Identification of N-Acetylgalactosamine in Carbohydrates of Xenopus laevis Testis
Upload
independentCategory
view
3download
0
Ba
DYDa
b
c
d
T
a
ARRAA
KB1LRS
1
htraeio
Nc
tU
0d
Toxicology Letters 194 (2010) 16–25
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
isphenol A may cause testosterone reduction by adversely affecting both testisnd pituitary systems similar to estradiol
aichi Nakamuraa, Yukie Yanagibaa, Zhiwen Duana, Yuki Itoa,1, Ai Okamuraa, Nobuyuki Asaedab,oshiaki Tagawab, ChunMei Li c,d, Kazuyoshi Tayac,d, Shu-Yun Zhanga,2, Hisao Naitoa,oni Hikmat Ramdhana, Michihiro Kamijimaa,1, Tamie Nakajimaa,∗
Department of Occupational and Environmental Health, Nagoya University Graduate School of Medicine, 65 Tsuruma-cho, Showa-ku, Nagoya 466-8550, Aichi, JapanSafety Assessment Laboratory, Sanwa Kagaku Kenkyusho Co., Ltd., 363 Shiosaki Hokusei, Inabe Mie 511-0406, JapanDepartment of Basic Veterinary Science, The United Graduate School of Veterinary Sciences, Gifu University, 1-1 Yanagido, Gifu 501-1193, JapanLaboratory of Veterinary Physiology, Department of Veterinary Medicine, Faculty of Agriculture, Tokyo University of Agriculture and Technology, 3-5-8 Saiwai-cho, Fuchu,okyo 183-8509, Japan
r t i c l e i n f o
rticle history:eceived 2 November 2009eceived in revised form 1 February 2010ccepted 1 February 2010vailable online 6 February 2010
eywords:isphenol A7�-Estradiol
a b s t r a c t
Bisphenol A (BPA) causes reproductive toxicities, but the mechanisms are still unclear. In the presentstudy, we sought to clarify these mechanisms in comparison with those of 17�-estradiol (E2). PrepubertalWistar/ST male rats (4 weeks old) were subcutaneously administered BPA (0, 20, 100 and 200 mg/kg/day)or E2 (10 and 100 �g/kg/day) for 6 weeks. Both BPA and E2 treatments decreased plasma and testiculartestosterone levels, and plasma luteinizing hormone (LH), but not E2 and follicle-stimulating hormonelevels, though E2 treatment increased its plasma level. In relation to the decreased testosterone levels,BPA and E2 decreased expressions of steroidogenic enzymes and cholesterol carrier protein in Leydigcells. Thus, decreased testosterone levels in plasma might have resulted from decreased expressions
eydig cellseproductive toxicityteroidogenesis
of these enzymes and protein as well as from decreased plasma LH levels. Interestingly, the changesin steroidogenic enzymes and carrier protein were observed at lower levels of exposure to BPA or E2than those inhibiting plasma LH levels. Microscopically, 200 mg/kg BPA and 100 �g/kg E2 significantlydecreased Leydig cell numbers in the testis. In addition, BPA and E2 also decreased expression of estrogenreceptor �-mRNA, which might be related to the decreased numbers of Leydig cells. Thus, BPA directlyaffects not only the Leydig cells but also the pituitary gland, but the former may be impaired at lower
than
exposure concentrations. Introduction
Effects of exogenous endocrine disruptors on human healthave been of great interest. These substances are thoughto cause adverse effects on development or function ofeproductive organs by binding to hormone receptors as
ndrogen or estrogen mimics (Matthews et al., 2001; Xut al., 2005). Especially, exposure to these chemicals dur-ng developing stages may cause dysfunctions or abnormalitiesf reproductive organs later at adult stages, which may∗ Corresponding author. Tel.: +81 52 744 2122; fax: +81 52 744 2126.E-mail address: [email protected] (T. Nakajima).
1 Current address: Department of Occupational and Environmental Health,agoya City University Graduate School of Medical Sciences, 1 Kawasumi, Mizuho-ho, Mizuho-ku, Nagoya 467-8601, Japan.
2 Current address: Laboratory of Pharmacology, National Institute of Environmen-al Health Sciences, National Institutes of Health, Research Triangle Park, NC 27709,SA.
378-4274/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.toxlet.2010.02.002
the latter.© 2010 Elsevier Ireland Ltd. All rights reserved.
lead to infertility or development of cancer (Timms et al.,2005).
BPA is thought to be a probable endocrine disruptor, becausethis chemical promoted proliferation of estrogen-responsive MCF-7 breast cancer cells in an in vitro system (Krishnan et al.,1993). This chemical is widely used in the manufacture of var-ious polycarbonate plastics, which are used in lacquer coatingsin food cans, food and beverage containers and dental sealants(Brotons et al., 1995; Olea et al., 1996). Thus, unpolymerised-BPA monomer or dimer including these materials is liquated intofoods and beverages by heating the cans or containers, which maybe absorbed by the human body. Indeed, 4–23 �g/can BPA wasdetected from canned food, 7–58 �g/g from canned beverages,and 90–913 �g/saliva one hour after application of certain dental
sealants (Brotons et al., 1995; Olea et al., 1996). In addition, BPA wasdetected to be 0.3–18.9 ng/ml, 8.3–8.7 ng/ml, and 1.0–104.9 ng/mlin serum, amniotic fluid and placental tissue of pregnant women,respectively. This chemical was also detected (0.2–9.2 ng/ml) infetal serum (Ikezuki et al., 2002; Schonfelder et al., 2002). Thus,
D. Nakamura et al. / Toxicology Letters 194 (2010) 16–25 17
Table 1Primers used for real-time PCR to quantify mRNA expression of biomolecules involved in testicular steroidogenesis.
Gene Accession no. Forward primer (5′–3′) Reverse primer (5′–3′)
HMG-CoA synthase X52625.1 CACAGCCGCAGTCTTCAATG TGGCGTTTCCTGAGGCATAHMG-CoA reductase BC064654.1 ACATCCGTCTCCAGTCCAAAA CAGGTTTCTTGTCGGTGCAAPBR NM 012515.1 GGCTTCATGGGAGCCTACTTT CAGGAGGGTTTCTGCAAGCTAStAR BC088859.1 CAGGTGTGGGCAAGGTGTTC TGCGGTCCACCAGTTCTTCP450scc J05156.1 ATCCAGCTTCTTTCCCAATCC AAAGCCCAAGTACCGGAAGTG3�-HSD (type I) M38178.1 CCGCTGCTGTCATTGATGTCT CGCAGGCCTCCAATATGTTCP45017� NM 012753.1 GCCCCAGATGGTGACTCAAA GGAGCTACCAGCATCTGCAAA17�-HSD (type III) AF035156.1 CCAGGTGCTGACCCCTTATTC ACTCATCGGCGGTCTTGGTAromatase NM 017085.1 ATGCTGAACCCCATGCACTAT AGGCCCATGATCAGCAGAAGT
ACACATGGGAGA
tr
at2et1dvdsepp
ormtaB
dcpwt
2
2
of
2
opwrh
2
s6cft
nology, Inc. (CA, USA). P450 side-chain cleavage enzyme (P450scc) polyclonalantibody was purchased from CHEMICON International Inc. (CA, USA). After block-ing with 3% skim milk, membranes were incubated with the primary antibodies
AR NM 012502.1ER� NM 012689.1ER� NM 012754.1GAPDH AB017801.1
hese results all suggest that the risk assessment of BPA is war-anted.
In laboratory animals, BPA induced an estrous cycle disordernd an increase in estrogen receptor-� (ER�) mRNA expression inhe hypothalamus in female rats (Khurana et al., 2000; Rubin et al.,001). Moreover, BPA administration induced cell proliferation ofpithelia of uterus and vagina, and hyperprolactinemia in ovariec-omized Fischer 344 rats (Khurana et al., 2000; Steinmetz et al.,997, 1998). On the other hand, in male animals, BPA treatmentecreased the weights of prostate, testes, epididymal and seminalesicle (Akingbemi et al., 2004; Takahashi and Oishi, 2003), and alsoecreased levels of luteinizing hormone (LH) and testosterone, andperm counts (Akingbemi et al., 2004; Herath et al., 2004; Rubint al., 2001; Welshons et al., 1999). In addition, BPA-induced mor-hological abnormality of ducts and fibroblast proliferation in therostate (Gupta, 2000; Ramos et al., 2001, 2003; Timms et al., 2005).
Because these abnormalities observed in the reproductivergans by BPA exposure are very similar to those of androgeneceptor (AR) knockout mice (Snyder et al., 2000), we measured AR-RNA in the testes and prostate in the BPA-treated rats. However,
here was no difference in the AR-mRNA levels between controlnd BPA-treated rats (unpublished data). Thus, the mechanism ofPA-induced reproductive toxicity remains known.
Here, we investigated the mechanism of BPA-induced repro-uctive toxicity, especially the decrease in testosterone levels, andompared with that caused by 17�-estradiol (E2), and also com-ared the reproductive organ-related functions in 10-week-old ratsith those of the baseline stage (at 4 weeks), the age at which BPA
reatment of rats was started.
. Materials and methods
.1. Chemicals
BPA (purity, >99%) and E2 were purchased from Tokyo Kasei (Tokyo, Japan). Cornil and other chemicals used in pathological fixation, and staining were purchasedrom Wako (Osaka, Japan).
.2. Animals
This study was conducted according to the Guidelines for Animal Experimentsf the Nagoya University Animal Center. Wistar/ST male rats (3 weeks old) wereurchased from Japan SLC (Shizuoka, Japan) and were housed in the animal centerith a 12 h light/dark cycle under a controlled temperature of 23–25 ◦C and 57–60%
elative humidity for 1 week to accommodate them to the new surroundings. Theyad free access to food (CE2, CLEA Japan Inc., Japan) and tap water.
.3. Treatments
Rats were randomly divided into seven groups (n = 8–9), and BPA and E2 wereubcutaneously injected 4 days (Monday, Tuesday, Thursday and Friday) a week forweeks (20, 100 and 200 mg BPA/kg/day; 10 and 100 �g E2/kg/day). These dosages
orresponded to 11.4, 57.1 and 114.2 mg/kg/day for BPA, and 5.7 and 57.1 �g/kg/dayor E2, when they were given BPA or E2 every day. In this experiment, there werewo control groups: one was rats sacrificed at 4 weeks old, that is, at the start
GTACCAGGGACCACGTTTT CCACAGATCAGGCAGGTCTTCTGACCCTTCACACCAAAGC CCAGCTCGTTCCCTTGGATTGATTGCGAAGAGTGGTA GCCTGACGTGAGAAAGAAGCAACATCATCCCTGCATCCA CCGTTCAGCTCTGGGATGAC
of BPA or E2 treatments, for investigation of the reproductive system at baseline(baseline control group); another was given a vehicle (corn oil) in the same wayas BPA or E2 (experimental control group). Sixteen hours after the last dosages,all rats except baseline controls were weighed and sacrificed by decapitation. Theblood was collected into heparinized tubes. The following organs were dissectedout and weighed as quickly as possible: epididymis, testes, seminal vesicle andprostate.
2.4. Hormone assay
Plasma was separated by centrifugation at 4 ◦C and stored at −80 ◦C until assayfor testosterone, LH, follicle-stimulating hormone (FSH) and E2. Testosterone andE2 levels in plasma were determined with RIA (radioimmunoassay) by SRL (Tokyo,Japan). Concentrations of LH and FSH in plasma were measured using NIDDK ratradioimmunoassay (RIA) kits (Torrance, CA, USA) for rat LH and FSH. As for theantisera, anti-rat LH-S-11 and anti-rat FSH-S-11 were used, while for iodinatedpreparation, rat LH-I-7 and FSH-I-7 were employed. For measurements of testiculartestosterone levels, a part (100 mg) of left testis was homogenized in 0.6 ml of 10 mMphosphate-buffered saline (PBS, pH 7.4). After centrifugation, testis testosterone wasthen extracted from aliquots of the supernatant (10 �l) with 2 ml of hexane–ethylicether mixture (3:2). Concentrations of testosterone in the testis were measured bya testosterone EIA kit (Cayman, Ann Arbor, USA).
2.5. Real-time quantitative PCR analysis
mRNA levels of several important enzymes, carrier proteins or receptorsinvolved in the testicular biosynthetic pathway of testosterone were measured.Total RNA was extracted using the RNeasy Mini kit (Qiagen, Germany) from theleft testis of rats dosed with BPA, E2 or vehicle only and of baseline control rats.One microgram of total RNA was reversely transcribed into cDNA in a 20 �l reactionsolution containing Oligo (dT)20 primers and SuperScriptTM III reverse transcriptase(Invitrogen, Tokyo, Japan) according to the manufacturer’s instructions. The primers(Table 1) were designed using Primer Express 1.0 software (Applied Biosystems,Foster, USA). Real-time quantitative PCR was performed in a 96-well plate using aABI PRISM 7000 Sequence Detection System (Applied Biosystems). Reactions werecarried out in a 25 �l volume containing 1× SYBR Green Master Mix (Applied Biosys-tems) and 100 nM of each forward and reverse primer. Reactions were run for 40cycles (denaturation at 95 ◦C for 15 s, annealing and extension at 60 ◦C for 1 min)following an initial 2 min step at 50 ◦C for enzyme activation and a 10 min incu-bation at 95 ◦C. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used asa housekeeping gene for data analysis. All genes of mRNA expression levels werenormalized to GAPDH expression levels.
2.6. Western blotting
Testis homogenates for all proteins adjusted to 20–30 �g protein were sub-jected to 10% SDS-PAGE and transferred to nitrocellulose membranes. The primarypolyclonal antibodies to steroidogenic acute regulatory protein (StAR), and P45017�-hydroxylase/C17–20 lyase (P45017�) were purchased from Santa Cruz Biotech-
to StAR, P450scc, and P45017� , respectively, followed by incubation with alka-line phosphatase-conjugated goat anti-rabbit IgG or donkey anti-goat IgG (JacksonImmunoResearch, West Grove, PA). 1-StepTM NBT/BCIP (Pierce Biotechnology, Rock-ford, IL) was used as the substrate of alkaline phosphatase. Each band was quantifiedby densitometry, using the Lane & Spot Analyzer version 5.0 (ATTO Corporation,Tokyo, Japan).
18 D. Nakamura et al. / Toxicology Letters 194 (2010) 16–25
Tab
le2
Bod
yan
dre
pro
du
ctiv
eor
gan
wei
ghts
inra
tsat
base
lin
e(4
wee
ks)
and
10w
eeks
.
Gro
up
Bas
elin
eco
ntr
ol(4
wee
ksol
d)
Exp
erim
enta
laco
ntr
ol(1
0w
eeks
old
)B
PA(1
0w
eeks
old
)aE2
(10
wee
ksol
d)a
20m
g/kg
100
mg/
kg20
0m
g/kg
10�
g/kg
100
�g/
kg
n9
99
98
99
Bod
yw
eigh
t(g
)99
.8±
4.78
*35
2±
24.4
355
±26
.732
5±
20.8
*30
9±
11.2
*30
4±
24.1
*27
3±
15.9
*
Test
es(g
)0.
397
±0.
035*
1.47
9±
0.05
81.
461
±0.
072
1.42
2±
0.11
21.
327
±0.
102*
1.27
9±
0.11
0*1.
013
±0.
099*
Test
es/b
.w.(
%)
0.39
8±
0.03
70.
421
±0.
025
0.41
2±
0.02
60.
430
±0.
031
0.42
4±
0.02
80.
413
±0.
030
0.36
6±
0.04
7*
Epid
idym
is(g
)0.
046
±0.
006*
0.38
6±
0.01
90.
367
±0.
024
0.35
0±
0.03
2*0.
318
±0.
031*
0.28
5±
0.04
4*0.
189
±0.
031*
Epid
idym
is/b
.w.(
%)
0.04
6±
0.00
5*0.
110
±0.
006
0.10
4±
0.00
70.
108
±0.
005
0.10
3±
0.00
90.
093
±0.
001*
0.06
7±
0.01
2*
Sem
inal
vesi
cle
(g)
0.08
0±
0.01
5*1.
190
±0.
140
1.23
8±
0.13
00.
771
±0.
12*
0.61
7±
0.17
9*0.
385
±0.
112*
0.23
9±
0.03
8*
Sem
inal
vesi
cle/
b.w
.(%
)0.
081
±0.
015*
0.33
8±
0.03
80.
351
±0.
049
0.23
5±
0.04
5*0.
198
±0.
052*
0.12
5±
0.03
0*0.
088
±0.
016*
Pros
tate
(g)
0.05
5±
0.01
6*0.
476
±0.
072
0.47
1±
0.07
40.
335
±0.
099*
0.32
0±
0.07
3*0.
167
±0.
065*
0.08
2±
0.02
6*
Pros
tate
/b.w
.(%
)0.
055
±0.
016*
0.13
6±
0.02
10.
133
±0.
019
0.10
2±
0.02
7*0.
074
±0.
023*
0.05
4±
0.01
7*0.
030
±0.
010*,
†
aEa
chgr
oup
was
give
nve
hic
le(c
orn
oil)
only
(exp
erim
enta
lcon
trol
),bi
sph
enol
A(B
PA)
or17
�-e
stra
dio
l(E2
)fo
r6
wee
ksby
subc
uta
neo
us
inje
ctio
n.
Val
ues
rep
rese
nt
mea
n±
S.D
.*
Sign
ifica
ntl
yd
iffe
ren
tfr
omex
per
imen
talc
ontr
ols
(P<
0.05
).†
Sign
ifica
nt
dec
reas
eco
mp
ared
wit
hva
lue
ofp
rost
ate/
b.w
.at
base
lin
eco
ntr
ols
(P<
0.05
).b.
w.,
bod
yw
eigh
t.
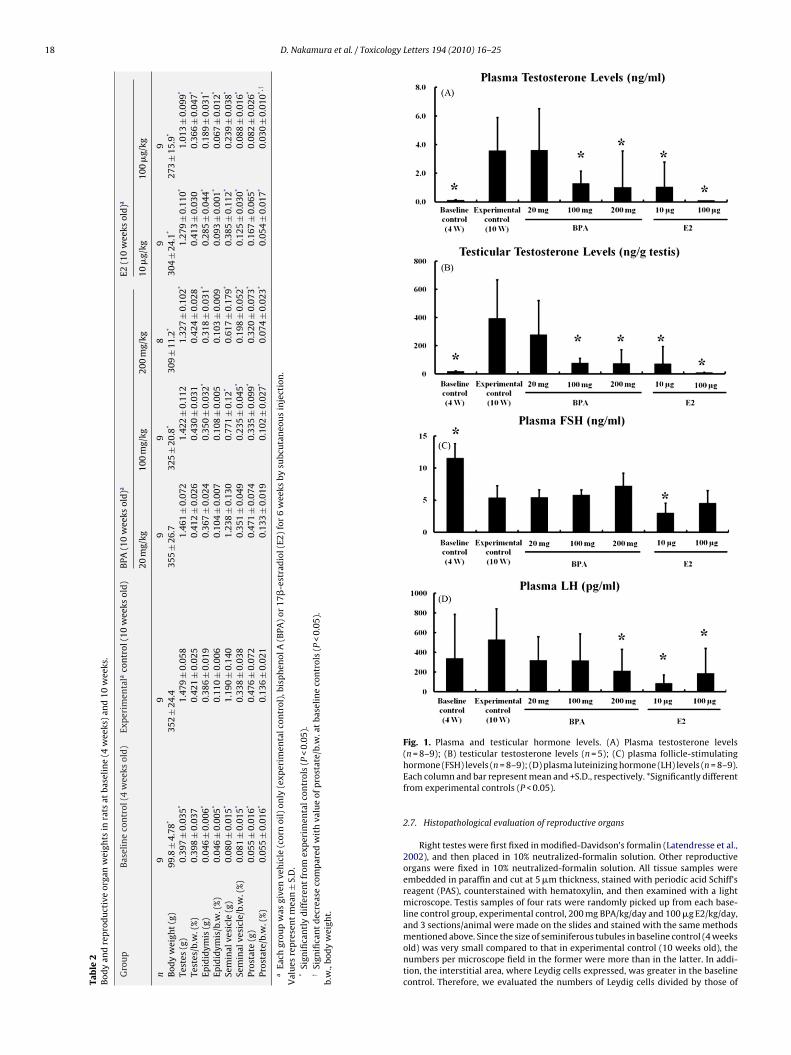
Fig. 1. Plasma and testicular hormone levels. (A) Plasma testosterone levels
(n = 8–9); (B) testicular testosterone levels (n = 5); (C) plasma follicle-stimulatinghormone (FSH) levels (n = 8–9); (D) plasma luteinizing hormone (LH) levels (n = 8–9).Each column and bar represent mean and +S.D., respectively. *Significantly differentfrom experimental controls (P < 0.05).2.7. Histopathological evaluation of reproductive organs
Right testes were first fixed in modified-Davidson’s formalin (Latendresse et al.,2002), and then placed in 10% neutralized-formalin solution. Other reproductiveorgans were fixed in 10% neutralized-formalin solution. All tissue samples wereembedded in paraffin and cut at 5 �m thickness, stained with periodic acid Schiff’sreagent (PAS), counterstained with hematoxylin, and then examined with a lightmicroscope. Testis samples of four rats were randomly picked up from each base-line control group, experimental control, 200 mg BPA/kg/day and 100 �g E2/kg/day,and 3 sections/animal were made on the slides and stained with the same methods
mentioned above. Since the size of seminiferous tubules in baseline control (4 weeksold) was very small compared to that in experimental control (10 weeks old), thenumbers per microscope field in the former were more than in the latter. In addi-tion, the interstitial area, where Leydig cells expressed, was greater in the baselinecontrol. Therefore, we evaluated the numbers of Leydig cells divided by those of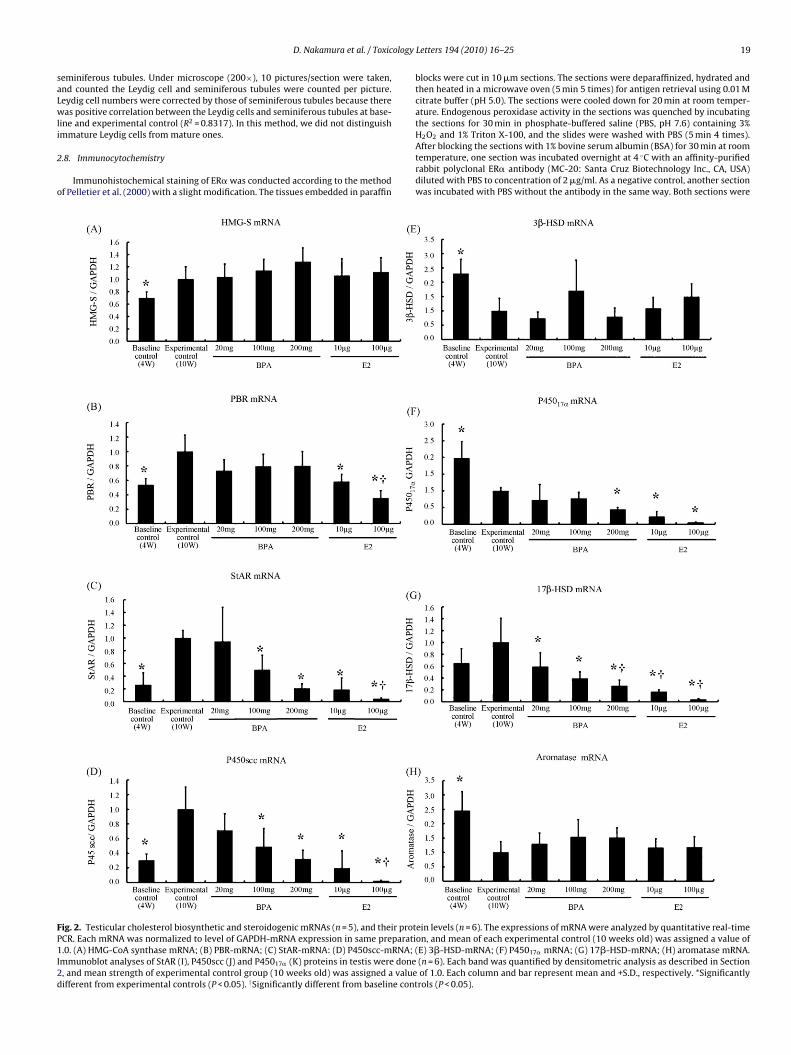
ology
saLwli
2
o
FP1I2d
D. Nakamura et al. / Toxic
eminiferous tubules. Under microscope (200×), 10 pictures/section were taken,nd counted the Leydig cell and seminiferous tubules were counted per picture.eydig cell numbers were corrected by those of seminiferous tubules because thereas positive correlation between the Leydig cells and seminiferous tubules at base-
ine and experimental control (R2 = 0.8317). In this method, we did not distinguish
mmature Leydig cells from mature ones..8. Immunocytochemistry
Immunohistochemical staining of ER� was conducted according to the methodf Pelletier et al. (2000) with a slight modification. The tissues embedded in paraffin
ig. 2. Testicular cholesterol biosynthetic and steroidogenic mRNAs (n = 5), and their protCR. Each mRNA was normalized to level of GAPDH-mRNA expression in same preparati.0. (A) HMG-CoA synthase mRNA; (B) PBR-mRNA; (C) StAR-mRNA: (D) P450scc-mRNA;mmunoblot analyses of StAR (I), P450scc (J) and P45017� (K) proteins in testis were done, and mean strength of experimental control group (10 weeks old) was assigned a valueifferent from experimental controls (P < 0.05). †Significantly different from baseline cont
Letters 194 (2010) 16–25 19
blocks were cut in 10 �m sections. The sections were deparaffinized, hydrated andthen heated in a microwave oven (5 min 5 times) for antigen retrieval using 0.01 Mcitrate buffer (pH 5.0). The sections were cooled down for 20 min at room temper-ature. Endogenous peroxidase activity in the sections was quenched by incubatingthe sections for 30 min in phosphate-buffered saline (PBS, pH 7.6) containing 3%
H2O2 and 1% Triton X-100, and the slides were washed with PBS (5 min 4 times).After blocking the sections with 1% bovine serum albumin (BSA) for 30 min at roomtemperature, one section was incubated overnight at 4 ◦C with an affinity-purifiedrabbit polyclonal ER� antibody (MC-20: Santa Cruz Biotechnology Inc., CA, USA)diluted with PBS to concentration of 2 �g/ml. As a negative control, another sectionwas incubated with PBS without the antibody in the same way. Both sections wereein levels (n = 6). The expressions of mRNA were analyzed by quantitative real-timeon, and mean of each experimental control (10 weeks old) was assigned a value of(E) 3�-HSD-mRNA; (F) P45017� mRNA; (G) 17�-HSD-mRNA; (H) aromatase mRNA.(n = 6). Each band was quantified by densitometric analysis as described in Sectionof 1.0. Each column and bar represent mean and +S.D., respectively. *Significantlyrols (P < 0.05).

20 D. Nakamura et al. / Toxicology Letters 194 (2010) 16–25
(Conti
tad3watw
Z
2
wcWnt
3
3
tBbgwt
taddadB
levels. E2 treatments also decreased the levels. BPA did not affect
Fig. 2.
hen washed in PBS 4 times each for 5 min, followed by incubating at room temper-ture for 30 min with biotin-labeled goat anti-rabbit IgG (CHEMICON International)iluted at 1:500 with PBS. After washing with PBS, the sections were incubated for0 min with streptavidin labeled-peroxidase (Nichirei, Tokyo, Japan), followed byashing with PBS (5 min 3 times) and Tris-buffered saline (pH 7.6, 5 min 1 time),
nd visualizing using Simple Stain DAB (Nichirei, Tokyo, Japan). The visualizing reac-ion was stopped by washing with distilled water. Each section was counterstainedith Mayer hematoxylin.
The photographs in the present study were taken by Fuji Digital Camera HC 300/CL (Olympus Japan Co.).
.9. Statistical analysis
All data were expressed as mean ± standard deviation (S.D.). Statistical analysesere performed by t-test when comparing two groups, and Dunnett’s test when
omparing more than three groups using JMP version 4.0 (SAS Institute Inc., USA).hen the values were not normally distributed, statistical analysis was done after
ormalization of the values by logarithmic or root transformation. P values of lesshan 0.05 were considered statistically significant.
. Results
.1. Body and organ weights
The body weights were 3.5-fold higher in the experimental con-rol rats compared to the baseline controls (Table 2). Low dose ofPA (20 mg/kg) did not influence the increase in the body weights,ut BPA more than the middle-dose group (100 mg/kg) and all E2roups (10 and 100 �g/kg) dose-dependently decreased the bodyeights compared to those of experimental control rats, suggesting
hat these treatments inhibited the increase in body weight.BPA and E2 also affected the reproductive organ weights: BPA
reatment at the high dose (200 mg/kg) decreased the testis weight,nd the middle (100 mg/kg) and high dosages reduced the epi-
idymis, seminal vesicle and prostate weights; all E2 dosagesecreased all reproductive organ weights investigated. In contrast,ll BPA dosages did not affect the ratios of testis/body, and epi-idymis/body weights, which were decreased by a high dose ofPA and both dosages of E2, respectively. BPA at more than middlenued).
dosages and all E2 dosages decreased the ratios of seminal vesi-cle/body and prostate/body weights. Interestingly, the high dose ofE2 decreased the ratios of seminal vesicle/body weights to the samelevel as those of baseline controls, and prostate/body weight to lev-els lower than those of baseline controls. These results suggest thatthese reproductive organs are vulnerable to the effects of the twochemicals, BPA and E2, compared to the testes and epididymis.
3.2. Hormone levels in plasma and testis
Plasma testosterone levels in baseline control rats were signif-icantly lower than those of experimental control rats: the formerlevels were one-thirty-sixth of those in the latter (Fig. 1A). BPAtreatments of more than the middle dosages decreased the lev-els to one-third of the experimental control rats. E2 treatmentsdose-dependently decreased the plasma testosterone levels, andsurprisingly, the high dose decreased the levels to the same as thosein the baseline controls.
Testicular testosterone levels were very similar to those inplasma: BPA treatments decreased the testicular testosterone lev-els more than the middle dosages and all E2 treatments (Fig. 1B).Plasma FSH and LH levels were also measured. Interestingly, plasmaFSH levels were lowered in experimental control rats compared tothose of baseline controls (Fig. 1C). BPA treatments did not influ-ence the levels, while the low dose of E2 significantly decreased thelevels.
In contrast to the FSH value, plasma LH levels were not differ-ent between the experimental control and baseline control rats(Fig. 1D). The high dose of BPA significantly decreased plasma LH
the plasma E2 levels, while E2 treatments increased the levelsdose-dependently: baseline control, 5.9 ± 4.39 pg/ml; experimen-tal control, 7.0 ± 1.30 pg/ml; 20 mg BPA, 7.1 ± 1.48 pg/ml; 100 mgBPA, 6.1 ± 1.34 pg/ml; 200 mg BPA, 12.0 ± 10.16 pg/ml; 10 �g E2,48.3 ± 22.69 pg/ml; 100 �g E2; 343.6 ± 140.01 pg/ml.

ology Letters 194 (2010) 16–25 21
3
te
3
s(ttlew
3
rmcmthdPd
iltededtws
3
iPtitdto
th(Ttaieseeat
Fig. 3. Testicular estrogen receptor (ER) �/� mRNA expressions (n = 5). (A) ER�mRNA; (B) ER� mRNA. Each mRNA was normalized to level of GAPDH-mRNA expres-sion in same preparation, and the mean of each experimental control (10 weeks
D. Nakamura et al. / Toxic
.3. Steroidogenic mRNA in testis
mRNA levels of several genes associated with testicular choles-erol biosynthesis, transport and testosterone biosynthesis werexamined by real-time quantitative PCR.
.4. Cholesterol biosynthesis
In testis, cholesterol is an essential substrate for testosteroneynthesis. Both levels of 3-hydroxy-3-methylglutaryl coenzyme AHMG-CoA) synthase and HMG-CoA reductase involved in choles-erol biosynthesis were measured. Neither BPA nor E2 influencedhese levels (data not shown). However, HMG-CoA synthase-mRNAevels were slightly lower in the baseline control rats than in thexperimental controls (Fig. 2A), but those of HMG-CoA reductaseere not (data not shown).
.5. Cholesterol transport
Peripheral benzodiazepine receptor (PBR) and StAR play a keyegulatory role in cholesterol transport from the outer to the inneritochondrial membrane, where P450scc is responsible for the
onversion of cholesterol to pregnenolone. In control groups, PBR-RNA expressions at the baseline controls were significantly lower
han those of experimental control rats (Fig. 2B). BPA treatments,owever, did not affect the levels, while E2 treatments dose-ependently decreased the levels; the low dose of E2 decreasedBR-mRNA to the same level as baseline control, and the high oneecreased the levels even lower than in the baseline control rats.
The expressions of StAR and P450scc-mRNA were also lowern the baseline control rats than in experimental controls, simi-ar to those of PBR (Fig. 2C and D). However, BPA dosages morehan 100 mg/kg and all E2 treatments significantly decreased thesexpressions in a dose-dependent manner. Interestingly, the highose of BPA and low-dose E2 decreased StAR and P450scc-mRNAxpressions to the same levels as baseline controls, and the highose of E2 did so to levels lower than the baseline controls. Thus,he effects of BPA treatments on the expressions of these genesere very similar to those of E2, but the toxic effects were clearly
tronger in E2 than BPA.
.6. Testosterone biosynthesis
In the smooth endoplasmic reticulum, pregnenolone producedn mitochondria is catalyzed by several enzymes to testosterone.regnenolone synthesized in mitochondria is, in turn, convertedo progesterone by 3�-hydroxysteroid dehydrogenase/�5-�4-somerase (3�-HSD), followed by P45017� to produce andros-endione. This hormone is also produced from pregnenolone viaehydroepiandrosterone (DEHA) by P45017� and 3�-HSD. Finally,estosterone is produced from androstendione by catalytic actionf 17�-HSD.
In contrast to the above-mentioned genes involved in choles-erol biosynthesis and transport, the levels of 3�-HSD-mRNA wereigher in the baseline controls than those of experimental controlsFig. 2E). However, BPA and E2 treatments did not affect the levels.he expressions of P45017� mRNA were also higher in baseline con-rol rats than experimental controls (Fig. 2F). The high dose of BPAnd E2 significantly decreased the expressions: the former chem-cal decreased the expressions by half compared to those in thexperimental controls, and all E2 dosages decreased the expres-
ions in a dose-dependent manner. There was no difference in thexpressions of 17�-HSD-mRNA between baseline control rats andxperimental controls (Fig. 2G). This gene was most sensitive to BPAnd E2: all dosages of these chemicals decreased dose-dependentlyhe levels of 17�-HSD-mRNA. The high dose of BPA decreased theold) was assigned a value of 1.0. Each column and bar represent mean and +S.D.,respectively. *Significantly different from experimental controls (P < 0.05).
levels more than half, and the high dose of E2 decreased themto one-tenth. The mRNA expression of aromatase, which convertstestosterone to E2, in the baseline control rats was significantlyhigher than the expression in experimental control rats (Fig. 2H).Neither BPA nor E2 influenced the expression.
3.7. Steroidogenic protein expression in testis
In order to confirm the effects of BPA or E2 on the steroido-genic mRNA levels, expressions of StAR, P450scc and P45017� wereinvestigated by Western blot analysis. BPA and E2 treatments didnot influence the expression of internal control, GAPDH. In agree-ment with the results in their mRNA levels, the expression of StARin rats at 10 weeks was stronger than that at 4 weeks; BPA and E2significantly decreased the StAR protein level (Fig. 2I). BPA and E2significantly decreased expressions of P450scc, which also agreedwith those of mRNA (Fig. 2J). However, the expression in baselinecontrol rats at 4 weeks was the same as that of experimental controlat 10 weeks, which was quite different from the result in the mRNAlevels. In P45017�, the expressions were significantly higher in ratsat 4 weeks than at 10 weeks, and BPA and E2 further decreased theexpressions, similar to those of the mRNA (Fig. 2K).
3.8. AR, ER˛/ˇ mRNA expression
AR and ER�/� play an important role for regulations of testos-terone synthesis and spermatogenesis (Pelletier et al., 2000). Nosignificant differences were noted in the expression of AR (datanot shown) and ER� mRNA (Fig. 3A) in testis between baselineand experimental controls. BPA and E2 did not influence the for-mer gene, but both chemicals decreased the expressions of the
latter one. In ER� mRNA, the expression was greater in the base-line control rats than in the experimental controls, while BPA andE2 treatment did not influence the gene levels (Fig. 3B).
22 D. Nakamura et al. / Toxicology Letters 194 (2010) 16–25
F A) SemX mese( . RS: rs
3
EiiiiaeasmssadsX
ig. 4. ER� localization in testis of experimental control, BPA and E2-treated rats. (II–XIII; (E) stages XIII–XIV; (F) mesenchyme of the BPA (200 mg/kg)-treated rat; (G)
H) Negative control of ER� staining, stages XII–XIV. Original magnification, 400×hows Leydig cell.
.9. ER˛ localization in testis
Since BPA and E2 treatments decreased the expressions ofR�-mRNA in the testis, the localization was investigated bymmunohistochemical analysis. Fig. 4A–E shows ER� localizationn the testis of experimental control rats. ER� expressions weremmunohistologically detected in Leydig cells, Sertoli cells andn both round and elongated spermatids. The round spermatidsppeared at stages I–VIII of seminiferous tubule in the testis; thelongated ones appeared at stages IX–VIII. ER� in Leydig cellsnd Sertoli cells were highly expressed at all stages. However, inpermatids, the ER� expression was detected in a stage-specificanner: although ER� was not expressed in round spermatids at
tages II–VI, it was expressed in elongated spermatids at these
tages (Fig. 4A). ER� started to express in the round spermatidst stages VII–VIII, whereas the expression of the receptor was notetected in elongated spermatids at these stages (Fig. 4B). Roundpermatids at stages VIII–X and elongated spermatids at stagesI–XIV constantly expressed ER� (Fig. 4C–E). The high dosage ofiniferous tubules at stages II–VI; (B) stages VII–VIII; (C) stages VIII–IX; (D) stagesnchyme of the E2 (100 �g/kg)-treated rat, stages II–VI. Note the low ER� expression.ound spermatids; ES: elongated spermatids. Arrowhead shows Sertoli cell. Arrow
BPA and E2 appeared to decrease the numbers of Leydig cells withER� expressed (Fig. 4F and G), but did not appear to influence theexpression of Sertoli cells in both round and elongated spermatidswith ER�.
3.10. Histopathological evaluation of reproductive organs
Under microscopical investigation, 20 and 100 mg/kg BPAdosages did not have an adverse effect on testis morphology com-pared with that of the experimental controls (Fig. 5A), while BPA200 mg/kg dose decreased the Leydig cell numbers (Fig. 5B and D),in agreement with the decreasing tendency of many genes associ-ated with Leydig cells (Fig. 2). The low dose of E2 (10 �g/kg) also
appeared to decrease the numbers, even more than at 200 mg/kgBPA. The 100 �g/kg E2 dose significantly decreased the numbers(Fig. 5C and D). Although the internal diameter of the seminiferoustubule atrophied at the higher dose of E2, no obvious changes inquantity or quality of component cells were observed.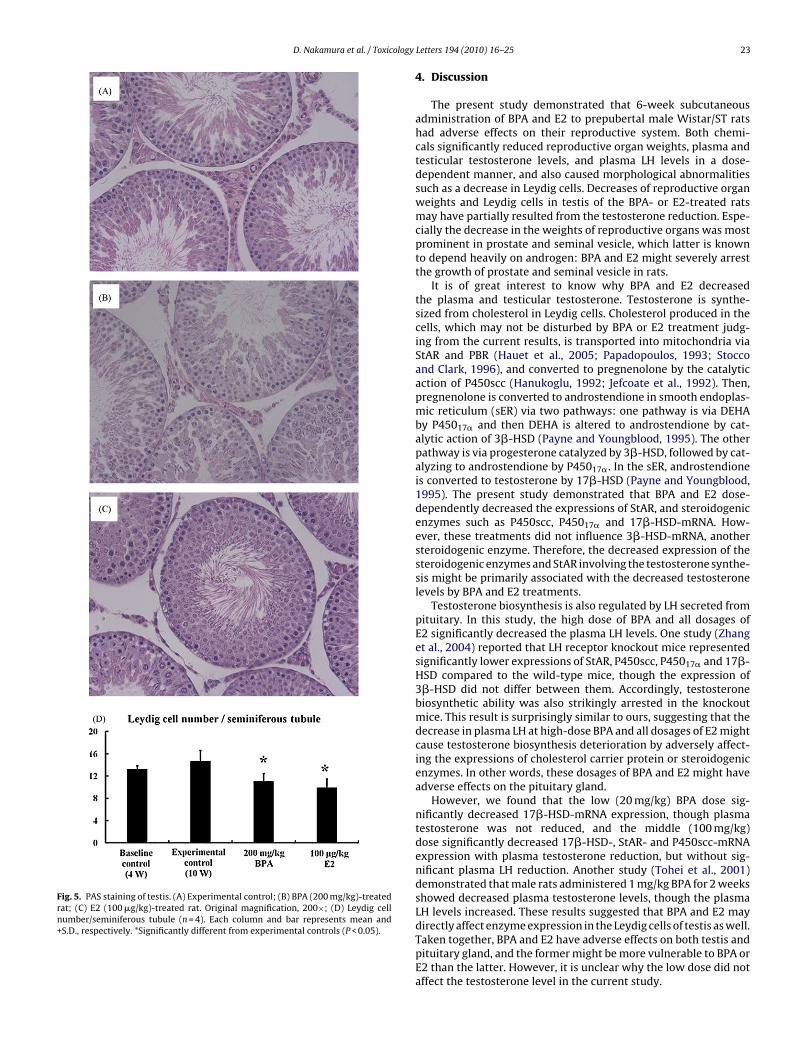
D. Nakamura et al. / Toxicology
Fig. 5. PAS staining of testis. (A) Experimental control; (B) BPA (200 mg/kg)-treatedrat; (C) E2 (100 �g/kg)-treated rat. Original magnification, 200×; (D) Leydig cellnumber/seminiferous tubule (n = 4). Each column and bar represents mean and+S.D., respectively. *Significantly different from experimental controls (P < 0.05).
Letters 194 (2010) 16–25 23
4. Discussion
The present study demonstrated that 6-week subcutaneousadministration of BPA and E2 to prepubertal male Wistar/ST ratshad adverse effects on their reproductive system. Both chemi-cals significantly reduced reproductive organ weights, plasma andtesticular testosterone levels, and plasma LH levels in a dose-dependent manner, and also caused morphological abnormalitiessuch as a decrease in Leydig cells. Decreases of reproductive organweights and Leydig cells in testis of the BPA- or E2-treated ratsmay have partially resulted from the testosterone reduction. Espe-cially the decrease in the weights of reproductive organs was mostprominent in prostate and seminal vesicle, which latter is knownto depend heavily on androgen: BPA and E2 might severely arrestthe growth of prostate and seminal vesicle in rats.
It is of great interest to know why BPA and E2 decreasedthe plasma and testicular testosterone. Testosterone is synthe-sized from cholesterol in Leydig cells. Cholesterol produced in thecells, which may not be disturbed by BPA or E2 treatment judg-ing from the current results, is transported into mitochondria viaStAR and PBR (Hauet et al., 2005; Papadopoulos, 1993; Stoccoand Clark, 1996), and converted to pregnenolone by the catalyticaction of P450scc (Hanukoglu, 1992; Jefcoate et al., 1992). Then,pregnenolone is converted to androstendione in smooth endoplas-mic reticulum (sER) via two pathways: one pathway is via DEHAby P45017� and then DEHA is altered to androstendione by cat-alytic action of 3�-HSD (Payne and Youngblood, 1995). The otherpathway is via progesterone catalyzed by 3�-HSD, followed by cat-alyzing to androstendione by P45017�. In the sER, androstendioneis converted to testosterone by 17�-HSD (Payne and Youngblood,1995). The present study demonstrated that BPA and E2 dose-dependently decreased the expressions of StAR, and steroidogenicenzymes such as P450scc, P45017� and 17�-HSD-mRNA. How-ever, these treatments did not influence 3�-HSD-mRNA, anothersteroidogenic enzyme. Therefore, the decreased expression of thesteroidogenic enzymes and StAR involving the testosterone synthe-sis might be primarily associated with the decreased testosteronelevels by BPA and E2 treatments.
Testosterone biosynthesis is also regulated by LH secreted frompituitary. In this study, the high dose of BPA and all dosages ofE2 significantly decreased the plasma LH levels. One study (Zhanget al., 2004) reported that LH receptor knockout mice representedsignificantly lower expressions of StAR, P450scc, P45017� and 17�-HSD compared to the wild-type mice, though the expression of3�-HSD did not differ between them. Accordingly, testosteronebiosynthetic ability was also strikingly arrested in the knockoutmice. This result is surprisingly similar to ours, suggesting that thedecrease in plasma LH at high-dose BPA and all dosages of E2 mightcause testosterone biosynthesis deterioration by adversely affect-ing the expressions of cholesterol carrier protein or steroidogenicenzymes. In other words, these dosages of BPA and E2 might haveadverse effects on the pituitary gland.
However, we found that the low (20 mg/kg) BPA dose sig-nificantly decreased 17�-HSD-mRNA expression, though plasmatestosterone was not reduced, and the middle (100 mg/kg)dose significantly decreased 17�-HSD-, StAR- and P450scc-mRNAexpression with plasma testosterone reduction, but without sig-nificant plasma LH reduction. Another study (Tohei et al., 2001)demonstrated that male rats administered 1 mg/kg BPA for 2 weeksshowed decreased plasma testosterone levels, though the plasmaLH levels increased. These results suggested that BPA and E2 may
directly affect enzyme expression in the Leydig cells of testis as well.Taken together, BPA and E2 have adverse effects on both testis andpituitary gland, and the former might be more vulnerable to BPA orE2 than the latter. However, it is unclear why the low dose did notaffect the testosterone level in the current study.
2 ology
sml2rmBEdfudIEiocrnbscahst
rai(is(ppsaH3tbtmlbsiiStomc
oarraetott
4 D. Nakamura et al. / Toxic
ERs play an important role for regulation of testosterone synthe-is. Testosterone levels were significantly greater in ER�-knockoutice than the wild-type mice, because of the significantly higher
evels of StAR and P45017�-mRNA in the former (Akingbemi et al.,003). Oral administration of low BPA (50 �g/ml) by drinking watereportedly induced expression of ER� mRNA, and repressed ER� -RNA in murine testis (Takao et al., 2003). In the present study,
PA did not change the expression of ER�, but decreased that ofR� in testis of rats at the middle and high dosages of BPA or allosages of E2. Though these paradoxes might have arisen from dif-erences of species, level and route of exposure, the reason remainsnclear. At least, ER� decreased by the two chemicals, BPA and E2,id not contribute to the decreased testosterone levels in this study.
n order to determine the cells that ER� expressions decreased,R� localization was investigated in testis by immunohistochem-stry. ER� expression has been found in spermatids, and Leydig cellsf rodent testis (Pelletier et al., 2000). In addition, ER� expressionould also be detected in Sertoli cells in this study, similarly to theeports by Baines et al. (2008) and Lucas et al. (2008). There wereo differences in ER� expression in Sertoli cells and spermatidsetween experimental controls and BPA or E2 treatments at anytages of seminiferous tubules. Microscopically, numbers of Leydigells were significantly decreased by the highest dosages of BPAnd E2. The decreased ER�-mRNA by BPA or E2 treatment mightave resulted from the decreased Leydig cells, where testosteroneynthesis is primarily involved. Taken together, BPA appeared noto induce ER� in the testes.
Finally, we investigated the development of testosterone-elated gene expression in the testes. The P45017�, 3�-HSD andromatase mRNAs investigated in the present study were lowern the experimental controls (10 weeks) than in baseline controls4 weeks). Mack et al. (2000) also reported similar results: theynvestigated plasma testosterone, and the genes involved in theynthesis in testis of male Sprague–Dawley rats at the immature21–28 days) and pubertal (45–60 days) stages, and showed thatlasma testosterone sharply increased from the immature to theubertal stage, whereas P45017� and 3�-HSD (I)-mRNA expres-ions decreased during these stages. Since these mRNA expressionsre inhibited via AR by testosterone (Burgos-Trinidad et al., 1997;eggland et al., 1997), decrease in the expressions of P45017� and�-HSD during the development may depend on the increasedestosterone levels. However, the former enzyme was decreasedy BPA or E2 treatments, whereas the latter was not. In contrast tohe above enzymes, expressions of StAR, P450scc, PBR, and HMG-S
RNAs were greater in the experimental controls than in base-ine controls, though the P450scc protein level was not differentetween them, for unknown reasons. Taken together, the expres-ions of some steroidogenic genes increased, and others decreasedn testis of mice between 4 and 10 weeks, while testosterone levelsncreased during this stage. Since BPA and E2 treatments decreasedtAR and steroidogenic enzymes excluding 3�-HSD (I) and HMG,he decreases might not result from the inhibition of the devel-pment of testosterone synthesis; rather their potential toxicitiesay inhibit the expressions in Leydig cells as well as decrease the
ells.Pottenger et al. (2000) showed the different bioavailability
f BPA among administration routes: the bioavailability by oraldministration was significantly lower than by the subcutaneousoute. Therefore, plasma BPA concentration might be higher inats of the present study compared to that by oral administrationt the same dose in the current study. However, in the present
xperiment, we focused on clarifying how BPA decreased testos-erone levels, and did not determine the adverse effect levels. Tour knowledge, this is the first report that BPA reduced plasmaestosterone levels by decreasing the expressions of the cholesterolransport protein, StAR and some steroidogenic enzymes, whichLetters 194 (2010) 16–25
may be caused by the direct effects on Leydig cells as well as pitu-itary gland, a mechanism similar to that of E2. Interestingly, theformer effects may be observed at a lower concentration of BPAthan the latter. Of course, further study is warranted to investigateanother factor involved in the reduction of testosterone by BPA.
Conflict of interest
Authors declare no conflict of interest.
Funding source
This study was supported in part by a Grant-in-Aid for ScientificResearch (B. 14370121 and 17310033) from the Japan Society forthe Promotion of Science (JSPS).
Acknowledgements
We wish to acknowledge the generous assistance of Ms. MaikoMiyata and Dr. Chul-Ho Lee in the Department of Occupationaland Environmental Health, Nagoya University Graduate School ofMedicine.
References
Akingbemi, B.T., Ge, R., Rosenfeld, C.S., Newton, L.G., Hardy, D.O., Catterall, J.F.,Lubahn, D.B., Korach, K.S., Hardy, M.P., 2003. Estrogen receptor-alpha gene defi-ciency enhances androgen biosynthesis in the mouse Leydig cell. Endocrinology144, 84–93.
Akingbemi, B.T., Sottas, C.M., Koulova, A.I., Klinefelter, G.R., Hardy, M.P., 2004. Inhibi-tion of testicular steroidogenesis by the xenoestrogen bisphenol A is associatedwith reduced pituitary luteinizing hormone secretion and decreased steroido-genic enzyme gene expression in rat Leydig cells. Endocrinology 145, 592–603.
Baines, H., Nwagwu, M.O., Hastie, G.R., Wiles, R.A., Mayhew, T.M., Ebling, F.J., 2008.Effects of estradiol and FSH on maturation of the testis in the hypogonadal (hpg)mouse. Reprod. Biol. Endocrinol. 6, 4.
Brotons, J.A., Olea-Serrano, M.F., Villalobos, M., Pedraza, V., Olea, N., 1995. Xenoe-strogens released from lacquer coatings in food cans. Environ. Health Perspect.103, 608–612.
Burgos-Trinidad, M., Youngblood, G.L., Maroto, M.R., Scheller, A., Robins, D.M., Payne,A.H., 1997. Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylase/C17-20 lyase gene (Cyp17) by androgens. Mol. Endocrinol. 11,87–96.
Gupta, C., 2000. Reproductive malformation of the male offspring following maternalexposure to estrogenic chemicals. Proc. Soc. Exp. Biol. Med. 224, 61–68.
Hanukoglu, I., 1992. Steroidogenic enzymes: structure, function, and role in regula-tion of steroid hormone biosynthesis. J. Steroid Biochem. Mol. Biol. 43, 779–804.
Hauet, T., Yao, Z.X., Bose, H.S., Wall, C.T., Han, Z., Li, W., Hales, D.B., Miller, W.L., Culty,M., Papadopoulos, V., 2005. Peripheral-type benzodiazepine receptor-mediatedaction of steroidogenic acute regulatory protein on cholesterol entry into Leydigcell mitochondria. Mol. Endocrinol. 19, 540–554.
Heggland, S.J., Signs, S.A., Stalvey, J.R., 1997. Testosterone decreases 3beta-hydroxysteroid dehydrogenase-isomerase messenger ribonucleic acid incultured mouse Leydig cells by a strain-specific mechanism. J. Androl. 18,646–655.
Herath, C.B., Jin, W., Watanabe, G., Arai, K., Suzuki, A.K., Taya, K., 2004. Adverse effectsof environmental toxicants, octylphenol and bisphenol A, on male reproductivefunctions in pubertal rats. Endocrine 25, 163–172.
Ikezuki, Y., Tsutsumi, O., Takai, Y., Kamei, Y., Taketani, Y., 2002. Determination ofbisphenol A concentrations in human biological fluids reveals significant earlyprenatal exposure. Hum. Reprod. 17, 2839–2841.
Jefcoate, C.R., McNamara, B.C., Artemenko, I.T.Y., 1992. Regulation of cholesterolmovement to mitochondrial cytochrome P450scc in steroid hormone synthesis.J. Steroid Biochem. Mol. Biol. 43, 751–767.
Khurana, S., Ranmal, S., Ben-Jonathan, N., 2000. Exposure of newborn male andfemale rats to environmental estrogens: delayed and sustained hyperpro-lactinemia and alterations in estrogen receptor expression. Endocrinology 141,4512–4517.
Krishnan, A.V., Stathis, P., Permuth, S.F., Tokes, L., Feldman, D., 1993. Bisphenol-A: anestrogenic substance is released from polycarbonate flasks during autoclaving.Endocrinology 132, 2279–2286.
Latendresse, J.R., Warbrittion, A.R., Jonassen, H., Creasy, D.M., 2002. Fixation of testes
and eyes using a modified Davidson’s fluid: comparison with Bouin’s fluid andconventional Davidson’s fluid. Toxicol. Pathol. 30, 524–533.Lucas, T.F., Siu, E.R., Esteves, C.A., Monteiro, H.P., Oliveira, C.A., Porto, C.S., Lazari, M.F.,2008. 17beta-Estradiol induces the translocation of the estrogen receptors ESR1and ESR2 to the cell membrane, MAPK3/1 phosphorylation and proliferation ofcultured immature rat Sertoli cells. Biol. Reprod. 78, 101–114.

ology
M
M
O
P
P
P
P
R
R
R
S
D. Nakamura et al. / Toxic
ack, S.O., Garrett, W.M., Guthrie, H.D., 2000. Absence of correlation betweenin situ expression of cytochrome P450 17alpha hydroxylase/lyase and 3beta-hydroxysteroid dehydrogenase/(Delta5-4) isomerase messenger ribonucleicacids and steroidogenesis during pubertal development in the rat testis. J. SteroidBiochem. Mol. Biol. 73, 19–28.
atthews, J.B., Twomey, K., Zacharewski, T.R., 2001. In vitro and in vivo interac-tions of bisphenol A and its metabolite, bisphenol A glucuronide, with estrogenreceptors alpha and beta. Chem. Res. Toxicol. 14, 149–157.
lea, N., Pulgar, R., Perez, P., Olea-Serrano, F., Rivas, A., Novillo-Fertrell, A., Pedraza,V., Soto, A.M., Sonnenschein, C., 1996. Estrogenicity of resin-based compositesand sealants used in dentistry. Environ. Health Perspect. 104, 298–305.
apadopoulos, V., 1993. Peripheral-type benzodiazepine/diazepam bindinginhibitor receptor: biological role in steroidogenic cell function. Endocr. Rev.14, 222–240.
ayne, A.H., Youngblood, G.L., 1995. Regulation of expression of steroidogenicenzymes in Leydig cells. Biol. Reprod. 52, 217–225.
elletier, G., Labrie, C., Labrie, F., 2000. Localization of oestrogen receptor alpha,oestrogen receptor beta and androgen receptors in the rat reproductive organs.J. Endocrinol. 165, 359–370.
ottenger, L.H., Domoradzki, J.Y., Markham, D.A., Hansen, S.C., Cagen, S.Z., WaechterJr., J.M., 2000. The relative bioavailability and metabolism of bisphenol A in ratsis dependent upon the route of administration. Toxicol. Sci. 54, 3–18.
amos, J.G., Varayoud, J., Sonnenschein, C., Soto, A.M., Munoz De Toro, M., Luque,E.H., 2001. Prenatal exposure to low doses of bisphenol A alters the periductalstroma and glandular cell function in the rat ventral prostate. Biol. Reprod. 65,1271–1277.
amos, J.G., Varayoud, J., Kass, L., Rodriguez, H., Costabel, L., Munoz De Toro, M.,Luque, E.H., 2003. Bisphenol a induces both transient and permanent histo-functional alterations of the hypothalamic–pituitary–gonadal axis in prenatallyexposed male rats. Endocrinology 144, 3206–3215.
ubin, B.S., Murray, M.K., Damassa, D.A., King, J.C., Soto, A.M., 2001. Perinatal expo-sure to low doses of bisphenol A affects body weight, patterns of estrous cyclicity,and plasma LH levels. Environ. Health Perspect. 109, 675–680.
chonfelder, G., Wittfoht, W., Hopp, H., Talsness, C.E., Paul, M., Chahoud, I., 2002.Parent bisphenol A accumulation in the human maternal–fetal–placental unit.Environ. Health Perspect. 110, A703–707.
Letters 194 (2010) 16–25 25
Snyder, R.W., Maness, S.C., Gaido, K.W., Welsch, F., Sumner, S.C., Fennell, T.R., 2000.Metabolism and disposition of bisphenol A in female rats. Toxicol. Appl. Phar-macol. 168, 225–234.
Steinmetz, R., Brown, N.G., Allen, D.L., Bigsby, R.M., Ben-Jonathan, N., 1997. The envi-ronmental estrogen bisphenol A stimulates prolactin release in vitro and in vivo.Endocrinology 138, 1780–1786.
Steinmetz, R., Mitchner, N.A., Grant, A., Allen, D.L., Bigsby, R.M., Ben-Jonathan,N., 1998. The xenoestrogen bisphenol A induces growth, differentiation, andc-fos gene expression in the female reproductive tract. Endocrinology 139,2741–2747.
Stocco, D.M., Clark, B.J., 1996. Regulation of the acute production of steroids insteroidogenic cells. Endocr. Rev. 17, 221–244.
Takahashi, O., Oishi, S., 2003. Testicular toxicity of dietarily or parenterally admin-istered bisphenol A in rats and mice. Food Chem. Toxicol. 41, 1035–1044.
Takao, T., Nanamiya, W., Nazarloo, H.P., Matsumoto, R., Asaba, K., Hashimoto, K.,2003. Exposure to the environmental estrogen bisphenol A differentially mod-ulated estrogen receptor-alpha and -beta immunoreactivity and mRNA in malemouse testis. Life Sci. 72, 1159–1169.
Timms, B.G., Howdeshell, K.L., Barton, L., Bradley, S., Richter, C.A., vom Saal, F.S.,2005. Estrogenic chemicals in plastic and oral contraceptives disrupt develop-ment of the fetal mouse prostate and urethra. Proc. Natl. Acad. Sci. U.S.A. 102,7014–7019.
Tohei, A., Suda, S., Taya, K., Hashimoto, T., Kogo, H., 2001. Bisphenol A inhibits testic-ular functions and increases luteinizing hormone secretion in adult male rats.Exp. Biol. Med. (Maywood) 226, 216–221.
Welshons, W.V., Nagel, S.C., Thayer, K.A., Judy, B.M., Vom Saal, F.S., 1999. Low-dosebioactivity of xenoestrogens in animals: fetal exposure to low doses of methoxy-chlor and other xenoestrogens increases adult prostate size in mice. Toxicol. Ind.Health 15, 12–25.
Xu, L.C., Sun, H., Chen, J.F., Bian, Q., Qian, J., Song, L., Wang, X.R., 2005. Evaluation
of androgen receptor transcriptional activities of bisphenol A, octylphenol andnonylphenol in vitro. Toxicology 216, 197–203.Zhang, F.P., Pakarainen, T., Zhu, F., Poutanen, M., Huhtaniemi, I., 2004.Molecular characterization of postnatal development of testicular steroido-genesis in luteinizing hormone receptor knockout mice. Endocrinology 145,1453–1463.
Copyright © 2022 FDOKUMEN