
Characterization and isolation of some genes of the shikimate pathway in sensitive and resistant...
9
This article was published in an Elsevier journal. The attached copy is furnished to the author for non-commercial research and education use, including for instruction at the author’s institution, sharing with colleagues and providing to institution administration. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Characterization and isolation of some genes of the shikimate pathway in sensitive and resistant...

This article was published in an Elsevier journal. The attached copyis furnished to the author for non-commercial research and
education use, including for instruction at the author’s institution,sharing with colleagues and providing to institution administration.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Characterization and isolation of some genes of the shikimatepathway in sensitive and resistant Centaurea jacea plants
after ozone exposure
A. Francini, C. Nali*, E. Pellegrini, G. Lorenzini
Dipartimento di Coltivazione e Difesa delle Specie Legnose ‘‘Giovanni Scaramuzzi’’, University of Pisa, Via del Borghetto 80, 56124 Pisa, Italy
Received 6 July 2007; accepted 8 July 2007
Characterization and isolation of PAL and CHS genes in Centaurea jacea exposed to O3.
Abstract
Centaurea jacea has been suggested as a potential bioindicator for ozone, but little is known about its intraspecific variation in sensitivity,especially at molecular level. The effects of ozone (200 ppb, 5 h) on sensitive and resistant lines of Centaurea have been investigated at the endof fumigation. Sensitive plants showed characteristic symptoms of injury in the form of diffuse discoloration stipples on leaves. A PCR-basedapproach was used to identify and isolate a partial-length cDNA coding for PAL and CHS genes. The northern analysis of PAL showed accu-mulation of transcript in both lines correlated with a typical increase of PAL activity (þ41 and þ91% in resistant and sensitive material, re-spectively, compared to controls). On the contrary, the transcripts of CHS, in resistant and sensitive plants, did not change after treatment.Total phenols were not affected by ozone, while anthocyanins were quickly utilised by resistant clone as antioxidant compounds.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Air pollution; Anthocyanins; Chalcone synthase; Phenols; Phenylalanine ammonia-lyase
1. Introduction
In all Europe, and especially in the Mediterranean basin,there is growing evidence of an increase in the concentrationin the troposphere of photochemically-produced ozone (O3);it is expected that it will continue to rise in the coming years.These expectances grow the interest in the effects of O3 onwild plants, a relatively poorly known topic.
Centaurea jacea L. (brown knapweed) is an herbaceousperennial plant widely distributed in grasslands; due to itsbroad climatic adaptation, this species is indigenous inmany European countries (Wagenitz, 1987). In recent years,biomonitoring network of the ICP Vegetation (United
Nations Economic Commission for Europe International Co-operative Programme on effects of air pollution on naturalvegetation and crops, http://icpvegetation.ceh.ac.uk/) hasused sensitive and resistant lines of brown knapweed as bio-monitoring system but, at the moment, these plants areknown only from a morphological and phenological pointof view.
Field observations of foliar symptoms have revealed the co-existence of O3 sensitive and tolerant individuals of C. jacea insingle populations (Bungener et al., 2003). However, it wasnot clear whether the observed differences were due tosmall-scale variations in environmental conditions or to mo-lecular variation, which may express phenotypic plasticitythat again depends upon environmental factors.
Plants respond to O3 stress by induction of several defencereactions. The active mechanisms of defence include increaseof transcripts of genes of the phenylpropanoid pathway such as
* Corresponding author. Tel.: þ39 050 221 0552; fax: þ39 050 221 0559.
E-mail address: [email protected] (C. Nali).
0269-7491/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.envpol.2007.07.007
Available online at www.sciencedirect.com
Environmental Pollution 151 (2008) 272e279www.elsevier.com/locate/envpol

Author's personal copy
those encoding phenylalanine ammonia-lyase (PAL, EC4.3.1.5) and chalcone synthase (CHS, EC 2.3.1.74) (Eckey-Kaltenbach et al., 1997). The first reaction of this biosyntheticpathway is the deamination of phenylalanine to cinnamic acidby PAL. In the next reaction, CHS condenses three malonyl-CoA molecules with cinnamoyl-CoA to produce chalcone.This condensation is the main branch in the pathway for theproduction of flavonoids (Ellis et al., 1989).
PAL, the key enzyme of phenylpropanoid pathway, seemsto be one of the preferential targets of O3 action in plants(Paakkonen et al., 1998). PAL appears to exist universally inhigher plants as a family of genes and the presence of PAL iso-forms is a common observation. Various structural and regula-tory properties of PAL genes from bean, parsley, raspberry,etc. have been reported (Logemann et al., 1995; Kumar andEllis, 2001; Paolacci et al., 2001).
The phenolic compounds produced by the phenylpropanoidpathway are powerful antioxidants in plant tissues under stressconditions. They are chemically heterogeneous compounds,such as flavonoids, lignins and tannins, which play a varietyof roles, e.g. defence against herbivores and pathogens, to ab-sorb high energy radiations and to reduce the growth of nearbycompeting plants (Taiz and Zeiger, 2002). Among flavonoids,the anthocyanins are highly water soluble pigments that accu-mulate in the vacuole (Chalker-Scott, 1999). Common injurysymptoms of the Asteraceae, such as C. jacea, show bronzestipples or leaf discoloration after only few days of field expo-sure to elevate O3 (Bungener et al., 2003) that are due to theproduction of anthocyanins (Koukol and Dugger, 1967). An-thocyanins production is beneficial to the plants in terms ofprotection of shade adapted chloroplasts from brief exposureto high solar radiation (Gould et al., 2000), protection ofleaves from photo-oxidative damage during senescence (Fieldet al., 2001). Furthermore, anthocyanins accumulate underUV-B (Steyn et al., 2002), drought (Balakumar et al., 1993),low temperature (Krol et al., 1995) and exposure to O3 (Footet al., 1996).
We report here the results about young C. jacea plants,one resistant and one sensitive to O3, exposed to this pollut-ant. In order to understand the metabolic and molecular re-sponses to O3, we identified genes encoding for keyenzymes of the phenylpropanoid metabolism (PAL andCHS), analysing also the specific activity of PAL and theamounts of secondary metabolites, such as total phenolsand anthocyanins.
2. Materials and methods
2.1. Plant material and cultivation
Two lines of C. jacea used in the ICP vegetation biomonitoring have pre-
viously been indicated as resistant (R) and sensitive (S ) (Hayes et al., 2006).
Seeds were sown in petri dishes equipped with three layers of moistened filter
paper. To improve germination rates, seeds were previously exposed to a 5-day
pre-chilling period at 4e6 �C under moist conditions. Germination took place
after approximately three days. The seedlings were transplanted into small
pots containing a mixture of peat and sand, and later, were transplanted into
(1 dm3) pot and placed in a controlled environment facility at a temperature
of 20 � 1 �C, a RH of 85 � 5% and a photon flux density at plant height of
500 mmol photons m�2 s�1 provided by incandescent lamps, during a 12 h
photoperiod. As most natural plants grow on soils that are nutrient-limited,
no fertilizers were added.
2.2. Ozone exposure
Uniform-sized plants (one month old) were placed in a controlled environ-
ment fumigation facility (Nali et al., 2005) under the same climatic conditions
as the growth chamber. Fumigation was performed in two Perspex chambers,
each measuring 0.90 � 0.90 � 0.65 m, continuously ventilated with charcoal-
filtered air (two complete air changes/min). O3 was generated by electrical dis-
charge using a Fisher 500 air-cooled apparatus (Zurich, Switzerland) supplied
with pure oxygen, and mixed with the inlet air entering the fumigation cham-
bers. Its concentration at plant height was continuously monitored with a photo-
metric analyzer (Monitor Labs, mod. 8810, San Diego, CA, USA) connected to
a computer. Plants were exposed to 200 ppb O3 (for O3, 1 ppb ¼ 1.96 mg m�3,
at 20 �C and 101.325 kPa) for 5 h in form of a square wave. Control plants were
exposed only to charcoal-filtered air in two more Perspex chambers identical to
those above mentioned.
Samples of leaves of both treated and control plants, were collected at the
end of the fumigation, frozen immediately in liquid N2 and kept at �80 �C un-
til needed.
2.3. Nucleic acid isolation
Total RNA was isolated from C. jacea leaf tissue using phenol-SDS extrac-
tion and lithium chloride precipitation based on the method by Verwoerd et al.
(1989). Extraction was performed in 0.1 M LiCl, 0.1 M TriseHCl (pH 8.0),
10 mM EDTA, 1% (w/v) SDS, and 50% (v/v) phenol:chloroform. After extrac-
tion, the RNA was precipitated in 2 M LiCl at 4 �C.
2.4. Reverse transcriptase-polymerase chainreaction (RT-PCR)
First-strand cDNA was prepared from 5 mg of total RNA using an oligo
(dT) primer and Superscript reverse transcriptase (Invitrogen, Carlsbad, CA).
The reaction was incubated at 37 �C for 1 h and stopped by heating to 94 �Cfor 10 min.
To amplify fragments that encode for PAL and CHS, RT-PCR was
performed using degenerate oligonucleotide primers designed to conserved
amino acid regions in other species obtained from GenBank (http://
www.ncbi.nlm.nih.gov/BLAST/).
Degenerate primers used for PAL and CHS genes isolation were respec-
tively: forward 50-CARTGGYTNGGNCCNCARATH-30; reverse 50-RCANGG
RTCRTCDATRTAN-30 and forward 50-ACATGCCTGGYGCYGAYTAYCA-
30; reverse 50-ACRCATGCRCTTGACATGTTTCCRTA-30; where Y ¼ C,T;
R ¼ A,G; H ¼ A,T,C; and N ¼ C,G,T,A.
PCR products were amplified from 300 to 500 ng of first-strand cDNA us-
ing the appropriate primers and Taq DNA polymerase (Hybaid, Ashford, UK).
All oligonucleotide primers were obtained from MWG-Biotech (Ebers-
berg, Germany). Reactions to amplify PAL and CHS were cycled at 94 �Cfor 1 min, 50 �C for 1 min and 30 s and 72 �C for 2 min (40 cycles).
2.5. Cloning and sequencing of RT-PCR products
The RT-PCR fragments obtained using the degenerate PAL and CHS
primers were cloned into pCR� 2.1-TOPO vector (Invitrogen) following the
manufacturer’s instructions. Plasmids were sequenced by BMR Universita
degli studi di Padova e CRIBI (Italy) and identified using the BLASTx
algorithm (Altschul et al., 1990).
2.6. Northern analysis
Denatured total RNA (20 mg) from leaves was separated at 100 V for 2e3 h in 1% (w/v) agarose gel containing 0.22 M formaldehyde and transferred
273A. Francini et al. / Environmental Pollution 151 (2008) 272e279

Author's personal copy
to BrightStar Plus membrane (Ambion, Italy) as described by Ingelbrecht et al.
(1998). Probe was labelled using biotin following the procedures described in
the Bright Star Psoralen-Biotin non-isotopic labelling kit manual (Ambion).
Hybridization, was approximately performed for 16e18 h, then the mem-
branes were washed in 2 � SSC (0.3 M NaCl/3 mM sodium citrate pH 7.2/
0.1% (w/v) SDS) at 65 �C for 15 min followed by a further two washes in
1 � SSC/0.1% SDS and 0.1 � SSC/0.1% SDS. Signals detection were deter-
mined using the Bright Star Bio-Detection system (Ambion) following the
protocol.
2.7. PAL activity, phenolic and anthocyanin content
The PAL activity was assayed in leaves (1 g) ground in pre-chilled
mortar with liquid nitrogen. The powder was immediately added to 3 ml
of extraction buffer containing 100 mM potassium borate (pH 8.8), with
14 mM 2-mercaptoethanol. The homogenate was then centrifuged at
13,000 � g for 30 min, and the supernatant was used as enzymatic extract.
The PAL assay was determined using a reaction mixture containing 100 ml
of enzymatic extract, 1 ml of 100 mM potassium borate buffer (pH 8.8)
and 200 ml of 100 mM L-phenylalanine. The reaction was incubated at
40 �C for 120 min, the reaction was stopped by adding 50 ml of 5 N
HCl. The cinnamic acid produced was measured at 290 nm. The molar
extinction coefficient for transcinnamic acid was determined to be
17,400 M�1 cm�1 (Cheng and Breen, 1991). Proteins were determined
according to Bensadoun and Weinstein (1976), using bovine serum albu-
min as standard. Total phenols were extracted from leaves according to
Gorinsteina et al. (2004) and estimated as by Singleton and Rossi
(1965). Fresh leaves were extracted in boiling 80% ethanol. Phenolic
extract was mixed with equal volume of 1 M Folin Ciocalteau and
1.5 ml of 7.5% of Na2CO3. Absorbance of the blue coloured solution
was measured at 760 nm. Total phenols were estimated by comparison
with the standard curve obtained with gallic acid.
Anthocyanins were extracted into methanolic HCl following the standard
AOAC method (Harborne, 1967) and the concentration of cyanidin-3-
glucoside equivalents (3 29,600) determined spectrophotometrically at
535 nm.
2.8. Statistics
A minimum of three plants per treatment was used in each of the three re-
peated experiments. Data was analyzed using two-way analysis of variance
(ANOVA) and comparison among means was determined by LSD post-test
(P � 0.05). Analyses were performed by NCSS 2000 Statistical Analysis
System Software.
3. Results
3.1. Isolation and sequence analysis of PAL and CHSfrom Centaurea jacea
RT-PCR, using degenerate PAL and CHS primers, was per-formed to amplify products from C. jacea resistant and sensitiveRNA. These primers correspond to two highly conserved re-gions of these genes found in other plants. Products ofapproximately 680 and 630 bp were amplified respectively forPAL and CHS (Fig. 1). Both products were cloned and, foreach gene, four of the positives were sequenced. Sequence anal-ysis indicated that all PAL and CHS clones were identical, witha length of 672 and 622 bp respectively (PAL; GenBank Acces-sion n� EF112475; CHS, GenBank Accession n� EF112474).
The sequences obtained were subsequently subjected toBLAST database searches at both a nucleic acid and aminoacid level. The results indicated that the isolated inserts en-codes a polypeptide with highest sequence identity to a numberof known PAL and CHS in plants. The most significantmatches are shown in Tables 1 and 2, while sequence identityto homologous sequences of PAL and CHS genes are alsoshown for comparative purposes.
The deduced amino acid sequence of the cloned fragments(named CjPAL1 and CjCHS1) are shown aligned with otherknown PAL and CHS genes in Fig. 2A,B. They illustratethat there are regions of homology between our sequencesand that found in other plants.
Blast analysis of CjPAL1 amino acid sequences (226 aa)showed a putative conserved domain PAL-HAL (phenylala-nine ammonia-lyase and histidine ammonia). It belongs to ly-ase class I_like superfamily of enzymes that catalyze similarbeta-elimination reactions and are active as homotetramers.
A phylogenetic tree was constructed based on multiplealignment of various available plant PAL and CHS amino
Table 1
Percentage sequence identities (at amino acid level) of the putative chalcone
synthase (partial-length) with other plants CHS
GenBank accession no. Organism Identity %
CAA86218 Gerbera hybrida 184/200 92
ABF69124 Chrysanthemum � morifolium 183/200 91
AAB72091 Vitis vinifera 179/200 89
AAQ19322 Triticum aestivum 175/200 87
AAK49457 Nicotiana tabacum 173/200 86
Sequence identities were calculated using the EMBOSS programme.
Table 2
Percentage sequence identities (at amino acid level) of the putative phenylal-
anine ammonia-lyase (partial-length) with other plants PAL
GenBank accession no. Organism Identity %
AAL55242 Lactuca sativa 222/226 98
CAA73065 Helianthus annuus 218/226 96
AAZ04477 Coffea canephora 208/226 92
AAQ19322 Arabidopsis thaliana 199/226 88
AAK49457 Nicotiana tabacum 173/200 86
Sequence identities were calculated using the EMBOSS programme.
L A B C D
679
622
Fig. 1. RT-PCR products amplified from Centaurea jacea total RNA using the
degenerate PAL and CHS primers. (L) Molecular size marker (1 Kb DNA lad-
der); (A and B) RT-PCR PAL products (679 bp) in Centaurea jacea resistant
and sensitive respectively; (C and D) RT-PCR CHS products (622 bp) in
Centaurea jacea resistant and sensitive respectively.
274 A. Francini et al. / Environmental Pollution 151 (2008) 272e279

Author's personal copy
acid sequences, thus allowing the relationships between differ-ent homologies to be examined (Figs. 3 and 4).
3.2. Expression analysis of chalcone synthase andphenilalanine ammonia-lyase under O3 fumigation
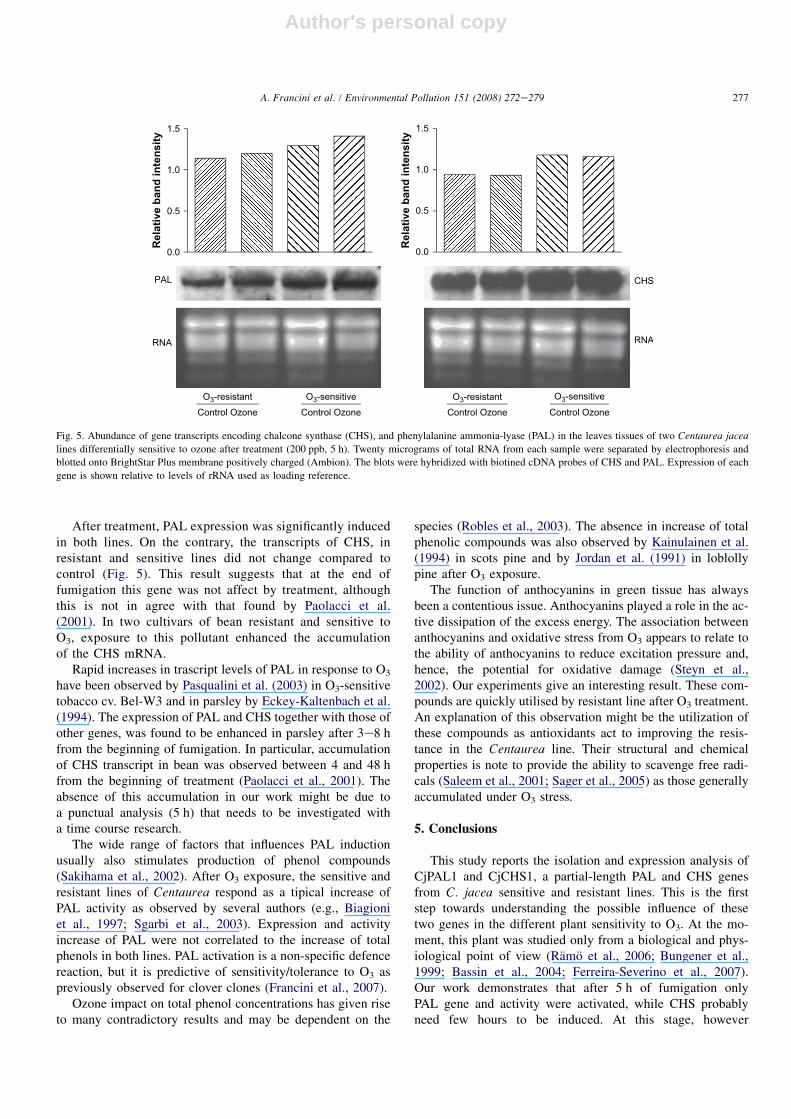
The biotined probes synthesized from the cDNA were usedin northern blots of total RNA extracted from sensitive and re-sistant treated and control plants. After 5 h of O3 exposureCjPAL1 transcripts increased in both lines of Centaurea(Fig. 5). On the contrary, CjCHS1 gene expression did notchange in both lines. In particular, the constitutive gene ex-pression levels of PAL and CHS in sensitive plants werehigher compared to the resistant one (Fig. 5).
3.3. PAL activity, anthocyanins and total phenols
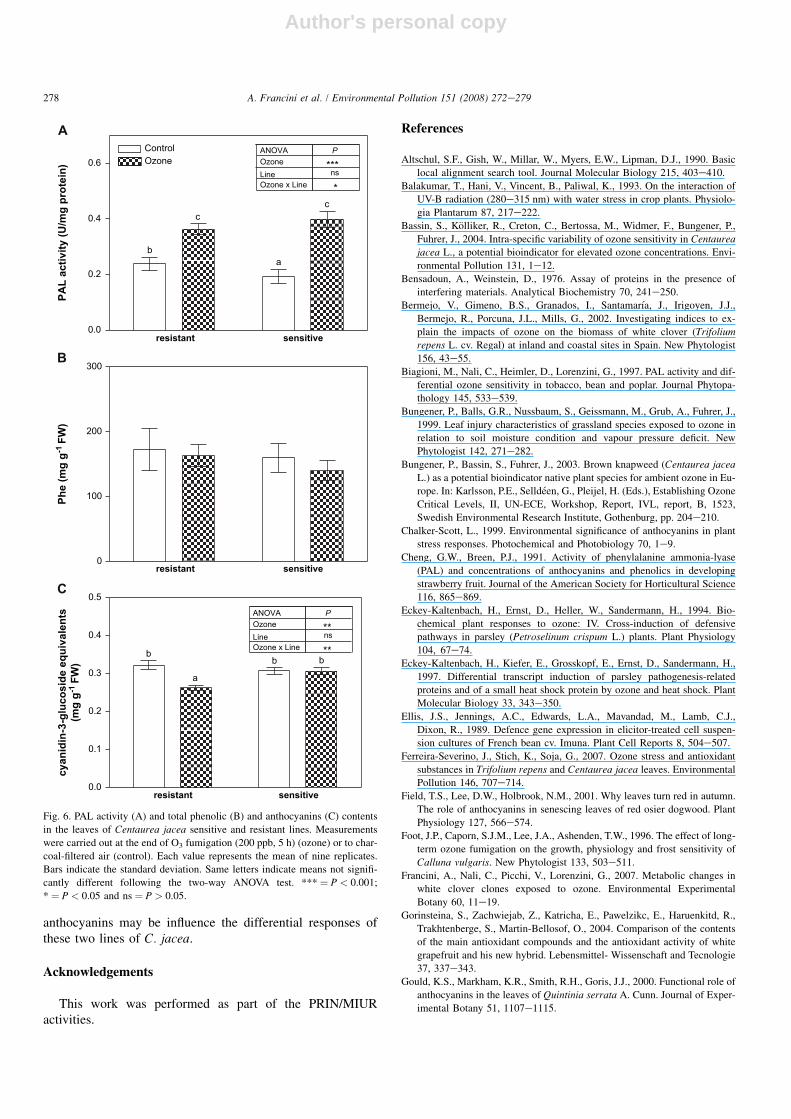
After O3 exposure, the sensitive line showed characteristicsymptoms of injury in the form of diffuse discoloration stip-ples on leaves. The O3 effect was also studied at biochemicaland physiological level, monitoring the PAL activity and sub-strates derived from its activation. Significant differences ofPAL activity were detected between the constitutive levels ofsensitive and resistant untreated plants (Fig. 6A). The treat-ment increased PAL activity in both lines (þ41 and 91% inresistant and sensitive plants, respectively, in comparisonwith controls). Total phenolic compounds content did notshow significant intra-line differences in treated and untreatedmaterial (Fig. 6B). The anthocyanins content significantly
B
A
Fig. 2. Sequence alignment of CjPAL1 (A) and CjCHS1 (B) with other phenylalanina ammonia-lyase and chalcone synthase. Sequences were aligned using the
CLUSTALW program (http://bioweb.pasteur.fr). *, fully conserved residues;., residues no conserved. GenBank accession nos. for PAL: Arabidopsis thalianaAAQ19322, Lactuca sativa AAL55242, Prunus avium AAC78457, Trifolium pratense BAE71252. GenBank accession nos. for CHS: Brassica napus
AAC31911, Cannabis sativa AAL92879, Gerbera hybrida CAA86218, Ruta graveolens CAC14059.
275A. Francini et al. / Environmental Pollution 151 (2008) 272e279

Author's personal copy
decreased by 20% in the resistant line after the fumigation,while did not show any change in the sensitive one (Fig. 6C).
4. Discussion
The increasing interest in understanding the differentialresponses of wild plants to O3 exposure induced scientists toinvestigate in this field, trying to identify the most O3-sensitivetaxa. For more than two decades, the framework of a UN/ECEInternational Cooperative Programme (ICP-Vegetation) for theConvention on Long-Range Transboundary Air Pollution, hasstudied the impacts of air pollutants (especially O3) on cropsand semi-natural vegetation.
Ozone-sensitive and resistant biotypes of white clover (Tri-folium repens cv. Regal) are currently used in this program, butalso studied in numerous works (e.g., Bermejo et al., 2002;Nali et al., 2004; Francini et al., 2007). An analogousapproach has been recently introduced in the UN-ECEprogramme using brown knapweed as bioindicator. The
utilization of this plant was already suggested by Bungeneret al. (1999, 2003). Visible injury in plain field commonlyused in mapping the occurrence of harmful O3 concentrationsneeded to be verify in controlled conditions especially whenplants are very poorly know as the case of C. jacea.
The phenomenological differential behaviour whensubjected to O3 stress of sensitive plants was confirmed inour experiment by formation of characteristic symptoms of in-jury in the form of diffuse discoloration stipples. The intraspe-cific variation that induced resistance and sensitivity inC. jacea under O3 treatment were studied for the first timein this work starting to a biomolecular approach.
The present report demonstrates the successful isolation,cloning and expression analysis of a putative partial-lengthC. jacea PAL and CHS genes. The isolated fragments weresubsequently cloned and sequenced. The sequences obtainedshowed highest homology to a number of PAL and CHS,thus suggesting that the isolated partial-length clone are aputative PAL and CHS genes.
Nucleotide Substitutions (x100)0
19.224681012141618
C.chinensisChrysanthemum x morifoliumChrysanthemum nankingenseGerbera hybridaCjCHS1Brassica napusCitrus sinensisRuta graveolensH.vulgareSorghum bicolorCannabis sativaJuglans nigra x Juglans regiaHypericum androsaemumAnthurium andraeanumIpomoea trifidaIpomoea nilScutellaria baicalensis
Fig. 4. Phylogenetic tree showing the relationships between chalcone synthase isolated in Centaurea jacea (CjCHS1) and sequences of chalcone synthase genes
isolated in other plant species using Clustal W alignment (slow/accurate, Gonnet).
Nucleotide Substitutions (x100)0
22.35101520
CjPAL1Lactuca sativaDaucus carotaPetroselinum crispumIpomoea batatasLycopersicon esculentumDigitalis lanataAstragalus membranaceusTrifolium pratenseUlmus americanaCamellia sinensisPrunus persicaPrunus aviumCoffea canephoraVitis viniferaArabidopsis thaliana
Fig. 3. Phylogenetic tree showing the relationships between phenylalanine ammonia lyase isolated in Centaurea jacea (CjPAL1) and sequences of phenylalanine
ammonia lyase genes isolated in other plant species using Clustal W alignment (slow/accurate, Gonnet).
276 A. Francini et al. / Environmental Pollution 151 (2008) 272e279
Author's personal copy
After treatment, PAL expression was significantly inducedin both lines. On the contrary, the transcripts of CHS, inresistant and sensitive lines did not change compared tocontrol (Fig. 5). This result suggests that at the end offumigation this gene was not affect by treatment, althoughthis is not in agree with that found by Paolacci et al.(2001). In two cultivars of bean resistant and sensitive toO3, exposure to this pollutant enhanced the accumulationof the CHS mRNA.
Rapid increases in trascript levels of PAL in response to O3
have been observed by Pasqualini et al. (2003) in O3-sensitivetobacco cv. Bel-W3 and in parsley by Eckey-Kaltenbach et al.(1994). The expression of PAL and CHS together with those ofother genes, was found to be enhanced in parsley after 3e8 hfrom the beginning of fumigation. In particular, accumulationof CHS transcript in bean was observed between 4 and 48 hfrom the beginning of treatment (Paolacci et al., 2001). Theabsence of this accumulation in our work might be due toa punctual analysis (5 h) that needs to be investigated witha time course research.
The wide range of factors that influences PAL inductionusually also stimulates production of phenol compounds(Sakihama et al., 2002). After O3 exposure, the sensitive andresistant lines of Centaurea respond as a tipical increase ofPAL activity as observed by several authors (e.g., Biagioniet al., 1997; Sgarbi et al., 2003). Expression and activityincrease of PAL were not correlated to the increase of totalphenols in both lines. PAL activation is a non-specific defencereaction, but it is predictive of sensitivity/tolerance to O3 aspreviously observed for clover clones (Francini et al., 2007).
Ozone impact on total phenol concentrations has given riseto many contradictory results and may be dependent on the
species (Robles et al., 2003). The absence in increase of totalphenolic compounds was also observed by Kainulainen et al.(1994) in scots pine and by Jordan et al. (1991) in loblollypine after O3 exposure.
The function of anthocyanins in green tissue has alwaysbeen a contentious issue. Anthocyanins played a role in the ac-tive dissipation of the excess energy. The association betweenanthocyanins and oxidative stress from O3 appears to relate tothe ability of anthocyanins to reduce excitation pressure and,hence, the potential for oxidative damage (Steyn et al.,2002). Our experiments give an interesting result. These com-pounds are quickly utilised by resistant line after O3 treatment.An explanation of this observation might be the utilization ofthese compounds as antioxidants act to improving the resis-tance in the Centaurea line. Their structural and chemicalproperties is note to provide the ability to scavenge free radi-cals (Saleem et al., 2001; Sager et al., 2005) as those generallyaccumulated under O3 stress.
5. Conclusions
This study reports the isolation and expression analysis ofCjPAL1 and CjCHS1, a partial-length PAL and CHS genesfrom C. jacea sensitive and resistant lines. This is the firststep towards understanding the possible influence of thesetwo genes in the different plant sensitivity to O3. At the mo-ment, this plant was studied only from a biological and phys-iological point of view (Ramo et al., 2006; Bungener et al.,1999; Bassin et al., 2004; Ferreira-Severino et al., 2007).Our work demonstrates that after 5 h of fumigation onlyPAL gene and activity were activated, while CHS probablyneed few hours to be induced. At this stage, however
0.0
0.5
1.0
1.5
0.0
0.5
1.0
1.5
O3-resistantControl Ozone Control OzoneControl Ozone Control Ozone
O3-sensitiveO3-resistant O3-sensitive
PAL
RNA
Relative b
an
d in
ten
sity
Relative b
an
d in
ten
sity
CHS
RNA
Fig. 5. Abundance of gene transcripts encoding chalcone synthase (CHS), and phenylalanine ammonia-lyase (PAL) in the leaves tissues of two Centaurea jacea
lines differentially sensitive to ozone after treatment (200 ppb, 5 h). Twenty micrograms of total RNA from each sample were separated by electrophoresis and
blotted onto BrightStar Plus membrane positively charged (Ambion). The blots were hybridized with biotined cDNA probes of CHS and PAL. Expression of each
gene is shown relative to levels of rRNA used as loading reference.
277A. Francini et al. / Environmental Pollution 151 (2008) 272e279
Author's personal copy
anthocyanins may be influence the differential responses ofthese two lines of C. jacea.
Acknowledgements
This work was performed as part of the PRIN/MIURactivities.
References
Altschul, S.F., Gish, W., Millar, W., Myers, E.W., Lipman, D.J., 1990. Basic
local alignment search tool. Journal Molecular Biology 215, 403e410.
Balakumar, T., Hani, V., Vincent, B., Paliwal, K., 1993. On the interaction of
UV-B radiation (280e315 nm) with water stress in crop plants. Physiolo-
gia Plantarum 87, 217e222.
Bassin, S., Kolliker, R., Creton, C., Bertossa, M., Widmer, F., Bungener, P.,
Fuhrer, J., 2004. Intra-specific variability of ozone sensitivity in Centaureajacea L., a potential bioindicator for elevated ozone concentrations. Envi-
ronmental Pollution 131, 1e12.
Bensadoun, A., Weinstein, D., 1976. Assay of proteins in the presence of
interfering materials. Analytical Biochemistry 70, 241e250.
Bermejo, V., Gimeno, B.S., Granados, I., Santamarıa, J., Irigoyen, J.J.,
Bermejo, R., Porcuna, J.L., Mills, G., 2002. Investigating indices to ex-
plain the impacts of ozone on the biomass of white clover (Trifoliumrepens L. cv. Regal) at inland and coastal sites in Spain. New Phytologist
156, 43e55.
Biagioni, M., Nali, C., Heimler, D., Lorenzini, G., 1997. PAL activity and dif-
ferential ozone sensitivity in tobacco, bean and poplar. Journal Phytopa-
thology 145, 533e539.
Bungener, P., Balls, G.R., Nussbaum, S., Geissmann, M., Grub, A., Fuhrer, J.,
1999. Leaf injury characteristics of grassland species exposed to ozone in
relation to soil moisture condition and vapour pressure deficit. New
Phytologist 142, 271e282.
Bungener, P., Bassin, S., Fuhrer, J., 2003. Brown knapweed (Centaurea jacea
L.) as a potential bioindicator native plant species for ambient ozone in Eu-
rope. In: Karlsson, P.E., Selldeen, G., Pleijel, H. (Eds.), Establishing Ozone
Critical Levels, II, UN-ECE, Workshop, Report, IVL, report, B, 1523,
Swedish Environmental Research Institute, Gothenburg, pp. 204e210.
Chalker-Scott, L., 1999. Environmental significance of anthocyanins in plant
stress responses. Photochemical and Photobiology 70, 1e9.
Cheng, G.W., Breen, P.J., 1991. Activity of phenylalanine ammonia-lyase
(PAL) and concentrations of anthocyanins and phenolics in developing
strawberry fruit. Journal of the American Society for Horticultural Science
116, 865e869.
Eckey-Kaltenbach, H., Ernst, D., Heller, W., Sandermann, H., 1994. Bio-
chemical plant responses to ozone: IV. Cross-induction of defensive
pathways in parsley (Petroselinum crispum L.) plants. Plant Physiology
104, 67e74.
Eckey-Kaltenbach, H., Kiefer, E., Grosskopf, E., Ernst, D., Sandermann, H.,
1997. Differential transcript induction of parsley pathogenesis-related
proteins and of a small heat shock protein by ozone and heat shock. Plant
Molecular Biology 33, 343e350.
Ellis, J.S., Jennings, A.C., Edwards, L.A., Mavandad, M., Lamb, C.J.,
Dixon, R., 1989. Defence gene expression in elicitor-treated cell suspen-
sion cultures of French bean cv. Imuna. Plant Cell Reports 8, 504e507.
Ferreira-Severino, J., Stich, K., Soja, G., 2007. Ozone stress and antioxidant
substances in Trifolium repens and Centaurea jacea leaves. Environmental
Pollution 146, 707e714.
Field, T.S., Lee, D.W., Holbrook, N.M., 2001. Why leaves turn red in autumn.
The role of anthocyanins in senescing leaves of red osier dogwood. Plant
Physiology 127, 566e574.
Foot, J.P., Caporn, S.J.M., Lee, J.A., Ashenden, T.W., 1996. The effect of long-
term ozone fumigation on the growth, physiology and frost sensitivity of
Calluna vulgaris. New Phytologist 133, 503e511.
Francini, A., Nali, C., Picchi, V., Lorenzini, G., 2007. Metabolic changes in
white clover clones exposed to ozone. Environmental Experimental
Botany 60, 11e19.
Gorinsteina, S., Zachwiejab, Z., Katricha, E., Pawelzikc, E., Haruenkitd, R.,
Trakhtenberge, S., Martin-Bellosof, O., 2004. Comparison of the contents
of the main antioxidant compounds and the antioxidant activity of white
grapefruit and his new hybrid. Lebensmittel- Wissenschaft and Tecnologie
37, 337e343.
Gould, K.S., Markham, K.R., Smith, R.H., Goris, J.J., 2000. Functional role of
anthocyanins in the leaves of Quintinia serrata A. Cunn. Journal of Exper-
imental Botany 51, 1107e1115.
resistant sensitive
0.0
0.2
0.4
0.6ControlOzone
ab
cc
Ozone x LinensLine
***OzonePANOVA
PA
L activity (U
/m
g p
ro
tein
)
resistant sensitive
0.0
0.1
0.2
0.3
0.4
0.5
a
bb b
cyan
id
in
-3-g
lu
co
sid
e eq
uivalen
ts
(m
g g
-1 F
W)
resistant sensitive
0
100
200
300
Ph
e (m
g g
-1 F
W)
*
Ozone x LinensLine**OzonePANOVA
**
A
B
C
Fig. 6. PAL activity (A) and total phenolic (B) and anthocyanins (C) contents
in the leaves of Centaurea jacea sensitive and resistant lines. Measurements
were carried out at the end of O3 fumigation (200 ppb, 5 h) (ozone) or to char-
coal-filtered air (control). Each value represents the mean of nine replicates.
Bars indicate the standard deviation. Same letters indicate means not signifi-
cantly different following the two-way ANOVA test. *** ¼ P < 0.001;
* ¼ P < 0.05 and ns ¼ P > 0.05.
278 A. Francini et al. / Environmental Pollution 151 (2008) 272e279

Author's personal copy
Harborne, J., 1967. Comparative Biochemistry of the Flavonoids. Academic
Press, New York, 383 pp.
Hayes, F., Mills, G., Harmens, H., Novak, K., Williams, P., 2006. ICP Vegeta-
tion Experimental Protocol for Monitoring the Incidences of Ozone Injury
on Vegetation. Natural Environment Research Council, 28 pp.
Ingelbrecht, I.L., Mandelbaum, C.I., Mirkov, T.E., 1998. Highly sensitive
northern hybridization using a rapid protocol for downward alkaline blot-
ting of RNA. Biotechniques 25, 420e425.
Jordan, D.N., Green, T.H., Chappelka, A.H., Lockaby, B.G., Meldahl, R.S.,
Gjerstad, D.H., 1991. Response of total tannins and phenolics in loblolly
pine foliage exposed to ozone and acid rain. Journal Chemistry Ecology
17, 505e513.
Kainulainen, P., Holopainen, J.K., Hyttinen, H., Oksanen, J., 1994. Effect of
ozone on the biochemistry and aphid infestation of scots pine. Phytochem-
istry 35, 39e42.
Koukol, J., Dugger, W.M., 1967. Anthocyanin formation as a response to
ozone and smog treatment in Rumex crispus L. Plant Physiology 42,
1023e1024.
Krol, M., Gray, G.R., Hurry, V.M., O’Quist, G., Malek, L., Huner, N.P.A.,
1995. Low-temperature stress and photoperiod affect an increased toler-
ance to photoinhibition in Pinus banksiana seedlings. Canadian Journal
Botany 73, 1119e1127.
Kumar, A., Ellis, B.E., 2001. The phenylalanine ammonia-lyase gene family in
Rubus idaeus: structure, expression and evolution. Plant Physiology 127,
230e239.
Logemann, E., Parniske, M., Hahlbrock, K., 1995. Modes of expression and
common structural features of the complete phenylalanine ammonia-lyase
gene family in parsley. Proceedings of the National Academy of Sciences
USA 92, 5905e5909.
Nali, C., Crocicchi, L., Lorenzini, G., 2004. Plants indicators of urban pollu-
tion (ozone and trace elements) in Pisa. Italy. Journal Environmental Mon-
itoring 6, 636e645.
Nali, C., Pucciariello, C., Mills, G., Lorenzini, G., 2005. On the different sen-
sitivity of white clower clones to ozone: physiological and biochemical pa-
rameters in a multivariate approach. Water Air and Soil Pollution 164,
137e153.
Paakkonen, E., Seppanen, S., Holopainen, T., Kokko, H., Karenlampi, S.,
Karenlampi, L., Kangasjarvi, J., 1998. Induction of genes for the stress
proteins PR-10 and PAL in relation to growth, visible injuries and stomatal
conductance in birch (Betula pendula) clones exposed to ozone and/or
drought. New Phytologist 138, 295e305.
Paolacci, A.R., D’Ovidio, R., Marabottini, R., Nali, C., Lorenzini, G.,
Abenavoli, M.R., Badiani, M., 2001. Ozone induces a differential accumu-
lation of phenylalanine ammonia-lyase, chalcone synthase and chalcone
isomerase RNA transcripts in sensitive and resistant bean cultivars. Austra-
lian Journal of Plant Physiology 28, 425e428.
Pasqualini, S., Piccioni, C., Reale, L., Ederli, L., Della Torre, G., Ferranti, F.,
2003. Ozone-induced cell death in tobacco cultivar Bel W3 plants. The
role of programmed cell death in lesion formation. Plant Physiology
133, 1122e1134.
Ramo, K., Slotte, H., Kanerva, T., Ojanpera, K., Manninen, S., 2006. Growth
and visible injuries of four Centaurea jacea L., ecotypes exposed to ele-
vated ozone and carbon dioxide. Environmental and Experimental Botany
58, 287e298.
Robles, C., Greff, S., Pasqualini, V., Garzino, S., Bousquet-Melou, A.,
Fernandes, C., Korboulewsky, N., Bonin, G., 2003. Phenols and flavonoids
in Aleppo pine needles as bioindicators of air pollution. Journal of
Environmental Quality 32, 2265e2271.
Sager, E.P.S., Hutchinson, T.C., Croley, T.R., 2005. Foliar phenolics in sugar
maple (Acer saccharum) as a potential indicator of tropospheric ozone
pollution. Environmental Monitoring and Assessment 105, 419e430.
Sakihama, Y., Cohen, M.F., Grace, S.C., Yamasaki, H., 2002. Plant phenolic
antioxidant and prooxidant activities: phenolics-induced oxidative damage
mediated by metals in plants. Toxicology 177, 67e80.
Saleem, A., Loponen, J., Pihlaja, K., Oksanen, E., 2001. Effects of long-term
open field ozone exposure on leaf phenolics of European silver birch
(Betula pendula ROTH). Journal of Chemical Ecology 27, 1049e1062.
Sgarbi, E., Baroni Fornasiero, R., Lins, A.P., Medeghini Bonatti, P., 2003. Phe-
nol metabolism differentially affected by ozone in two cell lines from
grape (Vitis vinifera L.) leaf. Plant Science 165, 951e957.
Singleton, V.L., Rossi, J.A., 1965. Colorimetry of total phenolics and phospho-
molybdic-phosphotungstic acid reagents. American Journal of Enology
and Viticulture 16, 144e148.
Steyn, W.J., Wand, S.J.E., Holcroft, D.M., Jacobs, G., 2002. Anthocyanins in
vegetative tissues: a proposed unified function in photoprotection. New
Phytologist 155, 349e361.
Taiz, L., Zeiger, E., 2002. Sinauer Associates, Inc. In: Plant Physiology, third
ed. Publishers, Sunderland, Massachusetts, pp. 283e308.
Verwoerd, C., Dekker, B.M.M., Hoekema, A., 1989. A small-scale procedure for
the rapid isolation of plant RNAS. Nucleic Acid Research 17, 2362e2363.
Wagenitz, G., 1987. Centaurea. In: Hegi, G. (Ed.), Illustrierte Flora von
Mitteleuropa. Blackwell, Berlin, pp. 1405e1413.
279A. Francini et al. / Environmental Pollution 151 (2008) 272e279




















