
Citrate enhances in vitro metastatic behaviours of PC3M human prostate cancer cells: Status of...
12
The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777 Citrate enhances in vitro metastatic behaviours of PC-3M human prostate cancer cells: Status of endogenous citrate and dependence on aconitase and fatty acid synthase Maria E. Mycielska ∗ , Timothy P. Broke-Smith, Christopher P. Palmer, Rachel Beckerman, Theodoros Nastos, Kamil Erguler, Mustafa B.A. Djamgoz Divison of Cell & Molecular Biology, Neuroscience Solutions to Cancer Research Group, Sir Alexander Fleming Building, Imperial College London, South Kensington Campus, London SW7 2AZ, UK Received 3 March 2006; received in revised form 14 April 2006; accepted 25 April 2006 Available online 9 May 2006 Abstract Prostate is a unique organ that produces and releases large amounts of citrate. This is reduced significantly in cancer and it is possible that citrate is (re)taken up and used as a metabolite to enhance cellular activity. The main purpose of this study was to determine how cytosolic citrate might affect in vitro metastatic cell behaviours (lateral motility, endocytosis and adhesion). Normal (PNT2-C2) and metastatic (PC-3M) human prostate cancer cells were used in a comparative approach. As regards intermediary metabolic enzymes, aconitase and fatty acid synthase, already implicated in prostate cancer, were evaluated. The level of intracellular citrate was significantly higher in PNT2-C2 cells under both control conditions and following preincubation in extracellular citrate. Supply of exogenous citrate enhanced endocytosis, lateral motility, decreased cell adhesion of PC-3M cells but failed to produce any effect on normal cells. Real-time PCR measurements showed that the mRNA levels of mitochondrial and cytosolic aconitases and fatty acid synthase were significantly higher in PC-3M cells. Correspondingly, aconitase activity was also higher in PC-3M cells. Using cerulenin (an inhibitor of fatty acid synthase), oxalomalate and fluorocitrate (inhibiting aconitases), we investigated the dependence of citrate-induced down-regulation of cellular adhesion on aconitase and fatty acid synthase activities. It was concluded: (1) that strongly metastatic PC-3M cells stored less/utilised more cytosolic citrate than the normal PNT2-C2 cells and (2) that cancer cells could metabolise cytoplasmic citrate via aconitase and fatty acid synthase to enhance their metastatic behaviour. © 2006 Elsevier Ltd. All rights reserved. Keywords: Prostate cancer; Citrate; Metabolism; Aconitase; Fatty acid synthase 1. Introduction Prostate produces, accumulates and releases large amounts of citrate (Costello & Franklin, 1991a). This is made possible by the rate-limiting enzyme mitochon- drial (m) – aconitase (ACNT), expression of which ∗ Corresponding author. Tel.: +207 594 5443; fax: +207 584 2056. E-mail address: [email protected] (M.E. Mycielska). is regulated by testosterone and prolactin (Costello & Franklin, 1991b) whilst its activity is reduced in the presence of intracellular Zn 2+ (Costello, Franklin, Liu, & Kennedy, 2000; Tsui, Chang, & Juang, 2006). Con- sequently, the concentration of citrate in prostatic fluid can be as high as 180 mM (Kavanagh, 1994). In a pre- vious study, we have shown that human normal prostate epithelial PNT2-C2 cells express a K + -dependent trans- porter designed primarily to release citrate into prostatic fluid (Mycielska & Djamgoz, 2004). Thus, citrate 3− was 1357-2725/$ – see front matter © 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocel.2006.04.008
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Citrate enhances in vitro metastatic behaviours of PC3M human prostate cancer cells: Status of...

The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777
Citrate enhances in vitro metastatic behaviours of PC-3M humanprostate cancer cells: Status of endogenous citrate and
dependence on aconitase and fatty acid synthase
Maria E. Mycielska ∗, Timothy P. Broke-Smith, Christopher P. Palmer,Rachel Beckerman, Theodoros Nastos, Kamil Erguler, Mustafa B.A. Djamgoz
Divison of Cell & Molecular Biology, Neuroscience Solutions to Cancer Research Group, Sir Alexander Fleming Building,Imperial College London, South Kensington Campus, London SW7 2AZ, UK
Received 3 March 2006; received in revised form 14 April 2006; accepted 25 April 2006Available online 9 May 2006
Abstract
Prostate is a unique organ that produces and releases large amounts of citrate. This is reduced significantly in cancer and it ispossible that citrate is (re)taken up and used as a metabolite to enhance cellular activity. The main purpose of this study was todetermine how cytosolic citrate might affect in vitro metastatic cell behaviours (lateral motility, endocytosis and adhesion). Normal(PNT2-C2) and metastatic (PC-3M) human prostate cancer cells were used in a comparative approach. As regards intermediarymetabolic enzymes, aconitase and fatty acid synthase, already implicated in prostate cancer, were evaluated. The level of intracellularcitrate was significantly higher in PNT2-C2 cells under both control conditions and following preincubation in extracellular citrate.
Supply of exogenous citrate enhanced endocytosis, lateral motility, decreased cell adhesion of PC-3M cells but failed to produceany effect on normal cells. Real-time PCR measurements showed that the mRNA levels of mitochondrial and cytosolic aconitasesand fatty acid synthase were significantly higher in PC-3M cells. Correspondingly, aconitase activity was also higher in PC-3Mcells. Using cerulenin (an inhibitor of fatty acid synthase), oxalomalate and fluorocitrate (inhibiting aconitases), we investigated thedependence of citrate-induced down-regulation of cellular adhesion on aconitase and fatty acid synthase activities. It was concluded:(1) that strongly metastatic PC-3M cells stored less/utilised more cytosolic citrate than the normal PNT2-C2 cells and (2) that cancercells could metabolise cytoplasmic citrate via aconitase and fatty acid synthase to enhance their metastatic behaviour.© 2006 Elsevier Ltd. All rights reserved.Keywords: Prostate cancer; Citrate; Metabolism; Aconitase; Fatty acid synthase
1. Introduction
Prostate produces, accumulates and releases largeamounts of citrate (Costello & Franklin, 1991a). Thisis made possible by the rate-limiting enzyme mitochon-drial (m) – aconitase (ACNT), expression of which
∗ Corresponding author. Tel.: +207 594 5443; fax: +207 584 2056.E-mail address: [email protected] (M.E. Mycielska).
is regulated by testosterone and prolactin (Costello &Franklin, 1991b) whilst its activity is reduced in thepresence of intracellular Zn2+ (Costello, Franklin, Liu,& Kennedy, 2000; Tsui, Chang, & Juang, 2006). Con-sequently, the concentration of citrate in prostatic fluidcan be as high as 180 mM (Kavanagh, 1994). In a pre-vious study, we have shown that human normal prostateepithelial PNT2-C2 cells express a K+-dependent trans-porter designed primarily to release citrate into prostaticfluid (Mycielska & Djamgoz, 2004). Thus, citrate3− was
1357-2725/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.doi:10.1016/j.biocel.2006.04.008

M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777 1767
transported together with 4 K+ outward; activity wasreduced in acidic pH due to change in the valency ofcitrate (Hering-Smith, Gambala, & Hamm, 2000).
Prostatic cells are non-citrate-oxidizing and the cit-rate: isocitrate ratio is high, 30–40:1 (Costello et al.,2000). This ratio as well as the absolute concentrationof citrate in prostatic fluid change significantly whenthe gland becomes cancerous; in metastasis particularly,the citrate level becomes similar to those found in othertissues (Costello, Franklin, & Narayan, 1999). This dra-matic fall in the concentration of prostatic citrate may bedue to several mechanisms including hormonal changesand reduced Zn2+ concentration (Costello et al., 2000).In a further study, we have shown that strongly metastatichuman prostate cancer PC-3M cells express not onlya K+-dependent ‘outward’ citrate transporter but alsoa Na+-dependent citrate transporter designed primarilyfor uptake of extracellular citrate (Mycielska, Palmer,Brackenbury, & Djamgoz, 2005).
Despite many studies showing the importance of cit-rate production and regulation in prostate, however, noth-ing is known about how high cytosolic citrate/citrateuptake might influence the epithelial cells’ behaviour.Here, for the first time, we have investigated the influenceof cytosolic citrate on cellular activities (motility, adhe-sion, endocytosis and proliferation) in a comparativeapproach to normal (PNT2-C2) and metastatic (PC-3M)human prostatic epithelial cells. We have studied alsopossible metabolic pathways through which taken-upcitrate could be metabolised and used to subserve suchcacaTbrb(gpcsd
2
2
dM
cells MCF-10A (used as control) were grown in DMEMmedium with 5% horse serum, 10 �g/ml insulin, 5 �g/mlhydrocortisone, 20 ng/ml EGF, 100 ng/ml cholera toxinand 4 mM glutamine. All cell lines were grown in 10 cmPetri dishes maintained in a humidified incubator with5% CO2 at 37 ◦C.
2.2. Enzyme spectrophotometry
Cellular uptake of citrate was measured by the spec-trophotometric “citrate lyase method” described ear-lier (Petrarulo, Facchini, Cerelli, Marangella, & Linari,1995; Mycielska and Djamgoz, 2004). Deproteination(to measure ‘free intracellular citrate’) was done using10% TCA. The amount of citrate in digested cells wasdetermined as before (Petrarulo et al., 1995). Back-ground medium absorption (at 330 nm) was checkedbefore the citrate lyase was added to the cuvettes andthis reading was subtracted from the final values; thenet absorption (Abs330) was directly proportional to theconcentration of citrate present (Petrarulo et al., 1995).Differences in net absorbances resulting from varioustreatments were denoted as “�Abs330”. Intracellular cit-rate in PNT2-C2 and PC-3M cells was measured usingthe same basic procedure. The amount of protein in agiven sample was determined using a BIO-RAD proteinassay following the manufacturer’s instructions.
2.3. NADH fluorescence
ellular activity. The key enzymes that metabolize citratere ACNT and ATP citrate lyase. The latter metabolisesitrate into acetylCoA which is the substrate for fattycid synthase (FAS) (Pitula et al., 2004; Baron, Migita,ang, & Loda, 2004). m-ACNT is well established toe involved as a rate-limiting enzyme in prostatic cit-ate production (Costello & Franklin, 1991b). FAS haseen shown to be overexpressed early in prostate cancerSwinnen et al., 2002) and may function as an onco-ene (Baron et al., 2004). In the present study, cells werereincubated in exogenous citrate in order to raise theytosolic levels. The role of ACNT and FAS in the con-equent enhancement of metastatic cell behaviours wereetermined using pharmacological blockers.
. Materials and methods
.1. Cell culture
PNT2-C2 and PC-3M cells were maintained asescribed previously (Mycielska and Djamgoz, 2004;ycielska et al., 2005a). Normal human breast epithelial
This was measured as described before (Mycielska,Krasowska, Grzywna, & Djamgoz, 2005). The cellswere plated into 35 mm Petri dishes at a density of2.5 × 105 48 h before treatments. One set of dishes wasleft untreated with just the growth medium changed; inthe other, the cells were preincubated in 10 mM Na+-citrate for 24 h. Immediately before the measurements,the cells were washed thoroughly twice in PBS andthen covered with 2 ml of 0.5 �M NADH. The cellswere placed under the microscope and left to settlefor about 2 min after which the enzymatic mixture wasadded (65 U of malic dehydrogenase and 150 U of citratelyase). The resulting decrease in the NADH fluores-cence, representing increased citrate concentration, wasrecorded for 10–20 min. The fluorescence signal wastransmitted through a DAPI (excitation D360/40x; emis-sion E420LPv2; Chroma Technology, Rockingham, VT,USA) filter to an infrared camera and then to a photo-multiplier controller which integrated the optical signalsevery 1 ms and recorded by a Cairn Photometric Acqui-sition System (Mycielska et al., 2005b). For time courseanalysis, every 100th point was re-sampled from the

1768 M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777
recordings and 10 such successive points were averagedto give a final data point (plotted) every 10 s (Mycielskaet al., 2005b). The obtained data were converted to per-centages where total decrease in NADH fluorescence(increase of citrate) was treated as 100%. The rest ofthe data was adjusted accordingly.
2.4. Assays of metastatic cell behaviour
1. Motility. Lateral motility of cells was quantified usinga wound-heal assay, as described previously (Silletti,Yao, Pienta, & Raz, 1995; Fraser et al., 2003). Briefly,cells were plated in 35 mm Petri dishes at an initialdensity of 1.5 × 105 cells/dish and allowed to settlefor 24 h. Initial wound widths (W0’s) were deter-mined immediately afterwards (0 h) by measuring thewidths of the wound at the 45 fixed points per dishusing a graticule on an inverted microscope (Zeiss,ID 03). The same sites were re-measured 24 and 48 hafterwards, with a change of medium at 24 h. Cel-lular motility was quantified as the “motility index”(MoI) defined as follows: MoI = 1 − (Wt/W0) whereWt is the width of the wound after 24 or 48 h. Accord-ingly, MoI = 0 indicated no movement of cells whilstMoI = 1 indicated complete wound closure. The pro-liferation assay used has been described in detailelsewhere (Grimes et al., 1995; Fraser, Ding, Liu,Foster, & Djamgoz, 1999). Proliferation was quan-tified using the colorimetric 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay
at least 3 different dishes were recorded. Single-celladhesion measurements from given dishes were aver-aged. In the text “n” denotes the number of dishesstudied. Each mean was compared with the controldata from the same set. Statistical significance wasevaluated using paired t-test.
3. Endocytosis. Endocytic membrane activity was stud-ied by uptake of the non-cytotoxic marker horseradishperoxidase (HRP) as described earlier (Mycielska,Fraser, Szatkowski, & Djamgoz, 2003; Onganer &Djamgoz, 2005). The cells were plated in a 96-welldish at a density of 1 × 105 cells per well. They werethen incubated with 0.5 mg/ml HRP (Sigma, type VI)for 35 min, followed by several washes in cold PBS.The cells were lyased in RIPA buffer and treatedwith diaminobenzidine (DAB) + H2O2. Absorbanceat 550 nm (Abs550) values were read on a MultiscanMCC/340 multi-well plate reader.
2.5. Real-time PCR
Total RNA was extracted from 100 mm dishes of PC-3M cells using StrataPrep Absolutely RNA Miniprepkits (Stratagene, La Jolla, CA). Approximate RNA quan-tity was visualised by spectrophotometric absorbance at260 nm. For each sample, 1 �g of RNA was reverse-transcribed into cDNA with Superscript II reverse-transcriptase (Invitrogen, Paisley, UK), using 250 ng ofrandom hexamer mix (Amersham Biosciences, Chal-font, UK) as a primer. Real-time PCR was performed
(Grimes et al., 1995). Spectrophotometric absorbanceat 570 nm was read 10 min after adding and mixingthe glycine buffer. Using standard curves, it was con-firmed that the readings were on the linear portion ofthe calibrations. Results were compiled as the mean ofat least three repeats for each condition and comparedwith control readings from three respective wells pro-cessed in parallel.
2. Adhesion. Single-cell adhesion was measured asdescribed by Palmer et al. (submitted). 5 × 104 cellswere plated in each 35 mm Petri dish. All drug treat-ments were as described above. Single-cell adhesionwas measured by applying negative pressure to sin-gle cells through the tip of a micro-pipette (diame-ter ∼50 �m). The values of the negative pressuresapplied were recorded online; adhesion was mea-sured as the “detachment negative pressure” (DNPR)normalised with respect to cell size (Palmer et al.,submitted). As there was no significant change in thecell sizes under the different treatment regimes, allthe measurements were converted in DNPR using thesame factor. For each condition, 30–100 cells from
using the DNA Engine Opticon 2 system (MJ Research,Waltham, MA). Duplicate PCR reactions were run foreach sample and �-actin was measured as the con-trol/reference gene to normalise the measured c-/m-ACNT and FAS mRNA expression levels (Diss et al.,2005). PCR reactions were performed in a total reac-tion volume of 20 �l, containing 10 �l Quantitect SYBRGreen PCR mix (Qiagen, Crawley, UK), 500 nM ofeach specific primer and 0.5 �l of cDNA. The PCR wasrun with an initial step of 95 ◦C for 15 min to activatethe HotStar Taq, followed by 40 cycles of 95 ◦C for30 s, 55 ◦C for 30 s and 72 ◦C for 30 s. Fluorescencefrom incorporation of SYBR green to double-strandedDNA was measured after each cycle. A ‘no-template’control was included for each PCR. Product compo-sition was verified with a melt curve in 0.3 ◦C stepsfrom 65 to 95 ◦C. The primers used were as follows:(1) c-aconitase: 5′-GGTGTGATCCCACTTGAATA-3′ and 5′-GGATCATGTAGTTGAGGATG-3′; (2) m-aconitase: 5′-GATCCACGAGACCAACCTGA-3′ and5′-TTGAGGGCACTGCCAGCGCG-3′; (3) FAS: 5′-CCAAGGCCAAGTACCATGGC-3′ and 5′-ACGCTC-

M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777 1769
ACGCGTGGCTCAGC-3′; and (4) �-actin: 5′-AGCC-TCGCCTTTGCCGA-3′ and 5′-CTGGTGCCTGGG-GCG-3′.
The threshold amplification cycles (CT’s) were deter-mined using the Opticon Monitor 2 software and anal-ysed by the 2−��CT method (Livak & Schmittgen,2001).
2.6. Aconitase activity assays
Mitochondrial and whole-cell fractions were used tomeasure ACNT activities according to the basic methodof Drapier and Hibbs (1996). Mitochodria were isolatedby incubating the cells in 0.25 M sucrose buffer withdigitonin. They were then transferred to RIPA bufferwith 1 mM PMSF, 1 �g/ml aprotinin, 1 �g/ml leupeptin,1 mM NaF and 1 g/ml pepstatin. For the whole-cell mea-surements, the cells were treated with the RIPA bufferwith additives only. The cells were then centrifuged at16,060 g (4 ◦C), the supernatant was removed and theamount of protein was determined using the BIO-RADprotein assay. Aconitase activity was measured in 50 mMTris–HCl buffer to which 100 �g of protein and 20 �lof 20 mM cis-aconitate were added. The readings wererecorded every 30 s.
2.7. Data analysis
In general, each experiment was repeated at least threetdu
3
ocati
3
(e“c““
evaluate and compare the “free” pool of citrate avail-able for use in metabolism. Under the control restingconditions, there was 40 ± 5.7% total (17 ± 1.2% free)more endogenous citrate in PNT2-C2 than in PC-3Mcells (p = 0.003 and 0.034, respectively; n = 5; Fig. 1A).The actual intracellular concentrations of citrate in nor-mal cells were 4.0 ± 0.48 and 0.84 ± 0.07 mmols/mg ofprotein (total and free, respectively). These were sig-nificantly lower in the case of PC-3M cells: 2.4 ± 0.10and 0.7 ± 0.01 mmols/mg protein of total and free cit-rate, respectively (p = 0.003 and 0.043 cf. PNT2-C2 cells,respectively; n = 6; Fig. 1A).
In response to long-term (24 and 48 h) preincubationin exogenous citrate, the two cell lines behaved differ-ently (Fig. 1B). There was no significant increase inthe total citrate content of PC-3M cells after preincuba-tion with 1 mM citrate up to 48 h (Fig. 1B). In contrast,after 24 h, PNT2-C2 cells showed a significant increaseby 12 ± 4.1% (p < 0.05; n = 4); there was no furtherincrease after 48 h (Fig. 1B). Increasing the extracellu-lar citrate concentration to 10 mM resulted in significantincreases in the total intracellular citrate levels of bothcell lines. In the case of PNT2-C2 cells, the increase wassignificant and markedly time-dependent: 30 ± 10.3%(24 h) and 92 ± 6.1% (48 h) (p < 0.05; n = 4 for both).Intracellular total citrate detected in PC-3M cells alsoincreased after preincubation in 10 mM citrate, but thiswas much less and not time-dependent: 20 ± 1.9% (24 h)and 22 ± 4.5% (48 h) (p < 0.05 versus control for each;p > 0.05 for 24 versus 48 h; n = 4 for both; Fig. 1B). As
imes, each involving three different dishes of cells. Allata are presented as means ± S.E.M. Paired t-test wassed mostly for the statistical analyses.
. Results
The first part of the study aimed to demonstrate uptakef extracellular citrate and its influence on metastaticell behaviours of normal prostatic epithelial PNT2-C2nd strongly metastatic PC-3M cells. In the second part,he involvement of ACNT and FAS in this process wasnvestigated.
.1. Uptake and release of citrate
As intracellular ions can be either in ‘bound’ forme.g. to proteins) and/or ‘free’ (i.e. active), we differ-ntiated between the two pools and present the data astotal citrate” (i.e. whole cellular citrate measured), “freeitrate” (i.e. citrate measured after deproteination) andbound citrate” (i.e. the value obtained by subtractingfree” from “total” citrate). This was done in order to
regards free citrate, in PC-3M cells, interestingly, therewas a significant 20.7 ± 4.1% increase after 24 h prein-cubation in 1 mM citrate (p = 0.005 versus control; n = 5;Fig. 1C). Increasing the extracellular citrate concentra-tion to 10 mM resulted in 25 ± 8.3% increase in intra-cellular free citrate of PC-3M cells after 24 h (p = 0.005;n = 5; Fig. 1C). Percentage increases in free citrate ofPNT2-C2 cells after 24 h preincuabtion in 1 and 10 mMwere 20 ± 7.2 and 51 ± 9.8%, respectively (p = 0.008and 0.002; n = 5; Fig. 1C).
Release of citrate was recorded over 20 min fromcontrol (i.e. non-pretreated) cells and cells pre-loadedwith citrate (Fig. 2). Control PNT2-C2 cells showeda slight decrease in fluorescence suggesting that basalrelease of endogenous citrate occurred (Fig. 2A); thiswas treated as 100% and used to compare data aver-ages (Fig. 2D). Release of citrate from PNT2-C2 cells,measured similarly following preincubation with 10 mMcitrate for 24 h, was much larger, reaching 337 ± 64.1%in steady state (p < 0.001; n = 15; Fig. 2A and D). Basalrelease of citrate from PC-3M cells was much smaller,40 ± 2.2% (n = 14; p = 0.0014) (Fig. 2B and D). Further-

1770 M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777
more, although PC-3M cells also released more citratefollowing preincubation with 10 mM citrate for 24 h, thisreached only 141 ± 16.3% (p < 0.001; n = 15; Fig. 2Band D). There was no significant release of citrate fromhuman normal breast epithelial MCF-10A cells underboth resting conditions and after 24 preincubation with10 mM citrate (Fig. 2C).
Thus, for the PC-3M cells, the intracellular citratelevel was more stable, and release of citrate was muchsmaller, compared with PNT2-C2 cells under identi-cal steady-state conditions (control or preloaded). Thesedata also suggested that the citrate taken-up by PC-3Mcells increased mainly the pool of free citrate and this sat-urated at low extracellular concentration (an increase inbound citrate was observed only at the higher concentra-tion). The citrate taken up could be utilised in driving thecells’ strong metastatic potential. The latter possibilitywas tested in the following series of experiments.
3.2. Effects of extracellular citrate on metastaticcell behaviours in vitro
The influence of extracellular citrate uptake on lat-eral motility, proliferation, adhesion and endocytosis ofPNT2-C2 and PC-3M cell lines were determined.
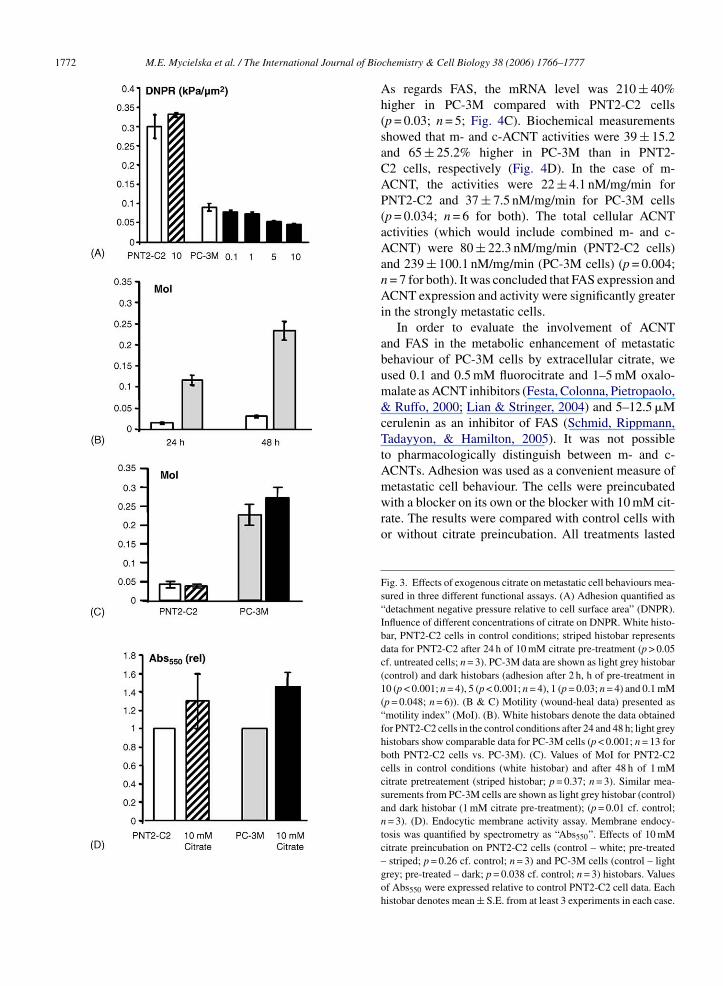
Adhesion. Control experiments showed that PNT2-C2 cells were ∼3-fold more adherent than PC-3M cells(DNPR = 0.30 ± 0.03 and 0.09 ± 0.01 kPa/�m2, respec-tively; p < 0.001; n = 4; Fig. 3A). Pretreatment with10 mM citrate for 48 h had no effect on the adhesion
of the PNT2-C2 cells (DNPR = 0.33 ± 0.01 kPa/�m2;p > 0.05 cf. untreated cells; n = 3; Fig. 3A). In contrast,PC-3M cells were sensitive and adhesion was reducedby extracellular citrate (Fig. 3A). This effect was dose-dependent and at the highest (10 mM) concentration ofcitrate used, adhesion was reduced to 51.4 ± 4.5% of nor-mal (DNPR = 0.046 ± 0.03 kPa/�m2; p < 0.001; n = 3;Fig. 3A).Fig. 1. Spectrophotometric measurements of intracellular resting cit-rate and citrate uptake into PNT2-C2 and PC-3M cells. (A) Spectropho-tometric measurements of intracellular citrate in control conditions inPNT2-C2 (white histobars – total citrate, dark squares – free citrate)and PC-3M (light grey histobars – total citrate, light grey squares –free citrate) cells. The data show values per 1 mg of protein. (B). Spec-trophotometric measurements of total intracellular citrate in PNT2-C2and PC-3M cells pre-incubated in 1 or 10 mM Na+-citrate for 24 or48 h. � total citrate denotes the change in citrate content expressed as apercentage of corresponding resting levels. Hatched histobars, data forPNT2-C2 cells. Black histobars, data for PC-3M cells. Each histobardenotes mean ± S.E. (n > 4). (C) Spectrophotometric measurements offree intracellular citrate in PNT2-C2 (dark squares) and PC-3M cells(light grey squares) pre-incubated in 1 and 10 mM Na+-citrate for 24 h.Each histobar denotes mean ± S.E. (n = 5).

M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777 1771
Lateral motility. Under control conditions, PC-3Mcells were significantly more motile than the PNT2-C2cells, as measured after 24 and 48 h (Fig. 3B). For bothmeasurement times, the motility index of PNT2-C2 wasonly ∼18% of that of PC-3M cells (p < 0.001; n = 13for both). Preincubation with 1 mM citrate did not pro-duce any significant effect on motility after 24 h in eithercell line (not shown). After 48 h, a significant increase
of 15 ± 6.8% was recorded in the motility of PC-3Mcells treated with 1 mM citrate (p = 0.01 cf. control; n = 3;Fig. 3C). In contrast, there was still no effect on PNT2-C2 cells after 48 h of citrate preincubation (Fig. 3C).Citrate uptake had no effect on the proliferation of bothcell lines in all of the conditions tested (not shown).
Endocytosis. The uptake of HRP was significantly(40 ± 14.2%) greater in PC-3M compared with PNT2-C2 cells (p < 0.05; n = 9; Fig. 3D). Preincubation in10 mM citrate resulted in a significant (45 ± 15.7%)increase in uptake in PC-3M cells (p = 0.038, n = 3;Fig. 3D). There was no effect on the PNT2-C2 cells(p = 0.26; n = 3; Fig. 3D).
In conclusion, although PNT2-C2 cells could takeup citrate (Fig. 1B), their metastatic behaviours werenot affected. In contrast, citrate uptake potentiated themetastatic behaviours of PC-3M cells.
3.3. Pharmacological tests of ACNT and FASinvolvement in citrate-stimulated reduction inadhesion
In this part of the study, we aimed to elucidatetwo main possible enzymatic pathways (ACNTs andFAS) which could be involved in the metabolism ofcitrate taken up from extracellular space and used inthe enhanced metastatic cell behaviour of PC-3M cells,focusing upon adhesion.
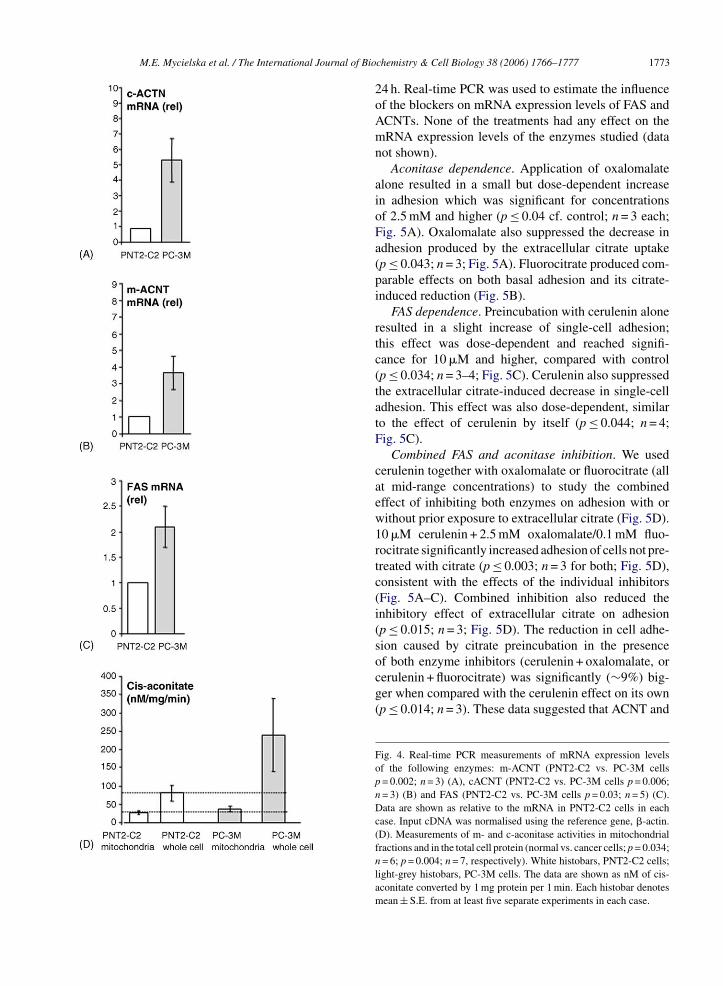
Enzyme expression and activities. mRNA levels of
c- and m-ACNTs in PNT2-C2 and PC-3M cells weremeasured by real-time PCR and found to be 533 ± 142and 366 ± 99% higher in PC-3M cells compared withPNT2-C2 cells (Fig. 4A and B). These differenceswere statistically significant (p = 0.002–0.006; n = 3).Fig. 2. Release of endogenous and exogenous citrate from PNT2-C2and PC-3M cell monolayers detected by NADH fluorescence. Thedecrease in the NADH fluorescence was proportional to the amountof citrate released. The data points were calculated as explained inMaterials and Methods. (A) Dark diamonds (line-1) show measure-ments from PNT2-C2 cells. Grey triangles (line-2) are data from cellspre-incubated in 10 mM Na+-citrate for 24 h. The final, steady-statevalue attained in (1) was treated as 100% and used to calculate therelative values of fluorescence (“F”) in all other measurements (A–D).(B) As in (A) for PC-3M cells. Dark diamonds (line-1) show measure-ments from PC-3M cells. Grey triangles (line-2) are data from cellspre-incubated in 10 mM Na+-citrate for 24 h. (C) As in (A) and (B)but for MCF-10A cells. Dark diamonds (line-1) show measurementsfrom MCF-10A; light grey triangles (line-2) are data from cells pre-incubated in 10 mM Na+-citrate for 24 h. (D). Averaged data for citraterelease from control PNT2-C2 cells (white histobar), control PC-3Mcells (light grey histobar), PNT2-C2 cells preincubated in 10 mM cit-rate (striped histobar) and PC-3M cells preincubated in 10 mM citrate(dark histobar). Each histobar denotes mean ± S.E. (n > 10).
1772 M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777
As regards FAS, the mRNA level was 210 ± 40%higher in PC-3M compared with PNT2-C2 cells(p = 0.03; n = 5; Fig. 4C). Biochemical measurementsshowed that m- and c-ACNT activities were 39 ± 15.2and 65 ± 25.2% higher in PC-3M than in PNT2-C2 cells, respectively (Fig. 4D). In the case of m-ACNT, the activities were 22 ± 4.1 nM/mg/min forPNT2-C2 and 37 ± 7.5 nM/mg/min for PC-3M cells(p = 0.034; n = 6 for both). The total cellular ACNTactivities (which would include combined m- and c-ACNT) were 80 ± 22.3 nM/mg/min (PNT2-C2 cells)and 239 ± 100.1 nM/mg/min (PC-3M cells) (p = 0.004;n = 7 for both). It was concluded that FAS expression andACNT expression and activity were significantly greaterin the strongly metastatic cells.
In order to evaluate the involvement of ACNTand FAS in the metabolic enhancement of metastaticbehaviour of PC-3M cells by extracellular citrate, weused 0.1 and 0.5 mM fluorocitrate and 1–5 mM oxalo-malate as ACNT inhibitors (Festa, Colonna, Pietropaolo,& Ruffo, 2000; Lian & Stringer, 2004) and 5–12.5 �Mcerulenin as an inhibitor of FAS (Schmid, Rippmann,Tadayyon, & Hamilton, 2005). It was not possibleto pharmacologically distinguish between m- and c-ACNTs. Adhesion was used as a convenient measure ofmetastatic cell behaviour. The cells were preincubatedwith a blocker on its own or the blocker with 10 mM cit-rate. The results were compared with control cells withor without citrate preincubation. All treatments lasted
Fig. 3. Effects of exogenous citrate on metastatic cell behaviours mea-sured in three different functional assays. (A) Adhesion quantified as“detachment negative pressure relative to cell surface area” (DNPR).Influence of different concentrations of citrate on DNPR. White histo-bar, PNT2-C2 cells in control conditions; striped histobar representsdata for PNT2-C2 after 24 h of 10 mM citrate pre-treatment (p > 0.05cf. untreated cells; n = 3). PC-3M data are shown as light grey histobar(control) and dark histobars (adhesion after 2 h, h of pre-treatment in10 (p < 0.001; n = 4), 5 (p < 0.001; n = 4), 1 (p = 0.03; n = 4) and 0.1 mM(p = 0.048; n = 6)). (B & C) Motility (wound-heal data) presented as“motility index” (MoI). (B). White histobars denote the data obtainedfor PNT2-C2 cells in the control conditions after 24 and 48 h; light greyhistobars show comparable data for PC-3M cells (p < 0.001; n = 13 forboth PNT2-C2 cells vs. PC-3M). (C). Values of MoI for PNT2-C2cells in control conditions (white histobar) and after 48 h of 1 mMcitrate pretreatement (striped histobar; p = 0.37; n = 3). Similar mea-surements from PC-3M cells are shown as light grey histobar (control)and dark histobar (1 mM citrate pre-treatment); (p = 0.01 cf. control;n = 3). (D). Endocytic membrane activity assay. Membrane endocy-tosis was quantified by spectrometry as “Abs550”. Effects of 10 mMcitrate preincubation on PNT2-C2 cells (control – white; pre-treated– striped; p = 0.26 cf. control; n = 3) and PC-3M cells (control – lightgrey; pre-treated – dark; p = 0.038 cf. control; n = 3) histobars. Valuesof Abs550 were expressed relative to control PNT2-C2 cell data. Eachhistobar denotes mean ± S.E. from at least 3 experiments in each case.
M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777 1773
24 h. Real-time PCR was used to estimate the influenceof the blockers on mRNA expression levels of FAS andACNTs. None of the treatments had any effect on themRNA expression levels of the enzymes studied (datanot shown).
Aconitase dependence. Application of oxalomalatealone resulted in a small but dose-dependent increasein adhesion which was significant for concentrationsof 2.5 mM and higher (p ≤ 0.04 cf. control; n = 3 each;Fig. 5A). Oxalomalate also suppressed the decrease inadhesion produced by the extracellular citrate uptake(p ≤ 0.043; n = 3; Fig. 5A). Fluorocitrate produced com-parable effects on both basal adhesion and its citrate-induced reduction (Fig. 5B).
FAS dependence. Preincubation with cerulenin aloneresulted in a slight increase of single-cell adhesion;this effect was dose-dependent and reached signifi-cance for 10 �M and higher, compared with control(p ≤ 0.034; n = 3–4; Fig. 5C). Cerulenin also suppressedthe extracellular citrate-induced decrease in single-celladhesion. This effect was also dose-dependent, similarto the effect of cerulenin by itself (p ≤ 0.044; n = 4;Fig. 5C).
Combined FAS and aconitase inhibition. We usedcerulenin together with oxalomalate or fluorocitrate (allat mid-range concentrations) to study the combinedeffect of inhibiting both enzymes on adhesion with orwithout prior exposure to extracellular citrate (Fig. 5D).10 �M cerulenin + 2.5 mM oxalomalate/0.1 mM fluo-rocitrate significantly increased adhesion of cells not pre-
treated with citrate (p ≤ 0.003; n = 3 for both; Fig. 5D),consistent with the effects of the individual inhibitors(Fig. 5A–C). Combined inhibition also reduced theinhibitory effect of extracellular citrate on adhesion(p ≤ 0.015; n = 3; Fig. 5D). The reduction in cell adhe-sion caused by citrate preincubation in the presenceof both enzyme inhibitors (cerulenin + oxalomalate, orcerulenin + fluorocitrate) was significantly (∼9%) big-ger when compared with the cerulenin effect on its own(p ≤ 0.014; n = 3). These data suggested that ACNT andFig. 4. Real-time PCR measurements of mRNA expression levelsof the following enzymes: m-ACNT (PNT2-C2 vs. PC-3M cellsp = 0.002; n = 3) (A), cACNT (PNT2-C2 vs. PC-3M cells p = 0.006;n = 3) (B) and FAS (PNT2-C2 vs. PC-3M cells p = 0.03; n = 5) (C).Data are shown as relative to the mRNA in PNT2-C2 cells in eachcase. Input cDNA was normalised using the reference gene, �-actin.(D). Measurements of m- and c-aconitase activities in mitochondrialfractions and in the total cell protein (normal vs. cancer cells; p = 0.034;n = 6; p = 0.004; n = 7, respectively). White histobars, PNT2-C2 cells;light-grey histobars, PC-3M cells. The data are shown as nM of cis-aconitate converted by 1 mg protein per 1 min. Each histobar denotesmean ± S.E. from at least five separate experiments in each case.

1774 M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777
FAS activities were both involved in the usage of extra-cellular citrate.
4. Discussion
Previous work on prostate cells has raised many ques-tions regarding the handling of citrate by epithelial cellsand the changes that occur during the process of metas-
tasis. The levels of citrate in prostatic fluids of normaland cancerous glands have been known for some time(Costello et al., 1999). Here, we used PNT2-C2 andPC-3M cells as models of normal and metastatic pro-static epithelial cells, respectively. Our results showedthe following: (1) Prostate cancer cells contained ∼40%less total intracellular citrate than normal epithelial cells.This difference was smaller (∼20%) when free intracel-lular citrate was compared. (2) Although both normal andcancer cells could uptake citrate, the intracellular level(total and free) remained higher in the former; citratetaken up by cancer cells was supplying mainly the ‘freepool’. (3) Both cell lines could release citrate but, again,this was much bigger in the case of normal cells. (4)Unlike normal cells, cancer cells were able to metaboliseand use exogenous citrate for enhancing metastaticcell behaviours. Thus, extracellular (and endogenous)
Fig. 5. Effects of ACNT and FAS inhibitors on PC-3M single-cell adhesion measurements (DNPR values). The basic pre-incubation/treatment conditions are summarised in the key at the bot-tom of the figure. These and their combinations are indicated belowindividual histobars for given experiments. (A) Influence of differentconcentrations of oxalomalate (3a–c) and 10 mM citrate (2). Light-grey histobars (related to left-hand axis) show basal (control) adhesiondata and effects of 1, 2.5 and 5 mM oxalomalate (3a–c). This was sig-nificant for concentrations of 2.5 mM and higher (p ≤ 0.04 cf. control;n = 3 each). Black histobars (related to right-hand axis) show percent-age changes in citrate-induced reduction in cell adhesion produced bydifferent concentrations of oxalomalate (p ≤ 0.043; n = 3; 3a–c + 2).(B) As in (A) but for fluorocitrate used at 0.1 and 0.5 mM (3a–b).
Application of fluorocitrate increased adhesion compared with con-trol conditions at 0.5 mM (p = 0.045; n = 3). Black histobars (related toright-hand axis) show percentage changes in citrate-induced reductionin cell adhesion produced by different concentrations of fluorocitrate(p < 0.001; n = 3 for both concentrations; 4a–b + 2). (C) As in (A) and(B) but for cerulenin (5, 10 and 12.5 �M; 5a–c). Application of ceru-lenin increased adhesion significantly for 10 �M and higher, comparedwith control conditions (p ≤ 0.034; n = 3–4). Black histobars (relatedto right-hand axis) show percentage changes in citrate-induced reduc-tion in cell adhesion produced by different concentrations of cerulenin(p = 0.045–0.005; n = 4 for all concentrations; 5a–c + 2). (D). Com-parison of the combined effect of blocking FAS and ACNT with theindividual effects of ACNT and FAS inhibitors. Data are plotted asin (A)–(C). Light-grey histobars (left-hand scale) represent DNPRvalues in control conditions, and following treatment with 10 �Mcerulenin (5b), 2.5 mM oxalomalate (3b), 0.1 mM fluorocitrate (4a),2.5 mM oxalomalate + 10 �M cerulenin (3b + 5b), and 0.1 mM flu-orocitrate + 10 �M cerulenin (4a + 5b) (p ≤ 0.003; n = 3 for 3b + 5band 4a + 5b respectively). Black histobars (right-hand scale) representchanges in citrate-induced reduction of cell adhesion in the presenceof citrate together with 10 �M cerulenin (5b + 2), 2.5 mM oxalomlate (3b + 2), 0.1 mM fluorocitrate (4a + 2) (statistics as in A, B and C). Thetwo last dark histobars show changes in citrate-induced reduction ofcell adhesion in the presence of combined 10 �M cerulenin + 2.5 mMoxalomlate (3b + 5b + 2) and 10 �M cerulenin + 0.1 mM fluorocitrate(3b + 4a + 2) (p ≤ 0.015; n = 3). Each histobar denotes mean ± S.E.from at least three separate experiments in each case.
M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777 1775
citrate produced cellular effects that were consistentwith enhanced metastatic behaviour – increased motil-ity (Fraser et al., 2003), increased membrane endocytosis(Mycielska et al., 2003) and decreased adhesion (Palmeret al., submitted). (5) ACNT and FAS expression/activitywere higher in prostate cancer compared with normalcells. (6) The enhancement of metastatic cell behavioursinduced by incubation in extracellular citrate in the for-mer involved FAS and ACNT.
4.1. Uptake and release of citrate
Overall, the total intracellular citrate content of PC-3M cells was ‘static’. Compared with PNT2-C2, PC-3Mcells had a low level of intracellular citrate and releasedlittle of it under all conditions studied (resting and citrate-preincubated). These results would confirm earlier sug-gestions that normal prostate epithelial cells would have“high intracellular citrate” whilst prostate cancer cellswould have “low intracellular citrate” (Kavanagh, 1994;Hochachka, Rupert, Goldenberg, Gleave, & Kozlowski,2002). Even after long-term preincubation in extracel-lular citrate, ∼70% more intracellular citrate was foundin PNT2-C2 cells compared with PC-3M cells. This dif-ference was reflected also in release of citrate from thecells. Although there was some release of endogenouscitrate from PC-3M cells, this again was much smallerthan the release from PNT2-C2 cells (Mycielska et al.,2005b). The free: bound citrate ratio in these two celllines was also different. Whilst normal cells had a ratiooet
rhmiwcNf
4a
pflwwm
side effects. In order to minimize such a possibility, theconcentration of citrate used was restricted to 1 mM inmost of the functional assays, particularly migration. Inany case, any chelation of Ca2+ would produce an oppo-site effect to the observed increase in motility (Tran,Hinman, Unger, & Sammak, 1999). The same would betrue for endocytosis which we have shown previously inprostate cancer cells to be Ca2+-dependent (Mycielskaet al., 2003). It is possible, therefore, that the effectof extracellular citrate in potentiating metastatic cellbehaviours, in fact, might have been underestimated.The effect of citrate on adhesion was dose-dependent,a significant change occurring at the level of 0.1 mM.We chose 10 mM citrate to study ACNT and FASinvolvement in citrate metabolism due to the larger andfaster effects that could be produced. Even at that con-centration, however, after 24 h preincubation, there wasno change in ACNT or FAS expression. Furthermore,the activities of the normal cells pre-incubated in 10 mMcitrate were not affected. We can assume, therefore, thatalthough some chelation might have occurred, it wasunlikely that this was significant in the citrate-inducedfunctional effects.
Metastatic cell behaviours in vitro. Pre-incubationin various concentrations of extracellular citrate didnot produce any changes in migration, proliferation,adhesion and secretory membrane activity of normalcells. This agrees with similar studies on chick embry-onic endothelial cells where citrate (and other oxidativemetabolites of comparable concentrations) were ineffec-
f ∼1:4, for cancer cells, this was higher: ∼1:2. Inter-stingly, citrate taken up by cancer cells supplied mainlyhe pool of free citrate.
The phenomenon of lower amount of intracellular cit-ate found in cancer cells may be due to these cells: (1)aving a lower ability to uptake and/or store citrate or (2)etabolizing taken-up citrate more efficiently in energiz-
ng their metastatic cell behaviours. The first possibilityould seem unlikely in the first instance since prostate
ancer cells were found earlier to functionally express aa+-dependent citrate inward transporter which should
acilitate uptake (Mycielska et al., 2005a).
.2. Extracellular citrate enhanced metastaticctivities selectively of PC-3M cells
Possible effects of chelation. In order to assess theossible influence of the high citrate level in prostaticuid (up to 180 mM) on prostate cells, functional studieshich involved ‘long-term’ preincubation in citrateere performed. However, as a trivalent anion, citrateight cause some chelation of cations and produce
tive in producing any change in migration or proliferation(Murray & Wilson, 2001). On the other hand, extracel-lular citrate enhanced the various metastatic behavioursof prostate cancer cells. The apparent insensitivity ofnormal cells to extracellular citrate is consistent withtheir characteristic low-level expression/activity of keymetabolic enzymes (Fig. 4) and cellular activity (Fig. 3).
4.3. Citrate - induced enhancement of metastaticcell behaviours was dependent on FAS and ACNTactivities
Aconitase. m-ACNT has been shown previously tobe a rate-limiting enzyme in Krebs cycle and is respon-sible for the high levels of citrate in normal prostaticfluid (Costello & Franklin, 1981). It has been suggestedthat this changes in prostate cancer where transforma-tion of prostate epithelial cells to a citrate-oxidizingstate increases the energy production capability and thisis essential to the process of malignancy and metasta-sis (Costello & Franklin, 1994). This agrees with ourdata showing that m-ACNT mRNA expression level

1776 M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777
and activity were significantly higher in the stronglymetastatic prostate cancer (PC-3M) cell line, comparedto normal epithelial prostate PNT2-C2 cells. Thus, cit-rate metabolism would be low and intracellular citratewould be high in normal cells, as our results demon-strated (Fig. 1A).
Significantly higher c-ACNT mRNA expression andenzyme activity were also observed in prostate cancercells. Although the precise physiological function(s) ofc-ACNT has not yet been clearly defined (Narahari,Ma, Wang, & Walden, 2000), the following observationswould suggest that it might play a role in metabolismof citrate in cancer cells: First, in PC-3M comparedwith PNT2-C2 cells, the activity of c-ACNT was ∼4times higher, whilst m-ACNT was higher by only ∼20%(Fig. 4D). Second, in PC-3M cells, c-ACNT versus m-ACNT activity was ∼6-fold higher. Furthermore, sincetransport through mitochondrial membranes is strictlyregulated (Murray & Wilson, 2001), oxalomalate andfluorocitrate, as well as citrate, would be less likely toaffect m-ACNT. Thus, the effects of ACNT inhibitionmeasured (Fig. 5A and B) would be more likely to reflectthe involvement of c-ACNT.
Fatty acid synthase. FAS, which is highly expressedin prostate cancer (Baron et al., 2004), catalyzes the syn-thesis of fatty acids from acetyl-CoA (which is producedby ATP citric lyase from citrate) and plays an impor-tant role in energy homeostasis by converting excesscarbon intake into fatty acids for storage. When nec-essary, fatty acids can provide energy via �-oxidation
hence, increased energy production might be ensuredthrough fatty acid synthesis and regulation of redox bal-ance in cells (Hochachka et al., 2002). Furthermore, ofthe metabolic substrates present in blood, citrate has thehighest concentration (Inoue, Zhuang, Maddox, Smith,& Ganapathy, 2002) and this and citrate in prostaticfluid may be used to promote metastatic cell behaviour.Again, as in the present study, increase in citrate-relatedmetabolic enzyme expression and activity has beenobserved in many different types of cancer (Huang, Zhu,Lu, Dai, & Jin, 2000; Menendez, Lupu, & Colmer, 2005).It is possible, therefore, that the phenomenon describedhere might be more widespread, involve additional com-ponents like fatty acid-binding proteins (Adamson et al.,2003) and thus form the basis of a more general mecha-nism in cancer.
Acknowledgements
This study was supported by a project grant and aVIP Award from the Wellcome Trust. We thank Prof.Norman Maitland (supported by the Yorkshire Can-cer Research) for the initial supply of the PNT2-C2cell line. Further support was provided by The Phys-iological Society (Vacation Studentship to RB) andThe British Council, TRNC (Chevening Scholarship toKE).
References
(Kuhajda et al., 2000). Cerulenin, chosen to inhibit FAS,has been shown previously to decrease fatty acid syn-thesis in several different systems, including prostatecancer, at concentrations similar to these used in thepresent study (Kuhajda et al., 2000). Inhibition of FASresulted in a significant and dose-dependent decrease inthe citrate-induced potentiation of prostate cancer cellactivity (Fig. 5C). This would suggest involvement ofFAS in the metabolism of extracellular citrate. In fact,the results of basal FAS (as well as ACNT) inhibition(Fig. 5A–C) are consistent with these enzymes havinga similar role in relation to endogenous citrate. Interest-ingly, combined FAS + ACNT inhibition of the citrate-induced effects was slightly but significantly greater thanFAS inhibition alone. This would suggest that ACNTand FAS were both involved and could be partially inter-dependent.
4.4. Concluding remarks
It has been known that cancer cells often are deprivedof oxygen and their increased demand for energy, and
Adamson, J., Morgan, E. A., Beesley, C., Mei, Y., Foster, C. S., Fujii,H., et al. (2003). High-level expression of cutaneous fatty acid-binding protein in prostatic carcinomas and its effect on tumori-genicity. Oncogene, 22, 2739–2749.
Baron, A., Migita, T., Tang, D., & Loda, M. (2004). Fatty acid syn-thase: a metabolic oncogene in prostate cancer? Journal of CellBiochemistry, 91, 47–53.
Costello, L. C., & Franklin, R. B. (1981). Aconitase activity, citrateoxidation, and zinc inhibition in rat ventral prostate. Enzyme, 26,281–287.
Costello, L. C., & Franklin, R. B. (1991a). Concepts of citrate produc-tion and secretion by prostate. 1. Metabolic relationships. Prostate,18, 25–46.
Costello, L. C., & Franklin, R. B. (1991b). Concepts of citrate produc-tion and secretion by prostate: 2. Hormonal relationships in normaland neoplastic prostate. Prostate, 19, 181–205.
Costello, L. C., & Franklin, R. B. (1994). Bioenergetic theory ofprostate malignancy. Prostate, 25, 162–166.
Costello, L. C., Franklin, R. B., & Narayan, P. (1999). Citrate in thediagnosis of prostate cancer. Prostate, 38, 237–245.
Costello, L. C., Franklin, R. B., Liu, Y., & Kennedy, M. C. (2000). Zinccauses a shift toward citrate at equilibrium of the m-aconitase reac-tion of prostate mitochondria. Journal of Inorganic Biochemistry,78, 161–165.
Diss, J. K., Stewart, D., Pani, F., Foster, C. S., Walker, M. M., Patel, A.,et al. (2005). A potential novel marker for human prostate cancer:

M.E. Mycielska et al. / The International Journal of Biochemistry & Cell Biology 38 (2006) 1766–1777 1777
Voltage-gated sodium channel expression in vivo. Prostate Cancer& Prostatic Diseases, 8, 266–273.
Drapier, J. C., & Hibbs, J. B., Jr. (1996). Aconitases: A class of met-alloproteins highly sensitive to nitric oxide synthesis. Methods inEnzymology, 269, 26–36.
Festa, M., Colonna, A., Pietropaolo, C., & Ruffo, A. (2000). Oxalo-malate, a competitive inhibitor of aconitase, modulates the RNA-binding activity of iron-regulatory proteins. Biochemical Journal,348, 315–320.
Fraser, S. P., Ding, Y., Liu, A., Foster, C. S., & Djamgoz, M. B. A.(1999). Tetrodotoxin suppresses morphological enhancement ofthe metastatic MAT-LyLu rat prostate cancer cell line. Cell & TissueResearch, 295, 505–512.
Fraser, S. P., Salvador, V., Manning, E. A., Mizal, J., Altun, S., Raza,M., et al. (2003). Contribution of functional voltage-gated Na+
channel expression to cell behaviors involved in the metastaticcascade in rat prostate cancer: I. Lateral motility. Journal of CellPhysiology, 195, 479–487.
Grimes, J. A., Fraser, S. P., Stephens, G. J., Downing, J. E., Lani-ado, M. E., Foster, C. S., et al. (1995). Differential expression ofvoltage-activated Na+ currents in two prostatic tumour cell lines:Contribution to invasiveness in vitro. FEBS Letters, 369, 290–294.
Hering-Smith, K. S., Gambala, C. T., & Hamm, L. L. (2000). Citrateand succinate transport in proximal tubule cells. American Journalof Physiology – Renal Physiolgy, 278, F492–F498.
Hochachka, P. W., Rupert, J. L., Goldenberg, L., Gleave, M., &Kozlowski, P. (2002). Going malignant: The hypoxia-cancer con-nection in the prostate. BioEssays, 24, 749–757.
Huang, P. L., Zhu, S. N., Lu, S. L., Dai, Z. S., & Jin, Y. L. (2000).Inhibitor of fatty acid synthase induced apoptosis in human coloniccancer cells. World Journal of Gastroenterology, 6, 295–297.
Inoue, K., Zhuang, L., Maddox, D. M., Smith, S. B., & Ganapathy,V. (2002). Structure, function, and expression pattern of a novelsodium-coupled citrate transporter (NaCT) cloned from mam-malian brain. Journal of Biological Chemistry, 277, 39469–39476.
K
K
L
L
M
M
M
expression to cell behaviors involved in the metastatic cascade inrat prostate cancer: II. Secretory membrane activity. Journal of CellPhysiology, 195, 461–469.
Mycielska, M. E., & Djamgoz, M. B. (2004). Citrate transport in thehuman prostate epithelial PNT2-C2 cell line: Electrophysiologicalanalyses. Journal of Physiology, 559, 821–833.
Mycielska, M. E., Palmer, C. P., Brackenbury, W. J., & Djamgoz,M. B. A. (2005). Expression of Na+-dependent citrate transportin a strongly metastatic human prostate cancer PC-3M cell line:Regulation by voltage-gated Na+ channel activity. Journal of Phys-iology, 563, 393–408.
Mycielska, M. E., Krasowska, M., Grzywna, Z., & Djamgoz, M. B.(2005). Endogenous and exogenous citrate transport and release inprostatic preparations: Semi-polarized two-dimensional culturesof human PNT2-C2 cells and isolated tubules and segments of ratprostate. Prostate, 65, 88–99.
Narahari, J., Ma, R., Wang, M., & Walden, W. E. (2000). The aconi-tase function of iron regulatory protein 1. Genetic studies in yeastimplicate its role in iron-mediated redox regulation. Journal ofBiological Chemistry, 275, 16227–16234.
Onganer, P. U., & Djamgoz, M. B. (2005). Small-cell lung cancer(human): Potentiation of endocytic membrane activity by voltage-gated Na+ channel expression in vitro. Journal of Membrane Biol-ogy, 204, 67–75.
Palmer, C. P., Mycielska, M. E., Burcu, H., Osman, K., Collins, T.,Beckerman, R., Perrett, R., Aydar, E. & Djamgoz, M. B. A. (inpreparation). A micro-pressure system for measuring single-celladhesion: Application to cancer cell lines of different metastaticpotential and voltage-gated Na+ channel expression.
Petrarulo, M., Facchini, P., Cerelli, E., Marangella, M., & Linari, F.(1995). Citrate in urine determined with a new citrate lyase method.Clinical Chemistry, 41, 1518–1521.
Pitula, J. S., Deck, K. M., Clarke, S. L., Anderson, S. A., Vasanthaku-mar, A., & Eisenstein, R. S. (2004). Selective inhibition of thecitrate-to-isocitrate reaction of cytosolic aconitase by phospho-
avanagh, J. P. (1994). Isocitric and citric acid in human prostatic andseminal fluid: Implications for prostatic metabolism and secretion.Prostate, 24, 139–142.
uhajda, F. P., Pizer, E. S., Li, J. N., Mani, N. S., Frehywot, G. L.,& Towsend, C. A. (2000). Synthesis and antitumor activity of aninhibitor of fatty acid synthase. Proceedings of National Academyof Sciences USA, 97, 3450–3454.
ian, X. Y., & Stringer, J. L. (2004). Inhibition of aconitase in astro-cytes increases the sensitivity to chemical convulsants. EpilepsyResearch, 60, 41–52.
ivak, K. J., & Schmittgen, T. D. (2001). Analysis of relativegene expression data using real-time quantitative PCR and the2(−��C(T)). Methods, 25, 402–408.
enendez, J. A., Lupu, R., & Colmer, R. (2005). Targeting fattyacid synthase: Potential for therapeutic intervention in her-2/neu-overexpressing breast cancer. Drugs News & Perspectives, 18,375–385.
urray, B., & Wilson, D. J. (2001). A study of metabolites as inter-mediate effectors in angiogenesis. Angiogenesis, 4, 71–77.
ycielska, M. E., Fraser, S. P., Szatkowski, M., & Djamgoz, M.B. (2003). Contribution of functional voltage-gated Na+ channel
mimetic mutation of serine-711. Proceedings of National Academyof Science U S A, 101, 10907–10912.
Schmid, B., Rippmann, J. F., Tadayyon, M., & Hamilton, B. S. (2005).Inhibition of fatty acid synthase prevents preadipocyte differentia-tion. Biochemical & Biophysical Research Communications, 328,1073–1082.
Silletti, S., Yao, J. P., Pienta, K. J., & Raz, A. (1995). Loss of cell-contact regulation and altered responses to autocrine motility factorcorrelate with increased malignancy in prostate cancer cells. Inter-national Journal of Cancer, 63, 100–105.
Swinnen, J. V., Roskams, T., Joniau, S., Van Poppel, H., Oyen, R.,Baert, L., et al. (2002). Overexpression of fatty acid synthase isan early and common event in the development of prostate cancer.International Journal of Cancer, 98, 19–22.
Tran, P. O., Hinman, L. E., Unger, G. M., & Sammak, P. J. (1999). Awound-induced [Ca2+]i increase and its transcriptional activationof immediate early genes is important in the regulation of motility.Experimental Cell Research, 246, 319–326.
Tsui, K. H., Chang, P. L., & Juang, H. H. (2006). Zinc blocks geneexpression of mitochondrial aconitase in human prostatic carci-noma cells. International Journal of Cancer, 118, 609–615.




















