
Chemotypical variations in Withania somnifera lead to differentially modulated immune response in...
11
Vaccine 30 (2012) 1083–1093 Contents lists available at SciVerse ScienceDirect Vaccine jou rn al h om epa ge: www.elsevier.com/locate/vaccine Chemotypical variations in Withania somnifera lead to differentially modulated immune response in BALB/c mice Susheela Kushwaha a,1 , Saptarshi Roy b,1 , Rita Maity b,1 , Asish Mallick b , Vishal K. Soni a , Prashant K. Singh a , Narayan D. Chaurasiya c , Rajender S. Sangwan c , Shailja Misra-Bhattacharya a,∗ , Chitra Mandal b,∗∗ a Division of Parasitology, Council of Scientific and Industrial Research (CSIR) – Central Drug Research Institute (CDRI), M.G. Marg, P.O. Box 173, Lucknow 226001, India b Cancer and Cell biology Division, CSIR – Indian Institute of Chemical Biology (IICB), 4, Raja S.C. Mullick Road, Kolkata 700032, India c Metabolic and Structural Biology Division, CSIR – Central Institute of Medicinal and Aromatic Plants (CIMAP), P.O. – CIMAP, Lucknow 226015, India a r t i c l e i n f o Article history: Received 10 June 2011 Received in revised form 30 November 2011 Accepted 5 December 2011 Available online 17 December 2011 Keywords: Chemotypes Immunomodulation Th1/Th2 cytokines Withania somnifera a b s t r a c t Withania somnifera (Ashwagandha) is a plant with known ethnomedicinal properties and its use in Ayurvedic medicine in India is well documented. The present investigation reports on immunomod- ulatory efficacy of aqueous-ethanol extracts of roots of three selected Withania somnifera chemotypes designated as NMITLI 101R, NMITLI 118R and NMITLI 128R. Each chemotype was administered 10–100 mg/kg orally to BALB/c mice once daily for 14 days. The immunomodulatory consequences were recorded by determining the humoral immune response with the help of hemagglutination, plaque form- ing cell assay and cellular response by measuring delayed type hypersensitivity reaction. Additionally, other immune parameters such as proliferation of T and B cells, intracellular and secreted Th1 and Th2 cytokines along with modulation in ROS production by peritoneal macrophages were monitored after feeding with lower doses (3–30 mg/kg/day) of these three chemotypes individually. NMITLI 101R incited both humoral and cellular immune response in terms of higher number of antibody producing cells and enhanced foot pad swelling at the 10 mg dose as also dose dependent B and T cell proliferations. Lev- els of intracellular and secreted cytokines post-NMITLI 101R treatment illustrated generation of mixed Th1/Th2 response that remained more polarized towards Th1. This chemotype also generated maximum reactive oxygen species. NMITLI 118R provoked comparatively reduced immune response in all humoral and cellular parameters at lower doses but induced highly polarized Th1 cytokine response. In con- trast, NMITLI 128R led to enhanced antibody production with minimal cellular response demonstrating marginally Th2 dominance at a lower dose. Taken together, it may therefore be concluded that there were distinct modulation in the immune response exhibited by the three chemotypes of Withania somnifera and NMITLI 101R appeared to possess a better immunostimulatory activity than the other chemotypes at lower doses. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction Herbal or semisynthetic compounds derived from plants are in the forefront as the candidate of future generation drugs. Ayurveda, one of the oldest medical prescriptions, had described different Abbreviations: DTH, delayed type hypersensitivity; FCS, fetal calf serum; HA titre, haemagglutination titre; PFC, plaque forming cells; SRBC, sheep red blood cells; WS, Withania somnifera; DCF-DA, 2,7- dichlorofluorescin diacetate. ∗ Corresponding author. Tel.: +91 522 2612411–18x4224, 4221; fax: +91 522 2623938/2623405. ∗∗ Corresponding author. Tel.: +91 33 2429 8861; fax: +91 33 2473 5197. E-mail addresses: shailja [email protected], shailja [email protected] (S. Misra-Bhattacharya), [email protected], chitra [email protected] (C. Mandal). 1 These authors should be considered as joint first authors. plant-based herbal products with properties of restraining many diseases by modulation of immune responses [1]. Immunomodu- lators derived from medicinal plants are critically being considered as an alternative and/or adjunct approach to conventional treat- ment against various disease. Extracts or molecules derived from plants or by chemical synthesis based on leads obtained from nat- ural products are opening new vistas for modern therapeutics by positively modulating the immune system against infections. In view of the relatively high cost and considerable side effects of allo- pathic medicines, researchers are now focussing on re-examining the traditional formulations used in Ayurveda and Homeopathy possessing minimal side effects [2]. Withania somnifera (WS), commonly known as Ashwagandha, Indian Ginseng or Winter cherry has often been compared with the Chinese Ginseng (Phanax gingeng) due to its wide array of health 0264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2011.12.031
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Chemotypical variations in Withania somnifera lead to differentially modulated immune response in...

Ci
SPCa
b
c
a
ARR3AA
KCITW
1
to
hW
f
(
0d
Vaccine 30 (2012) 1083– 1093
Contents lists available at SciVerse ScienceDirect
Vaccine
jou rn al h om epa ge: www.elsev ier .com/ locate /vacc ine
hemotypical variations in Withania somnifera lead to differentially modulatedmmune response in BALB/c mice
usheela Kushwahaa,1, Saptarshi Royb,1, Rita Maityb,1, Asish Mallickb, Vishal K. Sonia,rashant K. Singha, Narayan D. Chaurasiyac, Rajender S. Sangwanc, Shailja Misra-Bhattacharyaa,∗,hitra Mandalb,∗∗
Division of Parasitology, Council of Scientific and Industrial Research (CSIR) – Central Drug Research Institute (CDRI), M.G. Marg, P.O. Box 173, Lucknow 226001, IndiaCancer and Cell biology Division, CSIR – Indian Institute of Chemical Biology (IICB), 4, Raja S.C. Mullick Road, Kolkata 700032, IndiaMetabolic and Structural Biology Division, CSIR – Central Institute of Medicinal and Aromatic Plants (CIMAP), P.O. – CIMAP, Lucknow 226015, India
r t i c l e i n f o
rticle history:eceived 10 June 2011eceived in revised form0 November 2011ccepted 5 December 2011vailable online 17 December 2011
eywords:hemotypes
mmunomodulationh1/Th2 cytokinesithania somnifera
a b s t r a c t
Withania somnifera (Ashwagandha) is a plant with known ethnomedicinal properties and its use inAyurvedic medicine in India is well documented. The present investigation reports on immunomod-ulatory efficacy of aqueous-ethanol extracts of roots of three selected Withania somnifera chemotypesdesignated as NMITLI 101R, NMITLI 118R and NMITLI 128R. Each chemotype was administered10–100 mg/kg orally to BALB/c mice once daily for 14 days. The immunomodulatory consequences wererecorded by determining the humoral immune response with the help of hemagglutination, plaque form-ing cell assay and cellular response by measuring delayed type hypersensitivity reaction. Additionally,other immune parameters such as proliferation of T and B cells, intracellular and secreted Th1 and Th2cytokines along with modulation in ROS production by peritoneal macrophages were monitored afterfeeding with lower doses (3–30 mg/kg/day) of these three chemotypes individually. NMITLI 101R incitedboth humoral and cellular immune response in terms of higher number of antibody producing cells andenhanced foot pad swelling at the 10 mg dose as also dose dependent B and T cell proliferations. Lev-els of intracellular and secreted cytokines post-NMITLI 101R treatment illustrated generation of mixedTh1/Th2 response that remained more polarized towards Th1. This chemotype also generated maximumreactive oxygen species. NMITLI 118R provoked comparatively reduced immune response in all humoral
and cellular parameters at lower doses but induced highly polarized Th1 cytokine response. In con-trast, NMITLI 128R led to enhanced antibody production with minimal cellular response demonstratingmarginally Th2 dominance at a lower dose. Taken together, it may therefore be concluded that there weredistinct modulation in the immune response exhibited by the three chemotypes of Withania somniferaand NMITLI 101R appeared to possess a better immunostimulatory activity than the other chemotypesat lower doses.. Introduction
Herbal or semisynthetic compounds derived from plants are inhe forefront as the candidate of future generation drugs. Ayurveda,ne of the oldest medical prescriptions, had described different
Abbreviations: DTH, delayed type hypersensitivity; FCS, fetal calf serum; HA titre,aemagglutination titre; PFC, plaque forming cells; SRBC, sheep red blood cells; WS,ithania somnifera; DCF-DA, 2,7- dichlorofluorescin diacetate.∗ Corresponding author. Tel.: +91 522 2612411–18x4224, 4221;
ax: +91 522 2623938/2623405.∗∗ Corresponding author. Tel.: +91 33 2429 8861; fax: +91 33 2473 5197.
E-mail addresses: shailja [email protected], shailja [email protected]. Misra-Bhattacharya), [email protected], chitra [email protected] (C. Mandal).
1 These authors should be considered as joint first authors.
264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2011.12.031
© 2011 Elsevier Ltd. All rights reserved.
plant-based herbal products with properties of restraining manydiseases by modulation of immune responses [1]. Immunomodu-lators derived from medicinal plants are critically being consideredas an alternative and/or adjunct approach to conventional treat-ment against various disease. Extracts or molecules derived fromplants or by chemical synthesis based on leads obtained from nat-ural products are opening new vistas for modern therapeutics bypositively modulating the immune system against infections. Inview of the relatively high cost and considerable side effects of allo-pathic medicines, researchers are now focussing on re-examiningthe traditional formulations used in Ayurveda and Homeopathy
possessing minimal side effects [2].Withania somnifera (WS), commonly known as Ashwagandha,Indian Ginseng or Winter cherry has often been compared with theChinese Ginseng (Phanax gingeng) due to its wide array of health

1 ccine
aomibtwcttbp[nleswtei
cilicortmslrfbigmm
n[WaaclaN[isNatc
2
2
2wr
084 S. Kushwaha et al. / Va
ttributing properties. This Rasayana herb has been recognized asne of the most important medicinal plants in the traditional Indianedical systems for over 3000 years [3]. Considerable morpholog-
cal variations in both inter-habitat and intra-habitat nature haveeen observed in the plant populations of different geographic loca-ions [4]. However, the predominant class of its phytochemicals i.e.ithanolides possessed by the variants in the wild type and the
ultivars appear to be the same [5]. The variations with respecto discrete sets of individual members of withanolide group definehe distinct chemotypes. The plant has long been applied on humaneings as an anti inflammatory [6], chondroprotective [7], cardio-rotective [8], anti cancer [9,10], hypoglycaemic [11], anti oxidant12], hypolipidaemic [12], hepatoprotectant [12], anti stress [13],europrotective [14] or immunomodulatory agent [1]. Pharmaco-
ogical, chemical, immunological and toxicological studies havestablished that root extracts of WS contain several bioactiveteroidal lactone constituents based on an ergostane skeleton calledithanolides [5]. However, differences in composition and concen-
ration of constituents along with little modifications in chemicalntities in various chemotypes may have led to differences inmmunomodulatory properties.
Immunomodulation needs the orchestration of many immuneells with each other through signaling molecules to boost themmune system to fight against infection more efficiently. T-ymphocytes and the antigen presenting cells play a crucial rolen modulating immune response in vivo. The Th1 response washaracterized by IFN� production that induces B cells to producepsonizing antibodies and cell-mediated immunity, while the Th2esponse is characterized by the release of IL-4 that activates B cellso generate humoral immune response [15,16]. Th1 response is
ainly evoked against intracellular pathogens that cause immuneuppression while Th2 response is generated against extra cellu-ar pathogens [17–19]. This polarization of Th1 or Th2 immuneesponse results in establishment or elimination of the diseaserom the system. On the other hand, CD8+ T cells are responsi-le for elimination of pathogen-infected cells [18]. Macrophage
s the prime target of immunomodulators due to its capability ofenerating primary immune response. Macrophage exerts its antiicrobial activity by producing a number of reactive oxygen inter-ediate/species (ROI/ROS) [20,21].WS is one of the most attractive candidates due to its wide recog-
ition as a strong disease combating plant in traditional medicine22]. On the basis of the widespread geographical distribution of
S, chemotypical variations are likely to be present in this plantnd some of these may be genotypic in nature. Therefore, there is
dire need to investigate the pharmacological activity of varioushemotypes of WS. This entails cognate diversity in immunomodu-atory activity and efficacy as a formulation/drug directed towards
particular disease. Among many chemotypes of WS, NMITLI 101,MITLI 118 and NMITLI 128 are the three most important types
23]. It was therefore considered worthwhile to investigate themmunomodulatary activities of extracts of roots of these threeelected chemotypes designated as NMITLI 101R, NMITLI 118R andMITLI 128R. The present study describes the immune modulatingbility of these selected chemotypes with the help of a compara-ive analysis on humoral and cellular immune responses, Th1, Th2ytokines and activation of antigen presenting cells in BALB/c mice.
. Materials and methods
.1. Animals
Inbred female BALB/c mice of the age of 4–6 weeks weighing0–22 g were randomly distributed in groups (n = 5). The animalsere housed under standard conditions of temperature (23 ± 1 ◦C),
elative humidity (55 ± 10%) and 12 h/12 h light/dark cycles at
30 (2012) 1083– 1093
animal houses of both CSIR – Indian Institute of Chemical Biologyand CSIR – Central Drug Research Institute and fed with standardpellet diet and autoclaved water was given ad libitum. All the animalhandling and experimental protocols employed are approved byInstitutional Animal Ethics Committees from both Institutes. Boththe Institutes followed same protocol and each experiment wasindependently carried out at both Institutes under the umbrella ofIndian Government in the auspices of New Millenium Indian Tech-nology Leadership Initiative (NMITLI) project steered by CSIR. Atthe end, the results were validated by monitoring committee.
2.2. Reagents and antibodies
Medium RPMI-1640 containing Phenol Red,antibiotic–antimycotic cocktail (10× solution containing 10,000units penicillin, 10 mg streptomycin and 25 �g amphotericinB/ml), 2,7-dichlorofluorescin diacetate (DCF-DA), lipopolysaccha-ride (LPS) and HEPES were purchased from Sigma Chemical Co., St.Louis, MO, USA. Fetal calf serum (FCS) was from GIBCO, USA. Allantibodies along with antibody set for ELISA were purchased fromBecton and Dickinson (BD) Pharmingin, Mountain View, CA, USA.All other chemicals used were of analytical grade available locally.
2.3. Preparation of extracts of different chemotypes of WS
2.3.1. Source of plant materialA set of discrete chemotypes of WS were initially developed as a
consequence of large scale phytochemical screening of accessionsof the plant collected from almost all across major wild habitatsof India followed by genetic improvement of the selected linesin the identified core collection [23]. Of these, roots of 5 months’plants from three selected chemotypes NMITLI 101, NMITLI 118and NMITLI 128 were harvested in the month of June (summermonth) from their plantation raised (following standard agronomicpractices) at the Experimental Farm of CSIR-Central Institute ofMedicinal and Aromatic Plants at Lucknow 26.5◦N latitude, 85.5◦Elongitude and 120 m above sea level, India and used for preparingthe aqueous-ethanol extract.
2.3.2. Extraction and fractionationThe liquid nitrogen powder of fresh root tissue of each chemo-
type was extracted in two volumes (g/ml) of water:ethanol (75:25,v/v) for 24 h with occasional shaking followed by filtration throughtwo layers of muslin cloth and refiltration of the filtrate througha filter paper. The filtrate was saved and residue was reextractedtwo more times as above. The three filtrates were pooled, concen-trated in a flash evaporator at 50 ◦C followed by complete drynessin a freeze dryer. The lyophilized extract was weighed and analyzedby thin layer chromatography (TLC) and high performance liquidchromatography (HPLC) (Waters, Milford, MA, USA) essentially asreported earlier [24]. The root extracts of these chemotypes weredesignated as NMITLI 101R, NMITLI 118R and NMITLI 128R andimmunomodulatory potentials of these extracts were compared.
2.4. Chemotype doses and immunization
The extracts of all three chemotypes (NMITLI 101R, NMITLI 118Rand NMITLI 128R) suspended in double distilled water were admin-istered orally for 14 consecutive days in BALB/c mice while controlmice received water only. In preliminary studies, chemotypes wereevaluated at 10, 30 and 100 mg/kg body weight doses for theirimmunomodulatory activity using hemagglutination (HA) titre,
plaque forming cell (PFC) assay and delayed type hypersensitiv-ity (DTH) response by measuring footpad thickness. Additionally,extract and vehicle fed mice were immunized with sheep red bloodcells (SRBC) for measuring HA titre and DTH response. Once the
ccine
ie3p
2
2
Aihtasmo1sSbo
2
htht(abdsaTswc
2
oClrSswptt
2
iTp2a9(ta[
S. Kushwaha et al. / Va
mmunomodulatary activity was established by these parameters,xtracts were further explored at lower doses mainly 3, 10, and0 mg/kg body weight to dissect the sub-clones of B, T cells, antigenresenting cells and cytokine production by T cells.
.5. In vitro study
.5.1. Haemagglutination assayHumoral response was assessed in extract and vehicle fed mice.
ntibody production is the hallmark of humoral response. Accord-ngly, antibody level in serum samples was determined by theaemagglutination assay [25]. For this, sheep blood were asep-ically collected from a normal healthy sheep and mixed withlsever’s solution at 1:1 ratio. SRBC was separated by ficoll den-ity gradient centrifugation. The groups of extract and vehicle fedice were intraperitoneally immunized with SRBC (1 × 107/0.1 ml)
n day 9 since the start of oral feeding and blood were drawn on day5 and sera were separated and stored at −70 ◦C until used. In brief,erially diluted immunized sera in saline were mixed with v/v (1%)RBC suspension in U-bottom micro titre plates and overnight incu-ated at room temperature to observe agglutination. The endpointf agglutination was considered as HA titre.
.5.2. Plaque forming cell (PFC) assayAntibody producing cells increases in the blood during elevated
umoral response. Accordingly, PFC assay was used to evaluatehe number of antibody producing cells as another marker ofumoral response. Increase in the number of plaque representshe increase in number of antibody producing cells. Splenocytes1 × 108 cells/ml) were isolated from extract and vehicle fed miceseptically and suspended in RPMI + FCS (10%) medium. Agaroseottom layer (1.2% agarose in PBS) was prepared in plastic petriish (60 mm). Agarose (0.60% agarose in RPMI) containing SRBCuspension and spleen cells were overlaid on agarose layer at 37 ◦Cnd the petri dishes were incubated for 2 h at same temperature.he petridishes were reincubated with freshly collected guinea pigerum (10%) as a source of complement for another 2 h. The plaquesere counted and the values have been expressed as counts per 107
ells [19,26].
.5.3. Delayed type hypersensitivity (DTH) reactionIn parallel, another group of mice were fed to determine the sec-
ndary and more specific response i.e. cellular immune response.ellular response plays a lead role in immunomodulation. This cel-
ular immunity was monitored by evaluating DTH reaction. DTHesponse was monitored in extract and vehicle fed mice [19,27]. TheRBC immunized mice were challenged with SRBC (2 × 108/50 �l)uspensions on day 15 in right-hind footpad intradermally. Salineas injected in the contra lateral paw and the thickness of foot-ad was measured at 24, 48 and 72 h post SRBC challenge by a dialhickness gauge. The difference in the thickness of paw comparedo control was used as a measurement of DTH reaction.
.5.4. Measurement of Th1/Th2 cytokines by ELISACytokines play very important role by boosting different
mmune cells to exhibit immunomodulation. Secretion of Th1 andh2 cytokines were used as one of the major parameter for thisurpose. Splenocytes (5 × 105) from extract and vehicle fed mice in50 �l of complete RPMI-phenol red medium containing FCS (10%)long with antibiotic–antimycotic (1×) cocktail were placed on a6 well culture plate and incubated at 37 ◦C for 48 h by adding LPS
2.5 �g/ml). The culture supernatants (100 �l) were used for quan-ification of released IL-4 and IFN� by ELISA using commerciallyvailable antibodies following the instructions of the manufacturer28].30 (2012) 1083– 1093 1085
2.5.5. Estimation of ROS generation by peritoneal macrophagesMacrophage, the major antigen presenting cell, is a crucial
player in immune activation. ROS generation is the hallmark ofmacrophage activation. Intracellular ROS in peritoneal macrophagecells were determined through a fluorometric assay using2,7-dichlorofluorescin diacetate (DCF-DA) [29]. Briefly, freshly har-vested macrophage cells of both extract and vehicle fed mice wereadjusted to a concentration of 1 × 106 cells/ml in RPMI, washedin PBS thrice and transferred to FACS tube (1 × 106 cells/tube).Cells were incubated with DCF-DA at a final concentration of 1 �Mfor 15 min at 37 ◦C, washed twice in PBS, acquired and analyzedby measuring their fluorescence intensity on flow cytometer asdescribed above.
2.6. In vivo study
2.6.1. Measurements of B and T-lymphocyte sub populationsIn mammalian system, B and T cells are the key cells that are
responsible for modulation of immune response. Therefore it wasworthwhile to explore the status of B and T cells in the spleno-cytes by using specific cell surface markers in flow cytometry.Lymphocyte subset population was assessed using fluorochromeconjugated monoclonal antibodies against CD3, CD4, CD8 and CD19antigens by following the manufacturer’s protocol [19]. In brief,splenocyte suspension was prepared from the spleen of extractand vehicle fed mice on day 15 of the commencement of feed-ing. Cells (1 × 106) were initially blocked with mouse FcR, washedand labeled with FITC-anti-CD3 and PE-anti-CD19 separately for45 min in dark at 4 ◦C. Subsequently, FITC-anti-CD3 labeled cellswere further incubated separately with PE-anti-CD4 and PE-anti-CD8 respectively. Labeled cells were washed with PBS, suspendedin sheath fluid and analyzed by flow cytometer (BD Biosciences,USA) using CellQuestPro analysis software (BD Biosciences, USA)after gating the forward and side scatter settings to exclude debris.Results were reported as percentage of each cell population.
2.6.2. Measurement of intracellular cytokinesThe measurement of intracellular Th1 and Th2 cytokines in the
splenocytes was done by flow cytometry [29]. Briefly, splenocytes(2 × 106) were incubated with FITC-anti-mouse CD4 antibody for45 min and subsequently permeabilized using permeabilizationsolution (Cytofix/Cytoperm) for 10 min in dark. After washing inpermeabilization wash buffer, cells were dispensed into differenttubes and incubated separately with PE-anti-IL-4 and PE-anti-IFN�monoclonal antibody for 30 min at 4 ◦C. Cells were washed andfinally suspended in permeabilization wash buffer and processedsimilarly.
2.7. Statistical analysis
Data have been expressed as mean ± standard error (S.E.) andstatistical analysis was carried out by employing the Student’s t-testand analysis of variance by one way ANOVA (Dunnett’s multiplecomparison test). A conventional p < 0.05 was taken as evidenceof significant differences and p < 0.001 was considered as highlysignificant, while p > 0.05 was considered as not significant.
3. Results
3.1. NMITLI 101R, NMITLI 118R and NMITLI 128R showeddifferential existence of withanolides
The yield of extracts on fresh weight basis were 4.28%, 3.30% and4.08% and dry weight basis were 16.0%, 15.0% and 18.8% for NMITLI101R, NMITLI 118R and NMITLI 128R respectively. Withanolideprofiling of the root extracts of all the three chemotypes of WS was

1086 S. Kushwaha et al. / Vaccine 30 (2012) 1083– 1093
F le of wc (C). W2
c1tapkt1ba21wTe
3r
Masa
HA titre elicited by NMITLI 128R exhibited significant dose depen-dent increase of more than 2 fold in all three doses. On the other
Table 1Active constituents of NMITLI 101R, NMITLI 118R and NMITLI 128R.
Withanolide Withanolide content in root extractof chemotype (aua/mg extract)
NMITLI101R
NMITLI118R
NMITLI128R
Withanolide A 12.36 16.30 14.02Withanone 9.28 19.28 1.50Withaferin A 1.58 2.06 n.d.17-hydroxy withaferin A 7.58 n.d.b n.d.17-hydroxy, 27-deoxy withaferin A 3.02 5.16 n.d.3-OH withanone 1.44 n.d. n.d.
Variation in the signature profile of different active constituents in the chemotypes
ig. 1. HPLC profile. Comparative HPLC (reverse phase column, PDA detector) profihemotypes of Withania somnifera. NMITLI 101R (A), NMITLI 118R (B), NMITLI 128R7-deoxy withaferin A; 3-OH WN, 3-hydroxy withanone.
arried out by HPLC (Fig. 1). NMITLI 101R, NMITLI 118R and NMITLI28R showed different signature profiles in HPLC suggesting eacho be a characteristic mixture of constituent phytochemicals. As
compositional commonality of the extracts withanolide A wasredominatly present in all the three chemotypes (Fig. 1A–C). Theey compositional differences among the extracts included: (i) dis-inguished presence of detectable levels of withaferin A in NMITLI01R (Fig. 1A) and NMITLI 118R (Fig. 1B), (ii) withanone detected toe abundantly present in NMITLI 101R and NMITLI 118R had only
rudimentary presence in NMITLI 128R (Fig. 1C), (iii) 17-hydroxy7-deoxy withaferin A was detected in NMITLI 101R and NMITLI18R while 17-hydroxy withaferin A and 3-hydroxy withanoneere HPLC detectable only in NMITLI 101R among the chemotypes.
he relative quantitative amounts of the above withanolides in thextracts are presented in Table 1.
.2. NMITLI 101R and 128R showed augmentation of humoralesponse at lower dose
Mice were fed at 10, 30 and 100 mg/kg/body weight for 14 days.
ice were healthy as reflected in slight increase in their bodynd spleen weight (Table 2). All the three extracts of WS havehown an elevated HA titre in mouse sera than the control in vari-ble doses (Table 2). NMITLI 101R fed mice showed significant up
ithanolidal phytochemical constituents of aqueous-ethanol extract of roots of theFN, withaferin A; 17-OH WFN, 17-hydroxy withaferin A; Hy-d WFN, 17-hydroxy
regulation of HA titre of almost 2 fold even at 10 mg dose, whereastitre drastically decreased below control at 30 and 100 mg doses.
NMITLI 101R, NMITLI 118R and NMITLI 128R were profiled by HPLC. Comparativewithanolide complement characteristics of the roots of the three chemotypes arepresented in Fig. 1A–C.
a Peak area units ×106.b Not detected.

S. Kushwaha et al. / Vaccine
Tab
le
2Ef
fect
of
NM
ITLI
101R
, NM
ITLI
118R
and
NM
ITLI
128R
on
rela
tive
bod
y
and
sple
en
wei
ght,
HA
titr
e
and
PFC
.
Dos
e
(mg/
kg/d
ay)
Rel
ativ
e
tren
d
in
orga
n
wei
ght
HA
titr
e
PFC
Bod
y
Sple
en
101R
118R
128R
101R
118R
128R
0
(on
ly
veh
icle
)
20
±
0.83
0.21
2
±
0.01
5
640
±
128.
0
1049
±
98.6
7
416.
0
±
96.0
81.3
±
8.7
23.6
0
± 3.
14
121.
5
±
23.0
210
20.2
±
0.96
0.21
5
±
0.01
7
1126
±
150.
8*11
98
±
92.3
1
896.
0
±
128.
0**11
0
±
10.2
*30
.40
± 2.
57
219.
3
±
19.4
8*
30
20.6
±
0.87
0.27
5
±
0.02
5*61
4.4
±
102.
4
2119
±
75.2
**12
80.0
±
256.
0**13
4
±
15.6
*42
.63
± 3.
26*
236.
4
±
28.1
9*
100
21.1
±
0.97
0.28
3
±
0.02
9*33
2.8
±
76.8
0*22
50
±
112.
4**17
92.0
±
256.
0**16
0
±
24.3
*71
.63
±
4.19
**25
5
±
27.5
4*
Extr
act
and
veh
icle
trea
ted
mic
e
wer
e
wei
ghte
d
afte
r
14
day
s
of
feed
ing,
sacr
ifice
d
and
sple
en
wei
ght
mea
sure
d.
For
hae
mag
glu
tin
atio
n
assa
y
extr
acts
and
veh
icle
trea
ted
mic
e
wer
e
chal
len
ged
wit
h
SRB
C
(1
×10
7ce
lls/
100
�l)
on
day
9th
and
seru
m
coll
ecte
d
on
day
15th
. An
tibo
dy
leve
l on
the
seru
m
was
mea
sure
d.
For
PFC
assa
y
asep
tica
lly
isol
ated
sple
noc
ytes
from
con
trol
and
trea
ted
mic
e
wer
e
mix
ed
wit
h
SRB
C
and
over
laid
on
agar
ose
laye
r.
PFC
valu
es
wer
e
det
erm
ined
as
des
crib
ed
in
mat
eria
ls
and
met
hod
s.
The
valu
es
rep
rese
nt
the
mea
n
from
at
leas
t
thre
e
sep
arat
e
exp
erim
ents
±
stan
dar
d
erro
r
(n
=
5).
*p
<
0.05
was
sign
ifica
nt
com
par
ed
wit
h
the
con
trol
grou
p.
**p
<
0.00
1
was
sign
ifica
nt
com
par
ed
wit
h
the
con
trol
grou
p.
30 (2012) 1083– 1093 1087
hand, NMITLI 118R fed mice exhibited enhanced HA titre only athigher doses.
Humoral response was also measured by counting PFC afterSRBC mixed splenocytes were incubated with serum collected fromextract and vehicle fed mice (Table 2). NMITLI 101R exhibited∼1.5 fold increase in antibody producing cells at 10 mg dose andremained upregulated even at higher dose. Immune cells from fedmice with extract NMITLI 128R also showed enhanced PFC numberat lower dose. In contrast, NMITLI 118R showed significant increaseof humoral response only at higher doses.
3.3. NMITLI 101R showed higher DTH response at the 10 mg doseafter 24 h
The cellular response was monitored after immunization withSRBC by measuring DTH response (Table 3). NMITLI 101R showed∼1.5 fold increase in foot pad swelling at 10 mg dose even after24 h of SRBC challenge and response continued till 72 h challenge.On the contrary, there was no significant DTH response in NMITLI118R fed mice at 10 mg dose after 24 h than control. NMITLI 128Ralso did not show any effect in eliciting DTH response at earlier timepoints of SRBC challenge at lower doses. However, a 2 fold increasein footpad thickness was noticed only after 72 h at lower doses. Adrastic suppression in DTH response was observed 100 mg/kg doseat all time points.
3.4. NMITLI 101R is the most potential stimulator for Th1 and Th2cytokines in secretion
Modulation in T and B cells, intracellular as well as secretedcytokines and ROS generation was tested at lower doses(3–30 mg/kg/day). NMITLI 101R fed group exhibited significantincrease in IL-4 secretion at all three (3–30 mg) doses (p < 0.05,Fig. 2A). In contrast, NMITLI 128R was unable to increase significantIL-4 at 3 mg dose and NMITLI 118R could not induce this cytokineeven at highest dose.
Secretion of IFN� cytokine was significantly up regulated at allthree doses in NMITLI 101R fed mice (Fig. 2B). Both NMITLI 118Rand NMITLI 128R chemotypes were unable to increase significantIFN� production at lower dose (3.0 mg). However, enhanced IFN�secretion was observed only at higher doses by NMITLI 118R fedmice. NMITLI 128R fed mice marginally increased IFN� secretion athigher doses.
3.5. Macrophages of NMITLI 101R fed mice induced maximumROS
ROS are ultimate effecter molecules which are detrimentalfor pathogen and help the immune system to clear the infec-tion. Macrophages isolated from NMITLI 101R fed mice inducedmaximum amount of ROS than other chemotypes as fluorescenceintensity being higher than others (Fig. 3).
3.6. NMITLI 101R is leading in positive modulation of T and B cellproliferation
Splenocytes isolated from the treated mice showed differentialmodulation in number of T and B cells. The trend of upregulationof CD3+/CD4+ population of T cells in NMITLI 101R fed mice wasenhanced at 10 mg dose than control (Fig. 4A, p < 0.001). Similartrend was observed in mice fed both with extract NMITLI 118R andNMITLI 128R.
NMITLI 101R fed mice exhibited higher CD3+/CD8+ populationeven at 3 mg dose (p < 0.01). Both NMITLI 118R and NMITLI 128Rfailed to increase this cell population at this dose (Fig. 4B). The num-ber of cells enhanced only at higher doses in all three chemotypes
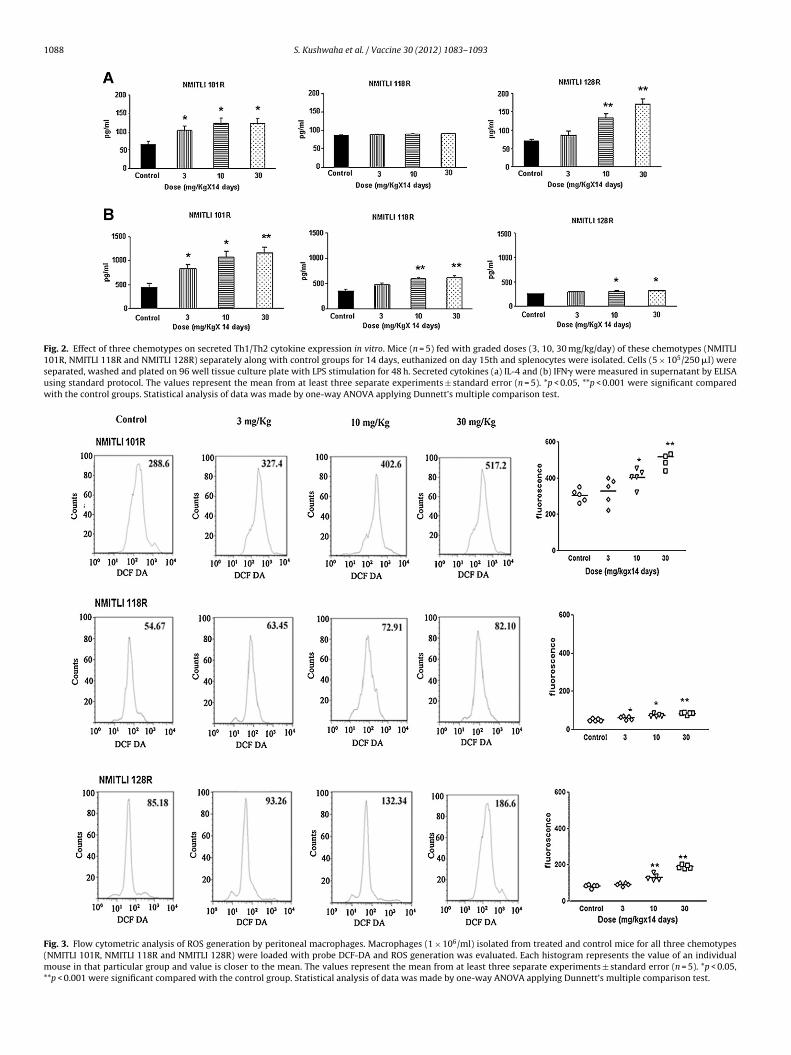
1088 S. Kushwaha et al. / Vaccine 30 (2012) 1083– 1093
Fig. 2. Effect of three chemotypes on secreted Th1/Th2 cytokine expression in vitro. Mice (n = 5) fed with graded doses (3, 10, 30 mg/kg/day) of these chemotypes (NMITLI101R, NMITLI 118R and NMITLI 128R) separately along with control groups for 14 days, euthanized on day 15th and splenocytes were isolated. Cells (5 × 105/250 �l) wereseparated, washed and plated on 96 well tissue culture plate with LPS stimulation for 48 h. Secreted cytokines (a) IL-4 and (b) IFN� were measured in supernatant by ELISAusing standard protocol. The values represent the mean from at least three separate experiments ± standard error (n = 5). *p < 0.05, **p < 0.001 were significant comparedwith the control groups. Statistical analysis of data was made by one-way ANOVA applying Dunnett’s multiple comparison test.
Fig. 3. Flow cytometric analysis of ROS generation by peritoneal macrophages. Macrophages (1 × 106/ml) isolated from treated and control mice for all three chemotypes(NMITLI 101R, NMITLI 118R and NMITLI 128R) were loaded with probe DCF-DA and ROS generation was evaluated. Each histogram represents the value of an individualmouse in that particular group and value is closer to the mean. The values represent the mean from at least three separate experiments ± standard error (n = 5). *p < 0.05,**p < 0.001 were significant compared with the control group. Statistical analysis of data was made by one-way ANOVA applying Dunnett’s multiple comparison test.
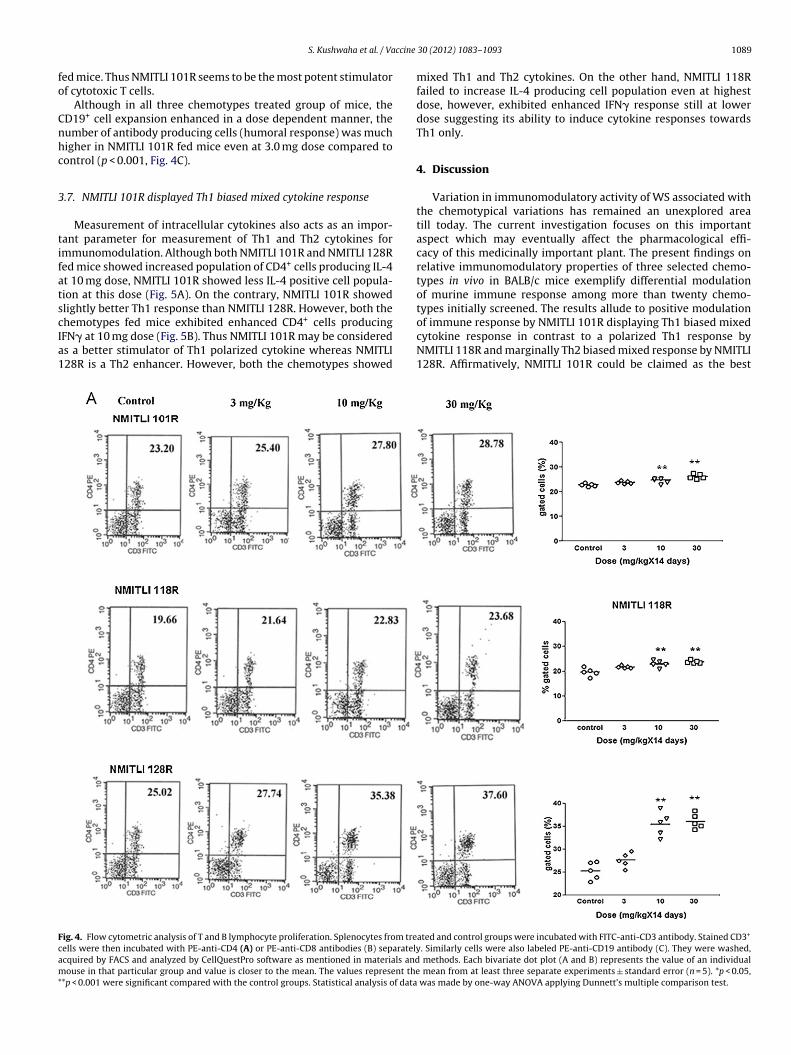
ccine
fo
Cnhc
3
tifatscIa1
Fcam*
S. Kushwaha et al. / Va
ed mice. Thus NMITLI 101R seems to be the most potent stimulatorf cytotoxic T cells.
Although in all three chemotypes treated group of mice, theD19+ cell expansion enhanced in a dose dependent manner, theumber of antibody producing cells (humoral response) was muchigher in NMITLI 101R fed mice even at 3.0 mg dose compared toontrol (p < 0.001, Fig. 4C).
.7. NMITLI 101R displayed Th1 biased mixed cytokine response
Measurement of intracellular cytokines also acts as an impor-ant parameter for measurement of Th1 and Th2 cytokines formmunomodulation. Although both NMITLI 101R and NMITLI 128Red mice showed increased population of CD4+ cells producing IL-4t 10 mg dose, NMITLI 101R showed less IL-4 positive cell popula-ion at this dose (Fig. 5A). On the contrary, NMITLI 101R showedlightly better Th1 response than NMITLI 128R. However, both the
hemotypes fed mice exhibited enhanced CD4+ cells producingFN� at 10 mg dose (Fig. 5B). Thus NMITLI 101R may be considereds a better stimulator of Th1 polarized cytokine whereas NMITLI28R is a Th2 enhancer. However, both the chemotypes showedig. 4. Flow cytometric analysis of T and B lymphocyte proliferation. Splenocytes from treells were then incubated with PE-anti-CD4 (A) or PE-anti-CD8 antibodies (B) separatelycquired by FACS and analyzed by CellQuestPro software as mentioned in materials andouse in that particular group and value is closer to the mean. The values represent the
*p < 0.001 were significant compared with the control groups. Statistical analysis of data
30 (2012) 1083– 1093 1089
mixed Th1 and Th2 cytokines. On the other hand, NMITLI 118Rfailed to increase IL-4 producing cell population even at highestdose, however, exhibited enhanced IFN� response still at lowerdose suggesting its ability to induce cytokine responses towardsTh1 only.
4. Discussion
Variation in immunomodulatory activity of WS associated withthe chemotypical variations has remained an unexplored areatill today. The current investigation focuses on this importantaspect which may eventually affect the pharmacological effi-cacy of this medicinally important plant. The present findings onrelative immunomodulatory properties of three selected chemo-types in vivo in BALB/c mice exemplify differential modulationof murine immune response among more than twenty chemo-types initially screened. The results allude to positive modulation
of immune response by NMITLI 101R displaying Th1 biased mixedcytokine response in contrast to a polarized Th1 response byNMITLI 118R and marginally Th2 biased mixed response by NMITLI128R. Affirmatively, NMITLI 101R could be claimed as the bestated and control groups were incubated with FITC-anti-CD3 antibody. Stained CD3+
. Similarly cells were also labeled PE-anti-CD19 antibody (C). They were washed, methods. Each bivariate dot plot (A and B) represents the value of an individual
mean from at least three separate experiments ± standard error (n = 5). *p < 0.05, was made by one-way ANOVA applying Dunnett’s multiple comparison test.
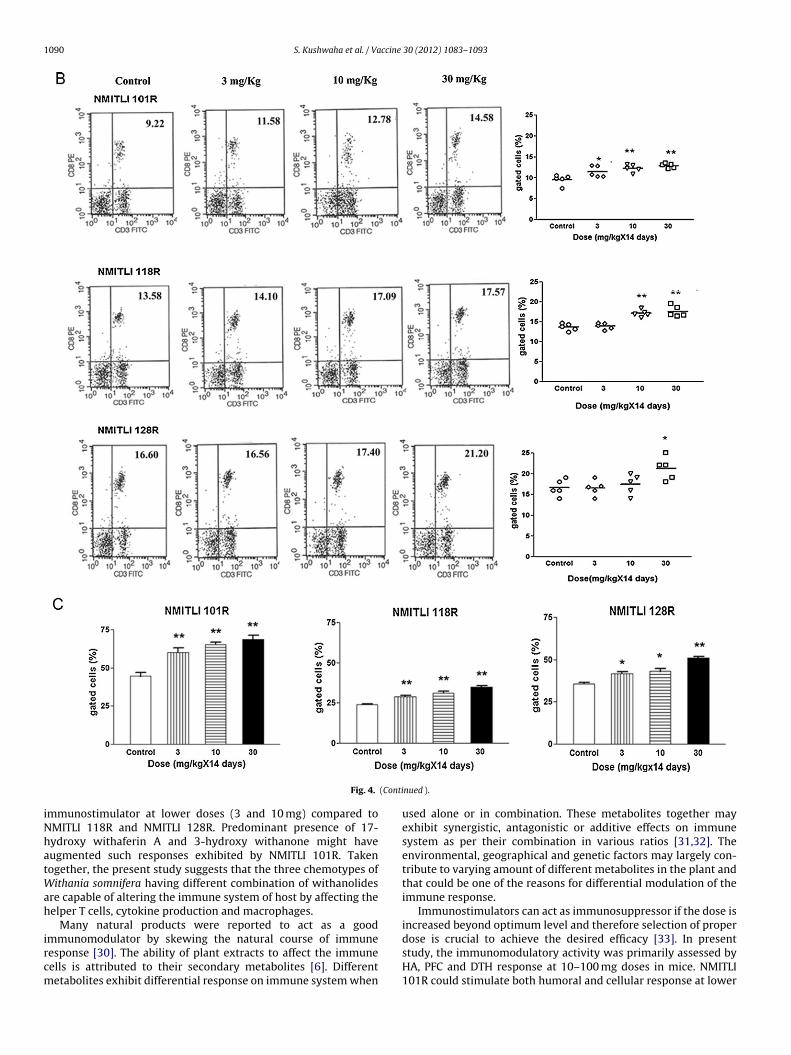
1090 S. Kushwaha et al. / Vaccine 30 (2012) 1083– 1093
(Conti
iNhatWah
ircm
Fig. 4.
mmunostimulator at lower doses (3 and 10 mg) compared toMITLI 118R and NMITLI 128R. Predominant presence of 17-ydroxy withaferin A and 3-hydroxy withanone might haveugmented such responses exhibited by NMITLI 101R. Takenogether, the present study suggests that the three chemotypes of
ithania somnifera having different combination of withanolidesre capable of altering the immune system of host by affecting theelper T cells, cytokine production and macrophages.
Many natural products were reported to act as a good
mmunomodulator by skewing the natural course of immuneesponse [30]. The ability of plant extracts to affect the immuneells is attributed to their secondary metabolites [6]. Differentetabolites exhibit differential response on immune system whennued ).
used alone or in combination. These metabolites together mayexhibit synergistic, antagonistic or additive effects on immunesystem as per their combination in various ratios [31,32]. Theenvironmental, geographical and genetic factors may largely con-tribute to varying amount of different metabolites in the plant andthat could be one of the reasons for differential modulation of theimmune response.
Immunostimulators can act as immunosuppressor if the dose isincreased beyond optimum level and therefore selection of proper
dose is crucial to achieve the desired efficacy [33]. In presentstudy, the immunomodulatory activity was primarily assessed byHA, PFC and DTH response at 10–100 mg doses in mice. NMITLI101R could stimulate both humoral and cellular response at lower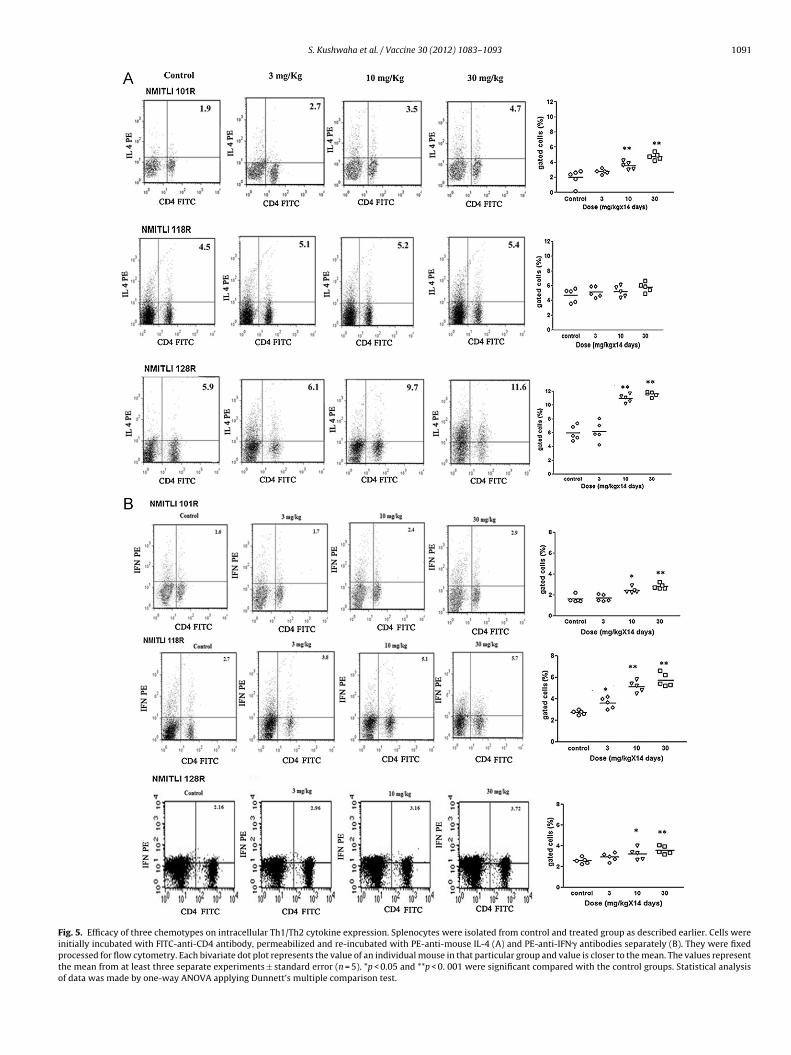
S. Kushwaha et al. / Vaccine 30 (2012) 1083– 1093 1091
Fig. 5. Efficacy of three chemotypes on intracellular Th1/Th2 cytokine expression. Splenocytes were isolated from control and treated group as described earlier. Cells wereinitially incubated with FITC-anti-CD4 antibody, permeabilized and re-incubated with PE-anti-mouse IL-4 (A) and PE-anti-IFN� antibodies separately (B). They were fixedprocessed for flow cytometry. Each bivariate dot plot represents the value of an individual mouse in that particular group and value is closer to the mean. The values representthe mean from at least three separate experiments ± standard error (n = 5). *p < 0.05 and **p < 0. 001 were significant compared with the control groups. Statistical analysisof data was made by one-way ANOVA applying Dunnett’s multiple comparison test.

1092 S. Kushwaha et al. / Vaccine
Tab
le
3C
han
ge
in
foot
volu
me
in
NM
ITLI
101R
, NM
ITLI
118R
and
NM
ITLI
128R
fed
mic
e.
Dos
e
(mg/
kg/d
ay)
Foot
pad
swel
lin
g
in
mm
at
dif
fere
nt
hou
rs
NM
ITLI
101R
NM
ITLI
118R
NM
ITLI
128R
24
48
72
24
48
72
24
48
72
0
(on
ly
veh
icle
)0.
60±
0.12
0.58
±
0.11
0.44
±
0.05
0.04
0
±
0.00
7
0.02
5
±
0.00
5
0.02
7
±
0.00
7
0.02
2
±
0.00
7
0.01
7 ±
0.01
1
0.01
0
±
0.00
410
0.97
±
0.09
*0.
68
±
0.07
0.73
±
0.12
*0.
015
±
0.00
5*0.
045
±
0.00
5*0.
030
±
0.00
5
0.02
4
±
0.00
7
0.02
2 ±
0.00
8
0.02
2
±
0.00
5*
30
0.84
±
0.02
*0.
66
±
0.14
0.47
±
0.02
0.07
8
±
0.01
0*0.
055
±
0.00
5**0.
032
±
0.00
2
0.03
0
±
0.01
2
0.03
0
±
0.01
2
0.02
7
±
0.00
6*
100
0.65
±
0.07
0.65
±
0.08
0.62
±
0.14
0.07
8
±
0.00
4**0.
056
±
0.00
7**0.
035
±
0.00
2
0.01
2
±
0.00
4
0.01
2
±
0.00
4
0.01
2
±
0.00
6
SRB
C
imm
un
ized
extr
acts
and
veh
icle
trea
ted
mic
e
wer
e
chal
len
ged
wit
h
SRB
C
(2
×10
8/5
0
�l)
susp
ensi
ons
in
righ
t-h
ind
foot
pad
intr
ader
mal
ly. T
hic
knes
s
of
foot
pad
was
mea
sure
d
at
24, 4
8,
72
h
pos
t
chal
len
ge
as
des
crib
ed
inm
ater
ials
and
met
hod
s
sect
ion
. Th
e
valu
es
rep
rese
nt
the
mea
n
from
at
leas
t
thre
e
sep
arat
e
exp
erim
ents
±
stan
dar
d
erro
r
(n
=
5).
*p
<
0.05
was
sign
ifica
nt
com
par
ed
wit
h
the
con
trol
grou
p.
**p
<
0.00
1
was
sign
ifica
nt
com
par
ed
wit
h
the
con
trol
grou
p.
30 (2012) 1083– 1093
doses whereas NMITLI 128R elicited only humoral response at thisdose. On contrary, NMITLI 118R exhibited humoral and cellularresponses only at higher dose. At lower doses potentiality wasobserved in both cellular and humoral parameters warrantingfurther evaluation of chemotypes on immune cells at lower dosesonly. Significant increase in B cell population was noticed intreated groups led by NMITLI 101R at the lower dose. After feedingat lower doses, significant up regulation of CD3+/CD4+ cells wasbrought about in all animals treated with all the three chemotypesthus denoting their ability to stimulate T helper cell populations,the major immune regulatory cells. This increase in CD3+CD4+
cell population may be due to cumulative effect of more thanone active constituent of chemotypes. The ratio of active con-stituents in particular chemotype would ultimately delineate thisCD3+CD4+ helper immune response into Th1 or Th2 type. UnlikeNMITLI 101R and NMITLI 118R, at 10 mg dose, NMITLI 128R did notsignificantly affect CD3+CD8+ population that may be correlatedwith the deviated phytochemical profile of the chemotype rootextracts. The withanolide phytochemical profile of NMITLI 101Rand NMITLI 118R had a relatively more matching of peaks resolvedby HPLC and it differed substantially from NMITLI 128R. Exceptwithanolide A and withanone, NMITLI 128R differed from NMITLI101R and NMITLI 118R due to the non-detectable presence ofother withanolides. Nevertheless, differential immunomodulatoryresponse could be equally well attributed to presence of somepolar withanolide moieties in NMITLI 101R and NMITLI 118R.This may be due to the nature of immunomodulation eitherstimulation or suppression depending on the concentration ofconstituents. Such low abundance compounds in the chemo-types are hydroxyl-derivatives of withaferin A (e.g. 17-hydroxywithaferin, 17-hydroxy-27-deoxy withaferin A) and withanone(3-hydroxy withanone) that appear in the early part (polar regionof the chromatogram) of the reverse-phase HPLC chromatogram.
Recent model suggests that if the cytokine balance is shiftedtowards a Th2 predominate state, then an enhanced humoralimmune response could result, possibly even at the expense ofTh1-dependent cell mediated immunity. The NMITLI 101R wasfound to up regulate both IFN� and IL-4 cytokines suggesting pres-ence of mixed Th1 and Th2 stimulating constituents. However,its relative immunomodulatory effect was skewed towards Th1as evidenced by significant increase in IFN�/IL-4 ratios, a com-monly used index of Th1/Th2 immunity. Th1 dominant responsemay be due to the abundance of withanone while 17-hydroxy with-aferin A and to some extent 3-hydroxy withanone might have beenresponsible for its mixed Th1 and Th2 nature. In contrast, higherIL-4/IFN� ratio demonstrated by NMITLI 128R indicated slightlyTh2 polarized immune response as also corroborated by predomi-nant humoral response seen by PFC and HA titre. Complete absenceof withaferin A, its derivatives (17-hydroxy 27-deoxy withaferin,17-hydroxy withaferin A) and 3-hydroxy withanone along withrudimentary presence of withanone may possibly be responsiblefor such immunomodulation. On the other hand, major Th1 dom-inant immune response exhibited by NMITLI 118R is reflected byan increase in IFN� production with no induction of IL-4 produc-tion as also evidenced from its IFN�/IL-4 ratio. Considerably higheramount of withanone than NMITLI 101R might be responsible forits sole Th1 response. Maximum amount of ROS was generated inmacrophages after NMITLI 101R treatment suggestive of a strongprimary immune response. It may be inferred that the increase inROS generation may be due to the presence of one or more activeconstituents or the combined effect of some other chemical con-stituents. The macrophage activation in NMITLI 101R chemotype
was further augmented by an increased IFN� secretion.Adjuvents are capable of modulating the degree and course ofimmune response through various pathways. Efficacy of a vac-cine can be enhanced by incorporation of pathogen-associated

ccine
mvbobmibtrAo[tdtaatpesa
A
ttpaDm
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
S. Kushwaha et al. / Va
olecular patterns in the formulation. Current generation ofaccines based on purified recombinant molecules is limitedy their poor immunogenicity. Accordingly, use of adjuvantsr immunomodulators along with vaccine is an obvious choiceased on the types of pathogen [34]. Alum, although holding theonopoly in the field of adjuvant is a weak inducer of cell mediated
mmunity, therefore, immunostimulators from plant origin coulde a better option. A triterpene saponin isolated from the soap barkree Quillaja saponaria, having immunomodulatory property is cur-ently being tested as adjuvant with wide array of vaccines [35].djuvant efficacy of aqueous extract of WS showed modulationf only humoral response to DPT vaccine in experimental system36]. Thus it may be envisaged that the use of various chemo-ypes of Withania will differentially skew the immune response inesired way once combined with vaccine antigens paving the wayo be used as adjuvant. These immunostimulants can also be usedlone or as adjunct to chemotherapeutic agents to treat diseasesccompanied with immunosupression such as AIDS, leishmaniasis,uberculosis and patients suffering from generalized immunosup-ression to provide a better treatment strategy. Further detailedvaluation of immunomodulatory activity of pure and active con-tituents is underway to directly correlate the pharmacologicalctivity with the chemical entity.
cknowledgments
The authors are grateful to Council of Scientific and Indus-rial Research (CSIR) for funds provided by CSIR under NMITLIo CSIR-CDRI, CSIR-IICB and CSIR-CIMAP. CM acknowledges sup-ort from Sir J.C. Bose National Fellowship, Department of Sciencend Technology, Govt. of India. We also thank Dr. C.M. Gupta, Ex-irector, CSIR-CDRI for his valuable suggestions and criticism. Theanuscript bears CSIR-CDRI communication no. 8171.
eferences
[1] Ziauddin M, Phansalkar N, Patki P, Diwanay S, Patwardhan B. Stud-ies on immunomodulatory effects of Ashwagandha. J Ethnopharmacol1996;50:69–76.
[2] Fulzele SV, Bhurchandi PM, Kanoje VM, Joshi SB, Dorle AK. Immunostimulantactivity of Ashtamangal ghrita in rats. Ind J Pharmacol 2002;34:194–7.
[3] Sharma V, Sharma S, Pracheta Paliwal R. Withania somnifera: a rejuvenatingayurvedic medicinal herb for the treatment of various human ailments. Int JPharm Tech Res 2011;3:187–92.
[4] Kumar A, Kaul MK, Bhan MK, Khanna PK, Suri KA. Morphological and chemicalvariation in 25 collections of the Indian medicinal plant, Withania somnifera (L.)Dunal (Solanaceae). Genet Resour Crop Evol 2007;54:655–60.
[5] Khajuria RK, Suri KA, Gupta RK, Satti NK, Amina M, Suri OP, et al. Separation,identification and quantification of selected withanolides in plant extracts ofWithania somnifera by HPLC-UV (DAD)-positive ion electrospray ionisation-mass spectroscopy. J Sep Sci 2004;27:541–6.
[6] Agarwal R, Diwanay S, Patki P, Patwardhan B. Studies on immunomodula-tory activity of Withania somnifera (Ashwagandha) extracts in experimentalimmune inflammation. J Ethanopharmocol 1999;67:27–35.
[7] Sumantran VN, Kulkarni A, Boddul S, Chinchwade T, Koppikar SJ, HarsulkarA, et al. Chondroprotective potential of root extracts of Withania somnifera inosteoarthritis. J Biosci 2007;32:299–307.
[8] Dhuley JN. Adaptogenic and cardioprotective action of Ashwagandha in rats
and frogs. J Ethnopharmacol 2000;70:57–63.[9] Mandal C, Dutta A, Mallick A, Chandra S, Misra L, Sangwan RS, et al. With-aferin A induces apoptosis by activating p38 mitogen-activated protein kinasesignaling cascade in leukemic cells of lymphoid and myeloid origin throughmitochondrial death cascade. Apoptosis 2008;13:1450–64.
[
[
30 (2012) 1083– 1093 1093
10] Mondal S, Mandal C, Sangwan R, Chandra S, Mandal C. WithanolideD induces apoptosis in leukemia by targeting the activation of neutralsphingomyelinase–ceramide cascade mediated by synergistic activation of c-Jun N-terminal kinase and p38 mitogen-activated protein kinase. Mol Cancer2010;9:239.
11] Udayakumar R, Kasthurirengan S, Mariashibu TS, Rajesh M, Ramesh AV, Kim SC,et al. Hypoglycaemic and hypolipidaemic effects of Withania somnifera root andleaf extracts on alloxan-induced diabetic rats. Int J Mol Sci 2009;10:2367–82.
12] Harikrishnan B, Subramanian P, Subash S. Effect of Withania Somnifera rootpowder on the levels of circulatory lipid peroxidation and liver marker enzymesin chronic hyperammonemia. E-J Chem 2008;5:872–7.
13] Dadkar VN, Ranadive NU, Dhar HL. Evaluation of antistress (adaptogen) activityof Withania somnifera (Ashwagandha). Ind J Clin Biochem 1987;2:101–8.
14] Kuboyama T, Tohda C, Komatsu K. Neuritic regeneration and synaptic recon-struction induced by withanolide A. Br J Pharmacol 2005;144:961–71.
15] O’Garra A, Robinson D. Development and function of T helper 1 cells. AdvImmunol 2004;83:133–62.
16] Naik S. Introduction to the immune system. J Indian Rheumatol Assoc2003;11:8–13.
17] Noelle RJ, Snow EC. Cognate interactions between helper T cells and B cells.Immunol Today 1990;10:361–8.
18] Lifson JD, Engleman EG. Role of CD4 in normal immunity and HIV infection.Immunol Rev 1989;109:93–117.
19] Malik F, Singh J, Khajuria A, Suri KA, Satti NK, Singh S, et al. A standardizedroot extract of Withania somnifera and its major constituent withanolide-Aelicit humoral and cell-mediated immune responses by up-regulation of Th1-dominant polarization in BALB/c mice. Life Sci 2007;80:525–38.
20] Syamsudin, Darmono, Kusmardi. The effect of Garcinia parvifolia Miq (activefraction) on phagocytosis by peritoneal macrophages during plasmodiumberghei infection in mice. Res J Immunol 2008;1:16–20.
21] Su YW, Chiou WF, Chao SH, Lee MH, Chen CC, Tsai YC. Ligustilide preventsLPS-induced iNOS expression in RAW 264.7 macrophages by preventing ROSproduction and down-regulating the MAPK, NF-�B and AP-1 signaling path-ways. Int Immunopharmacol 2011;11:1166–72.
22] Mishra LC, Singh BB, Dagenais S. Scientific basis for the therapeutic useof Withania somnifera (Ashwagandha): a review. Altern Med Rev 2000;5:334–46.
23] Chaurasiya ND, Sangwan RS, Misra LN, Tuli R, Sangwan NS. Metabolic clus-tering of a core collection of Indian ginseng (Withania somnifera) throughDNA, isoenzymes, polypeptide and withanolide profile diversity. Fitoterapia2009;80:496–505.
24] Chaurasiya ND, Uniyal GC, Lal P, Misra L, Sangwan NS, Tuli R, et al. Anal-ysis of withanolides in root and leaf of Withania somnifera by HPLC withphoto diode array and evaporative light scattering detection. Phytochem Anal2008;19:148–54.
25] Sharma V, Chatterjee M, Mandal C, Sen S, Basu D. Rapid diagnosis of Indianvisceral leishmaniasis using achatinin H, a 9-O-acetylated sialic acid bindinglectin. Am J Trop Med Hyg 1998;58:551–4.
26] Cunningham AJ, Szenberg A. Further improvements in the plaque techniquefor detecting single antibody forming cells. Immunology 1968;14:599–600.
27] Titus RG, Chiller JM. A simple and effective method to assess murine delayed-type hypersensitivity to proteins. J Immunol Methods 1981;45:65–78.
28] Khan S, Malik F, Suri KA, Singh J. Molecular insight into the immune up-regulatory properties of the leaf extract of Ashwagandha and identificationof Th1 immunostimulatory chemical entity. Vaccine 2009;27:6080–7.
29] Pathak M, Bano N, Dixit P, Soni VK, Kumar P, Maurya R, et al. Immunosuppres-sive activity of hexane and ethanolic extracts of Pterospermum acerifolium seedsin BALB/c mice. Med Chem Res 2011;20:1667–73.
30] Patil VV, Bhangale SC, Patil VR. Studies on immunomodulatory activity of Ficuscarica. Int J Pharm Sci 2010;2:97–9.
31] Barua CC, Gupta PP, Patnaik GK, Misra-Bhattacharya S, Goel RK, Kul-shrestha DK, et al. Immunomodulatory effect of Albizzia Lebbeck. Pharm Biol2000;38:161–6.
32] Kawakita SW, Giedlin HS, Nomoto K. Immunomodulators from higher plants. JNat Med 2005;46:34–8.
33] Daswani BR, Yegnanarayan R. Immunomodulatory activity of septilin, a poly-herbal preparation. Phytother Res 2002;16:162–5.
34] Lima KM, dos Santos SA, Rodrigues Jr JM, Silva CL. Vaccine adjuvant: it makesthe difference. Vaccine 2004;22:2374–9.
35] Sun HX, Xie Y, Ye YP. Advances in saponin-based adjuvants. Vaccine2009;27:1787–96.
36] Gautam M, Diwanay S, Gairola S, Shinde YS, Jadhav SS, Patwardhan B. Immuneresponse modulation to DPT vaccine by aqueous extract of Withania somniferain experimental system. Int Immunopharmacol 2004;4:841–9.




















