
N-terminal CFTR missense variants severely affect the behavior of the CFTR chloride channel

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
© The Histochemical Society, Inc.
0022-1554/04/$3.30
193
ARTICLE
Volume 52(2): 193–203, 2004Journal of Histochemistry & Cytochemistry
http://www.jhc.org
CFTR Localization in Native Airway Cells and Cell Lines Expressing Wild-type or F508del-CFTR by a Panel of Different Antibodies
Isabel Carvalho–Oliveira, Alexandra Efthymiadou, Rui Malhó, Paulo Nogueira, Maria Tzetis, Emmanuel Kanavakis, Margarida D. Amaral, and Deborah Penque
Centro de Genética Humana (ICO,MDA,DP), Observatório Nacional de Saúde (PN), Instituto Nacional de Saúde Dr Ricardo Jorge, Lisboa, Portugal; Department of Medical Genetics, University of Athens, Aghia Sophia Children’s Hospital, Athens, Greece (AE,MT,EK); and Departamento de Biologia Vegetal (RM), Departamento de Química e Bioquímica (MDA), Faculdade de Ciências de Lisboa, Universidade de Lisboa, Lisboa, Portugal.
SUMMARY
The intracellular localization of cystic fibrosis transmembrane conductanceregulator (CFTR) in native tissues is a major issue in the study of mutation, processing, andtrafficking effects in CFTR and in the evaluation of therapeutic strategies in cystic fibrosis(CF). This work evaluated the applicability of ten different antibodies (Abs) under variousfixation techniques for CFTR localization in fresh-brushed nasal epithelial cells collectedfrom CF patients homozygous for F508del and control individuals. In parallel, the same Abpanel was also tested on BHK cell lines overexpressing wild-type or F508del CFTR. The AbsMATG1061, 169, Lis1, MP-CT1, CC24-R, MAB25031, and MAB1660 gave the best detectionof CFTR in the apical region (AR) of nasal tall columnar epithelial (TCE) cells. The labelingpattern of these Abs was consistent with the postulated processing defect of F508del CFTRbecause only a minority of CF TCE cells present CFTR in the AR. In contrast, M3A7, MM13-4,and L12B4 weakly react with the AR and stain almost exclusively a
cis
-Golgi-like structure inthe majority of CF and non-CF airway cells. In BHK cells, all the Abs enabled distinction be-tween wild-type CFTR localization in cell membrane from F508del CFTR, which in thesecells is exclusively located in the endoplasmic reticulum. (
J Histochem Cytochem 52:193–203, 2004
)
C
ystic fibrosis
(CF) is the most common autosomalrecessive genetic disorder in whites. The disease is causedby mutations in the gene coding for the CF transmem-brane conductance regulator (CFTR), a glycoproteinthat functions as a cAMP-regulated chloride (Cl
�
) chan-nel in the apical membrane of epithelial cells (Riordan etal. 1989). Despite the great diversity of CF-causingmutations (to date, almost 1000 have been docu-mented), most individuals with CF carry at least oneallele with a deletion of a trinucleotide resulting in theloss of phenylalanine at position 508 (F508del) (Col-lins 1992). Data on CFTR expression and function are
mostly based on heterologous expression studies usingcultured human cells or cell lines
.
These studies indicatethat the F508del CFTR protein is misprocessed and mis-localized in the endoplasmic reticulum (ER) and is un-able to reach the cell surface (Cheng et al. 1990). Incontrast, data on the trafficking defect of F508delCFTR in fresh human native tissue are sparse becausethe procedures for sample collection are invasive andrisky and the resultant tissue is of poor quality. De-spite these difficulties, the CFTR expression in nativetissues appears to be low and can be modified accord-ing to the degree of tissue differentiation (Trezise andBuchwald 1991; Kartner et al. 1992; Brezillon et al.1995; Dupuit et al. 1995). Most existing CFTR anti-bodies (Abs) have been tested in CFTR-overexpressingcells and therefore may not have the sensitivity requiredfor native tissues.
Nevertheless, some studies have demonstrated mis-
Correspondence to: Deborah Penque, PhD, Centro de GenéticaHumana, Instituto Nacional de Saúde Dr Ricardo Jorge, 1649-016Lisboa, Portugal. E-mail: [email protected]
Received for publication February 18, 2003; accepted Septem-ber 24, 2003 (3A6023).
KEY WORDS
cystic fibrosis
CFTR
CFTR antibody
immunocytochemistry
nasal brushing
epithelial cells
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
194
Carvalho–Oliveira, Efthymiadou, Malhó, Nogueira, Tzetis, Kanavakis, Amaral, Penque
localization of F508del CFTR in sweat gland (Kartneret al. 1992) and in airway epithelium (Engelhardt andWilson 1992; Puchelle et al. 1992). Kälin et al. (1999)indicate that protein levels and localization are similarbetween F508del and normal CFTR protein when ex-pressed in airway (epithelial cells observed in nasalpolyps) and gut but not when expressed in sweatgland (Kälin et al. 1999). Using nasal epithelial cells,freshly obtained by brushing, we have confirmed thatF508del CFTR, albeit in a reduced number (22%)compared to F508del carrier (42%) and healthy cells(56%), does process through the ER and reaches thecell surface (Penque et al. 2000). The correct localiza-tion of F508del CFTR at the apical membrane in afraction of CF airway cells is nevertheless consistentwith the concept that the F508del mutation impairsthe intracellular trafficking of F508del CFTR protein.However, the trafficking defect of F508del CFTRcould also occur at a subcellular level other than ERretention and degradation.
In summary, recent findings clearly indicate thatadditional studies on native human tissues would leadto a better understanding of the processing, traffick-ing, and perhaps the function of normal and mutantCFTR. To establish the conditions under which nor-mal and mutated CFTR can be studied in native tis-sue, we have compared by different immunofluores-cence procedures, a panel of 10 commonly used CFTRAbs for CFTR detection in freshly brushed nasal epi-thelial cells collected from healthy individuals andfrom CF patients homozygous for F508del. In paral-lel, the same antibodies were tested in BHK cell linesstably expressing wild-type (wt) or F508del CFTR.
Materials and Methods
Individuals and Nasal Brushing Cells
After informed consent, nasal brushing cells were obtainedas previously described (Penque et al. 2000) from CF pa-tients homozygous for F508del (
n
�
7) and normal individu-als (
n
�
12) with no clinical signs of CF or F508del mutation,as determined by genomic DNA analysis. Genotypes of allindividuals studied were determined as described before(Duarte et al. 1996).
Cell Lines
Baby hamster kidney (BHK) cell lines expressing wt orF508del CFTR were cultivated as described (Chang et al.1993; Haardt et al. 1999). Cells were grown in an 8-wellchamber slide system (Nalge Nunc; Roskilde, Denmark) at acell density of 2
�
10
4
cells/per well before being analyzedby ICC.
Antibodies
A panel of the following anti-CFTR monoclonal (MAb) andpolyclonal (PAb) antibodies, previously shown to recognizeCFTR, was used: MATG 1061 MAb, raised against aa 503–
515 in the N-terminal (Transgène; Strasbourg, France) (Pu-chelle et al. 1992); 169 PAb, raised against a peptide com-prising amino acids (aa) 724–746 in exon 13 of the R do-main of CFTR (Crawford et al. 1991); Lis1 PAb, raisedagainst aa 1468–1480 in the C-terminus [produced in ourlaboratory (Farinha et al. 2002; and unpublished data)];MP-CT1 PAb raised against a CFTR epitope in the C-termi-nus (Lloyd Mills et al. 1992); CC24-R PAb, raisedagainst aa 693–716 of the R-domain (Picciotto et al. 1992);MAB25031 MAb, raised against exon 23 and 24, aa 1377–1480 of the C-terminus and MAB1660 MAb, raised againsta CFTR epitope in the R-domain (R&D Systems; Minneap-olis, MN); M3A7 MAb, raised against aa 1197–1480 of theC-terminal end of NBD2, L12B4 MAb, against aa 386–412of NBD1, and MM13-4 MAb, against aa 24–35 of theN-terminus (Chemicon International; Temecula, CA). IgGconcentrations and the corresponding dilutions used aregiven in Table 1.
The microtubule organizing center (MTOC)was stained by MAb anti-
�
-tubulin, (1:60). The MAb anti-
�
-COP (1:80) and MAb anti-Golgi 58K protein (1:60) wereused to stain the Golgi apparatus (Sigma Chemical; St Louis,MO). Secondary Abs were FITC-conjugated anti-rabbit IgG(Amersham Pharmacia Biotech; Poole UK), diluted 1:50 andFITC-conjugated anti-mouse IgG (Jackson ImmunoResearchLaboratories; West Chester, PA) diluted 1:100.
Immunocytochemistry
Nasal cells were recovered from the brushes and washed inPBS as previously described (Penque et al. 2000). For form-aldehyde fixation, cells were resuspended and fixed while insuspension in 4% (v/v) formaldehyde (Merck, Darmstadt,Germany; ref 104002), 3.7% (w/v) sucrose in PBS for 30min at 4C and resuspended in cold PBS. Cells were then ad-hered onto silane-coated glass slides (Sigma Chemical) bycentrifugation for 5 min at 2000 rpm in a Cytospin 3 centri-fuge (Shandon; Life Sciences International, Cheshire, UK).The cells adhered on slides were or were not additionallyfixed in methanol
�
20C for 5 min (formaldehyde/methanolfixation). For acetone or acetic acid fixation, both at
�
20Cfor 5 min, the cells were fixed only after being adhered ontoglass slides. For the following ICC analysis, two protocolswere used. The first, previously described by us (Penque etal. 2000), was applied for all Abs using different fixing solu-tions as described above. The second, as described by Dor-mer et al. (2001b), was tested for some Abs in cells that werefixed in formaldehyde.
Briefly, in our protocol, the cells on slides were permeabi-lized with 0.2% (v/v) Triton X-100 in PBS for 20 min,washed three times in PBS for 5 min each, and antigenblocked with 1% (w/v) BSA/PBS for 45 min before incuba-tion overnight at 4C with one of the anti-CFTR Abs (see Ta-ble 1). The cells were then washed three times with PBS for10 min each and incubated with the secondary Ab for 45min at RT. Or, as in Dormer’s protocol, the antigen wasblocked by goat serum 1:20 for 20 min at RT before incuba-tion with the primary Ab without cell permeabilization. Thewashes between the primary and secondary Ab incubationswere carried out using 1% Tween-20/PBS, repeated threetimes for 5 min each.
In both protocols, negative controls were performedomitting the primary Ab. The slides were mounted with
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
CFTR Localization by a Panel of Antibodies
195
Vectashield (Vector Laboratories; Burlingame, CA) contain-ing DAPI (4,6-diamino-2-phenylindole; Sigma Chemical) fornuclear staining and covered with a glass coverslip. Immuno-fluorescence was observed using an Olympus BX-51 with anOlympus X100 UplanApo (NA
�
1.4) objective fluores-cence microscope and images collected with a V-scan cooledcharged-coupled device (CCD) camera (Photonic Science;Millham, UK). Fluorescence analysis and image processingwere performed with Image Pro Plus 4.0 software (MediaCybernetics; Leiden, The Netherlands).
The ICC of BHK cell lines expressing wt or F508del wasperformed on chamber slides using our protocol (Penque etal. 2000).
Evaluation of Cell Types Recovered by Nasal Brushing
The freshly isolated human cells recovered from nasal brush-ings and spread on silane glass slides were stained by theMay–Grünwald–Giemsa (MGG) method and the epithelialcells classified exactly as described by Penque et al. (2000).
Statistical Analysis
For some Abs (see Table 2 and Figure 4),
about 100 TCEcells per slide were assessed for the presence or absence of
CFTR labeling at the apical region, exactly as described pre-viously (Penque et al. 2000). Data are summarized bymean
�
standard deviation (SD) and by mean
�
quartile de-viation (QD). Statistical significance comparisons betweenCF and non-CF cells were made using both the parametricStudent’s
t
-test and the nonparametric Mann–Whitney
U
-test for two independent samples (Sokal and Rohlf 1981).Coefficients with a
p
value less than 0.05 were consideredstatistically significant.
Results
Previously, we showed by immunofluorescence thatthree currently used anti-CFTR Abs raised against dif-ferent epitopes (169, MATG1061, and M13-1), wereable to detect CFTR at the apical region (AR) of tallcolumnar epithelial (TCE) cells freshly obtained fromnasal epithelia of CF and non-CF individuals (Penqueet al. 2000). Here we tested different cell fixationprocedures and compared, using the same protocol,whether eight additional anti-CFTR Abs (Table 1) arecapable of detecting and evaluating the presence ofCFTR in AR of those non-CF and CF TCE cells. Two
Table 1
Description of anti-CFTR antibodies tested in different fixation solutions
Abs Origin Fixation technique
a
Dilution IgG conc. (
�
g /ml)
MATG1061 (MAb) Transgene A, F, FM, AE, D 1:100 90169 (PAb) W. Guggino, Baltimore, MD A, F, FM, D 1:100 10–20MP-CT1 (PAb) R. Dormer, Cardiff, UK A, F, FM, AE, D 1:100 3CC24-R (PAb) H. DeJonge,
Rotterdam, The NetherlandsA, F, FM 1:100 2.3
Lis 1 (PAb) Our laboratory F, FM, AE, D 1:20/1:50/1:100 5–20MAB25031 (MAb) R&D Systems A, F, FM 1:10 10–20MAB1660 (MAb) R&D Systems A, F, FM 1:10/1:20 10–20M3A7 (MAb) Chemicon A, F, FM, AE 1:50/1:100 10–20MM13-4 (MAb) Chemicon A, F, FM, AE 1:50 20L12B4 (MAb) Chemicon F, FM, AE, D 1:50 20
a
A, acetone at
�
20C; F, formaldehyde at 4C; FM, formaldehyde at 4C plus methanol at
�
20C; AE, acetic acid/ethanol at
�
20C; D, Dormer et al. (2001a) protocolon formaldehyde-fixed cells.
Table 2
Significance tests for comparison in the proportion of TCE cells with CFTR in the AR for different anti-CFTR Abs
a
Ab Statistical parameters Controls CF patients Test
/p
value
MATG 1061 Mean
�
SD 58% (
�
4.5%) (
n
�
3) 21% (
�
1.6%) (
n
�
3) Student
t
/
p
0.001
b
Median
�
QD 58% (
�
0%) (
n
�
3) 21% (
�
0%) (
n
�
3) Mann–Whitney/
p
�
0.05169 Mean
�
SD 61% (
�
14.7%) (
n
�
4) 23% (
�
4.8%) (
n
�
3) Student
t
/
p
�
0.008
b
Median
�
QD 61% (
�
27.7%) (
n
�
4) 25% (
�
0%) (
n
�
3) Mann–Whitney/
p
�
0.034
b
MP-CT1 Mean
�
SD 57% (
�
10.2%) (
n
�
6) 18% (
�
5.0%) (
n
�
5) Student
t
/
p
0.001
b
Median
�
QD 60% (
�
17.4%) (
n
�
6) 20% (
�
9.4%) (
n
�
5) Mann–Whitney/
p
�
0.006
b
CC24-R Mean
�
SD 60% (
�
9.7%) (
n
�
4) 14% (
�
3.2%) (
n
�
3) Student
t
/
p
�
0.001
b
Median
�
QD 58% (
�
17.9%) (
n
�
4) 13% (
�
0%) (
n
�
3) Mann–Whitney/
p
�
0.034
b
Lis 1 Mean
�
SD 54% (
�
12.3%) (
n
�
8) 16% (
�
8.7%) (
n
�
5) Student
t
/
p
0.001
b
Median
�
QD 56% (
�
25.5%) (
n
�
8) 15% (
�
15%) (
n
�
5) Mann–Whitney/
p
�
0.003
b
MAB25031 Mean
�
SD 52% (
�
6.0%) (
n
�
5) 19% (
�
6.9%) (
n
�
4) Student
t/p0.001b
Median � QD 49% (�11.4%) (n�5) 17% (�13%) (n�4) Mann–Whitney/p�0.014b
MAB1660 Mean � SD 48% (�0.3%) (n�2) 21% (�9.5%) (n�3) Student t/p�0.032b
Median � QD 48% (�0%) (n�2) 15% (�0%) (n�3) Mann–Whitney/p�0.083
aTests were performed on formaldehyde fixed cells as described as Penque et al. (2000).bDifferences significant at p0.05.
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
196 Carvalho–Oliveira, Efthymiadou, Malhó, Nogueira, Tzetis, Kanavakis, Amaral, Penque
of the Abs used previously, MATG1061 and 169,were also included in this study. Four different fixa-tion procedures were tested: acetone �20C, formalde-hyde at 4C, formaldehyde at 4C followed by an addi-tional fixation in methanol at �20C (formaldehyde/methanol fixation), and acetic acid/ethanol fixation.Some Abs were also tested using the immunocytochem-istry protocol described by (Dormer et al. 2001a) innasal brushing cells fixation by formaldehyde fixationsolution. All 10 Abs (Table 1) were also used to evalu-ate the CFTR localization in BHK cell lines stably ex-pressing wt or F508del CFTR (see below).
CFTR Labeling in Nasal Cells UnderDifferent Procedures
CFTR labeling was first tested using our protocol(Penque et al. 2000) on acetone-, formaldehyde-,formaldehyde/methanol-, or acetic acid-fixed nasalcells obtained by brushing from control individuals.Fixation of cells in suspension, i.e., before adherenceonto glass slides, by either methanol, acetone, or ace-tic acid showed an increase in cell aggregation andshrinking, perhaps caused by cell membrane delipida-tion. Therefore, these fixation solutions were tested oncells already spun onto glass slides. The type of fixa-tion tested and the dilution for each one of the CFTRantibodies is described in Table 1. The results indi-cated that the fixation solutions did not significantlyalter the signal patterns obtained for the Abs tested(data not shown). The subsequent antibody tests weretherefore performed exclusively on formaldehyde-fixed cells. This fixation solution provides better cellmorphology preservation and can be used immedi-ately after obtaining the cells while convenientlyallowing storage at 4C for up to 1 week. Formalde-hyde-fixed cells were also analyzed by the protocol ofDormer et al. (2001b) using some CFTR Abs from thepanel (see Table 1). The results showed essentially thesame distribution of CFTR localization patterns asthose obtained by our protocol (see below).
CFTR Labeling in Control Nasal Brushing Cells
Considerable familiarity has been gained with the per-formance of each antibody. All Abs tested detectedCFTR in the AR of the nasal TCE cells of normalhealthy controls. However, the results shown in Fig-ure 1 reveal some differences in the Ab sensivity forCFTR detection in this region. The MATG1061 MAbvery clearly detects CFTR localized at the AR in themajority of nasal TCE cells (Figure 1B, arrowheads).With this MAb, almost no staining reaction is ob-served in the cytoplasm or in the nucleus of TCE cells(Figure 1B). The 169 PAb detects CFTR at the AR ofTCE cells (Figure 1C, arrowheads). However, cyto-plasmic labeling distribution and nuclear spot cross-
staining were also observed in TCE cells with this an-tibody. The PAb Lis1 clearly stains the AR of TCEcells (Figure 1D, arrowhead) but also exhibits somecrossreaction with cilia and nuclei of some cells (Fig-ure 1D). The PAbs MP-CT1 and CC24-R also providesuitable labeling for CFTR detection at the AR (Fig-ures 1E and 1F, arrowheads) despite some crossreac-tion with cilia (Figures 1E and 1F). Both Abs also staina cytoplasmic region near the nucleus in a small pro-portion of TCE cells (Figures 1E and 1F, arrows).These labeling patterns did not significantly changewhen we applied the protocol of Dormer et al. (2001b)(data not shown). CFTR at the AR is also detected byeither MAb MAB25031 or MAB1660 (Figures 1G and1H). However, the intensity of the background label-ing in the cytoplasm of TCE cells hinders a positiveclear signal at the AR (Figures 1E and 1F). In ourhands, when both Abs are combined in a single detec-tion procedure (see Materials and Methods), the CFTRat the AR is more clearly detected because the back-ground problem is reduced (Figure 1I, arrowhead).
In contrast to the Abs described above, MAbsM3A7, MM13-4, and L12B4 hardly detect CFTR atthe AR in a majority of the cell samples analyzed(Figures 1J–1L, arrowheads). Moreover, these MAbsstrongly stain an intracellular structure localized closeto the nucleus of TCE cells (Figures 1J–1L, arrows).This structure is also observed in basal cells. Thestaining pattern of this structure is very similar tothose obtained with MAbs against proteins of theGolgi apparatus, such as the anti-�-COP (Figure 1M,arrow) and anti-p58 Golgi proteins (Figure 1N, ar-row). Surprisingly, some control individuals (4/9 con-trol individuals) were consistently and repeatedly neg-ative (n�4 experiments) for those three CFTR MAbs(M3A7, MM13-4, L12B4). This observation corre-lated with a negative result for the anti-�-COP MAbfor those same individuals despite being positive forthe anti-p58 Golgi protein (results not shown). �-COPprotein is the major component of the COP-I proteincomplex, which is critical for vesicular trafficking be-tween the endoplasmic reticulum and the Golgi.�-COP is also a useful marker for the vesicotubularclusters at the cis face of the Golgi stack as well asthe cis Golgi itself (Oprins et al. 1993). Golgi p58, aformiminotransferase cyclodeaminase, is used as a se-lective marker for the cytoplasmic face of the Golgiapparatus (Gao et al. 1998). In nasal TCE cells, thestaining pattern of both Abs anti-�-COP and anti-p58are very similar in location and morphology, suggest-ing that both Golgi structures are very close to eachother in these cells. However, they seem to be distinctstructures because in the same individuals no reactionwas consistently obtained with anti-�-COP despite thepositive reaction to anti-p58 Golgi protein.
The intracellular structure stained by M3A7, L12B4,
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
CFTR Localization by a Panel of Antibodies 197
Figure 1 Immunolocalization of CFTR, �-COP, p58K, and �-tubulin in non-CF nasal brushing cells fixed with formaldehyde. Negative controlswere performed by incubation with the secondary Abs, anti-mouse IgG (1:100; A1) or anti-rabbit IgG (1:50; A2). Labeling with Abs MATG1061(1:100; B), 169 (1:100; C), Lis1 (1:50; D), MP-CT1 (1:100; E), CC24-R (1:100; F), MAB25031 (1:10; G), MAB1660 (1:10; H), MAB25031/MAB1660combined (1:5; I), M3A7 (1:50; J), L12B4 (1:50; K), and MM13-4 (1:50; L) detect CFTR at AR of TCE cells (arrowheads in B–L). The Abs M3A7,L12B4 and MM13-4 label mostly exclusively a Golgi-like intracellular structure in cells (arrows in J–L). This structure seems to be also weaklystained by the Abs Lis1, MP-CT1, and CC24-R (arrows in D–F). The staining pattern of this structure is very similar to that stained by the �-COPand p58K MAb markers for Golgi apparatus (arrows in M and N). The �-tubulin MAb detects the MTOC in the posterior site of the TCE cells(arrow in O) in a very distinct region of the Golgi apparatus. Bar � 15 �m.
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
198 Carvalho–Oliveira, Efthymiadou, Malhó, Nogueira, Tzetis, Kanavakis, Amaral, Penque
Figure 2 Immunolocalization of CFTR in F508del/F508del nasal brushing cells fixed with formaldehyde. Labeling with the Abs MATG1061(1:100; A), 169 (1:100; B), Lis1 (1:50; C), MP-CT1 (1:100; D), CC24-R (1:100; E), MAB25031 (1:10; F), MAB1660 (1:10; G), and MAB25031/MAB1660combined (1:5; H), M3A7 (1:50; I), L12B4 (1:50; J), and MM13-4 (1:50; K) (see Materials and Methods) detect CFTR at AR in a reduced numberof TCE cells (arrowheads in A–K). An intracellular region similar to Golgi apparatus is stained strongly by the Abs M3A7, L12B4, and MM13-4(arrows in I–K) and weakly by the Abs Lis1, MP-CT1, and CC-24 (arrows C–E). Bar � 15 �m.
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
CFTR Localization by a Panel of Antibodies 199
and MM13-4 Abs is localized in a very distinct placefrom the microtubule organizing center (MTOC) thatwas detected by anti-�-tubulin MAb in the posteriorregion of TCE cells (Figure 1O, arrow).
CFTR Labeling in F508del-Homozygous Nasal Brushing Cells
Nasal brushing cells of CF patients homozygous forF508del were examined using the 10 CFTR Abs that
had previously been tested on control nasal brushingcells (Table 1).
As shown in Figure 2, all 10 CFTR Abs producedessentially the same labeling pattern in CF nasal cellscompared to control samples. This means that no par-ticular intracellular staining that could be associatedwith the F508del trafficking defect was observed inTCE from F508del-homozygous CF patients. The in-tracellular staining observed in some CF TCE cells
Figure 3 Immunolocalization ofCFTR, �-COP, and p58K in wild-type(WT) and F508del CFTR BHK-express-ing cells fixed with formaldehyde. All10 CFTR Abs tested (MATG1061(1:100), 169 (1:100), Lis1 (1:50), MP-CT1(1:100), CC24-R (1:100), MAB25031(1:10), MAB1660 (1:10), MAB25031/MAB1660 (1:5) combined, M3A7(1:50), L12B4 (1:50), and MM13-4(1:50) label CFTR predominantly lo-cated at the plasma membrane of theWT CFTR BHK-expressing cells. InF508del CFTR BHK-expressing cells,using the same panel of CFTR Abs,the F508del CFTR is detected re-stricted to an area around the nu-cleus of the cells. In BHK cells, thestaining pattern of �-COP protein, acis-Golgi marker, localizes in smallvesicles spread into the cytoplasm,while p58 protein, a Golgi apparatusmarker, is concentrated in a juxtanu-clear region of the cells. Bar � 15 �m.
by guest on February 13, 2016jhc.sagepub.comDownloaded from
The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
200 Carvalho–Oliveira, Efthymiadou, Malhó, Nogueira, Tzetis, Kanavakis, Amaral, Penque
was also observed in some non-CF cells using, e.g., theAbs Lis1, MP-CT1, CC24-R, M3A7, L12B4, orMM13-4 (Figures 3C–3E, and 3I–3K, arrows). Themain difference between CF and non-CF cells seems tobe the number of TCE cells presenting CFTR at theAR (see the results of statistical analysis). As observedwith cells from some non-CF individuals, no reactionwas found with Abs M3A7, L12B4, or MM13-4 in2/5 F508del-homozygous CF patients.
CFTR Labeling in BHK Cells Stably Expressing Wt or F508del CFTR
Because the sensitivity in detecting CFTR in CF andnon-CF TCE cells differed for the 10 antibodies, espe-cially for the three MAbs M3A7, L12B4, and MM13-4,we examined these Abs for CFTR detection in wt- orF508del CFTR-overexpressing cells. All 10 antibodieswere able to distinguish wt CFTR localization fromF508del CFTR in BHK cells stably expressing wt orF508del CFTR, respectively (Figure 3). Whatever theAbs used, wt CFTR is predominantly present at thecell membrane of the cells (Figure 3 WT, arrows). Thelabeling pattern produced in wt cells exhibits longlines between cells, giving the idea that the cells arelonger in comparison to F508del cells. F508del CFTRis mostly restricted to an area around the nucleus (Fig-ure 3 F508del, arrowheads); therefore, cells seemrounder than wt cells. In contrast to the findings fromnasal TCE cell experiments, no intracellular Golgi-likestructures were detected with MAbs M3A7, L12B4and MM13-4 in CFTR-expressing BHK cells. TheseAbs are therefore very sensitive and suitable for CFTRdetection focused at the plasma membrane of cells ex-pressing wt CFTR (see Figure 3 WT, arrows).
The staining pattern of �-COP protein (a cis-Golgimarker) in BHK cells consists of punctate spots spreadthroughout the cytoplasm. In contrast, p58 protein,another Golgi apparatus marker, localizes in a con-centrated juxtanuclear region of BHK cells (Figure 3WT and F508del, �-COP and p58). In nasal TCEcells, both Golgi proteins localize closely in a region ofwhich the staining pattern is a unique intracellularstructure transversely located in the cell above the nu-cleus (see Figures 1M and 1N, arrows).
Statistical Analysis of TCEs with CFTR in the AR
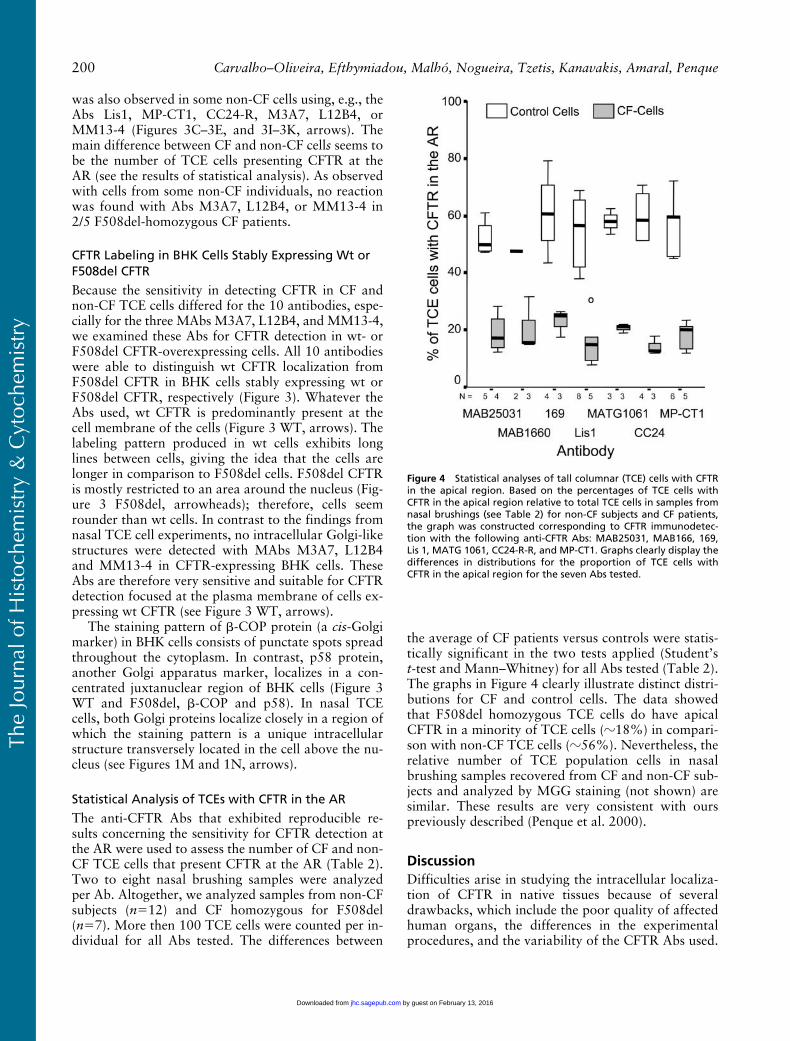
The anti-CFTR Abs that exhibited reproducible re-sults concerning the sensitivity for CFTR detection atthe AR were used to assess the number of CF and non-CF TCE cells that present CFTR at the AR (Table 2).Two to eight nasal brushing samples were analyzedper Ab. Altogether, we analyzed samples from non-CFsubjects (n�12) and CF homozygous for F508del(n�7). More then 100 TCE cells were counted per in-dividual for all Abs tested. The differences between
the average of CF patients versus controls were statis-tically significant in the two tests applied (Student’st-test and Mann–Whitney) for all Abs tested (Table 2).The graphs in Figure 4 clearly illustrate distinct distri-butions for CF and control cells. The data showedthat F508del homozygous TCE cells do have apicalCFTR in a minority of TCE cells (�18%) in compari-son with non-CF TCE cells (�56%). Nevertheless, therelative number of TCE population cells in nasalbrushing samples recovered from CF and non-CF sub-jects and analyzed by MGG staining (not shown) aresimilar. These results are very consistent with ourspreviously described (Penque et al. 2000).
DiscussionDifficulties arise in studying the intracellular localiza-tion of CFTR in native tissues because of severaldrawbacks, which include the poor quality of affectedhuman organs, the differences in the experimentalprocedures, and the variability of the CFTR Abs used.
Figure 4 Statistical analyses of tall columnar (TCE) cells with CFTRin the apical region. Based on the percentages of TCE cells withCFTR in the apical region relative to total TCE cells in samples fromnasal brushings (see Table 2) for non-CF subjects and CF patients,the graph was constructed corresponding to CFTR immunodetec-tion with the following anti-CFTR Abs: MAB25031, MAB166, 169,Lis 1, MATG 1061, CC24-R-R, and MP-CT1. Graphs clearly display thedifferences in distributions for the proportion of TCE cells withCFTR in the apical region for the seven Abs tested.
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
CFTR Localization by a Panel of Antibodies 201
Here we evaluated the applicability of a panel of 10different CFTR Abs to immunolocalize CFTR in CFand non-CF native airway epithelial cells as well as incell lines, BHK wt- or F508del CFTR-overexpressingcells. Cells recovered from the superficial nasal mu-cosa by brushing have been a useful native biologicalmaterial in the assessment of chronic respiratory dis-eases, including CF (Danel et al. 1996). The nasalbrushing technique is a noninvasive method that al-lows the easy sampling of many representative, well-preserved and dissociated cells from the superficial re-spiratory mucosa (Danel et al. 1996). As previouslydescribed (Bridges et al. 1991; Chapelin et al. 1996;Danel et al. 1996; Penque et al. 2000), 80–90% of na-sal cells recovered by brushing are epithelial cells. Inmost cases, nasal polyp surface epithelium representsremodeling with a degree of differentiation, i.e., squa-mous metaplasia that causes a decrease of CFTR ex-pression (Brezillon et al. 1995). In contrast, the epithe-lial cells contained in the nasal brush samples ofindividuals with no polyposis are good representa-tives of normal airway pseudostratified epitheliumand will not, as in nasal polyps, give rise to a CFTRexpression signal that could be associated with epi-thelial abnormalities. Fixing cells immediately afterrecovery from the brushes minimizes any possible al-terations of protein expression related to the stress ofcell dissociation.
MATG 1061 MAb and 169 PAb were previouslytested by us using nasal brushing cells (Penque et al.2000). In this study we confirmed that using a varietyof fixing solutions, both Abs are suitable for evalua-tion of CFTR in the apical region (AR) of CF and non-CF nasal TCE. MATG1061 showing markedly lessbackground staining than 169. Although MATG1061has been applied successfully in several studies of air-way cells (Puchelle et al. 1992; Jacquot et al. 1993;Brezillon et al. 1995; Dupuit et al. 1995; Kälin etal. 1999), Claass et al. (2000) demonstrated thatMATG1061 was nonspecific for CFTR in the sweatglands. However, these authors employed an IHCanalysis using an enhanced enzymatic alkaline phos-phatase–anti-alkaline phosphatase (APAAP) (Cordellet al. 1984) and not an immunofluorescence one.This method could increase nonspecific binding of theMATG1061 in the skin sections.
Using PAb MP-CT1 in nasal brushing cells, Dormeret al. (2001b) recently confirmed our previous results(Penque et al. 2000) showing that a reduced numberof CF TCE cells from CF-homozygous F508del pa-tients have F508del CFTR at the AR. The majority ofTCE cells from those CF patients presents a character-istically perinuclear location of F508del CFTR. UsingMP-CT1 PAb in formaldehyde-fixed cells, we con-firmed the results of Dormer et al. (2001b) concerningthe percentage of non-CF and F508del/F508del TCE
cells that present CFTR in the AR. Using MP-CT1PAb (or any of the other nine Abs) we did not observeCF TCE cells with a particular perinuclear F508delCFTR localization, even with the use of different fixa-tion solutions and/or Dormer’s methodology (Harriset al. 2001) (see CF repository in http://central.igc.gulbenkian.pt/cftr/index.html). Some cytoplasmic stain-ing is also observed in non-CF cells using those Abs.We could not detect apparent differences in the local-ization of CFTR and F508del CFTR in the nasal TCEcells. TCE cells that did not show CFTR labeling atAR, whether from CF or non-CF samples, showedweak widespread staining throughout the cell. Occa-sionally, we observed an intracellular structure barelystained by the Ab MP-CT1, but also by CC24-R andLis1, and more prevalently stained by the Abs M3A7,L12B4, and MM13-4 (see arrows in Figures 1 and 2)but those structures occur indistinctly in the apicalCFTR-positive or -negative cells of both CF and non-CF samples (see below). The MP-CT1 Ab also showedsome crossreaction with cilia under all conditionstested. The same result was observed with the PAbCC24-R, whose epitope is located in the R-domain ofCFTR (Picciotto et al. 1992) and Lis1, raised againstthe C-terminus (Farinha et al. unpublished data).However, PAbs showed good sensitivity in detectionof both wt and F508del CFTR in the AR of nasal TCEcells. The immunostaining patterns of CC24-R andLis1 PAbs were similar to that of MP-CT1 PAb, irre-spective of the fixing solution used. In our experi-ments, CC24-R PAb did not exhibit more sensitivityfor detecting F508del CFTR as observed by (Claass etal. 2000) in sweat glands from patients homozygousfor F508del. F508del CFTR is undetectable in sweatglands using several CFTR Abs but is observed intra-cellularly using CC24-R PAb (Claass et al. 2000).
The MAbs MAB25031 and MAB1660, previouslyproduced by Genzyme (24-1 MAb and 13-1 MAb)and now commercially available from R&D Systems,have been used for ICC analysis in CFTR-overexpress-ing cell lines (Gregory et al. 1990; Denning et al.1992). The MAB25031 MAb has also been used tolocalize CFTR in airway cells (Dupuit et al. 1995;Penque et al. 2000; Castillon et al. 2002). In the studyperformed in skin sections (Claass et al. 2000), bothMAbs MAB25031 and MAB1660 produced nonspe-cific labeling in sweat glands at the concentrationstested. Using a higher concentration, we show that,although the labeling produced by both Abs wasonly faintly visible (MAB25031 slightly better thanMAB1660), both MAbs are able to detect CFTR atthe AR of CF and non-CF TCE cells. The sensitivitycould be improved by the combination of both MAbsin the same reaction. Both MAbs, individually or com-bined, can distinguish wt from F508del CFTR in BHKcells overexpressing wt or F508del CFTR. However,
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
202 Carvalho–Oliveira, Efthymiadou, Malhó, Nogueira, Tzetis, Kanavakis, Amaral, Penque
the combination of both Abs did not improve the re-action in BHK cells (data not shown).
The MAbs M3A7 and L12B4 produced by (Kartneret al. 1992) are now commercially available, as well asMM13-4, from Chemicon. In many studies, M3A7and L12B4 showed specificity for CFTR immunoblot-ting, immuprecipitation, and immunocytochemistry inCFTR-overexpressing cells (Lukacs et al. 1994; Jensenet al. 1995; Kopito 1999; Farinha et al. 2002). This re-sult is confirmed here for BHK cells overexpressing wtor F508del CFTR. Although, Kartner et al. (1992)originally showed the mislocalization of F508delCFTR in sweat duct biopsies from CF patients (incomparison with wt CFTR in control skin biopsies),they failed to show CFTR localization with M3A7and L12B4 in airway tissue. In all conditions testedhere, M3A7, L12B4, and MM13-4 hardly detectedCFTR at the apical region of nasal epithelial cells,confirming the previous observation by Kartner et al.(1992). Our data show that these three MAbs reactpreferentially and very strongly with an intracellularstructure similar in location and morphology to theGolgi apparatus. Moreover, some CF and non-CF in-dividuals always show negative reaction for thoseCFTR Abs. This observation correlated with a nega-tive result for anti �-COP Abs in those same individu-als despite being positive for the anti-p58 Golgi pro-tein. By using M3A7 and L12B4, Kartner et al. (1992)showed individual variation concerning the detectionof dense staining bodies throughout the sweat coil ofsome CF and F508del carrier individuals. Kartner etal. (1992) attributed this variation to the ability ofsome cells to completely degrade trapped F508del inthe cell. Because we observed intracellular structuresalso labeling non-CF nasal TCE cells that do not ex-press F508del CFTR, the explanation advanced byKartner et al. (1992) does not fit our data. Neverthe-less, it has been described that most normal CFTRnever reaches the cell surface, being degraded intracel-lularly (see Kopito 1999 for review). Therefore, somenon-CF (and non-F508 carrier) individuals could alsoshow some variability in the ability to degrade normalCFTR. Although this is an interesting possibility,more experiments should be performed to clarify theseobservations. Caution must be taken in interpretingresults with those Abs while examining fresh nasal ep-ithelium.
In nasal TCE cells, the staining pattern obtainedwith anti-� COP and anti-p58 Abs suggests that cis-Golgi vesicles and Golgi structures are too close to bedistinguished by immunofluorescence methods. In con-trast, in BHK cells both structures are clearly distinctin location and morphology. This finding is probablydue to differences in the state of polarization of cellsand should be considered when trafficking studies areperformed in these cells.
In summary, all the CFTR Abs tested are able todistinguish wt from F508del CFTR location in BHKcells overexpressing wt or F508del CFTR. However,in native nasal epithelial cells obtained by brushing,7/10 Abs tested, MATG 1061 (provided by Trans-gène), 169, Lis1, MP-CT1, CC24-R, MAB1660, andMAB2560 (the last two commercially available fromR&D Systems) exhibited sensitivity for CFTR detec-tion in the AR. These Abs can be used to assess CFTRlocalization in the AR of native respiratory cells be-cause they provide a broad consensus, i.e., the Absshow that F508del-homozygous nasal TCE cells donot have apical CFTR in a minority of cells in compar-ison to non-CF nasal TCE cells, which exhibit CFTRin the majority of cells.
AcknowledgmentsSupported in part by a PRAXIS XXI PCOTI/1999/MGI/
35737 research grant. ICO is recipient of an FCT doctoralfellowship. AE was recipient of a travel grant from the Euro-pean CF Network (EU-QLK3-1999-00241).
We thank Dr C. Barreto (HSM, Lisbon), CF patients andcontrol individuals for their cooperation, Dr W. Guggino(Baltimore, MD) for 160 Ab, Dr H. DeJonge (Rotterdam,The Netherlands), for CC24-R Ab, Dr R.L. Dormer (Cardiff,UK), for the MP-CT1 Ab, Transgène (Strasbourg, France)for the MATG1061 Ab, C. Cunha (FCUL, Lisbon) for tech-nical assistance, and H. Davidson (Edinburgh, UK) for dis-cussion and revision of the manuscript.
Literature CitedBrezillon S, Dupuit F, Hinnrasky J, Marchand V, Kalin N, Tummler
B, Puchelle E (1995) Decreased expression of the CFTR proteinin remodeled human nasal epithelium from non-cystic fibrosispatients. Lab Invest 72:191–200
Bridges MA, Walker DC, Davidson AG (1991) Cystic fibrosis andcontrol nasal epithelial cells harvested by a brushing procedure[Letter]. In Vitro Cell Dev Biol 27A:684–686
Castillon N, Hinnrasky J, Zahm JM, Kaplan H, Bonnet N, CorlieuP, Klossek JM, et al. (2002) Polarized expression of cystic fibro-sis transmembrane conductance regulator and associated epithe-lial proteins during the regeneration of human airway surface ep-ithelium in three-dimensional culture. Lab Invest 82:989–998
Chang XB, Tabcharani JA, Hou YX, Jensen TJ, Kartner N, Alon N,Hanrahan JW, et al. (1993) Protein kinase A (PKA) still activatesCFTR chloride channel after mutagenesis of all 10 PKA consen-sus phosphorylation sites. J Biol Chem 268:11304–11311
Chapelin C, Coste A, Gilain L, Poron F, Verra F, Escudier E (1996)Modified epithelial cell distribution in chronic airways inflam-mation. Eur Respir J 9:2474–2478
Cheng SH, Gregory RJ, Marshall J, Paul S, Souza DW, White GA,O’Riordan CR, et al. (1990) Defective intracellular transport andprocessing of CFTR is the molecular basis of most cystic fibrosis.Cell 63:827–834
Claass A, Sommer M, De Jonge H, Kalin N, Tummler B (2000) Ap-plicability of different antibodies for immunohistochemical local-ization of CFTR in sweat glands from healthy controls and frompatients with cystic fibrosis. J Histochem Cytochem 48:831–837
Collins FS (1992) Cystic fibrosis: molecular biology and therapeuticimplications. Science 256:774–779
Cordell JL, Falini B, Erber WN, Ghosh AK, Abdulaziz Z, Mac-Donald S, Pulford KA, et al. (1984) Immunoenzymatic labelingof monoclonal antibodies using immune complexes of alkaline
by guest on February 13, 2016jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
CFTR Localization by a Panel of Antibodies 203
phosphatase and monoclonal anti-alkaline phosphatase (APAAPcomplexes). J Histochem Cytochem 32:219–229
Crawford I, Maloney PC, Zeitlin PL, Guggino WB, Hyde SC, Tur-ley H, Gatter KC, et al. (1991) Immunocytochemical localizationof the cystic fibrosis gene product CFTR. Proc Natl Acad SciUSA 88:9262–9266
Danel C, Erzurum SC, McElvaney NG, Crystal RG (1996) Quanti-tative assessment of the epithelial and inflammatory cell popula-tions in large airways of normals and individuals with cystic fi-brosis. Am J Respir Crit Care Med 153:362–368
Denning GM, Ostedgaard LS, Cheng SH, Smith AE, Welsh MJ(1992) Localization of cystic fibrosis transmembrane conduc-tance regulator in chloride secretory epithelia. J Clin Invest89:339–349
Dormer RL, Derand R, McNeilly CM, Mettey Y, Bulteau–PignouxL, Metaye T, Vierfond JM, et al. (2001a) Correction of delF508-CFTR activity with benzo(c)quinolizinium compounds throughfacilitation of its processing in cystic fibrosis airway cells. J CellSci 114:4073–4081
Dormer RL, McNeilly CM, Morris MR, Pereira MM, Doull IJ,Becq F, Mettey Y, et al. (2001b) Localisation of wild-type andDeltaF508-CFTR in nasal epithelial cells. Pflugers Arch 443(suppl 1):S117–120
Duarte A, Amaral M, Barreto C, Pacheco P, Lavinha J (1996) Com-plex cystic fibrosis allele R334W-R1158X results in reduced lev-els of correctly processed mRNA in a pancreatic sufficient pa-tient. Hum Mutat 8:134–139
Dupuit F, Kalin N, Brezillon S, Hinnrasky J, Tümmler B, Puchelle E(1995) CFTR and differentiation markers expression in non-CFand delta F 508 homozygous CF nasal epithelium. J Clin Invest96:1601–1611
Engelhardt JF, Wilson JM (1992) Gene therapy of cystic fibrosislung disease. J Pharm Pharmacol 44 (suppl 1):165–167
Farinha CM, Nogueira P, Mendes F, Penque D, Amaral MD (2002)The human DnaJ homologue (Hdj)-1/heat-shock protein (Hsp)40 co-chaperone is required for the in vivo stabilization of thecystic fibrosis transmembrane conductance regulator by Hsp70.Biochem J 366:797–806
Gao YS, Alvarez C, Nelson DS, Sztul E (1998) Molecular cloning,characterization, and dynamics of rat formiminotransferase cy-clodeaminase, a Golgi-associated 58-kDa protein. J Biol Chem273:33825–33834
Gregory RJ, Cheng SH, Rich DP, Marshall J, Paul S, Hehir K, Ost-edgaard L, et al. (1990) Expression and characterization of thecystic fibrosis transmembrane conductance regulator [see Com-ments]. Nature 347:382–386
Haardt M, Benharouga M, Lechardeur D, Kartner N, Lukacs GL(1999) C-terminal truncations destabilize the cystic fibrosistransmembrane conductance regulator without impairing its bio-genesis. A novel class of mutation [In Process Citation]. J BiolChem 274:21873–21877
Harris CM, McPherson M, Dormer RL. (2001) Assessment ofCFTR localisation in native epithelial cells obtained by brush-
ing. Virtual Repository in http://central.igc.gulbenkian.pt/cftr/index.html. Ref Type: Generic
Jacquot J, Puchelle E, Hinnrasky J, Fuchey C, Bettinger C, SpilmontC, Bonnet N, et al. (1993) Localization of the cystic fibrosistransmembrane conductance regulator in airway secretory glands[see Comments]. Eur Respir J 6:169–176
Jensen TJ, Loo MA, Pind S, Williams DB, Goldberg AL, Riordan JR(1995) Multiple proteolytic systems, including the proteasome,contribute to CFTR processing. Cell 83:129–135
Kälin N, Claass A, Sommer M, Puchelle E, Tummler B (1999)DeltaF508 CFTR protein expression in tissues from patients withcystic fibrosis. J Clin Invest 103:1379–1389
Kartner N, Augustinas O, Jensen TJ, Naismith AL, Riordan JR(1992) Mislocalization of delta F508 CFTR in cystic fibrosissweat gland. Nature Genet 1:321–327
Kopito RR (1999) Biosynthesis and degradation of CFTR. PhysiolRev 79:S167–173
Lloyd Mills C, Pereira MMC, Dormer RL, McPherson MA (1992)An antibody against a CFTR-derived synthetic peptide, incorpo-rated into living submandibular cells, inhibits beta-adrenergicstimulation of mucin secretion. Biochem Biophys Res Commun188:1146–1152
Lukacs GL, Mohamed A, Kartner N, Chang XB, Riordan JR, Grin-stein S (1994) Conformational maturation of CFTR but not itsmutant counterpart (delta F508) occurs in the endoplasmic retic-ulum and requires ATP. EMBO J 13:6076–6086
Oprins A, Duden R, Kreis TE, Geuze HJ, Slot JW (1993) Beta-COPlocalizes mainly to the cis-Golgi side in exocrine pancreas. J CellBiol 121:49–59
Penque D, Mendes F, Beck S, Farinha C, Pacheco P, Nogueira P,Lavinha J, et al. (2000) Cystic fibrosis F508del patients have api-cally localized CFTR in a reduced number of airway cells. LabInvest 80:857–868
Picciotto MR, Cohn JA, Bertuzzi G, Greengard P, Nairn AC (1992)Phosphorylation of the cystic fibrosis transmembrane conduc-tance regulator. J Biol Chem 267:12742–12752
Puchelle E, Gaillard D, Ploton D, Hinnrasky J, Fuchey C, BoutterinMC, Jacquot J, et al. (1992) Differential localization of the cysticfibrosis transmembrane conductance regulator in normal andcystic fibrosis airway epithelium. Am J Respir Cell Mol Biol7:485–491
Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzel-czak Z, Zielenski J, et al. (1989) Identification of the cystic fibro-sis gene: cloning and characterization of complementary DNA[published erratum appears in Science 1989; 29;245:1437]. Sci-ence 245:1066–1073
Sokal R, Rohlf F (1981) Biometry. The Principles and Practice ofStatistics in Biological Research. 2nd ed. New York: Freeman
Trezise AE, Buchwald M (1991) In vivo cell-specific expression ofthe cystic fibrosis transmembrane conductance regulator. Nature353:434–437
by guest on February 13, 2016jhc.sagepub.comDownloaded from
Copyright © 2022 FDOKUMEN




















