
Biology and Natural History of Caladenia
12
Review Biology and natural history of Caladenia Kingsley Dixon A,C and Raymond L. Tremblay B A Botanic Garden and Parks Authority, Kings Park and Botanic Garden, West Perth 6005, Western Australia and School of Plant Biology, The University of Western Australia, Nedlands 6009, Western Australia, Australia. B Department of Biology, 100 Carretera 908, University of Puerto Rico, Humacao Campus, Humacao, Puerto Rico 00792, USA and Crest-Catec, Center for Applied Tropical Ecology and Conservation, PO Box 23341, University of Puerto Rico, Río Piedras, Puerto Rico 00931-3341, USA. C Corresponding author. Email: [email protected] Abstract. The genus Caladenia comprises species that exhibit remarkable consistency in terms of growth form and phenological patterns. All taxa are herbaceous perennials, with the shoot arising annually from a single, usually spheroid, tuber and producing a single, hairy leaf. The tuber is annually replaced either side-by-side with the parent tuber or terminating a descending structure known as a dropper. The dropper is a depth-seeking mechanism that enables placement of the tuber at depth in the soil as a means to avoid predation by surface-foraging native mammals or away from the high temperatures and desiccating conditions during summer dormancy. The 3–5 attenuated roots produced at the junction between the tuber and shoot and produced late in the growing cycle and devoid of mycorrhiza suggest their functional significance may relate to water uptake. Mycorrhizal endophytes are confined to a hypertrophic stem region at the soil surface (collar) subtending the leaf that positions the collar directly in the organically rich zone at the soil surface. This morphology is a unique characteristic of several Australasian orchids in the tribe Diuridae. Mycorrhizal infection occurs rapidly, with maximum colonisation in concert with the onset of breaking rains. Pelotons are restricted to cortical cells, with fully developed pelotons throughout infected tissues within a week or so of soil wetting. Infection occurs as a ‘once-off’ event, with little evidence of secondary infection later in the growth cycle and no evidence of peloton digestion. Some taxa utilise vegetative propagation, often leading to localised clustering as for taxa in the ‘filamentosa’ complex or, extensive clonal mats as found in Caladenia flava and C. latifolia where daughter tubers are produced at the end of extending horizontal outgrowths. For the majority of taxa, plants remain dry-season (summer) dormant from a few months up to 7 months for arid-zone taxa, with shoot emergence from the tuber of temperate species thought to occur in response to a drop in the mean minimum temperature. Pollination biology of Caladenia is apparently through a process of deception, either as food or sexual mimics, with some taxa engaging in self- pollination. Here we review the natural history of Caladenia and acknowledge that much of our understanding is based on assumptions of the biology of terrestrial orchids in general and emphasise areas of research and biological enquiry that will be critical in the development of an effective conservation program for the genus. Introduction Caladenia represent a natural group of taxa in terms of morphological similarities – all species are herbaceous perennials, most have a period of summer (dry-season) dormancy where the perennating structure comprises a spherical to ovoid tuber. All taxa produce a single, often densely hairy leaf that is subtended at ground level (usually within the organic layer) by a swollen region occupied by mycorrhizal fungi. The related genera (some were once considered to be members of the genus Caladenia) Praecoxanthus, Cyanicula, Elythranthera and Pheladenia share similarities in morphology and phenology with Caladenia, linked to their common phylogenetic origin (Kores et al. 2001). Here we propose that the combination of morphological, phenological and ecological specialisations (mycorrhizal associates; pollinator specificity, particularly in sexually deceptive taxa) found in many Caladenia species may predispose plants to be highly vulnerable to a wide spectrum of environmental impacts (Koopowitz et al. 2003). Morphology The plant form of Caladenia species comprises an underground nutrient storage system (tuber), stem connecting the tuber to the soil surface and a single leaf produced at or near the soil surface (Fig. 1A). Plant size is variable from C. bryceana, where 2-cm flowers can be held on stems as short as 3 cm, to the tallest, C. excelsa, where plants can reach 90 cm, with flowers up to 15 cm across (Brown et al. 2008). Caladenia has a wide distributional range and phylogenetic spread through cool-temperate, subalpine–alpine, subtropical, mediterranean (cool moist winters, warm dry summers) to CSIRO PUBLISHING www.publish.csiro.au/journals/ajb Australian Journal of Botany, 2009, 57, 247–258 Ó CSIRO 2009 10.1071/BT08183 0067-1924/09/040247
Transcript of Biology and Natural History of Caladenia

Review
Biology and natural history of Caladenia
Kingsley DixonA,C and Raymond L. TremblayB
ABotanic Garden and Parks Authority, Kings Park and Botanic Garden, West Perth 6005, Western Australiaand School of Plant Biology, The University of Western Australia, Nedlands 6009, Western Australia, Australia.
BDepartment of Biology, 100 Carretera 908, University of Puerto Rico, Humacao Campus, Humacao,Puerto Rico 00792, USA and Crest-Catec, Center for Applied Tropical Ecology and Conservation,PO Box 23341, University of Puerto Rico, Río Piedras, Puerto Rico 00931-3341, USA.
CCorresponding author. Email: [email protected]
Abstract. The genus Caladenia comprises species that exhibit remarkable consistency in terms of growth form andphenological patterns. All taxa are herbaceous perennials, with the shoot arising annually from a single, usually spheroid,tuber and producing a single, hairy leaf. The tuber is annually replaced either side-by-sidewith the parent tuber or terminatinga descending structure known as a dropper. The dropper is a depth-seekingmechanism that enables placement of the tuber atdepth in the soil as a means to avoid predation by surface-foraging native mammals or away from the high temperatures anddesiccating conditions during summer dormancy. The 3–5 attenuated roots produced at the junction between the tuber andshoot and produced late in the growing cycle and devoid of mycorrhiza suggest their functional significance may relate towater uptake. Mycorrhizal endophytes are confined to a hypertrophic stem region at the soil surface (collar) subtending theleaf that positions the collar directly in the organically rich zone at the soil surface. Thismorphology is a unique characteristicof several Australasian orchids in the tribe Diuridae. Mycorrhizal infection occurs rapidly, with maximum colonisation inconcert with the onset of breaking rains. Pelotons are restricted to cortical cells, with fully developed pelotons throughoutinfected tissues within a week or so of soil wetting. Infection occurs as a ‘once-off’ event, with little evidence of secondaryinfection later in the growth cycle and no evidence of peloton digestion. Some taxa utilise vegetative propagation, oftenleading to localised clustering as for taxa in the ‘filamentosa’ complex or, extensive clonal mats as found inCaladenia flavaand C. latifoliawhere daughter tubers are produced at the end of extending horizontal outgrowths. For the majority of taxa,plants remain dry-season (summer) dormant froma fewmonths up to 7months for arid-zone taxa,with shoot emergence fromthe tuber of temperate species thought to occur in response to a drop in themeanminimum temperature. Pollination biologyofCaladenia is apparently through a process of deception, either as food or sexual mimics, with some taxa engaging in self-pollination. Here we review the natural history of Caladenia and acknowledge that much of our understanding is based onassumptions of the biology of terrestrial orchids in general and emphasise areas of research and biological enquiry thatwill becritical in the development of an effective conservation program for the genus.
Introduction
Caladenia represent a natural group of taxa in terms ofmorphological similarities – all species are herbaceousperennials, most have a period of summer (dry-season)dormancy where the perennating structure comprises a sphericalto ovoid tuber. All taxa produce a single, often densely hairy leafthat is subtended at ground level (usually within the organiclayer) by a swollen region occupied by mycorrhizal fungi. Therelated genera (some were once considered to be members of thegenus Caladenia) Praecoxanthus, Cyanicula, Elythranthera andPheladenia share similarities in morphology and phenologywith Caladenia, linked to their common phylogenetic origin(Kores et al. 2001). Here we propose that the combination ofmorphological, phenological and ecological specialisations(mycorrhizal associates; pollinator specificity, particularly in
sexually deceptive taxa) found in many Caladenia species maypredispose plants to be highly vulnerable to a wide spectrum ofenvironmental impacts (Koopowitz et al. 2003).
Morphology
The plant form of Caladenia species comprises an undergroundnutrient storage system (tuber), stem connecting the tuber to thesoil surface and a single leaf produced at or near the soil surface(Fig. 1A). Plant size is variable from C. bryceana, where 2-cmflowers can be held on stems as short as 3 cm, to the tallest,C. excelsa, where plants can reach 90 cm, with flowers up to15 cm across (Brown et al. 2008).
Caladenia has a wide distributional range and phylogeneticspread through cool-temperate, subalpine–alpine, subtropical,mediterranean (cool moist winters, warm dry summers) to
CSIRO PUBLISHING
www.publish.csiro.au/journals/ajb Australian Journal of Botany, 2009, 57, 247–258
� CSIRO 2009 10.1071/BT08183 0067-1924/09/040247
semi-arid climatic conditions. Caladenia taxa are also able totolerate a broad range of habitat conditions from litter-richsites, moss swards, nutrient-deficient podzols (<1% soilorganic matter), swamps and, for C. paludosa, a capacity tosurvive as a semi-aquatic (Fig. 2) (Brown et al. 2008). Theability of Caladenia to engage in extended periods of summerdormancy enables plants to survive in habitats that are seasonallyhostile and dry for periods of 6–7 months.
Caladenia taxa also exhibit a broad tolerance to light – fromfull sun to deep shade and, as with most plants, under low-light
conditions plants produce a highly etiolated leaf to compensatefor the lack of light. Depending on light availability, the clonalCaladenia species, C. flava and C. latifolia, exhibit the greatestplasticity in foliar morphology, most likely linked to the clonalhabit where shoots from the same individual may emerge in avariety of lighting conditions from deep shade to full sun.
A unique morphological characteristic of Caladeniaand related taxa (Pheladenia, Cyanicula, Glossodia andElythranthera) is the presence of a hypertrophic region at thebase of the leaf known as the collar (Ramsay et al. 1986; Dixon
P
C
DPT
RT
OT
Year 4 Year 5Year 2Year 1
(A)
(B) (C)
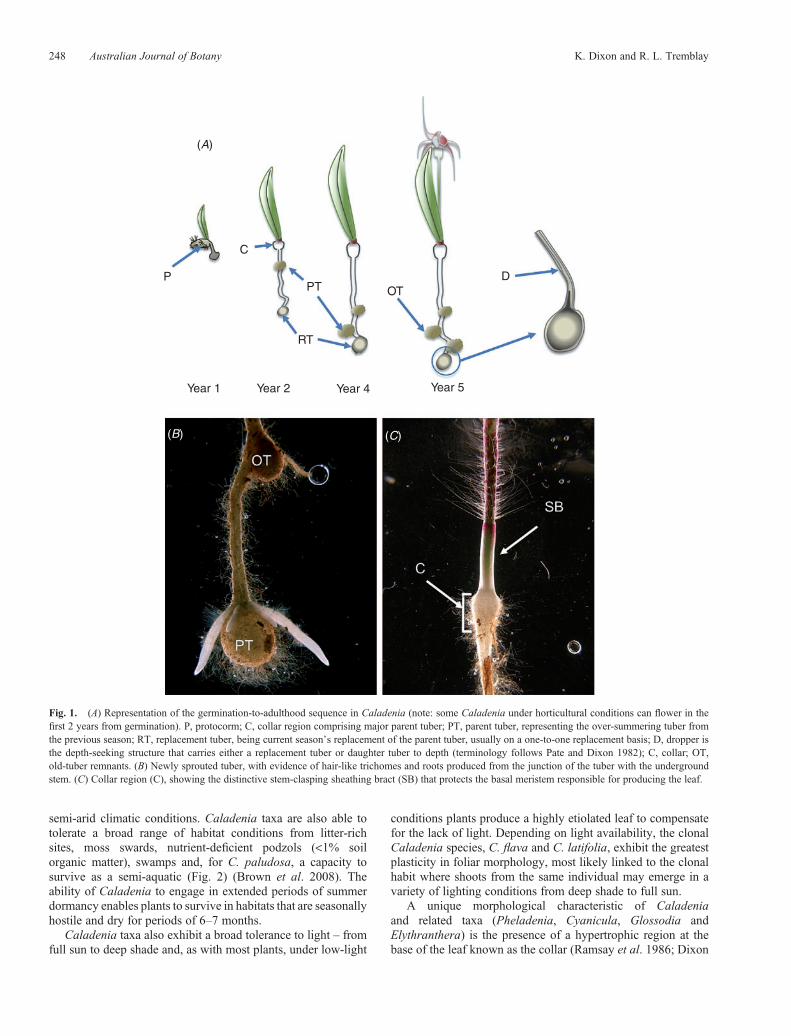
Fig. 1. (A) Representation of the germination-to-adulthood sequence in Caladenia (note: some Caladenia under horticultural conditions can flower in thefirst 2 years from germination). P, protocorm; C, collar region comprising major parent tuber; PT, parent tuber, representing the over-summering tuber fromthe previous season; RT, replacement tuber, being current season’s replacement of the parent tuber, usually on a one-to-one replacement basis; D, dropper isthe depth-seeking structure that carries either a replacement tuber or daughter tuber to depth (terminology follows Pate and Dixon 1982); C, collar; OT,old-tuber remnants. (B) Newly sprouted tuber, with evidence of hair-like trichomes and roots produced from the junction of the tuber with the undergroundstem. (C) Collar region (C), showing the distinctive stem-clasping sheathing bract (SB) that protects the basal meristem responsible for producing the leaf.
248 Australian Journal of Botany K. Dixon and R. L. Tremblay

1991). The collar varies from a few millimetres up to 6mm inlarger-growing Caladenia, is of stem origin and consists of astele and cortex of hypertrophic cells. Unlike most othermycorrhizal structures in the Orchidaceae, the collar regionin Caladenia is differentiated and is swollen beforemycorrhizal occupation of the cortical cells, in ‘anticipation’of mycorrhization when soil moistens.
Leaf morphology varies from thin, almost grass-like, tosomewhat broader and prominently keeled, to short, broad andflattened. Leaves may be scarcely to densely hairy. The exactrole of the hairs on the leaves of Caladenia species is unclear
but may be related to protection from grazing by invertebrates(the long hairs protecting the fleshy leaf lamina below).However, for seasonally arid sites or sites where rainfall issporadic, leaf hairs have been observed to act as structuresthat attract dew (Fig. 3). Dew-feeding of this type is wellknown in other plants of arid regions (Malek et al. 1999) andmay be important in Caladenia for providing supplementalmoisture during dry periods.
All Caladenia species produce a single leaf from a basalmeristem tightly encased in a stem-clasping bract (Fig. 1C).Once the underground shoot arising from the tuber reaches the
(A)
(C) (D)
(B)
(E )
Fig. 2. Habitats for Caladenia range include (A) coastal heathland, (B) forest, (C) swamps, (D) granite rocks and (E) semi-arid zone.
Biology and natural history of Caladenia Australian Journal of Botany 249

light, as found for tuberous Drosera species (see Dixon and Pate1978), the apex initiates a leaf primordium that effectivelyterminates any further stem extension. By the time the leaf isfully extended, the presence of a young, bracteate flower budtucked into the base of the leaf will indicate if flowering is tooccur. Once the leaf has fully expanded, the inflorescenceelongates beyond the sheathing leaf base.
Tuber form and function
The tuber of Caladenia species varies from small, sphericalstructures of less than 5mm to ovoid structures up to 15mm indiameter. Depending on species, the depth of burial of tubersranges from just a few centimetres up to 50 cm in larger-growingplants.
The anatomical nature of the spheroid tubers of terrestrialorchids, including the tubers ofCaladenia, has been the subject ofconsiderable debate. Dressler (1993) proposed that the storagetubers combined features of both roots and stems and werereferred to as root–stem tuberoids; however, this term is not ingeneral use in the orchid literature. The tubers of terrestrial
orchids are distinctive in morphology and anatomy anddifferent from the root and stem tubers of other geophyticplants (Pate and Dixon 1982). In Caladenia, the tuber lacksaxillary perennating buds, with the main bud situated on thetuber apex, meaning that damage to the apical bud may result inthe death of the plant.
Tubers attain depth by using elongated stem-like structuresknown as droppers that position the replacement tuber(or daughter tubers for clonal species) at depth. Droppers areof unclear anatomical origin even though they do represent anoutgrowth of bract tissue containing at its apex a dormant bud(Pate and Dixon 1982; Dressler 1993). The bract tissue forms apartially hollow structure that is capable of rapid extension intothe soil (Fig. 1A). How the dropper in Caladenia senses depth isunclear; however, once depth is attained, the dropper stalk ceasesto extend and the apex of the dropper begins enlarging andfilling with starch, nutrients and water to become the over-summering (dry-season) perennating structure (Pate and Dixon1982). The dropper may alter direction during elongation,resulting in a complex network of tuber connections forminga bead-like arrangement of senesced tuber remnants fromprevious growing seasons.
Seed germination and seedling development
The seed of Caladenia species is similar in form to the dust-likeseed typical of many other terrestrial orchids, comprising a thinpapery testa enclosing an undifferentiated pro-embryonicmass ofcells (Clements 1988). Germination ofCaladenia seed requires acombination of adequatemoisture, suitable temperatures (usually15–20�C) and the presence of a germination (symbiotic)-competent fungus (Batty et al. 2001; Ramsay and Dixon 2003).
Once a mycorrhizal association has been established (Huynhet al. 2004), the seed swells, testa splits and cell divisioncommences usually with intensive intracellular fungalinvestment (usually distal to the micropylar end of the seed)(Clements 1988). Under ideal conditions, irreversible loss ofdormancy occurs in Caladenia within weeks of the seedbecoming moist in the presence of a mycorrhizal fungus(Batty et al. 2001).
Under natural conditions, seed that is exposed to moistureand is not invested by a suitable endophyte usually leads to aloss of seed viability within 3–4 months (Batty et al. 2000). Theshort-lived nature of the soil seed bank of Caladenia points tothe need for annual investment of seed from eachflowering cycle,to ensure seedling germination occurs in favourable seasons. Theloss of a flowering cycle or seeding event because of predation,drought or as a result of spring burning may be crucial forconservation of Caladenia species, particularly rare species.
Little is known of the time taken from seed germination toflowering under habitat conditions. However, studies whereseedlings were germinated under artificial conditions foundflowering can occur 3 years after germination under laboratoryconditions and following transfer to soil under greenhouseconditions (Swarts 2007).
Mycorrhizal associations
Other authors in this Special Issue of Australian Journal ofBotany deal with mycorrhiza in detail, so here we confine
Fig. 3. Specialised hairs on the leaves ofCaladenia species may play a rolein attracting dew to the leaves as a means of supplementing moisture duringdry periods. Hairs on the leaf of C. longicauda ssp. eminens, showing thedistinctive basal cells and highly elongated terminal cell (left).
250 Australian Journal of Botany K. Dixon and R. L. Tremblay
the discussion to a description of the sites of infection andmorphological attributes of the infected plants and organs.
Although more than one endophyte has been found toassociate with Caladenia (Huynh et al. 2004), most studiesconfirm that Caladenia species possess a comparativelyhigh level of fungal specificity compared with other Australianterrestrial orchids (Warcup 1981; Ramsay et al. 1986; Clements1988; Huynh et al. 2004; Bonnardeaux et al. 2007). A recentstudy established a link between potential and ecologicalspecificity in the rare and threatened grand spider orchid,C. huegelii (Swarts 2007), showing for the first time that onefungus, isolated from the collar of an adult plant, remainedassociated with the species from seed germination to floweringplants, including individuals translocated to wild sites. This wasfurther emphasised in the study of Bonnardeaux et al. (2007)where the collar-infection syndrome inCaladenia involvedmorehighly specific endophytic fungi (characterised bymolecular andgermination-efficacy trials) than recorded for root-infectedorchids, including an invasive orchid species.
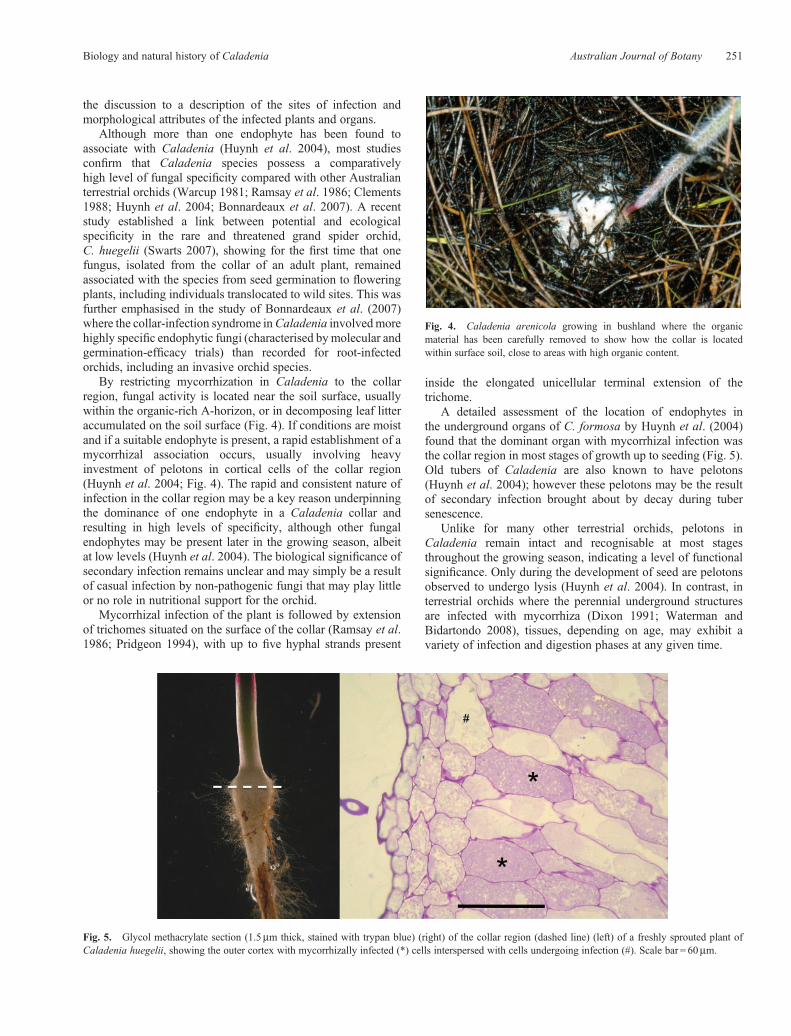
By restricting mycorrhization in Caladenia to the collarregion, fungal activity is located near the soil surface, usuallywithin the organic-rich A-horizon, or in decomposing leaf litteraccumulated on the soil surface (Fig. 4). If conditions are moistand if a suitable endophyte is present, a rapid establishment of amycorrhizal association occurs, usually involving heavyinvestment of pelotons in cortical cells of the collar region(Huynh et al. 2004; Fig. 4). The rapid and consistent nature ofinfection in the collar region may be a key reason underpinningthe dominance of one endophyte in a Caladenia collar andresulting in high levels of specificity, although other fungalendophytes may be present later in the growing season, albeitat low levels (Huynh et al. 2004). The biological significance ofsecondary infection remains unclear and may simply be a resultof casual infection by non-pathogenic fungi that may play littleor no role in nutritional support for the orchid.
Mycorrhizal infection of the plant is followed by extensionof trichomes situated on the surface of the collar (Ramsay et al.1986; Pridgeon 1994), with up to five hyphal strands present
inside the elongated unicellular terminal extension of thetrichome.
A detailed assessment of the location of endophytes inthe underground organs of C. formosa by Huynh et al. (2004)found that the dominant organ with mycorrhizal infection wasthe collar region in most stages of growth up to seeding (Fig. 5).Old tubers of Caladenia are also known to have pelotons(Huynh et al. 2004); however these pelotons may be the resultof secondary infection brought about by decay during tubersenescence.
Unlike for many other terrestrial orchids, pelotons inCaladenia remain intact and recognisable at most stagesthroughout the growing season, indicating a level of functionalsignificance. Only during the development of seed are pelotonsobserved to undergo lysis (Huynh et al. 2004). In contrast, interrestrial orchids where the perennial underground structuresare infected with mycorrhiza (Dixon 1991; Waterman andBidartondo 2008), tissues, depending on age, may exhibit avariety of infection and digestion phases at any given time.
Fig. 4. Caladenia arenicola growing in bushland where the organicmaterial has been carefully removed to show how the collar is locatedwithin surface soil, close to areas with high organic content.
Fig. 5. Glycol methacrylate section (1.5mm thick, stained with trypan blue) (right) of the collar region (dashed line) (left) of a freshly sprouted plant ofCaladenia huegelii, showing the outer cortex with mycorrhizally infected (*) cells interspersed with cells undergoing infection (#). Scale bar = 60mm.
Biology and natural history of Caladenia Australian Journal of Botany 251

Each year, Caladenia shoots emerge from the tuber and growwithin the remains of the stems from thepreviousyear, placing theshoot at the same point in the soil surface (Fig. 6). The abilityof Caladenia to emerge at the same point in the soil columnfrom year to year (thereby locating mycorrhizal tissues in thesame place each year) provides a level of localised enrichment ofmycorrhiza that results in the most favoured site for germinationusually being within close proximity to the parent plant(Batty et al. 2001). Hollick et al. (2007) found that it ispossible to artificially introduce endophytes into habitat siteswith fungal proliferation extending up to 50 cm from theinoculum source, a distance far greater than achieved forendophyte penetration when the inoculum source is derivedsolely from the parent plant.
Phenology of growth and development
Caladenia species usually initiate tuber sprouting under coolerconditions in concert with the onset of winter rainfall. Tubersprouting occurs from mid-autumn in temperate regions, withleaves emerging in late autumn to early winter. Leaves remainactive for 4–8 months, with leaf senescence occurring as a resultof rising temperatures and drying soil (Hopper and Brown 2000;Brown et al. 2008).
Apart from precocious autumn-flowering taxa such asC. drummondii that often flower in dry soil and are therefore
reliant on storedmoisture and nutrients from the over-summeringtuber, most Caladenia taxa flower from winter to early summer.Late-flowering taxa, such a C. corynephora, appear to utilisetuber reserves to support flowering and seed set often in dry soil.For autumn-flowering terrestrial orchids including Caladeniaspecies, it is not clear why the species flower at that time –
whether it is to do with pollinator availability or some ecologicaladvantage associated with production of seed early in thegrowing season.
Many taxa have elevated levels of flowering in response tofire, with some species such as C. reptans having populationsthat flower in profusion after fire (Brown et al. 2008). Post-fireflowering is most pronounced in food-deceptive taxa(e.g. C. flava) where the orchid presumably needs to attractpollinators in the post-fire environment by producing anabundance of flowers. It is thought that ethylene productionduring the passage of the fire front (Dixon and Barrett 2003)may be the agent most responsible for stimulating floralsynchrony in Caladenia species. The horticultural practice ofenclosing a ripened banana in a plastic bag with a pot of summer-dormantCaladenia orchid tubers may elicit a flowering responsesimilar to that of the post-fire environment. Alternatively, theremoval of competing species such as tall-growing grasses mayalso influence the probability of flowering. For example, selectedremoval of competitive plants from orchid habitats has beenshown to directly influence soil temperatures, in addition toaltering the nutrients released after fire that become availablefor the orchid rather than the competing species, as suggested forthe genus Prasophyllum (Coates et al. 2006).
The underground stem, tuber and roots are heavily investedwith trichomes; however, the function of these is not clear as thereis no evidence that these trichomes are directly involved in thesupport of themycorrhizal relationship (Fig. 1B). The productionof roots at depth, at the junctionof the tuber andunderground stem(Fig. 1B), may enable uptake of moisture to sustain the leaf,flowering and seed set, particularly where there is a lack ofrainfall, or for taxa from more arid areas where soil moisturemay reside for longer periods at depth than at the soil surface.Partitioning functional roles between the collar (nutrient uptake)and deeply buried roots (water uptake) may play an importantrole in ensuring the plant has sufficient moisture to supportflowering and seed development, even in the event of dryingof the soil surface. Equally, by sustaining the collar region untilplant senescence may ensure that mycorrhiza inoculum isabundant at the point of stem emergence in the following season.
For most species, seed production occurs in mid-spring tosummer (or late summer for late-flowering taxa), with seeddispersed onto dry soil and incorporated into the soil seedbank until germination occurs with the return of favourablesoil moisture and temperatures (Batty et al. 2000, 2001).
Floral and pollination biology
The genusCaladenia comprises a range of plant sizes and flowerforms, ranging from 30-cm giants in the subgenus Calonemasuch as Caladenia excelsa and members of the C. longicaudagroup, to the diminuitive C. bryceana. Complex floral structuresoccur in such species asC.multiclaviawhere themobile labellum,colourful striping and expanded columnwings add to the unusualform of the flower.
Fig. 6. The multiple layers of previous season’s underground stems havebeen prised open to reveal the stem for the current growing season. Scalebar = 1 cm.
252 Australian Journal of Botany K. Dixon and R. L. Tremblay

Floral form is directly related to pollination syndrome, withCaladenia species embracing self-pollination (C. graminifoliumexhibits high fruit : flower ratios), food-rewarding (C. nana;K. Dixon (pers. comm.) found C. paludosa has parts of thelabellum that exude a smear of sweet nectar – Fig. 7), food-deceptive (C. latifolia has brightly coloured and sweetly scentedflowers) to sexually deceptive systems (typified by theinsectiform labellum in C. barabarossa) (Salzmann et al.2006). Although studies are underway (P. Phillips, pers.comm.), it is apparent that sexual deceit as found inC. barbarossa (with a prominent insectiform, hinged labellum,akin to the labellum found in hammer orchids, Drakaea wherepollination rates are high) may be more successful in achievinghigher pollination success than is food deception.
A recent study (See 2006) found a continuum of pollinationsuccess across a range of orchid species, including Caladenia,from high levels in sexually deceiving taxa, to lower pollinationlevels in food-deceptive taxa. See (2006) also found that therare grand spider orchid, C. huegelii, exhibited very low seedset (<4%) across all populations investigated, including whenflowers were presented across a range of sites. Such lowsuccess in a sexually deceiving species (anecdotal reportscite male wasp visiting flowers of C. heugelii) may relate tothe rarity of the species, loss of habitat (a major contributingfactor in the rarity of the species) or, alternatively, suggestthat a form of bet-hedging is in operation where the prominentfloral display (large colourful petals and sepals) may beassociated with a degree of food-attraction (whereas thepresence of floral clubs and red-blotching of the labellum arelinked to sexual-deceit mechanisms (Brown et al. 2008)). Theconsequence of this form of bet-hedging may be a result ofthe sexual and food deception modes being only partiallyperfected and therefore not exactly matched to requirementsof a particular suite of dedicated pollinators. This may,therefore, result in lower than expected pollination success(See 2006) and may be a contributing factor in the rarity ofC. huegelii.
Once pollination has occurred, floral senescence occursquickly, with the ovary maturing during 4–6 weeks for speciesfrom temperate regions. The capsule splits to release up to 30 000seeds for the larger-flowered species; however, less than 1% ofseeds are thought to result in seedling establishment (Batty et al.2000, 2001).
Low pollination success in some Caladenia species may bebalanced by the ability to engage in belowground vegetativepropagation (Brown et al. 2008). Two forms of vegetative growthare recorded in Caladenia species (Fig. 8). Expansive clonalspread where daughter tubers are produced distally from theparent tuber (e.g. C. flava, C. latifolia) (Brown et al. 2008).Alternatively, in some taxa, proximal production of daughtertubers on short, vertical droppers will occasionally arise from justbelow the collar region, resulting in localised clustering of plants(e.g. C. filamentosa complex, C. reptans) (Pate and Dixon 1981;fig. 3.22, image 145i in Pate and Dixon 1982).
Population biology
The viability of a species depends on understanding thosevariables that influence survivorship of individuals andpopulations. This requires species to be perceived as
interacting within their environment in addition to thedynamics of individual growth, development and reproductionin time and space. A model that is helpful in integrating all ofthese variables has been proposed for plants (Schemske et al.1994) and for orchids (Tremblay and Otero 2009; Fig. 9).Naturally, in only a few cases (if ever) will we be able to learnall there is to know about a species to evaluate objectivelythe relative importance of each ecological and environmentalvariable that influences the survivorship of a species.Nevertheless, understanding the general concept ofinteractions often helps in making best guess estimates of therelative importance of particular variables. The challenge forevolutionary biologist as well as conservation biologists is tounderstand these processes (Tremblay et al. 2005). For example,how allele frequency influences inbreeding depression orhow changes in environmental conditions (i.e. climate change)influence recruitment? Only when these processes areunderstood, we will be able to start evaluating long-term trends.
The position of a flowering plant in the plant community mayexert a substantial influence on individual fitness in Caladeniaspecies. For example, when plants of Caladenia behrii are in thevicinity of the grass tree, Xanthorrhorea semiplana, thelikelihood of herbivory is reduced; however, the probability ofreproductive success is also greatly reduced (Petit and Dickson2005). In an experiment evaluating factors influencing success ofoutplanting of Caladenia into urban bushland, survivorship wasdirectly affected by ecological features such as slope, aspect andweed cover (Scade et al. 2006). Consequently, ecologicalcommunity context can have variable effects on lifetimereproductive success of individuals. Notwithstanding, differenttypes of ecological communities can influence successionaloutcome of orchids, including Caladenia species (Grant andKoch 2003).
Reproductive success is influenced, in part, by the environmentand morphological characters of the orchid. Dickson andPetit (2006) found that inflorescence height influences theprobability of a fruit set with taller inflorescences beingmore successful than shorter inflorescences. The preference bypollinators for inflorescence height may drive selection for specieswith higher inflorescences (Peakall and Beattie 1993, 1996).Reproductive success in Caladenia can also be influenced by thesize of the inflorescence; although not common, some specieshave multi-flowered inflorescences. For C. gracilis, which isreported to have up to six flowers in an inflorescence, multi-flowered individuals have higher rates of pollinaria removal,pollinaria deposition and fruit set, suggesting that selection ispossible for inflorescence size (Tremblay 2005). Clearly, aholistic approach can provide opportunities for developingthe science underpinning management, conservation andrestoration of Caladenia species. For example, integratinggenetics and pollinator behaviour provides new opportunitiesfor understanding the ecology and evolutionary processes inCaladenia (Peakall and Beattie 1996).
In general, Caladenia species face two major challenges –
enabling dormancy and ensuring a suitable mycorrhizalassociation for seed germination (and survival). Mycorrhiza-assisted germination is well established in many orchids(Bayman et al. 2002; Brundrett et al. 2003). Althoughpersistence of mycorrhizal associates is needed forgermination, it is unclear what level of reliance on mycorrhizal
Biology and natural history of Caladenia Australian Journal of Botany 253

(A) (B)
(C) (D)
(E ) (F )
Fig. 7. Floral variation in Caladenia is related to pollination syndrome, with food-deceptive taxa usually white or brightly coloured and scented; sexualdeceptive taxa are mostly dull coloured, with highly mobile or elastic-hinged labella and often with prominent terminal clubbing (arrow) on the perianth parts,and lack scent. (A) Self-pollinating C. graminifolium; (B) food-deceptive C. longicauda ssp. eminens; (C) nectar-producing C. paludosa (reflective part ofthe labellum is composed of dried, sweet exudates); (D) sexually deceptive C. barbarossa; (E) a wasp visiting C. falcate, with clubs on sepal segments (arrow)(image courtesy of B and B Wells). (F) Variation in sepal clubbing in Caladenia.
254 Australian Journal of Botany K. Dixon and R. L. Tremblay

infection is required to sustain the parent plant (Bayman et al.2002).
At any stage, plants (vegetative or flowering) can die orbecome dormant (Fig. 10). Consequently, the lack ofobservation of a plant from one year to the next does notassign it to death. To ascertain the integrity of the mycorrhizalassociation, plantswould require destructive sampling at differentlife-cycle and growth-cycle phases. This is a particular problemfor rare and endangered species (see other papers in this SpecialIssue). Plants may re-emerge either in the same growth stage,grow to the next stage, regress to a previous stage or fail to emerge(failure to emerge may not equate with a lack of undergroundgrowth and development). It seems that non-emergent‘dormancy’ (during the growing season) in Caladenia is onlyas long as 1 or 2 years. What are the proximal cues for the orchid
to remain in a stage, regress or become dormant? These areimportant components in the development of models ofCaladenia growth, development and survival.
Implications for conservation of Caladenia
Caladenia represent Australia’s most threatened terrestrialorchid genus in terms of the number of threatened species. Forexample, Caladenia represent 20 of the 36 orchids listed asthreatened for Western Australia (Anonymous 2008), whichreflects in part the ecological vulnerability of the growth,development and reproductive life-cycle stages for the genus.Ecological components of Caladenia and the enviroments inwhich they grow that directly impinge on survival include:
(A) (B)
(C) (D)
Fig. 8. Vegetative propagation inCaladeniaoccurs either through (A,B) productionof daughter plants at a certain distance from theparent plant (C. flava) or (C) where daughter plants are produced on droppers produced from the collar region (C. filamentosa).(D) An example of droppers (arrows) on C. longicauda subsp. eminens.
Biology and natural history of Caladenia Australian Journal of Botany 255
Determinate growth: Production of a single, determinateshoot means that damage or loss of the shoot duringemergence results in the plant failing to produce a leaf (withno potential for re-leafing to occur in the same growing), with
depletion of photosynthate into the replacement tuber.Longer term, repeated loss of the leaf may result in the deathof the plant.
Mycorrhizal associations: Production of the collar at the soilsurface as themain organ of nutrient transfer (via themycorrhiza)increases the vulnerability of the plant to soil disturbance,competition and drought particularly when compared to othermycorrhizal infection syndromes such as root-infected orchids(e.g. Diuris, Thelymitra, Prasophyllum) where root-inhabitingmycorrhiza are protected deep within the soil profile. Also thehigh degree of fungal specificity found in Caladenia may resultin the inability of Caladenia species to engage in ‘ecologicalsubstitution’ or ‘fungal swapping’ as found in other terrestrialorchids (Bonnardeaux et al. 2007) in the event of decline of soilmycorrhiza (Swarts and Dixon 2009). Whereas a high level oforchid mycorrhizal specificity may have assisted in the rapidradiation of the orchid species in combination with pollinatorspecificity (Otero and Flanagan 2006), in human-alteredenvironments, such levels of specificity may lead to decline inorchid abundance and possible extinction.
Pollination syndromes: For those species engaging in ahigh level of pollinator specificity (likely for many species),decline in pollinator abundance, seasonal flying times(human-induced climate change may limit overlap betweenorchid flowering time and pollinator activity) will result in a
Allele no., & frequency
Heterozygosity
evolutionary potential
Natural SelectionGenetic Drift
Ni (t ) Ni (t + 1)
Emmigrants
Environmental Stochasticity
Demographic Stochasticity
Temporal Stochasticity
Ecological Interaction
Reproductive system
pollen & resource limitation
inbreeding depression
Immigrants
S Sd J A
Fig. 9. Model of the interactions and variables that influence population size and viability (modified from Schemske et al.1994). Ni (t) = population at time 1 and Ni (t+ 1) = the population size at the next time step. S = seeds, Sd = seedlings,J = juveniles, A = adults.
Mycorrhizal ecology
Seedings
Death Death Death
Death
Vegetativeindividuals
Floweringindividuals
Dormant
Sexual reproduction
Fig. 10. Schematic diagram of the key life-cycle attributes of Caladenia.
256 Australian Journal of Botany K. Dixon and R. L. Tremblay

loss of reproductive output. Pollinator limitation is likely to beprevalent in Caladenia species given the high level of highlyspecific deception mechanisms recorded for the genus (Brownet al. 2008). Combined with a low incidence of vegetativepropagation for the genus (3% of Caladenia taxa engage invegetative propagation) results in a reliance on seed-basedrecruitment to replace loss of mother plants.
Translocation science: Conservation involving translocationactivities are compounded by complexities associated withsuccessful transfer of Caladenia species to soil. A combinationof low seedling survival and vulnerability of plants to grazingand predation when transferred to wild sites, as well assusceptibility to variability in moisture levels, results in low orerratic seedling success (Dixon 1992; Quay et al. 1995; Battyet al. 2001, 2002; Scade et al. 2006). Hollick et al. (2007)found that it is possible to substantially improve survival andgermination efficacy in Caladenia through the introductionof artificial inoculum and supplemental watering, offeringsome hope that with further research it will be possibleto overcome the difficulties associated with transfer of plantsto wild sites.
Habitat alteration: Managing the complex array of biotic andabiotic ecological interactions known in Caladenia particularlywhen ecosystems where Caladenia occur are often highlydegraded presents particular challenges in restoration ecology.This is the case for urban bushland areas that are vulnerable torapid ecological changes owing to invasive species, fire andhabitat alteration – all factors that rapidly and irrevocably tipthe ecological balance leading to the decline ofCaladenia species(Scade et al. 2006).
Climate change: The conservation of Caladenia isfurther compounded by the, as yet unknown, impacts ofglobal climate change. Given the high level of specific bioticinteractions (pollinator and mycorrhiza) and the limitedreproductive resilience of Caladenia (few species are clonal;annual seedling recruitment is low), the capacity for Caladeniaspecies to migrate to new, climatically suitable sites, particularlyin highly fragmented landscapes such as in southwest Australia,the home ofCaladenia species diversity,maymeanmany speciesface an uncertain future.
Clearly, future conservation challenges will involvedevelopment of representative seed and mycorrhiza genebanksas an emergency response to impending extinction events.Regrettably, for those species likely to suffer long-termimpacts from climate change, including loss of their home-range, the development of effective translocation programsthrough assisted migration (McLachlan et al. 2007) may be thelast resort for retaining Caladenia in the wild.
References
Anonymous (2008) Wildlife conservation (rare flora) notice 2008(2).Government Gazette, WA, The Government of Western Australia.
Batty AL, Dixon KW, Sivasithamparam K (2000) Soil seed bank dynamicsof terrestrial orchids. Lindleyana 15, 227–236.
Batty AL, DixonKW, BrundrettMC, SivasithamparamK (2001) Constraintsto symbiotic germination of terrestrial orchid seed in a mediterraneanbushland. New Phytologist 152, 511–520.doi: 10.1046/j.0028-646X.2001.00277.x
Batty AL, Dixon KW, Brundrett MC, Sivasithamparam K (2002) Orchidconservation and mycorrhizal associations. In ‘Microorganisms in plantconservation and biodiversity’. (Eds K Sivasithamparam, KW Dixon,RL Barrett) pp. 195–226. (Kluwer Academic Publishers: Dordrecht,The Netherlands)
Bayman P, Gonzalez EJ, Fumero JJ, Tremblay RL (2002) Are funginecessary? How fungicides affect growth and survival of the orchidLepanthes rupestris a in the field. Journal of Ecology 90, 1002–1008.doi: 10.1046/j.1365-2745.2002.00733.x
Bonnardeaux Y, Brundrett M, Batty AL, Dixon KW, Koch J,Sivasithamparam K (2007) Diversity of mycorrhizal fungi of terrestrialorchids: compatibility webs, brief encounters, lasting relationships andalien invasions. Mycological Research 111, 51–61.doi: 10.1016/j.mycres.2006.11.006
Brown AP, Dundas P, Dixon KW, Hopper SD (2008) ‘An illustrated guideto the orchids of Western Australia.’ (University of Western AustraliaPress: Perth)
Brundrett MC, Scade A, Batty AL, Dixon KW, Sivasithamparam K (2003)Development of in situ and ex situ seed baiting techniques to detectmycorrhizal fungi from terrestrial orchid habitats.Mycological Research107, 1210–1220. doi: 10.1017/S0953756203008463
Clements MA (1988) Orchid mycorrhizal associations. Lindleyana 3,73–86.
Coates F, Lunt I, Tremblay RL (2006) The effect of fire, rainfall, herbivoryon population dynamics of a threatened terrestrial orchid from south-eastern Australia, Prasophyllum correctum D.L., and consequences formanagement. Biological Conservation 129, 59–69.doi: 10.1016/j.biocon.2005.06.037
Dickson CR, Petit S (2006) Effect of individual height and labellum colouron pollination in Caladenia (syn. Arachnorchis) behrii (Orchidaceae)in the northern Adelaide region, South Australia. Plant Systematics andEvolution 262, 65–74. doi: 10.1007/s00606-006-0472-3
Dixon KW (1991) Seeder/clonal concepts in Western Australian orchids.In ‘Population ecology of terrestrial orchids’. (Eds TCE Wells,JH Willems) pp. 111–124. (SPB Academic Publishing: The Hague)
Dixon KW (1992) Orchids of Western Australia. An introduction to thepropagation and cultivation of Western Australian orchids. OrchidReview 100, 77–82.
Dixon KW, Barrett RL (2003) Defining the role of fire in south-westWesternAustralian plants. In ‘Fire in ecosystems of south-westWesternAustralia:impacts and management’. (Eds I Abbott, N Burrows) pp. 205–223.(Backhuys Publishers: Leiden, The Netherlands)
Dixon KW, Pate JS (1978) Phenology, morphology and reproductivebiology of the tuberous sundew, Drosera erythrorhiza Lindl.Australian Journal of Botany 26, 441–454. doi: 10.1071/BT9780441
Dressler R (1993) ‘Phylogeny and classification of the orchid family.’(Dioscorides Press: Portland, OR)
Grant CD, Koch J (2003) Orchid species succession in rehabilitated bauxitemines in Western Australia. Australian Journal of Botany 51, 453–457.doi: 10.1071/BT02127
Hollick PS, McComb JA, Dixon KW (2007) Introduction, growth andpersistence in situ of orchid mycorrhizal fungi. Australian Journalof Botany 55, 665–672. doi: 10.1071/BT06073
Hopper SD, Brown AP (2000) New genera, subgenera, combinations, andspecies in the Caladenia alliance (Orchidaceae: Diurideae). Lindleyana15, 120–126.
Huynh TT, McLean CB, Coates F, Lawrie AC (2004) Effect ofdevelopmental stage and peloton morphology on success in isolationof mycorrhizal fungi in Caladenia formosa (Orchidaceae). AustralianJournal of Botany 52, 231–241. doi: 10.1071/BT03099
Koopowitz H, Lavarack PS, Dixon KW (2003) The nature of threatsto orchid conservation. In ‘Orchid conservation’. (Eds KW Dixon,SP Kell, RL Barrett, PJ Cribb) pp. 25–42. (Natural HistoryPublications: Kota Kinabalu, Sabah, Malaysia)
Biology and natural history of Caladenia Australian Journal of Botany 257

Kores PJ, Molvray M, Weston PH, Hopper SD, Brown AP, Cameron KM,ChaseMW (2001) The phylogenetic analysis of Diurideae (Orchidaceae)based on plastid DNA sequence data. American Journal of Botany 88,1903–1914. doi: 10.2307/3558366
Malek E, McCurdy G, Giles B (1999) Dew contribution to the annual waterbalances in semi-arid desert valleys. Journal of Arid Environments 42,71–80. doi: 10.1006/jare.1999.0506
McLachlan JS, Hellman JJ, Schwartz MW (2007) A framework for debateof assisted migration in an era of climate change. ConservationBiology 21, 297–302. doi: 10.1111/j.1523-1739.2007.00676.x
Otero JT, Flanagan NS (2006) Orchid diversity – beyond deception.Trends in Ecology & Evolution 21, 64–65.doi: 10.1016/j.tree.2005.11.016
Pate JS, Dixon KW (1981) Plants with fleshy underground storage organs –a Western Australian survey. In ‘Biology of Australian plants’.(Eds JS Pate, JJ McComb) pp. 181–215. (University of WesternAustralia Press: Perth)
Pate JS, Dixon KW (1982) ‘Tuberous, cormous and bulbous plants –
biology of an adaptive strategy.’ (University of Western AustralianPress: Perth)
Peakall R, Beattie AJ (1993) Pollinators discriminate among floral heightsof a sexually deceptive orchid: implication for selection. Evolution 47,1681–1687. doi: 10.2307/2410212
Peakall R, Beattie AJ (1996) Ecological and genetic consequences ofpollination by sexual deception in the orchid Caladenia tentaculata.Evolution 50, 2207–2220. doi: 10.2307/2410692
Petit S, Dickson CR (2005) Grass-tree (Xanthorrhoea semiplana, Liliaceae)facilitation of the endangered pink-lipped spider orchid (Caladenia syn.Arachnorchis behrii, Orchidaceae) varies in South Australia. AustralianJournal of Botany 53, 455–464. doi: 10.1071/BT04034
PridgeonAM(1994)Systematic leaf anatomyofCaladeniinae (Orchidaceae).Botanical Journal of the Linnean Society 114, 31–48.
Quay L, McComb JA, Dixon KW (1995) Methods for ex vitro germinationof Australian terrestrial orchids. HortScience 30, 1445–1446.
Ramsay M, Dixon KW (2003) Propagation science, recovery andtranslocation of terrestrial orchids. In ‘Orchid conservation’.(Eds KW Dixon, SP Kell, RL Barrett, PJ Cribb) pp. 25–42. (NaturalHistory Publications: Kota Kinabalu, Sabah, Malaysia)
Ramsay RR, Dixon KW, Sivasithamparam K (1986) Patterns of infectionand endophytes associated with Western Australian orchids. Lindleyana1, 203–214.
Salzmann CC, Brown A, Schiestl FP (2006) Floral scent emission andpollination syndrome: evolutionary changes from food to sexualdeception. International Journal of Plant Sciences 167, 1197–1204.doi: 10.1086/508022
Scade A, Brundett MC, Batty AL, Dixon KW, Sivasithamparam K (2006)Survival of transplanted terrestrial orchid seedlings in urban bushlandhabitat with high or low weed cover. Australian Journal of Botany 54,383–389. doi: 10.1071/BT04025
Schemske DW, Husband BC, Ruckelshaus MH, Goodwillie C, Parker IM,Bishop JG (1994) Evaluating approaches to the conservation of rare andendangered plants. Ecology 75, 584–606. doi: 10.2307/1941718
See J (2006) Investigations into the pollination of Caladenia huegelii andits sister species C. arenicola by thynnid wasps (Tiphiidae: Thynninae).BSc (Honours) Thesis, The University of Western Australia, Perth.
Swarts ND (2007) Integrated conservation of the rare and endangeredterrestrial orchid Caladenia huegelii H.G.Reichb. PhD Thesis,The University of Western Australia, Perth.
Swarts ND, Dixon KW (2009) Terrestrial orchid conservation in the ageof extinction. Annals of Botany, in press. doi: 10.1093/aob/mcp025
Tremblay RL (2005) Larger is better: the effect of floral display onreproductive success in two populations of Caladenia (Stegostyla)gracilis R.Br. Muelleria 22, 77–85.
Tremblay RL, Otero JT (2009) Orchid conservation biology: predictingspecies dynamics and community interactions. In ‘Proceedings of thesecond scientific conference on Andean orchids’. (Eds AM Pridgeon,JP Suárez) pp. 197–207. (Universidad Teecnica Particular de Loja: Loja,Ecuador)
Tremblay RL, Ackerman JD, Zimmerman JK, Calvo R (2005) Variationin sexual reproduction in orchids and its evolutionary consequences:a spasmodic journey to diversification. Biological Journal of the LinneanSociety 84, 1–54. doi: 10.1111/j.1095-8312.2004.00400.x
Warcup JH (1981) The mycorrhizal relationships of Australian orchids.New Phytologist 87, 371–381.doi: 10.1111/j.1469-8137.1981.tb03208.x
Waterman RJ, Bidartondo MI (2008) Deception above, deception below:linking pollination and mycorrhizal biology of orchids. Journalof Experimental Biology 59, 1085–1096.
Manuscript received 6 October 2008, accepted 17 March 2009
258 Australian Journal of Botany K. Dixon and R. L. Tremblay
http://www.publish.csiro.au/journals/ajb




















