
Bryozoan-dominated benthos of Otago shelf, New Zealand: its associated fauna, environmental setting...
20
This article was downloaded by: [University of Otago] On: 20 April 2013, At: 15:45 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of the Royal Society of New Zealand Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tnzr20 Bryozoan-dominated benthos of Otago shelf, New Zealand: its associated fauna, environmental setting and anthropogenic threats ACL Wood a & PK Probert a a Department of Marine Science, University of Otago, Dunedin, New Zealand Version of record first published: 20 Apr 2013. To cite this article: ACL Wood & PK Probert (2013): Bryozoan-dominated benthos of Otago shelf, New Zealand: its associated fauna, environmental setting and anthropogenic threats, Journal of the Royal Society of New Zealand, DOI:10.1080/03036758.2012.756819 To link to this article: http://dx.doi.org/10.1080/03036758.2012.756819 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and- conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of Bryozoan-dominated benthos of Otago shelf, New Zealand: its associated fauna, environmental setting...

This article was downloaded by: [University of Otago]On: 20 April 2013, At: 15:45Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of the Royal Society of NewZealandPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnzr20
Bryozoan-dominated benthos of Otagoshelf, New Zealand: its associatedfauna, environmental setting andanthropogenic threatsACL Wood a & PK Probert aa Department of Marine Science, University of Otago, Dunedin,New ZealandVersion of record first published: 20 Apr 2013.
To cite this article: ACL Wood & PK Probert (2013): Bryozoan-dominated benthos of Otago shelf,New Zealand: its associated fauna, environmental setting and anthropogenic threats, Journal of theRoyal Society of New Zealand, DOI:10.1080/03036758.2012.756819
To link to this article: http://dx.doi.org/10.1080/03036758.2012.756819
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

REVIEW ARTICLE
Bryozoan-dominated benthos of Otago shelf, New Zealand: its associated fauna,
environmental setting and anthropogenic threats
ACL Wood* and PK Probert
Department of Marine Science, University of Otago, Dunedin, New Zealand
(Received 24 September 2012; accepted 2 December 2012)
The epibenthos of Otago shelf (south-eastern New Zealand, 458S, 1708E) is locally dominated bybryozoans that generate habitat both as living colonies and after death as sediment. Differencesin physico-chemical conditions across Otago shelf relate to the properties of different watermasses and to shore-parallel sediment facies. From 70 m water depth to the shelf break, large,heavily calcified bryozoans are abundant, and dominate an area of about 110 km2 on the mid-shelf, providing habitat at a range of scales. The presence of these bryozoans indicates a stablesubstratum, low levels of sediment input, relatively high mean water flow, and productive waters.Evidence indicates that bryozoans, particularly on the outer-shelf, are vulnerable to disturbancefrom bottom fishing. A better understanding of how bryozoans function as habitat is necessaryto predict effects of fishing impacts on these assemblages and we suggest ways this understandingmight be attained.
Keywords: Bryozoa; sediment; macroinvertebrate; habitat; fishing impact; biogenic; temperate;shelf; New Zealand
Introduction
Biologically generated habitats include seagrass
meadows (Bostrom et al. 2006), maerl beds
(Kamenos et al. 2004), oyster (Beck et al. 2011),
polychaete (Smith et al. 2005; Desroy et al.
2011) and coral reefs (Abele & Patton 1976;
Buhl-Mortensen & Mortensen 2005), and areas
dominated by large, heavily calcified bryozoans
(Wood et al. 2012). Most of these habitats are
relatively well understood ecologically, but
the few studies describing bryozoan-generated
habitats remain fairly site-specific, and an
overarching understanding of habitat-forming
bryozoan ecology is lacking.Bryozoans have generated reef-like habi-
tats since at least the Middle Ordovician, but
since the end of the Palaeozoic their ecologi-
cal importance in the tropics has decreased
(Taylor & Allison 1998). Today they occur at
continental shelf depths throughout temperate
and Antarctic zones, being particularly abun-
dant in New Zealand waters (Wood et al.
2012). Calcified bryozoan colonies ]5 cm in
three dimensions are considered habitat-forming
with respect to macrofauna when they dom-
inate areas of seabed greater than a few square
metres (Wood et al. 2012). Such habitats fre-
quently have a diverse associated fauna, ranging
in scale from small organisms living directly on
colonies (Stebbing 1971; McKinney & Jaklin
2000; Cocito et al. 2002) to more broadly asso-
ciated organisms (Fig. 1) living around bryozo-
an patches (e.g. Jones & Lockhart 2011), often
in concert with other habitat-forming organ-
isms (e.g. sponges, Gutt & Starmans 1998).
Habitat-forming bryozoans in New Zealand
are associated with commercial fishing grounds
for finfish (Vooren 1975; Jiang & Carbines
*Corresponding author. Email: [email protected]
Journal of the Royal Society of New Zealand, 2013http://dx.doi.org/10.1080/03036758.2012.756819
# 2013 The Royal Society of New Zealand
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

2002; Carbines et al. 2004) and shellfish
(Cryer et al. 2000; Michael & Cranfield 2001;
Cranfield et al. 2003). Like other biogenic
structures, habitat-forming bryozoans are vul-
nerable to anthropogenic impacts such as
pollution (Morgado & Tanaka 2001; Harmelin
& Capo 2002), climate warming and ocean
acidification (Rodolfo-Metalpa et al. 2010),
and physical impacts such as bottom fishing
(Saxton 1980; Cranfield et al. 1999; Jones &
Lockhart 2011). The latter is a particular
problem in New Zealand waters (Wood et al.
2012).This case study synthesizes our understand-
ing of one of the most intensively studied areas
of habitat-forming bryozoans: Otago shelf,
south-eastern New Zealand. We identify:
1) environmental conditions that allow bryozo-
ans to dominate the benthos; 2) distribution
and diversity of fauna associated with the
bryozoan-generated habitat at two scales;
3) effects of fishing at this site; and 4) future
research to support successful management.
Species names have been updated where neces-
sary (Appeltans et al. 2012).
Site overview
Otago shelf is a temperate, tectonically stable
area (Bradford-Grieve et al. 2006) on South
Island’s east coast (45853?S, 170848?E, Fig. 2).
The shelf is about 30 km wide, but is locally
narrowed by the promontory of Otago Peninsula,
and submarine canyons (Fig. 3) that incise the
shelf, reducing its width to as little as 10 km in
places. The generally smooth shelf gradient is
occasionally interrupted by rock outcrops and
post-glacial shorelines, the deepest dating from
the peak of the last glaciation c. 18,000�20,000
yr BP, when the shoreline ran along what is
now the shelf break (Carter et al. 1985). Today,
the canyons cause the shelf break to be
shallower (about 105 m) near the canyon heads
(Carter et al. 1985) and deeper (150 m) on the
intervening plateaux (Fig. 3).
Shelf environment
South Island lies across the Subtropical Front
(STF), a circumglobal convergence zone where
warm, saline Subtropical Water (STW) meets
cooler, fresher Subantarctic Water (SAW)
Figure 1 Larger macroinvertebrates such as Ophiocoma/Ophiopteris spp. using bryozoan colonies. Sedimentheterogeneity is visible as sandy substratum to the right of the photo and gravel to the left (photo: E Jones).
2 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013
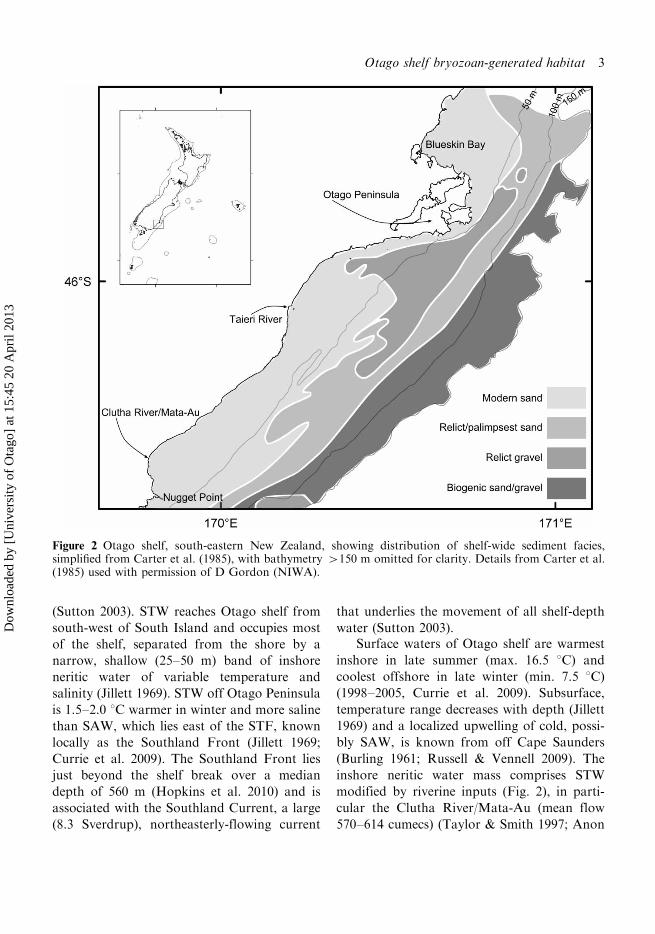
(Sutton 2003). STW reaches Otago shelf from
south-west of South Island and occupies most
of the shelf, separated from the shore by a
narrow, shallow (25�50 m) band of inshore
neritic water of variable temperature and
salinity (Jillett 1969). STW off Otago Peninsula
is 1.5�2.0 8C warmer in winter and more saline
than SAW, which lies east of the STF, known
locally as the Southland Front (Jillett 1969;
Currie et al. 2009). The Southland Front lies
just beyond the shelf break over a median
depth of 560 m (Hopkins et al. 2010) and is
associated with the Southland Current, a large
(8.3 Sverdrup), northeasterly-flowing current
that underlies the movement of all shelf-depth
water (Sutton 2003).Surface waters of Otago shelf are warmest
inshore in late summer (max. 16.5 8C) and
coolest offshore in late winter (min. 7.5 8C)
(1998�2005, Currie et al. 2009). Subsurface,
temperature range decreases with depth (Jillett
1969) and a localized upwelling of cold, possi-
bly SAW, is known from off Cape Saunders
(Burling 1961; Russell & Vennell 2009). The
inshore neritic water mass comprises STW
modified by riverine inputs (Fig. 2), in parti-
cular the Clutha River/Mata-Au (mean flow
570�614 cumecs) (Taylor & Smith 1997; Anon
Figure 2 Otago shelf, south-eastern New Zealand, showing distribution of shelf-wide sediment facies,simplified from Carter et al. (1985), with bathymetry �150 m omitted for clarity. Details from Carter et al.(1985) used with permission of D Gordon (NIWA).
Otago shelf bryozoan-generated habitat 3
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

2005). This river directly affects the salinity of
the inshore neritic water off the peninsula (Jillett
1969; Hawke 1989), which ranges between 33.72
and 34.89 at the surface (Jillett 1969; Croot &
Hunter 1998).The Southland Current conveys relatively
warm water along the coast, providing a buffer
from cold SAW for the temperate shelf assem-
blage, and transporting water-borne food to the
benthos. Most bryozoans are stenohaline
(Ryland 1970), so the relatively constant sali-
nity on the mid- and outer-shelf may be an
important variable describing the distribution
of bryozoan-dominated habitat, while the mix-
ing of subtropical, subantarctic and riverine
water masses has important consequences for
local primary productivity.
Food supply
Bryozoans are suspension feeders (McKinney
& Jackson 1989) and a good supply of phyto-
plankton is an important property of suitable
habitat for bryozoans (Taylor 2000). SAW is
generally rich in macronutrients, whereas STW
is relatively nutrient poor (Butler et al. 1992;
Boyd et al. 1999), and the STF where these
waters mix is known to be highly productive.
On Otago shelf, advection from SAW to STW
(reactive phosphorus (Hawke 1989; Croot &
Hunter 1998)), riverine supply (reactive silica
from the Clutha River/Mata-Au (Hawke 1989,
1992)) and resuspension of bottom sediments
(releasing iron (Croot & Hunter 1998)) provide
nutrients for a spring plankton bloom, which is
most productive on the mid-shelf (Pfannkuche
Figure 3 The continental shelf seaward of Otago Peninsula, including (shaded) the known distribution ofhabitat-forming bryozoans ascertained from the work of Probert et al. (1979), Batson (2000) and Jones(2006).
4 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

1998; Haywood 2004). SAW has a low con-centration of chlorophyll (0.3 mg L�1) year-round (Croot & Hunter 1998), consistent withiron and silica limitation (Boyd et al. 1999). Theblooms are associated with river-borne nutri-ents (Haywood 2004), particularly silica, whichstimulates phytoplankton growth on the inner-and mid-shelf (Hawke 1992; Boyd et al. 1999).Silica limitation may occur during a bloomwhen river flow is low (Hawke 1995), therebyaffecting the biological community of the wholeshelf. The shelf waters are undersaturated inCO2 with respect to the atmosphere (Currie et al.2009), causing CO2 to be consistently drawndown via photosynthesis, particularly in spring(see Zeldis et al. 2010).
Sediment inputs and facies
As suspension feeders, bryozoans are sensitiveto increased suspended sediment loads (Lagaaij& Gautier 1965; Ryland 1970; Best & Thorpe1996). Due to relatively limited terrigenousinputs, Otago shelf is a suitable habitat forbryozoans in this respect (Griffiths & Glasby1985; Hicks & Shankar 2003). Sediments thatreach Otago shelf are supplied mainly by theClutha/Mata-Au and Taieri Rivers (0.39 Mt y�1
and 0.32 Mt y�1, respectively (Hicks & Shankar2003)). (The Clutha delivered about 2.3 Mt y�1
before it was first dammed in the 1950s(Bradford-Grieve et al. 2006)). These modernsediments form a narrow, mobile, sandy facies(Carter et al. 1985) along the coast, although atleast 50% of sediment reaching Otago shelffrom the Clutha is mud (Carter et al. 1985). Themud is transported along the coast duringsoutherly weather, and in calm or northerlyweather is transported on to the mid-shelf,from where it is thought to be carried acrossthe shelf to the canyons (Andrews 1973; Carteret al. 1985). Sediment samples from the mid-shelf comprise 5%�6% mud (519%) (Andrews1973; Batson 2000). In most places the sand/mud facies extends to about 30 m water depth,but shelf sediments are heterogeneous and thedifferent facies overlap significantly (Andrews
1973; Carter et al. 1985). The full depth range ofthe inshore modern terrigenous sand/mud faciesis 0�70 m. Seaward, relict terrigenous gravelwith varying proportions of sand and mudoccurs from 30�80 m, predominantly relictpalimpsest sand from 30�120 m and biogenicsand/gravel from 50�120 m (Andrews 1973).
Gravels of the mid-shelf are mainly smooth,well-rounded quartz and quartzite with a meangrain size of 6 mm, and occasional pebbles upto 100 mm (Carter et al. 1985). The biogeniccomponent comprises molluscan and bryozoanfragments (Andrews 1973). Molluscan materialpredominates in most places, but bryozoan-generated sediments dominate the biogenicgravel fraction at 80�95 m off the peninsula(Batson 2000). The quartz gravel was suppliedby the Clutha River/Mata-Au during thePleistocene, and forms an almost continuousband along the mid-shelf from near the Cluthato just north of Otago Peninsula. Off thepeninsula, the gravel grades into a distinctrelict/palimpsest terrigenous sand facies, whichis coarser and more poorly sorted than themodern inshore sand (Carter et al. 1985). Thebiogenic sand/gravel of the outer-shelf com-prises minute bivalves and foraminifera, withlocally abundant bryozoan and molluscan frag-ments (Carter et al. 1985; Carter & Carter1986). Despite the transport of mud across theshelf, mid- and outer-shelf sediments are fairlyclean, so that suspension feeders, includingbryozoans, are at a low risk from clogging byfiner sediment fractions.
Water movement and sediment transport
Sufficient water movement is essential to thesuitability of habitat for bryozoans, but waveenergy is likely to be destructive to bryozoancolonies, particularly those on soft substrataliable to resuspension. On Otago shelf, habitat-forming bryozoans occur in water deeper thanc. 70 m (Batson 2000; Jones 2006), wheredisturbance from storm waves occurs aboutevery 25 years (Carter & Heath 1975). Despitetheir depth, Otago shelf bryozoans are probably
Otago shelf bryozoan-generated habitat 5
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

subject to strong lateral water movement,
comprising mean northeasterly flow (the South-
land Current), on which are superimposed the
effects of tides, weather and ground swell
generated in the Southern Ocean (Carter et al.
1985; Chiswell 1996). Semi-diurnal tides flood
to the north (tidal range 1.5 m [Carter et al.
1985]) and tend to enhance mean flow, as do
the variable effects of predominantly southerly
weather. A surface current speed of 7�15 cm s�1,
increasing to 26 cm s�1 during flood tide
(Carter et al. 1985; Hawke 1992; Chiswell
1996) probably represents a consensus of ‘nor-
mal’ conditions on the shelf. The current speed
increases near Otago Peninsula, where the body
of water is constricted by the projection of the
peninsula, with current speeds up to 100 cm s�1
measured near Cape Saunders (Russell &
Vennell 2009) and speeds up to 1.5 m s�1
possible (P. Russell, University of Otago, pers.
comm. 04/02/2011, see also Hydrographic De-
partment 1958). The near-bottom speed was
estimated as 2�6 cm s�1 (Carter et al. 1985),
but is difficult to measure accurately (P. Russell,
University of Otago, pers. comm. 04/02/2011).
The area of faster moving water caused by the
out-jutting peninsula is considered to have
important consequences for the benthos of the
region (Probert 1977), transporting a constant
supply of particulate organic matter (POM) to
the benthos and generating unfavourable con-
ditions for sediment deposition (Probert et al.
1979). Elsewhere in New Zealand, areas of
habitat-forming bryozoans are associated with
sites where current speed is high, for example in
Foveaux Strait to the south (Cranfield et al.
1999) and to the north of North Island (Cryer
et al. 2000). Bryozoan-dominated habitats
characteristically occur where current speeds
are elevated, or on topographic high points
(Wood et al. 2012). Such locations may increase
the delivery of POM to the bryozoans but also
have the effect of interrupting flow across the
seafloor, which may improve POM supply to
the benthos (Thrush & Dayton 2002).
Faunal assemblages
Broad, persistent species assemblages occur
across Otago shelf in association with the sedi-
ment facies. As for the sediments, the assem-
blages occur in broadly shore-parallel zones,
which overlap in depth range due to the inter-spersed nature of the sediments. An inshore
assemblage (and sub-assemblage associated with
muddier, slightly deeper sites) is restricted to
sandy sediments in water depths B30 m and
characterized by low species richness and con-
spicuous trochid gastropods (Probert et al.
1979; Swanson 1979; Probert & Wilson 1984).On the shallow mid-shelf (40�90 m), the
epifauna includes a mixture of species from
shallower and deeper water (Probert et al.
1979), while a distinct infaunal assemblage is
associated with the gravel (24.1%) and silt-clay
(6.6%) sediments (Probert & Wilson 1984).
Habitat-forming bryozoans are not part ofthese assemblages.
Between 45�130 m water depth, a distinctive
mid-shelf epifauna occurs on gravelly sediments
dominated by the habitat-forming bryozoan
Cinctipora elegans (Probert et al. 1979). Other
characteristic mid-shelf species are the habitat-forming bryozoans Hornera robusta, Hippo-
menella vellicata and other taxa including
Phyllochaetopterus socialis (Polychaeta), Poly-
syncraton mortenseni, Pyura picta (Ascidiacea),
Lophopagurus thompsoni (Paguridae), Crella
incrustans (Demospongiae) and Phellia auck-
landica (Actiniaria) (Probert et al. 1979). Jones(2006) confirmed this assemblage, adding the
large ophiuroids Ophiocoma/Ophiopteris spp.
and Ophiopsammus maculata, while Swanson
(1979) described an assemblage of ostracods
from the mid-shelf (40�95 m).The sandier outer-shelf sediments (from
95 m to the shelf break) also support character-istic assemblages (Probert et al. 1979; Swanson
1979; Jones 2006). Habitat-forming bryozoans
include Adeonellopsis spp., Celleporina grandis,
Hippomenella vellicata and Cellaria immersa
(Probert et al. 1979; Jones 2006), with other
characteristic epifauna including Sclerasterias
6 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

mollis (Asteroidea), Leptomithrax longipes,
Pylopagurus stewarti (Decapoda), Didemnum
spp. (Ascidiacea), Bunodactis chrysobathys
(Actiniaria) and Ophiopeza cylindrica (Ophiur-
oidea). Numerous mid-shelf species also occur
at these deeper sites, although often less fre-
quently than in shallower water (Probert et al.
1979; Jones 2006). Two infaunal assemblages
occur on the outer-shelf, but all species were
also found at shallower sites (Probert & Wilson
1984).
Habitat-forming bryozoans: distribution
overview
Large bryozoans are a ‘striking feature’ of the
mid- to outer-shelf benthos (Probert et al. 1979)
and two assemblages are apparent. Cinctipora
elegans dominates the mid-shelf (80�95 m),
with Hornera robusta, Celleporaria agglutinans
and smaller amounts of Hornera foliacea,
Cellaria immersa, Hippomenella vellicata and
Celleporina grandis; none of these species oc-
curs locally shallower than about 70 m (Probert
et al. 1979; Batson 2000; Jones 2006). On the
outer-shelf, Hippomenella vellicata, Cellaria
immersa and Celleporina grandis dominate
(Probert et al. 1979), but occur less frequently
than does Cinctipora elegans on the mid-
shelf. Cinctipora elegans does occur on the
outer-shelf, but rarely dominates (Probert et al.
1979). Celleporaria agglutinans also occurs on
the outer-shelf (Probert et al. 1979; Batson
2000; Jones 2006). Beyond the shelf break,
bryozoans that form habitat in shallower water
(Celleporaria agglutinans, Cellaria immersa,
Cellaria tenuirostris, Hippomenella vellicata and
Cinctipora elegans) occur in much lower abun-
dance than on the shelf (Probert et al. 1979;
Carter et al. 1985). On the northern side of
Blueskin Bay, Loutit (1975) reported only relict
bryozoans at these deeper sites. The species
distributions are considered to be stable over
periods of c. 30 years, given similarities between
reported assemblages (e.g. Probert et al. 1979;
Batson 2000; Jones 2006).
Along-shelf, the zone of bryozoan domi-nance is limited to roughly parallel with OtagoPeninsula. Habitat-forming bryozoans extendto the north side of Blueskin Bay (Fig. 3),including Celleporina grandis, Adeonellopsisspp., Cellaria immersa, Galeopsis polyporusand Tetrocycloecia spp. (Loutit 1975; Probertet al. 1979), although the bryozoans here arenot dominant (Batson 2000). South of SaundersCanyon (Fig. 3), habitat-forming bryozoansoccur at 70�120 m water depth (Batson 2000;Jones 2006), although the number of speciesand their abundance are lower, and speciesoccur over a wider depth range than at sites tothe north (Batson 2000).
Although the habitat-forming bryozoanson Otago shelf occur on the mid- and outer-shelf (Probert et al. 1979), the same speciesoccur elsewhere in shallower waters: Cinctiporaelegans at 12 m (Taylor et al. 2004) andAdeonellopsis sp. at 10�25 m (Smith et al.2001) on fiord walls in southwestern NewZealand. Celleporaria agglutinans occurs at20�40 m in the north of South Island at a sitewhere current speeds are relatively high (0.36m s�1) but wave energy is low (Grange et al.2003). These distributions, combined with thoseof the different sediment facies, suggest that thekey characteristics of substratum type arerelative stability and heterogeneity. On Otagoshelf, coarse gravels, living and dead molluscshells and bryozoans, and the chitinous-tubedwelling worm, Phyllochaetopterus socialis allprovide suitable, stable substrata for coloni-zation, even where sediments are sandier.Phyllochaetopterus socialis is particularly abun-dant on patches of fine sediment and mayfacilitate bryozoan colonization of sedimentson Otago shelf (Batson 2000). There are atpresent no studies of biologically mediatedsettlement/recovery on Otago shelf, but inFoveaux Strait (southern New Zealand), thestabilization of sediments by byssally attachedmolluscs is thought to enable other fauna, in-cluding habitat-forming bryozoans, to colonizeand eventually form epifaunal reefs (Cranfieldet al. 2004).
Otago shelf bryozoan-generated habitat 7
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

Habitat-forming bryozoans: species distributions
Habitat generated by bryozoans on Otago shelf
comprises patchy ‘thickets’, with up to six
species known from any one dredge sample,
and up to 16 species of habitat-forming bryozo-
ans known in total (Batson 2000; Batson &Probert 2000). The distributions of the eight
most common species are described here, for
transects centred around the narrowest part of
Otago shelf, spanning water depths of 60�110
m (Batson 2000; Jones 2006). For samples
taken in the 2000s, live habitat-forming bryozo-
ans were less common at 95�110 m water depthon the outer-shelf (Batson 2000; Jones 2006)
than on the mid-shelf, which differs somewhat
from samples taken in 1966�73 (Probert et al.
1979), in which Celleporaria agglutinans,
Cellaria immersa, Cellaria tenuirostris and
Cinctipora elegans occurred as frequently on
the outer-shelf as on the mid-shelf (althoughCinctipora elegans was noted as much less
abundant on the outer-shelf (Probert et al.
1979)). Both Batson (2000) and Jones (2006)
found an area where habitat-forming bryozo-
ans were unexpectedly rare or absent between
70�80 m in the north (described as southeastand east-southeast of Taiaroa Head), and a
band of high abundance along the shelf be-
tween 80�90 m. There, mean cover of bryozo-
ans was 19% on the central transect (sampled
by photography). Across the study area (all
depths combined) bryozoans attained 4.0%
mean cover (n�497 photographs) (Jones 2006).Cinctipora elegans dominates the benthos
between 70�90 m (Batson 2000). It is most
abundant in the north (Batson 2000) and
middle of the study area, and attained 56.4%
cover at one site, although mean cover across
all depths was 2.9% (Jones 2006). In the south,
Cinctipora elegans was restricted to 80 m waterdepth (Batson 2000; Jones 2006). Small colo-
nies of this species were characteristic of deeper
water (Jones 2006) and larger colonies occurred
on the mid-shelf (Batson 2000; Jones 2006).Hornera robusta (mean 0.32%, maximum
6.25% cover) and Adeonellopsis spp. (mean
0.30%, maximum 9.82% cover) were similarlyabundant, although Hornera robusta was mostabundant around 80 m in the north, andAdeonellopsis spp. were more abundant atdeeper stations to the south (Jones 2006).
Celleporaria agglutinans, Hornera foliaceaand Celleporina grandis were similarly abun-dant, each covering on average 0.05%�0.06%of the seafloor (Jones 2006). The abundance ofCelleporaria agglutinans peaked in the middleof the study area between 80�100 m. Jones(2006) found Hornera foliacea to be irregularlydistributed and absent from many sites, whileBatson (2000) considered it reasonably com-mon. Jones (2006) found that Celleporinagrandis was most abundant at 80 m in thesouth and at deeper stations in the north (90�100 m), while Batson (2000) found it to becommon or abundant at sites where the waterdepth was ]100 m.
Hippomenella vellicata was the second leastabundant habitat-forming bryozoan (0.03%cover (Jones 2006)), occurring more frequentlyin the north at 85�100 m, and at 80�90 m in thesouth (Batson 2000; Jones 2006). Least abun-dant was Cellaria immersa (mean 0.02% cover(Jones 2006)), which was most abundant at 100m in the north, but occurred in shallower waterin the south. This species is more usuallyassociated with sandy substrata (e.g. Nelsonet al. 1988a), so that more recent studies mighthave missed some of its potential range on theoutermost shelf. Probert et al. (1979) identifiedCellaria immersa as belonging to outer-shelf/upper canyon assemblages, although it wasfound at �75% of sites on the mid- andouter-shelf.
Overall, a number of environmental vari-ables combine to allow bryozoans to dominateareas of seafloor and to generate habitat formany other taxa. The Otago mid-shelf isrelatively stable in terms of temperature andsalinity, and the relative warmth of the STWmay enable warm- and cool-temperate speciesto coexist, thereby supporting the diversity ofhabitat-forming bryozoans on Otago shelf. Theproductivity associated with the mixing of
8 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

SAW, STW and riverine inputs serves tosupport the diverse benthic assemblage, butthere is no evidence that productivity nearOtago Peninsula is higher than to the northor south, away from where bryozoans arefound in high abundance. The SouthlandCurrent and the band of mid-shelf gravel withwhich the habitat-forming bryozoans are asso-ciated occur along much of Otago shelf, so ifthese two variables alone support the abundantbenthos, the bryozoans should occur all alongthe shelf. Available evidence supports thesuggestion of Probert et al. (1979), that theincreased current speed caused by constrictionof shelf waters by Otago Peninsula is sufficientto increase the rate of food supply to the ben-thos and simultaneously to minimize sedimen-tation, and that these characteristics underpinthe location of the Otago shelf habitat-formingbryozoans.
Sediment heterogeneity
The modern carbonate deposits of Otago shelfare relatively small (1500 km2 (Nelson et al.1988b)) and thin, being only a few centimetresdeep (Batson 2000), reflecting the short timesince the post-glacial transgression (Carter et al.1985). Relict molluscs dated at 10,000�12,000yrs BP collected in surficial sediments from themid-outer-shelf (Carter et al. 1985) indicate thelow sedimentation rate over the subsequentperiod. Today, bryozoan growth within the80�100 m depth range probably accounts forthe majority of carbonate production on Otagoshelf (Batson 2000), with minimum growth ratesranging from 0.8 mm y�1 (Celleporina grandis)to 5.0 mm y�1 (Hippomenella vellicata);Cinctiporaelegans grows a minimum of 2.0 mm y�1 (Jamet1999).
The general distribution of colony frag-ments in the sediment reflects the distributionof living bryozoans. In shallow water there arefew bryozoan fragments (Batson 2000), andbiogenic material mainly comprises abundant livegastropods and foraminifera (Andrews 1973).Seaward of 65 m water depth, the abundance
of bryozoan fragments in the gravel fraction
increases, being most abundant at 80�100 m,
where Cinctipora elegans comprises up to 22%
(by weight) of the gravel fraction, Hornera
robusta 2.6% and Hornera foliacea 0.4% (Bat-
son 2000). In deeper water (90�110 m), moll-
uscan fragments dominate, but Celleporina
grandis and Celleporaria agglutinans together
comprise 4.5% of the gravel fraction.
Adeonellopsis spp. fragments are fairly evenly
distributed on the mid- and outer-shelf, com-
prising 1.4%�1.5% of the gravel fraction at 80�120 m. Degraded fragments of Hippomenella
vellicata are rare (mean 0.15% at 80�95 m),
probably because of the way this species breaks
down (Batson 2000). Degraded bryozoan frag-
ments are scattered over a greater depth range
than live colonies, particularly near Cape
Saunders, where live colonies are abundant but
fragments less so, indicating that strong water
movement has removed fragments (Batson
2000).In addition to generating sediment, bryozo-
ans on Otago shelf interact with the environ-
ment to affect the distribution of existing
sediments. Within individual colonies fine
sediments build up, apparently resulting from
the baffling effect of colonies on water-borne
sediments (Wood 2005), and based on the
composition of associated macrofaunal assem-
blages, these sediments may provide habitat
for deposit feeders in an environment where
concentrations of fine sediments are otherwise
unusual (Wood 2005). At centimetre-scales,
sediment type varies around habitat-forming
colonies (Fig. 1), with patches of coarse gravel
alongside patches of clean sand, perhaps in the
lee of bryozoan colonies (Jones 2006). At
scales �10 m, large quantities of sediment
can accumulate. Cranfield et al. (2003) docu-
mented the release of millions of cubic metres
of biogenic sediment when epifaunal reefs
comprising (among other species) Cinctipora
elegans in Foveaux Strait were broken up by
dredging for oysters.
Otago shelf bryozoan-generated habitat 9
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

Bryozoans as habitat
Probert et al. (1979) noticed large bryozoans
on Otago shelf provided habitat for ‘many
small species’, while photographs of the sea-floor show larger invertebrates sheltering
around or on bryozoan colonies (Fig. 1) (Jones
2006). The distribution of macroinvertebratesvaries in relation to the different habitat-
forming bryozoan colony forms (Wood 2005).
For example, the bushy colonies of rough,cylindrical, bifurcating branches that comprise
Cinctipora elegans (Fig. 4) provide attachment
points and physical protection for ascidianswith soft tests (Corella eumyota), while
Hippomenella vellicata has a coarse foliaceous
‘honeycomb’ growth form that provides habitatfor mobile species (e.g. munidid squat lobsters)
able to use the large sheltered spaces this
bryozoan creates. The encrusting multilaminarcolonies of Celleporaria agglutinans often grow
over or around other structures and form
tubular ‘chimneys’ inside which accumulatefine sediments with which deposit feeders
(e.g. flabelligerid polychaetes) are associated.
Further, growth forms can change with colonysize: for example, the bifurcating branches of
Cinctipora elegans split at increasingly acute
angles as the colony grows, so that branchdensity increases with colony size, changing
the nature of the bryozoan-generated habitat
(Batson 2000). Overall, 138 non-colonial puta-tive species of macroinvertebrates are known
from these three habitat-forming species (Wood
2005). Mean species richness was found toincrease with colony size, and although distinct
assemblages of non-colonial organisms oc-
curred, most epifaunal species occurred on allhabitat-forming bryozoan species. The assem-
blages therefore comprised different abun-
dances of many of the same species. Thus theassemblage associated with Cinctipora elegans
had a greater abundance of nestling species
such as Amphipholis squamata (Ophiuroidea),Gregariella barbata, Neolepton antipodum
(Bivalvia) and Corella eumyota, while the
assemblage associated with Hippomenella
Figure 4 Different growth forms of three habitat-forming species. A, Cinctipora elegans. B,Hippomenellavellicata. C, Celleporaria agglutinans (photos: AWood).
10 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

vellicata and Celleporaria agglutinans includedspionid and phyllodocid polychaetes, Nemertea,an amphipod and a tanaidacean. Hippomenellavellicata also had greater abundance of Muni-didae sp. (Decapoda), Terebellidae sp. andHesionidae sp. (Polychaeta), while Kelliacycladiformis, Talochlamys sp., Cardita distorta(Bivalvia), P. socialis, a sipunculan, and flabel-ligerid polychaetes were more abundant oncolonies of Celleporaria agglutinans. Many ofthe fauna associated with Cinctipora elegans,particularly solitary ascidians, appear to ‘glue’the bryozoan branches together, increasing thestructural integrity of the habitat (Willan 1981;Cranfield et al. 1999; Batson 2000). Such inter-actions may shift the balance between habitatgrowth and (bio)erosion in favour of growth(e.g. Roberts 2005), although fauna are alsoresponsible for the breakdown of colony struc-ture: boring organisms (e.g. spionid and sabellidpolychaetes) were common inside habitat-forming colonies, particularly Celleporariaagglutinans (Wood 2005).
Thirty-four colonial taxa were identifiedfrom three habitat-forming species, occurringin greatest richness and abundance onHippomenella vellicata (Wood 2005). The mostabundant epifauna on Hippomenella vellicatawere epibiotic bryozoans (mean 1598 coloniesper colony) and serpulids (mean 164 individualsper colony). A separate study identified 1292epibiotic bryozoan colonies representing 53species on one colony of Hippomenella vellicata(Junge 1998). Epibiotic bryozoans were mostabundant in the middle and proximal parts ofcolonies. Sponges and colonial ascidians oc-curred mainly in the proximal parts, andhydroids more commonly in the distal parts(Wood 2005).
The varied particle sizes and shapes thatbryozoans form after death contribute to thehabitat complexity of the sediments. The het-erogeneous, poorly sorted facies of the mid-and outer-shelf were noted by Probert &Wilson (1984) as a possible explanation forthe species-rich infaunal assemblages of Otagoshelf, given that poorly sorted sediments tend
to support richer faunal assemblages (Gray
1974). The richness of ostracod assemblages
on Otago shelf was also positively correlated
with sediment coarseness and heterogeneity
(Swanson 1979), while for the New Zealand
region, Rowden et al. (2004) found a correla-
tion between heterogeneous bryozoan/mollus-
can-derived sediments and the diversity of all
bryozoans (see also Gappa 2000).In addition, an abundance of suspension
feeders on the seafloor may result in significant
energy transfer from the pelagic to the benthic
system (Batson 2000), thus altering the quality
of habitat for infauna in terms of food avail-
ability. Such a transfer may be detectable as
organic enrichment of sediments under the
bryozoans, perhaps generating decimetre-scale
heterogeneity in food availability, although at
the shelf scale, Probert & Wilson (1984) found
no particular infaunal assemblage occurred in
the zone of high bryozoan abundance.
Human impacts and conservation
Wood et al. (2012) found the most common
anthropogenic threats to habitat-forming
bryozoans globally were bottom fishing and
pollution, while others have shown the effects
of climate change (including ocean acidifica-
tion) are likely to affect bryozoan growth and
sedimentation, both for species present on
Otago shelf and elsewhere (Smith 2009; Rodolfo-
Metalpa et al. 2010). The most tangible anthro-
pogenic impacts on the temperate shelf in New
Zealand are sedimentation caused by changing
land use (Foster & Carter 1997; Bradford-Grieve
et al. 2006) and disturbance caused by fishing
gear (Jones 1992; Baird et al. 2011). On Otago
shelf, terrigenous sediments are transported
along the coast and seem, under present-day
conditions, unlikely to threaten the habitat-
forming bryozoans. The history of fishing on
Otago shelf is poorly known, but its possible
impacts on the benthos are likely to be numerous
and wide ranging (Jones 1992; Watling & Norse
1998; Auster & Langton 1999).
Otago shelf bryozoan-generated habitat 11
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

Limited fishing occurred on the mid- andouter-shelf from the nineteenth century untilthe 1950�60s (Saxton 1980). From the 1960sonwards, parts of the Otago shelf bryozoan-dominated area were trawled for various de-mersal fish species using nets with attachedfloats, intended to reduce the amount ofbenthic bycatch, thereby allowing densebryozoan areas to be fished more easily (Batson& Probert 2000). However, Saxton (1980) notedthat fishers of the Tasman Bay bryozoangrounds (northern South Island), who adoptedsimilar practices, found it impossible to fishover the bryozoans without causing damage,regardless of the arrangement of floats. Fishingintensity on the mid- and outer-Otago shelfvaried, as many fishers also worked the inshoregrounds for flatfish (usually at B50 m depth),as they do today (Batson & Probert 2000).Large quantities of bryozoans were occasion-ally taken, usually when fishing occurred be-tween 70�100 m depth, indicated by thedifficulty fishers sometimes had in landing thetrawl because of the weight of bryozoans(Batson & Probert 2000). During the 1960�70s, large foreign or joint venture vessels usingtrawls with steel bobbins fished the general areaof the bryozoan grounds. In addition tocommercial fishing, collections for researchpurposes have not been insignificant (Batson& Probert 2000). A voluntary ‘closed area’covering the zone of high bryozoan abundanceon the mid-shelf was agreed in the early 2000s,but the agreement is not publicly listed by theNew Zealand Ministry for Primary Industries(formerly the Ministry of Fisheries) and fishingactivities are not monitored at a sufficiently finescale that the effectiveness of the agreement canbe assessed.
The queen scallop Zygochlamys delicatula isabundant on the outer-shelf (Powell 1950;Probert et al. 1979) and has supported a small(380 tonne total allowable commercial catch)fishery since the late 1980s (Ministry of Fish-eries 2007). Although an association betweenhabitat-forming bryozoans and queen scallopshas been noted (Probert et al. 1979; Michael &
Cranfield 2001), most fishing occurs deeperthan the zone of high bryozoan abundance, atdepths of 110�200 m (Batson & Probert 2000;Jiang et al. 2004). Bryozoans have, however,been common at these depths in the recent past(Probert et al. 1979; Carter et al. 1985; Batson& Probert 2000). In the early days of thefishery, bryozoans dominated the bycatch(Michael & Cranfield 2001). Scallop fisherswere familiar with habitat-forming bryozoansand reported that significant quantities wereoccasionally caught on the outer-shelf, particu-larly south of Hoopers Canyon (Fig. 3) at 90�100 m depth (Batson & Probert 2000).
In a stock assessment for the Z. delicatulafishery, the outer-Otago shelf and upper slopewere surveyed at depths of 130�200 m usingindustry-standard fishing gear (Jiang et al.2004). The survey extended from Nugget Point(Fig. 2) to north of Karitane Canyon (total area740 km2). Sampling included an assessment of‘bycatch’ which comprised 94 putative species,of which 60 were invertebrates. Of 24.7 tonnestotal catch, 13.9 t was bycatch and 10.8 t ofthat comprised invertebrates. Potentially habitat-forming species comprised 63% (mass) of theinvertebrate bycatch (45% sponge, 10% soli-tary ascidian, 8% bryozoan). Eleven inverte-brate taxa were caught in quantities �100 kg,including ‘bryozoan general’, Hippomenellavellicata and Celleporina grandis; Cinctiporaelegans was collected only in small quantities,a total of 3.5 kg (0.03%).
Bycatch was from the total area surveyed,which included some ‘exploratory’ sites and insome areas scallop density was low and fishingwould be economically unviable. Even so,many species would be removed or damagedby scallop fishing, including habitat-formingbryozoans. For example, in the area borderingHoopers Canyon where Jiang et al. (2004)calculated commercial fishing was viable, habi-tat-forming bryozoans were fished at a rate ofc. 75 kg per half-hour of fishing (‘bryozoansgeneral’ 40.1 kg, Hippomenella vellicata 3.7 kg,Celleporina grandis 30.4 kg per half-hourtrawl).
12 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

Although there is no direct evidence thatbenthic habitat complexity has been reduced(the problem of the shifting baseline (e.g. Pauly1995)), Batson & Probert (2000) considered it‘reasonable to assume that fishing off OtagoPeninsula has adversely affected the bryozoangrounds’. Hippomenella vellicata and Celleporinagrandis, for example, were recorded as locallydominant species (Probert et al. 1979; Carter et al.1985) in depths targeted by scallop fishers onOtago shelf. With a (minimum) growth rate of5 mm y�1 (Jamet 1999), Hippomenella vellicatawould take c. 30 years to form colonies of the sizecollected on the mid-shelf by Wood (2005), whileCelleporina grandis (minimum growth rate 0.8mm y�1 (Jamet 1999)) could take more than 60years to reach 50 mm, the size defined as ‘large’ byBatson (2000) and considered as ‘habitat-forming’by Wood et al. (2012). Zygochlamys delicatulareaches legal size in about eight years (Michael &Cranfield 2001), meaning scallop fishing is likelyto occur more frequently than would allowpopulations of Hippomenella vellicata andCelleporina grandis to recover. In addition tothe habitat they generate, the habitat-formingbryozoans themselves are an important part ofNew Zealand’s biodiversity: at least 30 of NewZealand’s 45 known habitat-forming bryozoansare endemic, including Cinctipora elegans,Celleporaria agglutinans, Celleporina grandis,Adeonellopsis spp. and Hippomenella vellicata(Gordon et al. 2009).
In other parts of New Zealand, commercialfishing has variously destroyed whole or largeareas of bryozoan beds (Saxton 1980; Cran-field et al. 1999) or has been mitigated by areaclosures (Mace 1981; Cryer et al. 2000; Froude& Smith 2004). Foveaux Strait and Otagoshelf, the southernmost examples in NewZealand of this globally rare habitat type,remain both unprotected and largely unmoni-tored, despite calls to the contrary in the caseof Foveaux Strait (Carbines et al. 2004; Cran-field et al. 2004; Rowden et al. 2004). Studiesthat have reasoned or recorded that areas ofbiogenic habitat are avoided by fishers (Batson2000; Jiang et al. 2004; Hill et al. 2010) fail to
acknowledge the damage caused by ‘explora-tory tows’ and ‘fishing along the edge’ ofcomplex habitat (e.g. Saxton 1980; Hill et al.2010). The time taken for these habitats torecover from large-scale impacts is unknownfor Otago shelf, and is highly variable wheredata are available, ranging from one to morethan five decades in Foveaux Strait (Cranfieldet al. 2004).
Also concerning are the effects of climatechange on calcifying organisms. Bryozoangrowth results in the sequestration of carbon inthe sediment as carbonate, but ocean warmingand reduced seawater pH are likely to affectthis process (Rodolfo-Metalpa et al. 2010). Livebryozoans may continue to grow at reducedpH (Rodolfo-Metalpa et al. 2010), but deadbryozoan material dissolves more quickly(Smith 2009; Rodolfo-Metalpa et al. 2010),buffering pH (Gruber 2011) and decreasingsediment accumulation rates (Smith 2009).Bryozoans in southern New Zealand are likelyto be affected by reduced pH earlier than thosefarther north, because undersaturation of ara-gonite in seawater is predicted to occur therefirst, perhaps only 90 years from now (Gruber2011). Carbon sequestration by living coloniesand subsequent buffering of pH by dead colonyfragments may be an important service per-formed by bryozoans under future climateconditions. The Otago shelf bryozoan bedsrepresent a useful location where the effects ofclimate change on a southern fauna could bemonitored.
Summary
Habitat-forming bryozoans are particularlysuccessful where lateral water movement isfast and wave action is low. The apparentrestriction of bryozoans to water depths greaterthan c. 70 m on Otago shelf is likely to resultfrom destructive wave action and unsuitablesubstrata in shallower waters, rather than anydirect physiological effects of depth, since thesame species occur at shallower, more shelteredsites elsewhere. On Otago shelf, the speed of
Otago shelf bryozoan-generated habitat 13
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

the Southland Current increases around thepromontory of Otago Peninsula, so that POMmay be supplied to the benthos at a higher ratethan elsewhere on the shelf and high levelsof sedimentation are prevented. The relativewarmth of STW coupled with nutrients sup-plied by sedimentary, riverine and SAW inputssupport seasonal productivity in the well-mixedoverlying waters of the shelf, and this produc-tivity is likely to be important to supporting theabundant benthic fauna. Coarse terrigenoussediments of the mid-shelf, together with bio-genic sediments produced in situ (taphonomicfeedback), provide suitably stable substrata forthe settlement and growth of bryozoans andother suspension feeders, in an area where finesediment deposition has been low over ecolo-gical time.
The Otago shelf benthic assemblages occurin broadly shore-parallel zones, as the environ-ment changes across the shelf in terms ofsediment texture, water quality (temperatureand salinity), disturbance and depth. Thevarious habitat-forming bryozoans on Otagoshelf occur from c. 70 m to the shelf break,although Cinctipora elegans dominates the zoneof highest abundance between 70�100 m. Atdeeper sites, the bryozoans may be more diversebut less abundant. Associated fauna has beenstudied at two scales, although we recognizeboth fauna and habitat exist over a continuum.At the centimetre-decimetre scale, bryozoansexist as topography to be settled, rested orperhaps grazed upon, or sheltered alongside. Atthe millimetre-centimetre scale, bryozoans existas structures to be moved among, burrowedthrough, fed upon or attached to. At this smallscale, the various colony shapes of differentbryozoan species offer a variety of habitats, sothat the physical heterogeneity of the seabedresults to a large extent from the diversity andsize of habitat-forming bryozoans. The biodi-versity associated with the bryozoans maydepend on this variety of centimetre-scalehabitats.
On Otago shelf, where fishing of bryozoanthickets has been light compared with other
known bryozoan-dominated sites in New Zeal-and, recovery in terms of bryozoan settlementand growth is likely to be of the order ofdecades. Fishing on Otago shelf is expected tohave reduced the distribution of habitat domi-nated by bryozoans, and potentially decreasedthe structural complexity of the benthos, evenwithin areas where bryozoans remain in habi-tat-forming quantities. Studies are necessary toprovide parameters for future management.
Future research
The ecology of individual bryozoan species ispoorly known for the New Zealand fauna. Noor limited information exists describing bryozo-an reproduction, growth rates and settlementrequirements, so that settlement and growth ofhabitat-forming species cannot be consideredby management. The rate at which associatedorganisms may settle and their requirements forhabitat in terms of habitat size and quality aresimilarly unknown. A positive relationshipbetween overall colony size and diversity hasbeen demonstrated, both in Otago and else-where (McKinney & Jaklin 2000; Cocito et al.2002; Wood 2005), but which aspects ofbryozoan structure (e.g. height, surface area)are ecologically important? How might thisaffect the management of bryozoan-dominatedhabitats? On Otago shelf, many bryozoanspecies contribute to habitat complexity, butthe effect of this species-level habitat hetero-geneity on associated biodiversity has not beencompared with less heterogeneous sites, whichmay be important in terms of prioritizing sitesfor conservation strategies.
While studies indicate that the diversity ofassociated epifauna is related to the structuralcomplexity and heterogeneity of live bryozoans,the presence and nature of an effect of bryozo-ans on infaunal assemblages is largely un-known. Questions include the importance ofdirect contributions of bryozoans to the sedi-ment (dead colony fragments), the potentialfor organic accumulation around colonies asa result of benthic-pelagic coupling, and the
14 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

accumulation of fine sediments within/undercolonies as a result of baffling of suspendedsediments. Such information would greatlyimprove our understanding of the ecology ofbryozoan-dominated habitats and could beused to inform managers who are required tobalance commercial fishing impacts with theneed to protect associated and dependentspecies, biological diversity and habitats ofparticular significance for fisheries manage-ment (Section 9 of the New Zealand FisheriesAct 1996). For longer-term management, abetter understanding of carbonate productionand buffering, and carbon sequestration underthe combined effects of climate change wouldinform us about, and possibly help mitigate, thelikely effects of climate change.
Acknowledgements
We thank DP Gordon (NIWA) for allowing the
reuse of Fig. 4 from Carter et al. (1985), E Jones
(Golder Associates) for allowing us to reproduce an
image from her thesis, and S Fern and AM Smith
(both University of Otago) for helpful comments on
the manuscript. We acknowledge that Figs. 2 and 3
contain bathymetry information licensed under the
NIWA Open Data Licence v1.0: http://www.niwa.
co.nz/licences/niwa-open-data-licence-by-nn-nc-ver-
sion-1.
References
Abele LG, Patton WK 1976. The size of coral headsand the community biology of associated dec-apod crustaceans. Journal of Biogeography 3:35�47.
Andrews PB 1973. Late Quaternary continental shelfsediments off Otago Peninsula, New Zealand.New Zealand Journal of Geology and Geophy-sics 16: 793�830.
Anon 2005. NIWA’s use of Hydro2de. http://www.fluvial.ch/u/niwa_full_text.html (accessed 7February 2011).
Appeltans W, Bouchet P, Boxshall GA, De Broyer C,de Voogd NJ, Gordon DP, Hoeksema BW,Horton T, Kennedy M, Mees J, Poore GCB,Read G, Stohr S, Walter TC, Costello MJ eds2012. World Register of Marine Species. http://www.marinespecies.org (accessed 16 November2012).
Auster PJ, Langton RW 1999. The effects of fishingon fish habitat. American Fisheries SocietySymposium 22: 150�187.
Baird SJ, Wood BA, Bagley NW 2011. Nature andextent of commercial fishing effort on or nearthe seafloor within the New Zealand 200 n. mileExclusive Economic Zone, 1989�90 to 2004�05.New Zealand Aquatic Environment and Biodi-versity Report No. 73. 143 p.
Batson PB 2000. The Otago shelf bryozoan thickets:aspects of their distribution, ecology and sedi-mentology. Unpublished MSc thesis, Universityof Otago, Dunedin. 166 p.
Batson PB, Probert PK 2000. Bryozoan thickets offOtago Peninsula. New Zealand Fisheries Assess-ment Report 2000/46. 31 p.
Beck MW, Brumbaugh RD, Airoldi L, Carranza A,Coen LD, Crawford C, Defeo O, Edgar GJ,Hancock B, Kay M et al. 2011. Shellfish reefs atrisk globally and recommendations for ecosys-tem revitalization. Bioscience 61: 107�116.
Best MA, Thorpe JP 1996. The effect of suspendedparticulate matter (silt) on the feeding activityof the intertidal ctenostomate bryozoanFlustrellidra hispida (Fabricius). In: GordonDP, Smith AM, Grant-Mackie JA eds Bryozo-ans in space and time. Proceedings of the 10thInternational Bryozoology Association Confer-ence 1995. Wellington, NIWA. Pp. 39�45.
Bostrom C, Jackson EL, Simenstad CA 2006.Seagrass landscapes and their effects on asso-ciated fauna: A review. Estuarine, Coastal andShelf Science 68: 383�403.
Boyd P, LaRoche J, Gall M, Frew R, McKay RML1999. Role of iron, light, and silicate in control-ling algal biomass in subantarctic waters SE ofNew Zealand. Journal of Geophysical ResearchOceans 105 (C6): 13,395.
Bradford-Grieve J, Probert PK, Lewis K, Sutton P,Zeldis J, Orpin A 2006. New Zealand shelfregion. In: Robinson AR, Brink KH eds. Thesea. Cambridge, MA, Harvard University Press.Pp. 1451�1492.
Buhl-Mortensen L, Mortensen PB 2005. Distribu-tion and diversity of species associated withdeep-sea gorgonian corals off Atlantic Canada.In: Freiwald A, Roberts JM eds. Cold-watercorals and ecosystems. Berlin Heidelberg,Springer-Verlag. Pp. 849�879.
Burling RW 1961. Hydrology of circumpolar waterssouth of New Zealand. New Zealand Oceano-graphic Institute Memoir 10. 66 p.
Butler ECV, Butt JA, Lindstrom EJ, Teldesley PC,Pickmere S, Vincent WF 1992. Oceanographyof the Subtropical Convergence Zone around
Otago shelf bryozoan-generated habitat 15
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

southern New Zealand. New Zealand Journal ofMarine and Freshwater Research 26: 131�154.
Carbines G, Jiang W, Beentjes MP 2004. The impactof oyster dredging on the growth of bluecod Parapercis colias, in Foveaux Strait, NewZealand. Aquatic Conservation: Marine andFreshwater Ecosystems 14: 491�504.
Carter L, Heath RA 1975. Role of mean circulation,tides, and waves in the transport of bottomsediment on the New Zealand continental shelf.New Zealand Journal of Marine and Fresh-water Research 9: 423�448.
Carter L, Carter RM 1986. Holocene evolution ofthe nearshore sand wedge, South Otago con-tinental shelf, New Zealand. New ZealandJournal of Geology and Geophysics 29: 413�424.
Carter RM, Carter L, Williams JJ, Landis CA 1985.Modern and relict sedimentation on the SouthOtago continental shelf, New Zealand. NewZealand Oceanographic Institute Memoir 93.43 p.
Chiswell SM 1996. Variability in the SouthlandCurrent, New Zealand. New Zealand Journalof Marine and Freshwater Research 30: 1�17.
Cocito S, Ferdeghini F, Pisaroni S, Bedulli D 2002.Influence of colony morphology on associatedbiota diversity in four Bryozoa. In: WyseJackson PN, Buttler CJ, Spencer Jones MEeds. Bryozoan Studies 2001: Proceedings of the12th International Bryozoology AssociationConference. Lisse,TheNetherlands,AABalkema.Pp. 83�88.
Cranfield HJ, Michael KP, Doonan IJ 1999. Changesin the distribution of epifaunal reefs and oystersduring 130 years of dredging for oysters inFoveaux Strait, southern New Zealand. AquaticConservation: Marine and Freshwater Ecosys-tems 9: 461�483.
Cranfield HJ, Manighetti B, Michael KP, Hill A2003. Effects of oyster dredging on the distribu-tion of bryozoan biogenic reefs and associatedsediments in Foveaux Strait, southern NewZealand. Continental Shelf Research 23: 1337�1357.
Cranfield HJ, Rowden AA, Smith DJ, Gordon DP,Michael KP 2004. Macrofaunal assemblages ofbenthic habitat of different complexity and theproposition of a model of biogenic reef habitatregeneration in Foveaux Strait, New Zealand.Journal of Sea Research 52: 109�125.
Croot PL, Hunter KA 1998. Trace metal distribu-tions across the continental shelf near OtagoPeninsula, New Zealand. Marine Chemistry61: 185�201.
Cryer M, O’Shea S, Gordon D, Kelly M, Drury J,Morrison M, Hill A, Saunders H, Shankar U,Wilkinson M et al. 2000. Distribution andstructure of benthic invertebrate communitiesbetween North Cape and Cape Reinga. Finalresearch report for Ministry of Fisheriesresearch project ENV 9805 Objectives 1�4.154 p.
Currie KI, Reid M, Hunter K 2009. Interannualvariability of carbon dioxide drawdown bysubantarctic surface water near New Zealand.Biogeochemistry. doi: 10.1007/s10533-009-9355-3
Desroy N, Dubois SF, Fournier J, Ricquiers L,Mao PL, Guerin L, Gerla D, Rougerie M,Legendre A 2011. The conservation status ofSabellaria alveolata (L.) (Polychaeta: Sabellar-iidae) reefs in the Bay of Mont-Saint-Michel.Aquatic Conservation: Marine and FreshwaterEcosystems 21: 462�471.
Foster G, Carter L 1997. Mud sedimentation on thecontinental shelf at an accretionary margin*Poverty Bay, New Zealand. New ZealandJournal of Geology and Geophysics 40: 157�173.
Froude VA, Smith R 2004. Area-based restrictionsin the New Zealand marine environment. De-partment of Conservation MCU Report. 169 p.
Gappa JL 2000. Species richness of marine Bryozoain the continental shelf and slope off Argentina(south-west Atlantic). Diversity and Distribu-tions 6: 15�27.
Gordon DP, Taylor PD, Bigey FP 2009. PhylumBryozoa: moss animals, sea mats, lace corals. In:Gordon DP ed. The New Zealand inventory ofbiodiversity. Volume 1. Kingdom Animalia*Radiata, Lophotrochozoa, and Deuterostomia.Christchurch, Canterbury University Press. Pp.271�297.
Grange KR, Tovey A, Hill AF 2003. The spatialextent and nature of the bryozoan communitiesat Separation Point, Tasman Bay. MarineBiodiversity Biosecurity Report 4. 22 p.
Gray JS 1974. Animal-sediment relationships. Ocea-nography and Marine Biology: An AnnualReview 12: 223�262.
Griffiths GA, Glasby GP 1985. Input of river-derived sediment to the New Zealand continen-tal shelf: 1. Mass. Estuarine, Coastal and ShelfScience 21: 773�787.
Gruber N 2011. Warming up, turning sour, losingbreath: ocean biogeochemisty under globalchange. Philosophical Transactions of the RoyalSociety A 369: 1980�1996.
Gutt J, Starmans A 1998. Structure and biodiversityof megabenthos in the Weddell and LazarevSeas (Antarctica): Ecological role of physical
16 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

parameters and biological interactions. PolarBiology 20: 229�247.
Harmelin J-G, Capo S 2002. Effects of sewage onbryozoan diversity in Mediterranean rockybottoms. In: Wyse Jackson PN, Buttler CJ,Spencer Jones ME eds. Bryozoan Studies 2001:Proceedings of the 12th International Bryozool-ogy Association Conference. Lisse, The Nether-lands, AA Balkema. Pp. 151�157.
Hawke DJ 1989. Hydrology and near-surface nu-trient distribution along the South Otago con-tinental shelf, New Zealand, in summer andwinter 1986. New Zealand Journal of Marineand Freshwater Research 23: 411�420.
Hawke DJ 1992. Salinity variability in shelf watersnear Otago Peninsula, New Zealand, on a timescale of hours. New Zealand Journal of Marineand Freshwater Research 26: 167�173.
Hawke DJ 1995. Reactive silicate in shelf waters nearOtago Peninsula, New Zealand. Marine andFreshwater Research 46: 427�433.
Haywood GJ 2004. Some effects of river dischargesand currents on phytoplankton in the sea offOtago, New Zealand. New Zealand Journal ofMarine and Freshwater Research 38: 103�114.
Hicks DM, Shankar U 2003. Sediment from NewZealand rivers. NIWA Chart, MiscellaneousSeries No. 79.
Hill NAO, Michael KP, Frazer A, Leslie S 2010. Theutility and risk of local ecological knowledge indeveloping stakeholder driven fisheries manage-ment: The Foveaux Strait dredge oyster fishery,New Zealand. Ocean & Coastal Management53: 659�668.
Hopkins J, Shaw AGP, Challenor P 2010. TheSouthland Front, New Zealand: Variabilityand ENSO correlations. Continental Shelf Re-search 30: 1535�1548.
Hydrographic Department 1958. The New Zealandpilot. 12th edition. London, Admiralty Hydro-graphic Department. 500 p.
Jamet C 1999. Bryozoan colony growth rates: Aproxy for cool-water marine carbonate produc-tion rates. Unpublished Senior thesis, DickinsonCollege, Carlisle, Pennsylvania. 40 p.
Jiang W, Carbines G 2002. Diet of blue cod,Parapercis colias, living on undisturbed biogenicreefs and on seabed modified by oyster dredgingin Foveaux Strait, New Zealand. Aquatic Con-servation: Marine and Freshwater Ecosystems12: 257�272.
Jiang W, Gibbs MM, Hatton S 2004. Stock assess-ment of the queen scallop fishery in QSC3,QSC2002/01. Draft research report prepared forthe Ministry of Fisheries. 54 p.
Jillett JB 1969. Seasonal hydrology of the waters offthe Otago Peninsula, south-eastern New Zealand.New Zealand Journal of Marine and FreshwaterResearch 3: 349�375.
Jones CD, Lockhart SJ 2011. Detecting vulnerablemarine ecosystems in the Southern Ocean usingresearch trawls and underwater imagery. MarinePolicy 35: 732�736.
Jones E 2006. Bryozoan thickets on Otago shelf,New Zealand: a quantitative assessment of theepibenthos using underwater photography. Un-published MSc thesis, University of Otago,Dunedin.
Jones JB 1992. Environmental impact of trawling onthe seabed: A review. New Zealand Journal ofMarine and Freshwater Research 26: 59�62.
Junge C 1998. Bryozoen und Bryozoen-Riff-Struk-turen auf dem Kontinentalschelf von Otago/Neuseeland. Unpublished MSc/Diplomarbeitthesis, University of Hamburg, Hamburg. 182 p.
Kamenos NA, Moore GP, Hall-Spencer JM 2004.Maerl grounds provide both refuge and highgrowth potential for juvenile queen scallops(Aequipecten opercularis L.). Journal of Experi-mental Marine Biology and Ecology 313: 241�254.
Lagaaij R, Gautier YV 1965. Bryozoan assemblagesfrom marine sediments of the Rhone delta,France. Micropaleontology 11: 39�58.
Loutit TS 1975. Continental shelf sedimentation eastof Karitane Point, East Otago, New Zealand.Unpublished Honours thesis, University ofOtago, Dunedin. 92 p.
Mace J 1981. Separation Point closed. Catch 8:15�16.
McKinney FK, Jackson JBC 1989. Bryozoan Evolu-tion. Chicago, The University of Chicago Press.238 p.
McKinney FK, Jaklin A 2000. Spatial niche parti-tioning in the Cellaria meadow epibiont asso-ciation, northern Adriatic Sea. Cahiers deBiologie Marine 41: 1�17.
Michael KP, Cranfield HJ 2001. A summary of thefishery, commercial landings, and biology of theNew Zealand queen scallop, Zygochlamysdelicatula (Hutton, 1873). 24 p.
Ministry of Fisheries 2007. Queen Scallop (QSC 3)Plenary Document. Pp. 690�692.
Morgado EH, Tanaka MO 2001. The macrofaunaassociated with the bryozoan Schizoporellaerrata (Walters) in southeastern Brazil. ScientiaMarina 65: 173�181.
Nelson CS, Hyden FM, Keane SL, Leask WL,Gordon DP 1988a. Application of Bryozoazoarial growth-form studies in facies analysis
Otago shelf bryozoan-generated habitat 17
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

of non-tropical carbonate deposits in NewZealand. Sedimentary Geology 60: 301�322.
Nelson CS, Keane SL, Head PS 1988b. Non-tropicalcarbonate deposits on the modern New Zealandshelf. Sedimentary Geology 60: 71�94.
New Zealand Fisheries Act 1996. Public Act 1996No 88.
Pauly D 1995. Anecdotes and the shifting baselinesyndrome of fisheries. Trends in Ecology &Evolution 10: 430 p.
Pfannkuche J 1998. The optical properties of Otagowaters. Unpublished MSc thesis, University ofOtago, Dunedin. 94 p.
Powell AWB 1950. Mollusca from the continentalshelf, eastern Otago. Records of the AucklandInstitute and Museum 4: 73�82.
Probert PK 1977. Relict or recent shelf morphologyoff east Otago? Some relevant epifaunal obser-vations. New Zealand Journal of Geology andGeophysics 20: 397�398.
Probert PK, Wilson JB 1984. Continental shelfbenthos off Otago Peninsula, New Zealand.Estuarine, Coastal and Shelf Science 19:373�391.
Probert PK, Batham EJ, Wilson JB 1979. Epibenthicmacrofauna off southeastern New Zealand andmid-shelf bryozoan dominance. New ZealandJournal of Marine and Freshwater Research 13:379�392.
Roberts JM 2005. Reef-aggregating behaviour bysymbiotic eunicid polychaetes from cold-watercorals: Do worms assemble reefs? Journal of theMarine Biological Association of the UnitedKingdom 85: 813�819.
Rodolfo-Metalpa R, Lombardi C, Cocito S, Hall-Spencer JM, Gambi MC 2010. Effects of oceanacidification and high temperatures on thebryozoan Myriapora truncata at natural CO2
vents. Marine Ecology 31: 447�456.Rowden AA, Warwick RM, Gordon DP 2004.
Bryozoan biodiversity in the New Zealandregion and implications for marine conserva-tion. Biodiversity and Conservation 13: 2695�2721.
Russell P, Vennell R 2009. Observations of second-ary flow and upwelling at Cape Saunders, OtagoPeninsula. Proceedings of the 18th AustralasianCoasts and Ports Conference, Wellington, NewZealand.
Ryland JS 1970. Bryozoans. London, Hutchinson.175 p.
Saxton F 1980. Coral loss could deplete fish stocks.Catch 80: 15�16.
Smith AM 2009. Bryozoans as southern sentinels ofocean acidification: A major role for a minor
phylum. Marine and Freshwater Research 60:475�482.
Smith AM, McGourty CR, Kregting L, Elliot A2005. Subtidal Galeolaria hystrix (Polychaeta:Serpulidae) reefs in Paterson Inlet, StewartIsland, New Zealand. New Zealand Journal ofMarine and Freshwater Research 39: 1297�1304.
Smith AM, Stewart B, Key Jr MM, Jamet CM2001. Growth and carbonate production byAdeonellopsis (Bryozoa: Cheilostomata) inDoubtful Sound, New Zealand. Palaeogeogra-phy, Palaeoclimatology. Palaeoecology 175:201�210.
Stebbing ARD 1971. The epizoic fauna of Flustrafoliacea (Bryozoa). Journal of the Marine Bio-logical Association of the United Kingdom 51:283�300.
Sutton PJH 2003. The Southland Current: a sub-antarctic current. New Zealand Journalof Marine and Freshwater Research 37: 645�652.
Swanson KM 1979. The marine fauna of NewZealand: Ostracods of the Otago shelf. NewZealand Oceanographic Institute Memoir 78.56 p.
Taylor PD 2000. Origin of the modern bryozoanfauna. In: Culver SJ, Rawson PF eds. Bioticresponse to global change: The last 145 millionyears. Cambridge, Cambridge University Press.Pp. 195�206.
Taylor PD, Allison PA 1998. Bryozoan carbonatesthrough time and space. Geology 26: 459�462.
Taylor PD, Gordon DP, Batson PB 2004. Bathy-metric distributions of modern populationsof some common Cenozoic Bryozoa fromNew Zealand, and paleodepth estimation. NewZealand Journal of Geology and Geophysics 47:57�69.
Taylor R, Smith I 1997. The state of New Zealand’senvironment 1997. Ref. ME612. Ministry forthe Environment. Chapter 7, p. 20.
Thrush SF, Dayton PK 2002. Disturbance to marinebenthic habitats by trawling and dredging:implications for marine biodiversity. AnnualReview of Ecology and Systematics 33: 449�473.
Vooren CM 1975. Nursery grounds of tarakihi(Teleostei: Cheilodactylidae) around NewZealand. New Zealand Journal of Marine andFreshwater Research 9: 121�158.
Watling L, Norse EA 1998. Disturbance of theseabed by mobile fishing gear: A comparisonto forest clearcutting. Conservation Biology 12:1180�1197.
18 ACL Wood and PK Probert
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013

Willan RC 1981. Soft-bottom assemblages of Pater-son Inlet, Stewart Island. New Zealand Journalof Zoology 8: 229�248.
Wood ACL 2005. Communities associated withhabitat-forming bryozoans from Otago shelf,Southern New Zealand. Unpublished MSc the-sis, University of Otago, Dunedin.
Wood ACL, Probert PK, Rowden AA, Smith AM2012. Complex habitat generated by marinebryozoans: A review of its distribution, struc-
ture, diversity, threats and conservation.Aquatic Conservation: Marine and FreshwaterEcosystems 22: 547�563.
Zeldis J, Hicks M, Trustrum N, Orpin A, Nodder S,Probert K, Shankar U, Currie K 2010. NewZealand continental margins. In: Liu K-K,Atkinson L, Quinones R, Talaue-McMannus Leds. Carbon and nutrient fluxes in continentalmargins: A global synthesis. Berlin Heidelberg,Springer-Verlag. Pp. 273�287.
Otago shelf bryozoan-generated habitat 19
Dow
nloa
ded
by [
Uni
vers
ity o
f O
tago
] at
15:
45 2
0 A
pril
2013




















