
Carboxylesterase activity in earthworm gut contents: Potential (eco)toxicological implications
9
Carboxylesterase activity in earthworm gut contents: Potential (eco)toxicological implications Juan C. Sanchez-Hernandez a, ⁎, Christophe Mazzia b , Yvan Capowiez c , Magali Rault b a Laboratory of Ecotoxicology, Faculty of Environmental Science, University of Castilla-La Mancha, Avda. Carlos III s/n, 45071, Toledo, Spain b Universite d'Avignon et des Pays de Vaucluse, UMR 406 Abeilles et Environnement, Site AGROPARC, F-84914, Avignon Cedex 09, France c INRA, Unite PSH, Site AGROPARC, F-84914 Avignon Cedex 09, France abstract article info Article history: Received 12 June 2009 Received in revised form 21 July 2009 Accepted 22 July 2009 Available online 3 August 2009 Keywords: Gut secretion Lumbricus terrestris Luminal carboxylesterases Organophosphorus pesticides Pesticide-detoxifying esterases Carboxylesterases (CbEs) are key enzymes in pesticide detoxification. These esterases are involved in the biochemical mechanism for pesticide resistance in some pest species, and further they are considered an efficient protective mechanism against acute toxicity by organophosphate (OP) pesticides in mammals. To gain knowledge on the role of CbEs in pesticide toxicity and natural tolerance in earthworms, we performed an enzyme kinetic analysis to investigate whether these annelids are able to secrete them into their gut lumen. We determined levels of CbE activity and isozyme abundance in the gut wall and ingested soil collected from different portions of the gastrointestinal tract of Lumbricus terrestris. Moreover, modulation of enzyme activity by selected substrates (α-naphthyl acetate [α-NA], 4-nitrophenyl valerate [4-NPV] and 4-nitrophenyl acetate [4-NPA]) and OP pesticides was examined to compare the response between tissue and soil CbEs. We found a high CbE activity in the ingested soil extracts from the crop/gizzard (α-NA-CbE = 8.43± 2.76 U mg -1 protein and 4-NPA- CbE = 5.98 ± 2.11 U mg -1 protein) compared to the gut wall. Three lines of evidences suggest that the gut epithelium is the main source of this luminal CbE activity. First, the effect of substrate concentrations on CbE activity from both the ingested soil extracts and gut tissues resulted in similar apparent K m and V max values. Second, native PAGE gels revealed that some of the CbE isozymes in the gut tissue were also present in the soil extracts. Third, tissue and soil CbEs showed the same sensitivity to inhibition by OPs. The concentrations of insecticide causing 50% of esterase inhibition (IC 50 ) was comparable between tissue (IC 50 s range = 4.01–9.67 nM dichlorvos and 8480–6880 nM paraoxon) and soil (IC 50 s range=6.01–11.5 nM dichlorvos and 8400–7260 nM paraoxon). Our results suggest a set of (eco)toxicological implications and environmental applications derived from the ability of earthworms to secrete these pesticide-detoxifying enzymes. © 2009 Elsevier Inc. All rights reserved. 1. Introduction Earthworms occupy a notable place in terrestrial ecotoxicology. They are common standard organisms in soil toxicity testing to assess either harmful effects of new registered chemicals or toxicity of historically contaminated soils (van Straalen and van Gestel, 1998). Moreover, these organisms are demonstrated to be excellent bioindicators in the field monitoring of soil pollution by metals (Spurgeon and Hopkin, 1999), although some earthworm species are able to tolerate highly metal- contaminated soils (Spurgeon and Hopkin, 2000). The induction of metal-sequestrating proteins is among the more plausible causes for earthworm survival in heavily metal-polluted soils (Vijver et al., 2004). Likewise, pesticide-detoxifying enzymes such as carboxylesterases (CbE, EC 3.1.1.1) can make the pesticide ineffective against individuals that naturally have high levels of these enzymes (Stenersen, 2004). These esterases detoxify organophosphate (OP) and carbamate (CB) pesticides and synthetic pyrethroids (SPs) by two main ways; hydrolysis of the ester bond (CBs, SPs and those OPs containing this type of bond such as malathion) and binding of the pesticide (OPs) to the active site of CbE (Maxwell 1992; Chambers et al., 1994; Wheelock et al., 2005; Crow et al., 2007). Inhibition of CbE activity by OPs involves nucleophilic attack of the catalytic serine residue in the enzyme active site on the electronic deficient oxidized phosphorous to form a stable enzyme–inhibitor complex (Satoh and Hosokawa, 1998). This inhibition is considered a stoichiometric mechanism for decreasing the OP concentration at the target site, i.e., acetylcholinesterase (Maxwell, 1992; Chanda et al., 1997). Carboxylesterases have been described in earthworms (Haites et al., 1972; Øien and Stenersen, 1984). The intestinal tract of Lumbricus terrestris presents a wide variety of enzyme; among them, CbE activity is predominant in the peripherical chloragog tissue (Prento, 1987). Recently, Sanchez-Hernandez and Wheelock (2009) reported up to twelve CbE isozymes in the gastrointestinal tract, body wall muscle and reproductive tissues of L. terrestris, being particularly abundant in the crop, gizzard and foregut. The abundance of CbEs in the gastrointestinal Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511 ⁎ Corresponding author. Tel.: +34 925 268800; fax: +34 925 268840. E-mail address: [email protected] (J.C. Sanchez-Hernandez). 1532-0456/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.cbpc.2009.07.009 Contents lists available at ScienceDirect Comparative Biochemistry and Physiology, Part C journal homepage: www.elsevier.com/locate/cbpc
-
Upload
univ-avignon -
Category
Documents
-
view
1 -
download
0
Transcript of Carboxylesterase activity in earthworm gut contents: Potential (eco)toxicological implications

Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part C
j ourna l homepage: www.e lsev ie r.com/ locate /cbpc
Carboxylesterase activity in earthworm gut contents:Potential (eco)toxicological implications
Juan C. Sanchez-Hernandez a,⁎, Christophe Mazzia b, Yvan Capowiez c, Magali Rault b
a Laboratory of Ecotoxicology, Faculty of Environmental Science, University of Castilla-La Mancha, Avda. Carlos III s/n, 45071, Toledo, Spainb Universite d'Avignon et des Pays de Vaucluse, UMR 406 Abeilles et Environnement, Site AGROPARC, F-84914, Avignon Cedex 09, Francec INRA, Unite PSH, Site AGROPARC, F-84914 Avignon Cedex 09, France
⁎ Corresponding author. Tel.: +34 925 268800; fax: +E-mail address: [email protected] (J.C. San
1532-0456/$ – see front matter © 2009 Elsevier Inc. Aldoi:10.1016/j.cbpc.2009.07.009
a b s t r a c t
a r t i c l e i n f oArticle history:Received 12 June 2009Received in revised form 21 July 2009Accepted 22 July 2009Available online 3 August 2009
Keywords:Gut secretionLumbricus terrestrisLuminal carboxylesterasesOrganophosphorus pesticidesPesticide-detoxifying esterases
Carboxylesterases (CbEs) are key enzymes in pesticide detoxification. These esterases are involved in thebiochemicalmechanism for pesticide resistance in some pest species, and further they are considered an efficientprotective mechanism against acute toxicity by organophosphate (OP) pesticides in mammals. To gainknowledge on the role of CbEs in pesticide toxicity and natural tolerance in earthworms, we performed anenzyme kinetic analysis to investigate whether these annelids are able to secrete them into their gut lumen.Wedetermined levels of CbE activity and isozymeabundance in the gutwall and ingested soil collected fromdifferentportions of the gastrointestinal tract of Lumbricus terrestris. Moreover, modulation of enzyme activity by selectedsubstrates (α-naphthyl acetate [α-NA], 4-nitrophenyl valerate [4-NPV] and 4-nitrophenyl acetate [4-NPA]) andOP pesticides was examined to compare the response between tissue and soil CbEs.We found a high CbE activityin the ingested soil extracts from the crop/gizzard (α-NA-CbE=8.43±2.76 U mg−1 protein and 4-NPA-CbE=5.98±2.11 U mg−1 protein) compared to the gut wall. Three lines of evidences suggest that the gutepithelium is the main source of this luminal CbE activity. First, the effect of substrate concentrations on CbEactivity from both the ingested soil extracts and gut tissues resulted in similar apparent Km and Vmax values.Second, native PAGE gels revealed that some of the CbE isozymes in the gut tissue were also present in the soilextracts. Third, tissue and soil CbEs showed the same sensitivity to inhibition by OPs. The concentrations ofinsecticide causing 50% of esterase inhibition (IC50)was comparable between tissue (IC50s range=4.01–9.67 nMdichlorvos and 8480–6880 nM paraoxon) and soil (IC50s range=6.01–11.5 nM dichlorvos and 8400–7260 nMparaoxon). Our results suggest a set of (eco)toxicological implications and environmental applications derivedfrom the ability of earthworms to secrete these pesticide-detoxifying enzymes.
© 2009 Elsevier Inc. All rights reserved.
1. Introduction
Earthwormsoccupy anotable place in terrestrial ecotoxicology. Theyare common standard organisms in soil toxicity testing to assess eitherharmful effects of new registered chemicals or toxicity of historicallycontaminated soils (van Straalen andvanGestel, 1998).Moreover, theseorganisms are demonstrated to be excellent bioindicators in the fieldmonitoring of soil pollution by metals (Spurgeon and Hopkin, 1999),although some earthworm species are able to tolerate highly metal-contaminated soils (Spurgeon and Hopkin, 2000). The induction ofmetal-sequestrating proteins is among the more plausible causes forearthworm survival in heavily metal-polluted soils (Vijver et al., 2004).Likewise, pesticide-detoxifying enzymes such as carboxylesterases(CbE, EC 3.1.1.1) can make the pesticide ineffective against individualsthat naturally have high levels of these enzymes (Stenersen, 2004).These esterases detoxify organophosphate (OP) and carbamate (CB)
34 925 268840.chez-Hernandez).
l rights reserved.
pesticides and synthetic pyrethroids (SPs) by two main ways;hydrolysis of the ester bond (CBs, SPs and those OPs containing thistype of bond such as malathion) and binding of the pesticide (OPs) tothe active site of CbE (Maxwell 1992; Chambers et al., 1994; Wheelocket al., 2005; Crow et al., 2007). Inhibition of CbE activity by OPs involvesnucleophilic attack of the catalytic serine residue in the enzyme activesite on the electronic deficient oxidized phosphorous to form a stableenzyme–inhibitor complex (SatohandHosokawa, 1998). This inhibitionis considered a stoichiometric mechanism for decreasing the OPconcentration at the target site, i.e., acetylcholinesterase (Maxwell,1992; Chanda et al., 1997).
Carboxylesterases have been described in earthworms (Haites et al.,1972; Øien and Stenersen, 1984). The intestinal tract of Lumbricusterrestrispresents awide variety of enzyme; among them, CbE activity ispredominant in the peripherical chloragog tissue (Prento, 1987).Recently, Sanchez-Hernandez and Wheelock (2009) reported up totwelve CbE isozymes in the gastrointestinal tract, bodywall muscle andreproductive tissues of L. terrestris, being particularly abundant in thecrop, gizzard and foregut. The abundance of CbEs in the gastrointestinal

504 J.C. Sanchez-Hernandez et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
tract of earthworms leads to hypothesize if these enzymes represent anenzymatic barrier for reducing pesticide uptake from contaminatedingested soil, or even to speculate if these esterases can be secreted intothe gut lumen by earthworms themselves as this occurs with otherinvertebrates such as spiders (Mommsen, 1978), lepidopterans (Tur-unen and Chippendale, 1977), mosquitoes (Geering and Freyvogel,1974) or cockroaches (Cook et al., 1969).
The aims of this study were therefore to determine the presence ofCbEs in the earthworm gut content and to investigate whether gutwall is a source of luminal CbE activity. We measured the levels ofenzyme activity and isozyme abundance in tissue homogenates fromthe pharynx, crop/gizzard, foregut and midgut sections of thealimentary canal of L. terrestris. Soil samples collected from thesegut regions were also used for CbE determination. In addition, theeffect of selected substrates and inhibitors (OP insecticides) onenzyme activity was compared between tissue and soil CbE activities.These in vitro experiments provided solid evidences for a gutsecretion of these pesticide-metabolizing enzymes that suggestsome toxicological implications and environmental applications ofconcern in earthworm ecotoxicology.
2. Materials and methods
2.1. Reagents
The chemicals for CbE activity assays, α-naphthyl acetate (α-NA),4-nitrophenyl acetate (4-NPA), 4-nitrophenyl valerate (4-NPV), 4-nitrophenol, Fast Red ITR salt, Fast Blue RR salt and 4-methylumbelli-feryl acetate (4-MUBA) as well as lipase type II from porcine pancreas(100–400 U mg−1 protein) and lipase from Candida cylindracea(20.7 U mg−1 protein) were purchased from Sigma-Aldrich (Madrid,Spain). The OP pesticides (N98% purity) dichlorvos (2,2-dichlorovinyldimethyl phosphate) and paraoxon (O,O-diethyl O-(4-nitrophenyl)phosphate) were obtained from Dr. Ehrenstorfer (Augsburg, Ger-many). We used dichlorvos and paraoxon (the active oxonmetaboliteof parathion) as model pesticides because they are potent esteraseinhibitors (Chambers et al., 1994).
2.2. Experimental setup
Adults clitellate L. terrestris (3.4–5.0 g fresh mass) were obtainedfrom a commercial supplier (Decathlon, France) and kept individuallyin boxes containing 100 g of field soil (collected in uncontaminatedarea) during 1 week at 12–15 °C and darkness. Earthworms wereanesthetized by cooling (individuals were placed in petri dishes andkept on ice) to collect gut tissues and ingested soil. Dissection wasinitiated in the mid-dorsal line by a longitudinal incision. Pharynx,crop/gizzard, foregut (the gut segment between the gizzard and theclitellum) and midgut (the gut segment after the clitellum towardsthe anus) were removed and open separately to collect carefully theingested soil avoiding to scrape the epithelium. Soil present in the lastportion of the intestine (hindgut) corresponded to fresh cast.
Gut tissues, ingested soil, control soil (bulk soil from the box) andfresh cast were immediately transferred to 1.5-mL microcentrifugetubes and weighed. Tissue samples were homogenized in 10% (w/v)buffer containing 10 mM Tris–HCl (pH 7.3), 10 mM NaCl supplemen-ted with a cocktail of protease inhibitors (aprotinin, leupeptin andpepstatin=5 µg mL−1; antipain=1 µg mL−1; trypsin inhibitor=1 mg mL−1) using a conical hand-held homogenizer connected to abench-top overhead stirrer (IKA RW14 Basic). Fresh soil samples weresuspended in 10% (w/v) homogenization buffer and homogenized byrapid mechanical stirring for 1 min. Tissue homogenates and soilsuspensions were centrifuged at 3000 g for 10 min at 4 °C to obtainthe tissue post-mitochondrial supernatants (PMS) and the soilextracts, which were stored at −20 °C until analysis (within twomonths) with 10% glycerol as an enzyme stabilizing agent.
2.3. Carboxylesterase assays
Carboxylesterase activity was assayed spectrophotometrically usingthe substratesα-NA, 4-NPA and 4-NPV, and protocolswere adapted to amicroplate reader (Synergy HT, Bio-Tek). Carboxylesterase activityusingα-NAwas determined according to Gomori (1953), as adapted byBunyan et al. (1968). The reaction medium (200 µL, final volume)contained 25 mMTris–HCl (pH7.6), 1 mMCaCl2 and 2 mMα-NA, and itwas incubated for 10 min at 25 °C with the sample. The enzymaticformation of α-naphthol was stopped by the addition of 50 µL of 2.5%SDS in 0.1% Fast Red ITR/2.5% Triton X-100. Solutions were allowed tostand for 30 min at 22–23 ºC and dark. The absorbance of the naphthol–Fast Red ITR complex was read at 530 nm, and the concentration of α-naphthol was determined using a α-naphthol standard curve that wasmeasured under the same conditions. We used a reaction buffersupplementedwith CaCl2 because concomitant determination of Ca+2-dependent A-esterase activity (data not shown), however Ca+2 had noeffect on hydrolysis rate of α-NA.
Hydrolysis of 4-NPV and 4-NPA was determined as described byCarr and Chambers (1991) and Chanda et al. (1997), respectively. Thereaction mixture (250 µL, final volume including enzyme sample)contained 1 mM 4-NPV and 50 mMTris–HCl (pH 7.5) for assaying CbEactivity towards 4-NPV, or 5 mM4-NPA, 20 mMTris–HCl (pH 8.0) and1 mM EDTA when 4-NPA was used as substrate. In both methods, theformation of 4-nitrophenol was monitored for 5 min at 405 nm, 25 °Cand quantified using a 4-nitrophenol standard curve. Blanks (reactionmixture free of enzyme sample) were periodically checked fornonenzymatic hydrolysis of 4-NPV and 4-NPA, and no significantactivity was observed. Proteins were quantified according to theLowry method as modified by Markwell et al. (1978), and usingbovine serum albumin as the standard protein. All enzyme assayswere run in triplicate. Specific CbE activities were expressed as unitsper milligram of total proteins (U mg−1 protein). One unit of enzymeactivity is defined as one micromole of substrate hydrolyzed perminute under the experimental conditions described above.
2.4. Effect of substrate concentration on CbE activity
We examined the effect of the concentration of the substrates α-NA (range of tested concentrations=0.012–2.5 mM, final concentra-tion), 4-NPA (0.031–2.5 mM) and 4-NPV (0.012–5 mM) on tissue andsoil CbE activities. The kinetic parameters Km and Vmax associated tosubstrate-mediated modulation of CbE activity were estimated usingthe Michaelis–Menten equation, i.e., V0=Vmax×[S]/(Km+[S]). Con-centrations of substrate higher than 5 mM were not assayed becauseof substrate precipitation in the reaction medium.
2.5. Kinetic inhibition analysis
We studied the sensitivity of tissue and soil CbE activities todichlorvos and paraoxon. Themolar concentration of inhibitor causinga 50% reduction of initial enzyme activity (IC50) was calculated byplotting the logarithm of inhibitor concentration versus the percent-age of remaining CbE activity. Dose–response curves were fitted tothe four-parameter logistic equation y=min+(max−min)/1+(x/IC50)−HillSlope, where y is the percentage of residual CbE activitycompared to controls after a 30-min incubationwith the inhibitor,minis the y response to the highest concentration of inhibitor, max is yresponse to the lowest concentration of inhibitor, x is the logarithmicof inhibitor molar concentration and Hillslope describe the steepnessof the dose–response curve (Motulsky and Christopoulos, 2003).Esterase activity was assayed using α-NA solely because of the higherenzyme activity previously observed in both PMS and soil extracts.Aliquots of tissue or soil samples were incubated in the presence ofserial concentrations of dichlorvos (7×10−11−2.7×10−7 M) orparaoxon (7×10−12−2.7×10−4 M) for 30 min at 22–23 °C. Stock

505J.C. Sanchez-Hernandez et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
solutions (100 mM) of the OPs were prepared daily in dimethylsulfoxide (DMSO) and dilutions (b10−4 M, F.C.) weremade in distilledwater. The concentration of the solvent in the reaction medium waskept below 1.0%, which had no effect on CbE activity. Upon completionof the 30-min inhibition period, the residual CbE activity towards α-NA was assayed as described above. Inhibition kinetics were per-formed in triplicate.
We next determined the kinetic parameters (bimolecular inhibi-tion constant [Ki], phosphorylation constant [kp], and associationconstant [KA]) for esterase inhibition by dichlorvos and paraoxon asdescribed in Johnson and Wallace (1987). Aliquots of PMS or soilextract were incubated with 3 to 5 different concentrations of each OPpesticide. The residual CbE activity was periodically measured up tocomplete 10 min of incubation with dichlorvos, or 15 min withparaoxon. The interaction between CbE and the OP was stopped byadding 10 µL of α-NA, and the absorbance of the remaining esteraseactivity was read as described above. We calculated the kineticparameters describing the interaction between CbE and the OP byplotting the fraction of remaining CbE activity (Et/E0, where E0 is theinitial uninhibited esterase activity and Et is the residual esteraseactivity at time t) versus the time of incubation. These plots resultedin a linear relationship at each inhibitor concentration, where slope isthe apparent phosphorylation rate of CbE activity (−kapp). Thedouble-reciprocal plot between kapp values and the inhibitorconcentration was a linear regression with the slope, the y-interceptand the x-intercept as 1/Ki, 1/kp and -KA, respectively.
2.6. Native electrophoresis
Native polyacrylamide gel electrophoresis (PAGE) was performed onaBio-Radelectrophoresis unit (Bio-Rad,USA). Soil extracts andPMSwereloaded on 4% stacking and 12.5% resolving 0.75 mm polyacrylamide gel(25 mMTris, 192 mMglycine as running buffer), and electrophoresed ata constant voltage of 30 V for 30 min, and subsequently 150 V until thebromophenol blue tracking dye reached the bottom of the gel. Proteinbands were visualized by incubation (2–3 min at 22–23 °C) of the gelswith a staining solution containing 100 mM Na-phosphate buffer(pH=6.4), 0.5 mg mL−1 α-NA and 0.025 g of Fast Blue RR salt, whichwas prepared and filtered immediately before use.
We also examined the effect of dichlorvos on in-gel hydrolysis of CbEisozymes. Two post-electrophoresis gels were incubated with 50 mL1×10−7 M dichlorvos for 30 min at 22–23 ºC and continuous agitation,and subsequently the gels were washed with 100 mM Na-phosphatebuffer and stained for CbE activity. Likewise, relative mobility of CbEisozymes and in-gel sensitivity to dichlorvos were compared withcommercial lipases (pancreatic porcine lipase and C. cylindracea lipase),because the latter can also hydrolyze α-naphthyl esters (Gilham andLehner, 2005). In-gel hydrolysis stainingwas carried outwithα-NA/FastBlue RR salt (10-min incubation) and 4-MUBA (8-min incubation).Activity staining with 4-MUBA was performed according to Crow et al.(2007). A freshly prepared solution of 100 mM of Na-phosphate buffer(pH 6.5) containing 100 µM 4-MUBA (stock solution 20 mM in DMSO)was added to gels and stained bands were visualised in an UV-transilluminator. Stained gels were scanned using the Kodak DigitalScience 1D Image Analysis Software, version 2.03 (Rochester, NY, USA).
2.7. Statistical analysis
Significant differences in the mean CbE activity among the guttissues and soil extracts were tested using the non-parametric Kruskal–Wallis ANOVA test. Sigmoid regressions for estimating IC50s as well asthe Michaelis–Menten hyperbolic function for calculating apparent Km
and Vmaxwere obtained from the non-linear regression library includedin the SigmaPlot software (version 9.01, Systat Software, PointRichmond, CA, USA). Regression analysis was significant at a level ofprobability less than 0.05.
3. Results
3.1. Carboxylesterase activity levels
Mean (±standard deviation) of total proteins in the PMS were4.75±0.40 (pharynx), 5.24±0.81 (crop/gizzard), 6.92±1.09 (fore-gut) and 4.85±0.53 mg mL−1 (midgut), whereas the proteinconcentrations in the soil extracts were 2.02±0.58 mg g−1 freshweight (casts), 1.67±0.30 mg g−1 (control soil), 1.48±1.19 mg g−1
(soil from pharynx), 33.9±13.09 mg g−1 (soil from crop/gizzard),46.4±12.06 mg g−1 (soil from foregut) and 5.05±2.28 mg g−1 (soilfrom midgut). Carboxylesterase activity was found in all gut tissuesirrespective of the substrate used for enzyme assay (Fig. 1). However,there was a marked regional distribution of the esterase activity; thehighest specific activities were found in the crop/gizzard (0.93±0.10 U mg−1 protein, with α-NA, Fig. 1A) and foregut (0.85±0.14 Umg−1 and 0.66±0.12 U mg−1 with 4-NPA and 4-NPV, respectively,Fig. 1B and C). Ingested soil extracts collected at pharynx, crop/gizzard, foregut and midgut regions also showed CbE activity. Levelsof soil CbE activity from crop/gizzard section and using α-NA (8.43±2.76 U mg−1 protein) and 4-NPA (5.98±2.11 U mg−1 protein) wereone order ofmagnitude higher compared to thosemeasured in the gutwall (Fig. 1A and B). However, when we used 4-NPV, the levels ofesterase activity were comparable between tissue and soil or evenlower in the ingested soil from foregut and midgut compared to thecorresponding gut wall (Fig. 1C). In general, CbE activity reached apeak of maximum activity in the crop/gizzard, and decreased as soilprogressed through the intestinal tract. We detected no esteraseactivity in both fresh casts and the control soil.
Native PAGE revealed multiple esterase isozymes in the gut tissues(Fig. 1D).We distinguished up to eleven protein bands in the PAGE gelthat were numbered as ES1 to ES11 according to their relativemobility. The isozyme abundance and staining intensity variedmarkedly with the gut region. Native PAGE of ingested soil extractsfrom crop/gizzard, foregut and midgut showed two bands (ES6 andES7) that had the same relative mobility than those of the PMS. Thesetwo protein bands of soil extracts displayed a high hydrolytic activitytowards α-NA corroborating the levels of specific CbE activitymeasured in ingested soil extracts. Other three bands (ES9-ES11)with low mobility on PAGE gel were evident in the soil extracts fromcrop/gizzard and foregut, but weakly stained in the PMS from crop/gizzard.
3.2. Effect of substrate concentration on carboxylesterase activity
The CbE activity exhibited a Michaelis–Menten kinetic regardlessof the enzyme source (tissue or soil) (Fig. 2). We determined theapparent Km and Vmax directly from the hyperbolic equation(r2N0.90; Pb0.0001) when esterase activity was assayed with α-NAor 4-NPA. Nevertheless, the kinetic parameters for the hydrolysis of 4-NPV were calculated from the double-reciprocal Lineweaver–Burkplots (r2N0.96; Pb0.001) of both the initial velocity and substrateconcentration because of a deficient fitting of the experimental data tothe hyperbolic enzyme kinetic (Fig. 2C). Table 1 summarizes theapparent Km and Vmax values for the hydrolysis of the three substrates.The substrates α-NA and 4-NPA were more efficiently hydrolized bytissue (Vmax=0.27–0.77 U mg−1 protein) and soil (Vmax=6.16–12.5 Umg−1 protein) CbE activities compared to 4-NPV. Furthermore,the affinity of tissue CbE activity for the substrate followed the order4-NPV (Km=40.3 and 61.5 µM)Nα-NA (Km=75.6 and 79.6 µM)N4-NPA (Km=150 and 204 µM), whereas α-NA exhibited a higheraffinity for soil CbE activity. When comparing kinetic parametersbetween tissue and soil CbEs, Vmax increased markedly for soil CbEactivity using α-NA or 4-NPA, irrespective of the gut region, althoughthe apparent Km was similar. The contrary effect was observed when4-NPV was the substrate of the enzymatic reaction; Vmax did not vary

Fig. 1.Mean and standard deviation (n=7 earthworms) of carboxylesterase (CbE) activity in gut tissue and ingested soil in Lumbricus terrestris. Esterase activity wasmeasured usingthe substrates α-naphthyl acetate (α-NA, panel A), 4-nitrophenyl acetate (4-NPA, panel B) and 4-nitrophenyl valerate (4-NPV, panel C). Because tissue and soil CbE activitiestowards 4-NPV varied within the same rage of magnitude (0–0.8 U mg−1 protein), the same Y-axis was used for both plots. Panel D) Native PAGE of gut tissue supernatants andingested soil extracts from L. terrestris. Protein bands were detected and stained usingα-NA/Fast Blue RR. Lane 1: pharynx (41.16 µg protein), lane 2: crop/gizzard (43.32 µg protein),lane 3: foregut (58.52 µg protein), lane 4: midgut (45.52 µg protein), lane 5: soil extract collected from pharynx (0.48 µg protein), lane 6: soil extract from crop/gizzard (38.08 µgprotein), lane 7: soil extract from foregut (44.95 µg protein) and lane 8: soil extract from midgut (6.43 µg protein). ES depicts the esterase bands visualized in the gel. Diagram onpanel A) is a representation of gastrointestinal tract of earthworms (based on information in Brown and Doube, 2004; and Drake and Horn, 2007).
506 J.C. Sanchez-Hernandez et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
between tissue and soil CbEs, but the apparent Km was two- or three-fold higher for the soil CbE activity (Table 1). The catalytic efficiency ofCbE activity estimated as the Vmax/Km ratio was more than one orderof magnitude higher for soil CbE activity compared to that of guttissue, and it followed the order α-NAN4-NPA or 4-NPV for bothtissue and soil esterase activities.
3.3. Sensitivity of carboxylesterases to organophosphorus insecticides
We examined the sensitivity of both tissue and soil CbE activitiesusing α-NA to selected OP pesticides as well as their kineticparameters for enzyme inactivation. These in vitro experimentsprovided further indirect evidence for a luminal esterase activitysecreted by gut tissue. The sensitivity of tissue and soil CbE activitiesto the inhibitor followed a sigmoidal model (r2N0.97, Pb0.0001)regardless of the OP (Fig. 3). Dichlorvos proved a more potentinhibitor (IC50s=4.01 to 11.5 nM) than paraoxon (IC50s=6.88 to8.48 µM) (Table 2). IC50s were similar between tissue and soil CbEsmeasured at each gut region, except for the midgut tissue which
exhibited a CbE-IC50 of 0.77 nM for paraoxon (Fig. 3B). Thisunexpected result could be due to contamination of the midgutepithelium by the chloragog tissue. We decided therefore to avoid themidgut for further inhibition kinetic studies.
Fig. 4 shows the relationship between the fraction of remainingCbE activity (Et/E0) and the time of incubation with each OP pesticide.Linear regressions (r2N0.89, Pb0.05) enabled kapp values to becalculated, and plotted against the concentrations of the OP pesticide(Fig. 5). These representations allowed us to estimate the kineticconstants for CbE-OP interactions. In general, marked OP- and gutregion-specific differences were found in the kinetic constants for CbEinhibition. However, the inhibition kinetic constants were rathersimilar between tissue and soil CbEs of a particular intestinal region(Table 2). Considering dichlorvos interaction with CbE activity, KA andKi were higher in the foregut region than in crop/gizzard section,whereas the phosphorylation rate of CbE activity was faster in thecrop/gizzard. In the case of CbE inhibition by paraoxon, the differencesin the kinetic constants between the gut regions were not as markedas for dichlorvos.

Fig. 2. Effect of α-naphthyl acetate (α-NA) and 4-nitrophenyl acetate (4-NPA) concentrations on carboxylesterase (CbE) activity measured in gut tissue (panel A) and ingested soil(panel B) collected from crop/gizzard and foregut regions of Lumbricus terrestris. Kinetic parameters (apparent Km and Vmax) were estimated by fitting the data to the Michaelis–Menten equation. The apparent Km and Vmax associated to the hydrolysis of substrate 4-nitrophenyl valerate (4-NPV) was estimated by the Lineweaver–Burk double-reciprocal plot(panel C). Data are the average and standard deviation of three independent assays. Table 1 summarizes the kinetic parameters.
507J.C. Sanchez-Hernandez et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
We also investigated the contribution of lipases (EC 3.1.1.3) to thehydrolytic activity ofα-NA, 4-NPA and 4-NPV. The commercial lipasesfromporcine pancreas and from C. cylindraceawere run in native PAGEgels. Untreated and dichlorvos-treated post-electrophoresis gels werestained with α-NA/Fast Blue RR salt and 4-MUBA. These electropho-resis studies revealed that although commercial lipases were ableto hydrolysis both α-NA and 4-MUBA, they did not show the samerelative mobility that CbE isoforms found in L. terrestris (Fig. 6).Furthermore, 1×10−7 M dichlorvos caused a marked staining reduc-tion of CbE isozymes from gut tissue and soil extracts, but the intensityof the stained bands corresponding to lipase from C. cylindracea wasnot apparently decreased. Nevertheless, we believe that the proteinbands named as ES9 to ES11 of the soil extracts could be lipases
Table 1Kinetic parametersa for hydrolysis of α-naphthyl acetate (α-NA), 4-nitrophenyl acetate(4-NPA) and 4-nitrophenyl valerate (4-NPV) in the tissue homogenates and soilextracts from crop/gizzard and foregut of Lumbricus terrestris.
Crop/gizzard Foregut
Tissue Soil Tissue Soil
α-NAKm (µM) 79.6 42.2 75.6 48.9Vmax (U mg−1 protein) 0.77 6.16 0.32 11.8Vmax/Km
(mL min−1 mg−1 protein)9.67 146 4.23 241
4-NPAKm (µM) 150 232 204 213Vmax (U mg−1 protein) 0.67 9.61 0.27 12.5Vmax/Km
(mL min−1 mg−1 protein)4.46 41.4 0.96 58.7
4-NPVKm (µM) 40.3 113 61.5 110Vmax (U mg−1 protein) 0.15 0.43 0.13 0.44Vmax/Km
(mL min−1 mg−1 protein)3.72 3.80 2.11 4.00
Total proteins (mg g−1)b 65.5±10.1 33.9±13.1 86.5±13.7 46.4±12.0
a Kinetic parameters were estimated by fitting experimental data to the Michaelis–Menten equation or transformation to linearity using the double-reciprocal Line-aweaver–Burk plot in the case of 4-NPV. Data are the average of three independentassays (see Fig. 2).
b Protein concentration is the mean and standard deviation of seven samples.
because they displayed a relative mobility similar to those of standardC. cylindracea lipase and, further, they did not show a decrease of thestaining intensity in the dichlorvos-treated gel.
4. Discussion
4.1. Evidence for intestinal secretion of carboxylesterases
Carboxylesterase activity was found in the gut content of L. terrestris,and the hydrolytic activity towards α-NA and 4-NPA was one order ofmagnitude higher in the gut content than in the gut wall. Three lines ofevidences suggest that the source of this esterase is the gut epithelium.First, native PAGE gels of soil extracts collected from gut revealed twohighly stained protein bandswhich exhibited the same relativemobilitythat those observed in the gut tissue (Fig. 1D). Second, substrate-mediated modulation of CbE activity was comparable between tissueand ingested soil (Table 1). Third, tissue and soil CbE activities showed asimilar sensitivity to inhibition by OP insecticides (Fig. 3 and Table 2).High CbE activity levels are also present in the small intestine ofmammals (Satoh and Hosokawa, 1998). Likewise, an in vitro work bySanchez-Hernandez and Wheelock (2009) evidenced that CbE activityis well represented in the gastrointestinal tract of L. terrestris comparedto other tissues such as the body wall muscle or the reproductiveorgans.Why these esterases are abundant in the gastrointestinal tract ofL. terrestris can be explained by their likely involvement in the lipidmetabolism.
Intestinal secretion of CbEs seems a usual phenomenon inarthropods. For example, Turunen and Chippendale (1977) foundseveral CbE isozymes in the midgut fluids of the larval southwestercorn borer (D. grandiosella), which were sensitive to 10−5 M eserineand 10−4 M dichlorvos. Similarly, Geering and Freyvogel (1974)showed an unspecific esterase activity that increased its hydrolyticactivity in the midgut lumen of female mosquitoes (Aedes aegypti)after blood ingestion; although such an increase was not due to bloodesterases from the host. They suggested that secretion of theseesterases could be involved in the lipid metabolism and mobilization.A maximum of six CbE isozymes were distinguished by nativeelectrophoresis of the digestive secretion in cockroach, which alsocorresponded to those found in the midgut wall (Cook et al., 1969).Similarly, the digestive secretion of the spider Tegenaria atrica showedsix CbE isozymes, which could have also lipolytic activity (Mommsen,1978). A study by Prento (1987) revealed up to twenty different

Fig. 3. Invitro inhibitioncurves for tissueandsoil carboxylesterases (CbEs) fromcrop/gizzard,foregut andmidgut regions of the Lumbricus terrestris gut. Esterase activitywas assayed usingα-naphthyl acetate and inhibitors were dichlorvos (panel A) and paraoxon (panel B). Eachsymbol represents the mean and standard deviation of three independent assays.
508 J.C. Sanchez-Hernandez et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
enzymes (dehydrogenases, glucosidic enzymes, antioxidant enzymes,esterases, etc.) present in the midgut region of L. terrestris. In thatstudy, esterase activity using α-NA was particularly abundant in the
Table 2Kinetic constantsa for in vitro inhibition of carboxylesterase activityb by dichlorvos and par
Chemical Gut region Material IC50c (nM)
Dichlorvos Crop/gizzard Tissue 5.02Soil extract 6.62
Foregut Tissue 5.83Soil extract 8.60
Paraoxon Crop/gizzard Tissue 8236Soil extract 8409
Foregut Tissue 8029Soil extract 7423
a Inhibition kinetic parameters are the average of three independent assays.KA=associationb Enzyme activity was assayed using α-naphthyl acetate as substrate.c Concentration of organophosphorus insecticide required to inhibit 50% of esterase activ
chloragocyte fraction of the midgut, and it was suggested to beinvolved in the lipid metabolism.
Our results are in line with the previous findings by Prento (1987),and suggest an intestinal secretion of esterases in L. terrestris.However, caution must be taken into account because symbioticmicroorganisms could be involved in the presence of the high CbEactivity found in the earthworm gut content. Although symbiontscontribute to the digestive enzyme cocktail in the earthworm gut(Brown and Doube, 2004; Drake and Horn, 2007), a set of indirectevidences suggest that contribution of gut microflora to total CbEactivity should be minimal. Specific CbE activity of soil microorgan-isms is lower than that measured for soil extracts in this study. Forexample, Hatanaka et al. (1989) reported a CbE activity using 4-NPAof 0.028 U mg−1 protein in a Pseudomonas bacterium (measured incell extracts which were incubated at 37 ºC, pH=7.0 and 1.0 mM 4-NPA). This CbE activity is more than 2 orders of magnitude lower thanthat found in the soil collected from the crop/gizzard and foregutregions of L. terrestris, i.e., 5.98±2.11 and 5.30±1.92 Umg−1 protein,respectively. Despite contribution of microorganisms to luminal CbEactivity in earthworm cannot be totally excluded, interaction of thisesterase with dichlorvos or paraoxon is another line of evidence for aminimal contribution of symbionts to luminal CbE activity. Tissue andsoil CbEs showed similar IC50s and kinetic constants of inhibition bythese two OPs. These observations suggest therefore that the sameproteins account for the total CbE activity in both the gut content andthe gut tissue.
Another confounding factor is the participation of digestive enzymessuch as lipases to the hydrolysis of the selected substrates in this study.Although lipases preferentially cleave long-chain acylglycerols, they canalso hydrolyze naphthyl esters and 4-nitrophenyl esters (Gilham andLehner, 2005).Moreover, lipases canbe inhibitedbyOPpesticides (Krysanand Guss, 1971; Casida et al., 2008). For example, four of the six esteraseisozymes described by Cook et al. (1969) in the digestive secretion of thecockroach could be lipases because triglyceride hydrolysis decreased afterfull inhibition of these isozymes with 10−4 M dichlorvos. However,although lipases couldbepartially responsible for luminal esterase activityin L. terrestris as suggested by the electrophoretic outcomes, the substrate-and inhibitor-mediated modulation of tissue and soil esterase activitiespoint to CbEs as responsible for the high hydrolytic rate of α-NA and 4-NPA measured in the ingested soil extracts.
The soil CbE activity decreased progressively during the gutpassage of soil up to be almost undetectable in the fresh casts. Thereason could be the own digestive function of the midgut. Thissegment of the alimentary canal is highly specialized in the enzymaticdigestion of organic compounds and the presence of proteases couldcontribute to CbE degradation (Brown and Doube, 2004). Indeed, weobserved a significant decrease of the total protein concentration inthe ingested soil from the midgut (5.05±2.28 mg g−1 fresh soil)compared to that from the crop/gizzard (33.9±13.09 mg g−1 freshsoil). However, Tillinghast et al. (2001) found a decrease of theprotein content as well as the activity of some enzymes (proteasesand amylase) in the gut content of L. terrestris, but they did not
aoxon.
KA (µM−1) kp (min−1) Ki (µM−1 min−1)
0.62 6.04 3.780.42 6.72 2.843.65 1.88 6.893.80 1.75 6.660.0064 0.63 0.00400.0076 0.54 0.00420.0030 1.03 0.00300.0031 1.28 0.0040
rate constant; kp=phosphorylation rate constant andKi=bimolecular inhibition constant.
ity.

Fig. 4. Relationship between the remaining carboxylesterase (CbE) activity and the time after incubation with the organophosphate dichlorvos. Graph (A): Inhibition curves for crop/gizzard tissue. Graph (B): Inhibition curves for gut content collected from crop/gizzard region. Dichlorvos concentrations were: 27, 70, 130, 270, 340 and 700 nM. The slope of theregression lines is the apparent rate of CbE phosphorylation (−kapp). Symbols represent the mean and standard deviation of three independent assays. Circles are the CbE activitymeasured at time intervals between 0.5 and 140 s, whereas triangles represent those activity values measured at time intervals between 0.5 and 10 min.
509J.C. Sanchez-Hernandez et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
reproduce the same results when isolated gut contents wereincubated overnight. These observations suggest that loss of CbEactivity in our earthworms probably was not related to proteases. Onthe other hand, reabsorption of CbEs by the gut epithelium is not aconclusive explanation because a marked increase of the esteraseactivity in the midgut wall would occur compared to that from thecrop/gizzard. It does remain uncertain what mechanisms operate todecrease this esterase activity in the final portion of the L. terrestris; aloss of activity that also occurs for other enzymes such as cellulase(Nozaki et al., 2009).
4.2. (Eco)toxicological implications and environmental applications
Current results suggest a set of (eco)toxicological issues whenearthworms are used for toxicity testing of soil pollution or in fieldmonitoring of pesticide pollution. Earthworm exposure to soil pollu-
Fig. 5. Double-reciprocal plots of dichlorvos (panel A) and paraoxon (panel B) concentraline regressions is −1/Ki (Ki=bimolecular inhibition constant), intercept with y-axis is 1/constant). Inhibition kinetic constants are summarized in Table 2.
tants takes place by passive diffusion through their skin and ingestion ofcontaminated soil (Krauss et al., 2000). Intestinal absorption seems to bethe main uptake route for many hydrophobic contaminants in earth-worms, probably because these chemicals are strongly associated withthe organic matter in soil, reducing thereby the diffusion throughdermis. Indeed, Yu et al. (2006) showed that the high uptake ofchlorpyrifos by the earthworm Allolobophora caliginosa was due toactive ingestion of the pesticide present in the ingested soil, whilstuptake of chlorpyrifos by the dermal surface was negligible. Carbox-ylesterases participate actively in the detoxification of anti-ChEpesticides and SPs, either by hydrolysis or sequestering (irreversibleinhibition) the pesticide (Maxwell 1992; Sogorb and Vilanova, 2002;Crow et al., 2007). The high esterase activity levels found in the gutlumenof L. terrestrismightprovide anenzymatic barrier to the intestinaluptake of pesticides. This could lead to a certain natural tolerance ofearthworms to anti-ChE- and SP-contaminated soils. However, the
tions versus the apparent rate of carboxylesterase phosphorylation (kapp). Slopes ofkp (kp=phosphorylation rate constant) and x-intercept is −KA (KA=association rate
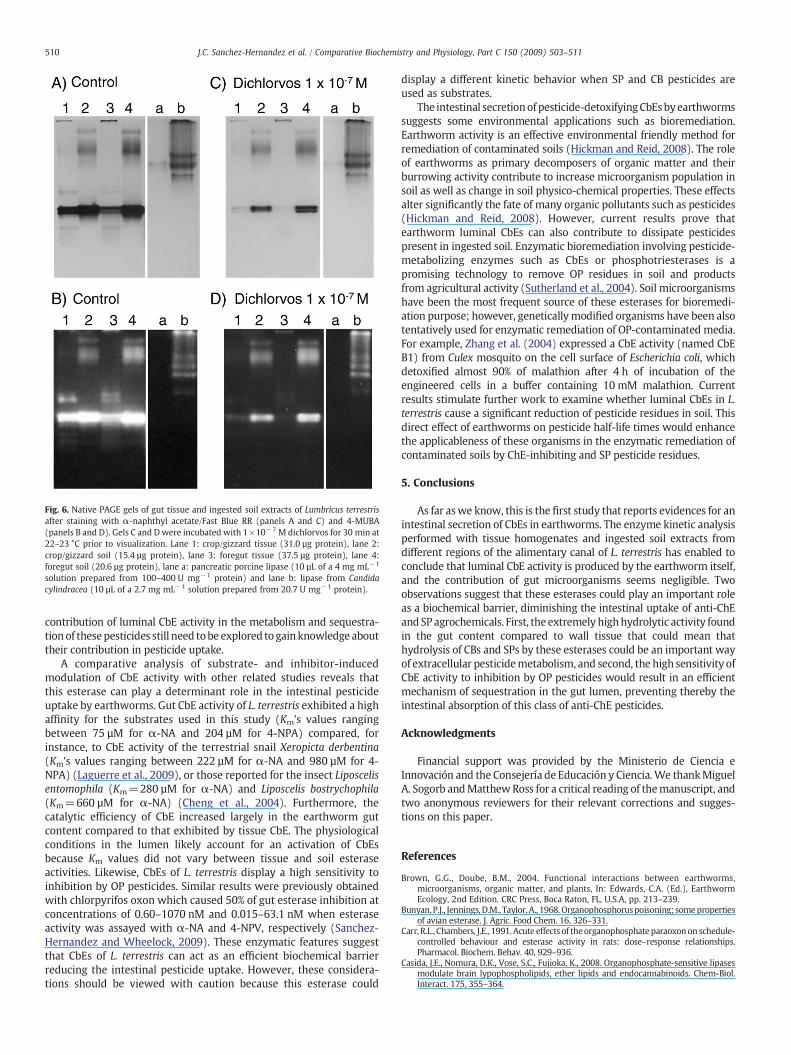
Fig. 6. Native PAGE gels of gut tissue and ingested soil extracts of Lumbricus terrestrisafter staining with α-naphthyl acetate/Fast Blue RR (panels A and C) and 4-MUBA(panels B and D). Gels C and Dwere incubated with 1×10−7 M dichlorvos for 30 min at22–23 °C prior to visualization. Lane 1: crop/gizzard tissue (31.0 µg protein), lane 2:crop/gizzard soil (15.4 µg protein), lane 3: foregut tissue (37.5 µg protein), lane 4:foregut soil (20.6 µg protein), lane a: pancreatic porcine lipase (10 µL of a 4 mg mL−1
solution prepared from 100–400 U mg−1 protein) and lane b: lipase from Candidacylindracea (10 µL of a 2.7 mg mL−1 solution prepared from 20.7 U mg−1 protein).
510 J.C. Sanchez-Hernandez et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
contribution of luminal CbE activity in the metabolism and sequestra-tion of thesepesticides still need tobe explored to gainknowledge abouttheir contribution in pesticide uptake.
A comparative analysis of substrate- and inhibitor-inducedmodulation of CbE activity with other related studies reveals thatthis esterase can play a determinant role in the intestinal pesticideuptake by earthworms. Gut CbE activity of L. terrestris exhibited a highaffinity for the substrates used in this study (Km's values rangingbetween 75 µM for α-NA and 204 µM for 4-NPA) compared, forinstance, to CbE activity of the terrestrial snail Xeropicta derbentina(Km's values ranging between 222 µM for α-NA and 980 µM for 4-NPA) (Laguerre et al., 2009), or those reported for the insect Liposcelisentomophila (Km=280 µM for α-NA) and Liposcelis bostrychophila(Km=660 µM for α-NA) (Cheng et al., 2004). Furthermore, thecatalytic efficiency of CbE increased largely in the earthworm gutcontent compared to that exhibited by tissue CbE. The physiologicalconditions in the lumen likely account for an activation of CbEsbecause Km values did not vary between tissue and soil esteraseactivities. Likewise, CbEs of L. terrestris display a high sensitivity toinhibition by OP pesticides. Similar results were previously obtainedwith chlorpyrifos oxon which caused 50% of gut esterase inhibition atconcentrations of 0.60–1070 nM and 0.015–63.1 nM when esteraseactivity was assayed with α-NA and 4-NPV, respectively (Sanchez-Hernandez and Wheelock, 2009). These enzymatic features suggestthat CbEs of L. terrestris can act as an efficient biochemical barrierreducing the intestinal pesticide uptake. However, these considera-tions should be viewed with caution because this esterase could
display a different kinetic behavior when SP and CB pesticides areused as substrates.
The intestinal secretionof pesticide-detoxifyingCbEsbyearthwormssuggests some environmental applications such as bioremediation.Earthworm activity is an effective environmental friendly method forremediation of contaminated soils (Hickman and Reid, 2008). The roleof earthworms as primary decomposers of organic matter and theirburrowing activity contribute to increase microorganism population insoil as well as change in soil physico-chemical properties. These effectsalter significantly the fate of many organic pollutants such as pesticides(Hickman and Reid, 2008). However, current results prove thatearthworm luminal CbEs can also contribute to dissipate pesticidespresent in ingested soil. Enzymatic bioremediation involving pesticide-metabolizing enzymes such as CbEs or phosphotriesterases is apromising technology to remove OP residues in soil and productsfrom agricultural activity (Sutherland et al., 2004). Soil microorganismshave been the most frequent source of these esterases for bioremedi-ation purpose; however, geneticallymodified organisms have been alsotentatively used for enzymatic remediation of OP-contaminated media.For example, Zhang et al. (2004) expressed a CbE activity (named CbEB1) from Culex mosquito on the cell surface of Escherichia coli, whichdetoxified almost 90% of malathion after 4 h of incubation of theengineered cells in a buffer containing 10 mM malathion. Currentresults stimulate further work to examine whether luminal CbEs in L.terrestris cause a significant reduction of pesticide residues in soil. Thisdirect effect of earthworms on pesticide half-life times would enhancethe applicableness of these organisms in the enzymatic remediation ofcontaminated soils by ChE-inhibiting and SP pesticide residues.
5. Conclusions
As far aswe know, this is the first study that reports evidences for anintestinal secretion of CbEs in earthworms. The enzyme kinetic analysisperformed with tissue homogenates and ingested soil extracts fromdifferent regions of the alimentary canal of L. terrestris has enabled toconclude that luminal CbE activity is produced by the earthworm itself,and the contribution of gut microorganisms seems negligible. Twoobservations suggest that these esterases could play an important roleas a biochemical barrier, diminishing the intestinal uptake of anti-ChEand SP agrochemicals. First, the extremely highhydrolytic activity foundin the gut content compared to wall tissue that could mean thathydrolysis of CBs and SPs by these esterases could be an important wayof extracellular pesticidemetabolism, and second, the high sensitivity ofCbE activity to inhibition by OP pesticides would result in an efficientmechanism of sequestration in the gut lumen, preventing thereby theintestinal absorption of this class of anti-ChE pesticides.
Acknowledgments
Financial support was provided by the Ministerio de Ciencia eInnovación and the Consejería de Educación y Ciencia.We thankMiguelA. Sogorb andMatthewRoss for a critical reading of themanuscript, andtwo anonymous reviewers for their relevant corrections and sugges-tions on this paper.
References
Brown, G.G., Doube, B.M., 2004. Functional interactions between earthworms,microorganisms, organic matter, and plants, In: Edwards, C.A. (Ed.), EarthwormEcology, 2nd Edition. CRC Press, Boca Raton, FL, U.S.A, pp. 213–239.
Bunyan,P.J., Jennings,D.M., Taylor,A., 1968.Organophosphoruspoisoning; somepropertiesof avian esterase. J. Agric. Food Chem. 16, 326–331.
Carr, R.L., Chambers, J.E., 1991. Acute effects of theorganophosphate paraoxonon schedule-controlled behaviour and esterase activity in rats: dose–response relationships.Pharmacol. Biochem. Behav. 40, 929–936.
Casida, J.E., Nomura, D.K., Vose, S.C., Fujioka, K., 2008. Organophosphate-sensitive lipasesmodulate brain lypophospholipids, ether lipids and endocannabinoids. Chem-Biol.Interact. 175, 355–364.

511J.C. Sanchez-Hernandez et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 503–511
Chambers, J.E., Ma, T.G., Boone, J.S., Chambers, H.W., 1994. Role of detoxificationpathway in acute toxicity levels of phosphorothionate insecticides in the rat. LifeSci. 54, 1357–1364.
Chanda, S.M., Mortensen, S.R., Moser, V.C., Padilla, S., 1997. Tissue-specific effects ofchlorpyrifos on carboxylesterase and cholinesterase activity in adult rats: an in vitroand in vivo comparison. Fundam. Appl. Toxicol. 38, 148–157.
Cheng, W.X., Wang, J.J., Ding, W., Zhao, Z.M., 2004. Inhibition kinetics on carbox-ylesterase and acetylcholinesterase of Liposcelis bostrychophila and Liposcelisentomophila (Psocop. Liposcelididae) of two insecticides. JEN 128, 292–297.
Cook, B.J., Nelson, D.R., Hipps, P., 1969. Esterases and phosphatases in the gastricsecretion of the crockroach, Periplaneta americana. J. Insect. Physiol. 15, 581–589.
Crow, J.A., Borazjani, A., Potter, P.M., Ross, M.K., 2007. Hydrolysis of pyrethroids by humanand rat tissues: examination of intestinal, liver and serum carboxylesterases. Toxicol.Appl. Pharmacol. 221, 1–12.
Drake, H.L., Horn, M.A., 2007. As the worm turns: the earthworm gut as a transienthabitat for soil microbial biomes. Annu. Rev. Microbiol. 61, 169–189.
Geering, K., Freyvogel, T.A., 1974. The distribution of acetylcholine and unspecificesterases in the midgut of female Aedes aegypti L. Comp. Biochem. Physiol., B 49,775–784.
Gilham, D., Lehner, R., 2005. Techniques to measure lipase and esterase activity in vitro.Methods 36, 139–147.
Gomori, G., 1953. Human esterases. J. Lab. Clin. Med. 42, 445–453.Haites, N., Don, M., Masters, C.J., 1972. Heterogeneity and molecular weight
interrelationships of the esterase isozymes of several invertebrate species. Comp.Biochem. Physiol., B 42, 303–332.
Hatanaka, Y., Inoue, Y., Murata, K., Kimura, A., 1989. Isolation and characterization ofcarboxylesterase from vinyl acetate-assimilating bacterium isolated from soil.J. Ferment. Bioeng. 67, 14–19.
Hickman, Z.A., Reid, B.J., 2008. Earthworm assisted bioremediation of organiccontaminants. Environ. Int. 34, 1072–1081.
Johnson, J.A., Wallace, K.B., 1987. Species-related differences in the inhibition of brainacetylcholinesterase by paraoxon and malaoxon. Toxicol. Appl. Pharmacol. 88,234–241.
Krauss, M., Wilcke, W., Zech, W., 2000. Availability of polycyclic aromatic hydrocarbons(PAHs) and polychlorinated biphenyls (PCBs) to earthworms in soils. Environ. Sci.Technol. 34, 4335–4340.
Krysan, J.L., Guss, P.L., 1971. Paraoxon inhibition of an insect egg lipase. Biochim.Biophys. Acta 239, 349–352.
Laguerre, C., Sanchez-Hernandez, J.C., Köhler, H.R., Triebskorn, R., Capowiez, Y., Rault, M.,Mazzia, C., 2009. B-type esterases in the snail Xeropicta derbentina: an enzymologicalanalysis to evaluate their use asbiomarkers of pesticide exposure. Environ. Pollut. 157,199–207.
Markwell, M.A.K., Haas, S.M., Bieber, L.L., Tolbert, N.E., 1978. Modification of Lowryprocedure to simplify protein determination in membrane and lippoproteinsamples. Anal. Biochem. 87, 206–210.
Maxwell, D.M., 1992. The specificity of carboxylesterase protection against the toxicityof organophosphorus compounds. Toxicol. Appl. Pharmacol. 114, 306–312.
Mommsen, T.P., 1978. Digestive enzymes of a spider (Tegenaria atrica Koch) — III.Esterases, phosphatases, nucleases. Comp. Biochem. Physiol., A 60, 377–382.
Motulsky, h.J., Christopoulos, A., 2003. Fitting Models to Biological Data using Linearand Nonlinear Regression. A Practical Guide to Curve Fitting. GraphPad SoftwareInc, San Diego, CA, U.S.A. 351 pp.
Nozaki, M., Miura, C., Tozawa, Y., Miura, T., 2009. The contribution of endogenouscellulase to the cellulose digestion in the gut of earthworm (Pheretima hilgendorfi:Megascolecidae). Soil Biol. Biochem. 41, 762–769.
Øien, N., Stenersen, J., 1984. Esterase of earthworms — II. Electrophoresis reveals thatEisenia fetida (Savigny) is two species. Comp. Biochem. Physiol., C 78, 277–282.
Prento, P., 1987. Distribution of 20 enzymes in the midgut region of the earthworm,Lumbricus terrestris L., with particular emphasis on the physiological role of thechloragog tissue. Comp. Biochem. Physiol., A 87, 135–142.
Sanchez-Hernandez, J.C., Wheelock, C.E., 2009. Tissue distribution, isozyme abundanceand sensitivity to chlorpyrifos-oxon of carboxylesterases in the earthworm Lumbricusterrestris. Environ. Pollut. 157, 264–272.
Satoh, T., Hosokawa, M., 1998. The mammalian carboxylesterases: from molecules tofunctions. Annu. Rev. Pharmacol. Toxicol. 38, 257–288.
Sogorb, M.A., Vilanova, E., 2002. Enzymes involved in the detoxification of organo-phosphorus, carbamate and pyrethroid insecticides through hydrolysis. Toxicol.Lett. 128, 215–228.
Spurgeon, D.J., Hopkin, S.P., 1999. Seasonal variation in the abundance, biomass andbiodiversity of earthworms in soils contaminated with metal emissions from aprimary smelting works. J. Appl. Ecol. 36, 173–183.
Spurgeon, D.J., Hopkin, S.P., 2000. The development of genetically inherited resistanceto zinc in laboratory-selected generations of the earthworm Eisenia fetida. Environ.Pollut. 109, 193–201.
Stenersen, J., 2004. Chemical pesticides: mode of action and toxicology. CRC Press, BocaRaton, Fl, U.S.A. 276 pp.
Sutherland, T.D., Horne, I., Weir, K.M., Coppin, C.W., Williams, M.R., Selleck, M., Russell,R.J., Oakeshott, J.G., 2004. Enzymatic bioremediation: from enzyme discovery toapplications. Clin. Exp. Pharmacol. Physiol. 31, 817–821.
Tillinghast, E.K., O'Donnell, R., Eves, D., Calvert, E., Taylor, J., 2001. Water-solubleluminal contents of the gut of the earthworm Lumbricus terrestris L. and theirphysiological significance. Comp. Biochem. Physiol., A 129, 345–353.
Turunen, S., Chippendale, G.M., 1977. Esterase and lipase activity in the midgut ofDiatraea grandiosella: digestive function and distribution. Insect Biochem. 7, 67–71.
Van Straalen, N.M., Van Gestel, C.A.M., 1998. Soil invertebrates and micro-organisms.In: Calow, P. (Ed.), Handbook of Ecotoxicology. Blackwell Science Ltd, Oxford, U.K.,pp. 251–277.
Vijver, M.G., van Gestel, C.A.M., Lanno, R.P., van Straalen, N.M., Peijnenburg, W.J.G.M.,2004. Internal metal sequestration and its ecotoxicological relevance: a review.Environ. Sci. Technol. 38, 4705–4712.
Wheelock, C.E., Shan, G., Ottea, J., 2005. Overview of carboxylesterases and their role inthe metabolism of insecticides. J. Pestic. Sci. 30, 75–83.
Yu, L.Y., Wu, X.M., Li, S.N., Fang, H., Zhan, H.Y., Yu, J.Q., 2006. An exploration of therelationship betweenadsorption andbioavailability of pesticides in soil to earthworm.Environ. Pollut. 41, 428–433.
Zhang, J., Lan,W., Qiao, C., Jiang, H., 2004. Bioremediation of organophosphorus pesticidesby surface-expressed carboxylesterase frommosquito on Escherichia coli. Biotechnol.Prog. 20, 1567–1571.




















