
Characterization of strains of Leuconostoc mesenteroides by analysis of soluble whole-cell protein...
11
Journal of Applied Microbiology 1997, 02, 578-588 Characterization of strains of Leuconostoc mesenteroides by analysis of soluble whole-cell protein pattern, DNA fingerprinting and restriction of ribosomal DNA F. Villani, G. Moschetti, G. Blaiotta and S. Coppola lstituto di Microbiologia Agraria e Stazione di Microbiologia industriale, Universita degli studi di Napoli ‘Federico II’, Portici, Italy 5850/07/96: received 16 July 1996, revised 17 September 1996 and accepted 24 September 1996 F. VILLANI, G. MOSCHETTI, G. BLAIOTTA AND s. COPPOLA. 1997. Of 215 leuconostocs isolated from field grass, natural whey cultures and water-buffalo milk, 178 were identified as Leuronostor mesenteroides ssp. mesenteroides while 37 strains could not be identified. Biochemical characterization allowed seven groups to be defined. Representative strains of each group and different habitat and nine reference strains were selected for further analyses. Protein profiles appeared suitable for species discrimination, but did not differentiate between the three subspecies of Leur. mesenteroides. The technique also showed some differences among equivocal strains. DNA fingerprinting for most strains of Leuc. mesenteroides ssp. mesenteroides examined showed a different restriction pattern from that of the type strain. Ribotyping was not useful for discriminating species and subspecies of the genus Leuconostor: Leur. mesenteroides ssp. mesenteroides and ssp. dextranirum showed the same ribopattern as Leuc. lartis while Leuc. mesenteroides ssp. rremoris exhibited a pattern distinct from all the other species examined. On the basis of ARDRA-PCR, two main groups could be distinguished: the larger group included Leur. mesenteroides, Leuc. lactis, Leuc. pseudomesenteroides and some unidentifiable strains; the second one included Leuc. citreum, Leur. fallax, IVeissella parumesenteroides and some unidentified strains. INTRODUCTION The genus Leuconostor (Leuc.) includes Gram-positive, catalase-negative micro-organisms with irregular coccoid morphology. Their distinction from gas-forming het- erofermentative lactobacilli has long been controversial. In fact, the two genera are now considered to be phylogenetically intermixed (Stackebrandt et al. 1983; Stackebrandt and Teuber 1988). Garvie (1981) defined the salient characteristics of the genus Leuconostor and recognized four species: Leuc. mesen- teroides (with the three subspecies, dextranirum, rvemoris and mesenteroides), Leuc. paramesenteroides, Leuc. lartis and Lrur. oenos, recently reclassified as Oenoroccus oeni (Dicks et al. 1995). Since 1989 six further species have been described: Leuc. carnosum and Leuc. gelidum (Shaw and Harding 1989); Correspundenre to: Profissor Salsatore Coppula, Istituto di .Uicrubzulugiu .4graria e Stazionr di ~2firruhiologia industriale, Unirersiti dz Napolz ‘Federico II ’, 800.75 Purtici, Italy (e-mail: suroppolia!ds.unzna.it). Leur. pseudomesenteroides and Leuc. citreum (Farrow et al. 1989), also known as Leuc. nmelibiosum (Schillinger et al. 1989; Takahashi et a/. 1992); Leur. argentinurn (Dicks et al. 1993); and an atypical leuconostoc named Leuc. fallax (Mar- tinez-Murcia and Collins 1991). More recently, Collins et al. (1993) proposed that Leur. paramesenteroides and related species be reclassified in a new genus, Ifkissella. Comparative analyses of 16s and 23s rDNA sequences revealed the presence of three distinct lines of descent: Leu- conostoc sensu strirto, the Leur. paramesenteroides group ( If kis- sella) and the species Leur. oenos (Yang and Woese 1989; Martinez-Murcia and Collins 1990, 1991; Collins et al. 1993; Martinez-Murcia et al. 1993). Leuconostocs are widespread in the natural environment, though probably because of their weak acidifying activity, they are not widely investigated. Apart from some spoilage roles, they are generally considered as flavouring agents in dairy, wine and vegetable fermentations. Their recognized antagonistic or synergistic properties in mixed microflora 0 1997 The Society for Applied Bacteriology
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Characterization of strains of Leuconostoc mesenteroides by analysis of soluble whole-cell protein...

Journal of Applied Microbiology 1997, 02, 578-588
Characterization of strains of Leuconostoc mesenteroides by analysis of soluble whole-cell protein pattern, DNA fingerprinting and restriction of ribosomal DNA
F. Villani, G. Moschetti, G. Blaiotta and S. Coppola lstituto di Microbiologia Agraria e Stazione di Microbiologia industriale, Universita degli studi di Napoli ‘Federico II’, Portici, Italy
5850/07/96: received 16 July 1996, revised 17 September 1996 and accepted 24 September 1996
F. VILLANI, G. MOSCHETTI, G . BLAIOTTA AND s. COPPOLA. 1997. Of 215 leuconostocs isolated from field grass, natural whey cultures and water-buffalo milk, 178 were identified as Leuronostor mesenteroides ssp. mesenteroides while 37 strains could not be identified. Biochemical characterization allowed seven groups to be defined. Representative strains of each group and different habitat and nine reference strains were selected for further analyses. Protein profiles appeared suitable for species discrimination, but did not differentiate between the three subspecies of Leur. mesenteroides. The technique also showed some differences among equivocal strains. DNA fingerprinting for most strains of Leuc. mesenteroides ssp. mesenteroides examined showed a different restriction pattern from that of the type strain. Ribotyping was not useful for discriminating species and subspecies of the genus Leuconostor: Leur. mesenteroides ssp. mesenteroides and ssp. dextranirum showed the same ribopattern as Leuc. lartis while Leuc. mesenteroides ssp. rremoris exhibited a pattern distinct from all the other species examined. On the basis of ARDRA-PCR, two main groups could be distinguished: the larger group included Leur. mesenteroides, Leuc. lactis, Leuc. pseudomesenteroides and some unidentifiable strains; the second one included Leuc. citreum, Leur. fa l lax , IVeissella parumesenteroides and some unidentified strains.
INTRODUCTION
The genus Leuconostor (Leuc.) includes Gram-positive, catalase-negative micro-organisms with irregular coccoid morphology. Their distinction from gas-forming het- erofermentative lactobacilli has long been controversial. In fact, the two genera are now considered to be phylogenetically intermixed (Stackebrandt et al. 1983; Stackebrandt and Teuber 1988).
Garvie (1981) defined the salient characteristics of the genus Leuconostor and recognized four species: Leuc. mesen- teroides (with the three subspecies, dextranirum, rvemoris and mesenteroides), Leuc. paramesenteroides, Leuc. lartis and Lrur . oenos, recently reclassified as Oenoroccus oeni (Dicks et al. 1995). Since 1989 six further species have been described: Leuc. carnosum and Leuc. gelidum (Shaw and Harding 1989);
Correspundenre to: Profissor Salsatore Coppula, Istituto di .Uicrubzulugiu .4graria e Stazionr d i ~2firruhiologia industriale, Unirersiti dz Napolz ‘Federico I I ’, 800.75 Purtici, Italy (e-mail: suroppolia!ds.unzna.it).
Leur. pseudomesenteroides and Leuc. citreum (Farrow et al. 1989), also known as Leuc. nmelibiosum (Schillinger et al. 1989; Takahashi et a/. 1992); Leur. argentinurn (Dicks et al. 1993); and an atypical leuconostoc named Leuc. fallax (Mar- tinez-Murcia and Collins 1991). More recently, Collins et al. (1993) proposed that Leur. paramesenteroides and related species be reclassified in a new genus, Ifkissella.
Comparative analyses of 16s and 23s rDNA sequences revealed the presence of three distinct lines of descent: Leu- conostoc sensu strirto, the Leur. paramesenteroides group ( I f kis- sella) and the species Leur. oenos (Yang and Woese 1989; Martinez-Murcia and Collins 1990, 1991; Collins et al. 1993; Martinez-Murcia et al. 1993).
Leuconostocs are widespread in the natural environment, though probably because of their weak acidifying activity, they are not widely investigated. Apart from some spoilage roles, they are generally considered as flavouring agents in dairy, wine and vegetable fermentations. Their recognized antagonistic or synergistic properties in mixed microflora
0 1997 The Society for Applied Bacteriology

CHARACTERIZATION OF LEUCONOSTOC MESENTEROIDES 579
populations has led to studies on their importance in some starter cultures (Cogan and Jordan 1994; Vedamuthu 1994).
Unequivocal criteria and reliable methods for identification and characterization of microbial strains are a primary requirement in industrial as well as in other fields of applied microbiology. The difficulties encountered in identifying Leuconostuc species reveal the need for additional charac- terization procedures that can be used in a reference labora- tory. Discrimination of strains of the same species by chromosomal DNA restriction endonuclease analysis (REA) is very useful for genetic characterization and has been applied to a variety of lactic acid bacteria (Tanskanen et al. 1990; Salzano et al. 1993; Moschetti et ul. 1995), but little infor- mation exists on the interstrain relationships of leuconostocs and their divergence from other species of the genus (Lam- oureux et al. 1993; Kelly et al. 1993; Tenreiro et al. 1994; Viti et al. 1995; Kelly et al. 1995).
Electrophoresis of whole-cell soluble proteins has been used to differentiate Leuronostoc species but is unable to distinguish the three subspecies of Lrur . mesenteroides (Dicks et al. 1990; Elliott and Facklam 1993; Tsakalidou et al. 1994). Molecular typing based on polymorphism of ribosomal DNA (rDNA) has potential as a taxonomic technique since rRNA genes are both ubiquitous and highly conserved (Grimont and Grimont 1986). Studies on a variety of micro-organisms including Haemophilus infiuenzae (Irino et al. 1988), Staphy- lococcus spp. (De Buyser et al. 1989; Thomson-Carter et al. 1989) and Streptococcus thermophilus (Salzano et al. 1994) have demonstrated that rRNA gene restriction pattern analysis provides a means of distinguishing between strains at species, subspecies or other taxonomic levels. The study of poly- morphism of ribosomal DNA, facilitated by restriction analy- sis of rRNA genes amplified by polymerase chain reaction (ARDRA-PCR), has also been used for the differentiation of species and subspecies (Gurtler et al. 1991; Jayarao et ul. 1991; Vaneechoutte et al. 1992).
In this paper we report the levels of genotypic relatedness among strains of leuconostocs, largely isolated from dairy products, by the combined use of analysis of soluble whole- cell protein pattern, DNA fingerprinting and restriction of ribosomal DNA, with the aim of determining their usefulness in differentiating between Leuronostoc species, subspecies and strains.
MATERIALS AND METHODS
Reference strains
The reference strains used are listed in Table 1. All cultures were obtained from the Deutsche Sammlung von Mikroor- ganismen (DSM, Braunschweig) and cultured in MRS broth (Oxoid) at 30°C.
Samples and isolation procedures
Bacteria were isolated from field grass (two samples), natural whey cultures (NWC) used as starters in the manufacture of water-buffalo Mozzarella cheese (five samples) and water- buffalo milk (one sample) by homogenizing 10 g or 10 ml in 90ml of quarter strength Ringer's solution in a stomacher. Suitable dilutions were then plated on high-sucrose Mayeux agar (Mayeux et al. 1962) and LUSM agar (Benkerroum et al. 1993). The plates were incubated aerobically at 30°C and examined a t 2, 3 and 5 d. A number of colonies equal to the square root of the total were picked at random using the Harrison disc described by Harrigan and McCance (1976). Individual isolates were maintained as deep cultures in MRS agar at 4°C.
Identification of bacterial isolates
All the isolates were tested initially for Gram reaction, cell morphology, catalase reaction (3?/0 HzOz), gas production from glucose (APT broth without sodium citrate but sup- plemented with l0/o dextrose and 0.5Oio sodium acetate) by the hot loop test (Sperber and Swan 1976), hydrolysis of arginine (MRS broth without glucose and meat extract but supplemented with 0.30/0 arginine and NH3 detected by addition of Nessler's reagent), and isomer of lactic acid by an enzymic method (Von Krush and Lompe 1982) using D- and L-lactate dehydrogenases (Boehringer Mannheim Italia SpA).
On the basis of the results, 215 isolates were identified as Leuronostor spp. and further characterized. The production of dextran from sucrose was determined on MRS agar with 596 sucrose replacing glucose. The ability to ferment various carbohydrates was determined using MRS broth without glucose and meat extract with 2O/o added carbohydrate and 0.00196 chlorophenol red (Schillinger and Lucke 1987).
The strains were identified according to the criteria of Garvie (1986), Holzapfel and Schillinger (1992) and Dicks et al. (1993).
SDS-PAGE of total soluble cell proteins
Cultures were grown on MRS agar plates at 30°C for 18 h. Soluble whole-cell proteins were prepared by suspending washed cell pellets ( z 100 mg wet weight) in 1 ml of sample treatment buffer (0.0625 mol l-' Tris-HC1, pH 6.8, 109'0 glycerol, 2% SDS, 596 P-mercaptoethanol) and disrupting the cells with glass beads (0.10-0.1 1 mm) on a vortex mixer for 6 min. After lysis, the cell suspensions were heated for 10 min at 95"C, cooled and pelleted at lO0Og for 10 min. The supernatant fluids (protein extract) were analysed by SDS- PAGE (10% in the running gel and 4.5% in the stacking gel) by the method of Laemmli (1970).
0 1997 The Society for Applied Bacteriology, Journal of Applied Microbiology82, 578-588
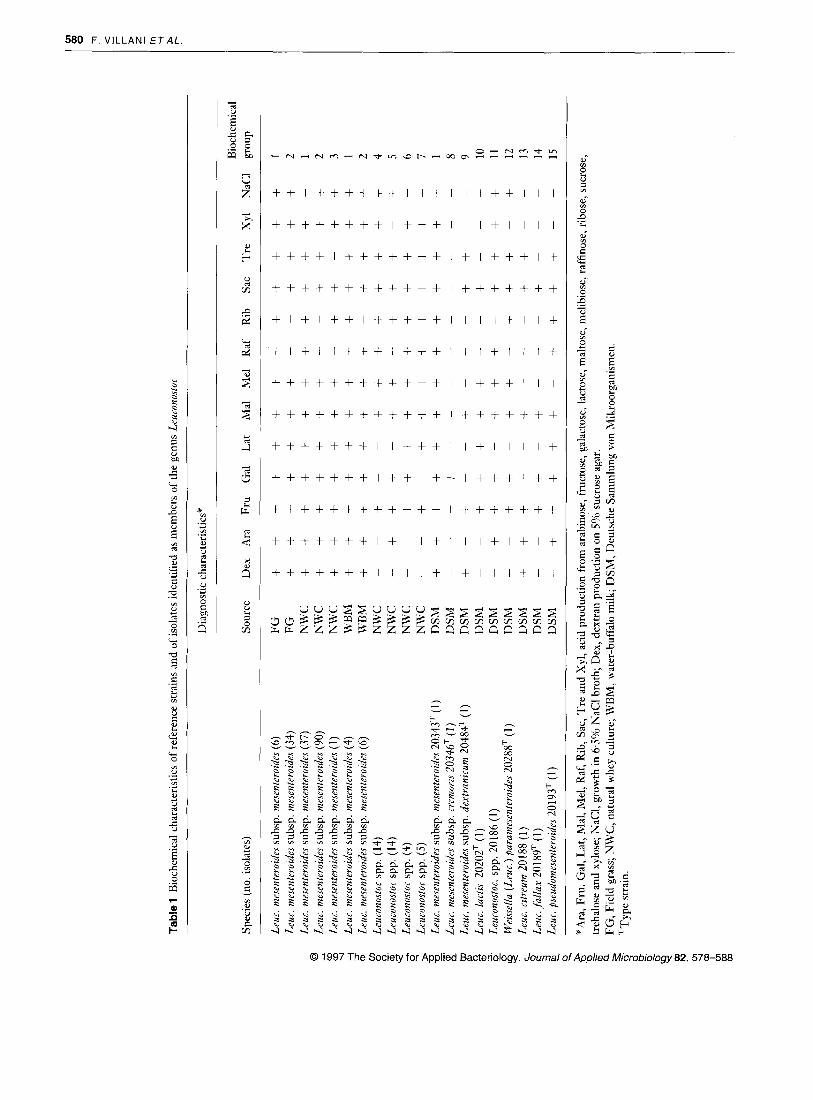
UI
-4
W
W
W d
Tab
le 1
Bio
chem
ical
char
acte
ristic
s of r
efer
ence
stra
ins
and
of is
olat
es id
entif
ied
as m
embe
rs o
f the
gen
us L
euco
nusto
6
Dia
gnos
tic c
hara
cter
istic
s"
Spec
ies (
no. i
sola
tes)
So
urce
D
ex
Ara
Fr
u G
al
Lat
M
a1
Me1
R
af
Rib
Sa
c T
re
Xyl
Leuc
. mes
ente
roid
es su
bsp.
mes
ente
roid
es (6
) L
eu. m
esen
tero
ides
subs
p. m
esen
tero
ides
(34)
Le
uc. m
esen
tero
ides
sub
sp. m
esen
tero
ides
(37)
Le
uc. m
esen
tero
ides
subs
p. m
esen
tero
ides
(90)
Le
uc. m
esen
tero
ides
subs
p. m
esen
tero
ides
(1)
Leuc
. mes
ente
roid
es su
bsp.
mes
ente
roid
es (4
) Le
uc. m
esen
tero
ides
subs
p. m
esen
tero
ides
(6)
Leuc
onos
toc
spp.
(14)
Le
ucon
osto
c sp
p. (1
4)
Leuc
onos
toc
spp.
(4)
Leuc
onos
toc
spp.
(5)
Leu
. mes
ente
roid
es su
bsp.
mes
ente
roid
es 2
0343
T (1
) L
eu. m
esen
tem
ides
subs
p. c
rem
ons 2
034b
T (1
) Le
uc. m
esen
tero
ides
subs
p. d
extr
unic
um 2
0484
T (1
) Le
uc. l
actis
202
02T
(1)
Leuc
onos
toc.
spp.
201
86 (1
) W
eiss
ella
(Leu
c.) p
aram
esen
tero
ides
202
88T (
1)
Leu
. citr
eum
201
88 (1
) Le
uc.fa
llux
2018
9T (1
) Le
uc. p
seud
omes
ente
roid
es 2
0193
T (1
)
FG
FG
N
WC
N
WC
N
WC
W
BM
W
BM
N
WC
N
WC
N
WC
N
WC
D
SM
D
SM
DS
M
DS
M
DS
M
DS
M
DSM
D
SM
D
SM
++
+
++
+
++
+
++
+
++
+
++
+
++
+
+ -
++
+ +
++
+
+-
+
+ -
++
+
++
+
+ -
++
--
--
--
--
_
--
--
--
+ + + + + + + + + - + + + + + + +
++
+
+
++
+
+
++
+
+
++
-
+
++
+
+
++
-
+
++
-
+
-+
+
-+
+
+
-
++
++
++
+
+-
++
+
++
++
++
+
+-
++
+
+-
++
-+
+
++
++
+
++
- +
++
+
++
++
+
+-
++
+-
+
++
++
+
++
++
++
+
++
++
+
--
--
_-
++
-
_-
+
-+
+-
-
++
- +
++
+
-+
++
-
++
-
++
--
-
++
++
-
--
-
--
~
Bio
chem
ical
N
aCl
grou
p
+I
$
2
+1
+
2
+3
+
1
+2
$
4
+5
+
6
+7
+
1
-8
-
9
-
10
+ 11
+
12
-
13
-
14
-
15
~
"Ara
, Fr
u, G
al, L
at, M
al, M
el, R
af, R
ib, S
ac, T
re an
d X
yl, a
cid
prod
uctio
n fr
om a
rabi
nose
, fru
ctos
e, g
alac
tose
, lac
tose
, mal
tose
, mel
ibio
se, r
affin
ose,
ribo
se, s
ucro
se,
treha
lose
and
xyl
ose;
NaC
1, g
row
th in
6.5
% N
aCl b
roth
; Dex
, dex
tran
prod
uctio
n on
5%
suc
rose
aga
r. F
G, F
ield
gra
ss; N
WC
, nat
ural
whe
y cu
lture
; WB
M, w
ater
-buf
falo
milk
; DSM
, Deu
tsch
e Sa
mm
lung
von
Mik
roor
gani
smen
. T
Typ
e str
ain.

CHARACTERIZATION OF LEUCONOSTOC MESENTEROlDES 581
Isolation of total DNA, restriction digestion and conventional electrophoresis
Total DNA was prepared by the method of Marmur (1961) from cells harvested by centrifugation (8000 rev min-I) of overnight cultures at 30°C in MRS broth. DNA (5 p g ) was digested with Hind111 (Promega) according to the supplier's instructions. Digests were separated by horizontal elec- trophoresis through 0.6% (w/v) agarose gel in TBE buffer (0.9 mol 1-' Tris-borate, 0.004 mol 1-' EDTA) at 2 .5 V cm-' for 12 h. After electrophoresis gels were stained with ethidium bromide.
Genomic DNA preparation, restriction digestion and PFGE
Genomic DNA of high molecular weight was prepared by the method of McClelland (1987). Inserts of intact DNA were dialysed in 20ml of T E (Tris 0.010 mol l-', EDTA 0,001 mol 1-I, pH 8.00) containing 0.1 mol 1 ' phe- nylmethylsulphonyl fluoride (PMSF) for 1 h at room tem- perature. After dialysis without PMSF, the samples were maintained in T E at 4°C for 30 min and then digested in 200 p1 of appropriate buffer supplemented with 40 U of <-lpaI (Promega) a t 37°C for 5 h.
Electrophoresis of the restriction digests was performed on the CHEF system (BioRad) with 100 (w/v) agarose gels and 0.5 x TBE (0.045 mol 1-' Tris-borate, 0.002 mol 1-' EDTA, pH 8.3) as running buffer, at 10°C. concatamers (Promega) and a 1 kb DNA ladder (Gibco BRL) were used as molecular size markers. Restriction fragments in the 5-200 kbp range were resolved in a single run, at a constant voltage of 6 V cm-', by a single phase procedure for 13 h with a pulse ramping between 2 s and 6 s followed by another single phase procedure for 4 h, with a pulse ramping of 10 s and 15 s.
Radioactive labelling of nucleic acids
Ribosomal 16.5 + 23s RNA from Escherichza colz (Serva) was used as described by Grimont and Grimont (1986). The reverse transcription mixture used to prepare the cDNA contained: 1 x reaction buffer (50nimol l-' Tris-HC1, pH 8.3; 75 mmol 1-' KC1; 3 mmol 1-' MgC12; 10 mmol 1 ' dithi- othreitol), 0.1 mmol 1-' each of dATP, dGTP, d T T P (Pro- mega), 12 U human placental RNase inhibitor (RNasina, Promega), 10 mg ml-' random primer d(Np,) (Promega), 2 p g RNA, 5 pl [u-"P]dCTP (Amersham) and 4 U reverse tran- scriptase enzyme (Promega). After incubation at 37°C for 1 h the reaction mixture was cooled on ice and stopped by addition of 1 pl of 0.5 mol 1-' EDTA, pH 8.0.
Southern transfer and hybridization
The electrophoresed gels containing Hind111 fragments were soaked in 0.5 mol 1-' NaOH-1 mol 1-' NaCl for 30 min,
neutralized in 0.5 moll-' Tris-1 moll ' NaCl, pH 7 4 , for 30 min and transferred to membranes (Zeta Probe, BioRad) by Southern blotting (Southern 1975). The DNA was fixed by baking at 80°C for 30 min under vacuum. The filters were prehybridized in 6 x SSC, 0.1% SDS, 5 x Denhardt and 125 pg ml-' denatured salmon sperm DNA (Sigma) at 65°C for 4 h. Hybridizations were performed overnight at 65°C in 6 x SSC, 0.lo/o SDS, 5 x Denhardt, 250pg ml-' denatured salmon sperm DNA and radioactive cDNA probe (about 8 x 10' cpm pg-I). Following incubation, the membranes were washed twice at 65°C in 3 x SSC, 0.lo.b SDS and then in 0.2% SSC, 0.1% SDS. Filters were autoradiographed with 'Kodak X-OMAT' film at -80°C with intensifying screen.
DNA amplification and restriction analysis
Synthetic oligonucleotide primers were selected and the PCR assay was carried out as described by Vaneechoutte et ul. (1992). The sequence of the oligonucleotides was 5' TGGCTCAGATTGAACGCTGGCGGC and 5' CCT- TTCCCTCACGGTACTGGT. The gene fragment ampli- fied (= 2 4 kbp) encompassed the 16S, the spacer region and part of 23s rDNA. The PCR assay was performed with 100 p1 containing 10 ng of bacterial DNA, 2.5 mmol 1 ~ ' MgC12, 200pmol 1-' of each of the four dNTPs, and 0.2pmol 1-' of each primer in Tny-buffer (Gibco BRL). After covering with 50 pl of paraffin oil, the reaction mixture was heated at 95°C for 5 min. After the addition of 2.5 U of T q polymerase (Gibco BRL), the PCR was carried out in a programmable heating incubator for 40 cycles (30 s at 95"C, 30 s at 55"C, 1 min at 72°C per cycle). Finally a 10 min extension period at 72°C was performed. The presence of specific PCR products was controlled by gel electrophoresis in 1% agarose gel con- taining ethidium bromide. The final amplification products were digested with Hind111 and H d I I (Promega), following the manufacturer's instructions. Restriction fragments were analysed by agarose ( l.SO/o w/v) gel electrophoresis, at 7 V cm-' during 2 h.
RESULTS
On the basis of published biochemical data, nine reference strains, obtained from the DSM collection, and the 215 iso- lates of Leuronostoc spp. from field grass, natural whey culture and water-buffalo milk were presumptively identified by the scheme shown in Fig. 1. Of the above 215 isolates, 178 strains (82.7%) were identified as Lruc. mesenterodes ssp. mesen- tevozdes with some differences in the fermentation of ribose, raffinose and trehalose while 37 strains could not be identified. The 215 strains and the nine reference strains could be allo- cated to 15 biochemical groups (Table 1). Representative strains of each group and different habitat were selected for further study.
0 1997 The Society for Applied Bacteriology, Journal of Applied Microbiology 82, 578-588

582 F. V lLLANl E T A L
Leuconostoc spp.: cocci or coccoidal rods, Gram-positive, gas from glucose, no NH, from arginine, D-Lactic acid from glucose
I Dextran formation + I
Dextran formation - I
I + sucrose L-a ra bi nose
maltose raffinose I I + I
fructose galactose I 1
- I + growthlat 37°C I I w
Leuc. carnosum Leuc. dexfracicum Leuc. cremoris Leuc. oenos I
ribose galactose
H - + I
I I
trehalose L-arabinose D-xylose
mannitol I Leuc. gelidum Leuc. mesenferoides Leuc. fallax Leuc. gelidum 1 I ,- 1 +, Leuc. mesenferoides
Leuc. amelibiosum Leuc. cifreum growth at 15°C growth in 6.5% NaCl
I I +
Leuc. lactis Leuc. argentinum
Leuc. pseudomesenferoides I Weissella (Leuc.) paramesenferoides
Leuc. mesenferoides
Fig. 1 Scheme for presumptive identification of Leuronostoc spp.
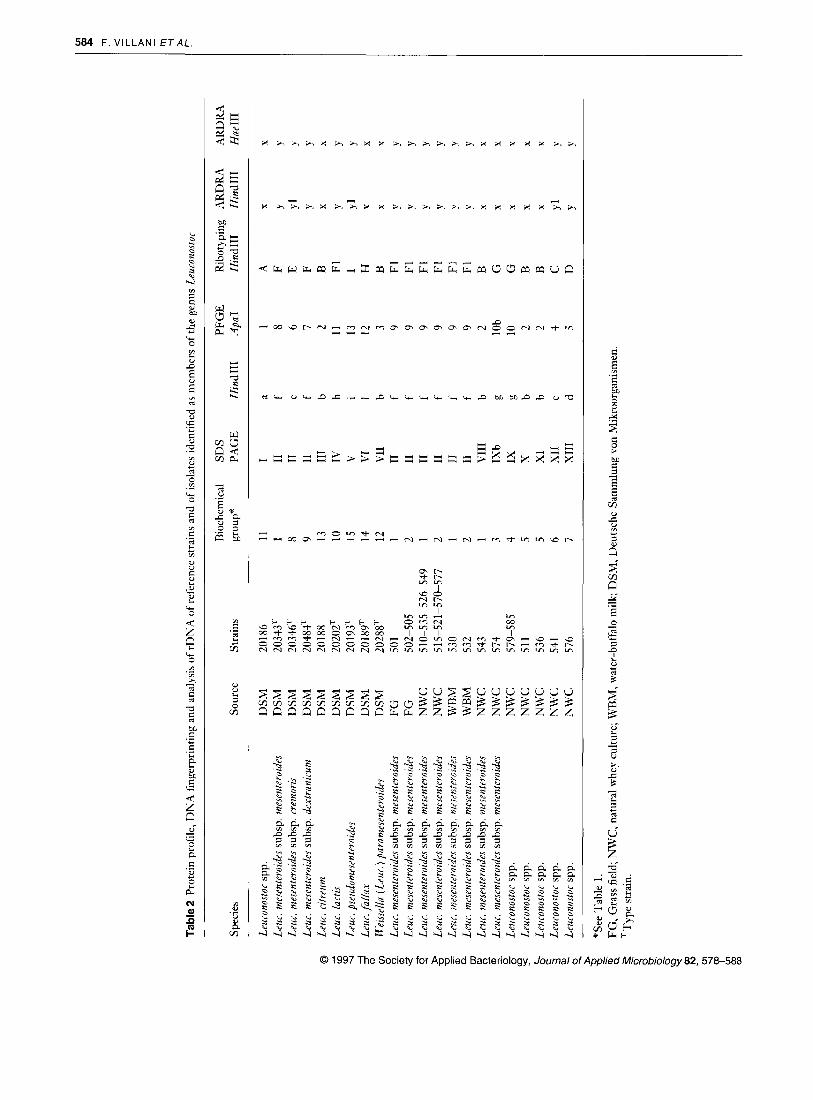
SDS-PAGE analysis of strains
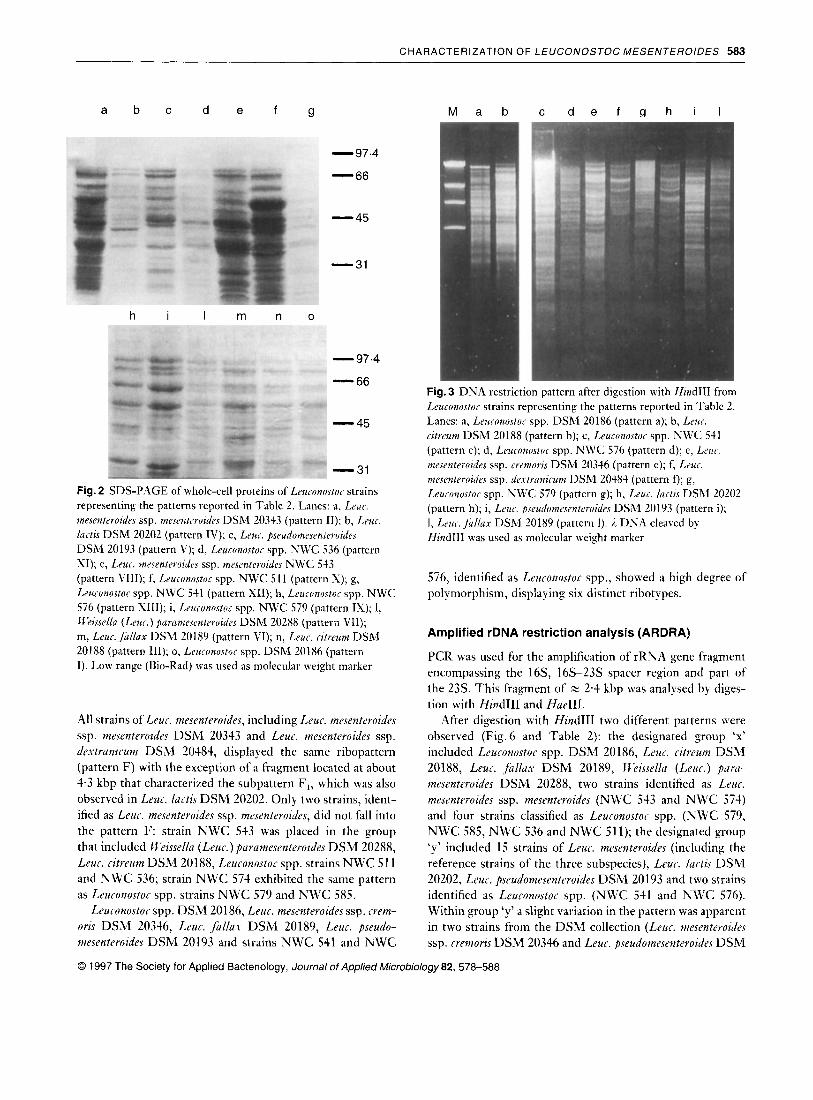
The 30 strains examined by SDS-PAGE exhibited 13 protein patterns as shown in Fig.2 and summarized in Table 2. Reference strains of Leuconostoc spp. DSM 20186, Leur. citreum DSM 20188, Leuc. lurtis DSM 20202, Weissellu (Leuc.) purumesenteroides DSM 20288, Leuc. fallax DSM 20189, Leuc. pseudomesenteroides DSM 20193, the strain identified as Leur. mesenteroides ssp. mesenteroides NWC 543 and the strains designated as Leuconostor spp. NWC 511, NWC 536, NWC 541 and NWC 576 displayed a unique and singular protein profile distinct from all the other strains examined. However, all the strains of Leur. mesenteroides (including the reference strains of the three subspecies) showed the same protein pattern, with the exception of NWC 574 that differed from strains NWC 579 and NWC 585, identified as Leuconostor spp., by a small variation in protein bands (patterns IX and IXb reported in Table 2).
High-frequency restriction endonuclease fingerprinting (REF)
Examination of the DNA of the 30 strains by high-frequency restriction fragments performed with HindIII resulted in the detection of 10 different patterns (Fig. 3 and Table 2). All strains of Leur. meseuteroides ssp. mesenteroides and ssp. dextranirum displayed the same restriction pattern, with the exception of strain NWC 574 that exhibited a pattern similar to Leuronostoc spp. strains NWC 579 and NWC 585. The strain NWC 543, identified as L e u . mesenteroides ssp. mesen- teroides, was included in the group containing Rieissellu (Leur. ) parumesenteroides DSM 20288, Leur. citreum DSM 20188 and Leuronostor spp. strains NWC 511 and NWC 536.
Leuronostor spp. DS 0 86, Leur. mesenteroides ssp. crem- oris DSM 20346, Leur. luctis DSM 20202, Leur. pseudo- mesenteroides DSM 20193, Leur. fallux DSM 20189 and the strains NWC 576 and NWC 541, identified as Leuronostor spp., showed unique and specific patterns distinct from those of all the other strains analysed.
Low-frequency restriction fragment analysis of genomic DNA
Results obtained after digestion of genomic DNA with ,4paI and separation by pulsed-field gel electrophoresis are depicted in Fig.4 and summarized in Table 2. The 30 strains could be allocated to 13 pattern groups, a result confirmed after digestion with SmaI (data not shown).
Thirteen strains of Leuc. mesenteroides ssp. mesenteroides, isolated from natural wheys, field grass and water-buffalo milk, displayed the same restriction pattern, while the strain DSM 20343 showed a unique fingerprint and the strain NWC 574 exhibited a pattern similar to Leuconostor spp. strains NWC 579 and NWC 585. Strain NWC 543 was included in the group containing Leuc. citreum DSM 20188 and Leu- conostor spp. NWC 5 11 and NWC 536. The remaining strains from the DSM collection and two strains identified as Leu- ronostor spp. (NWC 541 and NWC 576), showed specific and unique patterns distinct from those of all the other strains analysed.
Ribosomal RNA gene restriction fingerprinting
HindIII digests (Fig. 5) of the rRNA genes of the 30 strains produced nine distinct ribotypes as summarized in Table 2.
0 1997 The Society for Applied Bacteriology, Journal of Applied Microbiology 82, 578-588
CHARACTERIZATION OF LEUCONOSTOC MESENTEROIDES 583
a b c d e f g M a b c d e f a h i l
-97.4
- 66 - 45
- 31
h i I m n o
,(. . - 97.4
- 66 - 45
- 31
Fig. 2 SDS-PAGE of whole-cell proteins of Leuconustor strains representing the patterns reported in Table 2. Lanes: a, Leuc. mesentrroides ssp. mesentrroides DSM 20343 (pattern 11); b, Leur. lartis DSM 20202 (pattern IV); c, Leur. pseudomesenteroides DSM 20193 (pattern V); d, Leuconostor spp. NWC 536 (pattern XI); e, Leur. mesenteroides ssp. mrsenteruidrs NWC 543 (pattern VIII); f, Leurunustor spp. NWC 51 1 (pattern S); g, Leuionortor spp. NWC 541 (pattern X I ) ; h, Leuconostur spp. NWC 576 (pattern SIII); i, Leuronostoc spp. NWC 579 (pattern IS); 1, W2isselln (Lrur.) pclrnmesenteroides DSM 20288 (pattern VII); m, Leuc.fullnx DSM 20189 (pattern VI); n, Leur. cztreum DSM 20188 (pattern 111); o, Leuronostor spp. DSM 20186 (pattern I). Low range (Bio-Rad) was used as molecular weight marker
All strains of Leur. mesenteroides, including Leur. mesenteroides ssp. mesenteroides DSM 20343 and Leur. mesenteroides ssp. dertranir.um DSM 20484, displayed the same ribopattern (pattern F) with the exception of a fragment located at about 4.3 kbp that characterized the subpattern F,, which was also observed in Leur. lurtis DSM 20202. Only two strains, ident- ified as Leur. mesenteroides ssp. mesenteroides, did not fall into the pattern F: strain N W C 543 was placed in the group that included II‘eissella (Leur.) parnmesenteroides D S M 20288, Leur. ritreum DSM 20188, Leuronostor spp. strains N W C 51 1 and N W C 536; strain N W C 574 exhibited the same pattern as Leuronostor spp. strains N W C 579 and N W C 585.
Leuronostor spp. DSM 20186, Leur. mesenteroides ssp. rrem- oris DSM 20346, Leur. fallas DSM 20189, Leu. pseudo- mesenteroides DSM 20193 and strains N W C 541 and N W C
Fig. 3 DNA restriction pattern after digestion with HindIII from Leuronostoc strains representing the patterns reported in Tablc 2. Lanes: a, Leuconustor spp. DSM 20186 (pattern a); b, Lrur. ritreum DSM 20188 (pattern b); c, Leuronostor spp. NWC 541 (pattern c); d, Leuconustor spp. NWC 576 (pattern d); e, Lrur. mesenteroides ssp. rremoris DSM 20346 (pattern e); f, Lrur. mesrnteroides ssp. drxtraizirum DSM 20484 (pattern f); g, Leuronostoc spp. NWC 579 (pattern g); h, Leur. lartis DSM 20202 (pattern h); i, Leur. pseudumesentevoides DSM 20193 (pattern i); 1, Lrus. f u l la r DSM 20189 (pattern 1). 1 DNA cleaved by HindIII was used as molecular weight marker
576, identified as Leuronostor spp., showed a high degree of polymorphism, displaying six distinct ribotypes.
Amplified rDNA restriction analysis (ARDRA)
PCR was used for the amplification of rRNA gene fragment encompassing the 16S, 16s-23s spacer region and part of the 23s. This fragment of 2 2 4 kbp was analysed by diges- tion with HindIII and HaeIII.
After digestion with Hind111 two different patterns were observed (Fig.6 and Table 2): the designated group ‘x’ included Leuconostor spp. DSM 20186, Leur. ritreum DSM 20188, Leur. fallax DSM 20189, Ilkissrlla (Leur . ) pnrii- mesenteroides DSM 20288, two strains identified as Leur. mesenteroides ssp. mesenteroides (NWC 543 and N W C 574) and four strains classified as Leuronostoc spp. (NWC 579, N W C 585, N W C 536 and N W C 51 1); the designated group ‘y’ included 15 strains of Leur. mesenteroides (including the reference strains of the three subspecies), Leur. Iactis DSM 20202, Leuc. pseudomesentrroides DSM 20193 and two strains identified as Leuronostor spp. ( N W C 541 and N W C 576). Within group ‘y’ a slight variation in the pattern was apparent in two strains from the DSM collection (Leur. mesentrroirlrs ssp. rremoris DSM 20346 and Leur. pseudomesenteroide.~ DSM
0 1997 The Society for Applied Bacteriology, Journal of Applied Microbiology 82, 578-588
Tabl
e2 P
rote
in p
rofil
e, D
NA
fing
erpr
intin
g an
d an
alys
is o
f rD
NA
of r
efer
ence
stra
ins
and
of is
olat
es id
entif
ied
as m
embe
rs o
f the
gen
us L
euro
nosto
L -
Spec
ies
Sour
ce
Leuc
onos
tor
spp.
Le
uc. m
esen
tera
ides
subs
p. m
esen
teroi
des
Leuc
. mes
enter
oide
s su
bsp.
cre
moris
La
c. m
esen
teroi
des
subs
p. d
extr
anir
um
Leuc
. citr
eum
La
c. la
ctis
Leuc
. pse
udom
esen
teroi
des
Leuc
. fallax
LI eiss
ella
(Leu
r.) p
aram
esen
tero
ides
Le
uc. m
esen
teroi
des
subs
p. m
esen
terai
des
Leuc
. mes
enter
oide
s su
bsp.
mes
enter
oide
s Le
uc. m
esen
teroi
des
subs
p. m
esen
terai
des
Leuc
. mes
enter
oide
s su
bsp.
mes
enter
oide
s Le
uc. m
esen
terai
des
subs
p. m
esen
tero
zdes
Le
ur. m
esen
tero
ides
subs
p. m
esen
teroi
des
Leuc
. mes
enter
oide
s su
bsp.
mes
ente
roid
es
Leuc
. mes
enter
aide
s su
bsp.
mes
enter
oide
s Le
uron
osto
r sp
p.
Leur
onos
toc
spp.
Le
ucon
osto
r sp
p.
Leur
onos
toc
spp.
Le
uron
osto
c sp
p.
DS
M
DSM
D
SM
D
SM
D
SM
D
SM
D
SM
D
SM
D
SM
F
G
FG
N
WC
N
WC
W
BM
W
BM
N
WC
N
WC
N
WC
N
WC
N
WC
N
WC
N
WC
Stra
ins
2018
6 20
343T
20
346T
20
4WT
2018
8 20
202T
20
193T
20
189T
20
2881
50
1 50
2-50
5 51
0-53
5-52
&54
9 51
5-52
1-57
0-57
7 53
0 53
2 54
3 57
4 57
9-58
5 51
1 53
6 54
1 57
6
Bio
chem
ical
SD
S PF
GE
R
ibot
ypin
g A
RD
RA
A
RD
RA
gr
oup"
PA
GE
Hin
dIII
<.
lpaI
H
indI
II
Hin
dIII
H
ue11
1 ~
11
1 8 9 13
10
15
14
12
1 2 1 2 1 2 1 3 4 5 5 6 7
I a
I1
f I1
e
I1
f I11
b
IV
h V
1
VI
1 V
II
b I1
f
I1
f I1
f
I1
f I1
f
I1
f V
III
b IX
b g
IX
g x
b X
I b
XI1
C
XI11
d
1 A
8 F
6
E 7
F 2
B
11
F1
13
I 12
H
3
B 9
F1
9 F1
9
F1
9 F1
9
F1
9 F1
2
B
1 Ob
G
10
G
2 B
2
B
4 C
5
D
-
X Y Y Y Yl
X Yl
X X Y Y Y Y Y Y X X X X X Yl
Y
~
X Y P Y Y Y X X X Y Y Y p .v Y X X X X X Y Y
"See
Tab
le 1
. FG
, Gra
ss fi
eld;
NW
C, n
atur
al w
hey
cultu
re; W
BM
, wat
er-b
uffa
lo m
ilk; D
SM, D
euts
che
Sam
mlu
ng v
on M
ikro
orga
nism
en.
TT
ype
stra
in.

CHARACTERIZATION OF LEUCONOSTOC MESENTEROIDES 585
kb
23 - 9.6 - 6.6 -
M a b c d e f n h i I m n o D M kb
-150 -100 - 50
1200
a b c d e f g h i I kb
23 - 9.4- 6.6 - 4.3 -
2.3 - 2.0-
0.5-
Fig. 4 Pulsed-field gel electrophoresis of DNA restriction pattern after digestion with -$pa1 from Leuconustor strains representing the patterns reported in Table 2. Lanes: a, Leztzunustor spp. DSM 20186 (pattern 1); b, Leur. rnesenteroides ssp. mesenteruides DSM 20343 (pattern 8); c, Leuc. mesenterozdes ssp. rremoris DSM 20346 (pattern 6); d , Leuc. mesenteroides ssp. dextranzrum DSM 20484 (pattern 7); e, Leuc. ritreum DSM 20188 (pattern 2); f, Leur. mesenteroides ssp. mesenteroides NWC 570 (pattern 9); g, Leuconustor spp. NWC 579 (pattern 10); h, Leuc. mesenteroides ssp. mesenteroides NWC 574 (subpattern lob); i, L e u . lai.tis DSM 20202 (pattern 11); 1, Leuronostoc spp. NWC 576 (pattern 5); m, Leuronostor spp. NU'C 541 (pattern 4); n, U'eissella (Leur.) paramesenteroides DSM 20288 (pattern 3); 0,
Leur. fulliix DSM 20189 (pattern 12); p, Leur. pseudomesenteroides DSM 20193 (pattern 13). DNA cleaved by HindIII and 1 concatamers were used as molecular weight markers
20193) and in the strain NWC 541 identified as Leuronostor spp. (subgroup yl).
The results obtained after digestion of PCR products with Hue111 (Fig. 6 and Table 2) also exhibited two patterns com- parable to those obtained after digestion with HindIII.
DISCUSSION
As noted by other workers (Holzapfel and Schillinger 1992) the results confirm that traditional biochemical and physio- logical tests are unsatisfactory for the identification of leu- conostocs. More than 17% of the 215 isolates from natural whey starters, water-buffalo milk and field grass showed inconsistent biochemical properties, with reference to known species.
Leuronostor mesenferoides ssp. mesenteroides was by far the most frequently detected taxon, as 178 isolates could be ident- ified as belonging to it, with the acceptance of variability in ribose fermentation.
Natural whey cultures utilized as starter culture in the traditional manufacture of water-buffalo Mozzarella cheese exhibited the highest diversity; seven different biochemical groups were isolated. Of these, four could not be identified
Fig. 5 Autoradiograph of HindIII digests of DNA from Leuconostor strains representing the patterns reported in Table 2. Hybridization was performed with 32P-labelled cDNA probe of 16 + 23s rRNA from Esrhevirhia cola. Lanes: a, Leuronostor spp. DSM 20186 (pattern A); b, Leur. citreum DSM 20188 (pattern B); c, Leuconustor spp. NWC 541 (pattern C); d, Leuconustor spp. NWC 576 (pattern D); e, L a c . mesenterozdes ssp. rremoris DSM 20346 (pattern E); f, Leui.. mesenteroides ssp. dextranirum DSM 20484 (pattern F); g, Leur. mesenteroiiles ssp. mesenteroides NWC 570 (subpattern FI); h, Leuronostor spp. NWC 579 (pattern G); i, L e u . fullax DSM 20189 (pattern H); 1, Leur. pseudomesenteroides DSM 20193 (pattern I). 1 DNA cleaved by HindIII was used as molecular weight marker
according to the criteria of Garvie (1986), Holzapfel and Schillinger (1992) and Dicks et al. (1993).
As reported by other workers (Dicks et al. 1990; Elliott and Facklam 1993; Tsakalidou et al. 199.0, protein profiles appeared suitable for species discrimination, but not for dif- ferentiating the three subspecies of Leur. mesenteroides. This technique also highlighted some differences among strains belonging to Leuc. mesenteroides ssp. mesenteroides which posed identification problems.
High-frequency restriction endonuclease fingerprinting grouped the highest number of Leur. mesenteroides strains (with the exclusion of the two anomalous ones: NWC 543 and 574). However, it also grouped together other leu- conostocs such as Leur. ritreum, Keassel/a purumesenteroides and Leuronostor spp.
Surprisingly, low-frequency restriction fragment analysis of genomic DNA, performed by digestion with both,.lpuI and SmuI, appeared more discriminating than HindIII-digested DNA. Thus, all the species and subspecies of the reference strains were differentiated. Moreover, strains of Leur. mesen- teroides ssp. mesenteroides showed different restriction patterns compared with that of the type strain. Similar results were obtained by Kelly et al. (1995) using PFGE to differentiate Leuronostor strains from fermented rice cake.
Ribotyping was not useful for discriminating species or subspecies of the genus Leuconostor and gave results similar
0 1997 The Society for Applied Bacteriology, Journal of Applied Microbiology82, 578-588

586 F. VILLANI E r A L .
M a b c d e kb
3
2
1.6
1
0.50
0.39
0.34
0.29
0.22 f Fig. 6 Ethidium bromide-stained agarose gel (1.5%0) of PCR products of part of the rDNA of Leuconostor strains representing the patterns reported in Table 2. Digestion with Hind111 (lanes a, b, c) and HaeIII (lanes d, e). Lanes: a, Leur.onostor spp. DSM 20186 (pattern x); b, Leuc. mesentemides ssp. mesenteroides DSM 20343 (pattern y); c, Leuc. mesenteroides ssp. rremoris DSM 20346 (pattern yl); d, Leuronostor spp. DSM 20186 (pattern x); e, Leuc. mesenteroides ssp. mesenteroides DSM 20343 (pattern y). 1 kb DNA ladder was used as molecular weight marker
to those obtained with HindIII-digested DNA. Leuconostoc mesenteroides ssp. mesenteroides and ssp. dextranicum showed the same pattern as Leur. lactis while Leur. mesrnteroides ssp. cremoris exhibited a pattern distinct from the other species examined.
On the basis of ARDRA-PCR, two main groups could be distinguished: the major one, included Leur. mesenteroides, Leur. lactis, Leur. pseudomesenteroides and some unidentifiable strains; the second one, included Leuc. ritreum, Leur. fal lux, It ’eissellu and other unidentifiable strains.
The results indicate that molecular biology analyses are
0 1997 The SOC
useful to confirm and extend the identification of Leuronostoc strains based on conventional biochemical tests. This approach allows the identification of some equivocal isolates and discriminates unidentifiable strains that require further studies in view of the proposals for new species within this genus.
ACKNOWLEDGEMENTS
This work was supported by a grant from the National Research Council of Italy, Rome, Special Project RAISA, Sub-project 4, paper no. 2662. The authors thank Drs Maria Aponte, Giorgio Notaro and Giovanni Salzano for scientific and technical collaboration and Mark Walters for final proof- reading of the manuscript.
REFERENCES
Benkerroum, N., Misbah, M., Sandine, W.E. and Elaraki, A.T. (1993) Development and use of a selective medium for isolation of Leuconostor spp. from vegetables and dairy products. d-ipplaed and Enzironmental MirrobioloEy 59, 607-609.
Cogan, T.M. and Jordan, K.N. (1994) Metabolism of Leuconostnr bacteria. Journal of Dairy Science 77, 2704-2717.
Collins, M.D., Rodriguez, U., Ash, C., Aguirre, M., Farrow, J.A.E., Martinez-Murcia, A.J. et al. (1991) Phylogenetic analysis of the genus Lartobacillus and related lactic acid bacteria as determined by reverse transcriptase sequencing of 16s rRNA. FE,ZlS Micm- biology Letters 77, 5-12.
Collins, M.D., Samelis, J., Metaxopoulos, J. and Wallbanks, S. (1993) Taxonomic studies on some Leuconustor-like organisms from fermented sausages: description of a new genus It’eissella for the Leuronostor puramesenteroides group of species. Journal uf .4pplied Bacteriology 75, 59 5-603.
De Buyer, M.-L, Morvan, A,, Aubert, S., Grimont, F. and El Solh, N. (1989) Characterization of Stuph,ylurorrus species by ribosomal RNA gene restriction patterns. Journal of General hlicrobiology 135, 989-999,
Dicks, L.M.T., Van Vuuren, H.J.J. and Dellaglio, F. (1990) Tax- onomy of Leuconostor species, particularly Leuronostor oenos, as revealed by numerical analysis of total soluble cell protein patterns, DN14 base compositions and DNA-DNA hybrid- izations. International Journal ojS,~~stematic Bacteriology 40, 83- 91.
Dicks, L.M.T., Fantuzzi, L., Gonzales, F.C., Du Toit, M. and Dellaglio, F. (1993) Leuronostor argentinurn sp. nov., isolated from argentine raw milk. Internutional Journul of Systemutir Bar.- teriology 43, 347-351.
Dicks, L.M.T., Dellaglio, F. and Collins, M.D. (1995) Proposal to reclassify Leuconostor oenos as Oenororcus oeni (corrig) gen. nov., comb. nov. International Journal cf Systematic Bacteriology 45, 395-397.
Elliott, J.A. and Facklam, R.R. (1993) Identification of Leuronostoc spp. by analysis of soluble whole-cell protein patterns. Journul of Clinical hlirrobiology 3 1, 1030-1033.
:iety for Applied Bacteriology, Journal of Applied Microbiology 82, 578-588

C HA R ACT€ R I ZAT I 0 N 0 F L EUCO NOS TOC M ES EN TEROlD ES 587
Farrow, J.A.E., Facklam, R.R. and Collins, M.D. (1989) Nucleic acid homologies of some vancom ycin-resistant leuconostocs and description of Leuconostoc citreum sp.nov. and Leuconostoc pseudo- mesenteroides. International Journal of S,ystematic Bacteriology 39,
Gar\-ie, E.I. (1981) Sub-division within the genus Leuconostoc as shown by RNA-DNA-hybridization. Journal of General Micro- bioloKy 127, 209-212.
Garvie, E.I. (1986) Genus Leuconostoc. In Bergey ’s Manual of Sy- tematic Bacteriology ed. Sneath, P.H.A., Mair, N.S., Sharpe, M.E. and Holt, J.G. Vol. 2, pp. 1071-1075. Baltimore: Williams and Wilkins.
Grimont, F. and Grimont, P.A.D. (1986) Ribosomal ribonucleic acid gene restriction patterns as potential taxonomic tools. Annales Institut Pastew/ Microbiology 137B, 165- 175.
Gurtler, V., Wilson, V.A. and Mayall, B.C. (1991) Classification of medically important clostridia using restriction endonuclease site differences of PCR-amplified 16s rDNA. Journal qf’ General Microbiology 137, 2673-2679.
Harrigan, W.F. and McCance, M. (1976) Laboratocy Methods in Daivy and Food Microbiology. pp. 32&321. London: Academic Press.
Holzapfel, W.H. and Schillinger, V. (1992) The genus Leuconostoc. In The Prokaryotes ed. Balows, A., Triiper, H.G., Dworkin, M., Harder, W. and Schleifer, K. H. Vol. 2, pp. 1508-1534. New York: Springer-Verlag.
Irino, K., Grimont, F., Casin, I. and Grimont, P.A.D. (1988) The Brazilian Purpuric Fever Study Group: RNA gene restriction patterns of Haemophilus inflzenzae biogroup aegyptius strains associated with Brazilian purpuric fever. Journal ofclinical Mzcro- biolugy 26, 1535-1538.
Jayarao, B.M., Dore, J.J.E., Baumbach, G.A., Matthews, K.R. and Oliver, S.P. (1991) Differentiation of Streptococcus uberis from Streptococcus parauberis by polymerase chain reaction and restric- tion fragment length polymorphism analysis of 16s ribosomal DNA. Journal of Clinical Microbiology 29, 2774-2778.
Kelly, W.J., Huang, C.M. and Asmundson, R.V. (1993) Comparison of Leuconostoc oenos strains by pulsed-field gel electrophoresis. Applied and Environmental Microbiology 59, 3969-3972.
Kelly, W.J., Asmundson, R.V., Harrison, G.L. and Huang, C.M. (1995) Differentiation of dextran-producing Leuconostoc strains from fermented rice cake @to) using pulsed-field gel elec- trophoresis. International Journal of Food Microbiology 26, 345- 352.
Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227,680-685.
Lamoureux, M., Prevost, H., Cavin, J.F. and Divies, C. (1993) Recognition of Leuconostoc oenos strains by the use of DNA restriction profiles. Applied Microbiology and Biotechnology 39,
Marmur, J. (1961) A procedure for the isolation of deoxyribonucleic acid from microorganisms. Journal of Molecular Biology 3, 208- 218.
Martinez-Murcia, A. and Collins, M.D. (1990) A phylogenetic analysis of the genus Leuconnstoc based on reverse transcriptase sequencing of 16s rRNA. F E M S Microbiology Letters 70, 73-84.
Martinez-Murcia, A. and Collins, M.D. (1991) Phylogenetic analy-
279-283.
547-552.
sis of an atypical leuconostoc: description of Leuconostoc fallax sp. nov. F E M S Microbiology Letters 82, 55-60.
Martinez-Murcia, A.J., Harland, N.M. and Collins, M.D. (1993) Phylogenetic analysis of some leuconostocs and related organisms as determined from large sub-unit rRNA gene sequences: assess- ment of congruence of small- and large-subunit rRNA derived trees. Journal of Applied Bacteriology 74, 532-541.
Mayeux, J.V., Sandine, W.E. and Elliker, P.R. (1962) A selected medium for detecting leuconostoc organisms in mixed strains starter cultures. Journal of Dairy Science 45, 655-656.
McClelland, M., Jones, R., Patel, Y. and Nelson, M. (1987) Restricton endonuclease for pulsed field mapping of bacterial genomes. Nucleic Acids Research 15, 5985-6005.
Moschetti, G., Salzano, G., Villani, F., Pepe, O., Andolfi, R., Mau- riello, G. and Coppola, S. (1995) Characterization of dairy enter- ococci by restriction endonuclease and rDNA fingerprinting. Annali di Microbiologia ed Enzimologia 45, 1-12.
Salzano, G., Moschetti, G., Villani, F. and Coppola, S. (1993) Biotyping of Streptococcus thermophilus strains by DNA finger- printing. Research in Microbiology 144, 381-387.
Salzano, G., Moschetti, G., Villani, F., Pepe, O., Mauriello, G. and Coppola, S. (1994) Genotyping of Streptococcus thermophilus evidenced by restriction analysis of ribosomal DNA. Research in Microbiology 145, 651-658.
Schillinger, U. and Lucke, F.-K. (1987) Identification of lactobacilli from meat and meat products. Food Microbiology 4, 199-208.
Schillinger, U., Holzapfel, W. and Kandler, 0. (1989) Nucleic acid hybridization studies on Leuconostoc and heterofermentative lactobacilli and description of Leuconostoc amelobiosum sp. nov. Systematic and Applied Microbiology 12, 48-55.
Southern, E. (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. Journal of Molecular
Sperber, W.H. and Swan, J. (1976) Hot loop test for the deter- mination of carbon dioxide production from glucose by lactic acid bacteria. Applied und Environmental Microbiology 31,990-991.
Takahashi, M., Okada, S., Uchimura, T. and Kozaki, M. (1992) Leuconostoc amelibiosum Schillinger, Holzapfel and Kandler, 1989 is a later subjective synonym of Leuconostoc citreum Farrow, Fack- lam and Collins, 1989. International Journal of Systematic Bac- teriology 42, 649-65 1.
Tanskanen, E.I., Tulloch, D.L., Hillier, A.J. and Davidson, B.E. (1990) Pulsed-field gel electrophoresis of SmaI digests of lac- tococcal genomic DNA, a novel method of strain identification. Applied and Environmental Microbiology 56, 3105-31 11.
Tenreiro, R., Santos, M.A., Paveia, H. and Vieira, G. (1994) Inter- strain relationships among wine leuconostocs and their divergence from other Leuconostoc species, as revealed by low frequency restriction fragment analysis of genome DNA. Journal ofApplied Bacteriology 77, 271-280.
Thomson-Carter, F.M., Carter, P.E. and Pennington, T.H. (1989) Differentation of staphylococcal species and strains by ribosomal RNA gene restriction patterns. Journal of General Microbiulogj/
Tsakalidou, E., Manolopoulou, E., Kabaraki, E., Zoidou, E., Pot, B., Kersters, K. and Kalantzopoulos, G. (1994) The combined use of whole-cell protein extracts for the identification (SDS- PAGE) and enzyme activity screening of lactic acid bacteria iso-
Biology 98, 503-517.
135,2093-2097.
0 1997 The Society for Applied Bacteriology, Journal of Applied Microbiology82, 578-588

588 F. V lLLANl E T A L
latcd from traditional Greek dairy products. Systematic and .+plied .Clicrobiology 17, 44-58,
Vaneechoutte, M., Rossau, R., De Vos, P., Gillis, M., Janssens, D., Paepe, N. et al. (1992) Rapid identification of bacteria of the Comumonadaceae with amplified ribosomal DNA-restriction analysis (ARDRA). FEMS iZlicrobiology Lettevs 93, 227-234.
Vedamuthu, E.R. (1994) The dairy Leuconustor: use in dairy prod- ucts. Journml ofDaiyy Science 77,2725-2737.
Viti, C., Ventura, S., Granchi, L. and Giovannetti, L. (1995) Gen-
ome diversity in several strains of Leuconostoc oenos from the Chianti region. *.lnnali di Mirrobiologia ed Encimologia 45, 151- 158.
Von L u s h , U. and Lompe, A. (1982) Schnellest zum qualitativen Nachweis von L (+) und D (-) Milchsaure fur die Bestimmung van Milchesaurebakterien. MilchDissenschajl37, 65-68.
Yang, D. and Woese, C.R. (1989) Phylogenetic structure of the ‘Leuconustor’: an interesting case of rapidly evolving organism. Systematic and -4pplied Microbiology 12, 145-149.
0 1997 The Society for Applied Bacteriology, Journal of Applied Microbiology82, 578-588




















