
Preparation of PLLA/PEG Nanofibers by Electrospinning and Potential Applications
Upload
independentCategory
view
2download
0
Plant Science 162 (2002) 273–282
Changes in dehydrin expression associated with cold, ABA andPEG treatments in blueberry cell cultures
Cecile M. Parmentier-Line 1, Ganesh R. Panta 2, Lisa J. Rowland *Fruit Laboratory, USDA-ARS, Henry A. Wallace Belts�ille Agricultural Research Center, 10300 Baltimore A�e., Bldg. 010A, Belts�ille,
MD 20705, USA
Received 22 May 2001; received in revised form 15 October 2001; accepted 25 October 2001
Abstract
Cell cultures of highbush blueberry cultivar ‘Gulfcoast’ (Vaccinium corymbosum L. x Vaccinium darrowi Camp) were used todetermine if tissue cultures could be used as a surrogate for whole plants in studying dehydrin expression. Cell clusters in liquidmedium were subjected to treatments known to induce dehydrins in whole plants, and dehydrin protein and RNA levels weremonitored. Two dehydrins of 65 and 30 kDa were detected with a polyclonal antibody raised against the 65 kDa dehydrin ofblueberry. Using a full-length cDNA clone of blueberry dehydrin 1 as a probe, one mRNA of 0.75 kb, an appropriate size toencode the 30 kDa dehydrin, was detected on RNA blots. When cells were grown at 4 °C, abundance of the 30 kDa dehydrinincreased, but the 65 kDa dehydrin did not. Level of the 0.75 kb message increased temporarily then decreased with coldtreatment. Disparities between protein and transcript levels suggested that post-transcriptional mechanisms were involved inregulation of dehydrin expression. Some of the abscisic acid (ABA) concentrations tested induced expression of both dehydrins.Polyethylene glycol in the cultures repressed expression of both dehydrins and of the 0.75 kb mRNA. Results obtained from cellculture differed in too many ways from those on whole plants to make cell culture suitable for studying dehydrin expression inblueberry. © 2002 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Blueberry; Dehydrin; In vitro; Vaccinium
www.elsevier.com/locate/plantsci
1. Introduction
Growing plants are exposed to many environmentalstresses. Of the abiotic stresses, low temperature andwater stress have a particularly significant impact onplant growth, yield, and quality. It has been reportedthat low temperature and water stress are responsiblefor approximately 70% of all crop losses for whichindemnifications have been made to US farmers [1].
Plants react to such environmental stresses by initiat-ing a number of physiological and developmental
changes, which in turn require altered gene expression.Some stresses may utilize the same general mechanismof resistance. For example, it is widely accepted thatboth drought and freezing stress result in desiccation ofthe plant cell protoplasm [2]. During freezing stress, iceformation is generally initiated in the intercellularspaces. Because the chemical potential of ice is less thanwater, water moves from inside the cells to the intercel-lular spaces [3], resulting in desiccation of cell proto-plasm. Levitt [4] proposed that plants use a generalmechanism for desiccation resistance, regardless ofwhether desiccation results from drought or freezingstress. This early hypothesis has been supported bynumerous genetic and physiological studies. For exam-ple, a variety of genes are responsive to both droughtand low temperature [5]. Endogenous abscisic acid(ABA) levels increase in plant cells and tissues subjectedto low temperature [6] and water stress by high os-moticum, salt, or drought. Exogenous application of
* Corresponding author. Tel.: +1-301-504-6654; fax: +1-301-504-5062.
E-mail addresses: [email protected] (C.M. Parmentier-Line),[email protected] (G.R. Panta), [email protected] (L.J.Rowland).
1 Present address: Department of Natural Resource Sciences andLARC, University of Maryland, College Park, MD 20742, USA.
2 Present address: Department of Pharmacology, University ofTennessee Health Science Center, Memphis, TN 38163, USA.
0168-9452/02/$ - see front matter © 2002 Elsevier Science Ireland Ltd. All rights reserved.
PII: S 0 1 6 8 -9452 (01 )00563 -5

C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282274
ABA also induces expression of a number of genes thatrespond to drought and cold stress [5]. The phe-nomenon of cross-adaptation, where exposure to onetype of stress can provide tolerance to other types ofstress, supports Levitt’s hypothesis, as well. For exam-ple, plants exposed to low temperature can acquiredrought tolerance [7] and vice-versa [8,9].
Dehydrins are a family of plant proteins that areinduced by stimuli that have a dehydrative componentsuch as drought, low temperature, salinity, and ABA[10]. Also indicative of dehydrins is the presence of ahighly conserved lysine-rich 15-amino acid sequence(consensus sequence EKKGIMDKIKEKLPG), re-ferred to as the K segment, which is often repeatedseveral times [10]. Although there is not a clear under-standing of the entire sequence of events, dehydrins arethought to play a major role in protecting plants fromdehydrative stress. This is based on their presence inmany plant species, hydrophilic character, possiblecompatible solute-like properties, and correlation oftheir accumulation with cold and drought tolerances[10]. Indeed, there is emerging evidence for the partici-pation of dehydrins in the stabilization of cell mem-branes against dehydration-induced injury [3].
Previously, three dehydrins of 65, 60 and 14 kDawere identified as the predominant proteins present incold acclimated blueberry floral buds [11]. Levels wereshown to increase with cold acclimation and decreasewith deacclimation and resumption of growth. Further-more, the level of dehydrin accumulation in floral budsof several different blueberry genotypes has been shownto be closely associated with cold hardiness levels[11,12]. Expression studies have indicated that the blue-berry dehydrins are induced by cold stress in all organsexamined including floral buds, leaves, stems, androots, and by drought stress in primarily stems [11,13].To date, one full-length cDNA clone encoding the 60kDa dehydrin has been isolated from blueberry [14], ashave several other partial-length dehydrin cDNAs [15].
Cell culture systems have been widely used for physi-ological and gene expression studies because they offerpotential advantages over whole plants [16]. These ad-vantages include: (1) homogeneity of the cell popula-tion; (2) the ability to separate cellular response fromwhole plant response; (3) the ability to easily repeatexperiments and carry them out year-round. In addi-tion, the immediate environment of the cell culture canbe easily manipulated, allowing for more control overtreatments with chemicals. In fact, treatment of wholeplants with ABA for example can be problematic possi-bly due to inadequate uptake, rapid metabolism, and/ormicrobial degradation [17]. In contrast, cell cultureoffers a sterile system where the barriers to hormoneuptake are removed. Possible disadvantages are that aparticular response may be observed only in cell cultureor may not be observed because it requires a higher
level of tissue differentiation. A recent study [18]demonstrated that the same mechanisms were impli-cated in NaCl resistance in whole plants and tissuecultures of Mesembryanthenum crystallinum. The au-thors concluded that in ice plant, cell culture was anappropriate tool to study gene induction and physiolog-ical processes involved in salt resistance. In this study,we examined the expression of dehydrins and dehydrin-RNA in blueberry cell cultures in response to lowtemperature, ABA and polyethylene glycol (PEG) treat-ments. Expression of dehydrins was compared to thatseen previously with low temperature and droughtstress of whole plants.
2. Materials and methods
2.1. Blueberry cell cultures
Cell cultures were derived from shoot cultures ofblueberry highbush hybrid cultivar ‘Gulfcoast’ (Vac-cinium corymbosum L. x Vaccinium darrowi Camp),originally established in 1995 but regularly and rou-tinely subcultured [19]. Leaves and stems of a 10-day-old subculture were crudely chopped and placed in 25ml of liquid modified Woody Plant Medium [19] in 125ml Erlenmeyer flasks on a rotary shaker at 125–130rpm at room temperature. Calluses, formed after 2weeks, were broken up by pressing through a celldissociation c40 mesh screen (Sigma, St Louis, MO,USA). Cells were then placed in new medium andgrown at room temperature. Stock cultures were main-tained by transferring settled cell clusters into 25 ml offresh medium every 2 weeks.
2.2. Cold, ABA and PEG treatments
Cell clusters were grown in duplicate series in a coldroom at 4 °C in the dark. Cells were collected at thetime of subculture (0 day—no cold) and after 1, 2, 4, 9and 15 days at 4 °C. They were harvested by filtrationthrough Miracloth and washed with deionized water.Excess water was removed by blotting with Kimwipestissues. Samples were divided into 200 and 500 mg cellsFW, frozen in liquid nitrogen and kept at −80 °Cuntil analysis.
The ABA treatment consisted of cells subcultured atroom temperature in a fresh medium containing ABAat concentrations from 10−3 to 10−7 M. Cell clumpswere collected from duplicate flasks 1 and 2 weeks afteraddition of ABA. Control samples were collected at thesame times from cultures without ABA.
The response of cells to PEG was studied in cellssubcultured at room temperature in a medium contain-ing 10, 15, 20 or 25% (w/v) PEG for 10 or 17 days.Controls without PEG were also collected after 10 and

C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282 275
17 days of subculture. Cells were collected from dupli-cate flasks for each PEG concentration.
A control experiment was conducted to determine iftissue culture itself affected dehydrin expression. Cellswere collected from duplicate flasks at the time ofsubculture and after 1, 2, 4, 9 and 15 days of subcultureat room temperature.
Every treatment, control experiment included, wascarried out twice independently.
2.3. Protein extraction
Cell clusters (200 mg) were ground into fine powderwith liquid nitrogen and 50% (w/v) insoluble polyvinyl-polypyrrolidone in a pre-chilled mortar. Proteins werethen extracted by further grinding in 1.5 ml boratebuffer (50 mM sodium borate, 50 mM ascorbic acid, 1mM EDTA, 1% (v/v) �-mercaptoethanol, 1 mMPMSF, pH 9.0). The extract was centrifuged at 5000×g for 15 min, at room temperature. The resulting super-natant was centrifuged at 18 000×g at roomtemperature for 30 min. Proteins in the supernatantwere quantified using a protein assay kit (Bio-Rad,Hercules, CA, USA) based on the method of Bradford[20].
2.4. SDS-PAGE and western immunodetection
The appropriate volume of protein extract containing10 �g protein was diluted to 1 ml with purified water.Proteins were precipitated by adding 100 �l 0.15% (w/v)sodium deoxycholate (DOC) followed by 100 �l 72%(w/v) trichloracetic acid (TCA) [21]. Precipitates werecollected by centrifugation at 18 000×g at room tem-perature for 15 min. Pellets were rinsed twice withacetone and air-dried. Proteins were solubilized in 30 �lprotein solubilization buffer (62.5 mM Tris–HCl, pH6.8, 2% (w/v) SDS, 5% (v/v) �-mercaptoethanol, 10%(v/v) glycerol, 0.001% (w/v) bromophenol blue) [22].Samples were held in boiling water for 5 min andcooled to room temperature.
Proteins were separated by one-dimensional SDS-PAGE through 12.5% polyacrylamide gels, using theMini Protean II cell (Bio-Rad, Hercules, CA, USA).Fractionated proteins from unstained gels were elec-troblotted onto 0.45 �m nitrocellulose membrane usingthe Mini Trans Blot electrophoretic transfer cell (Bio-Rad, Hercules, CA, USA). Electroblotting was carriedout at 100 V for 75 min in Towbin buffer (192 mMglycine, 25 mM Tris base, 20% (v/v) methanol, pH 8.3)[23]. Membranes were blocked for 1 h at 30 °C in 3%gelatin in TBS buffer (25 mM Tris base, 137 mM NaCl,27 mM KCl, pH 7.4) and probed for 2 h at 30 °C withanti-65 kDa antibody (polyclonal antibody from gelpurified 65 kDa blueberry dehydrin, Organon Tech-nika, Durham, NC, USA) at 1:500 dilution in 1%
gelatin in TBS. The blots were washed three times inTTBS (0.002% (v/v) Tween 20 in TBS) and incubatedfor 1 h at 30 °C with goat anti-rabbit-alkaline phos-phatase-conjugated IgG (Boehringer Mannheim, Indi-anapolis, IN, USA) at 1:500 dilution in TBS.Membranes were washed three times in TTBS followedby one wash in TBS. Immuno-reacting proteins werevisualized with nitroblue tetrazolium (NBT, 0.3 mg/ml)and 5-bromo-4-chloro-3-indolyl phosphate (BCIP, 0.15mg/ml) in alkaline phosphatase buffer (100 mM Tris–HCl pH 9.5, 100 mM NaCl, 5 mM MgCl2) [24]. Im-munoblot analyses of dehydrins after the varioustreatments were repeated at least twice. Replicationsshowed similar trends in terms of increase or decreasein dehydrin levels. Therefore, the best single representa-tive blots were chosen for presentation.
2.5. Total RNA extraction
Frozen cells (500 mg) were ground in a pre-chilledmortar in the presence of liquid nitrogen. The frozenpowder was transferred to a centrifuge tube and mixedwith 3 ml extraction buffer (100 mM Tris base pH 8.5,20 mM EDTA pH 8.0, 1.4 M NaCl, 2% (w/v)polyvinylpyrrolidone, 2% (v/v) Igepal, 1.5% (w/v)DOC, 2% (w/v) thiourea, 1% (v/v) �-mercaptoethanol,4 M guanidine thyocianate, 300 mM LiCl, 1.5 mMMgCl2·6H2O) by vigorous vortexing [15]. An equalvolume of chloroform was added and again mixed byvortexing. The extract was centrifuged at 12 000×g at4 °C for 10 min. The aqueous upper phase was ex-tracted two more times with equal volumes of acidphenol:chloroform (5:1) pH 4.7 and once with chloro-form:isoamylalcohol (24:1). RNA was precipitatedovernight at −20 °C with the addition of 1/10 volume3 M NaCl and 2 volumes of 95% ethanol. PrecipitatedRNA was collected by centrifugation at 12 000×g at4 °C for 10 min. Pellets were washed with 70% ethanol,air-dried, and dissolved in 100 �l RNA storage solution(Ambion, Austin, TX, USA).
2.6. RNA blots
Total RNA (about 4 �g/lane) was separated by sizeon 1% agarose/formaldehyde gels and transferred ontoBrightstar-Plus™ nylon membranes (Ambion, Austin,TX, USA) by capillary transfer, using NorthernMax™blotting kit (Ambion, Austin, TX, USA). The RNAwas cross-linked to the membrane by baking at 80 °Cfor 15 min.
The probe DNA was derived from a 2.0 kb cDNAdehydrin clone, bbdhn1, which was isolated from ablueberry flower bud cDNA library [14]. The plasmidDNA containing the cDNA was purified from theEscherichia coli strain using the PEG method [25] anddigested with EcoRI and XhoI restriction enzymes

C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282276
(Promega Inc., Madison, WI, USA) to release theinsert. The digested DNA was separated on a 1%agarose gel. The 2.0 kb cDNA insert was excised fromthe gel, purified with Geneclean® Spin kit (Bio 101 Inc.,Carlsbad, CA, USA) and 32P-labeled to a specific activ-ity of 2.0×1010 cpm/�g by random priming (HighPrime kit, Boehringer-Mannheim). Membranes werepre-hybridized in the hybridization buffer (0.25 MNa2HPO4, 7% SDS) for 5 min and hybridized overnightat 60 °C. Washes were done at 60 °C in SSC/SDS withincreasing stringency (10 min in 1×SSC/1% SDS; 15min in 0.5×SSC/0.5% SDS; 15 min in 0.25×SSC/0.25% SDS; 15 min in 0.1×SSC/0.1% SDS). Blots wereexposed to X-ray film (Fuji Medical System, Stamford,CT, USA) with intensifying screens, at −80 °C.
2.7. Densitometry
Developed protein and RNA blots were imaged withScanMaker (Microtek, Redondo Beach, CA, USA) andbands quantified using a digital imaging system (KodakDigital Science 1D Image Analysis software, EastmanKodak Company).
3. Results
3.1. Dehydrin expression at the protein le�el
Levels of dehydrins in different treatments of blue-berry cell cultures of ‘Gulfcoast’ were analyzed by gelelectrophoresis followed by immunoblotting. The anti-serum used was raised against the 65 kDa dehydrinfrom flower buds of cold hardy blueberry plants. Thisantiserum cross-reacts to both 60 and 65 kDa dehydrinsin flower buds but not to the 14 kDa dehydrin [14]. The14 kDa protein was identified as a dehydrin previouslybased on its cross-reaction with polyclonal antibody[26] raised against a synthetic peptide of the conserveddehydrin K-box [11]. The 65, 60 and 14 kDa dehydrinsappear as the most abundant proteins from extracts ofwinter hardy flower buds [11]. In blueberry cell cultures,two major polypeptides with estimated molecularmasses of 65 and 30 kDa were identified on westernblots using the anti-65 kDa antibody. A 14 kDa proteinwas observed on Coomassie-stained gels but no strikingchanges in its levels were noticed with any of thetreatments. Therefore, changes in levels of only the 65and 30 kDa dehydrins were monitored.
3.1.1. AgingTo monitor if dehydrins were influenced by aging of
the culture alone, proteins were analyzed during onesubculture of cells at room temperature in liquidmodified Woody Plant Medium. The 30 kDa dehydrinwas present at a very low level and did not seem to
Fig. 1. Immunoblot of dehydrins from untreated blueberry cell cul-tures of ‘Gulfcoast’ cultivar. Cells were collected during a subcultureat room temperature. Ten microgram proteins were separated bySDS-PAGE, transferred to nitrocellulose membrane and probed withanti-65 kDa antibody. The arrows indicate the two dehydrins of 65and 30 kDa. St, pre-stained protein standard; Mm, molecular mass ofthe protein standards.
change during the culture (Fig. 1). The level of the 65kDa dehydrin on the other hand fluctuated during thesubculture at room temperature. It increased to peak atthe 4th day and then decreased to reach the initial levelby the 9th day of subculture. In addition to the 30 and65 kDa proteins, extra bands were apparent on theimmunoblot of the control experiment. Since thesepolypeptides were not consistently observed across thedifferent treatments, they were considered an artifact ofa longer color reaction development, necessary to visu-alize lower levels of proteins.
3.1.2. Cold treatmentWhen cells were kept at 4 °C in the dark, the level of
the 65 kDa dehydrin did not change during the 2-weektreatment (Fig. 2). However, the level of the 30 kDadehydrin increased as soon as cells were transferred tocold conditions (by 1 day) and increased more as thecultures were held longer at 4 °C.
Fig. 2. Immunoblot of dehydrins from cold-treated blueberry cellcultures of ‘Gulfcoast’ cultivar. Ten microgram proteins were sepa-rated by SDS-PAGE, transferred to nitrocellulose membrane andprobed with anti-65 kDa antibody. The arrows indicate the twodehydrins of 65 and 30 kDa. St, pre-stained protein standard; Mm,molecular mass of the protein standards.

C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282 277
Fig. 3. Immunoblot of dehydrins from ABA-treated blueberry cell cultures of ‘Gulfcoast’ cultivar. Cells were collected after 1 and 2 weeks ofculture at room temperature in the presence of different concentrations of ABA. Ten microgram proteins were separated by SDS-PAGE,transferred to nitrocellulose membrane and probed with anti-65 kDa antibody. The arrows indicate the two dehydrins of 65 and 30 kDa. St,prestained protein standard; Mm, molecular mass of the protein standards.
3.1.3. ABA treatmentThe response of cells to ABA was investigated after 1
and 2 weeks of co-culture with different concentrationsof ABA. With all ABA concentrations tested, the 1-week treatment resulted in an induction of the 30 kDadehydrin with respect to the control without ABA (Fig.3). With the 1-week treatment, the 65 kDa dehydrinappeared to be unchanged or only slightly induced. Ifcells were treated with ABA for 2 weeks, the highestlevels of both the 30 and 65 kDa dehydrins wereobserved with 10−5 and 10−6 M ABA (Fig. 3). At thehighest concentration tested (10−3 M) the immunoblotrevealed less reactive bands indicating a lower amountof proteins detected by the antibody. Indeed, the cellslooked browner and the total fresh weight collected at10−3 M was lower by 45–65% than for any of theother concentrations (results not shown). With 2 weeksat 10−4 M ABA the 30 kDa dehydrin did not appear tobe significantly induced or reduced while level of the 65kDa protein was slightly increased over control level.The lowest concentration of ABA (10−7 M) for 2weeks was ineffective in inducing any changes in levelsof the 30 or 65 kDa dehydrins when compared to thecontrol without ABA.
3.1.4. PEG treatmentCells were grown under conditions of induced water
stress using PEG added to the culture medium. After a10-day treatment with PEG at 10 and 25% (w/v), therewas a decrease in the levels of the 65 and 30 kDadehydrins when compared to the control without PEG(Fig. 4). The 15 and 20% PEG concentrations stimu-lated the accumulation of the 30 kDa dehydrin whilethe level of the 65 kDa dehydrin seemed unchanged orslightly reduced. In cells kept for a further 7 days withPEG, the levels of the 30 and 65 kDa dehydrins werenot affected until 20% PEG was reached (Fig. 4). Aswith the 10-day treatment, the 17-day treatment ofPEG applied at 25% repressed the levels of the twodehydrins.
3.2. Dehydrin expression at the RNA le�el
For each experiment, northern blots of total RNAare presented along with the corresponding ethidium-bromide stained total RNA from agarose/formaldehydegel electrophoresis. Densitometric scans of the gels wereperformed, along with the northern blots, to correct forany slight differences in amounts of RNA loaded. Theprobe used for the northern blots was a full-lengthdehydrin cDNA clone from flower buds of cold hardyblueberry plants, representing the bbdhn1 gene, whichencodes the 60 kDa dehydrin [14]. This cDNA hy-bridizes to two chilling-responsive transcripts of 2.0 and0.5 kb from flower bud RNA [14]. With blueberry cellcultures, the bbdhn1 probe hybridized to one transcriptof 0.75 kb.
3.2.1. AgingBefore examining the effect of different treatments,
we monitored how aging affected the dehydrin messagelevels. Blots from the control experiment revealed thatthe 0.75 kb transcript was present at a very low levelinitially and became practically undetectable with addi-tional days at room temperature (Fig. 5).
3.2.2. Cold treatmentAfter making corrections for the differences in
amount of RNA loaded based on the intensity of therRNA bands, it appeared that the level of the 0.75 kbdehydrin transcript was affected by the 4 °C treatment(Fig. 5). After a slight increase between 1 and 2 days,the level of the transcript remained lower than the levelat day 1. For both aging and cold experiments, controlsamples, i.e. samples collected at the time of subculture,was not represented on the blots shown. Because oflower RNA yield for these samples, there was notenough RNA for all the northern blots that were done.Moreover, none of the controls from the ABA andPEG experiments could replace the missing samplesbecause those controls were ‘concentration controls’collected at different times.
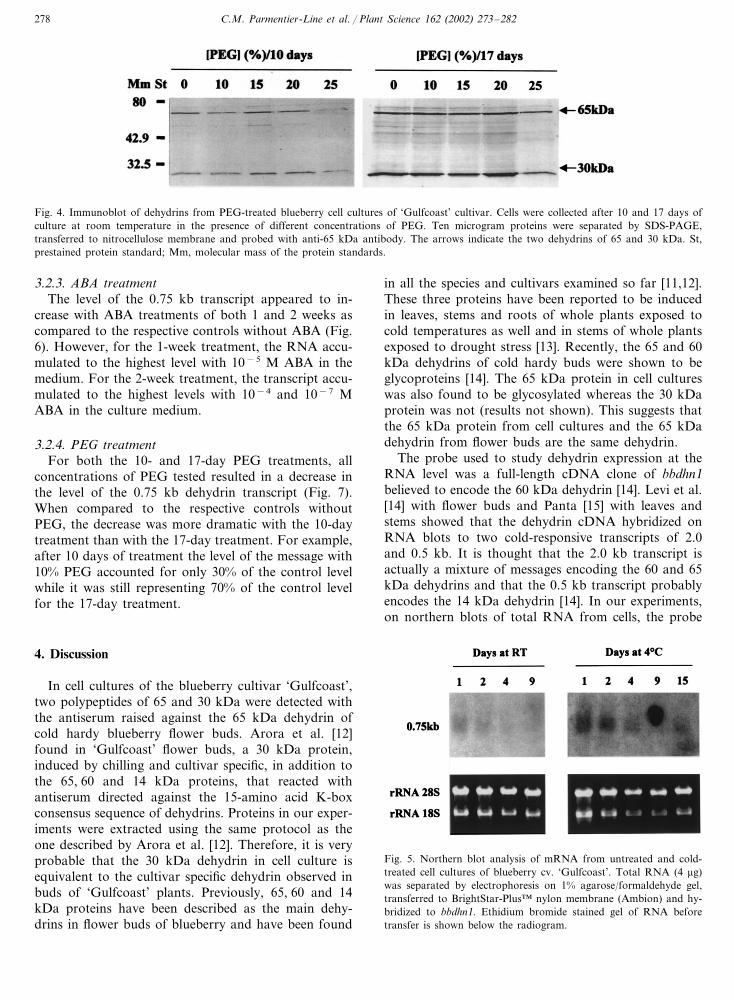
C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282278
Fig. 4. Immunoblot of dehydrins from PEG-treated blueberry cell cultures of ‘Gulfcoast’ cultivar. Cells were collected after 10 and 17 days ofculture at room temperature in the presence of different concentrations of PEG. Ten microgram proteins were separated by SDS-PAGE,transferred to nitrocellulose membrane and probed with anti-65 kDa antibody. The arrows indicate the two dehydrins of 65 and 30 kDa. St,prestained protein standard; Mm, molecular mass of the protein standards.
3.2.3. ABA treatmentThe level of the 0.75 kb transcript appeared to in-
crease with ABA treatments of both 1 and 2 weeks ascompared to the respective controls without ABA (Fig.6). However, for the 1-week treatment, the RNA accu-mulated to the highest level with 10−5 M ABA in themedium. For the 2-week treatment, the transcript accu-mulated to the highest levels with 10−4 and 10−7 MABA in the culture medium.
3.2.4. PEG treatmentFor both the 10- and 17-day PEG treatments, all
concentrations of PEG tested resulted in a decrease inthe level of the 0.75 kb dehydrin transcript (Fig. 7).When compared to the respective controls withoutPEG, the decrease was more dramatic with the 10-daytreatment than with the 17-day treatment. For example,after 10 days of treatment the level of the message with10% PEG accounted for only 30% of the control levelwhile it was still representing 70% of the control levelfor the 17-day treatment.
4. Discussion
In cell cultures of the blueberry cultivar ‘Gulfcoast’,two polypeptides of 65 and 30 kDa were detected withthe antiserum raised against the 65 kDa dehydrin ofcold hardy blueberry flower buds. Arora et al. [12]found in ‘Gulfcoast’ flower buds, a 30 kDa protein,induced by chilling and cultivar specific, in addition tothe 65, 60 and 14 kDa proteins, that reacted withantiserum directed against the 15-amino acid K-boxconsensus sequence of dehydrins. Proteins in our exper-iments were extracted using the same protocol as theone described by Arora et al. [12]. Therefore, it is veryprobable that the 30 kDa dehydrin in cell culture isequivalent to the cultivar specific dehydrin observed inbuds of ‘Gulfcoast’ plants. Previously, 65, 60 and 14kDa proteins have been described as the main dehy-drins in flower buds of blueberry and have been found
in all the species and cultivars examined so far [11,12].These three proteins have been reported to be inducedin leaves, stems and roots of whole plants exposed tocold temperatures as well and in stems of whole plantsexposed to drought stress [13]. Recently, the 65 and 60kDa dehydrins of cold hardy buds were shown to beglycoproteins [14]. The 65 kDa protein in cell cultureswas also found to be glycosylated whereas the 30 kDaprotein was not (results not shown). This suggests thatthe 65 kDa protein from cell cultures and the 65 kDadehydrin from flower buds are the same dehydrin.
The probe used to study dehydrin expression at theRNA level was a full-length cDNA clone of bbdhn1believed to encode the 60 kDa dehydrin [14]. Levi et al.[14] with flower buds and Panta [15] with leaves andstems showed that the dehydrin cDNA hybridized onRNA blots to two cold-responsive transcripts of 2.0and 0.5 kb. It is thought that the 2.0 kb transcript isactually a mixture of messages encoding the 60 and 65kDa dehydrins and that the 0.5 kb transcript probablyencodes the 14 kDa dehydrin [14]. In our experiments,on northern blots of total RNA from cells, the probe
Fig. 5. Northern blot analysis of mRNA from untreated and cold-treated cell cultures of blueberry cv. ‘Gulfcoast’. Total RNA (4 �g)was separated by electrophoresis on 1% agarose/formaldehyde gel,transferred to BrightStar-Plus™ nylon membrane (Ambion) and hy-bridized to bbdhn1. Ethidium bromide stained gel of RNA beforetransfer is shown below the radiogram.

C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282 279
Fig. 6. Northern blot analysis of mRNA from ABA-treated cell cultures of blueberry cv. ‘Gulfcoast’. Cells were collected after 1 and 2 weeks ofculture in presence of different concentrations of ABA. Total RNA (4 �g) was separated by electrophoresis on 1% agarose/formaldehyde gel,transferred to BrightStar-Plus™ nylon membrane (Ambion) and hybridized to bbdhn1. Ethidium bromide stained gels of RNA before transfer areshown below the radiograms.
hybridized to a 0.75 kb message that was not previouslyreported in flower buds [14] nor leaves and stems [15] ofblueberry plants, but based on the average molecularweight of amino acids (0.11 kDa), the transcript is of anappropriate size to encode the 30 kDa cultivar specificdehydrin detected in cells of ‘Gulfcoast’. Whether ornot this is the case still remains to be proven. Eventhough a 65 kDa dehydrin was detected on westernblots, there was no corresponding signal on the north-ern blots. Considering that the 2.0 kb transcript isprobably a mixture of two messages encoding the 60and the 65 kDa dehydrins, it is conceivable that thetranscript for the 60 kDa dehydrin accounts for most ofthe signal. Hence, because the 60 kDa dehydrin is notpresent in cell cultures, the message for the 65 kDadehydrin could be too low to be detected, explainingwhy the probe hybridized to only a 0.75 kb transcript.
When dehydrin levels were monitored during a sub-culture at room temperature, both 65 and 30 kDadehydrins were detected on western blots. Their pres-ence, even in the absence of treatment, would suggestthat these two dehydrins are constitutive in blueberrycell cultures of the ‘Gulfcoast’ cultivar. Some earlierstudies support the idea that dehydrins, like heat shockproteins, are a group of proteins that have constitutiveand inducible members. Close et al. [26] showed thatbarley seedling tissues contained some dehydrinswhether they were stressed or not. For example, theyfound in the scutellum of well hydrated seedlings a 60kDa dehydrin identical to the 60 kDa dehydrin ofstressed seedlings. In the aleurone layer of well hy-drated seedlings they detected mainly a 25 kDa dehy-drin but when the seedlings were stressed there wereseven additional bands detected by the antibody raisedagainst the 15-amino acid K-box consensus sequence ofdehydrins. Robertson and Chandler [27] identified inpea (Pisum sati�um L. ‘Greenfeast’) a 40 kDa proteinthat they referred to as a dehydrin cognate, which was
strictly constitutively expressed and whose abundanceand that of the corresponding mRNA changed littlewith dehydration treatment and ABA application. InArabidopsis, the dhnX gene was not induced by eitherlow temperature, drought or ABA and exhibited aconstitutive expression pattern [28]. The authors sug-gested that the constitutive protein could provide anearly protection before the stress responses are engaged.In experiments with tuber-bearing potato species [29],immunoblots revealed that for some species a 42 kDadehydrin could be detected in the leaves of non-accli-mated plants and that cold acclimation treatment, forall the species in the study, resulted in increased expres-sion of the 42 kDa dehydrin.
As with whole plants [11,13] subjecting cell culturesof blueberry to low non-freezing temperatures resultedin an increase in dehydrin levels. Earlier studies haveshown that in cell cultures derived from alfalfa [30],bromegrass [31,32] or peach [33], proteins accumulatedif the cultures were grown at 3 °C. However, theauthors did not specify if any of the induced proteinswere dehydrins. While the level of the 30 kDa dehydrincontinuously increased, the level of the 0.75 kb mRNAdecreased during the subculture, except for a shortincrease between 1 and 2 days of cold treatment. Sinceproteins and RNA were extracted from the same sam-ples, it is unlikely that the difference between proteinand transcript accumulation is an experimental error. Ithas also been seen with whole plants of blueberry [15]and with peach [34,35] that the levels of dehydrins andtheir corresponding mRNAs are not always commensu-rate with each other. For example, in leaves of the‘Bluecrop’ cultivar of blueberry, elevated levels of dehy-drins were still present after 5 weeks of cold treatmentwhile transcript levels had declined by 4 weeks [15]. Inpeach, as well, elevated levels of dehydrin PCA60 werestill present when the transcript level had already de-clined significantly [34]. This would indicate that post-

C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282280
transcriptional control mechanisms are involved in theregulation of dehydrin expression. Post-transcriptionalregulation could imply regulation at the RNA level—processing, transport, stability of RNA [36],— regula-tion at the translational level or regulation at theprotein level. Our experiments did not allow, and didnot intend for, us to specify the regulation level(s) thatoccurred in the cell culture, but since dehydrin abun-dance increased while the corresponding RNA abun-dance decreased, one might hypothesize an increase inthe translational efficiency and/or an increase in thestability of the protein.
Dehydrins have been shown to increase in responseto applied ABA [37–39] and to treatments like water ordehydration stresses [13,28,37,38,40,41]. Polyethyleneglycol (PEG), a non-penetrating osmotic agent thatlowers the water potential of the medium, is commonlyused to induce water stress in cultured cells of herba-ceous plants [42–44] as well as woody plants [45–47].In order to assess the response of blueberry cell culturesto ABA and water stress, we tested ABA and PEG inthe culture medium at different concentrations for 1and 2 weeks and 10 and 17 days, respectively. Some ofthe ABA concentrations induced expressions of bothdehydrins and of the 0.75 kb message, but strongdiscrepancies were observed between the 30 kDaprotein and mRNA levels. For example, for the 2-weektreatment, the two concentrations of ABA that showedthe highest increases at the RNA level were the oneswhere no differences were discernable at the proteinlevel. Amongst others, a study with yeast [48] revealedthe absence of strong correlation between the abun-dance of proteins and the corresponding mRNAs, indi-cating that genes are subjected to post-transcriptionalregulation. What regulation mechanism(s) is (are) in-volved with ABA treatment and whether or not it is(they are) the same as with cold treatment could not beanswered here. Unexpectedly, in our experiments, PEG
in the medium repressed dehydrins both at the proteinand RNA levels. One possible explanation for theseresults could be that the cell cultures failed to adjust tothe water stress. As a matter of fact, Tschaplinski et al.[49] with callus cultures of poplar showed that eventhough PEG in the medium provided an osmotic stress,the cultures did not display osmotic adjustment to thewater stress. Another possibility could be that dehy-drins in blueberry cell cultures do not respond to thestress and that other mechanisms and solutes such asproline, glycine–betaine, sucrose and fructans are in-volved in adjusting the osmotic potential of the cells[50].
From our experiments it appears that the 30 and 65kDa dehydrins of ‘Gulfcoast’ cell cultures differ in theirresponse to the various treatments used. The 30 kDadehydrin is induced by both cold and ABA in themedium whereas the 65 kDa dehydrin is not induced bycold and is induced by ABA only after 2 weeks oftreatment. Dehydrins are indeed a group of proteinswhose members differ in their responsiveness to differ-ent stimuli. WCS120 (50 kDa dehydrin in wheat) [51]and P-80 (80 kDa dehydrin in barley) [52] specificallyaccumulate in response to low temperature and are notinduced under drought or exogenous ABA. Anotherdehydrin gene in barley, Dhn5 that encodes for an 86kDa dehydrin, is specifically induced by low tempera-ture while Dhn1 and Dhn2 are not [53]. In wheat, severewater stress, osmotic stress and ABA treatment signifi-cantly induce the accumulation of WCOR410 proteins(28 kDa acidic dehydrin) but to a lower level than lowtemperature [54]. On the other hand, the expression ofrab18 is strongly induced in both water stressed andABA treated plants of Arabidopsis thaliana but is onlyslightly responsive to cold [28].
If we compare the results obtained with blueberrycell cultures to what is known about expression ofdehydrins in whole plants, many differences can be
Fig. 7. Northern blot analysis of mRNA from PEG-treated cell cultures of blueberry cv. ‘Gulfcoast’. Cells were collected after 10 and 17 daysof culture in presence of different concentrations of PEG. Total RNA (4 �g) was separated by electrophoresis on 1% agarose/formaldehyde gel,transferred to BrightStar-Plus™ nylon membrane (Ambion) and hybridized to bbdhn1. Ethidium bromide stained gels of RNA before transfer areshown below the radiograms.

C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282 281
noted. In cell cultures, the main dehydrin, whose leveland that of the corresponding mRNA increase withcold and ABA treatments, appears to be the 30 kDadehydrin. Even though the 65 kDa dehydrin is detectedin cell cultures, it does not appear to respond to coldtreatment whereas in plants it is one of the maindehydrins whose levels increase with cold treatment[11,13]. Similarly in bromegrass, exposure of plants tolow temperature resulted in expression of dehydringenes but low temperature treatment of cell cultures didnot [55]. Also, Arora and Wisniewski [33], in theirstudies with peach cell suspension cultures, observedincreases in the levels of numerous proteins with coldtreatment but not of a 60 kDa protein although thebest characterized cold acclimation-associated proteinin whole plants of peach is a 60 kDa dehydrin[34,35,56]. In whole plants, drought induces expressionof the 65 and 60 kDa dehydrins [13] while in ourexperiments PEG in the medium repressed the expres-sion of both the 30 and the 65 kDa dehydrins. It ispossible that the increase in the dehydrin levels seen inwhole blueberry plants with drought treatment is reallydue to increase in endogenous ABA levels which areknown to occur in response to cold and drought treat-ments of some plants [3]. Some types of drought-re-sponsive genes appear to be responsive to changes inABA levels alone and not to wilting itself [5]. In factour work with whole plants suggests that the blueberrydehydrins may be primarily responsive to changes inABA levels during drought stress rather than desicca-tion or wilting [13]. Blueberry cell cultures may notrespond to osmotic stress with increase in endogenousABA levels like whole plants apparently do. Changes inthe 60 and 14 kDa dehydrins were not observed in cellcultures of ‘Gulfcoast’ with any of the treatments or inanother variety examined (‘Tifblue’—data not shown).There may be some other factors in the signal transduc-tion pathway, absent from cell cultures, that are re-quired for induction of these other dehydrins. In lightof these differences, cell cultures do not appear to us asa suitable system for replacing whole plants for thestudy of dehydrin expression in blueberry. Tschaplinskiet al. [49] as well dismissed the use of callus culture forassessing water stress at the whole plant level. Baldwinet al. [57], based on their studies with tissue culturedplantlets of saskatoon berry, concluded that cautionshould be taken when using tissue culture as a systemto evaluate, in their case, regulation of cold acclimationin woody plants, in part, due to effects of phytohor-mones in the media.
References
[1] J.S. Boyer, Plant productivity and environment, Science 218(1982) 443–448.
[2] P.J. Steponkus, A unified concept of stress in plants, in: D.W.Rains, R.C. Valentine, A. Hollaender (Eds.), Genetic Engineer-ing of Osmoregulation, Plenum Press, NY, USA, 1980, pp.235–255.
[3] M.F. Thomashow, Plant cold acclimation: freezing tolerancegenes and regulatory mechanisms, Ann. Rev. Plant Physiol.Plant Mol. Biol. 50 (1999) 573–599.
[4] J. Levitt, Frost, drought and heat resistance, in: Protoplasmolo-gia Viii. Physiologie des Protoplasmes, Springer-Verlag, Vienna,1958, pp. 1–87.
[5] K. Shinozaki, K. Yamaguchi-Shinozaki, Molecular responses todehydration and low temperature: differences and cross-talkbetween two stress signaling pathways, Curr. Opin. Plant Biol. 3(2000) 217–223.
[6] T.H.H. Chen, P.H. Li, M.L. Brenner, Involvement of abscissicacid in potato cold acclimation, Plant Physiol. 71 (1983) 362–365.
[7] J. Levitt, C.Y. Sullivan, E. Krull, Some problems in droughtresistance, Bull. Counc. Isr. 80 (1960) 173–179.
[8] T. Anisko, O.M. Lindstrom, Seasonal changes in cold hardinessof Rhododendron L. ‘catawbiense Boursault’ grown under contin-uous and periodic water stress, J. Am. Soc. Hort. Sci. 121 (1996)301–306.
[9] P. Chen, P.H. Li, C.J. Weiser, Induction of frost hardiness in redosier dogwood stems by water stress, HortSci. 10 (1975) 372–374.
[10] T.J. Close, Dehydrins: emergence of a biochemical role of afamily of plant dehydration proteins, Physiol. Plant 97 (1996)795–803.
[11] M.M. Muthalif, L.J. Rowland, Identification of dehydrin-likeproteins responsive to chilling in floral buds of blueberry (Vac-cinium, section Cyanococcus), Plant Physiol. 104 (1994) 1439–1447.
[12] R. Arora, L.J. Rowland, G.R. Panta, Chill-responsive dehydrinsin blueberry: are they associated with cold hardiness or dor-mancy transitions?, Physiol. Plant 101 (1997) 8–16.
[13] G.R. Panta, M.W. Rieger, L.J. Rowland, Effect of cold anddrought stress on blueberry dehydrin accumulation, J. Hort. Sci.Biotech. 76 (2001) 549–556.
[14] A. Levi, G.R. Panta, C.M. Parmentier, M.M. Muthalif, R.Arora, S. Shanker, L.J. Rowland, Complementary DNA cloningsequencing and expression of an unusual dehydrin from blue-berry floral buds, Physiol. Plant 107 (1999) 98–109.
[15] G.R. Panta, Cloning and characterizing blueberry (Vacciniumsection Cyanococcus) dehydrins. Ph.D., University of Georgia,1999.
[16] M. Tal, Selection for stress tolerance, in: D.A. Evans, W.R.Sharp, P.V. Ammirato, Y. Yamada (Eds.), Handbook of PlantCell Culture, Macmillan Publishing Co., New York, 1983, pp.461–488.
[17] T.H.H. Chen, L.V. Gusta, Abscissic acid-induced freezing resis-tance in cultured plant cells, Plant Physiol. 73 (1983) 71–75.
[18] R. Vera-Estrella, B.J. Barkla, H.J. Bohnert, O. Pantoja, Saltstress in Mesembryanthemum crystallinum L. cell suspensionsactivates adaptive mechanisms similar to those observed in thewhole plant, Planta 207 (1999) 426–435.
[19] L.J. Rowland, E.L. Ogden, Use of a cytokinin conjugate forefficient shoot regeneration from leaf sections of highbush blue-berry, HortScience 27 (1992) 1127–1129.
[20] M.M. Bradford, A rapid and sensitive method for the quantita-tion of microgram quantities of protein utilizing the principle ofprotein-dye binding, Anal. Biochem. 72 (1976) 248–254.
[21] A. Bensadoum, D. Weinstein, Assay of protein in the presence ofinterfering material, Anal. Biochem. 70 (1976) 241–250.
[22] U.K. Laemmli, Cleavage of structural proteins during the assem-bly of the head of the bacteriophage T4, Nature 227 (1970)680–685.

C.M. Parmentier-Line et al. / Plant Science 162 (2002) 273–282282
[23] H. Towbin, T. Staehelin, J. Gordon, Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: proce-dure and some applications, Proc. Nat. Acad. Sci. USA 76(1979) 4350–4354.
[24] J. Sambrook, E.F. Frisch, T. Maniatis, Molecular Cloning: ALaboratory Manual, Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY, 1989.
[25] V.G. Nicoletti, D.F. Condorelli, Optimizing PEG method forrapid plasmid DNA purification: high yield from ‘Midi-Prep’,Biotechniques 14 (1993) 535–536.
[26] T.J. Close, R.D. Fenton, F. Moonan, A view of plant dehydrinsusing antibodies specific to the carboxy terminal peptide, PlantMol. Biol. 23 (1993) 279–286.
[27] M. Robertson, P.M. Chandler, A dehydrin cognate protein frompea (Pisum sati�um L.) with an atypical pattern of expression,Plant Mol. Biol. 26 (1994) 805–816.
[28] B.V. Welin, A. Olson, M. Nylander, E.T. Palva, Characteriza-tion and differential expression of dhn/lea/rab-like genes duringcold acclimation and drought stress in Arabidopsis thaliana,Plant Mol. Biol. 26 (1994) 131–144.
[29] B.H. Karlsson, J.P. Palta, Expression of heat-stable and putativedehydrin proteins during non-acclimated and cold-acclimatedconditions in several tuber-bearing Solanum species, J. Am. Soc.Hort. Sci. 124 (1999) 245–251.
[30] A.J. Robertson, L.V. Gusta, Abscisic acid and low temperatureinduced polypeptide changes in alfalfa (Medicago sati�a) cellsuspension cultures, Can. J. Bot. 64 (1986) 2758–2763.
[31] A.J. Robertson, L.V. Gusta, M.J.T. Reany, M. Ishikawa,Protein synthesis in bromegrass (Bromus inermis Leyss) culturedcells during the induction of frost tolerance by abscisic acid orlow temperature, Plant Physiol. 84 (1987) 1331–1336.
[32] A.J. Robertson, L.V. Gusta, M.J.T. Reany, M. Ishikawa, Iden-tification of proteins correlated with increased freezing tolerancein bromegrass (Bromus inermis Leyss cv. Manchar) cell cultures,Plant Physiol. 86 (1988) 344–347.
[33] R. Arora, M.E. Wisniewski, Ultrastructural and protein changesin cell suspension cultures of peach associated with low tempera-ture-induced cold acclimation and abscisic acid treatment, PlantCell Tissue Organ Cult. 40 (1995) 17–24.
[34] T. Artlip, M. Wisniewski, Tissue-specific expression of a dehy-drin gene in one-year old ‘Rio Oso Gem’ peach trees, J. Am.Soc. Hort. Sci. 122 (1997) 784–787.
[35] T.S. Artlip, A.M. Callahan, C.L. Bassett, M.E. Wisniewski,Seasonal expression of a dehydrin gene in sibling deciduous andevergreen genotypes of peach (Prunus persica [L.] Batsch), PlantMol. Biol. 33 (1997) 61–70.
[36] D.R. Gallie, Post-trancriptional regulation of gene expression inplants, Ann. Rev. Plant Physiol. Plant Mol. Biol. 44 (1993)77–105.
[37] M. Robertson, P.M. Chandler, Pea dehydrins: identification,characterisation and expression, Plant Mol. Biol. 19 (1992)1031–1044.
[38] B. Han, A.R. Kermode, Dehydrin-like proteins in castor beanseeds and seedlings are differentially produced in response toABA and water-deficit-related stresses, J. Exp. Bot. 47 (1996)933–939.
[39] S.P. Lee, T.H.H. Chen, Molecular cloning of abscisic acid-re-sponsive mRNAs expressed during the induction of freezingtolerance in bromegrass (Bromus inermis Leyss) suspension cul-ture, Plant Physiol. 101 (1993) 1089–1096.
[40] M. Bracale, M. Levi, C. Savini, W. Dicorato, M.G. Galli, Waterdeficit in pea root tips: effects of the cell cycle on the productionof dehydrin-like proteins, Ann. Bot. 79 (1997) 593–600.
[41] F. Cellier, G. Conejero, J.C. Breitler, F. Casse, Molecular andphysiological responses to water deficit in drought-tolerant anddrought-sensitive lines of sunflower. Accumulation of dehydrintranscripts correlates with tolerance, Plant Physiol. 116 (1998)319–328.
[42] R.A. Bressan, P.M. Hasegawa, A.V. Handa, Resistance of cul-tured higher plant cells to polyethylene glycol-induced waterstress, Plant Sci. Let. 21 (1981) 23–30.
[43] S. Bhaskaran, R.H. Smith, R.J. Newton, Physiological changesin cultured sorghum cells in response to induced water stress. 1.Free proline, Plant Physiol. 79 (1985) 266–269.
[44] K.M. Fallon, R. Phillips, Responses to water stress in adaptedand unadapted carrot cell suspension cultures, J. Exp. Bot. 40(1989) 681–687.
[45] G. Ben-Hayyim, Relationship between salt tolerance and resis-tance to polyethylene glycol-induced water stress in culturedcitrus cells, Plant Physiol. 85 (1987) 430–433.
[46] T. Leustek, E.G. Kirby, The influence of glutamine on growthand viability of cell suspension cultures of Douglas-fir afterexposure to polyethylene glycol, Tree Physiol. 4 (1988) 371–380.
[47] T. Leustek, E.G. Kirby, Selection and physiology of cell culturesof Douglas-fir grown under conditions of water stress, TreePhysiol. 6 (1990) 317–327.
[48] S.P. Gygi, Y. Rochon, B.R. Franza, R. Aebershold, Correlationbetween protein and mRNA abundance in yeast, Mol. Cell. Biol.19 (1999) 1720–1730.
[49] T.J. Tschaplinski, G.M. Gebre, J.E. Dahl, G.T. Roberts, G.A.Tuskan, Growth and solute adjustment of calli of Populus clonescultured on nutrient medium containing polyethylene glycol,Can. J. For. Res. 25 (1995) 1425–1433.
[50] A.D. Hanson, W.D. Hitz, Metabolic responses of mesophytes toplant water deficits, Ann. Rev. Plant Phys. 33 (1982) 163–203.
[51] M. Houde, J. Danyluk, J.F. Laliberte, E. Rassart, R.S. Dhindsa,F. Sarhan, Cloning, characterization, and expression of a cDNAencoding a 50-kilodalton protein specifically induced by coldacclimation in wheat, Plant Physiol. 99 (1992) 1381–1387.
[52] L.A. Bravo, T.J. Close, L.J. Corcuera, C.L. Guy, Characteriza-tion of an 80-kDa dehydrin-like protein in barley responsive tocold acclimation, Physiol. Plant 106 (1999) 177–183.
[53] K. Van Zee, F.Q. Chen, P.M. Hayes, T.J. Close, T.H.H. Chen,Cold-specific induction of a dehydrin gene family member inbarley, Plant Physiol. 108 (1995) 1233–1239.
[54] J. Danyluk, A. Perron, M. Houde, A. Limin, B. Fowler, N.Benhamou, F. Sarhan, Accumulation of an acidic dehydrin inthe vicinity of the plasma membrane during cold acclimation inwheat, The Plant Cell 10 (1998) 623–638.
[55] A.J. Robertson, A. Weninger, R.W. Wilen, P. Fu, L.W. Gusta,Comparison of dehydrin gene expression and freezing tolerancein Bromus inermis and Secale cereale grown in controlled envi-ronments, hydroponics, and the field, Plant Physiol. 106 (1994)1213–1216.
[56] R. Arora, M.E. Wisniewski, Cold acclimatiom in geneticallyrelated (sibling) deciduous and evergreen peach (Prunus persica[L.] Batsch): a 60-kilodalton bark protein in cold acclimatedtissues of peach is heat-stable and related to the dehydrin familyof proteins, Plant Physiol. 105 (1994) 95–101.
[57] B.D. Baldwin, M.S. Bandara, K.K. Tanino, Is tissue culture aviable system with which to examine environmental and hor-monal regulation of cold acclimation in woody plants?, Physiol.Plant 102 (1998) 201–209.
Copyright © 2022 FDOKUMEN




















