
Characterization in vivo and in vitro of a strain of Leishmania (Viannia) shawi from the Amazon...
7
Characterization in vivo and in vitro of a strain of Leishmania (Viannia) shawi from the Amazon Region Patrícia K.S. Ramos a , José A.P. Diniz a , Edilene O. Silva b, ⁎, Juarez A.S. Quaresma c , Elvira M. Saraiva d , Sérgio H. Seabra e , Geórgia C. Atella f , Wanderley de Souza g a Instituto Evandro Chagas, Unidade de Microscopia Eletrônica, Av. Almirante Barroso 492, Bairro Marco, 66090-000, Belém, Pará, Brazil b Universidade Federal do Pará, Instituto de Ciências Biológicas, Laboratório de Biologia Estrutural, Av. Augusto Corrêa 01, Bairro Guamá, 66075-110, Belém, Pará, Brazil c Universidade Federal do Pará, Núcleo de Medicina Tropical, Av. Generalíssimo Deodoro 92, Bairro Umarizal, 66075-970, Belém, Pará, Brazil d Universidade Federal do Rio de Janeiro, Instituto de Microbiologia Prof. Paulo de Góes, Departamento de Imunologia, Laboratório de Imunobiologia das Leishmanioses, Av. Carlos Chagas Filho 373, Bloco I, sala I2-052, Ilha do Fundão, 21941902, Rio de Janeiro, RJ, Brazil e Centro Universitário da Zona Oeste, Av. Manuel Caldeira de Alvarenga, 1203, Campo Grande, 23070-200 – Rio de Janeiro, RJ, Brazil f Universidade Federal do Rio de Janeiro, Instituto de Bioquímica Médica, Ilha do Fundão, 21941-590, Rio de Janeiro, RJ, Brazil g Universidade Federal do Rio de Janeiro, Instituto de Biofísica Carlos Chagas Filho, Laboratório de Ultraestrutura Celular Hertha Meyer, Ilha do Fundão, 21941-590, Rio de Janeiro, RJ, Brazil abstract article info Article history: Received 18 June 2008 Received in revised form 15 January 2009 Accepted 19 January 2009 Available online 5 February 2009 Keywords: Leishmania (Viannia) shawi Ultrastructure Complement resistance Carbohydrate Susceptibility Amazon Region The present study analyses complement resistance, cell surface carbohydrates expression, lipidic composition and morphology in vivo and in vitro, of Leishmania (Viannia) shawi, a parasite identified in the Amazon region, Pará state, in 1989. We demonstrated that promastigotes in the stationary (STAT) growth phase are more resistant to complement lysis than in the logarithimic (LOG) growth phase. Ultrastructural analyses and imidazol technique showed accumulation of lipids in STAT growth phase promastigotes, which was confirmed by biochemical approach. Light and electron microscopy of skin lesion in hamster footpads caused by promastigotes in STAT growth phase, 90 days post inoculation, showed amastigotes inside of macrophage and free in the tissue surrounded by collagen fibers as well as extensive inflammatory reaction with tissue destruction. We also demonstrated, using lectins by agglutination assays and flow cytometry, the presence of fucose, mannose and/or glucose carbohydrate residues on the surface of LOG and STAT promastigotes. The results constitute the first characterization essay combining biochemical and morphological approaches dedicated to LOG and STAT growth phase promastigotes of L. (V) shawi contributing for a better knowledge of this poorly studied species of the New World. © 2009 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Leishmaniasis comprises a spectrum of diseases found all over the world in tropical and subtropical regions [1]. In the Amazon Region of Brazil, American cutaneous leishmaniasis (ACL) is regarded as a zoonosis found in wild animals, humans and phlebotominae sandflies from the primary forest in the State of Pará, North of Brazil. In this area, the cutaneous disease is caused by the infection of seven recognized species of Leishmania [2]. One of these species, Leishmania (Viannia) shawi, was initially isolated from primates and other silvatic animals and is capable to infect man and to cause disease [3,4]. This specie was distinguished from others by biological, biochemical and serological characters such as morphology, isoenzyme profiles and reactivity with monoclonal antibodies [3]. In addition, both amasti- gote and promastigote forms are smaller than those from other species and subgenus, however, they have a peripylarian development in the sandfly vector. Therefore, every contribution that allows improving the knowledge about this parasite is important, because of its potential to cause clinical manifestations in humans [4]. In view of the growing importance of L. (V.) shawi as an agent of human leishmaniasis in the Amazon region we decided to make a characterization of some basic biological aspects of this organism. In the present study, we characterized logarithmic (LOG) and stationary (STAT) growth phase promastigote forms of L. (V.) shawi considering (a) complement resistance using normal human and guinea pig sera, (b) morphological features recognized by transmission electron microscopy, (c) lectin binding of surface-exposed carbohydrates, (d) total lipid composition, and (e) infectivity to hamsters. Parasitology International 58 (2009) 154–160 ⁎ Corresponding author. Laboratório de Biologia Estrutural, Universidade Federal do Pará, Av. Augusto Corrêa, 01 66075-900, Belém, PA, Brazil. Tel.: +55 91 3201 8232; fax: +55 91 3201 7601. E-mail address: [email protected] (E.O. Silva). 1383-5769/$ – see front matter © 2009 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.parint.2009.01.009 Contents lists available at ScienceDirect Parasitology International journal homepage: www.elsevier.com/locate/parint
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Characterization in vivo and in vitro of a strain of Leishmania (Viannia) shawi from the Amazon...

Parasitology International 58 (2009) 154–160
Contents lists available at ScienceDirect
Parasitology International
j ourna l homepage: www.e lsev ie r.com/ locate /par in t
Characterization in vivo and in vitro of a strain of Leishmania (Viannia) shawi from theAmazon Region
Patrícia K.S. Ramos a, José A.P. Diniz a, Edilene O. Silva b,⁎, Juarez A.S. Quaresma c, Elvira M. Saraiva d,Sérgio H. Seabra e, Geórgia C. Atella f, Wanderley de Souza g
a Instituto Evandro Chagas, Unidade de Microscopia Eletrônica, Av. Almirante Barroso 492, Bairro Marco, 66090-000, Belém, Pará, Brazilb Universidade Federal do Pará, Instituto de Ciências Biológicas, Laboratório de Biologia Estrutural, Av. Augusto Corrêa 01, Bairro Guamá, 66075-110, Belém, Pará, Brazilc Universidade Federal do Pará, Núcleo de Medicina Tropical, Av. Generalíssimo Deodoro 92, Bairro Umarizal, 66075-970, Belém, Pará, Brazild Universidade Federal do Rio de Janeiro, Instituto de Microbiologia Prof. Paulo de Góes, Departamento de Imunologia, Laboratório de Imunobiologia das Leishmanioses, Av. Carlos ChagasFilho 373, Bloco I, sala I2-052, Ilha do Fundão, 21941902, Rio de Janeiro, RJ, Brazile Centro Universitário da Zona Oeste, Av. Manuel Caldeira de Alvarenga, 1203, Campo Grande, 23070-200 – Rio de Janeiro, RJ, Brazilf Universidade Federal do Rio de Janeiro, Instituto de Bioquímica Médica, Ilha do Fundão, 21941-590, Rio de Janeiro, RJ, Brazilg Universidade Federal do Rio de Janeiro, Instituto de Biofísica Carlos Chagas Filho, Laboratório de Ultraestrutura Celular Hertha Meyer, Ilha do Fundão, 21941-590, Rio de Janeiro, RJ, Brazil
⁎ Corresponding author. Laboratório de Biologia EstruPará, Av. Augusto Corrêa, 01 66075-900, Belém, PA, Braz+55 91 3201 7601.
E-mail address: [email protected] (E.O. Silva).
1383-5769/$ – see front matter © 2009 Elsevier Irelanddoi:10.1016/j.parint.2009.01.009
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 June 2008Received in revised form 15 January 2009Accepted 19 January 2009Available online 5 February 2009
Keywords:Leishmania (Viannia) shawiUltrastructureComplement resistanceCarbohydrateSusceptibilityAmazon Region
The present study analyses complement resistance, cell surface carbohydrates expression, lipidiccomposition and morphology in vivo and in vitro, of Leishmania (Viannia) shawi, a parasite identified inthe Amazon region, Pará state, in 1989. We demonstrated that promastigotes in the stationary (STAT) growthphase are more resistant to complement lysis than in the logarithimic (LOG) growth phase. Ultrastructuralanalyses and imidazol technique showed accumulation of lipids in STAT growth phase promastigotes, whichwas confirmed by biochemical approach. Light and electron microscopy of skin lesion in hamster footpadscaused by promastigotes in STAT growth phase, 90 days post inoculation, showed amastigotes inside ofmacrophage and free in the tissue surrounded by collagen fibers as well as extensive inflammatory reactionwith tissue destruction. We also demonstrated, using lectins by agglutination assays and flow cytometry, thepresence of fucose, mannose and/or glucose carbohydrate residues on the surface of LOG and STATpromastigotes. The results constitute the first characterization essay combining biochemical andmorphological approaches dedicated to LOG and STAT growth phase promastigotes of L. (V) shawicontributing for a better knowledge of this poorly studied species of the New World.
© 2009 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Leishmaniasis comprises a spectrum of diseases found all over theworld in tropical and subtropical regions [1]. In the Amazon Region ofBrazil, American cutaneous leishmaniasis (ACL) is regarded as azoonosis found in wild animals, humans and phlebotominae sandfliesfrom the primary forest in the State of Pará, North of Brazil. In thisarea, the cutaneous disease is caused by the infection of sevenrecognized species of Leishmania [2]. One of these species, Leishmania(Viannia) shawi, was initially isolated from primates and other silvatic
tural, Universidade Federal doil. Tel.: +55 91 3201 8232; fax:
Ltd. All rights reserved.
animals and is capable to infect man and to cause disease [3,4]. Thisspecie was distinguished from others by biological, biochemical andserological characters such as morphology, isoenzyme profiles andreactivity with monoclonal antibodies [3]. In addition, both amasti-gote and promastigote forms are smaller than those from otherspecies and subgenus, however, they have a peripylarian developmentin the sandfly vector. Therefore, every contribution that allowsimproving the knowledge about this parasite is important, becauseof its potential to cause clinical manifestations in humans [4].
In view of the growing importance of L. (V.) shawi as an agent ofhuman leishmaniasis in the Amazon region we decided to make acharacterization of some basic biological aspects of this organism. Inthe present study, we characterized logarithmic (LOG) and stationary(STAT) growth phase promastigote forms of L. (V.) shawi considering(a) complement resistance using normal human and guinea pig sera,(b) morphological features recognized by transmission electronmicroscopy, (c) lectin binding of surface-exposed carbohydrates,(d) total lipid composition, and (e) infectivity to hamsters.

155P.K.S. Ramos et al. / Parasitology International 58 (2009) 154–160
2. Materials and methods
2.1. Animals
The hamsters used in our study were obtained from the animalcare facility at the Instituto Evandro Chagas (IEC) and maintained instandard cages. All animals procedures were in accordance with theNational Institutes of Health Guide for the Care and Use of LaboratoryAnimals (2nd Edition, 2002) and Brazilian laws, and were performedin biosafety level 2 facilities. The study was approved by the EthicsCommittee of IEC under the process number 014/2005/CEPAN/IEC/SVS/MS.
2.2. Parasites
Promastigotes of the L. (V.) shawi (MHOM/2001/BR/M19703)were obtained by inoculation into axenic medium of tissue fragmentsobtained from a patient with human cutaneous leishmaniasis in theState of Pará, Brazil and characterized by isoenzyme and monoclonalantibodies [4]. The infectivity of the parasites was maintained byregular passages in hamster (Mesocricetus auratus). The promastigoteswere grown in RPMI medium supplemented with 10% fetal bovineserum at 27 °C for up to 20 days. Parasites were centrifuged (2500 ×g,10 min, 24 °C), washed 3 times in RPMI 1640 and used throughout.
2.3. Hamster infection
Hamsters (Mesocricetus auratus) four weeks old were inoculated inthe right footpad with 2×106 LOG or STAT phases promastigotes andthe footpad thickness was evaluated with a paquimeter every evenweek up to three months after inoculation.
2.4. Histopathology
After 3 months, animals were killed in a CO2 chamber and footpadtissues were washed twice in PBS, fixed in 10% formalin and processedfor histopathology by paraffin inclusion. Blocks were cut at 10 µmthickness in a microtome (Leica Microsystems Inc, Germany),mounted in electrostatic slides and stained with hematoxylin-eosin.
2.5. Transmission electron microscopy
For electron microscopy of parasites grown in axenic cultures orfrom hamster lesions, the cells were rinsed in saline and fixed for60 min at room temperature in a solution containing 2.5% glutar-aldehyde and 5 mM CaCl2 in 0.1M cacodylate buffer, pH 7.2. Afterfixation, the cultures were rinsed in cacodylate buffer, post-fixed in asolution containing 1% osmium tetroxide, 0.8% potassium ferrocya-nide and 5 mM CaCl2, washed in the same buffer, dehydrated inacetone and embedded in Epon. Ultrathin sections were stained withuranyl acetate and lead citrate and observed using a Zeiss EM 900electron microscope.
For lipid localization, promastigotes were fixed in a solutioncontaining 2.5% glutaraldehyde and 2.5% freshly prepared formalde-hyde in 0.1M cacodylate buffer (pH 7.2), washed initially in cacodylatebuffer and then in 0.1 M imidazole buffer, pH 7.2. The parasites werethen post-fixed in a solution containing 2% OsO4 in 0.1 M imidazolebuffer, pH 7.5, for 2 h in dark conditions. After that, the specimenswere washed in imidazole buffer, dehydrated in acetone, andembedded in Epon 812 resin. Thin sections were observed with nocounter stain [5].
2.6. Complement lysis assay
Serum from two healthy volunteers who had never been exposedto Leishmania spp. and a pool of 12 guinea pig sera, were maintained
at−80 °C and used in all experiments. Serawere thawed immediatelybefore use and a twofold serial dilution in PBS containing 0.15 mMCaCl2 and 1 mM MgCl2 in 96-wells plate (TPP) was incubated withpromastigotes from the LOG phase (2 days) or STAT phase (6 days) at2×106 cells/well, and incubated at 37 °C. After 1 h, the plates wereplaced on ice and live parasites were counted in a heamocytometer. Asa control both sera were heat inactivated (30 min, 56 °C) and assayedas above. Results were expressed as the percentage of parasite survivalcompared with controls that were not exposed to sera.
2.7. Lectin agglutination test
Lectin agglutination assays were performed using two-fold serialdilutions of the different lectins in 25 µl of PBS. The same volume of aLOG and STAT phase promastigotes suspension (2×107 cells/ml) wasincubated with the different lectins during 1 h, at room temperature.Agglutination was determined by microscopic observation accordingto [6]. Parasites incubated without lectins were used as control. Theexperiments were repeated at least three times. Lectins fromConcanavalin A (ConA), Ulex europaeus I (UEA-I), Wheat germagglutinin from Triticum vulgaris (WGA), Peanut agglutinin from Ar-achis hypogaea (PNA), Ricinus communis I (RCA-I), soy bean-agglutinin(SBA), Dolichos biflorus-agglutinin and (DBA) (all from Vectorlaboratories, Peterborough, U.K).
2.8. Flow cytometry
Parasite suspensions were incubated with 10 µg/ml of lectin-FITC for 1 h at room temperature. Parasites were then washed,resuspended in PBS and analyzed by flow cytometry using aFACSCAN cytometer with CellQuest (Benckton Dickinson) supportsoftware for acquisition and analysis. The results were analysed byWinMDI 2.8.
2.9. Lipid composition analysis
Promastigotes were obtained from LOG and STAT growth phaseharvested by centrifugation (750 ×g, 15 min) and washed two timeswith phosphate buffered saline. Lipid extraction was performedaccording to [7]. The resulting pellet (108 parasites/ml) was mixedfor 2 h in a stoppered tube using chloroform–methanol–water solution(2:1:0.8, v/v/v), with intermittent agitation for 1 h. The mixture wascentrifuged and the supernatant was collected. The pellet wassubjected to a second lipid extraction (1 h). To the pooled supernatant,1.0 ml of water and 1.0 ml of chloroform were added. The mixture wasvigorously shaken. After centrifugation, the organic phase wasremoved and dried under a stream of nitrogen. Lipids were analyzedby HPTLC on Silica gel 60 plates (60F254, Merck, Darmstadt,Germany). Plates were developed in hexane–ethyl ether–acetic acid(60:40:1, v/v). For lipids visualization HPTLC plates were stained byspraying with Charring reagent [CuSO4 3%, H3PO4 8% (v/v)] andheating at 110 °C for 10 min [8]. The charred HTLC plates were thenanalyzed by densitometry (Image Total Lab software, USA). Each lipidspot was identified by comparison with lipid standards (Sigma AldrichCo., St. Louis, MO, USA), run in parallel.
3. Results
3.1. L. (V.) shawi LOG and STAT phase promastigotes presented differentsensitivity to complement lysis
The strain of L. (V.) shawi used in this work presented welldefined LOG and STAT phases in RPMI 1640 reaching LOG growthphase in the second day and beginning STAT phase at the sixth day(data not shown). To evaluate complement sensitivity of bothpromastigotes growth phases, 2 and 6 days old cultures were
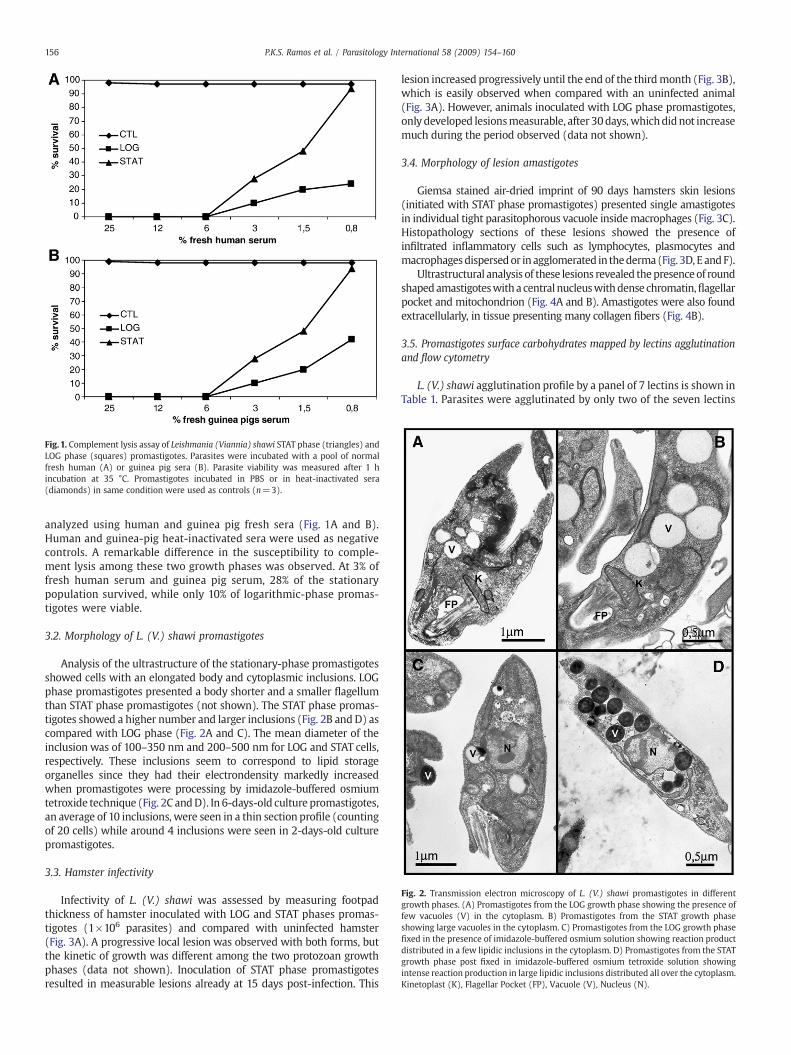
Fig. 1. Complement lysis assay of Leishmania (Viannia) shawi STAT phase (triangles) andLOG phase (squares) promastigotes. Parasites were incubated with a pool of normalfresh human (A) or guinea pig sera (B). Parasite viability was measured after 1 hincubation at 35 °C. Promastigotes incubated in PBS or in heat-inactivated sera(diamonds) in same condition were used as controls (n=3).
Fig. 2. Transmission electron microscopy of L. (V.) shawi promastigotes in differentgrowth phases. (A) Promastigotes from the LOG growth phase showing the presence offew vacuoles (V) in the cytoplasm. B) Promastigotes from the STAT growth phaseshowing large vacuoles in the cytoplasm. C) Promastigotes from the LOG growth phasefixed in the presence of imidazole-buffered osmium solution showing reaction productdistributed in a few lipidic inclusions in the cytoplasm. D) Promastigotes from the STATgrowth phase post fixed in imidazole-buffered osmium tetroxide solution showingintense reaction production in large lipidic inclusions distributed all over the cytoplasm.Kinetoplast (K), Flagellar Pocket (FP), Vacuole (V), Nucleus (N).
156 P.K.S. Ramos et al. / Parasitology International 58 (2009) 154–160
analyzed using human and guinea pig fresh sera (Fig. 1A and B).Human and guinea-pig heat-inactivated sera were used as negativecontrols. A remarkable difference in the susceptibility to comple-ment lysis among these two growth phases was observed. At 3% offresh human serum and guinea pig serum, 28% of the stationarypopulation survived, while only 10% of logarithmic-phase promas-tigotes were viable.
3.2. Morphology of L. (V.) shawi promastigotes
Analysis of the ultrastructure of the stationary-phase promastigotesshowed cells with an elongated body and cytoplasmic inclusions. LOGphase promastigotes presented a body shorter and a smaller flagellumthan STAT phase promastigotes (not shown). The STAT phase promas-tigotes showed a higher number and larger inclusions (Fig. 2B and D) ascompared with LOG phase (Fig. 2A and C). The mean diameter of theinclusion was of 100–350 nm and 200–500 nm for LOG and STAT cells,respectively. These inclusions seem to correspond to lipid storageorganelles since they had their electrondensity markedly increasedwhen promastigotes were processing by imidazole-buffered osmiumtetroxide technique (Fig. 2C andD). In 6-days-old culture promastigotes,an average of 10 inclusions, were seen in a thin section profile (countingof 20 cells) while around 4 inclusions were seen in 2-days-old culturepromastigotes.
3.3. Hamster infectivity
Infectivity of L. (V.) shawi was assessed by measuring footpadthickness of hamster inoculated with LOG and STAT phases promas-tigotes (1×106 parasites) and compared with uninfected hamster(Fig. 3A). A progressive local lesion was observed with both forms, butthe kinetic of growth was different among the two protozoan growthphases (data not shown). Inoculation of STAT phase promastigotesresulted in measurable lesions already at 15 days post-infection. This
lesion increased progressively until the end of the thirdmonth (Fig. 3B),which is easily observed when compared with an uninfected animal(Fig. 3A). However, animals inoculated with LOG phase promastigotes,only developed lesionsmeasurable, after 30days,whichdidnot increasemuch during the period observed (data not shown).
3.4. Morphology of lesion amastigotes
Giemsa stained air-dried imprint of 90 days hamsters skin lesions(initiated with STAT phase promastigotes) presented single amastigotesin individual tight parasitophorous vacuole insidemacrophages (Fig. 3C).Histopathology sections of these lesions showed the presence ofinfiltrated inflammatory cells such as lymphocytes, plasmocytes andmacrophagesdispersedor inagglomerated in thederma(Fig. 3D, E andF).
Ultrastructural analysis of these lesions revealed thepresenceof roundshapedamastigoteswitha central nucleuswithdense chromatin,flagellarpocket and mitochondrion (Fig. 4A and B). Amastigotes were also foundextracellularly, in tissue presenting many collagen fibers (Fig. 4B).
3.5. Promastigotes surface carbohydrates mapped by lectins agglutinationand flow cytometry
L. (V.) shawi agglutination profile by a panel of 7 lectins is shown inTable 1. Parasites were agglutinated by only two of the seven lectins

Fig. 3. Experimental infection in Hamster. A – Usual aspect of the uninfected hamster footpad. B – General view of primary lesion of hamster footpad presenting intense edema(arrow) three months after inoculation with 2×106 promastigotes of STAT growth phase L. (V.) shawi. C – Giemsa stained air dried imprint of the infected skin lesion showingamastigotes loading in individualized vacuoles inside macrophages (arrows). D–F – Histopathology of primary lesion of skin stained with hematoxilin-eosin. The epidermis presentsthe usual aspect with areas of rectifications (D) and derma show the presence of macrophages (arrows) dispersed and some times agglomerated (rectangle) (E–F).
157P.K.S. Ramos et al. / Parasitology International 58 (2009) 154–160
tested: Canavalia ensiformis (CON A), which agglutinated stationaryphase promastigotes with half the concentration used for LOG phasecells, and Ricinus communis I (RCA I), which agglutinated equally bothpromastigotes. All the other tested lectins were unable to agglutinatethe parasites at the highest concentration tested (500 µg/ml).
Flow cytometry analysis showed that lectins from Ulex europaeus-I(L-fucose binding) (Fig. 5A and B) Concanavalin A (D-mannose-binding) (Fig. 5C and D) were able to discriminate between LOGand STAT phase promastigotes. Both lectins labeled a higher numberof LOG phase promastigotes. Lectins from Helix pomatia, Griffoniasimplicifolia I (D-GalNAc binding), Euonymus europaeus (D-Gal
Fig. 4. Transmission electron microscopy of hamster footpad lesion three months after inamastigote (arrow) in the tissue. B – Amastigote is observed free and circunded by intense tecFlagellar Pocket, M mitochondrion, CF Collagen Fibers.
binding), Limax flavus (sialic acid binding) bound with low affinityto both, LOG and STAT phase promastigotes (data not shown).
3.6. Lipid composition analysis
The analysis of composition of the lipids of L. (V.) shawi showedthat the parasites contained a large variety of associated neutral lipids.The triacylglycerol (TG) and cholesteryl ester (CE) were the mainneutral lipids, corresponding to 34%, 37% (STAT growth phase) and29%, 27% (LOG growth phase). Although in an insignificant amounts,free fatty acid (FA) and an undetermined lipid (UDL) were also
fection with promastigotes from STAT growth phase of L. (V.) shawi. A – Leishmaniaidual destruction and collagen fibers deposition. N nucleus, NHC Nucleus of Host Cell, FP
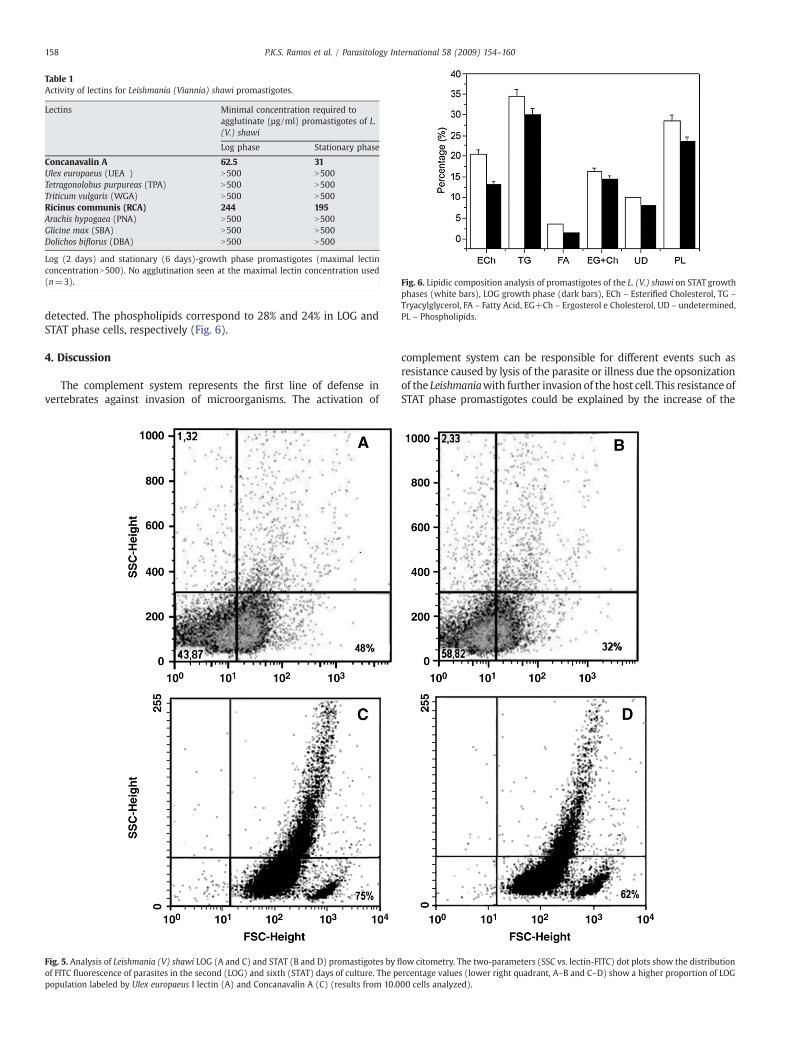
Table 1Activity of lectins for Leishmania (Viannia) shawi promastigotes.
Lectins Minimal concentration required toagglutinate (µg/ml) promastigotes of L.(V.) shawi
Log phase Stationary phase
Concanavalin A 62.5 31Ulex europaeus (UEA ) N500 N500Tetragonolobus purpureas (TPA) N500 N500Triticum vulgaris (WGA) N500 N500Ricinus communis (RCA) 244 195Arachis hypogaea (PNA) N500 N500Glicine max (SBA) N500 N500Dolichos biflorus (DBA) N500 N500
Log (2 days) and stationary (6 days)-growth phase promastigotes (maximal lectinconcentrationN500). No agglutination seen at the maximal lectin concentration used(n=3). Fig. 6. Lipidic composition analysis of promastigotes of the L. (V.) shawi on STAT growth
phases (white bars), LOG growth phase (dark bars), ECh – Esterified Cholesterol, TG –
Tryacylglycerol, FA – Fatty Acid, EG+Ch – Ergosterol e Cholesterol, UD – undetermined,PL – Phospholipids.
158 P.K.S. Ramos et al. / Parasitology International 58 (2009) 154–160
detected. The phospholipids correspond to 28% and 24% in LOG andSTAT phase cells, respectively (Fig. 6).
4. Discussion
The complement system represents the first line of defense invertebrates against invasion of microorganisms. The activation of
Fig. 5. Analysis of Leishmania (V) shawi LOG (A and C) and STAT (B and D) promastigotes by fl
of FITC fluorescence of parasites in the second (LOG) and sixth (STAT) days of culture. The ppopulation labeled by Ulex europaeus I lectin (A) and Concanavalin A (C) (results from 10.0
complement system can be responsible for different events such asresistance caused by lysis of the parasite or illness due the opsonizationof the Leishmaniawith further invasion of the host cell. This resistance ofSTAT phase promastigotes could be explained by the increase of the
ow citometry. The two-parameters (SSC vs. lectin-FITC) dot plots show the distributionercentage values (lower right quadrant, A–B and C–D) show a higher proportion of LOG00 cells analyzed).

159P.K.S. Ramos et al. / Parasitology International 58 (2009) 154–160
surface-exposed lipophosphoglycan (LPG) in this phase of developmentthat prevents the access of the C5b-9 membrane attack complex to theparasite membrane [9–11]. On the other hand, LPG activates comple-ment and promotes the binding of opsonizing fragments, allowingattachment and invasion in macrophage [12].
Thus, we consider interesting to compare complement sensitivityof LOG and STAT phase parasites. Our results demonstrated a time-dependent complement resistance of the promastigotes throughoutthe growth curve. These results are similar to those reported for otherLeishmania species showing that parasites from the STAT phase weremore resistant to the complement than promastigotes of LOG phase[9,3–15,11,16]. However, Noronha et al. (1998) [17] analyzing differentspecies of subgenera Leishmania and Viannia showed that parasites ofthe last subgenus do not express any resistance to complement duringall stages of their growth in vitro. These results are not in agreementwith our present observation, since that L (V.) shawi is resistant to thelytic action of complement during STAT growth. In agreement withour results, Pinto-da-Silva et al. (2002) [11] also find complementresistance by a member of subgenus Viannia, L. (V.) braziliensis.Further studies will be necessary to characterize other species fromViannia subgenus found in the Amazon region.
Evaluation of the infectivity of these two phases of developmentwas carried out in vivo. The lesions produced by promastigotes of STATgrowth phase were greater than LOG growth phase. These findingsshowed that the behavior of this strain is similar to that observed in otherspecies of the Viannia subgenus [18–19]. Recent studies with L. (V.)lainsoni, a closer specie of L. (V.) shawi from Amazon region, in exper-imental infected hamster showed the same results presentedhere, exceptfor the absence of collagen deposition around the amastigotes [19].
The importance of carbohydrates on cell surface of Leishmania hasbeen pointed out in previous reports [6,16,20–22]. The cell surfacecarbohydrates play an important role in the survival of Leishmania insidemacrophage and during development in the digestive tract of the vector[21]. In view of the importance of carbohydrates to Leishmania, wedecided to perform a comparative analysis of the cell surfacecarbohydrate residues in LOG and STAT phases cells. Our results showedthat the L. (V.) shawi was agglutinated by Con A and RCA, lectins thatbind to mannose and galactose residues, respectively. The STAT phasecells were agglutinated with a lower concentration of Con A than LOGphase cells indicating a greater number of residues of mannose on itssurface. However, this result was not confirmed by flow cytometryanalyses. Our results are similar to those found by some authors. Groglet al. (1987) [14] using STAT growth phase of L. (V.) braziliensis andNagakura et al. (1986) [23] using the same species. Andrade and Saraiva(1999) [22] analyzing cell-surface carbohydrates of different Leishmaniaspecies, using only STAT phase promastigotes, showed that promasti-gotes of L. donovani were agglutinated only by Con A.
An interesting aspect found in this report was that results obtainedby flow cytometry analysis. The lectin Ulex europeaus-I, which bindsfucose residues, was able to bind the surface of LOG and STAT phasecells, but its expression slightly decreased in stationary phase whencompared with log phase promastigotes. This finding suggests thepresence of fucose residues on promastigote surface recognized byUlex europeaus-I. Bandyopadhyay et al. (1991) [16] also demonstratedthe presence of fucose residues on Leishmania (L.) major usingagglutination tests. However, we only detected the presence of fucoseon the L. (V.) shawi by cytometry analysis what can be due to the highsensibility of this technique. Some authors reported agglutination ofLeishmania species by PNA, a lectin that binds galactose residues[21,24]. However, PNA did not agglutinate L. shawi.
Lipids are essential and highly abundant components of allorganisms. The lipid metabolism is important to the virulence, growth,replication anddifferentiation of Leishmania species in the host [25]. Theneutral lipids, as triacylglycerol and steryl esters, have been proposed toact as fatty acid reservoirs in trypanosomatids [26–27]. Our presentobservations showed that both LOGandSTATpromastigotes presented a
large number of cytoplasmic inclusions resembling structures contain-ing lipids. Cytochemical analysis confirmed the presence of lipids sincethe inclusions became electrondense when the cells were fixed in animidazole-buffered osmium tetroxide solution [5]. We also noticed thatSTAT promastigotes contained a larger number of the lipid inclusionsthan LOGpromastigotes. In addition, themeandiameter of the inclusionwas larger in STAT promastigotes. The biochemical analysis also showeda higher fatty acid concentration in the STAT phase than LOG phase cellssuggesting that this accumulation is essential to the multiplication andhost-parasite interaction. It is possible that accumulation of lipids bySTAT promastigotes is necessary to keep these cells, presumablyinfective promastigotes, metabolically active until their penetrationinto macrophages from the vertebrate host.
In conclusion, these studies showed that: (a) STAT growth phasepromastigotes of L. (V.) shawi was resistant to complement-mediatedlysis. (b) LOG and STAT phases displays characteristic features such asthe presence of fucose residues on its surface, that could be used todistinguish it from other Leishmania species of the NewWorld, and (c)accumulate lipids as cytoplasmic storage organelles.
Acknowledgements
We thank Dr. Fernando Tobias Silveira (Instituto Evandro Chagas,Belém, Pará, Brazil) for fruitful discussions and interaction andRaimundo Nonato Barbosa Pires, Antonio F.P. Martins and João AlvesBrandão for their technical assistance and animal care. This work wassupported by Programa Nacional de Cooperação Acadêmica (PROCAD/CAPES), Instituto Evandro Chagas, SEDECT/FAPESPA/MS/CNPq/UFPA/FADESP (Project n° 110/2004), MCT/CNPq/MS-SCTIE-DECIT 25/2006and Programa de Núcleos de Excelência (PRONEX) from CNPq/FAPERJ.
References
[1] WHO. Weekly epidemiological record, 77; 2002. p. 365–72.[2] Silveira FT, Ishikawa EAY, De Souza AAA, Lainson R. An outbreak of cutaneous
leishmaniasis among soldiers in Belém, Pará State, Brazil, caused by Leishmania(Viannia) lindenbergi n. sp. A new leishmanial parasite of man in the Amazonregion. Parasite 2002;9:43–50.
[3] Lainson R, Braga RR, De Souza, Povoa MM, Shaw JJ, Ishikawa EA, Silveira FT.Leishmania (Viannia) shawi sp. N., A parasite of monkeys, sloths and procyonids inAmazonian Brazil. Ann Parasitol Hum Comp 1989;65:200–7.
[4] Shaw JJ, Ishikawa EAY, Lainson R, Braga RR, Silveira FT. Cutaneous Leishmaniasis ofman due to Leishmania (Viannia) shawi Lainson, De Souza, Póvoa, Ishikawa &Silveira in Pará State, Brazil. Ann Parasitol Hum Comp 1991;66:243–6.
[5] Angermuller S, Fahimi HD. Imidazole-buffered osmium tetroxide: an excellentstain for visualization of lipids by transmission electron microscopy. Histochem J1982;14:823–35.
[6] Saraiva EMB, Andrade AFB, Pereira MEA. Cell surface carbohydrate of L. mexicanaamazonensis: differences between infective and non-infective forms. Eur J Cell Biol1986;40:219–25.
[7] Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. CanJ Biochem Physiol 1959;37:911–7.
[8] Ruiz JL, Ochoa B. Quantification in the subnanomolar range of phospholipids andneutral lipids by nanodimensional thin layer chromatography and image analysis.J Lipid Res 1997;38:1482–9.
[9] Puentes SM, Sacks DL, Da Silva RP, Joiner KA. Complement binding by twodevelopmental stages of L. major promastigotes varying in expression of a surfacelipophosphoglycan. J Exp Med 1988;167:887–902.
[10] Puentes SM, Da Silva RP, Sacks DL, Hammer CH, Joiner KA. Serum resistance ofmetacyclic stage of L. major promastigotes is due to release of C5-9. J Immunol1990;145:4311–6.
[11] Pinto-da-Silva LH, Camurate M, Costa KA, Oliveira SMP, Cunha-e-Silva NL, SaraivaEM. Leishmania (Viannia) braziliensis metacyclic promastigotes purified usingBauhinia purpurea lectins are complement resistant and highly infective formacrophages in vitro and hamsters in vivo. Int J Parasitol 2002;32:1371–7.
[12] Sacks DL. Metacyclogenesis in Leishmania promastigotes. Exp Parasitol 1989;69:100–3.[13] Franke KD, McGreevy PB, Katz SP, Sacks DL. Growth cycle-dependent generation of
complement-resistant Leishmania promastigotes. J Immunol 1985;134:2713–8.[14] Grogl M, Franke ED, McGreevy PB, Kuhn RE. Leishmania braziliensis: protein,
carbohydrate, and antigen differences between log phase and stationary phasepromastigotes in vitro. Exp Parasitol 1987;83:352–9.
[15] Howard MK, Sayers G, Miles MA. L. donovani metacyclic promastigotes: transforma-tion invitro, lectin agglutination, complement resistance and infectivity. ExpParasitol1987;64:147–79.
[16] Bandyopadhyay P, Ghosh DK, Aloke D, Ghosh KN, Chaudhuri PP, Das P, et al.Metacyclogenesis of Leishmania spp: species-specific in vitro transformation,

160 P.K.S. Ramos et al. / Parasitology International 58 (2009) 154–160
complement resistance, and cell surface carbohydrate and protein profiles. JParasitol 1991;77:411–6.
[17] Noronha FSM, Nunes AC, Souza KT, Melo MN, Ramalho-Pinto FJ. Differentialsensitivity of new World Leishmania spp. promastigotes to complement-mediatedlysis: Correlation with the expression of three parasite polypeptides. Acta Tropica1998;69:17–9.
[18] Lainson R, Shaw JJ, Souza AA, Silveira FT, Falqueto A. Further observations onLutzomyia ubiquitalis (Psychodidae: Phlebotominae), the sandfly vector of Leish-mania (Viannia) lainsoni. Mem Inst Oswaldo Cruz 1992;87:437–9.
[19] Correa JR, Soares MJ. Leishmania (Viannia) Lainsoni (Silveira et al., 1987):ultrastructural aspects of the parasite and skin lesion in experimental infectedhamster (Mesocricetus auratus). Parasitol Res 2007;100:1227–32.
[20] Sacks DL, Hieny S, Sher A. Identification of cell surface carbohydrate and antigenicchanges between noninfective and infective stages f L.major promastigotes.J Immunol 1985;135:564–9.
[21] Saraiva EM, Pimenta PFP, Brodin TN, Rowton E, Modi GB, Sacks DL. Changes inlipophosphoglycan and gene expression associated with the development ofLeishmania major in Phlebotomus papatasi. Parasitology 1995;111:275–87.
[22] Andrade AF, Saraiva EM. Lectin-binding properties of different Leishmania species.Parasitol Res 1999;85:576–81.
[23] Nagakura K, Tachibana H, Kaneda Y, Nakae T. Leishmania braziliensis: localizationof glycoproteins in promastigotes. Exp Parasitol 1986;61:335–42.
[24] Schottelius J. Lectin binding strain specific carbohydrates of the cell surfaces ofLeishmaniastrains from the old world. Z Parasitenkd 1982;66:237–47.
[25] Zufferey R, Mamoun CB. The initial step of glycerolipid metabolism in Leishmaniamajor promastigotes involves a single glycerol-3-phosphate acyltransferaseenzyme important for the synthesis of triacylglycerol but not essential forvirulence. Mol Microbiol 2005;56:800–10.
[26] Dixon H,Williamson J. The lipid composition of blood and culture forms of T. lewisiand T. rhodesiense compared with that of their environment. Comp BiochemPhysiol 1970;33:111–28.
[27] Florin-Christensen M, Florin-Christensen J, De Isola ED, Lammel E, Meinardi E,Brenner RR, et al. Temperature acclimation of Trypanosoma cruzi epimastigote andmetacyclic trypomastigote lipids. Mol Biochem Parasitol 1997;88:25–33.




















