
Characterization of the prime and non-prime active site specificities of proteases by...
25
Characterization of Protease Prime and Non-prime Active Site Specificities Using Proteome-derived Peptide Libraries and Tandem Mass Spectrometry Oliver Schilling 1 , Pitter F. Huesgen 2 , Olivier Barré 2 , Ulrich auf dem Keller 3 , and Christopher M. Overall 2,4 1 University of Freiburg Institute for Molecular Medicine and Cell Research Stefan-Meier-Strasse 17 D-79104 Freiburg Germany [email protected] 2 University of British Columbia Department of Biochemistry and Molecular Biology Department of Oral Biological and Medical Sciences Centre for Blood Research 4.401 Life Sciences Institute 2350 Health Sciences Mall Vancouver, British Columbia Canada V6T 1Z3 3 ETH Zürich Institute of Cell Biology Schafmattstrasse 18 CH-8093 Zürich Switzerland 4 To whom correspondence should be addressed. [email protected] phone: +1-604-822-2958 fax: +1-604-822-7742 web: http://www.clip.ubc.ca
-
Upload
fz-juelich -
Category
Documents
-
view
0 -
download
0
Transcript of Characterization of the prime and non-prime active site specificities of proteases by...

Characterization of Protease Prime and Non-prime Active Site
Specificities Using Proteome-derived Peptide Libraries
and Tandem Mass Spectrometry
Oliver Schilling 1, Pitter F. Huesgen 2, Olivier Barré 2, Ulrich auf dem Keller3, and Christopher M.
Overall 2,4
1University of Freiburg
Institute for Molecular Medicine and Cell Research
Stefan-Meier-Strasse 17
D-79104 Freiburg
Germany
2 University of British Columbia
Department of Biochemistry and Molecular Biology
Department of Oral Biological and Medical Sciences
Centre for Blood Research
4.401 Life Sciences Institute
2350 Health Sciences Mall
Vancouver, British Columbia
Canada V6T 1Z3
3ETH Zürich
Institute of Cell Biology
Schafmattstrasse 18
CH-8093 Zürich
Switzerland
4 To whom correspondence should be addressed.
phone: +1-604-822-2958
fax: +1-604-822-7742
web: http://www.clip.ubc.ca

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 2 of 21
SUMMARY
To link cleaved substrates in complex systems with a specific protease, the protease active site
specificity is required. Proteomic Identification of Cleavage Sites (PICS) simultaneously determines
both the prime and non�–prime side specificities of individual proteases through identification of
100s of individual cleavage sequences from biologically relevant peptide libraries. PICS also
identifies subsite cooperativity. To generate PICS peptide libraries, cellular proteomes are digested
with a specific protease such as trypsin. Following protease inactivation, primary amines are
protected. After incubation with a test protease, prime�–side cleavage fragments possess a free
neo N�–terminus, which is biotinylated for affinity�–isolation and identification by liquid
chromatography�–tandem mass spectrometry. The corresponding non�–prime sequences are
derived bioinformatically. The step�–by�–step protocol is improved from the primary paper (Schilling
and Overall, 2008, Nature Biotechnology 685�–694) and presents a web service for PICS data
analysis. We also introduce and validate PICS peptide libraries made from Escherichia coli.
Search terms: protease: specificity: cleavage site: peptide library; proteomics; degradomics, mass
spectrometry, WebPICS, subsite cooperativity
Related manuscripts
Schilling, O. and Overall, C. M.,
Proteome-derived, database-searchable peptide libraries for identifying protease cleavage sites
Nat. Biotechnol. 26 (6), 685 (2008).
DOI: 10.1038/nbt1408
A previous version of this workflow was published at
www.natureprotocols.com/2008/05/14/protease_subsite_profiling_wit.php
The present version presents substantial improvements to the workflow, introduces bacterial
proteomes as a source for peptide libraries, and introduces an internet based data analysis.

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 3 of 21
INTRODUCTION
Proteolysis is a ubiquitous post�–translational modification that is often integral in protein
maturation, modulation of function and activity, as well as in cell and tissue localization and
turnover. Protease specificity is fundamentally guided by enzyme substrate interactions, often in
an extended active site cleft 1. Profiling of protease cleavage site specificity is an essential step in
the biochemical characterization of proteases and so provides valuable information for the design
of specific peptide substrates useful for assay development and small molecule protease inhibitors
2. The interactions of protease active sites with peptidic or native protein substrates are described
according to Schechter and Berger 1: Enzyme binding subsites (S) for residues N�–terminal to the
scissile peptide bond are designated as S1, S2 and so on, while the corresponding substrate
peptide (P) residues are designated as P1, P2, etc. Binding pockets and substrate residues C�–
terminal to the scissile peptide bond are termed �“prime side�” and designated as S1�’, S2�’, etc., and
P1�’, P2�’, etc, respectively.
Proteomic Identication of protease Cleavage Sites (PICS) is a unique approach that utilizes
proteome derived peptide libraries to harness natural sequence diversity in order to probe for
cleavage sequence and enzyme specificity. When derived from a fully sequenced organism,
peptides in these libraries can be sequenced using readily available proteomics software.
Therefore the power of mass spectrometry can be utilized for the rapid and automated analysis of
cleaved peptide libraries, which cannot be done for complex synthetic peptide libraries 3. Moreover,
unlike synthetic peptide library approaches 4, including peptide microarrays and positional
scanning peptide libraries, PICS simultaneously determines both the prime and nonprime side
specificities and can also detect subsite cooperativity. In contrast to genetic screens such as
phage 5 or bacterial substrate display 6, PICS does not require laborious and costly iterative
selection schemes. Rather one to ten liquid chromatography tandem mass spectrometry (LC-
MS/MS) analyses are required per assay, which is cost effective, fast and can be performed
overnight by most core facilities.

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 4 of 21
The key to PICS is the use of natural proteomes to generate the diverse peptide libraries. Such
libraries expose the protease to natural sequence diversity while at the same time they are
amenable to MS/MS sequencing and database sequence analysis. To prepare complementary
PICS libraries, cellular proteomes are first digested with specific endoproteases such as trypsin
(cleaves C-terminal to basic amino acids), GluC (cleaves C-terminal to glutamate and occasionally
aspartate), or chymotrypsin (cleaves C-terminal to hydrophobic amino acids) (Fig. 1). After
digestion and protease inactivation, any disulfide bonds are reduced and free thiol groups are
chemically protected. Primary amines (N-terminal -amines and lysine -amines) are then blocked
by dimethylation. The use of the three different proteases to prepare three classes of libraries
ensures that basic residues are accessible in the peptides in the GluC and chymotryptic libraries.
Similarly, acidic residues are also positioned throughout the sequences in the tryptic and
chymotryptic libraries. Hence, cleavage data is not skewed by the C-terminal specificity of trypsin,
GluC or chymotrypsin.
Unlike most synthetic peptide libraries that typically exclude cysteine, PICS libraries contain all
amino acids with the caveat that the cysteines and lysines are blocked. While phage and bacterial
substrate displays typically include unprotected cysteine and lysine residues, PICS library
preparation can be modified to use reversible cysteine protection to allow for profiling of specificity
towards free cysteines 3. This strategy has however not yet been used in practice. Lysine
protection proceeds through dimethylation, a sterically small modification that preserves its basic
character. Proteases such as MMP 2 recognize dimethylated lysine as a basic residue and display
a positional preference (here in P2�’) that is corroborated by further studies employing unprotected
lysine 3,7.
In a PICS assay, the library is incubated with a test protease (Fig. 2). C-terminal cleavage
products possess free N-terminal -amines that can be affinity-tagged with a cleavable biotin,
allowing for streptavidin�–based isolation of the cleavage products. LC�–MS/MS identifies the
enriched C-terminal cleavage peptides. The corresponding sequences N�–terminal to the cleavage
site are derived using bioinformatics. PICS has been successfully used for aspartic�–, cysteine�–,

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 5 of 21
metallo�–, and serine proteases, yielding 10s �– 100s of cleavage sequences in a single assay that
profiles subsite specificity from up to P6 �– up to P6�’. A significant advantage of our approach is that
since individual peptide sequences are identified then subsite cooperativity can be characterized
as shown for the S3 and S1 binding pockets of HIV�–1 protease 3.
PICS only profiles the active site specificities of endoproteases and is not suited for the
characterization of exoproteases: Blocked N termini are often not recognized by aminopeptidases
whereas carboxypeptidases do not produce C-terminal peptides that are required for a PICS
analysis. Importantly, the PICS approach�—like any other peptidic specificity profiling technique�—
does not identify physiological substrates. Many proteases do not utilize optimal cleavage sites
and most potential cleavage sites predicted from consensus cleavage site data are shielded from
proteolysis in natively folded proteins. So, simple extrapolation to physiological cleavage events
based on consensus cleavage site data yields an unacceptably high number of false candidate
substrates 3. PICS specificity profiles are however useful for increasing the confidence of candidate
substrates identified by in vivo substrate screening experiments such as Terminal Amine Isotope
Labeling of Substrates 8. As shown for matrix metalloproteinase (MMP) 2 (Fig. 3), there is a close
match of the active site specificity profiles determined using native proteins with TAILS and
denatured peptides with PICS. Hence, for substrate discovery experiments analyzing complex
biological samples, comparing PICS specificity profiles to the TAILS data is helpful for ruling out
low confidence candidate substrates that might have been generated by secondary cleavage
events by unrelated proteases in the biological sample. An important use of protease specificity
profiles is in the development of specific inhibitors with the aim to determine unique specificity
features, especially when comparing related proteases 9. This is even more powerful in
combination with 3D structural information.
PICS libraries have been generated from HEK293 and CCL243 cells grown in suspension culture
as well as HT1080 and HEK293 adherent cells. Recently, PICS assays were also performed using
Escherichia coli proteome as a peptide library source. In validating E. coli libraries we compared
MMP-2 digest results with those generated previously. PICS assays of MMP-2 using human and

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 6 of 21
bacterial peptide libraries yielded comparable specificity profiles (Fig. 3). Interestingly, the primary
specificity determinants were more pronounced in the PICS profile using the bacterial peptide
library. Hence, we conclude that mammalian proteases can be profiled using bacterial peptide
libraries, which although less complex, still provide sufficient sequence diversity for protease
specificity propfiling.
The following step-by-step protocol describes in detail how to generate PICS peptide libraries and
how to perform a PICS assay (Fig. 4). Compared to the original PICS protocol 3, and also the
previously published step-by-step protocol on the community generated section of the Nature
Protocols Network (www.natureprotocols.com/2008/05/14/protease_subsite_profiling_wit.php), the
present step-by-step protocol is further streamlined and peer reviewed. We have simplified the
library preparation and optimized the entire workflow. In addition, we introduce bacterial proteome
sources for PICS peptide libraries. The bioinformatic data analysis and data visualization have also
been streamlined by a novel web�–based service. Using pre prepared libraries, a typical PICS
assay can now be performed within two days, including LC MS/MS analysis.
MATERIALS
REAGENTS
4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid (HEPES, Sigma-Aldrich)
Acetonitrile (Sigma-Aldrich)
CAUTION: Highly flammable and harmful. Wear gloves and protective glasses
CRITICAL: Softeners from laboratory plasticware may dissolve in acetonitrile. Avoid
prolonged exposure and heating. Use plasticware (e.g. from Eppendorf) with a track record
of acetonitrile compatibility.
Appropriate cell culture media
Bicinchoninic acid assay (Pierce)
Bradford protein assay (BioRad)
Chymotrypsin (1-chloro-3-tosylamido-7-amino-2-heptanone treated, Worthington)

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 7 of 21
Cyanoborohydride (ALD coupling solution, Sterogene)
CRITICAL Limited stability. Observe recommended storage conditions and shelf life time.
Dimethylsulfoxide (DMSO, Sigma-Aldrich)
CAUTION Wear gloves and protective glasses.
Dithiothreitol (DTT, Sigma-Aldrich)
CRITICAL Instable in water, always prepare fresh stock solution shortly before use.
DNase I (Sigma-Aldrich)
Ethylenediaminetetraacetic acid (EDTA, Sigma-Aldrich)
Formaldehyde (solution for molecular biology 36.5 38% in water, Sigma-Aldrich)
Formic acid (Sigma-Aldrich)
CAUTION Corrosive. Wear gloves and avoid inhalation of fumes.
GluC (Staphylococcus aureus V8 protease, Worthington)
Iodoacetamide (Sigma-Aldrich)
CAUTION Toxic. Wear gloves and protective glasses.
CRITICAL Instable in solution. Always prepare fresh stock solution immediately before use
and avoid exposure to light.
Methanol (MeOH, Fisher)
CAUTION: Highly flammable and toxic. Wear gloves and protective glasses.
Micro bicinchoninic acid assay (Pierce)
NaCl (Sigma-Aldrich)
Phenylmethylsulphonyl fluoride (PMSF, Sigma-Aldrich)
CAUTION Toxic. Wear protective equipment including full-face particle respirator when
preparing the solution.
CRITICAL Instable in water. Always prepare fresh stock solution in DMSO or MeOH
immediately before use.
Phosphate buffered saline (PBS)
Potassium phosphate buffer, adjusted to pH 2.7
Reversed-phase solid phase extraction cartridges (e.g. Sep-Pak, Waters, or equivalent)
RNase E (Sigma-Aldrich)

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 8 of 21
Sephadex G-10 resin (GE Healthcare)
Sodium dodecyl sulfate (SDS, Sigma-Aldrich)
Spin columns, ~10 m pore size, top and bottom plug (Product # 69705, Pierce)
Streptavidin sepharose (GE Healthcare)
CRITICAL Observe recommended storage conditions.
Sulfosuccinimidyl 2-(biotinamido)-ethyl-1,3-dithiopropionate (Sulfo-NHS-SS-biotin, Pierce)
CRITICAL Follow manufacturer�’s instructions for storage. Prepare freshly in DMSO. Allow
package to reach ambient temperature before opening. Do not store as solution.
Test protease for PICS assay
Trans-epoxysuccinyl-L-leucylamido(4-guanidino)butan (E-64,Sigma-Aldrich)
Trichloracetic acid (Sigma-Aldrich)
CAUTION Corrosive. Wear gloves and protective glasses, avoid inhalation of fumes.
Trifluoracetic acid (Sigma-Aldrich)
Trypsin (1-chloro-3-tosylamido-4-phenyl-2-butanone treated, Sigma-Aldrich)
Versene (Invitrogen) or self-prepared
EQUIPMENT
Standard laboratory equipment for cell culture, molecular biology, and protein chemistry
Centrifuge for volumes up to 50 ml at up to 20,000 g
Chromatography columns to pack up to 5 ml resin
Optional: Chromatography system (e.g. GE Healthcare Aekta or equivalent) with UV/VIS and
conductivity detector
Tandem mass spectrometry system coupled in�–line with a nano�–flow chromatography system
Personal computer with internet access
Spectrophotometer, e.g. NanoDrop (Thermo Scientific)
pH test strips (e.g. from Merck, pH range 6 9)
Polystyrene-divinylbenzene resin column (e.g. GE Healthcare or equivalent)
Table top centrifuge for 2 ml reaction tubes
Ultrasonication bath

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 9 of 21
Vacuum evaporation system (commonly referred to as speed-vac)
XK 26/20 columns (GE Healthcare)
REAGENT SETUP
Strains: Any cell line or other proteome source from an organism with a comprehensive proteome
database. We have successfully used human HEK293, HT1080 and CCL243 cell lines, as well as
E. coli DH5 .
PROCEDURE
PREPARATION OF PROTEOME–DERIVED PEPTIDE LIBRARY
1. Grow and harvest (cellular) proteome source. Follow the steps in Options A or B for
adherent and suspension cell cultures respectively.
A. Adherent cell line.
i. Grow cells to confluence. A 150 �– 175 cm2 cell culture plate typically yields 2
�– 6 mg of purified proteome�–derived peptide library. We recommend starting
library preparation with material from at least three 150 �– 175 cm2 cell
culture plates.
ii. Detach cells, preferably with protease-free detachment methods such as
incubation with Versene. Avoid scraping of cells since this can result in
mechanical cell rupture and reduced yields.
iii. Centrifuge cells at 400 x g, 4 °C for 5 min; decant supernatant and keep cell
pellet. Prolong centrifugation time if necessary.
B. Suspension cell culture, both eukaryotic and bacterial.
i. Grow cells to desired density.
ii. Centrifuge cells at 400 x g, 4 °C for 5 min; decant supernatant and keep cell
pellet. Prolong centrifugation time if required.
iii. Wash cell pellet with PBS and repeat centrifugation.
2. Determine weight of cell pellet.

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 10 of 21
3. Resuspend cell pellet in 5 volumes (assuming that 1 mg cell pellet 1 ml volume) of
hypotonic lysis buffer consisting of 10 mM HEPES, pH 7.5, and the protease inhibitors 1.0
mM PMSF, 10 M E-64, and 10 mM EDTA. Add PMSF directly or prepare a fresh 1 M stock
solution shortly before use since PMSF quickly degrades in water.
CAUTION PMSF is toxic and care should be taken not to inhale dust during stock solution
preparation. Handle according to manufacturer recommendations.
4. Lyse cells. For eukaryotic cells, use repeated aspiration (15 or more times) of the cell
suspension with a 27-gauge needle. Use a larger diameter needle if clogging occurs. For
bacterial cells use ultrasonication rod. Cell suspensions are lysed on ice by 10 cycles of 10
s sonication and 20 s cooling interval.
5. Centrifuge lysate at 20,000 x g for 20 min. Proceed with supernatant.
6. Determine protein concentration and total protein amount using the Bradford assay
method.
7. Adjust to 100 mM HEPES pH 7.5 (final concentration).
8. Add 5 mM DTT (final concentration) and incubate for 60 min at 25 ºC.
CRITICAL STEP Do not exceed temperatures of 37 °C if the reaction is performed in
plastic tubes. At higher temperatures, we have observed leeching of phtalates from plastic
tubes that cause high chemical background noise and peptide ion suppression in the LC-
MS/MS analysis, dramatically decreasing the number of identified peptides.
9. Add 20 mM iodoacetamide (final concentration) and incubate for 60 min at 25 ºC in the
dark.
CRITICAL STEP Do not exceed temperatures of 37 °C if the reaction is performed in
plastic tubes.
10. Quench unreacted iodoacetamide by addition of another 5 mM DTT (final concentration,
accumulated DTT concentration is now 10 mM) and incubate at 25 ºC for 15 min.
11. Precipitate proteins by addition of 15 % (v/v, final concentration) TCA.
12. Incubate on ice for 1 h.
PAUSE POINT Samples can be stored for several days at -20 ºC or -80 ºC after addition of
TCA.

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 11 of 21
13. Centrifuge at 20,000 x g at 4 °C for 10 min or at 8,500 x g at 4 °C for 1 h.
14. Wash pellet twice with minute amounts of �–20 °C cold methanol. If pellet loosens, briefly
centrifuge.
15. Briefly air dry pellet after the second wash step.
CRITICAL STEP Heating or overdrying makes subsequent resolubilization difficult and
causes sample losses.
16. Overlay pellet with ice-cold 20 mM NaOH. Use sufficient volume to reach an assumed 2.0
mg/ml protein concentration based on the total protein amount determined in step 6.
17. Ultrasonicate in a cooled ultrasconication bath for up to 60 min. Use ice slurry to keep
sample cold. The ultrasconication time can be doubled if the protein pellet is hard to re-
dissolve.
18. Bring to 200 mM HEPES, pH 7.5.
19. Centrifuge at 20,000 x g for 10 min or at 8,500 g for 30 min and collect the supernatant.
20. Determine the protein concentration and total protein amount using the Bradford assay
method.
21. Digest with either trypsin, chymotrypsin or GluC. Use a protease to proteome ratio of 1:100
(w/w) and incubate at 37 °C for 16 h.
22. Analyze a small aliquot of the digest by sodium dodecyl sulfate�–polyacrylamide gel
electrophoresis (SDS�–PAGE) to assess for completion of proteome digest. No major
protein bands > 10 kDa indicative of intact proteins or large protein fragments should be
observed after staining. The absence of large protein fragments indicates cleavage at most
potential cleavage sites and hence sufficient proteome digestion to generate a highly
assortment of peptides. If the proteome is not completely digested, add more of the
protease used in step 21 and incubate additional 3 �– 16 h at 37 °C and repeat step 22.
CRITICAL STEP Incomplete digest causes sample losses and generates peptides with
several missed cleavages that complicate peptide identification.
23. Add 1 mM PMSF (final concentration) to abolish activity of the digestion protease.
24. Add 1.0 M guanidine hydrochloride (final concentration).

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 12 of 21
25. Centrifuge at 20,000 x g at 4 °C for 10 min or at 8,500 x g for 1 h. Continue with
supernatant.
26. Add 5 mM DTT (final concentration) and incubate at 37 °C for 1 h.
27. Add 40 mM iodoacetamide (final concentration) and incubate at 37 °C for 1.5 h.
28. Add 15 mM DTT (final concentration, accumulated concentration is now 20 mM) and
incubate at 37 °C for 10 min.
29. Add 30 mM formaldehyde (final concentration) and 30 mM sodium cyanoborohydride (�“ALD
coupling solution�”, final concentration) and incubate at 25 °C for 2 h.
30. Add another 30 mM formaldehyde and 30 mM sodium cyanoborohydride (final,
accumulated concentrations are now 60 mM) and incubate at 25 °C for 16 h.
31. Add 100 mM glycine (final concentration) and incubate at 25 °C for 0.5 h.
32. Optional purification step: Batch separation of peptides from chemicals and buffer
exchange by G10 size exclusion chromatography. Use a G10 column equilibrated with 10
mM sodium phosphate, 20 % (v/v) acetonitrile, pH 2.7. Collect and combine early fractions
with high A280 and A214 absorption until an increase in conductivity is observed. Before
proceeding, remove acetonitrile by vacuum evaporation (the time varies according to
column and sample size).
PAUSE POINT Samples can be stored at -20 °C for several days after removal of
acetonitrile.
33. Acidify to 0.5 % (v/v) TFA and let degas by applying mild vacuum.
34. Purify by C18 solid phase extraction according to manufacturer�’s instruction with the
exception of eluting with 80 % (v/v) acetonitrile without added TFA. Several solid phase
extraction devices can be coupled in tandem to increase capacity.
35. After elution, determine peptide concentration by the bicinchoninic acid (BCA) assay. The
BioRad assay is not suitable since it detects proteins rather than peptides.
36. Remove acetonitrile from solid phase extraction eluate by vacuum evaporation. Check
remaining volume regularly. Based on the total peptide amount determined in step 35 and
based on the visually estimated volume, do not exceed a peptide concentration of 2 mg/ml.
Once a volume resulting in a peptide concentration of 2 mg/ml has been reached, refill with

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 13 of 21
water to approximately half of the original volume. Repeat this step four times and continue
with a calculated peptide concentration of 1.5 �– 2.0 mg/ml.
37. If peptide precipitation occurs, incubate in an ultrasconication bath for up to 3 h until the
precipitate redissolves.
38. Centrifuge at 20,000 x g for 10 min or at 8,500 g for 30 min and continue with supernatant.
39. Determine final peptide concentration.
40. Store peptide libraries in aliquots of 200 to 400 g at �–80 ºC.
PAUSE POINT
To confirm purity and integrity of the peptide library, 1 to 10 g of the library may be
desalted and analyzed by ESI-MS/MS as detailed in steps 70 to 79.
PROTEASE CLEAVAGE ASSAY AND ISOLATION OF CLEAVAGE PRODUCTS
41. Thaw peptide library. Use 200 g of peptide library if neither the test protease nor assay
buffer (see step 42) contains primary amines such as tris(hydroxymethyl)aminomethane
(Tris) or reducing substances such as DTT. Use 400 g of peptide library if either test
protease or assay buffer contains primary amines such as Tris or reducing substances
such as DTT.
42. Adjust appropriate buffer conditions for the test protease (pH, type of buffer, reducing
agents, co-factors). Do not use detergents, carrier protein or peptide additives which are
incompatible with MS analysis.
43. Adjust peptide library concentration to approximately 1 mg/ml.
44. Add active test protease; typical protease to library ratios are 1:1000 1:50 (w/w) with 1:100
(w/w) usually applied for a first enzyme characterization.
45. Incubate for 1 16 h at a temperature suitable for the protease under investigation.
46. Heat de-activate test protease.
47. Skip this step if both test protease and assay buffer do not contain primary amines such as
Tris or reducing substances such as DTT. If primary amines or reducing substances are
present, perform C18 solid phase extraction using cartridges with sufficient peptide binding
capacity and follow manufacturer's instructions. After elution, vacuum-evaporate C18 solid

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 14 of 21
phase extraction eluate to near-dryness and reconstitute sample in 200 l 200 mM HEPES,
pH 7.5.
48. Biotinylate cleavage products using 0.5 mM sulfo-NHS-SS-Biotin and incubate at 25 °C for
2 h.
CRITICAL STEP Sulfo-NHS-SS-Biotin is instable in aqueous solution. Prepare a 10 mM
sulfo-NHS-SS-Biotin solution (20x stock) in DMSO immediately prior to mixing with the
PICS sample. Follow manufacturer�’s instructions for storage and handling of sulfo-NHS-
SS-Biotin powder.
49. During this time, equilibrate high-capacity streptavidin-Sepharose (GE Healthcare) in 50
mM HEPES, 150 mM NaCl, pH 7.5. Use a slurry volume of 1.5-times the volume of the
PICS sample. This provides sufficient binding capacity for bound and unbound biotin in
PICS assays without further cleanup. Note that the commercially available high-capacity
streptavidin-Sepharose is provided as a 4-fold diluted slurry, so typically 1.2 ml diluted
slurry is used for each PICS assay. Buffer-equilibrate the slurry by at least 5 times repeated
centrifugation and resuspension in 50 mM HEPES, 150 mM NaCl, pH 7.5.
CRITICAL STEP If more than 0.5 mM sulfo-NHS-SS-Biotin were used in step 48, adjust the
volume of streptavidin-Sepharose slurry accordingly in order to provide enough binding
capacity for all biotin molecules.
50. Add buffer-equilibrated streptavidin-Sepharose to the biotinylated PICS sample. The
required volume of undiluted slurry is 1.5-times the volume of the PICS sample volume.
51. Incubate for 0.5 h at 22 °C. Apply mild agitation to keep the slurry in suspension.
PAUSE POINT The incubation time can be extended up to two days at 4 °C.
52. Pour slurry into a spin column of sufficient volume with a filter of ~10 m pore size. In a
typical PICS experiment, the total volume at this point is 550 600 l (200 l library + buffer
+ protease + sulfo-NHS-SS-Biotin solution + 300 l streptavidin-Sepharose).
53. Place column in a 2 ml reaction tube.
54. Centrifuge at 150 x g for 1 min. If the resin runs dry, re-apply flow-through and centrifuge
at a lower speed. If the supernatant has not entirely passed through resin, repeat
centrifugation.

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 15 of 21
55. Re-apply the first flow-through to the streptavidin-Sepharose resin and centrifuge a second
time.
56. Discard flow-through.
57. Wash resin with 500 l of 50 mM HEPES, 150 mM NaCl, pH 7.5.
58. Centrifuge (see step 54) and discard flow-through.
59. Repeat washing steps 57 and 58 ten times.
CRITICAL STEP Insufficient washing may lead to carry-over of uncleaved peptides
dramatically decreasing the number of cleavage site identifications.
60. Prepare fresh elution buffer: 50 mM HEPES, 20 mM DTT, pH 7.5.
61. Press bottom plug in column.
62. Apply 500 l of elution buffer and gently resuspend slurry with pipette tip.
63. Close column top with screw cap.
64. Incubate for 1 h at 25 °C with mild agitation.
65. Place column in a clean 2 ml reaction tube.
66. Remove bottom plug and loosen screw cap.
67. Centrifuge at 500 x g for 1 min and keep eluate.
68. Place column in a clean 2 ml reaction tube and apply 500 l of freshly prepared elution
buffer.
69. Centrifuge at 500 x g and keep eluate
70. Pool the two elution fractions.
71. Reversed-phase solid phase extraction of eluate using C18 cartridges or tips with peptide
binding capacity of 10 100 g. Proceed according to manufacturer�’s instructions.
72. Determine peptide concentration of C18 eluate by BCA assay.
LC–MS/MS AND DATA ANALYSIS
73. Analyze prime�–side cleavage products by LC-MS/MS. Standard LC-MS/MS settings for
peptide/proteome analysis are typically employed. Since these settings are machine
specific, they are outside the scope of this protocol.

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 16 of 21
74. Determine prime side sequences from LC MS/MS data using spectrum-to-sequence
assignment software such as Mascot 10 or X!Tandem 11. Set static modifications as
carboxyamidomethylation of cysteine residues (+57.02 Da), dimethylation of lysines
(+28.03 Da) and thioacylation of peptide N termini (+88.00 Da). Apply semi-style cleavage
searches since all enriched peptides are products of a double digestion with an N-terminal
cleavage by the test protease and a C-terminal cleavage by the library-generating
protease. Define tryptic specificity as cleavage C terminal to lysine or arginine and
chymotryptic specificity as cleavage C terminal to tryptophan, tyrosine, leucine and
phenylalanine. For GluC libraries, perform two separate searches: one with cleavage
C terminal to glutamate and one with cleavage C terminal to glutamate or aspartate. Allow
for up to three missed cleavages by the library-generating enzyme. This increases the
number of identified peptides because library peptides often retain potential cleavage sites
due to incomplete digest in step 22, especially in the case of GluC.
75. Use the PeptideProphet algorithm 12 of the Trans-Proteomic Pipeline 13 for secondary
validation of search results. Save PeptideProphet result as tab-delimited text file, typically
called �“interact.pep.xls�”.
76. In a web browser, navigate to http://clipserve.clip.ubc.ca/pics. This web service
automatically retrieves nonprime cleavage sequences, renders the list of cleavage sites
non redundant, provides graphical and tabular representation of subsite preferences, and
screens the cleavage sites for potential subsite cooperativity.
77. Follow instructions on the webpage. Choose the �“interact.pep.xls�” file for upload, enter the
name of the test protease without blank spaces, select digestion protease and proteome
source and select whether the dataset will be scanned for potential subsite cooperativity.
78. Click on the �“press�” button once and wait for the result page to appear. Data processing
can last several minutes.
79. Display results of PICS analysis. Follow the link �“Click here for your PICS results�” to display
the results of the PICS analysis as heatmap style graphics and tabulated values.
80. Obtain overview of potential subsite interactions. Follow the link �“Click here for subsite
cooperativity analysis results�” to get an overview of potential subsite interactions. Use

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 17 of 21
manual parsing to distinguish true subsite cooperativity from artifacts according to the
following rules: (a) amino acids with similar physicochemical properties (size, charge,
hydrophobicity) have similar effects on the positional preferences in the dependent subsite
while (b) amino acids with contrasting physicochemical properties have contrasting effects
on the positional preferences in the dependent subsite. (c) Lastly, changes in positional
preference should be interdependent (�“vice�–versa effect�”). For example, in HIV-1 protease,
large amino acids in P3 such as Leu and Phe increase the occurrence of small amino acids
in P1 such as Ala and Gly (and vice versa) while small amino acids in P3 decrease the
occurrence of small amino acids in P1 (and vice versa).
81. Download the results file. Follow the link �“Results (including html files) in one compressed
file for local use�” to download the results file for storage on a local computer.
82. Obtain sequence logo encompassing the cleavage site. Follow the link �“Cleavage sites for
logos�” to obtain a sequence logo encompassing the cleavage site. To generate an IceLogo
14, copy the cleavage site list from the displayed web page, and click on the external link
�“Link to IceLogo web page�” and follow the instructions.
TROUBLESHOOTING
Problem Solution
Viscous lysate when preparing the library Add DNAse I (1 g / mL final concentration) to
degrade DNA; a major cause of viscosity in cell
lysates. Be aware that DNAse I only has
marginal activity in lysis buffer since EDTA is
present. If necessary add 1 mM MgCl2 to
improve DNAse digestion.
Low number of cleavage products identified by
LC MS/MS
Control protease activity with an appropriate
assay
Check pH of sample
Use a higher protease:library ratio
Analyze data with non enzyme constrained
search

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 18 of 21
Use PICS library prepared with a different
digestion protease
PICS analysis server does not recognize
peptide list
In Microsoft Excel, save peptide list in text
format (Windows) and include header
�“peptides�”.
Selecting correct controls Possible control experiments include performing
a PICS assay and pull out using i) no protease
added during the incubation; ii) protease that is
inactivated through an active site mutation or by
application of a covalent inhibitor, or heat
denaturation of the protease before addition to
the library. Any identified cleavage products
represent background proteolysis, e.g. from
sample contamination. Few to none cleavage
products should be identified
TIMING
PICS library preparation: 3 days
PICS assay: 2 days
LC�–MS/MS analysis and PICS data handling: 1 day
For detailed timing see Figure 4.
ANTICIPATED RESULTS
One-dimensional LC�–MS/MS analysis of the peptide libraries should identify 100s of peptides, with
>90% of the peptides showing the expected cleavage site specificity of the protease used for
library generation at both the N- and C-terminus. Multidimensional LC�–MS/MS analysis of the
peptide libraries should identify 1,000s of peptides. The level of amine and thiol protection should
exceed 98%.

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 19 of 21
In a successful PICS experiment, 10s to 100s of cleavage site sequences representing the
specificity of the test protease are identified. The number of cleavage site identifications that are
required for a reliable specificity profile varies between proteases with strict specificity and
proteases with broad specificity. The canonical specificity of caspase 7 was accurately profiled with
as little as 37 cleavage site sequences 3. Proteases with broad specificity, such as MMPs, typically
require a larger number (> 100) of cleavage site sequences. Subsite cooperativity analysis typically
requires > 300 cleavage sites and should be verified in replicate experiments.
FIGURE LEGENDS
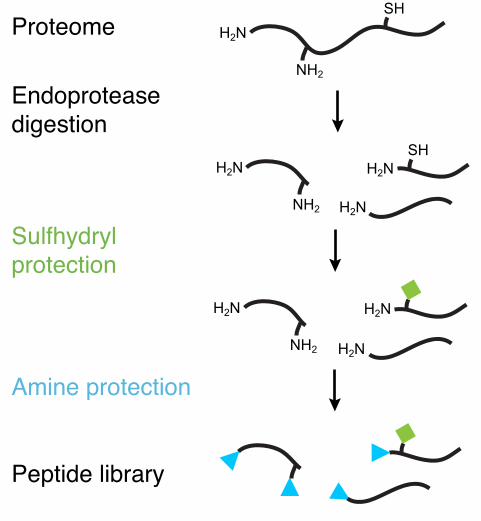
Figure 1: PICS library generation. Libraries are generated from cellular proteomes and thus
represent biological sequence diversity. Specific endoproteases such as Trypsin, GluC or
Chymotrypsin are used to digest proteomes into oligopepeptides. Then, thiols and primary amines
are chemically protected and the library is purified. See procedure text for details.
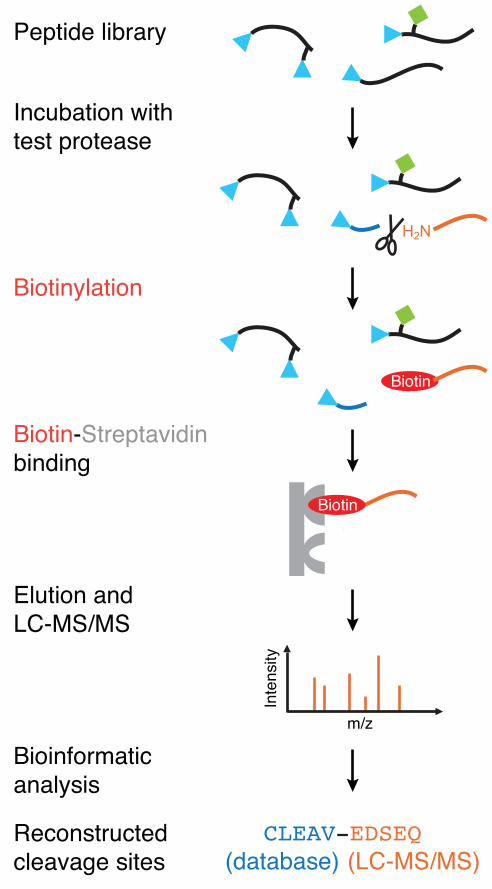
Figure 2: PICS specificity assay. PICS peptide libraries are incubated with a test endoprotease.
Prime�–side cleavage products possess free N�–termini that are tagged with a cleavable biotin,
allowing for specific isolation with immobilized streptavidin. Following elution, prime�–side cleavage
products are identified by LC�–MS/MS whereas the corresponding non�–prime sequences are
derived bionformatically through a web�–based service.
Figure 3: Comparison of MMP-2 specificity profiles from peptide-centric (PICS 3) and protein-
based (TAILS 8) approaches. Human or bacterial (E. coli) peptide libraries yield comparable
specificity profiles thereby validating the use of bacterial peptide libraries for PICS experiments
with eukaryotic enzymes. For the protein-centric experiment, MMP-2 was added in vitro to
concentrated cell conditioned medium to minimize secondary cleavage events, e.g. through
activation of further proteases by the cell. Both PICS and TAILS yield similar specificity profiles
illustrating that PICS profiling of a protease is useful to corroborate in vivo cleavage events.
Sequence logos were generated with iceLogo 14. While PICS has been shown to characterize

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 20 of 21
subsite preferences from P6 �– P6�’ 3, we present MMP�–2 specificity profiles from P4 �– P4�’ in order
to better focus on primary MMP-2 specificity determinants 3,7.
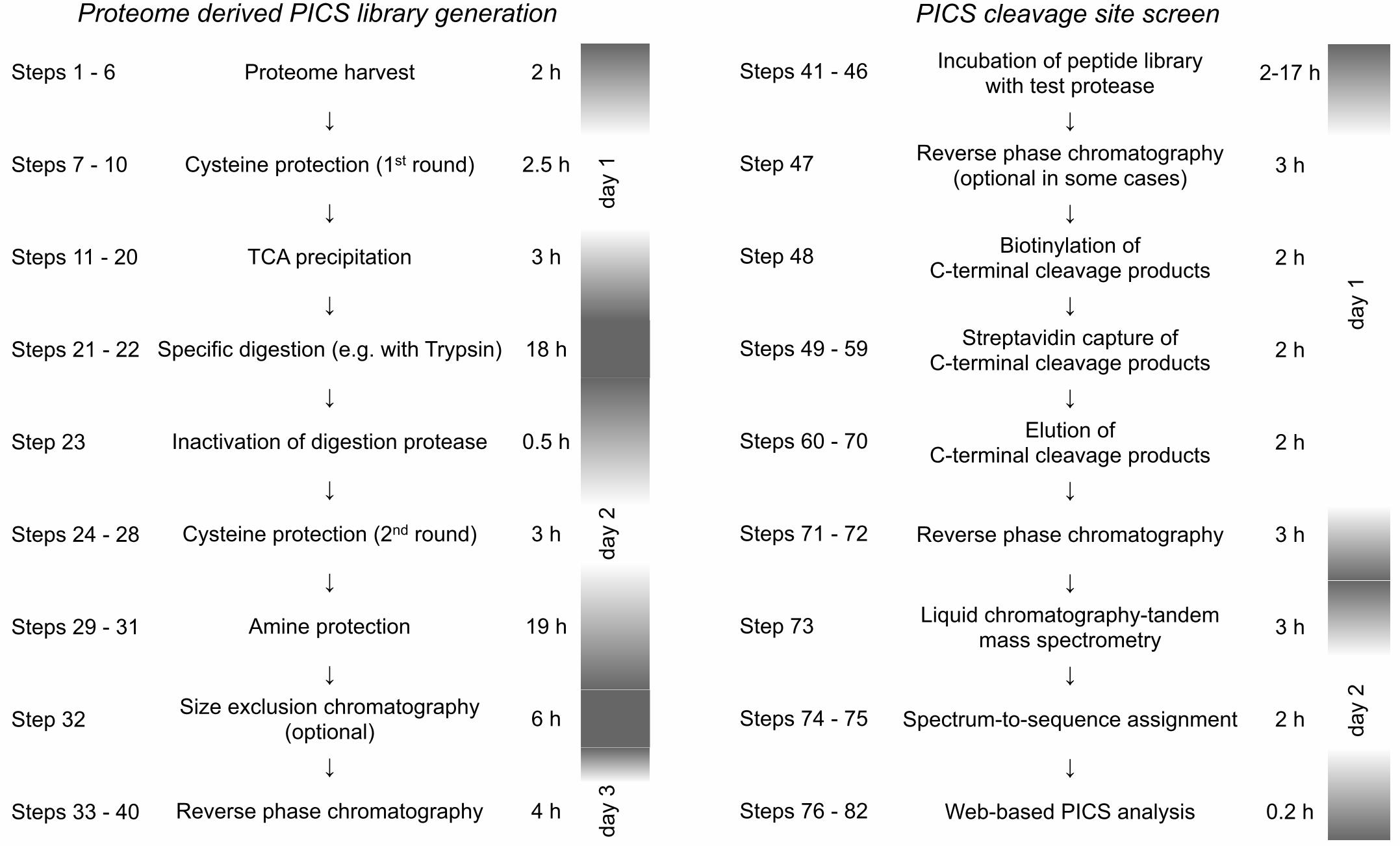
Figure 4: Timeline for PICS library generation and PICS specificity assay. Overall, PICS library
preparation and purification requires up to three days. A PICS specificity assay, including isolation
of cleavage products typically requires two days while LC�–MS/MS analysis and PICS data
handling requires a single day.
AUTHOR CONTRIBUTIONS STATEMENTS
OS and CMO designed and developed the PICS procedure. OS and UadK developed the web-
based analysis platform, PFH established bacterial libraries, and OS, PFH and OB optimized the
protocol. OS drafted the manuscript, OS and PFH designed the figures. All authors edited and
approved the manuscript.
ACKNOWLEDGMENTS
The authors thank Bettina Mayer for technical assistance in establishing the simplified PICS library
generation workflow. OS acknowledges support from the Deutsche Forschungsgemeinschaft
(DFG) (grants SCHI 871/1-1 and 871/2-1) and the Michael Smith Foundation for Health Research
(MSFHR). PFH is supported by Postdoctoral Fellowships of the German Academic Exchange
Service (DAAD) and the MSFHR. OB acknowledges support from the Swiss National Foundation
of Sciences (SNF) and the Canadian Institutes of Health Research (CIHR). CMO is supported by a
Canada Research Chair in Metalloproteinase Proteomics and Systems Biology. This work was
supported by a grant from the Canadian Institutes of Health Research (CIHR) and from a program
project grant in Breast Cancer Metastases from the Canadian Breast Cancer Research Alliance
(CBCRA) with funds from the Canadian Breast Cancer Foundation and the Cancer Research
Society as well as with an Infrastructure Grant from MSHFR.

Schilling et al. Protease Specificity Profiling With Proteome-derived Peptide Libraries
Page 21 of 21
COMPETING FINANCIAL INTERESTS
The authors declare no competing financial interests.
REFERENCES
1. Schechter, I. and Berger, A., On the active site of proteases. 3. Mapping the active site of papain; specific peptide inhibitors of papain Biochem Biophys Res Commun 32 (5), 898 (1968). 2. Schilling, O. and Overall, C. M., Proteomic discovery of protease substrates Curr. Opin. Chem. Biol. 11 (1), 36 (2007). 3. Schilling, O. and Overall, C. M., Proteome-derived, database-searchable peptide libraries for identifying protease cleavage sites Nat. Biotechnol. 26 (6), 685 (2008). 4. Auf dem Keller, U. and Schilling, O., Proteomic Techniques and Activity-based Probes for the System-wide Study of Proteolysis Biochimie in press (2010). 5. Matthews, D. J. and Wells, J. A., Substrate phage: selection of protease substrates by monovalent phage display Science 260 (5111), 1113 (1993). 6. Boulware, K. T. and Daugherty, P. S., Protease specificity determination by using cellular libraries of peptide substrates (CLiPS) Proc. Natl. Acad. Sci. U.S.A. 103 (20), 7583 (2006). 7. Turk, B. E., Huang, L. L., Piro, E. T., and Cantley, L. C., Determination of protease cleavage site motifs using mixture-based oriented peptide libraries. Nat. Biotechnol. 19 (7), 661 (2001). 8. Kleifeld, O. et al., Isotopic labeling of terminal amines in complex samples identifies protein N-termini and protease cleavage products Nat. Biotechnol. 28 (3), 281 (2010). 9. Sukuru, S. C. K. et al., A lead discovery strategy driven by a comprehensive analysis of proteases in the peptide substrate space Protein Sci, in press (2010). 10. Perkins, D. N., Pappin, D. J., Creasy, D. M., and Cottrell, J. S., Probability-based protein identification by searching sequence databases using mass spectrometry data Electrophoresis 20 (18), 3551 (1999). 11. Craig, R. and Beavis, R. C., TANDEM: matching proteins with tandem mass spectra Bioinformatics 20 (9), 1466 (2004). 12. Keller, A., Nesvizhskii, A. I., Kolker, E., and Aebersold, R., Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search Anal. Chem. 74 (20), 5383 (2002). 13. Pedrioli, P. G. A., Trans-proteomic pipeline: a pipeline for proteomic analysis Methods Mol. Biol. 604, 213 (2010). 14. Colaert, N., Helsens, K., Martens, L., Vandekerckhove, J., and Gevaert, K., Improved visualization of protein consensus sequences by iceLogo Nat. Methods 6 (11), 786 (2009).
SH
NH2
H2N
NH2
H2N
H2N
H2N
Proteome
Peptide library
Endoprotease digestion
Sulfhydryl protection
Amine protection
SH
NH2
H2N
H2N
H2N
Biotin
CLEAV-EDSEQ(database) (LC-MS/MS)
m/z
Inte
nsity
Peptide library
Incubation with test protease
Biotinylation
Biotin-Streptavidin binding
Elution and LC-MS/MS
Bioinformatic analysis
Reconstructedcleavage sites
H2N
Biotin

MMP-2 PICSHuman tryptic library
MMP-2 PICSE. coli tryptic library
P4 P3 P2 P1 P1’P2’P3’P4’
15
10
fold
enr
ichm
ent
over
nat
ural
abu
ndan
ce
0 P4 P3 P2 P1 P1’P2’P3’P4’
15
10
fold
enr
ichm
ent
over
nat
ural
abu
ndan
ce
0
MMP-2 TAILSHuman secreted proteins
P4 P3 P2 P1 P1’P2’P3’P4’
15
10
fold
enr
ichm
ent
over
nat
ural
abu
ndan
ce
0
Thursday, August 12, 2010
Proteome harvest 2 h
!
Cysteine protection (1st round) 2.5 h
!
TCA precipitation 3 h
!
Specific digestion (e.g. with Trypsin) 18 h
!
Inactivation of digestion protease 0.5 h
!
Cysteine protection (2nd round) 3 h
!
Amine protection 19 h
!Size exclusion chromatography
(optional) 6 h
!
Reverse phase chromatography 4 h
Proteome derived PICS library generation
day
1da
y 2
day
3
Incubation of peptide library with test protease 2-17 h
!Reverse phase chromatography
(optional in some cases) 3 h
!Biotinylation of
C-terminal cleavage products 2 h
!Streptavidin capture of
C-terminal cleavage products 2 h
!Elution of
C-terminal cleavage products 2 h
!
Reverse phase chromatography 3 h
!!Liquid chromatography-tandem
mass spectrometry 3 h
!
Spectrum-to-sequence assignment 2 h
! Web-based PICS analysis 0.2 h
PICS cleavage site screen
day
1da
y 2
Steps 1 - 6
Steps 7 - 10
Steps 11 - 20
Steps 21 - 22
Step 23
Steps 24 - 28
Steps 29 - 31
Step 32
Steps 33 - 40
Steps 41 - 46
Step 47
Step 48
Steps 49 - 59
Steps 60 - 70
Steps 71 - 72
Step 73
Steps 74 - 75
Steps 76 - 82
Saturday, October 9, 2010




















