
Enzymatic rotating biosensor for ciprofloxacin determination
Upload
independentCategory
view
4download
0
Gene 216 (1998) 255–265
Characterization of the enzymatic domains in the modular polyketidesynthase involved in rifamycin B biosynthesis by
Amycolatopsis mediterranei
Li Tang a,1, Yeo J. Yoon a, Cha-Yong Choi c, C. Richard Hutchinson a,b,*a School of Pharmacy, University of Wisconsin, Madison, WI 53706, USA
b Department of Bacteriology, University of Wisconsin, Madison, WI 53706, USAc Department of Chemical Technology, Seoul National University, Seoul, South Korea
Received 2 March 1998; received in revised form 5 June 1998; accepted 9 June 1998; Received by J. Wild
Abstract
Five clustered polyketide synthase (PKS) genes, rifA–rifE, involved in rifamycin (Rf ) biosynthesis in Amycolatopsis mediterraneiS699 have been cloned and sequenced (August, P.R. et al., 1998. Chem. Biol. 5, 69–79). The five multifunctional polypeptidesconstitute a type I modular PKS that contains ten modules, each responsible for a specific round of polyketide chain elongation.Sequence comparisons of the Rf PKS proteins with other prokaryotic modular PKSs elucidated the regions that have an importantrole in enzyme activity and specificity. The b-ketoacyl:acyl carrier protein synthase ( KS) domains show the highest degree ofsimilarity between themselves (86–90%) and to other PKSs (78–85%) among all the constituent domains. Both malonyl-coenzymeA (MCoA) and methylmalonyl-coenzyme A (mMCoA) are substrates for chain elongation steps carried out by the Rf PKS. Sinceacyltransferase (AT) domains of modular PKSs can distinguish between these two substrates, comparison of the sequence of allten AT domains of the Rf PKS with those found in the erythromycin (Er) (Donadio, S. and Katz, L., 1992. Gene 111, 51–60)and rapamycin (Rp) (Haydock, S. et al., 1995. FEBS Lett. 374, 246–248) PKSs revealed that the AT domains in module 2 ofRifA and module 9 of RifE are specific for MCoA, whereas the other eight modules specify mMCoA. Dehydration of the b-hydroxyacylthioester intermediates should occur during the reactions catalysed by module 4 of RifB and modules 9 and 10 ofRifE, yet only the active site region of module 4 conforms closely to the dehydratase (DH) motifs in the Er and Rp PKSs. TheDH domains of modules 9 and 10 diverge significantly from the consensus sequence defined by the Er and Rp PKSs, except forthe active site His residues. Deletions in the DH active sites of module 1 in RifA and module 5 in RifB and in the N- andC-terminal regions of module 8 of RifD should inactivate these domains, and module 2 of RifA lacks a DH domain, all of whichare consistent with the proposed biosynthesis of Rf. In contrast, module 6 of RifB and module 7 of RifC appear to contain intactDH domains even though DH activity is not apparently required in these modules. Module 2 of RifA lacks a b-ketoacyl:acylcarrier protein reductase (KR) domain and the one in module 3 has an apparently inactive NADPH binding motif, similar toone found in the Er PKS, while the other eight KR domains of the Rf PKS should be functional. These observations are consistentwith biosynthetic predictions. All the acyl carrier protein (ACP) domains, while clearly functional, nevertheless have active sitesignature sequences distinctive from those of the Er and Rp PKSs. Module 2 of RifA has only the core domains (KS, AT andACP). The starter unit ligase (SUL) and ACP domains present in the N-terminus of RifA direct the selection and loading of thestarter unit, 3-amino-5-hydroxybenzoic acid (AHBA), onto the PKS. AHBA is made by the products of several other genes inthe Rf cluster through a variant of the shikimate pathway (August, P.R. et al., inter alia). RifF, produced by the gene immediatelydownstream of rifE, is thought to catalyse the intramolecular cyclization of the PKS product, thereby forming the ansamacrolideprecursor of Rf B. © 1998 Elsevier Science B.V. All rights reserved.
Keywords: Modular PKS; Antibiotics; Sequence homology
* Corresponding author. Tel.: +1 608 2627582; Fax: +1 608 2623134; E-mail: [email protected] Li Tang and Yeo J. Yoon contributed equally to this work.
Abbreviations: aa, amino acid(s); ACP, acyl carrier protein; AHBA, 3-amino-5-hydroxybenzoic acid; AT, acyltransferase; CL, carboxylic acidcoenzyme A ligase; DEBS, 6-deoxyerythronolide B synthase; DH, dehydratase; Er, erythromycin; KR, b-ketoacyl ACP reductase; KS, b-ketoacylACP synthase; MCoA, malonyl-coenzyme A; mMCoA, methylmalonyl-coenzyme A; 6MSAS, 6-methylsalicylic acid synthase; PKS, polyketidesynthase; RAPS, rapamycin synthase; Rf, rifamycin(s); Rp, rapamycin; SUL, starter unit ligase.
0378-1119/98/$19.00 © 1998 Elsevier Science B.V. All rights reserved.PII: S0378-1119 ( 98 ) 00338-2

256 L. Tang et al. / Gene 216 (1998) 255–265
1. Introduction 2. Materials and methods
2.1. Cloning of the rif PKS genes and their disruptionRifamycins (Rf ) are naphthalenic ansamycin antibi-otics (Sensi et al., 1959) that strongly inhibit bacterial
The gene cloning and disruption experiments to obtainDNA-dependent RNA polymerases, thereby inhibitingthe Rf PKS genes and show that they are essential forprotein synthesis and cell growth, and also block theRf B biosynthesis were performed as described byaction of the RNA-dependent DNA polymerases ofAugust et al. (1998).retroviruses ( Wehrli and Staehelin, 1971; Wehrli, 1983).
They are major antibiotics for the treatment of tubercu-losis, leprosy and other mycobacterial infections 2.2. DNA sequencing and analysis(Mandell and Sande, 1980). Rf derivatives have alsoshown excellent activity against opportunistic pathogens Subclones derived from the original plasmids contain-in the AIDS complex of diseases ( Woodley and Kilburn, ing the rif PKS genes were prepared in pUC19 or1982; Dickinson and Mitchison, 1987), as well as against pGEM-3ZF(±) as described (August et al., 1998). EachHIV and related oncogenic viruses due to the inhibition DNA segment was sequenced independently on bothof the viral reverse transcriptase. strands using the Perkin Elmer Amplitaq Dye-
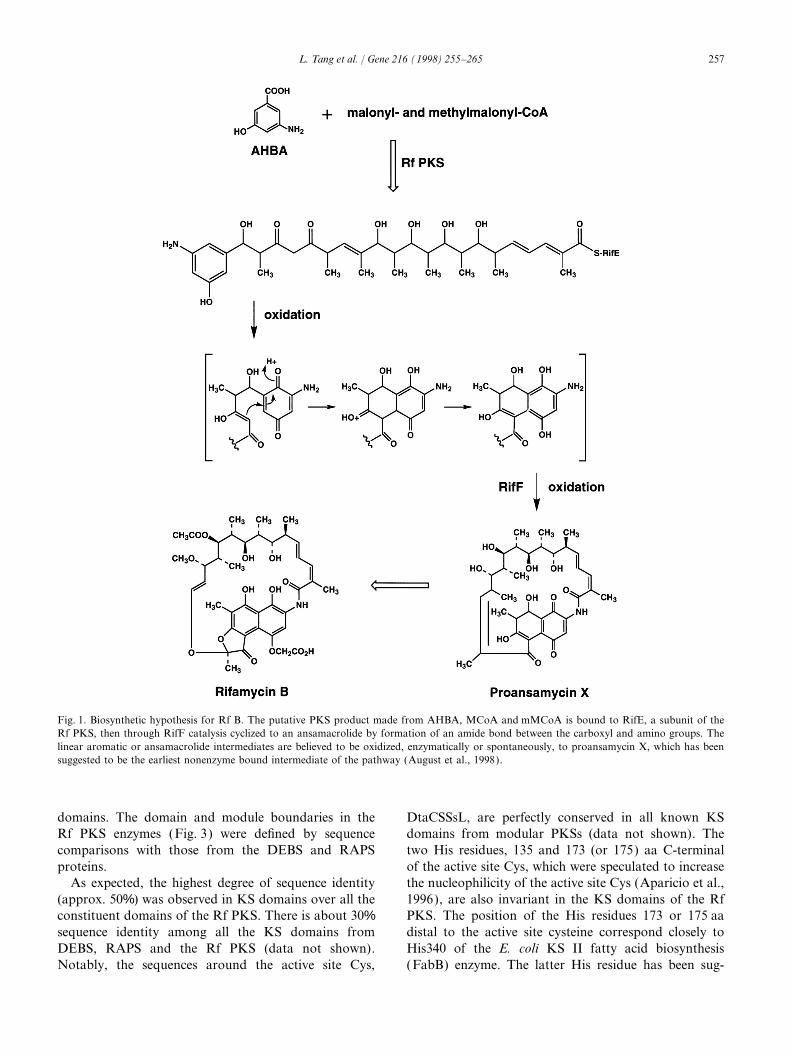
We have recently reported that five adjacent genes, Terminator sequencing system with ds DNA templatesrifA–rifE encoding the components of the modular type and run on an Applied Biosystem 373A or 377 auto-I polyketide synthase (PKS), are required for the biosyn- mated sequencer. Sequencing reactions were initiallythesis of Rf B (August et al., 1998). These multifunc- performed with universal forward and reverse primers,tional polypeptides use 3-amino-5-hydroxybenzoic acid followed by specific primers generated from the initial
sequence data. PCR clones were occasionally used as(AHBA) as a starter unit and consist of 10 modulestemplates to fill the gaps between clones. Alignment ofthat catalyse successive rounds of polyketide chain elon-sequence contigs was performed using the DNA Stargation to build an undecapolyketide. The enzyme-boundSeqMan software (Madison, WI, USA). Sequenceproduct is believed to be cyclized by the product of thehomology searches were performed using the NCBIrifF amide synthase gene (August et al., 1998) immedi-world wide blast server. The nt sequences were analyzedately downstream of rifE. Cytochrome P450-mediatedusing Wisconsin GCG programs, v. 8.1 (Devereux et al.,oxidation may also be involved to form the dihydro-1984). Multiple sequence alignments were performednaphthoquinone unit, resulting in the formation ofusing GCG programs and the DNA Star programproansamycin X, the putative ansamacrolide precursorMegAlign. The sequence data are available fromof Rf B (Fig. 1). The timing of dihydronaphthoquinoneGenBank under the accession number AF040570.formation is unknown.
Related modular type I PKSs, composed of largemultifunctional polypeptides, have been reported for thebiosynthesis of erythromycin (Er) (Donadio et al., 1991; 3. Results and discussionBevitt et al., 1992; Donadio and Katz, 1992), avermectin(MacNeil et al., 1992), oleandomycin (Swan et al., 3.1. b-Ketoacyl:acyl carrier protein synthase (KS),1994), rapamycin (Rp) (Schwecke et al., 1995; Aparicio acyltransferase (AT) and acyl carrier protein (ACP)et al., 1996), soraphen (Schupp et al., 1995) and FR008 domains(Hu et al., 1994). Complete sequence analysis of the Erand Rp biosynthesis genes encoding the subunits of The Rf PKS consists of five multifunctional enzymes6-deoxyerythronolide B synthase (DEBS) and rapa- with a total of ten modules (Fig. 2): modules 1–3 andmycin synthase (RAPS) established the likely bound- the loading domains are in the RifA protein, modulesaries and recognizable active site motifs for individual 4–6 in RifB, module 7 in RifC, module 8 in RifD, anddomains in DEBS and RAPS. Further examples of the modules 9–10 in RifE. The genetic organization ofmodular type I PKS must be analyzed to provide modules and the steps in which each module is used ininformation needed for a better understanding of the the biosynthetic pathway of Rf B are colinear, ascatalytic mechanism and to engineer the structure of in the case of DEBS (Donadio et al., 1991) and RAPSnovel polyketides by genetic manipulation of PKS- (Schwecke et al., 1995), although one of the threeencoding genes. Here, based on detailed analysis of the RAPS genes is transcribed convergently with respect toDNA sequence, we describe some of the characteristics the other two. In addition, the domain order in eachof the Rf PKS. Its modular organization and domain module of the Rf PKS is KS, AT, (dehydratase (DH )structures are significantly different from those of pre- and b-ketoacyl:ACP reductase ( KR), if they are pre-viously reported modular PKSs, especially the mode of sent) and ACP, as in animal FAS ( Wakil, 1989), DEBS
and RAPS. The Rf PKS has no enoyl reductase (ER)polyketide chain initiation and termination.
257L. Tang et al. / Gene 216 (1998) 255–265
Fig. 1. Biosynthetic hypothesis for Rf B. The putative PKS product made from AHBA, MCoA and mMCoA is bound to RifE, a subunit of theRf PKS, then through RifF catalysis cyclized to an ansamacrolide by formation of an amide bond between the carboxyl and amino groups. Thelinear aromatic or ansamacrolide intermediates are believed to be oxidized, enzymatically or spontaneously, to proansamycin X, which has beensuggested to be the earliest nonenzyme bound intermediate of the pathway (August et al., 1998).
domains. The domain and module boundaries in the DtaCSSsL, are perfectly conserved in all known KSdomains from modular PKSs (data not shown). TheRf PKS enzymes (Fig. 3) were defined by sequence
comparisons with those from the DEBS and RAPS two His residues, 135 and 173 (or 175) aa C-terminalof the active site Cys, which were speculated to increaseproteins.
As expected, the highest degree of sequence identity the nucleophilicity of the active site Cys (Aparicio et al.,1996), are also invariant in the KS domains of the Rf(approx. 50%) was observed in KS domains over all the
constituent domains of the Rf PKS. There is about 30% PKS. The position of the His residues 173 or 175 aadistal to the active site cysteine correspond closely tosequence identity among all the KS domains from
DEBS, RAPS and the Rf PKS (data not shown). His340 of the E. coli KS II fatty acid biosynthesis(FabB) enzyme. The latter His residue has been sug-Notably, the sequences around the active site Cys,

258 L. Tang et al. / Gene 216 (1998) 255–265
Fig. 2. Domain organization of the Rf PKS. Each domain is represented by a rectangle that is shaded according to the predicted enzymatic functionindicated at the bottom of the figure and with a length proportional to the size of the domain. Partial filling of DH and KR rectangles indicatesa domain that is predicted to be inactive.
gested to be an essential catalytic base in the active site His201 (Serre et al., 1995), is normally found in theMCoA specific AT domains (Haydock et al., 1995), butby analysis of the crystal structure of FabB (Huang
et al., 1998). in the AT of module 9 of the Rf PKS it is replaced byHis (Fig. 3). In AT domains specific for mMCoA, AsnAT domains from many modular PKSs can be divided
into two groups depending on their substrate specificity always substitutes this Gln (Haydock et al., 1995).Leu93 adjacent to the Ser92 nucleophile, whose mainfor the (2S )-mMCoA (methylmalonyl-coenzyme A) and
malonyl-coenzyme A (MCoA) chain extender units. The chain carbonyl accepts a H-bond from a water molecule,is always Gln in the AT domains specific for mMCoAspecificity has been correlated to aa residues of divergent
sequences from 33 to 13 aa N-terminal of the active site extender units among all modular PKS, including theRf PKS, while in the MCoA-specific AT domains of theSer in RAPS and DEBS (Haydock et al., 1995),
although in the absence of a molecular scale model it is Rf PKS and in the AT of module 14 of the RAPS thisGln is substituted by Ile and in the other MCoA-specificnot known whether the residues of these motifs are in
direct contact with the CoA substrate. The consensus AT domains of the RAPS this Gln is substituted by Val( Katz, 1997). Furthermore, Katz (1997) showed thatsequence, ETGYA( X7)QAXFGLL, suggested for AT
domains that incorporate MCoA extender units 30 residues upstream of the active-site Ser one finds aThr in more than 12 streptomycete PKS MCoA-specific(Haydock et al., 1995), does not fit the sequences of the
two AT domains in modules 2 and 9 of the Rf PKS AT domains and Val in more than two dozen mMCoA-specific AT domains. These relationships are also well(Fig. 3). Two other residues in the active site, identified
on the basis of the crystal structure of Escherichia coli maintained in the Rf PKS. Two other regions that allowa clearer assignment of the substrate specificity of ATmalonyl-CoA:ACP acyltransferase (Serre et al., 1995),
have been noted to be correlated with a specific extender domains were also noted. In AT domains specific forMCoA extender units, the three aa residues N-terminalunit (Haydock et al., 1995) and are well conserved in
the AT domains of the Rf PKS. Gln250 (E. coli number- of His201, which is an H-bond donor to the nucleophilicSer92 hydroxyl and a critical active site residue (Serreing), known as a H-bond acceptor of the active site

259L. Tang et al. / Gene 216 (1998) 255–265
Fig. 3. Alignment of the aa sequences of the 10 modules of the Rf PKS. Numbers at the end of each line refer to the aa sequence position in eachmodule. Major consensus sequences are boxed. Dashes refer to computer-generated gaps to maximize alignments. The extent of each domain isindicated by overlining of the sequences with black bars. The active site aa are marked by asterisks and the aa mentioned in the text are underlined.Computer-assisted sequence analyses were performed by the University of Wisconsin Genetics Computer Group (version 8.1) and DNASTARMegAlign programs.

260 L. Tang et al. / Gene 216 (1998) 255–265
Fig. 3. (continued )

261L. Tang et al. / Gene 216 (1998) 255–265
Fig. 3. (continued )

262 L. Tang et al. / Gene 216 (1998) 255–265
et al., 1995), are always HAFH201 in all hitherto knownprokaryotic modular PKSs. The AT of 6-methylsalicylicacid synthase (6MSAS) (Beck et al., 1990), which isspecific for MCoA, has the consensus sequence IAFH.In AT domains that incorporate mMCoA extender units,the corresponding segment of aa has the consensussequence YASH in all known PKSs. The aa residue nextto Gln250 is always Val in AT domains specific forMCoA and always replaced by Leu in AT domainsspecific for mMCoA (Haydock et al., 1995). Asexpected, the active site signature sequence GHSXGand two more active site residues, Gln11 and Arg117 inthe E. coli enzymes, which have been shown to beinvariant (Aparicio et al., 1996), are also found amongall the AT domains of the Rf PKS.
Surprisingly, all the ACP domains in the Rf PKShave the active site sequence, XGFDS (Fig. 3) where Xis either Ala, His, Val, Leu, Thr, or Ile. All the previouslyreported ACPs of modular PKS have Leu adjacent tothe Glu.
3.2. DH and KR domains
DH domains are found in all the Rf PKS modulesexcept for modules 2 and 3; the former module has onlythe core domains with a direct linkage between the ATand ACP domains. We predict that DH activity isrequired only in modules 4, 9 and 10 for biosynthesisof the Rf B precursor (Fig. 1). Interestingly, alignmentof the sequences revealed that the DH domainsin modules 9 and 10 have the apparent active sitemotif H( X3)G( X4)A and H( X3)D( X4)P, respectively(Fig. 3), where all hitherto identified DH domains fromother modular PKSs have the H( X3)G( X4)P consensussequence (Donadio and Katz, 1992; Aparicio et al.,1996). Apparently intact DH domains are present inmodules 6 and 7, even though DH activity is notexpected to be required at these steps. A molecular basisfor the inactivity of module 6 DH is not immediatelyapparent, but the Phe residue adjacent to the C-terminalPro in the predicted active site of module 7 could bethe basis for its proposed dysfunction, since Val has aquite different molecular volume than Phe and is presentin the active sites of modules 9 and 10 of the Rf PKS(Fig. 3), DEBS and RAPS. On the other hand, module8 should be nonfunctional due to the large deletions inits N- and C-terminal regions (Fig. 3). Six and seven aaare deleted from the active sites of the DH domains inmodules 1 and 5, respectively, which should be sufficientto inactivate them, although this deletion is differentfrom that of the nonfunctional DH domains of RAPSmodules 3 and 6 (Aparicio et al., 1996). Gly is presenton the C-terminal side of the active site Pro in all theknown functional DH domains, which may provideFig. 3. (continued)stabilization to the intermediate according to the crystal

263L. Tang et al. / Gene 216 (1998) 255–265
Fig. 4. Alignments of the sequences for the AMP-binding motifs found in the SUL domains of RifA and RAPS1 with those from prokaryotic andeukaryotic CLs. Numbers at the end of each line refer to the aa sequence position in each protein. Major consensus sequences are boxed. Dashesrefer to computer-generated gaps to maximize alignments. The putative AMP-binding motifs are underlined. B. subtilis and N. crassa are theacetate-CoA ligases of Bacillus subtilis (Grundy et al., 1993) and Neurospora crassa (Connerton et al., 1990), respectively.
structure of E. coli b-hydroxydecanoyl thiol ester dehy- which is also well conserved in the SUL domain ofdrase (Leesong et al., 1996). In the DH domains of RAPS (Aparicio et al., 1996). An ACP domain isRAPS and the Rf PKS that are clearly nonfunctional present between the SUL domain and the KS domainby active site deletion, this Gly residue is replaced by in module 1 of RifA. With the exception of the ERAla (in modules 2, 5, 11 and 12 of RAPS and module domain between the SUL and ACP domains in the5 of Rf RKS) or Asn (Fig. 3). This correlation does loading region of RAPS1 (Aparicio et al., 1996; Lowdennot extend to modules 6 or 7 of the Rf PKS, however. et al., 1996), the organization of loading domains isInterestingly, in Rf PKS modules 2 and 3 there is a quite similar in both RAPS1 and RifA. Consequently,trace of an aa sequence that is present in the DH the starter unit loading mechanism proposed for RAPSdomains, especially at their N-terminus. This observa- (Lowden et al., 1996) could be applicable to the Rftion could support the evolutionary hypothesis that PKS; i.e., the AHBA starter unit is first activated as itsmodular PKSs have evolved from an ancestral element acyl-adenylate by the SUL, loaded onto the thiol of thethat contained the full set of b-carbonyl reduction 4∞-phosphopantetheine prosthetic group of the ACPactivities, followed by loss of the functions not required domain and transferred to the thiol of the KS inat particular cycles of chain extension (Donadio and module 1.Katz, 1992). A well-conserved interdomain region between all the
All the Rf PKS modules contain KR domains exceptDH and KR domains and the one between the AT and
for module 2 (Fig. 2). The apparently functional KRKR domains in module 3 is present in all Rf PKSdomains contain the previously identified motif for anmodules, except module 2 in which the AT and ACPNADP(H) binding site, GXGXXG( X3)A, (Fig. 3) inare linked directly, as reported for RAPS module 14DEBS (Donadio and Katz, 1992) and the oleandomy-(Aparicio et al., 1996). A high degree of sequencecin PKS (Swan et al., 1994). The third invariant Glysimilarity could be detected when the DH/KR andin this motif is replaced by Ala in RAPS (AparicioAT/KR linker regions were compared (data not shown).et al., 1996). In the Rf PKS, the module 3 KR domainIt has been speculated that this linker region is involvedseems to be nonfunctional, since the sequences corre-in facilitating protein–protein interaction in animalsponding to an NADP(H ) binding motif are signifi-
cantly different from the consensus sequence, and a fatty-acid synthase ( Witkowski et al., 1991). Since thishighly conserved motif, aSRrG, found in all known inter-domain region between the AT/DH and DH/ERPKS, is deleted (Fig. 3). The invariant Arg in this domains is found within almost all modules of DEBS,motif has been reported to be one of the binding sites RAPS and the Rf PKS, it must have an important butfor the 2∞-phosphate group of NADPH (Scrutton et al., still unknown role. The N-termini of the five Rf PKS1990). These observations are consistent with the fact multienzymes include 30–34 aa segments in front of thethat the KR domain in DEBS module 3, which is KS domains, as reported for DEBS2, DEBS3, RAPS2nonfunctional (Donadio et al., 1991), lacks the same and RAPS3 (Aparicio et al., 1996). These N-terminalsegment. extensions have been suggested to facilitate the dimeriza-
tion of individual multifunctional polypeptides3.3. Starter unit loading domain, inter-domain linkers(Aparicio et al., 1996). There are also long C-terminaland extra-domain regionstails at the end of each multienzyme of the Rf PKS, asin the DEBS and RAPS systems, except for RifE. InSequence alignment data (Fig. 4) show that thethe latter protein, the ACP domain is followed by onlyN-terminal region of RifA contains a starter unit ligase4 aa, two of which overlap the predicted translation(SUL) domain with high similarity to both prokaryoticstart site of the following gene, rifF. A similar observa-(Grundy et al., 1993) and eukaryotic (Connerton et al.,tion has been made for 6MSAS, which only has a 6 aa1990) carboxylic acid:coenzyme A ligases (CLs), plus
the putative AMP-binding motif, mXXTSGtTGXPK, C-terminal tail (Beck et al., 1990).

264 L. Tang et al. / Gene 216 (1998) 255–265
Donadio, S., Katz, L., 1992. Organization of the enzymatic domains4. Conclusionsin the multifuctional polyketide synthase involved in erythromycinformation in Saccharopolyspora erythraea. Gene 111, 51–60.(1) The Rf PKS has a unique modular organization,
Donadio, S., Staver, M.J., McAlpine, J.B., Swanson, S.J., Katz, L.,which is significantly different from those of DEBS 1991. Modular organization of the genes required for complexand RAPS, since it contains two monomodular polyketide biosynthesis. Science 252, 675–679.
Grundy, F.J., Waters, D.A., Takova, T.Y., Henkin, T.M., 1993. Identi-enzymes.fication of genes involved in utilization of acetate and acetoin in(2) Two divergent sequence motifs that clearly distin-Bacillus subtilis. Mol. Microbiol. 10, 259–271.guish the substrate specificity of AT domains and
Haydock, S., Aparicio, J.F., Molnar, I., Schwecke, T., Konig, A.,are directly related to active site are present, asMarsden, A.F.A., Galloway, I.S., Staunton, J., Leadlay, P.F., 1995.
in RAPS. Divergent structural motifs correlated with the substrate specificity(3) In the apparently inactive DH domains of the Rf of (methyl )malonyl-CoA: acyl carrier protein transacylase domains
PKS, the active site sequences are distinctly different in modular polyketide synthase. FEBS Lett. 374, 246–248.Hu, Z., Bao, K., Zhou, X., Zhou, Q., Hopwood, D.A., Kieser, T.,from those reported for the inactive DHs of RAPS
Deng, Z., 1994. Repeated polyketide synthase modules involved inand DEBS.the biosynthesis of a heptaene macrolide by Streptomyces sp.(4) All the ACP domains in the Rf PKS have uniqueFR-008. Mol. Microbiol. 14, 163–172.
active site sequences when compared with those of Huang, W., Jia, J., Edwards, J., Dehesh, K., Schneider, G., Lindqvist,previously reported modular PKSs. Y., 1998. Crystal structure of b-ketoacyl-acyl carrier protein syn-
(5) The organization of the loading domain closely thase II from E. coli reveals the molecular architecture of condensingenzyme. EMBO J. 17, 1183–1191.resembles that of the RAPS1 protein, however, RifA
Katz, L., 1997. Manipulation of modular polyketide synthases. Chem.contains SUL and ACP domains only without anRev. 97, 2557–2575.ER domain between them, as in RAPS1.
Leesong, M., Handerson, B.S., Gilling, J.R., Schwab, J.M., Smith,Note added in proof J.L., 1996. Structure of a dehydratase-isomerase from the bacterialThe cloning and sequence analysis of the rifamycin pathway for biosynthesis of unsaturated fatty acids: two catalytic
PKS genes in A. mediterranei have been carried out activities in one active site. Structure 4, 253–264.Lowden, P.A.S., Bohm, G.A., Staunton, J., Leadlay, P.F., 1996. Theindependently by Schupp et al. (1998), whose report
nature of the starter unit for the rapamycin polyketide synthase.appeared while our paper was being reviewed.Angew. Chem. Int. Ed. Engl. 35, 2249–2251.
MacNeil, D.J., Occi, J.L., Gewain, K.M., MacNeil, T., Gibbons, P.H.,Ruby, C.L., Danis, S.J., 1992. Complex organization of the Strepto-myces avermitilis genes encoding the avermectin polyketide synthase.
References Gene 115, 119–125.Mandell, G.L., Sande, M.A., 1980. Drugs used in the chemotherapy
August, P.R., Tang, L., Yoon, Y.J., Ning, S., Muller, R., Yu, T.-W., of tuberculosis and leprosy. In: Goodman, A.G., Goodman, L.S.,Taylor, M., Hoffmann, D., Kim, C.-G., Zhang, X., Hutchinson, Gulman, A. (Eds.), The Pharmocological Basis of Therapeutics, 6thC.R., Floss, H.G., 1998. Biosynthesis of the ansamycin antibiotic ed. Macmillan, New York, pp. 1203–1206.rifamycin: deductions from the molecular anaysis of the rif biosyn- Schupp, T., Toupet, C., Cluzel, B., Neff, S., Hill, S., Beck, J.J., Ligon,thetic gene cluster of Amycolatopsis mediterranei S699. Chem. Biol. J.M., A Sorangium cellulosum (myxobacterium) gene cluster for the5, 69–79. biosynthesis of the macrolide antibiotic soraphen A: cloning, charac-
Aparicio, J.F., Molnar, I., Schwecke, T., Konig, A., Hydock, S.F., terization, and homology to polyketide synthase genes from actino-Khaw, L.E., Staunton, J., Leadlay, P.F., 1996. Organization of the mycetes. 1995. J. Bacteriol. 177, 3673–3679.biosynthetic gene cluster for rapamycin in Streptomyces hygroscopi- Schupp, T., Toupet, C., Engel, N., Goff, S., 1998. Cloning andcus: analysis of the enzymatic domains in the modular polyketide sequence analysis of the putative rifamycin polyketide synthase genesynthase. Gene 169, 9–16. cluster from Amycolatopsis mediterranei. FEMS Microbiol. Lett.
Beck, J., Ripka, S., Siegner, A., Schiltz, E., Schweiser, E., 1990. The 159, 201–207.multifunctional 6-methylsalicylic acid synthase gene of Penicillium Schwecke, T., Aparicio, J.F., Molnar, I., Konig, A., Khaw, L.E.,patulum. Its gene stucture relative to that of other polyketide
Hydock, S.F., Olinyk, M., Caffrey, P., Cortes, J., Lester, J.B., Bohm,synthases. Eur. J. Biochem. 192, 487–498.
G.A., Staunton, J., Leadlay, P.F., 1995. The biosynthetic gene clus-Bevitt, D.J., Cortes, J., Haydock, S.F., Leadlay, P.F., 1992.
ter for the polyketide immunosuppressant rapamycin. Proc. Natl.6-Deoxyerythronolide B synthase 2 from SaccharopolysporaAcad. Sci. USA 92, 7839–7843.erythraea. Cloning of the structural gene, sequence analysis and
Scrutton, N.S., Berry, A., Perham, R.N., 1990. Redesign of the coen-inferred domain structure of the mutifunctional enzyme. Eur. J.zyme specificity of a dehydrogenase by protein engineering. NatureBiochem. 204, 39–49.343, 38–43.Connerton, I.F., Fincham, J.R., Sandeman, R.A., Hynes, M.J., 1990.
Sensi, P., Margalith, P., Timbal, M.T., 1959. Rifomycin, a new anti-Comparison and cross-species expression of the acetyl-CoA synthe-biotic. Preliminary report. . Farmaco. ED. Sci. 14, 146–147.tase genes of the Ascomycete fungi Aspergillus nidulans and Neuros-
Serre, L., Verbee, E.C., Dauter, Z., Stuitje, A.R., Derewenda, Z.S.,pora crassa. Mol. Microbol. 4, 451–460.1995. The Escherichia coli malonyl-CoA:acyl carrier protein transa-Devereux, J., Haeberli, P., Smithies, O., 1984. A comprehensive set ofcylase at 1.5 A resolution. J. Biol. Chem. 270, 12961–12964.sequence analysis programs for the VAX. Nucl. Acids Res. 12,
Swan, D.G., Rodriguez, A.M., Vilches, C., Mendez, C., Salas, J.A.,387–395.1994. Characterization of a Streptomyces antibioticus gene encodingDickinson, J.M., Mitchison, D.A., 1987. In vitro activity of newa type I polyketide synthase which has an unusual coding sequence.rifamycins against rifampicin-resistant M. tuberculosis and MAIS
complex mycobacteria. Tubercule 68, 177–182. Mol. Gen. Genet. 242, 358–362.

265L. Tang et al. / Gene 216 (1998) 255–265
Wakil, S.J., 1989. Fatty acid synthase, a proficient multifunctional Witkowski, A., Rangan, V.S., Randhawa, Z.I., Amy, C.M., Smith, S.,1991. Structural organization of the multifuctional animal fatty-acidenzyme. Biochemistry 28, 4523–4530.
Wehrli, W., 1983. Rifampin: mechanisms of actions and resistance. synthase. Eur. J. Biochem. 198, 571–579.Woodley, C.L., Kilburn, J.O., 1982. In vitro susceptibility of Mycobac-Rev. Infect. Dis. 5, S407–S411.
Wehrli, W., Staehelin, M., 1971. Actions of rifamycins. Bacteriol. Rev. terium avium complex and Mycobacterium tuberculosis strains to aspiropiperidyl rifamycin. Am. Rev. Respir. Dis. 126, 58635, 290–309.
Copyright © 2022 FDOKUMEN




















