
Gold Matters - EX4DEV19 – Artisanal and Small-Scale Mining ...
Upload
independentCategory
view
2download
0
Journal of Dairy Research (1997) 64 409–421 Printed in Great Britain 409
Characterization of the lactic acid bacteria in artisanal dairy
products
B TIMOTHY M. COGAN*, MANUELA BARBOSA†, ERIC BEUVIER‡,BRUNA BIANCHI-SALVADORI§, PIER S. COCCONCELLI¶, ISABEL
FERNANDES†, JESUS GOMEZs, ROSARIO GOMEZ**, GEORGEKALANTZOPOULOS††, ANTONIO LEDDA‡‡, MARGARITA MEDINAs,
MARY C. REA* EVA RODRIGUEZs
*National Dairy Products Research Centre, Teagasc, Fermoy, Irish Republic† Instituto Nacional de Engenharia e Tecnologia Industrial, Azinhaga dos Lameiros,
P-1699 Lisbon, Portugal‡ Institut National de la Recherche Agronomique, Station de Recherches en Technologie
et Analyses Laitie[ res, BP 89, F-39801 Poligny, France§Centro Sperimentale del Latte, Strada per Merlino 3, I-20060 Zelo Buon Persico
(Mi), Italia¶ Istituto di Microbiologia, Universita[ Cattolica del Sacro Cuore, Via Emilia
Parmense, 84, I-29100 Piacenza, Italias Instituto Nacional de InvestigacioU n y TecnologıUa AgrarıUa y Alimentaria,
Apartado 8111, E-28080 Madrid, Espanh a** Instituto del Frio, Ciudad Universitaria, E-28040 Madrid, Espanh a
††Agricultural University of Athens, Iera Odos 75, 118 55 Athens, Greece‡‡ Istituto Zootecnico Caseario per la Sardegna, I-07040 Olmedo (Sassari), Sardinia,
Italia
(Received 20 December 1995 and accepted for publication 20 December 1996 )
S. In all, 4379 isolates from 35 products, including 24 artisanal cheeses, weresurveyed with a view to identifying strains that could be used as starters incommercial dairy fermentations. Of the isolates, 38% were classified as Lactococcus,17% as Enterococcus, 14% as Streptococcus thermophilus, 12% as mesophilicLactobacillus, 10% as Leuconostoc and 9% as thermophilic Lactobacillus. Acidproduction by the isolates varied considerably. Of the 1582 isolates of Lactococcusand 482 isolates of mesophilic Lactobacillus tested, only 8 and 2% respectivelyproduced sufficient acid to lower the pH of milk to ! 5±3 in 6 h at 30 °C. In contrast,53, 32 and 13% of Str. thermophilus, thermophilic Lactobacillus and Enterococcusisolates respectively reduced the pH to 5±3. These isolates were found only in someFrench, Italian and Greek cheeses. Bacteriocins were produced by 11% of the 2257isolates tested and 26 of them produced broad-spectrum bacteriocins which inhibitedat least eight of the ten target strains used, which included lactic acid bacteria,clostridia and Listeria innocua. The most proteolytic of the 2469 isolates tested wereStr. thermophilus from Fontina cheese followed by Enterococcus from Fiore Sardo andToma cheese and thermophilic Lactobacillus from all sources. Exopolysaccharideswere produced by 5±3% of the 2224 isolates tested.
In many Southern European countries cheeses are made from cows’, goats’, ewes’

410 T. M. C
and buffalo milk by farmers and shepherds on a small scale in the farmhouse or dairyusing traditional techniques. In this paper these cheeses will be referred to as‘artisanal ’. Commercial starter cultures are not normally used in making thesecheeses. Instead the cheesemaker relies on the lactic acid bacteria (LAB) naturallypresent in the milk, as adventitious contaminants, to grow and produce the lacticacid required for expulsion of the whey (moisture) from the coagulum duringcheesemaking. Alternatively, different types of natural cultures are used which areproduced by incubating milk or whey from the previous day under definedconditions. With some notable exceptions, e.g. Manchego, Cabrales, Maho! n andMajorero cheese (Nun4 ez & Martı!nez-Moreno, 1976; Nun4 ez, 1978; Sua! rez et al. 1983;Fontecha et al. 1990), little is known about the LAB involved in artisanal cheeses.Manchego, Cabrales, Maho! n and Majorero cheese usually contain high numbers oflactococci, leuconostocs, mesophilic lactobacilli and enterococci.
Proteolytic activity and lactic acid, bacteriocin and exopolysaccharide (EPS)production are important attributes of the starter bacteria used in commercialcheesemaking. Milk is deficient in many of the amino acids required by starterbacteria for growth. Consequently these bacteria must have a proteolytic system tohydrolyse the milk proteins to the amino acids and peptides required for growth.Proteolysis is also important in the development of flavour in cheese. The ability toproduce acid rapidly is probably the most important property of starter bacteria. Ithelps to reduce the pH which in turn increases the expulsion of whey from the curdand reduces the moisture content. The low pH and low moisture are important inreducing the propensity of the cheese for microbial spoilage. Bacteriocin productionby the starter LAB may also be important in this regard. Broad-spectrumbacteriocins produced by GRAS (generally regarded as safe) organisms such as LABhave potentially wide uses in the food industry especially if they are heat and acidstable and active at the neutral or slightly acidic pH values of many foods (for areview see DeVuyst & Vandamme, 1994). EPS production is not an importantproperty of the starters used in cheese manufacture but it is important in those thatare used to make fermented milks where the EPS increases the body and texture ofthe product.
This paper is a report on the identification of a large number of LAB, isolatedfrom artisanal products, to genus level and their characterization in terms of thecommercially important properties outlined above. Such strains should be useful inextending the number of commercially available cultures and in forming the basis ofnew starter cultures for the manufacture of fermented dairy products.
Origin of samples
Altogether 35 different products were used, including 24 cows’, ewes’ or goats’milk cheeses, 6 natural starters, 2 fermented milks and cows’, ewes’ and goats’ milk.The origin and the number of samples tested are shown in Table 1.
Isolation of strains
Thermophilic lactobacilli and streptococci were isolated on milk (100 g}l) agarcontaining 0±2 g bromcresol purple}l. The spread plate technique with anaerobicincubation at 42 °C was used for the lactobacilli and the pour plate technique withaerobic incubation at 42 °C for Streptococcus thermophilus. Mesophilic strains wereisolated on KCA agar (Nickels & Leesment, 1964), KCA agar containing X-Gal

Lactic acid bacteria in artisanal cheeses 411
Table 1. Location, source, age and number of products sampled in this study
Product Type of product Type of milk Source Laboratory
No. oflocationssampled
No. ofsamplestested
Age ofsamples,
d
Kefir Fermented milk Cow Ireland DPC 33 33 3Sa4 o Jorge Cheese Cow Azores DPC 5 5 4–5Sa4 o Joa4 o Cheese Cow Azores DPC 1 4 4–5Fresh Greek Cheese Ewe Greece AUA 1 11 5cheese
Kasseri Cheese Ewe Greece AUA 1 30 9–120Feta Cheese Ewe Greece AUA 2 7 2–365Galotyri Cheese Ewe Greece AUA 1 3 9, 30Scotta innesto Natural starter Ewe Sardinia IZCS 10 10 1Gioddu Yogurt Ewe Sardinia IZCS 4 4 2Fiore Sardo Cheese Ewe Sardinia IZCS 6 8 1, 130Casu axedu Cheese Goat Sardinia IZCS 10 10 2Mozzarella Natural starter Cow Italy CSL 1 2 1–3Caciotta Natural starter Cow Italy CSL 1 1 1–3Grana Natural starter Cow Italy CSL 1 4 1–2Pecorino Natural starter Ewe Italy CSL 1 3 1–3Caciotta Cheese Cow Italy CSL 1 6 1–30Fontina Cheese Cow Italy UCSC 1 11 1–2Toma Cheese Cow Italy UCSC 1 4 1–2Val Itelvi Cheese Cow Italy UCSC 1 2 1–2Serra da Estrela Cheese Ewe Portugal INETI 6 33 30–40Milk Ewe Portugal INETI 24 31 1–2Manchego Cheese Ewe Spain INIA 17 17 1–2Gredos Cheese Goat Spain INIA 12 15 1–2Majorero Cheese Goat Spain IFPL 4 8 2, 30Milk Goat Spain IFPL 2 4 1–2Fresh French Cheese Cow France INRA 25 40 1–8cheese
Munster Cheese Cow France INRA 2 4 1Reblochon-type Cheese Cow France INRA 1 2 1Vacherin-type Cheese Cow France INRA 1 1 1Chevret Cheese Cow France INRA 1 3 1Tomme Cheese Cow, ewe, goat France INRA 18 33 1–100Morbier Cheese Cow France INRA 1 1 120Comte! Cheese Cow France INRA 6 9 1Comte! Natural starter Cow France INRA 10 14 1Milk Cow France INRA 16 16 1
DPC, National Dairy Products Research Centre, Teagasc, Fermoy, Irish Republic ; AUA, AgriculturalUniversity of Athens, Greece; IZCS, Istituto Zootecnico e Caseario per la Sardegna, Olmedo, Sardinia, Italy;CSL, Centro Sperimentale del Latte, Zelo Buon Persico (MI) Italy; UCSC, Universita' Cattolica del Sacro Cuore,Piacenza, Italy; INETI, Instituto Nacional de Engenharia e Tecnologia Industrial, Lisbon, Portugal ; INIA,Instituto Nacional de Investigacio! n y Tecnologia Agraria y Alimentaria, Madrid, Spain; IFPL, Instituto delFrio, Consejo Superior de Investigaciones Cientificas, Ciudad Universitaria, Madrid, Spain; INRA, InstitutNational de la Recherche Agronomique, Poligny, France.
(Vogensen et al. 1987) or LM17 agar (Terzaghi & Sandine, 1975) at 30 °C. All strainswere stored frozen at ®80 °C in the laboratories in which they were isolated.
Identification
Thermophilic strains were grown in MRS broth (De Man et al. 1960) andmesophilic strains in LM17 broth. All strains were tested for their Gram reaction,catalase using 3 g H
#O
#}l, and shape by phase contrast microscopy of overnight
cultures. Growth of the coccal-shaped organisms was examined in LM17 afterincubation at 10 °C for 7 d, at 45 °C for 2 d, and in LM17 containing 20, 40 and65 g NaCl}l after incubation at 30 °C for 2–3 d (mesophiles) and in LM17 containing65 g NaCl}l after incubation at 42 °C for 2 d (thermophiles). Growth of the rod-

412 T. M. C
Table 2. Tests used in presumptive identification of the isolates in this study
Growth in NaCl of Growth at
Shape Heterofermentation 20 g}l 40 g}l 65 g}l 10 °C 15 °C 45 °C Identification
Cocci ® }® ® ND ® LactococcusCocci }® ND ND ® LeuconostocCocci ® ND EnterococcusCocci ND ® ® ® ND Streptococcus thermophilusRods ND ND ND ND ND ® Thermophilic LactobacillusRods }® ND ® Mesophilic Lactobacillus
, Positive; ®, negative; }®, positive or negative; ND, not determined.
shaped organisms was examined in MRS at 15 and 45 °C after 5 and 2 d respectively.All mesophilic strains were tested for heterofermentation or homofermentation ofsugars by the procedure of Abd-el-Malek & Gibson (1948). Based on these results, allGram-positive, catalase-negative strains were presumptively classified to genus levelaccording to the outline shown in Table 2.
Acid production
This was determined in heat-treated (5 min at 121 °C) reconstituted skim milk(RSM, 100 g}l) from each laboratory or in Nilac skim milk, (NIZO, NL-6710 Ede,The Netherlands). A 10 ml}l inoculum was used with incubation for 6 h at 30 °C formesophiles and 42 °C for the thermophiles. In this regard, enterococci wereconsidered to be thermophiles.
Proteolysis
This was measured by the o-phthaldialdehyde method (Church et al. 1983) afterincubation in RSM at 30 °C (mesophiles) or 42 °C (thermophiles) for 24 h. The resultswere calculated as the increase in A
$%!over the uninoculated control.
Bacteriocin production
This was measured by the overlay (O) and agar diffusion (AD) techniques. In theO technique, spots (10 µl) of log phase cells were spotted on the appropriate agar(MRS or LM17) and, after overnight growth at the appropriate temperature,overlaid with 5 ml of the target strain (C 10& cfu}ml) in sloppy agar (7±5 g}l). Theplates were incubated at the temperature appropriate for the target strain for 2–3 dand the diameter of the zone of inhibition measured. For the AD technique, the teststrain, grown in the appropriate broth, was centrifuged and the supernatantadjusted to pH 6±5 before filtration (Millex GV; Millipore, Bedford, MA 01730, USA).The target organism was seeded into the appropriate medium (C 10& cfu}ml). Wells(diam. 6 mm) were cut in the solidified agar and filled with 50 µl of test solution. Theplates were held at 4 °C for 2 h to allow diffusion of the bacteriocin into the agarbefore incubating them at the optimum temperature of the target strain for 1–2 d.
The target (indicator) organisms used were Lactobacillus casei ATCC 334, Lb.helveticus ATCC 15009, Lb. reuteri DSM 20016, Pediococcus pentosaceus FBB 63,Lactococcus lactis subsp. cremoris CNRZ 117, Listeria innocua BL 86}26, Str.thermophilus ST20, Enterococcus faecalis EF1, Clostridium sporogenes C22}10, and Cl.tyrobutyricum NCDO 1754. All of these were provided to each author by Dr P. S.Cocconcelli, Istituto di Microbiologia, Universita' Cattolica del Sacro Cuore, I-29100Piacenza, Italy. These strains have also been used to screen bacteriocin productionin the FLAIR and BRIDGE programmes of the EU. Str. thermophilus ST112 was

Lactic acid bacteria in artisanal cheeses 413
used instead of Str. thermophilus ST20 in some laboratories while Ln. mesenteroidesDB 1275 and Lc. lactis subsp. lactis IFPL 186 were also used in the IstitutoZootecnico e Caseario per la Sardegna and the Instituto del Frio respectively. Cl.sporogenes NCFB 1755 and Cl. tyrobutyricum NCFB 1791 were used instead of thetarget clostridia at the National Dairy Products Research Centre.
Polysaccharide formation
EPS production from lactose was determined by qualitatively measuring thedegree of ‘stringiness’ of cultures grown in RSM at 30 °C (mesophiles) or 42 °C(thermophiles) for 18 h. A positive result was recorded if the coagulated culture couldbe teased into a string with an inoculating loop. Strains that failed to coagulate milkin 18 h were rechecked in RSM containing yeast extract (3 g}l).
Characterization
Handling large numbers of isolates meant that relatively few tests would be doneon each one. The tests outlined in Table 2 can discriminate adequately between mostof the common genera of starter bacteria but they do not unequivocally distinguishbetween Enterococcus and Lactococcus since Ec. dispar and Ec. sulfureus do not growat 45 °C (Devriese et al. 1993) and some Lactococcus strains can grow in 65 g NaCl}l(Facklam & Collins, 1989; Pot et al. 1994, 1996); thus Lactococcus and Enterococcuscould be confused. However, it is likely that Ec. faecalis, Ec. faecium, Lc. lactis andLc. raffinolactis would be the commonest species of enterococci and lactococci foundin milk and cheese. These enterococci and lactococci would be distinguished fromeach other at the genus level by their growth responses at 45 °C and in 65 g NaCl}l(Table 2).
A total of 4379 isolates from 35 different sources were classified (Table 3). Mostof the cheeses examined were relatively young and would, therefore, be dominatedby the bacteria responsible for acid production. Most strains were identified asLactococcus (38%) followed in order by Enterococcus (17%), Str. thermophilus (14%),mesophilic Lactobacillus (12%), Leuconostoc (10%) and thermophilic Lactobacillus(9%). Only 26 (! 0±6%) of the strains could not be characterized by the tests usedand were considered to be atypical. For the purposes of this paper, Enterococcusstrains are considered to be thermophilic.
Strains from each of the above groups of LAB, except Leuconostoc, were found inKasseri, Feta and fresh cheese from Greece, in Tomme cheese from France, and inFontina and Toma cheese from Italy. Manchego, Gredos and Majorero cheese fromSpain and Serra da Estrela cheese from Portugal contained Lactococcus, Leuconostoc,mesophilic Lactobacillus and Enterococcus, which confirms previous results (Nun4 ez &Martı!nez-Moreno, 1976; Nun4 ez, 1978; Sua! rez et al. 1983; Fontecha et al. 1990).
Lactococcus strains were isolated from kefir, natural Mozzarella starter and all 24cheeses, except Galotyri, Caciotta, Morbier and Comte! (Table 3). Many of these alsocontained thermophilic LAB. The presence of mesophilic and thermophilic strains inthe same product is not unusual in artisanal cheeses, which are made withoutcommercial starters.
Mesophilic lactobacilli were isolated from 19 of the 35 products but in smallnumbers from most of them. However, they were a major component of themicroflora of Kasseri, Feta, Serra da Estrela, Gredos and Majorero cheese. Except forKasseri, large numbers of lactococci were also isolated from these cheeses. Cheese is

414 T. M. C
Table
3.Sou
rce
and
iden
tifica
tion
ofth
eis
olate
sfr
omart
isanaldair
ypro
duct
sto
genus
orsp
ecie
sle
vel
Pro
duct
Lact
ococ
cus
Leu
conos
toc
Ente
roco
ccus
Str
epto
cocc
us
ther
mop
hilus
Aty
pic
al
cocc
iM
esophilic
Lact
obaci
llus
Ther
mophilic
Lact
obaci
llus
Aty
pic
al
rods
Tota
ls
Kefi
r361
41
402
Sa4 o
Jorg
ech
eese
238
47
1286
Sa4 o
Joa4 o
chee
se11
11
Fre
shG
reek
chee
se17
36
55
310
121
Kass
erich
eese
590
90
47
60
292
Fet
ach
eese
100
28
324
4159
Galo
tyri
chee
se3
3Sco
tta
innes
to2
93
54
149
Gio
ddu
32
55
87
Fio
reSard
och
eese
17
30
47
Casu
axed
uch
eese
73
11
13
3100
Mozz
are
lla,natu
ralst
art
er57
11
41
8108
Caci
ott
a,natu
ralst
art
er30
30
Gra
na,natu
ralst
art
er73
73
Pec
ori
no,natu
ralst
art
er12
61
73
Caci
ott
a,ch
eese
106
106
Fonti
na
chee
se9
67
45
19
40
180
Tom
ach
eese
534
93
17
68
ValIt
elvich
eese
4152
77
10
243
Ser
rada
Est
rela
chee
se24
85
172
182
Ser
rada
Est
rela
milk
35
60
83
178
Manch
ego
chee
se169
101
59
31
360
Gre
dos
chee
se186
58
74
55
373
Majo
rero
chee
se47
11
74
10
107
249
Majo
rero
milk
48
20
32
100
Fre
shF
rench
chee
se105
19
12
61
2145
Munst
erch
eese
96
11
21
20
Reb
loch
on-t
ype
chee
se8
8V
ach
erin
-type
chee
se5
21
8Chev
ret
chee
se12
31
117
Tom
me
chee
se72
10
17
25
19
2127
Morb
ier
chee
se4
4Com
te! c
hee
se5
49
Com
te! n
atu
ralst
art
er11
57
932
Com
te! m
ilk
21
15
229
Tota
l1649
442
724
608
24
527
403
24379

Lactic acid bacteria in artisanal cheeses 415
an ideal medium for the growth of mesophilic lactobacilli (Laleye et al. 1990;Peterson & Marshall, 1990; McSweeney et al. 1993, 1994), but whether they areinvolved in ripening is not clear.
Significant numbers of enterococci were found in fresh Greek, Kasseri, Feta,Fontina, Fiore Sardo, Toma, Val Itelvi, Manchego, Gredos and Majorero cheese.Enterococci were more prevalent in the Sardinian cheeses, Fiore Sardo and Casuaxedu, which had mainly a mesophilic flora, than in the other Sardinian products,Scotta innesto and Gioddu, which had a thermophilic flora. Scotta innesto is anatural starter and Gioddu is a ewes’ milk yogurt.
The presence of enterococci in cheese has been questioned since some strains cancause serious health problems in humans, especially endocarditis and urinary tractinfections. However, they have a positive effect on flavour development in many ofthese cheeses. Their isolation in such large numbers is not surprising since they arenormal inhabitants of the intestine from where they contaminate the udder and themilk. They have been found previously in high numbers in artisanal cheeses (Nun4 ez& Martı!nez-Moreno, 1976; Nun4 ez, 1978; Sua! rez et al. 1983; Fontecha et al. 1990) andsome workers have shown their usefulness in flavour formation in Cheddar cheese(Jensen et al. 1975). Enterococci have several properties (e.g. heat and salt tolerance)which make them ideal starter cultures and many of them are also good acidproducers. Their role in cheese ripening should be re-evaluated.
Acid production
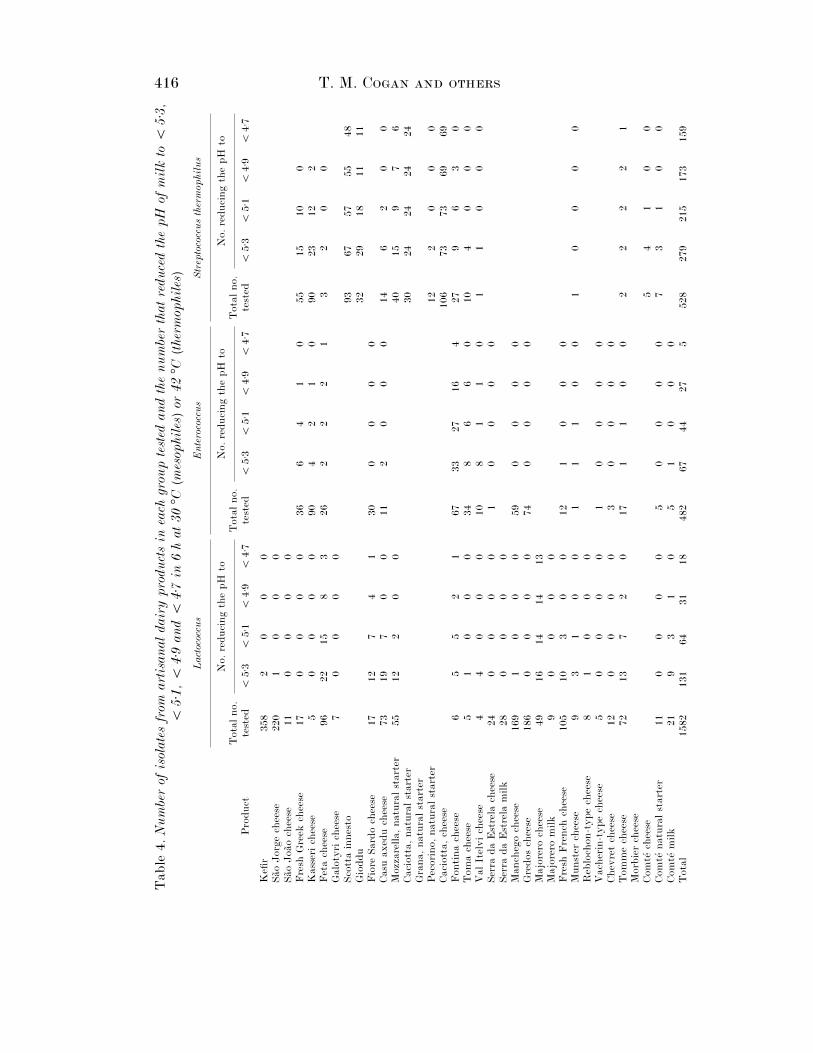
A good acid-producing starter culture will reduce the pH of milk from its normalvalue of C 6±6 to 5±3 in 6 h at 30 °C (mesophiles) or 42 °C (thermophiles) using aninoculum of 10 ml}l. Of the 442 Leuconostoc isolates tested for acid production onlysix reduced the pH of RSM below 5±9, five (one each from Sa4 o Jorge, Majorero andTomme cheese and two from ewes’ milk) to pH 5±5 and one (from ewes’ milk) to 5±4.Despite this leuconostocs are considered to play an important role in ripening of Serrada Estrela cheese. The acid-producing abilities of a further 3475 isolates aresummarized in Table 4. In interpreting these results and those in the rest of thisstudy it should be remembered that some isolates from the same source could beidentical.
The vast majority of Lactococcus isolates were poor acid producers: only 8±3% ofthe 1582 isolates tested reduced the pH of RSM to ! 5±3 (Table 4). This may bebecause lactococci that ferment lactose slowly contain both β-galactosidase (EC3.2.1.23) and phospho-β-galactosidase (EC 3.2.1.85), while those that ferment itrapidly contain only phospho-β-galactosidase (Farrow, 1980). Rapid acid-producingLactococcus were more prevalent in Mozzarella natural starter and Comte! milk andFeta, Fiore Sardo, Casu axedu, Fontina, Majorero and Tomme cheese than in theother cheeses. Some of these cheeses also contained thermophilic LAB. Within themesophilic Lactobacillus, only 2±1% of the isolates reduced the pH to ! 5±3 and mostof these were from Fontina cheese.
Rapid acid producers were much more prevalent in thermophilic than inmesophilic LAB, and were more common in some cheeses than in others. Forexample, large numbers of good, acid-producing Enterococcus were found particularlyin Fontina and Val Itelvi cheese while good acid-producing thermophilic Lactobacilluswere found in Scotta innesto, Gioddu and Fontina cheese and in natural Granastarters. In contrast, slow acid-producing thermophilic Lactobacillus were found inKasseri and Toma cheese and Mozzarella and Pecorino natural starters. Good acid-producing Str. thermophilus were common in all cheeses in which they were found.
416 T. M. C
Table
4.N
um
ber
ofis
olate
sfr
omart
isanaldair
ypro
duct
sin
each
grou
pte
sted
and
the
num
ber
thatre
duce
dth
epH
ofm
ilk
to!
5±3
,!
5±1
,!
4±9
and
!4±7
in6
hat30
°C(m
esop
hiles
)or
42
°C(ther
mop
hiles
)
Lact
ococ
cus
Ente
roco
ccus
Str
epto
cocc
us
ther
mop
hilus
Tota
lno.
No.re
duci
ng
the
pH
toT
ota
lno.
No.re
duci
ng
the
pH
toT
ota
lno.
No.re
duci
ng
the
pH
to
Pro
duct
test
ed!
5±3
!5±1
!4±9
!4±7
test
ed!
5±3
!5±1
!4±9
!4±7
test
ed!
5±3
!5±1
!4±9
!4±7
Kefi
r358
20
00
Sa4 o
Jorg
ech
eese
220
10
00
Sa4 o
Joa4 o
chee
se11
00
00
Fre
shG
reek
chee
se17
00
00
36
64
10
55
15
10
0K
ass
erich
eese
50
00
090
42
10
90
23
12
2F
eta
chee
se96
22
15
83
26
22
21
32
00
Galo
tyri
chee
se7
00
00
Sco
tta
innes
to93
67
57
55
48
Gio
ddu
32
29
18
11
11
Fio
reSard
och
eese
17
12
74
130
00
00
Casu
axed
uch
eese
73
19
70
011
20
00
14
62
00
Mozz
are
lla,natu
ralst
art
er55
12
20
040
15
97
6Caci
ott
a,natu
ralst
art
er30
24
24
24
24
Gra
na,natu
ralst
art
erP
ecori
no,natu
ralst
art
er12
20
00
Caci
ott
a,ch
eese
106
73
73
69
69
Fonti
na
chee
se6
55
21
67
33
27
16
427
96
30
Tom
ach
eese
51
00
034
86
60
10
40
00
ValIt
elvich
eese
44
00
010
81
10
11
00
0Ser
rada
Est
rela
chee
se24
00
00
10
00
0Ser
rada
Est
rela
milk
28
00
00
Manch
ego
chee
se169
10
00
59
00
00
Gre
dos
chee
se186
00
00
74
00
00
Majo
rero
chee
se49
16
14
14
13
Majo
rero
milk
90
00
0F
resh
Fre
nch
chee
se105
10
30
012
10
00
Munst
erch
eese
93
10
01
11
00
10
00
0R
eblo
chon-t
ype
chee
se8
10
00
Vach
erin
-type
chee
se5
00
00
10
00
0Chev
ret
chee
se12
00
00
30
00
0T
om
me
chee
se72
13
72
017
11
00
22
22
1M
orb
ier
chee
seCom
te! c
hee
se5
41
00
Com
te! n
atu
ralst
art
er11
00
00
50
00
07
31
00
Com
te! m
ilk
21
93
10
51
00
0T
ota
l1582
131
64
31
18
482
67
44
27
5528
279
215
173
159

Lactic acid bacteria in artisanal cheeses 417
Ther
mophilic
Lact
obaci
llus
Mes
ophilic
Lact
obaci
llus
Tota
lno.
No.re
duci
ng
the
pH
toT
ota
lno.
No.re
duci
ng
the
pH
to
Pro
duct
test
ed!
5±3
!5±1
!4±9
!4±7
test
ed!
5±3
!5±1
!4±9
!4±7
Kefi
rSa4 o
Jorg
ech
eese
Sa4 o
Joa4 o
chee
seF
resh
Gre
ekch
eese
10
43
13
00
00
Kass
erich
eese
60
21
047
00
00
Fet
ach
eese
41
10
11
00
00
Galo
tyri
chee
seSco
tta
innes
to54
40
34
34
33
Gio
ddu
55
45
21
10
7F
iore
Sard
och
eese
Casu
axed
uch
eese
32
22
213
00
00
Mozz
are
lla,natu
ralst
art
er8
00
00
Caci
ott
a,natu
ralst
art
erG
rana,natu
ralst
art
er73
17
80
0P
ecori
no,natu
ralst
art
er61
00
00
Caci
ott
a,ch
eese
Fonti
na
chee
se41
15
10
71
17
55
30
Tom
ach
eese
17
00
00
31
10
0V
alIt
elvich
eese
11
00
0Ser
rada
Est
rela
chee
se72
00
00
Ser
rada
Est
rela
milk
83
00
00
Manch
ego
chee
se31
00
00
Gre
dos
chee
se55
00
00
Majo
rero
chee
se108
31
10
Majo
rero
milk
10
00
00
Fre
shF
rench
chee
se1
00
00
Munst
erch
eese
10
00
0R
eblo
chon-t
ype
chee
seV
ach
erin
-type
chee
seChev
ret
chee
se1
00
00
Tom
me
chee
se1
10
00
19
00
00
Morb
ier
chee
se1
00
00
40
00
0Com
te! c
hee
se4
11
00
Com
te! n
atu
ralst
art
er9
10
00
Com
te! m
ilk
20
00
0T
ota
l401
129
81
54
43
482
10
74
0

418 T. M. C
The reason that good acid producers were not isolated uniformly from all cheeses isnot clear, but the results suggest that each cheese is a unique ecosystem.
Bacteriocin production
The O and AD methods were used to monitor bacteriocin production. The Otechnique was used in the National Dairy Products Research Centre, CentroSperimentale del Latte and INRA Poligny, and the AD technique in the Universita'Cattolica del Sacro Cuoro, the Agricultural University of Athens and the IstitutoZootecnico e Caseario per la Sardegna. Both techniques were used at the Instituto delFrio, Instituto Nacional de Engenharia e Tecnologia Industrial and InstitutoNacional de Investigacio! n y Tecnologia Agraria y Alimentaria. The O method iseasier to use where large numbers of strains have to be screened, but H
#O
#and lactic
acid can give false positives. In the AD method, neutralized supernatants were used,which overcame the problems due to lactic acid. A preliminary trial at InstitutoNacional de Investigacio! n y Tecnologia Agraria y Alimentaria and Instituto del Frioshowed that the number of bacteriocin-producing strains was reduced from 579 bythe O technique to 12 when the AD technique was used. Thus the latter techniqueis better in determining bacteriocin production despite the fact that it is morelaborious. These results suggest that few strains are genuine bacteriocin producersunder the conditions of the assay and that the numbers of bacteriocin-producingstrains checked by the O method may be overestimated. In the O technique used inthe National Dairy Products Research Centre, 10 µl proteinase K was spottedalongside the producer organism to eliminate nonspecific interference. Only strainsthat produced inhibitory zones that were inactivated by proteinase K (EC 3.4.21.14)were considered to be bacteriocin-producing.
Overall, 239 (11%) of the 2257 strains tested produced bacteriocins, most ofwhich were narrow-spectrum, inhibiting only one or two of the target strains, ormedium-spectrum, inhibiting three to seven of the target strains. A total of 26 broad-spectrum bacteriocins, which inhibited eight to ten of the target strains, wereidentified. Fifteen isolates (Lactococcus strains DPC 3147, DPC 3153, DPC 3178, DPC3205, DPC 3216, DPC 3244, DPC 3254, INRA 224, INRA 226 and INIA 515 andthermophilic Lactobacillus strains UCSC 20613, UCSC 3051, UCSC 3052, UCSC 3117and UCSC 3119) inhibited all ten target strains. The remaining eleven isolates haddifferent activities (Table 5). Broad-spectrum bacteriocins were found in all groupsof LAB except Str. thermophilus.
Little is known about the properties of the bacteriocins identified in the presentstudy but several are being actively studied. At least three different bacteriocinswere produced by the lactococcal isolates from kefir; a narrow-spectrum one thatinhibited only lactococci, a medium-spectrum one that inhibited Lb. casei, Lb.helveticus and Pd. pentosaceus and a broad-spectrum one that inhibited the ten targetstrains. That produced by DPC 3147 inhibited the ten target strains and numerousother strains of Lactococcus, Leuconostoc, Lactobacillus, Pediococcus, Str. thermophilusand Staphylococcus aureus (Ryan et al. 1996). This was not nisin, since it inhibited anisin producer. Production of the bacteriocin was plasmid encoded. It wasparticularly inhibitory to the non-starter LAB found in Cheddar cheese and is beingused as a tool to study the effect of non-starter LAB on cheese flavour. A novel phageresistance gene is also encoded on the same plasmid (Coakley et al. 1997). Strain IFPL105 (Table 5) has been identified as Lb. curvatus. The bacteriocin produced by thisstrain is heat-stable (partial inactivation occurs at 121 °C in 15 min), has a

Lactic acid bacteria in artisanal cheeses 419
Table
5.Sum
mary
ofth
eba
cter
ioci
n-p
roduci
ng
stra
ins
from
art
isanaldair
ypro
duct
sth
atin
hib
ited
eigh
tor
nin
eof
the
targ
etst
rain
s
Bact
erio
cin-p
roduci
ng
stra
in
Leu
conos
toc
Lact
obaci
llus
Ente
roco
ccus
Lact
ococ
cus
stra
ins
stra
ins
stra
ins
stra
ins
Targ
etst
rain
INR
A39
INR
A73
INR
A172
INR
A222
INR
A244
INR
A259
Ther
mophilic
CSL
9±4
Mes
ophilic
IFP
L105
INR
A296
INR
A307
INR
A309
Lact
obaci
llus
case
iA
TCC
334
®
®
®
Lb.
hel
veticu
sA
TCC
15009
Lb.
reute
riD
SM
20016
®
®
®
P
edio
cocc
us
pen
tosa
ceus
FB
B63
®
Lact
ococ
cus
lact
issu
bsp
.cr
emor
isCN
RZ
117
Lis
teri
ain
noc
ua
BL
86}2
6
®
Str
epto
cocc
us
ther
mop
hilus
ST
20
ND
ND
ND
ND
ND
ND
ND
ND
ND
Str
.th
erm
ophilus
ST
112
ND
ND
®
®E
nte
roco
ccus
faec
alis
EF
1
®
®
Clo
stri
diu
msp
orog
enes
C22}1
0
Cl.
tyro
buty
ricu
mN
CD
O1754
®
,In
hib
itio
n;®
,no
inhib
itio
n;N
D,not
det
erm
ined
.
Table
6.E
xop
olysa
cchari
de
pro
duct
ion
bydiff
eren
tis
olate
sof
lact
icaci
dba
cter
iafr
omart
isanalpro
duct
s
Lact
ococ
cus
Leu
conos
toc
Str
epto
cocc
us
ther
mop
hilus
Ther
mophilic
Lact
obaci
llus
Mes
ophilic
Lact
obaci
llus
Ente
roco
ccus
Labora
tory
Tota
lno.
test
edN
o.
posi
tive
Tota
lno.
test
edN
o.
posi
tive
Tota
lno.
test
edN
o.
posi
tive
Tota
lno.
test
edN
o.
posi
tive
Tota
lno.
test
edN
o.
posi
tive
Tota
lno.
test
edN
o.
posi
tive
INIA
355
16
159
086
0133
1A
UA
126
23
155
41
78
8157
10
CSL
57
11
0189
1142
01
0U
CSC
50
61
12
15
010
2D
PC
487
12
47
01
0IN
ET
I12
0T
ota
l1030
52
219
0350
43
232
991
0302
13
INIA
,In
stit
uto
Naci
onaldeIn
ves
tigaci
o! ny
Tec
nolo
gia
Agra
ria
yA
lim
enta
ria,M
adri
d,Spain
;A
UA
,A
gri
cult
ura
lU
niv
ersi
tyofA
then
s,G
reec
e;CSL
,Cen
tro
Sper
imen
tale
del
Latt
e,Zel
oB
uon
Per
sico
(MI)
,It
aly
;U
CSC,U
niv
ersi
ta'Catt
olica
del
Sacr
oCuore
,P
iace
nza
,It
aly
;D
PC,N
ati
onalD
air
yP
roduct
sR
esea
rch
Cen
tre,
Tea
gasc
,F
erm
oy,
Iris
hR
epublic;IN
ET
I,In
stit
uto
Naci
onalde
Engen
hari
ae
Tec
nolo
gia
Indust
rial,
Lis
bon,P
ort
ugal.

420 T. M. C
bacteriolytic mode of action, is plasmid encoded and inhibits several lactobacilli,enterococci, lactococci, pediococci, listeria, clostridia and bacilli (Casla et al. 1996).
Proteolysis
Another technologically important property of starter cultures is possession of aproteinase system. Proteinase activity is necessary for good growth of starterbacteria in milk and in hydrolysing casein during the ripening of cheese. Theproteolytic activity of 2469 strains was measured. Altogether 35 sets of correlationsbetween acid production and proteolysis were calculated for the different groups ofLAB from different products. Within the same group, the coefficients ofdetermination (r#) varied widely, e.g. from 0±00 to 0±49 for the lactococci, from 0±02to 0±59 for Str. thermophilus, from 0±00 to 0±06 for the thermophilic Lactobacillus andfrom 0±01 to 0±55 for Enterococcus (results not shown).
The most proteolytic organisms were Str. thermophilus strains from Fontinacheese followed by Enterococcus isolates from Fiore Sardo and Toma cheese andthermophilic lactobacilli from Grana, Gioddu, Toma, Scotta innesto and Fontina(results not shown). In addition, there was considerable variation in the proteolyticactivity within some bacterial groups from the same cheese, e.g. Str. thermophilusand the enterococci isolated from Toma and Fiore Sardo showed similar extremes.In contrast, there was relatively little variation in the proteolytic activity of thelactococci or the mesophilic or thermophilic lactobacilli, with the possible exceptionof the lactococcal isolates from Serra da Estrela (results not shown). The reason forthese contrasting results are not clear. However, they do suggest that strains of thesame genus or species isolated from the same source were different.
Exopolysaccharide production
Only 117 (5%) of the 2224 strains tested produced EPS (Table 6). This was notsurprising since EPS production, at least in lactococci, is normally plasmidassociated and is therefore easily lost on subculture.
Conclusions
Considerable variation was found in the types of LAB present in 35 Europeanartisanal products and in their ability to produce acid, EPS, bacteriocins andproteinases, suggesting that each cheese is a unique ecosystem. Some of these isolatesare potentially useful new starters for fermented dairy products and evaluation oftheir cheesemaking characteristics is currently being undertaken. In addition, theisolates producing broad spectrum bacteriocins are being studied to determine ifthese are novel bacteriocins and if they are of value in controlling the growth ofspoilage and pathogenic bacteria in foods.
The European Union is thanked for partly financing this project under ECLAIRcontract CT-91-0064. A data base of 3719 strains on FileMaker Pro for the PC isavailable from Dr P. S. Cocconcelli, Istituto de Microbiologia, Universita' CattolicaSacro Cuoro, I-29100 Piacenza, Italy.
A--M, Y. & G, T. 1948 Studies in the bacteriology of milk. I. The streptococci of milk. Journalof Dairy Research 15 233–248
C, D., R, T. & G! , R. 1996 Antimicrobial activity of lactic acid bacteria isolated from goat’smilk and artisanal cheeses : characteristics of a bacteriocin produced by Lactobacillus curvatus IFPL 105.Journal of Applied Bacteriology 81 35–41

Lactic acid bacteria in artisanal cheeses 421
C, F. C., S, H. E., P, D. H. & C, G. L. 1983 Spectrophotometric assay usingo-phthalaldehyde for determination of proteolysis in milk and isolated milk proteins. Journal of Dairy Science66 1219–1227
C, M., F, G. F. & R, R. P. 1997 Application and evaluation of the phage resistance- andbacteriocin-encoding plasmid pMRC01 for the improvement of dairy starter cultures. Applied andEnvironmental Microbiology 63 1434–1440
D M, J. C., R, M. & S, M. E. 1960 A medium for the cultivation of lactobacilli. Journal ofApplied Bacteriology 23 130–135
D, L. A., P, B. & C, M. D. 1993 Phenotypic identification of the genus Enterococcus anddifferentiation of phylogenetically distinct enterococcal species and species groups. Journal of AppliedBacteriology 75 399–408
DV, L. & V, E. J. 1994 Bacteriocins of Lactic Acid Bacteria: microbiology, genetics andapplications. London: Blackie
F, R. R. & C, M. D. 1989 Identification of Enterococcus species isolated from human infectionsby a conventional test scheme. Journal of Clinical Microbiology 27 731–734
F, J. A. E. 1980 Lactose hydrolysing enzymes in Streptococcus lactis and Streptococcus cremoris and alsoin some other species of streptococci. Journal of Applied Bacteriology 49 493–503
F, J., P! , C., J! , M., R, T., G! , C. & R, M. 1990 Biochemical andmicrobiological characteristics of artisanal hard goat’s cheese. Journal of Dairy Science 73 1150–1157
J, J. P., R, G. W., W, C. J. & V, E. R. 1975 Role of enterococci in Cheddarcheese: organoleptic considerations. Journal of Milk and Food Technology 38 142–145
L, L. C. S, R. E., L, B. H. & H, R. A. 1990 Quality attributes of Cheddar cheesecontaining added lactobacilli. Journal of Food Science 55 114–118
MS, P. L. H., F, P. F., L, J. A., J, K. N. & C, T. M. 1993 Contribution of theindigenous microflora to the maturation of Cheddar cheese. International Dairy Journal 3 613–634
MS, P. L. H., W, E. M., F, P. F., C, T. M., D, F. D. & C-G, M. 1994A procedure for the manufacture of Cheddar cheese under controlled bacteriological conditions and the effectof adjunct lactobacilli on cheese quality. Irish Journal of Agricultural and Food Research 33 183–192
N, C. & L, H. 1964 [Method for the differentiation and quantitative determination of starterbacteria.] Milchwissenschaft 19 374–378
N4 , M. 1978 Microflora of Cabrales cheese: changes during maturation. Journal of Dairy Research 45501–508
N4 , M. & M!-M, J. L. 1976 [Microbial flora of Manchego cheese. 1. Behaviour of themicrobial flora in artisanal Manchego cheese.] Anales del Instituto Nacional de Investigaciones Agrarias, SerieGeneral 4 11–31
P, S. D. & M, R. T. 1990 Non starter lactobacilli in Cheddar cheese. A review. Journal ofDairy Science 73 1395–1410
P, B., D, L. A., U, D., V, P., H, F. & K, K. 1996 Phenotypicidentification and differentiation of Lactococcus strains from animals. Systematic and Applied Microbiology 19213–222
P, B., L, W., K, K. & S, K.-H. 1994 Taxonomy of lactic acid bacteria. InBacteriocins of Lactic Acid Bacteria, pp. 13–90 (Eds L. deVuyst and E. J. Vandamme). London: Blackie
R, M. P., R, M. C., H, C. & R, R. P. 1996 An application in Cheddar cheese manufacture for astrain of Lactococcus lactis producing a novel broad-spectrum bacteriocin, lacticin 3147. Applied andEnvironmental Microbiology 62 612–619
S! , J. A., B, R. & I4 , B. 1983 Contribution to study of Maho! n cheese. III. Lactic acid bacteriaand enterococci. Chemie Mikrobiologie Technologie der Lebensmittel 8 52–56
T, B. E. & S, W. E. 1975 Improved medium for lactic streptococci and their bacteriophages.Applied Microbiology 29 807–813
V, F. K., K, T., L, J. J., K, B., E, D. & W N, E. W.1987 Improved direct differentiation between Leuconostoc cremoris, Streptococcus lactis subsp. diacetylactis,and Streptococcus cremoris}Streptococcus lactis on agar. Milchwissenschaft 42 646–648
Copyright © 2022 FDOKUMEN




















