
Applying the Contingent Valuation Method in Resource Accounting: A Bold Proposal
Upload
campus-paris-saclayCategory
view
1download
0
ORIGINAL INVESTIGATION
Brain activation by short-term nicotine exposurein anesthetized wild-type and beta2-nicotinic receptorsknockout mice: a BOLD fMRI study
S. V. Suarez & A. Amadon & E. Giacomini & A. Wiklund &
J.-P. Changeux & D. Le Bihan & S. Granon
Received: 14 May 2008 /Accepted: 10 September 2008# Springer-Verlag 2008
AbstractRationale The behavioral effects of nicotine and the role ofthe beta2-containing nicotinic receptors in these behaviorsare well documented. However, the behaviors altered bynicotine rely on the functioning on multiple brain circuitswhere the high-affinity beta2-containing nicotinic receptors(β2*nAChRs) are located.Objectives We intend to see which brain circuits areactivated when nicotine is given in animals naïve fornicotine and whether the β2*nAChRs are needed for itsactivation of the blood oxygen level dependent (BOLD)signal in all brain areas.Materials and methods We used functional magneticresonance imaging (fMRI) to measure the brain activationevoked by nicotine (1 mg/kg delivered at a slow rate for45 min) in anesthetized C57BL/6J mice and beta2 knockout(KO) mice.
Results Acute nicotine injection results in a significantincreased activation in anterior frontal, motor, and somato-sensory cortices and in the ventral tegmental area and thesubstantia nigra. Anesthetized mice receiving no nicotineinjection exhibited a major decreased activation in allcortical and subcortical structures, likely due to prolongedanesthesia. At a global level, beta2 KO mice were notrescued from the globally declining BOLD signal. Howev-er, nicotine still activated regions of a meso-cortico-limbiccircuit likely via alpha7 nicotinic receptors.Conclusions Acute nicotine exposure compensates for thedrop in brain activation due to anesthesia through the meso-cortico-limbic network via the action of nicotine onβ2*nAChRs. The developed fMRI method is suitable forcomparing responses in wild-type and mutant mice.
Keywords Psychostimulant . Addiction . Dependence .
Reward system . Anesthesia . Imaging
Introduction
Nicotine is a major addictive component of tobacco smokeand a strong psychostimulant. From a behavioral point ofview, nicotine has been involved in multiple cognitive,emotional, and psychomotor processes, either after acute orchronic exposure, in human (Heishman et al. 1994;Newhouse et al. 2004; Xu et al. 2007), primate (Valette etal. 2007), or rodent models (e.g., Stolerman et al. 1973;Marks et al. 1985; Gäddnäs et al. 2000; Besson et al. 2007;for review, see Matta et al. 2007). It activates the release ofseveral neurotransmitters (Li and Eisenach 2002; Rossi et al.2005; Villégier et al. 2006; Fallon et al. 2007), stimulatesdopaminergic neuron firing (Mameli-Engvall et al. 2006)and dopamine striatal release (Di Chiara 2000; Pietila and
PsychopharmacologyDOI 10.1007/s00213-008-1338-x
S. V. Suarez : J.-P. Changeux : S. Granon (*)Unité de Neurobiologie Intégrative du Système Cholinergique,URA CNRS 2182, Institut Pasteur, Département de Neuroscience,25 rue du Dr. Roux,75015 Paris, Francee-mail: [email protected]
A. Amadon : E. Giacomini :D. Le BihanService Hospitalier Frédéric Joliot,4 place du général Leclerc,91400 Orsay, France
A. WiklundSection of Anaesthesiology and Intensive Care Medicine,Department of Physiology and Pharmacology,Karolinska Institutet,Stockholm, Sweden

Ahtee 2000), and enhances the transcription of multipleimmediate early genes in various brain regions (Hiremagalurand Sabban 1995). The nicotinic receptors are pentamericoligomers resulting from the assembly of diverse subunits(Le Novère et al. 2002; Gotti and Clementi 2004) and arequite ubiquitously distributed in the brain (Champtiaux andChangeux 2004; Han et al. 2003; Gotti et al. 2007;Changeux and Edelstein 2005 for review), in neuronal types(Gotti and Clementi 2004 for review), and in cell compart-ments (Devillers-Thiéry et al. 2003). Therefore, assessing theregional brain activation at the circuit level responsible forthe effect of nicotine on a particular neuronal type and/or aspecific nicotinic receptor subtype is particularly challeng-ing. The receptors containing the beta2 subunit are the mostabundant in the mammal brain (Le Novère et al. 2002) andexhibit the highest affinity for nicotine (Picciotto et al.1995). We have previously shown that mice lacking thenicotinic receptors containing the beta2 subunit (β2−/−mice) did not self-administer nicotine either intravenously(Picciotto et al. 1998) or intracerebrally (Besson et al. 2006)but showed normal withdrawal syndrome when nicotine,given passively and chronically, was removed (Besson et al.2006). Moreover, it was recently shown that the nicotine-elicited activation of the dopaminergic neurons of the ventraltegmental area (VTA) is abolished in β2−/− mice (Mameli-Engval et al. 2006), whereas the dopaminergic receptoractivation, the dopamine release in projection areas, andsome of the behavioral defects identified in β2−/− mice(Granon et al. 2003) were recovered after re-expression ofthe β2*nAChRs within the VTA via a viral vector (Maskoset al. 2005). However, despite this important amount of dataconcerning the effect of nicotine on the brain and onβ2*nAChRs, two major questions remain: Would an acuteinjection of nicotine produce a general brain activationconsistent with the ubiquity of nicotinic receptors or would itmobilize only some particular brain circuits, such as thoseidentified by local methods (electrophysiology and micro-dialysis)? In other words, does the nicotine-elicited brainactivation require the β2*nAChRs in all circuits?
Prior imaging studies using magnetic resonanceimaging showed that it is a suitable and powerful tech-nique to localize in vivo drug actions (Stein 2001) and toimage nicotine action on the brain in rats (Choi et al.2006). Furthermore, it allows the comparison betweendifferent strains of mice (Small et al. 2000), and bloodoxygen level dependent (BOLD) can reveal patternslinked to cellular gene activation/deactivation in rats(Stark et al. 2006).
Here, we measured the BOLD activation evoked bysubcutaneous administration of nicotine in control andβ2−/− mice in order to investigate both the effect ofnicotine on regional brain activation in mice and thecontribution of the β2*nAChRs in this activation pattern.
Materials and methods
Subjects
All mice used in this study were 4- to 6-month-old malesand naive for any behavioral or pharmacological experi-ments. C57BL6/J wild-type (WT) mice and mice withdeletion of the β2 nicotinic subunit gene (β2−/−) wereobtained from Charles River Laboratories (France). Allanimals were transferred to the magnetic resonance imaging(MRI) facility at least 2 weeks before the experiments. Theconstruction of the genetically modified mice deleted[knockout (KO)] has been described previously (Picciottoet al. 1995). β2−/− mice were obtained from parentsbackcrossed for 19 generations (β2−/−) to the C57BL6/6Jparental strain. Mice were housed four to five per cageunder a 12-h light–dark cycle in rooms at a controlledtemperature (21°C) with free access to water and food. Allexperiments were carried out during the light phase,between 8 A.M. and 7 P.M. Experiments were carried outfollowing the guidelines of the European CommunitiesCouncil (86/609/EEC).
Fifteen mice were used during the pilot study designedto set up the functional MRI (fMRI) sequences. Twenty-seven mice were then subsequently used, of which six werediscarded because of major signal artifacts (four), move-ments during scanning (one), or premature death (one).
Twenty-one animals were therefore considered: sixC57bl/6 mice receiving nicotine, eight C57bl/6 micereceiving placebo (control), and seven KO receivingnicotine.
Drugs
Anesthesia
Mice were anesthetized with an initial 0.1 ml/10 gintraperitoneal injection of 2.5% xylazine (Rompun 2%)and 15% ketamine (Imalgen 1000) combination diluted in82.5% PBS. The anesthetic was then continuously deliv-ered subcutaneously via a catheter implanted at the nap ofthe neck (internal diameter 0.36 mm) connected to aHarvard pump at the slow rate of 0.1 ml/10 g/h.
Nicotine
Nicotine hydrogen tartrate salt (Sigma) was dissolved insterile saline solution (0.9% NaCl), and the pH wasadjusted to 7 with sodium hydroxide (NaOH). A solutionof nicotine 1 mg/kg (expressed as free base) was preparedand frozen so that the same concentration was used for eachmouse. An initial bolus of nicotine (0.1 ml/10 g) wasinjected via the subcutaneous catheter, and then, the
Psychopharmacology

nicotine solution was delivered continuously at the rate of0.1 ml/10 g/h during 45 min. The BOLD sequence started5 min after the initial injection of nicotine.
Placebo
The placebo solution was constituted of 0.9% NaCl only.
Design and procedures
MRI setup
Each mouse was weighed and anesthetized. The nap of theneck was then shaved, and a small incision was performedto implant subcutaneously two catheters (internal diameter0.36 mm), one for nicotine (or the placebo) and one for theanesthetic, securely taped with medical tape on the skin. Asensor for cardiac activity was placed under the chest of theanimal and connected to a monitoring device placed outsideof the scanner. The animal was then placed within aplexiglass home-made stereotaxic frame comprising tooth-support and specially shaped ear-bars designed to provide afixed and flat head without any pain or breathing difficulty(see Introduction of Paxinos and Franklin 2004). Ear-barstherefore did not enter the auditory meatus but ended with aflat surface covered with a soft material. A slim plastic leafwas placed on the skull and the eyes of the animal in orderto separate the head from the RF head coil. The animal wasplaced between the two curls of a home-made Helmoltz coil(see below). Finally, the animal was covered with a heatingblanket (circuit of hot water), and the entire stereotaxicframe was placed within the MRI machine.
General MRI and acquisition sequence
Studies were performed using a 3 T whole-body MRIsystem (Bruker) equipped with a gradient hardware reach-ing 45 mT/m in each direction, with a maximum slew rate of140 mT/m/ms. The gradient system had a 55-cm innerdiameter, leaving plenty of room for mouse placement.However, this clinical system was not optimized for smallanimal imaging. Limitations on the gradient hardwareprevented fast acquisitions at high (microscopic) resolution.A standard 2D T2*-weighted gradient-echo sequence withgradient-spoiled magnetization was used to acquire fMRIdata, both in control and under nicotine conditions. Theacquisition parameters were the following: TE=30 ms; TR=2450 ms, during which 40 250-μm-thick interlaced adjacentslices were acquired using a 85° SLR-type 3-lobe RF pulse(Pauly et al. 1991) for sharp selective excitation. The sliceswere selected perpendicular to the magnet axial direction,corresponding to the coronal plane of the mouse head. Theread direction followed the vertical axis, whereas the phase
encode direction was the mouse left-right axis. For every slice,128×256 points were acquired with an 8.9-kHz sampling rateand a 19.2×38.4-mm field-of-view. Thus, a 150-μm in-planeresolution was achieved. Four signal acquisitions wereaccumulated to double signal-to-noise, leading to a totalacquisition time of 42 min for each condition. Thus, for everymouse, a first brain image was acquired while injecting theanesthetic solution only. This constituted the baseline imageto be subtracted from the subsequent activation conditionimage. So, after a 10–15-min break to let the gradient coilscool down, a second image was acquired while injectingeither the nicotine or placebo solution in addition to theanesthetic.
Construction of the head coil for mice
We optimized our home-made coil as a compromisebetween the need for access to the mouse head, therequirement for a generated B1 field as homogeneous aspossible, and the need for high sensitivity accessibility. Wetherefore designed a Helmholtz-type double coil (seeFig. 1) with distributed capacities. The coupling wasinductive, and the ideal geometry of the double coil allowedeasy access to ear and tooth bars in order to restrict themovement of the mouse head. The diameter of the double
Fig. 1 Double coil designed for the mouse brain. The black loops areHelmoltz resonators (diameter 1.2 cm; space between the two loops1.2 cm). The silver-coated loop on top of the coil allows coupling andadaptation of impedance. The black box to which the loops areattached serves to adapt the frequency. The entire system was fixed onthe plexiglass stereotaxic frame
Psychopharmacology

coil was 12 mm, and the space between the two loops was12-mm wide to allow the head of the mouse to be placed inbetween. The ratio (close to 1) between the radius of thecoils and their spacing gave a good B1 homogeneity in thevolume targeted. The distribution of the capacitanceallowed the division of the resistance and decreased theloss, therefore, increasing the quality of the signal.
The electric circuit was the following:
wo ffi 1ffiffiffiffiffiffiffi
LCp ; Qv ¼ Lwo
Ravec wo ¼ 2pfo
At fo=125.120 MHz, the coil quality factor was measuredto be 105 when loaded with the mouse head (Q=140unloaded).
The coupling to realize the impedance adaptation wasinductive, with the coupling constant M ¼ k
ffiffiffiffiffiffiffiffiffi
LLop
and k afactor dependent of the distance.
At the resonance of the probe, we have an entryimpedance Zo ¼ M2w2
R .
Data analysis
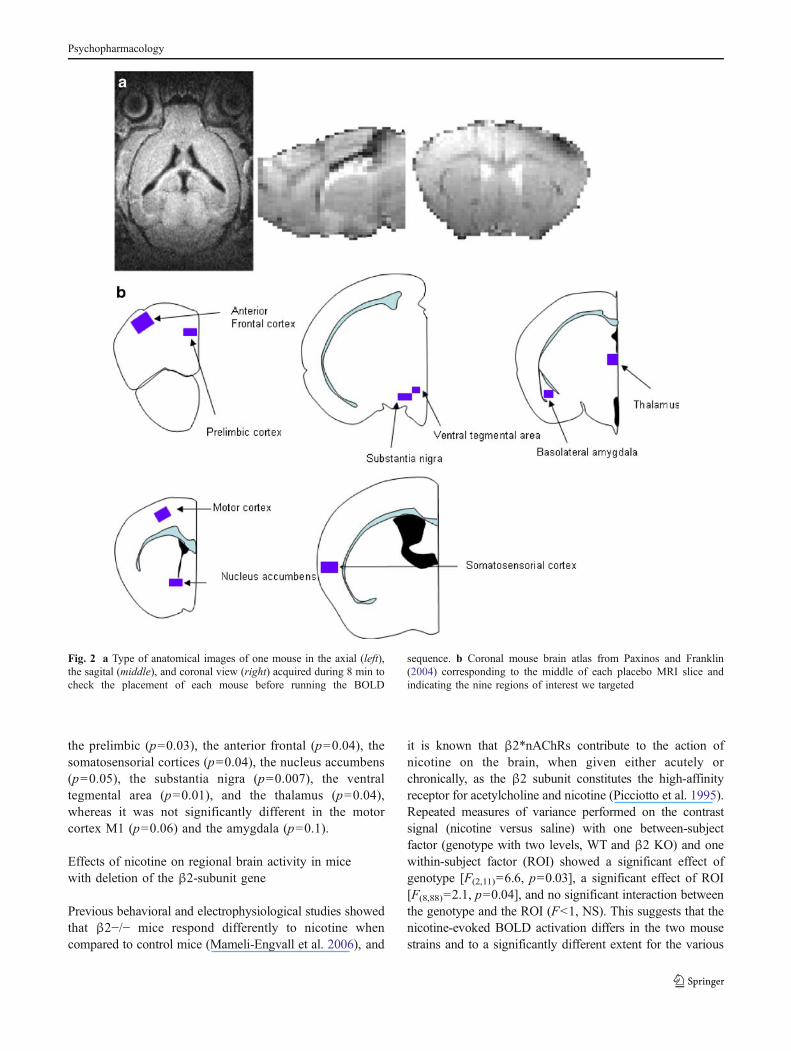
The mouse was placed in the scanner so that for eachmouse, an anatomical image can be obtained in the threedimensions (Fig. 2a).
Signal treatment
We used the Anatomist/BrainVISA software package(version 3.0.1 for Linux 2005; Rivière et al. 2000, 2003)to perform image post-processing and analysis.
For each mouse, a “Brain Only” region of interest (ROI)was created by extracting the brain contour of the baselinecondition. The extraction was performed by hand based on30 coronal slices. This ROI was used as a mask. The regionoutside the mask was subtracted from the baseline image.Then, the nicotine (or placebo) condition image wasaligned by hand with the Brain Only baseline image. Alinear spline interpolation was used to adjust the alignedimage. The region outside the Brain Only mask wassubtracted from the nicotine/placebo image. Finally, theactivation intensity in each ROI was extracted by subtract-ing the baseline image from the nicotine/placebo image.
The 3D picture of nicotine condition was aligned byhand with the Brain Only picture of placebo condition. Theregion outside the Brain Only ROI mask was then
subtracted from the nicotine 3D picture with “AimsMask”command. Finally, the activation intensity in each ROI wasextracted with the “AimsVoiStat” command.
Regions of interest
Nine 3D ROIs were delineated by hand on the placeboimage using Anatomist software, and a corresponding maskfor each ROI was generated (see Fig. 2b).
Statistical analysis
In C57bl/6 mice, the effect of nicotine on BOLD contrastswas analyzed by repeated measures of variance ANOVA(Statview 5.0) with “drug”, with two levels—saline andnicotine—as a main between-subject factor and ROI as amain within-subject factor, with nine levels—prelimbiccortex, anterior frontal cortex, somatosensorial cortex,motor M1 cortex, basolateral amygdala, nucleus accum-bens, thalamus, ventral tegmental area, and substantia nigra.In the control experiment, we investigated the role of β2-containing nicotinic receptors. Repeated measures ANOVAon nicotine-evoked BOLD contrasts were conducted with“genotypes” as a main between-subject factor with twolevels (WT and β2−/−) and ROI as a main within-subjectfactor (same ROIs as above). Studies of potential co-activation of different brain regions or of correlationbetween the baseline BOLD activity and the nicotine-evoked BOLD activation were analyzed with simple linearregression measures (StatView 5.0).
Results
Effects of nicotine on regional brain activity in C57BL/6mice
A continuous and slow subcutaneous delivery of nicotineresulted in a significant BOLD response in cortical andsubcortical areas in C57BL/6 mice, as compared to micereceiving only saline (Fig. 3a and b). Repeated measures ofvariance performed on the contrast signal obtained betweenthe second session (either saline or nicotine) and the firstsession (anesthetic) of the acquisition for each region ofinterest showed a significant nicotine effect [F(1,12)=5.9,p=0.03], no significant effect of ROI [F(8,96)=1.2, NS],and a significant interaction between the drug and theROIs [F(8,96)=2.3, p=0.03]. These results suggest thatnicotine caused brain activation in a different manner thansaline and that the signal recorded in the various ROIs wasaffected in a different manner by nicotine and saline. Posthoc Fisher test showed that the nicotine-evoked activationsignificantly differed from the baseline saline activity in
Psychopharmacology
the prelimbic (p=0.03), the anterior frontal (p=0.04), thesomatosensorial cortices (p=0.04), the nucleus accumbens(p=0.05), the substantia nigra (p=0.007), the ventraltegmental area (p=0.01), and the thalamus (p=0.04),whereas it was not significantly different in the motorcortex M1 (p=0.06) and the amygdala (p=0.1).
Effects of nicotine on regional brain activity in micewith deletion of the β2-subunit gene
Previous behavioral and electrophysiological studies showedthat β2−/− mice respond differently to nicotine whencompared to control mice (Mameli-Engvall et al. 2006), and
it is known that β2*nAChRs contribute to the action ofnicotine on the brain, when given either acutely orchronically, as the β2 subunit constitutes the high-affinityreceptor for acetylcholine and nicotine (Picciotto et al. 1995).Repeated measures of variance performed on the contrastsignal (nicotine versus saline) with one between-subjectfactor (genotype with two levels, WT and β2 KO) and onewithin-subject factor (ROI) showed a significant effect ofgenotype [F(2,11)=6.6, p=0.03], a significant effect of ROI[F(8,88)=2.1, p=0.04], and no significant interaction betweenthe genotype and the ROI (F<1, NS). This suggests that thenicotine-evoked BOLD activation differs in the two mousestrains and to a significantly different extent for the various
Fig. 2 a Type of anatomical images of one mouse in the axial (left),the sagital (middle), and coronal view (right) acquired during 8 min tocheck the placement of each mouse before running the BOLD
sequence. b Coronal mouse brain atlas from Paxinos and Franklin(2004) corresponding to the middle of each placebo MRI slice andindicating the nine regions of interest we targeted
Psychopharmacology

ROIs. Post hoc Student tests showed that β2 KO miceshowed significantly different (decreased) regional activationfrom WT mice in the anterior frontal (t=3.7, df=11, p=0.004), motor M1 (t=2.4, df=11, p=0.03), and the somato-sensorial cortices (t=2.6, df=11, p=0.02), as well as in thesubstantia nigra (t=2.6, df=11, p=0.02), the ventral tegmen-tal area (t=2.9, df=11, p=0.02), and the thalamus (t=2.5,df=11, p=0.03). The genotype effect was only marginal inthe prelimbic cortex (t=1.9, df=11, p=0.08), the nucleusaccumbens (t=2.1, df=11, p=0.06), and the amygdala (t=2.0, df=11, p=0.07). These results, illustrated on Fig. 3a andc, show that β2*nAChRs contribute significantly to the
acute nicotine-evoked brain activation in nearly all the brainregions investigated.
Control of physiological parameters in WT and β2−/− miceduring a 2-h anesthesia procedure
Outside of the scanner, we conducted a control experimentdevoted to check whether classical physiological parameters(Nonin 8500AV, Nonin Medical, MN, USA) such astemperature, respiratory frequency, O2 saturation, and cardiacfrequency would be differentially affected by a 2-h anesthesiain WT and β2−/− mice. We therefore measured these
Fig. 3 BOLD signal image of one mouse (a) and mean percentage ofBOLD signal contrasts calculated for the different ROIs (b and c).Each bar represents the mean of all animals of each group of the totalBOLD signal contrast±SEM. b Nicotine versus saline (n=6; black
bars) and saline versus saline (n=8; white bars) contrasts in controlmice. c Nicotine versus saline contrast in control (n=6; white bars)and in β2 KO (n=7; black bars) mice
Psychopharmacology

parameters in the same experimental conditions as the oneused in the MRI scanner (continuous xylazine/ketamineanesthesia, animals under a heating blanket, and 2-h duration)but on different WT and β2−/− animals as the one used inthe scanner. The measures were taken every 30 min for 2 h.As in the MRI scanner, no oxygen was added to the inspiredair.
Results show that oxygen saturation fluctuated between 76and 74 in WT mice and 71 to 75 in β2−/− mice from the firstmeasure taken 30 min after the beginning of the anesthesia tothe last one measured 120 min after the beginning. From thebeginning to the end of the experiment, cardiac frequencyfluctuated from 293 to 312 inWTmice and from 341 to 324 inβ2−/− mice, whereas respiratory frequency fluctuated from184 to 187 in WT mice and from 293 to 158 in β2−/− mice.Temperature fluctuated from 37 to 36 in WT mice and from39 to 34 in β2−/− mice. None of these physiologicalparameters were significantly different between the geno-types. Respiratory frequency was the most variable parame-ters between animals during anesthesia, and although notfalling to levels that would cause hypoxic damage to theorganism, is the parameter that was the most altered byanesthesia. These results suggest that none of these physio-logical constants can explain the BOLD response differencesbetween WT and β2−/−mice.
Regional correlation between the BOLD signalduring nicotine delivery and saline delivery in WTand β2−/− mice
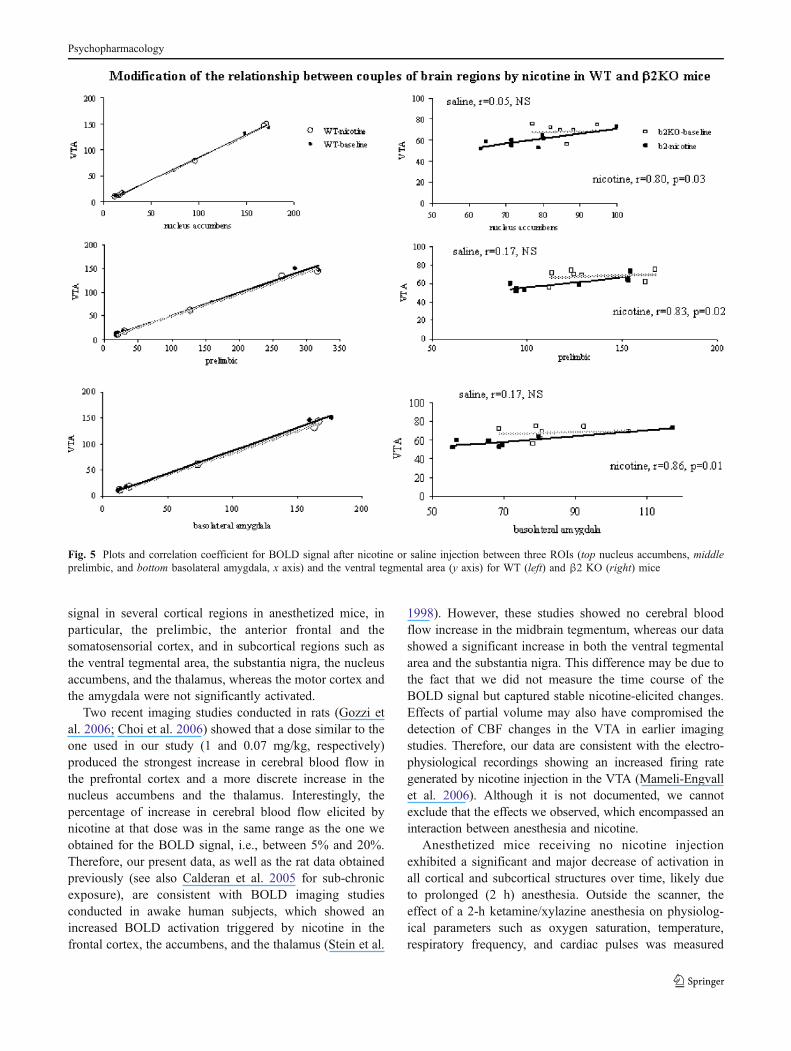
We observed a large interindividual variability for theabsolute value of the BOLD signal intensity, independentlyof the genotype considered. We therefore intended to seewhether nicotine produced a BOLD signal change corre-lating with the signal recorded during the baseline sessionor whether the nicotine-evoked signal was independent ofthe baseline level of activity. Therefore, statistical correla-tions between the nicotine and baseline activity werecalculated for pairs of ROIs selected for being directlyanatomically connected, in WT and β2−/− mice. In WTmice, the nicotine-evoked BOLD signal change waspositively and highly correlated to the baseline signal inevery ROI (Table 1), indicating that in normal mice, theBOLD response to nicotine occurs as a function of baselinelevels, independently of the region of interest considered, asillustrated on Fig. 4. By contrast, in β2−/− mice, nicotine-evoked signal was correlated to the baseline signal only inthe thalamus (Table 1).
Quite intriguingly, the BOLD signal in WT mice showedhigh positive correlation between any ROI, either whenbaseline signal is considered or when nicotine-evokedsignal is considered (not shown). This observation suggeststhat for WT mice, a low or a high baseline activity reflects a
general feature of the individual mouse, independently ofthe brain area analyzed, with nicotine producing a BOLDsignal increase as a constant proportion of the baseline. Bycontrast, in the β2−/− mice, only two pairs of brain regionsshowed significantly correlated BOLD baseline signals: theprelimbic area and the anterior frontal cortex on the onehand, and the thalamus and the amygdala on the other hand.These correlations became stronger under nicotine exposure(Table 2). More interestingly and unlike what was observedin WT mice, nicotine exposure in β2−/− mice triggeredpositive correlations (Table 2) between the BOLD signalrecorded in several pairs of brain region such as the VTAand the amygdala, the VTA and the nucleus accumbens,the VTA and the prelimbic cortex, the prelimbic cortexand the amygdala, the prelimbic cortex and the thalamus,and the nucleus accumbens and the amygdala (see Fig. 5),all of which pairs are linked functionally and anatomically.
Discussion
Three major findings were obtained in these experiments.First, our results show that nicotine administration in WTmice produces a localized BOLD response in cortical andsubcortical brain areas. The observation and quantificationof this response in the mouse suggests that fMRI is a usefulapproach to investigate the pharmacology and genetics ofthe nicotine-elicited brain activation. Second, we show thata continuous low-dose nicotine injection prevents the dropin BOLD signal observed during prolonged anesthesia.Third, the nicotine-elicited BOLD response is largelymediated by β2*nAChRs.
A bolus of nicotine given in the general circulationproduces both localized dopaminergic activation (e.g.,Champtiaux et al. 2003; Marubio et al. 2003; Maskos etal. 2005; Mameli-Engvall et al. 2006) and distributedeffects such as the release of other neurotransmitters, thesynthesis of transcription factors (Hiremagalur and Sabban1995; Fu et al. 2003; Rossi et al. 2005), the consumption of
Table 1 Statistical correlation between the nicotine-evoked BOLDsignal and the baseline BOLD signal in WT and β2 KO mice
Regions of interest WT β2 KO
Prelimbic cortex r=0.99, p<0.0001 r=0.61, p=0.15, NSAnterior frontal cortex r=0.99, p<0.0001 r=0.72, p=0.07, NSSomatosensorial cortex r=0.99, p<0.0001 r=0.71, p=0.08, NSM1 motor cortex r=0.99, p<0.0001 r=0.70, p=0.08, NSNucleus accumbens r=0.98, p<0.0002 r=0.29, p=0.53, NSVentral tegmental area r=0.99, p<0.0001 r=0.1, p=0.83, NSSubstantia nigra r=0.99, p<0.0001 r=0.48, p=0.27, NSThalamus r=0.98, p<0.0002 r=0.85, p=0.02Basolateral amygdala r=0.99, p<0.0001 r=0.84, p=0.02
Psychopharmacology

glucose (London et al. 1988; Marenco et al. 2000) showinga nicotine-evoked stimulation of neural activation, andbrain metabolism. However, the nicotine-stimulating effectis exerted both by β2*nAChRs and α7 subtypes expressedon GABAergic neurons and glutamatergic nerve endings,but also and mostly at nonsynaptic compartments, and canincrease the influx of both Na+ and Ca2+ (Pakkanen et al.2005). Thus, the regulation of nicotine brain activation islikely to be complex.
Current approaches for the study of nicotine’ actionallow fine temporal resolution, but in very localized areas(electrophysiology, microdialysis). fMRI has the potentialto capture a full image of nicotine neuromodulatoryactivity, likely to implicate different brain circuits, due tothe location of the various nicotinic receptors (Le Novère etal. 2002).
The main findings of our study are that acute nicotineadministration significantly increased the relative BOLD
Table 2 Evolution of the relationship of BOLD signal between couples of regions after nicotine exposure in β2 KO mice
Couples of regions of interest Baseline session Nicotine session
Prelimbic cortex and anterior frontal cortex r=0.90, p=0.006* r=0.93, p=0.002*Nucleus accumbens and ventral tegmental area r=0.051, p=0.91, NS r=0.80, p=0.03*Nucleus accumbens and substantia nigra r=0.04, p=0.93, NS r=0.19, p=0.68, NSSubstantia nigra and ventral tegmental area r=0.75, p=0.05* r=0.10, p=0.83, NSPrelimbic cortex and ventral tegmental area r=0.17, p=0.71, NS r=0.83, p=0.02*Prelimbic cortex and amygdala r=0.05, p=0.91, NS r=0.78, p=0.04*Ventral tegmental area and amygdala r=0.17, p=0.72, NS r=0.86, p=0.01*Nucleus accumbens and amygdala r=0.60, p=0.15, NS r=0.93, p=0.002*Thalamus and amygdala r=0.89, p=0.007* r=0.99, p<0.0001*Thalamus and prelimbic area r=0.21, p=0.65, NS r=0.81, p=0.03*
* a significant correlation between the BOLD activation of both regions of interest
Fig. 4 Plots and correlation coefficient between BOLD signal changes recorded during the first injection (saline, x axis) and the second injection(nicotine, y axis) for WT (white square) and β2 KO (black square) mice
Psychopharmacology
signal in several cortical regions in anesthetized mice, inparticular, the prelimbic, the anterior frontal and thesomatosensorial cortex, and in subcortical regions such asthe ventral tegmental area, the substantia nigra, the nucleusaccumbens, and the thalamus, whereas the motor cortex andthe amygdala were not significantly activated.
Two recent imaging studies conducted in rats (Gozzi etal. 2006; Choi et al. 2006) showed that a dose similar to theone used in our study (1 and 0.07 mg/kg, respectively)produced the strongest increase in cerebral blood flow inthe prefrontal cortex and a more discrete increase in thenucleus accumbens and the thalamus. Interestingly, thepercentage of increase in cerebral blood flow elicited bynicotine at that dose was in the same range as the one weobtained for the BOLD signal, i.e., between 5% and 20%.Therefore, our present data, as well as the rat data obtainedpreviously (see also Calderan et al. 2005 for sub-chronicexposure), are consistent with BOLD imaging studiesconducted in awake human subjects, which showed anincreased BOLD activation triggered by nicotine in thefrontal cortex, the accumbens, and the thalamus (Stein et al.
1998). However, these studies showed no cerebral bloodflow increase in the midbrain tegmentum, whereas our datashowed a significant increase in both the ventral tegmentalarea and the substantia nigra. This difference may be due tothe fact that we did not measure the time course of theBOLD signal but captured stable nicotine-elicited changes.Effects of partial volume may also have compromised thedetection of CBF changes in the VTA in earlier imagingstudies. Therefore, our data are consistent with the electro-physiological recordings showing an increased firing rategenerated by nicotine injection in the VTA (Mameli-Engvallet al. 2006). Although it is not documented, we cannotexclude that the effects we observed, which encompassed aninteraction between anesthesia and nicotine.
Anesthetized mice receiving no nicotine injectionexhibited a significant and major decrease of activation inall cortical and subcortical structures over time, likely dueto prolonged (2 h) anesthesia. Outside the scanner, theeffect of a 2-h ketamine/xylazine anesthesia on physiolog-ical parameters such as oxygen saturation, temperature,respiratory frequency, and cardiac pulses was measured
Fig. 5 Plots and correlation coefficient for BOLD signal after nicotine or saline injection between three ROIs (top nucleus accumbens, middleprelimbic, and bottom basolateral amygdala, x axis) and the ventral tegmental area (y axis) for WT (left) and β2 KO (right) mice
Psychopharmacology

every 30 min. Ketamine acts mainly as glutamate-NMDA,a dopamine-D2-receptor antagonist, and a partial D2agonist (Kapur and Seeman 2002). Anesthetic doses ofketamine in humans produce a dissociative state, in whichdisconnection of thalamocortical and corticocortical net-works leads to unconsciousness (Honey et al. 2005).Xylazine is an alpha2-adrenoceptor agonist with analgesicand sedative properties and has long been in veterinary use(Clarke and Hall 1969), and ketamine has been reported notto affect cerebral activity (Small et al. 2000).
Behavioral studies show that nicotine enhances arousalin WT but not in β2−/− mice, and β2*nAChRs areinvolved in the regulation of normal sleep (Léna et al.2004). It is therefore possible that an injection of nicotine inanesthetized mice causes arousal, and if β2*nAChRs arepresent, leading in turn to increased respiratory rate, betterblood oxygenation, and consequently, alterations in theBOLD signal. BOLD signal is the result of alteration inflow and blood volume and oxyhemoglobin concentrationand has been shown to reflect either direct changes inneural activity (Logothetis et al. 2001) or in afferent activity(Vafaee and Gjedde 2004). Activation of nAChRs receptorshas been previously shown to be involved in vasodilatationin the neocortex and hippocampus, via the excitation ofcholinergic fibers originating in the basal forebrain (forreview, see Sato and Sato 1992). Vasodilatation maytherefore increase significantly the regional BOLD signaltriggered by nicotine. However, the significant regionalnicotine-elicited activation we observe is unlikely to resultonly from a general and unspecific increase of cerebralvasculature, as some brain regions were not significantlyactivated by nicotine (see also Gozzi et al. 2006 fordiscussion).
The comparison between the BOLD signal obtained afternicotine injection and the one obtained without nicotinetherefore revealed an additive effect of nicotine over theeffect of anesthetics on brain activity. Two recent studiesshowed that general anesthesia with alpha-chloralose, aGABAA receptor agonist, produces a general and signifi-cant decrease of brain activity in rats (Luo et al. 2007; Chinet al. 2008) and interferes with the binding of somenicotinic agents (Chin et al. 2008) and with ethanolchallenge (Luo et al. 2007), although the ketamine/xylazinecombination and nicotine itself that we both used in thepresent study were not specifically tested. However, BOLDfMRI activation maps in anesthetized rats correspond tostaining for c-Fos derived from conscious animals, at least forareas that display positive BOLD signal (Stark et al. 2006).
In anesthetized rats, nicotine increases cortical cerebralblood flow, and this increase is abolished by blockade ofnicotinic, but not muscarinic, receptors in the brain (Uchidaet al. 1997). β2−/− mice showed no increased activation,either cortical or subcortical, after nicotine injection. How-
ever, regional co-activation concerning mainly the rewardsystem was evoked by nicotine, specifically in β2−/− mice.These data reveal that acute nicotine administration producescortical and subcortical activation, largely mediated byβ2*nAChRs. In WT mice, the baseline BOLD signalpredicts the dynamic range of the nicotine response. As thisprediction is largely not verified in β2 KO mice, we canpropose that it normally relies on β2*nAChRs. When thesereceptors are absent, however, nicotine may partially restorethe co-activation between directly connected brain regions,via non β2*nAChRs. By exclusion, this effect is likelymediated by α7-nAChRs (Besson et al. 2007). Nicotine-mediated co-activation has been studied only in slicepreparations and therefore refers to local circuits (LeMagueresse et al. 2006). Our present data reinforce andcomplete our previous behavioral and electrophysiologicaldata (Besson et al. 2007, 2008) showing that α7 receptorscontribute to such nicotine-mediated co-activation. Usingviral vector-mediated restoration, we demonstrated previous-ly that β2-containing receptors are involved in the acetyl-choline-mediated co-activation in the absence of nicotine(Maskos et al. 2005). Although the existence of suchnicotinic-mediated synchronization remains to be directlyshown, it has been recently evidenced that nicotinic receptor-mediated transmission contributes significantly to evokinglocal release of Ca2+ from internal stores in cell dendrites(Girod et al. 2003; Lohmann et al. 2002), a process involvedin synaptic plasticity and long-distance cellular communica-tion (Deisseroth et al. 1998). It must be noticed thatcorrelation between BOLD signals from different brain areashas been performed, to our knowledge, only once (Iidaka etal. 2001) and was interpreted as an index of the functionalco-operation between connected brain areas. This should befurther investigated, for example, using electrophysiologicalrecordings in multiple sites after nicotine administration. Ourdata may have implications in nicotine addiction, becausethe role of alpha7-containing receptors, unmasked after achronic exposure to the drug (Besson et al. 2007), is alsoshown here after a singly acute exposure by altering thesynchronization between different brain areas. Such process-es of synchronization, altered by β2-subunit deletion andrestored after nicotine exposure, might be part of the globalneuromodulatory effect produced by nicotine.
Our current procedure paves the way for further BOLDactivation studies in mice, and together with pharmacolog-ical manipulations, may provide unique information aboutthe neurotransmitters implicated in nicotine-elicited brainregional activation.
Acknowledgments This research was supported by the Associa-tion pour la Recherche sur les Nicotinanées, which provided a grantto S. Granon and a post-doc grant for S. Suarez. S. Granon was alsosupported by an ANR grant. D. LeBihan was supported by a grantfrom the cancéropôle d’Ile-de-France. The Pasteur Institute (Paris),
Psychopharmacology

the CNRS, and the CEA also contributed to this work. The authorsthank Gilles Bonvento and Vincent Lebon for their help in the setupof the experiments; Uwe Maskos, Philippe Faure, and ArnaudCressant for comments on the manuscript; and two anonymousreferees for their constructive reading and suggestions.
References
Besson M, David V, Suarez S, Cormier A, Cazala P, Changeux J-P,Granon S (2006) Genetic dissociation of two behaviors associ-ated with nicotine addiction: Beta-2 containing nicotinic recep-tors are involved in nicotine reinforcement but not in withdrawalsyndrome. Psychopharmacology (Berlin) 187:189–199
Besson M, Granon S, Mameli-Engvall M, Cloëz-Tayarani I,Maubourguet N, Cormier A, Cazala P, David V, Changeux JP,Faure P (2007) Long-term effects of chronic nicotine exposureon brain nicotinic receptors. Proc Natl Acad Sci USA 104:8155–8160
Besson M, Suarez SV, Cormier A, Changeux J-P, Granon S (2008)Chronic nicotine exposure has dissociable behavioural effects oncontrol and beta2−/− mice. Behav Genet (in press)
Calderan L, Chiamulera C, Marzola P, Fabene PF, Fumagalli GF,Sbarbati A (2005) Sub-chronic nicotine-induced changes inregional cerebral blood volume and transversal relaxation timepatterns in the rat: a magnetic resonance study. Neurosci Lett377:195–199
Champtiaux N, Changeux JP (2004) Knockout and knockin mice toinvestigate the role of nicotinic receptors in the central nervoussystem. Prog Brain Res 145:235–251, Review
Champtiaux N, Gotti C, Cordero-Erausquin M, David DJ, Przybylski C,Léna C, Clementi F, Moretti M, Rossi FM, Le Novère N,McIntosh JM, Gardier AM, Changeux JP (2003) Subunitcomposition of functional nicotinic receptors in dopaminergicneurons investigated with knock-out mice. J Neurosci 23:7820–7829
Changeux JP, Edelstein SJ (2005) Allosteric mechanisms of signaltransduction. Science 308:1424–1428
Chin C-L, Pauly JR, Surber BW, Skoubis PD, McGaraughty S,Hradil VP, Luo Y, Cox BF, Fox GB (2008) PharmacologicalMRI in awake rats predicts selective binding of α4β2 nicotinicreceptors. Synapse 62:159–168
Choi K-J, Mandeville JB, Chen IY, Kim YR, Jenkins BG (2006) Highresolution spatial mapping of nicotine action using pharmaco-logic magnetic resonance imaging. Synapse 60:152–157
Clarke KW, Hall LW (1969) “Xylazine”—a new sedative for horsesand cattle. Vet Rec 85:512–517
Deisseroth K, Heist EK, Tsien RW (1998) Translocation of calmod-ulin to the nucleus supports CREB phosphorylation in hippo-campal neurons. Nature 392:198–202
Devillers-Thiéry A, Bourgeois JP, Pons S, Le Sourd A, Pucci B,Changeux JP (2003) An in vitro study of the sub-cellulardistribution of nicotinic receptors. Biol Cell 95:373–381
Di Chiara G (2000) Role of dopamine in the behavioural actions ofnicotine related to addiction. Eur J Pharmacol 393:295–314
Fallon S, Shearman E, Sershen H, Lajtha A (2007) The effects ofglutamate and GABA receptor antagonists on nicotine-inducedneurotransmitter changes in cognitive areas. Neurochem Res32:535–553
Fu Y, Matta SG, Kane VB, Sharp BM (2003) Norepinephrine releasein amygdala of rats during chronic nicotine self-administration:an in vivo microdialysis study. Neuropharmacology 45:514–523
Gäddnäs H, Pietilä K, Ahtee L (2000) Effects of chronic oral nicotinetreatment and its withdrawal on locomotor activity and brainmonoamines in mice. Behav Brain Res 113:65–72
Girod R, Jareb M, Moss J, Role L (2003) Mapping of presynapticnicotinic acetylcholine receptors using fluorescence imaging ofneuritic calcium. J Neurosci Methods 122:109–122
Gotti C, Clementi F (2004) Neuronal nicotinic receptors: fromstructure to pathology. Prog Neurobiol 74:363–396
Gotti C, Moretti M, Gaimarri A, Zanardi A, Clementi F, Zoli M(2007) Heterogeneity and complexity of native brain nicotinicreceptors. Biochem Pharmacol 74:1102–1111
Gozzi A, Schwarz A, Reese T, Bertani S, Crestan V, Bifone A (2006)Region-specific effects of nicotine on brain activity: a pharma-cological MRI study in the drug-naïve rat. Neuropsychopharma-cology 31:1690–1703
Granon S, Faure P, Changeux JP (2003) Executive and socialbehaviors under nicotinic receptor regulation. Proc Natl AcadSci USA 100:9596–9601
Han ZY, Zoli M, Cardona A, Bourgeois JP, Changeux JP, Le Novère N(2003) Localization of [3H]nicotine, [3H]cytisine, [3H]epibatidine,and [125I]alpha-bungarotoxin binding sites in the brain of Macacamulatta. J Comp Neurol 461:49–60
Heishman SJ, Taylor RC, Henningfield JE (1994) Nicotine andsmoking: a review of effects on human performance. Exp ClinPsychopharmacol 2:345–395
Hiremagalur B, Sabban EL (1995) Nicotine elicits changes inexpression of adrenal catecholamine biosynthetic enzymes,neuropeptide Y and immediate early genes by injection but notcontinuous administration. Brain Res Mol Brain Res 32:109–115
Honey GD, Honey RA, Sharar SR, Turner DC, Pomarol-Clotet E,Kumaran D, Simons JS, Hu X, Rugg MD, Bullmore ET,Fletcher PC (2005) Impairment of specific episodic memoryprocesses by sub-psychotic doses of ketamine: the effects oflevels of processing at encoding and of the subsequent retrievaltask. Psychopharmacology 181:445–457
Iidaka T, Omori M, Murata T, Kosaka H, Yonekura Y, Okada T,Sadato N (2001) Neural interaction of the amygdala with theprefrontal and temporal cortices in the processing of facialexpressions as revealed by fMRI. J Cog Neurosci 13:1035–1047
Kapur S, Seeman P (2002) NMDA receptor antagonists ketamine andPCP have direct effects on the dopamine D(2) and serotonin 5-HT(2)receptors—implications for models of schizophrenia. MolPsychiatry 7:837–844
Le Magueresse C, Safiulina V, Changeux JP, Cherubini E (2006)Nicotinic modulation of network and synaptic transmission in theimmature hippocampus investigated with genetically modifiedmice. J Physiol 576:533–546
Léna C, Popa D, Grailhe R, Escourrou P, Changeux JP, Adrien J(2004) Beta2-containing nicotinic receptors contribute to theorganization of sleep and regulate putative micro-arousals inmice. J Neurosci 24:5711–5718
Le Novère N, Corringer PJ, Changeux JP (2002) The diversity of subunitcomposition in nAChRs: evolutionary origins, physiologic andpharmacologic consequences. J Neurobiol 53:447–456, Review
Li X, Eisenach JC (2002) Nicotinic acetylcholine receptor regulationof spinal norepinephrine release. Anesthesiology 96:1450–1456
Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A (2001)Neurophysiological investigation of the basis of the fMRI signal.Nature 412:150–157
Lohmann C, Myhr KL, Wong ROL (2002) Transmitter-evoked localcalcium release stabilizes developing dendrites. Nature 418:177–181
London ED, Connolly RJ, Szikszay M, Wamsley JK, Dam M (1988)Effects of nicotine on local cerebral glucose utilization in the rat.J Neurosci 8:3920–3928
Luo F, Li X, Treistman SN, Kim YR, King JA, Fox GB, Ferris CF(2007) Confounding effects of volatile anesthesia on CBVassessment in rodent forebrain following ethanol challenge. JMagnetic Res Imaging 26:557–563
Psychopharmacology

Mameli-Engvall M, Evrard A, Pons S, Maskos U, Svensson TH,Changeux JP, Faure P (2006) Hierarchical control of dopamineneuron-firing patterns by nicotinic receptors. Neuron 50:911–921
Marenco T, Bernstein S, Cumming P, Clarke PB (2000) Effects ofnicotine and chlorisondamine on cerebral glucose utilization inimmobilized and freely-moving rats. Br J Pharmacol 129:147–155
Marks MJ, Romm E, Bealer SM, Collins AC (1985) A test battery formeasuring nicotine effects in mice. Pharmacol Biochem Behav23:325–330
Marubio LM, Gardier AM, Durier S, David D, Klink R, Arroyo-Jimenez MM, McIntosh JM, Rossi F, Champtiaux N, Zoli M,Changeux JP (2003) Effects of nicotine in the dopaminergicsystem of mice lacking the alpha4 subunit of neuronal nicotinicacetylcholine receptors. Eur J Neurosci 17:1329–1337
Maskos U, Molles BE, Pons S, Besson M, Guiard BP, Guilloux J-P,Evrard A, Cazala P, Cormier A, Mameli-Engvall M, Dufour N,Cloëz-Tayarani I, Bemelmans A-P, Mallet J, Gardier AM, David V,Faure P, Granon S, Changeux J-P (2005) Nicotine reinforcementand cognition restored by targeted expression of nicotinicreceptors. Nature 436:103–107
Matta SG, Balfour DJ, Benowitz NL, Boyd RT, Buccafusco JJ,Caggiula AR, Craig CR, Collins AC, Damaj MI, Donny EC,Gardiner PS, Grady SR, Heberlein U, Leonard SS, Levin ED,Lukas RJ, Markou A, Marks MJ, McCallum SE, Parameswaran N,Perkins KA, Picciotto MR, Quik M, Rose JE, Rothenfluh A,Schafer WR, Stolerman IP, Tyndale RF, Wehner JM, Zirger JM(2007) Guidelines on nicotine dose selection for in vivo research.Psychopharmacology (Berlin) 190:269–319
Newhouse PA, Potter A, Singh A (2004) Effects of nicotinic stimulationon cognitive performance. Curr Opin Pharmacol 4:36–46
Pakkanen JS, Jokitalo E, Tuominen RK (2005) Up-regulation of beta2and alpha7 subunit containing nicotinic acetylcholine receptors inmouse striatum at cellular level. Eur J Nuerosci 21(10):2681–2691
Pauly JM, Le Roux P, Nishimura DG, Macovski A (1991) Parameterrelations for the Shinnar-Le Roux RF pulse design algorithm.IEEE Trans Med Imag 10:53–65
Paxinos G, Franklin KBJ (2004) The mouse brain in stereotaxiccoordinates, 2nd edn. Elsevier Academic, San Diego
Picciotto MR, Zoli M, Lena C, Bessis A, Lallemand Y, Le Novere N,Vincent P, Pich EM, Brulet P, Changeux JP (1995) Abnormalavoidance learning in mice lacking functional high-affinitynicotine receptor in the brain. Nature 374:65–67
Picciotto MR, Zoli M, Rimondini R, Léna C, Marubio LM, Pich EM,Fuxe K, Changeux JP (1998) Acetylcholine receptors containingthe beta2 subunit are involved in the reinforcing properties ofnicotine. Nature 391:173–177
Pietila K, Ahtee L (2000) Chronic nicotine administration in thedrinking water affects the striatal dopamine in mice. PharmacolBiochem Behav 66:95–103
Rivière D, Papadopoulos-Orfanos D, Régis J, Mangin J-F (2000) Astructural browser of brain anatomy. NeuroImage HBM 11:560
Rivière D, Régis J, Cointepas Y, Papadopoulos-Orfanos D, Cachia A,Mangin J-F (2003) A freely available Anatomist/BrainVISApackage for structural morphometry of the cortical sulci. Neuro-Image HBM 19:934
Rossi S, Singer S, Shearman E, Sershen H, Lajtha A (2005) Theeffects of cholinergic and dopaminergic antagonists on nicotine-induced cerebral neurotransmitter changes. Neurochem Res30:541–558
Sato A, Sato Y (1992) Regulation of regional cerebral blood flow bycholinergic fibers originating in the basal forebrain. Neurosci Res14:242–274
Small SA, Wu EX, Bartsch D, Perera GM, Lacefield CO, DeLaPaz R,Mayeux R, Stern Y, Kandel ER (2000) Imaging physiologicdysfunction of individual hippocampal subregions in humans andgenetically modified mice. Neuron 28:653–664
Stark JA, Davies KE, Williams SR, Luckman SM (2006) Functionalmagnetic resonance imaging and c-Fos mapping in rats followingan anorectic dose of m-chlorophenylpiperazine. NeuroImage31:1228–1237
Stein EA (2001) fMRI: a new tool for the in vivo localization of drugactions in the brain. J Analyt Toxicol 25:419–424
Stein EA, Pankiewicz J, Harsch HH, Cho JK, Fuller SA, Hoffmann RG,Hawkins M, Rao SM, Bandettini PA, Bloom AS (1998) Nicotine-induced limbic cortical activation in the human brain: a functionalMRI study. Am J Psychiatry 155:1009–1015
Stolerman IP, Fink R, Jarvik ME (1973) Acute and chronic toleranceto nicotine measured by activity in rats. Psychopharmacologia30:329–342
Uchida S, Kagitani F, Nakayama H, Sato A (1997) Effect ofstimulation of nicotinic cholinergic receptors on cortical cerebralblood flow and changes in the effect during aging in anesthetizedrats. Neurosci Lett 228:203–206
Vafaee and Gjedde 2004 (2004) Spatially dissociated flow-metabolismcoupling in brain activation. Neuroimage 21:507–515
Valette H, Dollé F, Saba W, Roger G, Hinnen F, Coulon C, Ottaviani M,Syrota A, Bottlaender M (2007) [18F]FPhEP and [18F]F2PhEP,two new epibatidine-based radioligands: evaluation for imagingnicotinic acetylcholine receptors in baboon brain. Synapse 61:764–770
Villégier AS, Salomon L, Blanc G, Godeheu G, Glowinski J, Tassin JP(2006) Irreversible blockade of monoamine oxidases reveals thecritical role of 5-HT transmission in locomotor response inducedby nicotine in mice. Eur J Neurosci 24:1359–1365
Xu J, Mendrek A, Cohen MS, Monterosso J, Simon S, Jarvik M,Olmstead R, Brody AL, Ernst M, London ED (2007) Effect ofcigarette smoking on prefrontal cortical function in nondeprivedsmokers performing the stroop task. Neuropsychopharmacology32:1421–1428
Psychopharmacology
Copyright © 2022 FDOKUMEN




















