
Central Command Regulation of Circulatory Function Mediated by Descending Pontine Cholinergic Inputs...
27
K. Goodchild James R. Padley, Natasha N. Kumar, Qun Li, Thomas B.V. Nguyen, Paul M. Pilowsky and Ann Cholinergic Inputs to Sympathoexcitatory Rostral Ventrolateral Medulla Neurons Central Command Regulation of Circulatory Function Mediated by Descending Pontine Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2007 American Heart Association, Inc. All rights reserved. is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Circulation Research doi: 10.1161/01.RES.0000257370.63694.73 2007;100:284-291; originally published online January 4, 2007; Circ Res. http://circres.ahajournals.org/content/100/2/284 World Wide Web at: The online version of this article, along with updated information and services, is located on the http://circres.ahajournals.org/content/suppl/2007/01/04/01.RES.0000257370.63694.73.DC1.html Data Supplement (unedited) at: http://circres.ahajournals.org//subscriptions/ is online at: Circulation Research Information about subscribing to Subscriptions: http://www.lww.com/reprints Information about reprints can be found online at: Reprints: document. Permissions and Rights Question and Answer about this process is available in the located, click Request Permissions in the middle column of the Web page under Services. Further information Editorial Office. Once the online version of the published article for which permission is being requested is can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Circulation Research in Requests for permissions to reproduce figures, tables, or portions of articles originally published Permissions: by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from by guest on September 29, 2013 http://circres.ahajournals.org/ Downloaded from
Transcript of Central Command Regulation of Circulatory Function Mediated by Descending Pontine Cholinergic Inputs...

K. GoodchildJames R. Padley, Natasha N. Kumar, Qun Li, Thomas B.V. Nguyen, Paul M. Pilowsky and Ann
Cholinergic Inputs to Sympathoexcitatory Rostral Ventrolateral Medulla NeuronsCentral Command Regulation of Circulatory Function Mediated by Descending Pontine
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2007 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000257370.63694.732007;100:284-291; originally published online January 4, 2007;Circ Res.
http://circres.ahajournals.org/content/100/2/284World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2007/01/04/01.RES.0000257370.63694.73.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from by guest on September 29, 2013http://circres.ahajournals.org/Downloaded from

Central Command Regulation of Circulatory FunctionMediated by Descending Pontine Cholinergic Inputs to
Sympathoexcitatory Rostral Ventrolateral Medulla NeuronsJames R. Padley, Natasha N. Kumar, Qun Li, Thomas B.V. Nguyen,
Paul M. Pilowsky, Ann K. Goodchild
Abstract—Central command is a feedforward neural mechanism that evokes parallel modifications of motor andcardiovascular function during arousal and exercise. The neural circuitry involved has not been elucidated. We haveidentified a cholinergic neural circuit that, when activated, mimics effects on tonic and reflex control of circulationsimilar to those evoked at the onset of and during exercise. Central muscarinic cholinergic receptor (mAChR) activationincreased splanchnic sympathetic nerve activity (SNA) as well as the range and gain of the sympathetic baroreflex viaactivation of mAChR in the rostral ventrolateral medulla (RVLM) in anesthetized artificially ventilated Sprague–Dawley rats. RVLM mAChR activation also attenuated and inhibited the peripheral chemoreflex and somatosympatheticreflex, respectively. Cholinergic terminals made close appositions with a subpopulation of sympathoexcitatory RVLMneurons containing either preproenkephalin mRNA or tyrosine hydroxylase immunoreactivity. M2 and M3 receptormRNA was present postsynaptically in only non–tyrosine hydroxylase neurons. Cholinergic inputs to the RVLM ariseonly from the pedunculopontine tegmental nucleus. Chemical activation of this region produced increases in muscleactivity, SNA, and blood pressure and enhanced the SNA baroreflex; the latter effect was attenuated by mAChRblockade. These findings indicate a novel role for cholinergic input from the pedunculopontine tegmental nucleus to theRVLM in central cardiovascular command. This pathway is likely to be important during exercise where a centrallyevoked facilitation of baroreflex control of the circulation is required to maintain blood flow to active muscle. (Circ Res.2007;100:284-291.)
Key Words: baroreflex � exercise � chemoreflex � somatosympathetic
A distinct pattern of tonic and reflex cardiovascularadjustments is mediated by central command to ensure
appropriate muscle and organ perfusion during differentarousal or behavioral states, such as sleep and exercise.1–3
Limited evidence implicates some regions within the ponsand hypothalamus that could provide descending input tocardiovascular control sites4–6; however, the neural circuitryand neurotransmitters involved are yet to be elucidated.
Activation of the central cholinergic system has a profoundeffect on cardiovascular and other autonomic functions.7–18
Systemic or central administration of acetylcholinesteraseinhibitors or muscarinic agonists increases blood pressure,7–11
lowers body temperature,12 and alters respiration.13,14 Pressorresponses can be evoked via activation of muscarinic recep-tors (mAChR) within several cardiovascular nuclei, includingthe posterior hypothalamus,7 nucleus of the solitary tract,15
and rostral ventrolateral medulla (RVLM).10,11 Effects ofcentral mAChR activation on cardiovascular reflexes are lesswell understood.8,16,17
Sympathoexcitatory and hypertensive effects of intrave-nously administered physostigmine are largely mediated byexcitation of RVLM neurons.10,11,18 The RVLM generatesbasal sympathetic vasomotor activity and is a critical synapticrelay in cardiovascular reflexes.19,20 Descending cholinergicprojections to the RVLM arise from neurons in the peduncu-lopontine tegmental nucleus (PPT),21 although local medul-lary neurons may also be a source of cholinergic input.22 Thefunction of this input into the RVLM is unknown. A densecholinergic terminal field is present within the RVLM,11,22,23
although supportive anatomical evidence that cholinergicterminals provide input to C1 or non-C1 spinally projectingneurons is lacking. Activation of the inhibitory M2 mAChRsubtype in the RVLM is thought to mediate pressor respons-es,10,11 but its cellular location or that of other mAChRsubtypes within the RVLM is unknown.
We hypothesized that cholinergic input to the RVLM fromthe PPT is involved in central command–mediated effects oncardiovascular function. Previous studies have shown that the
Original received October 13, 2006; revision received November 29, 2006; accepted December 20, 2006.From the Hypertension and Stroke Research Laboratories, Kolling Institute of Medical Research, Royal North Shore Hospital and School of Medical
Sciences, University of Sydney, Australia.Correspondence to Ann K. Goodchild, Hypertension and Stroke Research Labs, Building 10, Royal North Shore Hospital, St Leonards NSW 2065,
Australia. E-mail [email protected]© 2007 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000257370.63694.73
284

PPT is involved in initiation of movement and modulation ofmuscle tone during locomotion, exercise, and arousal.4,24,25
Additionally, the PPT connects albeit indirectly with bothmotor and sympathetic outflows.5
We aimed, firstly, to determine the role of the RVLM in theautonomic responses and effects on reflex control of thecirculation evoked by central mAChR activation. Secondly,we identified the mAChR subtypes involved by examininggene expression within phenotypically identified RVLMneurons and determined the exact sources of cholinergic inputto the RVLM. Finally, we determined the tonic and reflexcardiovascular effects generated by chemical stimulation ofthe PPT.
Materials and MethodsAn expanded Materials and Methods section is available in theonline data supplement at http://circres.ahajournals.org.
Studies were approved by the Animal Care and Ethics Committeeof Royal North Shore Hospital/University of Technology, Sydney.Following experimentation, rats were euthanized while under surgi-cal anesthesia by KCl (3 mol/L, 1 mL IV).
Male Sprague–Dawley rats (n�17) were anesthetized (urethane,1.2 g/kg, 10% IP; Sigma-Aldrich) and maintained at surgical depththroughout experiments. Rats were intubated and arterial (femoral orcarotid) and intravenous catheters inserted for arterial pressure (AP)measurement or drug administration. Splanchnic sympathetic activ-ity (SNA) and phrenic nerve activity (PNA) were recorded usingbipolar electrodes and in four rats tail blood flow (TBF) was alsorecorded using a laser Doppler flow probe (Oxford Optronics,Oxford, UK). In 2 rats electromyographic (EMG) activity from theleft biceps femoris was also recorded. All signals were acquiredonline using Spike 2 software (CED Ltd, Cambridge, UK) asdescribed previously.26,27 Microinjections were made into theRVLM as described previously26,27 in paralyzed (pancuroniumdibromide, 0.8 mg), nonvagotomized animals (n�13), or into thepons using coordinates of Paxinos and Watson28 in bilaterallyvagotomized animals before and after paralysis (n�4).
Cardiovascular reflexes were evoked as described previously26,27
using sequential injection of sodium nitroprusside (SNP) and phen-ylephrine (PE) (10 �g/kg), or intermittent electrical stimulation (0.5Hz, 100 sweeps; twin pulses at 2.5-ms intervals; pulse width, 1 ms)of a somatic tibial nerve (TN, 15 to 20 V) or the barosensory aorticnerve (5 to 10 V). Carotid chemoreceptors were activated by briefhypoxia (100% N2 for 15 seconds).
Intravenous drugs were dissolved in saline: atropine methylnitrate(mATR) (peripheral mAChR blocker; t1/2, 4 hours; 5 mg/mL;Sigma); oxotremorine sesquifumarate salt (OXO) (broad spectrummAChR agonist; t1/2, 1.6 hours; 0.5 mg/mL; Sigma); (-)scopolaminehydrobromide (SCOP) (broad spectrum mAChR antagonist; t1/2, 8hours; 5 mg/mL); SNP (Faulding); and PE (ICN Biomedicals Inc).Drugs for microinjection were dissolved in 10 mmol/L PBS:L-glutamic acid (monosodium salt; 100 mmol/L [5 nmol/50 nL];Sigma); (-)SCOP (60 mmol/L [3 nmol/50 nL]; Sigma); DL-homocysteic acid (DLH) (an excitatory amino acid; 100 mmol/L;MP Biomedicals); and (-)-bicuculline methiodide (a selectiveGABA-A receptor antagonist; 4 mmol/L; Sigma).
As described previously,29,30 injections of cholera toxin B subunit(CTB) (1%, retrograde tracer) were made centered in the interme-diolateral cell column (n�10) or RVLM (n�3) under anesthesia andanimals were recovered for 36 to 72 hours. Rats were transcardiallyperfused and brains processed for light and fluorescence immuno-histochemistry30,31 or combined in situ hybridization and immuno-histochemistry as described previously.32 Briefly, 50-�m sectionswere incubated with species-specific primary antibodies to detectCTB (goat; List OR rabbit; Virostat), vesicular acetylcholine trans-porter (vAChT) (Chemicon), tyrosine hydroxylase (TH) (Sigma,Australia), or neuron-specific nuclear protein (NeuN) (Chemicon).For detection of mRNA sections were hybridized with digoxigenin
(DIG)-labeled antisense riboprobe specific to preproenkephalin(PPE), M2 or M3 receptor (see the online data supplement) followedby incubation with DIG primary antibodies. For detection of proteinsections were incubated with biotinylated- or fluorophore-conjugated secondary antisera (1:500; Jackson ImmunoResearchLaboratories, Inc). For light microscopy, vAChT and CTB weredetected using enhanced diaminobenzidine reactions (nickel andimidazole).30
ProtocolsAnimals were pretreated with mATR (2 mg/kg), and OXO wasinjected intravenously (0.2 mg/kg); SCOP was then injected bilater-ally into the RVLM (9 nmol per side). Reflexes were tested beforeand after OXO in the absence or presence of SCOP. Injections ofDLH or bicuculline were made into the PPT and effects of SNPinjection and TN stimulation were tested before and after SCOPintravenous injection (2 mg/kg).
Close appositions between vAChT immunoreactivity (vAChT-IR)and CTB/TH-IR or non–TH-IR neurons in the RVLM were exam-ined at �100 magnification; serial cell counts of vAChT-IR appo-sitions or M2 receptor/CTB/TH were made from every fourthsection. For detailed data and statistical analysis, see the online datasupplement.
ResultsRVLM mAChR Mediate Sympathoexcitatory butNot Other Autonomic Effects Evoked by CentralmAChR ActivationCentral mAChR activation (OXO) significantly increasedAP, mean and postinspiratory-related discharge of SNA,heart rate (HR) and TBF and reduced PNA amplitude (Figure1A, 1B, and 1D). Bilateral injection of SCOP into the RVLM(Figure 1C) significantly attenuated the increase in AP (n�8,P�0.01), SNA (n�8, P�0.01), HR (n�8, P�0.05) andpost-inspiratory activation of SNA (n�4, P�0.05) but had noeffect on changes in TBF (n�4; P�NS) or PNA amplitude(n�4; P�NS) evoked by OXO (Figure 1D).
Spectral analysis of systolic AP (SAP) and SNA revealedan increase in low frequency (LF, �0.4 Hz) oscillationsfollowing OXO (0.3�0.1 versus 36.5�15.1 mm Hg2,P�0.05; 4.6�1.4 versus 47.5�17.9 SNA%2, P�0.05).Respiratory-related oscillations of SNA also tended to beincreased (P�0.051). SCOP injected bilaterally into theRVLM had no effect on baseline parameters but preventedthe increase in LF oscillations evoked by OXO (0.3�0.1versus 0.4�0.1 mm Hg2; 3.3�1.0 versus 7.3�2.5 SNA%,2
n�7, P�0.05) (Figure II in the online data supplement).
Activation of RVLM mAChR Facilitates theSympathetic Baroreflex and Inhibits theSomatosympathetic and ChemoreflexesOXO significantly enhanced the reflex sympathoexcitatoryand inhibitory responses evoked by equipotent doses of SNPand PE (Figure 2). This effect was reproducible followingrepeat injection of OXO (Figure 2A). OXO significantlyincreased the maximum plateau (146�4 versus 321�12%,P�0.01) and maximum gain of the SNA baroreflex (4.4�0.5versus 8.2�0.6%/mm Hg, P�0.05) (Figure 2B and 2C). Theoperating point (resting MAP) also shifted closer to the pointof maximum gain (Figure 2C).
Figure 3 shows the effects of OXO on cardiovascularreflexes before and after blockade of mAChR bilaterally in
Padley et al Cholinergic Inputs to RVLM in Central Command 285

the RVLM. OXO increased the magnitude of SNA inhibitionevoked by aortic nerve stimulation (166�13% control, n�6,P�0.01) or excitation following SNP administration (4�1versus 17�2% SNA/50 mmHg, n�4, P�0.01). In contrast,OXO inhibited both excitatory peaks of SNA evoked by TNstimulation (early peak 37�3% control, P�0.01, late peak41�5% control, n�9, P�0.01). Sympathoexcitatory andpressor responses to brief hypoxia were attenuated andinhibited, respectively (53�6% control, P�0.01;�33�2 mm Hg versus �17�5 mm Hg, n�7, P�0.01).Bilateral injection of SCOP into the RVLM reversed effectsof OXO on reflexes, such that they were mostly indistinguish-able from controls. The early peak of the somatosympatheticreflex was only partially restored (66�4% control, P�0.01).A repeat injection of OXO 30 minutes following SCOP failed
to elicit effects on any reflex similar to its initial robust effects(n�4 per group). The pressor response to brief hypoxia didnot return to normal after the initial dose of OXO. Groupeddata are illustrated in supplemental Figure I.
Cholinergic Terminals Closely ApposeSympathoexcitatory RVLM NeuronsvAChT-IR terminals were found throughout the VLM and incell bodies within the facial and ambigual motor nuclei,consistent with previous reports.33,34 A choline acetyltrans-ferase–positive cell group previously identified in the ventro-medial medulla22 was not present using vAChT labeling(supplemental Figure IV). vAChT-IR terminals were closelyapposed to CTB-labeled spinally projecting cells in theRVLM; 32.6�7.4% (379/1118 cells, n�3) of all CTB-IRneurons and 31.1�5.6% (66/206 cells, n�3) of TH-positiveCTB-IR cells (Figure 4A and 4B). vAChT-IR varicositiesalso formed perisomatic appositions with NeuN-positivenon-TH RVLM neurons that expressed PPE, as well as otherPPE-negative NeuN-positive cells (Figure 4C).
M2 and M3 Receptor mRNA Is Expressed inSpinally Projecting Non-TH Neurons inthe RVLMAll mAChR subtypes were expressed in an RVLM tissuepunch (supplemental Figure III). We analyzed the cellulardistribution of M2 receptor expression in the RVLM andfound that no spinally projecting TH neurons contained M2receptor mRNA (0/310, n�5) (Figure 5A). In contrast,23�4% of spinally projecting non-TH RVLM neurons didexpress M2 receptor mRNA (78/367, n�5) (Figure 5C). M3receptor mRNA was also expressed in some TH-IR/non-CTB-IR and some CTB-IR/non-TH neurons (Figure 5B and5D).
Direct Cholinergic Projections to the RVLM Fromthe PPTTo determine the source of cholinergic input to the RVLM,discrete injections of CTB were made unilaterally into theRVLM (Figure 6A) and cholinergic neurons were identifiedby vAChT-IR. CTB-IR neurons were found in regionspreviously described, including the parabrachial nucleus, theparaventricular nucleus of the hypothalamus (Figure 6B),central nucleus of the amygdala, and the cortex.21,35
vAChT-IR neurons were also found in regions previouslydescribed33,34 (Figure 6C and 6D). Neurons that were double-labeled for CTB and vAChT had a restricted distribution andwere confined within the PPT (Figure 6C and 6D).
Chemical Stimulation of the PPT Increases MuscleActivity and SNA and Facilitates the SympatheticBaroreflex via mAChR ActivationBilateral injection of bicuculline into the PPT evoked in-creases in AP and EMG activity (Figure 7A). EMG activitybut not the increase in AP was abolished by subsequentneuromuscular (NM) blockade (Figure 7A). Injection of DLHinto the PPT produced an increase in SNA and AP andincreased the magnitude of SNA excitation produced byinjection of SNP (Figure 7B). These effects could be evoked
Figure 1. The role of RVLM mAChR in mediating autonomiceffects of central mAChR activation in urethane-anesthetizedrats. A, Following pretreatment with mATR, OXO (0.2 mg/kg IV)evokes an increase in SNA, AP, and TBF but a reduction in PNAamplitude. Following identification of RVLM pressor sites(L-glutamic acid [glut]), SCOP (9 nmol per site) is injected intothe RVLM bilaterally substantially reducing the pressor, sympa-thetic, and HR effects but not the TBF or PNA response toOXO. Increases in HR are caused by sympathetic activation. B,Average SNA (bold line) and PNA (thin line) waveforms showingincrease in postinspiratory (P-I)-related discharge of SNA follow-ing OXO and blockade of this effect by SCOP injection in theRVLM. C, Injection sites in the RVLM (open circles) (only unilat-eral sites shown). D, Group data from 8 animals illustratingeffects seen in A and B. Data shown are mean�SEM; bpu indi-cates blood perfusion units; au, arbitrary units.
286 Circulation Research February 2, 2007

throughout the rostrocaudal extent of the PPT (�7 to 9 mmcaudal to Bregma). Smaller increases in SNA were evoked atsites dorsal or ventral but facilitation of baroreflex-evokedSNA responses was restricted to the PPT (6.5 to 7.5 mmventral) (Figure 7B). Transient alterations in PNA frequencywere observed following stimulation of the PPT and sur-rounding brain areas, whereas the somatosympathetic reflexwas unaffected. Prior blockade of central mAChR receptorswith SCOP intravenous injection prevented the facilitation ofbaroreflex-evoked SNA responses (n�3, P�0.05) but did notabolish sympathoexcitation produced by DLH injection intothe PPT (Figure 7C).
DiscussionThe novel findings of this study are (1) mAChR activationin the RVLM facilitates the sympathetic baroreflex andattenuates and inhibits the sympathetic chemoreflex andsomatosympathetic reflex, respectively; (2) identifiedsympathoexcitatory neurons in the RVLM receive cholin-ergic input and differentially express M2 and M3 receptorsubtypes; (3) chemical stimulation of the PPT, whichprovides the only direct cholinergic input to the RVLM,evokes a similar pattern of tonic and baroreflex SNAresponses to that seen following RVLM mAChR activa-tion. Neurons in the PPT control muscle tone during
Figure 2. Effects of central mAChR acti-vation on sympathetic baroreflex func-tion. A, OXO (0.2 mg/kg IV) evokes sig-nificant facilitation of the sympatheticreflex effects evoked by baroreceptorunloading (SNP) and loading (PE). Thiseffect is reproducible. B, Average 4-pa-rameter sympathetic baroreflex functioncurves generated from data (n�3) includ-ing that shown in A and C: their firstderivatives (error bars are omitted forclarity). Central mAChR activation withOXO shifts the SNA baroreflex to higherAP and SNA and increases its range andgain.
Figure 3. The role of RVLM mAChR inmediating effects on cardiorespiratoryreflex function following central mAChRactivation. A continuous recording ofMAP and SNA (bottom two rows) illus-trating differential effects of OXO (0.2mg/kg IV) on the baroreflex (aortic nervestimulation, top row), the somatosympa-thetic reflex (TN stimulation, secondrow), and the peripheral chemoreflex (N2
substitution, third and fourth rows). Thestimulus periods are indicated under theperistimulus-averaged SNA responses.OXO enhances SNA baroreflexresponses, inhibits both early and latepeaks of the somatosympathetic reflex,and attenuates sympathoexcitatory andpressor effects of the peripheral che-moreflex. These effects are blocked byprior injection of SCOP bilaterally in theRVLM. Mean changes in SNA are over-laid on the raw SNA signal to illustrateeffect of OXO on reflex responses tobaroreceptor unloading with SNP (*).Dotted horizontal lines indicate the con-trol level of SNA.
Padley et al Cholinergic Inputs to RVLM in Central Command 287

locomotion, exercise, and arousal.4,24,25 Our findings indi-cate that tonic and reflex cardiovascular adjustments arealso evoked from the PPT via direct cholinergic proj-ections to the RVLM. These data support our hypothesis
that cholinergic input to the RVLM is involved in centralcommand.
RVLM mAChRs mediate the increase in SNA and HRevoked by centrally acting OXO, in agreement with previousstudies.10,11 Postinspiratory-related discharge of SNA wasalso enhanced, indicating a direct effect on respiratory-relatedinputs to the RVLM36 that presumably contributes to themean increase in SNA evoked by OXO. This effect waselicited independently of the OXO-evoked depression ofphrenic amplitude, which is mediated by sites other than theRVLM.13,14 The OXO-evoked increase in TBF, presumablycontributing to the well-described hypothermic effect of thisdrug,12 is also mediated via other central sites. These mayinclude the preoptic area of the hypothalamus as it receivescholinergic input and carbachol microinjections here evokehypothermia.37
We showed, for the first time, that the reflex responses ofSNA to baroreceptor loading or unloading, demonstratedfollowing vasoactive drug administration and by direct stim-ulation of baroreceptor afferents, were markedly enhanced byOXO and were mediated by RVLM mAChRs. Furthermore,baroreflex-related LF oscillations of both SAP and SNA38
were enhanced.Our findings show that RVLM mAChR activation resets
the SNA baroreflex to higher pressures and increases itsrange and gain. Earlier studies showed that the pressor effectof bilateral carotid occlusion was greater after central orsystemic administration of physostigmine.8,17 Caputi et aldemonstrated an upward shift in baroreflex HR responseswithout changes in range or gain following intracerebroven-tricular injection of physostigmine.16 A limitation of thepresent study was that pretreatment with mATR to blockperipheral mAChR precluded analysis of vagally mediatedHR responses. Our findings indicate that RVLM mAChRactivation facilitates sympathetic vasomotor responses tobaroreflex activation, whereas cholinergic effects at brainsites important in reflex vagal control, including the nucleusambiguus,39 may evoke resetting of the HR baroreflex with-out changing its gain.
OXO evoked differential effects on cardiovascular reflexesvia RVLM mAChR activation. Baroreflex SNA responses
Figure 4. Neurons in the RVLM are closely apposed byvAChT-IR varicosities. A, RVLM neurons retrogradely labeledfrom the thoracic intermediolateral cell column (CTB-IR,brown reaction product) are surrounded by vAChT-IR varicos-ities (black reaction product) that make close appositionswith their cell bodies (arrowheads). Note the neurons in themore dorsal nucleus ambiguus that are immunoreactive forvAChT. B, CTB-IR cells or CTB-IR/TH-positive cells areclosely apposed by vAChT-IR (arrowhead or *, respectively).C, Neurons in the RVLM identified by the neuron-specificmarker NeuN that express PPE mRNA (arrows) are closelyapposed by vAChT-IR, as are PPE-negative neurons. Scalebars: 400 �m (A, left); 50 �m (A, right); 25 �m (B and C). pyrindicates pyramidal tract. IOL indicates inferior olivarynucleus; NAc, nucleus ambiguus pars compacta.
Figure 5. Cellular distribution of M2 (A and C) orM3 receptor mRNA (B and D) and TH-IR neuronsin the RVLM. M2 receptor mRNA is colocalized inRVLM neurons that project to the thoracic interme-diolateral cell column (CTB-IR) (C), but none ofthese contain TH-IR. M3 receptor mRNA isexpressed in some CTB-IR neurons (arrows) or inTH-IR cells lacking CTB-IR (arrowheads) (boxedarea in B shown in D). Scale bars: 250 �m (A andB); 100 �m (C and D).
288 Circulation Research February 2, 2007

mediated by direct inhibition or disinhibition of RVLMneurons40 were enhanced. Somatosympathetic and chemore-flex SNA responses mediated by direct excitation of RVLMneurons36 were inhibited and attenuated, respectively. Theclear inhibition of the somatosympathetic reflex suggests thatthese effects were not indirectly caused by raised sympatheticactivity. To our knowledge, a study in anesthetized cats alsoshowed that a somatosympathetic reflex evoked by intercos-tal nerve stimulation was inhibited by OXO.41 As singleRVLM neurons receive largely convergent input from barore-ceptors, peripheral chemoreceptors, somatic afferents, andcentral respiratory neurons,42–45 3 mechanisms are possible toexplain our data: OXO activates inhibitory presynapticmAChRs located on reflex inputs to RVLM neurons, postsyn-aptic excitatory mAChRs on RVLM neurons, or a combina-tion of both. Pre- and postsynaptic effects of carbachol onRVLM neurons have been demonstrated in vitro.18
Phenotypically identified sympathoexcitatory (CTB�TH)and putative sympathoexcitatory (CTB�non-TH, or PPE�)neurons in the RVLM were closely apposed by vAChT-IRvaricosities. This is the first anatomical evidence indicatingthat cholinergic terminals may synapse with sympathoexcit-atory RVLM neurons. Milner et al23 showed that cholineacetyltransferase IR terminals formed abundant synaptic con-tacts in the ventral medulla but these were rarely seen withTH-containing neurons. In the study by Milner et al, however,only caudal sections of the RVLM were examined (0.5 to 2.0mm caudal to the facial nucleus); these contain few spinallyprojecting neurons.29 Furthermore, compared with cholineacetyltransferase, immunoreactivity to vAChT as used heregives better cholinergic terminal labeling.33,34
Our results showed, for the first time, that the M2 receptorwas not expressed in TH neurons but was expressed in asubpopulation of spinally projecting non-TH neurons. M2receptor-preferring antagonists prevent pressor effects ofRVLM mAChR activation.11 The ligands used, however, donot display high affinity for any 1 particular subtype.46 If M2receptors do mediate OXO-evoked sympathoexcitatory re-
sponses, then they are most likely located presynaptically inthe RVLM or this effect is mediated by non-TH spinallyprojecting neurons. Huangfu et al have shown in neonatalRVLM that both C1 and non-C1 cells depolarized in responseto mAChR activation.18 Because vAChT-IR terminals ap-posed both classes of RVLM neurons, we sought evidence forexpression of other receptor subtypes. A subpopulation ofspinally projecting non-TH RVLM neurons also contain M3receptor mRNA. We have further demonstrated that mRNAfor all 5 mAChR subtypes was present in the RVLM,confirming earlier studies in WKY and SHR rats.47 Ourresults suggest that different or multiple mAChR subtypesmay be expressed by sympathoexcitatory RVLM neurons.
In agreement with Yasui et al,21 we found that theprojection from the PPT to the RVLM is cholinergic. Inaddition, we showed that the PPT is the only cholinergic cellgroup that provides input to the RVLM. Local inputs fromcholine acetyltransferase–positive neurons in the ventrome-dial medulla22 are not functionally cholinergic, as we foundthat these cells did not contain vAChT.
We demonstrated for the first time that chemical stimula-tion of the PPT facilitates baroreflex-evoked excitation ofSNA, mimicking effects of RVLM mAChR activation.Blockade of mAChR with SCOP prevented this effect but didnot completely abolish sympathoexcitation generated by PPTactivation. Electrical stimulation of the PPT increases AP,HR, and renal SNA (with a lesser increase in lumbar SNA) indecerebrate animals.48,49 Sympathoexcitatory responses arealso evoked from surrounding brain areas including thecuneiform nucleus.50 At present, we cannot explain the lackof effect of stimulating the PPT on other reflex responses thatare modified by activation of RVLM mAChR.
Disinhibition of the PPT increased EMG activity, consis-tent with studies that reported increases in muscle activityfollowing electrical or chemical stimulation of the PPT inanesthetized or decerebrate animals.24,48,49 Single cholinergicneurons in the PPT have dual connections with the motorcortex and stellate ganglion, as revealed by polysynaptic viral
Figure 6. Distribution of retrogradely labeled neu-rons in the hypothalamus (B) and pons (C and D)following injection of CTB into the pressor regionof the RVLM (A) (CTB green). Neurons double-labeled (arrows) for CTB and vAChT (red) werefound only within rostral and caudal parts of thePPT. C and D, Schematics adapted from Paxinosand Watson.28 Scale bars: 500 �m (A and B);100 �m (C); 200 �m (D). 3v indicates third ventri-cle; pyr, pyramidal tract; VII, facial nucleus.
Padley et al Cholinergic Inputs to RVLM in Central Command 289

tracing.5 The PPT may therefore be a key nodal point wherechanges in motor signals can be coordinated with descendingmodulation of sympathetic function. The simplest explanation ofour data is that stimulation of the PPT evokes muscular activityand releases acetylcholine activating RVLM mAChR pre-and/or postsynaptically located on sympathoexcitatory neurons,causing an increase in AP and SNA as well as increasing therange and gain of the sympathetic baroreflex.
Functional ImplicationsThe involvement of the PPT in initiating and modulatingmovement related to arousal and locomotion, including ex-ercise, is well recognized.4,24,25 The present findings indicatethat the cholinergic projection to the RVLM may be activatedin parallel to elicit tonic and reflex cardiovascular adjust-
ments that are appropriate to different behaviors. The patternof effects bears a striking similarity to those evoked by centralcommand during exercise.2,3,51
Exercise is accompanied by a resetting of baroreflexcontrol of SNA and HR to higher AP.2,51–53 This is thought tobe crucial to AP elevation at exercise onset and AP stabili-zation during exercise and can oppose other reflex influenceson circulation, including nociceptive and peripheral chemore-flexes.3 In addition to an increase in AP and SNA, theincrease in the range and gain of the sympathetic baroreflexas seen here strongly resembles that evoked during treadmillexercise in conscious rats.52 Studies showing complete sym-pathetic baroreflex function curves during exercise in humansare sparse, although some studies have demonstrated largeincreases in linear baroreflex gain of muscle SNA duringstatic exercise53 or no change during moderate intensity armcycling.54 In contrast, exercise appears to reset the cardiaccomponent of the baroreflex to higher AP without changingits gain,51 also resembling effects on the HR baroreflexevoked by central administration of physostigmine.16 Recentevidence indicates that the cardiac baroreflex is transientlyinhibited at exercise onset, which may facilitate immediatevagal withdrawal.55
In conclusion, our data indicate that the cholinergic projectionfrom the PPT to the RVLM is an integral component of thecentral command pathway that regulates circulatory functionduring exercise and possibly other arousal or behavioral states.
AcknowledgmentsWe thank Simon McMullan and Peter Burke for helpful discussions.
Sources of FundingSupported by the National Health and Medical Research Council ofAustralia (211023, 211196), Garnett Passe and Rodney WilliamsMemorial Foundation, North Shore Heart Research Foundation (6-05/06), and Northern Sydney Central Coast Area Health (2006:03). J.R.P.and N.N.K. receive Australian Postgraduate awards.
DisclosuresNone.
References1. White SW, Pitsillides KF, Parsons GH, Hayes SG, Gunther RA, Cottee
DB. Coronary-bronchial blood flow and airway dimensions in exercise-induced syndromes. Clin Exp Pharmacol Physiol. 2001;28:472–478.
2. Williamson JW, Fadel PJ, Mitchell JH. New insights into central cardio-vascular control during exercise in humans: a central command update.Exp Physiol. 2006;91:51–58.
3. Rowell LB, O’Leary DS. Reflex control of the circulation duringexercise: chemoreflexes and mechanoreflexes. J Appl Physiol. 1990;69:407–418.
4. Garcia-Rill E, Homma Y, Skinner RD. Arousal mechanisms related toposture and locomotion. 1. Descending modulation. Prog Brain Res.2004;143:283–290.
5. Krout KE, Mettenleiter TC, Loewy AD. Single CNS neurons link bothcentral motor and cardiosympathetic systems: a double-virus tracingstudy. Neuroscience. 2003;118:853–866.
6. Dampney RA, Horiuchi J, Killinger S, Sheriff MJ, Tan PS, McDowallLM. Long-term regulation of arterial blood pressure by hypothalamicnuclei: some critical questions. Clin Exp Pharmacol Physiol. 2005;32:419–425.
7. Buccafusco JJ, Brezenoff HE. Pharmacological study of a cholinergicmechanism within the rat posterior hypothalamic nucleus which mediatesa hypertensive response. Brain Res. 1979;165:295–310.
Figure 7. Coronal sections of the pons showing tracks andinjection sites of 4 mmol/L bicuculline (bic) (A) and 100 mmol/LDLH (B) (arrows) and cardiovascular responses evoked from thePPT. A, Disinhibition (bicuculline) of the PPT increases AP andEMG activity. B, Unilateral injection of DLH into the PPT (2), butnot more dorsally (1) or ventrally, (3) increases SNA and AP andenhances the reflex increase in SNA following SNP injection. C,Pretreatment with SCOP (2 mg/kg IV) blocks the facilitation ofthe baroreflex evoked by PPT stimulation (lowercase letters indi-cate reference levels of SNA: a, baseline; b, new level reachedfollowing DLH injection into the PPT; c, peak level reached fol-lowing SNP injection before SCOP). Data are mean�SEM.Schematics adapted from Paxinos and Watson.28 Cu indicatescuneiform nucleus; PAG, periaqueductal gray; scp, superior cer-ebellar peduncle.
290 Circulation Research February 2, 2007

8. Brezenoff HE, Carney K, Buccafusco JJ. Potentiation of the carotid arteryocclusion reflex by a cholinergic system in the posterior hypothalamicnucleus. Life Sci. 1982;30:391–400.
9. Brezenoff HE, Giuliano R. Cardiovascular control by cholinergic mech-anisms in the central nervous system. Annu Rev Pharmacol Toxicol.1982;22:341–381.
10. Kubo T. Cholinergic mechanism and blood pressure regulation in thecentral nervous system. Brain Res Bull. 1998;46:475–481.
11. Giuliano R, Ruggiero DA, Morrison S, Ernsberger P, Reis DJ. Cho-linergic regulation of arterial pressure by the C1 area of the rostralventrolateral medulla. J Neurosci. 1989;9:923–942.
12. Daws LC, Overstreet DH. Ontogeny of muscarinic cholinergic supersen-sitivity in the Flinders Sensitive Line rat. Pharmacol Biochem Behav.1999;62:367–380.
13. Nattie EE, Li AH. Ventral medulla sites of muscarinic receptor subtypesinvolved in cardiorespiratory control. J Appl Physiol. 1990;69:33–41.
14. Shao XM, Feldman JL. Acetylcholine modulates respiratory pattern:effects mediated by M3-like receptors in preBotzinger complexinspiratory neurons. J Neurophysiol. 2000;83:1243–1252.
15. Criscione L, Reis DJ, Talman WT. Cholinergic mechanisms in thenucleus tractus solitarii and cardiovascular regulation in the rat. EurJ Pharmacol. 1983;88:47–55.
16. Caputi AP, Rossi F, Carney K, Brezenoff HE. Modulatory effect of brainacetylcholine on reflex-induced bradycardia and tachycardia in consciousrats. J Pharmacol Exp Ther. 1980;215:309–316.
17. Park KH, Long JP. Modulation by physostigmine of head-up tilt- andbilateral carotid occlusion-induced baroreflexes in rats. J Pharmacol ExpTher. 1991;257:50–55.
18. Huangfu D, Schreihofer M, Guyenet PG. Effect of cholinergic agonists onbulbospinal C1 neurons in rats. Am J Physiol. 1997;272:R249–R258.
19. Pilowsky PM, Goodchild AK. Baroreceptor reflex pathways and neuro-transmitters: 10 years on. J Hypertens. 2002;20:1675–1688.
20. Guyenet PG. The sympathetic control of blood pressure. Nat RevNeurosci. 2006;7:335–346.
21. Yasui Y, Cechetto DF, Saper CB. Evidence for a cholinergic projectionfrom the pedunculopontine tegmental nucleus to the rostral ventrolateralmedulla in the rat. Brain Res 28. 1990;517:19–24.
22. Ruggiero DA, Giuliano R, Anwar M, Stornetta R, Reis DJ. Anatomicalsubstrates of cholinergic-autonomic regulation in the rat. J Comp Neurol.1990;292:1–53.
23. Milner TA, Pickel VM, Giuliano R, Reis DJ. Ultrastructural localizationof choline acetyltransferase in the rat rostral ventrolateral medulla:evidence for major synaptic relations with noncatecholaminergic neurons.Brain Res. 1989;500:67–89.
24. Pahapill PA, Lozano AM. The pedunculopontine nucleus and Parkinson’sdisease. Brain. 2000;123:1767–1783.
25. Bedford TG, Loi PK, Crandall CC. A model of dynamic exercise: thedecerebrate rat locomotor preparation. J Appl Physiol. 1992;72:121–127.
26. Miyawaki T, Goodchild AK, Pilowsky PM. Activation of mu-opioidreceptors in rat ventrolateral medulla selectively blocks baroreceptorreflexes while activation of delta opioid receptors blocks somato-sympathetic reflexes. Neuroscience. 2002;109:133–144.
27. Makeham JM, Goodchild AK, Pilowsky PM. NK1 receptor activation inrat rostral ventrolateral medulla selectively attenuates somato-sympathetic reflex while antagonism attenuates sympathetic chemoreflex.Am J Physiol Regul Integr Comp Physiol. 2005;288:R1707–R1715.
28. Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. 3rd ed.San Diego, Calif: Academic Press, Inc; 1996.
29. Phillips JK, Goodchild AK, Dubey R, Sesiashvili E, Takeda M, ChalmersJ, Pilowsky PM, Lipski J. Differential expression of catecholamine bio-synthetic enzymes in the rat ventrolateral medulla. J Comp Neurol.2001;432:20–34.
30. Goodchild AK, Llewellyn-Smith IJ, Sun QJ, Chalmers J, CunninghamAM, Pilowsky PM. Calbindin-immunoreactive neurons in the reticularformation of the rat brainstem: catecholamine content and spinal proj-ections. J Comp Neurol. 2000;424:547–562.
31. Springell DA, Powers-Martin K, Phillips JK, Pilowsky PM, GoodchildAK. Phosphorylated extracellular signal-regulated kinase 1/2 immunore-activity identifies a novel subpopulation of sympathetic preganglionicneurons. Neuroscience. 2005;133:583–590.
32. Li Q, Goodchild AK, Seyedabadi M, Pilowsky PM. Preprotachykinin AmRNA is colocalized with tyrosine hydroxylase-immunoreactivity inbulbospinal neurons. Neuroscience. 2005;136:205–216.
33. Arvidsson U, Riedl M, Elde R, Meister B. Vesicular acetylcholine trans-porter (VAChT) protein: a novel and unique marker for cholinergic
neurons in the central and peripheral nervous systems. J Comp Neurol.1997;378:454–467.
34. Schafer MK, Eiden LE, Weihe E. Cholinergic neurons and terminal fieldsrevealed by immunohistochemistry for the vesicular acetylcholine trans-porter. I. Central nervous system. Neuroscience. 1998;84:331–359.
35. Horiuchi J, Potts PD, Polson JW, Dampney RA. Distribution of neuronsprojecting to the rostral ventrolateral medullary pressor region that areactivated by sustained hypotension. Neuroscience. 1999;89:1319–1329.
36. Miyawaki T, Minson J, Arnolda L, Chalmers J, Llewellyn-Smith I,Pilowsky P. Role of excitatory amino acid receptors in cardiorespiratorycoupling in ventrolateral medulla. Am J Physiol. 1996;271:R1221–R1230.
37. Tanaka M, Nagashima K, McAllen RM, Kanosue K. Role of the med-ullary raphe in thermoregulatory vasomotor control in rats. J Physiol.2002;540:657–664.
38. Ringwood JV, Malpas SC. Slow oscillations in blood pressure via anonlinear feedback model. Am J Physiol Regul Integr Comp Physiol.2001;280:R1105–R1115.
39. Wang J, Irnaten M, Neff RA, Venkatesan P, Evans C, Loewy AD,Mettenleiter TC, Mendelowitz D. Synaptic and neurotransmitter acti-vation of cardiac vagal neurons in the nucleus ambiguus. Ann N Y AcadSci. 2001;940:237–246.
40. Lipski J, Kanjhan R, Kruszewska B, Rong W. Properties of presympa-thetic neurones in the rostral ventrolateral medulla in the rat: an intra-cellular study “in vivo’. J Physiol. 1996;490:729–744.
41. Baum T, Shropshire AT. Influence of a cholinergic agent, oxotremorine,on sympathetic reflexes. Eur J Pharmacol. 1978;52:243–249.
42. Guyenet PG, Darnall RA, Riley TA. Rostral ventrolateral medulla andsympathorespiratory integration in rats. Am J Physiol. 1990;259:R1063–R1074.
43. Miyawaki T, Pilowsky P, Sun QJ, Minson J, Suzuki S, Arnolda L,Llewellyn-Smith I, Chalmers J. Central inspiration increases barosensi-tivity of neurons in rat rostral ventrolateral medulla. Am J Physiol.1995;268:R909–R918.
44. Pilowsky P, Arnolda L, Chalmers J, Llewellyn-Smith I, Minson J,Miyawaki T, Sun QJ. Respiratory inputs to central cardiovascularneurons. Ann N Y Acad Sci. 1996;783:64–70.
45. Verberne AJ, Stornetta RL, Guyenet PG. Properties of C1 and otherventrolateral medullary neurones with hypothalamic projections in the rat.J Physiol. 1999;517:477–494.
46. Caulfield MP, Birdsall NJ. International Union of Pharmacology. XVII.Classification of muscarinic acetylcholine receptors. Pharmacol Rev.1998;50:279–290.
47. Gattu M, Wei J, Pauly JR, Urbanawiz S, Buccafusco JJ. Increasedexpression of M2 muscarinic receptor mRNA and binding sites in therostral ventrolateral medulla of spontaneously hypertensive rats. BrainRes. 1997;756:125–132.
48. Chong RK, Bedford TG. Heart rate, blood pressure, and running speedresponses to mesencephalic locomotor region stimulation in anesthetizedrats. Pflugers Arch. 1997;434:280–284.
49. Koba S, Yoshida T, Hayashi N. Differential sympathetic outflow andvasoconstriction responses at kidney and skeletal muscles during fictivelocomotion. Am J Physiol Heart Circ Physiol. 2006;290:H861–H868.
50. Verberne AJ. Cuneiform nucleus stimulation produces activation of med-ullary sympathoexcitatory neurons in rats. Am J Physiol. 1995;268:R752–R758.
51. Raven PB, Fadel PJ, Smith SA. The influence of central command onbaroreflex resetting during exercise. Exerc Sport Sci Rev. 2002;30:39–44.
52. Miki K, Yoshimoto M, Tanimizu M. Acute shifts of baroreflex control ofrenal sympathetic nerve activity induced by treadmill exercise in rats.J Physiol 1. 2003;548:313–322.
53. Kamiya A, Michikami D, Fu Q, Niimi Y, Iwase S, Mano T, Suzumura A.Static handgrip exercise modifies arterial baroreflex control of vascularsympathetic outflow in humans. Am J Physiol Regul Integr CompPhysiol. 2001;281:R1134–R1139.
54. Fadel PJ, Ogoh S, Watenpaugh DE, Wasmund W, Olivencia-Yurvati A,Smith ML, Raven PB. Carotid baroreflex regulation of sympathetic nerveactivity during dynamic exercise in humans. Am J Physiol Heart CircPhysiol. 2001;280:H1383–H1390.
55. Matsukawa K, Komine H, Nakamoto T, Murata J. Central commandblunts sensitivity of arterial baroreceptor-heart rate reflex at onset ofvoluntary static exercise. Am J Physiol Heart Circ Physiol. 2006;290:H200–H208.
Padley et al Cholinergic Inputs to RVLM in Central Command 291

Supplemental Information
Central command regulation of circulatory function mediated by descending
pontine cholinergic inputs to sympathoexcitatory RVLM neurons
James R Padley, Natasha N Kumar, Qun Li, Thomas BV Nguyen, Paul M Pilowsky,
Ann K Goodchild
Expanded Materials and Methods
Animals
Studies were approved by the Animal Care and Ethics Committee of Royal North
Shore Hospital / University of Technology Sydney. Following experimentation rats
were killed whilst under deep surgical anaesthesia by injection of potassium chloride
(KCl 3M, 1mL iv).
Anaesthesia, surgical procedures and recording
Male Sprague Dawley (SD) rats (n=17) were anaesthetised with urethane (ethyl
carbamate, Sigma-Aldrich, 1.2 g/kg, 10% ip). The withdrawal response to hindpaw
pinch, testing of corneal reflexes and pattern of phrenic nerve discharge were used to
monitor the depth of anaesthesia and level of neuromuscular blockade. Additional
anaesthetic (urethane, 30mg) was administered when required. Rats were intubated
and arterial (femoral or carotid) and intravenous catheters inserted for blood pressure
(AP) measurement or drug administration. The left splanchnic sympathetic and
phrenic nerves were isolated for recording 1 and in four rats tail blood flow (TBF) was
recorded via a laser Doppler flow probe (λ = 780 ± 10 nm, power = 0.5 – 1.0 mW,

Oxford Optronics, UK). All signals were amplified and recorded differentially
(Bioamplifier, CWE Inc., USA) and acquired at 1000 Hz online (Spike 2, Cambridge
Electronic Design, CED Ltd., UK) 1-3.
Microinjections into the RVLM were made following exposure of the dorsal surface
and the RVLM was identified as the site where injection of glutamate evoked a sharp
transient increase in AP (>50 mmHg) and splanchnic nerve activity (SNA) (>200 %)
1. Microinjections into the pons were made by placing rats in a flat-skull position and
exposing the parietal cortex bilaterally. In these rats both vagi were cut and in two
animals EMG from the left biceps femoris was recorded. Rats were paralysed with
pancuronium dibromide (0.8mg + 0.4mg/hr, 2mg/ml) and ventilated with 100% O2
mixed with room air adjusted to maintain end-tidal CO2 between 4 – 5 % of expired
gases (Capstar-100, CWE Inc., USA).
Activation of cardiovascular reflexes
Reflexes were evoked as described previously 2-4. Briefly, changes in AP were evoked
by sequential iv injection of sodium nitroprusside (SNP) and phenylephrine (PE) (10
µg/kg). Submaximal baroreflex inhibition of SNA was evoked by intermittent
electrical stimulation of the AN (0.5 Hz x 100, twin pulses separated by 2.5 ms, pulse
width 1 ms, 5-10 V). The somatosympathetic reflex was evoked by stimulation of the
TN (0.5 Hz x 100, twin pulses separated by 2.5 ms, pulse width 1 ms, 15-20 V).
Carotid chemoreceptor activation was achieved by substitution of the inhaled gas
mixture for 100% N2 for 15s.
Drugs used

The following drugs were used for iv injection; all were dissolved in saline (0.9%
NaCl, pH 7.4): atropine methylnitrate (mATR, a peripheral mAChR blocker, Sigma,
5mg/ml), oxotremorine sesquifumarate salt (OXO, a broad spectrum mAChR agonist,
Sigma, 0.5 mg/ml), (-) scopolamine hydrobromide (SCOP, a broad spectrum mAChR
antagonist, 5 mg/ml), SNP (Faulding) and PE (ICN Biomedicals Inc.). All drugs were
administered in a volume of 0.4 ml/kg.
The following drugs were used for microinjection; all were dissolved in phosphate
buffered saline (PBS, 0.01M, pH 7.4): l-glutamic acid (glut, monosodium salt, Sigma,
100mM (5nmol/50nl)); (-) scopolamine hydrobromide, a broad spectrum mAChR
antagonist (SCOP, Sigma, 60mM (3nmol/50nl)); DL-homocysteic acid (DLH, MP
Biomedicals USA, 100mM); (-)-bicuculline methiodide (Sigma, 4 mM); injection site
markers, either colloidal gold (Sigma, 20nM) or methylene blue (4%) 5, 6.
Experimental protocol
All animals were pretreated with mATR to block peripheral mAChR (2 mg/kg). In
nine rats, all reflexes were activated before and after central mAChR activation with iv
injection of OXO (0.2 mg/kg). SCOP was injected bilaterally into the RVLM to
determine if this reversed effects of OXO on SNA, AP and reflexes. Repeat doses of
OXO were administered and reflexes were again tested. In four rats reflexes were not
tested during experiments to examine effects of OXO on SNA, AP, phrenic nerve
activity (PNA), TBF and spectral parameters before and after SCOP injection into the
RVLM. In four additional rats, DLH (8 nmol) or bicuculline (0.2 nmol) was injected
into the pons at 6.5 – 9.5 mm caudal and 2 mm lateral to Bregma at varying depths
(5.5 – 9 mm ventral) 7 to examine site-specific effects on AP, SNA and SNA
baroreflex responses. In some animals effects on the SSR were also examined. Effects

on EMG were examined in two rats prior to neuromuscular blockade and nerve
recording. In three animals sites within the PPT where stimulation evoked increases in
SNA and SNA baroreflex responses were tested following SCOP (2 mg/kg)
administered intravenously to block central mAChR.
Data Analysis
All data were analysed offline using Spike 2 software (CED Ltd, UK). As previously
described 2, 3, the magnitude of respiratory modulation of SNA was quantified by peri-
phrenic averaging and SNA responses to intermittent AN or TN stimulation were
quantified by peri-stimulus averaging. Maximal changes in SNA and AP following
brief hypoxia were expressed as a percent of baseline SNA or AP (mmHg) taken over
10 s prior to the stimulus. Baroreflex function curves describing the SNA-MAP
relationship and their first derivatives for calculation of gain were calculated as
described previously 8. Spectral parameters were quantified at low (0.25-075 Hz) and
phrenic nerve frequencies (1-2.5 Hz) by fast Fourier transformation (size 256) of
systolic AP and SNA time series uniformly resampled at 10 Hz.
Retrograde labeling from the spinal cord or the RVLM
In 10 rats anaesthetised with sodium pentobarbital (60 mg/kg ip) and administered
carprofen (2.5 mg ip), injections of the retrograde tracing agent cholera toxin β
subunit (CTB, 1 %, 200 nl) were made bilaterally into the intermediolateral cell
column (IML) at T1/T2 as described previously 9, 10. In three additional rats, AP was
recorded from the superficial femoral artery and CTB (20 nl) was injected into the
RVLM identified functionally as a site where glut evoked a sharp rise in AP (>50
mmHg). Following recovery (36 hrs RVLM group, 72-96 hrs IML group), rats were

deeply anaesthetised with pentobarbital (70 mg/kg) and transcardially perfused with
tissue culture medium (ph 7.4, Dulbeco’s Modified Eagle’s Medium, Sigma) and 4 %
formaldehyde (0.1 M in phosphate buffer (PB)) (RVLM-injected group and 7 IML-
injected rats) or 0.9% saline and freshly prepared 4 % paraformaldehyde (0.1 M in
PB) (3 IML-injected rats).
Light and fluorescence immunohistochemistry, in situ hybridisation and PCR
Following overnight fixation, serial transverse sections (50 µm) of the entire brain or
medulla and spinal cord were cut using a vibrating microtome (Leica VT1000S). For
light or fluorescence immunohistochemistry, sections were washed serially in 50%
ethanol (30 min) and Tris-phosphate-buffered saline (TPBS; Tris-HCl 0.01 M, sodium
phosphate buffer 0.01 M, 0.9% NaCl, pH 7.4) (3 x 30 min). For fluorescence
microscopy, sections were incubated with three primary antibodies (see below)
diluted in 5% normal horse serum (NHS) prior to overnight incubation with
fluorophore-conjugated secondary anti-sera diluted 1:500 in 2% NHS. For light
microscopy, sections were reacted with species-specific primary and secondary
antibodies, which were detected sequentially using enhanced diaminobenzidine
reactions (nickel and imidazole) as described previously 11. RVLM and IML injection
sites were visualised by immunohistochemistry to CTB 10.
For immunohistochemical detection of mRNA as well as protein, floating brain
sections were processed with a combined method of in situ hybridisation (ISH) and
fluorescence immunocytochemistry 12. Sections were firstly hybridized with
preproenkephalin (PPE), M2 or M3 receptor antisense riboprobe, washed in
descending concentrations of salt, then reacted with primary antibodies against

digoxigenin (DIG, alkaline phosphatase conjugated) and other proteins (see below)
for 48 hrs at 4ºC. The proteins were then detected by fluorophore-conjugated
secondary antibodies (1:500, Jackson) and DIG-labelled in situ neurons by a
histochemical reaction using nitroblue tetrazolium and 5-bromo-4-chloro-3-indolyl
phosphate salts.
Primary antibodies used were as follows. Spinally projecting neurons were visualised
using goat anti-CTB (light 1:50 000, fluorescence 1:1000, List). Cholinergic perikarya
and terminals were visualised using rabbit anti-vAChT (light 1:500, fluorescence
1:800, Chemicon, USA). Catecholaminergic neurons were visualised using mouse
anti-TH (fluorescence 1:2000, Sigma, Australia). For visualisation of enkephalinergic
neurons in the RVLM sections were reacted with primary antibodies against DIG
(alkaline phosphatase conjugated sheep anti-DIG (1:1000, Roche, Germany), mouse
anti-neuron-specific nuclear protein (NeuN, 1:2000, Chemicon, USA) and rabbit anti-
vAChT (1:800). For visualisation of M2 or M3 receptor mRNA within the RVLM
sections were reacted with primary antibodies against DIG, mouse anti-TH (1:2000)
and rabbit anti-CTB (1:5000, Virostat, USA).
Secondary antibody mixtures varied and were as follows. All secondary antisera
(Jackson Immunoresearch) were diluted 1:500 in TPBS-M containing 2% NHS.
For visualisation of spinally projecting and catecholaminergic neurons in the RVLM,
sections were incubated overnight in a mixture of Fluorescein Isothiocyanate (FITC)-
conjugated donkey anti-rabbit IgG (for vAChT), 7-Amino-4-methylcoumarin-3-acetic
acid (AMCA)-conjugated donkey anti-mouse IgG (for TH) and Texas Red (TR)-
conjugated donkey anti-goat IgG (for CTB). For visualisation of enkephalinergic

neurons and cholinergic terminals in the RVLM, sections were incubated overnight in
Cy3-conjugated donkey anti-mouse IgG (for NeuN), FITC-conjugated donkey anti
rabbit (for vAChT) and DIG-labelled in situ neurons (for PPE mRNA) were detected
as described in detail previously 12. For visualisation of M2 or M3 receptor mRNA
colocalised with catecholaminergic and spinally projecting neurons in the RVLM,
sections were incubated for 24 hrs at 4ºC with Cy3-conjugated donkey anti-mouse
IgG (for TH) and FITC-conjugated donkey anti-rabbit IgG (for vAChT). DIG-labelled
neurons (for M2 or M3 mRNA) were detected as described previously 12. For
visualisation of cholinergic neurons projecting to the RVLM, sections were incubated
with FITC-conjugated donkey anti-rabbit IgG (for vAChT) and TR-conjugated
donkey anti-goat IgG (for CTB).
Synthesis of DIG-labelled riboprobes for M2, M3 receptor and PPE
DNA fragments for PPE was firstly amplified by PCR from rat brain cDNA using
forward and reverse primers with SP6 and T7 promoters attached at the 5′ end,
respectively. Homology analysis using nucleotide BLAST searches of the National
Centre for Biotechnology Information (NCBI, USA) were carried out to ensure that
the RNA probe sequence was specific to the mRNA of interest only. Antisense
sequences, riboprobe length and GeneBank accession source numbers for M2
receptor, M3 receptor and PPE mRNA are shown in Table 1.

Supplemental Table 1. Sp6- and T7-tagged primer sequences for DIG-labelled
riboprobes
Gene Primer GenBank Nucleotide sequence (5’-->3’)* Size
(bp)
fwd ATTTAGGTGACACTATAGAAGtgcctccgttatgaatctcc M2
rvse
NM_031
016 TAATACGACTCACTATAGGGAGAcgacccaactagttctacagt
750
fwd ATTTAGGTGACACTATAGAAGatcaaaggaaacgctgtgct M3
rvse
NM_012
527 TAATACGACTCACTATAGGGAGAgtcggctgtaaacaccacct
514
fwd GGATCCATTTAGGTGACACTATAGAAGctaaatgcagctac
cgcctg
PPE
rvse
BC08356
3
GAATTCTAATACGACTCACTATAGGGAGAttccatctcgg
gaacttcttt
743
*Upper case denotes Sp6 and T7 tag sequences and lower case denotes primer
sequence
The antisense and sense riboprobes were then transcribed in vitro using DIG-11-UTP
(Roche Applied Sciences) and Ampriscibe T7 or Sp6 transcription Kit (Epicenter,
USA).
All sections were mounted onto slides with Prolong anti-fade (Molecular Probes,
Invitrogen, Australia) or Vectorshield Hard Set (Vector laboratories, USA) and
viewed using a fluorescent microscope (Leica DML or Zeiss imager Z.1, Germany).
Images were acquired using a SPOT2 digital camera (Diagnostic Instruments).

Fluorescence images were merged using SPOT2 and contrast adjusted using Zeiss
Imaging software.
PCR detection of M1-M5 receptor mRNA in the RVLM
SD (n=3) were anesthetised and perfused as previously described 13. The brainstem
was cut into 350 µm sections in freezing conditions using a vibratome (Leica
VT1000S). The RVLM (located ventral to the nucleus ambiguus in the two sections
spanning approximately 700 µm immediately caudal to the facial nucleus) and the
cerebellum (posterior lobe) were immediately dissected into RNAlater cryoprotectant
reagent (Ambion Inc, USA) and stored at -70 ºC.
Total RNA was isolated and reverse transcribed to cDNA as described previously 13.
PCR (Corbett Rotorgene 3000) was performed in a reaction solution (20 µl)
composed of 1 µl of cDNA, 100 µM each primer (see Table 2), 2 mM MgCl2, 0.25
mM dNTPs, 10X immunobuffer, 0.125 U Immolase hot-start DNA polymerase
(Bioline, Australia) and SYBR Green I fluorescent stain (1:40 000 dilution, Molecular
Probes, USA). Samples of the amplified M1-M5 receptor gene products were
electrophoresed in an ethidium bromide-stained 2% agarose gel. Gene products were
recovered enzymatically using QiaQuick gel extraction kit (Qiagen, Australia) and the
DNA was sequenced using the ABI Prism 3730 platform (SUPAMAC, Australia) to
confirm specificity to the GenBank sequence.
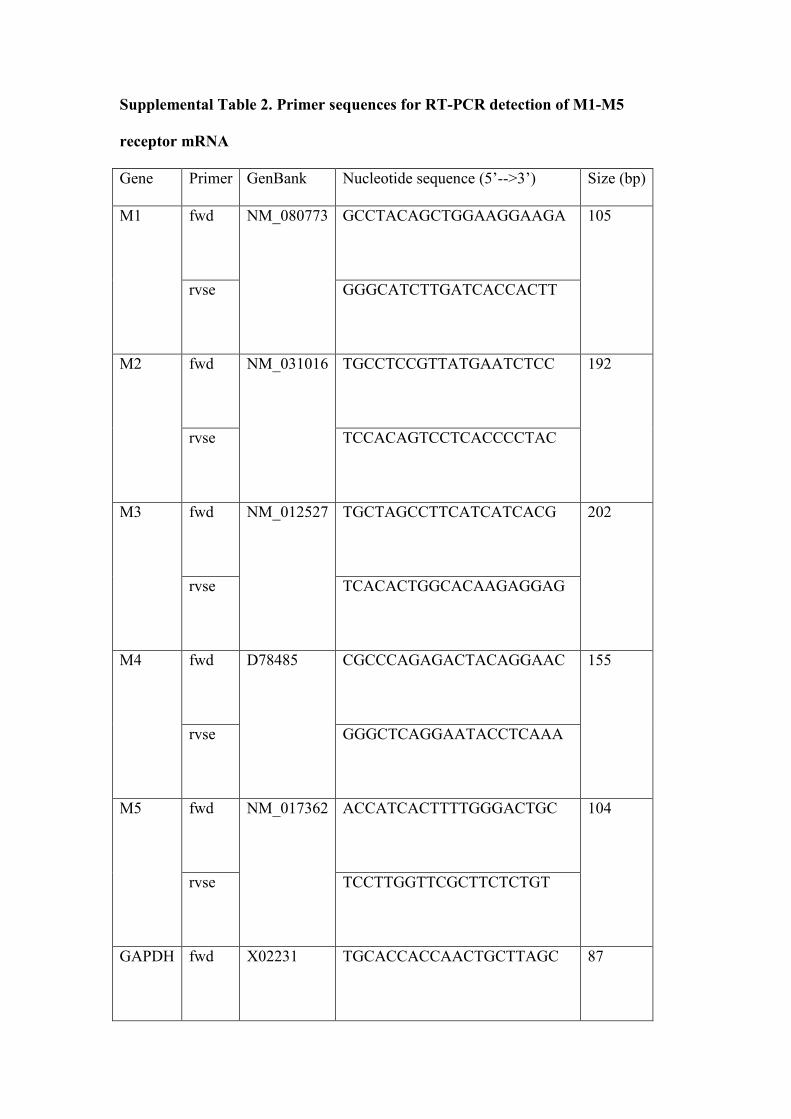
Supplemental Table 2. Primer sequences for RT-PCR detection of M1-M5
receptor mRNA
Gene Primer GenBank Nucleotide sequence (5’-->3’) Size (bp)
fwd GCCTACAGCTGGAAGGAAGA
M1
rvse
NM_080773
GGGCATCTTGATCACCACTT
105
fwd TGCCTCCGTTATGAATCTCC
M2
rvse
NM_031016
TCCACAGTCCTCACCCCTAC
192
fwd TGCTAGCCTTCATCATCACG
M3
rvse
NM_012527
TCACACTGGCACAAGAGGAG
202
fwd CGCCCAGAGACTACAGGAAC
M4
rvse
D78485
GGGCTCAGGAATACCTCAAA
155
fwd ACCATCACTTTTGGGACTGC
M5
rvse
NM_017362
TCCTTGGTTCGCTTCTCTGT
104
GAPDH fwd X02231
TGCACCACCAACTGCTTAGC
87

rvse GGCATGGACTGTGGTCATGAG
Detection of close appositions and cell counts
Light microscopic sections were viewed at high magnification (x 100) to identify close
appositions between vAChT-IR terminals (black reaction product) and CTB-labelled
neurons in the RVLM (brown reaction product). CTB- and TH-immunoreactive (IR)
cells were counted in every 4th section in the RVLM from the caudal pole of the facial
nucleus (VII) (Bregma -11.80 to -12.40 mm) 9 to determine the proportion of cells that
are closely apposed by vAChT-IR varicosities or express M2 receptor mRNA.
Expression of M3 receptor mRNA and close appositions from vAChT-IR was also
examined in NeuN-IR / PPE-positive cells.
Statistical analysis
All data are presented as mean ± standard error of the mean. A paired Students’ t-test
was used to calculate effects of treatment versus control or following SCOP and
P<0.05 was considered significant.

Supplemental Figure Legends
Figure S1 MAChR activation within the RVLM enhances the baroreflex and
inhibits the somatosympathetic and peripheral chemoreflex. Grouped data from nine
animals showing OXO-evoked effects on cardiorespiratory reflex function. OXO
enhances SNA baroreflex responses, inhibits both early and late peaks of the
somatosympathetic reflex and attenuates sympathoexcitatory and pressor effects of
the peripheral chemoreflex. Most reflexes return to control level following SCOP
injection bilaterally into the RVLM, although the early peak of the somatosympathetic
reflex remains attenuated and chemoreflex activation evokes no change in AP. Prior
injection of SCOP into the RVLM blocks all effects on cardiorespiratory reflexes
evoked by OXO. Data are mean ± SEM, *P<0.05, ** P<0.01, ns = non significant.
Figure S2 MAChR activation within the RVLM enhances low frequency (0.4 Hz)
and respiratory-related oscillations of SNA. Power spectra of SNA and SAP
(uniformly resampled at 10 Hz) are illustrated showing that the amplitude of the LF
peak is increased following OXO. Grouped data from seven animals are shown on
right illustrating the significant increase in LF power of SNA and SAP following
OXO and blockade of this effect by prior injection of SCOP bilaterally into the
RVLM. OXO also tended to increase HF oscillations of SNA but not SAP. Data are
mean ± SEM, *P<0.05, ** P<0.01, ns = non significant.
Figure S3 M1-M5 receptor subtypes are expressed in the RVLM. 2% Ethidium
bromide stained agarose gel with high intensity amplicons confirming the presence of
M1-M5 receptor mRNA in a tissue punch taken from the RVLM in an SD rat.

Figure S4 Comparison of ChAT and vAChT immunoreactivity in the ventral
medulla. Coronal sections of the medulla at the same level of the RVLM are shown at
low (left panel) and high power (right panel) from different SD rats that were perfused
transcardially. 50 µm brainstem sections were reacted for immunoreactivity against
ChAT (sheep, 1:500, Chemicon, USA) or vAChT (rabbit 1:500, Chemicon, USA) and
detected using nickel-intensified diaminobenzidine reactions 11. ChAT labelling in the
medulla identifies large motoneurons of the compact and loose formation of the
nucleus ambiguus that can be seen to send fine ChAT containing fibres ventrally into
the RVLM. Smaller ChAT-positive perikarya are clearly visible in the medial ventral
medulla. In comparison, vAChT labelling identifies ambiguual motoneurons and
cholinergic terminal fields in the RVLM but labelling in the ventromedial region is
clearly absent. Scale bars = 200 µm. Abbreviations: NAc, nucleus ambiguus pars
compacta; pyr, pyramidal tract; SpV, spinal trigeminal tract.

Supplemental References
1. Padley JR, Li Q, Pilowsky PM, Goodchild AK. Cannabinoid receptor
activation in the rostral ventrolateral medulla oblongata evokes
cardiorespiratory effects in anaesthetised rats. Br J Pharmacol. 2003;140:384-
394.
2. Miyawaki T, Goodchild AK, Pilowsky PM. Activation of mu-opioid receptors
in rat ventrolateral medulla selectively blocks baroreceptor reflexes while
activation of delta opioid receptors blocks somato-sympathetic reflexes.
Neuroscience. 2002;109:133-144.
3. Makeham JM, Goodchild AK, Pilowsky PM. NK1 receptor activation in rat
rostral ventrolateral medulla selectively attenuates somato-sympathetic reflex
while antagonism attenuates sympathetic chemoreflex. Am J Physiol Regul
Integr Comp Physiol. 2005;288:R1707-1715.
4. Miyawaki T, Goodchild AK, Pilowsky PM. Rostral ventral medulla 5-HT1A
receptors selectively inhibit the somatosympathetic reflex. Am J Physiol Regul
Integr Comp Physiol. May 2001;280(5):R1261-1268.
5. Verner TA, Goodchild AK, Pilowsky PM. A novel method for marking
microinjection sites using methylene blue and diaminobenzidine. J Neurosci
Methods. 2003;124:207-211.
6. Verner TA, Goodchild AK, Pilowsky PM. A mapping study of
cardiorespiratory responses to chemical stimulation of the midline medulla
oblongata in ventilated and freely breathing rats. Am J Physiol Regul Integr
Comp Physiol. 2004;287:R411-421.
7. Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 3rd ed. San
Diego: Academic Press, Inc.; 1996.
8. Padley JR, Overstreet DH, Pilowsky PM, Goodchild AK. Impaired cardiac
and sympathetic autonomic control in rats differing in acetylcholine receptor
sensitivity. Am J Physiol Heart Circ Physiol. 2005;289:H1985-92.
9. Phillips JK, Goodchild AK, Dubey R, Sesiashvili E, Takeda M, Chalmers J,
Pilowsky PM, Lipski J. Differential expression of catecholamine biosynthetic
enzymes in the rat ventrolateral medulla. J Comp Neurol. 2001;432:20-34.
10. Makeham JM, Goodchild AK, Pilowsky PM. NK1 receptor and the ventral
medulla of the rat: bulbospinal and catecholaminergic neurons. Neuroreport.
2001;12:3663-3667.
11. Goodchild AK, Llewellyn-Smith IJ, Sun QJ, Chalmers J, Cunningham AM,
Pilowsky PM. Calbindin-immunoreactive neurons in the reticular formation of
the rat brainstem: catecholamine content and spinal projections. J Comp
Neurol. 2000;424:547-562.
12. Li Q, Goodchild AK, Seyedabadi M, Pilowsky PM. Preprotachykinin A
mRNA is colocalized with tyrosine hydroxylase-immunoreactivity in
bulbospinal neurons. Neuroscience. 2005;136:205-216.
13. Kumar NN, Goodchild AK, Li Q, Pilowsky PM. An aldosterone-related
system in the ventrolateral medulla oblongata of spontaneously hypertensive
and Wistar-Kyoto rats. Clin Exp Pharmacol Physiol. 2006;33:71-75.

mATR
OXO
iv
SCOP R
VLM
OXO
post
SCOP
Ba
rore
ce
pto
r re
fle
xS
om
ato
sy
mp
ath
eti
c r
efl
ex
Ch
em
ore
ce
pto
r re
fle
x
120
20
(late peak)
**
*
120
20
Incr
ea
se in
SN
A(%
mA
TR
)
**
*
ns
120
20
(early peak)
Incr
ea
se in
SN
A(%
mA
TR
)
**
**
**
50
25
0
-25
Ch
an
ge
in M
AP
(mm
Hg
)
**
ns
Figure S1.
180
20
Re
du
ctio
n in
SN
A(%
mA
TR
)
****
(aortic nerve)
20
0
Incr
ea
se in
SN
A(%
SN
A/5
0m
mH
g)
***
(SNP)
mATR
OXO
iv
SCOP R
VLM
OXO
post
SCOP

mATR
OXO
iv
SCOP R
VLM
OXO
pos
t SCOP
2
2P
ow
er
(mm
Hg
or
SN
Aa
u) 100
10
1
0.1
100
10
1
0.1
LF
0.2
5-0
.75
Hz
HF
1-2
.5H
z
****
P=0.051
n.s.
SAP SNA
Figure S2.
frequency (Hz)
0.1 1
2p
ow
er
(mm
Hg
2 o
r S
NA
au
)100
-41x10
10
1
-31x10
-21x10
0.1
0.1 1 0.1 1
SAP
SNA
mATR OXO iv OXO iv post SCOP RVLM

Figure S3.

ChAT
vAChT
pyr
SpV
SpV
pyr
NAc
NAc
Figure S4.
ChAT
vAChT




















