
Rabies surveillance in bats in Northwestern State of São Paulo

A
cdSo
wata©
K
1
oRvpR
S
0d
Available online at www.sciencedirect.com
Virus Research 131 (2008) 33–46
Characterization of Rabies virus isolated from canids and identificationof the main wild canid host in Northeastern Brazil
Pedro Carnieli Jr. a,∗, Willian de Oliveira Fahl a, Juliana Galera Castilho a,Rafael de Novaes Oliveira a, Carla Isabel Macedo a, Ekaterina Durymanova a,
Rodrigo S.P. Jorge b, Ronaldo G. Morato b, Romualdo O. Spındola c,Lindenrberg M. Machado d, Jose E. Ungar de Sa e,
Maria Luiza Carrieri a, Ivanete Kotait a
a Instituto Pasteur de Sao Paulo, SP, Brazilb CENAP/IBAMA de Sao Paulo, SP, Brazil
c Fundacao Municipal de Saude, Teresina/Piauı, Brazild Laboratorio de Apoio Animal, Pernambuco, Brazil
e LACEN, Bahia, Brazil
Received 23 May 2007; received in revised form 9 August 2007; accepted 10 August 2007Available online 21 September 2007
bstract
The rabies cases in dogs and wild canids in Northeastern Brazil are a public and animal health problem. This paper describes the identities of theoding region of the N-gene of Rabies virus (RABV) isolated in canids from Northeastern Brazil. The genetic tree generated using the sequenceata described here divided the cluster BRAZILAN CANIDS into two subclusters (DOG-RELATED STRAINS and WILD CANID-RELATEDTRAINS) with identities greater than those already described. The two subclusters are sub-divided into geographic groups related to the originf the isolates, suggesting a long-standing ecological coexistence of the sequence types characteristic of the groups.
This article also analyzes the 513-nucleotide stretch of the mitochondrial DNA control region of rabies-positive canids from Northeastern Brazilith a view to identifying the main RABV host among them. Among the four species of wild canids found in the region, two (Cerdocyon thous
nd Pseudalopex vetulus) are frequently associated with rabies. Phylogenetic analysis of sequence data generated from mtDNA suggests that C.hous is the main wild canid host in the region. The results obtained in this study are in concordance with the zoology and ecology of wild canids,nd thus, help improve epidemiologic vigilance of rabies and allow a more targeted control of the disease.
2007 Elsevier B.V. All rights reserved.
epide
eTta
eywords: Rabies virus; Cerdocyon thous; Nucleoprotein; mtDNA; Molecular
. Introduction
The Rabies virus (RABV) belongs to the Lyssavirus genusf the Rhabdoviridae family. It is an enveloped, negative-senseNA virus with a linear genome approximately 12-kb long. The
irion consists of the following five proteins: nucleoprotein (N),hosphoprotein (P), matrix protein (M), glycoprotein (G) andNA-dependent RNA polymerase (L) (Fauquet et al., 2005).∗ Corresponding author at: Av. Paulista 393, Zip Code 01311-001, Sao Paulo,P, Brazil. Tel.: +55 11 32880088; fax: +55 11 32844924.
E-mail address: [email protected] (P. Carnieli Jr.).
gs2ra2
R
168-1702/$ – see front matter © 2007 Elsevier B.V. All rights reserved.oi:10.1016/j.virusres.2007.08.007
miology; Brazil
The L polymerase lacks a proofreading and post-replicationrror correction mechanism (Domingo and Holland, 1994).his characteristic, together with the evolutionary mechanisms
hat give rise to genetic variability within a population, suchs genetic drift, bottleneck and environmental selection, allowenetic variants of RABV to emerge and these are usually foundpatially and ecologically distributed (Kissi et al., 1999; Smith,002). The genetic variants of RABV responsible for enzooticabies may be spread to other sympatric susceptible domestic
nd wildlife species causing disease expansion (Kuzmin et al.,004).RABV reservoirs are those mammal species able to maintainabies virus variants enzootic within geographically discrete

3 s Res
aapttbgeelbl1aRuass
acgAF
bdcoArm11
tf2wwa1no(
tchic
iaow
raRoifv
msteoecnsiw
cAppgttft(htaM
aslNoscN
2
2
2
(were collected between 2002 and 2005 in Northeastern Brazil
4 P. Carnieli Jr. et al. / Viru
reas. In several instances these mammal species are welldapted to their biomes allowing them to occur in highopulation densities, which in turn make rabies enzootics sus-ainable over time. The terms used in the literature to describehese duets, reservoir-virus, are named ecotype or biotype. Aiotype may encompass several distinctive phylogeographicroups. These phylogeographic groups are regional virus lin-ages that, in the final analysis, demonstrate the molecularvolution of the virus (WHO, 2004), which is the result ofocal neutral variation and the geographic isolation imposedy mountain chains and large rivers. RABV geographical phy-ogroups are contained within this isolation (Bourhy et al.,999). RABV variants characteristic of terrestrial species thatre responsible for zoonotic rabies cycles are groupings ofABV lineages and represent subdivisions of the virus pop-lation in distinct habitats (Real et al., 2005). Nadin-Davis etl. (1999) suggest that the circulation of RABV lineages in hostubpopulations determines the virus’s phylogeographic relation-hips.
In Brazil, two rabies enzootics (urban and wild) are well char-cterized (Ito et al., 2001). The species responsible for theseycles are the dog and vampire bat, respectively, and the anti-enic variants harbored by these species have been identified asgV2 (dog) and AgV3 (Desmodus rotundus) (Diaz et al., 1994;avoretto et al., 2002).
In recent years there has been a marked decrease in the num-er of cases of rabies in dogs in Brazil. A total of 1746 cases ofog rabies were recorded in 1998 compared with a total of 67ases in 2006, a reduction brought about by the increased effortsf the National Program for the Control of Rabies in Domesticnimals. Between 1986 and 2006, a total of 758 cases of human
abies were recorded in Brazil. Dogs were responsible for trans-ission of 524 of these cases (69.1%), cats for 30 (4%), bats for
30 (17.1%), nonhuman primates for 14 (1.9%) and foxes for3 (1.7%) (SVS/MS, 2006).
Deforestation, which results in the destruction and fragmen-ation of habitats and roosts disturbances, is the main drivingorce wild animals into urban and periurban areas (Halpin et al.,007), a trend that in turn leads to increased contact betweenild animals and humans. The number of attacks on humans byild animals in Brazil in 1999 and 2005 notified to the authorities
nd for which the victims received medical attention were 35 and6334, respectively. Bats accounted for 29 and 11811 attacks,onhuman primates for 5 and 3360, and foxes and other speciesf lesser epidemiologic importance for 1 and 1163, respectivelySVS/MS, 2006).
With regard to Northeastern Brazil, which was the focus ofhis study, a direct relationship can be established between rabiesases in dog biotypes and dog-related biotypes, which we willenceforth refer to as wild canid biotypes. This is the only regionn Brazil in which RABV has been isolated recently in wildanids (SVS/MS, 2006).
Carnieli et al. (2006), studying rabies epidemiology in canids
n Northeastern Brazil, identified two rabies enzootic in thesenimals: one among domestic animals (dog biotype) and thether among wild canids (wild canid biotype), although thereas overlap between the two enzootic. Based on epidemiologici(M(
earch 131 (2008) 33–46
abies surveillance in the region (SVS/MS, 2006) and zoologicalnd ecological studies related to both species (Wilson andeeder, 1993), it was suggested that among the six speciesf wild canids in Brazil Cerdocyon thous (crab-eating fox)s the main wild RABV host species in Northeastern Brazil,ollowed by Pseudalopex vetulus (hoary fox) (=Dusicyonetulus).
Identification of wild canid species involved in rabies trans-ission in Northeastern Brazil has been carried out in a
ubjective, and thus, controversial, manner. Identification ofhe two species of wild canids cited so far, namely, the crab-ating fox, C. thous, and the hoary fox, P. vetulus, was basedn morphological characteristics and geographic records. How-ver, morphological similarities between the two species canonfuse researchers inexperienced in the identification of car-ivores. Additionally, the overlap in the distribution of the twouspected species may lead to erroneous identification when thiss based on the geographic area where the sampled individualas found.The crab-eating fox C. thous is a medium-sized (5–7 kg)
anid with generalist habits and widespread distribution in Southmerica. It is widely distributed in Brazil and only has a limitedresence in the Amazon forest. This animal has rostrum long andointed, head relatively short and narrow, pelage generally darkrey to black along dorsum down to midline; midline to ven-rum including legs grey or black; neck and underparts creamo buff white. The area in which the hoary fox (P. vetulus) isound is smaller than that of C. thous, as it is only found inhe Pantanal and the Brazilian Cerrado. P. vetulus is smaller2.5–4 kg) and less robust than C. thous, has more specializedabits, pelage woolly not bristly; rostrum length shorter relativeo rostrum width in C. thous with relatively large ears (Eisembergnd Redford, 1999; Emmons and Feer, 1999; Courtenay andaffei, 2004).The aim of this study was to investigate rabies epidemiology
mong domestic and wild canids in Northeastern Brazil. Thistudy compares the genetic identity of the RABV N-gene circu-ating in dog and wild canid biotypes by analyzing the complete-gene coding region and describes the genetic identificationf the wild canid species involved in rabies epidemiology byequencing and analysis of the hypervariable region of mito-hondrial DNA extracted from rabies-positive samples fromortheastern Brazil.
. Materials and methods
.1. RABV nucleoprotein gene
.1.1. SamplesA total of 52 brain samples were used: dog (n = 17), cat
n = 04), wild canid (n = 30) and human (n = 01) (Table 1). These
n several towns in the states of Paraıba (PB), PernambucoPE), Bahia (BA), Piauı (PI), Alagoas (AL), Sergipe (SE) and
aranhao (MA) and in the state of Para (PA) in Northern BrazilFig. 1).
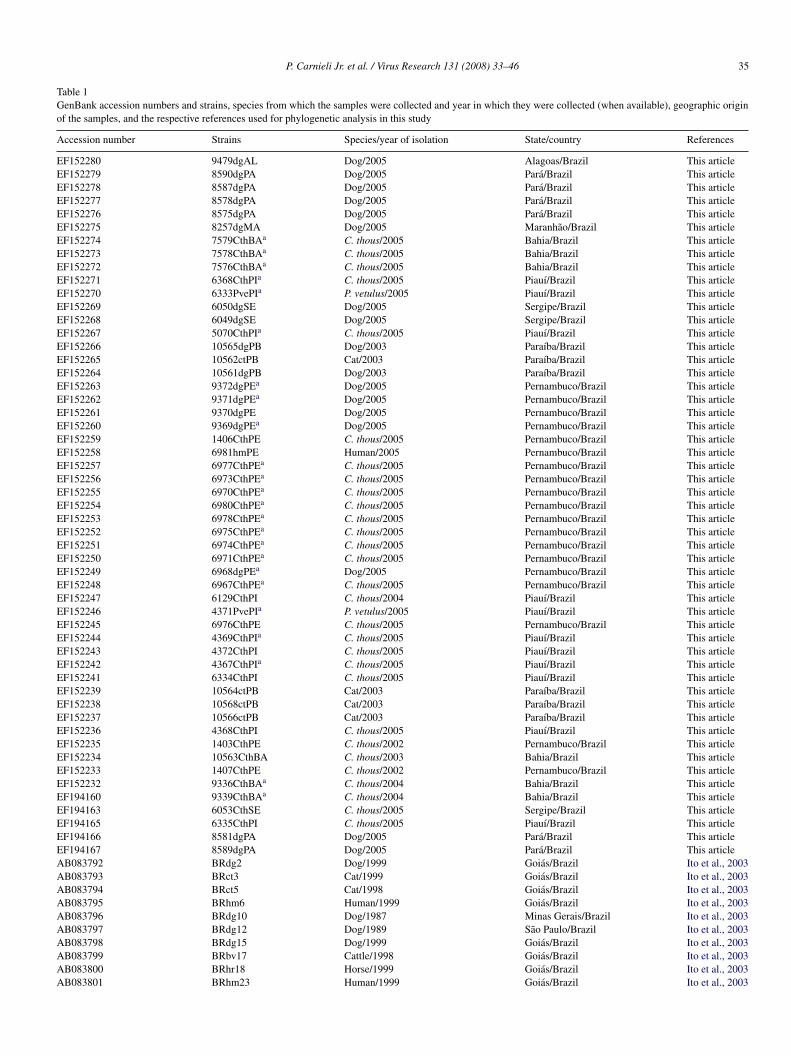
P. Carnieli Jr. et al. / Virus Research 131 (2008) 33–46 35
Table 1GenBank accession numbers and strains, species from which the samples were collected and year in which they were collected (when available), geographic originof the samples, and the respective references used for phylogenetic analysis in this study
Accession number Strains Species/year of isolation State/country References
EF152280 9479dgAL Dog/2005 Alagoas/Brazil This articleEF152279 8590dgPA Dog/2005 Para/Brazil This articleEF152278 8587dgPA Dog/2005 Para/Brazil This articleEF152277 8578dgPA Dog/2005 Para/Brazil This articleEF152276 8575dgPA Dog/2005 Para/Brazil This articleEF152275 8257dgMA Dog/2005 Maranhao/Brazil This articleEF152274 7579CthBAa C. thous/2005 Bahia/Brazil This articleEF152273 7578CthBAa C. thous/2005 Bahia/Brazil This articleEF152272 7576CthBAa C. thous/2005 Bahia/Brazil This articleEF152271 6368CthPIa C. thous/2005 Piauı/Brazil This articleEF152270 6333PvePIa P. vetulus/2005 Piauı/Brazil This articleEF152269 6050dgSE Dog/2005 Sergipe/Brazil This articleEF152268 6049dgSE Dog/2005 Sergipe/Brazil This articleEF152267 5070CthPIa C. thous/2005 Piauı/Brazil This articleEF152266 10565dgPB Dog/2003 Paraıba/Brazil This articleEF152265 10562ctPB Cat/2003 Paraıba/Brazil This articleEF152264 10561dgPB Dog/2003 Paraıba/Brazil This articleEF152263 9372dgPEa Dog/2005 Pernambuco/Brazil This articleEF152262 9371dgPEa Dog/2005 Pernambuco/Brazil This articleEF152261 9370dgPE Dog/2005 Pernambuco/Brazil This articleEF152260 9369dgPEa Dog/2005 Pernambuco/Brazil This articleEF152259 1406CthPE C. thous/2005 Pernambuco/Brazil This articleEF152258 6981hmPE Human/2005 Pernambuco/Brazil This articleEF152257 6977CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152256 6973CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152255 6970CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152254 6980CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152253 6978CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152252 6975CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152251 6974CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152250 6971CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152249 6968dgPEa Dog/2005 Pernambuco/Brazil This articleEF152248 6967CthPEa C. thous/2005 Pernambuco/Brazil This articleEF152247 6129CthPI C. thous/2004 Piauı/Brazil This articleEF152246 4371PvePIa P. vetulus/2005 Piauı/Brazil This articleEF152245 6976CthPE C. thous/2005 Pernambuco/Brazil This articleEF152244 4369CthPIa C. thous/2005 Piauı/Brazil This articleEF152243 4372CthPI C. thous/2005 Piauı/Brazil This articleEF152242 4367CthPIa C. thous/2005 Piauı/Brazil This articleEF152241 6334CthPI C. thous/2005 Piauı/Brazil This articleEF152239 10564ctPB Cat/2003 Paraıba/Brazil This articleEF152238 10568ctPB Cat/2003 Paraıba/Brazil This articleEF152237 10566ctPB Cat/2003 Paraıba/Brazil This articleEF152236 4368CthPI C. thous/2005 Piauı/Brazil This articleEF152235 1403CthPE C. thous/2002 Pernambuco/Brazil This articleEF152234 10563CthBA C. thous/2003 Bahia/Brazil This articleEF152233 1407CthPE C. thous/2002 Pernambuco/Brazil This articleEF152232 9336CthBAa C. thous/2004 Bahia/Brazil This articleEF194160 9339CthBAa C. thous/2004 Bahia/Brazil This articleEF194163 6053CthSE C. thous/2005 Sergipe/Brazil This articleEF194165 6335CthPI C. thous/2005 Piauı/Brazil This articleEF194166 8581dgPA Dog/2005 Para/Brazil This articleEF194167 8589dgPA Dog/2005 Para/Brazil This articleAB083792 BRdg2 Dog/1999 Goias/Brazil Ito et al., 2003AB083793 BRct3 Cat/1999 Goias/Brazil Ito et al., 2003AB083794 BRct5 Cat/1998 Goias/Brazil Ito et al., 2003AB083795 BRhm6 Human/1999 Goias/Brazil Ito et al., 2003AB083796 BRdg10 Dog/1987 Minas Gerais/Brazil Ito et al., 2003AB083797 BRdg12 Dog/1989 Sao Paulo/Brazil Ito et al., 2003AB083798 BRdg15 Dog/1999 Goias/Brazil Ito et al., 2003AB083799 BRbv17 Cattle/1998 Goias/Brazil Ito et al., 2003AB083800 BRhr18 Horse/1999 Goias/Brazil Ito et al., 2003AB083801 BRhm23 Human/1999 Goias/Brazil Ito et al., 2003

36 P. Carnieli Jr. et al. / Virus Research 131 (2008) 33–46
Table 1 (Continued )
U22477 9126MEX Dog/1991 Unknown/Mexico Ito et al., 2003U22478 9001GUY Dog/1990 Unknown/French Guiana Ito et al., 2003AB083806 BRvmbt33 Vampire bat/unknown Sao Paulo/Brazil Ito et al., 2003AB083807 BRvmbt34 Vampire bat/1998 Sao Paulo/Brazil Ito et al., 2003AB083816 BRvmbt47 Vampire bat/unknown Unknown/Brazil Ito et al., 2003DQ837486 YO6867 Dog/2005 Unknown/Israel UnpublishedDQ837485 YA6530 Fox/1996 Unknown/Israel UnpublishedDQ837484 VS3370 Fox/1995 Unknown/Israel UnpublishedDQ837482 TL8852 Jackal/2004 Unknown/Israel UnpublishedDQ837479 T6 Dog/2000 Unknown/Turkey UnpublishedDQ837473 SP1662 Wolf/1997 Unknown/Israel UnpublishedDQ837454 OD6220 Wolf/2005 Unknown/Israel UnpublishedDQ837418 HR1510 Fox/2004 Unknown/Israel UnpublishedL20675 91RABN0783 Antarctic fox/2001 Ontario/Canada Nadin-Davis et al., 1993NC 006429 GI:55770806 Mokola virus Paris/France Le Mercier et al., 1997AB207884 BRPfx6 Fox/2005 Paraıba/Brazil UnpublishedAB206419 BRPbt6 Fox/2005 Paraıba/Brazil UnpublishedAB206418 BRPbt5 Fox/2005 Paraıba/Brazil UnpublishedAB206417 BRPbt4 Fox/2005 Paraıba/Brazil UnpublishedAB206416 BRPpt3 Fox/2005 Paraıba/Brazil UnpublishedAB206415 BRPbt2 Fox/2005 Paraıba/Brazil UnpublishedAB206414 BRPbt1 Fox/2005 Paraıba/Brazil UnpublishedAB206413 BRPfx8 Fox/2005 Paraıba/Brazil UnpublishedAB206412 BRPfx7 Fox/2005 Paraıba/Brazil UnpublishedAB206411 BRPfx5 Fox/2005 Paraıba/Brazil UnpublishedAB206410 BRPfx4 Fox/2005 Paraıba/Brazil UnpublishedAB206409 BRPfx3 Fox/2005 Paraıba/Brazil UnpublishedAB206408 BRPfx2 Fox/2005 Paraıba/Brazil UnpublishedAB206407 BRPfx1 Fox/2005 Paraıba/Brazil UnpublishedDQ447958 brcth5692 C. thous/unknown Ceara/Brazil Favoretto et al., 2006DQ447957 brcth5697 C. thous/unknown Ceara/Brazil Favoretto et al., 2006DQ447956 brcth5695 C. thous/unknown Ceara/Brazil Favoretto et al., 2006DQ447951 brcth5361 C. thous/unknown Ceara/Brazil Favoretto et al., 2006DQ447948 brcth4122 C. thous/unknown Ceara/Brazil Favoretto et al., 2006M13215.1 Rabies virus, strain PV France Tordo et al., 1988DQ42112.1 Rabies virus, strain CVS France Tordo et al., 1986
a
2i
Mt
2
neuoid
2
wir(
((wsuCtoAgsrstBsn
Specimens genetically characterized at the mitochondrial locus.
.1.2. Fluorescent antibody test (FAT) and mousenoculation test (MIT)
All samples were diagnosed by FAT (Dean et al., 1996) andIT (Koprowski, 1996). The standard RABV used in all the
ests was the “Challenge Virus Standard” (CVS-31).
.1.3. Antigenic analysisA panel of eight monoclonal antibodies against the RABV
ucleoprotein (N-Mabs), kindly provided by the Centers for Dis-ase Control and Prevention (CDC), Atlanta, GA, USA, wassed for the antigenic characterization. An indirect immunoflu-rescence assay using prints of brain material taken from micenfected with the samples being studied was used, as previouslyescribed by Diaz et al. (1994).
.1.4. RT-PCR, DNA sequencing and phylogenetic analysisTotal RNA from the 52 brain samples was extracted
ith Trizol® (Invitrogen) according to the manufacturer’snstructions. Reverse transcriptase (RT) and polymerase chaineaction (PCR) were performed using sense primer 21GATGTAACACCTCTACAATG) and antisense primer 304
tr(w
TTGACGAAGATCTTGCTCAT), as previously describedOrciari et al., 2001). Amplified DNA fragments were purifiedith GFX PCR DNA and Gel Band Purification Kit (Amer-
ham BiosciencesTM) and submitted to sequencing reactionssing sense and antisense primers with BigDye Terminator v3.1ycle Sequencing Kit (Amersham BiosciencesTM) according
o the manufacturer’s instructions. The sequencing was carriedut in an Applied Biosystems 3130 DNA automated sequencer.
1393-nucleotide region corresponding to the nucleoproteinene located between nucleotides (nt) 89 and 1482 of the PVtrain (accession number M13215.1) was analyzed. Data fromaw sequencing were edited using CHROMAS software (ver-ion 2.24 Copyright© 1998–2004 Technelysium Pty Ltd.), andhe final sequences were aligned with sequences present in Gen-ank (Table 1) by the CLUSTAL/W method using BioEdit
oftware (Hall, 1999). The alignments were used to buildeighbor-joining distance-based DNA phylogenetic trees with
he Kimura-2 parameter correction model and 1000 bootstrapepetitions for statistical support using the Mega 2.1 programKumar et al., 2001). The nucleotide and amino acid identitiesere calculated using BioEdit software.
P. Carnieli Jr. et al. / Virus Research 131 (2008) 33–46 37
Fig. 1. Map of Brazil and its Biomes (IBGE, 2007). The abbreviations SP, MG, GO, TO, MT, MS, AM, PA, BA, PI, MA, CE, RN, PB, PE, AL and SE refer tothe states of Sao Paulo, Minas Gerais, Goias, Tocantins, Mato Grosso, Mato Grosso do Sul, Amazonas, Para, Bahia, Piauı, Maranhao, Ceara, Rio Grande do Norte,Paraıba, Pernambuco, Alagoas and Sergipe, which are referred to in the main text. The Northeast of Brazil is shown inside the dotted square. Rabies cases amongc ter. C( cyon tt n.org)
2m
2
cstaBaa
2
cm(CTpw10ttt1aacaa
osdtt((fp
3
3
3
F
3
tl46MnC
3
anids in the Northeastern region in the period 2004–2005 are shown on the cenSVS/MS, 2006). Maps of the current distribution of the crab-eating fox, Cerdohe right. Based in Canid Specialist Group and Mammal Assessment (www.iuc
.2. Identification of wild canid species usingitochondrial DNA
.2.1. SamplesTo identify the species of wild canids involved in the
anine rabies cycles in Northeastern Brazil, rabies-positive brainamples from wild canids and dogs were used (29 and 05, respec-ively) (Table 2). These samples were collected between 2004nd 2006 in a number of towns in the states of Pernambuco,ahia, Piauı and Alagoas. The RABV was identified, isolatednd antigenically typed as described in Sections 2.1.2 and 2.1.3bove.
.2.2. PCR, DNA sequencing and phylogenetic analysisThe 5′ portion of the Mitochondrial DNA (mtDNA)
ontrol region containing the first hypervariable seg-ent (HVS-I) was amplified using primers MTLPRO2
5′-CACTATCAGCACCCAAAGCTG) and CCR-DR1 (5′-TGTGACCATTGACTGAATAGC) (Ward et al., 1991).otal DNA was extracted from the samples using a standardhenol/chloroform protocol (Sambrook et al., 1989). PCRas carried out in an 80-�L reaction mixture containing0 �L 10× PCR buffer, 1.5 mM MgCl2, 0.2 �M dNTPs,.2 �M of each primer, 0.75 unit Taq polymerase and 5 �L ofemplate dilutions of the extracted DNA using an EppendorfTM
hermocycler. Thermocycling conditions for amplification ofhe control-region (HVS-I segment) DNA were as follows:0 cycles (touchdown amplification protocol), each including45-s denaturing step at 94 ◦C, 45-s annealing at 60–51 ◦C,
nd a 1.5-min extension at 72 ◦C; this was followed by 30ycles of 45-s denaturing at 94 ◦C, 30-s annealing at 50 ◦Cnd a 1.5-min extension at 72 ◦C to produce amplicons withn average length of 600 nt. The PCR protocol was based
se(
ases of rabies in dogs are shown in (A) and cases of rabies in wild canids in (B)hous (I) and the hoary fox, Pseudalopex vetulus (II) in Northeastern Brazil, on.
n Tchaicka et al. (2007), with minor modifications. DNAequencing and phylogenetic analysis were performed asescribed in Section 2.1.3. Canid species were identified usinghe following sequences: three from C. thous (C th bCth201, Ch bCth205, C th bCth51); two from Pseudalopex gymnocercusP gym bPgy37, P gym bPgy18); and three from P. vetulusP vet bPve02, P vet bPve032, P vet bPve08). In addition,our sequences from dogs were used as outgroups in thehylogenetic analysis (Table 2).
. Results
.1. RABV nucleoprotein gene
.1.1. FAT, MIT and RT-PCRAll the 52 RABV isolates used in this study were positive for
AT, MIT and RT-PCR.
.1.2. Antigenic typingThe RABV isolates in this study were typed and belong
o antigenic variant 2 (AgV2), with the exception of iso-ates 7576CthBA, 7578CthBA, 9336CthBA, 9339CthBA,369CthPI, 1406CthPE, 1407CthPE, 6974CthPE, 6977CthPE,968dgPE and 6981hmPE, which did not react to the C1 N-ab from the panel of monoclonal antibodies against the RABV
ucleoprotein. The group of isolates that did not react to N-Mab1 are here referred to as N-Mab C1 negatives (N-Mab C1−).
.1.3. Genetic analysis of the N-gene
The genetic tree in Fig. 2 was produced from the results ofequencing of the RABV N-gene isolated in canids in North-astern Brazil and other sequence data retrieved from GenBankTable 1). The topology of the genetic tree can be divided into
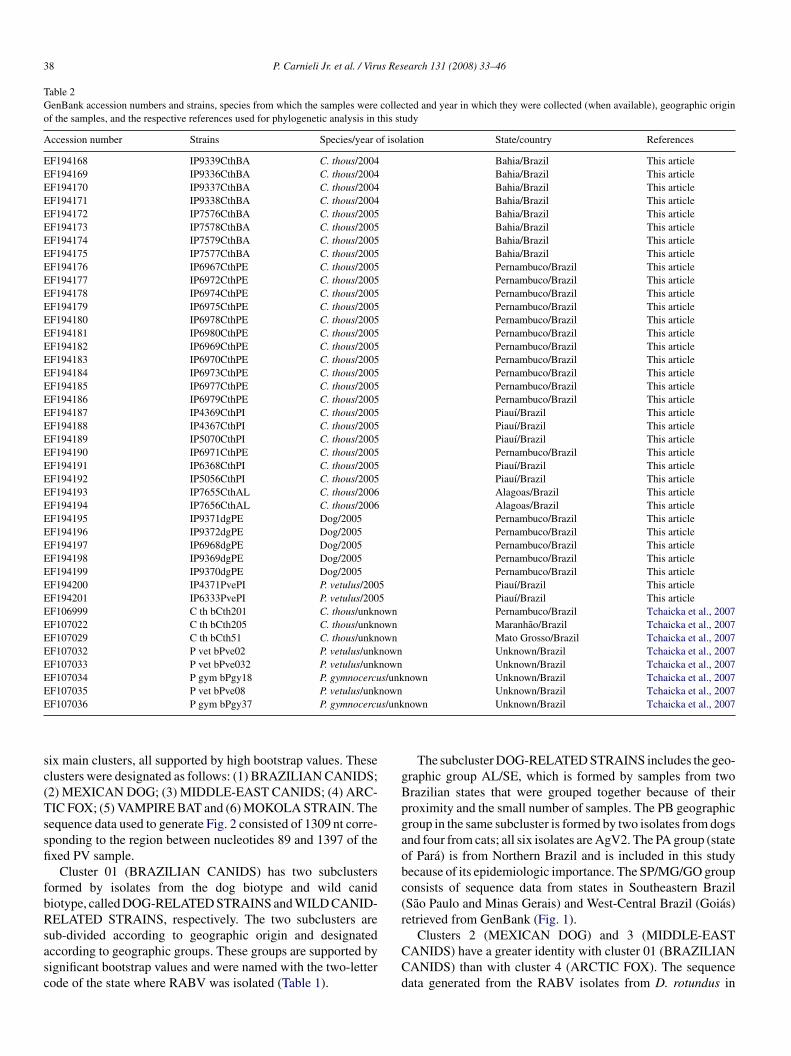
38 P. Carnieli Jr. et al. / Virus Research 131 (2008) 33–46
Table 2GenBank accession numbers and strains, species from which the samples were collected and year in which they were collected (when available), geographic originof the samples, and the respective references used for phylogenetic analysis in this study
Accession number Strains Species/year of isolation State/country References
EF194168 IP9339CthBA C. thous/2004 Bahia/Brazil This articleEF194169 IP9336CthBA C. thous/2004 Bahia/Brazil This articleEF194170 IP9337CthBA C. thous/2004 Bahia/Brazil This articleEF194171 IP9338CthBA C. thous/2004 Bahia/Brazil This articleEF194172 IP7576CthBA C. thous/2005 Bahia/Brazil This articleEF194173 IP7578CthBA C. thous/2005 Bahia/Brazil This articleEF194174 IP7579CthBA C. thous/2005 Bahia/Brazil This articleEF194175 IP7577CthBA C. thous/2005 Bahia/Brazil This articleEF194176 IP6967CthPE C. thous/2005 Pernambuco/Brazil This articleEF194177 IP6972CthPE C. thous/2005 Pernambuco/Brazil This articleEF194178 IP6974CthPE C. thous/2005 Pernambuco/Brazil This articleEF194179 IP6975CthPE C. thous/2005 Pernambuco/Brazil This articleEF194180 IP6978CthPE C. thous/2005 Pernambuco/Brazil This articleEF194181 IP6980CthPE C. thous/2005 Pernambuco/Brazil This articleEF194182 IP6969CthPE C. thous/2005 Pernambuco/Brazil This articleEF194183 IP6970CthPE C. thous/2005 Pernambuco/Brazil This articleEF194184 IP6973CthPE C. thous/2005 Pernambuco/Brazil This articleEF194185 IP6977CthPE C. thous/2005 Pernambuco/Brazil This articleEF194186 IP6979CthPE C. thous/2005 Pernambuco/Brazil This articleEF194187 IP4369CthPI C. thous/2005 Piauı/Brazil This articleEF194188 IP4367CthPI C. thous/2005 Piauı/Brazil This articleEF194189 IP5070CthPI C. thous/2005 Piauı/Brazil This articleEF194190 IP6971CthPE C. thous/2005 Pernambuco/Brazil This articleEF194191 IP6368CthPI C. thous/2005 Piauı/Brazil This articleEF194192 IP5056CthPI C. thous/2005 Piauı/Brazil This articleEF194193 IP7655CthAL C. thous/2006 Alagoas/Brazil This articleEF194194 IP7656CthAL C. thous/2006 Alagoas/Brazil This articleEF194195 IP9371dgPE Dog/2005 Pernambuco/Brazil This articleEF194196 IP9372dgPE Dog/2005 Pernambuco/Brazil This articleEF194197 IP6968dgPE Dog/2005 Pernambuco/Brazil This articleEF194198 IP9369dgPE Dog/2005 Pernambuco/Brazil This articleEF194199 IP9370dgPE Dog/2005 Pernambuco/Brazil This articleEF194200 IP4371PvePI P. vetulus/2005 Piauı/Brazil This articleEF194201 IP6333PvePI P. vetulus/2005 Piauı/Brazil This articleEF106999 C th bCth201 C. thous/unknown Pernambuco/Brazil Tchaicka et al., 2007EF107022 C th bCth205 C. thous/unknown Maranhao/Brazil Tchaicka et al., 2007EF107029 C th bCth51 C. thous/unknown Mato Grosso/Brazil Tchaicka et al., 2007EF107032 P vet bPve02 P. vetulus/unknown Unknown/Brazil Tchaicka et al., 2007EF107033 P vet bPve032 P. vetulus/unknown Unknown/Brazil Tchaicka et al., 2007EF107034 P gym bPgy18 P. gymnocercus/unknown Unknown/Brazil Tchaicka et al., 2007EF107035 P vet bPve08 P. vetulus/unknown Unknown/Brazil Tchaicka et al., 2007E s/unk
sc(Tssfi
fbRsasc
gBpgaobc(r
F107036 P gym bPgy37 P. gymnocercu
ix main clusters, all supported by high bootstrap values. Theselusters were designated as follows: (1) BRAZILIAN CANIDS;2) MEXICAN DOG; (3) MIDDLE-EAST CANIDS; (4) ARC-IC FOX; (5) VAMPIRE BAT and (6) MOKOLA STRAIN. Theequence data used to generate Fig. 2 consisted of 1309 nt corre-ponding to the region between nucleotides 89 and 1397 of thexed PV sample.
Cluster 01 (BRAZILIAN CANIDS) has two subclustersormed by isolates from the dog biotype and wild canidiotype, called DOG-RELATED STRAINS and WILD CANID-ELATED STRAINS, respectively. The two subclusters are
ub-divided according to geographic origin and designatedccording to geographic groups. These groups are supported byignificant bootstrap values and were named with the two-letterode of the state where RABV was isolated (Table 1).
CCd
nown Unknown/Brazil Tchaicka et al., 2007
The subcluster DOG-RELATED STRAINS includes the geo-raphic group AL/SE, which is formed by samples from tworazilian states that were grouped together because of theirroximity and the small number of samples. The PB geographicroup in the same subcluster is formed by two isolates from dogsnd four from cats; all six isolates are AgV2. The PA group (statef Para) is from Northern Brazil and is included in this studyecause of its epidemiologic importance. The SP/MG/GO grouponsists of sequence data from states in Southeastern BrazilSao Paulo and Minas Gerais) and West-Central Brazil (Goias)etrieved from GenBank (Fig. 1).
Clusters 2 (MEXICAN DOG) and 3 (MIDDLE-EASTANIDS) have a greater identity with cluster 01 (BRAZILIANANIDS) than with cluster 4 (ARCTIC FOX). The sequenceata generated from the RABV isolates from D. rotundus in
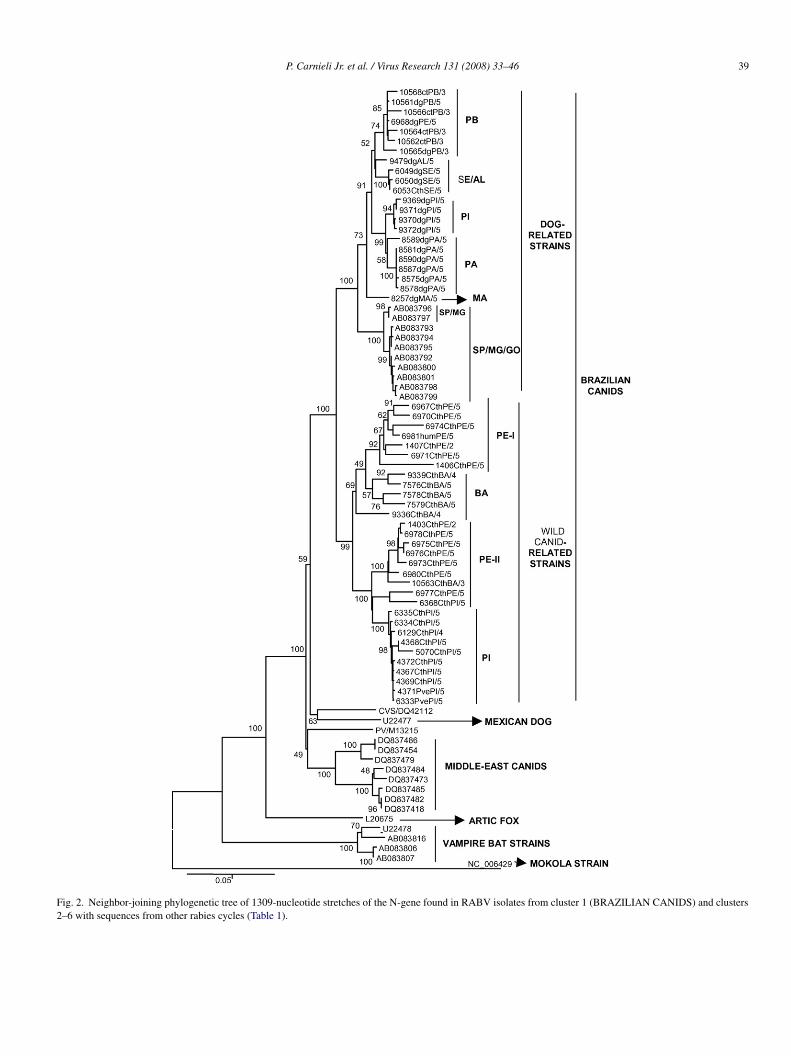
P. Carnieli Jr. et al. / Virus Research 131 (2008) 33–46 39
Fig. 2. Neighbor-joining phylogenetic tree of 1309-nucleotide stretches of the N-gene found in RABV isolates from cluster 1 (BRAZILIAN CANIDS) and clusters2–6 with sequences from other rabies cycles (Table 1).
40 P. Carnieli Jr. et al. / Virus Research 131 (2008) 33–46
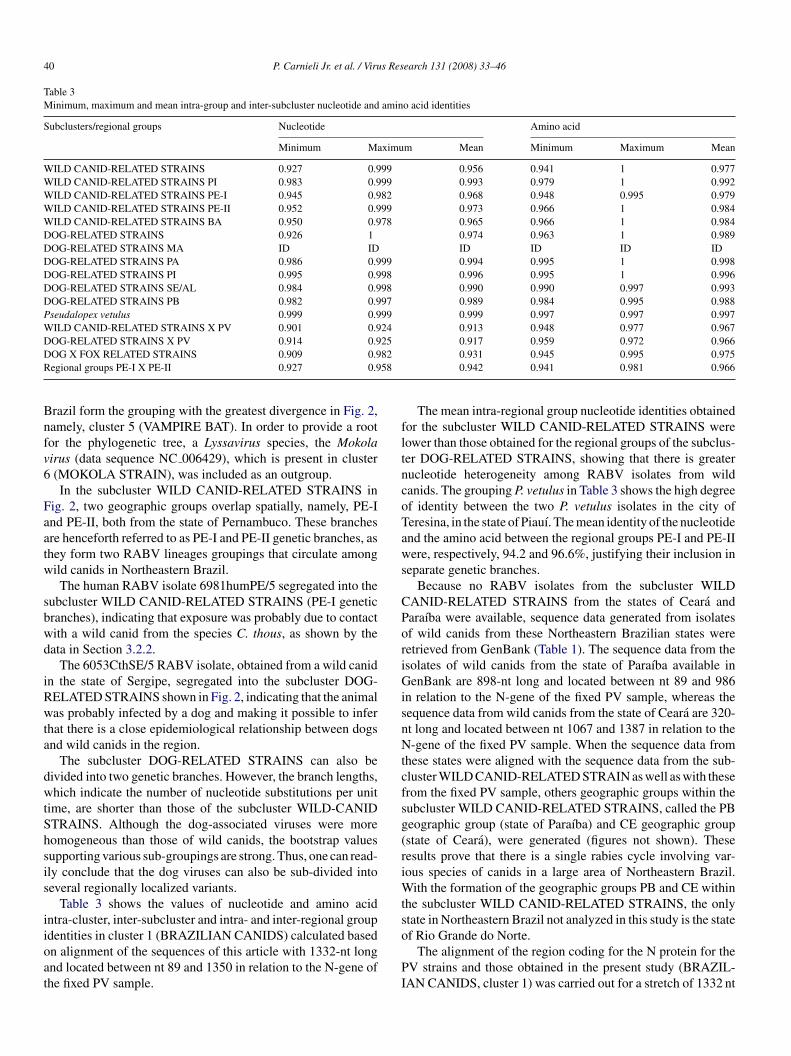
Table 3Minimum, maximum and mean intra-group and inter-subcluster nucleotide and amino acid identities
Subclusters/regional groups Nucleotide Amino acid
Minimum Maximum Mean Minimum Maximum Mean
WILD CANID-RELATED STRAINS 0.927 0.999 0.956 0.941 1 0.977WILD CANID-RELATED STRAINS PI 0.983 0.999 0.993 0.979 1 0.992WILD CANID-RELATED STRAINS PE-I 0.945 0.982 0.968 0.948 0.995 0.979WILD CANID-RELATED STRAINS PE-II 0.952 0.999 0.973 0.966 1 0.984WILD CANID-RELATED STRAINS BA 0.950 0.978 0.965 0.966 1 0.984DOG-RELATED STRAINS 0.926 1 0.974 0.963 1 0.989DOG-RELATED STRAINS MA ID ID ID ID ID IDDOG-RELATED STRAINS PA 0.986 0.999 0.994 0.995 1 0.998DOG-RELATED STRAINS PI 0.995 0.998 0.996 0.995 1 0.996DOG-RELATED STRAINS SE/AL 0.984 0.998 0.990 0.990 0.997 0.993DOG-RELATED STRAINS PB 0.982 0.997 0.989 0.984 0.995 0.988Pseudalopex vetulus 0.999 0.999 0.999 0.997 0.997 0.997WILD CANID-RELATED STRAINS X PV 0.901 0.924 0.913 0.948 0.977 0.967DOG-RELATED STRAINS X PV 0.914 0.925 0.917 0.959 0.972 0.966D 82R 58
Bnfv6
Faatw
sbwd
iRwta
dwtShsis
iioat
fltncoTaws
CPoriGisnNtcfsg(riWts
OG X FOX RELATED STRAINS 0.909 0.9egional groups PE-I X PE-II 0.927 0.9
razil form the grouping with the greatest divergence in Fig. 2,amely, cluster 5 (VAMPIRE BAT). In order to provide a rootor the phylogenetic tree, a Lyssavirus species, the Mokolairus (data sequence NC 006429), which is present in cluster(MOKOLA STRAIN), was included as an outgroup.In the subcluster WILD CANID-RELATED STRAINS in
ig. 2, two geographic groups overlap spatially, namely, PE-Ind PE-II, both from the state of Pernambuco. These branchesre henceforth referred to as PE-I and PE-II genetic branches, ashey form two RABV lineages groupings that circulate amongild canids in Northeastern Brazil.The human RABV isolate 6981humPE/5 segregated into the
ubcluster WILD CANID-RELATED STRAINS (PE-I geneticranches), indicating that exposure was probably due to contactith a wild canid from the species C. thous, as shown by theata in Section 3.2.2.
The 6053CthSE/5 RABV isolate, obtained from a wild canidn the state of Sergipe, segregated into the subcluster DOG-ELATED STRAINS shown in Fig. 2, indicating that the animalas probably infected by a dog and making it possible to infer
hat there is a close epidemiological relationship between dogsnd wild canids in the region.
The subcluster DOG-RELATED STRAINS can also beivided into two genetic branches. However, the branch lengths,hich indicate the number of nucleotide substitutions per unit
ime, are shorter than those of the subcluster WILD-CANIDTRAINS. Although the dog-associated viruses were moreomogeneous than those of wild canids, the bootstrap valuesupporting various sub-groupings are strong. Thus, one can read-ly conclude that the dog viruses can also be sub-divided intoeveral regionally localized variants.
Table 3 shows the values of nucleotide and amino acidntra-cluster, inter-subcluster and intra- and inter-regional group
dentities in cluster 1 (BRAZILIAN CANIDS) calculated basedn alignment of the sequences of this article with 1332-nt longnd located between nt 89 and 1350 in relation to the N-gene ofhe fixed PV sample.o
PI
0.931 0.945 0.995 0.9750.942 0.941 0.981 0.966
The mean intra-regional group nucleotide identities obtainedor the subcluster WILD CANID-RELATED STRAINS wereower than those obtained for the regional groups of the subclus-er DOG-RELATED STRAINS, showing that there is greaterucleotide heterogeneity among RABV isolates from wildanids. The grouping P. vetulus in Table 3 shows the high degreef identity between the two P. vetulus isolates in the city oferesina, in the state of Piauı. The mean identity of the nucleotidend the amino acid between the regional groups PE-I and PE-IIere, respectively, 94.2 and 96.6%, justifying their inclusion in
eparate genetic branches.Because no RABV isolates from the subcluster WILD
ANID-RELATED STRAINS from the states of Ceara andaraıba were available, sequence data generated from isolatesf wild canids from these Northeastern Brazilian states wereetrieved from GenBank (Table 1). The sequence data from thesolates of wild canids from the state of Paraıba available inenBank are 898-nt long and located between nt 89 and 986
n relation to the N-gene of the fixed PV sample, whereas theequence data from wild canids from the state of Ceara are 320-t long and located between nt 1067 and 1387 in relation to the-gene of the fixed PV sample. When the sequence data from
hese states were aligned with the sequence data from the sub-luster WILD CANID-RELATED STRAIN as well as with theserom the fixed PV sample, others geographic groups within theubcluster WILD CANID-RELATED STRAINS, called the PBeographic group (state of Paraıba) and CE geographic groupstate of Ceara), were generated (figures not shown). Theseesults prove that there is a single rabies cycle involving var-ous species of canids in a large area of Northeastern Brazil.
ith the formation of the geographic groups PB and CE withinhe subcluster WILD CANID-RELATED STRAINS, the onlytate in Northeastern Brazil not analyzed in this study is the state
f Rio Grande do Norte.The alignment of the region coding for the N protein for theV strains and those obtained in the present study (BRAZIL-AN CANIDS, cluster 1) was carried out for a stretch of 1332 nt

P. Carnieli Jr. et al. / Virus Research 131 (2008) 33–46 41
F neticw ure sD
bssaaRatL1cbCofitmw
3
3
si4ragt(
3
wgfftf
4
asotrrtdfpoe
ligthPc2m2
reidateDSh
ig. 3. Comparison of the predicted N sequences of some representative geas performed on the PV nucleoprotein of 450 amino acids; however, the figOG-RELATED STRAINS.
etween nt 89 and 1420 in relation to the N-gene of the fixed PVtrain. The putative amino acids alignment, one could observeome regions that characterize the DOG-RELATED STRAINSnd WILD CANID-RELATED STRAINS subclusters, as wells the geographic groups PE-I and PE-II (Fig. 3). The DOG-ELATED STRAINS subcluster has Cysteine (C), Isoleucine (I)nd Threonine (T) redisues at positions 08, 119 and 433, respec-ively. Regarding geographic group PE-I, one can find Serine (S),eucine (L), Isoleucine and Lysine (K) residues at positions 119,26, 410 and 418, respectively. The geographic group PE-II isharacterized by an Argenine (R) at position 35. It was not possi-le to identify an amino acid area characteristic of strains N-Mab1−. These strains are dispersed among the geographic clustersf the WILD CANID-RELATED STRAINS group. Only therst 18 nucleotides, related to the 6 first amino acids of the puta-
ive N proteins studied herein were not sequenced and in this areight be an explanation to the lack of reactivity of such strainsith N-Mab C1.
.2. Identification of wild canids using mitochondrial DNA
.2.1. FAT, MIT, RT-PCR and antigenic typingAll the 34 rabies-positive isolates used to identify wild canid
pecies (Table 2) were positive by FAT, MIT and RT-PCR. Thesolates 7576CthBA, 7578CthBA, 9336CthBA, 9339CthBA,369CthPI, 6974CthPE, 6977CthPE and 6968dgPE did noteact against N-Mab C1 from the panel of monoclonalntibodies against the RABV viral nucleoprotein and wererouped together as N-Mab C1−. Antigenic typing showedhat all the other samples belonged to antigenic variant 2AgV2).
.2.2. Phylogenetic analysisA 513-nt sequence of the mtDNA control region (CR)
as obtained from the 34 isolates (Table 2). The genetic treeenerated with these sequences (Fig. 4) can be divided into
our clusters: C. thous, P. gymnocercus, P. vetulus and Canisamiliaris. The cluster C. thous is made up of 27 sequences,he cluster P. vetulus of two and the cluster C. familiaris ofour.ldtp
groups that circulate in the Northeastern Brazil. The amino acid alignmenthows only stretches representatives of PE-I and PE-II groups and subcluster
. Discussion
Rabies control depends on governmental programs suchs mass vaccination of domestic animals and epidemiologicalurveillance. However, there is also a need for molecular typingf RABV to help understand the dynamics of maintenance ofhe virus in the wild, and thus, determine its phylogeographicelationships (WHO, 2004). Molecular typing should be car-ied out together with a geographic analysis of the region wherehe viruses were isolated because the emergence of infectiousiseases is influenced simultaneously by genetic and ecologicalactors. As the dynamics of an epizootic are reflected in the sub-opulations of the virus in the epizootic area, characterizationf these subpopulations allows spatial and temporal links to bestablished between them (Real et al., 2005).
The results presented in this paper show that there is a corre-ation between the genotypic divergence of the RABV lineagessolated in canid populations in Northeastern Brazil and theeographic distance between the different regional groups ofhe virus and provide an understanding of the dynamics ofow the virus circulates between dog and wild canid biotypes.hylogenetic analysis using pairwise clustering allows RABVycles to be identified in areas where rabies is persistent (Smith,002). In Brazil, because dog and C. thous populations constituteetapopulations, rabies persistence is more likely (Bingham,
005).The available epidemiologic data indicate that the temporal
elationship between RABV populations circulating in North-astern Brazil is not a recent one (SVS/MS). The rabies cyclen this region was not interrupted by mass immunization ofomestic animals, and cases of rabies in dogs and wild canidsre constantly being recorded. The long temporal relationshiphat exists between RABV subpopulations circulating in North-astern Brazil is reflected in the formation of the subclustersOG-RELATED STRAINS and WILD CANID-RELATEDTRAINS, which are divided by regional groups isolated byundreds of kilometers. The spatial organization of RABV
ineages can be explained by neutral evolution coupled withispersal and mixing of RABV lineages and the groupings ofhese sequences indicate ecological subdivisions of the virusopulation (Real et al., 2005).
42 P. Carnieli Jr. et al. / Virus Research 131 (2008) 33–46
Fig. 4. Neighbor-joining phylogenetic tree of 513-nucleotide stretches of the mtDNA control region found in BRAZILIAN CANIDS. The data sequences are showni m thec
saarpv
Bnasib
pt
reetcge
n Table 2. Note the rabies-positive samples 4371PvetPI/5 and 6333PvetPI/5 froanid reference sample sequences.
The results of the antigenic typing of isolates presented herehow that the dog and wild canid biotypes are closely related,s both were found to belong to antigenic variant 2. However,significant number of C. thous were N-Mab C1−. Similar
esults have been reported by a number of authors. For exam-le, Velasco-Villa et al. (2006) described five atypical antigenicariants in Mexico.
The establishment of regional RABV groups in Northeasternrazil reflects the diversity of RABV lineages, suggesting thatew, spatially separated variants coexist or emerged together,
s stated by Velasco-Villa et al. (2005). Kuzmin et al. (2004)uggest that regional differences, which separate the RABVnto geographic groups, are determined by adaptation of speciesiotypes to geographic biomes, and that species other than therbb2
state of Piauı in Northeastern Brazil in the cluster Pseudalopex vetulus. In bold
referred rabies host in a particular geographic area are normallyerminal hosts.
Limited genetic diversity of RABV can be determined by aecent bottleneck or by a limited variety of host species in thepizootic region (Smith, 2002). This is not the case in Northeast-rn Brazil, as rabies is endemic in this region, thus, excludinghe possibility of a recent bottleneck, and the fauna in the regiononsists of a large number of wild species, ensuring that theenotypic variation of the virus is maintained. As RABV lin-ages are perpetuated by certain species that act as regional
eservoirs of the virus (Rupprecht et al., 2002), the similarityetween the RABV lineages remains stable, allowing RABV toe track over space and animal reservoir (Nadin-Davis et al.,006).
s Res
DSwlfttdb(oatci(
R(eltmovn2cihwat
S1ecocadFfCefawao
DStS
tiib
wWtPNePeCiicgtdsarn
bRbaR
aeveto
fuPadetstCsths
P. Carnieli Jr. et al. / Viru
The average nucleotide identity value between the subclustersOG-RELATED STRAINS and WILD CANID-RELATEDTRAINS is greater than that obtained by Carnieli et al. (2006),ho found an average intercluster identity of 88.6% when ana-
yzing 160 nt of the RABV N-gene in other samples also isolatedrom canids in Northeastern Brazil. The discrepancy betweenhe two values (93.1% versus 88.6%) is due to the size ofhe sequences analyzed. Another possible explanation for thisifference is that Carnieli et al. (2006) only sequenced the car-oxy terminus region, where there is great nucleotide variabilityKissi et al., 1995). The values for the intra-subcluster identitiesbtained here indicate that there is a large degree of similaritymong the RABV populations isolated in dog and wild canid bio-ypes. The results also underline the need to analyze the wholeoding region of the N-gene of RABV isolated from animalsn which the virus is maintained in close or overlapping cyclesSmith, 2002).
In this study, greater genetic variability was observed amongABV isolates from wild canids than among those from dogs
Table 3). A similar result has already been reported by Carnielit al. (2006), suggesting that the homogeneity among RABV iso-ated from the dog biotype may be the result of the extensive con-act between the canine population, either as a result of human
ovement or the greater proximity of these animals to eachther. Another factor that may have led to the homogeneity of thisirus is the constant vaccination campaigns that eliminate a largeumber of RABV lineages from the canine population (Smith,002). In contrast, the large geographic distances between wildanid populations and the fragmentation brought about by thencreasing destruction of the natural habitat of these animals byumans prevent RABV lineages being homogeneous. Coupledith the geographic distance, the lack of vaccination campaigns
mong wild canids in Brazil allows greater RABV heterogeneityo be maintained among these animals.
The existence of the two subclusters DOG-RELATEDTRAINS and WILD CANID-RELATED STRAINS in clusterBRAZILIAN CANIDS can be interpreted as the result of an
cological subdivision of the virus population circulating amonganids in Northeastern Brazil. However, the synanthropic habitsf some wild canids, together with the fact that feral dogs areonstantly in contact with wild animals, as described in Suzannd Ceballos (2005), means that the expression “ecological sub-ivision of the virus population” cannot be used in this study.urthermore, the majority of isolates typed here were obtainedrom dog and wild canid biotypes from a single biome (theaatinga), which has a semiarid climate and savannah-type veg-tation, as a result of which animals have to travel in search ofood and water. The animals can travel relatively easily, as therere no major geographic barriers. There are mountain rangesith average heights of 1000 m, and the only big river that could
ct as a barrier, the Sao Francisco, only partly isolates some areasf the Northeast (IBGE, 2007).
There is a flow of RABV lineages between the subclusters
OG-RELATED STRAINS and WILD CANID-RELATEDTRAINS. The RABV isolate 6053CthSE/5, obtained from C.hous and characterized genetically here as DOG-RELATEDTRAIN, proves the existence of this relationship between the
mdr(
earch 131 (2008) 33–46 43
wo subclusters. Carnieli et al. (2006) also described the geneticdentification of a virus typical of the wild canid biotype isolatedn a dog, confirming the flow of RABV lineages between the twoiotypes.
In Northeastern Brazil the ecological niches of dogs andild canids, and the species C. thous in particular, are different.hereas dogs are found in very large numbers in urban areas, C.
hous lives in areas where there are fewer humans. The states ofaraıba and Pernambuco are located in the highest areas of theortheast, and, whether by chance or not, this area has the great-
st number of rabies-positive wild canids and where PE-I andE-II regional groups are identified. The interior of Northeast-rn Brazil is located in a xerophytic system (a biome known asaatinga), whereas the coastal strip, where most cases of rabies
n dogs are identified, has small areas of remnant forest belong-ng to another Brazilian biome, the Atlantic Forest (Fig. 1). Theoastal strip has a higher human population density, and thus, areater concentration of dogs. It is therefore reasonable to asserthat the RABV lineages that circulate between dogs and C. thousiverge as a result of the different use the species make of theame ecosystem and that they do not characterize ecologicaldaptation by the animals but rather geographic separation, aseported by Real et al. (2005), albeit with overlap of ecologicaliches.
In the present study, canid samples from essentially oneiome (the Caatinga) were used, and a regional variation inABV was observed, allowing ecological and spatial links toe established. This finding allowed the regional RABV groupsnalyzed in this study to be characterized as phylogeographicABV lineages.
The emergence of regional groupings of canids shown in thisrticle expresses the diversity of the RABV N-gene in Northeast-rn Brazil and corroborates a previous finding that intragenotypeariation (and, in consequence, RABV diversity) has been under-stimated (Velasco-Villa et al., 2005). Rabies control requireshat there be widespread consensus regarding the epidemiologyf this disease.
Bernardi et al. (2005) typed three RABV isolates from hoaryoxes (P. vetulus = D. vetulus) (the nomenclature of the Canidaesed here follows Sillero-Zubiri et al., 2004) in the state ofaraıba and, after analyzing the P gene, they suggested that thesenimals are responsible for a rabies cycle that is independent ofomestic animals in that region of the state. According to Satot al. (2006), P. vetulus is the main RABV host species amonghe wild canids of Northeastern Brazil, and C. thous plays aecondary role in rabies epidemiology. These results should bereated with caution, however, as according to Dalponte andourtenay (2004), the hoary fox is not normally found in the
tate of Paraıba (Ginsberg and Macdonald, 1990). Furthermore,he results of the genetic identification of the species describedere only identified the hoary fox in the Cerrado biome in thetate of Piauı, where the species is normally found.
When identifying wild canid species involved in rabies trans-
ission, confusion can arise because of the overlap in theistribution of the two main suspected species. C. thous iselatively common throughout its geographic distribution areaFig. 1) (Courtenay and Maffei, 2004; Eisemberg and Redford,

4 s Res
1dlaptcSGdseC
ttRiitwTa
cctmpmvmtct
asmcsblrdosct
utPaN
m
Hbaotil
wiccroeInc
oliCdcacbtltdittcslte
A
lVaMldai
4 P. Carnieli Jr. et al. / Viru
999; Emmons and Feer, 1999). Courtenay and Dalponte (2004)escribe the geographic distribution of the hoary fox, P. vetu-us, associated with the Cerrado habitats (a mosaic of grasslandsnd xerophytic vegetation of the central Brazilian plateau), anderipheral transitional zones including the dry open habitats ofhe Pantanal (in the state of Mato Grosso). They also describeonfirmed reports of the species in the states of Minas Gerais,ao Paulo, Mato Grosso do Sul, Mato Grosso, Tocantins andoias, southern and western Bahia and western Piauı in Serraa Capivara National Park (Fig. 1). Capture records of an extantpecimen held in Teresina Zoological Park suggest that its north-rn geographic limit is probably in northern Piauı (Costa andourtenay, 2003).
The two P. vetulus (hoary fox) (=D. vetulus) shown amonghe other animals in Table 2 were identified in the area wherehe species is normally found. There was probably spillover ofABV from C. thous to P. vetulus. Characterization of RABV
solated in the two P. vetulus revealed minimal differences indentity compared with the other C. thous RABV isolates fromhe PI regional group. Epidemiological data indicate that thereas a rabies outbreak among wild canids in 2005 in the city oferesina, Piauı (SVS/MS), where the P. vetulus were capturednd that this probably made the spillover possible.
One of the aims of this study was to identify species of wildanids by analyzing 513 nt of the mitochondrial DNA (mtDNA)ontrol region (HVS-I) extracted from rabies-positive animalissue. The occurrence of the wild RABV cycle identified here
akes it possible to affirm that other species of wild canidsresent in the region in lower densities, such as P. vetulus, theaned wolf Chrysocyon brachyurus and the bush dog Speothos
enaticus (Zuercher et al., 2004), as well as other terrestrialammals, may be endangered by the presence of the virus. Iden-
ification of the main wild canid species involved in the rabiesycle in Northeastern Brazil is vital if effective rabies control iso be implemented in the region.
In the present study, following sequencing and subsequentnalysis of mtDNA extracted from rabies-positive canids tis-ues from Northeastern Brazil, C. thous was identified as theain RABV host in the epidemiology of rabies among wild
anids in the region. This result agrees with the findings of thepecialists concerning the area where this species can normallye found and, when combined with the results of epidemio-ogic surveillance, allows greater confidence to be placed in theesults described here. In view of the fact that epidemiology inogs is associated with wild canids in the region, identificationf C. thous as a regional wild reservoir means that an ecologicaltudy of the animal can be discussed and carried out and steps toontrol rabies among these wild canids implemented later withhe relevant Brazilian institutions.
The results for the species identification described here allows to characterize C. thous as the rabies biotype species, i.e.,he species is the regional RABV reservoir among wild canids.. vetulus is probably an accidental host in the region studied,
nd thus, plays a less important role in rabies epidemiology inortheastern Brazil.Analysis of mtDNA was used in this study to determine theain RABV host species in wild canids in Northeastern Brazil.
R
B
earch 131 (2008) 33–46
owever, this technique has further applications and can alsoe used for epidemiologic surveillance when only the sample,nd not the carcass, is available for diagnosis, and identificationf the animal is not possible. Also, in cases where samples areaken from animals killed in road accidents, the carcass is oftenn very poor condition and cannot be visually identified. In theatter case, even dogs can be mistaken for wild canids.
The areas where the wild canids originated from (Table 2)ere compared with the geographic groupings of the RABV
solates (Fig. 2) in an attempt to correlate the origin of the wildanids with that of the RABV isolates. The results were incon-lusive, possibly as the result of two factors. First, to identifyeliably the region where the wild canids originated, other areasf the genome of these animals need to be sequenced. Tchaickat al. (2007) obtained positive results when they analyzed HSV-and the mtDNA cytochrome b gene together. Secondly, theumber of wild canids used was too small for us to be able toorrelate the origin of the virus with the hosts.
Rabies has emerged as the most common cause of diseaseutbreaks in wild canids worldwide (WHO, 2004), and a simi-ar situation can also be observed in Brazil. The results describedn this paper show that rabies in dogs is associated with rabies in. thous, the main species of wild canid involved in rabies epi-emiology in Northeastern Brazil. The RABV lineages that areharacteristic of both species overlap spatially but are separateds a result of the different ways these species exploit ecologi-al niches. This finding means that the C. thous population cane characterized as the regional rabies biotype species amongerrestrial carnivores in Northeastern Brazil. The RABV popu-ation in Northeastern Brazil is made up of regional strains, andhe virus subpopulation isolated in C. thous has greater geneticiversity than the virus subpopulation isolated in dogs, suggest-ng that the wild strain has expanded geographically. In additiono being a major pubic health concern, canine rabies is a potentialhreat to wild species. The high degree of susceptibility of wildanids to RABV highlights the need to adopt rabies control mea-ures among wild canids in Brazil. Preservation of wild canids, ateast in so far as rabies is concerned, requires specific programsailored to the particular social and economic circumstances ofach country.
cknowledgements
We wish to thank Jucelia de Jesus Pereira of the Epidemio-ogic and Biostatistical Laboratory, Department of Preventiveeterinary Medicine and Animal Health, USP for her valu-ble help; Dr. Eduardo Eizirik of the Center for Genomic andolecular Biology, Faculty of Biosciences, PUCRS for his col-
aboration in the genetic identification of canids; and Drs. Mariao Socorro C.O. Braga of the Federal University of Maranhaond Enock L.V. da Silva of LACEN-Sergipe, for kindly provid-ng samples of canids from the states in which they work.
eferences
ernardi, F., Nadin-Davis, S.A., Wandeler, A.I., Armstrong, J., Gomes, A.A.,Lima, F.S., Nogueira, F.R., Ito, F.H., 2005. Antigenic and genetic character-

s Res
B
B
CC
C
C
C
D
D
D
D
E
E
F
F
F
G
H
H
I
I
I
K
K
K
K
K
L
N
N
N
O
R
R
S
S
S
S
S
S
T
T
P. Carnieli Jr. et al. / Viru
ization of rabies viruses isolated from domestic and wild animals of Brazilidentifies the hoary fox as a rabies reservoir. J. Gen. Virol. 86, 3153–3162.
ingham, J., 2005. Canine rabies ecology in southern Africa. Emerg. Infect.Dis. 11 (9), 1337–1342.
ourhy, H., Kissi, B., Audry, L., Smreczak, M., Sadkowska-Todys, M., Kulonen,K., Tordo, N., Zmudzinski, F.J., Holmes, C.E., 1999. Ecology and evolutionof rabies virus in Europe. J. Gen. Virol. 80, 2545–2557.
anid Specialist Group & Mammal Assessment, 2007 (www.iucn.org).arnieli Jr., P., Brandao, P.E., Carrieri, M.L., Castilho, J.G., Macedo, C.I.,
Machado, M., Rangel, N., de Carvalho, R.C., de Carvalho, V.A., Monte-bello, L., Wada, M., Kotait, I., 2006. Molecular epidemiology of rabiesvirus strains isolated from wild canids in Northeastern Brazil. Virus Res.120 (1–2), 113–120.
osta, C.H.N., Courtenay, O., 2003. A new record of the hoary fox Pseudalopexvetulus in north Brazil. Mammalia 67, 593–594.
ourtenay, O., Dalponte, J.C., 2004. Hoary fox Pseudalopex vetulus, (Lund,1842). In: Sillero-Zubiri, C., Hoffmann, M., Macdonald, D.W. (Eds.),Canids: Foxes, Wolves, Jackals and Dogs. Status Survey and ConservationAction Plan. IUCN/SSC Canid Specialist Group, Gland, Switzerland andCambridge, UK, p. 430.
ourtenay, O., Maffei, L., 2004. Crab-eating fox Cerdocyon thous, (Linnaeus,1766). In: Sillero-Zubiri, C., Hoffmann, M., Macdonald, D.W. (Eds.),Canids: Foxes, Wolves, Jackals and Dogs. Status Survey and ConservationAction Plan. IUCN/SSC Canid Specialist Group, Gland, Switzerland andCambridge, UK, p. 430.
alponte, J.C., Courtenay, O., 2004. Hoary fox Pseudalopex vetulus, (Lund,1842). In: Sillero-Zubiri, C., Hoffmann, M., Macdonald, D.W. (Eds.),Canids: Foxes, Wolves, Jackals and Dogs. Status Survey and ConservationAction Plan. IUCN/SSC Canid Specialist Group, Gland, Switzerland andCambridge, UK, p. 430.
ean, D.J., Abelseth, M.K., Atanasiu, P., 1996. The fluorescent antibody test. In:Meslin, F.X., Kaplan, M.M., Koprowski, H. (Eds.), Laboratory Techniquesin Rabies, fourth ed. World Health Organization, Geneva, pp. 88–95.
iaz, A.M., Papo, S., Rodriguez, A., Smith, J.S., 1994. Antigenic analysisof rabies-virus isolates from Latin America and the Caribbean. ZentralblVeterinarmed B 41 (3), 153–160.
omingo, E., Holland, J.J., 1994. Mutation rates and rapid evolution of RNAviruses. In: Morse, S.S. (Ed.), The Evolutionary Biology of Viruses. RavenPress, New York, pp. 161–184.
isemberg, J.F., Redford, K.H., 1999. Mammals of the neotropics. The CentralNeotropics: Equador, Peru, Bolıvia, Brasil, 03. The University of ChicagoPress, Chicago, 609.
mmons, L.H., Feer, F., 1999. Neotropical rainforest mammals. In: A FieldGuide, second ed. The University of Chicago Press, Chicago, pp. 307.
auquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., 2005.Virus taxonomy. In: Eighth Report of the International Committee on Tax-onomy of Viruses. Academic Press, San Diego, CA.
avoretto, S.R., Carrieri, M.L., Cunha, E.M., Aguiar, E.A., Silva, L.H., Sodre,M.M., Souza, M.C., Kotait, I., 2002. Antigenic typing of Brazilian virussamples isolated from animals and humans, 1989–2000. Rev. Inst. Med.Trop. Sao Paulo. 44 (2), 91–95.
avoretto, S.R., de Mattos, C.C., de Morais, N.B., Carrieri, M.L., Rolim, B.N.,Silva, L.M., Rupprecht, C.E., Durigon, E.L., de Mattos, C.A., 2006. Rabiesvirus maintained by dogs in humans and terrestrial wildlife, Ceara State,Brazil. Emerg. Infect. Dis. 12, 1978–1981.
insberg, J.R., Macdonald, D.W., 1990. IUCN/SSC Canid Specialist Group.IUCN Publications, Gland, Switzerland and Cambridge, UK, 116 pp.
all, T.A., 1999. BioEdit: a user-friendly biological sequence alignment editorand analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41,95–98.
alpin, K., Hyatt, A.D., Plowright, R.K., Epstein, J.H., Daszak, P., Field,H.E., Wang, L., Daniels, P.W., Henipavirus Ecology Research Group, 2007.Emerging viruses: coming in on a wrinkled wing and a prayer. Clin. Infect.
Dis. 44 (5), 711–717.BGE–Instituto Brasileiro de Geografia e Estatıstica, 2007, http://www.ibge.gov.br/english/default.php.
to, M., Arai, T.Y., Itou, T., Sakai, T., Ito, H.F., Takasaki, T., Kuranet, I., 2001.Genetic characterization and geographic distribution of rabies virus isolates
T
earch 131 (2008) 33–46 45
in Brazil: identification of two reservoirs, dogs and vampire bats. Virology284, 214–222.
to, M., Itou, T., Shoji, Y., Sakai, T., Ito, H.F., Arai, T.Y., Takasaki, T., Kurane,I., 2003. Discrimination between dog-related and vampire bat-related rabiesviruses in Brazil by strain-specific reverse transcriptase-polymerase chainreaction and restriction fragment length polymorphism analysis. J. Clin.Virol. 26 (3), 317–330.
issi, B., Tordo, N., Bourhy, H., 1995. Genetic polymorphism in the rabies virusnucleoprotein gene. Virology 209 (2), 526–537.
issi, B., Badrane, H., Audry, L., Lavenu, A., Tordo, N., Brahimi, M., Bourhy,H., 1999. Dynamics of rabies virus quasispecies during serial passages inheterologous hosts. J. Gen. Virol. 80 (8), 2041–2050.
oprowski, H., 1996. The mouse inoculation test. In: Meslin, F.X., Kaplan,M.M., Koprowski, H. (Eds.), Laboratory Techniques in Rabies, fourth ed.World Health Organization, Geneva, pp. 80–87.
umar, S., Tamura, K., Jakobsen, I.E., Nei, M., 2001. MEGA 2: molecularevolutionary genetic analysis software. In: Evolutionary Genetics AnalysisSoftware. Arizona State University, Tempe.
uzmin, I.V., Botvinkin, A.D., McElhinney, L.M., Smith, J.S., Orciari, L.A.,Hughes, G.J., Fooks, A.R., Rupprecht, C.E., 2004. Molecular epidemiologyof terrestrial rabies in the former Soviet Union. J. Wildl. Dis. 40 (4), 617–631.
e Mercier, P., Jacob, Y., Tordo, N., 1997. The complete Mokola virus genomesequence: structure of the RNA-dependent RNA polymerase. J. Gen. Virol.78 (7), 1571–1576.
adin-Davis, S.A., Casey, G.A., Wandeler, A., 1993. Identification of regionalvariants of the rabies virus within the Canadian province of Ontario. J. Gen.Virol. 74 (5), 829–837.
adin-Davis, S.A., Sampath, M.I., Casey, G.A., Tinline, R.R., Wandeler, A.,1999. Epidemiol. Infect. 123, 325–336.
adin-Davis, S.A., Muldoon, F., Wandeler, A.I., 2006. Persistence of geneticvariants of the arctic fox strain of Rabies virus in southern Ontario. Can. J.Vet. Res. 70 (1), 11–19.
rciari, L.A., Niezgoda, M., Hanlon, C.A., Shaddock, J.H., Sanderlin, J.H.,Yager, P.A., Rupprecht, C.E., 2001. Rapid clearance of SAG-2 rabies virusfrom dogs after oral vaccination. Vaccine 19 (31), 4511–4518.
eal, L.A., Henderson, J.C., Biek, R., Snaman, J., Jack, T.L., Childs, J.E., Stahl,E., Waller, L., Tinline, R., Nadin-Davis, S., 2005. Unifying the spatial pop-ulation dynamics and molecular evolution of epidemic rabies virus. Proc.Natl. Acad. Sci. USA 102 (34), 12107–12111.
upprecht, C.E., Hanlon, A.C., Hemachudha, T., 2002. Rabies re-examined.Lancet Infect. Dis. 2, 327–343.
ambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning: A LaboratoryManual, second ed. Cold Spring Harbor 829 Laboratory Press, New York.
ato, G., Kobayashi, Y., Shoji, Y., Sato, T., Itou, T., Ito, F.H., Santos, H.P., Brito,C.J., Sakai, T., 2006. Molecular epidemiology of rabies from Maranhao andsurrounding states in the northeastern region of Brazil. Arch. Virol. 151 (11),2243–2251 (Epub 2006 June 16).
VS/MS–Secretaria de Vigilancia Sanitaria do Ministerio da Saude, 2006,http://portal.saude.gov.br/portal/saude.
illero-Zubiri, C., Hoffmann, M., Macdonald, D.W., 2004. Canids: foxes,wolves, jackals and dogs. In: Status Survey and Conservation Action Plan.IUCN/SSC Canid Specialist Group, Gland, Switzerland and Cambridge,UK.
mith, J.S., 2002. Molecular Epidemiology. In: Jackson, A.C., Wunner, W.H.(Eds.), Rabies. Academic Press, San Diego, pp. 79–111.
uzan, G., Ceballos, G., 2005. The role of feral mammals on wildlife infectiousdisease prevalence in two nature reserves within Mexico City limits. J. ZooWildl. Med. 36 (3), 479–484.
chaicka, L., Eizirik, E., Oliveira, T.G., Candido Jr., J.F., Freitas, T.R.O., 2007.Phylogeography and population history of the crab-eating fox (Cerdocyonthous). Mol. Ecol. 16, 819–838.
ordo, N., Poch, O., Ermine, A., Keith, G., Rougeon, F., 1986. Walking alongthe rabies genome: Is the large G-L intergenic region a remnant gene? Proc.
Natl. Acad. Sci. USA 83 (11), 3914–3918.ordo, N., Poch, O., Ermine, A., Keith, G., Rougeon, F., 1988. Completion ofthe rabies virus genome sequence determination: highly conserved domainsamong the L (polymerase) proteins of unsegmented negative-strand RNAviruses. Virology 165 (2), 565–576.

4 s Res
V
V
W
W
W
Zuercher, G.L., Swarner, M., Silveira, L., Carrillo, O., 2004. Bush dog Speothos
6 P. Carnieli Jr. et al. / Viru
elasco-Villa, A., Orciari, L.A., Souza, V., Juarez-Islas, V., Gomez-Sierra, M.,Castillo, A., Flisser, A., Rupprecht, C.E., 2005. Molecular epizootiology ofrabies associated with terrestrial carnivores in Mexico. Virus Res. 111 (1),13–27 (Epub 2005 April 25).
elasco-Villa, A., Orciari, L.A., Juarez-Islas, V., Gomez-Sierra, M., Padilla-Medina, I., Flisser, A., Souza, V., Castillo, A., Franka, R., Escalante-Mane,M., Sauri-Gonzalez, I., Rupprecht, C.E., 2006. Molecular diversity of rabies
viruses associated with bats in Mexico and other countries of the Americas.J. Clin. Microbiol. 44 (5), 1697–1710.ard, R.H., Frazier, B.L., Dew-Jager, K., Paabo, S., 1991. Extensive mitochon-drial diversity within a single 875 Amerindian tribe. Proc. Natl. Acad. Sci.88, 8720–8724.
earch 131 (2008) 33–46
ilson, D.E., Reeder, D.A.M., 1993. Mammal Species of the World: A Taxo-nomic and Geographic Reference, second ed. Smithsonian Institution Press,Washington.
HO—World Health Organization, 2004. Expert Consultation on Rabies.First Report (Technical Report Series, 931), World Health Organization,Geneva.
venaticus (Lund, 1842). In: Sillero-Zubiri, C., Hoffmann, M., Macdon-ald, D.W. (Eds.), Canids: Foxes, Wolves, Jackals and Dogs. Status Surveyand Conservation Action Plan. IUCN/SSC Canid Specialist Group, Gland,Switzerland and Cambridge, UK, p. 430.
Copyright © 2022 FDOKUMEN




















