcastle diseasevirus; (2) serum corticosterone levels; (3) het-erophil:lymphocyte ratios; and (4) delayed-typehypersensitivity (DTH) test to a foreign protein(phytohemagglutinin). DTH responses are antigen-specific, cell-mediated immune reactions (Dhab-har 1998). Experiments have shown that acutestress administered immediately before the in-troduction of an antigenic challenge signifi-cantly enhances a cutaneous DTH response,whereas chronic stress suppresses cutaneousDTH (Dhabhar 1998). Collette found that han-dled chicks had a significantly greater humoralresponse to Newcastle disease virus and a signif-icantly lower DTH response to a foreign proteinthan did non-handled chicks. According to cur-rent immunologic theory, antibody response is in-versely related to DTH response (Hassig et al.1996). Corticosterone levels were also lower inhandled chicks, but the difference between thetwo groups was not statistically significant.Handled and non-handled chicks were indistin-guishable with respect to heterophil:lymphocyteratios.
Although Davis and Millam (1997) andCollette et al. (2000) found that postnatal han-dling resulted in tame chicks, both studies re-ported a considerable variation in degree of tame-ness. Since a high degree of tameness may be val-ued by individuals seeking parrots as companionanimals, and tameness may reduce the degree ofstress parrots experience in captive environments,it is important to consider what factor, or factors,may have led to the observed degrees of tamenessin these studies.
Handler personality is one possibility. Sea-brook (1972) examined the influence of the
12 / Handler Attitude and Chick Development 115

herdsman’s personality on milk yields of dairycows. He found that “good” herders’ cows pro-duced as much as 20% more milk than cowsunder the care of other herders at similar facilitiesunder the same ownership. Seabrook defined agood herder as one with “knowledge of the be-haviour of the individual cows in the herd and theability to notice deviations from normal behav-iour” (p. 376). He found that patient herdsmenthat behaved consistently and showed considera-tion for their cows’ needs had cows with the bestmilk yield. Other traits of the good herders in-cluded confidence, self-reliance, and introvertedpersonalities. Other studies have found that pigsthat are fearful of humans after intentional aver-sive handling experiences had depressed growthrates and reproductive performance (see, e.g.,Hemsworth et al. 1981, 1986). This is probablydue to a chronic stress response, since even afteraversive handling ceased, and humans were nolonger present, pigs continued to exhibit elevatedlevels of plasma-free corticosteroids.
Empathy
Although it is difficult to predict what factor, orfactors, of human personality may influence ananimal’s fear response to a human, the ability toempathize might be one important trait. Peoplewho experience sympathy or empathy “are morelikely to help, comfort, or share with other peopleor animals” (Eisenberg 1988, p. 16). Conversely,cruelty toward other people or animals is consid-ered, by mental health professionals, to be the re-sult of a distortion in the development of empathy(Ascione 1993).
Definitions of empathy are varied but similar.For example, it has been defined as “the capacityto feel what another is feeling” (Zahn-Waxler etal. 1985, p. 22), or “the power of understandingand imaginatively entering into another’s feel-ings” (Fox 1985, p. 61), or “a vicariously inducedemotional reaction based on the apprehension ofanother’s state or condition that is similar to theother’s emotional state or consistent with theother’s situation” (Eisenberg 1988, p. 15).
Darwin described the empathy that developedbetween himself and his dog as an interactionwhereby an animal and a human intuit the other’sstate from behavioral cues (cited in Buck andGinsburg 1997). In animal handling, an empathichandler presumably makes accurate assessments
of an animal’s fearfulness or timidity in certainsituations. The empathic handler would thenmake adjustments in his or her behavior or the en-vironment in order to lessen the animal’s experi-ence of fear or apprehension. In contrast, a han-dler that fails to take an animal’s perspective andto recognize and correctly interpret the animal’scues may handle the animal inappropriately in re-lation to its internal state; if handled regularly bythe same individual, the animal may become con-ditioned to fear humans and to experience them asstressful stimuli.
Purpose
This study was undertaken to address an aspect ofemerging societal concern: human-animal inter-action. The purpose of this study was to examinethe relationship between the level of empathy ofthe handler, the attitude of the handler towardpets, and the degree of tameness and immunecompetence in handled parrots. It was hypothe-sized that parrots handled by more empathic han-dlers would exhibit a higher level of tameness andan increased ability to cope with stress than birdshandled by less empathic handlers. Since geneticfactors and quality of parrot parenting may alsoplay a role in tameness and immune competence,these variables were also examined.
A better understanding of the human factorsthat increase tameness and the ability to respondeffectively to stress is expected to (1) benefit avi-culturists who wish to rear highly marketable par-rots for the pet bird trade, (2) contribute to theoverall welfare of parrots that must co-exist withhumans in their captive environments, and (3) in-crease the knowledge of the effects that peoplehave on the behavior of captive parrots.
PARTICIPANTS, ANIMALS, MATERIALS,AND METHODS
Human Participants
In order to examine the effect of handler empathyon parrot tameness, volunteer handlers thatscored either high or low on the construct of em-pathy were solicited from the student populationof the University of California, Davis. Potentialparticipants were contacted by U.S. mail, elec-tronic mail, posters, and classroom solicitations.In order to avoid any confounding sex differ-ences, only female students were selected to par-
116 Manual of Parrot Behavior

ticipate in the study. The average age of the par-ticipants in this study was 21 years.
Procedure
Students interested in participating in the han-dling study were asked to call or e-mail the exper-imenter to request a personality test. Ninety-twostudents requested personality tests. Sixty-eightstudents returned completed personality tests.Seven students were selected to participate in thehandling study. The number of participants se-lected was based on the number of chicks avail-able to be handled. The ratio of handlers to chickswas 1:3.
Personality test scores were not revealed to anystudents. As compensation for the time requiredto complete the questionnaire, each student whoreturned a questionnaire was provided with a listof occupations engaged in by individuals withprofiles similar to their own. These occupationalprofiles are often used by professionals in careercounseling.
Selected participants were required to com-plete a health surveillance questionnaire and todiscuss their responses with a nurse at EmployeeHealth Services, University of California, Davis.Participants were also required by the Office ofthe Campus Veterinarian, University of Califor-nia, Davis, to obtain tuberculosis clearance (via atuberculin skin test) and to have a serum sampledrawn for archival purposes. Both procedureswere conducted by personnel at Employee HealthServices. There was no cost to the participants forthese medical procedures.
Instruments
16PFThe Sixteen Personality Factor Questionnaire(Institute for Personality and Ability Testing, Inc.,Champaign, Illinois) is a psychological assess-ment instrument designed by Raymond B. Cattell.In the 1940s, Cattell used factor analysis to re-duce 45 categories of words, commonly used inthe English language to describe human personal-ity, to 15 dimensions of personality (Karson &O’Dell 1976). Later, three less replicable factorswere discarded and four factors that were consid-ered important were added (Karson & O’Dell1976). Currently, the test measures 16 normal di-mensions of adult personality: warmth, reasoningability, emotional stability, dominance, impulsiv-
ity, conformity, social boldness, sensitivity, vigi-lance, imagination, privateness, apprehension,openness to change, self-reliance, perfectionism,and tension. Since the first-order factors are cor-related, or oblique factors, Cattell was able toconduct a second factor analysis and arrive atsecond-order factors. These factors summarizethe relationships found among the 16 first-orderfactors. The test measures five second-order fac-tors: extraversion, anxiety, tough-mindedness, in-dependence, and self-control. The 16PF consistsof a total of 187 questions.
The 16PF was employed in this study to meas-ure the level of one second-order personality fac-tor: tough-mindedness, a measure of empathy. Anindividual who scores high on tough-mindednesshas low scores on four first-order factors:warmth, emotional sensitivity, imagination, andopenness to change (Karson and O’Dell 1976).Karson and O’Dell (1976) explain that these peo-ple are much less likely to be controlled by theirfeelings than by their intellect. In contrast, a lowtough-mindedness score is associated with highscores on the first-order factors of warmth, emo-tional sensitivity, imagination, and openness tochange.
All tests were hand-scored. Females whoscored high (7.7–10) or low (1.0–2.0) on tough-mindedness were selected to handle parrot chicks.Due to the fact that very few female test-takersscored on the high end of the tough-mindednessscale, a larger point spread was allowed in thehigh tough-mindedness handlers’ scores than inthe low tough-mindedness handlers’ scores (2.3versus 1.0 point spread).
PET ATTITUDE SCALE
Prior to the start of handling, the Pet AttitudeScale (PAS; Templer et al. 1981; see Appendix12A) was administered to participants to measurefavorableness of attitudes toward pets. The PASwas selected because it is one of few publishedscales with reliability information: Cronbach’salpha coefficient is 0.93, and two-week test-reteststability is 0.92 (Lago et al. 1988). The PAS con-sists of 18 7-point Likert format questions; there-fore, 126 points are possible. A higher score rep-resents a more favorable attitude toward pets. Theitems on the PAS represent three factorially de-rived scales: love and interaction, pets in thehome, and joy of pet ownership.
12 / Handler Attitude and Chick Development 117

Animals and Housing
The animals in this study were the offspring ofwild-caught Orange-winged Amazon Parrots(Amazona amazonica). All chicks hatched in theanimal colony at the University of California,Davis. Sixteen breeding pairs were housed in twoadjacent rooms, one pair per cage, eight pairs perroom. Pairs were housed in 1 m x 1 m x 2 msuspended, wire welded cages. Each cage hadtwo 1 m x 4 cm x 8.5 cm wooden perchesmounted with metal brackets to the 2 m lengthsof the cage. One perch was mounted near thefront of the cage (the cage door end) and theother near the rear (the nest box end) of the cage.Birds were maintained in accordance with theUniversity of California, Davis, Animal Use andCare Provisions.
BREEDING CONDITIONS
Sixteen pairs of Orange-winged Amazons werestimulated to breed by exposure to long day-lengths and the provision of nest boxes. Nestboxes were presented to breeding pairs on May21, 1997. Stainless steel sheet metal “grandfatherclock” type (40 cm x 38 cm x 76 cm) nest boxeswere installed at the rear of each cage on its exte-rior surface. A 14 cm x 18 cm opening in the wireof the cage provided access to the nest box hole.The nest box hole was surrounded with a woodeninsert with a 9 cm diameter hole in it. The woodeninsert enabled the birds to enlarge the hole and, ina sense, chew through it into the nest box. Thenest box floors were lined with 8–12 cm of auto-claved, premium-grade pine shavings.
On the same day that nest boxes were pre-sented, light schedules began to be increased from10 hours light and 14 hours dark (10L:14D) to14L:10D. Light was increased by 30-minute in-crements each day (15 minutes in the A.M. and 15minutes in the P.M.) over a period of eight days.
Diet was changed from Roudybush mainte-nance pellets (provided during non-breeding peri-ods) to Roudybush breeder pellets (RoudybushInc., Sacramento, California) during breeding,laying, and rearing stages. Water was available adlibitum from nipple waterers.
CHICK FOSTERING
Due to the cannibalistic behavior of two breedingpairs (sire band #693 & dam band #458, and sire#187 & dam #338) in previous breeding seasons,
all fertile eggs were removed from the nest boxesof these pairs. The eggs were placed either withanother suitable breeding pair during the late in-cubation stage or in an incubator (RX2TT, serialno. MM, cat. no. 910-063, Lyon Electric Co.,Chula Vista, California) during the early incuba-tion stage and then transferred to another breed-ing pair during the late incubation stage.Incubator conditions were as follows: (1) averagewet and dry bulb temperatures were 87 and 99 de-grees Fahrenheit (30.6 and 37.2 Celsius), respec-tively; (2) relative humidity ranged from 60 to65%; and (3) eggs were rotated by automaticturner once per hour.
Handling Procedure
Handling of chicks began between 28 and 35 dayspost-hatch. Each handler was assigned threechicks to handle for the duration of the study.Clutchmates were not assigned to the same han-dler. Within each clutch, chicks were randomly as-signed to either a high or low tough-mindednesshandler. When clutches consisted of an even num-ber of chicks, equal numbers of chicks were as-signed to high and low tough-mindedness han-dlers. A total of 20 chicks were included in thehandling study: 11 chicks were assigned to low-empathy handlers and nine chicks were assignedto high-empathy handlers.
To allow for freedom of interaction betweenhandlers and chicks, handlers were provided onlybasic instructions about the handling procedure.They were shown how to isolate the parent birdsfrom the nest box by insertion of a metal plate be-tween the lower one-third and upper two-thirds ofthe grandfather style next box, and how to removethe chicks from the nest box and place them in ashallow, towel-lined plastic tub. The handlerswere instructed to handle each chick individuallyand to interact with the chick in any way theychose. Each chick was handled three times perweek and handling sessions lasted for a period of20 minutes. Handling continued until the time oftesting.
All chicks were also handled briefly by theprincipal investigator during leg band placement,weekly weight measurements, and twice-weeklynest box cleaning. During nest box cleaning, thechicks in a clutch were placed in a towel-lined,plastic tub while the soiled pine shavings were re-placed with clean shavings.
118 Manual of Parrot Behavior

Tameness Tests
CHICKS
After fledging (approximately 56 days post-hatch), a series of tameness measures was madeon each chick (see Chick Tameness Score Sheet,Appendix 12B). The tests consisted of both be-havioral and physiological measures. The behav-ioral measures included (1) response to extendedfinger/hand; (2) response to touch on head, cheek,or back; (3) response to a food offering (Cheeriosbrand breakfast cereal, General Mills Inc.,Minneapolis, Minnesota); (4) response to a novelobject (a 4 cm length of plastic drinking straw);and (5) proximity seeking (i.e., did the bird seekcloseness with a human or with another parrot ofthe same species?).
Two indirect measures of fear were made: res-pirations per minute and period of latency to defe-cation. These measures were selected becausethey were (1) non-invasive and (2) viewable at adistance by a second test administrator. Respira-tions per minute were measured after the bird wasremoved from the home cage and allowed to set-tle on the test perch for 60 seconds in full view ofthe test administrators. The latency to defecationtest was limited to 300 seconds. A score of 300was assigned to the bird if the latency period ex-ceeded 300 seconds.
Five tameness test trials were made on eachchick. The first trial was carried out by the bird’shandler. Two succeeding trials were conducted byeach of two individuals that the bird had never in-teracted with previously (i.e., a “stranger”). Dur-ing each trial, the bird’s behavior was recorded bytwo individuals: the person administering thetests to the bird and one additional individual. Thelatter also made timed measures of respirationrate and latency to defecation using a stopwatch.Test administrators were blind. When the two in-dividuals recording the bird’s behaviors disagreedabout a behavioral tameness measure, the lower(i.e., less tame) score was awarded to the bird.The five scores from each of the five trials wereaveraged to obtain one overall tameness score.Likewise, the five measures of respiration rateand latency to defecation were averaged.
PARENT BIRDS
Behavioral and physiological measures of tame-ness were also made on the wild-caught parentbirds in order to assess a possible genetic compo-
nent of fearfulness or tameness. The chick tame-ness tests were modified for use with the adultwild birds (see Appendix 12C). For example,most of the wild birds would not approach ahuman; therefore, a measure was made ofwhether or not the bird withdrew to the more dis-tant perch when a human stood in front of thecage. Also, responses to the opening of the cagedoor and the extension of a hand into the cagewere recorded. Respiration rate and latency todefecation were measured while the experimenterstood in front of the cage. Timing of latency todefecation began immediately after the experi-menter stepped in front of the cage. As with thechicks, the latency test was limited to 300 sec-onds: a score of 300 was assigned to the birdwhen the latency period exceeded 300 seconds.Respiration rate was measured after the experi-menter had been in full view of the birds for 60seconds.
Quality of Parenting Measures
Two variables were selected to represent meas-ures of quality of parenting: (1) chick rate ofweight gain, and (2) chick plumage condition.Each chick was weighed every seven days begin-ning on day 7 post-hatch. Weekly rate of absoluteweight gain over the first four weeks of life wasaveraged.
Since some breeding pairs had a history inprior breeding seasons of pulling out their chicks’feathers, this behavior was also recorded as ameasure of quality of parenting. Chick plumagecondition was scored either as feather-plucked ornon-plucked.
Immunoresponse Test
Immune status was assessed by response to a DTHtest to a foreign protein, phytohemagglutinin-M(USB Specialty Biochemicals, Cleveland, Ohio).This test was selected because Collette et al.(2000) found a significant DTH response in han-dled versus non-handled Amazon chicks. Follow-ing the completion of tameness measures, eachchick was placed in an animal carrier (26 cm x 28cm x 40 cm) to produce a condition of novel envi-ronment and mild immobilization stress. Chickswere retained in the carrier for a period of fiveminutes. Placement in an animal carrier was se-lected as a stressor because most captive animalswill be transported in carriers during their life-
12 / Handler Attitude and Chick Development 119

times; therefore, it allowed the inducement of astress that would occur under normal conditions incaptivity.
Twenty-four hours after initial placement in the carrier (at 7 A.M.), each chick was again re-tained in the carrier for five minutes. After this,wing web thickness was measured with a mi-crometer and marked with a black marker dot.Phytohemagglutinin-M (0.25 mg dissolved in0.05 ml sterile saline) was then injected subcuta-neously to the marker dot in the wing web. After12 hours (7 P.M.), the area was measured againfor change in tissue thickness.
Statistics
SAS (1990) was employed to conduct all statisti-cal analyses. The procedure used was PROCREG, version 6.12. Stepwise regression analysiswas utilized to analyze the data. Regressionanalysis allowed the development of an equationthat enabled the prediction of one variable fromthe knowledge of another variable.
At each stage in the stepwise regression proce-dure, the algorithm began with the calculation ofF-statistics for each of the predictors that werecurrently selected for the regression model. If apredictor did not meet a specified significancelevel (0.100 in this study), it was removed fromthe equation. Next, an attempt was made to add anew predictor by calculating an F-statistic foreach variable not currently in the equation. Ateach step in the model-building procedure, atmost, one term was removed from or added to themodel. If one or more terms were eligible to beremoved, the one with the largest significancelevel (p-value) was removed. If one or more termswere eligible to be added to the model, the onewith the smallest significance level was added.The stepwise regression procedure stopped whenpredictors were no longer added or deleted.
The procedure used to analyze the data in thisstudy differed in that handler tough-mindednesswas forced to be included in all regression mod-els. This was done because the purpose of thestudy was to determine whether handler tough-mindedness had an impact on tameness; there-fore, p-values were being sought regardless ofwhether they were or were not significant. Otherpredictors that were examined included handlerattitude toward pets, whether or not chicks werefeather-plucked, average rate of weight gain over
the first four weeks of life, and behavioral andphysiological measures of tameness in parentbirds.
RESULTS
Reproductive Response
Fourteen breeding pairs produced a total of 77eggs. At least 44 of the 77 eggs were fertile.Thirty-four eggs hatched (hatch rate = ~77%).The average number of chicks reared by a breed-ing pair was 2.3 chicks. This number includes fos-ter chicks (see the following section on chick fos-tering). Six fertile eggs suffered damage inflictedby the parent birds: one female (#352) fracturedher five eggs by pecking them when the nest boxdoor was opened; another pair (#369 and #172)punctured the shell of one of their eggs with a toe-nail. Although the holes and fractures werepatched with tape, all of the damaged eggs failedto hatch.
Candling revealed that 27 eggs were infertile.The fertility status of six other artificially incu-bated eggs was indeterminable; either the em-bryos died early or the eggs were infertile.
Twelve chicks that hatched later died. Fivehatchlings and one three-month-old chick (band#97-29) were killed by parent birds (sire #540 &dam #389, sire #436 & dam #602, and sire #183& dam #374). Three hatchlings died due toparental neglect (sire #519 & dam #350, and sire#513 & dam #352). One six-week-old chick diedfrom a respiratory infection (#97-22). The causeof death of two remaining hatchlings was un-known.
Six breeding pairs that experienced the loss oftheir first clutch produced a second clutch. Aver-age size of second clutches was four eggs. Onlyone pair (sire #436 & dam #602) had a survivingchick from the first clutch. One pair (sire #433 &dam #504) failed to produce any fertile eggs ineither the first or second clutch. Two pairs hadthe eggs in their first clutches removed to an in-cubator due a cannibalistic history by the pair(sire #693 & dam #458, and sire #187 & dam#338). One pair (sire #540 & dam #389) killedall of the chicks in the first clutch. Another pair(sire #513 & dam #352) fractured all of the eggsin the first clutch and, as a result, the embryosdied.
One breeding pair failed to produce any eggs,
120 Manual of Parrot Behavior

although a bowl was created in the nest box shav-ings. Another pair, later determined by geneticsexing to consist of two females, laid a total offive infertile eggs. Three other pairs producedonly infertile eggs.
Of the surviving 23 chicks, 11 were assigned tolow-empathy handlers and nine were assigned tothe high-empathy handlers. Three chicks (#97-22,#97-23, and #97-24) were excluded from thestudy because they suffered from a chronic respi-ratory infection that required administration oftwice-daily injectible and oral antibiotics; there-fore, it was believed that tameness outcomeswould be confounded by the frequent and aver-sive handling of these chicks.
Chick Fostering
Due to the cannibalistic behavior of two breedingpairs in previous breeding seasons (sire #187 &dam #338, and sire #693 & dam #458), their eggswere fostered to other possibly more suitablepairs. A third pair (sire #540 & dam #389) killedall of the chicks in their first clutch and the firstchick in their second clutch during the currentbreeding season. The female of this pair had beentreated for a respiratory infection during the incu-bation of her first clutch. The three remainingeggs in her second clutch were fostered to otherpairs.
Four pairs were selected to foster eggs fromthe late incubation stage. During early incuba-tion, some eggs remained with the biological par-ents and others were artificially incubated.Incubation in an incubator served a dual purpose:it provided an environment for the growth anddevelopment of embryos, and it also allowed thedeveloping chicks to be photographed during thevarious stages of development without any dis-turbance to adult birds. The photographs of theembryos were used in another study. Four of tenartificially incubated eggs hatched. Two of thesechicks were successfully reared by foster parentbirds.
A total of 13 fertile eggs were fostered (four ar-tificially incubated and nine naturally incubated).Eggs were fostered one week prior to the ex-pected date of hatch. Ten chicks hatched and eightchicks survived. Two pairs successfully com-pleted the incubation of foster eggs: chickshatched and were successfully reared. One ofthese pairs (sire #368 & dam #407) completed the
incubation of two eggs from the incubator. Theother pair (sire #495 & dam #545) successfullycompleted the late incubation stage of three eggsthat were previously incubated by the biologicaldam (#458). Neither foster pair had produced anyfertile eggs of their own. Their infertile eggs werereplaced with an equal number of fertile fostereggs.
Two other pairs successfully incubated fostereggs and chicks hatched; however, some chickswere killed or neglected (which resulted in death).One of these foster pairs (#379 & #646) producedno fertile eggs because both birds were later de-termined by genetic sexing to be females. Thefirst hatchling in their care was found dead on day2 post-hatch; therefore, the two remaining fostereggs were fostered to another pair (sire #183 &dam #374). The latter pair completed the incuba-tion of the eggs and successfully reared the twochicks for three months before killing the youngerchick. Another foster pair (sire #513 & dam #352)produced one hatchling of their own, but it wasfound dead on the same day that the first fosterchick hatched. A second foster hatchling in theircare survived only one week.
One chick hatched in the incubator five daysprior to the expected date of hatch. This chick wasfostered to a pair that was successfully raisingfour of their own chicks (sire #317 & dam #111).The pair’s youngest chick was close in age to thefostered chick. The foster chick failed to thriveand expired after seven days. Whether this wasdue to a biological anomaly that may have beenassociated with the chick’s early hatching orwhether it was due to the possibly overworkedparents’ ability to provide for one additional chickis unknown.
Tameness
EMPATHY
The global factor “tough-mindedness” was uti-lized as a measure of handler empathy. The tough-mindedness scale ranges from 1 to 10. A lowertough-mindedness score is associated with ahigher degree of empathy. Handler scores were1.6, 1.9, 2.0, 7.7 (two handlers with this score),8.7, and 9.3.
According to the stepwise regression models,handler tough-mindedness significantly predictedboth physiological measures of tameness. Tough-mindedness predicted chick respiration rate,
12 / Handler Attitude and Chick Development 121

p = 0.002, and inversely predicted latency to defe-cation, p = 0.070 (Figures 12.1 and 12.2, respec-tively).
In contrast, handler tough-mindedness did notsignificantly predict the behavioral measure oftameness, p = 0.947. Birds handled either byhigh- or low-empathy handlers did not demon-strate appreciable differences in behavioral tame-ness. They perched on experimenters’ hands, per-mitted human touch, and accepted food items andnovel objects from experimenters.
ATTITUDE
Templer’s PAS was employed to measure handlerattitude toward pets. A higher score is associatedwith a more positive attitude toward pets. A totalof 126 points are possible. Pet attitude scores ofthe handlers in this study were 108, 111, 112,113, 117 (two handlers with this score), and 123.
According to the stepwise regression models,
handler attitude did not significantly predict be-havioral tameness, p = 0.190, latency to defeca-tion, p = 0.500, or respiration rate, p = 0.883. Thecorrelation between pet attitude and tough-mindedness was weak, r = 0.101.
PARENTING VARIABLES
Quality of parenting was assessed by measures oftwo variables: (1) chick average rate of weeklyweight gain over the first four weeks of life, and(2) chick plumage condition. Three breedingpairs (sire #513 & dam #352, sire #519 & dam#350, and sire #436 & dam #602) were responsi-ble for feather plucking 25% of the chicks in thestudy.
A feather-plucked condition significantly pre-dicted both physiological measures of tameness:respiration rate, p = 0.005, and latency to defeca-tion, p = 0.048 (Figures 12.3 and 12.4, respec-tively). However, according to the sums of
122 Manual of Parrot Behavior
Figure 12.1. Chicks handled by low-empathyhandlers had elevated respiration rates (p =0.002 [coefficient = 1.282] by stepwise linearregression). Plots depict median (line), 25th, 75th percentiles (box), and 5th, 95th percentiles(circles).
Figure 12.2. Chicks handled by low-empathyhandlers had shorter latencies to defecation (p = 0.069 [coefficient = �9.713] by stepwiselinear regression) (plot key as in Figure 12.1).

squares, handler tough-mindedness was a betterpredictor of respiration rate than was a feather-plucked condition (sum of squares = 329.7 versus238.5, respectively). In contrast, a feather-plucked condition was a somewhat better predic-tor of latency to defecation than was handlertough-mindedness (sum of squares = 22,667 ver-sus 18,632, respectively).
In contrast to the effect of handler tough-mindedness on behavioral tameness, a feather-plucked condition significantly and inversely pre-dicted behavioral tameness, p = 0.042 (Figure 12.5).
Rate of weight gain did not significantly pre-dict any measure of tameness. Mean weekly chickweight gain over the first four weeks of life was84.55 grams/week ± 2.78 grams/week (mean ±SE).
PARENTAL TAMENESS
Paternal respiration rate inversely and signifi-cantly predicted latency to defecation in chicks, p= 0.013 (Figure 12.6). It was a more significantpredictor of latency to defecation than eitherfeather-plucked condition or handler tough-mindedness (sum of squares = 38,950 versus22,667 and 18,632, respectively).
Maternal behavioral tameness inversely andsignificantly predicted chick tameness, p = 0.007(Figure 12.7). Interestingly, the correlation be-tween foster chick tameness and biological damtameness was greater than the correlation be-tween foster chick tameness and foster dam tame-ness, r = �0.400 versus r = �0.249, respectively.
Cell-Mediated Immunity
According to the stepwise regression model, han-dler empathy did not predict immune response, p= 0.761. Birds handled either by high- or low-empathy handlers did not demonstrate any dis-
12 / Handler Attitude and Chick Development 123
Figure 12.3. Feather-picked chicks had higherrespiration rates than non-picked chicks (p =0.005 [coefficient = �8.06] by stepwise linearregression) (plot key as in Figure 12.1).
Figure 12.4. Feather-picked chicks had shorterlatencies to defecation than non-picked chicks(p = 0.048 [coefficient = �78.5] by stepwiselinear regression) (plot key as in Figure 12.1).

cernible difference in change in wing thicknessafter the administration of PHA-M. Mean wingthickness change in birds handled by low-empathyhandlers was 0.592 mm ±0.169 mm (n = 11) ver-sus 0.579 mm ± 0.261 mm (n = 7) in birds handledby empathic handlers. Two birds handled by em-pathic handlers were omitted from the computa-tions because one chick died before the test wasconducted and an improper measuring techniqueled to an erroneous measure on a second chick.
DISCUSSION
Tameness
HANDLER EMPATHY
The results of this study support, in part, the hy-pothesis that degree of handler empathy affectsparrot chick tameness. Level of handler empathywas found to significantly influence both physio-logical measures of fear examined in this study:
respiration rate and period of latency to defeca-tion. In the presence of humans, chicks that hadbeen handled by individuals with lower levels ofempathy exhibited higher rates of respiration andshorter periods of latency to defecation thanchicks that had been handled by high-empathyhandlers.
The surprising finding in this study is that birdshandled either by high- or low-empathy handlersexhibited overtly tame behaviors such as a will-ingness to perch on a human hand and permithuman touch. However, although birds handledby low-empathy handlers exhibited tame behav-iors, the results of this study suggest that thesebirds continued to experience fear on a physiolog-ical level when in the presence of humans. Thisoutcome is consistent with Hennessy andLevine’s (1979) finding that “some physiologicalresponses may habituate more slowly than overtbehavioral reactions” (qtd. in Levine et al. 1989,p. 344). Habituation is the process by which newstimuli or situations are compared with represen-tations in the CNS of previous events (Sokolov,
124 Manual of Parrot Behavior
Figure 12.5. Feather-picked chicks were lesstame than non-picked chicks (p = 0.042 [coeffi-cient = 1.13] by stepwise linear regression) (plotkey as in Figure 12.1).
Figure 12.6. Paternal respiration rate inverselypredicted chick latency to defecation (p =0.013 [coefficient = �6.92] by stepwise linearregression).
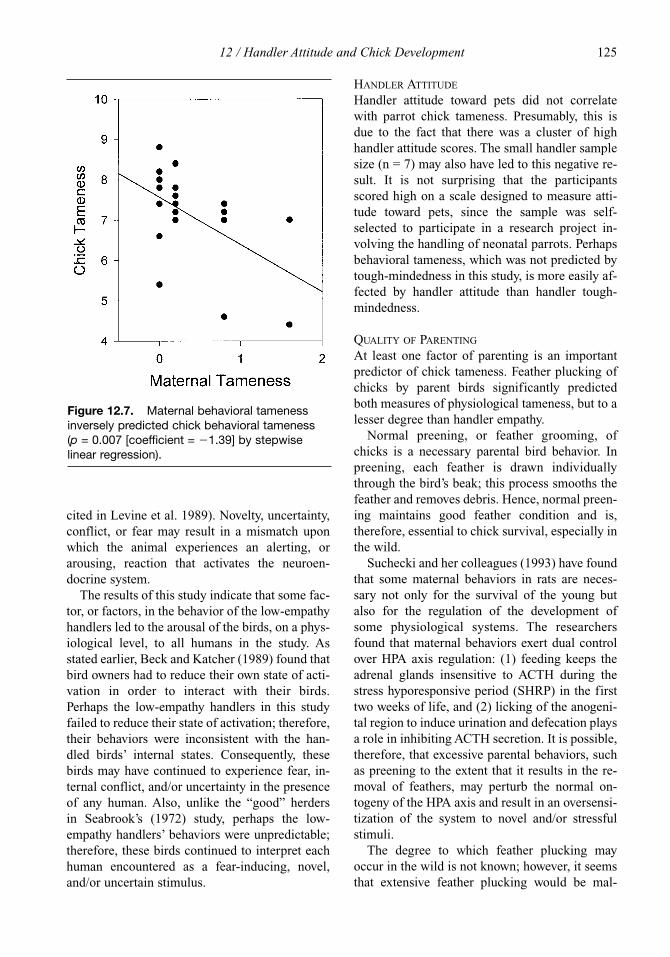
cited in Levine et al. 1989). Novelty, uncertainty,conflict, or fear may result in a mismatch uponwhich the animal experiences an alerting, orarousing, reaction that activates the neuroen-docrine system.
The results of this study indicate that some fac-tor, or factors, in the behavior of the low-empathyhandlers led to the arousal of the birds, on a phys-iological level, to all humans in the study. Asstated earlier, Beck and Katcher (1989) found thatbird owners had to reduce their own state of acti-vation in order to interact with their birds.Perhaps the low-empathy handlers in this studyfailed to reduce their state of activation; therefore,their behaviors were inconsistent with the han-dled birds’ internal states. Consequently, thesebirds may have continued to experience fear, in-ternal conflict, and/or uncertainty in the presenceof any human. Also, unlike the “good” herders in Seabrook’s (1972) study, perhaps the low-empathy handlers’ behaviors were unpredictable;therefore, these birds continued to interpret eachhuman encountered as a fear-inducing, novel,and/or uncertain stimulus.
HANDLER ATTITUDE
Handler attitude toward pets did not correlatewith parrot chick tameness. Presumably, this isdue to the fact that there was a cluster of highhandler attitude scores. The small handler samplesize (n = 7) may also have led to this negative re-sult. It is not surprising that the participantsscored high on a scale designed to measure atti-tude toward pets, since the sample was self-selected to participate in a research project in-volving the handling of neonatal parrots. Perhapsbehavioral tameness, which was not predicted bytough-mindedness in this study, is more easily af-fected by handler attitude than handler tough-mindedness.
QUALITY OF PARENTING
At least one factor of parenting is an importantpredictor of chick tameness. Feather plucking ofchicks by parent birds significantly predictedboth measures of physiological tameness, but to alesser degree than handler empathy.
Normal preening, or feather grooming, ofchicks is a necessary parental bird behavior. Inpreening, each feather is drawn individuallythrough the bird’s beak; this process smooths thefeather and removes debris. Hence, normal preen-ing maintains good feather condition and is,therefore, essential to chick survival, especially inthe wild.
Suchecki and her colleagues (1993) have foundthat some maternal behaviors in rats are neces-sary not only for the survival of the young butalso for the regulation of the development ofsome physiological systems. The researchersfound that maternal behaviors exert dual controlover HPA axis regulation: (1) feeding keeps theadrenal glands insensitive to ACTH during thestress hyporesponsive period (SHRP) in the firsttwo weeks of life, and (2) licking of the anogeni-tal region to induce urination and defecation playsa role in inhibiting ACTH secretion. It is possible,therefore, that excessive parental behaviors, suchas preening to the extent that it results in the re-moval of feathers, may perturb the normal on-togeny of the HPA axis and result in an oversensi-tization of the system to novel and/or stressfulstimuli.
The degree to which feather plucking mayoccur in the wild is not known; however, it seemsthat extensive feather plucking would be mal-
12 / Handler Attitude and Chick Development 125
Figure 12.7. Maternal behavioral tamenessinversely predicted chick behavioral tameness (p = 0.007 [coefficient = �1.39] by stepwiselinear regression).

adaptive and selected against. The basis of featherplucking or “overgrooming” in captivity is notunderstood, but repeated human handling may beone contributing factor. It is interesting that thechicks in one feather-plucked clutch (not includedin the study) required excessive, and intrinsicallyungentle, human handling for treatment of achronic respiratory infection. The chicks werefrequently returned to the nest box or home cagewith their plumage in disarray. If parental groom-ing of chicks is a response to soiled and/ormalaligned feathers, overgrooming could con-ceivably develop from repeated human handling.
It is not surprising that parenting factors influ-ence tameness. Since, in this study, chicks spentmore than 99% of their time in the care of theirparents, the opportunity for chick tameness to beinfluenced by parental factors was great. It islikely that numerous other parenting factors existthat may also affect chick tameness. The potentialexists to select breeding pairs that exhibit specificparenting qualities that increase chick tameness.Work to identify important parenting variables hasrecently begun with the analysis of videotapes,made during the course of the present study, ofparent and chick interaction in the nest box.
PARENTAL TAMENESS
One counterintuitive finding from this study isthat maternal behavioral tameness inversely andsignificantly predicted chick behavioral tame-ness. This result may be due to either post-hatchenvironmental factors or genetic factors or somecombination of the two. Perhaps an as-yet-unidentified difference in the maternal behaviorsof more and less tame mothers accounts for theinverse relationship of maternal and chick behav-ioral tameness. The results indicate that there maybe a genetic component, since foster chick tame-ness was more strongly correlated with biologicaldam tameness than with foster dam tameness.
Paternal respiration rate also significantly andinversely predicted chick latency to defecation:male parents with low respiration rates in the pres-ence of humans had chicks with long periods oflatency to defecation when humans were present.This outcome suggests that there may also be a ge-netic component to fearfulness. Taken together,these results suggest that chicks may inherit verydifferent factors of tameness from each parent.
Immune Status
Neither degree of handler empathy nor attitudetoward pets significantly predicted cell-mediatedresponse to a foreign protein challenge. Chickshandled either by high- or low-empathy handlersshowed similar changes in wing thickness. Asstated earlier, Collette et al. (2000) found thathandled parrot chicks had a lower cell-mediatedresponse and a higher antibody response to aNewcastle disease virus challenge than did non-handled chicks. This inverse relationship betweenthe two immune responses would be predicted ifthe birds had experienced handling as a stressfulstimulus (Hassig et al. 1996). The results of thepresent study support Collette’s finding thathuman handling is interpreted as stressful and ac-tivates the neuroendocrine system; however, theresults here show that handling and human pres-ence are interpreted as stressful only when post-natal handling has been carried out by a humanhandler with a low level of empathy.
CONCLUSION
Differences in empathy of humans providingneonatal handling of Amazon chicks produceddifferences in physiologic indices of stress, al-though no difference was detected in chick behav-ioral tameness or DTH response. Parental featherpicking predicted differences in both physiologicand behavioral responses to handling but notDTH response to a foreign protein challenge. Theresults suggest that adaptability of wild animalsto captivity might be improved by a better under-standing of the basis for differences in the effectsof quality of parenting and human handler per-sonality on behavioral development of parrots.
REFERENCES
Alcock, J. 1993. Animal behavior. Sunderland, MA:Sinauer Associates, Inc.
Ascione, F.R. 1993. Children who are cruel to animals:A review of research and implications for develop-mental psychopathology. Anthrozoos 6:226–247.
Beck. A.M., and A.H. Katcher. 1989. Bird-human in-teraction. Journal of the Association of Avian Veteri-narians 3:152–153.
Bodnoff, S.R., B. Suranyi-Cadotte, R. Quirion, andM.J. Meaney. 1987. Postnatal handling reducesnovelty-induced fear and increases [3H]flunitra-zepam binding in rat brain. European Journal ofPharmacology 144:105–107.
126 Manual of Parrot Behavior

Buck, R., and B. Ginsburg. 1997. “Communicative genesand the evolution of empathy.” In Empathic accuracy,ed. W. Ickes, pp. 17–43. New York: Guilford Press.
Collette, J.C., J.R. Millam, K.C. Klasing, and P.S.Wakenell. 2000. Neonatal handling of Amazon par-rots alters the stress response and immune function.Applied Animal Behavior Science 66:335–349.
Davis, W.L. (now Aengus), and J.R. Millam. 1997.Tameness produced by the neonatal handling of par-ent-raised orange-winged amazon chicks. Master’sthesis, University of California, Davis.
Denenberg, V.H., and M.X. Zarrow. 1971. “Effect ofhandling in infancy upon adult behavior and adreno-cortical activity: Suggestions for a neuroendocrinemechanism.” In Early childhood: The developmentof self regulatory mechanisms, ed. D.H. Walcher andD.L. Peters, pp. 40–74. New York: Academic Press.
Dhabhar, F.S. 1998. Stress-induced enhancement ofcell-mediated immunity. Annals of the New YorkAcademy of Sciences 840:359–372.
Duncan, I.J.H. 1992. “The effect of the researcher onthe behavior of poultry.” In The inevitable bond: Ex-amining scientist-animal interactions, ed. H. Davisand D. Balfour, pp. 285–294. Cambridge: Cam-bridge University Press.
Eisenberg, N. 1988. Empathy and sympathy: A brief re-view of the concepts and empirical literature. An-throzoos 2:15–17.
Fox, M.W. 1985. “Empathy, humaneness and animalwelfare.” In Advances in animal welfare science1984, ed. M.W. Fox and L.D. Mickley, pp. 61–73.Hingham, MA: Kluwer Academic Publishers.
Gross, W.B., and P.B. Siegel. 1979. Adaptation ofchickens to their handler, and experimental results.Avian Diseases 23:708–714.
Harris, J.M. 1989. Avian companions and the human-animal bond. Journal of the American VeterinaryMedical Association 195:1517–1518.
Hassig, A., L. Wen-Xi, and K. Stampfli. 1996. Stress-induced suppression of the cellular immune reac-tions: On the neuroendocrine control of the immunesystem. Medical Hypotheses 46:551–555.
Hemsworth, P.H., J.L. Barnett, and C. Hansen. 1981.The influence of handling by humans on the behav-iour, reproduction and corticosteroids in the juvenilefemale pig. Horm Behav 15:396–403.
Hemsworth, P.H., J.L. Barnett, and C. Hansen. 1986.The influence of handling by humans on the behav-iour, reproduction, and corticosteroids of male andfemale pigs. Applied Animal Behavior Science15:303–314.
Hennessy, J.W., and S. Levine. 1979. Stress, arousaland the pituitary-adrenal system: A psychoen-docrine model. Progress in Psychobiology and Phy-siological Psychology 8.
Jones, R.B., and J.M. Faure. 1981. The effects of regu-lar handling on fear responses in the domestic chick.Behavioral Processes 6:135–143.
Jones, R.B., and D. Waddington. 1993. Attenuation ofthe domestic chick’s fear of human beings via regu-lar handling: In search of a sensitive period. AppliedAnimal Behaviour Science 36:185–195.
Karsh, E.B. 1983. “The effects of early handling on thedevelopment of social bonds between cats and peo-ple.” In New perspectives on our lives with compan-ion animals, ed. A.H. Katcher and A.M. Beck, pp.22–28. Philadelphia: University of PennsylvaniaPress.
Karson, S., and J.W. O’Dell. 1976. Clinical use of the16PF. Champaign, IL: Institute for Personality andAbility Testing.
Lago, D., R. Kafer, M. Delaney, and C. Connell. 1988.Assessment of favorable attitudes toward pets: De-velopment and preliminary validation of self-reportpet relationship scales. Anthrozoos 1:240–254.
Levine, S. 1957. Infantile experience and resistance tophysiological stress. Science 125:405.
Levine, S. 1962. Plasma-free corticosteroid response toelectric shock in rats stimulated in infancy. Science135:795–796.
Levine, S., C. Coe, and S.G. Wiener. 1989. “Psycho-neuroendocrinology of stress: A psychobiologicalperspective.” In Psychoendocrinology, ed. R.F.Brush and S. Levine, pp. 341–377. San Diego: Aca-demic Press.
Lorenz, K. 1952. King Solomon’s ring. New York:Signet.
Lorenz, K.Z. 1953. Man meets dog. Baltimore: Penguin.Loughlin, C.A., and P.W. Dowrick. 1993. Psycholog-
ical needs filled by avian companions. Anthrozoos6:166–172.
Meaney, M.J., D.H. Aitken, S.R. Bodnoff, L.J. Iny, andR.M. Sapolsky. 1985a. The effects of postnatal han-dling on the development of the glucocorticoid re-ceptor systems and stress recovery in the rat.Progress in Neuro-Psychopharmacology & Biologi-cal Psychiatry 9:731–734.
Meaney, M.J., D.H. Aitken, S.R. Bodnoff, L.J. Iny, J.E.Tatarewicz, and R.M. Sapolsky. 1985b. Early post-natal handling alters glucocorticoid receptor con-centrations in selected brain regions. BehavioralNeuroscience 99:765–770.
Meaney, M.J., D.H. Aitken, and R.M. Sapolsky. 1987.Thyroid hormones influence the development ofhippocampal glucocorticoid receptors in the rat: Amechanism for the effects of postnatal handling onthe development of the adrenocortical stress re-sponse. Neuroendocrinology 45:278–283.
Meaney, M.J., D.H. Aitken, C. van Berkel, S. Bhat-nagar, and R.M. Sapolsky. 1988. Effect of neonatal
12 / Handler Attitude and Chick Development 127

handling on age-related impairments associatedwith the hippocampus. Science 239:766–768.
Meaney, M.J., J. Diorio, D. Francis, J. Widdowson, P.LaPlante, C. Caldji, S. Sharma, J.R. Seckl, and P.M.Plotsky. 1996. Early environmental regulation offorebrain glucocorticoid receptor gene expression:Implications for adrenocortical responses to stress.Developmental Neuroscience 18:49–72.
Mitchell, J.B., L.J. Iny, and M.J. Meaney. 1990. Therole of serotonin in the development and environ-mental regulation of type II corticosteroid receptorbinding in rat hippocampus. Developmental BrainResearch 55:231–235.
Mugford, R.A., and J.G. M’Comisky. 1975. “Some re-cent work on the psychotherapeutic value of cagebirds with old people.” In Pet animals and society,ed. R.S. Anderson, pp. 54–65. London: BailliereTindall.
Murphy, L.B., and I.J.H. Duncan. 1978. Attempts tomodify the responses of domestic fowl towardshuman beings: II. The effect of early experience.Applied Animal Ethology 4:5–12.
Nicol, C.J. 1992. Effects of environmental enrichmentand gentle handling on behaviour and fear responsesof transported broilers. Applied Animal BehaviourScience 33:367–380.
Scott, J.P., and J.L. Fuller. 1965. Genetics and the so-cial behavior of the dog. Chicago: University ofChicago Press.
Seabrook, M.F. 1972. A study to determine the influ-ence of the herdsman’s personality on milk yields. JAgric Labour Sci 1:45–59.
Selye, H. 1950. The physiology and pathology of expo-sure to stress. Montreal: Acta.
Smythe, J.W., W.B. Rowe, and M.J. Meaney. 1994.Neonatal handling alters serotonin (5-HT) turnoverand 5-HT2 receptor binding in selected brain re-gions: Relationship to the handling effect on gluco-corticoid receptor expression. Developmental BrainResearch 80:183–189.
Statistical Analysis Systems Institute, Inc. 1990.SAS/STAT user’s guide: Version 6, 4th ed. SAS Insti-tute Inc., Cary, NC.
Suchecki, D., P. Rosenfeld, and S. Levine. 1993.Maternal regulation of the hypothalamic-pituitary-adrenal axis in the infant rat: The roles of feedingand stroking. Developmental Brain Research75:185–192.
Templer, D.I., C.A. Salter, S. Dickey, R. Baldwin, andD.M. Veleber. 1981. The construction of a pet atti-tude scale. Psychological Record 31:343–348.
Toft, C.A. 1993. Genetics of captive propagation forconservation: A manual for aviculturists. Specialpublication No. 1, Psittacine Research Project, Dept.of Avian Sciences, University of California, Davis.
Vriends. M.M. 1984. Pet birds. New York: Simon andSchuster.
Zahn-Waxler, C., B. Hollenbeck, and M.R. Radke-Yarrow. 1985. “The origins of empathy and altru-ism.” In Advances in animal welfare science 1984,ed. M.W. Fox and L.D. Mickley, pp. 21–41. Hing-ham, MA: Kluwer Academic Publishers.
128 Manual of Parrot Behavior

For over 25 years, I have used the modeling tech-nique described in this chapter to teach GreyParrots (Psittacus erithacus) to use elements ofEnglish speech meaningfully and then used thiscommunication code to examine their cognitivecapacities. My oldest subject, Alex, exhibits abil-ities comparable to those of marine mammals and apes, and sometimes to those of four- to six-year-old children (Pepperberg 1999). These abili-ties are not, however, inferred from Alex’s re-sponses to various types of operant behaviortasks, as is common in animal research (Zentall1993) but, because he uses human speech, are de-termined from his direct vocal answers to directvocal questions, much as we study young chil-dren. Thus he demonstrates intriguing com-municative and cognitive parallels with humans,despite his phylogenetic distance. I doubt I taughtAlex and the other parrots these abilities de novo;rather, their achievements likely derive from exis-tent cognitive and neurological architectures usedin nature. This point is critical in discussing myresearch involving the importance of direct in-terspecies communication as an investigativetool, the capacities this communication form en-ables us to unveil, and the centrality of birds forstudying the evolution of communication andcognition.
THE IMPORTANCE OF INTERSPECIESCOMMUNICATION
Parrots’ vocal plasticity enables direct inter-species communication (Pepperberg 1981). Butwhy use this code rather than their natural systemto examine how communication and cognitionevolved and the extent to which their abilities
match those of other creatures? The answer lies inthe just-mentioned existent cognitive architecture.I believe parrots acquire elements of human com-munication that can be mapped or adapted to theircode. By observing what they do and do notlearn, we determine what is held in common withhumans and thus more about the avian system. Ido not believe parrots could, for example, learnaspects of reference (e.g., labels for specificclasses of objects such as apples, or for colorssuch as green or red) unless their own naturalcode has such referentiality. Although determin-ing referentiality in this manner is inferential, di-rect determination also has difficulties (seeCheney & Seyfarth 1992). I also believe thatstressing the avian system to see what aspects ofinput are needed for exceptional learning—thatwhich does not necessarily occur during normaldevelopment (Bandura 1971; e.g., acquiring an-other species’ code)—provides a detailed under-standing of the learning process. Because richerinput is needed to learn another species’ code (al-lospecific learning) than to learn one’s ownspecies’ code (conspecific learning; Pepperberg1985), we can determine how and whether “nur-ture” modifies “nature” (i.e., the innate predispo-sitions that facilitate conspecific learning andmay initially block allospecific learning), andthus uncover additional mechanisms for, and thefull extent of, communicative learning. Again,these mechanisms must be part of the existentcognitive architecture and not something taughtde novo, and are mechanisms that could bemissed without proper experimentation. And onlyby elucidating such mechanisms can we under-stand the complexity of the parrot system.
133
13Grey Parrot Cognition and Communication
Irene M. Pepperberg

Interspecies communication also has practicalapplications for studying cognition. It (1) directlystates the content of our queries—animals needn’tdetermine both the answer and nature of a ques-tion via trial and error; (2) incorporates researchshowing that social animals may respond morereadily and accurately within an ecologicallyvalid social context (see Menzel & Juno 1982);(3) allows facile data comparisons amongspecies, including humans; (4) allows rigoroustesting of the acquired communication code thatavoids expectation cuing (i.e., subjects choose re-sponses from their entire repertoire rather thanfrom a subset relevant only to a particular topic);and, most importantly, (5) is an open, arbitrary,creative code with enormous signal variety, en-abling animals to respond in novel, possibly inno-vative ways that demonstrate greater competencethan do operant paradigms’ required responses,and (6) thereby allows examination of the natureand extent of information animals perceive.Interspecies communication thus facilely demon-strates non-humans’ inherent capacities and mayenable more complex learning (see Pepperberg1981, 1999).
HOW GREYS LEARN: PARALLELS WITHHUMANS
My Greys’ learning sometimes parallels humanprocesses and suggests insights into how acquisi-tion of complex communication may haveevolved. Input that is referential, demonstratesfunctionality (i.e., is contextually applicable), andis socially rich allows parrots, like young children(e.g., Hollich et al. 2000), to acquire communica-tion skills effectively (Pepperberg 1981, 1985,1990a, 1994b, 1999; Pepperberg & McLaughlin1996; Pepperberg et al. 1998, 1999). Reference isgenerally defined as an utterance’s meaning—therelationship between labels and the objects towhich they refer—and is exemplified by our useof referential rewards (the bird receives the ob-jects it labels). Context/functionality involves theparticular situation in which an utterance is usedand effects of its use; initial use of a label to re-quest objects gives the bird a reason to learn theunique, unfamiliar sounds of English labels.Social interaction signals which environmentalcomponents should be noted, emphasizes com-mon attributes—and possible underlying rules—of diverse actions, and allows input to be contin-
uously adjusted to the learner’s level. Interactionalso engages the subject directly, provides contex-tual explanations of the reasons for the actions,and demonstrates actions’ consequences. I de-scribe the primary technique for training ourbirds, our results, and experiments to determinewhich input elements are both necessary and suf-ficient to engender learning.
The primary training technique, the model/rival (M/R) system (Pepperberg 1981), is based onthat of Todt (1975), who examined parrots’ sociallearning, and Bandura’s (1971) studies of how so-cial modeling affects human learning. The M/Rprocedure involves three-way interactions amongtwo human speakers and the avian student; it usessocial interaction to demonstrate targeted vocalbehavior. M/R training introduces new labels andconcepts and aids in shaping pronunciation
During M/R training, humans demonstrate in-teractive responses to be learned. Sessions beginwith a parrot observing two humans handling oneor more items in which the bird has shown an in-terest. The bird watches as one human “trains” theother. The trainer presents and asks questionsabout the item(s) (e.g., “What’s here?” “Whatcolor?”), and gives the human model praise andthe object(s) in question to reward correct an-swers referentially. Multiple exemplars of the ob-ject are used to help the bird generalize the labelbeyond a specific stimulus. Incorrect responses(like those the bird may make at the time) arepunished by scolding and temporarily removingthe item(s) from sight. Thus the second human isnot only a model for the parrot’s responses and arival for the trainer’s attention but also illustratesthe effects of an error: The model is asked to tryagain or talk more clearly if the response was (de-liberately) incorrect or garbled, thereby allowinga bird to observe corrective feedback (seePepperberg 1999). A bird is included in interac-tions and rewarded for successive approximationsto a correct response; thus training is adjusted toits level.
Unlike Todt’s procedure (and that of others; seePepperberg & Sherman 2000), our protocol alsoinvolves reversing roles of human trainer andmodel, and includes the parrot in interactions.Thus one person is not always the questioner andthe other the respondent, and we show how theprocedure effects environmental change (Pepper-berg 1981). Inclusion of role reversal in M/R
134 Manual of Parrot Behavior

training counteracts drawbacks associated withTodt’s method: His birds, whose trainers alwaysmaintained their respective roles, responded onlyto the human posing the questions. In contrast,our birds respond to, interact with, and learn fromall trainers.
Importantly, M/R training exclusively uses in-trinsic reinforcers. That is, the bird’s reward forproducing a label (“X”) is the object (X) to whichthe label or concept refers. This procedure en-sures the closest possible correlation of label orconcept to be learned and the object or task towhich it refers. Earlier unsuccessful programs forteaching psittacids to communicate with humans(e.g., Mowrer 1950) used extrinsic rewards. Thus,on the few occasions when those subjects cor-rectly labeled food or non-food items, or madeappropriate responses to various specific com-mands, they received a single, favored food thatneither directly related to, nor varied with, thelabel or concept being taught. Extrinsic rewards,however, delay label/concept acquisition by con-founding the label of the exemplar or concept tobe learned with that of the food (see Miles 1983;Pepperberg 1981; Pepperberg & Sherman 2000).My birds never receive extrinsic rewards. Too, useof intrinsic rewards demonstrates label function-ality: Initially, label use results in acquisition ofdesired objects, and thus provides a reason for thebird to acquire that label.
Occasionally, Alex receives a more general re-ward: Because he sometimes will not focus onobjects used to train a particular concept, wetaught him “I want X” (i.e., to separate labelingand requesting; Pepperberg 1988b) so his rewardis the right to request vocally something more de-sirable than what he identified. This protocol pro-vides flexibility but maintains referentiality: Alexnever, for example, automatically receives nutsfor identifying a cork. He must state “I want nut,”and trainers will not respond to requests until theappropriate prior task is completed. Thus birdslearn the label as a true identifier, not merely onan emotional level. Training “want” provides twoadditional advantages: First, trainers can distin-guish incorrect labeling from appeals for otheritems; that is, during a test birds unable to use“want” might not be making errors but could beasking for treats, and test scores might decline forreasons unrelated to competence. Second, birdscan be tested for a simple form of intentionality.
For example, if Alex asks for a certain object, heis rarely satisfied with a replacement and contin-ues to request the desired item (Pepperberg 1999).
RESULTS (FROM PEPPERBERG 2002b)
Labeling and Basic Requests, Overview
Through M/R training, Alex learned tasks thatwere once thought beyond the capability of all buthumans or, possibly, certain non-human primates(Premack 1978). He can label over 50 objects. Hehas functional use of “no” and phrases such as“come here,” “I want X,” and “Wanna go Y,”where X and Y are appropriate labels for objectsor locations (Pepperberg 1981, 1999). Trainers’incorrect responses to his requests (e.g., substitu-tion of something other than what he requested)generally result in his saying “No” and repeatingthe request (Pepperberg 1987c, 1988b). He labelsseven colors and identifies five shapes (as “two-,”“three-,” “four-,” “five-,” or “six-corner”; Pepper-berg 1983). He uses the labels “two,” “three,”“four,” “five,” and “six” to distinguish quantitiesof objects, including collections made of novelitems and randomly placed and heterogeneoussets of items (Pepperberg 1987b, 1994a). Hecombines vocal labels to identify proficiently, re-quest, refuse, categorize, and quantify over 100objects, including those varying somewhat fromtraining exemplars. His accuracy averages ~80%when tested on these abilities (Pepperberg 1981,1983, 1987b, 1987c, 1988a, 1994a, 1999).
Concepts of Category
Alex comprehends the concept of “category.” Helearned not only to label different hues andshapes, but also to categorize objects having bothcolor and shape with respect to either category byresponding to “What color?” or “What shape?”(Pepperberg 1983). He understands that “green,”for example, is a particular instance of the cate-gory “color,” and that, for objects with both colorand shape, specific instances of these attributes(e.g., “green,” “three-corner”) represent differentcategories. Thus, he learned that a set of responsesexists—color labels—that forms the class “color”and another set—shape labels—that forms theclass “shape” (Pepperberg 1996). His successshows understanding of higher-order class con-cepts, because color labels have no intrinsic con-nection to the label “color” nor do shape labels to
13 / Grey Parrot Cognition and Communication 135

the label “shape”. Our protocol requires Alex tocategorize the same exemplar with respect toshape at one time and color at another, and thusinvolves flexibility in changing the basis for clas-sification. Such capacity for reclassification isthought to indicate “abstract aptitude” (Hayes &Nissen 1956/1971).
Same/Different and Absence
In the 1970s, comprehension of same/differentwas singled out as requiring abilities not typicallyattributable to non-primates and specifically notto birds (Premack 1978, 1983; Mackintosh et al.1985). Researchers argued that understanding thesame/different concept is more complex thanlearning to respond to match-to-sample and nonmatch-to-sample or oddity-from-sample, orhomogeneity and non-homogeneity. The first re-quires use of arbitrary symbols to represent rela-tionships of sameness and difference between setsof objects and the ability to denote the attributethat is same/different (Premack 1983). Specifi-cally, Premack (1983) claims animals need sym-bolic representation—some elementary form oflanguage—to succeed. The other tasks, in con-trast, require only that subjects show savings inthe number of trials needed to respond to B and Bas a match (or as a homogeneous field) afterlearning to respond to A and A as a match (andlikewise by showing a savings in trials involvingC and D after learning to respond appropriately toA and B as non-matching or non-homogeneous).Subjects in match-to-sample and nonmatch-to-sample studies might even be respondingbased on “old” versus “new” or “familiar” versus“unfamiliar,” that is, on the relative number oftimes they experience A versus the number oftimes they see different Bs (Premack 1983).Subjects that understand same/different, however,not only know that two non-identical red objectsare related in the same way as are two non-identical blue objects—by color—but also knowthe red objects are related to each other differ-ently than are two non-identical square objects,and, moreover, can transfer this understanding toany attribute of an item (Premack 1978, 1983).Subjects likewise have to understand the conceptof difference.
Alex learned abstract concepts of “same” and“different” and to respond to the absence of infor-mation about these concepts if nothing is same or
different. Thus, given two objects that are identi-cal or that vary with respect to some or all of theattributes of color, shape, and material, Alex re-sponds with the appropriate category label as towhich attribute is “same” or “different” for anycombination (Pepperberg 1987a), or “none” ifnothing is same or different (Pepperberg 1988a).He responds accurately for novel objects, colors,shapes, and materials, including those he cannotlabel. Furthermore, Alex responds to the specificquestions, not merely on the basis of his trainingand physical attributes of the objects: He was stillabove chance when, for example, we asked“What’s same?” for a green wooden triangle anda blue wooden triangle. If he had ignored thequestion and responded based on prior training,he would have determined, and responded withthe label for, the one anomalous attribute (in thiscase, “color”). Instead, he produced one of thetwo appropriate answers (i.e., “shape” or “mah-mah” [matter]; Pepperberg 1987a).
Alex’s use of “none” is important because un-derstanding and commenting upon non-existence,or even the slightly more basic notion of absence,although seemingly simple, denotes a relativelyadvanced stage in cognitive and linguistic devel-opment (Brown 1973). Organisms react to ab-sence only after acquiring knowledge about theexpected presence of events, objects, or other in-formation in their environment, that is, only whendiscrepancy exists between the expected and ac-tual state of affairs (e.g., Skinner 1957; de Villiers& de Villiers 1979; Hearst 1984). (Such behavioris, however, qualitatively different from learningwhat stimulus leads to absence of reward—e.g.,Astley & Wasserman 1992—where subjects sim-ply learn what to avoid.) Non-humans have beentested on absence using Piagetian object perma-nence, and some do react to the disappearance ornon-existence of specific items they expect to bepresent (e.g., Funk 1996; Pepperberg et al. 1997).Evidence also exists in nature. Some songbirds,for example, react to absence of signs of territo-rial defense (e.g., song) from conspecific neigh-bors with positive acts of territorial invasion(Peek 1972; Krebs 1977; Smith 1979). Bloom(1970), however, suggests that not only compre-hension but also verbal production of terms re-lating to non-existence is necessary before anorganism is thought to have the concept of non-existence. Experimentally demonstrating this
136 Manual of Parrot Behavior

concept thus is difficult, even in humans, andAlex’s capacities are therefore notable.
Numerical Concepts
I then asked whether Alex could form a new cat-egorical class of labels for quantity, that is, learna concept of number. Could he now learn, for ex-ample, to reclassify a group of wooden objectsknown as “wood” or “green wood” as “fivewood”? To succeed, he would have to understandthat a new set of labels, “one,” “two,” “three,”“four,” “five,” and “six,” represented a novelclass: a means to categorize objects based on acombination of physical similarity within a groupand the group’s quantity, rather than only physicalcharacteristics of group members. He would alsohave to generalize this new class of numerical la-bels to novel items, objects in random arrays, andheterogeneous collections. Note that Koehler(1943, 1950, 1953) and colleagues (Braun 1952;Lögler 1959) had already demonstrated Greys’sensitivity to quantity and basic concepts ofnumerosity/numerousness; Koehler’s birds couldopen boxes randomly containing zero, one, or twobaits until they obtained a fixed number (e.g.,four). The number of boxes to be opened to obtainthe precise number of baits varied across trials,and the number sought depended upon independ-ent visual cues: black box lids denoted two baits,green lids three, and so forth; Koehler’s birds sup-posedly solved four different problems of thiskind. He did not state, however, if different col-ored lids were presented randomly in a single se-ries, and thus whether colors indeed “repre-sented” particular quantities (see Pepperberg1987b). Lögler (1959) transferred such behaviorto light flashes and flute notes, thus going fromsimultaneous visual representations to sequentialauditory ones. But could Alex, like Matsuzawa’s(1985) chimpanzee, go beyond these tasks anduse number as a categorical label?
Although Alex’s numerical understanding issimpler than that of human children (Fuson1988), he does comprehend some concept ofquantity. We have yet to show conclusively thatAlex can, for example, without training, transferfrom enumerating simultaneous visual displays toenumerating sequential auditory ones (e.g., trans-fer to count sequential metronome clicks to statehe has heard “three”), but he can label quantitiesof physical objects up to and including six
(Pepperberg 1987b). The object sets need not befamiliar, nor be in any particular pattern, such asa square or triangle. Furthermore, shown a het-erogeneous collection—of Xs and Ys—he can re-spond appropriately to either “How many X?” or“How many Y?” (Pepperberg 1987b). This abilityis beyond what might be considered subitizing inyoung children, who are generally given only ho-mogeneous sets (e.g., Starkey & Cooper 1995),and who, if asked about subsets, generally labelthe total number of objects in a heterogeneous setif, like Alex, they have been trained on homoge-neous sets exclusively (see Greeno et al. 1984).Shown a “confounded number set” (collections offour groups of items that vary in two colors andtwo object categories—e.g., blue and red woodand blue and red wool), Alex can label the num-ber of items uniquely defined by the combinationof one color and one object category (e.g., “Howmany blue wood?”). Although Alex’s mechanismsmay not be identical to those of humans, his accu-racy (Pepperberg 1994a) replicates that of hu-mans (Trick & Pylyshyn 1989); thus a non-primate, non-mammal shows competence that, ina chimpanzee, would be taken to indicate ahuman level of intelligence (Pepperberg 1999).
Relative Size
All research discussed until now involves forma-tion of categorical classes based at least indirectlyon absolute physical criteria, not relative con-cepts. Color and shape labels are symbolic andthus abstract, but refer to concrete entities (Pep-perberg 1996). Demonstrating that animals, andbirds in particular, can respond to relative con-cepts is difficult: Previous studies suggested thatresponse on an absolute basis was always used inpreference to response on a relative basis; the lat-ter response was apparent only if the former wasblocked (e.g., Page et al. 1989; Hulse et al. 1990;cf. Weisman & Ratcliffe 1989; Hurly et al. 1990).Might Alex’s categorical class training enable himto respond on a relative basis (e.g., bigger/smaller)? Such data would provide direct compar-isons with research on marine mammals (Schus-terman & Krieger 1986).
After M/R training on “What color bigger/smaller?” with a limited set of colors and objects(yellow, blue, green; cups, woolen felt circles,Play-Doh rods), we tested Alex on a variety of fa-miliar and unfamiliar items. Alex could indeed
13 / Grey Parrot Cognition and Communication 137

classify objects with respect to relative size; over-all test scores were 78.7% (Pepperberg &Brezinsky 1991). Although we did not examinewhether he could (or would) transfer this conceptto a different modality (e.g., amount of sound),whether acquisition of this concept might helphim learn a different relationship (e.g., relativedarkness), or how close in size two objects mustbe before he could not discriminate a difference,Alex demonstrated an understanding at leastequivalent to that of certain marine mammals(e.g., Schusterman & Krieger 1986). He trans-posed size relationships to stimuli outside thetraining domain (Pepperberg & Brezinsky 1991)and, because he responded with the label for anattribute other than size (i.e., color), we could re-move many absolute stimulus cues by using en-tirely novel objects (see Pepperberg 1987a,1987b, 1990a): unfamiliar shapes, materials,sizes, and colors, often ones he could not label(e.g., hand-dyed styrofoam stars; Pepperberg &Brezinsky 1991). Most interesting was that hecould also, without any training, indicate whenexemplars did not differ in size by responding“none” and answer questions based on object ma-terial rather than color (Pepperberg & Brezinsky1991). Thus he was not limited to respondingwithin a single dimension, he was attending toour questions, and he transferred informationlearned in one domain (the same/different study)to another. Such ability to transfer is, as justnoted, a mark of complex cognitive processing(Rozin 1976).
Comprehension via Recursive andConjunctive Tasks
We next compared Alex’s comprehension abilitieswith those of marine mammals trained in inter-species communication (e.g., Herman 1987;Schusterman & Gisiner 1988). Most work withcetaceans and pinnipeds uses the comprehensionmode; that is, researchers demonstrate how wellanimals understand the communication code byacting appropriately upon various commands. Incontrast, most work with non-human primates upuntil the early 1990s and all prior work with Alex,although clearly involving comprehension (e.g.,the difference between queries of “How many?”“What’s same/different?” “What color bigger/smaller?” etc. with respect to any two objects),emphasized how accurately subjects produce the
code (Gardner & Gardner 1969; Pepperberg1981; Miles 1983; Savage-Rumbaugh 1984; cf.Savage-Rumbaugh et al. 1993). To maintain ourvocal paradigm but provide the necessary com-parisons, students and I trained and tested Alex ona recursive task like that used with other animals(Pepperberg 1990b; also Granier-Deferre &Kodratoff 1986). In a recursive task, subjects arepresented with several different objects and oneof several different possible questions or com-mands concerning the attributes of these objects.Each question or command contains several parts,the combination of which uniquely specifieswhich object is targeted and what action is to beperformed. Question complexity is determined bycontext (number of different possible objectsfrom which to choose) and the number of its parts(e.g., number of attributes used to specify the tar-get and number of actions from which to choose).Subjects must divide the question into these partsand (recursively) understand each part to answercorrectly. Subjects demonstrate competence byreporting on only a single aspect (e.g., color,shape, or material) of, or performing one of sev-eral possible actions (fetching, touching) on, anobject that is one of several differently coloredand shaped exemplars of various materials. Alexwas shown many unique combinations of sevenexemplars and asked, “What color is object-X?”“What shape is object-Y?” “What object is color-A?” or “What object is shape-B?” His accuracy,above 80% (Pepperberg 1990b), was comparableto that of marine mammals and non-humanprimates.
Students and I took this work one step further,by adding a conjunctive condition to the recursivetask (Pepperberg 1992). Here Alex was againshown seven-member collections but was nowasked to provide information about the specificinstance of one category of an item uniquely de-fined by the conjunction of two other categories;for example, “What object is color-A and shape-B?”. Other objects on the tray exemplified one,but not both, of these defining categories. Alex’saccuracy was 76.5%, indicating he understood allelements in the question. Again, his data wascomparable to that of marine mammals.
FURTHER STUDIES ON ACQUISITION
M/R training successfully demonstrated whatinput elements enabled acquisition of some level
138 Manual of Parrot Behavior

of allospecific communicative competence butdid not elucidate which were necessary and suffi-cient. What happens if input lacks some ele-ments? Answering that question required addi-tional parrots, because Alex might have ceasedlearning simply because circumstances hadchanged, not because of the quality of the change.New, untrained subjects would be uninfluencedby prior experience. With three new Greys,Kyaaro, Alo, and Griffin, students and I begantesting the relative importance of three majorinput elements of M/R training: reference, con-text/function, and social interaction.
Initial Studies Eliminating Aspects of Input
I first gave Alo and Kyaaro three types of input:(1) audiotapes of Alex’s sessions, which werenon-referential, not contextually applicable, andnon-interactive; (2) videotapes of Alex’s sessions,which were referential, minimally contextuallyapplicable, and non-interactive; and (3) usualM/R training that was referential, contextually ap-plicable, and interactive. In the first two experi-ments, birds listened to or watched tapes in socialisolation. The first condition paralleled earlierstudies on allospecific song acquisition (Marler1970; Baptista & Petrinovich 1984, 1986; speechis not a natural parrot vocalization), and the sec-ond involved still-unresolved issues about avianvision and video (e.g., Ikebuchi & Okanoya 1999;Lea & Dittrich 1999). I counterbalanced labelsacross birds and ensured equivalent training timeacross sessions. Like songbirds, neither parrotlearned anything substantive from audio, norfrom videotapes. Both, however, learned to com-prehend and produce M/R-trained labels (Pepper-berg 1994b). But such studies were just the firststeps in determining strictures for allospecificacquisition.
Further Studies on Elements of Input
We then completed five more experiments totease out effects of input. In the first, juveniles’video sessions were repeated with “co-viewers”who merely ensured birds attended to the monitor(Pepperberg et al. 1998). Trainers provided socialapprobation for viewing and pointed to thescreen, making comments like “Look what Alexhas!” but did not repeat new labels, ask questions,or relate content to other training. The procedurewas based on data from young children that sug-
gested that video learning (e.g., from SesameStreet) increased when children watched with in-teractive co-viewers (Corder-Bolz & O’Bryant1978; Lemish & Rice 1986; Lesser 1974;Salomon 1977; Watkins et al. 1980; but see Riceet al. 1990). For this experiment, any attempt toproduce the label would be rewarded with vocalpraise, not the object. Thus social interaction waslimited and functional meaning was the same asin basic videotape sessions. In the second study,we increased the amount of interaction, so thetrainer now repeated the new labels and askedquestions (Pepperberg et al. 1999); the rationalewas additional data for children showing that ex-tent of interaction might affect video learning (St.Peters et al. 1989). In the third study, we ensuredthat lack of reward for an attempted targeted vo-calization did not prevent learning from video(Pepperberg et al. 1998): Using the basic video-tape protocol, we included a reward system thatenabled a socially-isolated parrot to receive theitem if it attempted to produce its label. The sys-tem was controlled by a student in another roomwho monitored the parrot’s utterances throughheadphones. In the fourth study, the M/R proce-dure was amended to eliminate some functional-ity and as much social interaction as possible(Pepperberg & McLaughlin 1996). Here we repli-cated studies with children that demonstrated theeffect of an adult jointly focusing (with the child)on the object being labeled: For children, lack ofjoint attention prevented label acquisition (e.g.,Baldwin 1995). In our study, a single trainer satwith her back to the bird, who was seated on aperch within reach of an object (e.g., key). Thetrainer repeated various phrases about the object(e.g., “Look, a shiny key!” “Do you want thekey?” etc.; sentence frames; Pepperberg 1981) butdid not make eye contact with the parrot, nor didshe present the object to the bird. Any attempt atthe targeted label received vocal praise. In thefifth study, we tested whether the bird might havebecome habituated to the single tape we used.Although the tape contained many interactionsand all the different responses that Alex and thetrainers made, the bird might have ignored thematerial after several sessions. We therefore repli-cated the study using live video input from Alex’ssessions (Pepperberg et al. 1999). In none of theseexperiments did parrots learn referential use of la-bels, but they did learn simultaneously presented
13 / Grey Parrot Cognition and Communication 139

M/R-trained labels. Although we are currentlyreplicating video studies using a liquid crystalmonitor to determine whether flicker-fusion ofthe standard cathode ray tube affects video learn-ing (Ikebuchi & Okanoya, 1999), our results sofar emphasize the importance of training that in-volves reference, demonstration of contextualuse/functionality, and social interaction if a parrotis to communicate and not simply mimic humanspeech.
Nevertheless, at least one set of conditions re-mained to be tested: What would happen if weeliminated just the modeling aspect, that is, ifonly a single student labeled the object, queriedthe bird, jointly attended to the object, and thusinteracted fully with the bird and the object?Griffin did not learn labels trained in this mannerafter 50 sessions (Pepperberg et al. 2000). Whenwe switched to M/R training, however, he pro-duced labels with complete clarity after two orthree sessions. Apparently, latent learning had oc-curred: Griffin had acquired the label but did notknow how to use it until he saw its use modeled.In other instances of similar switching (e.g., after50 video sessions to M/R training), birds needed~20 sessions before producing the label clearly(Pepperberg et al. 2000). Thus specific demon-stration of label functionality is also a vital aspectof the training.
Mutual Exclusivity: A More Subtle Form ofInput
In yet another study, students and I found thatcontext-dependent input engenders a form of mu-tual exclusivity (ME) during label learning byGrey Parrots, much like that of young children(Pepperberg & Wilcox 2000). For children, MErefers to their assumption during early wordlearning that an object has one, and only one,label (e.g., Liittschwager & Markman 1991,1994; Merriman 1991). Along with the whole ob-ject assumption (that a label likely refers to theentire object, not some attribute; Macnamara1982; Soja et al. 1985; Markman & Wachtel1988), ME supposedly guides children in initiallabel acquisition. ME may later help childrenovercome the whole object assumption by helpingthem interpret a novel word as something otherthan an object label (Markman 1990), but for veryyoung children, any second label for an object caninitially be more difficult to acquire than the first,
because the second label is viewed as an alterna-tive (Liittschwager & Markman 1991, 1994). ButME depends upon input: Children (Gottfried &Tonks 1996) and parrots like Alex, who receiveinclusivity data (X is a kind of Y; color labelstaught as additional, not alternative, labels for anitem, i.e., “Here’s a key; it’s a green key”), tend toaccept multiple labels for items and form hierar-chical relations. Thus Alex responds to “Whatcolor?” “What shape?” “What matter?” and“What toy?” for a wooden block (Pepperberg1990a, 1990b). If, however, subjects are givencolor or shape labels as alternative labels (i.e.,“Here’s a key; it’s green”), they resist using thesemodifiers for a previously labeled item. Griffin,trained in the latter manner, thus responded to“What color?” with previously learned object la-bels in over 50 training sessions (Pepperberg &Wilcox 2000). Results were not confined to mod-ifiers. If, for example, he lacked an object label—cup—he used its color label even if asked “Whattoy?” and had difficulty learning “cup” (Pepper-berg & Wilcox 2000). Thus even small changes ininput (e.g., “It’s a green key” versus “It’s green”)affect label acquisition in parrots in a way thatmatches young children.
Combinatory Learning
On the basis of primarily behavioral data, re-searchers (e.g., Greenfield 1991) argue that (1)parallel development of communicative and phys-ical object (manual) combinatorial abilities existsin young children, (2) these abilities initially havea common neural substrate, (3) a homologousgreat ape substrate allows for similar, if limited,co-emergence of these two abilities, and (4) suchabilities indicate a shared evolutionary history forcommunicative and physical behavior (Johnson-Pynn et al. 1999). Interestingly, we found compa-rable, if limited, parallel combinatorial develop-ment in a Grey Parrot (Pepperberg & Shive 2001).
Our juvenile Griffin spontaneously combinedphysical objects in similar proportions to sponta-neous label combinations. His two-label combi-nations (e.g., “want X,” “color + object”;Pepperberg & Wilcox 2000) preceded our study;we were thus unable to see if the two behaviorpatterns co-emerged. We did, however, track sys-tematic initiation of both three-item and three-label combinations. Both began in early 2000,though Griffin’s first—and then for several
140 Manual of Parrot Behavior

months only—successful three-object combina-tion occurred in June 1999. Interestingly (1) per-centages of physical and vocal combinations wereroughly equal; (2) despite several months of train-ing on x-corner wood/paper, vocal three-itemcombinations emerged only when he more fre-quently began to combine three objects; (3) suchvocal combinations were not exclusively thosetrained; and (4) combinations were performedwith his beak rather than his feet. Our data implythat a particular mammalian brain structure is not uniquely responsible for such behavior andthat co-occurrent combinations need not arisemanually.
Unlike the Cebus studied (Johnson-Pynn et al.1999), Griffin was not trained on item-basedtasks; we wanted to see if, as his vocalizations de-veloped complexity, spontaneous manipulativebehavior co-emerged. He rarely combined morethan two items, possibly because of difficulty si-multaneously grasping more than two items:Failed attempts at multiple combinations oc-curred because he lacked physical dexterity, notbecause he tried to form impossible combina-tions. However, the frequency with which hepicked up and manipulated paired caps/lids, andhis manipulations of other items, could be exam-ples of intermediate or transitory stages leading tomore difficult combinations. And, by limitingtraining on three-label combinations, we providedonly an example; we then could document sponta-neous utterances. Although we trained “x-cornerwood/paper” (x = 2, 5), “2-corner wood” was onlyone of 14 recorded three-label combinations.
We do not argue that Griffin’s behavior—oreven that of our most advanced subject, Alex(e.g., Pepperberg 1999)—constitutes anythingcomparable to human language or the complexcombinatory behavior of two- to three-year-oldhumans. We have, however, documented co-occurrence of vocal and physical-object combina-torial behavior not previously described in birds.We suggest that (1) our Greys’ behavior patternscompare to some of those of non-human pri-mates, (2) parallel communicatory and physicaldevelopment is not restricted to primates, and (3)neural structures involved in such behavior arenot unique to primates. Although avian neu-roanatomy and its relation to the mammalian lineis not yet well enough understood to determinespecific parallels among oscine, psittacine, and
mammalian structures, the responsible substratesare likely analogous and arose independentlyunder similar evolutionary pressures (e.g., Dea-con 1997); nevertheless, recent arguments (e.g.,Jarvis & Mello 2000; Medina & Reiner 2000)suggest that additional study is needed beforedefinite conclusions are made. Given the evolu-tionary distance between parrots and primates, wesuggest that the search for and arguments con-cerning responsible substrates and common be-havior should be approached with care and not berestricted to the primate line.
PARALLEL EVOLUTION OF AVIAN ANDMAMMALIAN ABILITIES?
In sum, I am not claiming isomorphism forhuman language and any animal communicationsystem, or for human and non-human cognition,but do argue that we must look across species forinformation on evolutionary pressures that helpedshaped existent systems (Pepperberg 1999). Suchpressures were exerted on more than primates andthe existence of complex analogous communica-tion systems and cognitive abilities demonstratethe need to accept their bases in analogous neuralarchitectures. Moreover, complex communicativesystems apparently require or at least co-evolvewith complex cognition: Communication is a so-cial function, but its complexity is based on thecomplexity of information that must be commu-nicated, processed, and received (Smith 1997);thus contingencies that shape intelligence (social,ecological, etc.) also likely shape communication.Humphrey (1976), among others, looking at pri-mates, proposed that intelligence (and presum-ably the need for cognitive processing) is a corre-late of a complicated social system and a longlife, that is, that intelligence resulted from selec-tion favoring animals that could remember andact upon knowledge of detailed social relationsamong group members; more generally, Rozin(1976) defined intelligence as flexibility in trans-ferring skills acquired in one domain to another.How these two patterns might also drive parrotcognitive skills and vocal behavior seems obvi-ous: Long-lived birds existing in complex socialsystems, not unlike those of some primates, useabilities honed for social gains to direct otherforms of information processing and vocal learn-ing. Add the need for categorical classes (e.g., todistinguish neutral stimuli from predators, poi-
13 / Grey Parrot Cognition and Communication 141

sonous from healthful foods, etc.), abilities bothto recognize and remember environmental regu-larities and adapt to unpredictable environmentalchanges over an extensive lifetime, and a commu-nication system that is primarily vocal, the capac-ities of parrots are not surprising. In fact, Marler(1996) proposed some similar parallels betweenbirds and primates, although not specifically forparrots. I believe we must look for parallels—es-sential commonalities—across many species todevelop theories about those behavioral elementsthat are essential to, and the evolutionary pres-sures that have shaped, complex communication(Pepperberg 1999). Whether avian communica-tion and human language (and avian and humancognition) demonstrate convergent evolution—where similar adaptive responses have independ-ently evolved in remotely associated taxa in asso-ciation with similar environmental pressures (Ball& Hulse 1998)—is unclear, but a common core ofskills likely underlies complex cognitive andcommunicative behavior in many species, even ifspecific skills have different manifestations ineach species.
ACKNOWLEDGMENTS
This chapter was based on articles published pre-viously (Pepperberg 2002a, 2002b) that summa-rized other cited material. Writing of this articlewas supported by the MIT School of Architectureand Planning and a grant from The AmericanFoundation. Research was supported by NSF(IBN 96-03803) and REU supplements, the JohnSimon Guggenheim Foundation, the Kenneth A.Scott Charitable Trust, the Pet Care Trust, theUniversity of Arizona Undergraduate BiologyResearch Program, and many, many donors to theAlex Foundation.
REFERENCES
Astley, S.L., and E.A. Wasserman. 1992. Categoricaldiscrimination and generalization in pigeons: Allnegative stimuli are not created equal. Journal ofExperimental Psychology: Animal Behavior Proc-esses 18:193–207.
Baldwin, D.A. 1995. “Understanding the link betweenjoint attention and language.” In Joint attention: Itsorigin and role in development, ed. C. Moore andP.J. Dunham, pp. 131–158. Hillsdale, NJ: Erlbaum.
Ball, G.F., and S.H. Hulse. 1998. Birdsong. AmericanPsychologist 53:37–58.
Bandura, A. 1971. “Analysis of social modelingprocesses.” In Psychological modeling, ed. A.Bandura, pp. 1–62. Chicago: Aldine-Atherton.
Baptista, L.F., and L. Petrinovich. 1984. Social interac-tion, sensitive phases, and the song template hypo-thesis in the white-crowned sparrows. AnimalBehaviour 32:172–181.
Baptista, L.F., and L. Petrinovich. 1986. Song devel-opment in the white-crowned sparrow: Social factorsand sex differences. Animal Behaviour 34:1359–1371.
Bloom, L. 1970. Language development: Form andfunction in emerging grammars. Cambridge, MA:MIT Press.
Braun, H. 1952. Uber das Unterscheidungsvermögenunbenannter Anzahlen bei Papageien. Zeitschrift fürTierpsychologie 9:40–91. [Concerning the ability ofparrots to distinguish unnamed numbers.]
Brown, R. 1973. A first language: The early stages.Cambridge, MA: Harvard University Press.
Cheney, D.L., and R.M. Seyfarth. 1992. Precis of “Howmonkeys see the world.” Behavioral and BrainSciences 15:135–182.
Corder-Bolz, C.R., and S. O’Bryant. 1978. Teacher vs.program. Journal of Communication 28:97–103.
Deacon, T.W. 1997. The symbolic species: The co-evolution of language and the brain. New York:Norton.
de Villiers, P.A., and J.G. de Villiers. 1979. Early lan-guage. Cambridge, MA: Harvard University Press.
Funk, M.S. 1996. Development of object permanencein the New Zealand parakeet (Cyanoramphus auri-ceps). Animal Learning & Behavior 24:375–383.
Fuson, K.C. 1988. Children’s counting and concepts ofnumber. New York: Springer-Verlag.
Gardner, R.A., and B.T. Gardner. 1969. Teaching signlanguage to a chimpanzee. Science 187:644–672.
Gottfried, G.M., and J.M. Tonks. 1996. Specifying therelation between novel and known: Input affects theacquisition of novel color terms. Child Development67:850–866.
Granier-Deferre, C., and Y. Kodratoff. 1986. Iterativeand recursive behaviors in chimpanzees duringproblem solving: A new descriptive model inspiredfrom the artificial intelligence approach. Cahiers dePsychologie Cognitive 6:483–500.
Greenfield, P.M. 1991. Language, tools and brain: Theontogeny and phylogeny of hierarchically organizedsequential behavior. Behavioral & Brain Sciences14:531–595.
Greeno, J.G., M.S. Riley, and R. Gelman. 1984.Conceptual competence and children’s counting.Cognitive Psychology 16:94–143.
Hayes, K.J., and C.H. Nissen. 1956/1971. “Highermental functions of a home-raised chimpanzee.” InBehavior of non-human primates, Volume 4, ed.
142 Manual of Parrot Behavior

A.M. Schrier and F. Stollnitz, pp. 57–115. NewYork: Academic Press.
Hearst, E. 1984. “Absence as information: Some impli-cations for learning, performance, and representa-tional processes.” In Animal cognition, ed. H.L.Roitblat, T.G. Bever, and H.S. Terrace, pp. 311–332.Hillsdale, NJ: Erlbaum.
Herman, L.M. 1987. “Receptive competencies oflanguage-trained animals.” In Advances in the studyof behavior, Volume 17, ed. J.S. Rosenblatt, C. Beer,M-C. Busnel, and P.J.B. Slater, pp. 1–60, New York:Academic Press.
Hollich, G.J., K. Hirsh-Pasek, and R.M. Golinkoff.2000. Breaking the language barrier: An emergentistcoalition model for the origins of word learning.Monographs of the Society for Research in ChildDevelopment 262:1–138.
Hulse, S.H., S.C. Page, and R.F. Braaten. 1990.Frequency range size and the frequency range con-straint in auditory perception by European starlings(Sturnus vulgaris). Animal Learning & Behavior18:238–245.
Humphrey, N.K. 1976. “The social function of intel-lect.” In Growing points in ethology, ed. P.P.G.Bateson and R.A. Hinde, pp. 303–317. Cambridge,UK: Cambridge University Press.
Hurly, T.A., L. Ratcliffe, and R. Weisman. 1990. Relativepitch recognition in white-throated sparrows, Zono-trichia albicollis. Animal Behaviour 40:176–181.
Ikebuchi, M., and K. Okanoya. 1999. Male zebrafinches and Bengalese finches emit directed songsto the video images of conspecific females projectedonto a TFT display. Zoological Science 16:63–70.
Jarvis, E.D., and C.V. Mello. 2000. Molecular mappingof brain areas involved in parrot vocal communica-tion. Journal of Comparative Neurology 419:1–31.
Johnson-Pynn, J., D.M. Fragaszy, E.M. Hirsh, K.E.Brakke, and P.M. Greenfield. 1999. Strategies usedto combine seriated cups by chimpanzees (Pantroglodytes), bonobos (Pan paniscus), and capuchins(Cebus apella). Journal of Comparative Psychology113:137–148.
Koehler, O. 1943. “Zähl”-Versuche an einemKolkraben und Vergleichsversuche an Menschen.Zeitschrift für Tierpsychologie 5:575–712. [“Num-ber” ability in a raven and comparative research withpeople.]
Koehler, O. 1950. The ability of birds to “count.”Bulletin of the Animal Behavior Society 9:41–45.
Koehler, O. 1953. “Thinking without words.” Proceed-ings of the XIVth International Congress of Zoology,pp. 75–88.
Krebs, J. 1977. “Song and territory in the great tit.” InEvolutionary ecology, ed. B. Stonehouse and C.Perrins, pp. 47–62. New York: Macmillan.
Lea, S.E.G., and W.H. Dittrich. 1999. What do birdssee in moving video images? Cahiers de Psycho-logie Cognitive/Current Psychology of Cognition18:765–803.
Lemish, D., and M.L. Rice. 1986. Television as a talk-ing picture book: A prop for language acquisition.Journal of Child Language 13:251–274.
Lesser, G.S. 1974. Children and television: Lessonsfrom Sesame Street. New York: Random House.
Liittschwager, J.C., and E.M. Markman. 1991. Mutualexclusivity as a default assumption in second labellearning. Paper read at the biennial meetings of theSociety for Research in Child Development, April,Seattle, OR.
Liittschwager, J.C., and E.M. Markman. 1994. Sixteen-and 24-month olds’ use of mutual exclusivity as adefault assumption in second-label learning.Developmental Psychology 30:955–968.
Lögler, P. 1959. Versuche zur Frage des “Zähl”-Vermögens an einem Graupapagei und Vergleich-sversuche an Menschen. Zeitschrift für Tierpsy-chologie 16:179–217. [Studies on the question of“number” sense in a grey parrot and comparativestudies on humans.]
Mackintosh, N.J., B. Wilson, and R.A. Boakes. 1985.Differences in mechanism of intelligence amongvertebrates. Philosophical Transactions of the RoyalSociety, London, Series B 308:53–65.
Macnamara, J. 1982. Names for things: A study ofhuman learning. Cambridge, MA: MIT Press.
Markman, E.M. 1990. Constraints children place onword meaning. Cognitive Science 14:57–77.
Markman, E.M., and G.F. Wachtel. 1988. Children’suse of mutual exclusivity to constrain the meaningsof words. Cognitive Psychology 20:121–157.
Marler, P. 1970. A comparative approach to vocallearning: Song development in white-crowned spar-rows. Journal of Comparative and PhysiologicalPsychology 71:1–25.
Marler, P. 1996. “Social cognition: Are primatessmarter than birds?” In Current ornithology, Volume13, ed. V. Nolan Jr. and E.D. Ketterson, pp. 1–32.New York: Plenum Press.
Matsuzawa, T. 1985. Use of numbers by a chimpanzee.Nature 315:57–59.
Medina, L., and A. Reiner. 2000. Do birds possess ho-mologues of mammalian primary visual, somatosen-sory, and motor cortices? Trends in Neurosciences23:1–12.
Menzel, E.W., Jr., and C. Juno. 1982. Marmosets(Saguinus fuscicollis): Are learning sets learned?Science 217:750–752.
Merriman, W.E. 1991. The mutual exclusivity bias inchildren’s word learning: A reply to Woodward andMarkman. Developmental Review 11:164–191.
13 / Grey Parrot Cognition and Communication 143

Miles, H.L. 1983. “Apes and language.” In Language inprimates, ed. J. de Luce and H.T. Wilder, pp. 43–61,New York: Springer-Verlag.
Mowrer, O.H. 1950. Learning theory and personalitydynamics. New York: Ronald Press.
Page, S.C., S.H. Hulse, and J. Cynx. 1989. Relativepitch perception in the European starling (Sturnusvulgaris): Further evidence for an elusive phenome-non. Journal of Experimental Psychology: AnimalBehavior Processes 15:137–146.
Peek, F.W. 1972. An experimental study of the territo-rial function of vocal and visual displays in the malered-winged blackbird (Agelaius phoeniceus).Animal Behaviour 20:112–118.
Pepperberg, I.M. 1981. Functional vocalizations by anAfrican grey parrot (Psittacus erithacus). Zeitschriftfür Tierpsychologie 55:139–160.
Pepperberg, I.M. 1983. Cognition in the African greyparrot: Preliminary evidence for auditory/vocalcomprehension of the class concept. AnimalLearning & Behavior 11:179–185.
Pepperberg, I.M. 1985. Social modeling theory: A pos-sible framework for avian vocal learning. Auk102:854–864.
Pepperberg, I.M. 1987a. Acquisition of the same/differ-ent concept by an African grey parrot (Psittacuserithacus): Learning with respect to categories ofcolor, shape, and material. Animal Learning &Behavior 15:423–432.
Pepperberg, I.M. 1987b. Evidence for conceptual quan-titative abilities in the African grey parrot: Labelingof cardinal sets. Ethology 75:37–61.
Pepperberg, I.M. 1987c. “Interspecies communication:A tool for assessing conceptual abilities in the greyparrot (Psittacus erithacus).” In Language, cogni-tion, and consciousness: Integrative levels, ed. G.Greenberg and E. Tobach, pp. 31–56. Hillsdale, NJ:Erlbaum.
Pepperberg, I.M. 1988a. Acquisition of the concept ofabsence by an African grey parrot: Learning with re-spect to questions of same/different. Journal of theExperimental Analysis of Behavior 50:553–564.
Pepperberg, I.M. 1988b. An interactive modeling tech-nique for acquisition of communication skills:Separation of “labeling” and “requesting” in a psit-tacine subject. Applied Psycholinguistics 9:59–76.
Pepperberg, I.M. 1990a. “An investigation into the cog-nitive capacities of an African grey parrot (Psittacuserithacus).” In Advances in study of behavior, ed.P.J.B. Slater, J.S. Rosenblatt, and C. Beer, pp.357–409. New York: Academic Press.
Pepperberg, I.M. 1990b. Cognition in an African greyparrot (Psittacus erithacus): Further evidence forcomprehension of categories and labels. Journal ofComparative Psychology 104:41–52.
Pepperberg, I.M. 1992. Proficient performance of aconjunctive, recursive task by an African grey parrot(Psittacus erithacus). Journal of ComparativePsychology 106:295–305.
Pepperberg, I.M. 1994a. Evidence for numerical compe-tence in an African grey parrot (Psittacus erithacus).Journal of Comparative Psychology 108:36–44.
Pepperberg, I.M. 1994b. Vocal learning in African greyparrots: Effects of social interaction. Auk111:300–313.
Pepperberg, I.M. 1996. “Categorical class formation byan African grey parrot (Psittacus erithacus).” InStimulus class formation in humans and animals, ed.T.R. Zentall and P.R. Smeets, pp. 71–90. Amster-dam: Elsevier.
Pepperberg, I.M. 1999. The Alex studies. Cambridge,MA: Harvard University Press.
Pepperberg, I.M. 2002a. Cognitive and communicativeabilities of grey parrots. Current Directions inPsychological Science 11:83–87.
Pepperberg, I.M. 2002b. In search of King Solomon’sring: Cognition and communication in grey parrots.Brain, Behavior & Evolution 59:54–67.
Pepperberg, I.M., and M.V. Brezinsky. 1991. Relationallearning by an African grey parrot (Psittacus eritha-cus): Discriminations based on relative size. Journalof Comparative Psychology 105:286–294.
Pepperberg, I.M., L.I. Gardiner, and L.J. Luttrell. 1999.Limited contextual vocal learning in the grey parrot(Psittacus erithacus): The effect of co-viewers onvideotaped instruction. Journal of ComparativePsychology 113:158–172.
Pepperberg, I.M., and M.A. McLaughlin. 1996. Effectof avian-human joint attention on allospecific vocallearning by grey parrots (Psittacus erithacus).Journal of Comparative Psychology 110:286–297.
Pepperberg, I.M., J.R. Naughton, and P.A. Banta. 1998.Allospecific vocal learning by grey parrots (Psit-tacus erithacus): A failure of videotaped instructionunder certain conditions. Behavioural Processes42:139–158.
Pepperberg, I.M., R.M. Sandefer, D. Noel, and C.P.Ellsworth. 2000. Vocal learning in the grey parrot(Psittacus erithacus): Effect of species identity andnumber of trainers. Journal of Comparative Psy-chology 114:371–380.
Pepperberg, I.M., and D. Sherman. 2000. Proposed useof two-part interactive modeling as a means to in-crease functional skills in children with a variety ofdisabilities. Teaching and Learning in Medicine12:213–220.
Pepperberg, I.M., and H.A. Shive. 2001. Hierarchicalcombinations by a grey parrot (Psittacus erithacus):Bottle caps, lids, and labels. Journal of ComparativePsychology 115:376–384.
144 Manual of Parrot Behavior

Pepperberg, I.M., and Wilcox, S.E. 2000. Evidence fora form of mutual exclusivity during label acquisitionby grey parrots (Psittacus erithacus)? Journal ofComparative Psychology 114:219–231.
Pepperberg, I.M., M.R. Willner, and L.B. Gravitz.1997. Development of Piagetian object permanencein a grey parrot (Psittacus erithacus). Journal ofComparative Psychology 111:63–75.
Premack, D. 1978. “On the abstractness of human con-cepts: Why it would be difficult to talk to a pigeon.”In Cognitive processes in animal behavior, ed. S.H.Hulse, H. Fowler, and W.K. Honig, pp. 421–451.Hillsdale, NJ: Erlbaum.
Premack, D. 1983. The codes of man and beast.Behavioral and Brain Sciences 6:125–167.
Rice, M.L., A.C. Huston, R. Truglio, and J. Wright.1990. Words from Sesame Street: Learning vocabu-lary while viewing. Developmental Psychology26:421–428.
Rozin, P. 1976. “The evolution of intelligence and ac-cess to the cognitive unconscious.” In Progress inpsychobiology and physiological psychology,Volume 6, ed. J.M. Sprague and A.N. Epstein, pp.245–280. New York: Academic Press.
Salomon, G. 1977. Effects of encouraging Israeli moth-ers to co-observe Sesame Street with their five-yearolds. Child Development 48:1146–1151.
Savage-Rumbaugh, E.S. 1984. “Acquisition of func-tional symbol use in apes and children.” In Animalcognition, ed. H.L. Roitblat, T.G. Bever, and H.S.Terrace, pp. 291–310. Hillsdale, NJ: Erlbaum.
Savage-Rumbaugh, S., J. Murphy, R.A. Sevcik, K.E.Brakke, S.L. Williams, and D.M. Rumbaugh. 1993.Language comprehension in ape and child.Monographs of the Society for Research in ChildDevelopment 233:1–258.
Schusterman, R.J., and R.C. Gisiner. 1988. Artificiallanguage comprehension in dolphins and sea lions:The essential cognitive skills. Psychological Record38:311–348.
Schusterman, R.J., and K. Krieger. 1986. Artificial lan-guage comprehension and size transposition by a
California sea lion (Zalophus californianus). Jour-nal of Comparative Psychology 100:348–355.
Skinner, B.F. 1957. Verbal behavior. New York:Appleton-Century-Crofts.
Smith, D.G. 1979. Male singing ability and territorialintegrity in red-winged blackbirds (Agelaiusphoeniceus). Behaviour 68:191–206.
Smith, W.J. 1997. “The behavior of communicating,after twenty years.” In Perspectives in ethology,Volume 12, ed. D.H. Owings, M.D. Beecher, andN.S. Thompson, pp. 7–53. New York: Plenum Press.
Soja, N., S. Carey, and E. Spelke. 1985. Constraints onword learning. Paper read at the biennial conventionof the Society for Research in Child Development,April, Toronto, CA.
Starkey, P., and R.G. Cooper. 1995. The development ofsubitizing in young children. British Journal ofDevelopmental Psychology 13:399–420.
St. Peters, M., A.C. Huston, and J.C. Wright. 1989.Television and families: Parental coviewing andyoung children’s language development, social be-havior, and television processing. Paper presented atthe Society for Research in Child Development,April, Kansas City, KS.
Todt, D. 1975. Social learning of vocal patterns andmodels of their applications in grey parrots.Zeitschrift für Tierpsychologie 39:178–188.
Trick, L., and Z. Pylyshyn. 1989. Subitizing and theFNST spatial index model. University of Ontario,COGMEM #44. [Based on paper presented at the30th Psychonomic Society Meeting, Atlanta, GA.]
Watkins, B., S. Calvert, A. Huston-Stein, and J.C.Wright. 1980. Children’s recall of television mate-rial: Effects of presentation mode and adult labeling.Developmental Psychology 16:672–674.
Weisman, R., and L. Ratcliffe. 1989. Absolute and rel-ative pitch processing in black-capped chickadees,Parus atricapillus. Animal Behaviour 38:685–692.
Zentall, T.R., ed. 1993. Animal cognition: A tribute toDonald A. Riley. Hillsdale, NJ: Erlbaum.
13 / Grey Parrot Cognition and Communication 145

Some parrots behave in friendly, sociable wayswhile others are flat-out unapproachable. Someparrots entertain themselves for hours in theircages while others scream incessantly. Observingthis kind of behavioral variability leads many ofus to ask some very important questions, such as,Why do parrots behave the way they do? Howshould we expect them to behave? Can they learnto behave as pets? Knowing the answers to thesequestions can make the difference between life-long success and failure to thrive for parrots incaptivity, particularly in our homes. However, tounderstand, predict, and change behavior we firstneed to know how it works.
BEHAVIOR ANALYSIS
Learning and behavior have been studied as a nat-ural science within the field of psychology forwell over a century. This science has come to beknown as behavior analysis. Pierce and Cheney(2004, p. 420) provide the following contempo-rary definition: “Behavior analysis is a compre-hensive experimental approach to the study of thebehavior of organisms. Its primary objectives arethe discovery of principles and laws that governbehavior, the extension of these principles overspecies, and the development of an applied tech-nology.”
Behavior can be investigated at many differentlevels of analysis, as with genetics, neurology,and pharmacology. The focus of behavior analysisis the environmental determinants of behavior,from which behavioral learning theory has beenformulated and continues to be refined. (The term“theory” is used technically to mean an estab-lished explanation accounting for known facts or
phenomena, as opposed to the non-technicalusage, which means an unproven guess or per-sonal opinion. Other theories of learning and be-havior are named according to their particular fo-cuses, such as cognitive theory and psychody-namic theory.)
Behavioral learning theory explains a secondkind of selection by consequences first recog-nized in natural selection (Skinner 1981).Whereas natural selection is the process of func-tional genomic adaptation of an entire speciesacross generations, learning is the process offunctional behavioral adaptation of a single indi-vidual within its lifetime. The two keystones oflearning theory are (1) learning is largely deter-mined by external environmental influences, and(2) the laws of learning are general in nature, tran-scending species and situations. In its simplestterms, according to each individual’s experienceinteracting with his or her environment, behaviorsthat “work” are repeated and behaviors that don’twork are modified or suppressed.
Over the last 60 years, the applied branch ofbehavior analysis has matured into a highly ef-fective technology to solve practical, real-worldbehavior problems. Its widespread applicabilitycontinues to expand, having already been demon-strated across seemingly diverse areas such asspecial education, industrial safety, and animalmanagement. Other names such as operant con-ditioning, behavior modification, and behaviortherapy refer to the same basic interventionstrategies; however, applied behavior analysis in-cludes a more rigorous and comprehensivecourse of action involving the scientific proce-dures of hypothesis generation (functional as-
147
14Behavior Analysis
and Parrot Learning
S.G. Friedman, Steve Martin, and Bobbi Brinker

sessment), testing (functional analysis), and eval-uation (measurement). Intervening to change be-havior in this systematic way allows us to solvebehavior problems with a high degree of preci-sion, replicability, and accountability. In thischapter the tools and techniques of applied be-havior analysis are discussed in reference to thecare and management of captive parrots, particu-larly those kept as pets.
THE ABCs OF BEHAVIOR
The fundamental unit of behavior analysis is thethree-term contingency, described by Skinner (ascited in Chance 1998, p. 38): “An adequate for-mulation of the interaction between an organismand its environment must always specify threethings: (1) the occasion upon which a responseoccurs, (2) the response itself, and (3) the . . . con-sequences.”
These three terms comprise the behaviorABCs: antecedent, behavior, and consequence.Behavior does not occur independently of the en-vironmental events that surround it, thereforethere is never just behavior. The smallest elementof behavior that can be meaningfully analyzed isan ABC unit, described further in the followingsections.
Antecedents
Antecedents are the stimuli, events, and condi-tions that immediately precede a behavior. Theyare functionally related to the behavior that fol-lows if the appearance of the behavior depends onthe presence of the antecedent stimuli. Antece-dents set the occasion for behavior rather thancause it. For example, an open hand presented toa parrot can be an antecedent for either steppingup or running away, depending on the conse-quences the parrot experienced for doing so in thepast. Thus, we can increase the probability that aparticular behavior will occur by carefully arrang-ing antecedents, but ultimately the animal makesa choice to behave as we have planned or in someother way. By definition, operant (i.e., voluntary)behavior acknowledges the individual’s power tooperate on his or her environment.
Behavior
In applied behavior analysis, behavior is what anorganism does that can be measured. The mainfocus is overt behaviors that can be operationally
defined and unambiguously observed. Birds dojump off perches, hang upside down, rouse theirfeathers, bite hands, ring bells, pin their eyes, andflare their tails. These behaviors can be unam-biguously observed and measured according todifferent dimensions of interest such as fre-quency, rate, duration, and intensity. Covert beha-viors, including thinking and feeling, are privateevents that can only be observed and measured bythe individual engaging in it. This makes parrots’covert behaviors impractical, if not impossible,behavior-changing targets at this time.
Psychological constructs, such as intelligence,neurosis, and confidence, are not behaviors. Gallet al. (2003, p. 621) define constructs in this way:“A concept that is inferred from commonalitiesamong observed phenomena that can be used toexplain these phenomena. In theory development,a concept that refers to a structure or process thatis hypothesized to underlie particular observedphenomena.”
Thus, constructs are what we think may be oc-curring inside an organism that explains why it isacting in particular ways. We don’t really perceiveintelligence, neurosis, or confidence with oursenses. What we perceive are overt behaviors suchas talking in context, plucking feathers, and goingto strangers without hesitation. Constructs are bestthought of as placeholders for internal processesas yet unknown involving nerves, brains, hor-mones, and muscles (Manning & Stamp Dawkins,1992). Unfortunately, constructs all too easilycome to be thought of as real entities residingsomewhere in the brain. This leads to what Gould(1981) calls the fallacy of reification and expla-natory fictions. The fact remains that even whenthe underlying physiological processes that sup-port behavior are understood, no account ofbehavior can be complete without the behavior-environment factor.
Vague labels, such as sweet, spoiled, and jeal-ous, are also not behavior. Labels typically de-scribe what people think a bird is rather than whatit does. For example, the label “is sweet” tells usnothing about the behavior we want to train ormaintain. We can’t train a bird to do sweet but wecan train a bird to step up for all family members.To improve our ability to understand, predict, andchange parrots’ behavior, the focus should be onobservable, measurable behaviors, not constructsor vague labels.
148 Manual of Parrot Behavior

Consequences
Consequences are the stimuli, events, and condi-tions that occur after a behavior and influence theprobability that the behavior will occur again.There is a functional relation between a specificbehavior and a consequence if the appearance ofthe consequence depends on the behavior occur-ring first. Social attention, items and activities,sensory feedback and escape from aversive eventsare all consequences that affect parrot behavior.Consequences are nature’s feedback about the ef-fectiveness of an individual’s behavior. In thisway, past consequences affect motivation for fu-ture behavior. This is the law of effect that states,“In any given situation, the probability of a be-havior occurring is a function of the conse-quences that behavior has had in that situation inthe past” (Chance 2003, p. 137).
Thus, parrots, like all animals, don’t just “suf-fer the consequences”—they learn from the con-sequences how to behave in the future, givensimilar antecedent circumstances. Learning byconsequences is a natural process that accountsfor behavior in both the free range and captivity.Even innate behavior (elicited automatically,without prior learning) is flexible according toconsequences. For example, although nest build-ing tends to be stereotypical within many species,we expect that birds improve in their abilities tobuild nests with experience.
FUNCTIONAL ASSESSMENT/ANALYSIS
The ABCs form the basis of an important toolcalled functional assessment, the hypothesis-generating phase of changing behavior. Aftercarefully observing and operationally definingthe target behavior (the one we want to under-stand, change, or both), functional assessment isthe next step in any behavior change program. Byhypothesizing the antecedents that set the occa-sion for a behavior and the consequences thatgive the behavior function, the chance of suc-cessfully changing behavior is greatly increased.For example, consider the following commonscenario: Sam Parrot has started refusing to steponto Grace’s hand from the top of his cage. Graceworries that Sam is trying to dominate her fromhis high perch, and she wonders if she shouldforce him down with a towel to show him who’sboss. It was suggested to her that she cut off the
cage legs so as not to trigger this innate responseagain.
A functional assessment of Sam’s prior step-upbehavior reveals a convincing alternative hypoth-esis to that posed by Grace: Sam refuses to stepup to avoid being locked in his cage, as indicatedby the following.
Setting: Sam Parrot is playing with his bell ontop of his cage.
A: Grace offers her hand.B: Sam steps up.C: Grace returns Sam to his cage.Prediction: Sam will step up less often in the
future.The hypothesis that Sam no longer steps up
from his cage top to avoid being locked in hiscage can be tested by changing the antecedents,the consequences, or both, and observing anyconcomitant changes in the frequency of Sam’sstep-up behavior. It is at this point that functionalassessment turns into functional analysis. Onepossible antecedent solution is to allow Sam ac-cess to the cage top only when there is sufficienttime for him to tire of being there. One possibleconsequence change is to offer a special treat asSam steps up and to have a special item in thecage to be discovered once he’s inside it.
The process of functional assessment allows usto generate highly specific and testable hypothe-ses about behavior-environment relations. Thequestion addressed with functional assessment isnot why does the bird behave this way, but ratherwhat valued consequence does the bird get by be-having this way; in other words, what’s the func-tion of the behavior? It is through changing an-tecedents and consequences that behaviorchanges. Since the environment in which captiveparrots live is largely controlled by their care-givers, changing parrot behavior is usually the re-sult of changing human behavior first.
THE PROBLEMS WITH DOMINANCE
With this foundation in place, we can better eval-uate two common misconceptions about behaviorthat have caused particular problems for parrotsand their owners. The first is that parrots arestrongly motivated by an innate drive or charactertrait to dominate their human caregivers. The sec-ond is that caregivers must establish and enforcesuperior rank over parrots to control them. Thesetwo misconceptions, and many others like them,
14 / Behavior Analysis and Parrot Learning 149

come to have a life of their own, independent ofsound scientific information about behavior. Theyappeal to conventional wisdom and our penchantfor quick fixes, but in the long run they pose seri-ous obstacles to appropriate learning solutionsand the behavioral health of captive parrots. Theimportant implications of these two fallacies arediscussed separately in the following sections.
Parrots and Dominance
Giving commands, following orders, and jockey-ing for position within linear social hierarchiesare common activities for most humans. Thesebehaviors are well supported by our educational,religious, sports, military, and corporate organi-zations throughout our lives. We are also prone toobserve, or think we observe, in other species thatwhich we most expect to see. This problem,known as observer-expectancy bias, is well docu-mented even among those who watch birds (seefor example, Balph & Balph 1983.) Perhaps thisaccounts for the widely held and persistent beliefamong parrot enthusiasts that parrots’ dominantnature impels them to refuse to step off cage tops(height dominance), to chase and bite humans andother animals while on the floor (floor domi-nance), to scream when the telephone is in use(phone dominance), and to lunge at feed doors(cage dominance). In the companion parrot arena,the different supposed forms of dominance thatparrots use to subjugate their caregivers goes on,ad infinitum.
In fact, even among scientists the term “domi-nance” is ambiguous and varies significantlyfrom report to report (an inherent problem withconstructs). In technical usage, dominance gener-ally describes some aspect of an animal’s priorityaccess to resources such as food, location, andmates, which is often achieved through agonisticcontrol of another animal. However, in Barrow’sAnimal Behavior Desk Reference (2001) there areseven different definitions of social dominance,including four subcategories, one of which hastwo subtypes. As reported by Barrow, “Hand(1986, p. 202) indicates that there is no agree-ment regarding how to define, or measure, socialdominance.”
To further complicate matters, Barnett (1981)suggests that “Dominance should be distin-guished from an animal’s superiority resultingfrom its being in its own territory. Dominance
should also be distinguished from being a leader”(p. 633). Moreover, a critical omission in manydiscussions of dominance is variables such aschanging motivations, contexts, and prior learn-ing history (see, e.g., the influence of context,Cloutier et al. 1995). This lack of scientific con-sensus about what dominance is should call intoquestion its usefulness for understanding andmanaging companion parrot behavior (as is cur-rently being done regarding the behavior ofwolves and dogs; see Mech 1999, 2000, and VanKerkhove 2004).
Although some people support the validity ofthe dominance model applied to pet parrots basedon free-range behavior, social hierarchies amongwild parrots have not been well documented.Other people support the validity of the domi-nance model based on the unnatural demands ofthe captive environment. No studies could be lo-cated on dominance relationships between parrotsand humans. One study, of a flock of 12 group-housed Cockatiels (Nymphicus hollandicus),lends support to the hypothesis that males tend tohold higher dominance ranks than females, basedon well-operationalized definitions of aggression,submission, and rank (Seibert & Crowell-Davis2001). These findings are consistent with thosereported by Weinhold with aviary-kept Blue-fronted Amazon Parrots (Amazona aestiva) (ascited in Seibert & Crowell-Davis 2001). Seibertand Crowell-Davis discussed several limitationsof their study that restrict the extent to whichthese conclusions can be generalized to otherflock-housed Cockatiels: Only one flock of 12Cockatiels was investigated; the genetic related-ness of the birds was unknown; and the data werecollected during mate selection and breeding sea-son. Further research is needed to assess the ex-tent to which these findings generalize to parrotskept as pets and to parrot-human interactions. Theimplications, if any, to companion parrot behaviormanagement appear to be remote.
The ubiquitous dominance interpretation ofcompanion parrot behavior has other problems aswell. First, the expectation that pet parrots aremotivated to win superior rank over their care-givers in some pecking order can serve as a self-fulfilling prophecy. As mentioned previously,when people have expectations about another in-dividual’s behavior, they act differently and tendto get what they expect. Second, since dominance
150 Manual of Parrot Behavior

is thought to be an invisible drive or charactertrait inside the bird, a dominance problem is a badbird problem. This provides a convenient excusefor getting rid of the bird rather than taking re-sponsibility for the circumstances (antecedentsand consequences) under which these behaviorsarise. Third, the dominance explanation predis-poses many caregivers to use forceful manage-ment strategies in order to counterdominate theirbirds and win the struggle for alpha organism.Fourth, the dominance explanation ends thesearch for proximal, environmental causes andsolutions. The very process of labeling a problemprovides a false sense of closure when in fact ithas only provided a name. Thus, the essentialprocesses of functional assessment and solutionbuilding are prematurely terminated and theknown and remediable relations between behav-ior and environment remain unexplored (Chance1998).
THE CASE FOR EMPOWERMENT
When the dominance construct is extended intoparrot management practices it takes the form of“show them who’s boss” and “never let themmake any important decisions.” These sugges-tions are ubiquitous in both popular magazinesand professional veterinary literature. However,much to the contrary, scientific evidence indi-cates that animals tend to thrive in environmentsin which they are not subjugated but rather havecontrol over significant life events (Schwartz etal. 2002). Given knowledge of how behaviorworks and sound training skills, parrots can beempowered instead of overpowered, without alter-ing our standards for good companion behavior.
One important demonstration of the emotionalgain that comes from having control over one’senvironment is experiments conducted by Watsonwith two groups of human babies only threemonths old (as cited in Schwartz et al. 2002).Under the pillows of the first group was a switchthat operated a mobile whenever the infantsturned their heads. The babies in the secondgroup had no control over their mobiles, althoughtheir mobiles automatically moved as much as thefirst group’s did. As expected according to the lawof effect, the frequency of head movements inonly the first group increased since doing so wasreinforced by the mobiles’ movement (i.e., themobiles’ movement depended on what the babies
did). However, other differences were observed inthe two groups of babies that were very surpris-ing. Initially, both groups of babies responded tothe moving mobiles by cooing and smiling, a rea-sonable measure of well-being. These happy re-sponses continued throughout the experiment forthose babies who controlled their mobiles but forthe babies who did not control their mobiles, thecooing and smiling quickly stopped. Apparently,controlling one’s consequences explains, at leastin part, what makes them reinforcing.
Another relevant line of research is the freefood phenomenon, also known as contrafreeload-ing. With contrafreeloading, animals choose toperform a learned response to obtain reinforcerseven when the same reinforcers are freely avail-able. For example, given a choice between work-ing for food and obtaining food for free, animalstend to choose to work, often quite hard, with abowl of free food placed right next to them. Thisphenomenon has been replicated with rats, mice,chickens, pigeons, crows, cats, gerbils, Siamesefighting fish, and humans (Osborne 1977); star-lings (Inglis & Ferguson 1986); AbyssinianGround Hornbills and Bare-faced Curassows(Gilbert-Norton, 2003); and captive parrots(Coulton et al. 1997). There are several interest-ing hypotheses explaining why this phenomenonoccurs. Contrafreeloading behavior may be moti-vated by innate foraging behaviors that are other-wise frustrated in captivity; animals may be en-gaging in information-seeking behaviors as theywork to predict the location of optimal foodsources; or they may be responding to the addi-tional reinforcement provided by stimuluschanges when one works for food, such as thesound of a hopper. Nonetheless, animals’ prefer-ence to behave in ways that impact their environ-ment is demonstrated once again. Animals arebuilt to behave, not to be passive.
A third area of scientific inquiry called learnedhelplessness adds additional support to the theorythat personal control over significant environ-mental events is necessary for animals to behavehealthfully. This phenomenon further demon-strates that a lack of control can have pathologicaleffects including depression, learning disabilities,emotional problems (Maier & Seligman 1976),and suppressed immune system activity (Lauden-slager et al. 1983). Learned helplessness occurswhen an animal with no prior escape history is
14 / Behavior Analysis and Parrot Learning 151

prevented from escaping severe, aversive stimuli.Under this condition, the animal eventually givesup attempting to escape and remains passive.Later when escape is made blatantly possible theanimal does not make the expected escape re-sponse, as if helpless. This research has beenreplicated with cockroaches (Brown et al. 1988),dogs, cats, monkeys, children, and adults (Over-mier & Seligman 1967). Further, Seligman’s(1990) research suggests that we can “immunize”learners from the effects of lack of control by pro-viding them with experiences in which their be-havior is effective, that is, in which they controltheir own outcomes. In this way, the effects of ex-posure to uncontrollable aversive stimuli, whichis inevitable in all our lives to some degree, can beminimized.
Based on these three related research areas, it isvery possible that a lack of control explains some,if not many, of the pathological behaviors we seein parrots such as self-mutilation, mate killing,and phobias. To the greatest extent possible, par-rots should be empowered to make important de-cisions, such as when to exit or enter their cagesor go on and off their caregiver’s hands. Parrots soempowered will likely experience greater behav-ioral and emotional health in captivity.
TOOLS AND TECHNIQUES FORBEHAVIOR CHANGE
Although a parrot’s biological history often takescenter stage, much of the time behavior problemsare the result of its learning history in captivity,which is composed of all the environmental eventsthat have affected the parrot’s behavior up to thepresent. When one stops to think about it, behav-ior problems can be reduced to two simple cate-gories: not doing something enough (e.g., step-ping up, staying put, and eating pellets) and doingsomething too much (e.g., screaming, biting, andchewing woodwork). Our responsibility is to suc-
cessfully increase desirable behavior and decreaseproblem behavior using the most positive, least in-trusive methods possible. Table 14-1 describes thissimple behavior support model, after which themajor strategies that compose the teaching tech-nology of behavior analysis are discussed.
Changing Behavior with Antecedent Strategies
There are three general categories of antecedentsthat precede behavior: discriminative stimuli, set-ting events, and establishing operations. A dis-criminative stimulus (SD, pronounced ess-dee), or“cue”, belongs to a special class of antecedentsthat signal that a certain response will be rein-forced (among all possible responses). A stimulusor event becomes an SD by being repeatedly pres-ent when a response is reinforced. For example,when the doorbell rings, we open the door ratherthan pick up the phone, hurry to the exits, orgather up our school books. We do so because inthe past, the doorbell has been consistently pairedwith reinforcement for opening the door and notfor those other behaviors. The strength of a stim-ulus to cue a behavior is related to the strength ofthe reinforcer that follows the behavior. For somebirds a perching stick comes to signal that step-ping up will be reinforced with activities outsidethe cage. A ringing phone signals that saying“hello” will be reinforced with gales of laughter,and a person approaching a cage with a bowl inhand signals that coming to the feed door will bereinforced with food.
Problem behaviors are cued by discriminativestimuli as well. The very same cues just describedcan just as easily signal that biting will be rein-forced if we remove the perching stick, return thephone to its base, or hastily install the food bowland retreat fast. Cues don’t only come from peo-ple. The setting sun, cage covers, and microwavescan function as cues for particular behavior too.The approach of one of the author’s (Friedman)
152 Manual of Parrot Behavior
Table 14-1 A simple model of behavioral support
Not enough behavior Too much behavior
Goal Increase/maintain current frequency Decrease/suppress current frequencyAntecedent changes Setting events Setting events
Establishing operations Establishing operationsAdding a cue Removing a cue
Consequence changes Reinforcement Punishment

Shih Tzu pups cued (antecedent) her UmbrellaCockatoo to call raucously (behavior), which wasthen reinforced by the Shih Tzu’s howling (conse-quence); thus, the frequency of the bird’s raucouscalling increased. (Turning the raucous parrot callinto a cue for the dog to return to its owner for abiscuit took care of the problem.)
Setting events also influence behavior. Theyare the context, conditions, or situational influ-ences that affect the contingencies that follow.Hands held too low, noisy environments, cagearrangements, and the number of people in theroom are all potential setting events that can af-fect the way in which a bird responds to an of-fered hand. The relation between setting eventsand problem behavior should be considered care-fully, as the setting is often one of the easiestthings to change.
Establishing operations (Michael 1982) tem-porarily alter the effectiveness of consequences.As further explained by Kazdin (2001), “Motiva-tional states, emotions, and environmental eventsare establishing operations because they momen-tarily alter the effectiveness of the consequencethat may follow behavior and influence the fre-quency of the behavior” (p. 454). The effective-ness of a consequence to increase the frequencyof a behavior is often related to its availability
(i.e., excess or deficit). For example, hunger andsatiation alter the reinforcing strength of foodtreats in opposite ways: A few sunflower seedsmay be a highly motivating consequence to a birdthat rarely has access to them but not motivatingat all to a bird that has unlimited access to themevery day.
Establishing operations can be used to alter thestrength of other non-food reinforcers as well. Forexample, a bird may be more motivated to stay ona play gym after some quality time with a favoritecaregiver. Chasing the family cat may be less re-inforcing after an energetic training session, andstepping onto a hand may be more reinforcingwhen the bird is on the floor. Table 14-2 lists ad-ditional examples of the many ways antecedentscan be carefully arranged to decrease the occur-rence of problem behaviors and increase desirablebehaviors.
Changing Behavior with ConsequenceStrategies
At the heart of good training is two-way commu-nication that results from the planned arrange-ment of contingencies. Contingencies are theif/then dependencies between behavior and itsconsequences. For example, to increase the fre-quency of quiet vocalizations we can offer the fol-
14 / Behavior Analysis and Parrot Learning 153
Table 14-2 Examples of antecedent behavior change strategies
Type of antecedent event Antecedent technique Problem behavior Application
Discriminative Add a cue for the right Lunges when cage is Cue bird to go to a farstimulus behavior. serviced. perch before servicing
cage.Remove a cue for the Bites shirt buttons. Don t-shirt before
problem behavior. handling bird.Setting events Decrease the response Refuses to go to others Set bird on counter
effort for the right from preferred person’s before offering non-behavior. shoulder. preferred hand.
Increase the response Chews door frame. Move play tree to center effort for the problem of room.behavior.
Establishing Increase reinforcer strength Resists returning to cage. Remove treat from diet operations for the right behavior. except when bird
enters cage. Decrease reinforcer strength Jumps off T-stand. Offer undivided attention
for the problem behavior. for 10 minutes before T-stand.

lowing contingencies: if the parrot vocalizes qui-etly, then a preferred person approaches, but if theparrot vocalizes loudly, then no attention follows.Unfortunately, the opposite contingencies areoften provided (i.e., if the parrot vocalizes loudly,then a preferred person approaches), inadver-tently giving function to problem behaviors likeexcessive screaming.
Contingencies empower learners to choosehow to operate on their environment. When a per-son offers a hand to a parrot, it chooses to step upor not depending on past consequences. If theparrot runs away, it communicates clearly thatpast consequences for stepping up are not suffi-ciently motivating at that moment to repeat thebehavior. Rather than force the bird to comply,this is the time to consider ways to alter the an-tecedents and consequences to change the behav-ior. The question to ask before making any requestof a parrot is, “Why should he”? and the answerlies in the consequences we consistently provide.
There are two broad categories of consequencetechniques: Reinforcement strengthens behaviorand punishment weakens it. Although the termsmean many different things in common usage,they have specific, technical meaning in the sci-ence of behavior that maximizes their usefulness.Behavioral strength can refer to different re-sponse dimensions such as frequency, rate, dura-tion, intensity, topography (form, e.g., a footbarely lifted off a perch versus a foot raised highin the air), and latency (the time lag between thecue and the onset of the behavior). To simplify thediscussion, frequency of behavior, the most often
used measure of behavioral strength, is discussedthroughout this section.
REINFORCEMENT
When a behavior doesn’t occur often enough wecan increase its frequency with reinforcement.Reinforcement is the procedure of contingentlyproviding consequences for a behavior that in-crease or maintain the frequency of that behavior.Positive reinforcement, sometimes called rewardtraining, is a reinforcement procedure in which abehavior is followed by the presentation of astimulus. Negative reinforcement, sometimescalled escape training, is a reinforcement proce-dure in which a behavior is followed by the re-moval of a stimulus. Technically, the terms “posi-tive” and “negative” refer only to the operation ofpresenting (+) or removing (�) a stimulus that, inthe case of reinforcement, functions to increase ormaintain the behavior it follows. However, it isgenerally accurate and often easier to assume thatpositive reinforcers have “positive” value to thelearner (something it works to get) and negativereinforcers have “negative” value (something itworks to escape). Examples of positive and nega-tive reinforcement are in Table 14-3.
Although both positive and negative reinforce-ments increase or maintain behavior, they can af-fect the manner in which a learner engages intraining quite differently: To get positive rein-forcers, learners often enthusiastically exceed theminimum effort necessary to gain them. Alterna-tively, to escape negative reinforcers, learnerstend to offer only the minimum behavior neces-
154 Manual of Parrot Behavior
Table 14-3 Examples of positive and negative reinforcement
Antecedent Behavior Consequence Future behavior
Positive Grace asks Sam to Sam hops onto Grace adds food Sam goes to perch reinforcement go to the back perch bowl through more (reward) perch feed door
Grace is working Sam nips her hand Grace scratches Sam nips hand on her computer Sam’s head more
Negative Grace offers left Sam steps up Grace puts down Sam steps up more reinforcement hand with towel towel(escape) in right hand
Grace offers perch Sam pulls back Grace removes Sam steps on perch while holding foot to step on thumb as Sam more Sam’s toes with perch steps downher thumb

sary to avoid the aversive stimuli. Moreover, theuse of aversive procedures has been repeatedlydemonstrated to increase learners’ escape behav-iors, aggression, apathy, and generalized fear(Azrin & Holz 1966). These side effects are detri-mental and are discussed further in the section onpunishment. As a result, positive reinforcement isthe gold standard of behavior-change procedures.It is powerful, effective, and is not associated withaversive fallout (Sulzer-Azaroff & Mayer 1991).
Factors Affecting Reinforcement
Several important factors affect reinforcement.The first is contingency, the degree to which de-livery of the reinforcer depends on the behavioroccurring first. Consistent pairing of the behaviorand the reinforcer clearly communicates the con-tingency between a behavior and a reinforcer.Without consistency, it’s difficult for a parrot tomake the connection between the two events,which slows down learning and produces incon-sistent behavior.
Contiguity refers to the temporal closeness ofthe behavior and the reinforcer. Reinforcers thatare delivered immediately after the behavior com-municate the contingency most clearly. Lattal(1995) demonstrated the importance of timing toeffective reinforcement with pigeons learning topeck a disk. With just a ten-second delay beforedelivering the food reinforcer, the pigeons neverlearned to peck the disk after 40 days of one-hourtraining sessions. When the delay was reduced toone second, the pigeons learned to peck the diskin less than 20 minutes.
Certain characteristics of the reinforcers suchas type and magnitude also affect reinforcement.Simmons (1924) found that rats reinforced at theend of a maze with bread and milk ran signifi-cantly faster than those reinforced with sunflowerseeds. In two studies comparing frequency andmagnitude, Schneider (1973) and Todorov et al.(1984) found that small, frequent reinforcerstended to be more effective than large, occasionalones. Research continues on the many other fac-tors that affect reinforcement such as task charac-teristics, task difficulty, relative availability, andlearning history.
Amid these general factors, individual differ-ences should be carefully considered when ar-ranging contingencies for desirable behavior. Aconsequence that is reinforcing to one parrot may
be neutral or aversive to another. Regardless ofthe teacher’s intentions, the proof of reinforce-ment is in the strength of the resulting behavior.Only by watching the data, the parrot’s behavior,can we know the extent to which it has been rein-forced. To determine an individual parrot’s rein-forcers, one can observe the bird’s favorite items,foods, activities, people, sounds, and locations.Establishing new reinforcers, a process discussedin the next section, keeps the list growingthroughout a learner’s lifetime.
Establishing New Reinforcers
The enormous degree of behavioral flexibility in-herent in many species is related to the capricious-ness of the environments in which they live.Indeed, if the environment remained constant, andtherefore predictable, all the behavior we wouldever need to survive could be genetically transmit-ted and elicited reflexively by particular triggeringstimuli. Instead, for parrots, as with humans,learning is the rapid-adaptation system that allowsthem to meet the demands of an unpredictable en-vironment in constant change. This extraordinarybehavioral flexibility includes the process bywhich neutral stimuli become reinforcers, calledsecondary or conditioned reinforcers.
Secondary reinforcers, such as praise, favoriteperches, and the sound of a clicker or whistle, arepreviously neutral stimuli that acquire their rein-forcing value by repeated pairing with existing re-inforcers. Primary, or unconditioned, reinforcers,such as food, water, and relief from heat or cold,are automatically reinforcing; that is, they requireno prior pairing or experience to function as be-havior increasing consequences. Primary rein-forcers are related to basic survival functions,which makes them a good starting point for con-ditioning secondary reinforcers.
Primary and secondary reinforcers have differ-ent advantages and disadvantages in the contextof training (Chance 2003). On one hand, primaryreinforcers are generally quite powerful and theyare not dependent on their association with otherreinforcers; but, they are few in number and moresusceptible to a temporary loss of effectivenessdue to satiation. For most parrots, the first fewsunflower seeds will be more motivating (astronger reinforcer) than the last few. On the otherhand, secondary reinforcers tend to hold theirvalue longer (satiate slower) and they can be de-
14 / Behavior Analysis and Parrot Learning 155

livered with less disruption, better contiguity, at agreater distance, and in a wider variety of situa-tions. However, secondary reinforcers tend to besomewhat weaker than primary reinforcers andtheir effectiveness relies on being paired withother reinforcers, at least some of the time. Bothkinds of reinforcers, in the greatest possible num-ber, add power to a trainer’s toolbox and increasethe quality of life for companion parrots.
Schedules of Reinforcement
Schedules of reinforcement are the rules that de-termine which particular instance of behavior willbe reinforced. Although it can be a complicatedtopic and beyond the scope of this chapter, thethree simple schedules that are most important tounderstanding and managing parrot behavior arediscussed briefly here. They are continuous, inter-mittent, and extinction schedules.
A continuous reinforcement (CRF) schedule isone in which each and every instance of the be-havior is reinforced (1:1). Given this perfect con-sistency, CRF provides the clearest communica-tion to the learner about what behavior is beingreinforced. As a result, the CRF schedule pro-duces rapid learning and is recommended for sta-bilizing and increasing existing behaviors andteaching new behaviors (Sulzer-Azaroff & Mayer1991).
At the other end of the spectrum is extinction(EXT), in which no instances of the behavior arereinforced (1:0). As the name suggests, when thereinforcer that previously maintained a behavioris withheld, the rate of that behavior predictablydecreases to pre-reinforcement levels (not neces-sarily total suppression).
Another category of simple schedules of rein-forcement is intermittent schedules. With inter-mittent schedules only some instances of the be-havior are reinforced, as opposed to all (CRF) ornone (EXT). Once a behavior is learned, an inter-mittent schedule produces persistent behavior inthe sense that it takes longer to extinguish thanbehaviors maintained on a continuous reinforce-ment schedule. Perhaps the clearest example ofthis partial reinforcement effect is the differentpatterns of responding that occur at vendingmachines versus slot machines. Given a continu-ous reinforcement history interacting with vend-ing machines, most people stop dropping coinsinto the slot after the first or second instance that
nothing comes out. But, given an intermittentreinforcement history with slot machines, mostpeople continue dropping coins into a slot per-sistently although rarely does anything ever comeout.
The partial reinforcement effect explains manyof the persistent misbehaviors we see in compan-ion parrots. The occasional time a lunge to thefeed door results in an escape to the top of thecage or a top decibel scream produces an exple-tive from a caregiver is often enough to produceenduring problem behaviors, due to the intermit-tent reinforcement schedule on which these be-haviors are maintained. The solution to each ofthese problems is not to ignore the behaviors bet-ter but to consider antecedent and consequencechanges to prevent them from happening in thefirst place and to reward alternative positive be-haviors instead.
All things considered, our birds benefit mostfrom our ability to catch them being good withthe highest possible rate of reinforcement. Oneimportant benefit of this approach is that the peo-ple who deliver dense schedules of reinforcementare more likely to become valued secondary rein-forcers themselves. A common axiom is, “You getwhat you reinforce.” Where problem behaviorsare concerned, what you get when you reinforceintermittently is persistent problems.
Implementing Reinforcement Effectively
Sulzer-Azaroff and Mayer (1991) present severalguidelines for maximizing the effectiveness of re-inforcement procedures that, when overlooked,account for ineffective behavior change programswith children. As these guidelines apply to alllearners and situations, they should be accountedfor carefully in our work with parrots. An adaptedlist of guidelines follows.
• Reinforce immediately until the behavior is oc-curring at a high steady rate, then gradually in-troduce delay.
• Reinforce every response initially until the be-havior is well established, and then graduallyintroduce intermittent reinforcement.
• Specify the conditions under which reinforcerswill be delivered (i.e., the cue and criterion forreinforcement) and incorporate other an-tecedent conditions (e.g., setting events and es-tablishing operations).
156 Manual of Parrot Behavior

• Deliver a quantity of reinforcers sufficient tomaintain the behavior without causing rapid sa-tiation.
• Select reinforcers appropriate to the individual.• Use a variety of reinforcers and reinforcing sit-
uations.• Provide opportunities to experience new rein-
forcers.• Eliminate, reduce, or override competing con-
tingencies.
Shaping New Behaviors
A behavior can’t be reinforced until it occurs,which could present a problem when one needs toteach a new behavior to a parrot. Waiting for thebehavior to occur by happenstance and capturingit with reinforcement might be an option, butsome behaviors occur too infrequently or not atall. The solution to this problem is known as shap-ing, technically called differential reinforcementof successive approximations. Shaping is the pro-cedure of reinforcing a graduated sequence ofsubtle changes toward the final behavior, startingwith the closest response the bird already does.The following are two examples of shaping plansfor teaching independent toy play and bathing.
Shaping Plan 1: Playing with Toys1. Final behavior: Independent toy play.2. Closest behavior bird already does: Looks at toy.3. Reinforcer for each approximation that meets
the criterion: Safflower seeds and praise.4. Tentative approximations:
a. Look at toyb. Move toward toyc. Touch beak to toyd. Pick up toy with beake. Touch foot to toyf. Hold toy with foot while manipulated with
beakg. Repeat previous approximation for longer
durations
Shaping Plan 2:Triggering the Bathing Response1. Final behavior: Step into shallow water dish.2. Initial behavior: Looks at water dish.3. Reinforcers for each approximation that meets
criterion: Applause and praise.4. Tentative steps:
a. Look at dishb. Face dish
c. Take a step toward dishd. Take two steps toward dishe. Walk up to dishf. Look at water in dishg. Lift foot next to dishh. Touch water in dish with footi. Step into dish with one footj. Step into dish with both feetk. Walk around in dish
Implementing a shaping procedure requiresnoticing the subtle, natural variation in the waybehaviors are performed within a response class(called an operant class). For example, a parrotnaturally lifts its foot a little differently every time(left or right, high or low, fast or slow, with toemovement or without, etc.). Typically this varia-tion is unimportant and it is simply classified asone behavior, or operant class, called lifting afoot. However, this subtle variation is exactlywhat allows us to shape a parrot to “wave” with afoot lifted fast, held high, and toes open andclosed.
Shaping starts by reinforcing the first approxi-mation every time it is offered, until it is per-formed without hesitation. Next, an even closerapproximation is reinforced, at which time rein-forcement for the first approximation is withheld.Once the second approximation is performedwithout hesitation, an even closer approximationis reinforced while withholding reinforcement forall previous approximations. In this way, the cri-terion for reinforcement is gradually shifted(graduated) closer and closer to the target behav-ior. Finally, every instance of the target behavioris reinforced.
If the learner experiences difficulty at any cri-terion, the trainer backs up and repeats the previ-ous successful step, or reinforces smaller approx-imations. Once an approximation is performedwithout hesitation, more variability can be gener-ated from which to select the next approximationby switching from continuous reinforcement tointermittent reinforcement (see the discussion ofextinction bursts later). Ultimately, it is the parrotwho determines the exact sequence and pace ofthe shaping plan. This is where sensitivity and ex-perience is required on the part of the trainer toobserve the nuances of behavior.
With shaping toy play and bathing, the toys andwater dish are the antecedents that set the occa-
14 / Behavior Analysis and Parrot Learning 157

sion for the respective behaviors. For other behav-iors, a cue from the trainer (discriminative stimu-lus) can be added to signal the behavior. To add acue, start by introducing it while the behavior isoccurring. Next, gradually deliver the cue earlierand earlier until it is signaled before the behavior.Last, reinforce only cued instances of the behav-ior and ignore all others. This will establish therelationship between the cue and behavior, calledstimulus control. When a behavior is said to beunder stimulus control, it is emitted after the cueand rarely or not at all when the cue is absent.
With shaping we can theoretically train any be-havior within the biological constraints of thelearner. Husbandry, medical, and enrichment be-haviors can be shaped to reduce stress and in-crease physical and mental stimulation. Birds canlearn such behaviors as raising each foot for nailtrims, going in and out of crates, staying calmwrapped in towels, flying to designated perches,and playing basketball. Shaping can also be usedto change different dimensions of existing behav-iors such as duration, rate, intensity, topography,and response time.
Not surprisingly, problem behaviors are oftenunwittingly shaped as well. We inadvertentlyteach our birds to bite harder, scream louder, andchase faster through the subtle mechanisms ofshaping. For better and for worse, shaping is end-lessly applicable to teaching captive parrots, mak-ing targeting the sharpest of all training tools. Itsuses are limited only by one’s imagination andcommitment to learning how to use it well.
Shaping Touch-to-Target
Regarding cats, Catherine Crawmer (2001) de-scribes the technique known as targeting this way:“If we could get a cat to touch his nose to a stickon cue what could we do with that behavior? Theanswer is a question: What couldn’t we do withit?” (p. 57).
Targeting is the behavior of touching a bodypart (e.g., beak, wing, or foot) to a designated ob-ject or mark and it is taught easily to parrots withshaping. By teaching birds how to target the endof a wooden dowel with their beaks, caretakerscan predict and control the birds’ movements. Forexample, an untamed bird can be taught to targeta stick while inside its cage, enabling the care-taker to safely increase interaction with the bird,deliver positive reinforcement, and establish two-
way communication. A bird that refuses to comeoff the top of its cage can be targeted to a perchinside it; a wary bird can be targeted into a travelcrate for veterinary visits; and an aggressive birdcan be quickly redirected to the target to distractit from biting. Also, enrichment behaviors can betaught with targeting such as turning in a circle,climbing up and down ladders, and ringing a bell.Target training is an important basic skill for allcompanion parrots, as it opens the door to allsorts of positive reinforcement and managementopportunities.
Differential Reinforcement of AlternativeBehaviors
Differential reinforcement is any training proce-dure in which certain kinds of behavior are sys-tematically reinforced and others are not. Shapingis one example of differential reinforcement; atany point in the shaping sequence reinforcementis delivered for one approximation and withheldfor all earlier ones. The process of withholding re-inforcers that previously maintained a behavior iscalled extinction and it results in an overall reduc-tion in the frequency of the behavior. Thus, differ-ential reinforcement is technically two proce-dures, positive reinforcement and extinction, thecombined effect of which is to increase the rein-forced behavior and extinguish (decrease) theunreinforced one.
The relevance of differential reinforcement pro-cedures to companion parrot behavior is enor-mous, specifically as an alternative to punishment.Punishment procedures focus solely on decreasingor suppressing behavior, teaching what not to do,which necessarily reduces the amount of positivereinforcement available to the bird. Instead, differ-ential reinforcement of alternative behavior fo-cuses on reinforcing appropriate replacement be-haviors, teaching what to do, while at the sametime the undesired behavior is ignored. Whenproperly implemented, the result is a high rate ofpositive reinforcement for the bird, and a low rateof the problem behavior for the teacher.
There are three things to consider when select-ing an alternative behavior for a differential rein-forcement procedure (Alberto & Troutman 2003).First, although the behavior targeted for reductionis a problem to people, it serves a legitimate func-tion to the parrot or it would not continue to ex-hibit the behavior. The function is either to gain
158 Manual of Parrot Behavior

something of value (positive reinforcement, e.g.,social attention, items or activities, sensory rein-forcement) or to remove something aversive (neg-ative reinforcement, e.g., escape), as whenscreaming gains attention from caregivers andlunging removes intruding hands. An alternativebehavior should be selected that replaces thefunction served by the problem behavior but in amore appropriate way. If the alternative behavioris incompatible with the problem behavior (i.e., ifboth behaviors can’t physically be performed atthe same time), the behavior change program willbe that much more powerful. For example, talkingis incompatible with screaming, and waiting on afar perch is incompatible with lunging at the feeddoor.
Second, the alternative behavior must result inthe same amount of or more reinforcement thanthe problem behavior, in order to successfullycompete with and replace it. This is predicted bythe matching law, which states “that the distribu-tion of behavior between alternative sources of re-inforcement is equal to the distribution of rein-forcement for these alternatives” (Pierce &Cheney 2004, p. 434). Thus, given a choice be-tween two alternatives, parrots will exhibit thebehavior that results in the greater reinforcement.Third, the alternative behavior should be one thebird already knows how to do; a well-establishedbehavior is more likely to be performed than onethat is newly acquired.
When alternative behaviors are strengthenedand maintained, differential reinforcement canprovide long-lasting results. As this method relieson positive reinforcement to reduce problem be-haviors by teaching birds what to do, it offers a
positive, constructive, and practical approach tomanaging parrots in captivity.
PUNISHMENT
As discussed previously, even with the most pro-ficient and proactive behavior management skills,the time will likely come when the frequency ofsome behavior needs to be decreased. Althoughthe following behavior reduction procedures maybe useful adjuncts to positive reinforcement, theyshould not be used alone (Kazdin 2001). Overall,punishment is used too frequently and less effec-tively than it should be, partly because it is suchan ambiguous concept. In behavior analysis it hasa specific, technical meaning: Punishment is theprocedure of contingently providing conse-quences for a behavior that decreases or sup-presses the frequency of that behavior. Positivepunishment is a behavior reduction procedure inwhich a behavior is followed by the presentation(+) of an aversive stimulus. Negative punishmentis a behavior reduction procedure in which a be-havior is followed by the removal (�) of positivereinforcers. Examples of positive and negativepunishment are listed in Table 14-4. As can beseen in the table, the frequency of the target be-haviors is decreased in each example as that de-fines punishment.
Like reinforcement, punishment is definedsolely by its effect on behavior. Punishment canbe said to have occurred only if the frequency ofthe target behavior decreases. Statements like“I’ve sprayed him a million times, punishmentdoesn’t work with parrots!” are nonsensical.There is no such thing as failed punishment (orreinforcement). When an attempt to reduce the
14 / Behavior Analysis and Parrot Learning 159
Table 14-4 Examples of positive and negative punishment
Antecedent Behavior Consequence Future behavior
Positive Grace passes Sam’s Sam charges again Grace sprays Sam charges bars punishment cage bars water at Sam less
Grace is on the Sam bites her hand Grace drops Sam Sam bites lesstelephone to the floor
Negative Grace offers hand Sam hangs on cage Time out—Grace Sam hangs on punishment door walks away for cage door less
a few minutesGrace enters home Sam whistles shrilly Extinction—Grace Sam whistles
remains silently shrilly lessout of sight

frequency of a behavior produces no immediatechange whatsoever, punishment has not occurredand different strategies should be implemented(Chance 2003). Although both positive and nega-tive punishment decrease or suppress behavior,positive punishment is associated with particu-larly adverse side effects discussed in the nextsection. It seems logical that having something ofvalue taken away (negative punishment) is ulti-mately less aversive, although not necessarily lesseffective, than having something noxious admin-istered (positive punishment). This makes nega-tive punishment the preferred strategy afterantecedent arrangements and differential rein-forcement of alternative behaviors.
Like reinforcement, punishing stimuli can beclassified as primary (automatic) or secondary(learned by association with existing punishers),and the effectiveness of punishment proceduresdepends on clear contingency, close contiguity,type, magnitude, and schedule of delivery, as wellas other factors.
The Problems with Positive Punishment
Positive punishment, such as shaking perches,banging cages, spraying, hitting, laddering, flash-ing lights, and plucking out feathers, is problem-atic for parrots and their relationship with humansfor several reasons. Like all learned behaviors,problem responses continue because they are re-inforced. When we implement punishment we notonly fail to teach what to do, we necessarily re-duce the amount of reinforcement previouslyavailable to the learner for misbehaving—a dou-ble negative of sorts, as punishment is added andreinforcement is subtracted. This makes it vitallyimportant to use punishment in conjunction withpositive reinforcement procedures to strengthendesirable behaviors and maintain a reinforcingenvironment. This guideline is called the fair pairrule (White & Haring 1976).
Another problem with punishment is the sever-ity required to produce lasting effects. Researchhas shown (e.g. Azrin & Holz 1966) that high-intensity punishment is more effective than eitherlow-intensity punishment or escalating levels ofpunishment. The intensity required to suppressparrots’ problem behaviors is often greater thanthat which meets acceptable standards of ethicalpractice or is comfortably administered by care-givers.
With negative reinforcement an aversive stimu-lus is present in the antecedent environment, theremoval of which reinforces the escape behavior.With positive punishment the aversive stimulus isadministered without escape, which sets the stagefor the detrimental side effects frequently ob-served with positive punishment. They are escapebehaviors; aggression and other emotional reac-tions; generalization of emotional reactions to un-related people, settings, and items; apathy (a gen-eral reduction of all behavior); and behavioralcontrast (the increase of the target behavior inother settings). These side effects are well estab-lished, having been broadly investigated for manydecades with countless species of animals (e.g.,Azrin et al. 1965; Sidman 1989); and they are star-tlingly common among captive parrots, many ofwhich show extreme aggression, apathy, and fear.
It is the narrow view that effectiveness is thesole criterion for choosing behavior-change pro-cedures that perhaps keeps so many people usingpunishment. Unfortunately, every time a problembehavior is successfully decreased with positivepunishment, the person delivering the punishmentis negatively reinforced for having used it. Ofcourse, this will result in an increased probabilitythat positive punishment will be used more. Yetbased on the nature of parrots’ problem behaviorsin captivity, the known detrimental side effects ofpositive punishment, and the power of reinforce-ment-based alternatives, there can be little justifi-cation for using positive punishment with captiveparrots.
Negative Punishment
The two negative punishment procedures relevantto parrot behavior are time out from positive rein-forcement (time out) and extinction. Time out isthe contingent, temporary removal of access to allpositive reinforcers and extinction is the contin-gent, permanent removal of the specific rein-forcer(s) maintaining the problem behavior. Bothprocedures can be very effective when used cor-rectly but they are frequently misunderstood andvery poorly implemented.
The effectiveness of time out is undermined byunclear contingency, slow contiguity (timing),and inadvertent reinforcement, also known as“bootleg” reinforcement (Chance 1998, p. 458).For example, chasing the bird, scolding, andmarching to distant cages can provide bootleg re-
160 Manual of Parrot Behavior

inforcement that renders time out ineffective. Un-der these conditions, the parrot has little chanceof perceiving clearly the contingent withdrawal ofpositive reinforcers, thereby obscuring the associ-ation between the offending behavior and beingreturned to its cage. Time out is more effectivewhen using the following guidelines:
• Plan the time out location ahead of time to en-sure that it can be managed with clear contin-gency and immediacy. For many tame parrots,simply turning away or being set down for ashort time is an effective time out from positivereinforcement.
• Increase the salience of the contingency be-tween the behavior and the consequence bykeeping the time out interval short (approxi-mately 30 seconds to a few minutes). Watch theclock or count out the seconds to track the timesystematically.
• Immediately after the time out interval, givethe bird the opportunity to practice the appro-priate behavior and reinforce it amply everytime it is exhibited.
• Allow time out to do all the work in decreasingthe problem behavior. There is no need forother consequences or emotional displays fromthe caregiver, which may provide bootleg rein-forcement for the problem behavior.
Extinction used in combination with positivereinforcement has already been discussed as it ap-plies to shaping and differential reinforcement ofalternative behavior. To implement extinction as asingle behavior reduction procedure, the rein-forcer that maintains the problem behavior shouldbe identified first by conducting a functional as-sessment (ABCs). In the case where the maintain-ing reinforcer is human attention, extinction istantamount to inviolate ignoring—the total andpermanent withholding of attention. Unfortu-nately, for some parrot behaviors like excessivescreaming, biting, and chewing unapproveditems, ignoring is easier to prescribe than to im-plement effectively.
As discussed by Alberto and Troutman (2003),careful consideration should be given to the fol-lowing points before using extinction to decreasea problem behavior. First, extinction tends to be aslow procedure. Once the maintaining reinforceris withheld, the behavior continues for an indeter-
minate amount of time. As discussed previously,behaviors with an intermittent reinforcement his-tory are the slowest to change, the most resistantto extinction. Second, the frequency, intensity,and/or duration of the behavior may sharply in-crease before a significant decrease in the prob-lem behavior occurs. This phenomenon is knownas an extinction burst. This predictable escalationis often beyond toleration for caregivers. As aresult, they abandon the program by providing at-tention, and the behavior is unintentionally re-inforced at the new level of intensity. Third, be-haviors associated with frustration, such asaggression, are commonly induced by extinction.For parrots, this may mean an increase in the fre-quency and intensity of already severe biting.Fourth, as with time out, bootleg reinforcementcan be a problem. Reinforcement can be deliveredby other pets, children, or even an echo in theroom. Further, some behaviors appear to be auto-matically reinforcing. When the maintainingreinforcer is not in the control of the trainer, ex-tinction cannot be effective.
The fifth point to consider is spontaneous re-covery, also known as resurgence (Sulzer-Azaroff& Mayer 1991). Resurgence is the reappearanceof the extinguished behavior after an extended pe-riod of time. Forewarned, the immediate reimple-mentation of strict extinction conditions will re-turn the behavior to its pre-recovery frequency.Sixth, the problem behaviors that caregivers ig-nore can be imitated by other parrots. This pro-duces additional behavior problems for caregiversto solve and increases the probability of bootlegreinforcement: One parrot’s imitative behaviorcan reinforce another parrot’s problem behavior.
On the whole, ignoring is most effective as apreventative strategy rather than a problem solu-tion. It offers a window of opportunity to avoidgiving the problem behavior function by with-holding reinforcement the very first time it is ex-hibited. Once a problem behavior is well estab-lished, differential reinforcement of alternativebehaviors is usually the better strategy.
CONCLUSION
The allure of companion parrots is often out-weighed by the collateral challenges of keepingthem in captivity. This is especially true when thewelfare of the animals is kept in the foreground.A basic understanding of how behavior works
14 / Behavior Analysis and Parrot Learning 161

combined with a practical, humane teaching tech-nology will help stem the tide of parrots adver-tised for resale in newspapers and relinquished toshelters and sanctuaries.
There are currently several popular belief sys-tems regarding how best to manage parrot behav-ior. When opinions differ, emotions are strong, andthe stakes are high, science should hold a highervalue than conventional wisdom and personalrecipes about behavior. Science demonstrates animportant association between behavioral healthand empowerment; that is, the personal power tocontrol significant environmental events. Over-powering parrots with forceful and coercive train-ing methods should be understood as stealingbehavior that could be given to us instead with fa-cilitative antecedents and positive reinforcement.Empowering captive parrots to the greatest extentpossible, within the context of appropriate trainingobjectives, may mitigate the behavioral pathologiesso prevalent among them.
Given a choice between different behavioral in-terventions, selecting the most positive, least in-trusive, effective strategy meets the highest stan-dard of ethical practice. Antecedent changes andpositive reinforcement procedures should alwaysbe tried before implementing negative punish-ment (removing positive reinforcers) or negativereinforcement (escape training). Positive punish-ment procedures, in which aversive stimuli areapplied, should be used rarely, if ever. Finally, allthree procedures—negative reinforcement, nega-tive punishment, and positive punishment—should only be used as an adjunct to positive rein-forcement strategies.
Taking full responsibility for parrots’ learningand behavior is the first and most important stepto supporting their behavioral health. Companionparrots offer their caregivers the opportunity toeducate themselves about behavior and signifi-cantly improve the quality of life for parrots incaptivity.
REFERENCES
Alberto, P.A., and A.C. Troutman. 2003. Applied be-havior analysis for teachers, 6th ed. Upper SaddleRiver, NJ: Merrill Prentice Hall.
Azrin, N.H., and W.C. Holz. 1966. “Punishment.” InOperant behavior: Areas of research and application,ed. W.K. Honig, pp. 380–447. New York: Appleton-Century-Crofts.
Azrin, N.H., R.R. Hutchinson, and R. McLaughlin.1965. The opportunity for aggression as an operantreinforcer during aversive stimulation. Journal ofthe Experimental Analysis of Behavior, 8:171–180.
Balph, D.F., and M.H. Balph. 1983. On the psychologyof watching birds: The problem of observer-expectancy bias. Auk 100:755–757.
Barnett, S.A. 1981. Modern ethology. New York:Oxford University Press.
Barrow, E.M. 2001. Animal behavior desk reference: Adictionary of animal behavior, ecology, and evolu-tion, 2nd ed. Boca Raton, FL: CRC Press.
Brown, G.E., G.D. Hughs, and A.A. Jones. 1988. Effectsof shock controllability on subsequent aggressive anddefensive behaviors in the cockroach (Periplanetaamericana). Psychological Reports 63:563–569.
Chance, P. 1998. First course in applied behavioranalysis. Pacific Grove, CA: Brooks/Cole Publish-ing Company.
Chance, P. 2003. Learning and behavior, 5th ed. Bel-mont, CA: Wadsworth/Thomson Learning.
Cloutier, S., J.P. Beaugrand, and P.C. Lague. 1995. Theeffect of prior victory or defeat in the same site asthat of subsequent encounter on the determination ofdyadic dominance in the domestic hen. BehavioralProcesses 35:293–298.
Coulton, L.E., N.K. Warren, and R.J. Young. 1997.Effects of foraging enrichment on the behavior ofparrots. Animal Welfare 6:357–363.
Crawmer, C. 2001. Here kitty, kitty: Catherine Craw-mer on training cats. Sand Lake, NY: Author.
Gall, M.D., J.P. Gall, and W.R. Borg. 2003. Educationalresearch, 7th ed. Boston: Allyn and Bacon.
Gilbert-Norton, L. 2003. Captive birds and freeload-ing: The choice to work [Electronic version]. Re-search News 4.
Gould, S.J. 1981. The mismeasure of man. New York:W.W. Norton & Company.
Inglis I.R., and N.J.K. Ferguson. 1986. Starlings searchfor food rather than eat freely available food. AnimalBehaviour 34:614–616.
Kazdin, A.E. 2001. Behavior modification in appliedsettings, 6th ed. Belmont, CA: Wadsworth/Thomson.
Lattal, K.A. 1995. Contingency and behavior analysis.Behavior Analyst 24:147–161.
Laudenslager, M.L., S.M. Ryan, R.C. Drugan, and R.L.Hyson. 1983. Coping and immunosuppression:Inescapable but not escapable shock suppresses lym-phocyte proliferation. Science 221:568–570.
Maier, S.F., and M.E.P. Seligman. 1976. Learned help-lessness: Theory and evidence. Journal of Experi-mental Psychology: General 105:3–46.
Manning, A., and M. Stamp Dawkins. 1992. An intro-duction to animal behavior, 4th ed. Cambridge:Cambridge University Press.
162 Manual of Parrot Behavior

Mech, L.D. 1999. Alpha status, dominance, and divi-sion of labor in wolf packs. Canadian Journal ofZoology 77:1196–1203.
Mech, L.D. 2000. Leadership in wolf, Canis lupus,packs. Canadian Field-Naturalist 114:259–263.
Michael, J. 1982. Distinguishing between discrimina-tive and motivational functions of stimuli. Journal ofAnalysis of Behavior 37:149–155.
Osborne, S.R. 1977. The free food (contrafreeloading)phenomenon: A review and analysis. AnimalLearning & Behavior 5:221–235.
Overmier, J.B., and M.E.P. Seligman. 1967. Effects ofinescapable shock upon subsequent escape andavoidance responding. Journal of Comparative andPhysiological Psychology 63:28–33.
Pierce, W.D., and C.D. Cheney. 2004. Behavior analy-sis and learning, 4th ed. Mahwah, NJ: LawrenceErlbaum Associates, Inc.
Schneider, J.W. 1973. Reinforcer effectiveness as afunction or reinforcer rate and magnitude: A com-parison of concurrent performance. Journal ofExperimental Analysis of Behavior 20:461–471.
Schwartz, B., E.A. Wasserman, and S.J. Robbins. 2002.Psychology of learning and behavior, 5th ed. NewYork: W.W. Norton & Company, Inc.
Seibert, L.M., and S.L. Crowell-Davis. 2001. Gendereffects on aggression, dominance rank, and affilia-
tive behaviors in a flock of captive cockatiels.Applied Animal Behavior Science 71:155–170.
Seligman, M.E.P. 1990. Learned optimism. New York:Knopf.
Sidman, M. 1989. Coercion and its fallout. Boston:Authors Cooperative.
Simmons, R. 1924. The relative effectiveness of certainincentives in animal learning. Comparative Psychol-ogy Monographs 7.
Skinner, B.F. 1981. Selection by consequences. Science213:501–504.
Sulzer-Azaroff, B., and G.R. Mayer. 1991. Behavioranalysis for lasting change. Orlando, FL: Harcourt,Brace, Jovanovich.
Todorov, J.C., E.S. Hanna, and M.C.N. Bittencourt deSa’. 1984. Frequency versus magnitude or rein-forcement: New data with a different procedure.Journal of the Experimental Analysis of Behavior4:157–167.
Van Kerkhove, W. 2004. A fresh look at the wolf-packtheory of pet dog social behavior. Journal of AppliedAnimal Welfare Science 4:279–285.
White, O.R., and N.G. Haring. 1976. Exceptionalteaching. Upper Saddle River, NJ: Merrill/PrenticeHall.
14 / Behavior Analysis and Parrot Learning 163

INTRODUCTION
Behavior problems are well recognized as a lead-ing cause of euthanasia or abandonment for dogsand cats.[1, 2] People become disillusioned witha pet that does not meet their expectations. Petbirds are not exempt from this type of problem.Behavior problems such as biting, screaming, andfeather picking are very common among petbirds. In fact, since they are still essentially un-changed from the wild state, these behavioralproblems may be even more common. Euthanasiais still a very uncommon request in pet birds.Owners and veterinarians alike are hesitant to enda bird’s life for their own convenience. As a result,these birds are often bounced from one home toanother, frequently ending up as breeding birds. Itis not difficult to find even very expensive birdsgiven away because of behavioral problems. Theauthor owns five psittacids with a combined valueof over $4,000 that were given up because of var-ious behavioral problems. The birds that are soldor given to a new home are the lucky ones. Othersare ignored or neglected because the owners areafraid of them, or they are put into basements orother isolated areas because they are too noisy.While the idea of placing these birds in breedingprograms may have merit in some instances, overmany generations, we may be selectively breed-ing for the least desirable behavioral traits. Theauthor has seen at least one African Grey Parrotpair whose offspring start feather picking beforethey ever leave the pet store. Obviously, this couldbe due to the early practices of the breeder or
many other factors. However, like many othertraits, behavior can have genetic and environmen-tal influences. If only those pets that develop un-desirable behaviors are brought into breeding pro-grams, this situation may become increasinglycommon.
Wild parrots are usually hatched in clutches oftwo to six, and the mother and father tend themconstantly. Following fledging and weaning, par-rots continue to learn from their parents and otherflock members. They follow the parent birdsaround the environment and learn what to fear,what not to fear, what to eat, how to find food,where to sleep, and how to interact with othermembers of the flock. In captivity, neonatal par-rots often are poorly socialized during the hand-feeding process. They are often housed in individ-ual brooders and their only interaction is whenthey are fed. Once weaned, many are immediatelysold or placed into new homes. Very infrequentlyare they “educated” regarding coping with thecaptive environment.
Behavior classes for owners and birds are onemeasure to both prevent and rehabilitate problemparrots. When a dog trainer began to offer obedi-ence training in the author’s practice, the idea of asimilar forum for addressing avian behavior is-sues was formed. By offering a class where sev-eral clients could be educated at the same time, agreater amount of information could be trans-ferred. The owners are given information thatthey can use at home to work with the bird. In ad-dition to the benefit of improving the relationship
165
15Behavior Classes in the
Veterinary Hospital: PreventingProblems Before They Start
Kenneth R. Welle

between bird and owners, a behavior class can af-ford an opportunity for introducing the bird tonovel situations. Many pet birds live their entirelives in one room of a house. These birds oftenbecome very intolerant of small changes in theirlives and will be easily stressed by inevitableevents.[3, 4] An organized class, with birds in-cluded, allows for socialization of birds, owners,and the veterinary staff in a much less intimidat-ing setting than the clinical examination.
Many of today’s bird owners are hungry for in-formation of any kind, and behavior is a hot topic.Even those owners who know quite a bit aboutavian behavior are anxious for more. The infor-mation regarding avian behavior is scattered inmany places. In addition, there is considerabledisagreement among “authorities” on bird behav-ior. This can lead to confusion and inappropriatehandling of birds. Advice about bird behavior isoften taken from untrained pet store personnel. Inmany areas, the avian veterinarian may be theonly authoritative resource for bird owners. Ofcourse, not every avian veterinarian is qualified toteach such a class. The veterinarian wishing tostart behavior training must thoroughly familiar-ize him- or herself with pet birds, their behavior,and their relationships with owners. The instruc-tor of such a class must be very confident han-dling birds. Some veterinarians restrain birds reg-ularly but have little experience handling them inan interactive setting. Counseling pet owners onbehavior is an art that must be studied prior tostarting.[5] It is often helpful to directly observethe interactions between a bird and its owner. Thiscan reveal a wealth of information about the rela-tionship that an owner may be unable or unwillingto recognize.
COURSE DESCRIPTION
The author’s behavior class was started in 1996.Since this time the course has evolved andchanged to adopt the changing opinions regardingthe best ways of preventing and treating commonbehavior problems. Because of the popularity ofobedience training for dogs, the term “avian obe-dience training” was originally adopted. Sincethis time the term “bird socialization class” hasbeen used in place of obedience training. Thecourse is five weeks long; the first week is forowners only, while the following four weeks arefor birds and owners. Each session is about one
hour long. During the first session, a videotape,Parrots: Look Who’s Talking (Nature, Thirteen-WNET, and PBS, New York) is shown. This tapeis very entertaining and informative and intro-duces some of the concepts discussed in the class.The remainder of the first session is used to begindiscussion of the course topics. Wild parrot be-havior, psittacine social structure, and environ-mental factors for pet birds are some of the firsttopics covered.[6–11] In the sessions where birdsare present, discussions and bird exercises aremixed to keep up the interest of both the birds andowners. Ten minutes of talking followed by tenminutes of exercises appears to work well. Table15-1 lists the topics and exercises covered in eachof the sessions. To complement the class, a writ-ten handout is given to the class members. At thecompletion of the course, each bird is given adiploma, which is easily generated on a personalcomputer. Clinics wishing to begin these classescan start with this format and then modify it tosuit their own practice. The author’s original classhandout is available through the Association ofAvian Veterinarians publication office.
CAVEATS
One concern with a class such as this is the trans-mission of infectious diseases. While this is notentirely preventable, requiring that all applicableinfectious diseases be screened for in all regis-trants can minimize the risk. The type of clientwho is interested in such a class generally willagree to this requirement. In the author’s practice,the requirements include an examination, a com-plete blood count, and chlamydophila screeningwithin the past year. Old world parrots are re-quired to have a negative screening for circovirusat some point. Any birds with problem behaviorsshould be worked up for medical causes of theseproblems before assuming that it is solely behav-ioral. Some types of behavioral problems aremore suitable for individual counseling than agroup setting. Clients should bring their owntowels and carriers. The perches are made ofpolyvinyl chloride pipes and covered with dispos-able self-adhesive elastic bandage so that they canbe sanitized in between uses. Alternately, ownerscan be required to provide their own perches. Pre-weaned babies are not permitted, as they are verysusceptible to bacterial infections. This is not aprogram that should be used for aviculture birds.
166 Manual of Parrot Behavior

It is intended for pet birds where the need for pro-tecting them must be tempered by the need to im-prove the quality of life. If more than one bird isin the household, the class participant should beconsidered a “new bird” again after the comple-tion of the class and all applicable quarantine pro-cedures should be followed.
The other concern with this program is the lia-bility. During the course of the sessions, ownersmay be bitten by their birds. As with clinical ex-aminations, the liability for such bites is with thepractice owner. However, having a technicianhandle the bird for the owner defeats the purpose.A waiver, indicating that owners are aware of therisk of being bitten, provides some protection inthe event of a severe bite. If younger family mem-bers participate, a parent should give consent.Bird to bird aggression could also result in liabil-ity claims. While this has never occurred in theauthor’s practice, it could easily ruin an otherwisepleasant class. To minimize risks, all birds are re-
quired to have trimmed wings and owners are notallowed to bring more than one bird for each han-dler. The room should allow the birds enoughspace that they do not feel threatened by the prox-imity of other birds or handlers.
COURSE CONTENT
Psittacine Socialization
During the class, it is emphasized that parrots arealtricial and that they have very long behavioraldevelopment.[3, 6] Hand-raised parrots do nothave the instincts needed to be well-adjusted pets.The owner’s family must take on the role of theflock and guide the development of the chick.While some owners feel that teaching obedienceto a bird is cruel and unnatural, the opposite istrue; it is cruel and unnatural to let them try tofigure things out for themselves.[3] It is empha-sized that pet parrots must be “educated” to prop-erly adapt to human habitats. Parrots and their rel-
15 / Behavior Classes: Preventing Problems Before They Start 167
Table 15-1 Sample outline for in-clinic avian behavior classes
Session Topic Exercises
1 Environment Step upPsittacine socialization Step downCommunication Stay
2 Development and learning Step upIndependence and confidence Step down
StayHooding
3 Interactive games Step upStep downStayHoodingCarrier
4 Bonding Step upStep downStayHoodingCarrierTowel play
5 Motivation and reinforcement Step upStep downStayHoodingCarrierTowel playTricks

atives are highly social and intelligent birds.There are 332 species of psittacine birds and thesocial structure of each of these is somewhat dif-ferent. These facts make it inherently difficult toprovide an appropriate social setting for thesebirds in the captive environment. When socialneeds are not met, companion birds often developbehaviors that are self-destructive or make themundesirable as pets.
A common misconception is that social behav-ior evolved because of a need for companionship.In reality the congregation of groups of animalswithin a species serves a much more utilitarianpurpose. Aptly put by Christine Davis, “In thewild, the flock is primarily a protective unit, andaffiliation with others is essential to the survivalof each individual. The need for social interac-tion, although extremely important, is secondaryto the instinct for survival.”[12] The presence ofconspecifics makes it much more difficult for apredator to successfully stalk and kill a given in-dividual. Even with a less altruistic perspective,the presence of other potential prey items in thevicinity makes it mathematically less likely for anindividual to be preyed upon. Behaviors nec-essary for relatively conflict-free interactionbetween individuals of a group evolved as a sec-ondary requirement. This fact makes social inter-action even more critical. Not only is a bird lonelywhen alone, it feels vulnerable to attack. A closeanalogy would be to compare the situation to aperson walking at night. Generally people feelmore secure and safe when with a group of peo-ple than when walking alone.
While descriptions of natural psittacine socialbehavior abound, most of these are based more ontelevision, magazines, captive observations, andconjecture rather than scientific observation ofparrots in the wild. Both the descriptions of par-rot behavior and their interpretation vary to someextent. Nonetheless, these descriptions currentlyprovide the best basis for determining what thehealthiest social environment will be for captivebirds.
Most parrots pair bond. Keas are polygamous.Asiatic Parakeets and Eclectus bond only forbreeding season. Most parrots separate in pairsduring breeding, while some, such as Quakers,Brown-throated Conures, and Patagonian Con-ures, are colony nesters. Outside breeding season,parrots live in small to very large flocks (A.U.
Luescher, unpublished data, March 1999). Inorder for any animal society to function, commu-nication and conflict-resolution mechanisms mustbe developed. For most social animals a form ofhierarchy develops to prevent true combat situa-tions. According to some, most parrot flocksfunction under a hierarchic arrangement.[12]Young parrots learn to submit to the leader. Ifthey try to share resources, they are bitten andchased away.[6] Others argue that while withinpairs there appears to be a dominance relation-ship, in flock situations no dominance has beendescribed (A.U. Luescher, unpublished data,March 1999). Because of their intelligence, parrotsocial structure may not be a linear dominance hi-erarchy. There may be a matter of creating al-liances more than of dominance. Also, dominancein one situation may not apply to another.[13]Whether it comprises a true dominance hierarchysystem or not, it certainly appears that parrotsavoid violent conflict by ritualized posturing andpositioning.
Vertical placement or height is frequently re-ferred to in avian behavior literature. It has beensaid that the dominance order can often be deter-mined by seeing dominant individuals in higherpositions.[12] Others claim that the apparent rela-tionship of height to dominance may be related tothe high spot being the most desirable perchrather than the height itself.[13] Others still main-tain that the notion of a flock leader and dominantbirds sitting higher than submissive birds is amyth entirely (A.U. Luescher, unpublished data,March 1999).
Most parrots are noisy and communicate vo-cally. They learn a flock dialect. They also com-municate visually with body language and color(A.U. Luescher, unpublished data, March 1999).Different vocalizations are used by parrots to sig-nal danger or food or to greet another bird.[13, 14]Vocalization is critical for keeping flocks together,especially in dense rain forest habitats. Calls areused to bring the flock together at the end of theday.[14] According to Davis, “Screaming, espe-cially at daybreak and dusk, is a normal activity inthe wild, essential in keeping track of companionsat those particular times and at various intervalsthroughout the day.”[12] Pairs engage in socialgrooming and mutual feeding to maintain theirbonds. These interactions are uncommon betweenother members of the flock (A.U. Luescher, un-
168 Manual of Parrot Behavior

published data, March 1999). Various behaviorsare used for communication with less intimateflock mates.[14]
• Beak clicking: greeting or warning• Beak grinding: contentment• Eye pinning: excitement, either good or bad• Facial feather twitching: startled or intrigued• Fluffing: prelude to preening or tension re-
leaser• Foot tapping: territorial defense• Tail fanning: courtship or aggressive display
Development and Learning
For many altricial species, including most of thepsittacine family, behaviors tend to be learned.This is true whether the bird is in the wild or incaptivity. Birds learn various skills more effi-ciently at certain stages of development, knownas sensitive periods or critical periods.[13] Onesuch behavior is imprinting, either filial imprint-ing (forming social attachments) or sexual (form-ing an image of a desirable mate).[13] This canlead to conflict when birds are raised by humans.As stated by Harrison in Avian Medicine:Principles and Applications, “Birds raised byhuman foster parents will imprint as people, notbirds. As they mature, their natural instincts tochoose a mate may cause objectionable behav-iors. . . . An imprinted bird will spend all of itstime attempting to drive unwanted individuals,other pets or objects out of its territory, while try-ing to find one chosen person with whom tomate.”[6] This species confusion is compoundedwhen the human caretaker fails to continue thebird’s education to include necessary skills forcoping with the artificial environment.
Once the bird fledges, it becomes versed in the“flock experience.”[6] Early learning can be ex-tremely critical for the long-term development ofthe bird. Any synapses that are not stimulated byearly sensory experiences may be “pruned” by thebrain and eliminated.[15] Play is used frequentlyto learn the environment and reaffirm flockposition.[14] When normal play behaviors ofWhite-fronted Amazons were studied, behaviorsincluded play solicitation, play biting, or playfighting, or those associated with pair bonding,such as allopreening and bill nibbling.[6] In cap-tivity, playing with a bird can provide an effectivemeans of teaching necessary skills to pet birds.
Structured training or play sessions can allowowner and pet to interact in an entertaining butstructured manner.[16] The objectives are to teachboth behaviors required for companionability andbehaviors that assure comfort, health, and happi-ness.[15] Play should focus on behaviors theanimal likes to do and encourage those that aredesirable.[16]
Developing a Healthy Social Environment forPet Birds
A healthy human-avian bond requires each side tobehave in a manner that promotes this bond.Necessary components of a well-adjusted andwell-behaved pet bird include respect and trust ofthe owner and other humans, independence, and aplatonic bond.
As wild animals, parrots have no concept of thehuman-animal bond and can only interact withhuman companions as flock members.[12]Initially this may be the role of a parent bird. InGuide to the Quaker Parrot, Athan writes, “A suc-cessful relationship between human and com-panion bird casts the human in the role of lovingparent. Because parrots have a strong instinct tounderstand what is expected of them, a loving, au-thority-based relationship is probably the bestway to maintain a long-term relationship betweenhuman and parrot.”[17] This relationship is rela-tively easy to accomplish. It is important to setrules for an authority-based relationship, in whichthe bird understands that the humans are theboss.[11] One of the rules pertains to height.Whether because of dominance, site-related ag-gression, or other unknown reasons, birds aremore aggressive when placed in high posi-tions.[12] Pet birds should be kept below the eyelevel and in front of the handler. Shoulders areparticularly bad locations because they preventthe person from keeping eye contact, from plac-ing hands properly for handling, and becausebites from this position can be particularly dan-gerous. The next aspect of maintaining the au-thority-based relationship is teaching some basiccommands and using them frequently. Steadyhands impart confidence on the part of the han-dler and also give steady footing to the bird. Birdsoften become infuriated when someone offers ahand to a bird only to pull it back when they reachout with their beak to test it. The step-up drill isone that every pet bird should know. In this drill
15 / Behavior Classes: Preventing Problems Before They Start 169

the command “step up” is given and the bird iscoaxed to step up onto the hand. Each time thisexercise is performed, respect and trust are built.It is not enough that the bird knows what the com-mand means. Practice must be maintained tomaintain that mutual trust and respect. Once trustand respect are established, other skills can betaught. Important social skills can include inter-action with other birds, playing with water, ex-ploring foods, meeting strangers, and manyothers.[15] These behaviors are learned throughimitation. In the home, parrots imitate their com-panions, whether avian or mammal. If they cannotcopy behaviors, they improvise. If the improvisedbehaviors are reinforced, the behavior can be-come a pattern. Likewise, desirable behaviors canbe encouraged by modeling those behaviors infront of the bird. The motivation to copy behav-iors can be enhanced by providing a competitivesituation in which the bird learns. The method isknown as the model-rival technique and has beenpioneered in birds by Dr. Irene Pepperberg.[11]
Independence and Confidence
Contrary to popular opinion, behavioral disordersare not always a result of inadequate interactionwith a pet bird. Very often, the interactions are ex-cessive or inappropriate and the bird never devel-ops the ability to be comfortable by itself. Birdsthat have not been taught independence are muchmore likely to develop stress-related behaviors.Most of the strategies a parrot uses to get along inlife are learned. Linden describes four distinct be-havioral stages, each with skills that must belearned in order to move to the next. Neonatesneed comfort, “neophytes” develop curiosity,fledglings develop coordination, and “thinklings”acquire decision-making capabilities.[18] Com-municating, eating, and learning how to play andwhat to be afraid of all must be learned for the birdto develop independence. While many types ofparrots are independent by nature, others mustlearn this from caregivers. When they meet a newsituation they look for cues as to how they shouldreact to the situation. If the caregiver is happy, the bird will not be afraid.[19] Parrots must betaught to be adventurous. Owners should modelplaying with the toys or add another bird or personto play, providing a model-rival.[19] These skillsare critical for the development of independent be-havior in pet birds. Many pet bird owners fear
spoiling a bird by responding to cries. With juve-nile birds displaying baby behaviors such as cry-ing, responding to the bird will not “spoil” the birdbut make it more secure.[19] Particularly in veryyoung birds, withholding food, attention, or othernecessities can make them doubt the safety of theirsituation. The common advice of ignoring a vocal-izing bird can contribute to a nuisance-screamingproblem. A bird that is ignored will continue toscream until something happens. Acknowledgingthe bird and maintaining some auditory contactcan minimize the begging and vocalizations. Vocalcues can be used to calm birds. The key is to planahead and consistently use particular words andexpressions in the same situations. This way theassociation between the word and the situation isclearly patterned. These words can then be used ina threatening situation to give a sense of secu-rity.[20] Vocal games can also promote independ-ence. Some games that are recommended to own-ers in the author’s practice include the following.These were devised from many suggestions andsources and are not necessarily original. They arewritten in the same way that they are presented inclient education materials.
HOUSE TOUR
Theory: In the wild, fledgling birds follow parentsand flock mates around their environment. Byseeing the response of the adult birds to variousstimuli, they learn what to eat, what to fear, whatto avoid, and so forth. This game is intended to dothe same thing.
How to play: The bird must be tame and mustknow the basic step-up command. Carry the birdon the hand and walk around the house. Point outeverything you see and say its name. Most impor-tantly, be very calm. By seeing that you are notupset, the bird will learn not to be. Don’t forget tointroduce all of the human and animal householdmembers. Also, do not neglect sounds. Take thebird near the source of some sounds and do thesame exercise. The bonus of this game is that talk-ing birds often learn how to identify people andthings in the house.
COLOR GAME
Theory: Parrots are very visually oriented and in-telligent creatures. This game helps stimulatetheir curiosity.
How to play: Take pieces of colored construc-
170 Manual of Parrot Behavior

tion paper. Say the color to the bird. Repeat for allof the other colors. Keep in mind that the birdsees colors slightly differently than you do butcan still distinguish them well. More advancedlessons will ask the bird what color. For even bet-ter results, do this game with another person infront of the bird. When the person gets the answercorrect, he/she is lavishly praised.
WHISTLE WHILE YOU WORK
Theory: In the wild, parrots vocalize to maintainaudio contact with members of their flock. Beingalone puts birds at greatly increased risk of preda-tion. Survival depends on maintaining contact. Ifthey cannot hear the response of the group, theythink they have lost contact, then they call louder.It is often said to never respond to a bird’s vocal-ization. Imagine the following scenario. You areat home alone and you hear someone come in thedoor. You think it is your spouse so you call outhis/her name but you get no response. You callagain and still no response. At this point you startto panic and get ready to call 911! This is what weare doing to the bird when we ignore its calls.
How to play: In order to take the flock contactinitiative away from the bird, announce where youare as you move about the house. This is espe-cially true if you are out of sight. Try whistling,humming, singing, or talking as you go.
TRICK TRAINING
Theory: Parrots are highly intelligent birds. Men-tal challenges can occupy some of the time theymay otherwise use for self-destructive behaviors.Additionally, trick training provides ammunitionfor counterconditioning. Tricks can and should berelatively natural behaviors that the bird learns todo on request.
How to play: Watch your bird for certain behav-iors that are interesting. Then start to give a cue, tryto get the bird to do the behavior, and reward evenmild attempts at performing it. As time goes on, re-quire a little better performance to receive a re-ward. Rewards can be verbal or food treats. Ideasthat may be useful include waving the foot, somer-sault on the perch or table, holding up wings, hold-ing up objects with the foot, or tearing up a toy.
Providing an Appropriate Environment
The environment can be very critical to the bird’sability to be independent. Birds may perceive
many things as dangerous that a large, predatoryspecies such as a human would not. Location inthe home should take this fact into account. Birdsshould be able to see approaching people and petsrather than having them appear from nowhere.Placing a cage on a wall right next to a door maybe more stressful for a nervous bird. Parrots mayeven be sensitive to the emotional “energies” ofthose around them and can suffer from this in-creased stress.[21] Such things as marital diffi-culties, spousal abuse, child abuse, and loss of aloved one, even one the bird does not know, mayindirectly affect some birds. These can be diffi-cult subjects to approach with clients and must behandled delicately. People should be advised toseek professional counseling if such factors areinvolved.
Bonding
The whole point of having a pet is to bond withthem in some fashion. While most birds’ roles ina household are simple companions, occasionallybirds are kept as surrogates for children, spouses,or parents. Sometimes birds, especially since theyare long-lived, can serve as a last link to a lovedone that has passed away.[22] When the behaviorof a parrot causes this bond to be severed, theresults can be devastating to the owner. Unfortu-nately, placement of these birds into these surro-gate roles can lead to an unhealthy social environ-ment. Ideally birds and owners interact as flockmembers, but not as mates. Social activities forbirds within a flock may include foraging, play-ing, flying as a group, and other relatively dy-namic activities. Allopreening and cuddling aredone primarily with the mate. In companionbirds, these interactions should be reserved forevenings and naptimes or when the bird is con-fronted with unfamiliar things.[19, 21] Manybirds will beg for attention. Behaviors birds use toseek attention include shaking toys, sneezing, softvocalization, displays, crouching and wing quiv-ering, staring, and screaming.[14] Interactive dy-namic attention given to a begging bird mightteach more appropriate behavior.[19] Healthy so-cial environments include social feeding (eatingwith the owners).[11] This can allow more social-ization in a more casual atmosphere. It also givesan opportunity to demonstrate independent eatingby eating in front of the bird and offering to sharefood.[19] Some species of psittacine birds are
15 / Behavior Classes: Preventing Problems Before They Start 171

noted to spend all of their waking hours with theirmates. “For such intensely social birds, life in anenclosure with no companionship must be the ul-timate ‘psychological torture.’”[6] Realistically,no one can provide 24-hour interaction with thebird. It is therefore best to avoid having the birddevelop a pair bond with a person. In addition,birds that develop pair bonds with an owner willtend to be more aggressive. It is a myth that somespecies are “one-person birds.” Interaction withonly one person in captivity is contradictory tothe psittacine social nature.[23] Birds should beencouraged to be accepting of multiple people. Ifreluctant, out-of-territory interactions such asstep ups, rescues, or outings are recom-mended.[19] Rescues involve such things as aperson who is not normally the favorite bringingthe bird for a veterinary visit or grooming.Following the procedure and in this “hostile” en-vironment, the person that the bird does not likestarts to look very safe and comfortable to thebird. Outings are similar but simply involve tak-ing the bird for a trip out of the home to an unfa-miliar area. Again, the person becomes the mostfamiliar thing to the bird. People disliked by abird should practice step-up exercises in an areaunfamiliar to the bird. This “home court advan-tage” will often result in a bird that behaves muchbetter for a person it would normally bite.
Parrots’ social behavior is both what makesthem endearing to people and what often leads toundesirable or self-destructive behaviors. By at-tempting to understand the ways in whichpsittacine birds interact with one another, bettermethods of socializing these birds in home envi-ronments are possible. A balance between affec-tion and independence is critical to psychologi-cally healthy pet birds.
Motivation and Reinforcement
The concepts of motivation and reinforcement areexplained to owners in the behavior class. Themotivation for undesirable behavior must be re-moved. Desirable behaviors must be reinforcedand undesirable ones must not be. Behaviors,both good and bad, exhibited by animals and peo-ple typically are initiated as a result of a particu-lar motivation. For parrots motivations are typi-cally simple things such as hunger, fear, play, so-cial climbing, reproduction, or other things thatwould benefit the bird in the wild. In behavior
modification, the motivation for a given behaviorshould be sought so that it can be removed or en-hanced, depending on the desirability of the be-havior. For example, the initial motivation forscreaming may be hunger, pain, flock cohesion,or fear. These motivations can be easily removedby feeding the bird, treating the painful stimulus,calling to the bird, or removing the source of fear.Behaviors, both good and bad, generally continueto occur because they are being reinforced.[24]Inadvertent reinforcement of undesirable behav-ior is common among pet owners, especially withparrots and dogs. The reinforcement for scream-ing may come in the form of an excited verbal re-sponse, physical attention, being left alone, oreven being given a treat of some kind. In additionto avoiding reinforcing bad behavior, it is impor-tant to reinforce good behavior. When a scream-ing bird is quiet, the owner should talk to or petthe bird.
Exercises
STEP UP
One of the basic needs of a pet bird owner is theability to transport the bird upon the hand. In orderfor this to occur the bird must learn to easily steponto the proffered hand. This is where the “stepup” command comes into play. The handler states“step up” and moves the hand toward the bird. Thehand must be very steady and positioned in a waythat it is easy for the bird to step up on it. If thebird is not schooled in this exercise, it may requiresome coaxing to get it up on the hand. Sometimesone foot can be lifted and placed up on the hand,which is then gently lifted up. The bird must thenbring the other foot up on the hand. As this processis repeated, the bird learns to step readily onto thehand. Some birds resist stepping up, not becausethey do not know what is requested but becausethey wish to exert control over the situation. Inthese cases, the handler should “follow through”with the hand, sweeping past the bird if it shouldhesitate. The motion is neither violent nor hesitant.It simply shows that the handler expects the bird to step up right away. This is one of the areaswhere these classes are very helpful. The instruc-tor can monitor the motions and mannerisms ofthe handler and offer constructive criticism anddemonstrate proper technique. It is remarkablehow differently the same bird responds to differenthandling techniques.
172 Manual of Parrot Behavior

STEP DOWN
Just as important to the bird owner is the ability toget the bird to step off of the hand. This is wherethe step down command comes in. The hand thatthe bird is perched upon is brought adjacent to theperch. The term “step down” is spoken. The birdis then encouraged to step onto the perch. The au-thor prefers to have the bird step backward ontothe perch. This makes the motion different fromthe “step up” motion, avoiding confusion. If thebird does not step onto the perch, the hand is low-ered below the level of the perch. If the hand is infront of the perch the tail will then bump theperch and most birds will step back onto theperch. If not, one foot can be gently placed on theperch, and the other will follow as the hand islowered.
STAY
This command is essential if parrots are to be al-lowed substantial time with the “flock” or family.The bird is simply placed upon the perch and theterm “stay” is spoken. The handler then moves ashort distance from the bird. Many birds will pos-ture at this point, indicating the desire to move offof the perch. At this point the “stay” command isrepeated. If the bird leaves the perch, it is replacedand the process is repeated. Increasing distancesand times are then used to teach the bird to stayupon a given site.
Hooding
Hooding is a term taken from the time-honoredfalconry technique of placing a leather cap orhood over the head of the bird to reduce its reac-tion to visual stimuli. Rather than placing aleather hood, in this case a hand is simply cappedover the head to guard the bird from perceivedthreats. Many birds will tolerate this immediately,while others will require desensitization. Ownersare told to proceed slowly to prevent making thebird more fearful.
Carrier
Placing a bird into a carrying cage is a commonmaintenance procedure. In some cases, however,it can be a traumatic event. The method combinesthe preceding techniques to put the bird easilyinto a front-loading pet carrier. The bird isstepped up onto one hand, the other hand iscupped over the head, and then the bird is placed
backward into the carrier. A “step down” and a“stay” command are given. When the bird stepsdown, the carrier can be closed.
Towel
Desensitizing pet birds to towels makes othercommon maintenance procedures less stressful.Clients are instructed to slowly accustom the birdto being touched with and eventually wrapped upin a towel. Again it is important to move slowly toprevent making the bird more fearful of towels.
Owners are instructed to practice these exer-cises for 15–20 minutes daily. The amount of timein the classes is insufficient for the birds to be-come proficient. The class time is really for in-struction of the owners. Their handling tech-niques should be honed so that the home trainingis more effective.
BENEFITS OF THE BEHAVIOR CLASSES
The performance level of the birds in the class hasbeen highly variable. Each bird, however, hasshown improvement throughout the class. Thebirds coming in with the poorest behavior oftenshow the most improvement. The owners gener-ally feel closer to the bird and have a better under-standing of the bird’s needs. Several have re-peated the class or had another family member gothrough the class.
The bird behavior class has been very well re-ceived by the public. About four sessions are runper year, and there has been no difficulty in fill-ing these slots. There have been between four andten members in each class. This class has been atremendous positive public-relations builder. Theauthor has improved his own exam room de-meanor by a greater understanding of parrot be-havior. A project with such positive aspects alsogives the instructor a lift in attitude. Clients bondwith their birds better as a result of the obedienceclasses. Better behavior on the part of the bird anda better understanding of parrot’s behavior on thepart of the owner keep the frustration of both to aminimum. Several owners have decided to keeptheir birds instead of selling or placing them.Several people have started to handle their birdsmuch more since taking the obedience class. Theyhave been able to establish dominance and nowthey can handle the bird without chasing it aroundor being bitten. When owners have to administertreatments to their birds, those that have been to
15 / Behavior Classes: Preventing Problems Before They Start 173

the class are able to comply with treatments bet-ter than before. One client that went through theclass had a conure that was very cage territorialand, whenever he was brought in for boarding, hiscage would be very dirty. After the owners hadtaken the class, they kept the cage much cleanerthan before and the bird was easier to care for inthe hospital. Most of the birds that have beenthrough obedience classes are much easier to han-dle for clinical examinations. The birds have beento the hospital without having unpleasant proce-dures, so they are not stressed by their mere pres-ence there. The other reason is that one of the ex-ercises in the class is to play with the bird using atowel. These birds often allow the towel to beplaced over them without a struggle. Obedienceclasses can also become a practice profit center.The material costs of running the class, includingperches, photocopying, and the videotape, areminimal. The only other cost is the time of theperson teaching the class. In the author’s practiceit is the veterinarian that teaches the class; how-ever, a well-trained and experienced techniciancould fill the instructor’s role. More importantthan the revenues from the class itself is the factthat clients who take this class bond with thepractice. They can see that their veterinarian isnot just interested in money or in the medical as-pects of birds. They can see that the bird’s well-being is considered important. This is somethingthey tell their friends and family .
REFERENCES
01. Neville, P.F. 1997. “Preventing problems via socialand environmental enrichment.” Proceedings NorthAmerican Veterinary Conference, pp. 31–32.
02. Anderson, R.K. 1990. Preventing needless deathsof pets: Putting dogs on their best behavior.Veterinary Forum, April, pp. 32–33.
03. Wilson, L. 1996. “Non-medical approach to thebehavioral feather plucker.” Proc Annu Conf AssocAvian Vet, pp. 3–9.
04. Voith, V.L., and P.L. Borchelt. 1985. Fears and pho-bias in companion animals. Compend Cont EducPract Vet 7 (3):209–218.
05. Evans, J.M. 1985. The Evans guide for counselingdog owners. New York: Howell Book House.
06. Harrison, G.J. 1994. “Perspective on parrot behav-ior.” In Avian medicine: Principles and applica-tions, ed. B.W. Ritchie, G.J. Harrison, and L.R.Harrison, pp. 96–108. Lake Worth, FL: WingersPublishing Inc.
07. Lafeber, T.J. 1983. Let’s celebrate pet birds. Odell,IL: Lafeber Co.
08. Ramey, K., N. Moore, and J.R. Millam. 1994.“Affiliative behavior in captive breeding amazonsparrots.” Proc Annu Conf Assoc Avian Vet, p. 434.
09. Jochim, L., and J.R. Millam. 1994. “Behavior oforange-winged amazon parrots before and afternest box presentation.” Proc Annu Conf AssocAvian Vet, p. 435.
10. Millam, J.R. 1994. “Environmental enrichmentstimulates egg laying in naive orange-wingedamazon parrots.” Proc Annu Conf Assoc AvianVet, p. 436.
11. Athan, M.S. 1993. Guide to a well-behaved parrot.Hong Kong: Barron’s.
12. Davis, C. 1997. “Behavior.” In Avian medicine andsurgery, ed. R.B. Altman, S.L. Clubb, G.M.Dorrestein, and K. Quesenberry, pp. 96–100.Philadelphia: W.B. Saunders Company.
13. Smith, I.L. 1999. “Basic behavioral principles forthe avian veterinarian.” Proc Annu Conf AssocAvian Vet, pp. 47–55.
14. Rach, J.A. 1998. Why does my bird do that: Aguide to parrot behavior. New York: Howell BookHouse.
15. Friedman, S.G., and B. Brinker. 2000. Early social-ization: A biological need and the key to compan-ionability. Original Flying Machine 2:7–8.
16. Horwitz, D. 1999. “Playtime: How to have funwith your pet.” Proc North Am Vet Conf, p. 31.
17. Athan, M.S. 1997. Guide to the quaker parrot.Hong Kong: Barron’s.
18. Linden, P.G. 1999. “Deliberate behavioral devel-opment: Stages, principles, and applications.” ProcAnnu Conf Assoc Avian Vet, pp. 269–271.
19. Athan, M.S., and D. Deter D. 2000. The Africangrey parrot handbook. Hong Kong: Barron’s.
20. Blanchard, S. 2000. Teaching your parrot self-soothing techniques. Pet Bird Report 9 (6):52–53.
21. Clark, P. 2000. The optimal environment: Part IV,the social climate. Pet Bird Report 9 (6):26–31.
22. Harris, J.M. 1997. “The human-avian bond.” InAvian medicine and surgery, ed. R.B. Altman, S.L.Clubb, G.M. Dorrestein, and K. Quesenberry, pp.995–998. Philadelphia: W.B. Saunders Company.
23. Wilson, L. 2000. “The one person bird-preventionand rehabilitation.” Proc Annu Conf Assoc AvianVet, pp. 69–73.
24. Case, L.P. 1994. “Motivation and reinforcement:The keys to solving behavior problems in dogs.”Proceedings Eastern Illinois Veterinary MedicalAssociation Spring Clinic, pp. 1–6.
174 Manual of Parrot Behavior

ESTABLISHING THECONSULTANT/CLIENT RELATIONSHIP
The most important facet to establishing a con-sultant/client relationship is to first create an openand honest rapport with the parrot owner. Withoutthis connection, it will be exceedingly difficultfor the behavior consultant or avian veterinarianto accurately access the situation, and withoutthis, it will be extremely difficult to assist in im-proving the situation. Judgmental language mustbe avoided at all costs, as this type of approachwill often encourage owners to be dishonest withthe interviewer in hopes of avoiding a negative re-sponse. This author (Wilson) finds that recount-ing her own mistakes with psittacine behavior canencourage owners to relax and talk openly.
IS THERE REALLY A PROBLEM?
Once a rapport has been established, the consult-ant must assess whether or not a problem actuallyexists. People often want to train their parrots tonot be parrots, such as wanting to teach them tobe quiet all the time, not to be destructive, and notto be so untidy. Parrots are inherently loud, de-structive, and messy creatures, and trying tochange them into something else is an exercise infutility. The end result is likely to be the bird los-ing its home.
Problems that require intervention with a con-sultation would include management problemssuch as an inappropriate environment (i.e., a par-rot living in a bar), or an inappropriate socialclimate (such as clients having combative rela-tionships with others in the environment). Addi-tionally, there are the problem behaviors them-
selves, such as self-inflicted damage (i.e., featherdestruction, self-mutilation), excessive fear, ag-gressive behaviors, and excessive vocalizations.
LEVEL OF COOPERATION FROMOTHERS IN THE ENVIRONMENT
The next step would be to evaluate the amount ofassistance the consultant can expect from theowners. Do all the people in the environmentwant to resolve the problem with the parrot, orwould they prefer the bird be gone? While someproblems, such as biting, can be resolved on aone-on-one basis, problems such as excessivescreaming require the cooperation of everyone inthe environment. If even one person rewards un-acceptable behaviors (such as the husband whowould give his wife’s African Grey Parrot [Psitta-cus erithacus] a piece of cheese to shut it up),then no improvement will be realized. Odds in-crease exponentially for a successful denouementif consultations include all family memberswhenever possible, especially with excessivenoise.
Many behaviors in pet parrots may not initiallyseem problematic to owners, until further ramifi-cations develop. A good example of this is theone-person bird. Many people are tremendouslyflattered by a bird’s exclusive affection, and actu-ally relish this situation, often (passively or other-wise) allowing the bird’s aggression toward othersin the household. These people usually do notseek assistance with this type of situation until thebird manifests further evidence of these problembehaviors, such as with feather destruction. Someowners will blatantly encourage aggression to-
175
16Clinical Evaluation of
Psittacine Behavioral Disorders
Kenneth R. Welle and Liz Wilson

ward others, for example, finding it amusing towatch a little bird chase their spouse, and manybehavior consultants believe that human laughteris a powerful reinforcement for the companionparrot.
An extremely common underlying factor withproblem behaviors in companion parrots is that ofinappropriate bond formation, as in overdepen-dencies and/or the formation of a “pair bond” or“mate bond” with an inappropriate species like aperson. Unfortunately, much of the lay literaturestill encourages this, and most owners see nothingwrong with “being their bird’s mate.” At least theysee nothing wrong until the confused and ex-tremely frustrated parrot begins manifesting un-acceptable behaviors such as feather destruction,excessive screaming, severe aggression, and, insome cases, even self-mutilation.
It is an unfortunate reality that many people donot wish for their baby parrots to mature. Theconcept of neoteny is a powerfully appealing one,and many owners try to maintain their relation-ships on the level of cuddling instead of allowingthe bird to mature and grow into the entity naturedesigned it to be. These people apparently hope tomaintain themselves as a “bird mommy” forever,and frequently comment that they “want theirsweet baby back” when their parrots start to ma-ture. However, this approach is guaranteed to leadto a dysfunctional relationship.
Fear can be a problem with companion parrots,whether the bird fears the person or vice versa.Like all companion animals, parrots will formdifferent relationships with different people intheir lives, and some relationships are more com-bative than others. While biting is a commonproblem with companion psittacids, it is often notdifficult to resolve, depending on the etiology.However, it is not easy for people who are afraidof parrots to control their charges. Parrots will notbe comfortable with people who are not comfort-able with them, and it is difficult to train parrotsto be tolerant of people who are awkward and illat ease around them. As prey animals, it is logicalthat they cannot relax when humans, for example,offer a hand to step up and then jerk the handaway when parrots reach with their beaks to sta-bilize the perch.
Occasionally, one encounters clients who, de-spite problem behaviors with their parrots, areaverse to the idea of modifying what they per-
ceive as “natural” behaviors. These people areoften under the mistaken impression that allpsittacine behaviors are instinctual, which is farfrom the case. Like most intelligent animals, par-rots have a tremendous capacity for learning andadapting their behavior in response to environ-mental input.[1] In the wild, other flock memberseducate these birds. In captivity, unless educatedby the humans with whom they live, psittacinebirds have no idea how to behave. As a result, theyoften develop displacement behaviors that hu-mans find distasteful, such as excessive scream-ing, feather destruction, and aggression.
A frustrating reality is the owner who has, con-sciously or otherwise, already decided to give upa parrot prior to seeking assistance in behaviormodification. In this situation, working with a be-havior consultant and/or avian veterinarian hasthe only function of assuaging the owner’s guiltby allowing him or her to believe that he or shehas “tried everything.” Not surprisingly, a behav-ior consultation will not change the outcome inthis situation, as the odds are against the ownerbeing sufficiently motivated to make the neces-sary changes to accomplish anything positivewith the bird.
TAKING THE BEHAVIORAL HISTORY
Unlike with medical problems, there is no testingthat one can do to assist in evaluating behavioraldisease. As a consequence, the behavioral historyand observation are the only diagnostic toolsavailable for evaluation of behavioral disor-ders.[2] However, problems arise when trying toevaluate psittacine behavior when the animal is ina veterinary clinic exam room, as the strange sur-roundings can substantially modify the animal’sbehavior. The same is true, to a lesser degree, ofthe behavior consultant doing an in-home visit,and this can somewhat limit the value of directobservation. However, within those limitationsmuch can be learned if the consultant is astute.
As with any properly done physical exam, theexaminer needs to work with consistency and astandardized form. A sample form is included inAppendix 16A. This enables the examiner to col-lect data in a somewhat organized manner and de-creases the odds of forgetting to ask an importantquestion. Some consultants prefer to send clients aform to fill out prior to the first appointment. Thiscan save a tremendous amount of time, which is its
176 Manual of Parrot Behavior

greatest advantage. This can also weed out thosewho are not sufficiently dedicated to followthrough, thereby saving time and frustration forthe consultant. However, much can be learnedwith spontaneous answers, especially when deal-ing with more than one person in the environment,as contradictions can be confusing but also ex-tremely revealing. As a consequence, other con-sultants prefer to ask the questions, watch and/orlisten carefully to the way questions are answered,and then transcribe the information. This methodis extremely time-consuming but can be quitevaluable, as the client’s tone of voice and bodylanguage, if present, can be quite telling.
EVALUATING THE ENVIRONMENT
For evaluating a parrot’s environment, photo-graphs can be useful, but nothing surpasses actu-ally being there and walking through the space.Consultants can also request a somewhat-to-scalesketched floor plan of the room or rooms in whicha parrot spends time (M.S. Athan, personal com-munication, 1998), which will aid in visualiza-tion. Clients are requested to label windows,walls, skylights, doors, and the bird’s cage, plusimportant pieces of furniture like the television,the sofa, and the client’s favorite chair. Clients arealso asked to use a dotted line to indicate the traf-fic flow through the room(s). For in-hospital con-sults, video tours are valuable. Watching thevideo with blueprint in hand can give an adequateimpression of the space in which a parrot lives. Inaddition, when it comes to truly evaluating a par-rot’s environment and its interactions with the hu-mans around it, this author (Wilson) has found theuse of videotapes to be priceless.
GETTING HONEST ANSWERS
It is critical that the examiner get as much infor-mation as possible about a situation, and this isnot always easy. Taking a good behavioral historyis quite an art form, requiring tact and a delicatetouch. The interviewer must avoid judgmental be-havior totally, as an accusatory approach can in-duce some clients to lie to avoid condemnation.As with any history taking, the interviewer mustalso avoid the use of leading questions, as thesecan encourage some clients to give the inter-viewer what they think he or she wishes to hear,rather than the truth.
It may be useful to start an interview with a re-
minder that the value of the consult is dependenton the clients being willing to provide completeand honest answers, and that it is not the inter-viewer’s job to judge—only to get as much infor-mation as possible to increase the likelihood ofbeing able to help improve the situation.
If the examiner is sympathetic and non-accusatory, the likelihood increases that honestanswers will result. For instance, in a situationwhere physical abuse was likely but unstated, aconsultant might talk about how awful it is to bearound, for example, a screaming parrot. By com-menting that the gut-wrenching noise makes ithard not lose one’s temper and throw something ata screaming parrot, the consultant might succeedin prompting the owner to admitting to havingdone just that.
OBSERVATION OF BEHAVIOR
Observation of the patient’s behavior can be themost challenging part of behavior work. The dif-ficulty is that the method used to observe the be-havior can add artifacts to the observations.Location and the very presence of the observermay influence the bird’s behavior. Observation inthe veterinary examination room can reveal how abird responds to a frightening situation. Birds thatcalmly preen their feathers while in the examina-tion room are generally birds that tolerate changewell and are not afraid of strangers. Aggressivebirds may be much more docile away from theirhome territory and will often allow handling bypeople they will ordinarily bite severely. Observa-tion of a bird in its home environment during ahouse call is somewhat better. Here the bird is infamiliar territory and will have fewer distractions.This method also allows the consultant a first-hand view of the environment. The consultant,however, will influence the behavior to some ex-tent. Most birds alter their behavior somewhat inthe presence of unfamiliar people.
Video recording the bird’s behavior may be themost effective means. Not only can the consultantview the interaction between bird and owner, butalso the camera can be set up to record the behav-ior of the bird when the owner leaves the area.This can be extremely useful information. Ani-mals with separation anxiety often exhibit signsof distress in the initial minutes following the exitof the owner. This is frequently observed invideos of pet birds.
16 / Clinical Evaluation of Psittacine Behavioral Disorders 177

Experience dictates the necessity of limitingthe amount of tape a client sends, and clients areasked to send no more than two hours of tape,with the explanation that otherwise, there won’tbe time to watch it all, so something importantmight be missed. The following is part of the in-formation this author (Wilson) sends to prospec-tive phone clients regarding what to film:
One of the primary purposes of the film isso I can watch a parrot’s body language, sotry to avoid things like filming your parrotagainst a sunny window during the day, etc.Telephotos are ideal, because then I can seethe bird more clearly without your having tostick the camera in its face. If you can’t filmin ideal circumstances, then just do the bestyou can. Whatever you send will be betterthan nothing. Just remember that the better Ican see the bird, the better I might be able tounderstand what is going on with it.
Also, film your parrot’s body language asit interacts with you in a variety of circum-stances (i.e., eating, playing, snuggling,etc.). Do the same with anyone else who rou-tinely interacts with the bird. Film the bird’sbody language when it is hanging out in thecage (i.e., eating, playing, etc.), both whenyou are in the room and when you are not.This part will require a tripod so the cameracan run without you. Film the bird’s bodylanguage as you interact with other familymembers.
Lastly, please film the view that the birdcan see from its cage, panning slowly aroundthe room. Be sure to set up the camera two tothree days prior to filming, so the bird willget accustomed to its presence. This is ex-tremely important because otherwise theparrot won’t relax and act normally.
The following case illustrates the value of eval-uating videotapes. A client thought his three-year-old female African Grey was screaming when heleft the room as a manifestation of a control issue,and from what the client described, the consultant(Wilson) agreed. As requested, the client filmedhis interactions with the Grey and then left thecamera running after he left the room. The clienthad watched the tape prior to sending it but didnot see anything that changed his interpretation of
the bird’s behavior. However, this author (Wilson)saw something totally different when she re-viewed the tape. Instead of showing irritation atthe human ignoring its calls, the young Grey waspanicking, chewing on her nails and flipping herwings, which this author feels can be evidence ofstress in a Grey. So instead of a control issue, thisbird was apparently suffering from a psittacineform of separation anxiety. If the author hadbased her recommendations solely on the client’sinterpretation of the situation, then she wouldhave given incorrect information as to how to re-solve the conflict.
OWNER’S BEHAVIOR
It is very important for the consultant to make ob-servations of the owner’s behavior as well. Veryoften, it is not the bird that is behaving poorly.Some owners simply do not know how to handletheir bird properly. Simple adjustments in themanner used by the owner can immediately solvesome problems. Owners that continually makeexcuses for the behavior of the bird or who makeunreasonable concessions to avoid being attackedmay be the cause of aggression in a bird. Whenthe bird sees that biting effectively controls theowner, the biting is reinforced. It is often startlingwhat owners think is reasonable. One of the au-thors (Welle) has had clients who admitted to tak-ing all phone calls in the garage to avoid beingbitten, or others who rolled on the floor to get abird off of the shoulder.
EVALUATING THE DATA
Once the data has been collected, it should becarefully evaluated for information that may leadto a clinically relevant conclusion. Veterinary be-havior as a clinical science is still in its infancy.This is particularly true with respect to pet birds.The etiologies of behavioral disorders, the mini-mum standards for the care of companionpsittacine birds, and the reference values for the“normal” responses of parrots to various situa-tions have not been established. These facts canmake the interpretation of the behavior historydifficult. Nonetheless, trends can sometimes beidentified. Associations between anamnestic fac-tors and certain behavioral traits sometimes canbe made. This is the means by which clinical sci-ences develop. More importantly, evaluation ofthe history allows a customized plan of environ-
178 Manual of Parrot Behavior

mental and behavioral modification to be formu-lated for the patient at hand.
SIGNALMENT
Signalment is as important in behavior as in othermedical fields. Species, age, and gender can giveclues to the type of problem that may be present.Parrots that mature rapidly (e.g., Budgerigars,Cockatiels) are somewhat less likely to developcertain behavioral traits than those that maturevery slowly (e.g., large cockatoos, macaws). Thesesmaller, rapidly developing species must learnmaintenance and social behaviors in a fairly shortperiod of time and therefore lack the extreme flex-ibility of learning that slow-developing speciespossess. The slightly more rigid learning patternprevents the development of some undesirabletraits. Some species such as cockatoos tend tobond by means of physical touch, allopreening,and other contact, while others, such as amazonparrots, tend to use vocalization, visual display,and other means to bond. These tendencies will at-tract different types of owners and will often leadto different types of behavior. Many species aregenetically identical to the wild counterparts whilea few (Budgerigars, Cockatiels, Peach-facedLovebirds) are truly domesticated. Some speciesappear to be more prone to develop aggression;others are more prone to anxiety or fear. Not all ofthese traits are genetically determined, and it is im-portant not to automatically label a problem basedsolely on the species, but the species can give aclue to the likely source of a behavioral disorder.
The age of the bird is also important. Some be-haviors may be normal phases of development ina young bird but are not normal later in life. Manyparrots will go through a phase of regurgitation asthe crop shrinks during weaning. Begging behav-iors are normal in nestling and fledgling birds butshould diminish as they mature. Many behavioraltraits are blamed on reproductive hormones.Before making any such claim, it must be deter-mined whether the patient has reached sexual pu-berty. For small psittacine birds, this will occurbefore they reach a year of age, while in mediumto large parrots, reproductive hormones are not asignificant factor until they reach two to fouryears or more in age. Likewise, behavior prob-lems are not likely to be primarily of sex hormoneorigin if they begin many years after the bird hasbeen sexually mature.
The gender of many psittacine patients is notknown. While in some instances it may have littlerelevance, some behaviors will be very dependenton the sex of the bird. The sex of the bird shouldbe determined when reproductive influence issuspected. During breeding season, wild maleparrots will generally find and guard a nest site.For a pet bird this natural trait may lead to territo-rial aggression.
DEVELOPMENT
More important than signalment is the back-ground of the bird. Parrots are altricial birds,completely helpless at hatching. Their behaviorpatterns are very open at this point and the periodbetween hatching and independence is critical.Early development may affect a bird’s confidenceand sense of security. It can determine a parrot’sintelligence, flexibility, and potential as a com-panion animal. During its time with parents, awild baby parrot learns what to eat, what to fear,how to travel, and how to interact with other flockmembers. The same is true in domestic-bred com-panion parrots, except that in many cases the birdhas human surrogate parents (see chapter 11).
Hand-rearing baby parrots has long been usedboth as a means of increasing production ofchicks and as a means of raising psittacine birdswith little fear of humans. While the techniquesfor hand-rearing physically healthy psittacids hasbeen evolving for a long time, the process of so-cializing and educating these birds has often beenignored. It is evident that hand-rearing of parrotsalone does not prevent fear of humans. Juvenilebirds must be socialized rather than assembly-linefed. Psittacine birds that are raised with no clutch-mates and receive minimal physical contact withthe caretaker will not often develop into well-ad-justed pets. These birds may not learn to socializewith other birds or with humans. Many will notlearn to play or explore their environment. Anysynapses that are not stimulated by early sensoryexperiences may be “pruned” by the brain andeliminated.[3] Many of these birds imprint on hu-mans as both parents and potential mates, whichcan lead to many problems in older parrots. Asstated by Harrison in Avian Medicine: Principlesand Applications, “Birds raised by human fosterparents will imprint as people, not birds. As theymature, their natural instincts to choose a matemay cause objectionable behaviors. . . . An im-
16 / Clinical Evaluation of Psittacine Behavioral Disorders 179

printed bird will spend all of its time attemptingto drive unwanted individuals, other pets or ob-jects out of its territory, while trying to find onechosen person with whom to mate.”[4] Thisspecies confusion is compounded when thehuman caretaker fails to continue the bird’s edu-cation to include necessary skills for coping withthe artificial environment. Psittacine birds thatare hand-reared and that are not sufficiently so-cialized with other conspecifics may have dif-ficulty in relating to other parrots in aviculturalsettings.[5]
In contrast, parent-raised psittacids that do notreceive any human contact until after they leavethe nest box are little different from a wild-caughtfledgling in behavior. They have no previous ex-perience with humans and will learn how to reactby watching and emulating their parents. If theparent birds have remained tame pets, the off-spring will often tame easily after fledging. Manybreeding birds, however, are either untamed orhave become aggressive or indifferent to humansduring the breeding cycle, and their offspring willbe much more difficult to tame. Perhaps an idealsituation is for baby birds to be raised by parentsbut handled regularly by humans prior to fledg-ing. While this may seem like a simple solution, itmay be difficult to accomplish. Some breedersmay abandon the nest if they are disturbed exces-sively, some may injure or kill their offspring intheir attempts to guard the nest box, and somemay severely injure the person attempting to in-vade the nest. Some situations in a bird’s historymay predispose it to anxiety or insecurity.Psittacine birds that are weaned too early, forcedto wean before they are ready, or sold unweanedto inexperienced handlers may have a difficulttime trusting humans. Pet parrots are often sold atvery young ages under the pretense of forming a“stronger bond” to the new owner. These birdshave already been removed from the natural par-ents, and then before they even have a chance tofinish growing, they are taken away from theirnew “parents.” It does not take these birds long tofigure out this pattern.
GROOMING
Veterinarians or other groomers of pet birds cancause substantial problems. Unfortunately, manyfeel that a wing trim, nail trim, or beak trim is aninnocuous procedure and will perform it without
instruction on technique or case selection.[6]Improperly performed these methods can lead tolong-term problems. First, the trauma of a painfulnail trim or beak trim can make even a hand-raised baby bird fearful of people. Secondly, a se-vere wing or nail trim can affect the balance andcoordination of the bird. Likewise, single wingtrims will unbalance a bird. Some birds fall fre-quently, leading to more fear and pain. When ex-cessive numbers of feathers are cut, the newfeathers that come in have no protection andtherefore are prone to damage. A vicious cycleensues, with terrified birds becoming fearful ofany human contact because they are afraid offalling and injury. Finally, flight is a learned skilland can lead to a greater sense of confidence, andeven intelligence.[7] Trimming the wings, espe-cially very early, can deprive birds of this neces-sary development.
Routine flight feather and nail clipping are nec-essary for most companion parrots, but the overlyaggressive handling and grooming of the pasthave been dramatically modified in the last fewyears, with more humane methods replacing thesepotentially destructive, outdated techniques.
The technique of swooping down from behindto capture a parrot in a towel is no longer re-quired. This technique, aptly named the “HarpyEagle Catch” by Blanchard, was developed whencaptive parrots were all wild-caught imports.[8]With hysterically terrified untamed birds, speedwas at a premium and it was critical to get the par-rots under control quickly, before they could in-jure themselves or their handlers. However, mostparrots in the United States today are domesti-cally raised and do not perceive humans as deadlypredators. Hence, the Harpy Eagle Catch is notonly unnecessary, it can be seriously damaging.Speer states that “The Harpy Eagle grab causesfear-induced behavioral disease”.[9]
Thanks to Blanchard’s work, one of the authors(Wilson) has developed what she calls the“Frontal Towel Approach.” This approach doesnot use a predatory attack to get control of a par-rot, so it is substantially less stressful for the bird.The eyes of prey animals like psittacine birds arelocated on the lateral aspect of the skull, so theirperipheral vision warns them of a forthcomingpredatory attack. Seeing the towel coming withthe Harpy Eagle Catch appears to throw a birdinto a full fight or flight response as it is captured.
180 Manual of Parrot Behavior

Once this systemic response is initiated, the birdwill frantically struggle to escape what it likelyperceives as a deadly situation.
The Frontal Towel Approach does not initiatethis terrified reaction. To place a tame parrotunder restraint using this method, the handlerdoes the following. Talking quietly to the bird (in-stead of the owner), the patient is stepped onto ahand, and the bird’s feet are gently but firmlypinned by the handler’s fingers (Figure 16.1).Continuing to talk softly and keeping the towellow, the handler catches one corner of the towel inthe fingers of the hand on which the bird is sitting(Figure 16.2), then smoothly brings the towel upand around behind the bird (Figure 16.3), andlowers it over the bird’s head (Figure 16.4). Theanimal is then gently put under restraint (Figure16.5). If the handler moves smoothly from thepinning of the feet to the towel’s approach, thisauthor (Wilson) has found that parrots pay littleor no mind to their feet being pinned, as thetowel’s approach consumes their attention.
16 / Clinical Evaluation of Psittacine Behavioral Disorders 181
Figure 16.1. The parrot is stepped onto thehand and the handler greets the bird in a friendlymanner.
Figure 16.2. The parrot's foot is gently butfirmly pinned with the thumb while the handlercontinues to talk softly and smile.
Figure 16.3. The handler catches one cornerof the towel between the fingers of the hand onwhich the bird is sitting. Friendly talk and smilescontinue.

Even a parrot that is terrified of towels cangenerally be captured utilizing this technique. Afrightened bird tends to flip backward from thehand as the towel approaches, but since the han-dler has the feet pinned, it cannot fall or escape.The towel is then wrapped around the upside-down bird, it is lowered to the table, and restraintis commenced as usual. While this approach isunquestionably more stressful than the previousone, the bird will still relax faster than it wouldwith a predatory capture. Since avian veterinari-ans have been discussing the dangers of stress for
decades, it seems reasonable to make every effortto avoid stressing the avian patient more thannecessary.
Important note: There is danger of iatrogenicfractures when small birds like Budgerigars (Melo-psittacus undulatus) and Cockatiels (Nymphicushollandicus) flip backward. In situations such asthose, this author (Wilson) immediately releasesthe feet so the bird escapes. Due to this potential,the toweling of small birds is often better accom-plished via the old style grab from behind.
For that very reason, many educated bird own-ers have patterned their birds to be unafraid ofbeing wrapped in towels. In the veterinary examroom, these owners will wish to towel their birdsthemselves, and then hand the towel-wrappedparrot to the veterinarian or veterinary technician.They will be justifiably disgruntled if a clinicianchooses instead to undo this training with apredatory grab. If the veterinarian is not comfort-able having the owner wrap the bird in this man-ner, then the veterinarian or technician can ac-complish the towel wrap in the same friendly,non-aggressive manner.
182 Manual of Parrot Behavior
Figure 16.4. Continuing to talk calmly with afriendly facial expression, the handler then usesthe other hand to slowly and smoothly bring theother end of the towel up . . .
Figure 16.5. And the parrot's head is gentlycovered. Full restraint can now commence.

It should be noted that, as with any restrainttechnique, practice is necessary for personnel tolearn how to accomplish this maneuver confi-dently and smoothly. New handling techniquesshould never be attempted with compromised pa-tients. Many hospitals have organized handlingworkshops for their personnel to develop confi-dence and proficiency with this capture methodusing their own pet birds prior to trying it withclients’ birds.
As far as this author (Wilson) is concerned, it isabsolutely unnecessary for tame birds to begrabbed from behind or by first darkening theroom. This author has been using this frontal ap-proach for over a decade and has yet to be bittenusing it. More importantly, most parrots seemdramatically less stressed by restraint when cap-tured in this manner.
PRIOR ENVIRONMENTS
Very often, birds that are presented for behaviorproblems are no longer with the family that origi-nally purchased them. It is very common for a newperson to adopt such a troubled bird assuming theproblems must be due to abuse or neglect. Whileit is true that prior owners and environment mayhave caused or contributed to the current problem,it is not always due to a lack of trying. When theproblem began before the current owner took pos-session of the bird, it is much more difficult toevaluate the root causes of the disorder. Some-times the information regarding the previoushome is available, but is often given in a somewhatslanderous fashion. The consultant should drawhis or her own conclusions about the prior situa-tion rather than taking the new owner’s word thatit was abusive. Parrots are social animals and needinteraction with other birds or with humans. Of-ten, due to fear, lack of time, or other factors, theseneeds are neglected. In rarer cases, teasing or evenphysical abuse may occur. These situations willvery often have severe effects on the behavior ofthe patient, with manifestations often dependingon the personality of the bird. Some may becomeaggressive, others fearful.
Just as common as neglect or abuse are situa-tions where overbonding or pair bonding with theprevious owner has occurred. This is particularlytrue with cockatoos, since people buy these birdsbecause they love to cuddle. They therefore in-dulge in cuddling and petting at the expense of
more active and independent play. The end resultis a bird that pair bonds to a human, expectingconstant and exclusive contact. When the personcannot meet these expectations, problems arise.When the new client adopts this bird, thinking theproblem was due to a lack of attention, the cyclestarts all over.
PHYSICAL ENVIRONMENT
An important part of the behavior consult is theevaluation of the physical environment of parrots.Factors that must be evaluated include size, con-struction, and cleanliness of the cage. Most birdsspend the majority of their time caged and poorconditions can be extremely stressful. The loca-tion and surroundings of the cage can be criticalas well. Some common household objects orsounds may be perceived as threatening to a bird.As prey species, parrots are naturally suspicious.Things that look completely innocuous to hu-mans, such as clocks, portraits, or animal pic-tures, may cause anxiety in predisposed birds.Very loud noises are stressful to some birds, buttotal silence may encourage more severe reac-tions to normal household noises. Children andother pets can be very threatening to a bird. Erra-tic movements, staring, predatory tendencies, orjust a lack of respect for personal space presentnot only a perception of threat but also potentiallya real danger. Lack of any visual stimuli may leadto boredom and inflexibility. Cigarette smoke orother air quality issues can also result in stressesas well as causing health problems. Some areasmay not allow a bird to “let down its guard.”Cages that are placed in the center of a room forcea bird to watch for danger in every direction. Thesame applies if the cage is placed against a largepicture window or glass doors. If a doorway is onthe same wall as the cage, surprise entry into theroom can startle a nervous bird. These findingscan add supportive evidence for anxiety-relatedbehavior problems.
Birds that are isolated from all family activitieswill not develop normal social behaviors. If birdsspend all of their time in one spot they tend to beeasily stressed by changes. These birds may alsobe more likely to become territorial of theircage.[10] Conversely, if time is spent in variousother locations, the bird may be more flexible.Perches or play gyms in an area of high activitycan suggest that the bird has more casual social-
16 / Clinical Evaluation of Psittacine Behavioral Disorders 183

ization and sensory variation. Sleep cages in aseparate room ensure sufficient dark and quiet forsleeping. Some birds commute regularly outsideof the home. Traveling can also provide stimula-tion but can be stressful as well. The conditionsunder which the patient travels and the personal-ity of the individual bird should be taken intoaccount.
SCHEDULE
Most parrots are tropical or subtropical, wheredays are divided into 12 hours of light and another12 hours of darkness. Birds getting less than eightto ten hours in a dark, quiet room are oftenstressed by sleep deprivation. Owners that simplycover the cage and yet continue to watch televi-sion in the room are deluding themselves intothinking that the bird is really sleeping.[11]
TOYS
There are several basic types of toys. Foot toys arethose that are manipulated by the bird’s feet;chewing toys are those that birds destroy or ma-nipulate with the beak; climbing toys are thosethat the bird climbs, hangs, or swings upon; puz-zle toys are those that require a bird to analyze,solve, and complete a task to receive a re-ward.[12] Some toys can serve as more than onetoy type. Ideally, at least one of each type shouldbe available to birds all of the time. When evalu-ating toys available to a patient, the type of toy, itssize, construction materials, and suitabilityshould be considered. Some toys intended aschew toys are too difficult for the bird to destroy,making it relatively uninteresting. The toysshould match the size and preferences of the bird.Also important is the frequency with which thetoys are changed. A playful bird will easily be-come bored with the same toys constantly avail-able. Birds vary in their enthusiasm regardingtoys. Birds that do not play with any toys mayhave never learned to play with toys. These birdsare often more “dependent” on the owners.
SOCIAL ENVIRONMENT
Social interactions should be evaluated as well.Birds that have interactions primarily with oneperson will often become “one person” birds andare more likely to pair bond with this individual.Birds that interact with several people are lesslikely to do so. These birds are often much more
independent. Nervous or anxious parrots andthose pair bonded to owners often tolerate “alonetime” poorly. This separation anxiety can lead toexcessive vocalization or feather picking.
BEHAVIOR DESCRIPTION ANDOBSERVATION
Finally, the owner’s description of the behavior ofthe bird should be evaluated. These descriptionsmay be tainted by the owner’s interpretation of thebehavior, so it is important to emphasize to theowner that raw observations are what is desired.The overall behavior should be described in addi-tion to any behavioral problems. The way inwhich the bird responds to various householdmembers as well as to strangers, new situations,and other stimuli can give a great deal of informa-tion about the personality type of the bird and thepossible motivation for various behaviors. Birdsthat are indifferent or frightened of people are un-likely to be using behaviors as a means of attract-ing attention. Likewise those that exhibit strongpositive responses to all household members areunlikely to be trying to avoid attention. Birds withpositive responses to some household memberswhile they have negative responses to others maybe overly dependent on one person. Birds that re-spond negatively to new people, places, or toysare often in a near-constant state of stress. Thepresence of certain sexual behaviors, such as re-gurgitation, masturbation, or nest building, cansupport a theory of a reproduction-associatedproblem.
A description of the problem behaviors and thecircumstances in which they occur is very impor-tant since many behaviors occur only at certaintimes or in certain situations. From a descriptionof the trigger, the behavior, and the consequencesof the behavior, the consultant must determinewhat is actually occurring. This may be very sim-ple in some cases, but can be very difficult inother situations. Owners often lack the ability toaccurately describe a behavior. Occasionally aparrot will be presented to a veterinarian becauseof seizures when actually the “fits” are masturba-tion. Some types of behavior, such as regurgita-tion, can be normal in some circumstances butabnormal in others. It is important to get as manydetails about the behavior as possible. In addition,the frequency, the severity, and the timing shouldbe recorded. Association with certain times or sit-
184 Manual of Parrot Behavior

uations may give a clue as to cause, but it alsohelps to document these factors so that progressof treatment plans can be evaluated.
The owner’s response to the behavior should bedocumented as well. In some cases the reactionsmay be reinforcing the behavior the owner wishesto eliminate. Reinforcement can come in manyforms, some of which do not match the motiva-tion the bird had for initiating the behavior. Food,drama, attention, or sometimes avoidance can bereinforcement in various situations.
More specific information to evaluate for spe-cific behaviors can help narrow down the prob-lem as well. For instance, with feather-destructiveparrots, it is important to find out what parts ofthe body are affected, whether the bird pickswhen alone or when around people, whether otheractivities are interrupted while picking, andwhether the picking seems to cause pain. Thisdata can be critical when trying to diagnose theproblem. A bird that only picks when alone maysuffer from separation anxiety; a bird that inter-rupts other activities may have a true skin disor-der that itches. These distinctions can help narrowdown the hundreds of possible causes of a com-plicated problem.
DRAWING CONCLUSIONS
When all of the data has been evaluated, someconclusions must finally be drawn. Behavior isnot an exact science and no two patients are ex-actly alike. Each diagnosis should be considereda theory. The validity of the theory can be testedby correcting the likely etiologic factors, applyingbehavior modification, and, in some cases,through the use of pharmaceutical therapy.
FOLLOW-UP
The evaluation does not end here. Follow-up tothe problem is critical. Owners should be asked tostart a journal of the bird’s behavior. The fre-quency, severity, and timing of the episodes of un-desirable behavior should be tracked. Any corre-sponding factors that are recognized should betracked as well. This journal can have several ben-efits. The first is that the effectiveness of treat-ment can be documented. Most behavior prob-
lems that are successfully treated do not simplystop one day. They gradually attenuate in fre-quency and severity over time. If there is norecord, owners may become discouraged and failto comply with recommendations for sufficienttime to resolve the behavior issues. The secondbenefit is that the additional data determined bythe record keeping may add information that canhelp further define the problem.
REFERENCES
01. Welle, K.R. 2002. “Psittacine social structure:Applications to companion birds.” Proc AnnuConf Assoc Avian Vet Specialty program.
02. Overall, K. 1997. Clinical behavioral medicine forsmall animals. Saint Louis: Mosby-Year Book,Inc., pp. 77–87.
03. Friedman, S.G., and B. Brinker. 2000. Early social-ization: A biological need and the key to compan-ionability. Original Flying Machine 2:7–8.
04. Harrison, G.J. 1994. “Perspective on parrot behav-ior.” In Avian medicine: Principles and applica-tion, ed. B.W. Ritchie, G.J. Harrison, and L.R.Harrison, pp. 96–108.Lake Worth FL: WingersPublishing Inc.
05. Styles, D.K. 2002. “Captive psittacine behavioralreproductive husbandry and management: Sociali-zation, aggression control, and pairing tech-niques.” Proc Annu Conf Assoc Avian Vet Spe-cialty program.
06. Speer, B.L. 2001. “The clinical consequences ofroutine grooming procedures.” Proc Annu ConfAssoc Avian Vet, Orlando, FL, August 22–24, pp.109–115.
07. Linden, P.G. 1998. “Behavioral development of thecompanion psittacine bird.” Proc Annu Conf AssocAvian Vet, pp. 139–143.
08. Blanchard, S. 1994. Trust building towel handlingtechniques. Pet Bird Report 14:36–37.
09. Speer, B.L. 2001. “The clinical consequences ofroutine grooming procedures.” Proc Annu ConfAssoc Avian Vet, Orlando, FL, August 22–24, pp.109–115.
10. Blanchard, S. 1990. Phobic parrots. Bird TalkMagazine 90/8 (8):64–73
11. Athan, M.S. 1993. Guide to a well behaved parrot.Hong Kong: Barron’s.
12. Wilson, L. 1996. “Non-medical approach to thebehavioral feather plucker.” Proc Annu Conf AssocAvian Vet, pp. 3–11.
16 / Clinical Evaluation of Psittacine Behavioral Disorders 185

This chapter will focus on the diagnostic workupof two behavioral problems commonly encoun-tered in avian practice, feather-damaging behav-ior and aggression.
FEATHER-DAMAGING BEHAVIOR
“Feather-damaging behavior” (FDB) is a termthat describes those conditions where the com-panion bird plucks out, destroys, or barbers itsown feathers (B. Speer, oral communication, Feb.2003). It is often referred to as feather picking,but picking may be only a component to the en-tire process, so FDB is a more inclusive term.However, FDB represents a symptom and is not adiagnosis for a disease process even though it isoften described in that manner. The term “FDB”should be used separately from the term “self-mutilation,” as this condition represents self-destruction of at least the surface of the skin. Bothconditions—FDB and self-mutilation—are oftenused as a diagnosis even though the true underly-ing cause and the diagnosis are obscure.
AGGRESSION
Aggression is often described in humans as a dis-turbed or psychological state, but in most animalsit tends to be species-specific behavior that con-fers survival benefits to the individual or socialgroup.[1] In a natural setting, the aggressive act isemployed to protect valuable resources or as aself-defensive measure. Aggression would be in-volved in maintaining the integrity of a socialgroup or acting to repel possible intruders.Aggression can be subdivided into a number ofcategories. According to Welle, in birds these cat-
egories would include fear aggression, possessiveaggression, protective aggression, and territorialaggression.[2] Others have different classifica-tion schemes. For example, fear, pain, and stress-induced aggression can be considered defensiveforms of aggression.[1] From the owner’s per-spective, the aggression takes the form of biting,as that is often why the owner brings the bird infor evaluation or considers sending the bird awayto a sanctuary.
Fear aggression can be defined as aggressionwith signs of fear associated with withdrawal orpassive and/or avoidance behaviors.[2] Theseare the birds that, when you approach, will bitewhen cornered and then retreat as quickly aspossible. Protective aggression is aggressionthat is directed toward another person who ap-proaches an object or, more commonly, a personthe aggressor wishes to control.[2] In the naturalsetting this would be the male amazon that de-fends its mate when another male approaches.This scenario is commonly expressed by a clientthat tells you that the bird “always bites my hus-band when he approaches me, Doc.” Protectiveaggression is consistently directed to a thirdparty in the presence of a certain individualwithout an actual threat.[2] Territorial aggres-sion is aggression that occurs in a defined arealike a cage. It often intensifies as the recipientapproaches and continues despite correction orattempt at interaction with the bird.[2] The mostcommon example is when the bird is asked tostep on the hand to get out of the cage, then thebird bites the hand—this is often described ascage aggression.
195
17Diagnostic Workup of
Suspected Behavioral Problems
Susan E. Orosz

DIAGNOSIS OF FDB AND AGGRESSION
The diagnostic workup of FDB and aggression re-quires a thorough history and clinical examina-tion. In human medicine, the history and physicalexamination represent approximately 80% of theinformation required to arrive at the diagnosis ofthe patient.[3]
The diagnostic tests ordered by the physicianare used to rule in or out the working diagnosis orthe differential diagnoses based on the findingsderived from the history and the physical exami-nation. In determining a behavioral diagnosis, thesame criteria are required—a thorough historyand examination. However, the examination is di-vided into two components, so it is more appro-priate to describe it as a clinical examination. Theclinical examination is divided into a physical ex-amination and a behavioral examination.
The History
History taking is described in another chapter(chapter 16) but is mentioned here because of itsimportance in determining a list of differential di-agnoses. The history must include a variety ofquestions not commonly asked in the traditionalveterinary format.[2] This requires more time anda different approach to history taking than isusual. As suggested, the large number of ques-tions that need to be addressed may require thatthe client(s) (and other people that play a role inthe life of the patient) fill out at least some of theinformation before the veterinary visit. It mayalso require that the behavioral consultant’s report(if available) be provided before the clinical ap-pointment. This is important so that the avian cli-nician can review the material and develop agame plan before the patient arrives.[4]
Rearing History—Possible Factor inDevelopment of FDB
In addition to the standard historical questions,the clinician—as well as the behavioral consult-ant—needs to ask the client questions to betterunderstand the rearing of the patient, as this canhave a major impact on the behaviors ob-served.[4] It has been suggested that some avianpatients with FDB may have a similar conditionto primates that exhibit self-injurious behavior(SIB). In a study on SIB in monkeys, mainte-nance of this behavior persisted or recurred de-
spite attempts to change the environment, sug-gesting that this behavior has value to the individ-ual.[5] Humans that self-wound expressed reliefpost-wounding of negative emotions that were ap-parently building up. This suggests that, in pri-mates, the act of SIB is followed by an immediatesense of relief, which may occur in companionbirds as well.[6, 7] Those studying primates withSIB noted that tension buildup was associatedwith an escalation of the animal’s heart rate. Thisoccurred within 30 seconds prior to the bitingepisode. It remained elevated during the bitingepisode and then would drop precipitously tobaseline between 30 and 60 seconds after theepisode was complete.[5] These data suggest thatthe self-injurious episode acts as a positive phys-iologic reinforcer of an abnormal behavior, mak-ing it difficult to quench or to provide relief withdrugs. If this same scenario exists for some birdsthat exhibit FDB, then this may explain the lackof responsiveness to medical management.
Companion birds, like primates, are also verysocial animals. The rearing of their young chicksappears to involve a large number of interactionsand learning experiences due to their altricial be-havior as well as their complex social behaviorsas psittacines. Bellanca and Crockett studied ab-normal behavior in pigtail macaques (Macacanemestrina), including SIB.[8] The amount oftime spent in single housing regardless of rearingmethods was linked to an increase in abnormalbehavior. They looked at differences in rearing asit pertained to degrees of abnormal behaviors andSIB, and response to enrichment as a treatment.Their data is of particular interest to avian practi-tioners as the method of raising monkeys in an in-fant laboratory has some similarities to currentavicultural practice of rearing chicks by hand,with only human interaction during the earlystages and some interaction with chicks of similarages. The monkeys were removed from theirmothers while under five days of age and rearedin single cages, although they could see and hearother infants.[8] At approximately 55 days of age,infants were allowed into a “playroom” with fourto six other infants of the same species and ap-proximate size and weight for 30-minute time pe-riods at least five days a week. Not until they wereweaned and at least six months of age were theygroup-housed. Anytime after that, they could beremoved to individual housing for projects.
196 Manual of Parrot Behavior

Results of the study showed that there were sta-tistically relevant increases in locomotor stereo-typies, self-abusive non-injurious behaviors thatincluded hair plucking, self-stimulation, andother abnormal behaviors in the nursery-rearedinfants as compared to mother-reared infants.[8]Non-injurious self-abuse is considered as “poten-tial SIB,” and if it continues, in many cases it doeslead to SIB. These behaviors appeared at non-specific times later in life after placement in soli-tary caging. This occurred despite the fact that themonkeys were caged in rooms with other mon-keys having visual, vocal, and auditory contact.For monkeys that had been mother-reared, thensingly caged, abnormal behaviors that developedwere much lower and were more effectivelytreated with enrichment, including social contactwith a same-species monkey. Those that had beennursery-reared were more likely to be unrespon-sive to enrichment or even socialization. Many ofthese primates that were not mother-reared ex-hibited self-abusive behaviors or other abnormalbehaviors that increased with time. In the obser-vations of non-human primate self-abusive andhair-plucking behaviors, most worsen with timeand many, depending on species, externalstresses, and individual temperament, becomeSIB conditions in some of the more common pri-mate species.[4]
What is different in the studies with primates,as compared with birds, is that no monkey wasisolated in a room without some sort of contact(visual, vocal, auditory) with other monkeys ofthe same species, as institutions are not allowed toplace a solitary animal in a room as primary hous-ing. Such isolation is contrary to the AnimalWelfare Act, Part D, Nonhuman Primates.[9]Many bird-owning households, on the other hand,have no other birds, let alone the same species, invisual, vocal, or auditory range of the newly pur-chased hand-raised bird. Often birds are weanedand placed into a household with no other birds.This would suggest that these situations may pre-dispose the single companion bird to abnormalbehaviors, as with primates.
It is important that the avian practitioner or thebehavioral consultant ask about this scenario onthe history form and take these concerns into ac-count when developing a working diagnosis ordetermining the response to treatment. This doesnot mean that the clinician or the behavioral con-
sultant should, when presented with a companionbird with FDB or any other behavioral problem,just throw up his or her hands and do nothing. Itshould instead provide one reason for the bird tohave a problem and/or not respond to treatment. Italso provides a backdrop to further understandthis problem and a way of looking at research toimprove a bird’s outcome when living with hu-mans. The primate data would also suggest thatbirds that were raised mostly by their parents andthen separated from them at weaning have a bet-ter chance of responding to treatment. Orange-winged Amazons (Amazona amazonica) that metthese criteria when singly housed and that devel-oped FDB improved with environmental enrich-ment.[10]
The amount of time birds spent foraging intheir natural environment may play an importantrole in understanding FDB, particularly withthose birds that are separated at weaning. InPuerto Rican Amazon Parrots (Amazona vittata),for example, four to six hours are spent each dayforaging for food and these birds often travel sev-eral miles between feeding sites.[11] In contrast,companion birds in our homes, like Orange-winged Amazon Parrots when provided a pelleteddiet, spend approximately 30–72 minutes eatingto meet their daily metabolic requirements with-out expending energy to travel to find food itemsor to manipulate them for consumption.[12]
These data suggest that companion bird speciesare highly motivated to forage for food over longperiods of time, just like their wild counter-parts,[10] and when foraging opportunities arenot present, then abnormal behaviors such asFDB may develop. Chickens when not providedwith foraging opportunities will also peck and dodamage to feathers as a displacement for the lackof foraging.[13, 14] Therefore, the time spent for-aging by the patient or interacting with toyswould be important to obtain during the history.
From a clinical point of view, the study by Drs.Meehan, Millam, and Mench suggests that certaintypes of enrichments have the potential forgreater use and hence success.[10] Orange-winged Amazon Parrots that were parent-raiseduntil weaning developed FDB when placed intoindividual cages at weaning.[10] These parrotswere divided into two groups after approximately11 months post-weaning when the FDB began.One group received foraging enrichments while
17 / Diagnostic Workup of Suspected Behavioral Problems 197

others were provided with physical enrichments.Foraging enrichments required that the parrotschew through barriers, manipulate objectsthrough holes, and sort through inedible materialand/or open containers to obtain food items whilethe normal ration was available. Physical enrich-ments included alternative perching sites, climb-ing or swinging opportunities, or providing move-able objects that could be manipulated with thefeet or beak.
The study showed that the parrots used theirenrichments within three days of introduction butthat the use of the physical enrichments declinedover time, while the use of the foraging ones re-mained stable.[10] Additionally, refeathering oc-curred within two weeks of introducing the itemsinto the cages. Six of eight parrots demonstratedan improved feather score, which suggests thatenrichment allows for foraging and reduces FDBbut does not eliminate it. However, this study in-dicates that these types of foraging activitiesshould be included as part of the clinical treat-ment regimen for FDB. This study also demon-strated that parrots in the enriched conditionworked to access supplemental food items an av-erage of 19–26% of their active time, even thoughtheir normal ration was available as a pellet. Thiswould support the findings in chickens that in abarren environment and/or one without foragingopportunities, foraging attempts may be redi-rected toward their plumage as with FDB.[13–15]The quality and the availability of the foragingenrichments in particular contribute to the effec-tiveness of the response to the FDB.
Foraging may be important in the neural devel-opment of parrots, as in primates. A strong rela-tionship was also found between the lack ofbehavioral opportunity for foraging and the devel-opment of stereotypic behaviors in parrots. Therewas evidence that there was an underlying neuralcompromise with stereotypies.[16] This suggeststhat neural dysfunction may result from a barrenenvironment or the lack of enrichments that theparrots would be engaged in long term. It may alsounderlie abnormal, repetitive behaviors.[10]
The hypothesis would be that FDB can repre-sent the avian counterpart to self-abusive non-injurious behavior and/or SIB in primates. If thiswere true, then feather damage could act as a cop-ing strategy and should be physiologically rein-forcing as with the change in heart rate (HR) in
primates. No information is currently available todetermine if there are HR changes in birds imme-diately prior to the feather-damaging event, butthis needs to be investigated. The hypothesiswould also include the effects of isolation andhand-rearing as an underlying factor in the devel-opment of the abnormal grooming behavior.[4]
This hypothesis suggests that new and impor-tant information be gathered during the historyabout hand-raising and weaning. This informationis important for understanding the possible role ofhand-raising practices and isolation of birds atweaning in the development of FDB. A form thattakes these factors into account is provided inTable 17-1 as a guide in designing your historyform. It is important that the clinical examinationincludes both a physical examination and a be-havioral evaluation.[2]
Clinical Examination
There are two important aspects to the physical ex-amination. The first is to document if there arephysical changes that can be observed by the avianclinician. This would be most obvious with FDB.The second is to determine the extent of thechanges observed. Documentation of these physi-cal exam findings helps determine if there are any infectious or metabolic changes that produce a secondary behavioral condition. A feather-scoring system has been developed and presented inthe literature for FBD and can be adapted for clini-cal use (see Table 17-2). It would be best to stan-dardize a feather scoring system so that all profes-sionals can then bring their data together to look atvarious parameters to further understand this prob-lem and enhance treatment success.
From a clinical perspective, it is important todetermine if there is damage to the feathers andwhere and how much damage has occurred. Theremay be situations where there is no visible dam-age to the feathers but the client reports that thebird is constantly chewing its feathers. Ownersmay be reporting normal behaviors that arethought to be abnormal. This most frequentlyhappens when owners don’t know how much timeis spent in daily grooming. The other situationmay occur when birds attempt to groom, but withlittle experience or learning from non-existentparents, they do not groom properly and spendmuch time doing a poor job. Owners may reportfeather loss when there is none. This most fre-
198 Manual of Parrot Behavior

199
Table 17-1 Feather damaging/self-mutilation behavior form
Date of Clinical Examination_________________Owner’s name_____________________________Patient’s name_______________ Species________________________ Age_____ Sex_____
Owner’s observations as told to clinician
Historical questions(Owner’s report) Yes No Unknown
Hen-reared
Hand-reared
Reared with siblings
Reared with similar aged but different species
Reared with non-similar aged birds
At what age was the patient taken from the nest? Description:
Age when singly housed Description:
Age when placed as a single bird Description:
Describe social contact during hand rearing Description:
Describe weaning process and procedures Description:
Locomotor behaviors noted: Yes No Unknown
Pacing back and forth on perch
Somersaulting in the cage
Bobbing head and body up and down
Head flipping back and forth
Running toes through beak
Stretching
Flapping of wings (i.e., flying in place)
Flaring the crest when stresses
Fluffing the feathers over the beak
Self-stimulation behaviors noted: Yes No Unknown
Head rubbing over uropygial gland and/or back
Overgrooming
Plucking over distal back and/or medial thighs
Cuddling foot or extremity
Vocalizations/screaming
Tail and/or vent flaring
Other:
(continued)

200
Table 17-1 Feather damaging/self-mutilation behavior form (continued)
Clinical examination (observations in clinic)
Feather-damaging behaviors Location: Side preference:
Chewing feather shafts
Chewing barbs
Chewing remiges; primaries, secondaries
Chewing rectrices
Plucking and removing coverts
Plucking and removing down
Plucking and removing remiges
Plucking and removing rectrices
Self-mutilation and where
Behavioral assessment (Record number of times for a 15-minute period)
Observer’s name:
Date: Yes Times/15 min Not observed
Aggression
Fear
Pacing back and forth on perch
Somersaulting in the cage
Bobbing head and body up and down
Head flipping back and forth
Socomotor behaviors noted: Yes No Unknown
Pacing back and forth on perch
Somersaulting in the cage
Bobbing head and body up and down
Head flipping back and forth
Running toes through beak
Stretching
Flapping of wings (i.e., flying in place)
Flaring the crest when stresses
Fluffing the feathers over the beak
Self-stimulation behaviors noted: Yes No Unknown
Head rubbing over uropygial gland and/or back
Overgrooming
Plucking over distal back and /or medial thighs
Cuddling foot or extremity
Vocalizations/screaming
Tail and/or vent flaring
Other:

quently happens when the owner sees the aptery-lae spaces between feather tracts or pterylae whenthe bird is wet.[17]
As indicated on the form provided, it is impor-tant to determine more precisely the lesions as totheir location, if there is a sidedness aspect to thepicking, what part of the feather is being dam-aged, if the entire feather is being plucked out,and what type of feathers are being damaged. Forexample, the bird may only chew at the barbs ofcoverts on the ventral surface of the pectoral mus-cles. A bird with this condition would have a 1.75score for the chest/flank back and legs portion ofthe scoring system but on the physical examina-tion sheet the location and the extent of the lesion
should be indicated. With all feather conditions,drawings and/or photos help document the lesionsand response to therapy and help with one’s mem-ory, particularly as most of these problems arelong-term situations.
The physical examination is often described asconsisting of two parts—observations followedby the actual hands-on palpation and auscultation.It is best to perform the observation and the be-havioral examination at the front end of the visitafter history taking. As indicated, often these ab-normal behaviors are not observed in the officebecause the birds will not do them in a situationwhere they are not completely comfortable.Home visits help remove that variable but often
17 / Diagnostic Workup of Suspected Behavioral Problems 201
Table 17-2 Feather-scoring system (from C.L. Meehan, J.R. Millam, and J.A. Mench.2003. Appl Anim Behav Sci 80:71–85)
Score Description
(a) Scoring system used for chest/flank, back, and legs0 All or most feathers removed, down removed and skin exposed, evidence of skin or tissue
injury0.25 All or most feathers removed, down removed and skin exposed, no evidence of skin or
tissue injury0.5 All or most feathers removed, some down removed, patches of skin exposed0.75 All or most feathers removed, down exposed and intact or feathers removed from more
than half of the area, some down removed, patches of skin exposed1.0 Feathers removed from less than half of the area, some down removed and skin exposed1.25 Feathers removed from more than half of the area, down exposed and intact1.5 Feathers removed from less than half of the area, down exposed and intact1.75 Feathers intact with fraying or breakage2.0 Feathers intact with little or no fraying or breakage
(b) Scoring system used for wings0 All or most primaries, secondaries, and coverts removed, down removed, skin exposed,
evidence of skin or tissue injury0.5 All or most primaries, secondaries, and coverts removed, down removed, skin exposed, no
evidence of injury1.0 More than half of coverts removed, down exposed and intact or more than half of
primaries and secondaries removed, down exposed and intact1.5 Fewer than half of coverts removed, down exposed and intact or fewer than half of
primaries and secondaries removed, down exposed and intact or primaries andsecondaries intact with significant breakage and fraying
2.0 Feathers intact with little or no fraying or breakage
(c) Scoring system used for tail0 All or most tail feathers removed or broken1 Some tail feathers removed or broken or significant fraying of tail feathers2 Feathers intact with little or no fraying or breakage

just the presence of a “visitor” extinguishes theabnormal behavior. As suggested, proper video-taping helps skirt this issue. However, there areother times when the stress of this “new” environ-ment brings out certain behaviors that the clini-cian can observe.
All abnormal behaviors of non-human primatesare also evaluated in a systematic way, to classifythem in categories such as affiliative, fearful, oragonistic.[4] The classification scheme allows forsystematic and regular objective evaluation for ef-fectiveness of treatment strategies. It also lends it-self to accurate documentation of the behaviorsand longitudinal assessments, unlike most evalua-tions of psittacine abnormal behaviors reported orobserved by practitioners. Analysis also allows forspecies-specific normal behaviors, such as regulargrooming or lip smacking in primates, to berecorded as percentage of time of the day, whichmay point to other learned behaviors (i.e., the ani-mal never learned proper behaviors from older in-dividuals). By classifying these behaviors, the vet-erinarian or behaviorist can work on training tomanage specific problems, to overcome fears,train out some aggressive tendencies, or encour-age affiliate behaviors.[4]
As indicated on the form provided in Table 17-1, locomotion-based behaviors are distinguishedby self-stimulating behaviors under FDB butshould be addressed for all abnormal behaviors aswell. The behavioral assessment should recordthose behaviors within a defined period of time(defined here, e.g., as 15-minute blocks). This isimportant so that true changes can be noted overtime and with treatments. We tend not to ap-proach behavioral examinations from this per-spective, but it is required when documenting ab-normal behaviors in primates. Without this rigor,it is very difficult to assess changes scientifi-cally.[4] However, primates are often videotapedin their normal caged environment and the behav-iors counted. For the avian patient, videotapingthem at home and having the professional reviewthe tape is best.
From the perspective of aggression, it may bevery difficult to assess territorial aggression in theexam room, as usually the companion bird is ag-gressive with its home cage. However, the tail fan-ning and flashing eyes of male amazons often in-dicate aggression. But often those signals aremissed by the client or others. If these are not un-
derstood, when the client asks the bird to step up,he or she is asking to get bitten.[18] This would bea normal behavior that is misinterpreted as abnor-mal, but the understanding of why the owner getsbitten needs to be addressed. A better understand-ing of the normal reproductive physiology wouldhelp the owner understand the common mistakesor miscues that are made in the home.[18]
Observations concerning physical changesshould be noted on the examination form. Thebird’s mentation, along with posture and stance,are important indicators of behavioral problemsor disease. For example, one amazon patient wasobserved in my practice that bore its weight onmostly the left side of the body while the rightside of the bird was mostly plucked over the back,hip, chest, and medial and lateral thigh. This pa-tient had severe arthritis of the right hip. Othersymptoms of disease need to be noted during theobservation phase. Pectoral muscle mass, featherquality, and how the contour feathers lay are im-portant to notice. They may indicate a tumor orfat deposition suggesting a lipoma. The numberand quality of the droppings need to be inspectedbecause increased urates suggest problems withthe kidneys, for example. The nares, beak, andeyes should be given scrutiny to look for abnor-malities. Although the form provided (Table 17-2)concentrates on the feathers and their quality, thecoloration, iridescence, and the lay of the feathersagainst the body wall are also important and needto be noted. It is important to perform both por-tions of the examination in a step-wise fashionand in the same order for each patient.
The second part of the physical examination isthe hands-on palpation and auscultation of the pa-tient. As suggested in other chapters, it is impor-tant to make the toweling and physical part asstress free as possible. If often helps to tell thebird what you are doing prior to the event and touse a slow and methodical wrapping of the patientas described previously (E. Wilson, personalcommunication, August 1999).[19] The handlingportion should be done in the same manner witheach patient so that no part is forgotten. However,the problem(s) of the patient dictates where theclinician will concentrate his or her physical in-vestigation. With skin and/or feather problems, ahead loop helps to magnify and examine the com-ponents of the integumentary system. Severalblood feathers in the affected area can be plucked
202 Manual of Parrot Behavior

and the pulp of the shaft used for a culture and/ora cytology. This should be done at the end of thehandling phase.
In terms of diagnostics, it is best to let the avianpatient calm down after its travel to the veterinaryhospital, then perform the history, the observationphase of the physical examination, and the behav-ioral phase if there will be limited handling. If aCBC or plasma for a biochemical analysis needsto be part of the workup for the particular patient,it is best to obtain the blood sample prior to a sig-nificant amount of time spent handling and timespent in the towel. This is important when inter-preting laboratory results as the stress of handlingand traveling can alter the CBC results, with thelymphocycte count tending to decrease and theheterophil count and the WBC increasing overnormal blood counts.[20] Elevation of several en-zymes including AST and LDH can occur withtoweling.[21] After the blood has been collected,the remainder of the physical examination can becompleted. It is often best to start at the head, ex-amine the oropharynx and take a choanal swab fora gram stain if one is performing a general healthcheck, and proceed caudally down the remainderof the body with special emphasis at examiningthe structures of the integumentary system. Theremay be a reason to perform a skin scraping or takefeathers for examination of parasites as well. Theexamination can then be completed with the re-mainder of the behavioral portion, if necessary. Itis always best to make observations relating to be-havior prior to toweling. However, the clinicianshould observe the patient immediately after han-dling. It is important to check the respiratory re-covery time and watch for any abnormal behav-iors, as stress may induce them.
DIFFERENTIAL DIAGNOSES FOR FDB
After the physical examination, one can narrowthe differential list into one of two possible butunfortunately overlapping general categories fordiseases that affect the integumentary system.[22,23] These would be inflammatory skin andfeather diseases and non-inflammatory skin andfeather diseases that are often assumed to be non-infectious. Feather-damaging behavioral condi-tions would be a subset of the latter category.Inflammatory skin diseases include viral, fungal,bacterial, and parasitic infections that may causedisease symptoms.[22] Trauma results in other in-
flammatory disease conditions. Common viraldiseases that affect the integumentary system ofpsittacine birds include papillomavirus, poly-omavirus, avipoxvirus, and psittacine beak andfeather disease (PBFD) virus or avian circovirus.Trauma includes several types of conditions in-cluding tail trauma, split sternum (often inAfrican Grey Parrots), other traumatic conditionsto the skin, and damage to blood feathers.[22, 24]
Papillomavirus is thought to result in wart-likedermal papillomas on unfeathered areas of the in-tegument. These lesions have been observed asmultiple cutaneous papillomas of the face on aTimneh African Grey Parrot, the legs of EuropeanChaffinches and Braming’s Finches, and the com-missures of the bills of Canaries.[22, 24, 25]Cutaneous herpesvirus lesions can be confusedwith dermal papillomas. Herpes lesions presentas wart-like lesions or thickenings of the skin andare often found on the feet or legs, particularly ofcockatoos and macaws. In these types of birdsthey may show up as a flat, raised plaque or aroughening of the skin with depigmentation. Thegross appearance is characteristic and diagnos-tic.[22, 24, 25] Biopsy with histopathology andsurgical incision can be performed but is not rec-ommended normally.
Polyomavirus often results in abnormal feath-ering of Budgerigars, particularly in chicks.Chicks affected between 7 and 15 days of ageoften have ascites and have changes of the pri-mary and secondary flight and tail feathers orthese feathers may be absent. Feathers demon-strate thickened shafts with hemorrhage. Featherabnormalities are uncommon in larger psittacinesbut may be found in those recovering from infec-tion. These larger species often present with sub-cutaneous hemorrhages, hemorrhages in thefeather shaft, and reduction in the numbers andchanges in the morphology of down and contourfeathers. Diagnosis is by biopsy with histopatho-logical examination for characteristic lesions. APCR probe is also available for diagnosis.[22, 24]
Avipoxvirus affects most of the families orgroups of birds with some cross-infection de-pending on the group.[25] Infections from avi-poxvirus produce symptoms that typically presentas either a wet or dry form. Canaries and other re-lated species can also present with a septicemicform. The dry or cutaneous form demonstratesraised papules, pustules, and/or nodules that can
17 / Diagnostic Workup of Suspected Behavioral Problems 203

rupture to form crusts or scabs on non-featheredareas. Wild-caught Blue-fronted Amazons andyoung Pionus parrots are considered the mostsusceptible for this disease. Diagnosis is based onthe gross appearance of the lesions and the pres-ence of large Bollinger bodies, which areeosinophilic cytoplasmic inclusions observedhistopathologically.[22, 24, 25]
PBFD affects a large number of species; how-ever, those most commonly affected include birdsfrom Africa, Asia, and Australia. The chronic formof the disease is associated with abnormal feather-ing, including dystrophic feathering with retainedfeather sheaths, blood in the umbilicus of thefeather shaft, clubbing of the feathers and otherabnormal feather shapes, multiple stress bars onthe feather vane, and annular constrictions of thebase of the feathers. Some birds may only exhibitfeather changes while others show only beak ab-normalities. Lesions of the beak include ulcerationand an unkempt appearance with elongation,beaks that are weak and fracture easily, andpalatine necrosis.[22, 24, 25] Beak lesions are ob-served more commonly in Sulfur-crested Cock-atoos (Cacatua sp.), Umbrella Cockatoos (Caca-tua alba), and Moluccan Cockatoos (Cacatuamoluccensis).[22] Lories and lorikeets appear tobe infected with a variant of PBFD. These birdsexhibit less obvious feather lesions and theirfeathers may only appear dull. Lovebirds mayalso present with dull feathering with brokenfeathers.[24] Diagnosis is made through histo-pathology of an affected feather in combinationwith a PCR probe to help confirm the disease.Blood can also be drawn to determine if there arecirculating viral particles that can be detectedwith the PCR probe. The PCR technology differsbetween lories and non-lories as the test has to bealtered to include these species. In some of thespecies not confirmed using PCR technology, di-agnosis is made by examination with the electronmicroscope of the biopsy sample.[26]
Fungal diseases affecting the integumentarysystem are not common in companion birds. It isdifficult to distinguish fungal isolates from theskin causing disease from those that are part ofthe normal flora. Fungal organisms associatedwith integumentary disease include the commondermatophytes Trichophyton sp. and Microspo-rum gypseum, along with other mycotic organ-isms Malassezia, Penicillium sp, Cryptococcus
neoformans, Aspergillus sp, Candida sp., Mucorsp., and Rhizopus sp. Some of these organismsproduce scaly skin encrustations, possible hyper-keratosis, and patchy feather loss often affectingthe head, neck, and breast. Cryptococcosis pres-ents more commonly with respiratory and/or neu-rological signs. When it affects the integumentarysystem it is associated with granulomatous le-sions of the face or beak. Candidal lesions ofteninvolve the rictus or commissures of the bill, theepidermal area around the nares, and the featherfollicles on various locations around the body.Penicillium and Aspergillus sp. are often isolatedfrom non-symmetrical erosive lesions of thebeak. These lesions may also be associated withneoplasia such as squamous cell carcinoma, nutri-tional deficiencies, or poor husbandry practices.
Diagnosis of fungal organisms on the surfaceof the skin may be through cytology, histopathol-ogy, and/or culture. Histologic examination fordermatophytes is important, as a positive cultureresult may be from a contaminant. A diagnosis ofMalassezia may be based on exfoliative cytologyof the characteristic footprint-shaped yeast cellswith confirmation by histopathology of the skinand/or feathers. Cryptococcosis can be confirmedby finding the characteristic yeast with its thick-ened capsule on histopathological or cytologicalexamination. Candida histologically presents as abasophilic yeast with a thin capsule on gramstains while Aspergillus and Penicillium sp. havebasophilic hyphae, which are much larger thangram-positive rods, with conidia.[22]
Bacterial infections may be primary or second-ary and may be focal or generalized. The mostcommon isolate from folliculitis is Staphylococ-cus sp., and it is associated with swelling and red-dening of the area.[22] Generalized bacterial der-matitis is associated with soft tissue swelling andwith flaking, reddening, or other discolorationsand crusting of the skin. These types of lesionsare often pruritic, resulting in self-trauma. Diag-nosis is based on gross appearance, histopath-ology, and/or culture results. Secondary bacterialinfections can often result post-trauma, includingFDBs that are psychologic in nature. Grosslythere may be swelling, necrosis, and/or cellulitis.It is more difficult with these secondary infec-tions to identify the causative organisms as resultsare based on histopathology and findings fromcultures.[22,24]
204 Manual of Parrot Behavior

Mycobacterial infections can appear as wart-like growths, flaking swellings, or granulomasoften on the head or in the oropharynx. Thesetend to not be pruritic and require biopsy withhistopathological observations using acid faststaining for diagnosis.[22]
There are a number of parasitic diseases thataffect the integumentary system in birds. They areoften species or at least genus dependent. Mitesare a common problem in chickens, with themharboring Knemidokoptes mutans, while Budgeri-gars and Canaries are infected with Knemido-koptes pilae. Other species affected with Kne-midokoptes have included Gouldian Finches, Cockatiels, amazon parrots, Ring-necked Para-keets, Scarlet-chested Parrots, Princess Parrots, andYellow-fronted Kakariki.[22] Mites are transmittedfrom bird to bird or by transmission in the nest tounfeathered chicks. Mites can often be diagnosedwith a skin scraping. Knemidokoptes mites areoften numerous and more easily observed com-pared with eternal mites that may not remain on thepatient long enough to find them.[22]
The red mites of various species of birds (Der-manyssus and Macronyssus sp.) can be observedon the external surface of the feathers of fowl andwild birds but may not cause feather damage.They have been reported in Canaries and occa-sionally in Budgies. These mites are noted as noc-turnal feeders and are not observed on the birdduring the day. They can be found by placing awhite cloth on the bottom of the cage for subse-quent examination. They take a blood meal atnight, so the patient may show only signs ofanemia without the presence of the mites on thefeathers.[22]
Myialges or the mite Metamichrolichtus nudusfeeds on the skin, burrowing into its surface toform pits. Birds with these mites are pruritic, withfeather loss around the head. They are often hy-perkeratotic, and erythematous as well. Grey-cheeked Parakeets (Brotogeris pyrrhopterus) aremore commonly affected. A louse or hippoboscidfly is required as a transport host to other birds forinfection to be spread. Quill mites are more com-monly associated with passerines. The Metami-chrolichtus mites may be observed on a skinscrape while the quill mites may be observed inthe powdery quill material or the broken featherswhen examined microscopically.[22]
Biting lice often produce pruritis, mild to mod-
erate feather damage, and hyperkeratosis depend-ing on the degree of infestation. They are morecommonly associated with gallinaceous birds,pigeons, and wild birds but have been reportedoccasionally in psittacine birds. Lice are hostspecific and do not survive long in the environ-ment. Lice eggs can be found on the primary andsecondary flight feathers or on feathers aroundthe vent.[22]
Non-infectious causes are often considered non-inflammatory but this may not be the case. Peri-vascular dermatitis is considered a non-infectious disease process that is diagnosed bybiopsy of skin for an infected area and a non-affected area. It is similar to lesions observed withhypersensitivity dermatitis in mammals withperivascular infiltrates of lymphocytes, plasmacells, eosinophils, and other inflammatorycells.[23] Histopathologic findings include thepresence of edema, vascular hypertrophy, hyper-plasia of the epidermis, and hyperkeratosis, de-pending on the length and severity of the lesions.These findings are consistent with other conditionsassociated with pruritis, suggesting that some birdswith FDB are pruritic and may express some typeof hypersensitivity reaction. Preliminary data sug-gest that there is a greater incidence in Blue-and-Gold Macaws with a lesser predisposition in ama-zon parrots and Eclectus Parrots.[23] It was foundto be uncommon in African Grey Parrots.
Another non-infectious cause of FDB is fromaberrant behaviors developed as a consequence ofhand-rearing. There may be other causes as wellthat result in the symptom of feather damage andthis condition has often been described as psy-chogenic feather picking.[23] Affected birds ap-pear clinically with symptoms similar to thosethat present with perivascular dermatitis.[23] It isparticularly important that biopsies are takenfrom both affected skin and unaffected sites.Inflammation can be observed histologically fromthe affected sites because of the trauma involvedin this psychogenic form. But in these birds, in-flammatory indicators are not observed in the“non-affected” biopsy samples, unlike those birdsthat exhibit perivascular dermatitis. Preliminarydata suggests that African Grey Parrots, cocka-toos, and Cockatiels are more likely to presentwith the psychogenic form of FDB in review ofbiopsy samples submitted.[23]
Feather dysplasia is another form of FDB and
17 / Diagnostic Workup of Suspected Behavioral Problems 205

can result from a variety of etiologies, some ofwhich are undetermined. This condition describesabnormal growth patterns of the developingfeathers and has been likened to the folliculardysplasia of the hair follicles of dogs.[23, 24]However, abnormal growth can result from a va-riety of diseases and may be of an infectious aswell as a non-infectious one. Birds may developfeather dysplasia as a consequence of a congeni-tal condition, an inherited problem, hormonal ormetabolic conditions, nutritional problems, spon-taneous situations, infectious agents such asPBFD virus, and other suspected but not con-firmed viral etiologies. For example, polyfolli-culitis syndrome is more commonly associatedwith lovebirds, Budgerigars, parrotlets, andCockatiels. It appears as multiple quills project-ing from a single feather follicle.[22] Its etiologyhas not been determined, but it is suspected to beviral in lovebirds.[22, 24]
DIFFERENTIAL DIAGNOSIS FORAGGRESSION
Aggression is often diagnosed through the clini-cal examination and the history provided by theowner. Commonly owners report that the bird isbiting someone in the family, often the spouse.Most of the time the bird has “chosen” the otherspouse as its mate and is just defending their rela-tionship. It is important to understand the cuesthat the human(s) and bird are providing eachother in this scenario to understand if the aggres-sion is from a bonding/mate problem.[18] Thereason for the aggression may be purely behav-ioral with no underlying physical problem, or itmay have a physical component. Often male ama-zons become aggressive around the time of thebreeding season, which would be normal behav-iorally but very difficult for a pet-oriented ownerto handle. Even though one might be able to de-termine the aggression is territorial or protectiveand might be from breeding activity, it would bebest to perform some diagnostics to confirm orrefute the working diagnosis. In this scenario, itwould be important to establish the sex of the birdand to run a hormone panel to determine if thebird is in the culmination phase of the breedingcycle. The hormone panel most commonly usedin birds is the ferret panel at the University ofTennessee, as it provides the male and female sexhormones along with some of the metabo-
lites.[27] The sex steroid hormones are notspecies dependent and can be measured in a vari-ety of species using the same type of testing pro-tocol, while the protein hormones cannot.
Another way to determine if the bird has in-creased sex hormone levels is to observe the sizeand quality of the gonads by endoscopy, inferringthat the hormone levels are elevated. Trainingowners to understand when birds are aggressive isthe first step in avoiding the problem of biting.The next step is to teach the bird and the ownerbasic commands and ways to discipline thebird.[28] Positive reinforcement and rewards forgood behavior help shape a positive attitude forthe companion bird with its owner. These may betaught by the clinician or the behavioral consult-ant with recheck appointments tailored for the in-dividual situation.
Other diagnostic tests for birds exhibiting ag-gression could be those that check the function ofthe liver and brain. Plasma biochemical analysesshould include an AST, CPK, albumin, total pro-tein, glucose, cholesterol, and bile acid levels tohelp determine normal liver function. Most com-monly, aggression from altered brain functionwould require a neurologic examination and otherdiagnostic tests such as a CT scan and/or electro-diagnostics. Pain may also make birds appear ag-gressive to the less knowledgeable, as they oftenbite when in pain. Therefore, the astute clinicianneeds to observe the bird carefully and monitorthe bird’s behavior in the exam room to determinethe diagnostic tests that need to be run.
DIAGNOSTIC TESTS FOR FEATHER-DAMAGING BEHAVIOR
Diagnostic tests should be tailored to the individ-ual bird and the findings from the history andclinical examination. They should be based on theworking diagnosis or the list of differential diag-noses. In the case of FDB, the diagnostic workupplan should center on diagnostics involving theintegumentary system. Although the inflamma-tory diseases of the integument overlap with thosein the non-inflammatory category, that does notmean that the shotgun approach of diagnostictesting is indicated. Most clients are exasperatedthat they spent a large amount of money with noresults. It is important to tailor the tests to the in-dividual bird and to explain to the owner what thetests may or may not show in helping to arrive at
206 Manual of Parrot Behavior

a diagnosis. Education is extremely important forthese owners. As clinicians, one should stress tothe owner that they should not expect the bird willgo home after one visit and grow back its feathersand look perfectly normal. Owners need to under-stand that many birds will stay the same, whileothers will improve but not return to “normal” de-pending on the diagnosis and the degree oftrauma to the skin and feather follicles. This isparticularly true at this point with our understand-ing of FDBs. Psychological FDB birds may im-prove for a while and then relapse. With attentionfrom clinicians and behavioral consultants, theymay improve again. It is important to look at thetriggers for FDB to find the underlying cause(s)for possible resolution. The bottom line is thatonce the companion bird starts damaging itsfeathers, it is not going to be a “quick fix.” It willrequire diligence on the part of the owner and willbe costly. It is important that the owner under-stands that on the front end.
Clinically speaking, a minimum database isoften referred to when describing a workup for aparticular disease condition. However, the mini-mum database for birds exhibiting FDB or otherintegumentary problems is not set in stone. Withmost disease processes, the minimum database isa CBC and plasma biochemical analyses with thepossible inclusion of a plasma electrophoresis(EPH).[29] The CBC is a general indicator ofoverall health and will not provide any cluesabout a specific disease. Most birds with FDBwill not have any changes in the components oftheir CBC. However, those that mutilate or causedamage to their skin may have an increased whiteblood cell count (WBC) with a heterophilia andthose that have lost a significant amount of bloodmay demonstrate anemia. In chronically ill birds,there may an increased WBC with a monocytosis.The CBC can also be used to monitor the re-sponse to treatment, particularly for those birdswith an infectious dermatitis.[29]
The plasma biochemical analysis is an indica-tor of general organ function. Those birds thathave been picking will often have an increased as-partate aminotransferase (AST) level because thisenzyme is released with liver damage and that ofmuscle cells that can occur with picking. Addi-tionally, picking birds can have an elevated crea-tinine phosphokinase (CPK), an enzyme that ele-vates only with muscle damage. Most commonly
in birds with FDB, both enzymes will be elevated.If only the AST level is elevated, then there isconcern that there may be liver damage. Liverdamage, in mammals, is known to result in pruri-tis and may be involved in the underlying cause ofthe problem.[29] Elevation of the AST value isalso observed in birds that have increased gonadalhormones. Uric acid levels are associated withrenal disease and may help explain some birdsthat pick only over their synsacrum. Other birdsthat are in the culmination phase of the breedingcycle often pick in this area, along with the lateralthighs and flanks. Lower than normal levels ofplasma proteins may be associated with liver dis-ease (where some are manufactured), problemswith filtration in the kidneys, and problems withingestion from the GI tract (observed in proven-tricular dilatation disease, PDD).[29]
Plasma electrophoresis provides general infor-mation about the health of the avian patient butdoes not provide a specific diagnosis. The EPHshould be used in conjunction with the CBC andplasma biochemical analysis. Many birds withFDB will have normal values, suggesting thatoften there is no specific infectious disease entitythat causes the underlying problem. However,those patients with an increased beta fraction areassociated with an acute infectious disease whilean increased gamma fraction is associated with achronic infection. This gamma fraction may oftenbe elevated with chronic pickers, especially thosewith damage to the skin.[29]
Sex Determination
It is important to know the sex of the bird as aguide to understanding the relationship to the dis-ease process. Sex determination can be per-formed by rigid endoscopy or by PCR or other re-cent technologies using the feather or blood.
Fecal Examination
Fecal examination can provide information aboutthe general health of the bird but does not indicatea diagnosis for most feather pickers except forthose with Giardia. Trophozoites can be observedon a slide of fresh droppings at 20–40X while thecysts are best seen with an iodine stain. GiardiaELISA tests designed for dogs may not be appro-priate for birds as the testing has not been con-firmed to cross-react between birds and dogs.Nematodes have been associated with “poor
17 / Diagnostic Workup of Suspected Behavioral Problems 207

doing” and pruritis, but most companion birds arenot exposed to the conditions for them to be aproblem.[24, 29]
Skin Scraping for Parasites
Skin scrapings for cytological examination forparasites is a common procedure in mammals thatare pruritic but uncommon in birds. Knemido-koptes sp. mites and Metamichrolichus nudus aretwo of the most common types of mites in avianspecies. The former species of mites do not typi-cally result in pruritis, while the second type,most common in Grey-cheeked Parakeets, ishighly pruritic.[22, 29] The same technique canbe used as in mammals to obtain the sample.However, care must be taken to not tear the skinas it is thinner than that of mammals.[17]
Heavy Metal Assays
There is anecdotal information that suggests thatheavy metal toxicosis, namely lead and/or zinc,may cause FDB. However, data on zinc toxicosissuggest that birds develop pancreatitis and do notbecome pruritic. Pruritis and feather damage donot seem to be a manifestation of lead toxico-sis.[30, 31]
BACTERIAL AND FUNGAL CULTURES
Bacterial and fungal cultures should be reservedto areas of the integument that are abnormal.Choanal and fecal cultures only reflect the mi-croorganisms from these regions and would implyoverall health but not specific problems that causethe bird to have FDB.[29]
In abnormal areas, samples for cultures can bepaired with Gram stains to make a diagnosis. Adeep culture is recommended over a superficialone. Often therapy can be initiated based on theGram stain while waiting for culture results, par-ticularly with fungal cultures. Histopathology canalso help in arriving at a diagnosis, and biopsysamples are recommended when a culture istaken.[22, 24]
Feather pulp cultures and Gram stains may beuseful in some patients with FDB, as the pickingmay cause damage that produces a secondary in-fection. The history and clinical examination mayhelp determine if the infection is secondary orprimary.[22] Often Gram stains of the pulp cavitymay be misdiagnosed as a fungal infection, as thenucleus of ruptured red blood cells can appear as
yeast-like organisms (S.E. Orosz, personal obser-vation, 2003).
Histopathology of the Skin or Feathers
A skin biopsy including a feather follicle, partic-ularly an immature one, is important for diagnos-ing an infectious or inflammatory condition.[22,23] It is important to take a biopsy of the skinfrom an affected area and one from a non-infected one. Samples should also be clearlymarked in order for the pathologist to make anappropriate interpretation. Results from thehistopathological examination can help rule in aswell as rule out a number of infectious causes ofFDB such as viral, bacterial, parasitic, and fungaldiseases. It is also important for determining ifthere is evidence of perivascular dermatitis.When there is evidence of disease while the cul-ture results are negative there can be a number ofexplanations for the cause. Negative aerobic cul-tures occur when an anaerobic organism is pres-ent, when the patient is on antibiotics, and whenthe cultures are not handled properly. When thehistopathologic results suggest the skin is nor-mal, then the diagnosis of FDB is more likely tobe psychological in origin.[22]
Intradermal Skin Testing
Early results developing the use of intradermalskin testing for birds have not been promising. Itis assumed that if these tests were developed tothe level as for mammals, then desensitizationmight also reduce the prevalence of FDB. Unfor-tunately, these tests have not indicated that aller-gens play a major role in FDB.[29]
Testing for Aspergillosis
Aspergillosis is not a common diagnosis for FDBand the Aspergillus organisms may affect thebeak more often than the feathers. Measuring thetiters or antibodies of Aspergillus sp. from bloodsamples does not help determine if this may bethe cause for integumentary problems. Fungalcultures, exfoliative cytology, and histopathologyof biopsy samples are more appropriate to make adiagnosis.[22, 24]
Viral PCR and Other Test Procedures
Psittacine birds with FDB are often tested usingviral PCR probes for avian polyomavirus andPBFD, as these two viruses can affect the feather-
208 Manual of Parrot Behavior

ing of psittacine birds.[22, 24, 25, 29] Polyoma-virus may cause feathering abnormalities of thesmaller species including Budgerigars andfinches, but the test performs poorly with thesesmaller species. The PCR test for PBFD may beconsidered in the old world species as these birdsare affected more commonly compared with thenew world species. The virus in the chronic formof the disease can produce feather and/or beakdystrophy, which may be mistaken for mutilationof the feathers. In smaller species and less com-mon ones, histopathology of a feather biopsy withits surrounding skin may be required, along withelectron microscopy of the sample.[26]
Whole Body Radiology and UltrasoundEvaluation
Whole body radiographs may be useful when apatient picks or mutilates over a particular area orto diagnose those with PDD. Some birds withPDD may also be pruritic and have FDB.[29]Dilation of the proventriculus can be observed ra-diographically as an organ shadow that passesfrom rostral to caudal and extends beyond theliver margin on the left side on a VD view. It maybe diagnosed using barium to coat the proven-triculus to delineate it if it extends beyond theliver margin. Additionally, birds with PDD havealtered motility of their proventriculus and giz-zard (ventriculus), and those changes can best beobserved using fluoroscopy.
Some birds with organ dysfunction may bepruritic, as is the case with liver disease, or maypick over a localized area where the organ is situ-ated, as with the kidneys. When there is suspectedorgan dysfunction, ultrasound evaluation of theorgan can be performed with a possible fine nee-dle aspirate as a biopsy.[29]
REFERENCES
01. Dodman, N.H. 1998. “Pharmacologic treatment ofaggression on veterinary patients.” In Psychophar-macology of animal behavior disorders, ed. N.H.Dodman and L. Schuster, pp. 41–63. Malden, MA:Blackwell Science.
02. Welle, K. 2003. “Evaluation, diagnosis, and modi-fication of behavioral disorders.” Proc MASAAVAvian Med Surg Conf, pp. 176–191.
03. Sackett, D.L., S.E. Straus, W.S. Richardson, W.Rosenberg, and R.B. Haynes. 2000. Evidence-based medicine: How to practice and teach EBM,2nd ed. London, England: Churchill Livingstone.
04. Orosz, S.E., and C.J. Delaney. 2003. “Self-injuri-ous behavior (SIB) of primates as a model forfeather damaging behavior (FDB) in companionpsittacine birds.” Proc Annu Conf Assoc Avian Vet:Avian Specialty Advanced Program—AnotherFeather Picker: That Sinking Feeling, pp. 39–50.
05. Novak, M.A. 2003. Self-injurious behavior in rhe-sus monkeys: New insights into its etiology, phys-iology, and treatment. Am J Primatology 59:3–19.
06. Jones, I.H., and B.M. Barraclough. 1978. Auto-mutilation in animals and its relevance to self-injury in man. Acta Psychiatr Scand 58:40–47.
07. Haines, J., C.L. Williams, K.L. Brain, and G.V.Wilson. 1995. The psychophysiology of self-mutilation. J Abnorm Psychol 104 (3):471–489.
08. Bellanca, R.U., and C.M. Crockett. 2002. Factorspredicting increased incidence of abnormal behav-ior in male pigtailed macaques. Am J Primatol 58(2):57–69.
09. USDA. 1985 and amendments. Animal WelfareAct, Subchapter A, Animal Welfare, Subpart D—Specifications for the Humane Handling, Care,Treatment, and Transportation of NonhumanPrimates; Paragraph 3.81, Environment enhance-ment to promote psychological well-being; AnimalPlant Health Inspection Service, USDA.
10. Meehan, C.L., J.R. Millam, and J.A. Mench. 2003.Foraging opportunity and increased physical com-plexity both prevent and reduce psychogenicfeather picking by young Amazon parrots. AppliedAnim Behav Sci 80:71–85.
11. Snyder, N.F.R., J.W. Wiley, and C.B. Kepler. 1987.The parrots of Luquillo: Natural history and con-servation of the Puerto Rican parrot. Los Angeles:Western Foundation of Vertebrate Zoology.
12. Oviatt, L.A., and J.R. Millam. 1997. Breeding be-havior of captive orange-winged amazon parrots.Exotic Bird Rep 9:6–7.
13. Huber-Eicher, B., and B. Wechsler. 1997. Featherpecking in domestic chicks: Its relation to dust-bathing and foraging. Anim Behav 54:757–768.
14. Huber-Eicher, B., and B. Wechsler. 1998. The ef-fect of quality and availability of foraging materi-als on feather pecking in laying hen chicks. AnimBehav 55:861–863.
15. El-lethey, H., T.W. Jungi, and B. Huber-Eicher.2001. Effects of feeding corticosterone and hous-ing conditions on feather pecking in laying hens(Gallus gallus domesticus). Physiol Behav 73:243–251.
16. Garner, J.P., C.L. Meehan, and J.A. Mench. Cage-induced stereotypy in parrots and brain dysfunc-tion: Parallels to schizophrenia and autism.Submitted.
17. Orosz, S.E. 2003. “Anatomy and physiology of theintegumentary system.” Proc Annu Conf Assoc
17 / Diagnostic Workup of Suspected Behavioral Problems 209

Avian Vet: Avian Specialty Advanced Program—Another Feather Picker: That Sinking Feeling, pp.3–12.
18. Speer, B.L. 2003. “Sex and the single bird.” ProcAnnu Conf Assoc Avian Vet, pp. 331–343.
19. Hooimeijer, J. 2003. “A practical behavior protocolfor dealing with parrots.” Proc Annu Conf AssocAvian Vet, pp. 177–181.
20. Speer, B.L., and P.H. Kass. 1995. “The influenceof travel on hematologic parameters in hyacinthmacaws.” Proc Annu Conf Assoc Avian Vet, pp. 43.
21. Orosz, S.E., J.M. Grizzle, J.W. Bartges, N.K.Zagaya, A.K. McGee, J. McGinn, and C. Cray.2000. “A critical care diet for use in parrots.” ProcAnnu Conf Assoc Avian Vet, pp. 7–9.
22. Reavill, D. 2003. “Inflammatory skin diseases.”Proc Annu Conf Assoc Avian Vet, pp. 13–24.
23. Garner, M.M. 2003. “Avian noninfectious skin dis-orders.” Proc Annu Conf Assoc Avian Vet, pp.21–24.
24. Koski, M.A. 2002. Dermatologic diseases inpsittacine birds: An investigational approach.Semin Avian Exotic Anim Med 11 (3):105–124.
25. Gerlach, H. 1994. “Avipoxvirus affects most of thefamilies or groups of birds with some cross infec-
tion depending on the group. Viruses.” In Avianmedicine: Principles and application, ed. B.W.Ritchie, G.J. Harrison, and L.R. Harrison LR, pp.862–948.. Lake Worth, FL: Wingers.
26. Woods, L.W., and K.S. Latimer. 2003. Circovirusinfection of nonpsittacine birds. J Avian Med Surg14 (3):154–163.
27. Pollock, C.G., and S.E. Orosz. 2002. Avian repro-ductive anatomy, physiology, and endrocrinology.Vet Clin North Am Exot Anim Pract 5 (3):441–474.
28. Speer, B.L. 2003. Avian medicine today: Settingthe standard. Proc Bayer Exotics Symp 25 (3A):21–31.
29. Rosenthal, K.L. 2003. “Diagnostics: Please letthere be an answer.” Proc Annu Conf Assoc AvianVet: Avian Specialty Advanced Program—AnotherFeather Picker: That Sinking Feeling, pp. 25–30.
30. Speer, B.L. 2003. “Zinc toxicosis: Separating factfrom fiction.” Proc 24th Annu Conf Avian MedSurg: MASAAV, pp. 156–159.
31. Levengood, J.M., G.C. Sanderson, W.L. Anderson,G.L. Foley, P.W. Brown, and J.W. Seets. 2000.Influence of diet on the hematology and serumbiochemistry of zinc-intoxicated mallards. J WildlDis 36 (1):111–123.
210 Manual of Parrot Behavior

Biting is one of the most common and definitelythe most serious behavior problem in any pet.Parrots are no exception. Psittacine aggression isone of the more serious of behavioral problemsfor pet bird owners. The strong jaws and hookedbill of parrots can inflict serious pain and do sub-stantial damage to the owners. Aggression in par-rots takes the form of biting or lunging at a per-son or another bird. Parrots will use biting as alast resort for protection and to guard resources.They also are very intelligent. Their experiencewill determine what behaviors are most effectiveto accomplish the goals they are looking toachieve. Some birds will learn that aggressive be-havior is very effective, while others may find an-other strategy works best for them.
While the aggression of an individual bird canhave overlapping and interacting etiologies, it ishelpful to categorize the problem in order to treatit. As in any other species, aggressive behaviorhas several etiologies.[1] Aggressive behavior eti-ologies in pet birds include fear, conditioning, ter-ritoriality, and attachment to a mate. Diagnosis ofany behavioral disorder is based upon the behav-ioral history, testing the bird’s response to varioussituations, and direct observation of the bird.Specifically, the signalment, a description of theenvironment and social interactions, and a de-scription of the aggression, the circumstances inwhich it occurs, and the owner’s reaction to thebehavior are critical to the assessment of a bitingbird.[2] It is particularly important to see thebird’s interaction with owners and other people.Aggressive birds will often behave much better inthe veterinary hospital or other unfamiliar placethan in their home environment. If all of the ob-servations are made away from the home terri-tory, the true behavior of the bird may not be
noted. A house call, or better yet, videotapes ofthe bird and owner at home are a much preferablemeans of assessing the behavior.
FEAR BITING
With parrots being a prey species, fear is one ofthe most common motivators for biting. A birdthat is frightened by people is simply trying to de-fend itself. There does not appear to be any age orsex predilection. This is technically not a behav-ior problem. Biting is a perfectly reasonable reac-tion to something thought to be a threat. The realproblem is the fact that the bird perceives some-one as a threat. The reasons that this occurs vary.Wild birds naturally fear humans. The process ofcapture, shipping, and sale does little to reducethese fears. These wild-caught birds, however, areless common than in years past. Most of those re-maining have had years to either reduce or con-firm their fears of humans. The vast majority ofpet birds are captive bred and have lived their en-tire lives around humans. While most Budgeri-gars and many other small psittacine birds areparent-reared, most larger parrots are raised byhumans. Their fears of humans are much moredifficult to explain. While overt abuse wouldreadily explain a bird’s fear of humans, there islittle evidence of such abuse in most cases.Certain species are more likely to develop fear ofpeople despite the fact that they have been raisedaround them. African Grey Parrots are wellknown for this problem. The factors involvedvary. Sometimes these birds are not socialized ad-equately when young. Some may have had exces-sively severe wing trims, causing them to besomewhat clumsy. These birds will then have fre-quent falls, feather and beak breakage, and otherpainful events that make them fear coming out of
211
18Aggressive Behavior in Pet Birds
Kenneth R. Welle and Andrew U. Luescher

their “safe haven” within their cage. The com-plexity of the early environment has an effect onemotional stability, too (see chapter 11), andmany hand-raised birds are kept in a severely re-stricted environment until fledging.
There are certain features to this type of bitingthat are relatively easy to recognize. Usually, thesebirds only bite when cornered or caught. They willnot often attack or chase someone. They may learnto attack rather than retreat when the cage door isopened, but they will generally try to get past thehandler rather than defend the cage. Fearful vocal-izations such as growling or screaming will oftenprecede or accompany the biting. Many will be“cage bound,” very reluctant to leave the cageeven on their own. When they are presented to theveterinary hospital, they will be extremelystressed, often vocalizing almost constantly. Theymay begin to cower, pant, or thrash about when thecage is approached. A stress leukogram is verycommon with these birds. Most affected birds willnot allow handling by anyone, although occasion-ally birds will be fearful of a particular person,gender, behavior, or a physical characteristic of aperson. One behavior, which is very likely to initi-ate biting, is when people offer a hand and thenjerk it back when the bird reaches out with thebeak. Many fear biters are somewhat clumsy andthey may have fallen when people have pulled thehand away from the beak.
Treatment of this problem requires a lot of pa-tience. Affected birds should be gradually desen-sitized to the presence of people. Depending onthe severity, initial contact may have to be limitedto basic care of the bird. If the bird accepts any-one, that person should do all of the basic main-tenance until the bird learns to accept others. Asthe bird begins to remain calm in the presence ofthe feared person, then eye contact and vocal in-teraction can be tried. In extreme cases, anxiolyticdrugs can be used to facilitate the early stages oftreatment. When basic non-contact communica-tion has progressed to a point where the bird iscalm, then handling can be tried. Since the bird isalready fearful, coaxing and reward should beused here. A favorite food item should be re-served for this purpose. It may be necessary to re-strict food slightly to encourage the bird to take atreat. Initially a treat should be placed on thefloor, in the food dish, or on the roost, and thehand withdrawn less and less far, so the bird gets
rewarded for approaching the hand. Then basicoperant conditioning can be used to pattern thebird to step out of the cage, and finally onto thehand. Clicker training would be an excellentmethod to deal with fearful birds (see chapter 20).The voice should remain calm and coaxing at alltimes. If the biting is severe, it may be necessaryto use a handheld perch or even a protective glovefor initial handling. Despite warnings aboutgloves causing greater fear, used properly, theycan be useful tools. Falconers have used glovesfor a thousand years, and they have never madetheir charges more fearful of humans. When usinghandheld perches or gloves, it is important thatthe bird is desensitized to them before stickingthem up to the bird’s chest. This can be achievedsimply by laying them a little closer to the cageevery day, and eventually moving them gently atdecreasing distances to the cage while the birdeats its favorite treat. The glove should only beused as a perch for the bird, never to grab or re-strain it. The advantage of the glove or perch isthat it can give the handler more confidence tohold the bird steady than a bare hand. A stabledowel or glove is much better than a hand that ispulled away. If there is any indication that gloveshave been used for restraint in the past, they shouldnot be used with that particular bird. Punishing afear biter is contraindicated. The biting behavior inthis situation is a normal response. The situation isabnormal and this is what should be addressed.
The prognosis for fear biting is fair. Successdepends on the patience and commitment of theowner. The speed of progress will depend on theseverity of the fear. Birds that have very general-ized fear of humans will require a greater lengthof time than those that simply fear hands. Theability of the owner to offer a steady hand to thebird is critical as well. If a stable perching surfacecannot be offered to the bird, then stepping upwill rarely occur.
CONDITIONED AGGRESSION
This type of aggression is often called dominanceaggression. Dominance aggression is a controver-sial classification. The term implies a social ordermaintained by subtle, species-typical cues thatserve to minimize aggression. This communica-tion is not possible between birds and people.However, birds can easily be conditioned to beaggressive by a person backing off or in other
212 Manual of Parrot Behavior

ways rewarding the aggression (see chapter 20).Fear aggression, mate-related aggression, and ter-ritorial aggression are very easily conditionedthat way. Also, if the bird was able to control theowner’s behavior for some time, and then theowner does not comply with the parrot’s expecta-tions, there is what is called a “frustration effect”that easily results in an outburst of aggression. Ifthen the owner does act according to the parrot’sexpectations, this aggression is reinforced. If theaggression is consistently successful, the bird be-comes very confident in using aggression, so afear aggression may change into confident, or of-fensive, aggression.
This type of aggression may occur in anyspecies but is particularly common in amazonsand macaws. It occurs more often in mature thanin juvenile birds. It has been suggested that malebirds are affected more commonly, but the gen-der of many birds is undetermined. In dogs, theanalogous problem is most commonly seen withowners with anthropomorphic, and thus incon-sistent, involvement with their pet, and in first-time dog owners.[3] In parrots it seems to occurmore commonly with birds belonging to peoplewith little experience with birds. It also occursmore frequently in larger species of birds. This ismore likely due to the fact that more people willbe intimidated by larger species than the fact thatthese birds are particularly aggressive. Affectedbirds have often learned to manipulate the ownerto their wishes. The owners’ behavior may be asuseful for a diagnosis as is the bird’s behavior.They often will make excuses for the bird’s be-havior, take extraordinary measures to preventupsetting the bird, or otherwise defer to thetantrums of the bird.
These birds will show a particular pattern ofaggression. The bird may bite certain familymembers but not others. When the interactionsare observed the ones bitten are less confidentand assertive with the bird than those that are notbitten. The biting does not appear to be influencedby the bond between bird and owner; the personbitten may be the bird’s favorite person. Bitingmay occur when the owner tries to get the bird offthe shoulder, put the bird in a cage, or remove thebird from the perch. These birds may occasionallybite when the owner stops petting them. The own-ers often blame themselves for the bite and willcomply.
For most animals, loss of control over the envi-ronment, alongside loss of the ability to predictwhat is going to happen, is very stressful. This ismost certainly true for intelligent animals such asparrots. They therefore strive to have control overthe environment. If the owners do not establishthe contingencies between the bird’s behavior andthe effects it has, the birds will. This is not an ab-normal behavior, but simply necessary for thebird’s well-being. Consistency in interaction withthe bird and training will give the bird control andmake the environment predictable in a mannerthat is also acceptable to the owners. The birdsimply has to do the right behavior to get what itwants, and the bird also learns when such behav-ior is going to pay off and when not. Aggressivebirds have been patterned to use aggressive be-havior to control the behavior of their owners,who have been patterned to comply. Both ownerand bird may be resistant to changing the situa-tion. After years of being bitten, the owner will behesitant to be assertive with the biting bird. Thebird will initially be a bit confused and will try tore-establish control. Aggression may get worsebefore it gets better. In some cases, the bird willbenefit from placement in a home with a moreconfident and consistent handler.
The wings should be trimmed to give the birdone less means of controlling situations. All han-dling should take place in neutral territory. Ifthere are any household members that can safelyhandle the bird, they should bring the bird to theneutral area. If no one can safely handle the bird,the owners should be schooled on safe towel cap-ture and restraint of the bird. Once in a neutralsite, out of the sight of the cage, step-up exercisesshould be practiced. The owners should becoached on confident and consistent handlingtechniques. If the owner cannot keep the hand infront of the bird without withdrawing from thebeak, then a perch or gloved hand should be used.All of the precautions regarding the use ofperches and gloves discussed previously shouldbe followed here as well.
The shoulders should be considered off-limits.This can be challenging in its own right if the birdis accustomed to riding on the owner’s shoulders.Hand position should be higher than the elbow sothe bird does not just climb up onto the shoulder.Birds are reluctant to slide down the arm to theelbow and then up the upper arm to the shoulder.
18 / Aggressive Behavior in Pet Birds 213

To prevent the bird from jumping to the shoulder,the feet may have to be restrained. Placing a towelor other object on the shoulder may discouragethe bird from jumping on the shoulder. If the birdjumps onto the shoulder with the towel on it, thetowel and bird are quickly removed.
The owners should be taught to recognizethreat behavior. When the bird displays threat be-havior, or a bite occurs, the bird should be givena command and rewarded for the appropriate be-havior. In this way the owner does not have to usea punishment but rather teaches the bird appropri-ate behavior. The beak should not be “grabbed” or“thumped,” as this can escalate the aggression.Birds should never be hit. Yelling or other types ofdrama can actually be entertaining for some birds,so the victim should always remain calm. Thevery effectiveness of the bite is what reinforcesthe behavior. Unless there is a significant anxietycomponent to the aggression, drug therapy is notindicated for this problem.
Prognosis is fair for the bird but guarded for therelationship between the owner and bird. Thelonger the duration of the problem, the more dif-ficult it is to reverse the problem. One client ofone of the authors (Welle) had a Scarlet Macawthat for eight years had controlled her by biting.Although the author established that this wouldnot work with him within one session, the macawstill viciously attacked the owner whenever shedid not comply with the bird’s wishes. It is diffi-cult for a bird with eight years of experienceshowing it can manipulate a person by biting tounlearn this behavior. However, the birds quicklylearn that a new person cannot be controlled, andso they will no longer try as hard.
TERRITORIAL AGGRESSION
Territorial aggression is particularly common incertain species. Quaker Parakeets are well knownfor this trait. Conures, miniature macaws, AfricanGrey Parrots, and amazon parrots are also proneto develop this problem. Breeding birds of allspecies tend to be territorial about their nest areaand cage (Figure 18.1). In these birds, this behav-ior is considered desirable. Guarding of the nestterritory is an important breeding cue. While thesex of many avian patients is unknown, it appearsthat territorial aggression is somewhat more com-mon in males than in females.
Diagnostic criteria for territorial aggression are
very simple. Territorial aggression occurs whenthe bird is in or on its cage, playpen, or other liv-ing area. If this is not the case, the aggressioncannot be territorial aggression. Aggressive be-havior must be limited to these areas. It is possi-ble, however, for a bird to exhibit both territorialaggression and other forms of aggression. Somefearful birds will appear to have territorial aggres-sion because they attack when a hand comes intotheir cage. These birds have learned that there isno retreat within the cage and therefore will at-tack a hand as it enters the cage. The way to dis-tinguish between the two is by observing the otherbehaviors. If fearful vocalizations occur, if the ag-gression occurs at locations other than the pri-mary living space and the bird is intolerant ofhandling under various situations, the problem ismore likely to be fear biting. However, fearfulbirds can attack in a confident way, without show-ing fear, if they have learned that the aggressionworks in getting the owner to withdraw. If the birdattacks without vocalizing or when the cage is notopened, but when away from the cage exhibits nofear or aggression, the problem is more likely ter-ritorial aggression.
Behavior modification for territorial aggres-sion is multifaceted. In order to achieve the spe-cific goals, general obedience training is essen-tial. The step-up command should be automaticfor the bird. This command is an important train-ing tool. To avoid injury to the owner, the birdshould be removed while servicing the cage.Owners should be schooled on atraumatic but se-
214 Manual of Parrot Behavior
Figure 18.1. Territorial behavior is normal inbreeder birds such as this Mitred Conure (Aratinga mitrata).

cure towel restraint of the bird if necessary. Birdsthat will not leave the cage without biting shouldbe caught and carried to a separate area. Somebirds can be safely handled following voluntaryexit from the cage. This will facilitate the othertraining measures.
An attempt should be made to make the birdless dependent on the cage. This helps both in theprevention and treatment of territorial aggression.Birds that spend most or all of their time in onecage can become viciously aggressive about de-fending it.[4] In the wild, birds roost in the samearea each night. During the day, they travel toother locations to forage, usually with their flock.A two-cage housing system helps provide a morenatural system (see chapter 26). A large, well-furnished cage or playpen should be used duringthe daytime to encourage activity. The cage shouldbe rearranged frequently to promote adaptabilityin the bird. During the night, a smaller roostingcage with rather spartan accommodations shouldbe used. Each morning the bird can be transportedto the larger cage and each evening to the roostingcage. If further measures are needed, the bird canbe meal fed twice daily in another location.
The bird should be integrated into the familysocial unit. Portable perching stations allow thebird to sit close to the activity of the family. This,combined with consistent handling and training,provides the bird with the social skills needed tobe well-adjusted pets. The two-cage housing sys-tem described previously forces owners to handlebirds at least twice daily to transport them.
Regularly scheduled handling and training ses-sions are important for maintaining socializationand control. The step up, rewarded with foodtreats, is especially important. A narrow T-standshould be used initially to train the bird. The T-stand is like a leash on a dog: it prevents the birdfrom moving too far. The T-stand is initiallyplaced in a location away from the cage (differentroom). As training progresses, the T-stand can bemoved closer and closer to the cage. Eventually,the step up can be practiced in the cage. This ne-cessitates a widely opening cage door. The train-ing should first be done by the person having theleast problems, and then by other people. Everynew person has to start over from the beginning,with the T-stand far away from the cage.
Psychotropic drugs are not generally indicatedin the treatment of territorial aggression in birds.
The chemical basis of aggression is unknown inbirds. Additionally, the prognosis for treatment ofterritorial aggression with behavioral modifica-tion alone is favorable.
MATE-RELATED AGGRESSION
Parrots are prone to developing unhealthy pairbonds with one of their owners. While a bird maybond to more than one person, only one is chosenfor a mate. However, if the chosen mate is theonly one who ever handles the bird, the aggressivetendencies will be much worse. Mate-related ag-gression occurs most commonly in hand-raisedbirds of larger parrot species. Amazon parrots,macaws, cockatoos, and Quaker Parrots are allcommonly affected. Male birds are most often af-fected. The behavior often begins or becomesmore serious at sexual maturity or during breed-ing season.
The aggression will most often occur in situa-tions where the bird’s favorite person is ap-proached by someone else, such as another fam-ily member. The aggression may be directed to-ward the rival or paradoxically redirected towardthe favorite person. Occasionally, an inanimateobject such as a telephone, or a cavity such as anopen drawer, may trigger the aggression. Unlikemost other types of aggression, these birds willoften attack or chase their victims. They are oftenexceptionally cuddly at other times, at least to-ward the favorite person.
To help minimize mate-related aggression, thebird should be socialized appropriately. The per-son to whom the bird has pair bonded shouldwork to develop a more platonic bond. Interac-tions should be more active and dynamic, avoid-ing cuddling and petting. The bird should not beallowed on the shoulder. This position in closeproximity to the face encourages pair bonding.Additionally, the damage done with a bite can bemuch greater. All members of the householdshould take the bird to novel, unfamiliar places sothey can be seen as the familiar, comforting fig-ure. In these locations, basic commands such as“step up” and “step down” should be practiced byall family members. The favored person shoulddo all of the unpleasant tasks such as grooming,and otherwise largely ignore the bird until theproblem is resolved. Only non-favored peopleshould give favorite treats.
The attacking bird should be gently captured in
18 / Aggressive Behavior in Pet Birds 215

a towel and placed in the cage or other controlledarea. Owners should be taught how to safely andeffectively towel restrain these birds. If the ownercan predict the aggression, the preferred personcould leave swiftly and therewith remove thecause of the aggression.
In some cases, the aggressive behavior can bereduced with hormonal therapy. Leuprolide ac-etate can reduce the sexual hormones and therebyreduce the intensity of the aggressive behaviorduring the initial phases of behavior modifica-tion. Orchiectomy can reduce aggressiveness insome cases.[5]
REDIRECTED AGGRESSION
Occasionally, the person who provided the stimu-lus for the bite is not the one bitten. If a birdcannot reach the person it intends to bite, it willsometimes bite whomever happens to be close. Re-directed aggression is not a diagnosis but refers toa mechanism that modifies other types of aggres-sion. Frequently, it is mate-related aggression thatgets redirected, in this case usually to the bondedperson because he or she is usually close by. Totreat the problem of redirected aggression, the ag-gression that is redirected needs to be addressed.
INTRASPECIFIC AGGRESSION
Aggression toward other birds represents anothercommon problem in companion and aviculturalbirds. Many bird owners have more than one birdand injuries of one by another are relatively com-mon. In aviculture, pairs must be kept together inthe breeding season in order to produce offspring,so safe congregation is crucial. In many cases thecauses for aggression between birds are the sameas aggression toward humans. Fear, territorial de-fense, and mate-related aggression can all play arole. Often, however, the problem is a result of alack of adequate flock social skills in the birds in-volved.[6] Many pet and breeding birds todayhave had little or no experience with other birds.They have not learned from parents or other flockmembers how to behave in a socially acceptablemanner. The result may be aggression exhibitedby or toward the poorly socialized bird. Whilesome authors advise that parrots should be social-ized specifically as replacement breeders, othersadvise keeping parrots as pets for only the firstseveral years and then placing them into breedingcollections.[7]
In pet birds, simply housing birds individuallyand supervising them when they are not confinedcan often avoid the problem. If cages are placedin proximity, the birds may become increasinglytolerant of the other bird. Care must be taken, es-pecially when one bird is loose and the other isconfined. It is very common for foot injuries tooccur when a bird climbs on the cage of anotherbird, resulting in a bite to the foot from the con-fined bird. Occasionally conflicts arise because oflimited access to resources. Mild aggression cansometimes be alleviated by providing extra re-sources, such as perching space, food and waterdishes, and toys. Overcrowding can easily lead toaggression and is a major stressor for any captiveanimal.
Aggression toward mates is a very commonproblem, especially in cockatoos (see chapter22). Male breeding parrots defend their territoryvigorously. If the proximity of another pair stim-ulates aggressive behavior, this can be redirectedtoward the female. Visual barriers, at least in thearea of the nest site, can help avoid this problem.It is also possible that the pair bond is dissolv-ing.[8] While the pair bond is usually permanent,occasionally parrots will leave a former mate for another. By allowing flocking in the non-breeding season, the birds can either re-establishthe pair bond or form a new pair prior to the nextseason.[6]
CONCLUSION
Like any behavioral disorder, improvement willusually occur slowly and gradually. Ownersshould be advised to keep a journal of their bird’sbehavior. The frequency, severity, and nature ofany occurrence of aggression should be logged. Areduction in the frequency or severity of aggres-sion indicates that the treatment plan is working.The patient should be re-evaluated every six toeight weeks and the client should be interviewedto determine if the treatment plan is being fol-lowed correctly. Alterations can be made if ade-quate progress has not been made. In cases wherethe problem has not improved, the diagnosisshould be reassessed. Depending upon the sever-ity of the problem and the personality of theowner, placement of the bird into a new home canbe considered. If this situation occurs, the veteri-narian should coach the new owner so that thesame cycle does not begin again.
216 Manual of Parrot Behavior

REFERENCES
1. Welle, K.R. 1998. “Psittacine behavior.” Proc An-nual Conf Assoc Avian Vet, pp. 371–377
2. Welle, K.R. 1999. Psittacine behavior handbook.Bedford, TX: Assoc Avian Vet Publication Office.
3. O’Farrell, V. 1995. “Effects of owner personalityand attitudes on dog behaviour.” In The domesticdog, ed. J. Serpell, pp. 153–158. Cambridge: Cam-bridge University Press.
4. Athan, M.S. 1993. Guide to a well-behaved parrot.Hong Kong: Barron’s, pp. 36–54.
5. Bennet, R.A. 2002. “Reproductive surgery inbirds.” Proc Atlantic Coast Veterinary Conference.
6. Styles, D.K. 2002. “Captive psittacine behavioral re-productive husbandry and management: Socializa-tion, aggression control, and pairing techniques.”Proc Annu Conf Assoc Avian Vet Specialty Program.
7. Clubb, S.L. 1998. Captive management of birds fora lifetime. JAVMA 212:1243–1245.
8. Clubb, K.J., S.L. Clubb, S. Phillips, and S. Wolf.1992. “Intraspecific aggression in cockatoos.” InPsittacine aviculture: Perspectives, techniques andresearch, ed. R.M. Schubot, K.J. Clubb, and S.L.Clubb, chapter 8. Loxahatchee, FL: AviculturalBreeding and Research Center.
18 / Aggressive Behavior in Pet Birds 217

The ability to vocalize is arguably both one of themost and least endearing traits in a pet parrot. Thepopularity of some species of parrots as pets,such as African Greys, derives from their abilityto speak, to mimic human speech and othernoises. However, parrots’ ability to vocalize andnormal patterns of vocalization often becomeproblematic for pet bird owners.
Naturalists have noted several features aboutpsittacine vocalizations in the wild that have rele-vance for the captive management of these birds.First is the daily pattern of vocalization. In mostspecies of parrots, especially those commonlykept as pets, such as amazons, macaws, and cock-atoos, the flock will be quiet from sunset until thenext dawn (Snyder 1987; Rowley 1990). At day-break, the flock will vocalize and fly around theroosting area before setting out to forage for theday in a different location. Again, as dusk ap-proaches, after the birds have returned to theroosting area, there is a period of vocalization.
In a household setting this pattern can lead toproblems, especially when birds vocalize at sun-rise and owners or their neighbors are not ready towake up. Owners may also complain of birds“screaming” when they come home from work.Vocalizations at this time of day may be due to acombination of factors—greeting the returning“flock member” and normal, pre-sunset, vocal-izations. The first step in treating these problemsis owner education. Pet parrot owners must beaware that their birds, no matter how bonded theyare to people and even if bred in captivity andhand-raised, are still wild animals. With the pos-sible exceptions of Cockatiels and Budgies, petparrots do not meet the basic definition of domes-ticated animals in that pet parrots are not geneti-cally different from wild parrots as a result of se-
lective breeding (Hurnik 1995). As a startingpoint in treating any pet bird behavior problem,owners must understand that they may be able tomodify the expression of their birds’ normal be-haviors but won’t be able to completely eradicatethese behaviors. In the case of morning andevening vocalizations, once owners understandthat their birds will still vocalize twice daily, thenthey are ready to start to shape those periods ofvocalization into more acceptable ones. Mostspecies of parrots that are kept as pets are tropicalor semi-tropical (Forshaw 1989). To replicate nat-ural conditions they should be receiving close to12 hours of light and 12 hours of dark each day.One easy way to control vocalizations is to con-trol the bird’s dark/light cycle. Covering cages orhaving birds sleep in a dark room allows ownersto set when “sunrise” occurs. Most parrots willnot vocalize in the morning until after daybreak.Likewise, owners may be able to shift eveningvocalizations by controlling when “sunset” oc-curs. Owners can also redirect these natural peri-ods of vocalization into more acceptable behav-iors by giving the bird another activity to performat these times. For example, in the evenings theowner can take advantage of the bird’s naturalproclivity to vocalize by using this time to teachthe bird new phrases or sounds to say. Alternately,owners can pre-empt and reduce some of thevocalizations by giving the bird a new toy or spe-cial food at the times that it is likely to vocalize.It is important that the toy or treat be given beforethe bird starts to vocalize. If this is not the casethen the bird may learn that by yelling it earns areward.
Naturalists have also noted several differenttypes of vocalizations from wild parrots. Theseinclude alarm calls, contact calls, food begging
219
19Parrot Vocalization
Laurie Bergman and Ulrike S. Reinisch

calls, and interspecific agonistic calls. Many ofthese vocalizations are learned by parrot chicksfrom their parents and flock mates. Naturallycross-fostered Galahs (Cacatua roseicapilla) thatwere reared by Pink Cockatoos (Cacatua lead-beateri) have contact calls like their foster par-ents, not like their own species. Budgerigars(Melopsittacus undulates) that are reared in isola-tion not only exhibit abnormal vocalizations butalso abnormal behaviors. Among these abnormalbehaviors is evidence of social bonding to inani-mate objects exhibited by warbling as thoughcourting selected objects (Farabaugh & Dooling1996). A hand-reared pet parrot will not have theexact same vocalizations as its wild relatives, butit will show similar patterns of vocalizations anduses of vocalizations.
Because almost all species of psittacines arehighly social, as flock dwellers parrots have de-veloped a variety of vocalizations that serve ex-clusively or primarily as contact calls. These arecalls that serve to identify where other membersof the flock are and help promote flock cohesion.Unfortunately, most pet parrots are not main-tained in flocks. Even if there are other birds inthe household parrots often form inappropriatepair bonds with their owners. When separatedfrom the owner these birds will vocalize. Initiallythey may give contact calls, which may progressto more distressed and anxious vocalizations ifthey receive no response to the calmer contactcalls. In some birds this vocalization may onlytake place when the owner leaves the house.Other birds may give contact calls every timetheir owners leave their sight. One way to addressthis problem is by having the owner maintain au-ditory contact with the bird when in another partof the house. For some birds this can be as simpleas hearing the owner whistling or talking while inanother room. The owner’s vocalizations need notbe in response to the bird’s contact calls but canbegin when he or she leaves the room before thebird begins to vocalize.
In more severe cases the bird should be treatedfor separation anxiety. This treatment begins byreducing the bird’s dependence upon the ownerwhile the owner is at home. This is accomplishedby limiting interactions with the bird, by makingthe interactions more structured through positivereinforcement-based training, and by providingthe parrot with alternatives to interacting with the
owner. These alternates can be toys, especiallyfood-dispensing toys or toys that can be chewedup or shredded, or special food treats, especiallychallenging foods like nuts that must be crackedor whole fruits. Other visual and auditory stimuli,like television or radio, can help give the parrotanother focus for its attention. The other part to aseparation anxiety treatment plan is using a seriesof very short departures to desensitize the bird tobeing left alone. These departures must be briefenough that the bird does not experience any anx-iety whatsoever. During these practice departuresthe parrot is provided with special food treats andtoys to give it opportunities for other more re-warding behaviors than calling out to its owner.
Vocalizations that serve as alarm calls have alsobeen identified in a number of parrot species. Insome species, such as Cockatiels (Nymphicus hol-landicus) and the Red-rumped Parrot (Psephotushaematonotus), several different types of alarmcalls that indicate varying levels of distress havebeen identified (Pidgeon 1981). In a pet situationowners may hear these calls in response to actualor perceived threats to the birds. Many owners areable to identify the function of these particular vo-calizations and realize that their birds are dis-tressed and not “just screaming.” This knowledgemay allow the owner to respond to the screamingby removing the stimulus that is causing thealarm. For example, one of the author’s (Bergman)Indian Ringneck Parakeets would only give analarm call if a dog was near his cage. The ownerlearned to call the dog whenever she heard thebird’s alarm call. Once the dog moved away fromthe bird’s cage the bird would stop calling andrelax.
Alarm calls can become problematic. By na-ture these calls tend to be loud and of a frequencythat carries well in order to warn the entire flockand possibly drive away threats (Fernandez-Juricic et al. 1998). A pet parrot that alarm callsfrequently can be quite disturbing to the peoplewho must live with or near the bird. Furthermore,a pet parrot that is alarm calling frequently is awelfare issue. These vocalizations are given whenthe bird is in distress. The presence of frequentalarm calls can indicate a poor husbandry situa-tion in which a bird is being kept in a distressingenvironment.
The first and most important step in treating aparrot with excessive alarm calls is to identify and
220 Manual of Parrot Behavior

remove the stimuli that are provoking the vocal-izations. These stimuli may be inanimate objectsin the bird’s immediate environment (e.g., toys,cage furniture) or anywhere within the bird’s lineof sight. Other possible stimuli are people orother animals. Remembering that pet parrots arewild animals that are a prey species in the wildcan help to identify the source of the bird’s alarm.Alarm calls may be elicited by seeing potentialpredators, dogs, cats, birds of prey, snakes, and soforth in the house, through a window, or in pic-tures or on television.
Often parrots will give alarm calls directed atunfamiliar or familiar but less-favored people.Provided the bird is not showing other signs ofmore significant fear or distress, such as attempt-ing to escape from the area, this behavior is besttreated by ignoring it. If the alarming person ap-proaches the bird, the bird may panic or act ag-gressively. If the owner or another favored personapproaches and attempts to calm or reassure thebird, then the bird is being rewarded for screamingout alarm calls. This will teach the bird that mak-ing alarm calls works to get the owner’s attentionand may result in a bird that alarm calls simply asan attention-seeking behavior and not out of dis-tress. Instead, the owner and the other peopleshould wait until the bird has stopped screamingand then reward the bird with a delicious treat oran object to play with or attention and affection.
Pet parrots may alarm call in response to beingin a new location or to changes in a familiar envi-ronment, such as new furniture or furniture thatwas moved. In this case “introducing” the bird toits environment is often sufficient to relieve thebird’s distress. The owner should walk around thearea with the bird, talking to the bird in an upbeatvoice, as though this tour is the most fun thingever to happen. The owner can touch things andgently show them to the bird. The key is for theowner to be relaxed and engaged with the envi-ronment and let the bird see that it is not threaten-ing. The parrot will then relax and choose thelevel of interaction with the environment that it iscomfortable with. The bird must not be pushedinto interactions for which it is not ready.
If the bird is panicked by the fear-provokingstimulus or is unresponsive to the previously de-scribed approaches, then systematic desensitiza-tion and counterconditioning should be used totreat the bird’s underlying fear and anxiety and
thereby reduce the problem vocalizations. Afterthe fear-provoking stimulus has been identifiedand removed from the bird’s environment, it isslowly reintroduced while the bird is rewarded forremaining calm and non-reactive. The stimulus isfirst introduced in a manner that does not causeany reaction. Gradually the level of stimulation isincreased. This can be done by altering the formof the stimulus. For example, a bird that was pan-icked by tie-dyed t-shirts was first introduced toplain t-shirts, then t-shirts with a small area of tie-dye, then t-shirts that were tie-dyed all over.Distance from the stimulus should also be used todecrease the intensity of the stimulus. The re-wards that the bird receives in the presence of thestimulus should be anything that the particularbird values highly. This can be food, toys, thingsto manipulate with its beak, petting, or being spo-ken to. Clicker training can also be a useful ad-junct to a desensitization and counterconditioningprotocol. In more extreme or refractory cases ad-junct drug therapy may be useful.
Problems can also arise from learnedvocalizations—words, phrases, and sounds thatparrots learn to mimic. Parrots may accidentallyor purposely be taught words or phrases that theirowners would prefer they do not say. These maybe as innocuous as an old flame’s name or as em-barrassing as obscenities. It is very likely thatbirds that are inadvertently exposed to expletivesmay repeat these words because they heard themsaid at a high volume and/or with a high level ofemotion. Parrots are often attracted to dramaticvocal displays from people and therefore mayfind these phrases memorable.
Many parrots are especially adept at mimick-ing electronic noises, such as the beeps from mi-crowaves, cell phones, and computers. Oftenthese sounds can become quite wearing on peoplewho have to hear them all day long. In addition,birds often learn that making these sounds causespeople to do things, like get up to check the mi-crowave or look for the ringing phone. The peo-ple’s behavior becomes rewarding for the birdsand encourages them to repeat these sounds moreoften.
To reduce these problematic learned vocaliza-tions owners must be prepared to be very patient.As is the case with many learned, rewarded be-haviors these vocalizations can be “unlearned” byignoring them via the process of extinction. Since
19 / Parrot Vocalization 221

parrots are so responsive to subtle body languageand facial expressions from people, owners mustbe prepared to be absolutely poker faced in thelight of whatever their birds may be saying.Owners should be taught about the phenomenonof extinction bursts, whereby the bird may repeatthe undesirable vocalization at a greater fre-quency or louder volume in response to the lackof reaction from the owner. Owners should alsobe aware that the slightest encouragement will re-sult in a resumption of the undesired behavior.This often becomes important when there are vis-itors at the house or other people caring for thebird. People who do not have to live with the par-rot may accidentally or purposely encourage thevery vocalizations the owners have been activelyworking to extinguish. If the bird is not respond-ing to the attempts at extinction a mild punish-ment may be added to the treatment plan. Thepunishment to be used, a form of social isolation,takes advantage of parrots’ natural gregarious-ness. The instant the bird begins the unwanted vo-calization all people present should turn awayfrom the bird or even leave the room, thereby“isolating” the bird for a brief (30 seconds) pe-riod. The idea is to teach the bird that the word orsound in question no longer gets a positive re-sponse; instead it drives people away.
Another situation of learned vocalizations cre-ating problems for pet parrot owners occurs aftera person in the household has had a cold. The par-rot begins to mimic the person’s coughs andsneezes. Often owners will present these birds toa veterinarian fearing that the bird has contractedthe person’s cold. These birds show no other signsof upper respiratory tract disease and their coughsand sneezes sound like human coughs andsneezes, not avian ones. If these vocalizations areworrisome for the owners ignoring them is thebest treatment.
The presenting complaint for problem vocal-izations is often simply that the bird is “scream-ing.” Although careful history taking may revealthat this is a problem that involves primarily con-tact calls (e.g., separation anxiety) or alarm calls(e.g., when there are visitors in the house), some-times it is difficult to pinpoint such a cause forthe problem vocalizations. In many such casesthe problem is primarily one of learned vocaliza-tions. The parrot has learned that people pay at-
tention to a screaming bird. Often the owners be-lieve that the attention they are providing is neg-ative and should stop the screaming. This oftenconsists of owners yelling at their birds or return-ing to the room where the bird is to “scold” thebird. Some owners will attempt other forms ofpunishment that are inappropriate and clearly in-effective if the problem continues. In cases of at-tention-seeking screaming the treatment is thesame as for the bird that is saying unwantedwords; attempt to extinguish the unwanted be-havior by ignoring it. Some owners can success-fully interrupt a screaming parrot by covering thebird’s cage. The cage should be uncovered imme-diately after the bird quiets down and the birdshould be given other things like toys or food tooccupy its time. Another way of reducing the vol-ume of a parrot’s vocalizations, whether atten-tion-seeking “screaming” or normal morning andevening vocalizations, is by teaching the bird towhisper. Reward the bird with attention, food, ortoys for speaking softly. This approach allows thebird to engage in a normal behavior but modifiesthat behavior into a form that is more acceptablefor life in captivity.
In rare cases parrots’ screaming becomes astereotypic behavior. Like all stereotypic behav-iors this can be thought of as an indication of in-adequate husbandry and a welfare problem. Thebird’s husbandry situation, including but not lim-ited to feeding, water source, caging, cage loca-tion, availability of toys, perch variety, light cycle,and contact with other birds/people, should be ad-dressed. Some of these birds might benefit fromtreatment with a psychotropic medication. Drugchoices would be similar to those used to treatpsychogenic feather picking.
REFERENCES
Farabaugh, S.M., and R.J. Dooling. 1996. “Acousticcommunication in parrots: Laboratory and fieldstudies of budgerigars, Melopsittacus undulates.” InEcology and evolution of acoustic communication inbirds, ed. D.E. Kroodsma and E.H. Miller,pp.97–117. Ithaca, NY: Cornell University Press.
Fernandez-Juricic, E., M.B. Maretlla, and E.V. Alvarez.1998. Vocalizations of the blue-fronted amazon(Amazona aestiva) in the Chancani Reserve, Cor-doba, Argentina. Wilson Bulletin 110 (3):352–362.
Forshaw, J.M. 1989. Parrots of the world, 3rd (rev.) ed.London: Blandford.
222 Manual of Parrot Behavior

Hurnik, J.F. 1995. Dictionary of farm animal behavior,2nd ed. Ames: Iowa State University Press.
Pidgeon, R. 1981. Calls of the galah Cacatua rose-icapilla and some comparisons with four otherspecies of Australian Parrots. Emu 81:158–168.
Rowley, I. 1990. Behavioral ecology of the galah,Eolophus roseicapillus: In the wheatbelt of West-
ern Australia. Chipping Norton, NSW: SurreyBeatty.
Snyder, N.F.R. 1987. The parrots of Luquillo: Naturalhistory and conservation of the Puerto Rican parrot.Los Angeles: Western Foundation of VertebrateZoology.
19 / Parrot Vocalization 223

Fear is a critical issue with parrots, especiallywhen trying to establish a relationship of trust ina companion animal situation. Fear is also likelya major cause of stress in companion psittacinebirds. Parrots are, after all, small prey animalsand humans are large predators.
Fear can be caused by a variety of stimuli in thecaptive environment. There is the low-grade feargenerated by a caged bird that is safely out ofreach but watched constantly by an intentlypredatory cat. There is the fear generated when acage-bound parrot is asked to leave the sanctuaryof its cage space; the fear that is generated whena shy parrot is required to step onto the hand of astranger; the fear caused to many psittacine birdsby the proximity of a hyperactive child. Addi-tionally, there is an anxious parrot’s fear when ahuman shows trepidation when reaching for it,and the fear caused by something new beingthrust into a parrot’s personal space.
In other words, fear can be the result of an end-less variety of things, many of which humans donot perceive as frightening. It is undoubtedly dif-ficult for a predator to perceive the world as preyanimals do. Unfortunately, many owners fail tocomprehend the validity of parrots’ fear re-sponses. As a result, instead of being patient andreassuring, they become irritated with frightenedbirds, apparently believing that the birds will “getover” their fear if the human forces the issue suf-ficiently. This bullying tactic is ineffective; in-deed, it can increase the birds’ fear exponentially.Instead, humans must be sensitive to a parrots’fears and, depending on the stimulus, adjust theenvironment to avoid frightening parrots or togradually desensitize the birds to the stimulus.
Additionally, psittacine birds tend to be neo-phobic, which is understandable for prey animals;
parrots undoubtedly survive longer in the wild ifthey approach new things with extreme caution.In the captive situation, this type of fear manifestsas terror of a new toy placed in the cage, theowner’s new hairdo, or a new picture on the wallnear the bird’s cage.
FEAR-BASED AGGRESSION
Fear manifests itself in the basic fight or flightformat, and parrots respond instinctively to a per-ceived threat by attempting to fly away. If flight isimpossible due to clipped wings and/or beingtrapped in a cage, the alternative is to fight, andthe birds will respond aggressively, which usuallymanifests as biting.
Fear-based biting falls into the category of thebest defense is a good offense. This situation willbe worsened if humans respond aggressively tothis behavioral strategy. Aggression begets ag-gression, and trying to get the fear-based biter toback down will only instill more fear and, hence,more aggression. Instead, people need to studythe situation and again look for techniques togradually desensitize the bird to the perceivedfear stimulus, or avoid the situation entirely.
Case Study: The “Suddenly Mean” Grey
Normally sweet and mild tempered, Lily, a 9-month-old African Grey hen (Psittacus erithacuserithacus) abruptly became hostile when herowner’s friends tried to handle her. She becameespecially antagonistic with the owner’s new boy-friend, striking quickly and biting hard.
Lily the Grey was biting from fear-based ag-gression. Frightened by unfamiliar people, sheneeded to be better socialized. If not identifiedand handled properly, a shy parrot like Lily canblossom into a determined fear biter. Lily’s person
225
20Parrots and Fear
Liz Wilson and Andrew U. Luescher

needed to reassure her that she was safe when in-teracting with others.
Introducing her to other people in neutral terri-tory with patience and sensitivity, the ownertaught Lily that new people are fun and interest-ing. Initially, she only expected Lily to step ontothe outsider’s hand politely, and then step imme-diately back onto her trusted caretaker’s hand.Lily’s good manners were then rewarded lavishlywith smiles and praise. Each time Lily did thissuccessfully, she discovered that positive thingshappened when she was compliant with new peo-ple. As a result, Lily gradually learned to enjoy in-teractions with strangers.
“PHOBIC” PARROTS
According to human psychology, a phobia is de-fined as “any unfounded or unreasonable dread orfear.”[1] Indeed, it is quite logical for a wild par-rot to be afraid of a predator, and blatant terror ata human’s approach would be expected in un-tamed psittacine birds. However, this is not thecase with domestically raised companion parrots.When there is no precipitating incident and thebird abruptly acts terrified of people, noises, orshadows, the use of the term “phobic” would ap-pear appropriate.
Dealing with the “phobic” or neurotically terri-fied parrot can be tremendously exasperating.The classic history involves an excitable youngbird that abruptly reacts to specific people as ifthey were deadly predators. This is especially dis-turbing when the primary object of terror is thepreviously loved owner, and the parrot flailsaround its cage, screaming and trying to escapewhen the owner approaches. Particularly distress-ing to the avian veterinarian is the bird thatdemonstrates severe anxiety or phobia as a directresult of a veterinary visit.
People are contacting parrot behavior consult-ants about increasing numbers of “phobic” birds,but this may indicate either an increase in thisphenomenon, increased recognition of the prob-lem, or an increase in the use of these terms.Certainly, there is much discussion on the Internetabout this behavior, and the term “phobic” is in-creasingly bantered about.
Unfortunately, many people perceive the word“phobic” as a synonym for the word “fear,” whichis totally inaccurate. A truly phobic bird is notsimply afraid of new toys or new people. Despite
being domestically bred and hand-raised by peo-ple, such a bird acts like a wild parrot upon theapproach of a deadly predator. A phobic parrot ishyperreactive to direct eye contact, often goinginto what appears to be a full-blown panic attackif people stare.[2] It is hyperreactive to sound,movement, and especially human hands. A phobicparrot has an invisible line around its territory,which identifies its comfort zone. Once invaded,the bird will thrash wildly in a frenzied flight re-sponse. As a result, broken blood feathers arecommon, and serious soft tissue damage can re-sult to keel and wing tips. In extreme cases, par-rots can actually pulverize their metacarpals andphalanges in repetitive frantic efforts to flee (L.Clark, personal communication, 1998).
Nervous psittacine birds are frequently appre-hensive of new things but unruffled during han-dling by trusted humans. These birds are not pho-bics. Confusing the issue further, there appear tobe degrees of phobic behaviors, ranging frommild to severe, with a gray area between a birdthat is simply very frightened and one that is bor-derline phobic.
Generally, aggressive birds are not truly phobic(J. Doss, personal communications, 1997–1998).Aggression and avoidance behaviors are two re-sponses to the same stimulus.[3] An apprehensiveparrot that views itself as being in jeopardy orvulnerable can either flee or attack. Since fear isexcessive in a phobic parrot, a truly phobic parrotwould always try to escape.
Phobic behaviors are more likely in certainspecies, including Poicephalus (i.e., Meyer’s [P.meyeri] and Senegal Parrots [P. senegalus]);small cockatoos like the Rose-breasted (Eolo-phus roseicapillus), Citron-crested (Cacatua sul-phurea citrinocristata), and Triton (C. s. triton);Eclectus Parrots (Eclectus roratus) and AfricanGreys (especially the Congo [Psittacus erithacuserithacus]). It is not surprising that these speciesalso are prone to feather destructive behaviors(FDB).
As a rule, phobic behaviors are seen more fre-quently with juvenile or adolescent parrots.Nevertheless, it is important to make a distinctionbetween an adolescent parrot demonstrating nor-mal pubescent challenges and the phobic. It is ahallmark of psittacine adolescence for young par-rots to balk at compliancy by running away fromhands that request the birds step up, refusing to
226 Manual of Parrot Behavior

exit the cage, or throwing themselves around thecage when people draw near.[4]
Theories abound about the etiology of phobicbehaviors. Infrequently, owners describe a spe-cific incident that appeared to trigger this behav-ior, but this probably is a stressor, not the actualetiology. The potential for phobic behaviors inhigh-strung species is likely to increase if neo-nates are maintained in too much light—for ex-ample, in glass aquariums under neon lighting ina pet store. Indeed, aviculturists note that neonatepsittacine birds actually gain weight faster if keptin the dark.[5] This overexposure to light whenparrots are very young appears to predispose par-rots to fear-based behaviors. Linden suggestseliminating the use of fluorescent lights aroundphobics, due to the increased sensitivity of avianvision.[6] Rearing parrot chicks alone in a re-stricted environment with little handling also con-tributes to exaggerated emotional responses laterin life.[7]
Physical and psychological abuse such as trau-matic capture and restraint techniques such asoverly aggressive toweling can predispose a par-rot to phobic behavior.[8] Indeed, overly aggres-sive toweling is considered a direct cause of “fear-induced behavioral disease.”[9] However,ethologists agree that aggressive handling or“punishment” is not the only reason that parrotsbecome phobic (A. Luescher, J. Oliva-Purdy, L.Seibert, personal communications, 2003). Thereis no history of abuse with most of these cases.
Case Study: The Non-Phobic Phobic
Care must be taken to accurately diagnose pho-bics, since they are handled so differently fromthe more common problem behaviors seen incompanion parrots. The first author (Wilson)worked with a “phobic” Yellow-naped Amazon(Amazona ochrocephala auropalliata) that turnedout to have an idiopathic medical problem thatpredisposed the bird to falling from the hand be-cause it could not grip properly with its feet.Multiple falls taught the bird a direct correlationbetween handling and pain. The result was a dra-matic fear response when people approached.Interestingly enough, the Amazon’s screamingand flailing was eliminated by the use of thedopamine antagonist haloperidol (Haldol™,Henry Schein) (D. Kupersmith, personal commu-nications, 1997–1998).
CLASSICAL CONDITIONING AND THEFEAR RESPONSE
Birds can learn by pairing neutral and unpleasantstimuli. An example would be association of theowner’s hand with restraint or the bad taste of amedication that the owner forced down the bird’sthroat. Another example would be fear shownwhen scolded because of previous punishment fol-lowing scolding. Classical or Pavlovian condition-ing results in the previously neutral stimulus be-coming aversive, that is, evoking a fear response.
Owners of high-strung birds must learn to relaxprior to approaching their animals. Movementsmust be deliberate and calm so they do notheighten the parrot’s anxieties. Hyper ownersoften exacerbate a difficult situation, pushing analready apprehensive bird into a full-blown pho-bic state. This is especially true in dramaticallyfrightening situations, such as natural disastersand veterinary office visits.
There have been multiple episodes in Califor-nia with psittacine birds responding to the horrorof an earthquake by becoming phobic with theowner. There have also been multiple situationswhere sensitive parrots become phobic with theirowners after a traumatic visit to the avian veteri-narian. In these situations, the suspicion is thatthe frightened bird is transferring its fear of thesituation to the owner (S. Blanchard, personalcommunications, 1996). When parrots are trau-matized, it is a natural inclination for the humanto rush over to reassure the bird, hysterically wor-ried about the animal’s safety. As a result, the per-son’s high anxiety terrorizes the bird even more,and its fear is transferred directly to the owner.This unfortunate devolution can cause a parrot tobecome phobic with the person it used to trustabove all.
THE VETERINARY EXAM ROOM:ADVICE TO THE CLIENTS
Many psittacine birds, especially youngsters,react very negatively to visits to the veterinary of-fice. It is important for clients to understand thatthey will make the situation worse if they are dis-tressed by necessary procedures. As a result, theycould not only terrify their animals more but alsodo serious harm to their bond with their parrots.Consequently, many experienced avian veterinar-ians suggest—or even insist—that procedures be
20 / Parrots and Fear 227

done away from the owners. However, care mustbe taken not to give the impression the veterinar-ian has anything to hide, nor should the clinicianautomatically remove the bird from the examroom without discussion with the owner. If clientschoose to be present, they should be counseledagainst petting their birds while under restraint, asserious bites can result. The owner should also nottell the parrot “It’s okay,” since being under re-straint is NOT okay as far as the bird is con-cerned. That phrase should only be used to reas-sure a bird when something is intimidating butnot actually dangerous (such as carrying a largeobject through the room). Using it when a bird isunder restraint risks negating the potential of thisphrase to reassure since it becomes associatedwith an aversive event.
To prevent any connection between ownersand traumatic office visits, seriously upset par-rots should be returned immediately into theircarriers, not into the arms of their humans.Clients can then verbally reassure their birdscalmly, without any physical contact. Oncehome, the owners should open the carrier doorand walk away. Continuing to reassure in a softvoice, they allow the birds to climb out on theirown. Clients should continue to soothe the birdsverbally and observe from a distance. Theyshould not approach the birds until the parrots’body language relaxes.
OPERANT CONDITIONING AND THEFEAR RESPONSE
A fear reaction can become conditioned throughavoidance conditioning or negative reinforcement(see chapter 16). This is especially true if theowner either retreats (if the owner was the threat)or removes the bird from the frightening stimulusand/or shelters the bird. The removal of the threat-ening stimulus is the reinforcement for the fearresponse. Behavior conditioned that way becomesvery reliable and resistant to extinction.
Birds may also learn to exhibit a fear responsein situations where they are not frightened, be-cause they learned that they can manipulate orcontrol a situation. For instance, by acting terri-fied and quivering, running away, or flailingabout, a bird invariably makes humans withdrawrather than allowing the bird to injure itself. Inthis way, a bird may prevent being picked up andput back into the cage. Therefore, exhibiting a
fear response can be a clear manifestation of a re-fusal to interact.
On the other hand, birds rarely learn to exhibita fear response through positive reinforcement.Therefore, giving food treats to a frightened birdis very unlikely to condition fearful behavior andis always appropriate (although if too frightenedthe bird may not take them), and the bird willlearn to associate the frightening (but in factharmless) situation with something pleasant.
It is important to go through a careful diagnos-tic process to determine the extent to which learn-ing contributes to the performance of fearful be-havior. Videotapes can be especially valuablewith these types of cases, as the parrot’s behaviorand body language can be observed when it is inits own environment. It is impossible to judge aparrot’s level of fear in its own environment byobservation in an alien setting such as a veteri-nary exam room. Indeed, fear behaviors canchange tremendously in a parrot’s own environ-ment when a stranger enters the space. For moreinformation regarding the use of videotapes, seechapter 16.
Case Study: The Umbrella That Was “Phobic”about Hands
This situation is exemplified by a consultation thefirst author (Wilson) did a few years ago, with aseven-year-old male Umbrella Cockatoo (Caca-tua alba) that had become “phobic about hands.”The bird was also “terrified” of the owner’s newboyfriend. Upon questioning, it turned out thatthe cockatoo would take food from people’s handsbut ran away when humans asked it to step up oncommand. In this situation, the intelligent birdhad learned that people would not press the issueif the cockatoo acted afraid, thus enabling the birdtotal freedom to do as he pleased.
Working with the bird away from its own terri-tory, the owner used positive reinforcement toconvince the cockatoo that following commandswas a good thing, and the owner’s problems withthe bird were resolved.
Fear reactions can also be enhanced throughlearning if the fear-evoking stimulus is of shortduration or if the bird can escape the stimulus. Inthese cases, from the bird’s point of view, the fearor escape reaction appears “successful” in avoid-ing the expected harm. Fear reactions conditionedin this way are very persistent.
228 Manual of Parrot Behavior

REHABILITATION OF THE PHOBIC ORNEUROTICALLY FRIGHTENED PARROT
Rehabilitation of phobic birds can be a painfullyslow process, but phobics can be helped, with ex-perience and exquisite patience.[10] However,misinterpretation and mishandling of birds withexcessive fear behaviors can reinforce terrifiedand frantic behavior. The use of anxiolytic drugsis often indicated to speed up the process and re-duce the anxiety to a level at which the bird is ca-pable of learning. The owner has to find a way togive the drugs in a non-traumatic way or applica-tion of the drugs itself will increase the level offear.
The first task of rehabilitation is to begin to re-establish a relationship of trust. Blanchard sug-gests the owner bring a chair as close to the cageas possible without frightening the bird and sitthere daily. Reading aloud quietly or singing(even badly) yields a positive reaction from thephobic psittacine bird (J. Doss, personal commu-nications, 1997; S. Blanchard, personal commu-nications, 1997).[11] This procedure uses thephenomenon of habituation: an animal will stopreacting to a neutral stimulus, that is, a stimulusthat does not have any pleasant or aversive conse-quences, through prolonged exposure. No directeye contact should be made but instead oneshould use what Blanchard calls “soft eyes,”where the owners look at the bird very briefly,then turn their eyes and face away (looking awaybecomes a negative reinforcement for being re-laxed; see chapter 16). This procedure often reas-sures the frightened bird.[12] Gradually the chaircan be moved closer and closer to the cage, andthe owner can look for increasingly longer timesat the bird. This procedure is called systematic de-sensitization and will likely be more expedientonce the bird can be rewarded for staying relaxed.
Food treats work well, if the parrot is not toofrightened to take treats from the owner. The par-rot can also be provided with its favorite food inits cage only during these sessions. In this way,the times during which the owner sits by the cagebecome associated with something very pleasant(counterconditioning).
Phobics are often terrified of strong light andare often more comfortable in lower light. An in-sufficient dark period can also increase arousaland reactivity, and owners should be recom-
mended to place the bird in a different cage forthe night in a dark, quiet room. Frightened parrotsare also easily alarmed by sounds, but soft musicoften soothes them more than total silence.
Allowing a phobic bird to regrow its wingfeathers frequently helps in building its self-confidence. Owners should be carefully in-structed on techniques as to how to keep the birdsafe while flighted (J. Doss, personal communica-tions, 1997). In some cases where fearful parrotsare flighted, trimming of the wings may becomenecessary, however (see later).
Rehabilitation should also entail letting thebird choose when and how it wishes interaction.Getting “in the bird’s face” and forcing the issuewill only make things worse. The bird needs toprogress at its own speed and cannot be hurried.
Because rehabilitating a phobic parrot can besuch a slow process, it is recommended thatclients start keeping a daily diary. By describingthe signs of a parrot’s fears in great detail, as wellas recording the miniscule signs of progress, own-ers are better able to see that actual progress ishappening, albeit slowly. These notes can greatlyassist clients later, when frustration mounts due tothe agonizingly protracted rehabilitation process.
Learned fears can be unlearned, although inbirds this may take a very much longer time than,for example, in dogs. Treatment of fear can beachieved through systematic desensitization,counterconditioning, and response substitution.The use of anxiolytic drugs as an adjunct to thebehavior modification technique may prove use-ful or even necessary.
SYSTEMATIC DESENSITIZATION
This is a technique used to reduce or eliminate aresponse (e.g., fear or aggression) to a stimulus.The animal is trained to quiescence. In a parrotthat may mean sitting quietly on a T-stand or theowner’s hand. The stimulus is then introduced at alow intensity (e.g., recording of a noise at low in-tensity, stranger from a distance) and the animalis rewarded for quiet behavior. Once the animalhas habituated to the stimulus at low intensity, theintensity is increased gradually, and the procedurerepeated. The increase in stimulus intensity has tobe so small that no fear response is ever elicited.
Although being in full flight may generally in-crease a bird’s self-confidence, it may be neces-sary to at least temporarily clip a bird in order to
20 / Parrots and Fear 229

desensitize it to a frightening stimulus and pre-vent self-reinforcing precipitous escape reactions.
Systematic desensitization can only be used ifthe stimulus can be identified, reproduced, andits intensity controlled. The handler has to beable to present the stimulus initially at lowenough an intensity that the bird does not react.Furthermore, the naturally occurring stimulushas to be avoided until behavior modification hasbeen completed. If these conditions cannot bemet, drug desensitization can be useful. In thisprocedure, an anxiolytic drug is given at a dosethat allows the bird to function normally andwithout fear. The dose is very gradually reduced,which is the equivalent of increasing the intensityof a threatening stimulus slightly. The bird needsexposure to the stimuli he should be comfort-able with throughout the treatment. If the drugdose is reduced gradually enough, the bird nevershows fear and can eventually be taken off drugsaltogether.
COUNTERCONDITIONING (INCLASSICAL CONDITIONING)
This refers to a procedure to change the meaningof a previously conditioned stimulus. For exam-ple, a previously fear-evoking but harmless stim-ulus such as the sound of the vacuum cleanerwhen always paired with food becomes a condi-tioned stimulus for food. Fearful behavior (theprevious conditioned response to the stimulus) isreplaced by a pleasant emotional response.
RESPONSE SUBSTITUTION (OFTENINCORRECTLY CALLED“COUNTERCONDITIONING”)
This refers to changing the meaning of a discrim-inatory stimulus (i.e., the fear-evoking situation).The aim is to replace undesirable behavior withdesirable behavior in a given situation. This isachieved by controlling the contingencies so thatthe undesirable behavior is no longer successfuland the desirable behavior is reinforced.
The situation in which undesirable behaviorusually occurs becomes a discriminatory stimulusfor desirable behavior. The desirable behaviorshould be incompatible with the undesirable be-havior (e.g., bird stepping up instead of biting).
Response substitution is often used in conjunc-tion with desensitization (e.g., training for quies-cence instead of fearful or aggressive behavior,
while exposing the bird to increasingly intensivestimuli).
TRAINING
Clicker training is another way of increasing thebird’s self-confidence and reducing its fear, espe-cially of the owner. It can be done hands-off froma distance in a non-threatening way. Clicker train-ing allows for consistent and thus predictable,stress-free interaction between bird and owner. Itallows for efficient communication between birdand owner and provides the bird with the abilityto predict and control its environment, thereby in-creasing self-confidence and well-being. Mostbirds really enjoy being trained that way. The onlyproblem might be how to give food treats as re-wards to a frightened parrot.
PREVENTION
The methods used to encourage a psittacine birdto develop its full capability as a companion ani-mal are the same ones used to prevent the devel-opment of high-strung or phobic personalities.The development of these skills begins in baby-hood with the aviculturist and hand feeder andcontinues with the future owners. These tech-niques include
• the nurturing of self-confidence and individualpotential during early development
• normal fledging, then gradual clipping prior tosale (if necessary). This enhances a youngbird’s self-confidence[13]
• abundance feeding and gradual weaning basedon the bird’s development, not the human’sconvenience
• the establishment of clear and consistent be-havioral guidelines in the new home[14]
• the encouragement of self-sufficiency throughindependent play
For more detail on these subjects, see chapter 11.
CONCLUSION
Anxious parrots need to learn that they are safeand can trust the humans around them to keepthem secure. Only through infinite patience canpathologically frightened parrots be rehabilitated,and trying to bully a frightened bird through itsfear will make the situation exponentially worse.When working with cases of neurotically terrified
230 Manual of Parrot Behavior

birds, often the true function of the clinician is tosupport the owners while they make the painstak-ingly slow journey back to a trusting relationshipwith their beloved companion parrots.
REFERENCES
01. American Psychiatric Association. 1994. The diag-nostic and statistical manual of mental disorders,4th ed. Washington, DC: APA, p. 443.
02. Blanchard, S. 1998. “Phobic behavior in compan-ion parrots.” Proc Ann Conf Int Avicult Soc,Orlando, FL.
03. Dodman, N.H. 1998. “Pharmacologic treatment ofaggression on veterinary patients.” In Psychophar-macology of animal behavior disorders, ed. N.H.Dodman and L. Schuster, pp. 41–63. Malden, MS:Blackwell Science.
04. Wilson, L. 1998. “Phobic psittacines: An increas-ing phenomenon?” Proc Ann Conf AAV, pp.125–131.
05. Silva, T. 1991. Psittaculture: The breeding, rearingand management of parrots. Ontario Canada:Silvio Mattacchione & Co, p. 63.
06. Parrot Education and Adoption Center. 2002. “Askthe Experts” roundtable discussion. San Diego, CA.
07. Sheehan, K.L. 2001. The effects of environmentalenrichment and post-natal handling on the devel-opment, emotional reactivity and learning abilityof juvenile nanday conures (Nandayus nenday).MS thesis, Purdue University.
08. Wilson, L. 1998. “Phobic psittacines: An increas-ing phenomenon?” Proc Ann Conf AAV, pp.125–131.
09. Speer, B. 2001. “The clinical consequences of rou-tine grooming procedures.” Proc Ann Conf AAV,pp. 109–115.
10. Wilson, L. 2000. “Behavior problems in pet par-rots.” In Manual of avian medicine, ed. G. Olsenand S. Orosz, pp. 124–147. Philadelphia: Mosby.
11. Leinneweber, T. My ms. duncan, African ark. Un-published.
12. Blanchard, S. 1992. Soft eye, evil eye. Pet bird re-port 92/2 (2):1–5.
13. Cravens, E. 1996. The progressive wing clipmethod. Birdkeeping Naturally.
14. Wilson, L. 1995. “Behavior problems in adoles-cent parrots.” Proc Ann Conf Assoc Av Vet, pp.415–418.
20 / Parrots and Fear 231

INTRODUCTION
Psittacine species have become popular as com-panion animals because they demonstrate manykinds of social behavior that humans find famil-iar and enjoyable. Such endearing traits as beauty,intelligence, and mimicry have led humans toadopt them as members of their family.Recognizing that some of these traits are innateand genetically determined will allow for a newand better understanding of companion psittacinebehavior and enable us to better predict responsesof different species to the conditions of pet birdcare. Developing an appreciation for how andwhy innate behaviors can be triggered by specificactions or conditions will hopefully lead to betterand healthier lives for captive psittacine birds.
Understanding innate behaviors of companionpsittacines requires an appreciation for the intri-cate physiologic and hormonal events that adapt aspecies to an environmental niche. Seasonalevents such as migration and molting have beenobserved, investigated, and found to be initiatedby environmental events and driven by endocrinemechanisms.[1–3] Observations of psittacinespecies in the wild and the experience of avicul-turists and pet bird owners support a similar pat-tern of environmental triggers capable of incitingreproductive behaviors. Recent advances in neu-rophysiology are unraveling the intricate proc-esses of initiation and expression of innate be-haviors in response to environmental stimuli.Although this research has focused specificallyon the mechanisms of context-driven song, it isallowing for a completely novel view of how in-nate potential can lead to the expression of spe-cific behaviors in certain situations.[4–6]
Reproductive behaviors observed in the wild,such as pair bonding, courtship regurgitation,cavity seeking, nest building, territorial aggres-sion, and copulation, are often displayed in ahome setting, though with human “flocks.” Fe-males of some species lay large numbers of eggsover extended periods of time to a point of com-plete physical collapse and failure. The reproduc-tive drive can also lead to behaviors that renderthe bird difficult or impossible to live with. Thesebehaviors often include incessant screaming, sud-den aggression toward favored (or not favored)humans, and destructive attempts to excavatenests in closets, couches, and drawers. As manyspecies are represented in the pet trade, there ex-ists extreme variability as to when behaviors willstart, which will be displayed, and how long theymay continue.[7]
In the wild, the expression of these largely re-productive behaviors would be regulated by envi-ronmental conditions, pair bond formation, andthe social hierarchy of the flock. Environmentalcues such as photoperiod, temperature, rainfall,available food supply, the presence of nesting ma-terial, or the presence of a mate can stimulate re-productive activity in birds.[3, 8, 9] Pair bondformation is enhanced with regurgitation, copula-tion, nest-site inspection, feeding, and mutualpreening.[10] When favorable environmentalconditions are present, opportunistic breeders canbecome active. Other species may not even breedon a yearly basis. The hypothalamic-pituitary-gonadal (HPG) axis, responsible for controllingreproductive development and subsequent repro-ductive behaviors, is triggered when environmen-tal conditions are appropriate for the species.
233
21Problem Sexual Behaviors
of Companion Parrots
Fern Van Sant

Behaviors that enhance the pair bond are alsorecognized as triggering cues for reproductive ac-tivity.[10] In other words, the hormones flowwhen triggered by favorable conditions. In thecase of companion psittacine birds, under con-strained circumstances, many of the conditions ofpet bird care are capable of triggering reproduc-tive behaviors.
Unfortunately, as companion birds, parrots arenot subjected to the normal limiting factors oftheir native environment and behavioral interac-tions with a flock. In fact, pair bonded ownersoften indulge their birds, providing them withseveral of the environmental and behavioral cuesthat can trigger the HPG axis. Owners often feeda varied, nutrient-rich diet daily (with possible in-creases in fat and protein), provide a nest (cage)and nesting material (newspaper), and, inadver-tently, provide themselves as a perceived mate.They sometimes feed the bird warm, soft food,much as might occur during courtship feeding.Owners encourage the pair bond formation by“preening” the birds with petting (often includingthe tail and back areas) and allowing excessivepreening to their person. In short, the owners in-advertently encourage reproductive behavior overa long period. The abundance of these environ-mental cues and pair bond activities in a homesetting may encourage the early development ofboth clinical and behavioral problems, stemmingfrom the reproductive drive.
Attempting to explain the sexual behaviors ofcompanion psittacine birds without understand-ing their biology is impossible. With continuedinvestigation, we are likely to find that the biolog-ical niche that supports the existence of a speciesalso directs the timing of sexual behavior and re-production. That said, it is possible to piece to-gether what we have observed throughout the last30 years and overlay that information with gen-eral reproductive biology, avian science, andpsittacine biology. We will then begin to interpretbehaviors carefully.
This discussion will identify and address sig-nificant clinical and behavioral problems thatappear to develop over time because of chronichormonal stress from the abundance of environ-mental cues and pair bond enhancing behaviorsprovided in many homes. Clinical conditions in-clude those obviously related to reproductivephysiology such as chronic egg production, egg
peritonitis, and yolk embolis.[11, 12] An effortwill be made to explain many conditions offeather loss and feather-destructive behaviors thatseem to be the result of abnormal, long-term, hor-monal stress.[13] Other clinical conditions in-cluding degenerative changes of pelvic and ab-dominal muscle that may result in herniation orprolapse are typical of chronic hormonal stress oroverproduction. Chronic egg laying may draincalcium stores, predisposing the hen to dystociaand osteoporosis.[10] Dramatic and abnormalconditions of bone are frequently seen in femalebirds demonstrating reproductive behaviors overprotracted periods of time.[10]
By carefully examining the many complicatedinteractions between psittacine birds and theirvarious natural environments, we will be betterable to interpret social and sexual behaviors ofcaptive companion psittacine birds. This im-proved understanding will lead to more effectivemedical intervention when necessary and hope-fully improved preventative care.
PSITTACINE BEHAVIORAL BIOLOGY
Psittacine species are thought to derive from anancient class of birds. It is considered likely thatthese birds have a more than 30-million-year his-tory, with their roots in the ancient landmass ofGondwanaland. As landmasses slowly drifted andcontinents formed, the evolutionary processesthat drove speciation slowly produced the threefamilies and 332 species of the order Psittaci-formes that we now know.[14, 15]
Of the 332 species known today, there are cleardistinctions between psittacine birds based on ge-ographical distribution. Neotropical species(from Central, South, and rarely North America)account for nearly two-thirds of all psittacinespecies. At first glance, they appear to be a di-verse group of birds, but Neotropical speciesshare many physical and behavioral traits and rep-resent only one subfamily (Psittacinae). In con-trast are the 109 species from Australia, NewZealand, and the Philippines that represent allthree subfamilies of psittacine species (Loriinae,Cacatuinae, and Psittacinae). This incredible di-versity of species is thought to result from the rel-atively long isolation of these geographic areas aswell as the absence of pressure from mammals.Only 34 species are found in Africa, India, andSoutheast Asia.[14–16]
234 Manual of Parrot Behavior

If any single evolutionary tendency of thesebirds were to be singled out, it would have to betheir flexibility to adapt to an impressive range ofhabitats. In order to understand the needs and be-haviors of companion parrots, we must acknowl-edge the individual adaptations that species havemade to survive in their distinctive environments.Viewing psittacine species from the perspectiveof evolution offers insights into many of theunique adaptations and behaviors that we recog-nize as characteristic of parrots. Most psittacinespecies display an uncanny ability to learn, so-cialize, and adapt. Some of these behaviors havebeen studied extensively.[17–19] Other physicaland behavioral traits such as feather color, vocal-izations, food preferences, learning abilities, pho-toperiod responsiveness, and breeding behaviormay reflect adaptations to very different climatesand environments over millennia. Once devel-oped, an evolutionary perspective may offer abetter understanding of many attributes and be-haviors that owners and behaviorists have foundpuzzling or difficult.
Innate Behaviors
Innate, or “hardwired,” behaviors are inherited,genetically driven, and species-specific like songsand nest design. These are observed in individu-als raised independently of conspecifics. Learnedbehaviors are those acquired by impressionableindividuals. These behaviors include imprinting,flying, food identification, and navigation. As amember of a flock, each individual bird mustlearn a complex set of vocalizations and body sig-nals to maintain cohesiveness. Individuals in aflock manifest an innate ability to conform.[15,20] Innate and learned behaviors support thegroup dynamic and facilitate social order.Although the mechanisms of expression of innateand learned behaviors at a molecular level remainpoorly defined, ongoing research by Erich Jarvisat Duke University is identifying the roles of spe-cific parts of the brain that direct certain innatebehaviors under a set of external conditions.[4–6]He is also unraveling the ways that complex be-haviors, both innate and learned, are triggered andexpressed in an individual.
Reproductive Behaviors
Many of the commonly observed and commonlymisunderstood behaviors of companion psittacine
species are reproductive in nature and stem fromactivation of the HPG axis, as well as from pairbond formation.[10] These behaviors include cav-ity seeking (the drive to find a small dark hide-away), nest preparation (shredding of paper orother bedding), bonding (an innate drive to dis-play an affinity for a single individual), sexual re-gurgitation (regurgitation of food to a bonded birdor human), and even copulation (birds of eithersex displaying receptive postures or actual copu-lation). The seasonally aggressive behaviors ofmany male amazon parrots are another exampleof innate sexual behaviors demonstrated season-ally and apparently triggered by environmentalconditions. Some clinical presentations, such ascloacal prolapses in male cockatoos, which seemto be linked to abnormal eating habits, may bemade worse by chronic reproductive stimulation(F. Van Sant, personal observation, January 2004).
Reproductive behaviors may best be under-stood by defining the events that trigger them andthe general context of the biology in which theyoccur. In short, what are the environmental cuesand other triggers that stimulate reproductive be-havior in a particular species? And what are theirbiological effects? If we can appreciate the influ-ence of environment and pair bond formation inwild species, we will better understand the quirkyreproductive behaviors and personalities of com-panion birds. Further, we may be able to use thisinformation to help modify problem reproductivebehaviors of companion psittacines.
Endocrine Regulation of ReproductiveDevelopment and Behavior
The set of innate and learned behaviors that drivereproduction and the physiologic mechanismsthat underlie the process are complex. Reduced tosimplest terms, common themes emerge. In birds,specific, and usually age-related, changes signalreproductive competency. The physiologic regula-tion of these events is controlled by the hormonalcenters of the brain and the ovaries through theHPG axis, and it is these endocrine pathways thatregulate reproduction.[10] Similar mechanisms inthe male result in the production of testosterone,the hormone that directs the reproductive drive ofmales.[21] The HPG, responsible for controllingreproductive development and expression of be-haviors, is triggered by various environmentalconditions appropriate for the species.[3, 8, 9]
21 / Problem Sexual Behaviors of Companion Parrots 235

Pair bond enhancing behaviors can trigger repro-ductive activity as well.[10]
As in mammals, the HPG axis in birds regu-lates reproductive events, with several differencesin birds. Because the capacity for flight demandsthat weight be minimized, females have only oneactive ovary. Birds further minimize their weightby maintaining inactive and atrophied gonadsduring most of the year. Only when conditions areappropriate for breeding will the HPG axis stim-ulate activation of the ovary and testes.[3, 22]During breeding season, these organs may weighmany times more than the inactive gland.[22]This adaptation necessitates a mechanism for ef-fectively signaling the body that breeding seasonhas arrived. For example, in temperate climatezones, day length is a common trigger.[2]
Species Variation
Matching reproductive activity to appropriate en-vironmental conditions benefits the survival ofthe species. Environmental triggers help ensurethat food and other resources are available tomake reproductive activity successful. Alongthese same lines, when conditions are such thatfood and water are scarce, the biologic behaviorsthat promote and sustain reproduction are nottriggered.
In some species, the initiating events are mini-mal and the response is swift. In others, there is asnowball effect that requires a complex syn-chrony of events that build progressively and cu-mulatively to nesting and oviposition. Cockatiels,Budgerigars, and other small psittacines native toharsh, water-limited environments respondswiftly to rainfall.[14, 15] The hormonal cascadesthat drive reproduction are initiated quickly by thehypothalamus. Pairing, courtship, nesting, and ul-timately oviposition are driven rapidly by theHPG axis. Gestation is short and the young ma-ture to independence quickly.
In contrast, many species of Neotropical par-rots, particularly macaws and amazon parrots,have a low rate of reproduction.[23, 24] Researchconducted by Charles Munn demonstrates that themacaws of Peru maintain stable pair bonds. Forthese species food is rarely a limiting factor, butnest cavity availability appears to be the ultimatelimiting factor. When food is present, then nestcavity availability becomes the most importantfactor in stimulating reproductive behavior.
Finding, excavating, and preparing a cavity ele-vates sexual hormones in these birds, preparingbirds for copulation and oviposition.[24, 25]
Studies by Millam with another Neotropicalspecies, Orange-winged Amazon Parrots (Ama-zona amazonica), have shown that testosteroneproduction builds and peaks as cavities are exca-vated.[26] The increased levels of testosteronemay drive the sexual regurgitation (courtshipfeeding) that acts to nourish the female and pro-vides the nutrients necessary to support ovulation.
ENVIRONMENTAL CUES FORREGULATION OF SEXUAL BEHAVIORAND MIGRATION
Once an understanding of parrots’ natural re-sponse to environmental cues is developed, it be-comes possible to look at many of the “quirky”behaviors they may exhibit in captivity. For exam-ple, companion psittacine birds demonstrate thesame sexual behaviors seen in their wild counter-parts but necessarily adapt them to the conditionsfound in the typical home. Companion parrotswill shred their bedding and cage liners, nest infood dishes, woo their images in mirrors, and ex-cavate cavities under furniture and in drawers.Companion birds will court and then copulatewith toys. Females of certain species can producean alarming number of eggs over months or evenyears.
It is imperative that the role of “normal” devel-opment be considered. As social species, parents,other adults, conspecifics in the flock, and eventhe competition of nest mates provide many be-havioral mechanisms to moderate drives.
Innate drives are also tempered by environ-mental factors. In arid areas, the limiting factorof water exerts a powerful influence on the abil-ity to survive. In moderate tropical climates, en-vironmental factors can seasonally affect foodsupplies and habitat. In the stable home environ-ment with regular and plentiful food supplies, itthen becomes a distinct possibility that innatedrives will go unchecked and often escalate. Inthe absence of limiting factors and plentiful at-tention, the home setting may contribute to be-havioral or clinical problems. When these condi-tions are coupled with a lifestyle that is devoid ofintense physical activity in animals that haveevolved specifically to fly, there is little wonderthat problems arise.
236 Manual of Parrot Behavior

Seasonal Breeding
In tropical and subtropical locations, avianspecies have adopted several triggers to signalbreeding season such as cessation or arrival ofseasonal rains.[15, 16] Defining and understand-ing different triggers to reproductive behavior andtheir effect on different species is necessary be-fore we can attempt to modify the behavior ofcompanion psittacines. Equally important is anunderstanding of how the process needs to beself-limiting and turn itself off.
With rare exceptions, psittacine species are di-urnal. Although these species, distributed amongfive continents, have established themselves in awide array of environmental niches, most speciescan be relied upon to demonstrate regular patternsof moving and feeding through specific areas.The pineal gland contains photoreceptors thatsense light independently and through visualpathways, and it is important in maintaining thecircadian rhythms of psittacines.[27] It appearsthat the pineal gland may also play an importantrole in mediating seasonal breeding.[27]
Molting
The process of molting involves an orderly re-placement of an individual’s feathers and usuallyoccurs as an annual event. Molting typically in-volves cessation of breeding activities and regres-sion of the gonads. Although the mechanism ofmolt is poorly defined, it is thought to be medi-ated by the pituitary, thyroid, and adrenal glands.Molt can be forced in several species by the ad-ministration of thyroxine, prolactin, and proges-terone. Sudden environmental events such as de-creased day length and restricted food and watercan also precipitate a molt.[2, 16]
Seasonal Migration
Seasonal migration to follow food supply is seenin a few parrot species. The Great Green Macaw(Ara amigua) seasonally migrates from coastallowlands in Costa Rica to higher elevation forestsin Nicaragua.[13] Two species of grass parakeetsmigrate over the Bass Straits of Australia across120 miles of open water to Tasmania to breed eachyear.[15] The Patagonian Conure (Cyanoliseuspatagonus) migrates during harsh winters towarmer areas of Uruguay. Species living at highaltitudes in the Andes migrate vertically to lower
altitudes during winter.[28] Thick-billed Parrots(Rhynchopsitta pachyrhyncha-pachyrhyncha) usedto migrate from northwestern Mexico to southernArizona where they found abundant piñon pinenuts.
Light changes may trigger seasonal migra-tions. Fluctuating seasonal photoperiods, subtlechanges in light wavelengths, or temperature fluc-tuations seem to initiate hormonal events directingmigration. Corticosterone and prolactin, triggeredby changes in the pituitary gland, are integral inthe hormonal regulation of migration.[2]
NORMAL SEXUAL BEHAVIORS OF WILDPSITTACINE SPECIES
An understanding of normal sexual behaviors ofwild psittacine birds is essential if we are to un-derstand, and in turn prevent, the development ofproblematic sexual behaviors in companion par-rots. Part of the solution is to recognize reproduc-tive behaviors and environmental triggers in thewild, then to recognize those behaviors as ex-pressed in a home setting.
Formation of Pair Bonds
Most psittacine birds are monomorphic and manyform stable pair bonds. The pair bond offers a dis-tinct advantage in undertaking the all-consumingcommitment to hatching and rearing altricialyoung. Birds using this reproductive strategy aretypically ready to breed without the need for elab-orate courtship rituals. When environmental cuessuch as climate and available food supplies trig-ger the HPG axis, the birds are primed for repro-ductive behavior and courtship rituals can ensue,in greater or lesser degree. Although stable pairbonds are common among all avian species, theseadaptations are well suited to the tropical and sub-tropical distribution of psittacine birds and reflectthe huge biologic investment required to raise al-tricial young.[1, 16]
Vocalization
Specific vocalizations signaling sexual readinesshave been observed in many species of parrots.Cockatiel males court females with shrill whistlesand ready females answer with incessant chatter.Orange-chinned Parakeet (Brotogeris jugularis)pairs exhibit a back-and-forth chatter so well co-ordinated, it sounds like it is coming from a sin-gle bird.[15] Some cockatoos begin their court-
21 / Problem Sexual Behaviors of Companion Parrots 237

ship with loud vocalizations that grow progres-sively quieter as the pair mates, the female layseggs, and the pair begins the work of incuba-tion.[28] Budgerigar males woo hens with a war-ble that continues in four-minute bursts that maycontinue for hours. In Budgerigars, specific areasof the brain have been identified as similar to thewell-studied song nuclei of passerines. The war-ble of the male Budgerigar seems to be a learnedtestosterone-driven behavior.[29]
Courting Behavior
Because many psittacine birds maintain stable pairbonds, courting behaviors usually involve a varietyof simple moves like hopping, bowing, strutting, ortail wagging.[14, 15] Excited or aroused psittacinebirds may exhibit pinning of the pupil and blush-ing. Macaws and Palm Cockatoos (Proboscigeraterrimus) can display a rush of color in their facialskin. Many Australian and Indonesian birds haveimpressive crests that can be used in very demon-strative ways. Head bowing, an invitation for mu-tual preening, is often intensified during breedingand courtship and seems to convey sexual signals.Physical contact intensifies dramatically duringbreeding season. Although social preening is com-mon among parrots, there are many nomadicspecies that reserve physical contact for breedingand fighting. For species that usually maintain adiscrete critical distance between individuals, thephysical contact of courtship is a powerful mecha-nism to synchronize males and females for suc-cessful breeding.[15, 16, 28] In a home setting,psittacine owners may inadvertently excite theirbirds through excessive physical contact, therebyencouraging reproductive behavior.
Courtship Feeding
Some species of parrots, particularly macaws andamazon parrots, will use courtship feeding, moti-vating the hormonal cascades that culminate in egglaying by both stimulating hormonal changes andproviding the caloric abundance that contributes toegg production.[15, 24] It is notable that manypsittacine owners feed soft, warm food to their par-rots, inadvertently mimicking courtship feedingand encouraging reproductive behaviors.
Nesting
There is amazing variability among the 332species of parrots as to how they claim and pos-
sess a nest site, and in their willingness to defendit. In the Neotropics, nest cavities are a limitingfactor to breeding. Because the cavity plays sucha key role in successful breeding of Neotropicalspecies, it is not surprising that pairs of birds willincorporate a great deal of care and effort to pre-pare it. Large cockatoos, specifically BlackPalms, have been observed drumming with sticksin displays that appear to claim territory and woomates.[14, 15] In a home setting, birds may seekcavities, such as under a chair or in a closet, orthey may use their cage as a nest.
Work done in Peru by Charles Munn identifiednest cavities as the primary limiting condition forreproduction in Scarlet (Ara macao) and Green-winged Macaws (Ara chloroptera).[24, 25] Whenartificial cavities were provided high in thecanopy, pairs of macaws quickly set up house-keeping. Biologists have noted that severalspecies of macaws and amazons have establishedstable shared ownership of nest sites.[24]
A few species of parrots (most notably MonkParakeets [Myopsitta monachus]) are colonybreeders that construct elaborate nests. EclectusParrots have been observed breeding in colonies.Several males may work together to support anesting female.[28, 30]
Several species of lovebirds (Agapornisspecies) transport leaf litter, bark, and twigs forelaborate nest construction. Peach-faced Love-birds (Agapornis roseicollis) tuck long pieces ofnest material across their back secured by thefeathers of the lower back. Lovebirds with whiteeye rings carry nest material in their mouths.Other species carry small pieces tucked undertheir body feathers.[15] William C. Dilger inves-tigated these behaviors three decades ago, findingthat both the method of carrying material and nestdesign had a clear genetic basis.[31] In a homesetting, birds may shred newspaper as a substitutenest material.
Copulation
Clear differences are observed in the physical pos-ture assumed during copulation in Neotropicaland old world birds. To achieve the cloacal contactof copulation, Neotropical males will mount thefemale with one foot holding on to a perch. In con-trast, the males of old world species will mountthe back of the hen with both feet. Copulation usu-ally lasts about a minute. Lovebirds have been ob-
238 Manual of Parrot Behavior

served copulating for up to six minutes. Toachieve internal fertilization, breeding must betimed to imminent ovulation. Oviposition, or lay-ing, usually follows ovulation by roughly 12–24hours. Generally a clutch of determinate size willbe laid before incubation begins.[14, 15, 32]
Laying of Eggs
Most large parrots lay small clutches of one tothree eggs. Parrot eggs are relatively small and areincubated for a fairly long period of time. The veryaltricial young are tiny and helpless at hatch. Thevalue of a strong, well-protected nest is untoldconsidering the substantial investment in time andenergy that large psittacine birds devote to theiroffspring. Female macaws assume the duty of in-cubation while the male ensures that she is fed andprotected. Cockatoos share incubation duties.Many smaller nomadic parrots use a differentstrategy, producing large clutches with relativelyshort incubation times. Young develop quickly,fledge, and mature to independence.[14, 15, 28]
Thermo-regulation is crucial in ensuring suc-cessful incubation.[33] In hot, humid, tropicalareas, it can be assumed that adaptive physiologicmechanisms maintain stable body temperatures ina brooding parent. Thermo-regulation of avianspecies has been investigated and seems to becontrolled by the dorsal hypothalamus and pe-ripheral receptors. The propatagium of the tho-racic limb has long been considered to be a prin-cipal site of thermo-regulation in flight and atrest. Other sites likely to play a role in criticalthermo-regulation are legs, abdomen, and possi-bly feet.[26, 33]
CLINICAL PRESENTATIONS
The clinical implications of feather picking andother feather-destructive behaviors have been thefocus of a great deal of attention by avian veteri-narians, parrot behaviorists, and bird owners. Bydefault, many of these conditions are dismissed as“normal” because systemic pathology cannot bediagnosed.[34] There is no single predisposingcause but rather a very complex mesh of genetic,environmental, and medical factors. Several clin-ical presentations of feather loss seem to regularlyoccur in concert with hormonally driven behav-iors. In many instances, reversal of hormonaldrives will be followed by regrowth of healthyskin and feathers. In principle, it follows that
chronic reproductive behaviors are hormonallydriven. In nature, parrots are “turned on” to breedby a set of well-defined environmental factors in-cluding seasonal changes, abundant food, avail-able mates, and available cavities. The conditionsof abundant food, bonded owners, cages that maytrigger cavity-nesting behavior, and considerablephysical contact seem to initiate breeding behav-iors that may subject the bird to chronic reproduc-tive stimulation. Without the naturally occurringenvironmental pressure of dwindling food sup-plies, changing environmental conditions, andcompetition for resources that limit breeding be-havior in wild populations, breeding behaviorsand hormonal activation persist unchecked. Whenthese behaviors continue in an unrelenting fash-ion, the physiologic occurrences timed to occurduring non-breeding season, such as molting, donot occur.
Molting follows breeding in the lives of mostbirds. Because both events are physically taxing,timing is important. Large psittacine birds havebeen observed to have a one- or two-year moltcycle synchronized with gonadal cycles. Largeparrots reproducing year-round in avicultural sit-uations have been noted to not molt in the samemanner as non-breeding or free living con-specifics.[35] Delayed molts and failure to moltare common complaints of companion parrotowners. Also common are sets of clinical signsthat have been observed to occur in companionparrots showing chronic breeding behavior.
Neotropical Parrots
Mature bonded female amazon parrots commonlydemonstrate chronic breeding behaviors such asposturing, shredding, and cavity seeking. Birdswith these signs are typically well nourished andoften obese. Feather loss over the trunk, legs,back, and patagium are common. Many seempruritic. New growth is sparse and often removedby the bird as pinfeathers erupt. Environmentalchanges are less rewarding in these birds as it ap-pears that the owner has become the bonded mateand the cage has become the perceived nest.Endoscopic exams of these birds usually reveal amature but abnormal ovary.[36] Lacking is theusual cascade of developing oocytes. Instead, asolitary, mature follicle is usually found. Therapywith leuprolide acetate (Lupron®) by itself isoften insufficient in restoring the normal cascade.
21 / Problem Sexual Behaviors of Companion Parrots 239

Lupron® can shut down the reproductive cyclebriefly, but if reproductive triggers remain un-changed, clinical symptoms can reappear.Successful therapy can be achieved with weightloss. Individual birds seem to have a weightthreshold under which chronic breeding behav-iors cease. This clinical recommendation is sup-ported by observations of wild amazon parrotsthat describe a weight increase triggered by foodavailability and male regurgitation and feedingpreceding ovulation.[15, 26]
Female macaws may demonstrate cavity seek-ing and protracted egg laying. In this context,Lupron® is a very useful therapeutic tool, provid-ing a means to diminish both. A less common butvery significant presentation in bonded femalemacaws is an inflammatory process of the medialleg. These birds exhibit agitation and foot stomp-ing. Close exam of the medial vascular pattern ofthe legs will usually reveal dilation and inflamma-tion of the vessels. The legs are often hot. Thiscondition may develop into a crusty dermatitis.Therapeutic Lupron® coupled with HCG and anon-steroidal anti-inflammatory are helpful.Discontinuing any warm food is imperative as thatpractice seems very capable of mimicking materegurgitation and inciting hormonal stimulation.
Male amazon parrots have been observed todemonstrate seasonal aggression, territorial dis-plays, and recurring focal inflammatory lesionsof the feet. These conditions can be successfullytreated with Lupron® and HCG. Cool water bathsto the legs are helpful.
Examining the cooling mechanisms of parrotsmay provide insights into clinical presentations ofinflamed legs and feet. Thermo-regulation is acritical component of successful incubation.Hormonal regulation of vascular channels is welldefined in mammalian and avian physiology andcould be assumed to be a critical factor here.
African Parrots
African Grey Parrots frequently demonstratesigns of breeding behavior. These can occur year-round, especially when there is a bonded owner.Warm food and abundant food will often triggerregurgitation behaviors. Commonly coupled withthese behaviors is feather picking of the trunk andlegs. Control of calories, especially those ob-tained from rich simple carbohydrates like pastaand cookies, is very helpful. Of serious conse-
quence is the tendency of African Grey Parrots todevelop severe non-responsive dermatitis of thepatagium. When closely monitored, these condi-tions appear to start with the loss of down andcontours of the wing web. Superficial vesselsusually appear dilated. If allowed to continue,secondary opportunistic yeast and bacterial infec-tions may develop. Control of chronic dermatitiscan be difficult, but early intervention and envi-ronmental and behavioral corrections that focuson decreasing hormonal stimulation are usuallysuccessful.
Considering the critical distance observed be-tween African Grey Parrots in the wild and inaviary situations, the role of physical affectiondemonstrated by frequent petting and body con-tact with an owner should be considered to be apowerful predisposing factor.
Indonesian/Australian Parrots
Three species of companion cockatoos com-monly demonstrate clinically significant changesthat may relate to hormonally driven behaviors.Umbrella Cockatoos, Moluccan Cockatoos(Cacatua moluccensis), and Goffin’s Cockatoos(Cacatua goffini) are frequently presented forfeather loss, feather picking, and dermatitis.Female cockatoos are commonly handled andcuddled by adoring owners. Lavishing physicalattention to the bird’s head, crest, and trunk usu-ally elicits posturing and often orgasmic-likeshuddering. In many cases, these birds exhibitthese behaviors for years. Clinical impressions ofthese birds seem to reveal a progressive and seri-ous pattern of degenerative changes includingdermatitis, loss of cloacal tone, and anemia. Theavailability of an estrogen panel assay at theUniversity of Tennessee Clinical EndocrinologyService made possible quantification of hor-monal levels in these species. TherapeuticLupron®, HCG, and a concerted change in thebirds’ environment result in dramatic recovery.
It now seems very likely that the role of intensephysical contact is capable of eliciting hormonalresponses, starting early and continuing progres-sively for years. The perception that “no amountof attention is ever enough for the social cocka-too” has been difficult to overcome. The missingpiece of information is the naturally occurring en-vironmental conditions that serve to make thesebehaviors seasonal rather than constant. In the
240 Manual of Parrot Behavior

case of the Goffin’s Cockatoo, seasonal shifts inthe Tanimbar Islands are dramatic. As the islandsare located in the geographical rain-shadow ofAustralia, there is a distinct periodicity to rainfall.Nine months of the year bring regular rainfall andabundant water. For three months of the year rain-fall amounts drop precipitously. Inherent in thispattern is an environmental shift that is not con-ducive to reproduction and challenging for sur-vival. For birds as companions, these conditions,or seasonal stresses, never occur and breeding be-haviors continue in an unrelenting fashion.
Male Moluccan Cockatoos, particularly thosethat have endured sedentary life and seed diets forten or more years, have a high incidence of self-mutilation over the sternum. Considered a behav-ioral problem by many or the result of abjectboredom, perhaps this should instead be investi-gated as a physiologic event. Building on the ex-perience of successful clinical intervention ofthese cases where lifestyle changes including dietand exercise have been used in concert withLupron® and symptomatic medicine, it might bepossible to hypothesize underlying factors involv-ing vascular changes, tissue perfusion dependenton physical conditioning, and even chronic hor-monal stress.
THERAPEUTIC REMEDIES
Avian veterinary medicine has become adept atdeveloping effective therapeutic modalities forthe most common manifestations of hormonaldysfunction. Experienced clinicians routinelyhandle medical emergencies related to ovulation.Protracted egg laying, as is commonly exhibitedin Budgerigars, Cockatiels, and Umbrella Cock-atoos, is routinely treated with Lupron®, agonadotropin-releasing hormone (GnRH) ago-nist, that acts by down-regulating pituitary GnRHreceptors.[7, 37] These therapies, though effectivein the short term, may become inadequate as along-term solution. When faced with the recur-rent nature of these problems, many companionpsittacine owners become impatient with the needfor return visits and expensive injections.
Based on Millam’s research, recommendationsto restore Cockatiels to a short-day photoperiodas a remedy to chronic egg laying became com-mon.[38] Other management-based recommen-dations, such as removal of the nest box andseparation from the male, also became common-
place. Many veterinarians counseled clients aboutthe role of physical contact in hormonal stimula-tion. But explanations about the mechanics ofhow to prevent recurrence are often poorly re-ceived and viewed as undesirable or impossible.Reinforcing these recommendations with inter-esting information about the bird’s origins andadaptations has proven to be an extremely usefultool.[37]
Adjust Photoperiod
If there is one single positive change that compan-ion bird owners can make, it is returning the birdto a regularly recurring photoperiod. Whether inthe wild or in captivity, most birds demonstrate aremarkable periodicity to their days. Restorationof a regular recurring day and night cycle usuallyresults in a happier and healthier companion bird.Ideally the photoperiod would begin at dawn whenmost birds, covered or not, sense the new day andbegin to stir. As most birds are from equatorial andsubequatorial latitudes where day length isroughly 12 hours year-round, establishing a rou-tine that follows a 12-hour day with a 12-hournight is ideal. Birds have in their brains a finelytuned, light sensitive pineal gland. This gland islikely the mechanism by which birds set their cir-cadian rhythm. There is some evidence to supportthe theory that seasonal shifts are sensed by therate of change of day length (like those that occurin spring and fall) rather than just keying off of asingle day length. Many owners initially anticipatea hardship or a loss of interactive time but insteadfind that the bird adapts within days to the newroutine and quickly demonstrates that the changeis a benefit.
Control Shredding
One of the easiest remedies to derail reproductivedrives is also one of the most powerful. Shreddingof paper, cardboard, or other bedding materialseems to mimic the intrinsic behaviors of nestpreparation. Typically regarded as benign, playfulactivity, shredding instead seems to promote re-productive activity, boosting hormone levels.[9,15, 26] As most cages are equipped with gratesthat prevent access to the cage floor, this behavioris usually easy to control. In cases where a grateis not provided, all liners can be removed and thecage tray simply rinsed daily. In many cases, thisbehavior may be one of the earliest warning signs
21 / Problem Sexual Behaviors of Companion Parrots 241

of reproductive activity. Preventing access toshredable substrate may quickly defuse reproduc-tive activity.
Curtail Cavity Seeking
As reproductive drives escalate, many companionbirds begin to roam and explore, seeking a cavity.The perceived cavity may be a closet, a drawer, ora box. Many birds have attempted to set up house-keeping under a chair or a couch. Owners havefound chair stuffing excavated and carpets rippedup by companion parrots driven to find a suitablenest site. Cavity seeking should be viewed as a se-rious escalation of hormonally driven behavior.Many Neotropical species will become very terri-torial and fiercely guard their homestead. Cavityseeking is often a sign of imminent ovulation inthe female. Millam investigated the importance ofthis hormonal drive in the cascade of physiologicchanges that lead to oviposition. His studiesdemonstrated that testosterone levels crescendo totheir peak levels in male Orange-winged Ama-zons during cavity exploration.[26] This informa-tion dovetails perfectly with the observed impor-tance of cavity availability to many Neotropicalspecies, including amazon parrots and macaws.Curtailing this behavior by not allowing the birdto wander is a simple and powerful solution.Female birds that are permitted or encouraged toestablish ownership of a “cavity” will often begina long stint of unrelenting egg production. Oftenthese female birds will lay several lifetimes worthof eggs and become quite stressed and—eventu-ally—quite ill from the physiologic demands ofegg production and incubation.
Watch Physical Contact
The role of physical contact, usually in the formof affectionate petting, can become extremely im-portant in inciting and fueling hormonal behav-iors. Physical contact seems most powerful incockatoos, Cockatiels, and Budgies. Cockatoos infact have been recommended and sold as com-panion birds that will thrive on attention andprobably suffer derangement without it. Manybirds train their owners early on in the best tech-niques to cuddle and adore them. These birds de-light in having their crests stroked and will oftenelicit attention by lowering their head. Many fe-male cockatoos demonstrate orgasmic pantingand shaking while caressed by owners. The sig-
nificance of this behavior is often missed by own-ers, who may interpret it as anything from aseizure to a sinus infection. Owners who havebeen warned about the risks of behavioral prob-lems and feather picking that may result from alack of attention are often devastated to see thesebehaviors develop in their companion birds.
The role of physical contact in most adult birdsis reserved for courting and breeding behaviors. Itis not surprising to find that species that cravephysical attention as companion birds are thesame ones that incorporate more physical contactinto their courtship rituals. Indeed, the crest ofcockatoos has been conjectured to be an impor-tant lure. The bowing and head-lowering behav-iors commonly demonstrated by cockatoos arelikely signals between the male and female. Thesesignals probably serve to synchronize the behav-iors of the female and male to time nesting, cop-ulation, and ovulation. The remedy, of course, isto decrease physical contact with susceptiblebirds. As these patterns are often very difficult forpet owners to break, efforts must be made toframe the change in understandable terms. Unfor-tunately, most owners will only start to listen oncedegenerative signs develop.
Adjust Feeding
Contrasting the patterns of food gathering in wildparrots with the feeding styles of companion par-rots reveals important and dramatic information.Foraging is thought to occupy a considerable partof every wild parrot’s day. Even in environmentssuch as rain forests where food is relatively abun-dant, parrots have been observed spending con-siderable time locating and foraging for food.Some biologists have hypothesized that the im-pressive capacity of the parrot to learn and re-member may have evolved in response to the needto locate and recognize a huge variety of foodsspread out over considerable distances, alongwith the amount of time necessary to cover thesedistances.[24] When this is coupled with the rea-sonable expectation that many parrots will betriggered to breed by seasonally abundant foodsupplies, the impact on companion parrots of reg-ularly delivered meals is immense.
When contrasted with wild parrots, the ac-cepted tradition of two or three daily meals pre-pared and delivered, often augmented with treatsand goodies, leaves little else to do during the day
242 Manual of Parrot Behavior

but digest. Companion parrots are notorious fordisrupting meal times, as they demand favoritemorsels from the table. As many owners thinktheir bird will balance its own diet and life is eas-ier at table times when the bird is happy, manybirds end up eating the foods that feel best goingdown. Bread, rice, pasta, sweets, and butter haveuniversal appeal but will not nourish a parrot in amanner that will provide sustainable health.
Many simple methods can remedy these com-mon problems. Varying the foods offered andfeeding only the amounts readily consumed cansimplify life and result in a healthier bird.Organic formulated diets deliver nourishing fareeasily and quickly. Supplementation with organicvegetables offers variety. As birds typically feedtwice a day, early morning and late afternoon,meal-feeding for a finite time (one to two hours)twice a day can have numerous benefits. Meal-feeding restores one more facet to the regular pe-riodicity observed in wild parrots. Meal-feedingavoids a lot of waste and seems to return food toa mode of sustenance instead of entertainment.Techniques that encourage foraging are becomingmore popular. Hiding morsels of food amongrocks in a bowl can keep a parrot thinking andmoving. Stainless steel skewers offer a handy wayto hang vegetables. Careful timing of feeding canturn a loud morning or evening “power hour” intoa quiet mealtime.
Provide Opportunities for Exercise
Parrots have evolved to fly. Most of the uniquephysiologic adaptations of a bird are geared tothis very physical activity. Flight demands vigor-ous health. From physical structure to mecha-nisms of gas exchange in their respiratory systembirds, including parrots, are all about flight. Thedrive to exercise is seen in companion birds.Many aviculturists advise allowing a young birdto successfully fledge as an important develop-mental milestone.[39] Certainly flight in mosthomes is indeed more hazardous than a physicallyunchallenging sedentary life, but the fact remainsthat real sustainable health in parrots requires ex-ercise and physical conditioning. Flight can betaught to any bird. Taking off is innate, landingand navigating are learned. The opportunity to flycan sometimes be offered in the home, or outdoorenclosures can offer an alternative. There is nodoubt that allowing flight is risky in many or most
situations and owners should educate themselvesthoroughly before choosing this option. Whetherexercised indoors or outdoors, clipped or flighted,physical activity is imperative for sustainablehealth. It is worth considering that a lack of phys-ical exercise and the subsequent disuse (atrophy)of the poorly defined but unique cardiovascularadaptations that allow for flight might predisposean individual to untoward effects of chronic hor-monal stimulation of the circulatory system.
Use Clicker Training
Clicker training has emerged as a very easy andrewarding way to enhance life and learning inmany animals. When the clicker is used with afood reward to develop new lines of communica-tion between a parrot owner and a bird, the capac-ity of the bird to learn can be astounding. Mostowners can be trained to teach simple behaviorsquickly and easily. The real beauty of this tech-nique is that it offers redirection of attention to-ward learning and exercise. Clicker training hasthe potential to replace quality-shared time be-tween an owner and a parrot with skills and learn-ing instead of cuddling. In this way, inappropriateand generally unintended overbonding with acompanion bird with its attendant reproductivebehavioral cues can be reduced or eliminated.
Address Problematic Bird Husbandry
In many homes, owners have adopted the notionof consistent cage arrangement, a wide variety offoods chosen by preference instead of nutritivevalue, and quality time usually consisting of latenights and cuddling. Unfortunately, these well-intended practices have likely contributed in amajor way to undermining the chances of a long-term successful pet relationship. However, sus-tainable health may be possible using knowledgegained from the natural history and innate behav-ioral tendencies of companion parrots. Changingenvironmental conditions, minimizing pair bondformation, and recognizing and addressing repro-ductive behaviors early could change the lives ofbirds—and their owners—dramatically.
CONCLUSION
Reproductive problems are common in avian vet-erinary medicine and frequently recur after med-ical intervention. While many of these disordersare perceived as behavioral problems, birds are
21 / Problem Sexual Behaviors of Companion Parrots 243

likely being cued reproductively by environmen-tal conditions present in the home that activate theHPG axis. Understanding the pathophysiology ofthese reproductive behaviors provides a likelymeans to prevent chronic hormonal stimulation.Individual species have their own triggers for re-productive activity, and environmental manipula-tion and attention to husbandry issues/pair bondformation may remove these triggers. Prevention,rather than intervention, may be the key.Enhancing our view of individual species with arenewed appreciation for their natural history andinnate behaviors will be necessary for success.
REFERENCES
01. Short, L.L. 1993. The lives of birds. New York:Henry Holt and Co.
02. Meyer, D.B. 1986. “Pineal gland.” In Avian physi-ology, ed. P.D. Sturkie, pp. 501–505. New York:Springer-Verlag.
03. Johnson, A.L. 2000. “Reproduction in the female.”In Sturkie’s avian physiology, 5th ed., ed. G.C.Whittow, pp. 569–596. San Diego: AcademicPress.
04. Jarvis, E.D., C. Scharff, M. Grossman, J.A. Ramos,and F. Nottebohm. 1999. For whom the bird sings:Context-dependent gene expression. Neuron21:775–788.
05. Jarvis, E.D., and C.V. Mello. 2000. Molecularmapping of brain areas involved in parrot vocalcommunication. J Comp Neurol 419:1–31.
06. Jarvis, E.D., S. Ribeiro, J. Vielliard, M. DaSilva, D.Ventura, and C.V. Mello. 2000. Behaviorally-driven gene expression reveals hummingbird brainsong nuclei. Nature 406:628–632.
07. Ottinger, M.A., J. Wu, and K. Pelican. 2002. Neu-roendocrine regulation of reproduction in birdsand clinical applications of GnRH analogues inbirds and mammals. Semin Avian Exotic Pet Med11 (2):71–79.
08. Speer, B.L. 2003. “Sex and the single bird.” ProcAnnu Conf Assoc Avian Vet, pp. 331–343.
09. Millam, J.R. 1997. “Reproductive physiology.” InAvian medicine and surgery, ed. R.B. Altman, S.L.Clubb, G.M. Dorrestein, and K. Quesenberry, pp.12–28. Philadelphia: .W.B Saunders.
10. Pollock, C.G., and S.E. Orosz. 2002. Avian repro-ductive anatomy, physiology and endocrinology.Vet Clin North Am Exotic Anim Pract 5:441–474.
11. Joyner, K.L. 1994. “Theriogenology.” In Avianmedicine: Principles and application, ed. B.W.Ritchie, G.J. Harrison, and L.R. Harrison, pp.748–804. Lake Worth, FL: Wingers Publishing.
12. Romagnano, A. 1996. Avian obstetrics. SeminAvian Exotic Pet Med 5 (4):180–188.
13. Rosskopf, W.J., and R.W. Woerpel. 1996. “Featherpicking and therapy of skin and feather disorders.”In Diseases of cage and aviary birds, ed. W. Ross-kopf and R. Woerpel, pp. 397–405. Baltimore:Williams and Wilkins.
14. Forshaw, J.M., and W.T. Cooper. 1989. Parrots ofthe world, 3rd (rev.) ed. Willoughby, NSW, Aus-tralia: Weldon Publishing.
15. Sparks, J., and T. Soper. 1990. Parrots: A naturalhistory. New York: Facts on File, Inc.
16. Coyle, P.G., Jr. 1987. Understanding the life ofbirds. Lakeside, CA: Summit Publications.
17. May, D.L. 1996. The behavior of African grey par-rots in the rainforest of the Central AfricanRepublic. Psittascene 8:8–9.
18. Pepperberg, I.M. 1994. Numerical competence inan African grey parrot. J Comp Psych 108:36–44.
19. Pepperberg, I.M. 1992. Proficient performance ofa conjunctive, recursive task by an African greyparrot (Psittacus erithacus). J Comp Psych106:295–305.
20. Elphick, C, J.B. Dunning Jr., and D.A. Sibley.2001. The Sibley guide to bird life and behavior.New York: Alfred A. Knopf.
21. Hudelson, S.A. 1996. Review of the mechanismsof avian reproduction and their clinical applica-tions. Semin Avian Exotic Vet Med 5 (4):189–198.
22. Johnson, A.L. 1986. “Reproduction in the male.”In Avian physiology, ed. P.D. Sturkie, pp. 432–451.New York: Springer-Verlag.
23. Powell, G., and R. Bjork. 1996. Ara ambigua:Preliminary observations on the Costa Rican pop-ulation and their lowland forest habitat and on theirconservation. Washington, DC: RARE Center forTropical Conservation.
24. Munn, C.A. 1994. Macaws: Winged rainbows.National Geographic 185 (1):118–140.
25. Munn, C.A. “Macaw biology and ecotourism” or“When a bird in the bush is worth two in the hand.”In New world parrots in crisis, solutions from con-servation biology, ed. S.R. Bessinger and N.F.R.Snyder, pp. 47–72. Washington DC: SmithsonianInstitution Press.
26. Millam, J.R. 1999. Reproductive management ofcaptive parrots. Vet Clin North Am Exotic AnimPract 2:93–110.
27. King, A.S., and J. McLelland. 1984. Birds: Theirstructure and function. London, UK: BailliereTindall.
28. Arndt, T. 1986. Parrots: Their life in the wild. Brom-litz, Germany: Horst Muller-Verlag Walsrode.
29. Nespor, A.A., M.J. Lukazewicz, R.J. Dooling, andG.F. Ball. 1996. Testosterone induction of male-
244 Manual of Parrot Behavior

like vocalizations in female budgerigars (Melo-psittacus undulates). Horm Behav 30 (2):162–169.
30. Sweeney, R.G. 1993. The Eclectus: A completeguide. Ontario, Canada: Silvio Mattacchione andCo.
31. Dilger, W.C. 1962. The behavior of lovebirds. SciAm 206:88–98.
32. Millam, J.R., B. Zhang, and M.E. el Halawani.1996. Egg production in cockatiels (Nymphicushollandicus) is influenced by number of eggs innest after incubation begins. Gen Comp Endo-crinol 101 (2):205–210.
33. Whittow, G.C. 1986. Regulation of body tempera-ture. In Avian physiology, ed. P.D. Sturkie, pp.221–252. New York, NY: Springer-Verlag.
34. Rosenthal, K.L. 2003. “Cytology, histology, andmicrobiology of feather pulp and follicles offeather pickers.” Proc Annu Conf Assoc Avian Vet,pp. 27–31.
35. Cooper, J.E., and G.J. Harrison. 1994. “Derma-tology.” In Avian medicine: Principles and appli-
cation, ed. B.W. Ritchie, G.J. Harrison, and L.R.Harrison, pp. 607–639. Lake Worth, FL: WingersPublishing.
36. Bowles, H.L. 2002. Reproductive diseases of petbird species. Vet Clin North Am Exot Anim Pract 5(3):489–506.
37. Millam, J.R., and H.L. Finney. 1994. Leuprolideacetate can reversibly prevent egg laying in cock-atiels (Nymphicus hollandicus). Zoo Biol13:149–155.
38. Shields, K.M., J.T. Yamamoto, and J.R. Millam.1989. Reproductive behavior and LH levels ofcockatiels (Nymphicus hollandicus) associatedwith photostimulation, nest-box presentation, anddegree of mate access. Horm Behavior 23 (1):68–82.
39. Linden, P.G. 1999. “Fledging and flight for aviancompanions.” Proc Annu Conf Assoc Avian Vet,pp. 61–65.
21 / Problem Sexual Behaviors of Companion Parrots 245

SUMMARY
Mate trauma is considered a syndrome of captivepsittacine birds. It is thought to be induced by thecircumstances of captivity and how they inter-play with the natural behaviors of breeding psit-tacines, that is, their breeding seasonality. Matetrauma occurs predominately in sexually maturecockatoo pairs. It is most common in the white,or light-colored, cockatoo species. In the author’sexperience, mate trauma appears to be most com-mon in domestically raised pairs, early in thebreeding season, and early in the morning. Typi-cally, the male bird is the aggressor and he mostoften attacks the female in the head, neck, andbeak region. The beak is typically the most seri-ously and permanently affected, and thus needslong-term—often lifelong—therapeutic trim-ming and filing. Unfortunately, in addition to thecockatoos, numerous other psittacines and somepasserine and rhamphastidae species also experi-ence mate trauma in captivity. Fortunately forthese species, the incidence of attacks is muchlower. In a select few, the female bird is the ag-gressor.
This chapter will review the syndrome of matetrauma, complete with a discussion of its pro-posed etiologies, preventative management, andtreatment. The chapter will also briefly review theanatomy and the prehensile function of the avianbeak.
MATE TRAUMA
The majority of large psittacine aviculturists inAmerica and Europe have witnessed mate or in-traspecific aggression.[1–5, 7] It is generally ac-cepted that breeding seasonality plays a signifi-cant role in the occurrence of mate aggressionand/or trauma in captivity. During the breeding
season, reproductively active psittacine birds be-come more possessive of their cages and, espe-cially, of their nest boxes; hence, aggressive dis-plays increase. For this reason, visual barriers onthe nest box side of the cage are imperative.[7] Itshould be noted that mate aggression resulting insevere injury or death—also referred to as matetrauma—does not occur in the wild nor is it rec-ognized in aviculture in Australia.[7]
The specific etiology of mate trauma is notknown. However, numerous theories exist, in-cluding asynchrony of reproductive activity, terri-tory and/or food defense, invasion of personalspace, confinement in too small a cage, displacedaggression due to territorial issues, behavioral in-teractions with birds in nearby cages or humansoutside the cage, hormonal imbalances, dietaryimbalances, and so forth.[5, 7] Mate traumas ap-pear to occur sporadically and without provoca-tion. A productive bonded pair may suddenly ex-perience fatal mate trauma for no apparentreason. The pair can be domestic, wild-caught, ormixed. It can be composed of young or old birdsor both. The pair could be recently set up or havea proven history of many years’ duration. The fre-quency of mate trauma is highest in breedingcockatoo species during the spring and early sum-mer months. In the author’s experience, matetrauma occurs most commonly in domesticallyraised pairs, early in the breeding season, andearly in the morning.
This intraspecific aggression is most commonin white cockatoos. In the author’s experience, theWhite Cockatoos are more commonly affectedthan pink, salmon, or black cockatoos. The mostaggressive white cockatoo is the Red-ventedCockatoo (Figure 22.1). This species is closelyfollowed by the Lesser Sulfur-crested Cockatoo
247
22Mate Trauma
April Romagnano

(Figure 22.2). The Red-vented Cockatoos willoften kill and then attempt to eat their dead cage-mates, and thus it is safest to keep the matingpairs of this species in separate cages during thenon-breeding season.
Mate trauma has also been seen in macaws,amazons, conures, Eclectus (Figure 22.3), Ring-necked Parakeets, Senegals, and thick-billedpsittacine parrots. Many species of soft bills, suchas toucans, turacos, doves, and finches, also ex-hibit intraspecific aggression. In the majority ofspecies affected, the male attacks the female ex-cept in the case of the Eclectus Parrot, where thefemale is known to be the aggressor. Occasion-ally, individual females of different species mayattack their male mates, but this phenomenon isnot necessarily species specific. In the author’sexperience, other female psittacine birds that havebeen noted to attack their male cagemates areRing-necked Parakeets, lovebirds, Senegal Par-rots, and Palm Cockatoos.
THE MATE TRAUMA ATTACK
Male cockatoos are the most common aggressors.The typical mate trauma attack occurs at dawn.The etiology of these aggressive attacks isthought to be multifactorial. It is the author’sopinion that it is triggered, to some extent, by hor-monal changes since the majority of attacks occurin the spring and early summer. Presumably, if themale bird’s aggressive advances are ignored orthwarted (i.e., disturbed by other birds or people,misinterpreted, or not fully acted out due to cagespace limitations), the male tends to lash out athis female cagemate. Since the female bird canrun but not escape the male’s offensive aggressiveadvances, she is forced to defend herself. The au-thor feels that the male becomes further enragedand increases the frequency of his attacks. Thus itappears that the male then relentlessly chases thefemale, attacking her whenever possible. The au-thor has treated approximately 100 mate traumas.If the female is rescued alive, in addition to being
248 Manual of Parrot Behavior
Figure 22.1. The most aggressive whitecockatoo is the Red-vented Cockatoo like theone pictured here. See also color section.
Figure 22.2. The aggressive Red-ventedCockatoo is closely followed in level of aggres-sion by the Lesser Sulfur-crested Cockatoo likethe one pictured here. See also color section.

wounded and moribund, she is typically ravenousand has lost a significant amount of weight. Thissuggests most mate traumas are not an isolatedevent and are more likely a series of offensive at-tacks by the male. If the female is not too old, ingood health, and flighted, she will initially be ableto evade the male’s aggressive attacks. However,without human intervention, captive females be-come exhausted and succumb. Attacks can occuranywhere in the cage, but are more likely to occurin areas of increased contact such as the feedingand drinking area or the nest box. These aggres-sive attacks essentially run the female ragged, andboth prostration and starvation set in. The cagedfemale thus becomes easy prey and is ultimatelymauled by the male. Usually, after the initial matetrauma attack occurs, the male goes back formore, unless an astute aviculturist rescues the fe-male. Most fatal attacks occur within the nestbox, since it is the easiest place for the male tocorner the female. If the female is not rescued, the
male will continue to attack his female cagemateuntil she is rendered completely moribund and fi-nally dies.
Mate trauma is virtually unknown in Aus-tralia.[7] It is presumed that in the wild, the unre-sponsive female simply flies away, escaping mateaggression and trauma completely. In captivity,as previously mentioned, mate trauma attackstypically continue until prostration, death, orhuman intervention occurs. Male cockatoos arerelentless, inflicting physical damage predomi-nately to their mate’s head, face, mouth, and beak(Figure 22.4).
Although beak trauma can also occur duringparental feedings of neonatal or juvenile birds,between neonatal birds and young juvenile birds,and between weaning birds housed together inoutdoor aviaries or in cages within a home, thesebeak traumas are often accidental or are isolatedsquabble incidents. Many result from parents (tooaggressively) feeding their babies or their babiesinnocently pumping on each other’s soft pliableuncalcified and unkeratinized beaks. The authorhas only once witnessed what appeared to be aweaning bird cagemate trauma. The mate traumaappeared to be deliberate and resulted in the deathof the attacked female. This trauma occurred in ajuvenile pair of unrelated Palm Cockatoos housedin a large outdoor nursery flight cage. The delib-erate nature of this cagemate trauma appears to be
22 / Mate Trauma 249
Figure 22.3. Eclectus, like those pictured here,are also capable of mate trauma. See also colorsection.
Figure 22.4. Severe beak damage in a whiteCockatoo, killed by mate trauma.

the exception and not the rule when the involvedbirds are immature and not set up for breeding.The author has also witnessed this level of fatalaggression in bonded, same-sex pairs that wereunknowingly set up and in groups of mate traumamales housed together. Same-sex aggression andtrauma involving male cockatoos has been wit-nessed by other aviculturists and avian veterinari-ans where birds were set up in a breeding colonyor known aggressive male were accidentallyplaced with another breeding pair or with conspe-cific males.
PREVENTATIVE MANAGEMENT
To best manage psittacine birds, it is necessary toconsider their life stages.[3] A parrot’s life stagescan be divided into neonatal and juvenile, adultreproductive, and adult post-reproductive stages.The adult reproductive stage is the longest periodof the bird’s life and when the majority of matetraumas occur.[3]
The adult reproductive life stage is also themost aggressive stage, when birds are the mostvocal and the least tame. A calmer, better pet-quality bird is characterized by the neonatal andjuvenile stage and the post-reproductive stage.[3]Birds are the most affectionate and playful whenyoung and then return partially to this state asquiet, calmer, geriatric birds, even if they spentthe interim years in breeding.[3]
Various methods of mate trauma preventionhave been reported.[1–5, 7] The author feels mostcomfortable with the simplest methods, which arenon-surgical and the least invasive. These includeroutine bilateral wing and maxilla beak tip trim-ming and filing (to roundness) of known aggres-sive male birds.
These procedures must be performed in thepre-breeding season, that is, the fall.
Although these beak and wing filing and trim-ming methods are temporary, the results usuallylast for up to four months, getting the pair throughbreeding season—the prime time for mate traumaaggression.
Other more invasive mate trauma preventionmethods include acrylic beak balls or rubber beakbumpers being surgically applied to the tip of themale’s maxilla. These methods were explored inthe 1990s and offered effective, albeit temporary(two weeks to three months), protection for fe-male cagemates.[1–4]
The rubber beak bumpers lasted longer thanthe acrylic beak balls. This same author devel-oped a modified surgical debeaking procedure foraggressive male psittacine birds.[1–4] In this pro-cedure, the bird is placed under anesthesia and themaxilla tip is cut off with the cutting wheel of adremel drill.[4] This procedure was found to betemporary, as the cut maxilla tip regrew in allcases but was often reduced in size. When com-pared to the acrylic beak balls or rubber beakbumpers, this procedure lasted the longest.[4]That author further noted that although disfigure-ment of the beak occurred, the process was tem-porary and could save pairs. Another author re-ported a more aggressive surgical technique thatwas intended to be permanent; it is called lowerbeak and complete mandible bisection or split-ting.[5] This surgery was viewed, by that author,as a salvage procedure to be performed only whenall other methods failed to protect the femalefrom an attacking male. The author stated that heperformed this surgery on only the most aggres-sive male cockatoos, so the pair could be kepttogether. However, any surgical procedure thatpermanently disfigures an animal is indeed con-troversial, as stated by that, and this, author.[5] Incases of very aggressive males, it is recom-mended to resort to the other preventative meth-ods such as permanent pair separation. In somecases, the pair must be sacrificed to save thefemale.
Other preventative measures include removingor changing the nest box. Special nest boxes canbe made with two entrances and baffled interiors.This type of nest box design may limit box attackson females. An extension of this design is twocages attached together, side by side. Each cagehas its own nest box with two entrances. Thecages communicate by two entrances (or exits),one at the front and one at the back of the bi-cagesetup. Other special cage designs focus on ele-vated escape areas for the flighted female that areless accessible to the wing-clipped males. To makethese areas completely inaccessible to the clippedmales they must also be skirted by cockatoo-safe sheet metal to prevent the clipped male fromclimbing to the female’s elevated escape areas.Temporary pair separation is usually necessaryand occasionally breaks the cycle of aggression.This separation can be for a minimum time ofseveral weeks to several months or years depend-
250 Manual of Parrot Behavior

ing on the severity of the mate trauma attack andthe time needed by the female to fully recover.Permanent separation, however, is the ultimateway to break the cycle of aggression and assurethe safety of the female.
Repair of the involved birds is yet another pre-ventative measure. However, in cases of recurrentmate traumas, very severe mate traumas, or matedeath the aggressive males should be retired alto-gether from breeding. The pair should be sepa-rated and the males placed in new homes either asdisplay birds in zoological collections or pet birdsin home situations. It is the author’s experiencethat some of these males do indeed make goodpets in single-pet homes. The surviving femaleswith extensive beak lesions may be retired to res-cue or pet bird home situations. Future owners ofthese severely affected female mate trauma birdsmust be willing to be diligent with the bird’s vet-erinary care for life. If repair of the birds is an op-tion, mate trauma males should be given larger,more aggressive female mates and the femalesgiven smaller, less aggressive male mates.
TREATMENT
To properly treat mate trauma, which typically in-volves various types of severe beak trauma, theavian veterinarian must be very familiar with theavian beak. The avian beak function and anatomymust be fully understood. The two following sec-tions on prehensile function of the beak andanatomy of the beak will outline the vast impor-tance of this organ to the healthy psittacine bird.
Moribund mate trauma females do best whenminimally handled. A quick careful physical ex-amination should be followed by parenteral fluidadministration.
Take care to also check that the bird is not eggbound by performing gentle abdominal palpation.Only warmed fluids should be used on matetrauma birds. After their quick physical examina-tions, the females should be treated immediatelywith intravenous (IV) dextrose and crystalloidfluids, subcutaneous (SQ) dextrose and crystal-loid fluids, plus a shock dose of SQ dexametha-sone. If possible, the female’s wounds should bevery carefully and quickly cleansed. She shouldimmediately be placed into a dark incubatorwhere heat and oxygen are provided. Next, de-pending on the extent of beak trauma, soft foods,such as mixed commercial baby bird formula,
soaked commercial bird pellets, and large juicypieces of fruit, should be offered. No watershould be offered initially. A water bowl in thecage of an extremely weak bird is dangerous sincethe bird could drown if it falls in. The weak trau-matized female should be kept quiet during herinitial period of stabilization.
After the bird is stabilized, the avian veterinar-ian can safely work on her wounds and addressher beak trauma in a conservative fashion. Pleasetake note that in the vast majority of cases, acry-lics are not indicated. The application of acrylicsto an ultimately infected bite wound is a recipe forabscess formation. Wounds in the avian beak, liketurtle and tortoise shell wounds, do best whenmanaged as open wounds. The female’s beakwounds should be cultured and treated with par-enteral and topical antibiotics and antifungals asindicated. Careful flushing and repeat debride-ment over time are an important part of the slowhealing process.
Despite the fact that female mate trauma vic-tims typically have permanent grotesquely disfig-ured beak lesions, they learn to adapt and eat ontheir own quite quickly. Their lesions, however,often require lifelong beak trimming and filing byan avian veterinarian. This is because most beaktraumas result in malocclusion, which over timecauses horrific unnatural overgrowths of affectedportions of the keratin of the upper or lower beak.In order to attempt to align the working horn, thehard keratin occlussal surface, the cutting edge orthe tomium, trimming and filing is imperative forlife so these birds can eat to the best of their abil-ity. Beak trauma birds are destined to have over-grown beaks secondary to their beak trauma-induced malocclusion. It is the author’s opinionthat birds altered in this way are more likely to bedepressed, timid, and non-confrontational but farewell if they survive the initial attack.
If these birds are subsequently kept alone as ei-ther educational or pet birds, they can still havehappy and healthy lives.
PREHENSILE FUNCTION OF THE BEAK
The psittacine beak has numerous prehensilefunctions and hence must be in the best possibleform at all times.
Since birds do not have lips and teeth and be-cause their thoracic limbs are specialized forflight, their beaks have evolved to have signifi-
22 / Mate Trauma 251

cant and extensive prehensile function. Thehighly evolved prehensile function of the avianbeak is aided by the dexterous tongue and flexiblefeet of the various avian species.[6]
Avian beak functions are numerous and includethe procurement of food and water and the prepa-ration of food. The beak’s cutting edge, ortomium, serves to shave the bird’s food versuschewing it. The beak also serves in grooming,sexual display, courtship, nest building, egg turn-ing, evaporative heat loss (panting), locomotion,and ultimately, defense.[6]
The adult avian beak is also a formidableweapon. It is known as the rhamphotheca and is ahard keratin structure that covers the rostral as-pect of the upper and lower jawbones of the bird.The upper beak is called the rostrum maxillarisand the lower beak the rostrum mandibularis.[6]
On the other hand, the growing neonatal andjuvenile beak is a pliable and malleable ap-pendage until calcification and keratinizationoccur. The beak of a young chick can thereforechange from perfect occlusion to complete mal-occlusion in a day. The majority of beak problemsin the very young are divided into five categories:lateral deviations of the maxilla (scissor beak),compression deformities of the mandible, prog-nathism (pug beak), subluxation of the premax-illa, which occurs more often in juveniles, andbeak trauma, which can occur in birds of any age.The first two malformations are most common inmacaws, the third in cockatoos, and the fourth injuvenile macaws.[6]
While chicks are young and the beak is still pli-able and fast-growing, physical therapy and trim-ming and filing, along with medical treatment,are indicated post-beak trauma and extensive sur-gery may not be necessary. After calcification andkeratinization, however, in addition to frequentclipping and filing, surgical acrylic implants orextensions may be needed to properly align acrushed and damaged beak. Hence after calcifica-tion and keratinization beak trauma repair may re-quire rhamphothotics to correctly repair thebeak’s alignment and integrity. However, this isoften not necessary even in the worst of traumacases, as psittacines are very adaptable and learnto eat and drink with even the most severe of beaktraumas.[6] Further, acrylic patches, or implants,applied over infected beak wounds foster bacter-ial and fungal abscess formation.
Physiologically the rhamphotheca is an ever-growing dynamic tissue, which contains abundantkeratin and, in adults, free calcium phosphate. Ittakes approximately six months to replace itselfin adults, making beak healing and repair a veryslow process.[6]
ANATOMY OF THE BEAK
The avian beak is composed of bone, dermis, epi-dermis, a transitional layer, and the keratinizedepidermis (horn or rhamphotheca). The rostrummaxillaris (upper beak) and the rostrum mandi-bularis (lower beak) are all-inclusive terms for thekeratin, soft tissue (dermis), and boney structuresof the indicated beak.[6]
The cancellous boney structure of the upperbeak includes the premaxilla, nasal, and frontalbones. The premaxilla thickens at the tip inpsittacines forming the bill tip organ, which is in-nervated by cranial nerve VI (deep ophthalmicnerve). This organ contains highly sensitive re-ceptors important in food collection, eating, andpreening. The lower cancellous boney beak con-sists of the mandible, which joins the quadratebone.[6] It also contains a bill tip organ in certainspecies, such as parrots, geese, and ducks. Theexact location and development of the bill tiporgan varies among avian species. In parrots it isbelieved that the bill tip organ is most developedin the lower beak. The bill tip organ is extremelysensitive and this should be considered when ma-nipulating the tip of both the upper and lowerbeak. To improve the sensory capacity of the billtip organ, the lower beak should always be in-cluded in routine grooming of the overgrownbeak.
The soft tissues of the beak include the rham-photheca, which is divided into the harder work-ing horn and the softer covering horn. Rostrallythe rhamphotheca changes into the working horn,a hard keratin occlusal surface ending in the cut-ting edge, or the tomium. The maxilla’s keratin isknown as the rhinotheca, and the mandible’s ker-atin is called the gnathotheca. The cere is knownas the base of the rhinotheca, which remains soft.The rhinotheca joins the frontal bone via a kineticjoint, or the nasal-frontal hinge. This synovialjoint is mobile in psittacines and fixed in raptors.The gnathotheca is adjoined to the mandible viaconnective and muscular tissue attachments.[6]
The rhinotheca grows in the cranioventral
252 Manual of Parrot Behavior

plane and the gnathotheca grows more quickly inthe craniodorsal plane. The rhamphotheca is com-posed of thickened layers of stratum corneumoriginating from the stratum germinativum. Itcontains abundant keratin and, in adults, free cal-cium phosphate. Baby birds’ beaks are pliable be-cause the free calcium phosphate has not yet beenlaid down, or calcified.[6]
CONCLUSION
Mate trauma is a syndrome induced by the con-fines and limitations of captivity. It apparentlydoes not occur in the wild, and given this fact, theauthor feels aviculturists and avian veterinariansshould go above and beyond to treat these abusedbirds to the best of our collective abilities. We, asavian veterinarians, should also attempt to ad-dress the emotional healing of these woundedbirds and fight for their future protection from re-peat abuse situations by encouraging their re-moval from breeding in captivity when necessary.Further, the author feels we, as avian veterinari-ans, should also educate new, and remind old-time, aviculturists of the horrors of mate traumaand the importance of an intact, fully functionalprehensile beak appendage.
ACKNOWLEDGEMENTS
The author would like to thank Dr. Scott G. Mar-tin and Dr. Tarah Hadley for their expert editorialcomments and advice.
REFERENCES
1. Clubb, K., S. Clubb, A. Phillips, S. and Wolf S.1992. “Intraspecific aggression in cockatoos.” InPsittacine aviculture, perspectives, techniques andresearch, ed. R.M. Schubot, K.J. Clubb, and S.L.Clubb, pp. 25-1–25-5. Loxahatchee, FL: AviculturalBreeding and Research Center.
2. Clubb, S.L. 1997. “Avicultural medicine and flockhealth management.” In Avicultural medicine andsurgery, ed. R.B. Altman, S.L. Clubb, G.M. Dorres-tein, and K. Quesenberry, pp. 101–116. Philadel-phia: W.B. Saunders Co.
3. Clubb, S.L. 1998. Captive management of birds fora lifetime. JAVMA 212 (8):1243–1245.
4. Clubb, S.L. 1998. Management of psittacines to re-duce mate aggression and trauma. PAAV, pp.133–138.
5. McDonald, S E. 2000. Beak altering procedures todisarm aggressive male cockatoos. Exotic DVM2.2:29–31.
6. Romagnano, A. 1998. “Beak malformations andcorrections.” ABVP Symposium, pp. 37–40.
7. Styles, D. 2001. “Captive psittacine behavioral re-productive husbandry and management: Sociali-zation, aggression control, and pairing techniques.”PAAV (Speciality Advanced Program), pp. 3–19.
22 / Mate Trauma 253

Feather picking is one of the most common andchallenging behavior problems of captive psitta-cine birds. It has been described as an exaggera-tion of normal preening behavior. However, thenormal preening patterns of most psittacinespecies have not been studied, and normal birdsmay spend a significant portion of their daily timebudget engaged in preening behavior (Spruijt etal. 1992). Regardless of the duration, in thecourse of normal preening, no damage to thefeathers or skin should occur.
Rosskopf and Woerpel (1996) define featherpicking as a condition in which the bird damagesits feathers or skin or prevents the normal growthof feathers. Feather picking in birds is associatedwith feather chewing or removal, with or withoutself-inflicted soft tissue damage (Galvin 1983;Nett & Tully 2003). Soft tissue damage to skin ormuscle occurs in some birds, with a higher inci-dence in cockatoo species (Rosenthal 1993)(Figures 23.1, 23.2, and 23.3). The term “feather-picking disorder” will be used in this chapter todescribe the syndrome involving a variety ofself-directed feather and soft tissue destructivebehaviors with no apparent underlying medicaletiology.
Clinical signs vary widely, ranging from mildlocalized feather damage to completely denudedareas and soft tissue excoriations. Chewed feathershave an irregular appearance, with the ramus of thefeather shaft split longitudinally. Damage is in-flicted to areas of the body that are accessible to thebird, with normal, unaffected head and crest feath-ers (Harrison 1986). The distribution of featherloss or damage is highly variable. According toRosenthal (1993), the inner thighs and sternum are
often affected. Nett and Tully (2003) report that themost commonly affected sites are the chest, underthe wings, and over the rump, with feather chewingprimarily affecting primary feathers. According toPerry (1994), the condition often lacks bilateralsymmetry, especially in the early stages.
SPECIES PREDISPOSITIONS
African Grey Parrots (Psittacus erithacus), ma-caws (Ara spp.), cockatoo species, conure spe-cies, Eclectus Parrots (Eclectus roratus), andGrey-cheeked Parakeets (Brotogeris spp.) are re-portedly predisposed to feather-picking disorder,while it is less common in Budgerigars (Melo-psittacus undulatus), Cockatiels (Nymphicus hol-landicus), and amazon parrots (Amazona spp.)(Rosskopf & Woerpel 1996). Jenkins (2001) in-cludes Monk Parakeets (Myiopsitta monachus) asbeing predisposed to feather-picking disorder.According the Rosskopf and Woerpel (1991) andBriscoe et al. (2001), African Grey Parrots are themost commonly presented species for feather-picking disorder.
DIAGNOSIS OF FEATHER-PICKINGDISORDER
Feather damage, removal, or loss may result froma variety of medical, environmental, or behavioralcauses (Harrison 1994; Welle 1999). Self-traumato feathers or skin is a non-specific symptom thathas many possible etiologies. Medical disorders,nutritional deficiencies, toxin exposure, and envi-ronmental irritants should be considered in anycase of feather picking, and primary medicalproblems and medical complications resultingfrom self-inflicted trauma need to be addressed.
255
23Feather-Picking Disorder
in Pet Birds
Lynne M. Seibert

All birds with feather abnormalities warrant athorough history and physical examination, withappropriate diagnostic testing (Rosskopf & Woer-pel 1996; Welle 1999). Any treatment plan shouldaddress both the physical and mental needs of thepatient.
Medical differentials that should be consideredinclude endoparasitism, systemic diseases, infec-tious diseases (circovirus, polyomavirus), malnu-trition, and neoplasia. Giardia has been associatedwith feather picking, particularly in Cockatiels(Fudge & McEntee 1986). Less common causesof feather picking include ectoparasitism, pri-mary infectious dermatitis or folliculitis, and en-docrine imbalances (Rosenthal 1993).
The influence of pruritis, allergies, and hyper-sensitivity on feather-picking behavior is unclear,although advances are being made in diagnostictesting for allergies in psittacine patients (Colom-bini et al. 2000; Macwhirter et al. 1999).
A thorough physical examination is necessary.Feather abnormalities will be confined to areasthat the bird can reach. There may be evidence ofchewed feathers, damaged soft tissue, missingfeathers, or feather regrowth.
Diagnostic testing may include fecal examina-tion, skin scrapings, hematology, biochemical
analysis, radiographs, bacterial or fungal cultureand sensitivities, feather pulp and skin cytology,feather follicle and skin biopsies, and viral testing(Koski 2002; Rosenthal 1993). Air quality, hu-midity level, and exposure to toxins, chemicals,or smoke should also be addressed.
Taking a detailed behavioral history for theavian patient is a time-consuming, but extremelyimportant, component of the diagnostic process.Information obtained from the history will aid notonly in the diagnoses but also in designing a com-prehensive treatment plan. To increase efficiency,clients can complete a questionnaire prior to theappointment, and additional questions can bebased on this information. A thorough behavioralhistory should include information regarding theonset of feather picking, environmental changesassociated with the onset of the behavior, durationof the condition, the time of day when the behav-ior is most intense, progression of signs, housingconsiderations, social interactions with otherbirds and human caregivers, a description of theabnormal behaviors, any seasonal patterns thatmay be apparent, and responses to previous treat-ments. Additional information should be obtainedabout the eliciting stimuli, how the behavior isterminated, whether or not the behavior can be in-
256 Manual of Parrot Behavior
Figure 23.1. Cockatoo with feather-pickingdisorder. See also color section.
Figure 23.2. Self-inflicted soft tissue lesions in a Moluccan Cockatoo with feather-pickingdisorder.

terrupted, and how the caregiver reacts to the be-havior. Early history information should deter-mine whether the bird was parent- or hand-raised,the source of the bird, the age at weaning, earlyadverse experiences, previous homes, and contactwith other birds and humans during possible sen-sitive periods of development. Caregivers shouldbe carefully questioned about nutrition, whichfoods are offered, which are eaten, and the quan-tities consumed.
CONSEQUENCES OF FEATHER PICKING
Feathers are essential for flight and provide in-sulation, mechanical and thermal protection,proprioception, and waterproofing (Lucas &Stettenheim 1972; Nett & Tully 2003). The con-sequences of feather picking may be strictlyaesthetic, but more serious sequelae may also re-sult. Molting increases the metabolic demandsfor the bird, and chronic feather replacement canincrease disease susceptibility and alter ther-moregulatory ability (Nett & Tully 2003; Ross-kopf & Woerpel 1996). Blood loss from trau-matized blood feathers and damaged soft tissuecan become significant (Galvin 1983). Feather-less areas may bruise easily, and secondary in-fections are also possible. Permanent damage to
the feather follicle may prevent normal regrowthof feathers.
POSTULATED CAUSES OF FEATHER-PICKING DISORDER
The causes of feather-picking disorder have notbeen determined. Many potential risk factorshave been suggested, but the exact etiology re-mains unknown. The common perception that in-appropriate husbandry or boredom is responsiblefor the development of feather-picking problemsraises concerns for caregivers about the quality oflife of affected birds.
Boredom is the most commonly cited cause offeather-picking disorder (Galvin 1983; Lawton1996; Rosenthal 1993). This is an overly simplis-tic explanation for a complicated disorder thatlikely develops as a result of multiple factors, in-cluding developmental influences, socializationdeficits, neurochemical abnormalities, environ-mental issues, temperament traits, hormonal in-fluences, genetics, undiagnosed medical condi-tions, and learning factors.
Foraging is an activity that consumes a signifi-cant amount of time for free-ranging psittacinebirds. Ad libitum provision of food for captivepsittacine birds, as well as restrictions in freemovement, may predispose some birds to developbehavior problems (Lawton 1996; Rosskopf &Woerpel 1996). Environmental enrichment ef-forts for avian species have addressed the impor-tance of providing opportunities for foraging(Bauck 1998; VanHoek & King 1997).
Optimal environments for captive psittacinebirds should also provide opportunities for thebird to engage in species-typical behaviors, in-cluding locomotor activities. Chronic confine-ment does not provide the necessary opportuni-ties for the bird to engage in normal behaviors.
Inappropriate or inadequate social interactionsare also common issues for captive birds and maybe problematic for some birds with feather-picking disorder. Pet birds are often isolated fromconspecifics and must rely on human caregiversfor social interactions. The majority of psittacinespecies kept in captivity are highly social, livingin stable flocks under natural conditions, and maynot acclimate well to a solitary lifestyle. Sepa-ration anxiety should be considered in birds thatengage in feather-picking behavior while aloneand in birds that show distress upon being left
23 / Feather-Picking Disorder in Pet Birds 257
Figure 23.3. Feather chewing in a Sulfur-crested Cockatoo with feather-picking disorder.

alone. Undesired exposure or contact with familypets, other birds, natural predators, or familymembers has also been implicated in some casesof feather picking.
Stress may also cause feather-picking behav-iors to develop. Preening is commonly performedas a displacement behavior in situations in whichconflicting behavioral systems are activated or instates of motivational ambivalence (Spruijt et al.1992). Preening has been associated with comfortor dearousal and may therefore be performed intimes of stress (Delius 1988). An unpredictableenvironment may be associated with increasedstress (Rosenthal 1993). Crowding can also con-tribute to stress, as well as poor husbandry. Anyabrupt change to the environment should also beconsidered as a potential stressor and contributingfactor. Environmental changes of potential impor-tance may include additions or deletions to thefamily, changes in the physical environment,schedule changes, housing changes, moving thebird, or climate changes. Rehoming of birds cancontribute to stress when important social bondsare broken and new social bonds must be formed.
Exaggerated or prolonged reproductive behav-ior has been suggested as a possible cause offeather picking (Rosenthal 1993). Reproductivebehavior changes can occur on a seasonal basis orthroughout the year, particularly in birds that areexposed to unnatural photoperiods. Some avianspecies naturally remove feathers from their ab-domen during the breeding season to enhancecontact with their eggs, creating a brood patch(Oppenheimer 1991). Feather removal during thebreeding season that involves ventral areas maybe associated with reproductive behavior. Featherpicking that persists past the breeding season isconsistent with a feather-picking problem. Birdsthat are exposed to prolonged light photoperiods,consume high fat diets, or display sexual behav-iors should be considered at risk for feather pick-ing, and these issues should be addressed.According to Rosskopf and Woerpel (1996),cockatoos, macaws, conures, and Grey-cheekedParakeets are likely to present with reproductivelyinduced feather picking.
Additional considerations for birds suspectedof reproductive-related feather picking include in-appropriate mate pairings, frustrated mating in-stincts, lack or loss of a mate, or the presence ofother nesting birds in the environment.
Sleep deprivation can have detrimental effectson the welfare of captive birds. Pet birds are oftenhoused in common areas of the home and coveredwhen the caregivers retire for the evening, or atdusk. The high level of vigilance that is character-istic of prey species may prevent adequate rest inthese environments. In addition to the stress asso-ciated with inadequate rest, exposure to lengthyphotoperiods can increase the incidence of unde-sirable reproductive behaviors.
Psittacine infants are altricial, requiring a rela-tively long period of maternal care. Newlyhatched birds in captive breeding programs areoften removed from their parents and weaned byhuman surrogates in an effort to promote ade-quate socialization to humans. However, detri-mental effects of early maternal deprivation havebeen documented in other species (Ruppenthal etal. 1976; Suomi et al. 1976) and should be consid-ered in planning captive breeding programs forpsittacine birds (Jenkins 2001). If birds reared byhuman surrogates do not possess adequate copingskills as adults, then alternative rearing methodsshould be considered. Aengus and Millam (1999)have documented successful socialization of par-ent-raised Orange-winged Amazon Parrots withneonatal handling. One group was handled dailyfor 10–30 minutes, while the control group washandled only to obtain their weights. The handledgroup was significantly tamer than the controlgroup, indicating that brief handling can promotetameness in parent-raised birds.
Iatrogenic causes of feather picking have alsobeen suggested, including feather trauma fromsloppy wing trims, small cages, or improperlyplaced perches (Davis 1991). Birds may also pickat injury sites (Rosenthal 1993).
With chronic feather picking, the initial precip-itating causes may no longer be present or rele-vant, but the abnormal behaviors persist (Cooper& Harrison 1994). Neurochemical abnormalitiesmay exist that support the persistence of these be-havioral patterns, even in the absence of stressorsand environmental deficits. Based on findings inother species, neurotransmitters of interest wouldinclude dopamine, serotonin, and opioids.
COMPARISON TO HUMANPSYCHIATRIC DISORDERS
Feather-picking disorder has been compared tohuman disorders involving obsessive-compulsive
258 Manual of Parrot Behavior

behavior and loss of impulse control. Obsessive-compulsive disorder (OCD) is classified by TheDiagnostic and Statistical Manual of Mental Dis-orders, 4th ed. (DSM-IV) as an anxiety disorder.Obsessive-compulsive disorders are characterizedby obsessions, intrusive thoughts or images, andcompulsions, repetitive behaviors performed inan attempt to prevent or reduce anxiety. Excessivehand washing is an example of a commonly re-ported compulsive behavior in humans. Stein(1996) and Stein et al. (1992) have suggested thathand washing in human OCD patients shares be-havioral similarities with repetitive grooming be-haviors in animals, including feather picking inbirds.
Bordnick et al. (1994) discuss the similaritiesbetween feather-picking disorder in birds and tri-chotillomania, or hair pulling, in humans. Tricho-tillomania is an impulse control disorder charac-terized by hair removal that results in alopecia. Itcan involve any region of the body where hairgrows. Tension increases as the individual at-tempts to resist the urge to pull hair. Manipulationof the hair is common, including hair twirling,chewing or mouthing the hair, and manual manip-ulation of removed hairs. Trichophagia may alsooccur. Similarly, birds with feather-picking disor-der have been reported to chew, manipulate, andsometimes consume removed feathers.
Garner et al. (2003) compared the stereotypicbehaviors of captive Orange-winged Amazon Par-rots with those of human patients suffering fromautism or schizophrenia. Performance of stereo-typic behaviors by the parrots correlated withpoor performance on a learning task in which thesubjects tended to repeat rather than switch incor-rect responses for subsequent trials. These re-sponses were consistent with the performance ofhuman patients with basal ganglia dysfunction.
TREATMENT OF FEATHER-PICKINGDISORDER
Treatment recommendations should be specificfor each individual based on a thorough historyand accurate assessment of the contributing fac-tors. Feather-picking disorder can be difficult totreat, and outcomes are often discouraging(Galvin 1983; Harrison 1994). Treatment shouldaddress underlying medical issues, nutritional de-ficiencies, environmental factors, behavior modi-fication, and, in some cases, restraint options.
Housing should address the needs of the birdand provide adequate stimulation as well as asense of safety. Cages that are kept in busy areasof the home may not provide adequate privacy,while cages kept in lower traffic areas may notprovide sufficient stimulation. Cages should belarge enough to allow movement, but all birdsshould have supervised time outside of the cage.Perches should provide stable surfaces in a vari-ety of diameters and materials. Perches should becarefully placed such that feathers are not dam-aged when the bird turns or moves in the cage.
Air quality can be addressed by providing reg-ular access to fresh air, use of air purifiers and hu-midifiers, and restricting the bird’s exposure tochemicals, scented oils, Teflon, and cigarettesmoke. The importance of regular bathing oppor-tunities for healthy skin and feathers has alsobeen discussed (Lawton 1996; Rosskopf & Woer-pel 1996).
Most birds will manipulate objects (toys), andpersonal preferences should dictate the types oftoys that are offered. Some toys should providethe bird with an opportunity to chew, such asrawhide, pieces of non-toxic wood, alfalfa cubes,or cardboard. Toys should not overcrowd the en-closure and can be rotated on a regular basis tomaintain the bird’s interest.
Foraging devices that require captive birds towork for food have been used for environmentalenrichment purposes (Coulton et al. 1997).Foraging enrichments that require subjects to ma-nipulate objects, sort through inedible materials,chew through barriers, and open containers werefound to reduce feather-picking behavior inamazon parrots (Meehan, Millam, & Mench2003). In addition to preventing the developmentof feather-chewing behaviors in the enrichedgroup, the same enrichments were used to reversethe development of feather picking in controlbirds. Environmental enrichments have beenfound to compensate for some of the effects ofearly maternal deprivation in rodent species(Bredy et al. 2003; Francis et al. 2002).
Treatment for feather-picking disorder may in-volve provision of appropriate social contacts.Meehan, Garner, and Mench (2003) found thatisosexual pairing of captive Orange-winged Ama-zon Parrots (Amazona amazonica) effected thedevelopment of abnormal behaviors. Pair-housedbirds used enrichment devices more than singly
23 / Feather-Picking Disorder in Pet Birds 259

housed cohorts and spent less time screaming, lesstime preening, and were more active. Pair-housingwas protective against the development of stereo-typic behaviors and neophobia. Local avian groupshave attempted to provide opportunities for social-ization with other birds through weekly or monthlygatherings of bird owners and their birds.
When reproductive causes are suspected, theenvironment can be modified to decrease repro-ductive behaviors. Shorter photoperiods, removalof toys or mirrors that promote masturbation orregurgitation, and separation from other nestingbirds have been suggested (Harrison & Davis1986). Interactions with the caregiver should belimited to activities that do not promote reproduc-tive behaviors. Providing an appropriate mate hasalso been suggested.
Sleep requirements need to be addressed for af-fected birds (Galvin 1983; Wilson 1999). A two-cage system, including a day cage and sleepingcage in a quiet room, will provide the bird with aquiet secluded sleeping area. A regular scheduleshould be maintained, with seasonal variationswhen appropriate.
Behavior modification exercises have been rec-ommended for the treatment of feather-pickingdisorder. Positive reinforcement of appropriatebehaviors, with food treats, praise, or an object re-ward, can be used when the bird is playing inde-pendently or manipulating appropriate objects.Training sessions to teach commands will providemental stimulation, increase bird-caregiver inter-actions, and allow the owner to redirect inappro-priate behaviors more effectively (Jenkins 2001).Desensitization and response substitution tech-niques can also be employed. Desensitizationinvolves benign gradual exposures to stress-inducing stimuli at a rate that does not provoke astress response. Desensitization techniques canbe combined with response substitution in whichthe bird is directed to perform an appropriate ac-tivity that is incompatible with the undesirablebehavior, such as performing commands, takingfood treats, or playing with a favorite toy.
Acupuncture has been used to treat feather-picking disorder. Worell and Farber (1993) treated28 birds (12 species) in an open trial. The subjectsunderwent weekly or biweekly acupuncture treat-ments, ranging from one to 42 treatments. Someof the subjects responded well to acupuncturetreatments, but more studies are needed to assess
required duration of treatment, frequency of treat-ment, and species differences in responses toacupuncture therapy.
The use of restraint devices has been advocatedby some and dismissed by others. Most practi-tioners agree that restraint devices should be fit-ted in the hospital to ensure that they can be wornsafely, and that they should be used only whennecessary to prevent serious self-injury (Lawton1996; Rosskopf & Woerpel 1996). Some birdsreact violently to restraint devices.
A variety of collars are available, includingElizabethan-style collars, made from radi-ographic film or plastic, and tube collars madefrom pipe insulator or acrylic (Figures 23.4, 23.5,23.6, and 23.7). The author has used a sweater-type restraint device made from a thick cottontube sock successfully in a variety of species.Cockatoos seem to be particularly accepting ofthis form of restraint and rarely need to be hospi-talized for fittings (Figure 23.8).
PHARMACOLOGICAL AGENTS
Few drugs have been systematically evaluated foradjunctive treatment of feather-picking disorder.They should be used with caution until more in-formation is available about toxicity, dosage re-quirements, and efficacy. Pharmacological thera-pies have historically been considered only as alast resort (Johnson-Delaney 1992). Dopamineantagonists, serotonergic agents, opioid antago-nists, tricyclic antidepressants, benzodiazepines,
260 Manual of Parrot Behavior
Figure 23.4. Combination collar constructed ofacrylic and plastic, weighing approximately 80grams.

261
Figure 23.5. Tube collar constructed from pipeinsulation weighing less than 10 grams.
Figure 23.6. Umbrella Cockatoo wearing thecollar in Figure 23.4.
Figure 23.7. Umbrella Cockatoo wearing thecollar in Figure 23.5.
Figure 23.8. Moluccan Cockatoo wearing asock sweater. See also color section.

and hormonal therapies have been used for thetreatment of feather picking, but few controlledstudies are available (Table 23-1). Welle (1998)has reviewed the psychotropic drugs used in avianspecies. Use of psychotropic drugs in psittacinespecies constitutes extra-label use and informedconsent of caregivers is recommended.
Serotonergic agents are the most commonlyprescribed pharmacological treatment for tricho-tillomania in humans and compulsive disorder innon-human species (Christianson & Crow 1996;Diefenbach et al. 2000). Fluoxetine (Prozac) is aselective serotonin reuptake inhibitor that en-hances serotonergic function via selective inhibi-tion of neurotransmitter reuptake at the serotonin
transporter. Fluoxetine has been used to treatfeather picking in birds at doses ranging from 1 to4 mg/kg q 24 h, but anecdotal reports would sug-gest that it is not very effective (Jenkins 2001).Fluoxetine, along with behavior and environmen-tal modifications, was used to treat repetitive toechewing in a Cockatiel at a dose of 1 mg/kg q 24h (Seibert 2004).
Tricyclic antidepressants enhance serotonergicand noradrenergic transmission by blocking theirreuptake into the presynaptic neuron. In addition,they produce anticholinergic and antihistaminiceffects. Doxepin (Sinequan) and amitriptyline(Elavil) have been used to treat feather-pickingdisorder.
262 Manual of Parrot Behavior
Table 23-1 Dosages for drugs used in the treatment of feather-picking disorder
Drug Dosage
Amitriptyline 1.0–5.0 mg/kg PO q 12–24 h (Lawton)
Butorphanol 0.2–1.0 mg/kg IMa
Chorionic gonadotropin (hCG) 500–1,000 IU/kg IM on days 1, 7, 14, 21, then q 28 da
Clomipramine 1 mg/kg PO q 12 hb
Diazepam 0.6 mg/kg IM; 1 mg/6 oz waterb
1.25–2.5 mg/120 ml watera
2.5–4.0 mg/kg PO q 6–8 ha
Diphenhydramine 2.0–4.0 mg/kg PO q 12 h0.5 mg/240 ml watera
Doxepin 0.5–1.0 mg/kg PO q 12 ha
Fluoxetine 0.4 mg/kg PO q 24 h (Lawton)2.0 mg/kg PO q 12 ha
Haloperidol 0.15–0.4 mg/kg PO q 12 h (Iglauer, Rasim)1.0–2.0 mg/kg IM q 21 da
Hydroxyzine 2.0–2.2 mg/kg PO q 8 ha
4 mg/120 ml watera
Leuprolide acetate 700–800 mcg/kg IM q 14 d for 3 doses, then q 4 mo
Naltrexone 1.5 mg/kg PO q 12 h (Turner)aCarpenter, Mashima, Rupiper, Exotic Animal Formulary, 2001.bAAHA Exotic Animal Formulary.

Clomipramine is a tricyclic antidepressant withhigh selectivity for serotonin reuptake inhibition. Itwas the first medication to be approved by theFood and Drug Administration (FDA) for the treat-ment of obsessive-compulsive disorder in humans.
Ramsey and Grindlinger (1992) used clomipra-mine to treat feather-picking disorder in an opentrial with 11 psittacine birds. Doses were gradu-ally increased to a maintenance dose of 0.5 mg/kgq 12 h. Only two of the 11 birds responded. ACongo African Grey Parrot (Psittacus erythacus)was treated for feather picking and self-mutilationwith an initial clomipramine dose of 4 mg/kg q 12h by Juarbe-Diaz (2000). The dose was eventuallyincreased to 9.5 mg/kg q 12 h. Feather regrowthoccurred, and the caregiver felt that the featherpicking was adequately controlled three monthsafter the final dosage adjustment.
Ritchie and Harrison (1994) recommendedusing a low dose of clomipramine initially with agradual increase over a four- to five-day period.However, they stated that clinical impressionssuggest that this drug is rarely effective in con-trolling mutilation behavior in birds.
A placebo-controlled clomipramine trial incockatoos with feather-picking disorder revealedsignificant improvement in treated birds at a doseof 3 mg/kg q 12 h (Seibert et al., in press). Noside effects were reported in cockatoos during thetrial. However, a Green-winged Macaw beingtreated by the author with clomipramine at a doseof 3 mg/kg q 12 h died on day 2 of treatment.
Haloperidol (Haldol) is a butyrophenone an-tipsychotic agent that exerts its behavioral effectsby blocking dopaminergic activity. Doses rangingfrom 0.1–0.4 mg/kg q 12 h have been used effec-tively (Iglauer & Rasim 1993; Lennox &VanDerHeyden 1993). It is recommended thatlower doses be used in cockatoos, African GreyParrots, and Quaker Parrots (0.05 mg/kg q 24 h).Side effects include sedation, agitation, inap-petence, incoordination, severe depression, ano-rexia, and hyperexcitability. An injectable form isavailable, haloperidol decanoate, and has beenused at a dose of 1 to 2 mg/kg IM q 14–21 d.
Opioid antagonists may improve feather-picking behavior through their ability to block therelease of endogenous opioids. Naltrexone(Trexan) was used to successfully treat featherpicking at a starting dose of 1.5 mg/kg q 12 h ina variety of species (Turner 1993).
Benzodiazepines have been used for the short-term alleviation of symptoms and to aid in accept-ance of restraint devices. Benzodiazepines exerttheir effects by binding to a site on the GABA re-ceptor and enhancing GABA-ergic activity.
Feather-picking disorder is a complicated syn-drome commonly seen in several pet bird species.Multiple etiologies should be considered beforedeveloping a treatment plan. Medical conditions,nutritional deficiencies, behavioral abnormalities,and environmental situations must be assessed foreach patient. Effective treatment depends on thor-ough consideration of relevant factors, treatmentof underlying or complicating medical issues,correction of environmental deficits, and imple-mentation of behavior modification exercises.The use of restraint devices should be reservedfor severe cases involving mutilation of soft tis-sue. Use of psychoactive medications should in-clude informed caregiver consent and carefulconsideration of potential risks. Several areaswarrant further investigation, such as the impactof early maternal deprivation on adult behavior,the safety of psychoactive drug use in psittacinespecies, and the risk factors for feather-pickingdisorder to guide preventive counseling.
REFERENCES
Aengus, W.L., and J.R. Millam. 1999. Taming parent-reared orange-winged amazon parrots by neonatalhandling. Zoo Biology 18:177–187.
American Psychiatric Association. 1994. Diagnosticand statistical manual of mental disorders, 4th ed. Washington, DC: American Psychiatric Asso-ciation.
Bauck, L. 1998. Psittacine diets and behavioral enrich-ment. Sem Avian Exotic Pet Med 7 (3):135–140.
Bordnick, P.S., B.A. Thyer, and B.W. Ritchie, BW.1994. Feather picking disorder and trichotillomania:An avian model of human psychopathology. J BehavTher and Exp Psychiat 25 (3):189–196.
Bredy, T.W. R.A. Humpartzoomian, D.P. Cain,and M.J.Meaney. 2003. Partial reversal of the effect of mater-nal care on cognitive function through environmen-tal enrichment. Neuroscience 118 (2):571–576.
Briscoe, J.A., L. Wilson, and G. Smith. 2001. “Non-medical risk factors for feather picking in pet par-rots.” Proc Assoc Avian Vet, p. 131.
Christianson, G.A., and S.J. Crow. 1996. The character-ization and treatment of trichotillomania. J ClinPsychiatry 57 (S8):42–49.
Colombini, S., C.S. Foil, G. Hosgood, and T.N. Tully.2000. Intradermal skin testing in Hispaniolan par-
23 / Feather-Picking Disorder in Pet Birds 263

rots (Amazona ventralis). Veterinary Dermatology11 (4):271–276.
Cooper, J.E., and G.J. Harrison. 1994. “Dermatology.”In Avian medicine: Principles and application, ed.B.W. Ritchie, G.J. Harrison, and L.R. Harrison, pp.607–639. Lake Worth, FL: Wingers Publishing Inc.
Coulton, L.E., N.K. Waran, and R.J. Young. 1997.Effects of foraging enrichment on the behavior ofparrots. Animal Welfare 6:357–363.
Davis, C.S. 1991. Parrot psychology and behaviorproblems. Vet Clin North Am Small Anim Pract 21(6):1281–1288.
Delius, J.D. 1988. Preening and associated comfort be-havior in birds. Ann NY Acad Sci 525:40–55.
Diefenbach, G.J., D. Reitman, and D.A. Williamson.2000. Trichotillomania: A challenge to research andpractice. Clin Psychol Rev 20 (3):289–309.
Francis, D.D., J. Diorio, P.M. Plotsky, and M.J. Meaney.2002. Environmental enrichment reverses the effectsof maternal separation on stress reactivity. Journalof Neuroscience 22 (18):7840–7843.
Fudge, A.M., and L. McEntee. 1986. “Avian giardiasis:Syndromes, diagnosis and therapy.” Proc AssocAvian Vet, pp. 155–164.
Galvin, C. 1983. “The feather picking bird.” In Currentveterinary therapy VIII small animal practice, ed. byR.W. Kirk, pp. 646–652. Philadelphia: W.B.Saunders Co.
Garner, J.P., C.L. Meehan, and J.A. Mench. 2003.Stereotypies in caged parrots, schizophrenia andautism: Evidence for a common mechanism. BehavBrain Res 145 (1–2):125–134.
Harrison, G.J. 1986. Feather disorders. Vet Clin ofNorth Am Caged Bird Med 14 (2):179–199.
Harrison, G.J. 1994. “Perspective on parrot behavior.”In Avian medicine: Principles and application, ed.B.W. Ritchie, G.J. Harrison, and L.R. Harrison, pp.96–108. Lake Worth, FL: Wingers Publishing Inc.
Harrison, G.J., and C. Davis. 1986. “Captive behaviorand its modification.” In Clinical avian medicineand surgery, ed. G.J. Harrison, pp. 20–28. Phila-delphia: W.B. Saunders Co.
Iglauer, F., and R. Rasim. 1993. Treatment of psy-chogenic feather picking in psittacine birds with adopamine antagonist. J Sm Anim Pract 34:564.
Jenkins, J. 2001. Feather picking and self-mutilation inpsittacine birds. Vet Clin N Amer Exotic Anim Prac 4(3):651–667.
Johnson-Delaney, C. 1992. “Feather picking: Diagnosisand treatment.” Proc Assoc Avian Vet, p. 82.
Juarbe-Diaz, S.J. 2000. Animal behavior case of themonth. J Am Vet Med Assoc 216 (10):1562–1564.
Koski, M.A. 2002. Dermatologic diseases in psittacinebirds: An investigational approach. Sem AvianExotic Pet Med 11 (3):105–124.
Lawton, M.P.C. 1996. “Behavioural problems.” InBSAVA Manual of Psittacine Birds, ed. P.H. Beynon,N.A. Forbes, and M.P.C. Lawton, pp. 106–114.Ames: Iowa State University Press.
Lennox, A.M., and N. VanDerHeyden. 1993. “Halo-peridol for use in treatment of psittacine self-mutilation and feather plucking.” Proc Assoc AvianVet, pp. 119–120.
Lucas, A.M., and P.R. Stettenheim. 1972. Avian anat-omy: Integument. Washington, DC: U.S. Govern-ment Printing Office.
Macwhirter, P., R. Mueller, and J. Gill. 1999. “Ongoingresearch report: Allergen testing as a part of diag-nostic protocol in self-mutilating psittaciformes.”Proc Assoc Avian Vet, pp. 125–128.
Meehan, C.L., J.P. Garner, and J.A. Mench. 2003.Isosexual pair housing improves the welfare ofyoung amazon parrots. Appl An Behav Sci 81:73–88.
Meehan, C.L., J.R. Millam, and J.A. Mench. 2003.Foraging opportunity and increased physical com-plexity both prevent and reduce psychogenic featherpicking by young amazon parrots. Applied AnimalBehavior Science 80 (1):71–85.
Nett, C.S., and T.N. Tully. 2003. Anatomy, clinical pres-entation, and diagnostic approach to feather pickingpet birds. Compendium 25 (3):206–218.
Oppenheimer, J. 1991. “Feather picking: Systemic ap-proach.” Proc Assoc Avian Vet, pp. 314–315.
Perry, R.A. 1994. “Perspective on parrot behavior.” InAvian medicine: Principles and application, ed.B.W. Ritchie, G.J. Harrison, and L.R. Harrison, pp.96–108. Lake Worth, FL: Wingers Publishing Inc.
Ramsey, E.C,, and H. Grindlinger. 1992. “Treatment offeather picking with clomipramine.” Proc AssocAvian Vet, pp. 379–382.
Ritchie, B.W., and G.J. Harrison. 1994. “Formulary.” InAvian medicine: Principles and application, ed.B.W. Ritchie, G.J. Harrison, and L.R. Harrison, p.461. Lake Worth, FL: Wingers Publishing Inc.
Rosenthal, K. 1993. “Differential diagnosis of featherpicking in pet birds.” Proc Assoc Avian Vet, Nash-ville, TN, pp. 108–112.
Rosskopf, W.J., and R.W. Woerpel. 1991. Pet avian con-ditions and syndromes of the most frequently pre-sented species seen in practice. Vet Clinics N Am SmAnim Pract 21 (6):1189–1211.
Rosskopf, W.J., and R.W. Woerpel. 1996. “Featherpicking and therapy of skin and feather disorders.”In Diseases of cage and aviary birds, 3rd ed., ed.W.J. Rosskopf and R.W. Woerpel, pp. 397–405.Baltimore: Williams and Wilkins.
Ruppenthal, G.C., G.L. Arling, H.F. Harlow, G.P.Sackett, and S.J. Suomi. 1976. A 10-year perspectiveof motherless-mother monkey behavior. Journal ofAbnormal Psychology 85 (4):341–349.
264 Manual of Parrot Behavior

Seibert, L.M. 2004. Animal behavior case of themonth. J Am Vet Med Assoc 224 (9):1433–1435.
Seibert, L.M., Crowell-Davis, S.L., G.H. Wilson, andB.W. Ritchie. In press. Placebo-controlled clomi-pramine trial for the treatment of feather picking dis-order in cockatoo (Cacatua) species. J Am AnimHosp Assoc.
Spruijt, B.M., J.A. Van Hooff, and W.H. Gispen. 1992.Ethology and neurobiology of grooming behavior.Physiol Rev 72 (3):825–852.
Stein, D.J. 1996. The neurobiology of obsessive-compulsive disorder. Neuroscientist 2 (5):300–305.
Stein, D.J., N. Shoulberg, K. Helton, and E. Hollander.1992. The neuroethological approach to obsessive-compulsive disorder. Comp Psychiatry 33 (4):274–281.
Suomi, S.J., M.L. Collins, H.F. Harlow, and G.C. Rup-penthal. 1976. Effects of maternal and peer separa-tions on young monkeys. Journal of Child PsycholPsychiatry 17 (2):101–112.
Turner, R. 1993. “Trexan (naltrexone hydrochloride)use in feather picking in avian species.” Proc AssocAvian Vet, pp. 116–118.
VanHoek, C.S., and C.E. King. 1997. Causation and in-fluence of environmental enrichment on featherpicking of the crimson-bellied conure (Pyrrhuraperlata perlata). Zoo Biol 16 (2):161–172.
Welle, K.R. 1998. “A review of psychotropic drug ther-apy.” Proc Assoc Avian Vet, pp. 121–124.
Welle, K.R. 1999. “Clinical approach to feather pick-ing.” Proc Assoc Avian Vet, pp. 119–124.
Wilson, L. 1999. Sleep: How much is enough for a par-rot? Pet Bird Report 43:60–62
Worell, A.B., and W.L. Farber. 1993. “Use of acupunc-ture in the treatment of feather picking inpsittacines.” Proc Assoc Avian Vet, Nashville, TN,pp. 121–126.
23 / Feather-Picking Disorder in Pet Birds 265

INTRODUCTION
Behavioral problems are one of the leadingcauses of pet relinquishment and/or euthanasia.The use of behavioral drugs has been advised inthe past to be considered only as a “last resort.”[1]It was believed that once on medication, birdswould have to remain on that drug permanently.In reality, appropriate usage of behavioral drugsmay mean the difference between a healthyhuman/bird bond and relinquishment or euthana-sia. Psittacines with behavioral disorders oftenbenefit from timely and appropriate use of behav-ioral drugs. When triggers are identified, anddrugs are used as an adjunct to behavioral and en-vironmental modification, birds may be success-fully weaned off medication.
MECHANISM OF ACTION
Most behavioral drugs exert their effects throughactions on neurotransmitters in the central nerv-ous system. Neurotransmitters are responsible fortransmission of impulses between neuronal andnon-neuronal cells. Alterations in neurotransmit-ter levels and receptors can lead to neurologicaland behavioral disorders. Behavioral drugs can beused to modify these neurotransmitters and resultin physiological and behavioral changes. Under-standing basic neurotransmitter pathophysiologyand how alterations in the levels of neurotransmit-ters affect behavior is beneficial in selection ofbehavioral drugs.
Neurotransmitters are released from the pre-synaptic neuron into the synaptic cleft and act onthe postsynaptic cell. Stimulation of neurotrans-mitter receptors on the postsynaptic cell can haveexcitatory or inhibitory effects. Continuous stim-
ulation by neurotransmitters or drugs (agonists)can hyposensitize or down-regulate receptors,while receptors that are not stimulated by neuro-transmitters or blocked by drugs (antagonists) canbecome hypersensitive or up-regulated. Someneurotransmitters can also act on distant targetsand thus act as neurohormones. Depending on theneurotransmitter, enzymes in the synaptic cleft orin the presynaptic neuron may inactivate theseneurotransmitters. Reuptake inhibitors block re-uptake and may prolong the effect of the neuro-transmitter. This is the basis for the proposedmechanism of action of psychoactive drugs.Specific mechanisms of action are largely un-known.
NEUROTRANSMITTERS
Neurotransmitters of importance are serotonin(5HT or 5-hydroxytryptamine), norepinephrine(NE), epinephrine (E), dopamine (DA), gamma-aminobutyric acid (GABA), and acetylcholine(Ach). Serotonin, norepinephrine, and dopamineare classified as monoamines.
Monoamines may be divided into two classes,catecholamines and indoleamines. They are con-centrated in the midbrain, hypothalamus, and lim-bic system. The catecholamines include norepi-nephrine, epinephrine, and dopamine. They arederived from the amino acid tyrosine. The in-doleamines include serotonin and melatonin andare derived from dietary tryptophan.
Serotonin
Serotonin is found mainly in cells of the midlineraphe and plays a role in the sleep-wake cycle,mood and emotion, and the suppression of impul-
267
24Psittacine Behavioral
Pharmacotherapy
Kenneth M. Martin

sive behavior. Serotonin is inactivated by reup-take or breakdown by monoamine oxidase(MAO). Serotonin depletion has been associatedwith depression, increased anxiety, irritability andaggression, and impulsiveness. Increasing or nor-malizing brain serotonin levels is beneficial incompulsive and stereotypic disorders and someforms of anxiety and aggression. Serotoninexcess has been associated with confidence,calmness, flexibility, and resilience. Selectiveserotonin reuptake inhibitors and tricyclic antide-pressants increase serotonin availability by de-creasing reuptake.
Norepinephrine
Norepinephrine is formed from the hydroxylationof dopamine. Norepinephrine is broken down bymonoamine oxidase A (MAO A) and by catechol-O-methyltransferase (COMT). Norepinephrinedepletion has been associated with depression,while excess has been associated with mania.Drugs that inhibit MAO and norepinephrine re-uptake are beneficial in treating depression inhumans.
Epinephrine
Epinephrine is secreted from the adrenal gland inresponse to norepinephrine to cause sympatheticeffects. There are alpha and beta adrenergic re-ceptors. Alpha adrenergic activation leads tovasoconstriction, increased cardiac contraction,iris dilation, intestinal relaxation, pilomotor con-traction, contraction of the bladder and intestinalsphincters, and inhibition of the parasympatheticnervous system.[2] Beta one adrenergic stimula-tion increases cardiac output, while beta twoadrenergic stimulation causes vasodilation, bron-chodilation, intestinal relaxation, uterine relax-ation, and dilation of coronary blood vessels.[2]Beta blocking drugs are beneficial in reducing thephysiological signs of fear.
Dopamine
Dopamine is produced from L-dopa and stored inthe presynaptic vesicles. Dopamine is a precursorfor norepinephrine. Dopamine is inactivated bymonoamine oxidase (primarily MAO B) and bycatechol-O-methyltransferase (COMT). Dopa-mine depletion produces behavioral quieting,depression, and extrapyramidal signs (muscle tre-mors, tics, and motor restlessness), while dopa-
mine excess has been associated with compulsiveand stereotypic behavior.
Gamma-Aminobutyric Acid
Gamma-aminobutyric acid (GABA) is an in-hibitory neurotransmitter that is widely distrib-uted in the central nervous system. It is synthe-sized from the amino acid glutamate. GABAdepletion and dysregulation has been associatedwith seizure activity, Parkinson’s disease, andfears and phobias. Benzodiazepines and barbitu-rates are GABA agonists and are useful in treat-ing these disorders.
Acetylcholine
Acetylcholine is the most widely distributed neu-rotransmitter in the body and is excitatory infunction. Acetylcholine is synthesized from ac-etate and choline and inactivated by acetyl-cholinesterase. Acetylcholine activates two differ-ent types of receptors referred to as muscarinicand nicotinic receptors. Muscarinic receptors arefound in cells stimulated by the postganglionicneurons of the parasympathetic nervous systemand postganglionic cholinergic neurons of thesympathetic nervous system.[2] Muscarinic stim-ulation leads to arteriole vasodilation, decreasedheart rate and cardiac output, and stimulation ofthe digestive system. Nicotinic receptors arefound in the synapses between pre- and postgan-glionic neurons of the sympathetic and parasym-pathetic nervous system.[2] Depletion of acetyl-choline is associated with cognitive decline andAlzheimer’s disease. Blockade of muscariniccholinergic receptors is responsible for anti-cholinergic side effects (dry mouth, dry eye, pupildilation, tachycardia, constipation, and urinary re-tention) of tricyclic antidepressants.
DRUG SELECTION ANDCONSIDERATION
There are currently no behavioral drugs licensedfor use in psittacine birds. Owners should be in-formed of use that is considered “extra-label”with respect to the manufacturer’s recommenda-tions and sign an appropriate consent form beforethe drug is dispensed. In addition, many drugsmust be compounded before they can be adminis-tered to avian patients. The effects of compound-ing on storage and stability must be considered.Administration of behavioral drugs can also be
268 Manual of Parrot Behavior

problematic, depending on the owner’s ability tohandle the avian patient and/or the bird’s failureof oral acceptance of the drug. Taste aversion andthe inability of masking bitter-tasting drugs withfruit flavorings may be encountered. Often, drugsmay be injected into fruits, such as a grape, ormixed with yogurt or fruit-based baby food to fa-cilitate oral administration.
Dosages and regimens of behavioral drugs areoften extrapolated from veterinary and human lit-erature and anecdotal reports in avian species. Formany drugs, accurate dosages and controlled stud-ies have not been performed. Dosage variation canbe found in the veterinary literature. Wheneverpossible, drug dosages and regimens given in thischapter will be based on veterinary references (seeTable 24-1). Ultimately, the prescribing veterinar-ian is responsible for appropriate selection,dosage, and administration of behavioral drugs inthe specific clinical situation and patient.
Drug selection requires an accurate diagnosisof the behavioral problem and knowledge of drugefficacy and safety in the human and veterinaryliterature. Veterinarians should become familiarwith indications, contraindications, proposed me-chanism of action, common side effects, and thepotential for adverse effects before using behav-ioral drugs. The bird species, age, sex, health, thecost of the medication, and ease of administeringthe medication are important aspects of drug se-lection. The safest drug that will be effective intreating the behavioral problem should be usedfirst. If drug treatment fails or unacceptable sideeffects occur, one should consider dosage adjust-ment, if the drug was used for an adequate dura-tion for effect, and re-evaluate the original diag-nosis. The benefit/risk ratio of other drug optionsmay be considered.
Ideally, a complete blood count and general bio-chemistry should be preformed to rule out under-lying medical problems and establish a baselinefor future testing. Regular monitoring of bloodwork should be performed based of the bird’shealth and the potential for adverse effects fromthe drug. Medical conditions should be treatedconcurrently with any behavioral problems. Longafter the medical condition has resolved, behav-ioral disorders may continue. This is especiallytrue for compulsive disorders. In addition, medi-cal conditions lower the tolerable threshold foranxiety-related behavioral disorders.
Behavioral drugs should be used only as an ad-junct to behavioral and environmental modifica-tion. Lack of concurrent behavioral and environ-mental modification may lead to drug treatmentfailure. The drug may become less effective inmodifying behavior with time. The first timeusing a drug with behavioral modification is thebest chance for cure. In addition, without concur-rent behavioral and environmental modification,the problem is likely to reoccur when the drug isdiscontinued.
Drug desensitization is often used when thestimulus cannot be controlled or avoided, or whenthere is overwhelming fear/anxiety or aggression.In this manner, behavioral drugs are used to facil-itate learning. The drug is used to decrease arou-sal and/or anxiety to a level that allows learning tocope with the stimuli or situation. After a suffi-cient learning period has occurred, the drugshould be weaned gradually. Abrupt discontinu-ance is likely to result in withdrawal syndromecharacterized by rebound anxiety and/or aggres-sion and reoccurrence of the behavioral problem.Generally, it is recommended that these drugs beweaned by reducing the total daily dosage by 25%every three weeks while maintaining the samedosing frequency. If during the weaning processbehavioral problems reoccur, the dosage shouldbe increased to the previous effective dose andtreatment duration should be extended.
DRUG CLASSES
Benzodiazepines
Benzodiazepines (BDZ) potentiate the effects ofGABA, an inhibitory neurotransmitter, and arebeneficial in treating conditions of fear, phobia,and anxiety, including fear aggression. Benzo-diazepines have been used to facilitate social inter-action and increase friendliness. Caution shouldbe taken when used in aggression cases. Fear usu-ally is conducive to defensive aggression, and ben-zodiazepines have the potential to disinhibit ag-gression, making the aggression more offensive.Benzodiazepines also have anticonvulsant effectsfor the treatment of acute seizure disorders. Theyhave a rapid onset of action. Side effects includesedation, ataxia, muscle relaxation, increased ap-petite, paradoxical excitation, and memorydeficits. Benzodiazepines may interfere withlearning and affect behavioral modification and
24 / Psittacine Behavioral Pharmacotherapy 269

270
Table 24-1 Psittacine behavioral pharmacotherapy
Drug Class Dosage Route Indications
Diazepam BDZ 2.5–4.0 mg/kg q6–8h34 PO Sedation(Valium®, Roche, 1.25–2.50 mg/120 ml PO Acute fearsNutley, NJ, USA) water14 Acute anxiety FPD
0.6 mg/kg q8–12h5 IM, IV AnticonvulsantAmitriptyline TCA 1–5 mg/kg q12h7 PO Pruritic FPD
(Elavil®, AstraZeneca, Chronic anxiety Wilmington, DE, USA) disorders
Doxepin TCA 1–2 mg/kg q12h7 PO Pruritic FPD (Sinequan®, Pfizer, (preferred drug)New York, NY, USA) 1–2 mg/lb q 12h40 Chronic anxiety
disordersClomipramine TCA, 3–5 mg/kg q12–24h7 PO Compulsive disorder
(Anafranil® or Selective Compulsive FPDClomicalm®, Novartis, Chronic anxiety East Hanover, NJ, disordersUSA)
Nortriptyline (Pamelor®, TCA 2 mg/120 ml drinking PO Chronic anxiety Sandoz, East Hanover, water14 disordersNJ, USA) Anxiety-induced FPD
(seldom used)Fluoxetine (Prozac®, SSRI 1–4 mg/kg q24h7 PO Compulsive disorder
Eli Lilly, Indianapolis, Dosages have been used Compulsive FPDIN, USA) up to 20 mg/kg/day7 Global fear, phobia
AggressionParoxetine (Paxil™, SSRI 1–2 mg/kg q12–24h16 PO Compulsive disorder
GlaxoSmithKline, Compulsive FPDTriangle Research Global fear, phobiaPark, NC, USA) Aggression (preferred
drug)Chlorpromazine Antipsychotic 0.1–0.2 mg/kg once14 IM Tranquilization
(Thorazine®, Low potency 1 ml solution X/4 oz PO Compulsive FPD used SmithKline Beecham, drinking water14 with CarbamazepinePhiladelphia, PA, USA) 0.2–1.0 ml/kg solution PO Solution X = 125 mg
X q12–24h14 chlorpromazine/31ml simple syrup14
Haloperidol (Haldol®, Antipsychotic 0.2–0.9 mg/kg q24h7 PO Compulsive FPDOrthoMcNeil, DA antag- 1–2 mg/kg q4wk24 IM Self-mutilation Raritan, NJ, USA) onist 6.4 mg/L drinking PO (preferred drug)
High potency water19
0.08 mg/kg sid5 POLower dosage in Quaker
Parakeets and cockatoos41
Diphenhydramine Antihistamine 2–4 mg/kg q12h24,41 PO Pruritic FPD(Benadryl®, Parke- 0.5 mg/240 ml drinking Davis, Morris Plains, water14
NJ, USA)

271
Hydroxyzine (Atarax®, Antihistamine 2 mg/kg q12h14 PO Pruritic FPDPfizer, New York, NY, USA)
Phenobarbital (Solfoton®, Anticonvulsant 1–7 mg/kg q8–12h24 PO Seizure disorderECR Pharm, Richmond, 6–10 mg/120 ml POVA, USA) drinking water14
Carbamazepine Anticonvulsant 3–10 mg/kg q24h14 PO Seizure disorder(Tegretol®, Novartis, 20 mg/120 ml PO Compulsive/impulsive East Hanover, NJ, USA) drinking water14 disorder
Naltrexone (Revia®, Narcotic an- 1.5 mg/kg q12h32 PO Acute compulsive DuPont, Wilmington, tagonist FPDDE, USA)
Butorphanol (Torbugesic®, Narcotic 2–4 mg/kg q8h33 PO, IM, Moderate/severe painFort Dodge, IA, USA) agonist IV (preferred drug)
Buprenorphine Narcotic 0.1 mg/kg q12h34 IM, IV Moderate/severe painBuprenex®, Reckitt & agonistColman, Richmond, VA, USA)
Carprofen (Rimadyl®, NSAID 2–10 mg/kg q24h34 SQ, IM, Mild/moderate painPfizer, New York, IVNY, USA)
Ketoprofen (Orudis®, NSAID 2 mg/kg q24h34 IM Mild/moderate painWyeth-Ayerst, Philadelphia, PA, USA, or Ketofen®,Fort Dodge, Fort Dodge, IA, USA)
Chorionic gonadotropin Hormone 500–1,000 U/kg IM Hormonal FPD(APL®, Wyeth-Ayerst, q4–6wk35 Chronic egg pro-Philadelphia, PA, USA) duction
Sexual behaviorLeuprolide acetate Hormone 100–1,000 ug/kg IM Hormonal FPD
(Lupron®, Tap Pharma- q2wks36 (for Chronic egg pro-ceuticals, Lake Forest, 3 treatments) ductionIL, USA) �300g bird—750ug/kg Sexual behavior
>300g bird—500ug/kg (antiandrogenic)Medroxyprogesterone Hormone 5–50 mg/kg q4–6wk34 SQ, IM Chronic egg pro-
acetate (Depo-Provera®, ductionPharmacia & Upjohn, 150g bird—0.05mg/g Sexual behavior Kalamazoo, MI, USA) 300–700g bird— (suggest use once)
0.03mg/g>700g bird—0.025mg/g
Table 24-1 Psittacine behavioral pharmacotherapy (continued)
Drug Class Dosage Route Indications

training. At low dosages, they act as sedatives, atmoderate dosages they have anxiolytic effects, andat high dosages benzodiazepines act as hypnoticsand facilitate sleep. Tolerance to sedation, ataxia,and muscle relaxation may develop with time.There is the potential for injury to birds fromfalling when perched. Generally, benzodiazepinesappear to be very safe drugs, but idiosyncratic he-patic failure has been reported in cats.[3]
Benzodiazepines are best used for short-termtreatment of acute and intermittent behavioraldisorders. When not used as a monotherapy, ben-zodiazepines may be combined with tricyclic an-tidepressants or selective serotonin reuptake in-hibitors. Dosages should be adjusted accordingly.When prescribing benzodiazepines, one mustconsider that they are controlled substances andhave the potential for human abuse.
Diazepam (Valium®)
Diazepam is available as an injectable solution (5mg/ml) for IM or IV administration or as a solu-tion (1 or 5 mg/ml) for PO administration. It isalso available in 2 mg, 5 mg and 10 mg tablets forcompounding. Three mg of diazepam is soluble in1 ml of water. Diazepam is tasteless initially, but abitter aftertaste soon develops. Diazepam may beused to control seizure disorders. It can be used totreat fears, phobias, anxiety, and possibly fear ag-gression. Diazepam is helpful in treating acuteseizure disorders (0.05–2.0 mg/kg IM or IV).[4]Diazepam may be beneficial in acute anxiety-induced feather picking that is not compulsive innature (0.6 mg/kg IV, IM).[5] Diazepam may alsobe useful to facilitate tolerance to Elizabethan col-lars in acute cases of self-mutilation. When usedin combination therapy with tricyclic antidepres-sants or selective serotonin reuptake inhibitors thestarting dosage should be reduced. Caution is war-ranted in patients with hepatic disease.
Antidepressants
Antidepressant drug classes include tricyclics, se-lective serotonin reuptake inhibitors, and mono-amine oxidase inhibitors. To the author’s knowl-edge, MAO inhibitors have not been used in avianpatients. Indications and side effects vary basedon sensitivity to and specificity for neurotrans-mitters. Generally, these drugs are best used totreat chronic anxiety disorders and cannot be usedon an “as needed” basis.
Tricyclic Antidepressants
Tricyclic antidepressants (TCAs) block the reup-take of norepinephrine and serotonin and arecompetitive antagonists at the muscarinic acetyl-choline, histamine H1, and alpha1 and alpha2adrenergic receptors. Increasing the availabilityof 5HT, NE, and DA, TCAs are indicated fortreating fear, phobia, anxiety, and aggression.Chronic neuropathic pain may also be amelio-rated by TCAs. The onset of action is usually de-layed, with clinical effects usually seen by three tofour weeks. Clomipramine is most selective forserotonin reuptake inhibition, making it the onlytricyclic effective in treating canine compulsivedisorders.[6] Tricyclic antidepressants producevarious degrees of sedation based on anticholin-ergic and antihistaminic effects. Amitriptylineand doxepin produce the strongest antihistaminiceffects, making them useful when sedation andantipruritic effects are desired. Sedation, tasteaversion, gastrointestinal upset, and anticholiner-gic side effects may occur, precluding use. Theyare contraindicated with alterations of blood glu-cose, glaucoma, seizures, cardiac disease, andconcurrent use of thyroid medication. Exercisecaution with concurrent use of serotonergicdrugs.
Amitriptyline (Elavil®)
Amitriptyline is available as a 10 mg, 25 mg, 50mg, 75 mg, 100 mg, and 150 mg tablet for oraladministration, as well as an injectable (10mg/ml) for IM administration. Amitriptyline hasbeen suggested for feather picking, yet is rarelyeffective.[5] The drug is ineffective in treatingdisorders of compulsive etiology. It is effective intreating global fear, separation anxiety, and gen-eralized anxiety disorders. Amitriptyline is a po-tent H1 blocker secondary to doxepin and may bebeneficial in cases of pruritis. Amitriptyline isfreely soluble in water or alcohol and has a bitterand burning taste. Amitriptyline inhibits 5HT andNE reuptake. Nortriptyline is the active metabo-lite. Suggested dosage is 1–5 mg/kg PO bid.[7]
Doxepin (Sinequan®)
Doxepin is available in 10 mg, 25 mg, 50 mg, 75mg, 100 mg, and 150 mg capsules for oral admin-istration, as well as a 10 mg/ml oral suspension. Itis freely soluble in alcohol. Doxepin is a moder-
272 Manual of Parrot Behavior

ate inhibitor of NE and a weak inhibitor of 5HT.Doxepin is the most antihistaminic TCA and maybe effective in treating pruritic feather picking. H1antagonism of doxepin is 800 times more potentthan the antihistaminic effect of diphenhy-dramine.[1] Doxepin is the most sedating TCAand should be the antipruritic drug of choice.Anecdotal reports in African Grey Parrots andcockatoos suggest the drug is beneficial in reduc-ing agitation, aggression, and fear, while increas-ing appetite, friendliness, play, and vocalizationsat a dosage of 0.5–1.0 mg/kg PO bid.[8] Sug-gested dosage ranges from 1–4 mg/kg PO bid.Use the lowest effective dosage to control thebehavior.
Clomipramine (Anafranil® or Clomicalm®)
Clomipramine is available in 25 mg, 50 mg, and75 mg oral capsules. Clomicalm® is available in20 mg, 40 mg, and 80 mg tablets. It is freely sol-uble in water and bitter tasting. Clomicalm, whichis approved for treating separation anxiety indogs, is contraindicated for treating aggressionper drug label and has been known to increase ag-gression. Clomipramine has been used in humanand veterinary patients to effectively control com-pulsive disorders.[9, 10] In humans, clomipra-mine was the first drug to be FDA approved fortreating obsessive-compulsive disorders. Clomi-pramine is the most serotonin selective of theTCAs. It may be effective in treating compulsivefeather picking and less effective when there is acomponent of self-mutilation. Sedation and re-gurgitation may be a common side effect. Regur-gitation may be prevented if given with food.Bitter taste may prevent oral administration insome birds. One study found clomipramine (1.0mg/kg PO sid, or divided bid) effective in onlythree of 11 feather-picking psittacines treated forfour weeks; however, all birds had a positive atti-tude change.[11] Another placebo-controlledstudy found clomipramine (3 mg/kg PO bid)effective in eight of 11 treated feather-pickingcockatoos (six pickers, five mutilators) at sixweeks.[12] Seven of the eight birds began to im-prove at three weeks, with significantly greaterimprovement noted at six weeks. One clomipra-mine treated bird (mutilator) was worse at sixweeks, and the remaining two treated birds wereunchanged. No adverse events were reported dur-ing the study period. A raspberry syrup and 2%
carboxymethyl cellulose suspension of clomipra-mine (4 mg/ml) was used. This study suggests thedrug may be beneficial at higher dosages and fora longer duration of treatment. Suggested dosageis 3–5 mg/kg PO sid to bid.[7]
Nortriptyline (Pamelor®)
Nortriptyline is available as a 10 mg, 25 mg, 50mg, and 75 mg capsule or as a 10 mg/5 ml sus-pension for oral administration. Nortriptyline hasbeen used in humans to treat depression. It maybe beneficial in some cases of feather picking thatare anxiety induced. Hyperactivity is a commonside effect and dosage should be adjusted accord-ingly. Suggested dosage is 1 ml (10 mg/5 ml sus-pension) per 4 ounces drinking water or 2 mg/120ml drinking water.[13, 14]
Selective Serotonin Reuptake Inhibitors
Selective serotonin reuptake inhibitors (SSRIs)increase the availability of serotonin by blockingreuptake. They are indicated for compulsive dis-orders, fear, phobia, anxiety, and aggression.Because of their mood-stabilizing effect, SSRIsshould be the drugs of choice for affective oranxiety-induced aggression. SSRIs are believedto be the least likely drugs to disinhibit aggres-sion. Onset of action is believed to be delayed,reaching peak in three to four weeks. Disorders ofcompulsive etiology may take longer to respond.SSRIs have a safer side effect profile than theTCAs, with minimal anticholinergic effects. Sideeffects may include sedation and gastrointestinalsigns such as anorexia and nausea. Contraindica-tions include seizures and alterations in bloodglucose. Caution needs to be exercised when usedconcurrently with serotonergic drugs.
FLUOXETINE (PROZAC®)Fluoxetine is available in 10 mg, 20 mg, and 40mg capsules, 10 mg and 20 mg tablets, and in a 4mg/ml mint-flavored liquid for oral administra-tion. Approximately 50 mg is soluble in 1 ml ofwater. Fluoxetine is odorless and tasteless. Fluox-etine is highly selective for serotonin and has lit-tle effect on other neurotransmitters, such as nor-epinephrine and dopamine. Fluoxetine is the leastsedating SSRI, yet in some cases may lead to in-creased anxiety and agitation. Norfluoxetine isthe active metabolite. Fluoxetine (2–3 mg/kg sidto 3 mg/kg bid) was evaluated in 24 feather-
24 / Psittacine Behavioral Pharmacotherapy 273

picking psittacines of various species.[15] In all12 psittacines that completed the trial, significantimprovements were noted after two weeks. Allbirds relapsed after four weeks, yet respondedpositively to a dosage increase at that time.Benefits only lasted a limited time, requiring in-creased dosage. Temporary ataxia and lethargywere reported. Suggested dosage is 1–4 mg/kgPO sid adjusted to effect. Dosages up to 20mg/kg/day have been used.[7]
PAROXETINE (PAXIL™)Paroxetine is available in a 10 mg, 20 mg, 30 mg,and 40 mg tablet, as well as a 2 mg/ml orange-flavored suspension for oral administration.Paroxetine is a highly selective 5HT reuptake in-hibitor. Minimal anticholinergic effects have beennoted with usage, making it more sedating thanfluoxetine. Paroxetine is beneficial in treatingcompulsive disorder, social phobia, and panic andanxiety disorders in people and animals. The au-thor has found paroxetine extremely beneficial intreating phobic psittacines. It is the author’s pre-ferred drug for treating compulsive feather pick-ing. Suggested dosage is 1–2 mg/kg PO sid tobid.[16]
Antipsychotics
Antipsychotics have also been referred to as neu-roleptics or dopamine receptor antagonists. Anti-psychotics have been used in humans to treat schi-zophrenia and other psychotic disorders, such asmania, severe agitation, and violent behavior.[17]Antipsychotics are classified based on chemicalstructure and potency. Low-potency antipsy-chotics are commonly used in veterinary medicineas tranquilizers. High-potency antipsychotics(e.g., haloperidol) are probably more effective intreating behavioral disorders. The antipsychotichaloperidol has been used experimentally to re-duce compulsive behavior in many animal spe-cies.[10] High-potency dopamine antagonistshave been used successfully to treat compulsivefeather-picking disorders in psittacids.[18–20]Potential side effects include hypotension, de-creased seizure threshold, bradycardia, ataxia, ex-trapyramidal signs such as muscle tremors or tics,and motor restlessness. Low-potency antipsy-chotics (e.g., chlorpromazine) have more non-neurological (cardiotoxic, epileptogenic) side ef-fects and are more sedating. High-potency
antispychotics (e.g., haloperidol) cause more neu-rological side effects and are least sedating.
Chlorpromazine (Thorazine®)
Chlorpromazine, a phenothiazine, is available in a2 mg/ml (syrup) oral suspension, 25 mg/ml injec-tion, and 10 mg, 50 mg, 100 mg, and 200 mgtablets. One gram is soluble in 1 ml of water and1.5 ml of alcohol. Idiosyncratic aggression hasoccurred in dogs with the use of the pheno-thiazine acepromazine. Dosages necessary totreat aggression suppress all other forms of be-havior, including social and exploratory behavior.Phenothiazines are a poor choice for aggressionin animals. In addition, phenothiazines lackspecificity as dopamine antagonists and interactwith serotonin and norepinephrine. Chlorpro-mazine’s major effect is sedation. Anticholiner-gic side effects and extrapyramidal reactions(Parkinson’s-like side effects) may occur. Ataxia,regurgitation, and drowsiness have been reportedin birds. Chlorpromazine has been suggested forfeather-picking birds.[14] It is suggested to dis-continue usage within 30 days. Efficacy report-edly diminishes in 14–30 days when given orally.In cockatoos, it has been suggested for use onceIM in combination with the anticonvulsant carba-mazepine.[14]
Haloperidol (Haldol®)
Haloperidol, a butyrophenone, is available as a 2mg/ml solution for oral administration and a 50 or100 mg/ml injectable decanoate for IM adminis-tration. Haloperidol is colorless, odorless, taste-less, and water soluble. Haloperidol has been ef-fective in treating compulsive and aggressivestates in people and animals. Haloperidol hasbeen used effectively to treat compulsive featherpicking.[18–20] It appears to work best in cocka-toos and in cases of self-mutilation, suggestingdifferent etiologies in various species.[21] The ef-fect of the injectable decanoate may last three tofour weeks. Quaker Parakeets and Umbrella andMoluccan Cockatoos appear to be sensitive,therefore lower dosages should be used (0.08mg/kg sid).[5] Anorexia, ataxia, or vomiting mayoccur but usually resolve in 24–48 hours.[18]Dosage should be increased or decreased 0.01 mlevery two days to effect. Successful long-termusage (seven to nine years) to control featherpicking has been reported in a Moluccan Cock-
274 Manual of Parrot Behavior

atoo and Yellow-naped Amazon Parrot with fewside effects.[18] Similarly, two African GreyParrots with feather-picking disorder were suc-cessfully treated with haloperidol for approxi-mately seven months.[19] Anecdotal reports ofdeath in a Hyacinth Macaw and Red-belliedMacaw suggest caution in these species.[20]
OTHER AGENTS
Antihistamines
Antihistamines block the physiologic effects ofhistamine. H1 receptors are responsible for pruri-tus, increased vascular permeability, release ofhistamine mediators, and recruitment of inflam-matory cells.[22] Antihistamines are beneficial inthe treatment of pruritus, self-trauma, and anxi-ety. H1 receptor antagonists have sedative, anti-nausea, anticholinergic, antiserotinergic, and localanesthetic effects.[22] Caution with concurrentanticholinergic agents, CNS depressants, and pa-tients with hepatic disease is indicated. Paradoxi-cal excitation and anxiety are rare side effects.
Diphenhydramine (Benadryl®)
Diphenhydramine is available in 25 mg and 50 mgcapsules for oral administration as well as a 10 or50 mg/ml injectable for IM or IV administration.One gram is soluble in 1 ml of water or 2 ml of al-cohol. Diphenhydramine has antihistaminic, seda-tive, and antidepressant activity. Atropine-like an-ticholinergic side effects may occur. In humans ithas been used to treat neuroleptic-induced parkin-sonism.[17] Diphenhydramine may be beneficialin pruritic feather-picking birds. Suggested dosageis 2–4 mg/kg PO bid, or 0.5 mg/240 ml drinkingwater.[14]
Hydroxyzine (Atarax®)
Hydroxyzine is available in a 10 mg, 25 mg, 50mg, and 100 mg tablet, as well as a 2 mg/ml solu-tion for oral administration. It is available as aninjection (25 mg/ml) for IM administration. It isvery soluble in water and freely soluble in alco-hol. Hydroxyzine may also inhibit mast cell de-granulation. Hydroxyzine has been used as ananxiolytic agent and may be beneficial in pruriticfeather-picking birds. One case report suggestedhydroxyzine combined with eicosapentaenoicacid (DermCaps) was benefical in treating afeather-picking Red-lored Amazon Parrot.[23]
Suggested dosage is 2.2 mg/kg PO tid, or 4mg/100–120 ml drinking water.[14]
Anticonvulsants
Anticonvulsants have few behavioral applicationsunless there is an epileptic component. Occa-sionally, it is difficult to differentiate compulsivedisorders from focal seizures. Response to ther-apy may be diagnostic. Benzodiazepines havealso been used as anticonvulsants for status epi-lepticus. They are not preferred for long-termmanagement of seizure disorders. Side effects ofbarbiturates are similar to those of benzodiaze-pines but with a lower therapeutic index. Cautionis indicated with concurrent CNS drugs (antipsy-chotic and antidepressant) because of increasedCNS and respiratory depression, and in patientswith hepatic disease.
Phenobarbital (Solfoton®)
Phenobarbital is available as a 15 mg, 16 mg, 30mg, 60 mg, and 100 mg tablet, a 3 mg/ml elixir,and 4 mg/ml solution for oral administration. Onegram is soluble in approximately 1,000 ml ofwater and 10 ml of alcohol. It is beneficial intreating seizure disorders. It has sedative, anti-spasmodic, and anticholinergic effects. Side ef-fects may include depression, vomiting, andataxia. Phenobarbital is a controlled substancewith human abuse potential. Suggested dosagesvary from 1–7 mg/kg PO bid to tid.[24, 25] Oneshould use the lowest dosage that controls the dis-order. Alternately, 6–10 mg/120 ml drinkingwater or 2–3.2 mg/kg PO bid has been suggestedfor amazon parrots and idiopathic epilepsy.[14]
Carbamazepine (Tegretol®)
Carbamazepine is available in a 100 mg and 200mg tablet or as a 20 mg/ml suspension for oral ad-ministration. Structure is similar to the tricyclicantidepressant imipramine. Carbamazepine hasbeen used to treat seizures, depression, mania,and explosive aggressive states in people.[17] Inbirds it may be useful in the treatment of compul-sive disorders and aggression due to anxiety orfrustration. Carbamazepine is slightly sedating,mildly anticholinergic, and does not cause musclerelaxation. Contraindications include renal, he-patic, cardiovascular, or hematological disorders.Carbamazepine has been suggested by some to bethe preferred drug for psittacine feather pick-
24 / Psittacine Behavioral Pharmacotherapy 275

ing.[14] Usually, the drug is combined with chlor-promazine or haloperidol for the initial two weeksof treatment. Suggested dosage is 3–10 mg/kg POsid or 20 mg/120 ml drinking water.[14]
NARCOTIC ANTAGONISTS
Narcotic antagonists have been used to treat ste-reotypies in zoo and companion animals.[26–29]In humans, it has been used to treat self-injuriousbehavior and addictions. Opiate peptides are re-leased during stress and activate the dopaminesystem, which may lead to compulsive andstereotypic behaviors.[31] Endogenous opioidsmay induce analgesia and block pain, allowingself-mutilation to occur. Therefore, narcotic an-tagonists may be effective in reducing compulsiveand stereotypic behaviors of recent origin. Clin-ical suppression of compulsive disorder may beshort-lasting and only beneficial in acute presen-tations.[27–30]
Naltrexone (Revia®)
Naltrexone is a synthetic opiate antagonist that isavailable in 50 mg oral tablets. 100 mg of naltrex-one is freely soluble in 1 ml of water. Caution isindicated in patients with hepatic disease. In onestudy, naltrexone was effective in reducingfeather picking in 26 of 42 cases. The use of re-straint collars in the study makes it difficult tocritically assess the effectiveness of the drug.[21]A 50 mg tablet can be mixed with 10 cc sterilewater and is apparently stable if refrigerated forup to three months. Suggested dosage is 1.5mg/kg or three to four drops oral bid for a Sulfuror Umbrella Cockatoo-sized bird.[32]
NARCOTIC AGONISTS
Narcotic agonists, or opiates, are useful in treat-ing moderate to severe pain in birds. Birds mayrespond to pain by trying to escape, becomingrestless and anxious, vocalizing and struggling, orbecoming aggressive. Acute pain may also mani-fest as ruffled feathers and immobility. Beha-vioral signs of chronic pain may include inappe-tence, weight loss, lack of grooming orovergrooming a painful site, or feather pickingover a specific body area or region.[33] In casesof self-injurious behavior and mutilation, a trialwith an effective pain reliever may be warranted.Recent studies suggest that kappa opioids, such asbutorphanol, may be more effective analgesics in
birds than mu agonists, such as buprenor-phine.[33] Respiratory, cardiac, and CNS depres-sion may be increased with concurrent use ofother CNS depressants. Opiates are controlledsubstances with the potential for human abuse.
Butorphanol (Torbugesic®)
Butorphanol, a kappa opioid, is available as a 1mg/ml, 2 mg/ml, and 10 mg/ml injectable for IMadministration. Butorphanol (1–3 mg/kg) hasbeen found to be an effective analgesic in AfricanGrey Parrots, cockatoos, and Hispaniolan Par-rots.[33] One study found the drug an ineffec-tive analgesic at 1 mg/kg for amazon parrots.Dosages of 6 mg/kg may have hyperalgesic ef-fects in some birds. Suggested dose is 2–4 mg/kgIV, IM, or PO tid.
Buprenorphine (Buprenex®)
Buprenorphine, a mu agonist, is available as a 0.3mg/ml injection. One study in African GreyParrots found no significant analgesic effect withlarge doses. Suggested dosage is 0.1 mg/kg IV orIM bid.[34]
Nonsteroidal Anti-inflammatory Drugs
Nonsteroidal anti-inflammatory drugs (NSAIDs)are analgesic, anti-inflammatory, and antipyretic.Carprofen and ketoprofen are the most commonlyused NSAIDs in avian medicine for mild to mod-erate pain.[32] NSAIDs are synergistic and moreeffective when combined with other analgesicagents. In mammals, gastrointestinal ulcerationand bleeding may result from drug-induced inhi-bition of prostaglandin synthesis. Use caution indehydrated patients because of increased renalcomplications. Information on dosages in birdshas been established empirically.
Carprofen (Rimadyl®)
Carprofen is available as a 50 mg/ml injectablefor IM, IV, or SC administration, or as a 25 mg,50 mg, 75 mg, and 100 mg tablet for oral admin-istration. Carprofen is insoluble in water andfreely soluble in ethanol. Carprofen at 1 mg/kgSQ has been shown an effective analgesic inbroiler chickens, reaching peak plasma levels inone to two hours and raising pain thresholds for atleast 90 minutes.[33] Carprofen is a specificCOX-2 inhibitor, making its primary effect anti-inflammatory, and thus sparing prostaglandins.
276 Manual of Parrot Behavior

Suggested dosage is 2–10 mg/kg IV, IM, or SQsid.[34]
Ketoprofen (Orudis® or Ketofen®)
Ketoprofen is available for IM, IV, or SC injection(100 mg/ml), as well as a 12.5 mg, 25 mg, 50 mg,and 75 mg capsule for oral administration. Keto-profen is practically insoluble in water and freelysoluble in alcohol. Suggested dosage is 2 mg/kgIM sid.[34]
Hormones
Hormones have historically been used to treatsome forms of anxiety and aggression in animals.In birds, they have a non-specific calming effectand have been advocated for treating dominanceand/or sexual aggression among birds and pluck-ing.[35] Progestins are antiandrogenic, causeCNS depression, and increase appetite. Severeside effects include diabetes mellitus, bone mar-row suppression, adrenocortical suppression, andcarcinomas.
Chorionic Gonadotropin (APL®)
Human chorionic gonadotropin (hCG) is avail-able for IM injection (500 units/ml, 1,000units/ml, and 2,000 units/ml). The drug has beenused to inhibit egg laying. It has been reportedmoderately successful for aggressive and feather-plucking female birds. It has been reported lesssuccessful in male birds with the exception ofEclectus males.[35] Dosage protocols suggest500–1,000 units/kg IM.[35] If no response is seenwithin three days, the dosage may be repeated. Ifno response after a second injection, the drug isunlikely to be effective. Injections may be re-peated every four to six weeks. Often the drug isless effective with time, requiring a shorter dosingfrequency or making usage impractical.
Leuprolide Acetate (Lupron®)
Leuprolide is available as an injection (5 mg/ml).It is a luteinizing hormone (LH)-releasing hor-mone that has an effect on lowering follicle-stimulating hormone (FSH), LH, testosterone,and estrogen through negative feedback. The drughas been used to treat chronic egg laying, cysticovarian and oviduct disease, egg yolk peritonitis,granulomas of the ovary and oviduct, cloacalprolapse, continued ovulation after salpingohys-terectomy, feather picking, aggression, and per-
sistent sexually induced regurgitation. Anecdotalreports suggest 73% overall improvement, withresolution in 89% of chronic egg-laying psit-tacids.[36] Dosage recommendations vary from100–1,000 g/kg IM every two weeks for threetreatments.[36]
Medroxyprogesterone Acetate (Depo-Provera®)
Medroxyprogesterone is available as a 100 mg/mland 400 mg/ml injectable. It has been used for ex-cessive egg production in Cockatiels and to detersexual behavior, including feather plucking. Sideeffects, such as lethargy, inappetence, weightgain, polyuria and polydipsia, hepatopathy, anddeath, preclude routine usage.[35] Suggesteddosage is 5–50 mg/kg IM or SQ every four to sixweeks—150g bird—0.05mg/g, 300–700g bird—0.03mg/g, and >700g bird—0.025mg/g.[34]
COMBINATION THERAPY
Behavioral drugs are occasionally used in combi-nation to enhance their effectiveness. The mostsensible choice is the combination of benzodi-azepines with antidepressants. Benzodiazepinesmay be added to antidepressants when the effec-tiveness of the antidepressant is waning. In addi-tion, benzodiazepines are useful because of theirrapid onset of action when waiting for the de-layed effect of antidepressants. Caution shouldbe taken because concurrent administration maylead to increased CNS depression. The dosage ofthe benzodiazepine should be lowered to avoidthis complication. In humans, one study foundthat combined administration of fluoxetine andalprazolam, a benzodiazepine, resulted in a 30%increase in the plasma benzodiazepine concen-trations.[37]
Fluoxetine and amitriptyline have been usedconcurrently in human and canine patients.Fluoxetine potentiates the effects of amitriptylineand the intermediate metabolite nortriptyline.[38]Caution should be taken with concurrent usage oftricyclic antidepressants and specific serotoninreuptake inhibitors (SSRIs and TCAs) because ofthe risk of serotonin syndrome. Serotonin syn-drome is a serious and potentially fatal condition.In order of appearance as the condition worsens,signs include diarrhea; restlessness; extreme agi-tation, hyperreflexia, and autonomic instabilitywith possible rapid fluctuations in vital signs;myoclonus, seizures, hyperthermia, uncontrol-
24 / Psittacine Behavioral Pharmacotherapy 277

lable shivering, and rigidity; and delirium, coma,status epilepticus, cardiovascular collapse, anddeath.[17] Treatment is often supportive.
Phenothiazines may be combined with benzo-diazepines or SSRIs with additive sedative ef-fects. Phenothiazines and TCAs should not beused in conjunction because both have sedativeand anticholinergic effects. The anticonvulsantcarbamazepine has been used in combinationwith antipsychotics (chlorpromazine and halo-peridol) during the initial treatment (first twoweeks) of compulsive feather-picking disordersand self-mutilation.[14] Combined admini-stration of paroxetine and phenobarbital mayresult in a decreased plasma concentration ofparoxetine.[39]
CONCLUSIONS
Behavioral pharmacotherapy is a beneficial ad-junct to behavioral and environmental modifica-tion in many situations. Most avian patients willbenefit from timely and appropriate use of behav-ioral drugs. Behavioral results are often expe-dited, and outcomes are often improved with con-comitant use of drugs. Unfortunately, scientificliterature for use in treating behavioral problemsof birds is limited at this time and consists prima-rily of anecdotal reports and uncontrolled studies.At present, studies are often of small sample sizeand varied species, making it difficult to drawstatically significant conclusions. Further clinicaltrials are necessary.
REFERENCES
1. Johnson-Delaney, C. 1992. Feather picking: Diag-nosis and treatment. Journal of the Association ofAvian Veterinarians 6 (2):82.
2. Guyton, A.C. 1991. Basic neuroscience: Anatomy& physiology, 2nd ed. Philadelphia: W.B. Saun-ders, p. 277.
3. Center, S.A., T.H. Elston, P.H. Rowland, D. Rosen,B.L. Reitz, I.E. Brunt, I. Rodan, J. House, S.Banks, L. Lynch, L. Dring, J. Levy. 1996. Ful-minant hepatic failure associated with oral admin-istration of diazepam in 11 cats. Journal of Veteri-nary Emergency and Critical Care 6:618–625.
4. Quesenberry, K. 1994. “Avian neurological disor-ders.” In Saunders manual of small animal prac-tice, ed. S.J. Birchard and R.G. Sherding, pp.1312–1316. Philadelphia: W.B. Saunders.
5. Ritchie, B.W., and G.J. Harrison. 1994. “Formu-lary.” In Avian medicine: Principles and applica-
tion, B.W. Ritchie, G.J. Harrison, and L.R. Harri-son, pp. 457–478. Lake Worth, FL: WingersPublishing, Lake Worth, Florida.
6. Hewson, C.J., J.M. Parent, P.D. Conlon, A.U. Lue-scher, R.O. Ball. 1998. Efficacy of Clomipraminein the treatment of canine compulsive disorder.Journal of the American Veterinary MedicalAssociation 213:1760–1766.
7. Seibert, L.M. 2003. “Psittacine feather picking.”Proceedings, Western Veterinary Conference.
8. Johnson, C.A. 1987. “Chronic feather picking: Adifferent approach to treatment.” Proceedings,First International Conference on Zoological andAvian Medicine, pp. 125–142.
9. Luescher, A.U. 2002. “Compulsive behaviour.” InBSAVA manual of canine and feline behaviouralmedicine, ed. D. Horwitz, D. Mills, and S. Heath,pp. 229–236. Quedgeley, Gloucester, England:BSAVA.
10. Luescher, A.U. 1998. “Compulsive behavior: Rec-ognition and treatment.” Proceedings, Am AssocZoo Vet/Am Assoc Wildlife Vet, pp. 398–402.
11. Ramsey, E.D., and H. Grindlinger. 1994. Use ofclomipramine in the treatment of obsessive behav-ior in psittacine birds. J Assoc Avian Vet 8:9–15.
12. Seibert, L.M., S.L. Crowell-Davis, G.H. WilsonGH, and B.W. Ritchie. 2004. Placebo-controlledclomipramine trial for the treatment of featherpicking disorder in cockatoos. Journal of the Ame-rican Animal Hospital Association 40:261–269.
13. McDonald, S.E. 1989. Summary of medicationsfor use in psittacine birds. Journal of the Associ-ation of Avian Veterinarians 3 (3):120–127.
14. Carpenter, J.W., T.Y. Mashima, and D.J. Rupiper.2001. Exotic animal formulary, 2nd ed. Phila-delphia: W.B. Saunders, pp. 171–174.
15. Mertens, P.A. 1997. “Pharmacological treatmentof feather picking in pet birds.” Proceedings, FirstInternational Conference on Veterinary BehavioralMedicine, Birmingham, England, pp. 209–211.
16. Martin, K.M. 2004. “Behavioral approach to psit-tacine feather picking.” Proceedings, Associationof Avian Veterinarians.
17. Sadock, B.J., and V.A. Sadock. 2001. Kaplan &Sadock’s pocket handbook of psychiatric drugtreatment, 3rd ed. Philadelphia: LippincottWilliams & Wilkins, p. 198.
18. Lennox, A.M., and N. VanDerHeyden. 1999.“Long-term use of haloperidol in two parrots.”Proceedings, Association of Avian Veterinarians,pp. 133–137.
19. Iglauer, F., and R. Rasim. 1993. Treatment of psy-chogenic feather picking in psittacine birds with adopamine antagonist. Journal of Small AnimalPractice 34:564–566.
278 Manual of Parrot Behavior

20. Lennox, A.M., and N. VanDerHeyden. 1993.“Haloperidol for use in treatment of psittacine self-mutilation and feather plucking.” Proceedings,Association of Avian Veterinarians, pp. 119–120.
21. Welle, K.R. 1998. “A review of psychotropic drugtherapy.” Proceedings, Annual Conference of theAssociation of Avian Veterinarians, pp. 121–123.
22. Scott, D.W., W.H. Miller, and C.E. Griffin. 1995.“Dermatologic therapy.” In Muller & Kirk’s smallanimal dermatology, 5th ed., pp. 211–218.Philadelphia: W.B. Saunders.
23. Krinsley, M. 1993. Use of Dermcaps liquid andhydroxyzine hcl for the treatment of feather pick-ing. Journal of the Association of Avian Veteri-narians 7 (4):221.
24. Gould, W.J. 1995. Caring for pet birds’ skin andfeathers. Veterinary Medicine 90 (1):53–63.
25. Bennett, R.A. 1996. “Common avian emergen-cies.” Proceedings, Fifth International VeterinaryEmergency and Critical Care Symposium, pp.698–703.
26. Kenny, D.E. 1994. Use of naltrexone for the treat-ment of psychogenically induced dermatoses infive zoo animals. Journal of the AmericanVeterinary Medical Association 205:1021–1023.
27. Dodman, N.H., and L. Shuster. 1998. Psychophar-macology of animal behavior disorders. Oxford,England: Blackwell Science, pp. 209–211.
28. Brown, S.A., S. Crowell-Davis, T. Malcom, P.Edwards. Naloxone-responsive tail-chasing in adog. Journal of the American Veterinary MedicalAssociation 1987:190;884–886.
29. Dodman, N.H., L. Shuster, S.D. White, M.H.Court, D. Parker, and R. Dixon. 1988. Use ofnarcotic antagonists to modify stereotypic self-licking, self-chewing, and scratching behavior indogs. Journal of the American Veterinary MedicalAssociation 193:815–819.
30. White, S.D. 1990. Naltrexone for the treatment ofacral lick dermatitis in dogs. Journal of the Ameri-can Veterinary Medical Association 196:1073–1076.
31. Landsberg, G., W. Hunthausen, and L. Ackerman.Handbook of behavior problems of the dog andcat, 2nd ed. Philadelphia: W.B. Saunders, p. 138.
32. Turner, R. 1993. “Trexan (naltrexone hydrochlo-ride) use in feather picking avian species.”Proceedings, Association of Avian Veterinarians,pp. 116–118.
33. Paul-Murphy, J. 2003. “Managing pain in birds.”Proceedings, Managing Pain Symposium.
34. Doolen, M. 1996. “Appendix—Formulary.” InBSAVA manual of psittacine birds, ed. P.H.Beynon, N.A. Forbes, and M.P.C. Lawton, pp.228–234. Ames: Iowa State University Press.
35. Lightfoot, T.L. 2001. “Feather ‘Plucking.’” Pro-ceedings, Atlantic Coast Veterinary Conference.
36. Zantop, D.W. 2000. “Using leuprolide acetate tomanage common avian reproductive problems.”Proceedings, International Conference on Exotics(2,3), p. 70.
37. Lasher, T.A., J.C. Fleishaker, R.C. Steenwyk, andE.J. Antal. 1991. Pharmacokinetic pharmacody-namic evaluation of the combined administrationof alprazolam and fluoxetine. Psychopharma-cology 104:323–327.
38. Mills, D.S., and B.S. Simpson BS. 2002. “Psycho-tropic agents.” In BSAVA manual of canine and fe-line behavioural medicine, ed. D. Horwitz, D.Mills, and S. Heath, pp. 237–244. Quedgeley,Gloucester, England: BSAVA.
39. Aranow, R.B., J.I. Hudson, H.G. Pope, T.A. Grady,T.A. Laage, I.R. Bell, and J.O. Cole. 1989.Elevated anti-depressant plasma levels after addi-tion of fluoxetine. Am J Psychiatry 146:922–913.
40. Jenkins, J.R. 2001. “Feather picking and self-mutilation in psittacine birds.” In Veterinary clinicsof North America: Exotic animal practice, ed. T.L.Lightfoot, pp. 651–667. Philadelphia: W.B.Saunders.
41. Tully, T.N. “Formulary.” In Avian medicine andsurgery, ed. R.B. Altman, S.L. Clubb, G.M. Dorre-stein, and K. Quesenberry, pp. 671–673. Philadel-phia: W.B. Saunders.Drug solubility was obtained from Plumb, D.C.1999. Veterinary drug handbook, 3rd ed. Ames:Iowa State University Press.
24 / Psittacine Behavioral Pharmacotherapy 279

INTRODUCTION
Psittacine birds have been kept in captivity sinceat least the fifth century B.C. In the 1800s, Britishrecords showed that several different species werebred successfully in captivity. However, it wasn’tuntil modern methods of travel, such as the com-mercial airplane, arose that larger psittacine birdsbegan to be propagated in captivity in earnest. Inthe 1970s, importation of psittacine birds for thepurpose of captive breeding was a lucrative indus-try. These birds were usually wild-caught animalsthat were arbitrarily thrown together in pairs or incolonies and expected to reproduce. When theydid not, it was generally assumed that somethingwas medically wrong or that the birds were pastfertile age, and since replacement breeder importswere readily available, little attention was given tosolving the mysteries of poor production. In theearly 1980s, the Association of Avian Veterina-rians was established, providing a forum for ideason the improvement of avian health and avicul-ture. In the early 1990s, with the passage of theWild Bird Conservation Act, the flow of importedparrots slowed to a trickle. This necessitated moreadvanced avicultural practices to increase domes-tic production and greater attention was given tothe husbandry and behavioral issues behind infer-tility. The realization dawned among aviculturistsand veterinarians alike that birds must not only behealthy but also “happy” in order to reproduceconsistently. Unfortunately, there have been rela-tively few studies of free-ranging or captivepsittacine birds that might better define their nor-mal behavior and promote reproductive effi-ciency (Millam 1994; Styles 2001). Most of whatis thought to be known today is anecdotal infor-
mation and observations made by aviculturists,avian veterinarians, and a few behaviorists.
SOCIALIZATION
It has long been discussed among aviculturiststhat for many psittacine species, wild-caughtpairs make better breeding stock than domesti-cally bred birds. Although there have been nostudies to confirm this, the observation may havesome validity. Proper socialization is critical tobreeding success among birds. Birds learn vari-able socialization skills at different stages of theirlives. These stages are known as sensitive or crit-ical periods (Smith 1999). Obviously, a wild-caught adult parrot will have experienced themost normal social development through thesestages. However, it is possible to reproduceproper socialization forums for young parrots incaptivity and these birds may be more adapted tothe stress of living in close proximity to humansand appear to reach puberty earlier, making thempotentially better breeding stock. Indeed, recentanecdotal reports from aviculturists with F2, 3,and 4 generation birds support this premise.Ideally, young birds that are intended for breedingshould be parent-raised and then placed in flightswith other juveniles of the same species so thatflock socialization may occur. If hatchlings andfledglings are not properly socialized with theirown species, then poor mating success or otherproblems may result later in life (Meehan et al.2003; Immelmann 1972; Rajecki et al. 1977).Critical periods vary based on species and thetype of learning that occurs. For example, filialimprinting and sexual imprinting occur duringdifferent critical periods (Smith 1999; Vidal
281
25Behavior of Captive Psittacids
in the Breeding Aviary
G. Heather Wilson

1980). Young cockatoos may not recognize par-ental vocalization until approximately five weeksafter hatching (Rowley 1980). There are somespecies that may breed successfully after beinghand-raised and kept as a companion animalwithout proper flock socialization, such as ama-zons and many of the smaller species. However,there are others that will very rarely breed underthese conditions, such as African Greys.
Most psittacine birds are social species andspend the non-breeding season in flocks, althoughsome birds, like the Kakapo, may be solitary.During the breeding season, most free-rangingpsittacine birds break off from the flock in pairs,although some nest in colonies, such as Pata-gonian Conures and Quaker Parakeets. Althoughmost larger parrots do form long-term pair bondsand are monogamous, there are some exceptions,such as the Kea, which is polygamous. Free-ranging male amazon parrots have been docu-mented to break up a nesting pair and then matewith the hen (Wiley 1980). In Asiatic Parakeets,such as the Indian Ringneck, and Eclectus Parrots,the pair bonds only last for one reproductive sea-son. There have been two reports of breeding andnesting behavior in captive Cockatiel flocks thatdemonstrate pairs, triads, and extra-pair mating dooccur (Seibert & Crowell-Davis 2001; Harrison1995). This same behavior has been recorded infree-ranging Echo Parakeets (Thorsen et al. 1998).
COURTSHIP BEHAVIOR
Courtship is an important prelude to breeding inall avian species. Allopreening and allofeedingmay be seen in bonded pairs throughout the yearbut often escalate prior to breeding (Figures 25.1aand b). Previous observations suggest that inamazon parrots, lovebirds, and the genus Melo-psittacus, the preening is confined to the head andneck region, while in Aratinga, Brotogeris, Ara,and Cacatua the area preened includes the head,wings, and tail (Harrison 1995). A receptive henwill signal her readiness to mate by leaning for-ward and fanning her tail. Amazons and someother species mount from the side, placing onefoot on the hen’s back, but in Cockatiels, love-birds, lories, and many other species the malemounts by placing both feet on the hen’s back.
If a new pair is to be introduced, it is generallyadvisable to put both birds simultaneously in anew cage. If this is not possible, altercations may
be decreased by timing the introduction to coin-cide with the non-breeding season, allowing thehen to occupy the breeding cage and the cock asmaller cage hung low on the outside of the largercage, and performing wing, beak, and nail trimson the male prior to introduction into the hen’scage.
Psittacine birds have complex methods of com-munication, including color, posturing, and vo-calization. Vocalization may play an integral partin the courtship of some species. It has been the-orized that contact call imitation in adult Bud-gerigars contributes to pair bond formation and
282 Manual of Parrot Behavior
Figure 25.1a and 25.1b. A bonded pair ofMilitary Macaws engaged in allopreening andallofeeding. See also color section.

maintenance (Hile et al. 2000). Vocalizationswere found to be distinct among pairs of PuertoRican Amazon Parrots in the wild (Wiley 1980).
In most species, the cock is generally dominantto the hen. In one study of the agonistic and affil-iative behaviors of a flock of 12 captive Cocka-tiels that were colony housed, males were shownto rank significantly higher than females in thesocial hierarchy based on dyadic agonistic inter-actions (Seibert & Crowell-Davis 2001). How-ever, there are exceptions to this scenario, as withpsittaculid parakeets and Eclectus Parrots. Inthese species the hen is thought to be dominantand it is preferable to keep an older cock with ayounger hen for optimal breeding success and toprevent damage to the male. In all psittacine birdscommonly kept in captivity, it is the hen thatchooses a male deemed suitable for mating. Whenbirds are forced paired, aggression or lack ofbreeding success may occur. Recent observationssuggest improved breeding success when mem-bers of a species or genera are flocked together inthe non-breeding season and allowed to choose
their own mates and are then paired off for thebreeding season into separate cages (Figure 25.2).
CAGING AND NEST SELECTION
When considering cage design and nest box con-struction, the natural behavior as well as the healthof the species should be considered. For example,is the species predominately arboreal, such asmost amazons, or does it spend a significant por-tion of its time in terrestrial activity, like many ofthe Australian cockatoos? This may provide guid-ance in selecting either a suspended welded-wireenclosure or one that allows access to the ground,despite increased health risks associated with thistype of caging (Figure 25.3). Additionally, mostparrots have evolved as a prey species, thereforethe ability to “escape” at least a short distance iscritical for the psychological comfort of most ofthese birds. Cages that provide perches that arehigh above human heads, and that allow for someflight distance, will result in birds that are lessstressed and more likely to perceive the enclosureas “safe” nesting territory (Figure 25.4).
25 / Behavior of Captive Psittacids in the Breeding Aviary 283
Figure 25.2. Flocking birds of the same species or genera in the non-breeding season may increaseproduction by allowing voluntary mate selection. See also color section.

284
Figure 25.3. An example of a captive environment that allows some natural behaviors, such asground foraging, which can improve breeding success in some species such as this Major MitchellCockatoo.

Appropriate perching is also vital to reproduc-tive success. Secure wooden branches that allowthe bird to wrap its foot approximately halfwayaround the diameter are usually ideal. Woodenperches naturally provide good footing, are easilyreplaceable and provide good nesting materialand safe chewing substrate (Figure 25.5).
It has been proposed that nest box presentationis the most effective stimulus for eliciting egg lay-ing in psittacine birds (Millam 1994). A wide vari-ety of nest boxes have been promoted in aviculture.Sometimes providing a choice of a couple of dif-ferent styles to birds may help increase nest satis-faction and thus production. Most parrots are notnest builders (although there are some exceptions,such as lovebirds and Hanging Parrots) but rathernest in cavities, usually in trees, in the wild.Therefore, wooden boxes with dark inner cavitiesare usually preferred. Many species, such as somemacaws, prefer either broad perches or horizontalboxes that allow them to mate in the nest. Others,
such as African Greys or the Poicephalus species,may prefer the “boot” type box that allows de-creased light penetration into the egg-laying area.Still others, like many cockatoo species, may pre-fer the “grandfather clock” or vertical type box thatwould more closely approximate the length of atree trunk (Figure 25.6). Many species excavatedown a tree trunk nest several feet before feelingsafe enough to lay and captive nests must approxi-
25 / Behavior of Captive Psittacids in the Breeding Aviary 285
Figure 25.4. An enclosure that is suspendedhigh above the ground, like this nesting Blue-fronted Amazon pair’s cage, can decrease stressand increase chick production.
Figure 25.5. Inappropriate perching, asdemonstrated by the slick, overly wide branchthis Red-lored Amazon is trying to balance on,can interfere with breeding success.
Figure 25.6. A choice of nest box styles andmaterials may help encourage a pair to go tonest. This pair of Hyacinth Macaws routinely“work” the grandfather-style box early in theseason and then lay eggs in the horizontal boxlater in the season.

mate this (Figure 25.7). Additionally, some avicul-turists prefer a “T” type box, especially with ag-gressive cockatoos, with two holes that allow thehen to escape aggressive advances from the cock.Nest boxes should be placed on the cage in such asway as to minimize disruptions (i.e., away fromfood bowls and general foot traffic) but allow easyinspection from outside the cage (Figure 25.8).
Nest substrate may also play a vital role in in-creased production. While some species may becontent to nest on recycled paper products, othersmay need more natural bark chips or wood thatcan be chewed down to the proper consistency.There have been many observations that suggestnot only may this be stimulatory for the pair butthat it may also help to decrease mate aggressionin some species, as the cock is initially busypreparing the nesting cavity and therefore givesthe hen more time to reach breeding condition.Having a narrow entrance to the box that must bechewed open to allow the pair access may alsoserve this same purpose.
The location of the cage in relation to othercages has also been shown to affect breeding be-havior. From a health aspect, it is important not tohouse species from different continents in thesame air space. This premise may also have someimportant psychological impact. For example,African Greys are a relatively quiet species andbreeding success has been shown to increase whenthese birds are housed away from noisier birds,
such as amazons or cockatoos. There may even beproblems when housing birds within the samegenus together. For example, most cockatoos,which are a highly territorial species, need largerbreeding space. For these birds, as well as otherspecies like African Greys, visual barriers betweencages kept in close proximity may increase repro-ductive success in captivity (Figure 25.9).Alternatively, some conures (Aratinga or Pyrrhua)or Budgerigars are communal and don’t appear tobe disrupted by visual contact with neighboringpairs. In fact, having other mating pairs of thesame species in the vicinity is thought to stimulatebreeding for many species. These birds may evenbe propagated in colony breeding situations, al-though it is advisable to have an equal number ofmales and females (Figure 25.10).
Ideally, cages should be located outdoors toallow access to a natural photoperiod and full-spectrum light. Circadian rhythms affect bodytemperature, hormone output, metabolism, andreproductivity (Ryan 1999). In birds, there are atleast three distinct input pathways for light to acton circadian rhythms (Campbell & Murphy1998). Although some species may reproduce de-spite lack of natural photoperiod, it probably doesplay a supportive role in the breeding success ofmost pairs (Millam 1994). It has been shown thatspermatogenesis in the Budgerigar can be ar-rested by reducing the birds’ photoperiod to eighthours of daylight or less (Humphreys 1975). Inmost cases, even breeding pairs that are imported
286 Manual of Parrot Behavior
Figure 25.7. Deep, dark natural nesting cavi-ties, such as this hollowed out palm log, areoften preferred nesting sites, as demonstratedby this Alexandrine Parakeet hen on five fertileeggs. See also color section.
Figure 25.8. Keeping the nesting box awayfrom high traffic areas, such as the path traveledwhen doing daily feeding, may increase breedingsuccess. See also color section.

into the United States and housed in outdooraviaries will adjust their breeding season to thespring and summer months of the NorthernHemisphere. There are some exceptions to this,such as Eclectus Parrots, which may breed year-round, and Alexandrine Parakeets and some Poi-cephalus parrots, which may breed in the winter(Figure 25.11). Interestingly, it has been notedthat in the wild, Eclectus Parrots do not exhibitbreeding seasonality but rather breed whenever anesting site becomes available (Sparks & Soper1990). Stimulation of egg laying and increase inplasma luteinizing hormone (LH) secretion wasfound to be highest in Cockatiel hens provided
with full mate contact, nest box access, and ap-propriate photostimulation (Shields et al. 1989).It has also been shown that the plumage of mostparrots reflects near ultraviolet (UVA) wave-lengths (300–400 nm) and exhibits UVA-inducedfluorescence. This appears to play a role in mateselection, possibly making access to full-spectrum light important in facilitation of pairbonding (Pearn et al. 2001).
Environmental stimuli that may serve as cuesto the onset of the avian breeding season are notlimited to light alone. Circannual rhythms, whichcontrol when certain species of birds know to mi-grate or mate, are probably controlled by otherfactors, such as temperature, humidity, nutrition,and social interaction. All these variations aremore difficult to control in captive indoor settingsand can affect breeding success (Millam 1994).
PARENTING
In most species, it is the hen that is responsible forincubation of the eggs. There are some exceptionsto this, such as Cockatiels, macaws, and conures,in which the cock incubates during the day and thehen at night. In all psittacine species studied thusfar the cock assists in the rearing of the young(Figure 25.12). Some species or individuals ap-pear to have better parenting skills than others andcross-fostering is commonly employed in avicul-ture. It is advisable to remain within the species ifthis is a common practice, both from a disease per-spective and behavioral development. This is justone reason why aviaries that concentrate on onegenus or species are often more successful thanthose that collect multiple species and attempt topropagate all of them. A flock average of fivefledged chicks/pair/year is considered an optimumproduction rate that neither “wears out” breedingpairs nor underutilizes them.
BEHAVIORAL ABNORMALITIES
Most psittacine birds, whether imported or do-mestically bred, kept as companion animals orused as breeding stock, are not considered domes-ticated but rather captive wild animals. Addi-tionally, unlike some other companion species,such as dogs or cats, psittacine birds are a preyspecies. The forced proximity of captive parrotsto humans and the effectively curtailed ability inmost cases of these animals to flee when threat-ened have produced a wide variety of behavioralproblems. Most of these will be addressed else-
25 / Behavior of Captive Psittacids in the Breeding Aviary 287
Figure 25.9. Complete visual barriers betweenthe cages of territorial species, such as theselovebirds, can improve production.
Figure 25.10. Conure species may only requirevisual barriers around the next box itself.

where in this text. However, there are some be-havioral abnormalities that impair effective repro-duction that must be mentioned here. These in-clude destruction of eggs, nest abandonment,excessive aggression toward mates or chicks, andstereotypical behaviors that consume the individ-ual to the exclusion of breeding.
Enclosure size, placement, and height as men-tioned previously may all play a role in preventingor alleviating these deviant behaviors. Addition-ally, proper nutrition (also described in anotherchapter) plays a critical role and should be tailoredto each species or individual. Environmental en-richment is also vital and should be given the sameattention in either breeding or companion birds.Disruptions by human or other predators may pre-cipitate these maladaptive behaviors and should bekept to a minimum (Figure 25.13).
288 Manual of Parrot Behavior
Figure 25.11. Large flights, like this Alexandrine Parakeet enclosure, with multiple nest boxes mayimprove fertility in colonial species.
Figure 25.12. A Double Yellow-headedAmazon Parrot displaying normal territorial ag-gression in defense of the nest box where thehen is on eggs. See also color section.

289
Figure 25.13. Environmental enrichment is important for all captive psittacine birds, even those inbreeding aviaries.

CONCLUSION
As the reproductive lifespan of birds imported inthe 1970s and 1980s draws to a close, the success-ful reproduction of their progeny becomes morecritical to the perpetuation of aviculture. Giventhe importance of normal reproductive behaviorin facilitating fertility and hatchability inpsittacine birds, it is hoped that aviculturists andavian veterinarians will continue to demonstratesupport in the study of these fascinating and com-plex creatures.
REFERENCES
Campbell, S.S., and P.J. Murphy. 1998. Extraocular cir-cadian phototransduction in humans. Science279:396–398.
Harrison, G.J. 1995. “Perspective on parrot behavior.”In Avian medicine: Principles and application, ed.B.W. Ritchie, G.J. Harrison, and L. Harrison, pp.96–105. Lake Worth, FL: Wingers Publishing.
Hile, A.G., T.K. Plummer, and G.F. Striedter. 2000.Male vocal imitation produces call convergence dur-ing pair bonding in budgerigars, Melopsittacus un-dulatus. Anim Behav 59 (6):1209–1218.
Humphreys, P.N. 1975. Ultrastructure of the budgerigartestis during a photoperiodically induced cycle. CellTissue Research 159 (4):541–550.
Immelmann, K. 1972. “Sexual and other long term as-pects of imprinting in birds and other species.” InAdvances in the study of behavior, ed. D.S. Lehr-man, R.A. Hinde, and E. Shaw, pp. 147–174. NewYork: Academic Press.
Meehan, C.L., J.P. Garner, and J.A. Mench. 2003.Isosexual pair housing improves the welfare ofyoung amazon parrots. Applied Animal BehaviorScience 81:73–88.
Millam, J.R. 1994. “U.C. Davis amazon breeding proj-ect.” Proc Assoc Avian Vet, pp. 403–408.
Pearn, S.M., A.T. Bennett, and I.C. Cuthill. 2001.Ultraviolet vision, fluorescence and mate choice in a
parrot, the budgerigar Melopsittacus undulatus.Proc R Soc Lond B Biol Sci 268 (1482):2273–2279.
Rajecki, D.W., S.J. Suomi, E.A. Scott, and B.Campbell. 1977. Effects of social isolation and so-cial separation in domestic chicks. DevelopmentalPsychology 13 (2):143–155.
Rowley, I. 1980. Parent-offspring recognition in acockatoo, the galah, Cacatua roseicapilla. Austr JZool 28 (3):445–456.
Ryan, T. 1999. “Use of light in aviculture and avianmedicine.” Proc Assoc Avian Vet, pp. 179–186.
Seibert, L.M., and S.L. Crowell-Davis. 2001. Gendereffects on aggression, dominance rank, and affilia-tive behaviors in a flock of captive adult cockatiels(Nymphicus hollandicus). Applied Animal BehaviorScience 71:155–170.
Shields, K.M., J.T. Yamamoto, and J.R. Millam. 1989.Reproductive behavior and LH levels of cockatiels(Nymphicus hollandicus) associated with photostim-ulation, nest-box presentation, and degree of mateaccess. Hormones and Behavior 23 (1):68–82.
Smith, I.L. 1999. “Basic behavioral principles for theavian veterinarian.” Proc Assoc Avian Vet, pp. 47–55.
Sparks, J., and T. Soper. 1990. Parrots: A natural his-tory. New York: Facts on File.
Styles, D.K. 2001. “Captive psittacine behavioral re-productive husbandry and management: Socializa-tion, aggression control, and pairing techniques.”Proc Assoc Avian Vet (Avian Specialty AdvancedProgram), pp. 3–14.
Thorsen, M., R. Shorten, G. Buchanan G, et al. 1998.“Echo parakeet management report from the Mau-ritian Wildlife Foundation.” Proc Internatl AvicultSoc, pp. 1–31.
Vidal, J.M. 1980. The relations between filial and sex-ual imprinting in the domestic fowl: Effects of ageand social experience. Anim Behav 28 (3):880–891.
Wiley, J.W. 1980. “The Puerto Rican parrot (Amazonavitatta): Its decline and the program for its conserva-tion.” Proc ICBP Parrot Working Group Meeting,pp. 133–159.
290 Manual of Parrot Behavior

PARROTS AS PETS
Parrots are wild animals. Most species, althoughnow domestically bred, are no more than one ortwo generations away from their wild ancestors.This not only means that parrots still have the be-havioral repertoire of the wild birds but most ofall that they become socially mature. Our domes-tic companion animals, and especially dogs, makegood pets because they are neotenized. Thatmeans they retain juvenile characteristics intoadulthood and never become socially maturewhen compared to their wild relatives. They staybabies and readily accept our caregiving all theirlives.
Parrots, on the other hand, become socially ma-ture. That is the time when many people start hav-ing problems with them. We believe that reachingsocial maturity is much more important for thedevelopment of behavior problems than attainingsexual maturity. The two are not the same, andsexual maturity may be achieved earlier than so-cial maturity. This is obvious in human teenagers,in puppies, and likely applies to parrots as well. Ifthe owner has not put a great deal of effort intoearning the parrot’s trust and affection, and hasnot communicated clear contingencies relative tothe bird’s behavior (i.e., set consistent rules), be-havior problems often ensue.
This is often accepted as a fact of life. One ofthe authors (Luescher) remembers discussingdifferent species with a breeder and being toldthat one species was better as a pet because itstayed nice longer, implying that sooner or later
all parrots lost their pet qualities. This does notneed to be the case, as many happy long-termrelationships between owners and parrots last.We hope that this and other chapters in this bookwill help set the groundwork for a happy, long-lasting, and enjoyable relationship between par-rots and owners.
Parrots are highly intelligent and remember fora long time (see chapter 13). Parrots are a preyspecies and their first reaction to novel stimuliwill be fear and avoidance. This is important toremember when new toys or other cage furniture,house decorations, or other pets are introduced.As a prey species they are very intolerant of phys-ical or emotional abuse. In nature, physical re-straint would likely mean death for a parrot and istherefore very frightening for pet birds unlessthey have been desensitized to it. If they are phys-ically or emotionally abused even just once, it cantake a very long time for them to overcome theirfear and distrust.
Parrots are high-input companions and need astable and consistent environment. Most speciesare also very long-lived. Before deliberatingwhat species to obtain, potential owners shouldask themselves if they are really ready and will-ing to have a bird. They should think about beingable to provide the necessary environment andcare for a long time to come. If they are youngand plan on having children, they may not be ableto make that commitment. Certain rules have tobe followed to accommodate a parrot in a youngfamily.[1]
291
26Housing and Management
Considerations for Problem Prevention
Andrew U. Luescher and Liz Wilson

Because parrots are high-input companions, anowner should generally only get one bird at a time(the exception may be when an owner wants topair-house two birds of a small species, is willingand able to work with each separately, and can ac-cept that there may be some territorial or mate-related aggression later on). There is a tendencyin the United States for the number of birds perbird-owning households to increase. We supposewith all the different, beautiful species available,a collector instinct is easily awakened in manyowners. Recently one of the authors (Luescher)talked to people who bought 14 parrots in fivemonths. As long as they are all babies, all seemswell. However, it is not humanly possible (unlessmaybe for a professional trainer who can devoteall day to the birds) to train all these birds to thepoint that there won’t be any problems once theybecome adults. A parrot needs daily one-on-oneattention and training if he or she is to make agood pet.
Parrots are often obtained for the wrong rea-sons. Many birds are impulse bought because oftheir beauty, their potential ability to talk, theircuteness as babies, and their apparent ability tomake the owner feel “cool.” Many people have noidea what they are getting into when they bringhome a parrot. Most people do not understandwhat kind of work is involved when they purchasetheir first parrot, and well over half of ownerswould not replace their parrot if it died.
The reality is that most humans are not pre-pared—physically or psychologically—to sharetheir life with a wild animal. People generallywant a pet animal that considers them (thehuman) to be the center of the universe, an animalthat offers unconditional love. Indeed, dogs arethe only animals on this planet that fulfill theserequirements. Parrots do not fulfill these obliga-tions. Parrots are not dogs with feathers, nor arethey feathered children. They are simply them-selves, and they are unable to make the compro-mises that the average human expects from acompanion animal.
CHOICE OF SPECIES
Generally, most people are attracted to the largeand expensive species because of the status theyconvey. At avicultural meetings the influence andcredibility of people seem to be directly corre-lated to the price of their birds. However, few peo-
ple are capable of offering a Hyacinth Macaw orMoluccan Cockatoo the accommodation and careit needs. Most people would be better off with asmall species such as a Budgie, Cockatiel, orQuaker Parakeet. Most of all, these small birdsare better off as pets because they can be providedwith more room, more appropriate play gyms,and so forth. They also are not as intimidating,make less of a mess, and do less damage whenthey bite or chew. Their pet quality is equal to orbetter than that of many large and expensivebirds. Small birds are therefore less likely to berelegated to a solitary, deprived life in the garageor basement.
When choosing a species, size, manageability,personality, and ability to cope with life as a soli-tary bird should be the key factors. The amountand kind of noise a bird makes will in part dependon the species, as will the amount of dust. Talkingability and beauty are certainly factors to consideras well, but they should be considered secondaryto the others. A plain-colored bird that does nottalk may be more likely to provide lasting joy thana beautiful talking bird that becomes emotionallyunstable because the owner can’t manage it.
CAGING
Parrots will naturally use different habitats fordifferent activities. They often forage in areas thatare quite different from the place where theyroost. Therefore, pet parrots should have at leasttwo cages, one for the night and one for the day.Using the two-cage system or “multiple habitathousing” has several advantages. It forces theowners to take the bird out of its cage and handleit at least twice a day. It likely reduces cage terri-toriality. It makes it much easier to place the birdin the cage before leaving for work and makes lifemuch more interesting and more natural for thebird.
The night cage doesn’t need to be big and canbe sparsely furnished, but it has to provide secu-rity, that is, near a wall, not too low, maybe partlysheltered. The night cage should be placed in aquiet room away from the main living/activityarea, where it will be dark after 9 or 10 P.M., sothat the bird can get a regular and sufficient darkperiod.
The day cage needs to be as large as possible.In particular, the day cage should be horizontalrather than vertical. In a vertically oriented cage,
292 Manual of Parrot Behavior

the bird will sit at the top most of the time andmove around little. In horizontal cages, it canmove around without having to give up height andmay therefore be much more active. A horizontalcage also allows for arrangement of toys andother furniture in places where they are not likelyto be soiled. For small species, it may even allowsome flight inside the cage. If fitted with aproper-size door, it will also allow enough elbowroom for the owner to train the up and down com-mands in the cage. Cages should be rectangularrather than round. Round cages allow no corner tosit in for a nervous bird. They are also usuallyrather high but small in diameter. The round, tall“tower” design is the most misguided cage de-sign. Unfortunately, most cages offered in mostpet stores are not suited for housing birds.
Cage placement is an important issue, as thiscan dramatically influence a parrot’s stress levelsand, therefore, the stress levels of the humansaround it. The ideal location is dependent on thepersonality of the particular bird. An extrovertedparrot that is caged away from human activities islikely to scream excessively as it calls to other“flock members.” Anxious, high-strung parrotsmay show stress behaviors like feather destruc-tion if caged in the middle of a high traffic area,especially if the cage shares a wall with a door. Ifso, the bird is continually jolted by people appear-ing abruptly. Many birds enjoy a window view,but cage placement directly in front of a windowcan cause stress if the animal cannot hide or avoiddirect sunlight.
In general, the day cage should therefore be lo-cated where there is an appropriate amount of ac-tivity for the bird’s personality. However, it shouldbe placed against a wall so that the bird has atleast one direction toward which it does not needto be vigilant. Threatening stimuli should beavoided. For some birds, these may include appar-ently innocuous things such as clocks or pictures.The cage should be placed so that the bird doesnot get surprised and startled by a person sud-denly entering its visual field. By placing thecage partially against a window and partiallyagainst a solid wall, the bird is allowed the choiceto be exposed to the outside or not and to avoiddirect sunlight. This appears critical to decreasingstress in the companion parrot.
Cages can be made easily and cheaply fromgalvanized wire. In a species that is not generally
a strong chewer, and if the cage is big enough andfurnished with enough things that are more attrac-tive to chew on, this may be an acceptable solu-tion. Zinc toxicity is a concern, at least in somespecies, and with poor-quality galvanization.[2]However, it has to be kept in mind that mostbreeder birds are kept in galvanized cages, usu-ally without problem.
Having a play gym on top of the cage is dis-couraged because it makes the cage dark and maybe conducive to territorial aggression (the playgym being the same area as the cage). It alsotempts the owner to let the bird in and out of thecage on its own and discourages regular handling.The bird can also not enjoy any change in sceneryif it is only either in or on top of the cage. If it al-ways remains in the same constant environment,it may become overreactive to any change.
Cages should open wide: the door should coveralmost the whole front. Otherwise it will be moredifficult to remove a bird, and the bird will not betaken out as often. Also, small doors make train-ing the parrot in the cage (training to step up anddown in the cage) difficult. Thus, owners tend tojust open the door and let the bird leave and enterthe cage of its own free will. However, this willlikely cause problems with territorial aggression.
Commonly, many cage companies equip theircages with huge perches that are too large for anybut the largest psittacine species. Appropriatelysized perches will allow a bird to wrap its feetmost of the way around, thereby providing astronger, safer grip. Due to its resistance to psitta-cine beaks, companies often use manzanita forperches, and this wood creates problems for manyparrots. Extremely smooth and slippery, it can in-crease the potential for accidents. Naturalbranches with bark provided as roosts providemore foot comfort due to softness and irregularthickness and provide a substrate for chewing.Natural branches can be arranged so that the foodand water, as well as the perches themselves,don’t get soiled. While contamination with wildbird droppings is a concern, one of the authors(Luescher) knows of many large breeding facili-ties, including Loro Parque, that provide freshbranches on a regular basis without problems.Some trees are toxic in themselves or sprayedwith toxic chemicals, so only branches of knownorigin and species, without obvious contamina-tion with bird droppings, should be used. Soft
26 / Housing and Management Considerations for Problem Prevention 293

wood provides much more recreational qualitiesthan non-destructible hardwood. Trees that one ofthe authors (Luescher) has experience with arepine, soft maple, tulip, and various poplar species,as well as wild grapevines.
Cages need to be designed so they can becleaned and sanitized easily. Flooring where thedroppings fall through (cage grates) is probablypreferable for most species.
For prey species, hiding places are also impor-tant, and parrots should be allowed the choice tobe invisible to others in the environment. Hidingplaces can entail anything from branches attachedwith electrical ties or cotton string to the outsideof the cage to produce a pseudo thicket (M.S.Athan, personal communications, 2000), to asheet thrown over one corner, to wooden sheltersattached to the side of the cage. Enclosed shelterscan work well for young parrots, but they areoften perceived as nests as parrots mature. Matureparrots can be accommodated with incompleteshelters such as a three-sided and roofed con-struction with a perch but lacking a front andfloor (see chapter 18 on aggression).
Cage height is important. If caged too low, aninsecure parrot can become seriously frightened.If caged too high, some individuals may be morefearful and more difficult to handle. Many parrotsappear most comfortable when allowed to perchat human chest or shoulder level.
PLAY GYMS
Play gyms can be bought for lots of money but areoften uninteresting and provide a very limitednumber of roosting places. Play gyms are muchmore interesting when large, with varied types ofperches, and destructible (or holding lots of de-structible toys). Unfortunately, many commer-cially available play gyms are small due to priceconsiderations, and they are made out of inde-structible material such as manzanita, plastic, oreven stainless steel.
Play gyms can easily be made (at least forsmall species) from under-bed plastic storagetrays and branches or vines. Plastic storage traysof various sizes are useful. Four fresh branchesare each placed with the thick end in one corner,leaned toward each other, and tied together abovethe center of the tray (Figure 26.1). They providenovelty, interesting shape, and destructibility tothe bird at a minimal price. Since they are very
cheap, a number of them can be set up in severalrooms of the house, so it will be possible to takethe bird along from room to room and have a des-ignated place for him or her. One can be placedon a side table beside the dining room table, andthe bird can be trained to stay on it while the fam-ily eats, receiving his or her own treats on the playgym.
A particularly nice play gym or indoor tree canbe made from a chimney flue and naturalbranches. On a vinyl floor cover (or the like), thechimney flue is placed upright. The branches arethen stuck into it like into a vase (no water isadded, of course). Soon, the birds will climb upand down in their tree and remove all the leavesand small twigs. The floor underneath will soonlook like a forest floor but can easily be cleanedup. Their enjoyment is well worth the trouble. Asimilar idea is to use a Christmas tree stand andput a branch in it. Obviously, all these gyms haveto be stable, especially for large species and whenchildren or other pets are around. Once these playgyms don’t look good any more, the branches canbe used to start a fire in the fireplace, and newbranches are brought in.
ROUTINE
Birds like routine. This does not mean that thingshave to happen at the same time each day. In fact,it is a good thing to get the bird used to some vari-ation in routine from an early age, so that it is lessstressful for them when the routine is broken.However, they like important events, such asfeeding or being taken out of the cage, to happen
294 Manual of Parrot Behavior
Figure 26.1. A beautiful Scarlet Macaw (Aramacao) at Palmitos Park.

at the same place in the routine. This way, eventsbecome predictable. Just as important is that thebirds learn that at certain times in the routine theyare not getting any attention, and they will be con-tent being ignored at those times.
As a basic routine, birds should be removedfrom the night cage in the morning, taken alongthrough the house to some of their play gyms,maybe have breakfast with the owners (but theyshould also be trained that sometimes they can’tparticipate in meals, for when guests are over),and eventually be placed in the day cage. To facil-itate placing the bird into the cage, a treat or for-aging device should be placed in the cage. Usinga separate day and night cage makes it much eas-ier anyway to remove a bird from its cage and toplace it into the other cage. In the evening a sim-ilar routine should be practiced, and the birdshould be placed in the night cage at a fairly reg-ular time.
Parrots, especially youngsters, need a suffi-ciently long dark period each night—probably asmany as ten hours. Many companion parrotspecies are equatorial in origin, and thereforeevolved in an ecosystem that provided 12 hoursof darkness, year-round. As previously men-tioned, due to their social nature, parrots areoften caged in high traffic areas, and this oftenputs them in the same room with the family TV.When questioned about sleep, owners generallycount the hours from the cage being covered untilit is uncovered the following morning. However,logic indicates that a prey species is unlikely tosleep deeply when a predator (human) is movingnear its roost. As a consequence, the real measureof the bird’s sleep period is the time between thelast person leaving the room at night and the timethe first person arises in the morning. Behavioralmanifestations of sleep deprivation would in-clude hyperactivity, aggression, excessivescreaming (especially after sunset), and featherdestruction.[3]
The feeding schedule contributes to routine aswell. Free choice feeding is the most commonmethod of feeding pet birds. While this is proba-bly the simplest method and assures adequate ac-cess to food, it is not really natural. Wild parrotsspend a substantial part of their time foraging forfood. The activities involved in their foraging be-havior are the same behaviors we associate withplay behavior in pet birds. Chewing, manipulat-
ing, and investigating are all part of the process offinding food. In captivity, birds have enormousquantities of calorie-dense food placed in easilyaccessible bowls at all times. Dr. Ted Lafeberoften recommended meal-feeding pet birds.[4]He felt that meal-feeding might encourage greateractivity by allowing birds to “forage” when foodwas not directly in front of them. In addition, hefelt that when given some time with no food infront of them, birds might be somewhat more ad-venturous about trying new food items. A com-promise may be found by offering pellets freechoice and fresh items in the morning andevening, when the birds’ foraging activities natu-rally peak.
Other factors that can be important regardingfeeding schedules regard the freshness of thefood. Fresh food items can spoil after a few hoursand sometimes will attract insects. The health im-plications of this are obvious, but also some birdswill be very upset when they see flies or other in-sects flying around their food dishes.
TOYS
Toys are a valuable means of encouraging devel-opment in pet birds. The activities involved in toyplay can promote learning, relieve stress, and oc-cupy idle time.
Natural branches keep birds better occupiedand meet their motivation to forage and chewmore than commercial toys. Some commercialtoys with tassels are conducive to facilitate redi-rected social grooming and therefore may helpmeet this special need. Foraging devices are veryuseful. Some are commercially available but theycan also be homemade. Seeds can be pressed intoa squash or placed between the leaves of a half ofa cabbage. They could also be strewn betweenlarge pebbles. However, size is important. One ofthe authors (Luescher) tried this with too small apebble size. His house looked like a gravel pitwhen he came home because the birds flung allthe pebbles out of the cages. Sometimes a wholeear of sweet corn in the husks can be provided.Large chunks of vegetables and fruit can be givenon skewers (see Figures 26.2 to 26.14).
Safety has to be kept in mind with toys. Sometoys with threads can result in a bird strangling it-self. Also, toys with rings or large chain links,through which the bird can put its head, should beavoided. To keep toys interesting, they should be
26 / Housing and Management Considerations for Problem Prevention 295

296
Figure 26.2. A Maroon-bellied Conure(Pyrrhura frontalis). See also color section.
Figure 26.3. A Red-masked Conure (Aratingaerythrogenys). See also color section.
Figure 26.4. A homemade, roomy cage for twolarger conures (Sharp-tailed Conures, Aratingaacuticaudata).
Figure 26.5. An appropriate play gym for smallto medium-sized birds, made from a chimneyflue and natural branches. See also colorsection.

297
Figure 26.6. Two Sharp-tailed Conures (A.acuticaudata) enjoying natural branches. Seealso color section.
Figure 26.9. Some birds enjoy joining theowner in the shower on a commercially availableshower perch. See also color section.
Figure 26.7. A play gym made from a chimneyflue and natural branches after the birds spentsome time on it. See also color section.
Figure 26.8. A smaller play gym made from astorage container and wild grape vines. See alsocolor section.

rotated (i.e., only a few provided at once and ex-changed for others every few days). Toys towardwhich the bird redirects social preening shouldstay in the cage permanently, however, as long asthe bird is not aggressive when near them.
GROOMING
In most situations, the wing feathers of pet birdswill have to be clipped for safety reasons. Overlyaggressive and cosmetically pleasing grooming
styles can, however, be devastating for a bird.Clipping style should be individually adjusted toeach bird so that the bird can still fly, although notupward. Clipping should always be symmetricalto allow the bird to control the direction of theflight. Clipping too severely can result in trau-matic falls and loss of self-confidence. It can alsoincrease the danger that the parrot may fall into oronto something harmful or become a victim ofanother pet, such as a dog.
298 Manual of Parrot Behavior
Figure 26.10. Two different devices to sprayparrots. The regular spray bottle on the left isadequate for individual birds. The pressuresprayer on the right is most suitable for largernumbers of birds.
Figure 26.11. Some birds prefer to wet them-selves by taking a bath in a water dish. In a largecage, a water dish can be positioned so thatfecal contamination is minimized.
Figure 26.12. A Mitred Conure (Aratingamitrata) with a “foot toy,” a toy commerciallyavailable for cats. See also color section.
Figure 26.13. Large pieces of fruits andvegetables provided on a skewer encourageforaging.

TRAINING AND INTERACTION
Petting, cuddling, kissing, and allowing birdsonto the shoulders are activities that will tend toencourage an inappropriate pair bond betweenowner and bird. Conversely, some activities en-courage a bird to flock bond. Toy play, training,verbal games, dancing, and exercise are all activ-ities that encourage a healthier bond. Time spenton a perch with the family without being held isan extremely valuable activity for pet birds. Thiscan encourage the bird to be more independent.As flock animals, wild parrots spend very littletime alone. The protective presence of the flockgives them a much-needed sense of security in thedangerous world in which they exist. Nonethe-less, confident pet parrots tolerate a certainamount of “alone time” well.
Training is discussed extensively in chapter 15.The only thing that shall be emphasized here isthat training goes a long way toward problem pre-
vention. Trained birds can be controlled betterand are more likely to stay out of trouble. Mostimportantly, training makes the interaction be-tween owner and bird much more consistent, pre-dictable, and stress-free. A relationship based onteaching and learning is much healthier than onebased on petting, cuddling, and, inadvertently,sexual stimulation. The basic commands used inbird training are the “up” and “down” commands.They should be practiced many times every day,and the bird should be rewarded most times forperforming the behaviors throughout its life. If abird is in full flight, it should also be trained to“come” reliably when told. In addition, manytricks can be taught (for the bird, all trained be-haviors are tricks). Training should start early inlife, ideally with the breeder, and continue for therest of the parrot’s life on a daily basis. No treatsshould be given to a parrot without a specific pur-pose. Giving treats outside of a training context(in the widest sense, including, e.g., a desensitiza-tion procedure) will result in missed opportunitiesto train, make the interaction between owner andparrot less consistent and thus more stressful, andresult in begging.
REFERENCES
1. Doane, B.M., and T. Qualkinbush. 1994. My parrot,my friend. New York: Howell Book House, p. 20.
2. Holz, P., J. Phelan, R. Slocombe, A Cowden, M.Miller, and B. Gartrell. 2000. Suspected zinc toxi-cosis as a cause of sudden death in orange-belliedparrots (Neophema chrysogaster). J Avian MedSurg 14:37–41.
3. Wilson, L. 1999. Sleep: How much is enough for aparrot? Pet Bird Report 43:60–63.
4. Lafeber, T.J. 1998. Let’s celebrate pet birds. Odell,IL: Lafeber Co.
26 / Housing and Management Considerations for Problem Prevention 299
Figure 26.14. Half of a Savoy cabbage withseeds strewn in between its leaves.

INTRODUCTION
The issue of captive parrot welfare is both timelyand ethically significant. Parrots are kept in a va-riety of captive situations including breeding fa-cilities, wholesale/retail facilities, zoos, shelters,sanctuaries, entertainment venues, and conserva-tion programs. In addition, the pet parrot industryhas grown tremendously over the past decade, andparrots are now the third most popular companionanimals in the United States (AVMA 2002).Recent estimates from the American VeterinaryMedical Association reveal that there are 10.1million parrots in 4.6 million homes in this coun-try alone (AVMA 2002). As they become moreand more popular, it is becoming increasingly ev-ident that ensuring a good state of welfare forcaptive parrots is not an easy task.
Parrots and other birds whose environmentaland behavioral needs are not met in captivity mayengage in a variety of distressing abnormal be-haviors, including incessant screaming, stereo-typic pacing, and feather picking that becomes soextreme that the bird denudes itself and evencauses skin or tissue injury (Grindlinger 1991;Davis 1991; van Hoek & ten Cate 1998; Meehan& Mench 2002; Meehan et al. 2002). These birdsmay also become excessively fearful or aggres-sive, directing their aggression toward familymembers or family friends. Improper care and en-vironmental conditions may also result in psycho-logical and physiological problems includingacute illness, injury, and even sudden death.Considering the life span of many birds (Budgeri-gars may live 15 years, Cockatiels 25–30 years,and large parrots up to 70 years), problems likethese may prove to be overwhelming for compan-ion bird owners, leading to the bird being abused,neglected, sold, given away, released, or relin-
quished to a shelter or sanctuary. A 2001 censusof ten parrot rescue and sanctuary organizationscompiled by the Gabriel Foundation revealed thatthere were over 2,000 parrots and small birdshoused at these organizations (J. Murad, personalcommunication, 2001), and there is anecdotal ev-idence of an escalation in the numbers of birdsbeing relinquished by private owners. For captiveparrots, there are now many of the same overpop-ulation problems as for dogs and cats—too manyunwanted animals and not enough qualifiedhomes. Part of the problem is that there is notenough information available to parrot owners re-garding parrot welfare, including what parrotsneed in order to maintain good welfare and howpoor welfare can be assessed and improved.
The main objective of this chapter is to exam-ine some of the welfare problems facing parrotsin a companion context in order to gain an under-standing of their causes and potential solutions.Many of the principles discussed in this chapterwill also be relevant to parrots housed in zoos,aviaries, and other captive settings. First, how-ever, we will discuss some of the concepts associ-ated with the study of animal welfare in general.
WHAT IS ANIMAL WELFARE?
In the last few decades, there has been increasingdiscussion about human ethical obligations to-ward animals, which has been reflected in a grow-ing body of legislation, particularly in Europe, de-signed to protect the welfare of farm, laboratory,and companion animals. Although it is beyondthe scope of this chapter to provide details of thevarious ethical theories about human-animal rela-tionships that have been proposed, all are basedon a common assumption: that animals have in-terests (e.g., an interest in avoiding pain and suf-
301
27Captive Parrot Welfare
Cheryl Meehan and Joy Mench

fering) that deserve consideration by humans.This assumption is now also widely accepted bymost people, at least in developed countries, al-though there is still considerable disagreementabout the nature of animals’ interests and howthey should be weighed against human interests.
This ethical concern for the quality of life ofanimals has led to the development of the field ofanimal welfare science. The goal of this field is todevelop methods for assessing and improving thewelfare of animals in a variety of settings includ-ing laboratories, zoos, farms, shelters, and privatehomes. Welfare is viewed as something intrinsicto the individual animal, that is, the animal’s stateof being well or “faring” well. That state of well-being has both a physical and psychological com-ponent and can be influenced by many factors in-cluding the animal’s housing, care, physicalhealth, and interactions with humans.
There are three broad approaches to the scien-tific study of animal welfare (Duncan & Fraser1997). “Feelings-based” approaches equate ananimal’s welfare with its subjective experience.Proponents of this approach place primary em-phasis on the reduction of pain and suffering andthe provision of comfort and pleasure. Dawkins(1988, p. 209) sums up the spirit of this approachwith this quote: “To be concerned about animalwelfare is to be concerned with the subjectivefeelings of animals, particularly the unpleasantsubjective feelings of suffering and pain.” Obser-vation of behaviors such as avoidance, aggres-sion, and vocalizations can be used to gain infor-mation about the internal experiences of animalswith respect to fear, pain, and distress. Determi-nation of what animals prefer and what theywould rather avoid are also important for evaluat-ing subjective feelings.
A second approach, known as the “functioning-based” approach, uses the biological state of theanimal as the main criteria by which to judge wel-fare. Normal functioning of biological and behav-ioral systems is essential to good welfare fromthis perspective. Thus, disease, injury, failure toreproduce, and the performance of abnormal be-haviors are considered evidence of compromisedwelfare.
A third approach operates under the assump-tion that in order to provide good welfare for acaptive animal we should allow it to perform all“natural” behaviors and raise it in an environment
that is as close to a natural environment as possi-ble. Research approaches appropriate to this viewinclude using the behavioral repertoire of animalsin the wild as a guide for evaluating the welfare ofcaptive counterparts of the same or similar spe-cies. Thus, from this perspective, parrots in cap-tivity that are unable to perform behaviors such aslocomotion, social interaction, and foraging forfood would have compromised welfare.
These different approaches to conceptualizingand assessing animal welfare are not necessarilymutually exclusive. For example, problems thataffect the functioning of animals can also affecttheir feelings: a parrot that injures itself whenfeather picking will have reduced biological func-tioning and also experience pain as a result of theinjury. However, there are other situations inwhich the relationships among indicators of wel-fare are not straightforward. Laying hens kept incages without nests pace and vocalize franticallybefore laying their eggs, which is an indicator ofdistress, but show no obvious impairment of re-production or health (Appleby et al., in press).
Fraser et al. (1997) suggest that the three ap-proaches can most usefully be integrated by un-derstanding that all animals possess adaptationsthat have arisen during the course of evolution. Inthe wild, these adaptations are reflected in an an-imal’s normal behavioral repertoire. Some adap-tations, however, may not be necessary in the cap-tive environment because their original functionis achieved in some other way. For example, adap-tations used to regulate body temperature duringcold weather are not necessary if animals are keptin comfortable, temperature-controlled environ-ments. Depriving the animal of the ability to per-form the behavior associated with such an adap-tation will not affect the animal’s quality of lifeunless the behavior is also motivated by a strongaffective (emotional) experience, like hunger or adesire to escape. In this situation, the animal’sfeelings, and hence its welfare, will be compro-mised even if its biological functioning is not. Forexample, captive animals are sometimes feed re-stricted because otherwise they become obesedue to lack of exercise. While this actually pro-motes good physical health, the animal still feelshungry and may even develop abnormal behav-iors, like chewing on cage bars or ingesting non-food items (pica), in an attempt to diminish itshunger.
302 Manual of Parrot Behavior

Welfare problems can also arise if animals lackthe adaptations necessary for particular features ofthe captive environment. Atmospheric ammonialevels can be high in farm and laboratory environ-ments, but most animals do not have evolutionaryadaptations to environmental pollutants and mayshow no avoidance reactions to high ammonia en-vironments, even though exposure eventually re-sults in damage to the eyes and respiratory system,and hence impairment of biological functioning.Lastly, welfare problems can arise when animalspossess the appropriate adaptations, but the adap-tations are inadequate to the degree of challengeimposed within the captive environment. In thiscase, there is likely to be a high correspondencebetween feelings and functioning (e.g., feeling hotand showing signs of heat stress). These examplesillustrate how important it is to assess multiplemeasures and responses to gain the most accuratepicture of the animal’s welfare.
THREATS TO PARROT WELFARE
The main threats to the welfare of captive parrotsfall into three categories: husbandry, environ-ment, and human interactions. Husbandry-relatedthreats include such things as poor nutrition (fora full discussion, see chapter 6), unsanitary con-ditions, lack of veterinary care, improper light/dark cycles, inappropriate temperature, and lackof opportunity for bathing. Environmental issuesinclude lack of space, threats to safety such asdangerous objects, improper bar spacing or po-tential for escape, improper perch size, barrencages, social isolation, and lack of opportunityfor privacy. Threats due to human interactions in-clude abuse, neglect, improper taming/trainingtechniques, and sexual bonding between parrotsand human companions.
In most cases, these threats will manifest them-selves in the parrots through physical, psycholog-ical, or behavioral indicators associated with poorwelfare. Physical indicators include disease, in-jury, parasites, obesity, and malnutrition. Apathyand anhedonia (loss of interest in things the par-rot once enjoyed such as favorite foods and toys)are potential psychological indicators of poorwelfare. Behavioral indicators include abnormalbehaviors such as stereotypy (repetitive, invariantperformance of a behavior, such as pacing),feather picking, incessant screaming, and exces-sive aggression or fearfulness. Sometimes these
indicators are observed shortly after the threat towelfare has been imposed. For example, an injurymay result immediately after an unsafe toy isplaced in the cage. However, other indicators ofpoor welfare may not be obvious for days, weeks,or even years. During the intervening time, theparrot may be attempting to cope with the threatand may show no outward signs of poor welfare.For example, sick birds often do not show anysigns of illness until they are in an advanced dis-ease state, and birds may live for long periods oftime in barren conditions before developing ab-normal behaviors. Despite the fact that outwardlythese parrots appear and act normally, biologicaland behavioral changes may be occurring thatwill eventually result in overt signs of poor wel-fare. At that point, reversing these changes, andthus returning the parrot to a state of good wel-fare, may be difficult or impossible. For this rea-son, it is imperative that owners thoroughly un-derstand the threats to their parrots’ welfare andstrive to eliminate these threats from their parrots’lives as soon as possible. Prevention of poor wel-fare is the best solution to poor welfare.
While there are many threats to companionparrot welfare, space does not permit a thoroughdiscussion of them all. As such, the remainder ofthis chapter will be focused on the behavioral ef-fects of environmental threats to welfare. We willaddress the connection between the captive envi-ronment and several of the most common behav-ioral problems experienced by parrots. We willalso provide information that will assist compan-ion parrot owners with the design of a captive en-vironment that can successfully prevent abnormalbehaviors from occurring, and sometimes evendecrease or abolish already-established abnormalbehaviors.
THE CAPTIVE ENVIRONMENT ANDPARROT WELFARE
As previously mentioned, animals have evolvedto respond adaptively through behavior tochanges in their environment. They have alsoevolved the ability to change their environment tomeet their needs and goals. In the course of evo-lution, animals have been selected, in consider-able part, based upon the strength of these abili-ties. In the wild, behavioral skills are usedregularly in the course of capturing or gatheringfood, avoiding predators, finding or creating shel-
27 / Captive Parrot Welfare 303

ter, and choosing or attracting mates. In captivity,most needs of animals are met without their directparticipation—food arrives at regular intervalspre-killed, cooked, or peeled; shelter is providedand often is immutable; social groups are selectedby humans; and predators are kept away by barsand glass. On one hand, a controlled environmentmay seem ideal as it provides freedom from foodscarcity, predation, and extreme climatic changes.But at the same time, many captive environmentsare quite limited with respect to space, complex-ity and behavioral opportunity when compared tothe environments of free-living individuals. Thisis the paradox of life in captivity—in an attemptto reduce exposure to environmental pressures,we prevent parrots from exhibiting many of theirnatural behaviors, creating a mismatch betweenthe parrot and its environment. If this mismatch is significant, normal behavioral expression isprevented and abnormal behaviors can developinstead.
In order to understand how the captive environ-ment might affect the welfare of parrots, it is nec-essary that we first understand the behavior ofparrots in the wild and how this diverges from thebehavior of parrots in captivity. It is also neces-sary to understand parrots in the context of do-mestication to appreciate how behaviorally closecaptive parrots are to their wild counterparts.
PARROTS IN THE WILD
In the wild, parrots are constantly engaged withtheir environment whether it is via social interac-tions, foraging activities, territory defense, nestselection, or predator avoidance. When broughtinto captivity and placed under close environmen-tal control, the opportunity to perform many ofthese behaviors is reduced.
Two of the most severely constrained classes ofbehavior in captive parrots are foraging and loco-motion. Parrots in the wild regularly travel severalmiles between feeding sites, and once they arriveengage in a rich suite of local search, food selec-tion, and food manipulation behaviors. For exam-ple wild Puerto Rican Amazons (Amazona vit-tata) spend approximately four to six hours perday foraging and are known to ingest the fruit,leaves, bark, vines, and/or other portions of atleast 58 species of indigenous plant material(Snyder et al. 1987). A study performed in 1985by Magrath and Lill characterized the activity
budget of a wild psittacine, the Crimson Rosella,in its native Australian habitat. Although the ac-tivity budget differed seasonally, young Rosellasin this study spent a mean time of 67% in feed-ing/foraging and only 7% of their time resting. Itis likely that other parrot species invest similaramounts of time in feeding behavior. In somecases the investment is made in searching forfood and in some cases in processing it. For ex-ample, the Gang-gang Cockatoo (Callocephalonfimbriatum) habitually feeds on the very well-protected nuts of the eucalyptus tree, expending aconsiderable portion of its time simply exposingthe edible portion of the item (Bauck 1999). Incontrast, most parrots in captivity do not travelbetween feeding sites, do not have to select differ-ent foods to balance their diet, and have little op-portunity to manipulate objects to obtain food.Thus, captive Orange-winged Amazon Parrots(Amazona amazonica) spend only 30–72 minutesa day in feeding behaviors when fed a pelleteddiet (Oviatt & Millam 1997). Many captive feed-ing methods allow minimal environmental inter-action and greatly reduce the amount of work andenergetic cost involved in feeding activities.Because of the importance of these behaviors inthe repertoire of wild parrots, it is possible thatcaptive parrots are highly motivated to search for,access, and process food items.
Wild parrots also exploit a complex physicalenvironment and, in addition to flight, show anumber of physical and behavioral adaptations tothis habitat. For instance, they utilize their grap-pling beak and grasping feet to negotiate treetopsand unstable fruit-bearing branches, are adeptclimbers of vertical surfaces, and are equallygraceful traversing the underside of branches(Sparks & Soper 1990). In captivity, parrots arerarely able to fly and are usually severely con-strained in the other locomotor behaviors theycan perform due to the design of their cage envi-ronment.
For most species of parrot, sociality is a con-stant feature throughout the lifetime of an individ-ual, although the characteristics of social group-ings can change daily and seasonally. Forexample, during the breeding season, wild ama-zon parrots tend to form small groups consistingof a pair and their young, but outside of the breed-ing season they can become highly gregariousand flock size can grow significantly (Gilardi &
304 Manual of Parrot Behavior

Munn 1998). The social environment of a parrotalso changes during development. As nestlings,parrots are restricted to social interactions withparents and clutchmates. As the birds fledge andbecome more independent, social interactions in-crease in diversity and complexity as the youngbirds are introduced into larger groups. In captiv-ity, companion and laboratory parrots are oftensocially isolated from conspecifics, although par-rots in zoos may be afforded more opportunity forsocial interaction.
It is evident from this review that foraging, var-ied locomotion, and social interactions are inte-gral components of the behavioral repertoire ofwild parrots. The next section demonstrates whythese same behaviors are important to captiveparrots as well.
PARROTS AND DOMESTICATION
Domestication is a process of adaptation to cap-tivity that includes both genetic changes occur-ring over generations as well as environmentallyinduced developmental events (such as taming)that occur within the lifetime of an individual(Price 1984). Because of their long lifespan (up to70 years or more), and the recent growth in pop-ularity of parrots as pets, many parrots currentlyin captivity were born to wild parents or areamong the first few generations of captive born.Unlike cats and dogs, which have been bred tolive as human companions for thousands of years,pet parrots are only a few generations out of thewild. Thus, although parrots may live in a varietyof captive situations, they can’t be considered do-mesticated. Since parrots are very early along inthe domestication process, it is likely that the ca-pacity to perform the behaviors seen in wildcounterparts remains (Price 1999). Even long-domesticated species such as Norway rats (Boice1981), rabbits (Vastrade 1986), and pigs (Wood-Gush et al. 1983) behave in a manner that isnearly identical to that of their wild ancestorswhen housed in “semi-natural” conditions.Although pet parrots are very similar behaviorallyand biologically to their wild counterparts, thetypical captive environment is a far cry from theenvironments parrots inhabit in the wild.
Recognizing that the wild can’t be re-createdinside a cage, the challenge to those interested inimproving parrot welfare by improving their envi-ronment is twofold: first, to determine which en-
vironmental elements are critical to normal be-havioral development and second, to developpractical methods of introducing these elementsin the captive context, a process referred to as en-vironmental enrichment.
ENVIRONMENTAL ENRICHMENT
The concept of environmental enrichmentevolved from a recognition that restrictive andbarren captive environments can impair behav-ioral and physiological development and con-comitantly reduce welfare. Shepherdson (1998,pg. 1) broadly defines environmental enrichmentas “an animal husbandry principle that seeks toenhance the quality of captive animal care byidentifying and providing the environmental stim-uli necessary for optimal psychological and phys-iological well-being.”
Common environmental enrichment strategiesinvolve changing the types of food or the methodsby which food is provided, providing social stim-ulation, changing structural features of animals’enclosures to provide cover or to encourage loco-motion, or exposing animals to novelty to facili-tate exploration. In mammals, appropriate enrich-ment has been shown to have many beneficialeffects, including facilitating a more normal anddiverse behavioral repertoire, decreasing stress,improving immune function, improving learningability, and decreasing fearfulness and the per-formance of abnormal behaviors (e.g., Renner &Rosenzweig 1987; Carlstead & Shepherdson1999; Bayne et al. 2002; Newberry 1995).
Although environmental enrichment is a poten-tially promising strategy for improving the wel-fare of captive and companion parrots, little isknown about the elements necessary for effectiveenvironmental enrichment for avian species ingeneral, and particularly for parrots (e.g., Birchall1990; Shepherdson 1993; King 1993). In fact,avian species are hugely under-represented whenit comes to innovations and research regardingenvironmental enrichment (King 1993). Themammalian bias present in the field of environ-mental enrichment may be due to the closer evo-lutionary relationship between humans and othermammals, the popular appeal of megavertebratessuch as carnivores and primates, or the perceivedlack of intelligence possessed by avian species(King 1993). Whatever the reason, birds in cap-tivity are potentially suffering due to lack of at-
27 / Captive Parrot Welfare 305

tention to their environmental enrichment needsor due to misguided attempts at enrichment thatare not based on scientific research.
In order to develop an enrichment program thatwill improve the welfare of captive parrots, it isnecessary to first identify those behaviors that aremost important to optimal behavioral develop-ment. Principles of environmental enrichment de-veloped through research with other species canbe used to inform the design of enrichment pro-grams for parrots. What is critical is that the en-richments be biologically relevant to the animaland be demonstrated to have positive effects onwelfare. For example, enrichment strategies thattake into consideration the motivational state andbehavioral skills of the particular species in ques-tion are generally more successful than strategiesthat are not based on species-appropriate hy-potheses (Newberry 1995). Thus, providingsingly housed rhesus monkeys with a foraging/grooming board consisting of a piece of Plexi-glass covered with artificial fleece with particlesof food treats rubbed into it improves welfare byincreasing foraging and grooming behaviors andreducing the performance of abnormal behaviors(Bayne et al. 1991), while providing sticks anddog toys has no effect on the performance of ab-normal behaviors in the same species (Line et al.1991). Similarly, the species-specific strategies ofintroducing live fish into the pool for fishing catsand hiding food in a brush pile in the enclosurefor leopard cats significantly reduce the perform-ance of abnormal behaviors (Shepherdson 1993).
For some animals, the physical design of thecage, rather than the feeding methods, must bemodified to reduce abnormal behaviors. Again,these modifications must be relevant to the ani-mal to be effective. For example, provision ofcover prevents stereotypies in young bank voles(Cooper et al. 1996), although increased cagespace does not (Odberg 1987). Weidenmayer(1997) was successful in preventing nearly all de-velopment of stereotypy in gerbils reared with ac-cess to a burrow as long as the burrow was pairedwith a tunnel-shaped entrance, which created aconfiguration similar to that of a wild gerbil bur-rowing system.
For species that have evolved in a social milieu,social stimulation may provide the most effectiveform of environmental enrichment because socialstimulation has dynamic qualities and is seldom
constant or completely predictable (e.g., Novak &Suomi 1988; Mendl & Newberry 1997). Al-though most species of parrots are social in thewild (Sparks & Soper 1990), companion parrotsare not often socially housed. In fact, recommen-dations in the popular literature on parrot caresuggest that pet parrots not be pair-housed be-cause they will become less desirable as pets(e.g., Blanchard 1999), although this has not beendemonstrated empirically. However, it has beensuggested that isolation from conspecifics maycontribute to the development of many abnormalbehaviors including excessive screaming, stereo-typies, fearfulness, and aggression (e.g., Wester-hof & Lumeij 1987; Lantermann 1993), so socialenrichment may, in fact, be an effective tool forimproving the welfare of companion parrots.
From a naturalistic standpoint, re-creating so-cial groups of parrots that are seasonally and de-velopmentally appropriate in a captive settingwould be considered the best approach to socialhousing. However, naturalistic social environ-ments necessarily involve increased risk of infec-tion as well as potential injury or stress due to ag-gressive encounters. In addition, the goals andrestrictions of most captive situations are notcompatible with the space, resource, and hus-bandry requirements of naturalistic social group-ings. Thus, pair-housing has been used in manycontexts and with many species as an alternativeto naturalistic social environments or individualhousing in an attempt to enrich the captive envi-ronment and improve welfare (e.g. Barnett et al.1984; Hughes et al. 1989; Reinhardt 1991; Chu etal., in press). Pair-housing can be a practical alter-native to single-housing for parrots because thespace requirements per pair are not extensive andnormal husbandry practices do not need to bechanged significantly.
There has been very little research on behav-ioral aspects of parrot welfare and of effective en-vironmental enrichment. In the following sec-tions, we therefore discuss some experimentalevidence derived from studies conducted at ourOrange-winged Amazon Parrot colony at theUniversity of California, Davis, that suggests thatenriching three aspects of the captive environ-ment can have significant positive effects on par-rot welfare. The three forms of enrichment testedin the studies outlined are providing foraging op-portunity, increasing the physical complexity of
306 Manual of Parrot Behavior

the cage, and allowing for social contact. The for-aging enrichments we utilized required the par-rots to perform behaviors such as chewingthrough barriers, sorting through inedible mate-rial, maneuvering objects through holes, or open-ing containers in order to access the food items.Physical enrichments provided alternate perchingsites, climbing or swinging opportunities, or mov-able objects that could be manipulated with thebeak and/or feet. In the cases where social enrich-ment was utilized, some parrots were housed insame-sex, same-age, non-related pairs, while oth-ers were singly housed. We found that these threeforms of enrichment successfully prevented andreversed the performance of several common ab-normal or undesirable behaviors and, as such, im-proved the welfare of the parrots involved. Thebehaviors studied were psychogenic feather pick-ing, fearfulness, aggression toward human han-dlers, stereotypic behavior, and screaming.
Psychogenic Feather Picking
Many owners of companion parrots have beenfaced with the frustrating dilemma of how to helpa feather-picking bird. In fact, it has been esti-mated that one in ten captive parrots performsself-directed psychogenic feather-picking behav-ior (Grindlinger 1991). It takes just one look at aparrot denuded or with self-inflicted skin andfeather damage for it to become obvious thatthere is great cause for concern. Psychogenicfeather-picking behavior develops or persists inthe absence of medical causes, and observationalevidence suggests that it may be associated with anumber of management factors such as inade-quate diet, social isolation, and lack of environ-mental stimulation (e.g., Mertens 1997). How-ever, despite the severity and prevalence of thisproblem, there has been very little research on theenvironmental correlates of feather picking.
Although there have been few systematic stud-ies of feather-picking behavior in psittacines,there has been a significant amount of researchon feather pecking, a similar behavior commonlyperformed by domestic fowl (Mench & Keeling2001). In chickens feather pecking is generallydirected at other birds, while in parrots picking isgenerally self-directed, but this difference maysimply reflect differences in management. Chick-ens are generally socially housed, while parrotsare often caged alone. When parrots are housed in
social groups, feather picking can also be directedat cagemates or nestlings.
Feather pecking by chickens is strongly associ-ated with the performance of foraging behavior(e.g., Nicol et al. 2001; Klein et al. 2000; Huber-Eicher & Wechsler 1997, 1998; Blokhuis 1986).In chickens, normal foraging behavior consists ofpecks directed at both edible and inedible sub-strates, but if chickens are housed such thatground pecking is prevented, then pecks may in-stead be directed at the feathers of conspecifics(Blokhuis 1986; Huber-Eicher & Wechsler 1997).Chickens may also consume the feathers that theymanage to pull out (McKeegan & Savory 1999).Provision of non-nutritive foraging material suchas long straw and polystyrene blocks is effectivein both preventing and reducing feather-peckingbehavior by chicks (Huber-Eicher & Wechsler1997). In addition, hens provided with foragingmaterial show significantly lower rates of featherpecking than those kept without foraging material(Wechsler & Huber-Eicher 1997). Thus, featherpecking is considered by many to be redirectedforaging behavior (e.g., Hoffmeyer 1969; Blok-huis 1989; Huber-Eicher & Wechsler 1997;Wechsler & Huber-Eicher 1997).
Given the putative role of foraging behavior inthe development of feather pecking by chickens, itis possible that a similar relationship exists be-tween foraging behavior and feather picking inparrots. This may mean that captive parrots arehighly motivated to perform the behaviors associ-ated with food procurement in the wild and thatthis motivation may persist despite the fact thatcaptive feeding methods meet their nutritionalneeds. There is some evidence that captive parrotsprefer to access food utilizing their foraging skillseven when “free” food is available (Coulton et al.1997). Thus, the act of foraging may be a behav-ioral need for parrots and the absence of foragingopportunity may result in frustration and redirec-tion of foraging-like activities toward the feathers.
In our study of Orange-winged AmazonParrots (Meehan et al. 2002), one group of youngbirds (between four and five months old) wasgiven the opportunity to utilize their foragingskills to access food while the other was not. Bothgroups of parrots were given pellets, fruits, nuts,seeds, and vegetables daily; the difference was inthe manner of presentation. The “control” groupreceived these foods cut up and served in a dish,
27 / Captive Parrot Welfare 307

as is common practice with captive parrots, whilethe “enriched” group received these foods in spe-cially designed feeders (see Figure 27.1). Thesefeeders significantly increased the diversity offeeding behaviors the parrots performed as wellas increased the amount of time they spent infeeding activities. Parrots in the enriched groupalso received enrichments that increased thephysical complexity of the cage. Plumage condi-tion served as an indirect measurement of feather-picking behavior.
The effect on feather-picking development wasdramatic. Over the course of the one-year study,six of eight parrots in the control group developedfeather-picking behavior (as evidenced by signif-icant decreases in feather score), while none ofthe birds in the enriched group showed any signsof developing the disorder (as evidenced by sta-bility or improvement in feather score). Thus, thisstudy demonstrates that presenting food in a man-ner that requires parrots to utilize their foragingskills and increasing the physical complexity of
the cage are effective in the prevention of feather-picking behavior. This is excellent news to own-ers of young parrots as it provides a method ofpreventing the development of this problematicbehavior. However, a second component of thestudy was required to determine if this method iseffective in reversing established feather picking.
In the second phase of the study, the parrotsthat had once been in the control condition weregiven access to the feeding devices and physicalenrichments used in the original study. After fourmonths, all six parrots that had feather pickedshowed significant improvements in feather con-dition, indicating that feather-picking behaviorhad significantly decreased. These results suggestthat making appropriate changes to the environ-ment can effectively treat parrots with establishedfeather-picking behavior. The parrots in this studyhad been performing the behavior for about oneyear before it was reversed, however, and it re-mains to be seen if the same course of action willbe effective with parrots that have performed thisbehavior for several years.
Taken as a whole, these results support the hy-pothesis that a captive environment that does notsupport the appropriate expression of foragingbehaviors can contribute to the development ofpsychogenic feather-picking behavior. By provid-ing foraging devices and physical enrichments itis possible to both prevent and reverse this disor-der. Since we introduced foraging and physicalenrichments together, rather than separately, it isdifficult to tell the relative importance of eachtype of enrichment from our study. It is possiblethat the combination of physical and foraging en-richments is necessary for the preventative andreversal effects on feather picking we observed.However, since the foraging enrichments wereused for significantly greater periods of time thanthe physical enrichments, our data certainlydemonstrate a link between foraging opportunityand the development and performance of featherpicking in parrots.
Fearfulness and Aggression
Hyperaggressiveness and excessive fear reactionsare often identified as indicators of poor welfare.Excessive aggression can result in injury and caninhibit social interactions within groups of par-rots or between captive parrots and their caretak-ers. In addition, fear is generally considered an
308 Manual of Parrot Behavior
Figure 27.1. Examples of the enrichmentsutilized in the U.C. Davis experiments. See alsocolor section.

undesirable emotional state (Jones 1997) and ex-aggerated fear reactions such as escape attemptsand panic can result in wasted energy or injury(Jones & Waddington 1992). Parrots that are ex-cessively fearful or aggressive are much more dif-ficult to care for and as a result are more likely tobe abused, neglected, or relinquished by ownerswho become frustrated by this behavior.
For parrots, novelty is often a potent stimulusfor excessive fearful or aggressive reactions.Unpredictable yet benign environmental changessuch as new people, new surroundings, or intro-duction of new toys or food items can inducescreaming, biting, cowering, shaking, and fleeing.It has been shown in other birds and mammalsthat the degree of fearful and aggressive reactionsis associated with the quality of the captive envi-ronment, in that animals reared in barren environ-ments with little exposure to novelty and little op-portunity for environmental interaction oftendisplay exaggerated responses compared to ani-mals reared in more complex environments. Forexample, rats reared in environments lackingcomplexity show an increased fear response tonovel situations and increased aggression towardnovel individuals (Fernandez-Teruel et al. 1997;Escorihuela et al. 1994; Renner 1987), whilechickens reared in this type of environment showincreased fear responses to novel places and novelobjects (Jones 1982; Jones & Waddington 1992).
Environmental enrichment that increases ex-posure to novelty during development has beensuccessfully employed to modulate fear responsesin both birds and mammals as evidenced by in-creased activity in novel environments(Fernandez-Teruel et al. 1997; Escorihuela et al.1994; Renner 1987) and decreased fear responsesto novel objects (Jones 1982; Jones & Waddington1992). There is also evidence that enrichment ofthe physical environment reduces fearfulness ofhumans (e.g., Jones & Waddington 1992; Nicol1992; Pearce et al. 1989).
Fear reactions to novel objects may be exagger-ated in parrots whose cage environment is barrenor unchanging, and environmental enrichmentmay reduce fear reactions to novelty by exposingparrots to an environment where novelty is expe-rienced on a consistent basis. We compared theresponses of parrots from enriched and barrenconditions to novel objects (Meehan & Mench2002). In one study, the enrichments used pro-
vided foraging opportunity and increased thephysical complexity of the cage. Parrots in the en-riched condition received a novel combination ofone foraging and one physical enrichment eachweek for 16 weeks. In a second study, the re-sponses to novel objects of parrots reared in pairswere compared with those of parrots that weresingly housed (Meehan et al. 2003). In this study,all parrots received the physical and foraging en-richment protocol previously described, leavingsocial enrichment as the variable of interest. Totest for the effect on fearfulness of novelty in theirenvironment, a novel object was placed in the par-rot’s home cage in a position that required the birdto approach the object in order to interact with it.
In the first study, parrots from the enrichedcondition had significantly shorter latencies to in-teract with novel objects than did parrots from thebarren cages. Parrots from the barren cages usu-ally responded initially to the novel object by re-treating from it. They would then engage in a se-ries of approach/retreat sequences, eventuallycontacting the object. The initial reaction of theparrots in the enriched condition was also to re-treat from the object. However, shortly after theyhad retreated and visually inspected the objectthey would begin to interact with the object with-out the approach/retreat sequences characteristicof the control group. Thus, the latency to interactwith the novel object was significantly shorter inthe enriched group than it was in the controlgroup. The data suggest that the accelerated ap-proach toward novel objects in the enriched groupreflects decreased fearfulness of the object ratherthan an increased tendency to engage in ex-ploratory behavior, since the parrots in the en-riched condition also had shorter overall durationof interaction and shorter bout lengths of interac-tion with the objects.
Pair-housing also had an effect on fear re-sponses to novel objects. The single parrots hadsignificantly longer latencies to interact with thenovel objects than the paired parrots (when thepaired parrots were tested together as a pair).Thus, having a social partner present appears toameliorate the fear-evoking effect of a novel stim-ulus. The fear-diminishing effect of a social part-ner has been observed in chickens (Jones &Merry 1988), monkeys (Coe et al. 1982), and rats(Taylor 1981). In each of these cases, as in ourstudy, a decreased fear response to novelty is ob-
27 / Captive Parrot Welfare 309

served when the animals are tested with a socialpartner as compared to when they are testedalone. Thus, in addition to an inanimate enrich-ment protocol, pair-housing is recommended asan effective method for creating a dynamic cageenvironment and reducing fearfulness of environ-mental novelty in captive parrots.
Interactions with Human Handlers
In most situations where companion parrots arekept, positive interactions with humans are val-ued. Parrots that are excessively fearful of or ag-gressive toward humans are generally not consid-ered desirable as pets, in aviculture, as researchsubjects, or as part of zoo exhibits. However, bit-ing and aggression are some of the most com-monly reported complaints by parrot owners.Aggressive or fearful responses toward humansmay be due to a variety of experiential or devel-opmental factors, including trauma and hormonalchanges. In addition, how parrots are housed maycontribute to this behavior. While the popular par-rot literature suggests that housing parrots to-gether in pairs, rather than singly, may negativelyimpact their interactions with human handlers(e.g., Blanchard 1999), recent experimental evi-dence suggests that pair-housing of young parrotsin same-sex pairs may actually improve, ratherthan hinder, interactions with human handlers(Meehan et al. 2003).
Responses to familiar and unfamiliar humanhandlers were tested in parrots that were housedeither individually or in same-sex pairs (Cramton1998; Aengus & Millam 1999). Testing involvedextending a finger toward the parrot, touching theparrot’s back, touching the parrot’s head, offeringfood, and observing flight distance when the par-rot was placed next to the handler. Two familiarhandlers as well as two unfamiliar handlers testedeach parrot. The familiar handlers were peoplewho had been working regularly in handling ses-sions with the parrots, while the unfamiliar han-dlers were people who were trained in parrot han-dling but who had never interacted directly withthe test parrot.
Pair-housing did not cause parrots to behavemore aggressively or fearfully to human handlers.In fact, parrots from both groups showed an im-provement in responses to familiar handlers overthe course of the year, and overall there was nodifference in responses between the two groups.
This suggests paired parrots are equally suitableas pets in this regard as singly housed parrots.
When tested with unfamiliar handlers, therewas a difference in responses between the twogroups. In this case, the singly housed parrots re-sponded more aggressively and fearfully than didthe paired parrots. This difference in responsemight be explained by the fact that the paired par-rots had more opportunities to explore changes intheir environment and thus were more likely to re-spond inquisitively rather than fearfully to thenovel human stimulus. In addition, social interac-tion and play facilitate behavioral flexibility andenhance the ability to transfer learning tasks (e.g.,Morgan 1973; Morgan et al. 1975; Einon et al,1978). Thus, because paired parrots had the op-portunity to interact with a pairmate and spentmore time in play activities than the single par-rots, they may have been better able to transfer thelearning experience of the regular handling ses-sions to the experience with the novel human.This ability is relevant to many captive contextsbecause parrots are likely to encounter novel hu-mans, such as visitors or veterinarians, regularly.
Stereotypic Behavior
Stereotypies are defined as behavior patterns thatare repetitive, invariant, and have no obvious goalor function (Mason 1991). While the precise eti-ology of these behaviors is not yet understood, ithas been suggested that stereotypies develop inanimals housed in environments that are subopti-mal in one or more dimensions (Mason 1991).For example, in many species, stereotypic behav-iors are significantly more evident in environ-ments that do not provide sufficient sensory stim-ulation (Mason 1991), opportunity to interactwith objects or conspecifics (Carlstead 1998), orthat leave the animal with little control over itssurroundings (Markowitz & Aday 1998).
Stereotypy development may be related to thefrustration of specific motivational systems. Forexample, a number of experiments demonstrate arelationship between stereotypies and restrictionof feeding or foraging behaviors, locomotion, orsocial contact. Feed restriction is closely associ-ated with stereotypy performance in pigs (Ter-louw et al. 1991), chickens (Savory et al. 1992),and sheep (Mardsen & Wood-Gush 1986). How-ever, feeding-related stereotypies can occur evenwhen food intake is not restricted. In these cases,
310 Manual of Parrot Behavior

it is thought that motivation to perform foragingbehaviors underlies stereotypy performance. Thisidea is supported by evidence that, in both mam-mals and birds, stereotypy performance is re-duced when opportunities to work in order to lo-cate, access and consume food items are provided(e.g., Keiper 1969; Kastelstein & Wiepkema1989; Line et al. 1989; Carlstead et al. 1991).
Some stereotypies are thought to develop fromfrustrated locomotor behavior. Hediger (1955)suggested that the stereotyped pacing common tozoo animals might develop from normal pa-trolling behaviors that are thwarted due to limitedspace. Increasing the complexity of the cage envi-ronment and providing opportunity for additionalperching and swinging behaviors reduced stereo-typic route tracing in Canaries (Keiper 1969),suggesting that this behavior might be related tofrustrated locomotor behaviors.
Finally, motivation for social contact may un-derlie some forms of stereotypy. For example,frustrated motivation for maternal contact isthought to be associated with some primate stereo-typies such as rocking and self-clasping (Marriner& Drickamer 1994), and providing horses with ei-ther a mirror or social contact reduces stereotypicweaving behaviors (Nicol 1999).
Thus, there is ample evidence that the frustra-tion of highly motivated behaviors is involved inthe development and performance of some formsof stereotypy. However, developmental evidencesuggests that stereotypy is not simply a behavioralresponse to an inappropriate environment butrather the product of an abnormal developmentalprocess resulting in both physiological and behav-ioral impairment. For example, stereotypieschange over time in both form (Meyer-Holzapfel1968; Cronin et al. 1984; Mason 1993) and fre-quency (e.g., Mason 1993; Würbel et al. 1998;Powell et al. 2000). In addition, stereotypies maybecome more difficult to reverse over time(Kiley-Worthington 1977; Cronin et al. 1984;Cooper et al. 1996) and eventually may becomeestablished in the behavioral repertoire of animalssuch that they remain unchanged even when theenvironment is modified. Thus, the fact that thenature and form of stereotypy change with time,even when the captive environment remains con-stant, indicates that stereotypy is the result of en-vironmentally induced qualitative changes in theanimal (Garner 1999; Würbel 2001).
Similar behaviors are also extremely commonin a number of human mental disorders. Stereo-typies are performed by approximately 70% ofchronic schizophrenic patients (Owens et al.1982) and are core symptoms of both Tourette’ssyndrome and autism (American Psychiatric As-sociation 1994). Recent evidence suggests that,like in human patients, stereotypy in caged ani-mals reflects a general disinhibition of behav-ioral control mechanisms (Garner & Mason2002; Garner et. al. 2003). Thus, it is possiblethat stereotypies seen in captive animals are theresult of environmentally induced neurologicaldeficits similar to those seen in human psychi-atric disorders.
The frequency of stereotypy in the captive par-rot population at large has not been estimated.Many parrot owners are not aware of this class ofabnormal behavior since stereotypies are oftendifficult to recognize without prior experienceand training. In addition, parrots may only per-form stereotypies when they are alone (which iswhy videotaping is necessary for stereotypy re-search), and thus owners may never witness theirparrots performing stereotypies. While the stereo-typies performed by parrots take many forms,they can, for the most part, be classified into threemain categories: oral stereotypies, locomotorstereotypies, and object-directed stereotypies.Oral stereotypies include such behaviors as spotpecking, sham chewing, bar biting, or tonguerolling. Locomotor stereotypies include routetracing and pacing. Object-directed stereotypiesinvolve repetitive, invariant manipulation of ob-jects such as toys, feeders, and waterers. The de-gree to which these behaviors are invariant intheir repetitions is illustrated in the following se-ries of pictures captured from a videotape of aparrot performing a route trace (see Figure 27.2).
There have been few studies of stereotypy inparrots, but those that have been completed impli-cate both lack of foraging opportunity and limitedphysical complexity in the cage environment inthe development of these behaviors. When parrotsare housed in cages that lack foraging opportunityand physical complexity, stereotypy reliably de-velops. For example, 96% of parrots in a colonyhoused in these conditions performed stereotypy,and individuals spent between 5% and 85% oftheir active time performing these behaviors(Meehan 2002).
27 / Captive Parrot Welfare 311

Enriching the environment with foraging de-vices and increasing the physical complexity ofcages significantly decreased the development ofstereotypy in young Orange-winged AmazonParrots (Meehan et al., submitted). Parrots in thecontrol condition spent significantly more of theiractive time performing stereotypies than did par-rots in the enriched condition. However, the de-gree of environmental modification used in thisstudy was not sufficient to completely prevent thedevelopment of stereotypic behavior. At the endof 48 weeks the parrots in the enriched conditionperformed stereotypies an average of 4% of theiractive time. This is a common outcome of studies
examining the role of specific environmental fac-tors in stereotypy development (e.g., Odberg1987; Würbel et al. 1998; Powell et al. 2000). Forexample, increasing the physical complexity ofcages with twigs prevents most, but not all,stereotypy development in young voles (Odberg1987). Similarly, the combination of feeding sun-flower seeds and increasing physical complexityhas a significant effect on the amount of time deermice spend performing stereotypy but does noteliminate the development of these behaviors al-together (Powell et al. 2000). These results indi-cate that additional research is necessary to deter-mine the specific environmental qualities needed
312 Manual of Parrot Behavior
Figure 27.2. The invariance of stereotypy. These frames are taken from a sequence of video inwhich the corner flipping stereotypy was repeated six times. Note the identical foot positions andbody posture. (a) Frame 40, (b) Frame 105, (c) Frame 162, (d) Frame 219, (e) Frame 267, (f) Frame319. The frame rate is 15 frames per second, giving a mean ± SD interval of 3.72 ± 0.43 secondsbetween frames. Reproduced with permission from Behavioral Brain Research.

to completely eliminate stereotypy from the be-havioral repertoire of parrots.
Parrots in the control condition developed bothlocomotor and oral stereotypies, while those inthe enriched condition developed almost exclu-sively locomotor stereotypies. This suggests thatspecific forms of stereotypy may be associatedwith the absence of specific environmental ele-ments (Mason & Mendl 1997). In this case, if oralstereotypies were associated with frustration offoraging behaviors, then the foraging enrichmentsmay have successfully eliminated this frustration.If the development of locomotor stereotypies wasassociated with limited space, prevention offlight, or lack of social contact, then this wouldexplain why these behaviors were not preventedby the physical enrichments we provided. There isevidence in Canaries that the development of oralstereotypies is related to lack of opportunity toperform foraging behaviors, while the develop-ment of locomotor stereotypies is related to a lackof space and physical complexity (Keiper 1969).Thus, additional experiments assessing the effec-tiveness of increased flight space and social hous-ing are needed to determine appropriate environ-mental remedies for locomotor stereotypy.
Screaming
The vocal behavior of captive animals is increas-ingly utilized as an index of welfare (e.g., Wearyet al. 1998; Grandin 1998, 2001). For example,Boinski and colleagues (1999) found that the rateof alarm vocalizations was positively correlatedwith the amount of time male capuchins per-formed stereotypic and redirected behaviors andnegatively correlated with the amount of normalbehavior performed. In addition, a positive rela-tionship between fecal cortisol levels and vocal-ization rate was found, indicating that monkeysthat vocalized more may have been morestressed. Finally, modification of the cage envi-ronment in the form of toys and foraging materi-als resulted in a significant decrease in monkeys’vocalization rate.
Incessant screaming is the second (after messi-ness) most common complaint of parrot owners(Kidd & Kidd 1998) and is one of the precipitat-ing factors for parrot neglect and relinquishment(Reynolds 1998). Much communication betweenparrots is vocal, and some authors have suggestedthat loud, high-pitched squawks are used as alarm
calls when individuals are in danger or distress(Alderton 1992) or as a contact call betweengroup members (Sparks & Soper 1990). However,when these vocalizations become prolonged andrepetitive, they are considered abnormal and maybe indicative of boredom (Davis 1991) or, similarto capuchin monkeys, stress (Boinski et al. 1999).
We found that housing young amazons in same-sex pairs effectively reduces the amount of timespent in prolonged vocalizations (loud vocaliza-tion bouts lasting more than two seconds) whencompared with singly housed parrots of the sameage. Over the course of one year, paired parrotsspent an average of 4.1% of their active timescreaming, while singly housed parrots spent anaverage of 9.5% of their active time screaming(Meehan et al. 2003). It is important to note thatall parrots in this study had visual and vocal con-tact with other parrots, but only the paired parrotscould interact physically. More research in thisarea is necessary to provide a basis for interpret-ing the significance of differences in vocal pat-terns with respect to welfare. However, the factthat social housing impacts the degree to whichyoung parrots perform prolonged vocalizations in-dicates that lack of physical interaction with socialpartners may be a factor in the development of thisbehavior. This finding is of great practical impor-tance because it suggests that pair-housing is aneffective way to ameliorate the performance ofthis common and frustrating behavioral problem.
PRACTICAL CONSIDERATIONS FORIMPROVING PARROT WELFARETHROUGH ENVIRONMENTALENRICHMENT
This chapter has highlighted some of the impactsthat environmental enrichment can have on thebehavioral development of companion parrots.Effective environmental enrichment can improvethe welfare of parrots by increasing the perform-ance of behaviors such as foraging, locomotion,and social interaction, while concomitantly re-ducing or eliminating the performance of abnor-mal behaviors such as feather picking, screaming,and stereotypy. While enrichment strategies musttake into consideration the behavior and ecologyof the species in question as well as the age andhistory of the individual parrot, there are severalimportant principles of environmental enrichmentthat will apply across all contexts.
27 / Captive Parrot Welfare 313

Enrichment must provide a dynamic captiveenvironment. This means that the parrots must begiven the opportunity to act, react, and interactwithin their home environs. Providing novel en-richments on a consistent basis can help to ac-complish this goal. Even the most wonderfullyenriched cage loses its effectiveness when par-rots are exposed to it for long periods of time.The exact timing of enrichment rotation will de-pend on the context and the particular parrot.Some parrots will require a day or so of habitua-tion to novel enrichments and, in this case,weekly rotation would be advised. On the otherhand, some parrots are extremely inquisitive andwould be excellent candidates for daily introduc-tion of novel items. Social companions also canprovide a dynamic quality to the environment.Ideally, all parrots should have the opportunity tointeract physically with other parrots in compati-ble social groups. However, more research isneeded to determine the best approach to provid-ing social enrichment to parrots throughout theirlifespan.
The life-history stage of parrots should betaken into consideration when planning to intro-duce social enrichment. If parrots are introducedto social groups prior to reaching sexual maturity,it is possible that the compatibility of thesegroups may change as the parrots mature. If theonset of sexual maturity results in reduced com-patibility or aggression, then social groupingsmay need to change as the parrots age. Older par-rots may be more suited to mixed-sex groups. Infact, there is some evidence that forming larger,mixed-sex groups of parrots who are sexually ma-ture can be accomplished successfully. An infor-mal study was conducted at U.C. Davis with fivepairs of parrots ranging in age from five to eightyears old. These parrots had been pair-housed forthree years in male/female pairs but had not suc-cessfully reproduced during any of the threebreeding seasons in which they were paired. Alarge flight cage was constructed and attached tothe home cages of the pairs, which allowed re-searchers to control access to the flight cage. Theflight was furnished with the same foraging andphysical enrichments described earlier and accesswas provided to all ten parrots simultaneously.The parrots were monitored regularly to assesslevels of aggression and decrease the risk of in-jury. After several weeks of interaction, all par-
rots, with the exception of one male (who was re-moved from the group) co-existed peacefully inthe large cage. The parrots utilized the enrich-ments extensively, despite the fact that they hadno prior experience with these types of devices.At this point, nest boxes were introduced and theparrots were allowed to “self-select” pairmates.There was some aggression involved in thisprocess, but the parrots were closely monitoredand the aggression was not severe enough tocause injury or warrant separation. Interestingly,two of the original pairs were maintained, onenew pair was formed, and one threesome (onemale and two females) was formed. Of these,three pairs laid eggs and two successfully rearedchicks. Of the pairs that raised chicks, one was anoriginal pair and one was a newly formed pair. Itshould be noted that social enrichment is an im-portant component of parrot husbandry whetheror not the parrots in question are to be used forbreeding purposes. Young parrots are very moti-vated to interact with conspecifics and will crossbarriers to access social partners, while they willnot cross these same barriers to access food ortoys (C. Meehan, unpublished data).
A successful environmental enrichment pro-gram can significantly improve environmentalquality and positively affect the welfare of captiveparrots. In order to be successful, an enrichmentprotocol must be well-conceived and must pro-vide parrots the opportunity to utilize theirconsiderable behavioral skills. A combination offoraging, physical, and social enrichment is rec-ommended for all parrots held in captivity. En-vironmental enrichment of this sort results indecreased screaming, feather picking, and stereo-typy; increased physical activity and play behav-ior; reduced fear responses to novelty; and im-proved reproductive success without impartingsignificant risk of illness and injury or jeopardiz-ing the ability of parrots to relate positively withhumans. Thus, by providing an environment richwith opportunities to perform a wide variety ofbehaviors, owners can make marked improve-ments in the welfare of their parrots. Creating adynamic, enriched environment in captivity re-quires significantly more time and creativity thanhousing parrots in barren cages. However, raisinga parrot that is physically healthy, behaviorallyactive, and free from distressing abnormal behav-iors is well worth the effort.
314 Manual of Parrot Behavior

REFERENCES
Aengus, W.L., and J.R. Millam. 1999. Taming parent-reared orange-winged amazon parrots by neonatalhandling. Zoo Biology 18:177–187.
Alderton, D. 1992. Parrots. London: Whittet Books.American Psychiatric Association 1994. Diagnostic
and statistical manual of mental disorders, 4th ed.Washington, DC: American Psychiatric Association.
American Veterinary Medical Association. 2002. U.S.pet ownership and demographics sourcebook.Schaumburg, IL: Center for Information Manage-ment, pp. 1–2.
Appleby, M.C., B.O.Hughes, and J.A. Mench. In press.Poultry behaviour and welfare. Wallingford, Oxon,U.K.: CAB International.
Barnett, J.L., G.M. Cronin, C.G. Winfield, and A.M.Dewar 1984. The welfare of adult pigs: The effectsof five housing treatments on behaviour, plasma cor-ticosteroids and injuries. Applied Animal BehaviorScience 12:209–232.
Bauck, L. 1999. Nutritional problems in pet birds.Semin Avian Ex Pet Med 4:3–8.
Bayne, K.B., B.V. Beaver, J.A. Mench, and D.B.Morton. 2002. “Laboratory animal behavior.” InLaboratory animal medicine, 2nd ed., pp.1240–1264. New York: Academic Press.
Bayne, K., H. Mainzer, S. Dexter, G. Campbell, F.Yamada, and S. Suomi. 1991. The reduction of ab-normal behaviors in individually housed rhesusmonkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology23:23–36.
Birchall, A. 1990. Who’s a clever parrot, then? NewScientist, 24 February, pp. 38–43.
Blanchard, S. 1999. “The pet bird report.” www.petbirdreport.com/myths.html.
Blokhuis, H.J. 1986. Feather pecking in poultry: Its re-lation with ground pecking. Applied AnimalBehaviour Science 16:63–67.
Blokhuis, H.J. 1989. The development and causation offeather pecking in the domestic fowl. Ph.D. thesis,Agricultural University, Wageningen, the Netherlands.
Boice, R. 1981. Captivity and feralisation. Psycho-logical Bulletin 89:407–421.
Boinski, S., S.P. Swing, T.S. Gross, and J.K. Davis.1999. Environmental enrichment of brown capuchin(Cebus apella): Behavioral and plasma and fecalcortisol measures of effectiveness. American Jour-nal of Primatology 48:49–68.
Carlstead, K. 1998. “Determining the causes of stereo-typic behaviors in zoo carnivores.” In Secondnature: Environmental enrichment for captive ani-mals, ed. D.J. Shepherdson, J.D. Mellen, and M.Hutchins, pp. 172–183. Washington, DC: Smithson-ian Institution.
Carlstead, K., J. Seidensticker, and R. Baldwin. 1991.Environmental enrichment for zoo bears. ZooBiology 10:3–16.
Carlstead, K., and D. Shepherdson. 2000. “Alleviatingstress in zoo animals with environmental enrich-ment.” In The biology of animal stress: Basic princi-ples and implications for welfare, ed. G.P. Mobergand J.A. Mench, pp. 337–354. Wallingford, Oxon,U.K.: CABI Publishing.
Chu, L-r., J.P. Garner, and J.A. Mench. In press. A be-havioral comparison of New Zealand white rabbit(Oryctolagus cuniculus) housed individually or inpairs in standard cages. Applied Animal BehaviourScience.
Coe, C.L., D. Franklin, E.R. Smith, and S. Levine.1982. Hormonal responses accompanying fear andagitation in the squirrel monkey. Physiology andBehavior 29:291–294.
Cooper, J.J., F. Odberg, and C. Nicol. 1996. Limitationson the effectiveness of environmental improvementin reducing stereotypic behaviour in bank voles(Clethrionomys glareolus). Applied Animal Be-haviour Science 48:237–248.
Coulton, L.E., N.K. Warran, and R.J. Young. 1997.Effects of foraging enrichment on the behaviour ofparrots. Animal Welfare 6:357–363.
Cramton, B. 1998. Master’s thesis. Department ofAvian Science. University of California, Davis.
Cronin, G.M., P.R. Wiepkema, and G.J. Hofstede.1984. “The development of stereotypies in tetheredsows.” In Proceedings of the International Congresson Applied Ethology in Farm Animals, ed. J.Unshelm, G. van Putten, & K. Keeb, pp. 97–100.Darmstadt, Germany: KTBL.
Davis, C.S. 1991. Parrot psychology and behaviorproblems. Veterinary Clinics of North America:Small Animal Practice 21:1281–1288.
Dawkins, M.S. 1988. Behavioral deprivation: A centralproblem in animal welfare. Applied AnimalBehaviour Science 20:209–225.
Duncan, I.J.H., and D. Fraser. 1977. “Understandinganimal welfare.” In Animal welfare, ed. M.C.Appleby and B.O. Hughes, pp. 19–32. Wallingford,Oxon, U.K.: CAB International.
Einon, D.F., M.J. Morgan, and C. Kibbler. 1978. Briefperiods of socialization and later behavior in the rat.Developmental Psychobiology 11:213–225.
Escorihuela, R.M., A. Tobena, and A. Fernandez-Teruel. 1994. Environmental enrichment reversesthe detrimental action of early inconsistent stimula-tion and increases the beneficial effects of postnatalhandling on shuttlebox learning in adult rats.Behavioural Brain Research 61:169–173.
Fernandez-Teruel, A., R.M. Escorihuela, B. Castellano,B. Gonzalez, and A. Tobena. 1997. Neonatal han-dling and environmental enrichment effects on emo-
27 / Captive Parrot Welfare 315

tionality, novelty/reward seeking and age-relatedcognitive and hippocampal impairments: Focus onthe Roman rat lines. Behavior Genetics 27:513–526.
Fraser, D., D.M. Weary, E.A. Pajor, and B.N. Milligan.1997. A scientific conception of animal welfare thatreflects ethical concerns. Animal Welfare 6:187–205.
Garner, J.P. 1999. The aetiology of stereotypy in cagedanimals. Ph.D. thesis, University of Oxford, U.K.
Garner, J.P., and G.J. Mason. 2002. “Cage stereotypy,psychiatric task performance, behavioural flexibil-ity, and apparent cognitive ability in songbirds: Theimpact of environmental enrichment.” In The 2ndInternational Symposium on Comparative CognitiveScience, ed. T. Matsuzawa, p. 72. Kyoto University,Kyoto and Inuyama, Japan.
Garner, J.P., C.L. Meehan, and J.A. Mench. 2003.Stereotypies in caged parrots, schizophrenia andautism: Evidence for a common mechanism.Behavioural Brain Research 145 (1–2):125–134.
Gilardi, J.D., and C.A. Munn. 1998. Patterns of activity,flocking and habitat use in parrots of the PeruvianAmazon. Condor 100 (4):641.
Grandin, T. 1998. The feasibility of using vocalizationscoring as an indicator of poor welfare during cattleslaughter. Applied Animal Behaviour Science56:121–128.
Grandin, T. 2001. Cattle vocalizations are associatedwith handling and equipment problems at beefslaughter plants. Applied Animal Behaviour Science71:191–201.
Grindlinger, H. 1991. Compulsive feather picking inbirds. Archives of General Psychiatry 48:857.
Hediger, H. 1955. The psychology and behaviour of an-imals in zoos and circuses. London: Butterworths.
Hoffmeyer, I. 1969. Feather pecking in pheasants—anethological approach to the problem. Danish ReviewGame Biology 6:1–36.
Huber-Eicher, B., and B. Wechsler. 1997. Feather peck-ing in domestic chicks: Its relation to dustbathingand foraging. Animal Behaviour 54:757–768.
Huber-Eicher, B., and B. Wechsler. 1998. The effect ofquality and availability of foraging materials onfeather pecking in laying hen chicks. AnimalBehaviour 55:861–873.
Hughes, H.C., S. Campbell, and C. Kenney. 1989. Theeffects of cage size and pair housing on exercise ofbeagle dogs. Laboratory Animal Science 39(4):302–305.
Jones, R.B. 1982. Effects of early environmental en-richment upon open-field behavior and timidity inthe domestic chick. Developmental Psychobiology15:105–111.
Jones, R.B. 1997. “Fear and distress.” In Animal wel-fare, ed. M.C. Appleby and B.O. Hughes, pp. 75–87.New York: CAB International.
Jones, R.B., and B.J. Merry. 1988. Individual or pairedexposure of domestic chicks to an open field: Somebehavioral and adrenocortical consequences.Behavioral Processes 16:75–86.
Jones, R.B., and D. Waddington. 1992. Modificationsof fear in domestic chicks, Gallus gallus domesticus,via regular handling and early environmental enrich-ment. Animal Behaviour 43:1021–1034.
Kastelstein, R.A., and P.R. Wiepkema. 1989. A diggingtrough as occupational therapy for Pacific walruses(Odobens rosmarus divergens) in human care.Aquatic Mammals 15:9–17.
Keiper, R.R. 1969. Causal factors of stereotypies incaged birds. Animal Behaviour 17:114–119.
Kidd, A.H, and R.M. Kidd. 1998. Problems and bene-fits of bird ownership. Psychological Reports83:131–138.
Kiley-Worthington, M. 1977. Behavioural problems offarm animals. London: Oriel Press.
King, C.E. 1993. Environmental enrichment: Is it forthe birds? Zoo Biology 12:509–512.
Klein, T., E. Zeltner, and B. Huber-Eicher. 2000. Aregenetic differences in foraging behaviour of layinghen chicks paralleled by hybrid-specific differencesin feather pecking? Applied Animal BehaviourScience 70:143–155.
Lantermann, W. 1993. Social deprivation in amazonparrots. Kleintierpraxis 38:511–520.
Line, S.W., H. Markowitz, K.N. Morgan, and S. Strong.1989. Evaluation of attempts to enrich the environ-ment of singly caged non-human primates. InAnimal care and use in behavioral research:Regulations, issues and applications, ed. J. Driscoll,pp. 103–117. Beltsville, MD: Animal WelfareInformation Center, National Agricultural Library.
Line, S.W, K.N. Morgan, and H. Markowitz. 1991.Simple toys do not alter the behavior of aged rhesusmonkeys. Zoo Biology 10:473–484.
Magrath, R.D., and A. Lill. 1985. Age related differ-ences in behavior and ecology of crimson rosellasduring the non-breeding season. Austr Wildl Res12:299–306.
Mardsen, D., and D.G.M. Wood-Gush. 1986. A note onthe behaviour of individually penned sheep regard-ing their use for research purposes. AnimalProduction 42:157–159.
Markowitz, H., and C. Aday. 1998. Power for captiveanimals: Contingencies and nature. In Second na-ture: Environmental enrichment for captive ani-mals, ed. D.J. Shepherdson, J.D. Mellen, and M.Hutchins, pp. 47–58. Washington, DC: SmithsonianInstitution.
Marriner, L.M., and L.C. Drickamer. 1994. Factors in-fluencing stereotyped behavior of primates in a zoo.Zoo Biology 13:267–275.
316 Manual of Parrot Behavior

Mason, G.J. 1991 Stereotypies: A critical review.Animal Behaviour 41:1015–1037.
Mason, G.J. 1993. Age and context affect the stereotyp-ies of caged mink. Behaviour 127 (3–4):191–229.
Mason, G.J., and M. Mendl. 1997. Do the stereotypiesof pigs, chickens and mink reflect adaptive speciesdifferences in the control of foraging? Appl AnimBehav Sci 53:45–58.
McKeegan, D.E.F., and C.J. Savory. 1999. Feather eat-ing in layer pullets and its possible role in the aetiol-ogy of feather pecking damage. Applied AnimalBehaviour Science 65:73–85.
Meehan, C.L. 2002. Environmental enrichment and be-havioral development of orange-winged amazonparrots (Amazona amazonica): Applications to ani-mal welfare. PhD diss., University of California,Davis.
Meehan, C.L., J.P. Garner, and J.A. Mench. 2003.Isosexual pair housing improves the welfare ofyoung amazon parrots. Applied Animal BehaviourScience 81 (1):73–88.
Meehan, C.L., J.P. Garner, and J.A. Mench. Submitted.Environmental enrichment and cage stereotypy inorange-winged amazon parrots (Amazona amazon-ica): Insights into developmental processes.Developmental Psychobiology.
Meehan, C.L., and J.A. Mench. 2002. Environmentalenrichment effects fear and exploratory responses inyoung amazon parrots. Applied Animal BehaviourScience 79 (1):77–90.
Meehan, C.L., J.R. Millam, and J.A. Mench. 2003.Foraging opportunity and increased physical com-plexity both prevent and reduce psychogenic featherpicking by young amazon parrots. Applied AnimalBehaviour Science 80 (1):71–85.
Mench, J.A., and L.J. Keeling. 2001. The social behav-iour of domestic birds. In Social behaviour in farmanimals, ed. L.J. Keeling and H.W. Gonyou, pp.177–211. Wallingford, Oxon, U.K.: CAB Inter-national.
Mendl, M., and R.C. Newberry. 1997. “Social condi-tions.” In Animal welfare, ed. M.C. Appleby andB.O.Hughes, pp. 191–203. New York: CAB Inter-national.
Mertens, P.A. 1997. “Pharmacological treatment offeather picking in pet birds.” In Proceedings of theFirst International Conference on VeterinaryBehavioral Medicine, ed. D.S. Mills and S.E. Heath,pp. 209–213. Potters Bar, U.K.: UFAW.
Meyer–Holzapfel, M. 1968. “Abnormal behaviors inzoo animals.” In Abnormal behavior in animals, ed.M.W. Fox, pp. 476–503. London: Saunders.
Morgan, M.J. 1973. Effects of pre-weaning environ-ment on learning in the rat. Animal Behaviour21:429–442.
Morgan, M.J., D.F. Einon, and D. Nicholas. 1975.Effects of isolation rearing on behavioral inhibitionin the rat. Quarterly Journal of ExperimentalPsychology 27:615–634.
Newberry, R.C. 1995. Environmental enrichment:Increasing the biological relevance of captive envi-ronments. Applied Animal Behaviour Science44:229–243.
Nicol, C.J. 1992. Effects of environmental enrichmentand gentle handling on behaviour and fear responsesof transported broilers. Applied Animal BehaviourScience 33:367–380.
Nicol, C.J. 1999. Understanding equine stereotypies.Equine Veterinary Journal, Supplement 28:20–25.
Nicol, C.J., A.C. Lindberg, A.J. Phillips, S.J. Pope, L.J.Wilkins, and L.E. Green. 2001. Influence of priorexposure to wood shavings on feather pecking, dust-bathing and foraging in adult laying hens. AppliedAnimal Behaviour Science 73:141–155.
Novak, M.A., and S.J. Suomi. 1988. Psychologicalwell-being of primates in captivity. AmericanPsychologist 10:765–773.
Odberg, F.O. 1987. The influence of cage size and en-vironmental enrichment on the development ofstereotypies in bank voles (Clethrionomys glareo-lus). Behavioural Processes 14:155–173.
Oviatt, L.A., and J.R. Millam. 1997. Breeding behaviorof captive orange-winged amazon parrots. ExoticBird Report 9:6–7.
Owens, D.G.C., E.C. Johnstone, and C.D. Frith. 1982.Spontaneous involuntary disorders of movement—their prevalence, severity, and distribution inchronic-schizophrenics with and without treatmentwith neuroleptics. Archives Of General Psychiatry39:452–461.
Pearce, G.P., A.M. Patterson, and A.N Pearce. 1989.The influence of pleasant and unpleasant handlingand the provision of toys on the growth and behav-iour of male pigs. Applied Animal BehaviourScience 23:27–37.
Powell, S.B., H.A. Newman, T.A. McDonald, P.Bugenhagen, and M.H. Lewis. 2000. Developmentof spontaneous stereotyped behavior in deer mice:Effects of early and late exposure to a more complexenvironment. Developmental Psychobiology 37:100–108.
Price, E.O. 1984. Behavioral aspects of animal domes-tication. Quarterly Review of Biology 59, 1–32.
Price, E.O. 1999. Behavioral development in animalsundergoing domestication. Applied Animal Be-haviour Science 65:245–271.
Reinhardt, V. 1991. Group formation of previouslysingle-caged adult rhesus macaques for the purposeof environmental enrichment. Journal of Experi-mental Animal Science 34:110–115.
27 / Captive Parrot Welfare 317

Renner, M.J. 1987. Experience-dependent changes inexploratory behavior in the adult rat (Rattus norvegi-cus): Overall activity level and interactions with ob-jects. Journal of Comparative Psychology 101:94–100.
Renner, M.J., and M.R. Rosenzweig. 1987. Enrichedand impoverished environments: Effects on brainand behavior. New York: Springer-Verlag.
Reynolds, M. 1998. The welfare of pet parrots.Psittascene 10:1–3.
Savory, C.J., E. Seawright, and A. Watson. 1992.Stereotyped behaviour in broiler breeders in relationto husbandry and opioid receptor blockade. AppliedAnimal Behaviour Science 32:349–360.
Shepherdson, D.J. 1993. “Environmental enrichment:An overview.” Proceedings, Annual Conference ofthe American Association of Zoological Parks andAquariums, pp. 100–103.
Shepherdson, D.J. 1998. “Tracing the path of environ-mental enrichment in zoos.” In Second nature:Environmental enrichment for captive animals, ed.D.J. Shepherdson, J.D. Mellen, and M. Hutchins, pp.1–12. Smithsonian Institution.
Snyder, N.F.R., J.W. Wiley, and C.B. Kepler. 1987. Theparrots of Luquillo: Natural history and conserva-tion of the Puerto Rican parrot. Los Angeles:Western Foundation of Vertebrate Zoology.
Sparks, J., and T. Soper. 1990 Parrots: A natural his-tory. New York: Facts on File.
Taylor, G.T. 1981. Fear and affiliation in domesticatedmale rats. Journal of Comparative PhysiologicalPsychology 95:685–693.
Terlouw, E.M.C., A.B. Lawrence, and A.W. Illius.1991. Influences of feeding level and physical re-striction on development of stereotypies in sows.Animal Behaviour 42:981–991.
van Hoek, C.S., and C. ten Cate. 1998. Abnormal be-havior in caged birds kept as pets. Journal of AppliedAnimal Welfare Science 1:51–64.
Vastrade, F.F. 1986. The social behavior of free-rangingdomestic rabbits (Oryctolagus Cuniculus L.).Applied Animal Behaviour Science 16:165–177.
Weary, D.M., L.A. Braithwaite, and D. Fraser. 1998.Vocal responses to pain in piglets. Applied AnimalBehaviour Science 56:161–172.
Wechsler, B., and B. Huber-Eicher. 1997. The effect offoraging material and perch height on feather peck-ing and feather damage in laying hens. AppliedAnimal Behaviour Science 58:131–141.
Weidenmayer, C. 1997. Causation of the ontogeneticdevelopment of stereotypic digging in gerbils.Animal Behaviour 53:461–470.
Westerhof, I., and J.T. Lumeij. 1987. “Feather pickingin the African grey parrot.” Proceedings of theEuropean Symposium on Bird Diseases, Beerse,Belgium. Netherlands Association of AvianVeterinarians, pp. 98–103.
Wood-Gush, D.G.M., A. Stolba, and C. Miller 1983.“Exploration in farm animals and animal hus-bandry.” In Exploration in animals and humans, ed.J. Archer & L.I.A. Birke, pp. 198–209. London: VanNostrand Reinhold.
Würbel, H. 2001. Ideal homes? Housing effects on ro-dent brain and behaviour. Trends in Neurosciences,24(4):207–211.
Würbel, H., R. Chapman, and C. Rutland 1998. Effectof feed and environmental enrichment on develop-ment of stereotypic wire-gnawing in laboratorymice. Applied Animal Behaviour Science, 60:69–81.
318 Manual of Parrot Behavior

Abnormal grooming behavior, 198Abstract aptitude, 136Abyssinian Ground Hornbills, 151Abyssinian Lovebird (A. taranta), 70, 71Accipiter species, 14Acetylcholine (Ach), 267, 268Acrylic beak balls, 250Acupuncture, 260Acute infectious disease, 207Adrenocorticotropin (ACTH), 114Aengus, W.L., 257African Grey hen (Psittacus erithacus erithacus), 225African Grey Parrot (Psittacus erithacus), 165, 205,
219, 226, 240, 263, 282and busyness, 107caging and nesting, 286diagnoses for feather-damaging behavioral, 203fear biting, 211feather-picking disorder, 255healthy neophytes, 97mineral requirements, 54protein requirements, 53territorial aggression, 214
African parrots, 240Aggressive behavior, 175, 308–310
conditioned aggression, 212–214encounters, 44fear biting, 211–212intraspecific aggression, 216mate-related aggression, 215–216Psittacine birds, 44redirected aggression, 216territorial aggression, 214–215
Agonistic display behaviors, 44–45Air quality, 259Alarm calls, 220–221Alberto, P.A., 161Alderton, D., 72Alexandrine Parakeet, 287
caging and nesting, 286parenting, 288
Alex Foundation, 142
Allofeeding, 46, 69, 282Allopreening, 46, 282Alternative behaviors, differential reinforcement of,
158–159Altricial birds, 95Amazona fauna, 14Amazon parrot (Amazona), 14, 22, 205, 255
breeding efforts for, 22–23clutch initiation, 17conditioned aggression, 213feeding visits, 17feral flock of, 101–102fledging age, 17incubation periods, 17mate-related aggression, 215in wild, 13–26
deficits in breeding effort, 22–23feeding behavior and rates, 18–20relationships of species conspicuousness and nest
accessibility, 20–22site and pair fidelity and cavity reuse, 17–18timing of nesting seasons, 16–17values of sociality among, 14–16
American Foundation, The , 142American Veterinary Medical Association, 301American Zoo, 23Amitriptyline (Elavil), 262, 270, 272Animal Behavior Desk Reference, 150Animal Welfare Act, Part D, Nonhuman Primates, 197Anticonvulsants, 275Antidepressants, 272Antihistamines, 275Anti-inflammatory, non-steroidal, 240Antipsychotics, 274Anxiety-related behavior problems, 183Anxiolytic drugs, 212, 229Appeasement, 44Approach/retreat sequences, 309Appropriate-aged pine (Pinus caribea), 16Aquarium Association, 23Ara species, 46, 282Aratinga species, 46, 282
319
Index

Arizona Game and Fish Department, 23Arrowood, P.C., 46, 67Asiatic Parakeets, 168, 282Aspartate aminotransferase (AST), 207Aspergillus organisms, 208Association of Avian Veterinarians, 166, 281Athan, M.S., 169Atmospheric ammonia levels, 303Atrophied gonads, 236Attention-seeking screaming, 222Audiogram, 35Auditory epithelium, 35Austral Conures, 6Australian cockatoos, 283Australian parrots, 240–241Australian psittacines, 72, 74Autism, 311Avian Medicine: Principles and Applications, 169, 179Avipoxvirus, 203
Bacterial infections, 204Bahama Amazons (Amazona leucocephala baha-
mensis), 71Bahama Parrot, 16, 17, 20, 71, 82Bandura, A., 134Bare-faced Curassows, 151Barnett, S.A., 150Barn Owl, 36Barrow, Wylie, 23, 150Bathing, 157–158, 303Beak
anatomy of, 252–253clicking, 169functions, 252gape, 45grabbing, 214grinding, 169lesions, 204prehensile function of, 251–252spar, 45thumping, 214trauma, 251
Beck, A.M., 125Begging, 179Behavior. See also aggressive behavior; behavioral
disorders; sexual behavior problemsabnormal grooming behavior, 198abnormalities in captive psittacids, 287–289affiliative behaviors, 67analysis and parrot learning, 147–163
ABCS of behavior, 148behavior analysis, 147–148case of empowerment, 151–152functional assessment/analysis, 148problems with dominance, 148–151
punishment, 159–161reinforcement, 154–159tools and techniques for behavior change, 152–154
begging behaviors, 179biologic behaviors, 236changing, 152–154classes in veterinary hospitals, 152–154
benefits of, 173–174caveats, 166–167course content, 167–173course description, 166
comfort behavior, 59–62description, 184–185drugs for, 269effects of nutrient deficiencies/excesses, 55–56grooming behavior in Budgerigars, 61hand-reared vs. parent reared
cockatiels, 88orange-winged Amazon parrots, 88
history, 176–177, 256learning theory, 147modification exercises, 260observation, 177–178, 184–185owner’s, 178patterns, 179pharmacotherapy, 267–279
combination therapy, 277–278drug classes, 269–275drug selection and consideration, 268–269mechanism of action, 267narcotic agonists, 276–277narcotic antagonists, 276neurotransmitters, 267–268other agents, 275–276
social, 43–48, 85support, 152tameness, 124veterinary behavior, 178
Behavioral disorderspsittacine, clinical evaluations, 175–185
behavioral history, 176–177behavior description and observation, 184–185development, 179–180drawing conclusions, 185establishing consultant/client relationship, 175evaluating data, 178–179evaluating environment, 177follow-up, 185grooming, 180–183honest answers, 177identifying problems, 175level of cooperation, 175–176observation of behavior, 177–178owner’s behavior, 178physical environment, 183–184
320 Index

prior environments, 183schedule, 184signalment, 179social environment, 184toys, 184
Behavioral problemsdiagnostic workup of, 195–210
aggression, 195bacterial and fungal cultures, 208–209diagnosis of FDB and aggression, 196–203diagnostic tests for feather-damaging behavior,
206–208differential diagnoses for FDB, 203–206differential diagnosis for aggression, 206feather-damaging behavior, 195
Behavior History Form, 186–193Bellanca, R.U., 196Benefit/risk ratio of drug options, 269Benzodiazepines (BDZ), 263, 269, 272Bergman, Laurie, 59–62, 219–222Beta blocking drugs, 268Bill nibbling, 169Bill touching, 67Biochemical analysis, 203Biologic behaviors, 236Biopsy, 203Bird husbandry, 243Bird mommy, 176Bird socialization class, 166Biting, 211–212Black-billed Amazons, 70Black-billed Parrot, 19, 20Black Cockatoo, 6, 8, 73Blanchard, S., 180, 229Blood loss, 256Blue-and-Gold Macaw (A. ararauna), 50, 71, 108, 205Blue-fronted Amazon Parrots (Amazona aestiva), 105,
150, 204Blue-throated macaw (A. glaucogularis), 30, 50Bollinger bodies, 204Bonding, 171–172, 206, 235Bootleg reinforcement, 160, 161Bordnick, P.S., 259Boredom, 256Braming’s Finches, 203Branches, 295Breeding, 22–23
conditions, 118facilities, 301programs, 89seasonal, 237
Breeding-season food, 20Brinker, Bobbi, 147–162Briscoe, J.A., 255Broad-winged Hawk, 15
Brotogeris species, 46, 282Brown-headed Parrots (Poicephalus cryptoxanthus), 65Brown-throated Conures, 168Brue, Randy, 23Budgerigar (Melo-psittacus undulatus), 59
aggressive behavior, 211caging and nesting, 286courtship and copulation, 69, 70courtship behavior, 282–283diagnoses for feather-damaging behavioral, 203,
205, 206feather-picking disorder, 255feeding strategies and diet ingredients, 50female, 69grooming, 182hand-rearing, 84hearing, 34–36mating pairs and groups, 46mating systems, 64, 66nesting, 72, 74pair relationships, 67, 68signalment, 179species variation, 236taste and smell, 37therapeutic remedies, 241viral PCR and other test procedures, 209vocalization, 220, 238
Budgies, 219, 242, 292Buffon’s macaw (A. ambigua), 50Buprenorphine (Buprenex), 271, 276Burrowing Parrot (Cyanoliseus patagonus), 64, 65,
66, 68, 71, 73Butorphanol (Torbugesic), 262, 276Butterfield, P.A., 46Butyrophenone antipsychotic agent, 263
Cacatua, 73, 282Caging, 215, 259
aggression, 195bound, 212captive psittacids, 283–287for problem prevention, 292–294
Calculated nutrient requirements, 53–54Callocephalon, 73Calorie-dense food, 295Calyptorhynchid feeding apparatus, 4Calyptorhynchus, 73Campus Veterinarian, 117Canaries, 36, 203, 205Canary-winged Parakeet (Brotogeris versicolurus), 46,
64, 65, 67, 71Candida, 204Captive-bred parrots, reintroduction of, 31Captive parrot welfare, 301–318
captive environment and parrot welfare, 303–304
Index 321

Captive parrot welfare (continued)environmental enrichment, 305–313
fearfulness and aggression, 308–310interactions with human handlers, 310psychogenic feather picking, 307–308screaming, 313stereotypic behavior, 310–313
parrots and domestication, 305practical considerations for improving, 313–314threats to parrot welfare, 303what it is, 301–303wild parrots, 304–305
Captive psittacids in breeding aviary, behavior of, 281–290behavioral abnormalities, 287–289caging and nest selection, 283–287courtship behavior, 282–283parenting, 287socialization, 281–282
Carbamazepine (Tegretol), 271, 275–276Carolina Parakeet (Conuropsis carolinensis), 16Carprofen (Rimadyl), 271, 276–277Casuarinas, 5Catechol-O-methyltransferase (COMT), 268Category, concepts of, 135–136Cattell, Raymond B., 117Cavity-nesting birds, 18, 239Cavity-poor environments, 18Cavity reuse, 17–18Cavity seeking, 235Cavity-type nest boxes, 79, 81Cayman Brac Parrot (Amazona leucocephala
hesterna), 15, 16, 21Cebus studied, 141Cell-mediated immunity, 123–124Chaffinches, 203Chapman, G., 84Chemosensory perception, 51Cheney, C.D., 147Chicks
fostering, 118, 121tameness, 119, 125
Chick Tameness Score Sheet, 130Chlorpromazine (Thorazine), 270, 274Choanal cultures, 208Chorionic gonadotropin (APL), 262, 271, 277Chronic confinement, 257Chronic egg production, 234Chronic reproductive stimulation, 239Chronic schizophrenic patients, 311Cigarette smoke, 183Circannual rhythms, 287Circovirus, 256Citron-crested Cockatoo (Cacatua sulphurea
citrinocristata), 226Classes, drug, 269–275
Classical conditioning, 227, 230Clicker training, 221, 243Clipping style, 297Clomicalm, 273Clomipramine (Anafranil), 262, 263, 270, 273Cloutier, S., 45Clutch initiation, for Amazona parrots, 17Cocatua species, 46Cockatiel (Nymphicus hollandicus), 4, 8, 88
chicks, 54, 79courtship and copulation, 69courtship behavior, 283diagnoses for feather-damaging behavioral, 205, 206dietary vitamin A, 55dominance, 150feather-picking disorder, 256feeding strategies and diet ingredients, 50grooming, 182hearing, 35housing and management considerations for, 292implications for, 53mating systems, 46, 64, 65pair relationships, 67, 68parental care, 73parenting, 287physical contact, 242positions of higher status, 44predisposition of, 255reproductive behavior, 84signalment, 179species variation, 236taste and smell, 37taste threshold of, 51–53therapeutic remedies, 241vitamins for, 54vocalization, 219, 220
Cockatoo (C. leadbeateri), 84, 205, 242feather-picking disorder, 255, 263intraspecific aggression, 216mate-related aggression, 215
Cockatoo neophytes (Cacatua), 100Cognitive development, 136Collars, 72, 260Collette, J.C., 115, 126Color game, 170–171Combination therapy, 277–278Combinatory learning, 140–141Comfort behavior, 59–62Communication, Grey Parrot, 133–145
further studies on acquisition, 138–141how learn parallels with humans, 134–135importance of interspecies communication, 133–134parallel evolution of avian and mammalian abilities,
141–142results from Pepperberg 2002B, 135–138
322 Index

Companion parrots, 89–90Conditioned aggression, 212–214Confidence, 170–171Confidence and independence, 170–171Confounded number set, 137Congo African Grey Parrot (Psittacus eythacus), 263Conjunctive tasks, 138Consejo Nacional de Ciencia y Tecnologia (CONA-
CYT), 23Conservation, parrot, 27–30Consultant/client relationship, 175Continuous reinforcement (CRF) schedule, 156Contrafreeloading behavior, 151Conure species
feather-picking disorder, 255parenting, 287territorial aggression, 214
Copulation, 68–70, 238–239Corellas (Cacatua tenuirostris), 72Corticotropin-releasing hormone (CRH), 114Counseling pet owners, 166Counterconditioning, 229, 230Courtship, 68–70Cramton, Brenda, 113–126Cranial nerve, 252Cravens, Eb, 101Crawmer, Catherine, 158Creatinine phosphokinase (CPK), 207Crimson Rosella (Platycercus elegans), 64, 74, 304Cross-gender allopreening, 46Crowell-Davis, S.L., 44, 45–46, 150Cruz, Javier, 23Cryptococcus neoformans, 204Crystalloid fluids, 251Cuban Parrot (Amazona leucocephala), 13, 15, 19Cuban Parrots on Grand Cayman (Amazona leuco-
cephala caymanensis), 15Cutaneous herpesvirus lesions, 203
Davis, C.S., 115, 117, 168, 306Dawkins, M.S., 302Deep ophthalmic nerve, 252Defecation, 122, 124Delayed-type hypersensitivity (DTH), 115Depigmentation, 203Dermanyssus, 205Desensitization techniques, 260Detection abilities, 36Diagnostic and Statistical Manual of Mental
Disorders, The, 4th (DSM-IV), 259Diazepam (Valium), 262, 270, 272Diets, high fat, 257Digestive anatomy, 49–50Dilger, William C., 65, 238Diphenhydramine (Benadryl), 262, 270, 275
Direct interspecies communication, 133Domestication and parrots, 305Domestic fowl, 56Domestic gallinaceous hens, 44Dominance, 150–151
aggression, 212and parrots, 150–151problems with, 148–151of Psittacine birds
indicators of, 44measuring of, 44–45relationships, 43–44
relationship function, 44Dooling, Robert J., 33–38Dopamine (DA), 267, 268Dopamine antagonist haloperidol, 227Doss, J., 229Double-eyed Fig Parrots (Opopsitta diophthalma), 72Double yellow-headed Amazon (Amazona ochro-
cephala tresmariae), 101, 102Doxepin (Sinequan), 262, 270, 272–273Drugs
for behavior, 269benefit/risk ratio of drug options, 269beta blocking drugs, 268classes for, 269–275desensitization, 269for feather-picking disorder, 261Food and Drug Administration (FDA), 268nonsteroidal anti-inflammatory drugs (NSAIDs), 268selection, 268–269
Duke University, 235Dutorphanol (Torbugesic), 271Dynamic cage environment, 310Dynamic captive environment, 314Dystrophic feathering, 204
Echo Parakeet, 30, 282Eclectus Parrots (Eclectus roratus)
bond, 168courtship behavior, 283diagnoses for feather-damaging behavioral, 205mate trauma, 248, 249neophytes exercise, 98neophytes preen, 100nesting, 238phobic parrots, 226socialization, 282
Egg laying, 22, 239Egg peritonitis, 234Ejido, Tutuaca, 23Ekstrom, J., 69Electroencephalographic (EEG) studies, 59Electromyographic (EMG) studies, 59Electronic noises, mimicking, 221
Index 323

ELISA tests, 207Elizabethan-style collars, 260Empathy, 116, 121–122Employee Health Services, 117Empowerment, 151–152Enclosed shelters, 294Endocrine regulation, 235–236Endoparasitism, 256Energy requirements, 53Enkerlin-Hoeflich, Ernesto C., 13–23Environmental conditions, 236Environmental enrichment, 288–289, 305–313
fearfulness and aggression, 308–310improving parrot welfare through, 313–314interactions with human handlers, 310psychogenic feather picking, 307–308screaming, 313stereotypic behavior, 310–313
Environments, 94–95, 171, 183–184cues, 236–237evaluating, 177inappropriate, 175irritants, 255social, 184stimuli, 287
Eolophus, 73Epinephrine (E), 267, 268Erosive lesions, non-symmetrical, 204Erratic movements, 183Escape, 154Escape training, 162Ethological studies, 34European Starlings, 44, 52Exaggerated reproductive behavior, 257Exercises, 98, 105, 172, 243Extinction (EXT), 156Extra-pair mating, 46Eye pinning, 169
Facial feather twitching, 169Farber, W.L., 260Fear, 175, 308–310
and aggression, 195, 213biting, 211–212, 225excessive, 175fear-diminishing effect, 309fear-evoking stimulus, 228fearful vocalizations, 212, 214fear-induced behavioral disease, 227fear-provoking stimulus, 221parrots, 225–231
classical conditioning and fear response, 227counterconditioning in classical conditioning, 230fear-based aggression, 225–226operant conditioning and fear response, 227–228
phobic parrots, 226–227prevention, 230rehabilitation of phobic or neurotically frightened
parrot, 229response substitution, 230systematic desensitization, 229–230training, 230veterinary exam room, 227–228
response, 227–228Feather destructive behaviors (FDB), 185, 195,
196–206, 208, 226, 234Feather-picking disorder, 255–265
comparison to human psychiatric disorders,258–259
consequences of, 257diagnostic of, 255–257pharmacological agents, 260–263postulated causes of, 257–258species predispositions, 255treatment of, 259–260
Feather scoring system, 198, 201Fecal examination, 207–208Feeding
adjusting, 242–243behavior of Psottacines, 18–20behavior of Rhynchopsitta parrots, 18–20behavior of wild Amazona, 18–20for courtship, 238food-dispensing toys, 220food reinforcer, 155schedules, 295
Feelings-based approaches, 302Female Budgerigars, 69Female domestic gallinaceous hens, 44Female Speckled Parrotlets, 44Filial imprinting, 169Finches, 209Fledging, 17, 104–110
benefits of foraging, 106benefits of preen, 107benefits of touch, 106–107companions who never fledge or fly, 109fledgling aviary, 104–105height inconvenience, 107–109mental benefits of fledging learning, 105–106physical benefits of exercise, 105social benefits of interacting, 105
Flock formation, Psittacine birds, 43Floor-laying birds, 79, 82Fluffing, 169Fluoxetine (Prozac), 262, 270, 273–274Flying, 45, 109, 225Follicular dysplasia, 206Food and Drug Administration (FDA), 263Footprint-shaped yeast, 204
324 Index

Foot tapping, 169Foot toys, 184Foraging, 198, 256
benefits of, 106devices for, 259, 295
Forshaw, J.M., 72Francis, D.D., 88Franck, D., 45, 46, 66Fraser, D., 302Freidman, S. G., 94, 147–162Frontal Towel Approach, 180, 181Frustration effect, 213F-statistic, 120Fungal abscess formation, 252Fungal diseases, 204Fungal organisms, 204
GABA-ergic activity, 263Gabriel Foundation, 301Galahs (Cacatua roseicapilla), 84, 220Galahs (Eolophus roseicapillus), 61, 64, 65, 66, 70,
71, 73Gall, M.D., 148Gallinaceous hens, 44Games, 170–171Gamma-aminobutyric acid (GABA), 267, 268Gamma fraction, 207Gang-gang Cockatoo (Callocephalon fimbriatum), 4,
8, 69, 304Garner, J.P., 259Garnetzke-Stollmann, K., 45, 46, 66Gender effects, 44, 45Gestation, 236Giardia, 207Gilbertson, T.A., 51Glossy Black Cockatoo (Calyptorhynchus lathami), 4,
64, 65, 72Gnam, R.S., 13Goffin’s Cockatoos (Cacatua goffini), 240, 241Golden Conure (Aratinga guarouba), 65, 66, 73, 74Golden-shouldered Parrots (Psephotus chrysoptery-
gius), 71Gonadotropin-releasing hormone (GnRH), 241Gondwanaland, 234Gould, S.J., 148Gouldian Finches, 205Graham, Jennifer, 33–38Gram stains, 208Grandry corpuscles, 37Granulomatous lesions, 204Greater Antilles, 14Greater Vasa Parrot, 66, 69, 70Great Green Macaw (Ara amigua), 237Green-rumped Parrotlet (Forpus passerinus), 64, 68,
69, 70, 72, 73, 74
Green-winged Macaw (Ara chloroptera), 30, 50, 106,238, 263
Grey-cheeked parakeets (Brotogeris pyrrhopterus),46, 205, 208, 255
Greylag goslings, 113Grey Parrot, 97, 263, 282
caging and nesting, 286cognition and communication, 133–145
evolution of avian and mammalian abilities,141–142
interspecies communication, 133–134learning parallels with humans, 134–135results from Pepperberg 2002B, 135–138
feather-picking disorder, 255Greys (P. erithacus), 100Grigor, P.N., 45Grindlinger, H., 263Grooming, 61, 180–183, 298–299Gross, W.B., 114Ground-nesting psittacine, 82Ground Parrot (Pezoporus wallicus), 50, 71Guide to the Quaker Parrot, 169Gustation, 50–53Gyms, 294
Habitat housing, multiple, 292Hair cell function, 35Halfmoon Conure (Aratinga canicularis), 59Haloperidol (Haldol), 262, 263, 270, 274–275Hamilton, Robert B., 23Handling, 86, 88, 113–114, 115–116, 118, 124–125,
310Hand-reared (H), 86, 88
baby parrots, 179behavioral impacts and implications for captive
parrots, 83–92Cockatiels, 85, 90maternal separation, 85Orange-winged Amazons, 89
Hands, phobia of, 228Hardwired behaviors, 235Hardy, J.W., 44, 45, 67Harpy Eagle Catch technique, 180–181Harrison, C.J.O., 46, 169, 179Harrison, G.J., 263Harvest concessions, 29Healthy social environments, 171Hearing, 34–37Heart rate (HR), 198Hemorrhages, 203Hennessy, J.W., 124Hens, female domestic gallinaceous, 44Herbst corpuscles, 37Herpes lesions, 203Herring Gulls, 52
Index 325

Heterophil count, 203Higgins, P.J., 72High-input companions, 291–292High-strung birds, 227Hippoboscid fly, 205Hispaniolan Parrot (Amazona ventralis), 13, 19, 71, 72Histopathology, 208Homberger, Dominique G., 3–12Hooded parrot (P. dissimilis), 50Hooding, 173Horizontal cages, 293Hormones, 240, 277Hostile environment, 172Housing, 116–120, 292Human-avian bond, 113, 169Human psychiatric disorders, 258–259Husbandry, 243, 303Hyacinth, 28, 29–30Hyacinth Macaw (Anodorhynchus hyacinthinus), 28,
50, 95, 292Hydroxyzine (Atarax), 262, 271, 275Hyperaggressiveness, 308Hyperkeratosis, 204Hypersensitivity dermatitis, 205Hypothalamic-pituitary-adrenal (HPA), 83, 114Hypothalamic-pituitary-gonadal (HPG) axis, 233
Iatrogenic fractures, 182Immune function effects, 54–55Immune status, 126Immunoresponse test, 119–120Imperial Parrot (Amazona imperialis), 13, 14, 21, 72Incubation periods, for Amazona parrots, 17Independence, 170–171Indian Ringneck Parakeets, 220, 282Indonesian parrots, 240–241Infectious diseases, 166–167, 207, 256Inflammatory skin, 203Interactive dynamic attention, 171Interspecies communication, 133–134Intradermal skin testing, 208Intraspecific aggression, 216, 247Intravenous (IV) dextrose, 251Intrinsic reinforcers, 135Island-like forest habitats, 14Isosexual allopreening, 46
Jamaican Black-Billed Parrot (Amazona agilis), 13, 20Japanese quail, 38Jarvis, Erich, 235Jersey Wildlife Preservation Trust, 30John Simon Guggenheim Foundation, 142Judgmental language, 175
Kakapo (Strigops habroptilus), 9, 63, 66, 69, 73, 74Karson, S., 117
Katcher, A.H., 125Kazdin, A.E., 153Keas (Nestor notabilis), 71, 282Kenneth A. Scott Charitable Trust, 142Keratin, 252Keratinized epidermis (horn or rhamphotheca), 252Ketofen, 277, 282Ketoprofen (Orudis), 271, 277King Parrots, 69Knemidokoptes, 205, 208Korbel, Ruediger, 33–38Koutsos, Elizabeth A., 49–56
Labeling, 135, 148Lammertink, Martjan, 23Lattal, K.A., 155Laughing Gulls, 52Laurival Lima, 30Learning and development, 169Lear’s macaw (A. leari), 50Lesions, beak, 204Lesser Antillean species, 14–15, 16, 22Lesser Sulfur-crested Cockatoo, 247Leuprolide acetate (Lupron), 216, 239, 262, 271, 277Levine, S., 124Levinson, S.T., 46Lice, 205Light/dark cycles, 303Lilac-crowned Amazons (Amazona finschi), 19, 101,
102Linden, Phoebe Greene, 93–110, 170Linguistic development, 136Liver damage, 207Locomotion-based behaviors, 202Lorenz, K., 113Lories, 204Lorikeets, 204Loro Parque, 83, 293Lourival Lima, 28Louse fly, 205Lovebird (Agapornis species), 70, 71, 204, 206, 238,
248Low-intensity punishment, 160Luescher, Andrew U., 93–110, 168, 211–216,
225–231, 291–299Lupron, 240, 241Luteinizing hormone (LH), 287Lymphocycte count, 203
Macaw (Ara), 238conditioned aggression, 213feather-picking disorder, 255mate-related aggression, 215parenting, 287
Macronyssus, 205Madagascar (Agapornis cana), 70, 71
326 Index

Major Mitchell’s Cockatoo (Cacatua leadbeateri), 64,65, 66, 71, 284
Malassezia, 204Mallard ducks, 113Malnutrition, 256Mammalian abilities, 141–142Marler, P., 142Maroon-bellied Conure (Pyrrhura frontalis), 296Maroon-fronted Parrot (R. terrisi), 14, 15, 16–17, 18,
71Martin, Kenneth M., 267–278Martin, Steve, 147–162Martinez del Rio, 51Masello, J.F., 72Masked Lovebird (Agapornis personata), 65, 71Mate bond, 176Mate-related aggression, 213, 215–216Maternal tameness, 125Mate trauma, 247–253
anatomy of beak, 252–253mate trauma attack, 248–250prehensile function of beak, 251–252preventative management, 250–251treatment, 251
Mating systems, 64–66Matson, Kevin David, 49–56Matsuzawa, T., 137Mayer, G.R., 156McElroy, Katy, 98Meal-feeding, 243Meaney, M.J., 83Meanness, 225–226Mechanoreceptors, 38Medical disorders, 176, 255Medroxyprogesterone acetate (Depo-Provera), 271,
277Meehan, Cheryl, 197, 259, 301–314Melopsittacus, 46, 282Mench, Joy, 197, 259, 301–314Merkel cell receptors, 37Metamichrolichtus mites, 205Metamichrolichus nudus, 208Mexican Amazona, 16Mexican Commission for Biodiversity Research
(CONABIO), 23Mexican Fund for Nature Conservation (FMCN), 23Microsporum gypseum, 204Migration, 236–237Millam, J.R., 115, 197, 236, 257Mimicking electronic noises, 221Minerals, requirements, 54Mites, 205, 208Mitred Conure (Aratinga mitrate), 297MIT School of Architecture and Planning, 142Model/rival (M/R) system, 134–135, 170Molting, 237, 256
Moluccan Cockatoo (Cacatua moluccensis), 97, 98,204, 240, 241, 292
Monk Parakeet (Myiopsitta monachus), 64, 65, 69, 70,71, 72, 73, 74, 255
Monoamine oxidase (MAO), 268Monoamine oxidase A (MAO A), 268Monterrey Institute of Technology (ITESM), 23Moribund mate trauma, 251M/R training, 137, 138, 139, 140Mucor, 204Multiple habitat housing, 292Munn, Charles, 236, 238Mycobacterial infections, 205Mycoplasma gallisepticum, 114Myers, S.A., 89Myialges, 205Myrtaceae, 5
Naltrexone (Revia), 262, 263, 271, 276Narcotic antagonists, 276–277Near-sea-level environment, 20Negative punishment, 159–161Negative reinforcement, 154, 160Neonates, 95–96, 170Neophobia, 87–88Neophytes, 96–100, 170Neoplasia, 256Neotropical species, 13, 16, 234, 236, 238, 239–240,
242Nertriptyline (Pamelor), 273Nest
accessibility, 20–22preferences, 79–82preparation, 235selection, captive psittacids, 283–287
Nesting, 70–72, 238, 284determining unseen schedules, 102–103seasons for Amazona, 16–17seasons for Rhynchopsitta parrots, 16–17
Nett, N.C., 255Neurochemical abnormalities, 257Neurotically frightened parrot, 229Neurotransmitters, 267–268Neutral territory, 213Newcastle disease, 115Nibbling, bill, 169Night cage, 292Night Parrots (Geopsittacus occidentalis), 71Noise, 183Non-contact communication, 212Non-existence, concept of, 136Non-inflammatory skin, 203Non-injurious self-abuse, 197Non-steroidal anti-inflammatory, 240Nonsteroidal anti-inflammatory drugs (NSAIDs), 276Non-symmetrical erosive lesions, 204
Index 327

Norepinephrine (NE), 267, 268Nortriptyline (Pamelor), 270Nothofagaceae, 5NSF supplement, 142Nuisance-screaming problem, 170Nutrition, 49–58
digestive anatomy, 49–50effects of nutrient deficiencies excesses, 54–56gustation in psittancines, 50–53nutrient deficiencies excesses, 54–56nutrient requirements, 53nutrient-rich diet daily, 234nutritional deficiencies, 204, 255nutritional enrichment, 56physiology, 49–50psittacine nutrition, 49wild-type diets, 49Nymphicus, 73
Obedience training, 166Object-directed stereotypes, 311Obsessive-compulsive disorder (OCD), 258–259O’Dell, J.W., 117Orange-bellied Parrot, 9Orange-chinned Parakeet (Brotogeris jugularis), 44,
67, 237Orange-fronted Conure (Aratinga canicularis), 35, 36,
37, 64, 66, 67, 68, 69, 70, 71Orange-fronted Parakeets, 44Orange-winged Amazon Parrot (Amazona amazonica),
88animals and housing, 118companion parrots, 89curtail cavity seeking, 242feather-picking disorder, 258feeding strategies and diet ingredients, 50hand-rearing, 84psychogenic feather picking, 307rearing history, 197social preferences, 85species variation, 236stereotypic behavior, 312vocal behavior, 86in wild, 304
Orosz, Susan E., 195–209Ortiz-Maciel, S. Gabriela, 23Otto, Roger, 23Out-of-territory interactions, 172Overcrowding, 216
Pachyrhyncha, 14Packard, Jane, 23Pair bonding, 17–18, 46, 61, 66–68, 168, 169, 176,
235, 237Paired Cockatiels, 66
Pair-housing, 259, 306, 309, 310Palm Cockatoo (Probosciger aterrimus), 4, 70, 72,
238, 248, 249Papillomavirus, 203Paradise Parakeets (Psephotus pulcherrimus), 71Paradoxical sleep (PS), 59–60Parasites, 208Parental vocalization, 282Parenting, 72–74, 84–89, 119, 122–126, 287Parent Tameness Score Sheet, 131Paroxetine (Paxil), 270, 274Parrotlets, 206Parrots: Look Who’s Talking (Nature), 166Patagonian Conure (Cyanoliseus patagonus), 168,
237, 282Pavlovian conditioning, 227PCR technology, 204Peach-faced Lovebird (Agapornis roseicollis), 71,
179, 238Pearly-eyed Thrashers, 18Peck threat, 45Penicillium, 204Pepperberg, Irene M., 86, 133–142, 170Perching stations, 215Peregrine Falcons (Falco peregrinus), 14Perez, Tila, 23Peripheral receptors, 239Perivascular dermatitis, 205, 208Perry, R.A., 255Pet Attitude Scale, 117, 129Pet Care Trust, 142Pet owners, counseling, 166Pet trade, 20–22Pharmacological agents, 260–263Phenobarbital (Solfoton), 271, 275Phobia, 89, 226–228Physical abuse, 227Physical contact, 242Physical enrichments, 198Physical examination, 202Physiologic regulation, 235Physiology, 49–50Phytohemagglutinin-M, 120Pierce, W.D., 147Pigtail macaques (Macaca nemestrina), 196Pineal gland, 237Pink Cockatoo (Cacatua leadbeateri), 4, 8, 220Pionus parrots, 204Pituitary gland, 237Placebo-controlled clomipramine trial, 263Plant-based (folivorous) diets, 49Plasma biochemical analyses, 206, 207Plasma electrophoresis (EPH), 207Play gyms, 294Podocarpaceae, 5
328 Index

Poicephalus parrot, 287Poisonwood (Metopium toxiferum), 16Polyfolliculitis syndrome, 206Polyomavirus, 203, 209, 256Positive punishment, 159–160Positive reinforcement, 154, 220Post-beak trauma, 252Postnatal handling, 115–116Power, D.M., 44, 45, 46Precocial birds, 95Predatory tendencies, 183Preening, 99–100, 107Princess Parrots, 205Probosciger, 73Procellariiform tubenose seabirds, 51Progesterone, 237Prolactin, 237Proteaceae, 5Protective aggression, 195Protein/amino acids, 53–54Pruritis and feather damage, 208Pseudo thicket, 294Psittacid feeding apparatus, 4Psittaciformes
evolutionary origin of, 3–6subdivision and classification of, 6–8
Psittacine beak and feather disease (PBFD), 203Psittacine behavioral biology, 234–236Psittacine behavioral disorders
clinical evaluations, 175–185behavioral history, 176–177behavior description and observation, 184–185development, 179–180drawing conclusions, 185establishing consultant/client relationship, 175evaluating data, 178–179evaluating environment, 177follow-up, 185grooming, 180–183honest answers, 177identifying problems, 175level of cooperation, 175–176observation of behavior, 177–178owner’s behavior, 178physical environment, 183–184prior environments, 183schedule, 184signalment, 179social environment, 184toys, 184
Psittacine behavioral pharmacotherapy, 270–271Psittacine birds. See also psittacine behavioral
disorderscomparative insights from, 13–26
deficits in breeding effort, 22–23
feeding behavior and rates, 18–20relationships of species conspicuousness and nest
accessibility to exploitation in pet trade,20–22
site and pair fidelity and cavity reuse, 17–18values of sociality among, 14–16
nutrition, 49social behavior of, 43–48socialization, 167–169
Psychiatric disorders, human, 258–259Psychogenic feather picking, 205, 307–308Psychological abuse, 172, 227Psychotropic drugs, 215Puerto Rican Amazon Parrots (Amazona vittata), 9
behavior of wild Amazona, 13courtship and copulation, 69, 70courtship behavior, 283deficits in breeding effort, 22–23feeding behavior and rates, 19mating systems, 47, 64, 65nesting, 71, 72pair relationships, 67, 68rearing history, 197site and pair fidelity and cavity reuse, 18values of sociality among, 16in wild, 304
Pug beak, 252Punishment, 159–161, 227
Quadratomandibular, 3–4Quaker Parakeets, 214, 282, 292Quaker Parrot, 168, 215, 263Quillfeldt, 72Quill mites, 205Quota-based harvest systems, 28
Radio, 220Rainbow Lorikeet (Trichoglossus haemotodus), 54,
64, 65, 69Ramsey, E.C., 263Receptor nerve fibers, 37Red-bellied macaw (A. manilata), 50Red-crowned Parrot (Amazona viridigenalis), 13, 16,
17, 18, 19, 20, 21, 65Red-Faced Lovebird (Agapornis pullaria), 71Red-faced parrot (H. pyrrhops), 50Red-fronted macaw (A. rubrogenys), 50Redirected aggression, 216Red-lored Parrot (Amazona autumnalis), 13, 16, 19,
20, 21Red-masked Conure (Aratinga erthrogenys), 296Red mites, 205Red-necked Parrot (Amazona arausiaca), 13, 14Red-rumped Parrot (Psephotus haemantonotus), 220Red-tailed Amazon (A. brasiliensis), 50
Index 329

Red-tailed Black Cockatoo (Calyptorhynchusbanksii), 4, 5, 8, 68, 72
Red-tailed Hawks (Buteo jamaicensis), 14, 15, 31Red-vented Cockatoo, 247, 248Red-winged Blackbirds (Agelaius phoeniceus), 51Refusal, 228Regents parrot (P. anthopeplus), 50Reinforcement, 154–159, 172Reinisch, Ulrike S., 59–62, 219–222Relationships, 136REM (rapid eye movement), 59–60Removal of stimulus, 154Repetitive behaviors, 198Reproductive behavior, 63–78, 84–85, 233, 235
courtship and copulation, 68–70exaggerated, 257mating systems, 64–66nesting, 70–72pair relationships, 66–68parental care, 72–74
Reproductive hormones, 179Reproductive-related feather picking, 257Reproductive response, 120–124Respiration rate, 122Retina, 33Reuptake inhibitors, 267REU supplement, 142Reward training, 154Rhamphotheca, 8, 252Rhinotheca, 252–253Rhizopus, 204Rhynchopsitta (R. phillipsi) parrot, 13–26
deficits in breeding effort, 22–23diet during breeding timing, 17feeding behavior and rates, 18–20nests, 22site and pair fidelity and cavity reuse, 17–18species conspicuousness and nest accessibility,
20–22timing of nesting seasons, 16–17values of sociality among, 14–16
Rhynchopsitta pachyrhyncha, 13Rhynchopsitta terrisi, 15Ring-necked Parakeets, 205, 248Ritchie, B.W., 263Rock Parakeets (Neophema petrophila), 71Rojas, Emilio, 23Romagnano, April, 247–253Rose-breasted Cockatoo (Eolophus roseicapillus),
226Rosenthal, K., 255Rosskopf, W.J., 255, 257Rostrum mandibularis (lower beak), 252Rostrum maxillaries (upper beak), 252Roudybush breeder pellets, 118
Roudybush maintenance pellets, 118Round cage, 293Rowley, I., 84Rozin, P., 141R. pachyrhyncha, 22R. terrisi, 22Rubber beak bumpers, 250Ruffini ending, 37Rushen, J., 44
Sacramento Zoo, 23Same-sex aggression, 250Sam Parrot, 149Sandell, M.I., 44Scaly-breasted Lorikeet (T. chlorolepidotus), 64, 65,
69Scaly-headed parrot (P. maximiliani), 50Scarlet-chested Parrots, 205Scarlet Macaw (Ara macao), 30, 50, 214, 238Scheid, Steve, 23Schindlinger, Michael, 23Schizophrenic patients, 311Schneider, J.W., 155Scissor beak, 252Sclerotic endocarps, 6Scolding, 227Screaming, 176, 313Seabrook, M.F., 125Seasonal breeding, 237Seasonal migration, 237Seed-shelling procedure, 4, 6Seibert, Lynne M., 43–47, 150, 255–263Selective serotonin reuptake inhibitors (SSRIs),
273–274Self-abuse, non-injurious, 197Self-injurious behavior (SIB), 175, 196, 255Self-mutilation, 175, 176Self-reinforcing precipitous escape reactions, 230Self-select pairmates, 314Seligman, M.E.P., 152Sellers, Christine, 96Semi-natural conditions, 305Senegal Parrot (Poicephalus senegalus senegalus), 84,
226, 248Sensitive periods, 169Sensory capacities, 33–42
hearing, 34–37taste and smell, 37touch, 37–38vision, 33–34
Serotonergic agents, 262Serotonin, 267–268Serpell, J.A., 45, 67Sex determination, 207Sex steroid hormones, 206
330 Index

Sexual behavior problems, 233–245clinical presentations, 239–241environment cues for regulation of sexual behavior
and migration, 236–237normal sexual behaviors of wild psittacine species,
237–239psittacine behavioral biology, 234–236therapeutic remedies, 241–243
Sexual imprinting, 169Sexual monomorphism, 64Sexual regurgitation, 235, 236Sharp-tailed Conures (Aratinga acuticaudata), 296,
297Shaver Starcross chickens, 114Shedding, controlling, 241–242Shelf-type nest box, 81Shelters, 294Shiflett, Jim, 23Siegel, P.B., 114Signal-to-noise ratios, 35Simmons, R., 155Sixteen Personality Factor Questionnaire (16PF), 117Skinner, B.F., 148Skin problems, 202, 208Sleep, 59–62
deprivation, 257requirements, 260sleep-wake cycle, 267
Slender-billed Cockatoos, 104Slender-billed Conures, 6Slow advance, 45Slow wave sleep (SWS), 59–60Smith, H.G., 44Snyder, Noel F.R., 13–23Socialization, 134, 171, 184, 256
among Amazona, Psittacines, and Rhynchopsittaparrots, 14–16
behavior, 43–48, 85benefits of interacting, 105in captive psittacids, 281–282developing healthy environment, 169–170environment, 184inadequate and inappropriate interaction, 256
Sparks, J.H., 45Spectacled Parrotlet (Forpus conspicillatus), 46, 64,
65, 66, 67, 68, 69, 70Speer, B.L., 180Spix’s macaw (C. spixii), 30, 50Spoon, Tracey R., 63–74Squamous cell carcinoma, 204Staphylococcus ., 204Starling (Sturnus vulgaris), 56, 183Stay command, 173Stein, D.J., 259Step down command, 173, 215
Step up command, 172, 214, 215Step-up exercises, 169–170, 213Stimulus, presentation and removal of, 154St. Lucia Parrot (Amazona versicolor), 13Stress hyporesponsive period (SHRP), 125Stress-related behaviors, 170Subcutaneous (SQ) dextrose, 251Subcutaneous hemorrhages, 203Submissive signals, 44Subtropical locations, 237Suchecki, D., 125Sulfur-crested Cockatoos (Cacatua), 204, 256Sulzer-Azaroff, B., 156Sunrise/sunset, 219Swooping down, 180Systematic desensitization, 229–230Systemic diseases, 256
Tail fanning, 169Tamaulipas, 14Tameness, 119, 121–126Taming/training techniques, 303Target training, 158Tarvin, K.A., 44Taste, 37, 51–52Taylor, Ali, 23, 86Television, 220Temperate-zone birds, 79Temperature, 303Tennessee Clinical Endocrinology, 240Terrisi, 13, 14Territorial aggression, 195, 213, 214–215Testing
for Aspergillosis, 208functional analysis, 148intradermal skin, 208procedures, 208–209taste, 51
Therapeutic remedies, sexual behavior problems,241–243
Therapy, combination, 277–278Thermo-regulation, 239, 240Thick-billed Parrot (Rynchopsitta pachyryncha), 14,
15, 16, 20, 31, 68, 70, 237Thick-billed psittacine parrots, 248Thinklings, 170Thoracic limb, 239Thyroid gland, 237Thyroxine, 237Time out, 160Timneh African Grey Parrot, 203Todorov, J.C., 155Todt’s method, 134–135Touch, 37–38, 67, 99, 106–107, 158Tough mindedness scale, 121
Index 331

Tourette’s syndrome, 310Towel-wrapped parrot, 182Toxin exposure, 255Toys, 157, 184, 220, 259, 295–298Trade, parrot, 27–30Traditional veterinary format, 196Training, 171, 230, 243, 299Trichoglossus (lorikeet) species, 45, 67Trichophagia, 259Trichophyton, 204Trichotillomania, 259Trick training, 171Tricyclic antidepressants (TCAs), 262, 263, 268,
272Triton Cockatoo (C. s. triton), 226Trophozoites, 207Tropical locations, 237Troutman, A.C., 161T-stand, 215, 229T type box, 286Tube collars, 261Tully, T.N., 255Two-cage system, 215, 292
Ultrasound evaluation, 209Ultraviolet (UV) light, 34, 287Umbrella Cockatoo (Cacatua alba), 153, 204, 228,
240, 241Unihemispheric sleep, 59University of Arizona Undergraduate Biology
Research Program, 142University of California, 117Unsanitary conditions, 303Uric acid levels, 207U.S. Fish and Wildlife Service/Secretaria de Medio
Ambiente y Recursos Naturales (USFWS/SEMARNAT), 23
Valentine, 86Van Sant, Fern, 233–244Vasa Parrot (Coracopsis vasa), 66, 70, 72, 73, 74Venegas, Diana, 23Ventriculus, 209Vertical box (VB), 80Video recording, 34, 177, 178Viral PCR, 208–209Vision, parrots, 33–34Visual acuity, 33Visual barriers, 216, 287Visual stimuli, 183Vitamins, 54, 55Vocalization, 85–87, 219–223, 237–238
excessive, 175vocal cues, 170
vocal games, 170vocal production, 34
Vulturine parrot (P. fulgidus), 50
Watch and/or listen method, 177Water-limited environments, 236Welle, Kenneth R., 165–174, 175–185, 195, 211–216Whistling, 171White blood cell count (WBC), 207White Cockatoo, 4, 8White-fronted Amazon Parrot (Amazona albifrons),
19, 44–45, 46, 64, 68, 70, 169White-tailed Black Cockatoo (Calyptorhynchus fu-
nereus), 4, 64, 65, 68, 69, 70, 72, 73Whole body radiology, 209Wholesale/retail facilities, 301Wild Bird Conservation Act, 83, 89, 281Wild fledglings, 101–104Wild guava (Tetrazygia bicolor), 16Wildlife Conservation Society, 29Wildlife Foundation, 23Wildlife Preservation Trust, 30Wild nests, 81–82Wild parrots, 3–11, 304–305
evolutionary origin of Psittaciformes, 3–6status of, 8–10subdivision and classification of Psittaciformes, 6–8
Wild-type diets, 49Wiley, James W., 13–23Wilson, Liz, 175–185, 225–231Wing flapping, 45Woerpel, R.W., 255, 257Wooden shelters, 294Woolfenden, G.E., 44Worell, A.B., 260World Parrot Trust, 30Wright, Timothy F., 33–38
Yellow-billed Amazon Parrot (Amazona collaria), 13,72
Yellow-fronted Kakariki, 205Yellow-headed Amazon Parrot (Amazona oratrix), 13,
16, 17, 18, 19, 20, 21, 22–23, 65, 288Yellow-naped Amazon (Amazona ochrocephala
auropalliata), 19, 67, 110, 227Yellow-tailed Cockatoo, 4Yellow-tinted mammalian lens, 34Yolk embolis, 234
Zann, R., 68Zebra Finches, 36Zinc toxicity, 293
332 Index




















