
Biological and structural comparison of recombinant phospholipase D toxins from Loxosceles...
13
Toxicon 50 (2007) 1162–1174 Biological and structural comparison of recombinant phospholipase D toxins from Loxosceles intermedia (brown spider) venom $ Rodrigo Ota´vio S. Ribeiro a , Olga Meiri Chaim a,b , Rafael Bertoni da Silveira a,c , Luiza Helena Gremski a,b , Youssef Bacila Sade a , Ka´tia Sabrina Paludo a,d , Andrea Senff-Ribeiro a , Juliana de Moura e , Carlos Cha´vez-Olo´rtegui f , Waldemiro Gremski a,g , Helena B. Nader b , Silvio Sanches Veiga a, a Department of Cell Biology, Federal University of Parana´, Jardim das Ame´ricas 81531-990, Curitiba, Parana´, Brazil b Department of Biochemistry, Federal University of Sa˜o Paulo, Sa˜o Paulo, Brazil c Department of Structural, Molecular Biology and Genetics, State University of Ponta Grossa, Parana´, Brazil d Medical Clinic Discipline, Department of Medicine, Federal University of Sa˜o Paulo, Sa˜o Paulo, Brazil e Department of Basic Pathology, Federal University of Parana´, Jardim das Ame´ricas 81531-990, Curitiba, Parana ´, Brazil f Department of Biochemistry and Immunology, Federal University of Minas Gerais, Belo Horizonte, Minas Gerais, Brazil g Catholic University of Parana´, Health and Biological Sciences Institute, Curitiba, Parana´, Brazil Received 5 July 2007; received in revised form 6 August 2007; accepted 7 August 2007 Available online 14 August 2007 Abstract The clinical features of brown spider bites are the appearance of necrotic skin lesions, which can also be accompanied by systemic involvement, including weakness, vomiting, fever, convulsions, disseminated intravascular coagulation, intravascular hemolysis and renal disturbances. Severe systemic loxoscelism is much less common than the cutaneous form, but it may be the cause of clinical complications and even death following envenomation. Here, by using three recombinant dermonecrotic toxins, LiRecDT1, LiRecDT2 and LiRecDT3 (the major toxins found in the venom), we report the biological, immunological and structural differences for these members of this toxin family. Purified toxins evoked similar inflammatory reactions following injections into rabbit skin. Recombinant toxin treatments of MDCK cells with LiRecDT1 and LiRecDT2 changed cell viability, as evaluated by neutral red uptake and assessment of cell morphology through inverted microscopy, whereas LiRecDT3 caused only residual activity. Differences in cell cytotoxicity triggered by recombinant toxins were confirmed through a human red blood lysis assay, during which LiRecDT1 and LiRecDT2 caused a high degree of hemolysis compared to LiRecDT3, which induced only a small hemolytic effect. ARTICLE IN PRESS www.elsevier.com/locate/toxicon 0041-0101/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.toxicon.2007.08.001 $ Ethical statement: We wish to confirm that there are no known conflicts of interest associated with this publication and there has no significant financial support for this work that could have influenced its outcome.We confirm that we have given the consideration to the protection of intellectual property associated with this work and that there are no impediments to publication, including the timing of publication, with respect to intellectual property. In so doing we confirm that we have followed the regulations of our institutions concerning intellectual property.We further confirm that any aspect of the covered in this manuscript that has involved either experimental animals has been conducted with the ethical approval of all relevant bodies and that such approvals are acknowledged within the manuscript. Corresponding author. Fax: +55 41 3266 2042. E-mail address: [email protected] (S.S. Veiga).
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Biological and structural comparison of recombinant phospholipase D toxins from Loxosceles...

ARTICLE IN PRESS
0041-0101/$ - se
doi:10.1016/j.to
$Ethical stat
significant finan
protection of in
publication, wit
concerning intel
animals has be
manuscript.�CorrespondiE-mail addre
Toxicon 50 (2007) 1162–1174
www.elsevier.com/locate/toxicon
Biological and structural comparison of recombinantphospholipase D toxins from Loxosceles intermedia
(brown spider) venom$
Rodrigo Otavio S. Ribeiroa, Olga Meiri Chaima,b, Rafael Bertoni da Silveiraa,c,Luiza Helena Gremskia,b, Youssef Bacila Sadea, Katia Sabrina Paludoa,d,Andrea Senff-Ribeiroa, Juliana de Mourae, Carlos Chavez-Olorteguif,Waldemiro Gremskia,g, Helena B. Naderb, Silvio Sanches Veigaa,�
aDepartment of Cell Biology, Federal University of Parana, Jardim das Americas 81531-990, Curitiba, Parana, BrazilbDepartment of Biochemistry, Federal University of Sao Paulo, Sao Paulo, Brazil
cDepartment of Structural, Molecular Biology and Genetics, State University of Ponta Grossa, Parana, BrazildMedical Clinic Discipline, Department of Medicine, Federal University of Sao Paulo, Sao Paulo, Brazil
eDepartment of Basic Pathology, Federal University of Parana, Jardim das Americas 81531-990, Curitiba, Parana, BrazilfDepartment of Biochemistry and Immunology, Federal University of Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
gCatholic University of Parana, Health and Biological Sciences Institute, Curitiba, Parana, Brazil
Received 5 July 2007; received in revised form 6 August 2007; accepted 7 August 2007
Available online 14 August 2007
Abstract
The clinical features of brown spider bites are the appearance of necrotic skin lesions, which can also be accompanied by
systemic involvement, including weakness, vomiting, fever, convulsions, disseminated intravascular coagulation,
intravascular hemolysis and renal disturbances. Severe systemic loxoscelism is much less common than the cutaneous
form, but it may be the cause of clinical complications and even death following envenomation. Here, by using three
recombinant dermonecrotic toxins, LiRecDT1, LiRecDT2 and LiRecDT3 (the major toxins found in the venom), we
report the biological, immunological and structural differences for these members of this toxin family. Purified toxins
evoked similar inflammatory reactions following injections into rabbit skin. Recombinant toxin treatments of MDCK cells
with LiRecDT1 and LiRecDT2 changed cell viability, as evaluated by neutral red uptake and assessment of cell
morphology through inverted microscopy, whereas LiRecDT3 caused only residual activity. Differences in cell cytotoxicity
triggered by recombinant toxins were confirmed through a human red blood lysis assay, during which LiRecDT1 and
LiRecDT2 caused a high degree of hemolysis compared to LiRecDT3, which induced only a small hemolytic effect.
e front matter r 2007 Elsevier Ltd. All rights reserved.
xicon.2007.08.001
ement: We wish to confirm that there are no known conflicts of interest associated with this publication and there has no
cial support for this work that could have influenced its outcome.We confirm that we have given the consideration to the
tellectual property associated with this work and that there are no impediments to publication, including the timing of
h respect to intellectual property. In so doing we confirm that we have followed the regulations of our institutions
lectual property.We further confirm that any aspect of the covered in this manuscript that has involved either experimental
en conducted with the ethical approval of all relevant bodies and that such approvals are acknowledged within the
ng author. Fax: +55 41 3266 2042.
ss: [email protected] (S.S. Veiga).

ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–1174 1163
Additionally, biological differences for recombinant toxins were corroborated through mice lethality experiments, which
showed animal mortality after LiRecDT1 and LiRecDT2 treatments, but an absence of lethality following LiRecDT3
exposure. Moreover, in experiments for edema, both the LiRecDT1 and the LiRecDT2 toxins evoked similar results,
causing edema following toxin exposure, whereas LiRecDT3 caused only residual effects. Characterization of antigenic
cross-reactivity using sera against crude venom toxins by immunoWestern blotting and immunodot blotting with
recombinant LiRecDT1, LiRecDT2 and LiRecDT3 compared among themselves pointed to a higher cross-reactivity for
LiRecDT1 compared to LiRecDT2 and LiRecDT3, corroborating structural and antigenic differences for these three
toxins. Finally, evidence for structural differences among the recombinant toxins was strengthened by circular dichroism
spectra, which suggested that the toxins were folded, and not aggregated or denatured proteins.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Brown spider; Venom; Phospholipase D; Recombinant toxins
1. Introduction
Accidents involving brown spiders (Loxosceles
genus) have been associated with clinical manifesta-tions characterized by dermonecrosis with gravita-tional spreading at and near the bite site, and to alesser extent with systemic involvement, such asacute renal failure, intravascular hemolysis, throm-bocytopenia and disseminated intravascular coagu-lation. The spiders of the Loxosceles genus have aworldwide distribution, and accidents have beendescribed on all continents (da Silva et al., 2004;Hogan et al., 2004; Swanson and Vetter, 2006).
Brown spider venom is a colorless and crystallineliquid, and has a complex molecular compositionwith several different toxins. Through proteomicapproaches, it was determined that the venom iscomposed of heterogeneous proteins varying inmolecular mass, but with a predominance of toxinsin the region of 3–40 kDa (da Silveira et al., 2002; deCastro et al., 2004; Machado et al., 2005). Amongthe several toxins described in the venom, thedermonecrotic toxin is capable of stimulatingdermonecrosis, hemolysis, platelet aggregation,renal disturbance, increase in vascular permeability,and dysregulated inflammatory response (da Silvaet al., 2004; Hogan et al., 2004; Swanson and Vetter,2006; Chaim et al., 2006). This toxin was previouslybiochemically characterized in the venom of Lox-
osceles reclusa as sphingomyelinase D, based on thegeneration of ceramide-1-phosphate from sphingo-myelin (Futrell, 1992; da Silva et al., 2004), butbased on biochemical approaches it has now beendiscovered that the enzyme has a broad substratespecificity, hydrolyzing glycerophospholipids andsphingophospholipids to generate lysophosphatidicacid or ceramide-1-phosphate. As a result, it hasbeen suggested that the toxin might more accurately
be described as a phopholipase D to account for itsbroader lipid hydrolyse activity (Lee and Lynch,2005).
Today, by using molecular biology and proteomictechniques, it has been demonstrated that thedermonecrotic toxin is a member of a larger familyof related molecules displaying similar molecularmasses, and amino acid and immunological homo-logies. This conclusion is based on previous studieswith L. reclusa venom, which reported the existenceof 4 isoforms of dermonecrotic toxins with aminoacidic homologies and phospholipase D and der-monecrotic activities (Futrell, 1992). Additionally,two related toxins in the venom of L. gaucho havebeen described that were able to induce dermone-crosis in rabbits (Cunha et al., 2003). Recently,studies with the venom of L. boneti have reportedthe existence of three toxins with amino acidhomology characterized as dermonecrotic toxins(Ramos-Cerrillo et al., 2004). Through two-dimen-sional electrophoresis, mass spectrometry andEdman chemical amino acid sequencing, the venomof L. gaucho has been found to contain at least 11toxins with a high percentage of homology todermonecrotic toxins (Machado et al., 2005).Finally, by using molecular biology methodologies,Binford et al. (2005) analyzed L. arizonica venomand reported three cDNA sequences with highhomology to dermonecrotic toxins. In addition,using cDNA libraries from the L. intermedia venomgland, the cloning and expression of differentisoforms of members of dermonecrotic toxin familyhave been described (Kalapothakis et al., 2002;Chaim et al., 2006; da Silveira et al., 2006, 2007a).These proteomic and molecular biology datacorroborate and strengthen the idea of the existenceof an intraspecies family of dermonecrotic toxinsand suggest that the noxious activities induced by

ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–11741164
dermonecrotic toxins reflect a synergistic mechan-ism for different toxin isoforms found in the wholevenom.
Herein, by using three previously describedrecombinant dermonecrotic toxin isoforms, LiR-ecDT1, LiRecDT2 and LiRecDT3 (Chaim et al.,2006; da Silveira et al., 2006, 2007a), obtained froma cDNA library of the L. intermedia venom gland,we report differences in structures and immunolo-gical and biological activities of these toxins,suggesting that, despite molecular amino acididentities, members of this family possess differentactivities.
2. Methods
2.1. Reagents
Polyclonal antibodies to L. intermedia wholevenom toxins were produced in rabbits (Harlowand Lane, 1988). IgGs were purified from serumusing a mixture of Protein-A and Protein-GSepharose beads (Amersham Biosciences, Piscat-away, USA) as recommended by the manufacturer.Whole venom from L. intermedia was extractedfrom spiders captured in the wild following Feitosaet al. (1998).
2.2. Recombinant protein expression
The venom gland cDNA library was builtfollowing Chaim et al. (2006) and da Silveira et al.(2006). GenBank data deposition information forL. intermedia cloned cDNAs are: LiRecDT1,DQ218155, LiRecDT2, DQ266399 and LiRecDT3,DQ267927. For negative control, a recombinanttoxin with similar molecular mass and obtainedfrom the same cDNA library was used. This toxinwas characterized as an ‘‘Astacin-like metallopro-tease’’ (da Silveira et al., 2007b) and it causes nodermonecrosis evidenced by dermonecrotic assay,following rabbit injection into the skin (data notshown). The cDNAs encoding for the putativemature dermonecrotic proteins LiRecDT1,LiRecDT2 and LiRecDT3 were amplified by PCRusing primers designed to contain Xho I restrictionsites at the 50-ends (forward primers) and BamH Isites (reverse primers). The PCR products weredigested with Xho I and BamH I restriction enzymesand subcloned into pET-14b (Novagen, Madison,USA) digested with the same enzymes. The recom-binant constructs were expressed as fusion proteins
with a 6x His-Tag at the N-terminus and a 13 aminoacid linker including a thrombin site between the 6xHis-Tag and the mature protein. The expressionconstructs were transformed into Escherichia coli
BL21(DE3)pLysS cells and plated on LB platescontaining 100 mg/ml ampicillin and 34 mg/ml chlor-amphenicol. Single colonies of the LiRecDT1,LiRecDT2 and LiRecDT3 constructs were inocu-lated into LB broth (100 mg/ml ampicillin and 34 mg/ml chloramphenicol) and grown overnight at 37 1C.These cultures were diluted 1:100 into 1L fresh LBbroth/ampicillin/chloramphenicol and incubated at37 1C until they achieved OD550 nm ¼ 0.5. Recombi-nant expression was induced by the addition of0.025mM IPTG (isopropyl b-D-thiogalactoside)and cells were incubated for 3.5 h at 30 1C. Cellswere harvested by centrifugation (4000g, 7min,4 1C), resuspended in 40ml of extraction buffer(50mM sodium phosphate pH 8.0, 500mM NaCl,10mM imidazole, 1mg/ml lysozyme) and frozen at�20 1C overnight.
2.3. Protein purification
Cells were thawed and disrupted by 6 cycles of10 s sonications at low intensity. Lysed material wascentrifuged (20,000g, 20min, 4 1C), and the super-natants were incubated with 1ml Ni2+-NTAagarose beads for 1 h at 4 1C. The suspensions wereloaded onto a column, and the packed gel waswashed with a buffer containing 50mM sodiumphosphate pH 8.0, 500mM NaCl and 20mMimidazole. Recombinant proteins were eluted with10ml of elution buffer (50mM sodium phosphatepH 8.0, 500mM NaCl, 250mM imidazole) and 1mlfractions were collected and analyzed by 12.5%SDS-PAGE under reducing conditions. Fractionswere pooled and dialyzed against phosphate buf-fered saline (PBS).
2.4. Animals
Adult Swiss mice (25–30 g) and adult rabbitsweighing approximately 3 kg from the CentralAnimal House of the Federal University ofParana were used for in vivo experiments withwhole venom and recombinant toxins. All proce-dures involving animals were carried out in accor-dance with ‘‘Brazilian Federal Laws’’, following theEthical subcommittee on research animal careagreement number 126 of the Federal Universityof Parana.

ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–1174 1165
2.5. Histological methods for light microscopy
Rabbit skin from animals that intradermallyreceived recombinant toxins was collected follow-ing anesthetization with ketamine (Agribands,Campinas, Brazil) and acepromazine (Univet, SaoPaulo, Brazil), and was then fixed in ‘‘ALFAC’’(absolute ethanol 85%, formaldehyde 10% andglacial acetic acid 5%) for 16 h at room tempera-ture. After fixation, samples were dehydrated in agraded series of ethanol before paraffin embedding(for 2 h at 58 1C). Then, thin sections (4 mm)were processed for histology. Tissue sections werestained by hematoxylin and eosin (HE) (Cullinget al., 1985).
2.6. Cell cytotoxicity assays
MDCK cells (‘‘Madin-Darby Canine Kidney’’epithelial cells) were grown in monolayer culturesin DMEM-F12 medium containing penicillin(10,000 IU/ml) supplemented with 10% fetal calfserum (FCS). The cultures were kept at 37 1C in ahumidified atmosphere plus 5% CO2. Releaseof cells was performed by treating with a2mM solution of ethylenediaminetetraacetic acid(EDTA) in cation-free/PBS and 0.05% trypsinfor a few minutes. After counting, the cells wereresuspended in an adequate volume of mediumsupplemented with FCS and allowed to adhere andgrow for 24 h. Cells were then evaluated in thepresence or absence of toxins (10 and 50 mg/ml).During the experiments, the plates were photo-graphed at 24 and 48 h using an inverted micro-scope (Leica-DMIL, Wetzlar, Germany), andchanges in cell morphology were evaluated. Alter-natively, cytotoxicity assays were carried out on96-well plates (TPP, Trasadingen, Switzerland).Cells (5� 103 cells/well) were plated and allowedto adhere and grow for 24 h before incubationwith recombinant toxins at concentrations of10 and 50 mg/ml for 24 and 48 h in hexaplicate.After toxin incubation, the measurement oftoxicity was performed by estimation of viabilityby neutral red uptake (Merck, Darmstadt,Germany) as described by Freshney (2000).The same experimental conditions were usedwith a control group except that the mediumcontained adequate amounts of vehicle (PBS)rather than toxins. The cell viability of the controlgroup (in the absence of toxins) was normalizedto 100%.
2.7. Hemolytic assay
To detect the hemolytic activity of recombinantproteins (LiRecDT1, LiRecDT2 and LiRecDT3),an in vitro assay was performed as described byOliveira-Lima and Dias-da-Silva (1970). Briefly,200 ml of ORh+ human erythrocytes with lectureof the hemoglobin absorbance at 540 nm corre-sponding to 0.210 was diluted in TBS-gel (25mMtriethanolamine hydrochloride 125mM NaCl,0.5mM MgSO4, 0.15mM CaCl2, 0.05% gelatin,pH 7.4) and incubated with 200 ml of L. intermedia
crude venom (positive control) or recombinantproteins at different concentrations (25 to 3 mg/ml)for 1 h at 37 1C under conditions of constantgentle shaking. The blood suspension was washedthree times with TBS gel by successive centrifuga-tions (440g/4min), and the erythrocytes wereresuspended in 400 ml of TBS-gel and reincubatedwith 100 ml of ORh+ normal serum for 1 h at 37 1C.The percent hemolysis of the supernatants andcontrols was estimated from the absorbance at540 nm of the hemoglobin released. Total lysis(100%) was obtained with water, and unlyzedORh+ human erythrocytes incubated under thesame conditions, with TBS-gel and serum but in theabsence of venom or recombinant toxins, were usedas control.
2.8. Foot pad edema-forming activity
Analysis of foot pad edema followed Rucavado etal. (1999). Groups of five mice were injectedsubcutaneously (s.c.) in the right foot pad withLiRecDT1, LiRecDT2 and LiRecDT3 or withvenom (5 mg dissolved in 20 ml PBS). Control micewere injected with 20 ml PBS. Edema was estimatedby the change in foot-pad thickness, measured inmicrometers.
2.9. Mouse intraperitoneal injections
Mouse mortality studies were conductedwith adult Swiss (25–30 g) animals. Wholevenom or purified recombinant toxins (50 and100 mg/kg of mice) were each injected intraperito-neally into five mice. A recombinant toxin withoutdermonecrotic activity was used as a negativecontrol. The mice were observed for 24 and 48 hafter injection, and survival was assessed at 1 hintervals.

ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–11741166
2.10. Gel electrophoresis and immunoblotting
Protein content was determined by the CoomassieBlue method (BioRad, Hercules, USA) followingBradford (1976). For protein analysis, 12.5%SDS-PAGE under reduced conditions followingLaemmli (1970) was performed. The molecularmass markers were acquired from Sigma. Forimmunoblottings, toxins (1.0 mg) were electricallytransferred to nitrocellulose filters followingSDS-PAGE (Towbin et al.,1979), or alternativelybound directly to nitrocellulose sheets (dotted)(Harlow and Lane, 1988) and immunostained byusing purified IgGs which react to whole venomtoxins under the same conditions for all samples.Results from both Western and dot blottingwere quantified with the aid of a Chemidoc System(Bio Rad).
2.11. Circular dichroism spectra measurements
For circular dichroism spectra measurements, therecombinant toxins LiRecDT1, LiRecDT2 andLiRecDT3 were dialyzed at 4 1C against 10mMsodium phosphate buffer, pH 7.4, and spectra wererecorded in a Jasco J-810 spectropolarimeter (JascoCorporation, Tokyo, Japan) using a 1mm gapcuvette. Each spectrum (0.5 nm interval) was theaverage of 8 measurements, performed at a rate of50 nmmin�1 using a response time of 8 s and abandwidth of 1 nm. The temperature was keptconstant at 25 1C. The secondary structures oftoxins were estimated from the spectra using theContin method in Dicroprot 2000 1.0.4 software(Deleage and Geourjon, 1993).
2.12. Statistical analysis
Statistical analyses were performed using analysisof variance (ANOVA) with post-hoc Tukey test inthe GraphPad InStat program version 3.00 forWindows 2000. Differences were considered signifi-cant when pp0.05.
3. Results
3.1. Expression, purification and control for
functionality of recombinant toxins
The recombinant toxins LiRecDT1, LiRecDT2and LiRecDT3 were expressed as N-terminal His-Tag fusion proteins in E. coli BL21(DE3)pLysS
cells. Soluble fractions of bacterial lysates were usedto purify toxins by Ni2+-chelating chromatographyas described previously (Chaim et al., 2006; daSilveira et al., 2006). Fig. 1A depicts the SDS-PAGEprofile of the purified toxins under reducing condi-tions. As shown, at the conditions used, moleculesare soluble and are purified. Additionally, with theaim to avoid criticism related to the lack of activityof toxins during expression and purification, toxinswere injected intradermally into rabbit skin. As anegative control, a recombinant toxin withoutinflammatory activity (da Silveira et al., 2007b),but purified under the same conditions as forLiRecDT1, LiRecDT2 and LiRecDT3 was used.Tissue samples were collected 24 h following toxinexposure and histologically analyzed by lightmicroscopy. Fig. 1B shows the inflammatory find-ings, emphasizing neutrophils diffusely spread with-in the dermis of animals exposed to dermonecrotictoxins, supporting the existence of biologicalactivities for these molecules.
3.2. Comparative cytotoxicity of recombinant
dermonecrotic toxin isoforms against epithelial cells
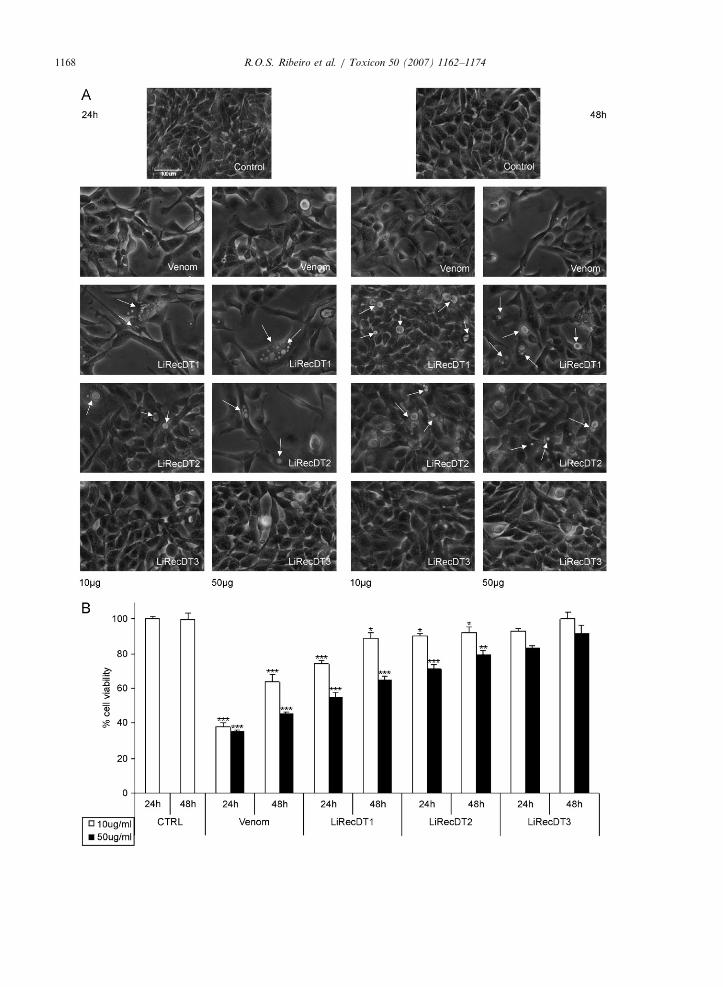
In order to ascertain the cytotoxicity of recombi-nant dermonecrotic toxins against cells in cultureand to support the biological activities of thesetoxins, which may account for cellular injuries aftertreatment with native dermonecrotic toxin frombrown spider venom, we performed experimentsusing kidney epithelial cells (MDCK) andL. intermedia crude venom or three different recom-binant dermonecrotic toxin isoforms LiRecDT1,LiRecDT2 and LiRecDT3. Experiments were car-ried out by observing morphology and cellularviability after toxin exposure. As shown in Fig. 2A,treatment with the recombinant toxin LiRecDT1caused the appearance of cytoplasmic vacuolizationand altered cellular spreading and cell–cell adhe-sion, as well as changes in cell morphology, in atoxin concentration-dependent manner (arrows).LiRecDT2 treatment induced similar cellular altera-tions when compared to the LiRecDT1 isoform, butapparently in a lower degree, whereas LiRecDT3had only a residual activity against cells even whenhigher toxin concentrations were used. Cellularviability experiments assayed through neutral reduptake indicated a significant alteration of epithelialcells following LiRecDT1 and LiRecDT2 treat-ments compared with LiRecDT3 exposure, whichsuggested only a residual activity for LiRecDT3
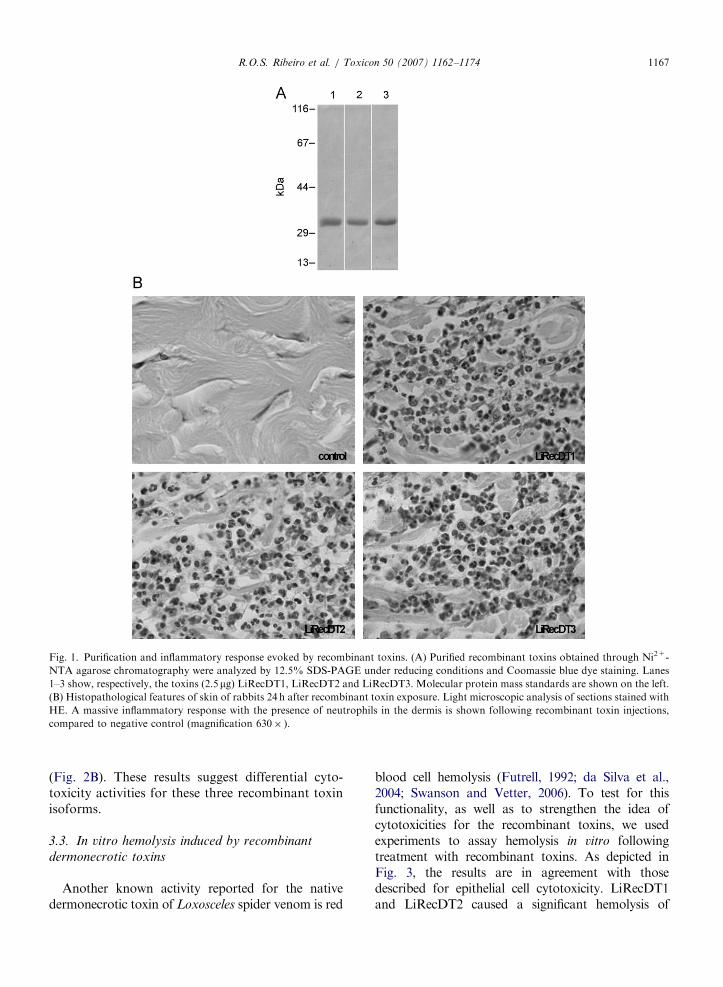
ARTICLE IN PRESS
Fig. 1. Purification and inflammatory response evoked by recombinant toxins. (A) Purified recombinant toxins obtained through Ni2+-
NTA agarose chromatography were analyzed by 12.5% SDS-PAGE under reducing conditions and Coomassie blue dye staining. Lanes
1–3 show, respectively, the toxins (2.5mg) LiRecDT1, LiRecDT2 and LiRecDT3. Molecular protein mass standards are shown on the left.
(B) Histopathological features of skin of rabbits 24 h after recombinant toxin exposure. Light microscopic analysis of sections stained with
HE. A massive inflammatory response with the presence of neutrophils in the dermis is shown following recombinant toxin injections,
compared to negative control (magnification 630� ).
R.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–1174 1167
(Fig. 2B). These results suggest differential cyto-toxicity activities for these three recombinant toxinisoforms.
3.3. In vitro hemolysis induced by recombinant
dermonecrotic toxins
Another known activity reported for the nativedermonecrotic toxin of Loxosceles spider venom is red
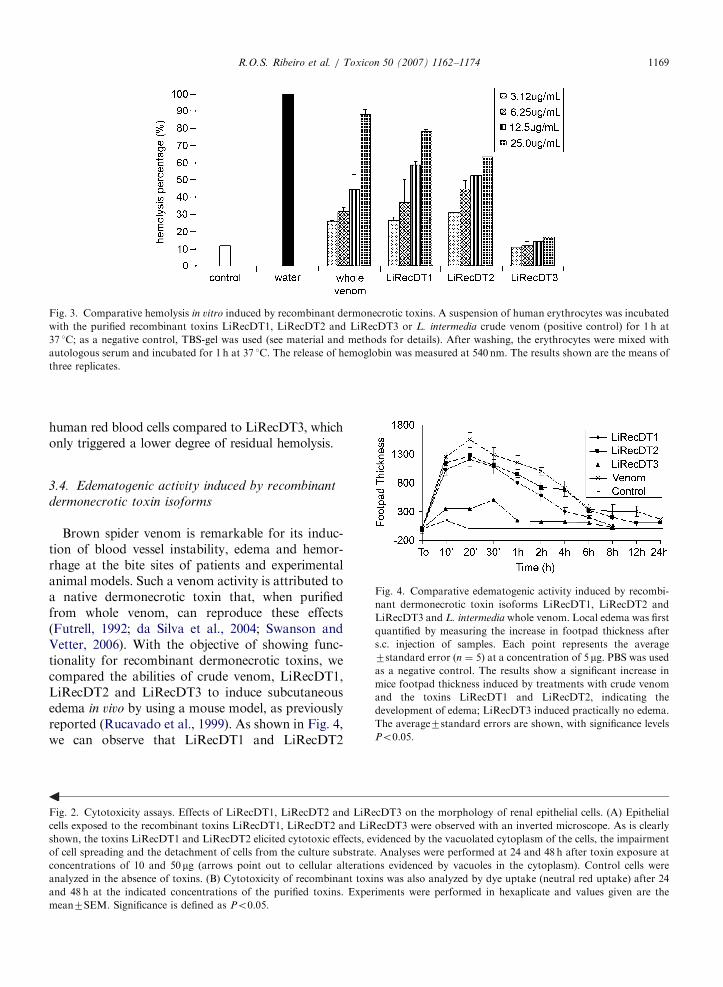
blood cell hemolysis (Futrell, 1992; da Silva et al.,2004; Swanson and Vetter, 2006). To test for thisfunctionality, as well as to strengthen the idea ofcytotoxicities for the recombinant toxins, we usedexperiments to assay hemolysis in vitro followingtreatment with recombinant toxins. As depicted inFig. 3, the results are in agreement with thosedescribed for epithelial cell cytotoxicity. LiRecDT1and LiRecDT2 caused a significant hemolysis of
ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–11741168
ARTICLE IN PRESS
Fig. 3. Comparative hemolysis in vitro induced by recombinant dermonecrotic toxins. A suspension of human erythrocytes was incubated
with the purified recombinant toxins LiRecDT1, LiRecDT2 and LiRecDT3 or L. intermedia crude venom (positive control) for 1 h at
37 1C; as a negative control, TBS-gel was used (see material and methods for details). After washing, the erythrocytes were mixed with
autologous serum and incubated for 1 h at 37 1C. The release of hemoglobin was measured at 540 nm. The results shown are the means of
three replicates.
R.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–1174 1169
human red blood cells compared to LiRecDT3, whichonly triggered a lower degree of residual hemolysis.
Fig. 4. Comparative edematogenic activity induced by recombi-
nant dermonecrotic toxin isoforms LiRecDT1, LiRecDT2 and
LiRecDT3 and L. intermedia whole venom. Local edema was first
quantified by measuring the increase in footpad thickness after
s.c. injection of samples. Each point represents the average
7standard error (n ¼ 5) at a concentration of 5 mg. PBS was used
as a negative control. The results show a significant increase in
mice footpad thickness induced by treatments with crude venom
and the toxins LiRecDT1 and LiRecDT2, indicating the
development of edema; LiRecDT3 induced practically no edema.
The average7standard errors are shown, with significance levels
Po0.05.
3.4. Edematogenic activity induced by recombinant
dermonecrotic toxin isoforms
Brown spider venom is remarkable for its induc-tion of blood vessel instability, edema and hemor-rhage at the bite sites of patients and experimentalanimal models. Such a venom activity is attributed toa native dermonecrotic toxin that, when purifiedfrom whole venom, can reproduce these effects(Futrell, 1992; da Silva et al., 2004; Swanson andVetter, 2006). With the objective of showing func-tionality for recombinant dermonecrotic toxins, wecompared the abilities of crude venom, LiRecDT1,LiRecDT2 and LiRecDT3 to induce subcutaneousedema in vivo by using a mouse model, as previouslyreported (Rucavado et al., 1999). As shown in Fig. 4,we can observe that LiRecDT1 and LiRecDT2
Fig. 2. Cytotoxicity assays. Effects of LiRecDT1, LiRecDT2 and LiRecDT3 on the morphology of renal epithelial cells. (A) Epithelial
cells exposed to the recombinant toxins LiRecDT1, LiRecDT2 and LiRecDT3 were observed with an inverted microscope. As is clearly
shown, the toxins LiRecDT1 and LiRecDT2 elicited cytotoxic effects, evidenced by the vacuolated cytoplasm of the cells, the impairment
of cell spreading and the detachment of cells from the culture substrate. Analyses were performed at 24 and 48 h after toxin exposure at
concentrations of 10 and 50 mg (arrows point out to cellular alterations evidenced by vacuoles in the cytoplasm). Control cells were
analyzed in the absence of toxins. (B) Cytotoxicity of recombinant toxins was also analyzed by dye uptake (neutral red uptake) after 24
and 48 h at the indicated concentrations of the purified toxins. Experiments were performed in hexaplicate and values given are the
mean7SEM. Significance is defined as Po0.05.

ARTICLE IN PRESS
Table 1
Effects of whole venom and recombinant toxins on mice
mortality
Mortality (%) 50 mg 100mg
24 h 48 h 24 h 48 h
Whole venom 3/5 (60%) – – –
LiRecDT1 2/5 (40%) 3/5 (60%) 3/5 (60%) –
LiRecDT2 3/5 (60%) 3/5 (60%) 3/5 (60%) –
LiRecDT3 0/5 (0%) 0/5 (0%) 0/5 (0%) 0/5 (0%)
Negative control 0/5 (0%) 0/5 (0%) 0/5 (0%) 0/5 (0%)
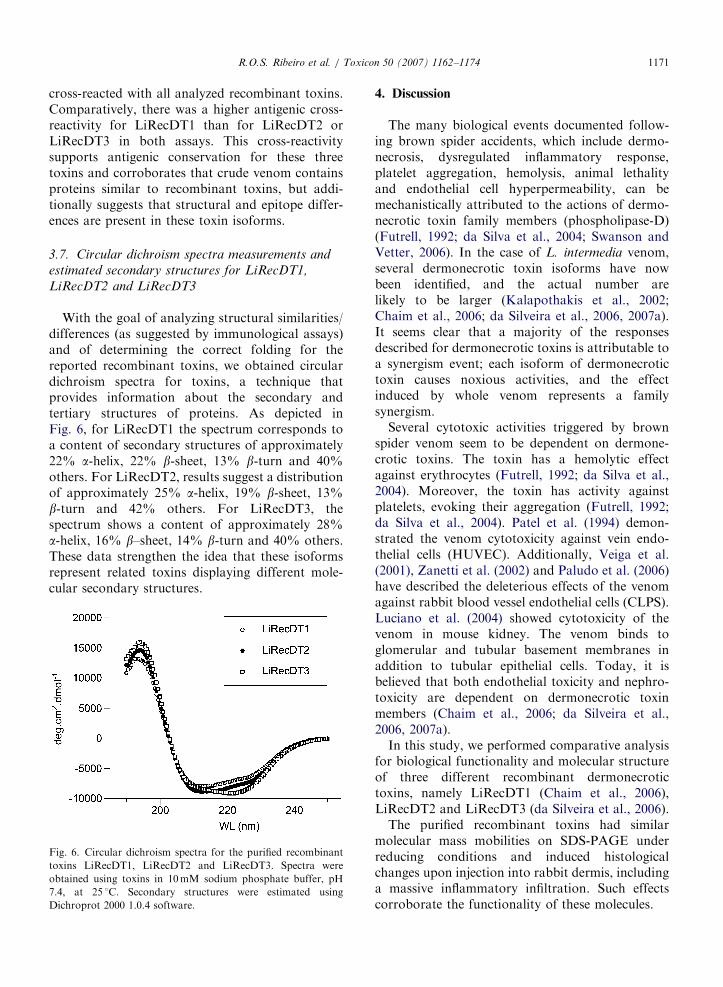
Fig. 5. Immunological cross-reactivity of the recombinant
dermonecrotic toxins LiRecDT1, LiRecDT2 and LiRecDT3.
Crude venom and the purified recombinant toxins LiRecDT1,
LiRecDT2 and LiRecDT3 at a concentration of 1.0mg were
separated by 12.5% SDS-PAGE under reducing conditions,
transferred onto nitrocellulose membranes and exposed to
purified antibodies against crude venom toxins (A). Molecular
mass marker is shown on the left of figure. Additionally, antibody
capture assays (Dot blotting) were carried out using crude venom
and the recombinant toxins LiRecDT1, LiRecDT2 and LiR-
ecDT3 (1.0mg) immobilized on nitrocellulose membranes (B).
Results of blottings were densitometrically quantified and
graphically plotted. In both cases (Western blotting and Dot
blotting), cross-reactivity occurred among antibodies to crude
venom toxins and recombinant toxins, with a major intensity for
LiRecDT1 and LiRecDT2 and a lesser intensity for LiRecDT3.
R.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–11741170
evoked edema in a time-dependent manner, whereasLiRecDT3 displayed only residual activity, in afashion identical to that seen for epithelial cytotoxi-city and hemolysis, as described above.
3.5. Comparative mouse mortality induced by
recombinant toxin isoforms
It is known that mouse mortality due to Lox-
osceles spider venom is attributed to the dermone-crotic fraction of the venom (Futrell, 1992; Barbaroet al., 1996; da Silva et al., 2004). Therefore, todemonstrate the recombinant toxins’ functionalities,Swiss mice received intraperitonial injections ofpurified LiRecDT1, LiRecDT2 and LiRecDT3.Whole venom was used as a positive control, and arecombinant toxin without dermonecrotic or inflam-matory activities obtained in a similar way asdescribed for the dermonecrotic molecules was usedas a negative control (n ¼ 5 per treatment). Thedeath:survival ratio was determined after 24h andafter 48h. Whole venom, LiRecDT1 and LiRecDT2induced mice mortality even at the lower doseassayed after 24h, while the negative control andLiRecDT3 showed no lethal effect (Table 1).
3.6. Comparative immunological cross-reactivity of
recombinant dermonecrotic toxin isoforms
Additionally, with the aim of evaluating somestructural relationship for the recombinant toxinsand of analyzing their importance as immunologicalantigens, we checked recombinant toxins (normal-ized for their protein contents) for antigenic cross-reactivity with a polyclonal antiserum against nativewhole venom toxins by an immunoWestern Blottingassay (denatured epitopes on the antigens) (Fig. 5A)and by immunoDot Blotting (nondenatured epi-topes on the antigens) (Fig. 5B). Reactions were
developed and stopped under the same laboratoryconditions for all samples. Results from the immuno-blotting were densitometrically graded with theaid of a Chemidoc System (Bio Rad). Antibodies
ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–1174 1171
cross-reacted with all analyzed recombinant toxins.Comparatively, there was a higher antigenic cross-reactivity for LiRecDT1 than for LiRecDT2 orLiRecDT3 in both assays. This cross-reactivitysupports antigenic conservation for these threetoxins and corroborates that crude venom containsproteins similar to recombinant toxins, but addi-tionally suggests that structural and epitope differ-ences are present in these toxin isoforms.
3.7. Circular dichroism spectra measurements and
estimated secondary structures for LiRecDT1,
LiRecDT2 and LiRecDT3
With the goal of analyzing structural similarities/differences (as suggested by immunological assays)and of determining the correct folding for thereported recombinant toxins, we obtained circulardichroism spectra for toxins, a technique thatprovides information about the secondary andtertiary structures of proteins. As depicted inFig. 6, for LiRecDT1 the spectrum corresponds toa content of secondary structures of approximately22% a-helix, 22% b-sheet, 13% b-turn and 40%others. For LiRecDT2, results suggest a distributionof approximately 25% a-helix, 19% b-sheet, 13%b-turn and 42% others. For LiRecDT3, thespectrum shows a content of approximately 28%a-helix, 16% b–sheet, 14% b-turn and 40% others.These data strengthen the idea that these isoformsrepresent related toxins displaying different mole-cular secondary structures.
Fig. 6. Circular dichroism spectra for the purified recombinant
toxins LiRecDT1, LiRecDT2 and LiRecDT3. Spectra were
obtained using toxins in 10mM sodium phosphate buffer, pH
7.4, at 25 1C. Secondary structures were estimated using
Dichroprot 2000 1.0.4 software.
4. Discussion
The many biological events documented follow-ing brown spider accidents, which include dermo-necrosis, dysregulated inflammatory response,platelet aggregation, hemolysis, animal lethalityand endothelial cell hyperpermeability, can bemechanistically attributed to the actions of dermo-necrotic toxin family members (phospholipase-D)(Futrell, 1992; da Silva et al., 2004; Swanson andVetter, 2006). In the case of L. intermedia venom,several dermonecrotic toxin isoforms have nowbeen identified, and the actual number arelikely to be larger (Kalapothakis et al., 2002;Chaim et al., 2006; da Silveira et al., 2006, 2007a).It seems clear that a majority of the responsesdescribed for dermonecrotic toxins is attributable toa synergism event; each isoform of dermonecrotictoxin causes noxious activities, and the effectinduced by whole venom represents a familysynergism.
Several cytotoxic activities triggered by brownspider venom seem to be dependent on dermone-crotic toxins. The toxin has a hemolytic effectagainst erythrocytes (Futrell, 1992; da Silva et al.,2004). Moreover, the toxin has activity againstplatelets, evoking their aggregation (Futrell, 1992;da Silva et al., 2004). Patel et al. (1994) demon-strated the venom cytotoxicity against vein endo-thelial cells (HUVEC). Additionally, Veiga et al.(2001), Zanetti et al. (2002) and Paludo et al. (2006)have described the deleterious effects of the venomagainst rabbit blood vessel endothelial cells (CLPS).Luciano et al. (2004) showed cytotoxicity of thevenom in mouse kidney. The venom binds toglomerular and tubular basement membranes inaddition to tubular epithelial cells. Today, it isbelieved that both endothelial toxicity and nephro-toxicity are dependent on dermonecrotic toxinmembers (Chaim et al., 2006; da Silveira et al.,2006, 2007a).
In this study, we performed comparative analysisfor biological functionality and molecular structureof three different recombinant dermonecrotictoxins, namely LiRecDT1 (Chaim et al., 2006),LiRecDT2 and LiRecDT3 (da Silveira et al., 2006).
The purified recombinant toxins had similarmolecular mass mobilities on SDS-PAGE underreducing conditions and induced histologicalchanges upon injection into rabbit dermis, includinga massive inflammatory infiltration. Such effectscorroborate the functionality of these molecules.

ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–11741172
Comparatively, the cytotoxicity observed follow-ing treatment of MDCK epithelial cells with theserecombinant toxins differed according to toxinused. Experiments carried out using MDCK cellstreated in culture with LiRecDT1 showed apotent toxin activity, causing a disturbance of cellmorphology, the appearance of intracellularvacuoles and changes in cellular adhesion to culturesubstratum, cell–cell contacts and cellular spread-ing. Likewise, LiRecDT1 also inhibited cellularviability in a concentration- and time-dependentway, further demonstrating a direct cytotoxicity.MDCK cell treatment with LiRecDT2 showedcytotoxicity, but in a lower degree than thoseevoked by LiRecDT1 in a time- and concentra-tion-dependent manner, whereas LiRecDT3 treat-ment caused only a residual cytotoxicity, suggestingdifferences in biological activities among these toxinisoforms.
Differential functionality for dermonecrotic toxinisoforms was further demonstrated by their hemo-lytic activities. As reported, erythrocyte cytotoxicitywas seen following LiRecDT1 and LiRecDT2treatments, but only a residual activity was reportedafter LiRecDT3 exposure. Similarly, mice lethalityexperiments using recombinant toxins, in whichLiRecDT1 and LiRecDT2 caused animal mortalityand LiRecDT3 was unable to evoke lethality,corroborated the cell cytotoxicity assays.
Additionally, edematogenic assays followingin vivo exposure to recombinant toxins in micedemonstrated similar results for LiRecDT1 andLiRecDT2 toxins, which resembled those evoked bycrude venom. LiRecDT3 caused only a residualeffect. Such results are in agreement with thosedescribed for histopathological analyses in the skinof rabbits using LiRecDT1 and LiRecDT2, includ-ing diffuse edema of the dermis, proteinaceousexudation, and a massive and diffuse collection ofinflammatory cells into the dermis (da Silveira et al.,2006). Apparently, by inducing an inflammatoryresponse and leukocyte infiltration into the dermisof mice (Sunderkotter et al., 2001; da Silva et al.,2004), venom toxins can increase vessel permeabil-ity. Nevertheless, toxin isoform LiRecDT3, in spiteof causing an inflammatory reaction in the dermisfollowed injection, did not trigger edematogenicactivity, suggesting that a direct toxin effect uponendothelia could cause endothelial cell activationand cytotoxicity and generate an increase in vesselpermeability by causing disturbances in the bloodvessel wall, thus explaining the increase of edema.
Such a hypothesis is in agreement with resultspreviously described suggested that endothelialcytotoxicity followed venom exposure (Patel et al.,1994; Veiga et al., 2001; Paludo et al., 2006).
The molecular mechanism by which brown spiderdermonecrotic toxins cause cellular injuries iscurrently under investigation. Although no directcorrelation between phospholipase activity and thenoxious effects evoked after dermonecrotic toxinexposure is known, we can speculate on somecorrelations for such effects, since LiRecDT1 andLiRecDT2 possess high phospholipase activitycompared to LiRecDT3, which has low activity(da Silveira et al., 2006). This hypothesis isstrengthened by the fact that brown spider venomphospholipases can generate lysophosphatidic acidor sphingosine-1-phosphate, which are known toinduce several pathological responses, includingpro-inflammatory responses, platelet aggregationand endothelial hyperpermeability (Futrell, 1992;da Silva et al., 2004; Moolenaar et al., 2004; Leeand Lynch, 2005). Interestingly, while LiRecDT1,LiRecDT2 and LiRecDT3 have different activities,the amino acid residues involved in catalysis forphospholipase activity (Murakami et al., 2005) areconserved. A putative explanation for the differ-ences described could be that the amino acidresidues of the toxins near to the catalytic domainshare high sequence identity for LiRecDT1 andLiRecDT2, but not for LiRecDT3 (da Silveira et al.,2006). An explanation for the differences describedcould be inferred from phylogenetic tree analysis,which suggests that LiRecDT1 and LiRecDT2belong to related sub-groups, whereas LiRecDT3belongs to a different sub-group together withL. boneti and L. laeta toxins (da Silveira et al.,2006).
Based on crystallographic results obtained with arecombinant dermonecrotic toxin isoform fromL. laeta venom, as well as on structural andsequence alignment comparisons (Murakami et al.,2006), a classification system for dermonecrotictoxin isoforms has been proposed, which takes intoaccount their phospholipase D activities. Accordingto the authors, brown spider venom dermonecrotictoxins can be divided into molecules of class I(those containing just one intra-chain disulfide bondand one extended hydrophobic loop), as is the casefor the L. laeta isoform analyzed, and class IImolecules, which contain an additional disulfidebond linking the catalytic loop to a second flexibleloop. The class II phospholipases can be subdivided

ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–1174 1173
into class IIa and class IIb (according to their abilityor inability to hydrolyze sphingomyelin, respec-tively). Based on this classification, LiRecDT1 andLiRecDT2 belong to class IIa since they possess twoputative disulfide bonds and are able to hydrolyzesphingomyelin, whereas LiRecDT3 is classified intoclass IIb since it poorly hydrolyzes sphingomyelin(da Silveira et al., 2006).
In agreement with the above-discussed ideas, thecircular dichroism results from spectra obtainedwith LiRecDT1, LiRecDT2 and LiRecDT3 stronglysuggest a structure–function relationship amongthese three toxins. For LiRecDT1 and LiRecDT2,experiments demonstrated significant amino acidicsecondary structure homology, whereas LiRecDT3showed significant differences, with more aminoacids present in alpha helical structures and fewerpresent in beta sheets. From circular dichroismspectra, we can conclude a correct folding forrecombinant toxins reported. The results are typicalof folded non-denatured and non-aggregated pro-teins, which eliminates the criticism that non-foldedstructures may be present for the recombinanttoxins. On the basis of CD spectra, we mayspeculate that the decrease of biological effectsdisplayed by LiRecDT3 despite the presenceof a catalytic site in this recombinant toxin isinfluenced by differences in secondary structures.For LiRecDT3, these secondary structure differ-ences may lead to differences in tertiary structureand alter interaction with lipid substrates andbiological activity.
Different isoforms of the same toxin mayrepresent a biological mechanism of defense againstenvironmental influences, which under differentconditions may inhibit some toxins but not others.In addition, this family of toxins represents asynergistic adaptation to paralyze and kill insectprey as well as a defense. Additionally, an isoformlike toxin 3 may represent a mutated moleculethat, despite the presence of a catalytic domain, hasits biological activities influenced by neighboringamino acid residues.
On the basis of the above results, we haveidentified biological and structural differences forthree recombinant dermonecrotic toxins whichbelong to the phospholipase-D family. These toxins,despite sharing high amino acid similarities andhaving a common catalytic site, have differentbiological properties in areas such as cytotoxicity,mouse lethality and edematogenic effects, and havedifferences in their secondary structures and epitope
contents. Such differences can account for thenatural properties triggered by brown spider wholevenom.
Acknowledgments
This work was supported by grants from CNPq,CAPES, Fundac- ao Araucaria-PR, MEC-SESU-Brazil.
References
Barbaro, K.C., Sousa, M.V., Morhy, L., Eickstedt, V.R., Mota,
I., 1996. Compared chemical properties of dermonecrotic and
lethal toxins from spiders of the genus Loxosceles (Araneae).
J. Protein Chem. 15, 337–343.
Binford, G.J., Cordes, M.H.J., Wells, M.A., 2005. Sphingomye-
linase D from venoms of Loxosceles spiders: evolutionary
insights from cDNA sequences and gene structure. Toxicon
45, 547–560.
Bradford, M.M., 1976. A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein–dye binding. Anal. Biochem. 72,
248–254.
Chaim, O.M., Sade, Y.B., da Silveira, R.B., Toma, L.,
Kalapothakis, E., Chavez- Olortegui, C., 2006. Brown spider
dermonecrotic toxin directly induces nephrotoxicity. Toxicol.
Appl. Pharmacol. 211, 64–77.
Culling, C.F.A., Allison, R.T., Barr, W.T., 1985. Cellular
Pathology Technique. Butterworth & Co Ltd, London,
England.
Cunha, R.B., Barbaro, K.C., Muramatsu, D., Portaro, F.C.V.,
Fontes, W., Sousa, M.V., 2003. Purification and characteriza-
tion of Loxnecrogin, a dermonecrotic toxin from Loxosceles
gaucho brown spider venom. J. Protein Chem. 22, 135–146.
da Silva, P.H., da Silveira, R.B., Appel, M.H., Mangili, O.C.,
Gremski, W., Veiga, S.S., 2004. Brown spider and loxosce-
lism. Toxicon 44, 693–709.
da Silveira, R.B., Filho, J.F.S., Mangili, O.C., Veiga, S.S.,
Gremski, W., Nader, H.B., 2002. Identification of proteases in
the extract of venom glands from brown spider. Toxicon 40,
815–822.
da Silveira, R.B., Pigozzo, R.B., Chaim, O.M., Appel, M.H.,
Dreyfuss, J.L., Toma, L., 2006. Molecular cloning and
functional characterization of two isoforms of dermonecrotic
toxin from Loxosceles intermedia (Brown spider) venom
gland. Biochimie 88, 1241–1253.
da Silveira, R.B., Pigozzo, R.B., Chaim, O.M., Appel, M.H.,
Silva, D.T., Dreyfuss, J.L., 2007a. Two novel dermonecrotic
toxins LiRecDT4 and LiRecDT5 from Brown spider
(Loxosceles intermedia) venom: from cloning to functional
characterization. Biochimie 89, 289–300.
da Silveira, R.B., Wille, A.C.M., Chaim, O.M., Appel, M.H.,
Silva, D.T., Franco, C.R.C., 2007b. Identification, cloning,
expression and functional characterization of an astacin-like
metalloprotease toxin from Loxosceles intermedia (brown
spider) venom. Biochem. J. 406, 355–363.
de Castro, C.S., Silvestre, F.G., Araujo, S.C., Gabriel, M.Y.,
Mangili, O.C., Cruz, I., 2004. Identification and molecular

ARTICLE IN PRESSR.O.S. Ribeiro et al. / Toxicon 50 (2007) 1162–11741174
cloning of insecticidal toxins from the venom of the brown
spider Loxosceles intermedia. Toxicon. 44, 273–280.
Deleage, G., Geourjon, C., 1993. An interactive graphic program
for calculating the secondary structure content of proteins
from dichroism spectrum. Comput. Appl. Biosci. 2, 197–199.
Feitosa, L., Gremski, W., Veiga, S.S., Elias, M.C.Q.B., Graner,
E., Mangili, O.C., 1998. Detection and characterization of
metalloproteinases with gelatinolytic, fibronectinolytic and
fibrinogenolytic activities in brown spider (Loxosceles
intermedia) venom. Toxicon 36, 1039–1051.
Freshney, R.I., 2000. Culture of Animal Cells: A Manual of Basic
Techniques, fourth ed. Wiley-Liss, Inc., New York, USA.
Futrell, J., 1992. Loxoscelism. Am. J. Med. Sci. 304, 261–267.
Harlow, E., Lane, D., 1988. Antibodies: A Laboratory Manual.
Cold Spring Harbor, New York, USA.
Hogan, C.J., Barbaro, K.C., Winkel, K., 2004. Loxoscelism: old
obstacles, new directions. Ann. Emerg. Med. 44, 608–624.
Kalapothakis, E., Araujo, S.C., Castro, C.S., Mendes, T.M.,
Gomez, M.V., Mangili, O.C., 2002. Molecular cloning,
expression and immunological properties of LiD1, a protein
from the dermonecrotic family of Loxosceles intermedia
spider venom. Toxicon 40, 1691–1699.
Laemmli, U.K., 1970. Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227,
680–685.
Lee, S., Lynch, K.R., 2005. Brown recluse spider (Loxosceles
reclusa) venom phospholipase D (PLD) generates lysopho-
sphatidic acid (LPA). Biochem. J. 391, 317–323.
Luciano, M.N., Silva, P.H., Chaim, O.M., Santos, V.P., Franco,
C.R.C., Soares, M.F.S., 2004. Experimental evidence for a
direct cytotoxicity of Loxosceles intermedia (brown spider)
venom on renal tissue. J. Histochem. Cytochem. 52, 455–467.
Machado, L.F., Laugesen, S., Botelho, E.D., Ricart, C.A.,
Fontes, W., Barbaro, K.C., 2005. Proteome analysis of brown
spider venom: identification of loxnecrogin isoforms in
Loxosceles gaucho venom. Proteomics 5, 2167–2176.
Moolenaar, W.H., van Meeteren, L.A., Giepmans, B.N., 2004.
The ins and outs of lysophosphatidic acid signaling.
BioEssays 26, 870–881.
Murakami, M.T., Fernandes-Pedrosa, M.F., Tambourgi, D.V.,
Arni, R.K., 2005. Structural basis for metal ion coordination
and the catalytic mechanism of sphingomyelinases D. J. Biol.
Chem. 8, 13658–13664.
Murakami, M.T., Fernandes-Pedrosa, M.F., De Andrade, S.A.,
Gabdoulkhakov, A., Betzel, C., Tambourgi, D.V., 2006.
Structural insights into the catalytic mechanism of sphingo-
myelinases D and evolutionary relationship to glyceropho-
sphodiester phosphodiesterases. Biochem. Biophys. Res.
Commun. 342, 323–329.
Oliveira-Lima, A., Dias-da-Silva, W., 1970. Imunologia, Imuno-
patologia, Alergia—Metodos, Capıtulo 6—Provas Sorologi-
cas com Complemento. 1a. ed., Guanabara Koogan, Rio de
Janeiro, Brasil.
Paludo, K.S., Gremski, L.H., Veiga, S.S., Chaim, O.M.,
Gremski, W., Buchi, D.F., 2006. The effect of brown spider
venom on endothelial cell morphology and adhesive struc-
tures. Toxicon 47, 844–853.
Patel, K.D., Modur, V., Zimmerman, G.A., Prescott, S.M.,
McIntyre, T.M., 1994. The necrotic venom of brown recluse
induces dysregulated endothelial cell-dependent neutrophil
activation. J. Clin. Invest. 94, 631–642.
Ramos-Cerrillo, B., Olvera, A., Odell, G.V., Paniagua-Solis, J.,
Alagon, A., Stock, R.P., 2004. Genetic and enzymatic
characterization of sphingomyelinase D isoforms from the
North American fiddleback spiders Loxosceles boneti and
Loxosceles reclusa. Toxicon 44, 507–514.
Rucavado, A., Sanchez, E.F., Franseschi, A., Magalhaes, A.,
Gutierrez, J.M., 1999. Characterization of the local tissue
damage induced by LHF-II, a metalloproteinase with weak
hemorrhagic activity isolated from Lachesis muta muta snake
venom. Toxicon 37, 1297–1312.
Sunderkotter, C., Seeliger, S., Schonlau, F., Roth, J., Hallmann,
R., Luger, T.A., 2001. Different pathways leading to
cutaneous leukocytoclastic vasculitis in mice. Exp. Dermatol.
10, 391–404.
Swanson, D.L., Vetter, R.S., 2006. Loxoscelism. Clin. Dermatol.
24, 213–221.
Towbin, H., Staehelin, T., Gordon, J., 1979. Electrophoretic
transfer of proteins from polyacrylamide gels to nitrocellulose
sheets: procedures and some applications. Proc. Natl. Acad.
Sci. USA 76, 4350–4354.
Veiga, S.S., Zanetti, V.C., Franco, C.R.C., Trindade, E.S.,
Porcionatto, M.A., Mangili, O.C., 2001. In vivo and in vitro
cytotoxicity of brown spider venom for blood vessel
endothelial cells. Thromb. Res. 102, 229–237.
Zanetti, V.C., da Silveira, R.B., Dreyfuss, J.L., Haoach, J.,
Mangili, O.C., Veiga, S.S., 2002. Morphological and bio-
chemical evidence of blood vessel damage and fibrinogenoly-
sis triggered by brown spider venom. Blood Coag. Fibrinol.
13, 135–148.




















