
Characterization of rubber tree microRNA in phytohormone response using large genomic DNA libraries,...
13
1 3 Mol Genet Genomics (2014) 289:921–933 DOI 10.1007/s00438-014-0862-0 ORIGINAL PAPER Characterization of rubber tree microRNA in phytohormone response using large genomic DNA libraries, promoter sequence and gene expression analysis Supanath Kanjanawattanawong · Sithichoke Tangphatsornruang · Kanokporn Triwitayakorn · Panthita Ruang‑areerate · Duangjai Sangsrakru · Supannee Poopear · Suthasinee Somyong · Jarunya Narangajavana Received: 6 March 2014 / Accepted: 5 May 2014 / Published online: 26 May 2014 © Springer-Verlag Berlin Heidelberg 2014 provided approximately 2.42 haploid genome equivalents of coverage based on the estimated 2.15 gb rubber tree genome. Based on these library sequences, the precursors of 1 member of rubber tree-specific miRNAs and 12 mem- bers of conserved miRNAs were successfully identified. A panel of miRNAs was characterized for phytohormone response by precisely identifying phytohormone-respon- sive motifs in their promoter sequences. Furthermore, the quantitative real-time PCR on ethylene stimulation of rub- ber trees was performed to demonstrate that the miR2118, miR159, miR164 and miR166 are responsive to ethylene, thus confirmed the prediction by genomic DNA analysis. The cis-regulatory elements identified in the promoter regions of these miRNA genes help augment our under- standing of miRNA gene regulation and provide a founda- tion for further investigation of the regulation of rubber tree miRNAs. Keywords BAC library · Ethylene · Expression · Fosmid · miRNA · Rubber tree Introduction The para rubber tree (Hevea brasiliensis Muell. Arg.) is the tree crop most commonly cultivated worldwide for the production of natural rubber (NR) latex, of which 93 % is produced in Asia (Rahman et al. 2013; Gébelin et al. 2012). As production of NR is important for world industriali- zation, demand has been rising annually and is projected to increase by 25 % in the next decade (ANPRC 2010). Therefore, increasing NR output to match world demand is currently a crucial challenge for producers. NR latex pro- duction level depends on the genetic make-up of cultivated clones, environmental factors and harvesting conditions Abstract The para rubber tree is the most widely culti- vated tree species for producing natural rubber (NR) latex. Unfortunately, rubber tree characteristics such as a long life cycle, heterozygous genetic backgrounds, and poorly understood genetic profiles are the obstacles to breeding new rubber tree varieties, such as those with improved NR yields. Recent evidence has revealed the potential impor- tance of controlling microRNA (miRNA) decay in some aspects of NR regulation. To gain a better understanding of miRNAs and their relationship with rubber tree gene regulation networks, large genomic DNA insert-containing libraries were generated to complement the incomplete draft genome sequence and applied as a new powerful tool to predict a function of interested genes. Bacterial artifi- cial chromosome and fosmid libraries, containing a total of 120,576 clones with an average insert size of 43.35 kb, Communicated by S. Hohmann. Electronic supplementary material The online version of this article (doi:10.1007/s00438-014-0862-0) contains supplementary material, which is available to authorized users. S. Kanjanawattanawong · J. Narangajavana (*) Department of Biotechnology, Faculty of Science, Mahidol University, Rama 6 Rd, Rajthewee, Bangkok 10400, Thailand e-mail: [email protected] S. Tangphatsornruang · P. Ruang-areerate · D. Sangsrakru · S. Poopear · S. Somyong (*) National Center for Genetic Engineering and Biotechnology, 113 Thailand Science Park, Phahonyothin Rd, Klong 1, Klong Luang, Pathum Thani 12120, Thailand e-mail: [email protected] K. Triwitayakorn Institute of Molecular Bioscience, Mahidol University, Nakhon Pathom 73170, Thailand
Transcript of Characterization of rubber tree microRNA in phytohormone response using large genomic DNA libraries,...

1 3
Mol Genet Genomics (2014) 289:921–933DOI 10.1007/s00438-014-0862-0
ORIGINAL PAPER
Characterization of rubber tree microRNA in phytohormone response using large genomic DNA libraries, promoter sequence and gene expression analysis
Supanath Kanjanawattanawong · Sithichoke Tangphatsornruang · Kanokporn Triwitayakorn · Panthita Ruang‑areerate · Duangjai Sangsrakru · Supannee Poopear · Suthasinee Somyong · Jarunya Narangajavana
Received: 6 March 2014 / Accepted: 5 May 2014 / Published online: 26 May 2014 © Springer-Verlag Berlin Heidelberg 2014
provided approximately 2.42 haploid genome equivalents of coverage based on the estimated 2.15 gb rubber tree genome. Based on these library sequences, the precursors of 1 member of rubber tree-specific miRNAs and 12 mem-bers of conserved miRNAs were successfully identified. A panel of miRNAs was characterized for phytohormone response by precisely identifying phytohormone-respon-sive motifs in their promoter sequences. Furthermore, the quantitative real-time PCR on ethylene stimulation of rub-ber trees was performed to demonstrate that the miR2118, miR159, miR164 and miR166 are responsive to ethylene, thus confirmed the prediction by genomic DNA analysis. The cis-regulatory elements identified in the promoter regions of these miRNA genes help augment our under-standing of miRNA gene regulation and provide a founda-tion for further investigation of the regulation of rubber tree miRNAs.
Keywords BAC library · Ethylene · Expression · Fosmid · miRNA · Rubber tree
Introduction
The para rubber tree (Hevea brasiliensis Muell. Arg.) is the tree crop most commonly cultivated worldwide for the production of natural rubber (NR) latex, of which 93 % is produced in Asia (Rahman et al. 2013; Gébelin et al. 2012). As production of NR is important for world industriali-zation, demand has been rising annually and is projected to increase by 25 % in the next decade (ANPRC 2010). Therefore, increasing NR output to match world demand is currently a crucial challenge for producers. NR latex pro-duction level depends on the genetic make-up of cultivated clones, environmental factors and harvesting conditions
Abstract The para rubber tree is the most widely culti-vated tree species for producing natural rubber (NR) latex. Unfortunately, rubber tree characteristics such as a long life cycle, heterozygous genetic backgrounds, and poorly understood genetic profiles are the obstacles to breeding new rubber tree varieties, such as those with improved NR yields. Recent evidence has revealed the potential impor-tance of controlling microRNA (miRNA) decay in some aspects of NR regulation. To gain a better understanding of miRNAs and their relationship with rubber tree gene regulation networks, large genomic DNA insert-containing libraries were generated to complement the incomplete draft genome sequence and applied as a new powerful tool to predict a function of interested genes. Bacterial artifi-cial chromosome and fosmid libraries, containing a total of 120,576 clones with an average insert size of 43.35 kb,
Communicated by S. Hohmann.
Electronic supplementary material The online version of this article (doi:10.1007/s00438-014-0862-0) contains supplementary material, which is available to authorized users.
S. Kanjanawattanawong · J. Narangajavana (*) Department of Biotechnology, Faculty of Science, Mahidol University, Rama 6 Rd, Rajthewee, Bangkok 10400, Thailande-mail: [email protected]
S. Tangphatsornruang · P. Ruang-areerate · D. Sangsrakru · S. Poopear · S. Somyong (*) National Center for Genetic Engineering and Biotechnology, 113 Thailand Science Park, Phahonyothin Rd, Klong 1, Klong Luang, Pathum Thani 12120, Thailande-mail: [email protected]
K. Triwitayakorn Institute of Molecular Bioscience, Mahidol University, Nakhon Pathom 73170, Thailand

922 Mol Genet Genomics (2014) 289:921–933
1 3
(Duan et al. 2013; Vinod et al. 2010). Unfortunately, rubber tree characteristics such as a long life cycle, heterozygous genetic backgrounds, and poorly understood genetic pro-files are obstacles to breeding new rubber tree varieties.
One possible strategy for maximizing NR production is, therefore, through improved understanding and subsequent manipulation of gene controlling NR biosynthesis and pos-sible regulators such as microRNAs (miRNAs). miRNAs are small, non-coding RNA molecules (around 22 nucleo-tides in length), which have been proven to involve in many functional processes in plants such as leaf, shoot, root and floral development, as well as biotic and abiotic stress responses, by targeting mRNAs for cleavage or transla-tional repression (Gray 2004). The central role of miRNAs in the control of key agronomic traits makes them appeal-ing biotechnological targets for engineering plant varieties with improved output. Recently, novel and conserved miR-NAs, and their target genes have been identified in rubber trees by Lertpanyasampatha et al. (2012, 2013) and Gébe-lin et al. (2012, 2013a, b). Of these, some miRNA targets involving in NR biosynthesis including mevalonate kinase (miR159, miR319), Phosphomevalonate kinase (miR172, miR894, miRn15) and Rubber elongation factor (miR159, miR535) were predicted (Gébelin et al. 2012, 2013a). In addition, some miRNA targets involving in phytohormones response such as ARF (miR160), abscisic acid (ABA) insensitive protein (miR166) and gibberellin 20-oxidase (miR408) were also reported (Gébelin et al. 2013a, b). Information on these miRNAs together with their pro-moters should be investigated to pave the way for further genetic manipulation. So far, miRNA promoter analyses have been reported for important economically crops such as rice (Cui et al. 2009), the model plant Arabidopsis (Zhao and Li 2013; Zhao et al. 2013), and humans (Vrba et al. 2013; Ozsolak et al. 2008).
Integration analysis of miRNA promoter and plant hor-mones (or phytohormones) treatment has been recently investigated in only a few model plants (Chen et al. 2012; Zhao et al. 2012; Liu et al. 2009) with the aim to manipu-late hormone-responsive miRNA-mediated gene expression regulation in the future. These phytohormones regulate cel-lular processes such as gene expression, cellular division and growth, and are generally categorized into five classes: auxins, gibberellins (GAs), cytokinins, ABA and ethyl-ene, although a variety of additional compounds such as jasmonic acid (JA) and brassinosteriods (BRs) have been found to play roles as phytohormones as well (Liu et al. 2009; Gray 2004).
The identification of miRNA sequences and analysis of their promoters rely on access to genomic information. Recently, Rahman et al. (2013) reported a rubber tree draft genome sequence spanning approximately 1.1 gb of the estimated 2.15 gb haploid genome. Due to the current lack
of complete genomic information, large insert fragment libraries can provide a powerful supplement and tool for advancing genomic research in several areas, such as physi-cal mapping, positional cloning and genome sequencing.
There are two most efficiency methods of choice for the generation of these large insert fragment libraries. The first method is the bacterial artificial chromosome (BAC) cloning system, which has become dominant over others due to its ease for manipulation and insert copy stability (Gonthier et al. 2010). BAC libraries have been invaluable tools for the genome-wide genetic dissection of complex organisms for diverse applications, such as instance map-based cloning in barley (Schulte et al. 2011) and coffee (Cacao et al. 2013), physical mapping in the common bean (Liu et al. 2010), comparative genomics in wheat (Sehgal et al. 2012), and elucidation of adaptive species divergence among lake whitefish (Jeukens et al. 2011). The second method is the fosmid system which, despite the smaller insert size, is easier to handle than BAC and introduces no potential insert bias due to the use of end-repaired sheared DNA rather than DNA digested with a specific restriction endonuclease (Zhang et al. 2007; Kim et al. 2003). The smaller insert sizes have not been limita-tions in many studies, as a region of interest is generally not so large that one fosmid clone can cover it (Kim et al. 2003). Fosmid libraries have contributed to structural and functional genomics studies such as viral meta-genome sequencing (Solonenko et al. 2013), moth EST sequenc-ing and comparative mapping (Kamimura et al. 2012), and sheep rumen xylanase gene screening (Wang et al. 2012). The generation of rubber tree BAC and fosmid libraries should enable a more detailed analysis of rubber tree miRNA coding sequences and their promoters.
Here, we report the use of these libraries in the identi-fication and characterization of rubber tree miRNA genes and their responsiveness to phytohormones. The cis-regu-latory elements identified in the promoter regions of these miRNA genes help augment our understanding of miRNA gene regulation related to NR rubber production, growth and development of the rubber tree.
Materials and methods
Plant materials
A self-breeded H. brasiliensis RRIM 600 clone was cre-ated by Chachoengsao Rubber Research Center (CCRC), Rubber Research Institute of Thailand (RRIT), Thailand. Young leaves from the RRIM 600 clone were collected and immediately frozen in liquid nitrogen, followed by short-term storage at −80 °C until use. For genotype con-firmation, 12 EST-SSR markers selected for their high

923Mol Genet Genomics (2014) 289:921–933
1 3
polymorphism information content (PIC) value were used for amplification and comparison of the RRIM 600 sample against the reference sample previously used for transcrip-tome sequencing by Triwitayakorn et al. (2011) (Online Resource 1).
BAC library construction
High-molecular weight (HMW) DNA isolation was per-formed using methods modified from Zhang (2000), Peterson et al. (2002), and Epicentre® (2002). A 100 g of frozen leaves was homogenized, transferred to ice-cold homogenization buffer (HB) (10 mM Trizma base, 80 mM KCl, 10 mM EDTA, 1 mM spermine, pH 9.4–9.5) with 0.15 % β-mercaptoethanol and 0.5 % Triton X-l00, centri-fuged for pellet collection, and washed again in HB. Aga-rose plugs were prepared as described by Peterson et al. (2002). The plugs were incubated in lysis buffer (1 % w/v sodium lauryl sarcosine, 0.1 mg/mL proteinase K, 0.1 % w/v ascorbic acid, 6 mM EGTA, 200 mM l-lysine, and 0.13 % w/v sodium diethyldithiocarbamate) at 50 °C for 24 h, and were transferred to fresh lysis buffer for a sec-ond incubation at 50 °C for 24 h. Plugs were transferred again to fresh lysis buffer before incubation at 4 °C for another 24 h. The plugs were cleaned by serial washing on ice for 1 h/step with 0.5 M EDTA, pH 9.0–9.3; 0.5 M EDTA, pH 8.0; and 0.05 M EDTA, pH 8.0, respectively. The cleaned plugs were stored in 0.05 M EDTA, pH 8.0 at 4 °C until use.
For DNA preparation, the plugs were washed with TE (10 mM Tris–HCl, pH 8.0; 1 mM EDTA, pH 8.0) sup-plemented with 0.1 mM phenylmethyl sulfonyl fluoride (PMSF), followed by digestion with HindIII (1 unit per half plug). Fragments were separated in 1.0 % agarose gels by pulsed-field gel electrophoresis (PFGE) under the following conditions: 12.5 °C, 80 (pump settings), 120° angle, 6 V/cm, initial pulse time 90 s and final pulse time 90 s for 18 h. Gel areas containing 100–300 kb DNA fragments were excised and gel separation repeated a second time to further purify the insert DNA and remove small DNA fragments.
Size-selected DNA fragments (100–200 and 200–300 kb) were ligated to the pCC1BAC™ vector (Epicentre Biotechnologies, USA). Ligation products were desalted using an agarose cone (0.1 M glucose/1.0 % agarose), elec-troporated into TransforMAX™ EPI300™ E. coli (Epi-centre Biotechnologies), and cultured overnight at 37 °C on blue/white colony selection LB medium containing 12.5 μg/mL of chloramphenicol, 80 μg/mL X-gal, and 100 μg/mL IPTG. A total of 38,400 individual BAC colo-nies were hand-picked, grown in 384-well microtiter plates at 37 °C for 16 h to set up a master plate library, replicated using a 384-pin replicator to generate a copy plate library, and stored at −80 °C.
Fosmid library construction
Rubber tree genomic DNA was prepared by the cetyltri-methyl ammonium bromide (CTAB) method with some modifications. Five grams of leaf sample was ground in liquid nitrogen and the resulting fine powder was trans-ferred to a new 50 mL tube containing 15 mL heated CTAB buffer (0.1 M Tris HCl, pH 8.0; 1.4 M NaCl; 0.02 M EDTA and 2 w/v of CTAB) containing 500 µL of 20 % SDS. The resulting solution was incubated at 65 °C for 30 min and cleaned in a 15 mL solution con-taining chloroform and isoamyl alcohol at a 24:1 ratio. After centrifugation at 13,000×g for 20 min at 4 °C, the upper phase was transferred to a new 50 mL tube and the DNA was precipitated by 1 volume of cold isopro-panol. To collect the DNA pellet, the mixture was cen-trifuged at 10,000×g for 15 min at 4 °C and the pellet was washed with 70 % ethanol. The mixture was finally centrifuged at 10,000×g for 7 min at 4 °C. The DNA pellet was dissolved in 1 mL of TE containing RNaseA (10 µg/mL). Around 150 µg of sheared DNA was frac-tionated in a 1.0 % agarose gel on the PFGE apparatus containing 0.5× TBE buffer using the following settings: 12.5 °C, 120° angle, 6 V/cm, 1–40 s linear ramp for 18 h. End-repair, ligation, phage packaging and infection steps were performed as described by the CopyControl™ Fos-mid Library Production Kit (Epicentre Biotechnologies). The vectors were included in the kit, pre-cut at Eco72I (CopyControl™ pCC1FOS™) and end-dephosphorylated. EPI300™-T1R Phage T1-resistant E. coli was used for phage infection.
A total of 82,176 individual fosmid colonies were hand-picked, grown in 384-well microtiter plates at 37 °C for 16 h to set up a master plate library, replicated using a 384-pin replicator to generate a copy plate library, and stored at −80 °C.
Insert size estimation
BAC and fosmid library clones were randomly selected and induced to high copy number using 1,000× Copy Control Induction Solution (provided with the pCC1BAC cloning kit). DNA extraction was performed by the alkaline lysis extraction procedure with some modifications (Peterson et al. 2002). In brief, cells were suspended in Solution 1 (50 mM Glucose; 10 mM EDTA, pH 8.0; 25 mM Tris HCl, pH 8.0) at room temperature for 5 min and mixed with Solution 2 (0.2 N NaOH, 1 % SDS) at room temperature for 5 min. The Solution 3 (3 M potassium acetate, 28.45 % glacial acetic acid) was then added and the mixture left on ice for 7 min prior to centrifugation at 13,000×g for 20 min to collect the supernatant. DNA was precipitated, digested with NotI, and separated by PFGE. DNA fragments were

924 Mol Genet Genomics (2014) 289:921–933
1 3
visualized by SYBR® Gold Nucleic Acid Gel Stain (Inv-itrogen, USA), and their sizes were estimated using the PFGE Marker I (New England Bio Labs, USA).
DNA pooling
Bacterial artificial chromosome or fosmid clones on each plate were replicated by a 384-pin replicator, grown over-night in the presence of 1,000× Copy Control Induction Solution, and pooled. The pooled cells were centrifuged and DNA isolated using the modified alkaline lysis proto-col described above. A total of 100 and 214 BAC and fos-mid DNA pools (from each 384-well plate) were checked for DNA quality and quantity by gel electrophoresis and spectrophotometry.
BLASTn sequence verification
Randomly selected BAC and fosmid clones were induced to high copy number and subsequently sequenced by Mac-rogen (Korea). End sequences were trimmed to remove vector and low-quality sequences and analyzed for simi-larity with H. brasiliensis sequences in the NCBI Gen-Bank database using BLASTn. The BAC library was also assessed for chloroplast and mitochondria genome sequence contamination using BLASTn (Figueira et al. 2012; Zhang et al. 2007).
Primer design
In vivo and/or in vitro accumulation patterns of mature miRNA sequences of 10 rubber tree-specific miRNAs and 19 conserved miRNAs previously reported by Lertpan-yasampatha et al. (2012, 2013) and Gébelin et al. (2012) were used (Online Resource 2 and 3). These mature miRNA sequences were compared against the rubber tree genome database (Triwitayakorn et al. 2011) by BLAST to identify miRNA precursor (pre-miRNA) sequences. All candidate pre-miRNAs were confirmed to be non-coding gene sequences by comparison against the NCBI database with BLAST, then evaluated against the miRNA crite-ria, namely: (1) the potential to form secondary structures using UNAFold (Markham and Zuker 2008) with minimal folding free energy index (MFEI) (Zhang et al. 2006), (2) the presence of no more than four consecutive mismatches between miRNA and miRNA*, (3) base-pairing of at least 17 of the 25 nucleotides centered around the miRNA, and (4) a hairpin length of at least 75 nucleotides (Barvkar et al. 2013; Zhang et al. 2006).
miRNA-specific primers for the ten rubber tree-spe-cific miRNAs and six selected conserved miRNAs were designed based on confirmed pre-miRNA sequences and then tested for specific amplification with genomic DNA. Primers used to identify positive BAC/fosmid clones con-taining miRNA sequences are listed in Table 1.
Table 1 miRNA-specific primers used in BAC/fosmid DNA screening
a Mature miRNA sequences derived from Lertpanyasampatha et al. (2012)b Primers were designed by searching for mature miRNA sequences against the rubber tree transcriptome sequence database (Triwitayakorn et al. 2011) to identify pre-miRNAs
miRNAa miRNA-specific primer (5′ → 3′)b PCR product size (bp)
Forward primer Reverse primer
miR6166 GGAGGAGCCTCCCTTCCTC TACCAAGAGGAAGAAGAAGGA 112
miR6167 CAAAGCTTCCACCTGGGTAG GGTGCGGCTCCTGGTGTGG 129
miR6168 ATTCCTCTCTTGGTGCG TTTGTTTGTTTCCTCTCTT 129
miR6169 TTCCAGCATGAAATTTAATT TGTTCTCGCTTATTGCTT 160
miR6170 GAAAACAAAGAGTCTCTC AAAATGTGGGGGAATCGT 140
miR6171 AGGGGTGTATGGCATATTAG CTCTTTCCTCGCGTGGCTC 160
miR6172 ATAAAAAGAAACTCTTGCC GTTTTCTTUCTTUUUCACG 140
miR6173 GCAGTATCCATCGTTAAA TGGTCCGTTTCGTCCCGTG 100
miR6174 TAAGAAAGCAAGGATGATAG GCCCCTTTTTGGCCTGC 160
miR6175 TAATGTGACCCCCTCTTGA CTGTTGGGTGTGGTTCCCT 142
miR159 CAAAACTTATGACTAAATC GAGTAGTACAGGTGAAAG 243
miR160 CTCACCTCCCTGTATGCC CCAGGCATAATCCATTTCA 180
miR164 TCACCATTTTAGGACCTCACC CCCCCTTCCTCAACAACAAT 83
miR166 TGAATTCAACACCATCTTCA GCTGATGATCAATCTACTCTC 153
miR167 TCTTCTTGCCGTGCACCACTA TTTTTCATGCTACCAGCAACA 137
miR2118 GCGAGTAAAGGTTTCTCCCTT AATTTAGGCAGGGTGTGATCA 210

925Mol Genet Genomics (2014) 289:921–933
1 3
PCR screening for miRNA sequences
Each BAC/fosmid DNA pool was screened by PCR with the specific primers indicated in Table 1. PCR was per-formed in a 15 μL reaction mix containing: 1× PCR buffer, 0.2 mM MgCl2, 0.2 mM dNTP mix, 0.25 unit of Taq DNA polymerase (Thermo Fisher Scientific, USA), 0.3 pmol each of the forward and reverse primers, and tem-plate DNA. The PCRs were run with the following condi-tions: 94 °C for 5 min, followed by 35 cycles of 94 °C for 30 s, 58 °C for 30 s and 72 °C for 1 min. For positive pools, their corresponding plates were used as templates for col-ony PCR in 384-well PCR plates containing 10 μL PCR mixes prepared as described above. An ABI Veriti® 384-well thermal cycler (Applied Biosystems, USA) was used to perform PCRs using the following conditions: 94 °C for 10 min, followed by 45 cycles of 94 °C for 1 min, 55 °C for 1 min and 72 °C for 1 min, ending at 4 °C for 5 min, and maintained at −20 °C. Positive BAC/fosmid clones were visualized by 1.0 % agarose electrophoresis using 1× TBE buffer.
Sequencing
The identified positive BAC/fosmid clones were induced to high copy number as described above and 1,000 ng of total DNA from each was extracted for 454-pyrosequenc-ing as described by Triwitayakorn et al. (2011). Sequence data were trimmed to remove vector sequences and assem-bled, and all candidate contigs were confirmed to be pre-miRNAs by the four miRNA criteria listed above.
Identification of miRNAs response to phytohormones
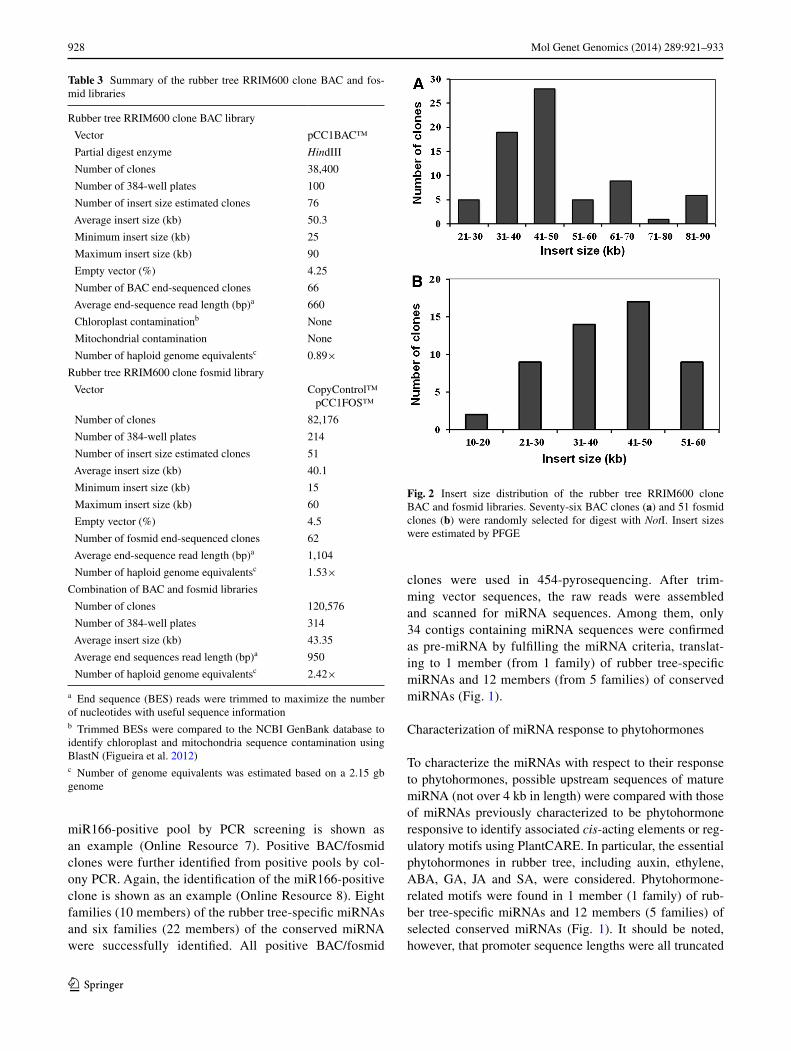
Upstream sequences (up to 4 kb) of confirmed pre-miR-NAs coding sequences were analyzed for their cis-acting elements or regulatory motifs, including phytohormone response motifs (auxin, ethylene, ABA, GA, JA and SA), environment response motifs, biotic and abiotic stress response motifs, and tissue-specific expression using the PlantCARE (Lescot et al. 2002) (Fig. 1). The results were compared with selected miRNAs previously determined to be phytohormone-responsive (Lertpanyasampatha et al. 2012, 2013; Gébelin et al. 2012). For ethylene response analysis, all miRNA promoter lengths were truncated to 1,000 bp (Table 2).
Quantitative real-time PCR (qRT-PCR)
Four-month-old budded RRIM600 clones with similar physical characteristics from an RRIT-certified propaga-tion farm were completely sprayed with 2.5 % ethephon plus 0.05 % Tween-20 at 0 and 24 h. The second and third
leaves from the apical shoot were collected individually at daytime (0, 4, 24, and 48 h after the first treatment) to avoid the effects of changing light conditions. The experiment was performed with three replicates for each time point.
Total RNA was isolated using the mirVana™ miRNA isolation kit (Ambion, USA). The concentration and qual-ity of RNA were determined by spectrophotometer and gel electrophoresis, and the total RNA was treated with RNase-free DNase (Ambion). Poly A polymerase and ATP were used to polyadenylate all miRNAs, followed by first-strand cDNA synthesis on 100 ng of total RNA using the Ncode™ miRNA First-Strand cDNA Synthesis and qRT-PCR kits (Invitrogen).
Ethylene-responsive and nonresponsive miRNA expres-sion was detected with 5′ miRNA-specific primers based on the first 14–15 nt of the mature miRNA sequences (Online Resource 4). Approximately, 20 ng/μL rubber tree cDNA was used for qRT-PCR using the SYBR Green PCR Master Mix (Applied Biosystems) on an ABI 7500 machine (Applied Biosystems). Relative quantities of can-didate miRNA transcripts, normalized to a validated inter-nal standard gene (miR160), were automatically calculated as 2−△△Ct by the ABI 7500 software using default Ct (cycle threshold) settings. All first-strand cDNA syntheses and RT-PCR amplifications were performed in triplicate for each gene per treatment. Standard error and ANOVA were used for statistical and significance analyses, respectively, taking into account all three-by-three replicates (Patanun et al. 2012).
Results
BAC and fosmid library construction
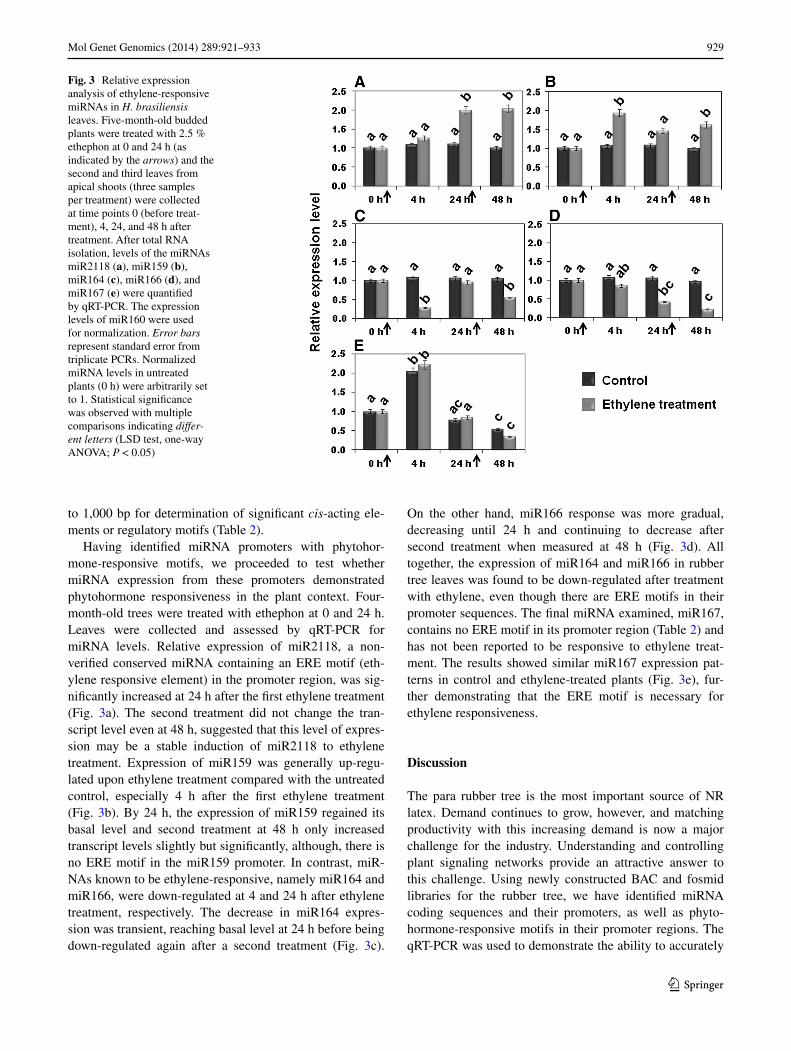
To generate the rubber tree BAC library, approximately 70 DNA plugs per 30 g of fresh leaf weight were par-tially digested with HindIII and fragments of 100–200 and 200–300 kb ligated to the pCC1BAC™ vector. The rubber tree BAC library containing 38,400 clones was arrayed in 100 384-well microtiter plates. NotI digestion of 76 ran-dom clones was used to estimate the average insert size of the library at 50.3 kb, with sizes ranging from 25 to 90 kb (Table 3; Fig. 2a). Based on the estimated 2.15 gb rub-ber tree genome size (Rahman et al. 2013), library cover-age was estimated to be 0.89 haploid genome equivalent. To enhance the coverage of our library resource, a fosmid library was also constructed. This library contains 82,176 clones arrayed in 214 384-well microtiter plates and has an average insert size of 40.1 kb (Fig. 2b). Coverage was estimated at 1.53 haploid genome equivalents. Combined, the BAC and fosmid libraries provide coverage of 2.42 haploid genome equivalents and were calculated to cover

926 Mol Genet Genomics (2014) 289:921–933
1 3
around 91.2 % of the genome using the formula of N = ln (1 − P)/ln (1 − [I/GS]) (Sambrook et al. 2001).
Library characterization
Library quality was initially examined by end-sequence analysis. The end sequences of 66 and 62 randomly selected BAC and fosmid clones were trimmed to remove vector and low-quality sequences, resulting in an average end-sequence read length of 950 bp. All rubber tree end sequences were matched to the rubber tree genome data-base (Triwitayakorn et al. 2011) with an E value thresh-old of ≤e−25 and 82–100 % sequence identity (data not shown). In addition, BAC library quality was also exam-ined by assessing chloroplast and mitochondria genome sequence contamination using BLASTn searches against
the NCBI GenBank database (Figueira et al. 2012). The results showed no similarity hits with any chloroplast or mitochondria gene sequences (Table 3). These preliminary characterizations of the rubber tree BAC and fosmid librar-ies suggested that the libraries had sufficient quality to be used further application, including promoter analysis of genes of interest.
Identification of miRNA precursors in the rubber tree genome database
The BAC and fosmid libraries generated were then applied to identify and characterize rubber tree-specific miRNAs and conserved miRNAs promoter regions. Mature miRNA sequences of 10 rubber tree-specific miRNAs and 19 con-served miRNAs (Online Resource 2) were searched against
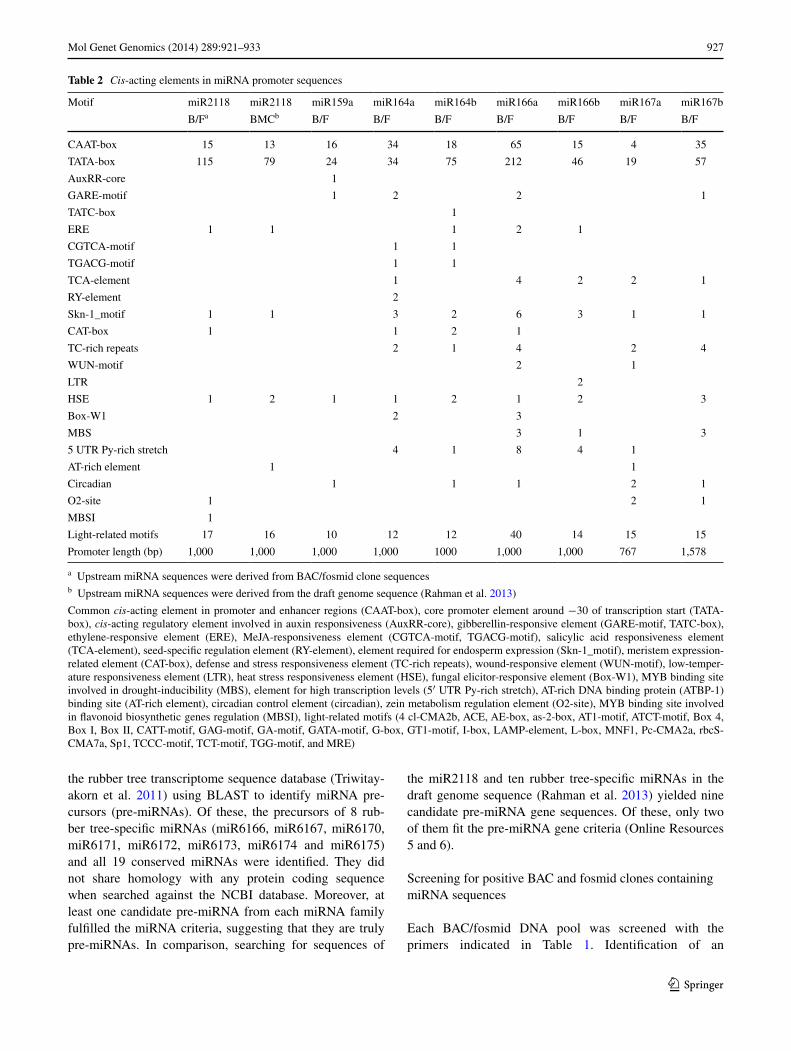
Fig. 1 Flowchart of process to identify phytohormone responsiveness of rubber tree-specific miRNAs and conserved miRNAs using an integration of BAC/fosmid libraries, promoter analysis and gene expression analysis. Underlined miRNA IDs indicate sequences identi-fied to have ethylene-responsive gene expression activity in this study. Phytohormone-related motifs found in miRNA gene promoters are indicated in parentheses. Italicized numbers indicate the lengths (in bp) of analyzed promoter sequences
927Mol Genet Genomics (2014) 289:921–933
1 3
the rubber tree transcriptome sequence database (Triwitay-akorn et al. 2011) using BLAST to identify miRNA pre-cursors (pre-miRNAs). Of these, the precursors of 8 rub-ber tree-specific miRNAs (miR6166, miR6167, miR6170, miR6171, miR6172, miR6173, miR6174 and miR6175) and all 19 conserved miRNAs were identified. They did not share homology with any protein coding sequence when searched against the NCBI database. Moreover, at least one candidate pre-miRNA from each miRNA family fulfilled the miRNA criteria, suggesting that they are truly pre-miRNAs. In comparison, searching for sequences of
the miR2118 and ten rubber tree-specific miRNAs in the draft genome sequence (Rahman et al. 2013) yielded nine candidate pre-miRNA gene sequences. Of these, only two of them fit the pre-miRNA gene criteria (Online Resources 5 and 6).
Screening for positive BAC and fosmid clones containing miRNA sequences
Each BAC/fosmid DNA pool was screened with the primers indicated in Table 1. Identification of an
Table 2 Cis-acting elements in miRNA promoter sequences
a Upstream miRNA sequences were derived from BAC/fosmid clone sequencesb Upstream miRNA sequences were derived from the draft genome sequence (Rahman et al. 2013)
Common cis-acting element in promoter and enhancer regions (CAAT-box), core promoter element around −30 of transcription start (TATA-box), cis-acting regulatory element involved in auxin responsiveness (AuxRR-core), gibberellin-responsive element (GARE-motif, TATC-box), ethylene-responsive element (ERE), MeJA-responsiveness element (CGTCA-motif, TGACG-motif), salicylic acid responsiveness element (TCA-element), seed-specific regulation element (RY-element), element required for endosperm expression (Skn-1_motif), meristem expression-related element (CAT-box), defense and stress responsiveness element (TC-rich repeats), wound-responsive element (WUN-motif), low-temper-ature responsiveness element (LTR), heat stress responsiveness element (HSE), fungal elicitor-responsive element (Box-W1), MYB binding site involved in drought-inducibility (MBS), element for high transcription levels (5′ UTR Py-rich stretch), AT-rich DNA binding protein (ATBP-1) binding site (AT-rich element), circadian control element (circadian), zein metabolism regulation element (O2-site), MYB binding site involved in flavonoid biosynthetic genes regulation (MBSI), light-related motifs (4 cl-CMA2b, ACE, AE-box, as-2-box, AT1-motif, ATCT-motif, Box 4, Box I, Box II, CATT-motif, GAG-motif, GA-motif, GATA-motif, G-box, GT1-motif, I-box, LAMP-element, L-box, MNF1, Pc-CMA2a, rbcS-CMA7a, Sp1, TCCC-motif, TCT-motif, TGG-motif, and MRE)
Motif miR2118 miR2118 miR159a miR164a miR164b miR166a miR166b miR167a miR167b
B/Fa BMCb B/F B/F B/F B/F B/F B/F B/F
CAAT-box 15 13 16 34 18 65 15 4 35
TATA-box 115 79 24 34 75 212 46 19 57
AuxRR-core 1
GARE-motif 1 2 2 1
TATC-box 1
ERE 1 1 1 2 1
CGTCA-motif 1 1
TGACG-motif 1 1
TCA-element 1 4 2 2 1
RY-element 2
Skn-1_motif 1 1 3 2 6 3 1 1
CAT-box 1 1 2 1
TC-rich repeats 2 1 4 2 4
WUN-motif 2 1
LTR 2
HSE 1 2 1 1 2 1 2 3
Box-W1 2 3
MBS 3 1 3
5 UTR Py-rich stretch 4 1 8 4 1
AT-rich element 1 1
Circadian 1 1 1 2 1
O2-site 1 2 1
MBSI 1
Light-related motifs 17 16 10 12 12 40 14 15 15
Promoter length (bp) 1,000 1,000 1,000 1,000 1000 1,000 1,000 767 1,578
928 Mol Genet Genomics (2014) 289:921–933
1 3
miR166-positive pool by PCR screening is shown as an example (Online Resource 7). Positive BAC/fosmid clones were further identified from positive pools by col-ony PCR. Again, the identification of the miR166-positive clone is shown as an example (Online Resource 8). Eight families (10 members) of the rubber tree-specific miRNAs and six families (22 members) of the conserved miRNA were successfully identified. All positive BAC/fosmid
clones were used in 454-pyrosequencing. After trim-ming vector sequences, the raw reads were assembled and scanned for miRNA sequences. Among them, only 34 contigs containing miRNA sequences were confirmed as pre-miRNA by fulfilling the miRNA criteria, translat-ing to 1 member (from 1 family) of rubber tree-specific miRNAs and 12 members (from 5 families) of conserved miRNAs (Fig. 1).
Characterization of miRNA response to phytohormones
To characterize the miRNAs with respect to their response to phytohormones, possible upstream sequences of mature miRNA (not over 4 kb in length) were compared with those of miRNAs previously characterized to be phytohormone responsive to identify associated cis-acting elements or reg-ulatory motifs using PlantCARE. In particular, the essential phytohormones in rubber tree, including auxin, ethylene, ABA, GA, JA and SA, were considered. Phytohormone-related motifs were found in 1 member (1 family) of rub-ber tree-specific miRNAs and 12 members (5 families) of selected conserved miRNAs (Fig. 1). It should be noted, however, that promoter sequence lengths were all truncated
Table 3 Summary of the rubber tree RRIM600 clone BAC and fos-mid libraries
a End sequence (BES) reads were trimmed to maximize the number of nucleotides with useful sequence informationb Trimmed BESs were compared to the NCBI GenBank database to identify chloroplast and mitochondria sequence contamination using BlastN (Figueira et al. 2012)c Number of genome equivalents was estimated based on a 2.15 gb genome
Rubber tree RRIM600 clone BAC library
Vector pCC1BAC™
Partial digest enzyme HindIII
Number of clones 38,400
Number of 384-well plates 100
Number of insert size estimated clones 76
Average insert size (kb) 50.3
Minimum insert size (kb) 25
Maximum insert size (kb) 90
Empty vector (%) 4.25
Number of BAC end-sequenced clones 66
Average end-sequence read length (bp)a 660
Chloroplast contaminationb None
Mitochondrial contamination None
Number of haploid genome equivalentsc 0.89×Rubber tree RRIM600 clone fosmid library
Vector CopyControl™ pCC1FOS™
Number of clones 82,176
Number of 384-well plates 214
Number of insert size estimated clones 51
Average insert size (kb) 40.1
Minimum insert size (kb) 15
Maximum insert size (kb) 60
Empty vector (%) 4.5
Number of fosmid end-sequenced clones 62
Average end-sequence read length (bp)a 1,104
Number of haploid genome equivalentsc 1.53×Combination of BAC and fosmid libraries
Number of clones 120,576
Number of 384-well plates 314
Average insert size (kb) 43.35
Average end sequences read length (bp)a 950
Number of haploid genome equivalentsc 2.42×
Fig. 2 Insert size distribution of the rubber tree RRIM600 clone BAC and fosmid libraries. Seventy-six BAC clones (a) and 51 fosmid clones (b) were randomly selected for digest with NotI. Insert sizes were estimated by PFGE
929Mol Genet Genomics (2014) 289:921–933
1 3
to 1,000 bp for determination of significant cis-acting ele-ments or regulatory motifs (Table 2).
Having identified miRNA promoters with phytohor-mone-responsive motifs, we proceeded to test whether miRNA expression from these promoters demonstrated phytohormone responsiveness in the plant context. Four-month-old trees were treated with ethephon at 0 and 24 h. Leaves were collected and assessed by qRT-PCR for miRNA levels. Relative expression of miR2118, a non-verified conserved miRNA containing an ERE motif (eth-ylene responsive element) in the promoter region, was sig-nificantly increased at 24 h after the first ethylene treatment (Fig. 3a). The second treatment did not change the tran-script level even at 48 h, suggested that this level of expres-sion may be a stable induction of miR2118 to ethylene treatment. Expression of miR159 was generally up-regu-lated upon ethylene treatment compared with the untreated control, especially 4 h after the first ethylene treatment (Fig. 3b). By 24 h, the expression of miR159 regained its basal level and second treatment at 48 h only increased transcript levels slightly but significantly, although, there is no ERE motif in the miR159 promoter. In contrast, miR-NAs known to be ethylene-responsive, namely miR164 and miR166, were down-regulated at 4 and 24 h after ethylene treatment, respectively. The decrease in miR164 expres-sion was transient, reaching basal level at 24 h before being down-regulated again after a second treatment (Fig. 3c).
On the other hand, miR166 response was more gradual, decreasing until 24 h and continuing to decrease after second treatment when measured at 48 h (Fig. 3d). All together, the expression of miR164 and miR166 in rubber tree leaves was found to be down-regulated after treatment with ethylene, even though there are ERE motifs in their promoter sequences. The final miRNA examined, miR167, contains no ERE motif in its promoter region (Table 2) and has not been reported to be responsive to ethylene treat-ment. The results showed similar miR167 expression pat-terns in control and ethylene-treated plants (Fig. 3e), fur-ther demonstrating that the ERE motif is necessary for ethylene responsiveness.
Discussion
The para rubber tree is the most important source of NR latex. Demand continues to grow, however, and matching productivity with this increasing demand is now a major challenge for the industry. Understanding and controlling plant signaling networks provide an attractive answer to this challenge. Using newly constructed BAC and fosmid libraries for the rubber tree, we have identified miRNA coding sequences and their promoters, as well as phyto-hormone-responsive motifs in their promoter regions. The qRT-PCR was used to demonstrate the ability to accurately
Fig. 3 Relative expression analysis of ethylene-responsive miRNAs in H. brasiliensis leaves. Five-month-old budded plants were treated with 2.5 % ethephon at 0 and 24 h (as indicated by the arrows) and the second and third leaves from apical shoots (three samples per treatment) were collected at time points 0 (before treat-ment), 4, 24, and 48 h after treatment. After total RNA isolation, levels of the miRNAs miR2118 (a), miR159 (b), miR164 (c), miR166 (d), and miR167 (e) were quantified by qRT-PCR. The expression levels of miR160 were used for normalization. Error bars represent standard error from triplicate PCRs. Normalized miRNA levels in untreated plants (0 h) were arbitrarily set to 1. Statistical significance was observed with multiple comparisons indicating differ-ent letters (LSD test, one-way ANOVA; P < 0.05)

930 Mol Genet Genomics (2014) 289:921–933
1 3
predict ethylene responsiveness of miRNA with ERE pro-moter motifs. These results bring the field a step forward toward maximizing desirable miRNA-regulated agronomic traits through phytohormone manipulation in rubber tree.
Recent research has made great progress in under-standing the intersection between miRNA pathways and phytohormone responses. In particular, there are many reports regarding interactions between miRNAs and phy-tohormones and the impact of these interactions on rubber tree growth, development, and yield (Gébelin et al. 2012; Dusotoit-Coucaud et al. 2010; Lertpanyasampatha et al. 2013). In this study, we identified the precursors of 8 rub-ber tree-specific miRNAs (miR6166, miR6167, miR6170, miR6171, miR6172, miR6173, miR6174 and miR6175) and 19 conserved miRNAs which fulfilled the miRNA criteria (Zhang et al. 2006; Barvkar et al. 2013) using our BAC and fosmid libraries. On the other hand, confirmed precursors of only one member from one family were found in the draft genome database of Rahman et al. (2013) (Online Resource 5). These observations suggest that our libraries provide unique information not available in the draft genome.
Among the identified precursor sequences, the promoter regions of rubber tree-specific miRNAs and conserved miRNAs were found to contain phytohormone-respon-sive elements, such as ERE, auxin-responsive elements (AuxRR-core and AuxRR-core) and gibberellin-related motifs (GARE-motif, P-box and TATC-box). Promoter analyses also revealed numerous TATA-box motifs in all miRNA promoters, providing support for the hypothesis put forth by Zhao and Li (2013) that TATA-box and CAAT-box enrichment in miRNA promoter regions may reflect their fundamental roles in miRNA expression.
These genomic data were confirmed by analysis of miRNA gene expression in response to ethylene. Up-reg-ulation of the non-verified miR2118 and the conserved miR159 was found in ethylene-treated plants. Studying expression of the target genes would deepen our under-standing of the function of these miRNA and how they relate to ethylene treatment. It should be noted that the ERE motif was not found in the miR159 promoter region in this study. This may be because only the first 1,000 bp of each 5′ promoter sequence was subjected to promoter analysis for the purpose of uniformity in our analyses, due to the variation in promoter sequence lengths. Neverthe-less, miRNA promoters can be as long as 5,000 bp (Heik-kinen et al. 2008; Cui et al. 2009), although only the first 2,000 bp are typically analyzed (Cui et al. 2009; Zhou et al. 2007). Furthermore, studies in Arabidopsis have shown that 90 % of predicted cis-regulatory elements are located within the first 800 bp of promoter sequences (Zhao and Li 2013; Megraw et al. 2006). Interestingly, although an ERE motif was not found in the miR159 promoter,
auxin-responsive and gibberellin-related elements were present. Crosstalk between ethylene, auxin and gibberel-lins has been suggested to play a role in plant growth and development. For instance, Chen et al. (2012) investigated the crosstalk between ethylene and auxin in the control of lateral root development of Medicago truncatula, while Achard et al. (2004) reported that gibberellins enhanced the transcript level of miR159 and are involved in the control of flowering time and anther development. Consequently, we surmise that these auxin-responsive and gibberellin-related elements were responsible for modulating miR159 response to ethylene in the absence of the ERE motif.
Among the previously characterized miRNAs, miR159, miR164, and miR166 have already been reported to be eth-ylene responsive (Duan et al. 2013; Gébelin et al. 2012). The rose plant was observed to exhibit down-regulation of miR164 by ethylene treatment, while expression from NAC, its target gene, was up-regulated (Pei et al. 2013a). Similarly, the expression of miR166 responded to ethyl-ene treatment by decreasing in expression level, varying inversely with expression of its target (beta-galactosidase) (Pei et al. 2013b). Along with miR167, the promoters of 12 members of these 5 miRNA families were found to have at least one phytohormone-related motif. It should be noted that pre-miRNA sequences extracted from available data-base sequences formed weaker stem-loop structures than those derived from our BAC and fosmid sequences (Online Resource 6), further underlining how our libraries offer a powerful new tool for rubber tree genome studies.
As a basis for comparison, miR167 was chosen to be a negative control for ethylene response. As far as the litera-ture is concerned, there have been no reports on miR167 expression in response to ethylene and no ERE motif is present in its promoter sequence. As there is a circadian response motif on the miR167 promoter, however, it was not unexpected to find up-regulation of miR167 in both control and ethylene-treated leaves at the 4 h time point (4 p.m.) Rhythmic control of miRNA expression has signifi-cant consequences for circadian timing and appears to be conserved from plants to mammals (Hansen et al. 2011). In particular, miR160b and miR167d have been identified to be regulated by circadian rhythm in Arabidopsis (Hansen et al. 2011; Hazen et al. 2009).
To date, several genomic resources such as genetic mark-ers, genetic linkage maps and genome sequences have been developed with the aim to facilitate molecular breeding for NR latex yield improvement. Even with these resources, however, information about this complex genome is still limited. In this study, the first rubber tree BAC library was created to further enhance our understanding of the genetic background of the rubber tree. This library consists of 38,400 clones with an average insert size of 50.3 kb pro-viding 0.89 haploid genome equivalent of coverage based

931Mol Genet Genomics (2014) 289:921–933
1 3
on an estimated 2.15 gb genome size (Rahman et al. 2013). Less than 5 % of the clones are empty vectors, an accept-able rate for a high-quality BAC library (Luo and Wing 2003; Schulte et al. 2011). On the other hand, the 50.3 kb average insert size (despite several cycles of size improve-ment) compares less favorably against the 80–150 kb aver-age insert size of BAC libraries constructed from genomes of other organisms (Cacao et al. 2013; Figueira et al. 2012; Gschwend et al. 2011; Roche et al. 2002). In the absence of any other reports on rubber tree BAC library construc-tion, it is difficult to assess whether or not the insert size limitation was imposed by the nature of the rubber tree itself. Different plant varieties exhibit different amounts of cell wall hydrolases and enzymes, polyphenolic substances, and/or polysaccharides, which may affect DNA digestion or cloning and thereby result in smaller insert sizes (Zhang et al. 1995, 2012). One report concerning the construction of Drosophila BAC libraries revealed average insert sizes between 50 and 150 kb (Osoegawa et al. 2007).
Due to the relatively small insert sizes of our rubber tree BAC library, we additionally constructed the fosmid library to both increase the variety of DNA fragments in the library and to enhance genome coverage. The rubber tree fosmid library contains 82,176 clones with an average insert size of 40.1 kb, which is standard for fosmid library construc-tion (Zhang et al. 2007; Kim et al. 2003; Peterson et al. 2002). Combining BAC and fosmid libraries resulted in 2.42 haploid genome equivalents of coverage, which is cal-culated to cover around 91.2 % of the genome.
The analysis of BAC end sequences (BESs) is generally used for the characterization of BAC libraries by identify-ing contamination of organellar genomes (Figueira et al. 2012; Schulte et al. 2011). The BESs of our rubber tree BAC library did not show any matches with either chlo-roplast or mitochondria sequences. Moreover, 100 % of BAC and 84 % of fosmid end sequences showed 82–100 and 86–100 % sequence identity to the rubber tree tran-scriptome (Triwitayakorn et al. 2011) and the draft genome sequences (Rahman et al. 2013), with an E value thresh-old of ≤e−25. Interestingly, 16 % of fosmid end sequences did not correspond with any sequence within any published database, suggesting that our BAC and fosmid librar-ies may enable identification and study of unknown rub-ber tree sequences. Therefore, beyond its application here toward the understanding of phytohormone regulation of miRNAs, these libraries should provide a valuable resource for genomic research for rubber tree and may be applied to other genome-wide studies, such as physical mapping, gene walking, and whole genome sequencing.
In this study, we characterized a panel of miRNAs for phytohormone response by precisely identifying pre-miRNA sequences and their promoters as well as phytohor-mone-responsive motifs in these promoters. Furthermore,
these findings were verified by testing with ethylene stim-ulation of rubber trees, demonstrating that the miR2118, miR159, miR164 and miR166 are responsive to ethylene, as forecasted by our genomic analysis strategy. In addition, we successfully generated alternative tools for the research community—the first BAC and fosmid rubber tree librar-ies, representing 2.42 haploid genome equivalents. Overall, our results provide a foundation for further investigation of the regulation of rubber tree miRNAs. The optimiza-tion of desirable agronomic traits of the rubber tree may be achieved by adjusting miRNA levels by phytohormone exposure. Better understanding of miRNA-controlled gene networks would help unravel the complex biology of the rubber tree, broadening the possibilities for rubber tree improvement.
Acknowledgments The plant materials used in BAC and fosmid library construction were provided by Ms. Kanikar Teerawatanasuk, CRRC-Rubber Research Institute of Thailand (CRRC-RRIT), Depart-ment of Agriculture. We would like to express special gratitude to Ms. Samaporn Teeravechyan, National Center for Genetic Engineer-ing and Biotechnology (BIOTEC) (Thailand) for her help in English editing. This work was supported by BIOTEC, RRIT, and Mahidol University.
References
Achard P, Herr A, Baulcombe DC, Harberd NP (2004) Modulation of floral development by a gibberellin-regulated microRNA. Devel-opment 131:3357–3365
Association of Natural Rubber Producing Countries (ANRPC) (2010) Average annual yield in member countries in a monthly bulletin of market trends and statistics. Nat Rubber Trends Stat 2:11
Barvkar VT, Pardeshi VC, Kale SM, Qiu S, Rollins M, Datla R, Gupta VS, Kadoo NY (2013) Genome-wide identification and charac-terization of microRNA genes and their targets in flax (Linum usitatissimum). Planta 237:1149–1161
Cacao SMB, Silva NV, Domingues DS, Vieira LGE, Diniz LEC, Vin-ecky F, Alves GSC et al (2013) Construction and characterization of a BAC library from the Coffea arabica genotype Timor hybrid CIFC 832/2. Genetica 141:217–226
Chen L, Wang T, Zhao M, Zhang W (2012) Ethylene-responsive miR-NAs in roots of Medicago truncatula identified by high-through-put sequencing at whole genome level. Plant Sci 184:14–19
Cui X, Xu SM, Mu DS, Yang ZM (2009) Genomic analysis of rice microRNA promoters and clusters. Gene 431:61–66
Duan C, Argout X, Gébelin V, Summo M, Dufayard JF, Leclercq J, Kuswanhadi PP et al (2013) Identification of the Hevea brasilien-sis AP2/ERF superfamily by RNA sequencing. BMC Genomics 14:30
Dusotoit-Coucaud A, Kongsawadworakul P, Maurousset L, Viboonjun U, Brunel N, Pujade-Renaud V, Chrestin H, Sakr S (2010) Eth-ylene stimulation of latex yield depends on the expression of a sucrose transporter (HbSUT1B) in rubber tree (Hevea brasilien-sis). Tree Physiol 30:1586–1598
Epicentre® (2002) CopyControl™ BAC cloning kit. Product manual. http://genomeprojectsolutions.com/pdfs/pcc1BAC.pdf. Accessed 1 May 2011
Figueira TRS, Okura V, Silva FR, Silva MJ, Kudrna D, Ammiraju JSS, Talag J, Wing R, Arruda P (2012) A BAC library of the

932 Mol Genet Genomics (2014) 289:921–933
1 3
SP80-3280 sugarcane variety (Saccharum sp.) and its inferred microsynteny with the sorghum genome. BMC Res Notes 5:185
Gébelin V, Argout X, Engchuan W, Pitollat B, Duan C, Montoro P, Leclercq J (2012) Identification of novel microRNAs in Hevea brasiliensis and computational prediction of their targets. BMC Plant Biol 12:18
Gébelin V, Leclercq J, Argout X, Chaidamsari T, Hu S, Tang C, Sarah G, Yang M, Montoro P (2013a) The small RNA profile in latex from Hevea brasiliensis trees is affected by tapping panel dry-ness. Tree Physiol 31:1084–1098
Gébelin V, Leclercq J, Chaorong T, Songnian H, Tang C, Montoro P (2013b) Regulation of MIR genes in response to abiotic stress in Hevea brasiliensis. Int J Mol Sci 14:19587–19604
Gonthier L, Bellec A, Blassiau C, Prat E, Helmstetter N, Rambaud C, Huss B, Hendriks T, Berges H, Quillet MC (2010) Construction and characterization of two BAC libraries representing a deep coverage of the genome of chicory (Cichorium intybus L., Aster-aceae). BMC Res 3:225
Gray WM (2004) Hormonal regulation of plant growth and develop-ment. PLoS Biol 2:1270–1273
Gschwend AR, Yu Q, Moore P, Saski C, Chen C, Wang J, Na JK, Ming R (2011) Construction of papaya male and female BAC libraries and application in physical mapping of the sex chromo-somes. J Biomed Biotechnol. doi:10.1155/2011/929472
Hansen KF, Sakamoto K, Obrietan K (2011) MicroRNAs: a potential interface between the circadian clock and human health. Genome Med 3:10
Hazen SP, Naef F, Quisel T, Gendron JM, Chen H, Ecker JR, Bor-evitz JO, Kay SA (2009) Exploring the transcriptional landscape of plant circadian rhythms using genome tiling arrays. Genome Biol 10:17
Heikkinen L, Asikainen S, Wong G (2008) Identification of phylo-genetically conserved sequence motifs in microRNA 5′ flanking sites from C. elegans and C. briggsae. BMC Mol Biol 9:105
Jeukens J, Boyle B, Kukavica-Ibrulj I, St-Cyr J, Le Vesque RC, Ber-natchez L (2011) BAC library construction, screening and clone sequencing of lake whitefish (Coregonus clupeaformis, Salmoni-dae) towards the elucidation of adaptive species divergence. Mol Ecol Resour 11:541–549
Kamimura M, Tateishi K, Tanaka-Okuyama M, Okabe T, Shibata F, Sahara K, Yasukochi Y (2012) EST sequencing and fosmid library construction in a non-model moth, Mamestra brassicae, for comparative mapping. Genome 55(11):775–781
Kim CG, Fujiyam A, Saitou N (2003) Construction of a gorilla fos-mid library and its PCR screening system. Genomics 82:571–574
Lertpanyasampatha M, Gao L, Kongsawadworakul P, Viboonjun U, Chrestin H, Liu R, Chen X, Narangajavana J (2012) Genome-wide analysis of microRNAs in rubber tree (Hevea brasiliensis L.) using high-throughput sequencing. Planta 236(2):437–445
Lertpanyasampatha M, Pramoolkit P, Kongsawadworakul P, Viboon-jun U, Chrestin H, Narangajavana J (2013) Conserved microR-NAs and their targets in rubber tree and the related Euphorbia-ceous species. J Plant Biochem Biot 22(1):52–61
Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, VandePeer Y, Rouzé P, Rombauts S (2002) PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res 30(1):325–327
Liu Q, Zhang YC, Wang CY, Luo YC, Huang QJ, Chen SY, Zhou H, Qu LH, Chen YQ (2009) Expression analysis of phytohormone-regulated microRNAs in rice, implying their regulation roles in plant hormone signaling. FEBS Lett 583:723–728
Liu SY, Yu K, Huffner M, Park SJ, Banik M, Pauls KP, Crosby W (2010) Construction of a BAC library and a physical map of a major QTL for CBB resistance of common bean (Phaseolus vul-garis L.). Genetica 138(7):709–716
Luo M, Wing RA (2003) An improved method for plant BAC library construction. Methods Mol Biol 236:3–20
Markham NR, Zuker M (2008) UNAFold: software for nucleic acid folding and hybridization. Methods Mol Biol 453:3–31
Megraw M, Baev V, Rusinov V, Jensen ST, Kalantidis K, Hatzigeor-giou AG (2006) MicroRNA promoter element discovery in Arabidopsis. RNA 12:1612–1619
Osoegawa K, Vessere GM, Shu CL, Hoskins RA, Abad JP, Pablos B, Villasante A, Jong PJ (2007) BAC clones generated from sheared DNA. Genomics 89(2):291–299
Ozsolak F, Poling LL, Wang Z, Liu H, Liu XS, Roeder RG, Zhang X, Song JS, Fisher DE (2008) Chromatin structure analyses identify miRNA promoters. Genes Dev 22:3172–3183
Patanun O, Lertpanyasampatha M, Sojikul P, Viboonjun U, Naranga-javana J (2012) Computational identification of microRNAs and their targets in cassava (Manihot esculenta Crantz.). Mol Bio-technol 53:257–269
Pei H, Ma N, Tian J, Luo J, Chen J, Li J, Zheng Y, Chen X, Fei Z, Gao J (2013a) An NAC transcription factor controls ethylene-reg-ulated cell expansion in flower petals. Plant Physiol 163:775–791
Pei H, Ma N, Chen J, Zheng Y, Tian J, Jing L, Zhang S, Fei Z, Gao J (2013b) Integrative analysis of miRNA and MRNA profiles in response to ethylene in rose petals during flower opening. PLoS One 8:5
Peterson D, Tomkins J, Frisch D, Wing R, Paterson A (2002) Con-struction of plant bacterial artificial chromosome (BAC) libraries: an illustrated guide. J Agric Genomics 5:1–100
Rahman AYA, Usharraj AO, Misra BB, Thottathil GP, Jayasekaran K, Feng Y, Hou S et al (2013) Draft genome sequence of the rubber tree Hevea brasiliensis. BMC Genomics 14:75
Roche D, Conner JA, Budiman MA, Frisch D, Wing R, Hanna WW, Ozias-Akins P (2002) Construction of BAC libraries from two apomictic grasses to study the microcolinearity of their apospory-specific genomic regions. Theor Appl Genet 104:804–812
Sambrook J, Fritsch EF, Maniatis T (2001) Molecular cloning: a labo-ratory manual, 3rd edn. CSH Laboratory Press, New York
Schulte D, Ariyadasa R, Shi B, Fleury D, Saski C, Atkins M, Jong P, Wu CC, Graner A, Langridge P, Stein N (2011) BAC library resources for map-based cloning and physical map construction in barley (Hordeum vulgare L.). BMC Genomics 12:247
Sehgal SK, Li W, Rabinowicz PD, Chan A, Simkova H, Dolezel J, Gill BS (2012) Chromosome arm-specific BAC end sequences permit comparative analysis of homoeologous chromosomes and genomes of polyploid wheat. BMC Plant Biol 12:64
Solonenko SA, Ignacio-Espinoza JC, Alberti A, Cruaud C, Hallam S, Konstantinidis K, Tyson G, Wincker P, Sullivan MB (2013) Sequencing platform and library preparation choices impact viral metagenomes. BMC Genomics 14:320
Triwitayakorn K, Chatkulkawin P, Kanjanawattanawong S, Sraphet S, Yoocha T, Sangsrakru D, Chanprasert J et al (2011) Transcrip-tome sequencing of Hevea brasiliensis for development of micro-satellite markers and construction of a genetic linkage map. DNA Res 18:471–482
Vinod KK, Suryakumar M, Chandrasekhar TR, Nazeer MA (2010) Temporal stability of growth and yield among Hevea genotypes introduced to a non-traditional rubber growing region of peninsu-lar India. Ann For Res 53(2):107–115
Vrba L, Munoz-Rodrıguez JL, Stampfer MR, Futscher B (2013) MiRNA gene promoters are frequent targets of aberrant DNA methylation in human breast cancer. PLoS one 8:1
Wang J, Sun Z, Zhou Y, Wang Q, Ye J, Chen Z, Liu J (2012) Screen-ing of a xylanase clone from a fosmid library of rumen micro-biota in Hu sheep. Anim Biotechnol 23(3):156–173
Zhang HB (2000) Construction and manipulation of large-insert bac-terial clone libraries. In: Manual. Department of Soil and Crop Sciences and Crop Biotechnology Center, Texas A&M University

933Mol Genet Genomics (2014) 289:921–933
1 3
Zhang HB, Zhao X, Ding X, Paterson AH, Wing RA (1995) Prepa-ration of megabase-size DNA from plant nuclei. Plant J 7(1):175–184
Zhang BH, Pan XP, Cox SB, Cobb GP, Anderson TA (2006) Evidence that miRNAs are different from other RNAs. Cell Mol Life Sci 63:246–254
Zhang L, Bao Z, Cheng J, Li H, Huang X, Wang S, Zhang C, Hu J (2007) Fosmid library construction and initial analysis of end sequences in Zhikong Scallop (Chlamys farreri). Mar Biotechnol 9:606–612
Zhang M, Zhang Y, Scheuring CF, Wu CC, Dong JJ, Zhang HB (2012) Preparation of megabase-sized DNA from a variety of organisms using the nuclei method for advanced genomics research. Nat Protoc 7:467–478
Zhao X, Li L (2013) Comparative analysis of microRNA promoters in Arabidopsis and rice. Genomics Proteomics Bioinform 11:56–60
Zhao JP, Jiang XL, Zhang BY, Su XH (2012) Involvement of micro-RNA-mediated gene expression regulation in the pathological development of stemcanker disease in Populus trichocarpa. PLoS One 7(9):e44968
Zhao X, Zhang H, Li L (2013) Identification and analysis of the prox-imal promoters of microRNA genes in Arabidopsis. Genomics 101:187–194
Zhou X, Ruan J, Zhang W, Wang G (2007) Characterization and iden-tification of microRNA core promoters in four model species. PLoS Comput Biol 3:3




















