Characterization of serotonin transporter in blood lymphocytes of rats. Modulation by in vivo...
10
Characterization of serotonin transporter in blood lymphocytes of rats. Modulation by in vivo administration of mitogens Nemesio Ceden ˜o 1 , Mary Urbina, Francisco Obrego ´ n, Lucimey Lima * Laboratorio de Neuroquı ´mica, Centro de Biofı ´sica y Bioquı ´mica, Instituto Venezolano de Investigaciones Cientı ´ficas, Apdo. 21827, Caracas 1020-A, Venezuela Received 18 June 2004; received in revised form 16 September 2004; accepted 17 September 2004 Abstract Serotonin transporter sites were characterized in blood lymphocytes of rats. Pharmacological characteristics of drug interactions were in concordance with recent studies in nervous and human immune cells. The potency order of inhibition of [ 3 H]paroxetine binding was imipramineNcitalopramNalaproclateNserotonin. Selective inhibitors of dopamine or noradrenaline transporters did not inhibit it. The specific binding of [ 3 H]paroxetine was higher at intermediate than at low concentrations, and the plot of free vs. specific binding had a sigmoid shape. The affinity constant or K d , 1.77 nM, was in close agreement with data obtained from kinetic studies (K d =1.33 nM), which evidences that the equilibrium was reached. In addition, serotonin transporter was evaluated by lipopolysaccharide or concanavalin A administration in vivo (0.1 mg/kg, i.p., 18 h). After the treatment with lipopolysaccharide, no changes were observed in the numbers of sites or B max or in the affinity, K d . The treatment with concanavalin A showed a significant reduction in B max and reduction in K d . Additionally, serotonin and 5- hydroxyindoleacetic acid levels were determined in plasma and lymphocytes by high-performance liquid chromatography. Treatment with lipopolysaccharide produced a significant increased of serotonin levels in lymphocytes without changes in 5-hydroxyindoleacetic acid level; in plasma, it produced an increase in serotonin and 5-hydroxyindolacetic acid levels. In addition, serotonin synthesis was evaluated by adding 300 AM of tryptophan in the medium, which significantly increased serotonin levels in control lymphocytes. Moreover, the concentrations of 5-hydroxyindoleacetic acid was enhanced significantly, both in plasma and lymphocytes in the presence of tryptophan after treatment with lipopolysaccharide. The administration of concanavalin A significantly decreased plasma levels of serotonin, as well as the concentrations of serotonin and 5-hydroxyindoleacetic acid in lymphocytes. These results demonstrate the presence of serotonin transporter in lymphocytes of rat blood, the capacity for serotonin synthesis in lymphocytes, and the modulation of these parameters by systemic administration of mitogens. The findings of this work contribute to understanding the immunological role of serotonin and the communication of immune and nervous systems. D 2004 Elsevier B.V. All rights reserved. Keywords: Concanavalin A; Lipopolysaccharide; Lymphocytes; Serotonin; Serotonin synthesis; Serotonin transporter 1. Introduction Evidences for communication between immune and nervous systems have been described, in which pro- nounced effects of neurotransmitters on immune functions are associated (Haas and Schauenstein, 1997). Serotonin (5-HT) is one of those neurotransmitters affecting the immune system (Jackson et al., 1985). During the last 20 years, 5-HT has been reported to play an inhibitory role in the regulation of the immune system (Jackson et al., 1985; Slauson et al., 1984; Sternberg et al., 1986; Pellegrino and Bayer, 2000). On the other hand, new proofs suggest that 5-HT up-regulates natural killer (NK), T and B cells proliferation (Hellstrand and Hermmodsson, 1990; Iken et al., 1995; Laberge et al., 0165-5728/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.jneuroim.2004.09.010 * Corresponding author. Tel.: +58 212 504 1213; fax: +58 212 504 1295. E-mail address: [email protected] (L. Lima). 1 Present address: Universidad Ro ´ mulo Gallegos, Departamento de Ciencias Morfolo ´gicas y Funcionales, San Juan de los Morros, Gua ´rico 2301, Venezuela. Journal of Neuroimmunology 159 (2005) 31 – 40 www.elsevier.com/locate/jneuroim
Transcript of Characterization of serotonin transporter in blood lymphocytes of rats. Modulation by in vivo...

www.elsevier.com/locate/jneuroim
Journal of Neuroimmunolo
Characterization of serotonin transporter in blood lymphocytes of rats.
Modulation by in vivo administration of mitogens
Nemesio Cedeno1, Mary Urbina, Francisco Obregon, Lucimey Lima*
Laboratorio de Neuroquımica, Centro de Biofısica y Bioquımica, Instituto Venezolano de Investigaciones Cientıficas,
Apdo. 21827, Caracas 1020-A, Venezuela
Received 18 June 2004; received in revised form 16 September 2004; accepted 17 September 2004
Abstract
Serotonin transporter sites were characterized in blood lymphocytes of rats. Pharmacological characteristics of drug interactions were in
concordance with recent studies in nervous and human immune cells. The potency order of inhibition of [3H]paroxetine binding was
imipramineNcitalopramNalaproclateNserotonin. Selective inhibitors of dopamine or noradrenaline transporters did not inhibit it. The specific
binding of [3H]paroxetine was higher at intermediate than at low concentrations, and the plot of free vs. specific binding had a sigmoid shape.
The affinity constant or Kd, 1.77 nM, was in close agreement with data obtained from kinetic studies (Kd=1.33 nM), which evidences that the
equilibrium was reached. In addition, serotonin transporter was evaluated by lipopolysaccharide or concanavalin A administration in vivo
(0.1 mg/kg, i.p., 18 h). After the treatment with lipopolysaccharide, no changes were observed in the numbers of sites or Bmax or in the
affinity, Kd. The treatment with concanavalin A showed a significant reduction in Bmax and reduction in Kd. Additionally, serotonin and 5-
hydroxyindoleacetic acid levels were determined in plasma and lymphocytes by high-performance liquid chromatography. Treatment with
lipopolysaccharide produced a significant increased of serotonin levels in lymphocytes without changes in 5-hydroxyindoleacetic acid level;
in plasma, it produced an increase in serotonin and 5-hydroxyindolacetic acid levels. In addition, serotonin synthesis was evaluated by adding
300 AM of tryptophan in the medium, which significantly increased serotonin levels in control lymphocytes. Moreover, the concentrations of
5-hydroxyindoleacetic acid was enhanced significantly, both in plasma and lymphocytes in the presence of tryptophan after treatment with
lipopolysaccharide. The administration of concanavalin A significantly decreased plasma levels of serotonin, as well as the concentrations of
serotonin and 5-hydroxyindoleacetic acid in lymphocytes. These results demonstrate the presence of serotonin transporter in lymphocytes of
rat blood, the capacity for serotonin synthesis in lymphocytes, and the modulation of these parameters by systemic administration of
mitogens. The findings of this work contribute to understanding the immunological role of serotonin and the communication of immune and
nervous systems.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Concanavalin A; Lipopolysaccharide; Lymphocytes; Serotonin; Serotonin synthesis; Serotonin transporter
1. Introduction
Evidences for communication between immune and
nervous systems have been described, in which pro-
0165-5728/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.jneuroim.2004.09.010
* Corresponding author. Tel.: +58 212 504 1213; fax: +58 212 504
1295.
E-mail address: [email protected] (L. Lima).1 Present address: Universidad Romulo Gallegos, Departamento de
Ciencias Morfologicas y Funcionales, San Juan de los Morros, Guarico
2301, Venezuela.
nounced effects of neurotransmitters on immune functions
are associated (Haas and Schauenstein, 1997). Serotonin
(5-HT) is one of those neurotransmitters affecting the
immune system (Jackson et al., 1985).
During the last 20 years, 5-HT has been reported to
play an inhibitory role in the regulation of the immune
system (Jackson et al., 1985; Slauson et al., 1984;
Sternberg et al., 1986; Pellegrino and Bayer, 2000). On
the other hand, new proofs suggest that 5-HT up-regulates
natural killer (NK), T and B cells proliferation (Hellstrand
and Hermmodsson, 1990; Iken et al., 1995; Laberge et al.,
gy 159 (2005) 31–40

N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–4032
1996; Eugen-Olsen et al., 1997). Recently, inhibition of
fish T-cell proliferation by p-clorophenylalanine, an
inhibitor of 5-HT synthesis, and its reversal by exogenous
5-HT has been showed, which suggest that 5-HT is
implicated in T cell proliferation stimulated by phytohe-
maglutinin (PHA) (Ferriere et al., 1999). Nonetheless, a
biphasic dose–response effect of 5-HT on immune cells
activity has been described (Kut et al., 1992). Low dose
of 5-HT has a stimulatory effect whereas high dose has a
suppressive effect (Stefulj et al., 2001).
High concentrations of 5-HT are associated with capital
immune functions. For example, 5-HT induces release of
interleukine-16 (IL-16), cellular recruitment and release of
chemotactic factors and cytokines, as observed in the
inflammatory response (Laberge et al., 1996), and defends
NK cells from injure at inflammatory sites (Betten et al.,
2001). Additional evidences suggest that 5-HT might be
potentially participating in the development of cell-
mediated immune responses (Askenase, 1988; Matsuda
et al., 1997). Moreover, various reports further document
a wide range of 5-HT immune functions mediated through
interaction with 5-HT1A receptor subtype (Iken et al.,
1995; Eugen-Olsen et al., 1997), which has been
pharmacologically demonstrated on activated human T
cells (Aune et al., 1993), in rainbow trout peripheral
blood lymphocytes (Ferriere et al., 1996), after mitogenic
stimulation, in activated B and T lymphocytes of mice
(Abdouh et al., 2001), rats (Sempere et al., 2003), and
humans (Urbina et al., 1999) resting lymphocytes.
Thus, 5-HT is capable of producing different effects on
the immune system; however, a bewildering number of
signals are required in order to stimulate immune cells
(Mossner et al., 2001). One of the important character-
istics in 5-HT signaling is the precise function of 5-HT
transporter (5-HTT) that modulates the signaling and
determines the magnitude and length of serotonergic
responses. In fact, 5-HTT is present in the central nervous
system and in non-neuronal cells. For instance, 5-HTT
has been reported in human blood platelets (Rudnick,
1997), goldfish retina (Lima and Schmeer, 1994), adrenal
chromaffin cells (Blakely et al., 1995), and in the crypt
epithelial cells in the gastrointestinal tract (Jackson et al.,
1985; Wade et al., 1996). Similarly, 5-HTT has been
reported in immune cells; for example, murine (Bonnet et
al., 1984), rainbow trout (Ferriere et al., 1999), and
human lymphocytes (Faraj et al., 1997; Urbina et al.,
1999; Lima and Urbina, 2002). This emphasizes the
potential importance of 5-HT as immunomodulator. More-
over, 5-HTT was reported as a biochemical and pharma-
cological-modulated marker in lymphocytes of depressed
patients under pharmacological treatment (Lima and
Urbina, 2002; Urbina et al., 1999).
Due to the potential importance of 5-HT as an
immunomodulator, the aim of this work was the character-
ization of 5-HTT in blood lymphocytes of rats, which is an
unchallenged field. In addition, other objectives were to
determine 5-HT and 5-hydroxyindolacetic acid (5-HIAA)
concentrations in lymphocytes and in plasma, to study the in
vivo modulation of 5-HTT by mitogens, such as lip-
opolysaccharide (LPS) from E. coli serotype 0111:B4 and
concanavalin A (Con A), and to evaluate 5-HT synthesis
from tryptophan in lymphocytes of the rats.
2. Materials and methods
2.1. Chemicals
[3H]Paroxetine (17.1 Ci/mmol) was purchased from New
England Nuclear (Boston, MA, USA). The following
compounds were obtained from Sigma (St. Louis, MO,
USA): 5-HT creatine sulfate, desipramine HCl, imipramine
HCl. Alaproclate, GBR 12909, and maleate of nomifensine
were purchased from Research Biochemical. NycoPrep
gradient lymphocyte medium was obtained from Nycomed
Pharma (Oslo, Norway). All other chemicals were standard
laboratory reagents of analytical grade.
2.2. Experimental animals
Male Sprague–Dawley rats (Rattus norvegicus) ranging
in weight from 200 to 250 g were obtained from the
Instituto Venezolano de Investigaciones Cientıficas (IVIC)
hatchery. The animals were housed individually and were
kept under conventional conditions with commercial rat
food and tap water ad libitum.
2.3. Isolation of lymphocytes
Blood was taken by intracardial puncture, collected in
tubes and centrifuged at 500�g for 10 min in order to
separate white and red blood cells from the plasma. Next,
the white cell layer were withdrawn with a Pasteur pipette
into a 50-ml polypropylene graduated conical tube with
sodium phosphate saline buffer 0.1 M, pH 7.4 (PBS).
This preparation was centrifuged at 500�g for 30 min at
room temperature over a density gradient of NycoPrep.
Mononuclear cell layer was obtained by gradient centri-
fugation, harvested, washed twice with PBS, and centri-
fuged at 500�g for 10 min. To achieve a purified
lymphocyte preparation with a minimal monocyte con-
tamination, the resulting pellet was diluted with Roswell
Park Memorial Institute Medium 1640 (RPMI) free of
fetal bovine albumin and incubated into a culture flask for
45 min at 37 8C and 5% of CO2. After the incubation,
lymphocytes, which are non-adherent cells (~80% of
cells), were dislodged from adherent monocytes (~20%
of cells) by gentle agitation, transferred to plastic tubes
and washed twice. Finally, the pellet was resuspended in
50 mM Tris–HCl, pH 7.4, and the cell viability of
isolated lymphocytes was determined by Trypan blue
exclusion test, and was greater than 90%.

N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–40 33
2.4. Lymphocyte membrane preparation
Lymphocytes pellet (80–90% CD3+) were resuspended
in Tris–HCl 50 mM, pH 7.4, and homogenized in a
Tissumizer (Tekmar, Cincinnati, OH). Then, the preparation
was centrifuged in a refrigerated Sorvall RC-5 at 38,000�g
for 10 min at 4 8C. This step was repeated three times, and
the final pellet was resuspended in 50 mM Tris–HCl, 120
mM NaCl, 5 mM KCl, pH 7.4, and stored at �80 8C.Proteins were measured by the method of Lowry et al.
(1951) using albumin as standard.
2.5. Radioligand binding assays
In order to determine the proper cell concentration, an
experiment with various dilutions of membranes was
performed. Inhibition experiments were carried out with
the aim of determining concentration of drug producing
50% inhibition of binding (IC50). The selectivity of binding
inhibition interaction was performed by incubation of 100 Alof membrane preparation (106 cell/ml) in a final volume of
0.5 ml, with a concentration of 2 nM of [3H]paroxetine, in
the presence of 15 concentrations (0.1–1 mM) of the
following drugs: imipramine, alaproclate and citalopram,
and 5-HT, serotonin reuptake inhibitors (SSRIs); nomifen-
sine and GBR12090, selective inhibitors of dopamine
transporter; and desipramine, inhibitor of noradrenaline
uptake. The reaction was stopped by placing the tubes on
ice and adding cold buffer solution. Separation of the
ligand-binding site complex from the free ligand was carried
out by rapid filtration over Whatman GF/A glass fiber
filters. The filters were washed twice with 5 ml of cold
buffer using a vacuum equipment system (Brandel M-48),
which took less than 20 s, placed on vials, and dried.
Toluene/Omnifluor (0.4%), 4 ml, was added and radio-
activity was measured in a liquid scintillation counter
(Packard Tricarb Model 1900 CR, USA), efficiency 65%.
Saturation experiments were performed and the follow-
ing parameters were determined: the transporter density
(Bmax), the affinity, which is usually expressed as the
dissociation constant at equilibrium (Kd), and the Hill
coefficient (nH) (Bylun and Yamamura, 1990). Saturation
experiments were done with 100 Al of membrane prepara-
tion (106 cells/ml), and in the presence of increasing
concentrations of [3H]paroxetine (0.1–7 nM). The specific
binding was determined with 100 AM imipramine, a
tricyclic antidepressant and inhibitor of 5-HT uptake. The
tubes were prepared in duplicates and were incubated at 25
8C for 90 min since the equilibrium was reached during this
period.
Kinetic experiments were performed in order to deter-
mine the apparent association constant (kobs), and to
measure when the equilibrium was reached (Bylun and
Yamamura, 1990). Lymphocyte membranes were incubated
with 2 nM [3H]paroxetine; specific binding was defined in
the presence of 100 AM alaproclate. Four tubes were used
for each point, two for total binding and two for nonspecific
binding. Incubation times were 5, 10, 15, 20, 30, 60, and 90
min, respectively, at 25 8C. To determine dissociation rate
constant (k2), the membrane preparation was incubated for
90 min and the displacement of ligand bound at equilibrium
was assessed by the addition of 100 AM imipramine in 1 ml
of buffer solution (100 AM).
2.6. Determination of serotonin and 5-hydroxyindoleacetic
acid in plasma and lymphocytes
5-HT and 5-HIAA were determined by reversed-phase
high-performance liquid chromatography (HPLC) with
electrochemical detection. Lymphocytes were collected as
described above, resuspended and homogenized in 500 Al ofmobile phase, which was 0.02 M sodium acetate, 0.0125 M
citric acid, 1 mM ethilendiamine tetra acetic acid (EDTA),
1.52 mM octanylsulfonate, pH 3.9, plus 10% acetonitrile.
After centrifugation, 40 Al of the supernatant was injected
into the chromatographic system. The quantity of 5-HT and
5-HIAA in the lymphocytes was calculated from the area
under the curve of samples and external standards, and
expressed as ng/ml or ng/106 cells, respectively.
2.7. Modulation of serotonin transporter by lipopolysac-
charide and concanavalin A
Rats were treated with LPS 0.1 mg/kg (Linthorst and
Reul, 1998) or with Con A (Canavalia ensiforme, Jack
Bean, Sigma), which were administrated intraperitoneally
(i.p.) diluted in PBS. Controls received the vehicle. After 18
h, the blood samples were taken and lymphocyte mem-
branes were prepared according to the methodology
previously described. The transporter capacity was calcu-
lated by saturation experiments.
2.8. Serotonin synthesis in lymphocytes of rats treated with
lipopolysaccharide by high-performance liquid
chromatography
5-HT synthesis in lymphocytes was evaluated. Previ-
ously, a curve time vs. tryptophan concentration was
performed in order to obtain the optimal incubation time
and tryptophan concentration. Lymphocytes were incu-
bated for 10, 20, 30, and 60 min in the presence or
absence of 100, 200, 300, and 500 AM tryptophan at 37
8C. Then, lymphocytes of rats were incubated in RPMI
free of fetal bovine serum for 20 min at 37 8C in the
presence of 300 AM tryptophan and incubated into a
culture flask for 20 min at 37 8C and 5% of CO2. After
the incubation, lymphocytes were transferred to plastic
tube and washed twice. Finally, the pellet was resus-
pended and homogenized as described to determine 5-HT
intracellular concentration by HPLC. In order to evaluate
the effect of LPS on 5-HT synthesis in lymphocytes, rats
were treated i.p. with LPS, 0.1 mg/kg, as described above.
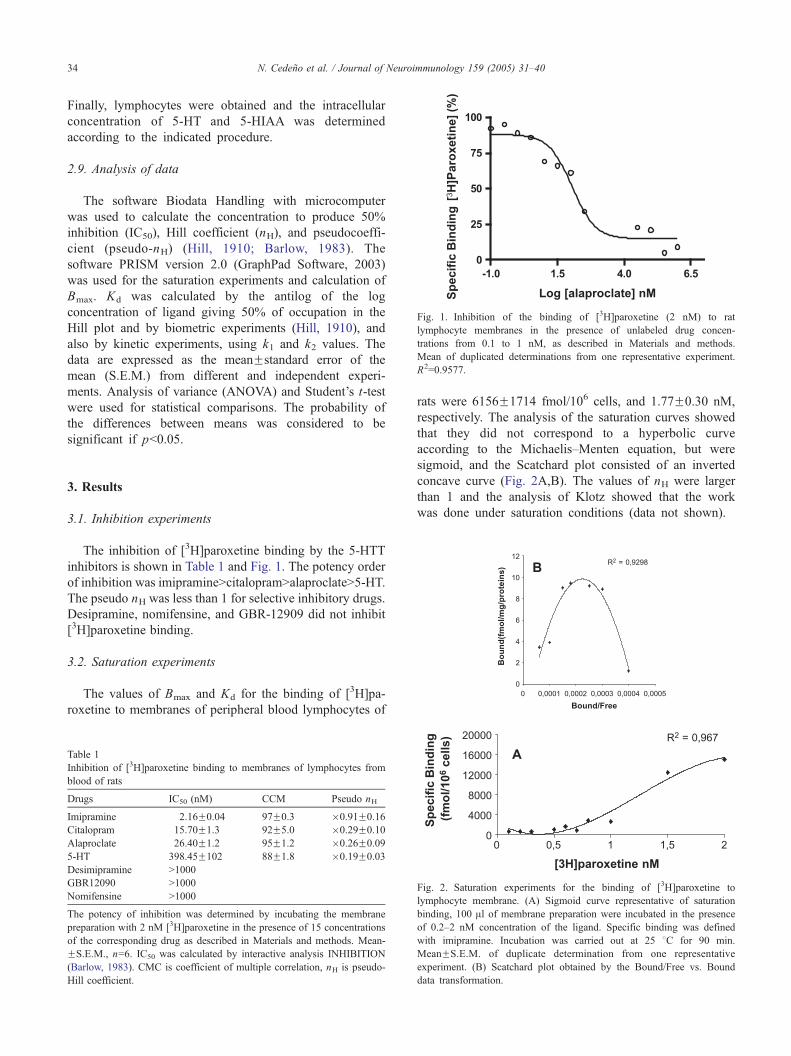
Fig. 1. Inhibition of the binding of [3H]paroxetine (2 nM) to rat
lymphocyte membranes in the presence of unlabeled drug concen-
trations from 0.1 to 1 nM, as described in Materials and methods.
Mean of duplicated determinations from one representative experiment.
R2=0.9577.
N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–4034
Finally, lymphocytes were obtained and the intracellular
concentration of 5-HT and 5-HIAA was determined
according to the indicated procedure.
2.9. Analysis of data
The software Biodata Handling with microcomputer
was used to calculate the concentration to produce 50%
inhibition (IC50), Hill coefficient (nH), and pseudocoeffi-
cient (pseudo-nH) (Hill, 1910; Barlow, 1983). The
software PRISM version 2.0 (GraphPad Software, 2003)
was used for the saturation experiments and calculation of
Bmax. Kd was calculated by the antilog of the log
concentration of ligand giving 50% of occupation in the
Hill plot and by biometric experiments (Hill, 1910), and
also by kinetic experiments, using k1 and k2 values. The
data are expressed as the meanFstandard error of the
mean (S.E.M.) from different and independent experi-
ments. Analysis of variance (ANOVA) and Student’s t-test
were used for statistical comparisons. The probability of
the differences between means was considered to be
significant if pb0.05.
3. Results
3.1. Inhibition experiments
The inhibition of [3H]paroxetine binding by the 5-HTT
inhibitors is shown in Table 1 and Fig. 1. The potency order
of inhibition was imipramineNcitalopramNalaproclateN5-HT.
The pseudo nH was less than 1 for selective inhibitory drugs.
Desipramine, nomifensine, and GBR-12909 did not inhibit
[3H]paroxetine binding.
3.2. Saturation experiments
The values of Bmax and Kd for the binding of [3H]pa-
roxetine to membranes of peripheral blood lymphocytes of
Table 1
Inhibition of [3H]paroxetine binding to membranes of lymphocytes from
blood of rats
Drugs IC50 (nM) CCM Pseudo nH
Imipramine 2.16F0.04 97F0.3 �0.91F0.16
Citalopram 15.70F1.3 92F5.0 �0.29F0.10
Alaproclate 26.40F1.2 95F1.2 �0.26F0.09
5-HT 398.45F102 88F1.8 �0.19F0.03
Desimipramine N1000
GBR12090 N1000
Nomifensine N1000
The potency of inhibition was determined by incubating the membrane
preparation with 2 nM [3H]paroxetine in the presence of 15 concentrations
of the corresponding drug as described in Materials and methods. Mean-
FS.E.M., n=6. IC50 was calculated by interactive analysis INHIBITION
(Barlow, 1983). CMC is coefficient of multiple correlation, nH is pseudo-
Hill coefficient.
rats were 6156F1714 fmol/106 cells, and 1.77F0.30 nM,
respectively. The analysis of the saturation curves showed
that they did not correspond to a hyperbolic curve
according to the Michaelis–Menten equation, but were
sigmoid, and the Scatchard plot consisted of an inverted
concave curve (Fig. 2A,B). The values of nH were larger
than 1 and the analysis of Klotz showed that the work
was done under saturation conditions (data not shown).
Fig. 2. Saturation experiments for the binding of [3H]paroxetine to
lymphocyte membrane. (A) Sigmoid curve representative of saturation
binding, 100 Al of membrane preparation were incubated in the presence
of 0.2–2 nM concentration of the ligand. Specific binding was defined
with imipramine. Incubation was carried out at 25 8C for 90 min.
MeanFS.E.M. of duplicate determination from one representative
experiment. (B) Scatchard plot obtained by the Bound/Free vs. Bound
data transformation.
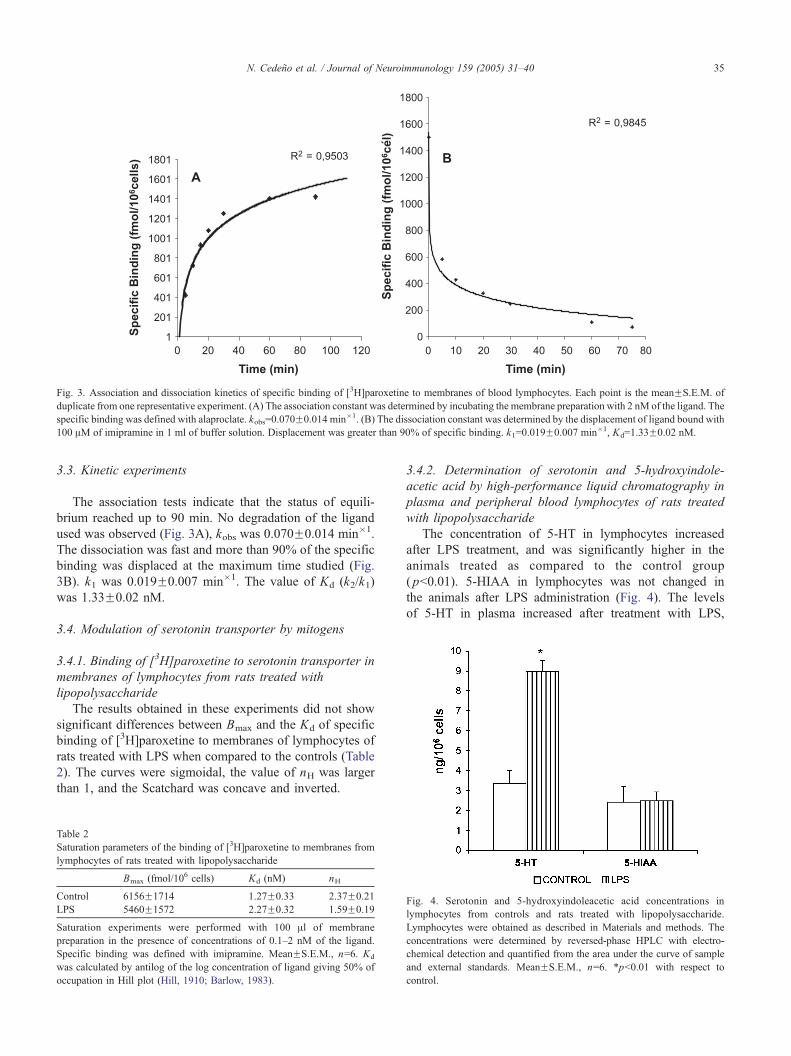
Fig. 3. Association and dissociation kinetics of specific binding of [3H]paroxetine to membranes of blood lymphocytes. Each point is the meanFS.E.M. of
duplicate from one representative experiment. (A) The association constant was determined by incubating the membrane preparation with 2 nM of the ligand. The
specific binding was defined with alaproclate. kobs=0.070F0.014 min�1. (B) The dissociation constant was determined by the displacement of ligand bound with
100 AM of imipramine in 1 ml of buffer solution. Displacement was greater than 90% of specific binding. k1=0.019F0.007 min�1, Kd=1.33F0.02 nM.
N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–40 35
3.3. Kinetic experiments
The association tests indicate that the status of equili-
brium reached up to 90 min. No degradation of the ligand
used was observed (Fig. 3A), kobs was 0.070F0.014 min�1.
The dissociation was fast and more than 90% of the specific
binding was displaced at the maximum time studied (Fig.
3B). k1 was 0.019F0.007 min�1. The value of Kd (k2/k1)
was 1.33F0.02 nM.
3.4. Modulation of serotonin transporter by mitogens
3.4.1. Binding of [3H]paroxetine to serotonin transporter in
membranes of lymphocytes from rats treated with
lipopolysaccharide
The results obtained in these experiments did not show
significant differences between Bmax and the Kd of specific
binding of [3H]paroxetine to membranes of lymphocytes of
rats treated with LPS when compared to the controls (Table
2). The curves were sigmoidal, the value of nH was larger
than 1, and the Scatchard was concave and inverted.
Table 2
Saturation parameters of the binding of [3H]paroxetine to membranes from
lymphocytes of rats treated with lipopolysaccharide
Bmax (fmol/106 cells) Kd (nM) nH
Control 6156F1714 1.27F0.33 2.37F0.21
LPS 5460F1572 2.27F0.32 1.59F0.19
Saturation experiments were performed with 100 Al of membrane
preparation in the presence of concentrations of 0.1–2 nM of the ligand.
Specific binding was defined with imipramine. MeanFS.E.M., n=6. Kd
was calculated by antilog of the log concentration of ligand giving 50% of
occupation in Hill plot (Hill, 1910; Barlow, 1983).
3.4.2. Determination of serotonin and 5-hydroxyindole-
acetic acid by high-performance liquid chromatography in
plasma and peripheral blood lymphocytes of rats treated
with lipopolysaccharide
The concentration of 5-HT in lymphocytes increased
after LPS treatment, and was significantly higher in the
animals treated as compared to the control group
( pb0.01). 5-HIAA in lymphocytes was not changed in
the animals after LPS administration (Fig. 4). The levels
of 5-HT in plasma increased after treatment with LPS,
Fig. 4. Serotonin and 5-hydroxyindoleacetic acid concentrations in
lymphocytes from controls and rats treated with lipopolysaccharide.
Lymphocytes were obtained as described in Materials and methods. The
concentrations were determined by reversed-phase HPLC with electro-
chemical detection and quantified from the area under the curve of sample
and external standards. MeanFS.E.M., n=6. *pb0.01 with respect to
control.
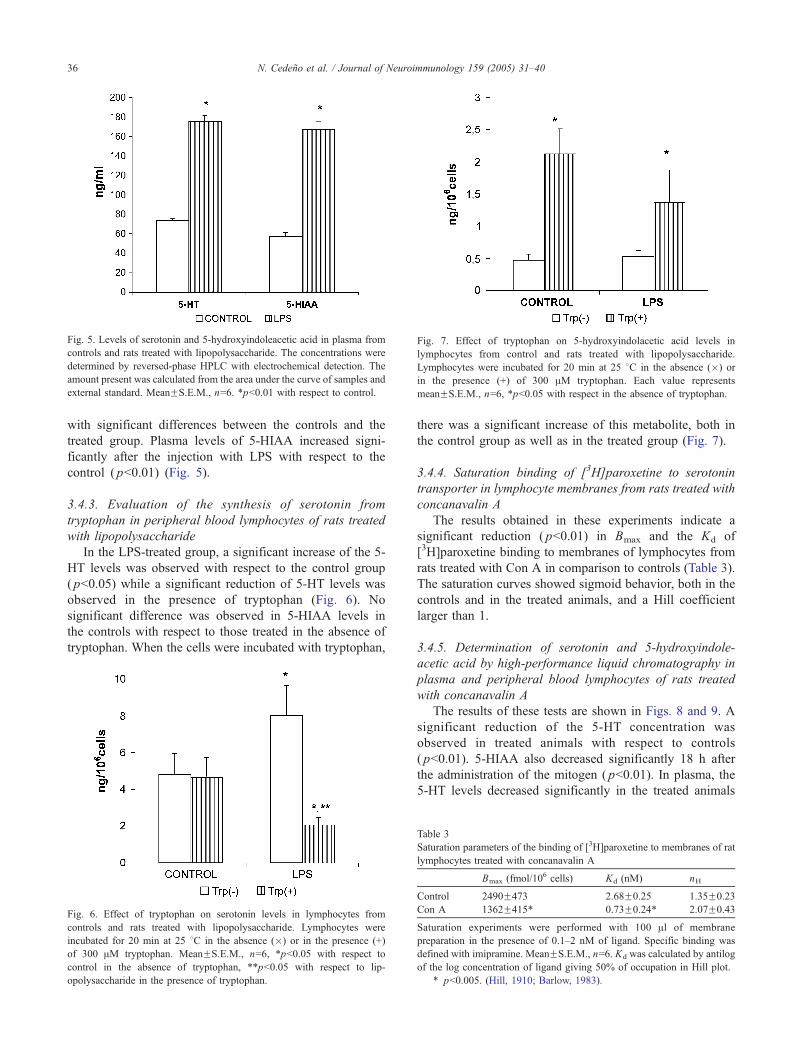
Fig. 7. Effect of tryptophan on 5-hydroxyindolacetic acid levels in
lymphocytes from control and rats treated with lipopolysaccharide.
Lymphocytes were incubated for 20 min at 25 8C in the absence (�) or
in the presence (+) of 300 AM tryptophan. Each value represents
meanFS.E.M., n=6, *pb0.05 with respect in the absence of tryptophan.
Fig. 5. Levels of serotonin and 5-hydroxyindoleacetic acid in plasma from
controls and rats treated with lipopolysaccharide. The concentrations were
determined by reversed-phase HPLC with electrochemical detection. The
amount present was calculated from the area under the curve of samples and
external standard. MeanFS.E.M., n=6. *pb0.01 with respect to control.
N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–4036
with significant differences between the controls and the
treated group. Plasma levels of 5-HIAA increased signi-
ficantly after the injection with LPS with respect to the
control ( pb0.01) (Fig. 5).
3.4.3. Evaluation of the synthesis of serotonin from
tryptophan in peripheral blood lymphocytes of rats treated
with lipopolysaccharide
In the LPS-treated group, a significant increase of the 5-
HT levels was observed with respect to the control group
( pb0.05) while a significant reduction of 5-HT levels was
observed in the presence of tryptophan (Fig. 6). No
significant difference was observed in 5-HIAA levels in
the controls with respect to those treated in the absence of
tryptophan. When the cells were incubated with tryptophan,
Fig. 6. Effect of tryptophan on serotonin levels in lymphocytes from
controls and rats treated with lipopolysaccharide. Lymphocytes were
incubated for 20 min at 25 8C in the absence (�) or in the presence (+)
of 300 AM tryptophan. MeanFS.E.M., n=6, *pb0.05 with respect to
control in the absence of tryptophan, **pb0.05 with respect to lip-
opolysaccharide in the presence of tryptophan.
there was a significant increase of this metabolite, both in
the control group as well as in the treated group (Fig. 7).
3.4.4. Saturation binding of [3H]paroxetine to serotonin
transporter in lymphocyte membranes from rats treated with
concanavalin A
The results obtained in these experiments indicate a
significant reduction ( pb0.01) in Bmax and the Kd of
[3H]paroxetine binding to membranes of lymphocytes from
rats treated with Con A in comparison to controls (Table 3).
The saturation curves showed sigmoid behavior, both in the
controls and in the treated animals, and a Hill coefficient
larger than 1.
3.4.5. Determination of serotonin and 5-hydroxyindole-
acetic acid by high-performance liquid chromatography in
plasma and peripheral blood lymphocytes of rats treated
with concanavalin A
The results of these tests are shown in Figs. 8 and 9. A
significant reduction of the 5-HT concentration was
observed in treated animals with respect to controls
( pb0.01). 5-HIAA also decreased significantly 18 h after
the administration of the mitogen ( pb0.01). In plasma, the
5-HT levels decreased significantly in the treated animals
Table 3
Saturation parameters of the binding of [3H]paroxetine to membranes of rat
lymphocytes treated with concanavalin A
Bmax (fmol/106 cells) Kd (nM) nH
Control 2490F473 2.68F0.25 1.35F0.23
Con A 1362F415* 0.73F0.24* 2.07F0.43
Saturation experiments were performed with 100 Al of membrane
preparation in the presence of 0.1–2 nM of ligand. Specific binding was
defined with imipramine. MeanFS.E.M., n=6. Kd was calculated by antilog
of the log concentration of ligand giving 50% of occupation in Hill plot.
* pb0.005. (Hill, 1910; Barlow, 1983).
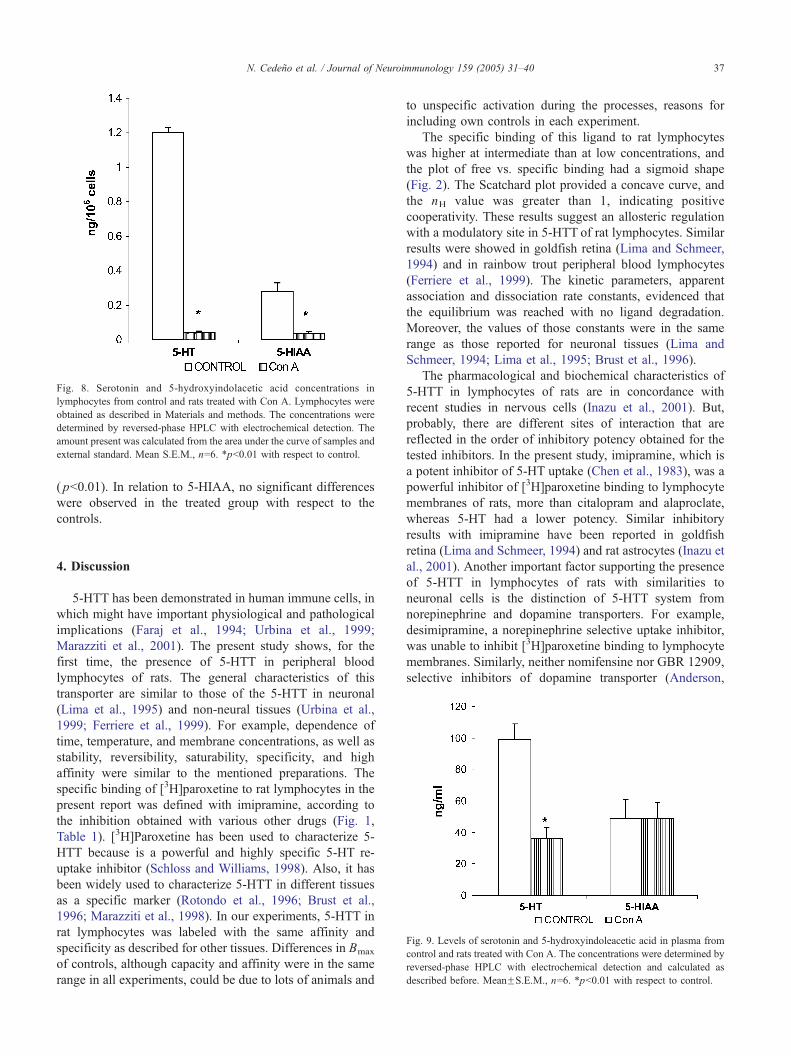
Fig. 8. Serotonin and 5-hydroxyindolacetic acid concentrations in
lymphocytes from control and rats treated with Con A. Lymphocytes were
obtained as described in Materials and methods. The concentrations were
determined by reversed-phase HPLC with electrochemical detection. The
amount present was calculated from the area under the curve of samples and
external standard. Mean S.E.M., n=6. *pb0.01 with respect to control.
N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–40 37
( pb0.01). In relation to 5-HIAA, no significant differences
were observed in the treated group with respect to the
controls.
Fig. 9. Levels of serotonin and 5-hydroxyindoleacetic acid in plasma from
control and rats treated with Con A. The concentrations were determined by
reversed-phase HPLC with electrochemical detection and calculated as
described before. MeanFS.E.M., n=6. *pb0.01 with respect to control.
4. Discussion
5-HTT has been demonstrated in human immune cells, in
which might have important physiological and pathological
implications (Faraj et al., 1994; Urbina et al., 1999;
Marazziti et al., 2001). The present study shows, for the
first time, the presence of 5-HTT in peripheral blood
lymphocytes of rats. The general characteristics of this
transporter are similar to those of the 5-HTT in neuronal
(Lima et al., 1995) and non-neural tissues (Urbina et al.,
1999; Ferriere et al., 1999). For example, dependence of
time, temperature, and membrane concentrations, as well as
stability, reversibility, saturability, specificity, and high
affinity were similar to the mentioned preparations. The
specific binding of [3H]paroxetine to rat lymphocytes in the
present report was defined with imipramine, according to
the inhibition obtained with various other drugs (Fig. 1,
Table 1). [3H]Paroxetine has been used to characterize 5-
HTT because is a powerful and highly specific 5-HT re-
uptake inhibitor (Schloss and Williams, 1998). Also, it has
been widely used to characterize 5-HTT in different tissues
as a specific marker (Rotondo et al., 1996; Brust et al.,
1996; Marazziti et al., 1998). In our experiments, 5-HTT in
rat lymphocytes was labeled with the same affinity and
specificity as described for other tissues. Differences in Bmax
of controls, although capacity and affinity were in the same
range in all experiments, could be due to lots of animals and
to unspecific activation during the processes, reasons for
including own controls in each experiment.
The specific binding of this ligand to rat lymphocytes
was higher at intermediate than at low concentrations, and
the plot of free vs. specific binding had a sigmoid shape
(Fig. 2). The Scatchard plot provided a concave curve, and
the nH value was greater than 1, indicating positive
cooperativity. These results suggest an allosteric regulation
with a modulatory site in 5-HTT of rat lymphocytes. Similar
results were showed in goldfish retina (Lima and Schmeer,
1994) and in rainbow trout peripheral blood lymphocytes
(Ferriere et al., 1999). The kinetic parameters, apparent
association and dissociation rate constants, evidenced that
the equilibrium was reached with no ligand degradation.
Moreover, the values of those constants were in the same
range as those reported for neuronal tissues (Lima and
Schmeer, 1994; Lima et al., 1995; Brust et al., 1996).
The pharmacological and biochemical characteristics of
5-HTT in lymphocytes of rats are in concordance with
recent studies in nervous cells (Inazu et al., 2001). But,
probably, there are different sites of interaction that are
reflected in the order of inhibitory potency obtained for the
tested inhibitors. In the present study, imipramine, which is
a potent inhibitor of 5-HT uptake (Chen et al., 1983), was a
powerful inhibitor of [3H]paroxetine binding to lymphocyte
membranes of rats, more than citalopram and alaproclate,
whereas 5-HT had a lower potency. Similar inhibitory
results with imipramine have been reported in goldfish
retina (Lima and Schmeer, 1994) and rat astrocytes (Inazu et
al., 2001). Another important factor supporting the presence
of 5-HTT in lymphocytes of rats with similarities to
neuronal cells is the distinction of 5-HTT system from
norepinephrine and dopamine transporters. For example,
desimipramine, a norepinephrine selective uptake inhibitor,
was unable to inhibit [3H]paroxetine binding to lymphocyte
membranes. Similarly, neither nomifensine nor GBR 12909,
selective inhibitors of dopamine transporter (Anderson,

N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–4038
1989; Tella and Goldberg, 2001; Moron et al., 2002),
blocked the binding of the radioligand.
5-HTT in lymphocytes of rat might be under same
genetic control as in other tissues. For instance, in human
brain and platelets, 5-HTT is codified by the same gene,
human chromosome 17 (Blakely et al., 1991; Lesch et al.,
1994, 1996), which was also cloned and evaluated in human
placenta (Ramamoorthy et al., 1993).
LPS (Hancock, 1991) induces a variety of inflammatory
and toxic effects (Mathew, 1998). When evaluating the
capacity of 5-HTT in lymphocytes from blood of rats after
treatment with LPS, no changes were observed in the
number of sites or the affinity of the ligand for 5-HTT
(Table 1), even though changes of the serotonergic system in
the brain by peripheral administration of LPS have been
shown (Linthorst and Reul, 1998).
Treatment with LPS produced a significant increase of
5-HT levels in lymphocytes in peripheral blood of rats
(Fig. 4). It might be that high concentrations of 5-HT in
lymphocytes could mediate some of the effects in
response to the administered dose of LPS. In immune
system cells, the immunoregulatory effect of 5-HT seems
to be partially mediated through 5-HT1A receptors
(Hellstrand and Hermmodsson, 1990; Aune et al., 1993;
Sempere et al., 2003; Eugen-Olsen et al., 1997). In rats,
5-HT and the 5HT1A agonist, 8-hydroxy-2(di-n-propilami-
no)tetralin, produce an increase in the proliferation of
mitogen-stimulated B cells (Iken et al., 1995). These
mechanisms, among others, could be taking place in
lymphocytes of LPS-treated rats.
The high 5-HT concentration observed in rat lympho-
cytes may be due to the activation of its synthesis through
stimulation with LPS, like that exercised by INF-g in
human mononuclear cells (Arzt et al., 1988). This
reaffirms the property of immune system cells of
producing not only cytokines, but also neuropeptides,
hormones, and neurotransmitters (Blalock et al., 1992;
Blalock, 1994). When evaluating the synthesis of this
indolamine, a significant increase in 5-HT levels was
observed in lymphocytes of rats that were treated with
LPS. This might be influenced by an increase in the
synthesis of 5-HT, or by affection of 5-HT recapturing
mechanisms, even though the number of carriers was not
significantly changed.
When adding tryptophan to the medium, a lowering of
the 5-HT levels was observed in the lymphocytes of rats
treated with LPS. Possibly this decrease in the intra-
cellular concentration of 5-HT is the result of a greater
release of 5-HT because of the effect of LPS and the use
of tryptophan available in the medium for 5-HT synthesis.
Probably, once the synthesis of 5-HT is activated and
released to the extracellular medium, it will favor either
clonal expansion (Finocchiario et al., 1988; Jackson et al.,
1985; Kut et al., 1992) or the presence of other immune
cell lines that may mediate a greater release of 5-HT
(Matsuda et al., 1997). 5-HIAA levels in lymphocytes
were not changed after the injection of LPS, and this
supports the idea of an increase in the origin of 5-HT, as
well as a possible reduction of catabolism. The concen-
trations of 5-HIAA increased significantly both in plasma
and lymphocytes in the presence of tryptophan after
treatment with LPS, although the intracellular 5-HIAA
levels did not change after administering LPS in the
absence of tryptophan. These results reflect a higher
metabolic replacement of 5-HT or increased turnover rate,
produced by LPS, evidenced when supplying the substrate
to the cells.
The general effects of LPS on T cells does not appear
to be direct, because it is not thymus-dependent, while
the proliferative response of this cell stock increases after
stimulation with Con A (Chen et al., 1995). Increased
expression of the 5-HT1A receptor has been shown in T
cells activated with PHA, which could be related to the
lymphoproliferative effect of 5-HT (Aune et al., 1993;
Iken et al., 1995). In our results, the number of binding
sites showed a significant reduction; however, the affinity
was increased by Con A, which might suggest the
presence of an autocrine mechanism through which
lymphocytes regulate their own activity. This autocrine
regulation mechanism by lectins has been described for
different subpopulations of human immune cells (Beuth
et al., 1992). In addition to this mechanism, possibly the
populations of lymphocytes modulated by the various
treatments are not the same, and the obtained results
correspond to the conjoint interaction of many factors.
Moreover, the administration of Con A significantly
decreased plasma levels of 5-HT, as well as the
intracellular concentrations of 5-HT and 5-HIAA in
lymphocytes. These results might be favoring lymphocyte
proliferation, the induction of the expression of surface
receptors that are closely related to the activation of
immune system cells, such as IL-2 and 5-HT1A receptors
(Waldmann, 1986; Oshima et al., 1991; Aune et al.,
1993; Sempere et al., 2003). Both receptors would play
a critical role in the evolution of specialized T cell
populations, necessary to establish the normal response
of those cells. If the administration of Con A favors
cellular proliferation per se, lymphocytes would regulate
their activity via an autocrine way involving 5-HT, in
which the expression of the number of 5-HTT could
play a modulatory role.
The observations reported in the literature when
combined with the results obtained in this work
support the idea of a strong immunomodulatory role
of 5-HT through producing certain specific actions
that promote and regulate immune responses.
Acknowledgements
This work was supported by Grant S1-3490 from
Fondo Nacional de Investigacion, Tecnologıa e Innova-

N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–40 39
cion (FONACIT). We appreciate the secretarial assistance
of Mrs. Isabel Otaegui. Nemesio Cedeno was a Graduate
Student of Centro de Estudios Avanzados (CEA) from
Instituto Venezolano de Investigaciones Cientıficas
(IVIC).
References
Abdouh, M., Storring, J.M., Riad, M., Paquette, Y., Albert, P.R., Drobetsky,
E., Kouassi, E., 2001. Transcriptional mechanisms for induction of 5-
HT1A receptor mRNA and protein in activated B and T lymphocytes.
J. Biol. Chem. 276, 4382–4388.
Anderson, P.H., 1989. The dopamine uptake inhibitor GBR 12909:
selective and molecular mechanism of action. Eur. J. Pharmacol. 166,
493–504.
Arzt, E.S., Fernandez-Castelo, S., Finocchiaro, L.M., Criscuolo, M.E.,
Dıaz, A., Askenase, P.W., 1988. Effector and regulatory mechanism in
delayed-type hypersensitivity. In: Middleton Jr., E., Reed, C.E., Ellis,
E.F., Atkinson, N.F. (Eds.), En Allergy: Principles and Practice (2nd
ed.). C.V. Mosby, St. Lluis.
Askenase, P.W., 1988. Effector and regulatory mechanism in delayed-type
hypersensitivity. In: Middleton Jr., E., Reed, C.E., Ellis, E.F., Atkinson,
N.F. (Eds.), En Allergy: Principles and Practice (2nd ed.). C.V. Mosby,
St. Lluis.
Aune, T.M., McGrath, K.M., Sarr, T., Bombara, M.P., Kelley, K.A.,
1993. Expression of 5-HT1A receptors on activated human T cells.
Regulation of cAMP levels and T cell proliferation by 5-hydroxy-
tryptamine. J. Immunol. 151, 1175–1183.
Barlow, R.W., 1983. Biodata Handling with Microcomputers. Elsevier
Science Publishers, Amsterdam, pp. 125–143.
Betten, A., Dahigren, C., Hermodsson, S., Hellstrand, K., 2001. Serotonin
protects NK cells against oxidatively induced functional inhibition and
apoptosis. J. Leukoc. Biol. 70, 65–72.
Beuth, J., Ko, H.L., Gabius, H.J., Burrichter, H., Oette, K., Pulverer, G.,
1992. Behavior of lymphocyte subsets and expression of activation
markers in response to immunotherapy with galactoside-specific lectin
from mistletoe in breast cancer patients. Clin. Investig. 70, 658–661.
Blakely, R., Berson, H.E., Fremeau, R.T., Caron, M.G., Peek, M.M., Prince,
H.K., Bradley, C.C., 1991. Cloning and expression of a functional
serotonin transporter from rat brain. Nature 354, 66–70.
Blakely, R.D., Wise, K.R., Schroeter, S., 1995. Localization of the
serotonin transporter to rodent adrenal chromaffin cells: support for a
role of 5-HT in adrenal physiology. Abstr.-Soc. Neurosci. 21, 316.1.
Blalock, J.E., 1994. The syntax of immune-neuroendocrine communication.
Immunol. Today 15, 504–511.
Blalock, J.E., Eds, K., Ishizaka, Lachman, P.J., Waskman, B.H., 1992.
Production of peptide hormones and neurotransmitters by the immune
system. Chem. Immunol. 52, 1–24.
Bonnet, M., Lespinats, G., Burtin, C., 1984. Histamine and serotonin
suppression on lymphocyte response phytohemagglutinin and alloge-
neic cells. Cell. Immunol. 83, 280–291.
Brust, P., Bergmann, R., Johannsen, B., 1996. High-affinity binding of
[3H]paroxetine to caudate nucleus and microvessels from porcine brain.
NeuroReport 31, 1405–1408.
Bylun, D.V., Yamamura, H.I., 1990. Methods in Neurotransmitter Receptor
Analysis. Reaven Press, New York.
Chen, S.S., Tung, J.S., Good, R.A., Hadden, J.W., 1983. Changes in surface
antigens of immature thymocytes under the influence of T cell growth
and thymic factors. Proc. Natl. Acad. Sci. 80, 5980–5985.
Chen, C., David, A., Nunnerley, H., Michel, M., Dawson, L., Berry, H.,
Dobbs, J., Fahy, T., 1995. Adverse life events and breast cancer: case-
control study. Med. J. 311, 1527–1530.
Eugen-Olsen, J., Afzelius, P., Andersen, L., Iversen, J., Kronborg, G.A.,
Aebech, P., Nielsen, J.O., Hofmann, B., 1997. Serotonin modulates
immune function in T cells from HIV-seropositive subjects. Clin.
Immunol. Immunopharmacol. 84, 115–121.
Faraj, B.A., Olkowski, Z.L., Jackson, R.T., 1994. Expression of high
affinity serotonin transporter in human lymphocytes. Int. J. Immuno-
pharmacol. 16, 561–567.
Faraj, B.A., Olkowski, Z.L., Jackson, R.T., 1997. Prevalence of high
serotonin uptake in lymphocytes of abstinent alcoholics. Biochem.
Pharmacol. 53, 53–57.
Ferriere, F., Khan, N.A., Troutaud, D., Deschaux, P., 1996. Serotonin
modulation of lymphocyte proliferation via 5-HT1A receptors in
rainbow trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 20,
273–283.
Ferriere, F., Khan, N.A., Meyniel, J.P., Deschaux, P., 1999. 5-Hydroxytrypt-
amine-induce calcium-channel gating in rainbow trout (Oncorhynchus
mykiss) peripheral blood lymphocytes. Biochem. J. 323, 251–258.
Finocchiario, L.M., Artz, S., Fernandez, M., Criscuolo, A., Dıaz, S.,
Finkielman, S., Nahrod, V.E., 1988. Serotonin and melatonin synthesis
in peripheral blood mononuclear cells stimulated by interferon-gamma
as part of immunomodulatory pathway. J. Interf. Res. 8, 705–711.
Haas, H.S., Schauenstein, K., 1997. Neuroimmunomodulation via limbic
structures—The neuroanatomy of psychoimmunology. Prog. Neurobiol.
51, 195–222.
Hancock, R.E.W., 1991. Bacterial outer membranes: evolving concepts.
Specific structures provide gram-negative bacteria with several unique
advantages. ASM News 57, 175.
Hellstrand, K., Hermmodsson, S., 1990. Enhancement of human natural
killer cell cytotoxicity by serotonin: role of non-T/CD16+ NK cells,
accessory monocytes, and 5HT1A receptors. Cell. Immunol. 127,
199–214.
Hill, A.V., 1910. The possible effects of the aggregation of molecules of
hemoglobin on its dissociation curves. J. Physiol. 1, iv–vii.
Iken, K., Chheng, S., Fargin, A., Goulet, A., Kouassi, E., 1995. Serotonin
upregulates mitogen-stimulated B lymphocyte proliferation through 5-
HT1A receptors. Cell. Immunol. 163, 1–9.
Inazu, M., Takeda, H., Ikoshi, H., Sugisawa, M., Uchida, Y., Matsumiya, T.,
2001. Pharmacological characterization and visualization of the glial
serotonin transporter. Neurochem. Int. 39, 39–49.
Jackson, J.C., Cross, R.J., Walker, R.F., Markesbery, W.R., Brooks, W.H.,
Roszman, T.L., 1985. Influence of serotonin on the immune response.
Immunology 54, 503–511.
Kut, J.P., Young, R.M., Crayton, J.W., Wright, M.A., Young, M.E.,
1992. Regulation of murine T-lymphocyte function by spleen cell-
derived and exogenous serotonin. Immunopharmacol. Immunotoxicol.
14, 783–796.
Laberge, S., Cruikshank, W.W., Beer, D.J., Center, D.M., 1996. Secretion
of IL-16 (lymphocyte chemottractant factor) from serotonin-stimulated
CD8+ T cells in vitro. J. Immunol. 156, 310–315.
Lesch, K.-P., Balling, U., Gross, J., Strauss, K., Wolozin, B.L., Murphy,
D.L., Riederer, P., 1994. Organization of the human serotonin
transporter gene. J. Neural Transm. 95, 157–162.
Lesch, K.P., Bengel, D., Heils, A., Sabol, S.Z., Greenberg, B.D.,
Petri, S., Benjamin, J., Muller, C.R., Hamer, D.H., Murphy, D.L.,
1996. Association of anxiety-related traits with a polymorphism in
the serotonin transporter gene regulatory region. Science 274,
1527–2530.
Lima, L., Schmeer, C., 1994. Characterization of serotonin transporter in
goldfish retina by binding of [3H]paroxetine in the uptake of
[3H]serotonin: modulation by light. J. Neurochem. 62, 528–535.
Lima, L., Urbina, M., 2002. Serotonin transporter modulation in blood
lymphocytes from patients with major depression. Cell. Mol. Neurobiol.
22, 797–804.
Lima, L., Trejo, E., Urbina, M., 1995. Serotonin turnover rate, [3H]Parox-
etine binding sites, and 5-HT1A receptors in the hippocampus of rats
subchronically treated with clonazepam. Neuropharmacology 34,
1327–1333.
Linthorst, A., Reul, J.M.H., 1998. Brain neurotransmission during
peripheral inflammation. Ann. N.Y. Acad. Sci. 840, 139–152.

N. Cedeno et al. / Journal of Neuroimmunology 159 (2005) 31–4040
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randal, R.J., 1951. Protein
measurement with folin phenol reagent. J. Biol. Chem. 193, 265–275.
Marazziti, D., Rossi, A., Giannaccini, G., Baroni, S., Lucacchini, A.,
Cassano, G., 1998. Presence and characterization of the serotonin
transporter in human resting lymphocytes. Neuropsychopharmacology
19, 154–159.
Marazziti, D., Ori, M., Nardini, M., Rossi, A., Nardi, I., Cassano, G.B.,
2001. mRNA expression of serotonin receptors of type 2C and 5A in
human resting lymphocytes. Neuropsychobiology 43, 123–126.
Mathew, P., 1998. Specificity and function of lipopolysaccaride antibodies.
In: Szentivanyi, A., Friedman, H.Y., Nowotny, A. (Eds.), En Immuno-
pharmacol. and Pathophysiol. of Bacterial Endotoxins, vol. II. Plenum
Press, New York, pp. 347–374.
Matsuda, H., Ushio, H., Geba, G.P., Askenase, P., 1997. Human platelets
can initiate T cell-dependent contact sensitivity through local serotonin
release mediated by IgE antibodies. J. Immunol. 158, 2891–2897.
Moron, J.A., Brockington, A., Wise, R.A., Rocha, B.A., Hope, B.T., 2002.
Dopamine uptake through the norepinephrine transporter in brain
regions with low levels of the dopamine transporter: evidence from
knock-out mouse lines. J. Neurosci. 22, 389–395.
Mossner, R., Daniel, S., Schmitt, A., Albert, D., Lesch, K.P., 2001.
Modulation of serotonin transporter function by interleukin-4. Life Sci.
68, 873–880.
Oshima, Y., Ko, H.L., Beuth, J., Burrichter, H., Oette, K., Pulverer, G.,
1991. Activation of mononuclear immune cells in response to
staphylococcal lipoteichoic acid. Zentralbl. Bakteriol. 275, 374–381.
Pellegrino, T.C., Bayer, B.M., 2000. Specific serotonin reuptake inhibitor-
induced decreases in lymphocyte activity require endogenous serotonin
release. Neuroimmunomodulation 8, 179–187.
Ramamoorthy, J.D., Leibach, F.H., Maeshh, V.B., Ganapathy, V., 1993.
Partial purification and characterization of the human placental
serotonin transporter. Placenta 14, 449–461.
Rotondo, A., Giannaccini, G., Betti, L., Chiellini, G., Marazziti, D., Martin,
C., Lucacchini, A., Cassano, G.B., 1996. The serotonin transporter from
human brain: purification and partial characterization. Neurochem. Int.
28, 299–307.
Rudnick, G., 1997. Active transporter of 5-hydroxytryptamine by plasma
membrane vesicles isolated from human blood platelets. J. Biol. Chem.
252, 2170–2174.
Schloss, P., Williams, D.C., 1998. The serotonin transporter: a primary
target for antidepressant drugs. J. Psychopharmacol. 12, 115–121.
Sempere, T., Cedeno, N., Urbina, M., Lima, L., 2003. 8-[3H]-hydroxy-2-
(di-n-propylamino)tetralin binding sites in blood lymphocytes of rats
and the modulation by mitogens and immobilization. J. Neuroimmunol.
138, 8–16.
Slauson, D.O., Walker, C., Kristensen, F., Wang, Y., De Weck, A.L., 1984.
Mechanisms of serotonin-induced lymphocyte proliferation inhibition.
Cell. Immunol. 84, 240–252.
Stefulj, J., Cicin-Sain, L., Schauenstein, K., Jernej, B., 2001. Serotonin and
immune response: effect of the amine on in vitro proliferation of rat
lymphocytes. Neuroimmunomodulation 9, 103–108.
Sternberg, E.M., Trial, J., Parker, C.W., 1986. Effect of serotonin on
murine macrophages: suppression of Ia expression by serotonin and its
reversal by 5-HT2 serotononergic receptor antagonists. J. Immunol.
137, 276–282.
Tella, S.R., Goldberg, S.R., 2001. Subtle differences in the discriminative
stimulus effects of cocaine and GBR-12909. Prog. Neuro-psychophar-
macol. Biol. Psychiatry 25, 639–656.
Urbina, M., Pineda, S., Pinango, L., Carreira, I., Lima, L., 1999.
[3H]Paroxetine binding to human peripheral lymphocyte membranes
of patients with major depression before and after treatment with
fluoxetine. Int. J. Immunopharmacol. 21, 631–646.
Wade, P.R., Chen, J., Jaffe, B., Kassem, I.S., Blakely, R.D., Gershon, M.D.,
1996. Localization and function of a 5-HT transporter in crypt epithelia
of the gastrointestinal tract. J. Neurosci. 16, 2352–2364.
Waldmann, T.M., 1986. The structure, function and expression of
interleukin-2 receptor on normal and malignant lymphocytes. Science
232, 727–732.