
Characterization of genetically modified human retinal pigment epithelial cells developed for in...
10
Characterization of Genetically Modified Human Retinal Pigment Epithelial Cells Developed for In Vitro and Transplantation Studies Naheed Kanuga, 1 Helen L. Winton, 1 Laurence Beauche ´ne, 2 Ahmet Koman, 2,3 Anne Zerbib, 2 Stephanie Halford, 4 Pierre-Olivier Couraud, 2,5 David Keegan, 6 Pete Coffey, 7 Raymond D. Lund, 6,8 Peter Adamson, 1 and John Greenwood 1 PURPOSE. To develop, by specific genetic modification, a differ- entiated human retinal pigment epithelial (RPE) cell line with an extended life span that can be used for investigating their function in vitro and for in vivo transplantation studies. METHODS. Primary human RPE cells were genetically modified by transfecting with a plasmid encoding the simian virus (SV)40 large T antigen. After characterization, two cell lines, designated h1RPE-7 and h1RPE-116, were chosen for further investigation, along with the spontaneously derived RPE cell line ARPE-19. Factors reported to be important in RPE and photoreceptor cell function and survival in vivo were exam- ined. RESULTS. Both h1RPE-7 and h1RPE-116 cells exhibited epithelial morphology, expressed cytokeratins, and displayed junctional distribution of ZO-1, p100-p120 and -catenin. The cells ex- pressed mRNA for RPE65 and cellular retinaldehyde-binding protein (CRALBP) and the trophic and growth factors brain- derived neurotropic factor (BDNF), ciliary neurotrophic factor (CNTF), basic fibroblast growth factor (bFGF), pigment epithe- lium– derived factor (PEDF), nerve growth factor (NGF), plate- let-derived growth factor (PDGF)-, insulin-like growth factor (IGF)-1, and vascular endothelial growth factor (VEGF). Se- creted BDNF, bFGF, and VEGF, but not CNTF, were identified in cell supernatants. The cell lines constitutively expressed HLA-ABC, CD54, CD58, and CD59. After activation with IFN- both HLA-ABC and CD54 were upregulated, and the expres- sion of HLA-DR was induced. Both cell lines failed to express CD80, CD86, CD40, or CD48 in vitro and in a mixed lympho- cyte reaction were unable to induce T-cell proliferation. Fas ligand (CD95L) was not detected in vitro by RT-PCR. Similar results were obtained with the ARPE-19 cell line. CONCLUSIONS. RPE lines h1RPE-7 and h1RPE-116 retain many of the morphologic and biochemical characteristics of RPE cells in vivo and may serve as a source of cells for in vitro analysis of RPE cell function, as well as for orthotopic transplantation studies. (Invest Ophthalmol Vis Sci. 2002;43:546 –555) R etinal pigment epithelial (RPE) cells perform a complex array of functions that are necessary in maintaining retinal photoreceptor homeostasis. Positioned strategically between the neural retina and the vascular choriocapillaris, the RPE constitutes the outer aspect of the blood–retinal barrier and controls the passage of metabolites to and from the circulation. Dysfunction of the RPE cell layer can lead to devastating effects on retinal function and in some diseases, such as age-related macular degeneration (AMD), 1–3 can lead to photoreceptor cell death. Our detailed understanding of the cellular function of human RPE cells remains limited, particularly with respect to their role in ocular disease. Much of our understanding has arisen from in vitro investigations of primary or early-passage cells harvested from donor eyes and more recently from the development of a number of human RPE cell lines that are phenotypically more stable in long-term culture. The limited life span of normal human RPE cells represents a substantial obstacle for cell and molecular analysis of these cells, and therefore such cell lines are considered to be an important resource. One cell line in particular, spontaneously arising ARPE-19, has been a valuable source of human RPE cells, despite the unknown nature of the immortalizing event conferring immor- tality. However, the exact profile of RPE-specific characteristics of this cell line remain unknown, and there is a strong case for the development of further RPE cell lines that may express specific characteristics of interest. Furthermore, in in vivo RPE transplantation studies, particularly if these are ultimately in- tended for treatment of patients, a known genetic modification may be more desirable than an undefined genetic alteration. We have previously reported that the subretinal transplan- tation of RPE cells into the Royal College of Surgeons (RCS) rat delays photoreceptor cell death 4,5 and, along with others, 6–9 have proposed that this may be an effective treatment strategy for diseases such as AMD. To overcome some of the limitations imposed by using primary cultures harvested from donor eyes, such as inadequate supply and difficulty in screening for patho- gens, the use of cell lines as a source of RPE cells for orthotopic transplantation may prove advantageous. Despite some major potential problems associated with this approach, such as the cells’ safety and ability to function effectively in vivo, it remains a promising strategy. In this study we therefore set out to develop additional human RPE cell lines with extended life span by transfection with the simian virus (SV)40 large T antigen. The generation of a range of cell phenotypes after From the Divisions of 1 Cell Biology, 4 Molecular Genetics, and 6 Pathology, Institute of Ophthalmology, University College London, London, United Kingdom; 2 Neurotech SA, Evry, France; 5 Institut Co- chin de Ge ´ne ´tique Mole ´culaire, Centre National de la Recherche Sci- entifique, Unite ´ Propre de Recherche 415, Paris, France; and the 7 Department of Psychology, University of Sheffield, Sheffield, United Kingdom. Present affiliations: 3 Department of Molecular Biology and Genet- ics, Bogazici University, Istanbul, Turkey; and the 8 Moran Eye Center, University of Utah, Salt Lake City, Utah. Supported by Neurotech SA, Ba ˆtiment Ge ´nopole-Industries, Evry, France. Submitted for publication July 9, 2001; revised October 11, 2001; accepted October 24, 2001. Commercial relationships policy: E (POC, LB, AK, AZ); F (RDL, PC, PA, JG); P (PA, RDL, JG); N (NK, HLW, SH, DK). The publication costs of this article were defrayed in part by page charge payment. This article must therefore be marked “advertise- ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact. Corresponding author: John Greenwood, Endothelial and Epithe- lial Cell Biology Research Unit, Division of Cell Biology, Institute of Ophthalmology, University College London, Bath Street, London EC1V 9EL, UK; [email protected]. Investigative Ophthalmology & Visual Science, February 2002, Vol. 43, No. 2 546 Copyright © Association for Research in Vision and Ophthalmology
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Characterization of genetically modified human retinal pigment epithelial cells developed for in...

Characterization of Genetically Modified Human RetinalPigment Epithelial Cells Developed for In Vitro andTransplantation Studies
Naheed Kanuga,1 Helen L. Winton,1 Laurence Beauchene,2 Ahmet Koman,2,3
Anne Zerbib,2 Stephanie Halford,4 Pierre-Olivier Couraud,2,5 David Keegan,6
Pete Coffey,7 Raymond D. Lund,6,8 Peter Adamson,1 and John Greenwood1
PURPOSE. To develop, by specific genetic modification, a differ-entiated human retinal pigment epithelial (RPE) cell line withan extended life span that can be used for investigating theirfunction in vitro and for in vivo transplantation studies.
METHODS. Primary human RPE cells were genetically modifiedby transfecting with a plasmid encoding the simian virus(SV)40 large T antigen. After characterization, two cell lines,designated h1RPE-7 and h1RPE-116, were chosen for furtherinvestigation, along with the spontaneously derived RPE cellline ARPE-19. Factors reported to be important in RPE andphotoreceptor cell function and survival in vivo were exam-ined.
RESULTS. Both h1RPE-7 and h1RPE-116 cells exhibited epithelialmorphology, expressed cytokeratins, and displayed junctionaldistribution of ZO-1, p100-p120 and �-catenin. The cells ex-pressed mRNA for RPE65 and cellular retinaldehyde-bindingprotein (CRALBP) and the trophic and growth factors brain-derived neurotropic factor (BDNF), ciliary neurotrophic factor(CNTF), basic fibroblast growth factor (bFGF), pigment epithe-lium–derived factor (PEDF), nerve growth factor (NGF), plate-let-derived growth factor (PDGF)-�, insulin-like growth factor(IGF)-1, and vascular endothelial growth factor (VEGF). Se-creted BDNF, bFGF, and VEGF, but not CNTF, were identifiedin cell supernatants. The cell lines constitutively expressedHLA-ABC, CD54, CD58, and CD59. After activation with IFN-�both HLA-ABC and CD54 were upregulated, and the expres-sion of HLA-DR was induced. Both cell lines failed to expressCD80, CD86, CD40, or CD48 in vitro and in a mixed lympho-cyte reaction were unable to induce T-cell proliferation. Fas
ligand (CD95L) was not detected in vitro by RT-PCR. Similarresults were obtained with the ARPE-19 cell line.
CONCLUSIONS. RPE lines h1RPE-7 and h1RPE-116 retain many ofthe morphologic and biochemical characteristics of RPE cellsin vivo and may serve as a source of cells for in vitro analysis ofRPE cell function, as well as for orthotopic transplantationstudies. (Invest Ophthalmol Vis Sci. 2002;43:546–555)
Retinal pigment epithelial (RPE) cells perform a complexarray of functions that are necessary in maintaining retinal
photoreceptor homeostasis. Positioned strategically betweenthe neural retina and the vascular choriocapillaris, the RPEconstitutes the outer aspect of the blood–retinal barrier andcontrols the passage of metabolites to and from the circulation.Dysfunction of the RPE cell layer can lead to devastating effectson retinal function and in some diseases, such as age-relatedmacular degeneration (AMD),1–3 can lead to photoreceptorcell death. Our detailed understanding of the cellular functionof human RPE cells remains limited, particularly with respectto their role in ocular disease. Much of our understanding hasarisen from in vitro investigations of primary or early-passagecells harvested from donor eyes and more recently from thedevelopment of a number of human RPE cell lines that arephenotypically more stable in long-term culture. The limitedlife span of normal human RPE cells represents a substantialobstacle for cell and molecular analysis of these cells, andtherefore such cell lines are considered to be an importantresource.
One cell line in particular, spontaneously arising ARPE-19,has been a valuable source of human RPE cells, despite theunknown nature of the immortalizing event conferring immor-tality. However, the exact profile of RPE-specific characteristicsof this cell line remain unknown, and there is a strong case forthe development of further RPE cell lines that may expressspecific characteristics of interest. Furthermore, in in vivo RPEtransplantation studies, particularly if these are ultimately in-tended for treatment of patients, a known genetic modificationmay be more desirable than an undefined genetic alteration.
We have previously reported that the subretinal transplan-tation of RPE cells into the Royal College of Surgeons (RCS) ratdelays photoreceptor cell death4,5 and, along with others,6–9
have proposed that this may be an effective treatment strategyfor diseases such as AMD. To overcome some of the limitationsimposed by using primary cultures harvested from donor eyes,such as inadequate supply and difficulty in screening for patho-gens, the use of cell lines as a source of RPE cells for orthotopictransplantation may prove advantageous. Despite some majorpotential problems associated with this approach, such as thecells’ safety and ability to function effectively in vivo, it remainsa promising strategy. In this study we therefore set out todevelop additional human RPE cell lines with extended lifespan by transfection with the simian virus (SV)40 large Tantigen. The generation of a range of cell phenotypes after
From the Divisions of 1Cell Biology, 4Molecular Genetics, and6Pathology, Institute of Ophthalmology, University College London,London, United Kingdom; 2Neurotech SA, Evry, France; 5Institut Co-chin de Genetique Moleculaire, Centre National de la Recherche Sci-entifique, Unite Propre de Recherche 415, Paris, France; and the7Department of Psychology, University of Sheffield, Sheffield, UnitedKingdom.
Present affiliations: 3Department of Molecular Biology and Genet-ics, Bogazici University, Istanbul, Turkey; and the 8Moran Eye Center,University of Utah, Salt Lake City, Utah.
Supported by Neurotech SA, Batiment Genopole-Industries, Evry,France.
Submitted for publication July 9, 2001; revised October 11, 2001;accepted October 24, 2001.
Commercial relationships policy: E (POC, LB, AK, AZ); F (RDL, PC,PA, JG); P (PA, RDL, JG); N (NK, HLW, SH, DK).
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be marked “advertise-ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Corresponding author: John Greenwood, Endothelial and Epithe-lial Cell Biology Research Unit, Division of Cell Biology, Institute ofOphthalmology, University College London, Bath Street, London EC1V9EL, UK; [email protected].
Investigative Ophthalmology & Visual Science, February 2002, Vol. 43, No. 2546 Copyright © Association for Research in Vision and Ophthalmology

expression of SV40 large T antigen allows for the systematicselection of lines that exhibit both normal RPE characteristicsand a nontransformed phenotype. Furthermore, this approachenables us to generate a pathogen-free cell line generated froman individual with a recorded lineage. The cell lines generatedin this study were then tested alongside ARPE-19 cells forRPE-specific and epithelial characteristics and for factors thatare believed to be important in cell survival and in the supportof photoreceptor cells in vivo.
MATERIALS AND METHODS
ReagentsHam’s F-10, Myoclone heat-inactivated fetal calf serum (FCS; sourcedfrom the United States) and penicillin-streptomycin were purchasedfrom Gibco Life Technologies (Paisley, Scotland, UK). FuGene 6 trans-fection reagent was obtained from Roche Molecular Biochemicals(Mannheim, Germany). Antibodies for immunocytochemistry werehamster polyclonal anti-SV40 large T antigen (a gift from MireilleViguier, Institut Cochin de Genetique Moleculaire, Paris, France); rab-bit polyclonal ZO-1 (Zymed, Cambridge Bioscience, Cambridge, UK);monoclonal antibodies RGE-53, specific for cytokeratin 18, and RCK-102, specific for cytokeratins 5 and 8 (Eurodiagnostica, Arnhem, TheNetherlands); and NCL-5D3, specific for cytokeratins 8 and 18 (Novo-castra Laboratories, Newcastle-upon-Tyne, UK); �-catenin (clone 14)and p100-p120 (clone 98; Transduction Laboratories, Nottingham,UK); and rabbit polyclonal von Willebrand factor (Dako, High Wy-combe, UK). Monoclonal antibodies used for flow cytometry wereanti-human HLA-ABC (clone W6-32), CD48 (clone MEM-102), CD58(clone BRIC-5), CD59 (clone MEM-43), CD80 (B7-1; clone DAL-1) andCD86 (B7-2; clone BU63), all from Serotec (Oxford, UK); HLA DR(clone G46-6) and CD40 (clone 5C3) from BD Pharmingen (Oxford,UK); and CD54 (ICAM-1; clone BBIG-11) from R&D Systems (Oxford,UK). Tetrarhodamine isothiocyanate (TRITC) and FITC-conjugatedgoat anti-mouse and goat anti-rabbit secondary antibodies were ob-tained from Jackson ImmunoResearch Laboratories (West Grove, PA).Primers for RT-PCR were purchased from Oswel (Southampton, UK),and other PCR reagents were from Perkin Elmer (Beaconsfield, UK).Unless specified, all other reagents were obtained from Sigma (Poole,UK) and were of the highest grade available.
Isolation and Culture of Human RPE CellsAfter local ethics committee approval and according to national guide-lines and in compliance with the tenets of the Declaration of Helsinki,a human eye was obtained from a 50-year-old female white donor, whounderwent exenteration due to conjunctival disease, and the anteriorsegment, iris, and lens were removed. The posterior segment wasplaced in sterile culture medium consisting of Ham’s F-10, 20% FCS, 2mM L-glutamine, 100 IU/mL penicillin, 100 �g/mL streptomycin, and 1�g/mL amphotericin-B and was stored overnight in the dark at 4°C tofacilitate separation of the retina from the RPE. Vitreous and neuralretina were gently teased away from the optic nerve, and the eye cupwas dissected into three segments. Cloning rings were placed on theexposed RPE cell layer, and cells were rinsed with 0.02% EDTA for 5minutes. Trypsin-EDTA (0.25%/0.02%) was pipetted into the cloningrings and incubated at 37°C for 45 minutes to detach RPE from Bruch’smembrane. RPE cells, released by gentle trituration, were plated ontoFCS-coated tissue culture flasks, and after 10 minutes, culture mediumwas added. Medium was changed every 2 to 3 days, and all cells werecultured at 37°C in 5% CO2. For comparative studies, primary culturesof human RPE cells were generated from additional donor eyes, usingthe same procedure, and the spontaneously arising human RPE cellline, ARPE-19 was obtained from ATCC (CRL-2302; Rockville, MD) andgrown as previously reported.10
Genetic Modification of Human RPE CellsThe primary human RPE culture, derived from the single donor de-scribed earlier, was transfected with RSV puro (Neurotech S.A., Evry,
France; which encodes a puromycin-selectable marker) and either aconstruct encoding SV40 large T antigen (pVim Twt) or SV40 large Tantigen that was deleted for the small T antigen (pVim T�t). Theconstructs were generously provided by Denise Paulin, University ofParis, Paris, France).11 This construct was chosen, because expressionof the SV40 large T antigen is known to result in a stable cell pheno-type and an associated extension of life span, but without induction ofcellular transformation. Transfection was performed in six-well platesseeded with 2 � 104 cells/cm2. pVimTwt or pVimT�t (0.9 �g) wascombined with 0.1 �g of RSV puro in 100 �L serum-free mediumcontaining 3 �L transfection reagent (FuGene; Roche). After a 48-hourincubation, cells were trypsinized and replated into Petri dishes andcultured in medium containing 1 �g/mL puromycin to select fortransfected cells. Surviving colonies were removed from Petri disheswith cloning rings and cultured in 20% FCS, 2 mM L-glutamine, 100 IUpenicillin with 100 �g/mL streptomycin, and 1 �g/mL puromycin. Onehundred twenty-six cell lines with large T�t and 42 cell lines withwild-type large T antigen were generated. Each cell line was assessedby phase-contrast microscopy for ability to form contact-inhibitedmonolayers of an epithelial cobblestone phenotype and, using mor-phologic criteria, 18 cell lines were selected for further characteriza-tion.
Characterization of RPE Cells
Immunocytochemistry. Primary human RPE cultures, theARPE-19 cell line, and the 18 selected genetically modified cell lineswere cultured to confluence on eight-well chamber slides (Gibco) forimmunocytochemical characterization. Cells were fixed in either 3.7%paraformaldehyde for 10 minutes or, for cytokeratin detection, ice-coldmethanol-acetone (1:1). Cells were permeabilized with 0.25% TritonX-100 and PBS for 10 minutes and blocked for 15 minutes with 10%goat serum and PBS. Primary antibodies were diluted in blockingsolution, added to the cells and incubated for 1 hour at room temper-ature. Detection was achieved using FITC- or TRITC-conjugated sec-ondary antibodies (1:50) added for 1 hour in the dark. After washing,cells were mounted, and the cell expression and distribution recordedby epifluorescence and confocal laser scanning microscopy. Omissionof the primary or secondary antibody in each case served as thenegative control. On the basis of morphologic appearance and theexpression of cytokeratins and SV40 large T and junctional molecules,two cell lines, generated as described earlier, were selected for furtheranalysis. One cell line, designated h1RPE-7, originated from the SV40large T antigen deleted for the small T antigen–transfected parent line,and another cell line, designated h1RPE-116, originated from transfec-tion with the SV40 large T wild-type antigen. All subsequent analysesdescribed in the following sections were performed on these two celllines between passages 13 and 22.
Ultrastructure. RPE cells were grown to confluence on 24-welltissue culture plates and fixed with one-half strength Karnovsky’sfixative (2% formaldehyde, 2% glutaraldehyde, 0.2 M sodium cacody-late buffer, and 6.5 mM calcium chloride). The monolayer was post-fixed in 1% osmium tetroxide for 1 hour, washed, and dehydratedthrough ascending concentrations of ethanol. For transmission elec-tron microscopy (TEM), the monolayers were embedded in resin andultrathin sections cut and counterstained with uranyl acetate and leadcitrate. The sections were viewed in a transmission electron micro-scope (model 1010; JEOL, Herts, UK). For scanning electron micros-copy (SEM), the monolayers were critical-point dried with CO2 andsputter coated with 20 nm of gold and then viewed on a scanningelectron microscope (model 6100; JEOL).
Reverse Transcription–Polymerase Chain Reaction.Expression of mRNAs encoding for discriminating markers, for growthand trophic factors deemed to be important in photoreceptor cellfunction and survival, and for Fas ligand (FasL; CD95L) were examinedusing reverse transcription–polymerase chain reaction (RT-PCR). Thethree cell lines—h1RPE-7, h1RPE-116, and ARPE-19—were investi-gated for the expression of mRNA for the discriminative cellular mark-
IOVS, February 2002, Vol. 43, No. 2 Genetically Modified Human RPE Cells 547

ers RPE65 and cellular retinaldehyde-binding protein (CRALBP). Thegrowth and trophic factors investigated were pigment epithelium–derived factor (PEDF), brain-derived neurotrophic factor (BDNF), basicfibroblast growth factor (bFGF), ciliary neurotrophic factor (CNTF),platelet-derived growth factor (PDGF)-�, insulin-like growth factor(IGF-1), nerve growth factor (NGF), and vascular endothelial growthfactor (VEGF). Oligonucleotide primers complimentary to the 5� and 3�ends were used in RT-PCR studies (Table 1).
For identification of discriminating markers and growth and trophicfactors, total RNA was extracted from 2-week postconfluent monolay-ers of h1RPE-7, h1RPE-116, and ARPE-19 cells. For identification ofFasL, total RNA was extracted from confluent cell monolayers ofh1RPE-7, h1RPE-116, and ARPE-19 cell lines and primary RPE cells, aswell as from human T cells. In all cases, total RNA was extracted usinga kit (RNeasy; Qiagen, West Sussex, UK). Oligo (dT)12-18 (1 �g ) primedtotal RNA was reverse transcribed using avian myoblastosis virus re-verse transcriptase (AMV-RT) supplied with a first-strand synthesissystem for RT-PCR (SuperScript; Life Technologies). PCR reactionscontained 1 �L cDNA, 67 mM Tris-HCl (pH 8.8), 16 mM (NH4)2SO4, 1.5mM MgCl2, 0.2 mM dNTPs, 10 pmol sense primer, 10 pmol antisenseprimer, 1.25 U DNA polymerase (BioTaq; Bioline, London, UK) in a50-�L reaction. PCR cycle parameters were conducted with a pream-plification denaturation at 94°C for 4 minutes, followed by 35 cycles ofdenaturation at 94°C for 30 seconds, annealing at the primer pair–specific annealing temperature (Table 1) for 30 seconds, and extensionat 72°C for 30 to 90 seconds, depending on the amplicon size ex-pected. PCR amplification of �-actin was routinely used as a control toassess integrity of RNA and cDNA. Ten-microliter samples of the am-plification reactions were resolved on 1% to 2% TAE agarose gels andthe products visualized by ethidium bromide staining. To verify au-thenticity, amplicons were excised from gels, repurified, and subjected
to DNA sequence analysis. Amplicons were cycle sequenced, usingsense primers and analyzed on an automated DNA sequencer (ABI 377Prism; Perkin Elmer).
Secreted Growth and Trophic Factors. Cell culture super-
natants were collected from either RPE cells derived from the originaldonor or from h1RPE-7, h1RPE-116, and ARPE-19 cell lines 2 weeksafter reaching confluence. Cell supernatants were collected after 5days of conditioning and centrifuged to remove particulates. Theconcentrations of bFGF, BDNF, CNTF, and VEGF (R&D Systems),human interferon (hIFN)-� (Amersham PLC, Amersham, UK) and hu-man interferon (hIFN)-� (Biosource International, Camarillo, CA) weremeasured using commercially available enzyme-linked immunosorbentassays (ELISA) in accordance with the manufacturer’s protocol.
Flow Cytometry. The expression of potentially important im-
munologic molecules was evaluated in h1RPE-7, h1RPE-116, andARPE-19 cell lines and in third-passage human RPE cells by flowcytometry. Cells were grown in six-well tissue culture dishes and wereleft either untreated or treated with 100 to 1000 U/mL IFN-� (R&DSystems) for 48 or 72 hours. After treatment, the cells were preparedas a single-cell suspension by treating with 0.5 mL of 1 mg/mL colla-genase for 10 to 15 minutes at 37°C. The cells were pelleted bycentrifugation and washed by resuspension in 1 mL sterile PBS. Eachcell suspension was then pelleted and resuspended in 100 �L PBS,with or without the appropriate primary antibody and incubated on icefor 1 hour. Two PBS washes were performed and the cells wereincubated with a secondary FITC-conjugated antibody for 1 hour on icein the dark, washed with PBS, and resuspended in 200 to 500 �L PBSfor flow cytometric analysis. The cell surface expression of majorhistocompatibility complex (MHC) class I (HLA-ABC) and class II (HLA-DR) antigens; the costimulatory molecules intercellular adhesion mol-
TABLE 1. Primer Sequences used for PCR
GenePrimer Sequence (5�-3�)
Position in mRNAAnnealingTemp (°C)
ProductSize (bp)
RPE-65 5� ATG TCT ATC CAG GTT GAGRPE-65 3� TCA AGA TTT TTT GAA CAG 52 1601
RPE-65 5� CCT TTC TTC ATG GAG TCT TTGRPE-65 3� ATT GCA GTG GCA GTT GTA TTG 52 390
CRALBP 5� ATG TCA GAA GGG GTG GGCRALBP 3� TCA GAA GGC TGT GTT CTC A 60 953
PEDF 5� GGA CGC TGG ATT AGA AGG CAGPEDF 3� TTG TAT GCA TTG AAA CCT TAC AGG 65 1490
BDNF 5� ATG ACC ATC CTT TTC CTT ACT ATG GTBDNF 3� TCT TCC CCT TTT AAT GGT CAA TGT AC 52 741
bFGF 5� GCC TTC CCG CCC GGC CAC TTC AAG GbFGF 3� GCA CAC ACT CCT TTG ATA GAC ACA A 55 179
CNTF 5� TGG CTA GCA AGG AAG ATT CGTCNTF 3� ACG AAG GTC ATG GAT GGA CCT 65 468
PDGF� 5� CCT GCC CAT TCG GAG GAA GAGPDGF� 3� TTG GCC ACC TTG ACG CTG CG 65 225
IGF-1 5� ATG TCC TCC TCG CAT CTC TTCIGF-1 3� CCT GTA GTT CTT GTT TCC TGC 65 337
NGF 5� TCG GCA TAC AGG CGC AAC CANGF 3� CCT GCT TGC CAT CCA TGG TC 55 612
VEGF 5� TTG CCT TGC TGC TCT ACC TCVEGF 3� AAA TGC TTT CTC CGC TCT GA 65 424
FasL 5� GGA TTG GGC CTG GGG ATG TTT CAFasL 3� TTG TGG CTC AGG GGC AGG TTG TTG 67 344
�-Actin 5� GAG CAC AGA GCC TCG CCT TTG C�-Actin 3� GGA TCT TCA TGA GGT AGT CAG TCA GG 65 620
548 Kanuga et al. IOVS, February 2002, Vol. 43, No. 2

ecule (ICAM)-1 (CD54), CD40, B7-1 (CD80), and B7-2 (CD86); and thepotential CD2 ligands CD48, CD58, and CD59 were evaluated by flowcytometry (FACScan; BD Biosciences, Oxford, UK). The data wereanalyzed on computer (CellQuest software; BD Biosciences).
Assessment of Tumorigenicity
To assess the tumorigenic capacity of h1RPE-7 and h1RPE-116 cells invivo, 10 � 106 cells suspended in 200 �L PBS and 10 mM glucose wereinjected subcutaneously (SC) into irradiated athymic Swiss nude mice(n � 10 animals for each clone). As a positive control, U87 humanglioblastoma cells were also injected SC at the same cell density (n �5 animals). Animals were assessed by daily gross examination and after3 and 15 weeks by histologic analysis of the graft site. At 3 or 15 weeksafter injection the nude mice were perfusion fixed with 4% parafor-maldehyde and PBS. The tissue was then postfixed in 4% paraformal-dehyde and PBS for 2 hours at 4°C, followed by cryoprotection in 20%sucrose and PBS overnight at 4°C and finally embedded in optimalcutting temperature compound (TissueTek; Miles Laboratories,Elkhart, IN). Tissue was frozen for 30 seconds at �40°C in isopentaneand liquid nitrogen and stored at �20°C until sectioned. Sections werecut (14 �m) and processed for hematoxylin and eosin staining. Tissuesections were finally dehydrated and mounted in resin (Eukitt; AgarScientific, Essex, UK). Animal care was in accordance with the ARVOStatement for the Use of Animals in Ophthalmic and Vision Research.Home Office (United Kingdom) regulations for the care and use oflaboratory animals and the provisions of the UK Animals (ScientificProcedures) Act (1986) were observed at all times.
Anchorage-independent growth, which is a function of cell trans-formation and tumorigenicity, was assessed using a soft agar colonyassay. Low melting temperature agarose (0.5% in culture medium;Seaplaque; Novara Group, Ltd, Staffs, UK) was plated onto 24-wellplates and left to gel. The cell lines h1RPE-7 and h1RPE-116 and thehuman glioblastoma cell line U87 were then suspended in a secondlayer of 0.33% agarose (7 � 104 cells per well), plated into individualwells, and placed at 4°C for 5 minutes to set. Media were then addedto each well and incubated at 37°C with regular changes. The wellswere examined by phase-contrast microscopy every 2 to 3 days for upto 40 days for the presence of cell aggregates or foci.
T-Cell Proliferation Assay:Mixed-Lymphocyte Reaction
Human peripheral blood mononuclear cells (PBMCs) were harvestedfrom a healthy human donor by density-gradient centrifugation (Ficoll-Paque; Pharmacia Biotech AB, Uppsala, Sweden). The PBMCs werethen plated onto plastic Petri dishes for 45 minutes at 37°C-5%CO2 toallow monocytes to adhere. The nonadherent cells were then re-moved, and the enriched population of T lymphocytes was cocultured(2 � 105 cells/well) with either stimulated (IFN-� at 100 U/mL for 48hours) or unstimulated human RPE cell lines that had previously beenirradiated (240 Gy). The RPE cell lines, h1RPE-7, h1RPE-116, andARPE-19 were plated at various concentrations, 2 � 104, 1 � 104, 0.5 �104, and 0.25 � 104 cells/well in triplicate in 96-well plates in 200 �LRPMI 1640 (Life Technologies) supplemented with 10% FCS, penicillin100 U/mL, and streptomycin 100 �g/mL. Two irradiated (120 Gy),allogeneic human B-cell lines (MOU, 9050; SA, 9001; from the TenthInternational Histocompatibility Workshop, New York, 1987) wereused as a positive control at the same concentrations as the RPE cells.The allogeneic T-cells were harvested, and proliferation was assessedon day 6, after an 18-hour pulse with [3H]-thymidine (0.5 �Ci/well), bymeasuring incorporated [3H]-thymidine by liquid scintillation spectros-copy. Background uptake of [3H]-thymidine into the irradiated cellsalone was subtracted from the harvested T-cell data.
RESULTS
Morphology
Primary cultures of human RPE cells grew as contact-inhibitedmonolayers and exhibited a cobblestone epithelial morphologywith areas of dense pigmentation that were gradually lost asthe cells divided in culture (Fig. 1A). The two human RPE celllines, h1RPE-7 (Fig. 1B) and h1RPE-116 (Fig. 1C), also grew ascontact-inhibited monolayers and exhibited an epithelial mor-phology but were devoid of pigmentation. Ultrastructurally,the two human RPE cell lines produced their own basal lamina,had a heterogeneous expression of apical microvilli (Figs. 2A,2B), and exhibited junctional specialization suggestive of tightjunctions (Figs. 2C–E).
Immunocytochemical Characterization
Cells of epithelial origin express cytokeratin intermediate fila-ments, which are considered a characteristic marker of theepithelial phenotype. Primary RPE cells, and both h1RPE-7 andh1RPE-116 cells showed positive immunoreactivity for apanel of anti-human cytokeratin antibodies including cloneRCK102 directed against cytokeratins 5 and 8 (all epithelia) andclones RGE53 (cytokeratin 18) and NCL5D3 (cytokeratins 8 and18; Figs. 1D–F), which recognize nonstratified epithelium.ARPE-19 were also found to be positive for these cytokeratins,
FIGURE 1. Phase-contrast photomicrographs of (A) a primary-donorRPE cell culture (day 3) and the derived cell lines, (B) h1RPE-7 (passage15) and (C) h1RPE-116 (passage 17). Cytokeratin-8 and -18 expressionin third-passage donor RPE cells (D), h1RPE-7 (E), and h1RPE-116 (F).ZO-1 and �-catenin expression in third-passage donor RPE cells (G andJ, respectively), passage 16 h1RPE-7 cells (H and K, respectively), andpassage 17 h1RPE-116 (I and L, respectively). Bars, (A–C) 100 �m;(D–F) 50 �m; (G–L) 20 �m.
IOVS, February 2002, Vol. 43, No. 2 Genetically Modified Human RPE Cells 549

whereas all RPE cells tested were negative for the endothelialcell marker, von Willebrand factor (data not shown). The celllines h1RPE-7 and h1RPE-116 cells were also confirmed aspositive for the SV40 large T antigen (data not shown).
The barrier function and polarization of RPE cells is consid-ered to be highly dependent on the integrity of specializedjunctions. Analysis of junctional proteins revealed strong pe-ripheral staining of ZO-1 in both primary RPE and the cell linesh1RPE-7 and h1RPE-116 (Figs. 1G–I). The staining pattern ofZO-1 at the point of cell–cell contact was largely continuousbut exhibited occasional discontinuities. The adherens junc-tional proteins, �-catenin (Fig. 1J–L) and p100-p120 (data notshown) were also expressed at the cell border, but in a morediffuse pattern characteristic of these molecules. Similar resultswere obtained for the cell line ARPE-19 (data not shown).
Reverse Transcription–PolymeraseChain Reaction
The discriminatory molecules RPE-65, an RPE cell–specific mol-ecule that is thought to play an important role in the RPE-photoreceptor vitamin A cycle, and CRALBP, which is involvedin the regeneration of visual pigment, were both detected byRT-PCR in h1RPE-7, h1RPE-116, and ARPE-19 cell lines produc-ing amplicons of the predicted size (Fig. 3) and correct se-quence.
A panel of growth and trophic factors with potential toaffect both RPE and photoreceptor cell function and survivalwere also investigated by RT-PCR. Positive mRNA expressionwas observed for CNTF, BDNF, bFGF, NGF, PDGF-�, PEDF,IGF-1, and VEGF (Fig. 3). Sequence analysis of all RT-PCRproducts showed amplified sequences were 100% concordantwith published human sequences (data not shown).
Fas and FasL are a receptor-ligand pair involved in theinduction of apoptotic cell death. FasL is a type II transmem-brane protein and a member of the TNF family that binds to Fas
(CD95) and induces apoptosis of the Fas-expressing cells. Theexpression of FasL by grafted RPE cells, therefore, may be acritical determinant in their ability to attenuate the rejectionprocess through FasL-mediated apoptosis of host T cells aftertransplantation. The cell lines h1RPE-7, h1RPE-116, andARPE-19 as well as a third-passage human RPE cell culture,were all found to be negative for FasL mRNA. However, T cellsassessed under identical stringent conditions were clearly pos-itive for FasL (Fig. 4), demonstrating that in vitro, these RPE celllines do not express FasL.
Secretion of Growth and Trophic Factors
ELISAs were performed on serial dilutions of cell culture su-pernatants, conditioned for 5 days, from a minimum of threeindependent experiments. BDNF, bFGF, and VEGF were posi-tively detected in primary RPE, h1RPE-7, h1RPE-116, andARPE-19 cells (Fig. 5). Secretion of bFGF by both h1RPE-7 andh1RPE-116 was 30-fold higher than in primary RPE andARPE-19 cells (P � 0.001; Fig. 5B), whereas secretion of VEGF,was much lower than in primary RPE cells (P � 0.001; Fig. 5C).CNTF, hIFN-�, and hIFN-� were not detected above the limit ofthe assays.
Tumorigenicity Testing
After subcutaneous transplantation of h1RPE-7 or h1RPE-116cells into irradiated athymic mice, neither cell line formedtumors over a 15-week period. Conversely, the human U87glioblastoma cell line formed large tumors by 3 weeks aftertransplantation (data not shown).
The anchorage-independent growth of both h1RPE-7 andh1RPE-116 was determined by growing cells in soft agar. Thesecell lines failed to form aggregates or colony foci over a 40-dayculture period (data not shown), indicating that they are bothnontransformed and nontumorigenic.
FIGURE 2. Electron micrographs of h1RPE-7 cells. (A) Scanning elec-tron micrograph showing microvilli on apical surface. (B) Transmis-sion electron micrograph showing microvilli (arrows). (C–E) Trans-mission electron micrographs depicting junctional specialization(arrowheads) and actin filaments radiating from the junctional region(arrows). Bars, (A) 1 �m; (B–E) 500 nm.
FIGURE 3. RT-PCR analysis of discriminatory molecules and growthand trophic factors in RPE cells. RNA was extracted from confluentflasks of h1RPE-7, h1RPE-116, and ARPE-19 cells. RT was performed on1 �g RNA. Lane 1: commercially supplied marker; lane 2: h1RPE-7;lane 3: h1RPE-116; lane 4: ARPE-19. cDNA (1 �L) was used for all PCRreactions and �-actin was used as a positive control throughout. Se-quencing was undertaken to verify authenticity of all PCR products.
550 Kanuga et al. IOVS, February 2002, Vol. 43, No. 2
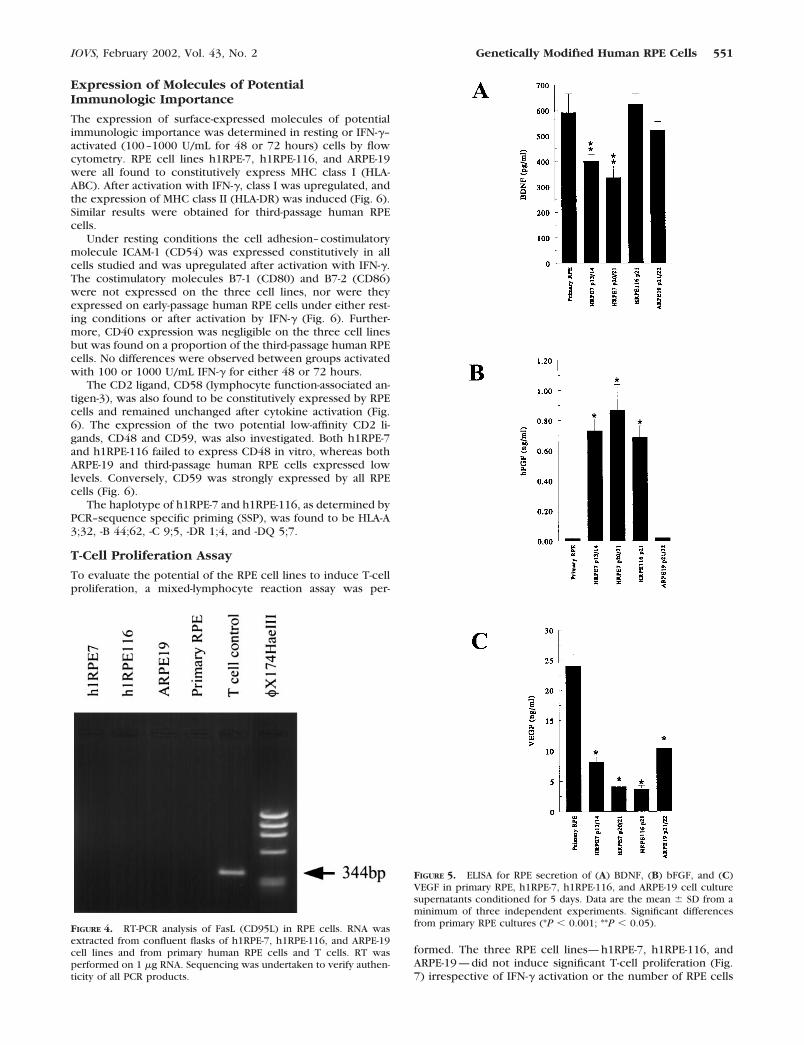
Expression of Molecules of PotentialImmunologic Importance
The expression of surface-expressed molecules of potentialimmunologic importance was determined in resting or IFN-�–activated (100–1000 U/mL for 48 or 72 hours) cells by flowcytometry. RPE cell lines h1RPE-7, h1RPE-116, and ARPE-19were all found to constitutively express MHC class I (HLA-ABC). After activation with IFN-�, class I was upregulated, andthe expression of MHC class II (HLA-DR) was induced (Fig. 6).Similar results were obtained for third-passage human RPEcells.
Under resting conditions the cell adhesion–costimulatorymolecule ICAM-1 (CD54) was expressed constitutively in allcells studied and was upregulated after activation with IFN-�.The costimulatory molecules B7-1 (CD80) and B7-2 (CD86)were not expressed on the three cell lines, nor were theyexpressed on early-passage human RPE cells under either rest-ing conditions or after activation by IFN-� (Fig. 6). Further-more, CD40 expression was negligible on the three cell linesbut was found on a proportion of the third-passage human RPEcells. No differences were observed between groups activatedwith 100 or 1000 U/mL IFN-� for either 48 or 72 hours.
The CD2 ligand, CD58 (lymphocyte function-associated an-tigen-3), was also found to be constitutively expressed by RPEcells and remained unchanged after cytokine activation (Fig.6). The expression of the two potential low-affinity CD2 li-gands, CD48 and CD59, was also investigated. Both h1RPE-7and h1RPE-116 failed to express CD48 in vitro, whereas bothARPE-19 and third-passage human RPE cells expressed lowlevels. Conversely, CD59 was strongly expressed by all RPEcells (Fig. 6).
The haplotype of h1RPE-7 and h1RPE-116, as determined byPCR–sequence specific priming (SSP), was found to be HLA-A3;32, -B 44;62, -C 9;5, -DR 1;4, and -DQ 5;7.
T-Cell Proliferation Assay
To evaluate the potential of the RPE cell lines to induce T-cellproliferation, a mixed-lymphocyte reaction assay was per-
formed. The three RPE cell lines—h1RPE-7, h1RPE-116, andARPE-19—did not induce significant T-cell proliferation (Fig.7) irrespective of IFN-� activation or the number of RPE cells
FIGURE 4. RT-PCR analysis of FasL (CD95L) in RPE cells. RNA wasextracted from confluent flasks of h1RPE-7, h1RPE-116, and ARPE-19cell lines and from primary human RPE cells and T cells. RT wasperformed on 1 �g RNA. Sequencing was undertaken to verify authen-ticity of all PCR products.
FIGURE 5. ELISA for RPE secretion of (A) BDNF, (B) bFGF, and (C)VEGF in primary RPE, h1RPE-7, h1RPE-116, and ARPE-19 cell culturesupernatants conditioned for 5 days. Data are the mean � SD from aminimum of three independent experiments. Significant differencesfrom primary RPE cultures (*P � 0.001; **P � 0.05).
IOVS, February 2002, Vol. 43, No. 2 Genetically Modified Human RPE Cells 551
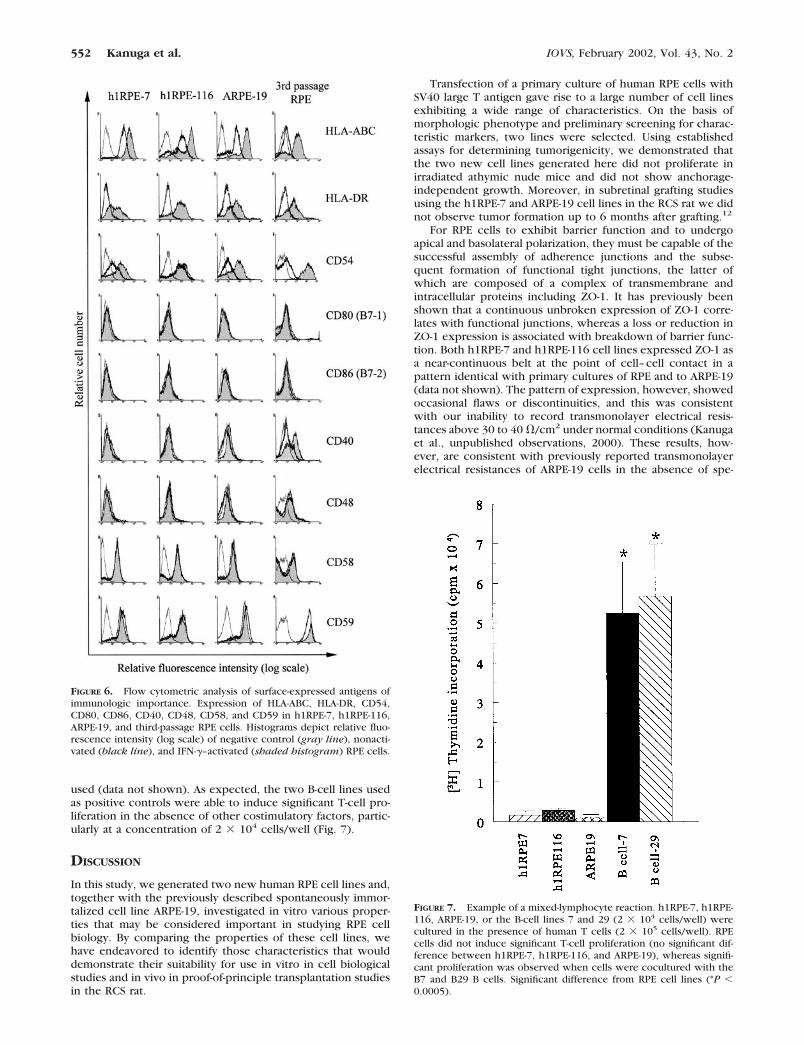
used (data not shown). As expected, the two B-cell lines usedas positive controls were able to induce significant T-cell pro-liferation in the absence of other costimulatory factors, partic-ularly at a concentration of 2 � 104 cells/well (Fig. 7).
DISCUSSION
In this study, we generated two new human RPE cell lines and,together with the previously described spontaneously immor-talized cell line ARPE-19, investigated in vitro various proper-ties that may be considered important in studying RPE cellbiology. By comparing the properties of these cell lines, wehave endeavored to identify those characteristics that woulddemonstrate their suitability for use in vitro in cell biologicalstudies and in vivo in proof-of-principle transplantation studiesin the RCS rat.
Transfection of a primary culture of human RPE cells withSV40 large T antigen gave rise to a large number of cell linesexhibiting a wide range of characteristics. On the basis ofmorphologic phenotype and preliminary screening for charac-teristic markers, two lines were selected. Using establishedassays for determining tumorigenicity, we demonstrated thatthe two new cell lines generated here did not proliferate inirradiated athymic nude mice and did not show anchorage-independent growth. Moreover, in subretinal grafting studiesusing the h1RPE-7 and ARPE-19 cell lines in the RCS rat we didnot observe tumor formation up to 6 months after grafting.12
For RPE cells to exhibit barrier function and to undergoapical and basolateral polarization, they must be capable of thesuccessful assembly of adherence junctions and the subse-quent formation of functional tight junctions, the latter ofwhich are composed of a complex of transmembrane andintracellular proteins including ZO-1. It has previously beenshown that a continuous unbroken expression of ZO-1 corre-lates with functional junctions, whereas a loss or reduction inZO-1 expression is associated with breakdown of barrier func-tion. Both h1RPE-7 and h1RPE-116 cell lines expressed ZO-1 asa near-continuous belt at the point of cell–cell contact in apattern identical with primary cultures of RPE and to ARPE-19(data not shown). The pattern of expression, however, showedoccasional flaws or discontinuities, and this was consistentwith our inability to record transmonolayer electrical resis-tances above 30 to 40 �/cm2 under normal conditions (Kanugaet al., unpublished observations, 2000). These results, how-ever, are consistent with previously reported transmonolayerelectrical resistances of ARPE-19 cells in the absence of spe-
FIGURE 6. Flow cytometric analysis of surface-expressed antigens ofimmunologic importance. Expression of HLA-ABC, HLA-DR, CD54,CD80, CD86, CD40, CD48, CD58, and CD59 in h1RPE-7, h1RPE-116,ARPE-19, and third-passage RPE cells. Histograms depict relative fluo-rescence intensity (log scale) of negative control (gray line), nonacti-vated (black line), and IFN-�–activated (shaded histogram) RPE cells.
FIGURE 7. Example of a mixed-lymphocyte reaction. h1RPE-7, h1RPE-116, ARPE-19, or the B-cell lines 7 and 29 (2 � 104 cells/well) werecultured in the presence of human T cells (2 � 105 cells/well). RPEcells did not induce significant T-cell proliferation (no significant dif-ference between h1RPE-7, h1RPE-116, and ARPE-19), whereas signifi-cant proliferation was observed when cells were cocultured with theB7 and B29 B cells. Significant difference from RPE cell lines (*P �0.0005).
552 Kanuga et al. IOVS, February 2002, Vol. 43, No. 2

cialized media.10 The adherens junction components, p100-p120 and �-catenin, were also found to be strongly expressedin vitro at the point of cell–cell contact in the two cell lines ina manner identical with that observed in primary RPE cells andARPE-19. Although the junctional expression of these mole-cules is a prerequisite for the formation of fully functionaljunctions, their appearance per se does not necessarily in-dicate functional junctions, because this is dependent on thecorrect assembly of the junctional complex as a whole.Although we did not investigate the associated question ofcell polarization in this study, it is interesting to note thatwhen grafted subretinally into RCS rats, both h1RPE-7 andARPE-19 cell lines are able to integrate with the host RPE,synthesize pigment granules, and express them at theirapical face, which suggests some degree of polarization.12
Clearly, further work is required to evaluate the capacity ofthese cells to exhibit functional polarity and the factorscontrolling this process.
The participation of RPE cells in the visual cycle is welldocumented. These cells are engaged in the synthesis andstorage of retinyl esters, isomerization of all-trans-retinoids to11-cis-retinoids and the eventual conversion of retinol to reti-nal. The expression of genes implicated in retinal vitamin Ametabolism is therefore an essential property for any RPE cellline. RPE65, a tissue-specific, highly conserved protein, ispresent at high levels in vivo and is thought to play an impor-tant role in the RPE-photoreceptor vitamin-A cycle.13,14 In thisstudy, we demonstrated mRNA expression of both the full-length and nested transcript of RPE65 in h1RPE-7 and h1RPE-116 cells. We also demonstrated the presence of mRNA forCRALBP in the two cell lines. This important protein is in-volved in the regeneration of visual pigment and has beenimplicated in autosomal recessive retinitis pigmentosa.15 Thefinding that these genes are also expressed in ARPE-19 isconsistent with a previous report.10
Recent observations have dramatically illustrated the effectsof either a single growth and trophic factor or a combination ofthese on photoreceptor viability in dystrophic animal models.In particular bFGF, BDNF, and CNTF have been shown to beneuroprotective in the retina.16–21 The capacity of the RPEcells to produce these factors was therefore investigated andbFGF, BDNF, and CNTF were all expressed at the transcrip-tional level, although only bFGF and BDNF were detected inconditioned supernatant. bFGF secretion by h1RPE-7 andh1RPE-116 cells was found to be significantly higher than thatproduced by primary RPE or ARPE-19 cells and in concentra-tions that are physiologically relevant.22,23 These studies dem-onstrate the capacity of these cells to produce potentiallyimportant neurotrophic factors that may protect photorecep-tors and enhance survival, although it remains to be shownwhether these factors are produced once the cells are graftedinto the retina.
The eye has been described as an immune-privileged site,with its immunity attributed in part to the expression of FasL,particularly at the site of the blood–ocular barriers.24 FasL is atype II integral membrane protein homologous with the TNFreceptor and is important for protection against inflammatoryprocesses. In the eye it has been reported to be constitutivelyexpressed on RPE cells where it promotes apoptosis of invad-ing Fas-positive immune cells.25,26 Thus, FasL expression byRPE cells may play an essential role in local immune regulationand in a transplantation setting in the prevention of immunerejection of transplanted cell lines.27 The in vitro detectionof FasL on RPE, however, has been problematic, because ofreported problems associated with certain FasL antibod-ies.28,29 In immunocytochemical assays we were also unableto show the presence of surface expressed FasL on RPE cells
in vitro (data not shown), and this prompted us to restrictour investigation to the use of RT-PCR for the detection ofFasL. The three cell lines that we studied, as well as theearly-passage human RPE cells, were all found to be negativefor FasL at the transcriptional level. It is likely, therefore,that, as previously reported,29 FasL is expressed only on RPEcells in vivo. It remains to be determined, however, whetherthese RPE cell lines are able to express this importantimmunomodulatory molecule when reintroduced subreti-nally.
The expression of MHC class II molecules on RPE has alsobeen the subject of much conjecture in relation to their abilityto act as antigen-presenting cells. Furthermore, in transplanta-tion studies it has been suggested that RPE allografts are vul-nerable to rejection as a result of the host immune responsebeing directed against transplantation autoantigens.30,31 Usingimmunocytochemical (not shown) and flow cytometric analy-sis, we have shown that the RPE cell lines did not express MHCclass II constitutively but that it was induced after cytokineactivation. These results are consistent with previous reportsdescribing MHC expression in human RPE cells.32 The expres-sion of costimulatory molecules would also determine whetherthese cells are potentially capable of inducing T-cell activation.We found that, under in vitro conditions, the three cell linesexpressed the adhesion–costimulatory molecule ICAM-1(CD54) but did not express B7-1 (CD80) or B7-2 (CD86) ineither resting or IFN-�–activated cells. These results are gener-ally consistent with other studies performed on primary orearly-passage cultures of RPE cells, in which CD54 has beenshown to be expressed,33 but not CD80 and CD86.33–35 How-ever, unlike a previous study in human fetal RPE cells,34 wewere unable to confirm the expression of CD40 on the RPE celllines, although we observed a significant population of positivecells in the early-passage human RPE cells. This may indicatethat the cell lines have lost the capacity to express CD40.
The alternative CD2-mediated pathway for T-cell ac-tivation has been suggested as a possible mechanism bywhich RPE cells may also activate T cells.35 We have shownthat the three cell lines, h1RRPE-7, h1RPE-116, and ARPE-19,express similar levels of the CD2 ligand, CD58, underboth resting conditions and after IFN-� activation. BothCD4836,37 and CD5938,39 have also been proposed as possi-ble low-affinity ligands for human CD2, although their func-tional relevance in humans has been questioned.35,40 How-ever, in the rat, both CD48 and CD59 may act as majorligands for CD235 and, in that these cell lines have beengrafted into rat retina,12 we investigated the expressionof these ligands on the RPE cells. Both CD58 and CD59were expressed constitutively, whereas CD48 was ex-pressed only weakly on ARPE-19 and early-passage RPE cells.It remains unclear whether in the in vivo setting either CD58or CD59 is able to activate T cells through this alternativepathway.
Although the cell lines were found to express MHC class II,the absence of costimulatory molecule expression suggeststhat they would not, at least in vitro, be able to induce T-cellproliferation. This was confirmed in a mixed lymphocyte reac-tion where the cell lines were unable to induce T-cell prolifer-ation unlike a B-cell line. Should these properties be retainedafter transplantation of these cells into the subretinal space, itwould suggest that in an allogeneic setting there would be alimited immune response, thus improving the longevity andviability of grafted cells.
In this study, the cell lines h1RPE-7 and h1RPE-116 retainedmany of the phenotypic features of RPE cells in vivo. Thesuitability of these newly generated cell lines, along with thatof ARPE-19, for in vitro research of RPE function are currently
IOVS, February 2002, Vol. 43, No. 2 Genetically Modified Human RPE Cells 553

under way.41 In addition, we have recently reported that sub-retinal transplantation of these cells in the RCS rat is able topreserve vision as assessed using histologic, behavioral, andphysiological criteria.12,42 These observations highlight thevalue of generating carefully characterized human RPE celllines for both in vitro and in vivo studies.
Acknowledgments
The authors thank Peter Munro for generous help with the electronmicroscopy; Pascal Chaux for valuable critical comment; and JohnHungerford, Phillip Luthert, and Alan Bird (Moorfields Eye Hospital andInstitute of Ophthalmology, London, UK) for their efforts in the pro-vision of human donor tissue.
References
1. Zarbin MA. Age-related macular degeneration: review of pathogen-esis. Eur J Ophthalmol. 1998;8:199–206.
2. Campachario PA, Soloway P, Ryan SJ, Miller JW. The pathogenesisof choroidal neovascularization in patients with age-related macu-lar degeneration. Mol Vis. 1999;5:34; available at http://www.molvis.org/molvis/v5/p34/.
3. Fine SL, Berger JW, Maguire MG, Ho MC. Age-related maculardegeneration. N Engl J Med. 2000;342:483–492.
4. Lund RD, Kwan ASL, Keegan DJ, Sauve Y, Coffey PJ, Lawrence JM.Cell transplantation as a treatment for retinal disease. Prog RetinalEye Res. 2001;20:415–449.
5. Whiteley SJ, Litchfield TM, Coffey PJ, Lund RD. Improvement ofthe pupillary light reflex of Royal College of Surgeons rats follow-ing RPE cell grafts. Exp Neurol. 1996;140:100–104.
6. Li L, Turner JE. Inherited retinal dystrophy in the RCS rat: preven-tion of photoreceptor degeneration by pigment epithelial celltransplantation. Exp Eye Res. 1988;47:911–917.
7. Gouras P, Lopez R, Kjeldbye H, Sullivan B, Brittis M. Transplanta-tion of retinal epithelium prevents photoreceptor degeneration inthe RCS rat. Prog Clin Biol Res. 1989;314:659–671.
8. Little CW, Castillo B, DiLoreto DA, et al. Transplantation of humanfetal retinal pigment epithelium rescues photoreceptor cells fromdegeneration in the Royal College of Surgeons rat retina. InvestOphthalmol Vis Sci. 1996;37:204–211.
9. Sheedlo HJ, Li L, Turner JE. Functional and structural characteris-tics of photoreceptor cells rescued in RPE-cell grafted retinas ofRCS dystrophic rats. Exp Eye Res. 1989;48:841–854.
10. Dunn KC, Aotaki-Keen AE, Putkey FR, Hjelmland LM. ARPE-19, Ahuman retinal pigment epithelial cell line with differentiated prop-erties. Exp Eye Res. 1996;62:155–169.
11. Vicart P, Testut P, Schwartz B, Llorens-Cortes C, Perdomo JJ, PaulinD. Cell adhesion markers are expressed by a stable human endo-thelial cell line transformed by the SV40 large T antigen undervimentin promoter control. J Cell Physiol. 1993;157:41–51.
12. Lund RD, Adamson P, Sauve Y, et al. Subretinal transplantation ofgenetically modified human cell lines attenuates loss of visualfunction in dystrophic rats. Proc Natl Acad Sci USA. 2001;98:9942–9947.
13. Hamel CP, Tslou E, Pfeffer BA, Hooks JJ, Detrick B, Redmond TM.A developmentally regulated microsomal protein specific for thepigment epithelium of vertebrate retina. J Biol Chem. 1993;34:414–425.
14. Redmond TM, Yu S, Lee E, et al. Rpe65 is necessary for productionof 11-cis-vitamin A in the retinal visual cycle. Nat Genet. 1998;20:344–351.
15. Maw MA, Kennedy B, Knight A, et al. Mutation of the geneencoding cellular retinaldehyde-binding protein in autosomal re-cessive retinitis pigmentosa. Nat Genet. 1997;17:198–200.
16. LaVail MM, Unoki K, Yasumara D, Matthes MT, Yanucopoulos GD,Steinberg RH. Multiple growth factors, cytokines and neurotro-phins rescue photoreceptors from the damaging effects of con-stant light. Proc Natl Acad Sci USA. 1992;89:11249–11253.
17. Faktorovich EG, Steinberg RH, Yasumara D, Matthes MT, LaVailMM. Basic fibroblast growth factor and local injury protect photo-
receptors from light damage in the rat. J Neurosci. 1992;12:3554–3567.
18. Hackett SF, Schoenfeld CL, Freund J, Gottsch JD, Bhargarve S,Campochiaro PA. Neurotrophic factors, cytokines and stress in-crease expression of basic fibroblast growth factor in retinal pig-mented epithelial cells. Exp Eye Res. 1997;64:865–873.
19. Unoki K, LaVail MM. Protection of the retina from ischaemic injuryby BDNF, CNTF, and bFGF. Invest Ophthalmol Vis Sci. 1994;35:907–915.
20. Carwile ME, Culbert RB, Sturdivant RL, Kraft TW. Rod outer seg-ment maintenance is enhanced in the presence of bFGF, CNTF andGDNF. Exp Eye Res. 1998;66:791–805.
21. Chong NHV, Alexander RA, Waters L, Barnett KC, Bird AC, LuthertPJ. Repeated injections of a CNTF analogue lead to long-termphotoreceptor survival in hereditary retinal degeneration. InvestOphthalmol Vis Sci. 1999;40:1298–1305.
22. Qian X, Davies AA, Goderie SK, Temple S. FGF2 concentrationregulates the generation of neurons and glia from multipotentcortical stem cells. Neuron. 1997;18:81–93.
23. Desire L, Courtois Y, Jeanny J-C. Endogenous and exogenousfibroblast growth factor 2 support survival of chick retinal neuronsby control of neuronal bcl-xL and bcl-2 expression through afibroblast growth factor receptor 1- and ERK-dependent pathway.J Neurochem. 2000;75:151–163.
24. Griffith TS, Brunner T, Fletcher SM, Green DR, Ferguson TA. Fasligand-induced apoptosis as a mechanism of immune privilege.Science. 1995;270:1189–1192.
25. Jørgensen A, Wiencke AK, la Cour M, et al. Human retinal pigmentepithelial cell-induced apoptosis in activated T cells. Invest Oph-thalmol Vis Sci. 1998;39:1590–1599.
26. Kaplan HJ, Leibole MA, Tezel T, Ferguson TA. Fas ligand (CD95ligand) controls angiogenesis beneath the retina. Nat Med. 1999;5:292–297.
27. Wenkel H, Streilein JW. Evidence that retinal pigment epitheliumfunctions as an immune-privileged tissue. Invest Ophthalmol VisSci. 2000;41:3467–3473.
28. Smith D, Sieg S, Kaplan D. Technical note: aberrant detection ofcell surface Fas ligand with anti-peptide antibodies. J Immunol.1998;160:4159–4160.
29. Kæstel CG, Madsen HO, Prause JU, et al. Lack of FasL expressionin cultured human retinal pigment epithelial cells. Exp Clin Im-munogenet. 2001;18:34–41.
30. Zhang X, Bok D. Transplantation of retinal pigment epithelial cellsand immune response in the subretinal space. Invest OphthalmolVis Sci. 1998;39:1021–1027.
31. Algvere PV, Berglin L, Gouras P, Sheng Y, Kopp ED. Transplanta-tion of RPE in age-related macular degeneration: observations indisciform lesions and dry RPE atrophy. Graefes Arch Clin ExpOphthalmol. 1997;235:149–158.
32. Liversidge JM, Sewell HF, Forrester JV. Human retinal pigmentepithelial cells differentially express MHC class II (HLA, DP, DRand DQ) antigens in response to in vitro stimulation with lympho-kine or purified IFN-gamma. Clin Exp Immunol. 1998;73:489–494.
33. Rezai KA, Semnani RT, Patel SC, Ernest JT, van Seventer GA. Theimmunogenic potential of human fetal retinal pigment epitheliumand its relation to transplantation. Invest Ophthalmol Vis Sci.1997;38:2662–2671.
34. Willermain F, Caspers-Velu L, Baudson N, et al. Role and expres-sion of CD40 on human retinal pigment epithelial cells. InvestOphthalmol Vis Sci. 2000;41:3485–3491.
35. Liversidge J, Dawson R, Hoey S, McKay D, Grabowski P, ForresterJV. CD59 and CD48 expressed by rat retinal pigment epithelialcells are major ligands for the CD2-mediated alternative pathway ofT cell activation. J Immunol. 1996;156:3696–3703.
36. Arulanandam ARN, Moingeon P, Concino MF, et al. A solublemultimeric recombinant CD2 protein identifies CD48 as a lowaffinity ligand for human CD2: Divergence of CD2 ligands duringthe evolution of humans and mice. J Exp Med. 1993;177:1439–1450.
554 Kanuga et al. IOVS, February 2002, Vol. 43, No. 2

37. Sandrin MS, Mouhtouris E, Vaughan HA, Warren HS, Parish CR.CD48 is a low affinity ligand for human CD2. J Immunol. 1993;151:4606–4613.
38. Hahn WC, Menu E, Bothwell AL, Sims PJ, Bierer BE. Overlappingbut nonidentical binding sites on CD2 for CD58 and a secondligand CD59. Science. 1992;256:1805–1807.
39. Deckert M, Kubar J, Bernard A. CD58 and CD59 molecules exhibitpotentializing effects in T cell adhesion and activation. J Immunol.1992;148:672–677.
40. Van der Merwe PA, Barclay AN, Mason DW, et al. Human cell-adhesion molecule CD2 binds CD58 (LFA-3) with a very low
affinity and an extremely fast dissociation rate but does not bindCD48 or CD59. Biochemistry. 1994;33:10149–10160.
41. Zhang J, Sathia PJ, Kanuga N, Aylward GW, Adamson P, Green-wood J. Modification of human retinal pigment epithelial (RPE)cell phenotype following growth on different substrates [ARVOAbstract]. Invest Ophthalmol Vis Sci. 2001;42(4):S758. Abstract nr4065.
42. Coffey PJ, Girman S, Wang SM, et al. Long-term preservation ofcortically dependent visual function in dystrophic RCS rats bytransplantation of immortalized retinal pigment epithelial cells.Nature Neurosci. 2002;5;53–56.
IOVS, February 2002, Vol. 43, No. 2 Genetically Modified Human RPE Cells 555




















