
Biodiversity and Biogeography of Cacti
32
Transcript of Biodiversity and Biogeography of Cacti


The Cactac
plant family
sions and
knowledge
to their high
with data o
of the speci
eas of all ca
fied and ch
diversity. O
versity patt
pollination
logeny of t
extremely
threat and
This volume
analysis of o
eae with c. 1
y of the arid
molecular
of the phylog
h value as orn
n ecological p
ies are availab
actus species
haracterized s
Overall diversit
erns of all tax
syndromes a
he Cactaceae
small distrib
insufficient c
e presents the
one of the larg
1435 species
regions of th
studies res
geny and taxo
namental plan
preferences an
ble. In this vo
are mapped
seven geogra
ty patterns of
xonomic subg
are presented
e. More than
bution ranges
coverage by
e most compr
ger plant fam
are the mos
he Americas.
ulted in an
onomy of this
nts, countless
nd geographic
lume, the dist
. On this basi
aphical cente
f the family, a
roups, growth
and mapped
50% of the s
s, resulting i
existing prote
rehensive biog
ilies.
st important
Recent revi-
n improved
s group. Due
publications
c occurrence
tribution ar-
s, we identi-
rs of cactus
s well as, di-
h forms, and
on the phy-
species have
in potential
ected areas.
geographical

Text zwei
c. 205 pag
all Cactus
c. 60 colo
habitats
Analyses a
growth fo
Based on
BARTHLOTT,
KOROTKOVA,
J. (2015): B
7. 205 page
x 29.7 cm
1437-2517.
Cactaceae:Generic diversity (128 gGenera per 2500 km†
1 - 23 - 45 - 67 - 89 - 1213 - 1516 - 1819 - 22
sprachig Engl
ges, c. 333 co
species
or figures inc
and diversity
rms, and polli
the most rece
W., BURSTE
N., MIEBACH,
iogeography
es, 333 maps,
(A4), Hardco
Bezugsadress
gen.)
ish und Deuts
lored maps sh
cl. 44 fotos o
maps for all
ination syndro
ent phylogene
EDDE, K., GE
A., RAFIQPOOR
and Biodiver
, ca. 60 photo
over – Price:
se (to order b
1500 km' Barthlott et al. 2013
Map 26
sch
howing the di
of cacti specie
tribes and ge
omes
etic studies
EFFERT, J.L.,
R, M.D., STEIN,
sity of Cacti.
os and other
39,-€ + tran
stribution of
es and their
enera, major
IBISCH, P.L.,
A. & MUTKE,
Schumannia
figures. 21.0
nsport, ISSN
u

Wilhelm Barthlott, Kerstin Burstedde, Jan Laurens Geffert, Pierre L. Ibisch, Nadja Korotkova,Andrea Miebach,M.Daud Rafiqpoor,Anke Stein & Jens Mutke
Biogeography and biodiversity of cactiBiogeographie und Biodiversität der Kakteen
Ein Sonderheft derDeutschen Kakteen-Gesellschaft e.V.
und der Gesellschaft Österreichischer Kakteenfreunde
Schumannia 72015
herausgegeben vonDetlev Metzing
Schumannia

AbstractThe Cactaceae with c. 1,435 species are the most important plant family of the arid regions of the Americas. Recent revisions and molecu-lar studies resulted in an improved knowledge of their phylogeny and taxonomy. Due to their high value as ornamental plants, countlesspublications with data on ecological preferences and geographic occurrence of the species are available. In this volume, the distribution ar-eas of all cactus species are mapped. On this basis, we identified and characterized seven geographical diversity centers. Overall diversitypatterns of the family, as well as, diversity patterns of all taxonomic subgroups, growth forms, and pollination syndromes are presented andmapped on the phylogeny of the Cactaceae. More than 50% of the species have extremely small distribution ranges, resulting in potentialthreat and insufficient coverage by existing protected areas.This volume presents the most comprehensive biogeographical analysis of oneof the larger plant families, illustrated by 333 colored maps and 55 color figures on c. 200 pages.
KurzfassungDie Cactaceae sind mit rund 1.435 Arten die bedeutendste Pflanzenfamilie der amerikanischen Trockengebiete.Aktuelle Revisionen undmolekulare Analysen haben das Verständnis von Taxonomie und Phylogenie erheblich verbessert.Aufgrund des weltweiten Interesses andieser Zierpflanzengruppe liegen unzählige Publikationen zu den ökologischen Ansprüchen und Standorten vor. In diesem Band wurdendieVerbreitungsmuster allerArten erfasst und auf farbigenKarten dargestellt.Auf dieserGrundlage wurden siebenDiversitätszentren iden-tifiziert und näher charakterisiert. Diversitätsmuster der Gesamtfamilie wie auch aller taxonomischen Untergruppen sowie z. B.Wuchsfor-men und Bestäubungssyndromen werden präsentiert und unter Einbeziehung phylogenetischer Analysen diskutiert. Über 50 % der Kak-teen haben sehr kleine Verbreitungsareale, womit in vielen Fällen eine potentielle Gefährdung und unzureichendeAbdeckung durch exis-tierende Schutzgebiete einhergeht. Illustriert durch insgesamt 333 farbige Karten und 55 farbige Abbildungen auf ca. 200 Seiten wird diebisher umfangreichste biogeographischeAnalyse einer größeren Pflanzenfamilie vorgelegt.
ResumenBiogeografía y Biodiversidad de las Cactáceas. Cactaceae con aprox. 1435 especies, es la familia más importante de plantas en las zonas ári-das de América. Revisiones y estudios moleculares recientes han mejorado el conocimiento de la taxonomía y filogenia de este grupo. De-bido a su importancia como plantas ornamentales, están disponibles un sinnúmero de publicaciones sobre las preferencias ecológicas y lasdistribuciones geográficas de las especies. En este volumen están representadas gráficamente las áreas de distribución geográfica de todaslas especies de cactos. Con base en esta información hemos identificado siete centros geográficos de diversidaden el grupo. En la filogeniade Cactaceae se representan los patrones de diversidad general de la familia, al igual que los patrones de diversidad de sus subgrupos taxo-nómicos, formas de crecimiento y síndromes de polinización.Más del 50% de las especies presentan ámbitos de distribución geográfica muyrestringidos, resultando en una amenaza potencial y presencia insuficiente en las áreas protegidas en existencia. Este volumen presenta elanálisis biogeográfico más comprensivo de una de las familias más grandes de plantas, ilustrado con 55 figuras y 333 mapas a color en aprox.200 páginas.
205© Schumannia 7 2015
Abstract / Kurzfassung / Resumen
Corresponding Author / Korrespondenzautor:Prof. Dr.Wilhelm BarthlottNees Institute for Biodiversity of Plants, University of BonnVenusbergwerg 2253115 Bonn – Germanye-Mail: [email protected]

Contents/Inhalt
Foreword and acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6Vorwort und Danksagungen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9Einleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
2 Cactus ecology and biogeography . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13Ökologie und Biogeographie der Kakteen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
2.1 Cacti and their habitats . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13Kakteen und ihre Lebensräume . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
2.2 Biogeography and palaeo-history . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13Biogeographie und Florengeschichte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
3 Phylogeny, evolution and systematics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Phylogenie, Evolution und Systematik . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
3.1 Finding the closest relatives of cacti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Die nächstenVerwandten der Kakteen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
3.2 Age estimates for the Cactaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Schätzungen desAlters der Cactaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
3.3 Current understanding of phylogenetic relationships within Cactaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Das aktuelle Verständnis der phylogenetischen Beziehungen innerhalb der Cactaceae . . . . . . . . . . . . . . . . . . . . . . . . . 27
3.4 Basal cactus lineages and the origin of cacti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Frühe Kakteen-Linien und der Ursprung der Kakteen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
3.5 Relationships within Cactoideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25Beziehungen innerhalb der Cactoideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
3.6 Concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26Schlussbemerkungen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
4 Mapping the diversity of cacti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30Kartographische Darstellung der Kakteen-Diversität . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
4.1 Main data sources . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30Wichtige Datenquellen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
4.2 Generating the distribution maps . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31Erstellung derVerbreitungskarten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
4.3 Data analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31Datenanalyse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
4.4 Quality and reliability of data . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32Qualität und Zuverlässigkeit der Daten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
4 © Schumannia 7 2015

5© Schumannia 7 2015
5 Patterns of diversity and endemism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37Muster von Diversität und Endemismus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
5.1 General overview . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37Allgemeiner Überblick . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
5.2 Centres of diversity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37Diversitätszentren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
5.3 Range-sizes of Cacti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41Arealgrößen von Kakteen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56
5.4 Spatial distribution of growth forms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44RäumlicheVerbreitung derWuchsformen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.5 Spatial distribution of pollination syndromes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47RäumlicheVerteilung der Bestäubungssyndrome . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
5.6 Diversity pattern of genera, tribes and subfamilies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48Diversitätsmuster der Gattungen,Triben und Unterfamilien . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66
5.7 Diversity gradients . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50Diversitätsgradienten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67
6 Conservation and hotspots: cactus diversity in change . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68Kakteen-Diversität imWandel und Naturschutz . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73
6.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68Einleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73
6.2 Threats to Cactaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69Gefährdungen der Kakteen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
6.3 Cacti as threats . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73Kakteen als Bedrohung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
6.4 Country profiles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73Länderprofile . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
7 Distribution maps of Cactaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82Verbreitungskarten der Cactaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89
7.1 Subfamily: Pereskioideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82Unterfamilie: Pereskioideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89
7.2 Subfamily:Maihuenioideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82Unterfamilie:Maihuenioideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89
7.3 Subfamily: Opuntioideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82Unterfamilie: Opuntioideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89
7.4 Subfamily: Cactoideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83Unterfamilie: Cactoideae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 90
8 References/Literatur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 198
Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 202
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 205Kurzfassung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 205Resumen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 205

6 © Schumannia 7 2015
Cacti are, just like bromeliads or humming-birds, creatures of the NewWorld.About 1440species populate the Americas; some of themare globular and about two centimetres small(Blossfeldia), while others are columnar andup to 14 metres tall (Carnegiea). Their distri-bution ranges from the boreal coniferousforests in theNorth (south-west Canada) up tothe grasslands of the Pampa in the south.Cactiinhabit diverse habitats, from the coast to thehighmountains, from deserts up to the rainfor-est canopy.The often densely spined succulents (called“herbal crystals” by the poet Adalbert Stifter)are characterized by magnificent flowers. Notonly scientists but also plant enthusiasts are fas-cinated by this plant family.They are covered byimportant collections, national and interna-tional cactus and succulent societies, about adozen journals and an enormous amount of lit-erature. Hence, cacti are one of the most well-studied major plant families.The first modern monographs in the new mil-lennium have been presented byEdward F.An-derson as“The cactus fanily” (ANDERSON 2001,German editions as “Das große Kakteen-Lexikon”, ANDERSON 2005, 2011) and DavidHUNT (2006).The amount of published data isoverwhelming, but a modern biogeographicoverview for the family is still missing. Thereare early biogeographic studies based on thedata available at that time. Karl SCHUMANN(1899a) published “Die Verbreitung der Cac-taceae im Verhältnis zu ihrer systematischenGliederung” [Distribution of Cactaceae in re-lation to their systematic classification] inBerlin, and Curt Backeberg published his „ZurGeschichte der Kakteen“ [History of the cacti](BACKEBERG 1942b) and „Verbreitung undVorkommen der Cactaceae” [Distribution andoccurrence of Cactaceae] (BACKEBERG 1944).Today, however, these pioneering treatmentsare outdated,especially with regard to the pres-ent state of the knowledge of phylogeny, sys-tematics, and molecular analyses.This analysis of biogeography and biodiversityof the cacti combines current knowledge with anew objective in the context of modern biodi-versity research. The family is analysed basedon systematic-taxonomic knowledge incorpo-rating geographic and phylogenetic data. Ini-tially, distributionmapswere created for 1416 ofthe 1438 accepted species. From these maps, di-versity maps were generated for the relevanttaxonomic levels from genera to the entire fam-
ily, and the different centres of diversity werecircumscribed.At the same time,macroecolog-ical relations, distribution of growth forms, pol-lination syndromes, and other aspects wererecorded, spatially visualized, and correlatedwith general biogeographical patterns and eco-regions. Issues of conservation were examinedunder consideration of land use data and exist-ing protected areas.This provides an importantfoundation for the conservation of Cactaceaeand their habitats.The plan for this publication emerged in theearly 1990’s. First modern maps of phytodiver-sity had already been published byBARTHLOTT(1983) for the Cactaceae–Rhipsalideae. As awell-known and extensively studied group,compared to other large families, theCactaceaewill act as a role-model for the analysis of bio-diversity and biogeography of a larger an-giosperm group.The monograph presented here is thus basedon three decades of preparatory work,as well asthe collaboration of many colleagues and stu-dents, who are either involved as authors or ac-knowledged for their contributions. SimoneStrecker (1992) submitted a diploma thesis(“Arealgeographie der Kakteen”), which wasbased on a manual compilation of diversitymaps of all Cactaceae genera. In subsequentyears, the data were complemented and editedby Pierre Ibisch, utilizing vegetation maps andhis fieldwork in Bolivia.He also supervised pre-liminary studies and the digital processing ofdistribution data by Georg Rauer, which wassupported by a research grant from theDeutsche Kakteen-Gesellschaft (the “GermanCactus Society“)—at that time still without truegeographic information systems (GIS).As partof a large collaborative project (BIOTA)funded by theGerman FederalMinistry for Ed-ucation and Research, a working group (“BIO-MAPS“) has been established in Bonn from2001 to 2010 to map the global geographic di-versity of angiosperms.Besides regional analy-ses, world maps of biodiversity of all land plantgroups were generated (e.g. BARTHLOTT & al.1996, 2005, KIER & al. 2009, KREFT & al. 2010,MUTKE & al. 2011). Therefore, the infrastruc-ture and the aforementioned geographic infor-mation systems were available for digitizationand spatial analysis of the cactus data. Thisstarted in 2007 and was carried out by two par-allel diplom theses of Anke Stein and KirstenBurstedde (nee Hahne) that were finished inspring 2009. In 2011 Andrea Miebach com-
pleted a bachelor thesis focusing on the evolu-tion of functional traits in the Cactaceae. JensMutke coordinated this work since 2007 withgreat dedication and expertise.In preparation for publication, all of the mapswere carefully revised and uniformly laid out(among others by Laurens Geffert, AndreaMiebach and Lina Samira Meiling (geb.Mathar).BorisO.Schlumpberger provided dataon pollination ecology from his on-going re-search project. Nadja Korotkova analysed thephylogenies based on molecular data, and ge-ographer M. Daud Rafiqpoor edited the spa-tial aspects of the distribution maps.Almost allof the above collaborators were, at least tem-porarily, funded by our long-term project „Bio-diversität im Wandel“ of the Academy of Sci-ences and Literature in Mainz.With the financial support of theAcademy of Sci-ences and Literature in Mainz, we established asmall working group of the International Organ-ization for Succulent Plant Study (IOS) at theend of our long-term mapping work and met forthe first time in 2009 during the IOS Inter-Con-gress 2009 in Bonn. The colleagues carefullychecked and revised the distribution maps pro-vided by us, and our special thanks go to DavidHunt, Nigel Taylor, Martin Lowry, Charles Gra-ham, Paul Hoxey, and Ralf Bauer.The list of people who contributed to the pub-lication presented here is long.We are indebtedto all of them. However, this particularly ap-plies to the Academy of Sciences and Litera-ture in Mainz: Without a sustainable institu-tional framework of our long-term project„Biodiversität im Wandel„ (1999–2014), thecomplex project ofmapping the cacti could nothave proceeded.The most important historical work on bio-geography of cacti was published in 1899 inBerlin,written by Professor Karl Moritz Schu-mann, who founded the German Cactus Soci-ety (DKG) in 1892.This society has accompa-nied and supported our project from the be-ginning, and we are very much obliged to it.We are especially grateful for the editorDetlevMetzing, who worked with great competenceand intensity at the completion of this manu-script. It is a fortunate coincidence that thiswork is published in the “Schumannia”, namedafter the founder of the DKG.
Bonn, in January 2015
Wilhelm Barthlott
Foreword and acknowledgements
Wilhelm Barthlott

Presumably, cacti (Melocactus) were alreadyamong the curiosities that Christopher Colum-bus presented as gifts to Queen Isabella ofCastile after his first voyage to the New Worldin 1492. Carl Linnaeus (1707–1778) already hada surprisingly clear picture of the delineation ofthis “family”. In his “Species Plantarum”, pub-lished 1753, he listed 22 cactus species. Basedon his Sexual System of classification, he as-sumed a close relationship between morpho-
logically very different taxa, such as Melocac-tus andOpuntia, but alsoPereskia, and he com-bined all these in the genus Cactus. With thestudies ofAdrian Haworth (1768–1833), JosephSalm-Dyck Reifferscheidt (1773–1861), Au-gustin Pyrame de Candolle (1778–1841), andLudwig G. K. Pfeiffer (1805–1877), the system-atic investigation in the middle of the 19th cen-tury was already far advanced. This was be-cause these plants had stimulated the interest
of botanists and plant enthusiasts at an earlystage, caused by their “bizarre” and “exotic“appearance, combined with modest cultivationrequirements of many species.In this era, systematic recording of species wasat the heart of research. This did not only ap-ply to cacti, but also to all other plant families.Plant geography (and ecology) started earlywith the exceptional singular“Essai sur la Géo-graphie des Plantes” (Paris 1805) by the vi-
9© Schumannia 7 2015
1 Introduction
Wilhelm Barthlott
Fig. 1: The overlapping distribution ranges of ten of the Mexican globular cactus genera accepted by Curt BACKEBERG (1944), which he noticeably summari-zed under the name Boreoechinocacti. The lines are distribution boundaries of the genera. The Chihuahua centre of diversity (cf. Map11) is already clearlyvisible, as already in the „Karte der Verdichtung der Kakteenvorkommen“ [map of densification of cactus occurrences ](BACKEBERG 1958: Abb. 4) (from:BACKEBERG 1944).Abb. 1: Die überlagerten Areale von zehn der von Curt Backeberg akzeptierten Gattungen mexikanischer Kugelkakteen (BACKEBERG 1944), die er bezeich-nenderweise unter dem Begriff Boreoechinocacti zusammenfasst. Die Linien sind die Arealgrenzen der Gattungen. Schon deutlich ist auf dieser Karte wieauch auf der „Karte der Verdichtung der Kakteenvorkommen“ (BACKEBERG 1958: Abb. 4) das Chihuahua-Diversitätszentrum der Familie (vgl. Karte 11)zu erkennen (aus: BACKEBERG 1944).

2.1 Cacti and their habitats
Cacti are one of the most characteristic plantgroups of the Americas, being almost endemicto the New World. only one single epiphyticspecies,Rhipsalis baccifera, has disjunct popu-lations in the old World.Latin America stands out for a high diversityof different landscapes and vegetation zones(Fig. 4). It may seem surprising to the non-spe-cialist that habitats of cacti represent a largerange of environmental conditions, as well.They grow from coastal sand dunes up to some5000 metres elevation in the high Andes, andfrom southern Canada to southern Argentina,spanning a latitudinal range of more than 100degree. Factors limiting the distribution rangeof the family at high latitudes are extreme coldand in most cases humid conditions with shortdays during winter. In addition, more humidparts of the mediterranean type winter rainfallareas in northern California and south-centralChile are avoided by most cacti. This seemssomewhat surprising, as especially somespecies of Opuntia have been widely natural-ized in the mediterranean Basin, includinglarge parts of southern Europe and northernAfrica. one important parameter limiting cac-tus growth in winter rainfall areas might be theinfluence of frequent fires. Fire also limits theestablishment of cacti in many grass lands andsavannah ecosystems.Even though cacti are an iconic element ofNew World deserts, the family inhabits a widerange of ecosystems (Figs. 5–21); cacti evengrow in humid tropical rainforests. However,this is more of an exception and mainly true forepiphytic species, some shrubby or arborescentPereskia in south-east Brazil and central Amer-ica, and the specialists growing on isolated rockoutcrops (inselbergs) representing dry, azonalhabitat islands within rainforests. In general,cacti are bad competitors under humid, fertileconditions. Thus, most species are found inecosystems with limited water availability atleast during some months of the year, particu-lar edaphic conditions such as the rock out-crops, saline, gypsum, serpentine substrates, orsand fields. This includes open desert to semi-desert vegetation types, as well as thorny shruband deciduous to semi-deciduous dry forests. It
has been demonstrated by several authors thatmany cacti species show higher rates of estab-lishment and survival in the shadow of trees orshrubs as “nurse plants” in semi-arid habitats(e.g., LARREA-ALCáZAR & al. 2008, mANDuJANo
& al. 2002).on an ecoregional basis, the mexican pine-oak forests of the Sierra madre oriental andoccidental as well as of the Trans-mexicanVolcanic belt belong to the areas with thehighest Cactus species richness, as well as themeseta Central matorral and the Chihuahuadesert in mexico, the Central Andean PunaEcoregion sensu WWF (oLSoN & al. 2001) inPeru and Bolivia, and the Bolivian montanedry forests.
2.2 Biogeography and palaeo-history
Apart from the early studies by SCHumANN
(1899a) and BACKEBERG (1942) there are fewbiogeographic studies for the cactus family(but compare e.g. HERNáNDEZ & GómEZ-HI-NoSTRoSA 2011, TAyLoR & ZAPPI 2004, THIEDE
1998). unfortunately, the analysis of biogeo-graphic patterns of cacti is hampered by theabsence of a fossil record. Not even the age ofthe family is known. Recent molecular clockapproaches using fossils of related taxa indi-cate a relatively short evolutionary historycompared to other flowering plant families(compare chapter 3.2).most cactus diversity today is linked to arid orsemi-arid conditions.The species rich arid re-gions of western North America (cf. Fig. 22)and mexico are dominated by the almost ex-clusively North American cactus tribes Cac-teae and Phyllocacteae, as well as many spe-cies of opuntioideae. These northern centresof cactus diversity are separated from the dryCaribbean Coast of Colombia and Venezuelaby the rainforests of southern Central Amer-ica with some linkage through the CaribbeanIslands.The humid rainforests of the Amazon Basinand of the Chocó region in western Colombiaseparate these northern hemisphere cactushabitats from suitable areas further south (cf.Fig. 4). Dry valleys of the Andes in westernSouth America are to some degree connected
to the more southern regions of the AtacamaDesert at the Peruvian and Chilean PacificCoast. However, the geologically relativeyoung and somewhat wetter Northern Andesespecially in Colombia harbour only very fewcactus species.The Atacama Desert with manyendemic species especially of the tribe Cereeaeis separated from the species rich Bolivian An-dean dry valleys and the western Chaco by thehigh Andes reaching altitudes of more than5000 m a.s.l. This Bolivian and northern Ar-gentinian South Central Andes centre of cac-tus diversity has the highest diversity of differ-ent tribes and genera, as well as growth formsof cacti. It is separated from the cactus diver-sity centre in the Caatinga in eastern Brazil bythe grasslands of the Cerrado, which are rela-tively poor in cacti, probably due to the fre-quent occurrence of fires. The high species di-versity in the south-east Brazilian rainforest ofthe mata Atlantica is a particular case owedmainly to the presence of many epiphytic spe-cies of the tribe Rhipsalideae, as well as speciesin azonal habitats like rock outcrops or coastalsand dunes.A detailed description of the maincentres of Cactus diversity is given in chapter5.2. For an overview on the biogeography andecology of cacti of Eastern Brazil see TAyLoR &ZAPPI (2004).The lack of data on the evolutionary historyof the Cactaceae makes it difficult to relatethe current distribution patterns to palaeo-geographic conditions and events. Both BACKE-BERG (1942b) as well as LEuENBERGER (1986)conclude an origin of the Cactaceae and ofthe most basal taxon in the phylogenetic tree,the subfamily Pereskoideae with the onlygenus Pereskia, in northern South America.According to recent molecular studies thisgenus is a grade with two clades at the basisof the cactus phylogeny (compare NyFFELER &EGGLI 2010b and Fig. 23). Today, the cladisti-cally basalmost clade of Pereskia species oc-cur in northern Venezuela, the Caribbean Is-lands, and Central America—only P. aureifo-lia occurs in south-east Brazil. In contrast, thesecond clade of Pereskia species is almost re-stricted to south-east Brazil and the centralAndes of Bolivia and Peru—with P. aculeatahaving a disjunct distribution area in the Car-ribean, as well.
13© Schumannia 7 2015
2 Cactus ecology and biogeography
Jens Mutke

3.1 Finding the closest relatives of cacti
The Cactaceae belong to the order Caryo-phyllales in which they are part of a group thatcontains most of the succulent families of theorder:Cactaceae,Anacampserotaceae,Basella-ceae, Didiereaceae, Halophytaceae, montia-ceae, Portulacaceae, and Talinaceae (CuENouD
& al. 2002, SCHÄFERHoFF & al. 2009).The place-ment of Cactaceae within the Caryophyllales(or in the former Centrospermae) dates backto the 19th century, and molecular phylogeneticstudies have undoubtedly confirmed thatCactaceae are part of the Caryophyllales. Nev-ertheless, the closest relatives of cacti havebeen identified with confidence only recently.A first hypothesis on the putative close relativesof the cacti was suggested by THoRNE (1976),who assumed a close relationship of Cactaceaeand Didiereaceae, Basellaceae and Portula-caceae and created the suborder Portulacinaefor this alliance. Recent studies have confirmedthat Cactaceae are closely associated with thePortulacaceae, and even nested within them(APPLEQuIST & WALLACE 2001, CuENouD & al.2002, HERSHKoVITZ & ZImmER 1997). NyFFELER
(2007) could finally identify the Portulacaceaetribe Anacampseroteae as the closest relativesof cacti. The Portulacaceae were found to bepolyphyletic and several nomenclatural changesbecame necessary. As one result, the Ana-campserotae are now treated as the familyAnacampserotaceae and are the sister family ofthe Cactaceae (NyFFELER & EGGLI 2010a).
3.2 Age estimates for the Cactaceae
Since Cactaceae are a New World family, ab-sent from Africa despite of suitable habitats(besides one Rhipsalis species and introducedopuntias), it is generally assumed that theyevolved in SouthAmerica after the break-up ofGondwana during the late Jurassic and earlyCretaceous. This suggests a limiting age of100–90 million years when South America andAfrica were already separated (mAuSETH 1990).An immediate age estimate for the Cactaceaeis not possible since no fossils are known.The most comprehensive hypothesis on theevolutionary history of the Cactaceae waslinked to the geological history of South Amer-ica and was proposed by BACKEBERG (1942b).
He assumed an origin of the family in the trop-ical zone of mesoamerica during the Creta-ceous, app. 130 million years, thus suggestingthat the Cactaceae are a rather ancient group.much later, HERSHKoVITZ & ZImmER (1997) sug-gested the Cactaceae being a young group ofonly 30 million years of age. Still, they did notprovide any plausible basis for this estimate.The most recent evidence for the age of theCactaceae comes from molecular dating of theCaryophyllales, yielding timeframes of 116–104million years (ANDERSoN & al. 2005, WIKSTRom
& al. 2001). No age estimates using a molecu-lar clock particularly for the Cactaceae wereavailable until recently, due to the lack of fos-sils, which are used as calibration points in suchanalyses. Age estimates for Cactaceae there-fore still have to rely on known fossils of moredistantly related taxa.Thus, the molecular clockcalibrations only allow just setting an approxi-mate timeframe when the family evolved.Twostudies using this approach yielded differentresults: the age of the cacti was inferred as19.1–3.1 million years (oCAmPo & CoLumBuS
2010) or 35 million years (ARAKAKI & al. 2011).
3.3 Current understanding ofphylogenetic relationshipswithin Cactaceae
The Cactaceae as a family were never seriouslyquestioned, but hypotheses on relationshipswithin the family were always troublesome.Convergent evolution is frequent in this fam-ily (WALLACE & GIBSoN 2002), making inter-pretation of morphological features challeng-ing. Columnar cacti, small globular cacti, andepiphytes, as well as adaptations to majorpollinator groups (bats, birds) have evolvedseveral times independently and the flowermorphology is rather uniform, providing fewvaluable characters. Furthermore, due to phe-notypic plasticity as a response of the environ-mental conditions, individual taxa are mor-phologically variable, making interpretation ofmorphological characters troublesome.molecular phylogenetic methods contributedmuch to the understanding of flowering plantevolution and phylogenetics. But the relation-ships within the Cactaceae are still insuffi-ciently understood, and few molecular phylo-genetic studies have been conducted compared
to other popular plant families such as orchi-daceae or Bromeliaceae.The first DNA sequence data for Cactaceaewere published by WALLACE (1995). A morecomprehensive phylogenetic tree derived fromthe chloroplast marker rbcL was provided byWALLACE & GIBSoN (2002). The first compre-hensive molecular phylogenetic study for theCactaceae was done by NyFFELER (2002) andwas based on datasets of the chloroplast re-gions trnK/matK and trnL-F. This study couldnot entirely clarify relationships within thefamily but identified major groups and fur-thermore revealed the para- or polyphyly of sometribes and genera, e.g. Pereskia, Notocacteae,Browningieae, Hylocereeae and Lepismium.The most comprehensive phylogenetic hy-potheses for the Cactaceae at present are basedon datasets of the plastid regions trnK/matK,the rpl16 intron and trnL-F and the nucleargene ppc (HERNáNDEZ-HERNáNDEZ & al. 2011)or on trnK/matK (BáRCENAS & al. 2011). Todate, the study of BáRCENAS & al. (2011) is theCactaceae study with the most species included(666 species).The results of these studies are animprovement compared to the previous ones,but the results are still not satisfactory and therelationships within Cactaceae remain partlyunresolved.
3.4 Basal cactus lineages and theorigin of cacti
In the following, the relationships are sum-marised as currently understood based on theaforementioned studies and further studies ofsingle tribes and genera. The first branching isthe subfamily Pereskioideae (only Pereskia,Fig. 26). As revealed by molecular data, Pere-skia is not a monophyletic group but forms agrade in the Cactaceae phylogeny (Figs. 24 &25) with mesoamerican and Caribbean Pere-skia species as the first branching group fol-lowed by the Andean Pereskia, which is sisterto the rest of the family (BuTTERWoRTH & WAL-LACE 2005, EDWARDS & al. 2005).Pereskia has traditionally been interpreted asthe most ancestral cactus since the works ofSCHumANN (1899b) and many of the Pereskiacharacters, which have been regarded as ple-siomorhic within Cactaceae. These are thewoody stem, the presence of leaves (in contrast
24 © Schumannia 7 2015
3 Phylogeny, evolution, and systematics
Nadja Korotkova

26 © Schumannia 7 2015
Fig. 25: Phylogenetic tree of the Cactaceae showing relationships bet-ween all Cactaceae genera based on the phylogenetic studies availableso far. The topology is based on HERNANDEZ-HERNANDEZ & al. (2011), NYFFELER(2002), GRIFFITH & PORTER (2009) (Opuntioideae), BUTTERWORTH & al. (2002)(Cacteae), KOROTKOVA & al. (2010, 2011) (Lymanbensonieae and Rhipsali-deae), RITZ & al. (2012) (Tephrocacteae) and SCHLUMPBERGER & RENNER (2012)for Echinopsis and allies. The suprageneric classification and the generafollow NYFFELER & EGGLI (2012), with modifications based on KOROTKOVA & al.(2010, 2011), RITZ & al. (2012) and SCHLUMPBERGER & RENNER (2012). Statisticalsupport for the main clades on tribal level is indicated with *** for maxi-mum support and ** for low support. Support for the higher nodes isnot shown for better readability of the tree. Genera with (?) have so farnot been sampled in a phylogenetic analysis.Abb. 25: Stammbaum der Cactaceae, gezeigt sind verwandtschaftlicheBeziehungen zwischen allen Cactaceae-Gattungen, basierend auf allenbisher verfügbaren phylogenetischen Studien. Die Topologie basiert aufHERNANDEZ-HERNANDEZ & al. (2011), NYFFELER (2002), GRIFFITH & PORTER (2009)(Opuntioideae), BUTTERWORTH & al. (2002) (Cacteae), KOROTKOVA & al. (2010,2011) (Lymanbensonieae und Rhipsalideae), RITZ & al. (2012) (Tephroca-cteae) und SCHLUMPBERGER & RENNER (2012) für Echinopsis und Verwandte.Die supragenerische Klassifizierung und die Gattungen folgen NYFFELER &EGGLI (2012), mit Änderungen basierend auf KOROTKOVA & al. (2010), RITZ &al. (2012) und SCHLUMPBERGER & RENNER (2012). Die statistische Unterstüt-zung für die wichtigsten Gruppen auf Tribus-Ebene wird mit *** fürmaximale Unterstützung und ** für geringe Unterstützung angegeben.Unterstützung für die höheren Knoten wird zur besseren Lesbarkeit desBaumes nicht gezeigt. Gattungen mit (?) wurden bisher in keiner phylo-genetischen Analyse untersucht.

4.1 Main data sources
Given the fact that cacti have always fascinateda large number of scientists, gardeners, andplant enthusiasts, the taxonomic concepts usedvary a lot. Already Karl SCHumANN in (1899a)starts his work on the “Verbreitung der Cac-taceae” with a discussion on the problem of ahigh degree of synonomy, nomina nuda, etc.Around 14,000 species names of cacti had al-ready been published in the 1970ies (BARTH-LoTT 1977). These included a vast amount of
synonyms. Recent compilations of acceptedspecies for the family list between 1,431 speciesin 124 genera (HuNT 2006) and 1,896 species in127 genera (ANDERSoN 2001, 2005). For ourdatabase, we used a slightly updated version ofthe “New Cactus Lexicon” species list (Hunt,pers. comm.).This list includes a few additionalnames compared to the “New Cactus Lexicon”.Distribution maps are presented in this volumefor only 1,416 species, since some species onthe original list are now regarded as synonymsin our current database and since the natural
range of some widely naturalized taxa is insuf-ficiently known (e.g.,Opuntia ficus-indica). In-fraspecific taxa are not considered and are allincluded in the respective species distributionmaps.Infrafamilial classification follows the tribesand subfamilies of NyFFELER & EGGLI (2010b),with minor modifications for the epiphytic Ly-manbensonieae based on the results of Ko-RoTKoVA & al. 2010 (cf. Figs. 24–25).For our analyses, we used additional data re-garding pollination syndromes (Schlumpber-
30 © Schumannia 7 2015
4 Mapping the diversity of cacti
Jens Mutke, Kirsten Burstedde, Jan Laurens Geffert, Andrea Miebach, M. Daud Rafiqpoor,Anke Stein & Wilhelm Barthlott
Fig. 32: Comparison of the diversity maps of Rhipsalinae from BARTHLOTT (1983) and Rhipsalideae from this work. The basic outline of the diversitypattern was already published in 1983 based on a very limited database.Abb. 32: Vergleich der Diversitätskarten der Rhipsalinae aus BARTHLOTT (1983) und Rhipsalideae aus dieser Arbeit. Das Grundmuster der Diversitäts-verteilung wurde schon 1983 basierend auf relativ wenigen Daten erkannt.

lie im Zentrum des Interesses einer ganzen An-zahl von Fachbotanikern, aber noch mehr vonPflanzenliebhabern (siehe Kap.1) steht:Es liegteine beinahe unüberschaubare Fülle von pu-blizierten Daten zu Fundorten in Fachpublika-tionen vor, vorbildhaft sei “mapping the Cactiof mexico“ (HERNáNDEZ & GómEZ-HINoSTRoSA
2011) genannt. Zudem liegen für beinahe allegrößeren Gattungen moderne Revisionen undtaxonomische Bearbeitungen bis hin zu mo-dernen Gesamtbearbeitungen der Familie(HuNT 2006) vor.Darüber hinaus werden in denzahlreichen nationalen Fachzeitschriften (z. B.in der Bundesrepublik Deutschland „Kakteenund andere Sukkulenten“ und den uSA „Ca-ctus and Succulent Journal“) beinahe monat-lich ergänzende Fundortdaten publiziert.Es sollaber angemerkt werden, dass die Datenlage im
Detail doch heterogen ist. Gattungen, die kei-nen hohen Sammlerwert besitzen (z. B. Opun-tia), sind schlechter erfasst als ausgesprocheneLiebhaber-Gruppen (z. B. Mammillaria). Viel-leicht eher politisch bedingt ist die Datenlagefür unterschiedliche geographische Regionenebenfalls heterogen: Südostbrasilien und me-xiko sind hervorragend bearbeitet, für Kuba istdie Datenlage eher spärlich.In noch höherem maße gilt dies für die Erstel-lung der Diversitätskarten, bei denen sich klei-nere Fehler innerhalb der Arten-Arealkartender erfassten Arten aufheben. Zum Vergleichhaben wir in Abb. 32 unsere, auf einer relativengen Datenbasis beruhende Diversitätskarteder Rhipsalideen von 1983 der jetzt erarbeite-ten Karte gegenübergestellt. Wenn man diedurch die unterschiedliche Projektion beding-
ten verschiedenen umrisslinien der Kontinen-te herausrechnet, sind die Karten beinahedeckungsgleich. Insofern ist voraussehbar, dasses z. B. an der Biodiversitätskarte der gesamtenFamilie (Karte 7) auch bei sich zunehmend ver-besserter Datenlage in den nächsten Jahrzehn-ten kaum eine Änderung mehr ergeben wird.Zusammenfassend lässt sich feststellen, dassdie Familie der Cactaceae durch die vorlie-genden publizierten Daten ein herausragendesBeispiel für die Analyse der Biogeographieund Biodiversität einer größeren Pflanzenfa-milie überhaupt darstellen. Es gibt wohl keinezweite Familie entsprechender Größe inner-halb des ganzen Pflanzenreiches, die eine ver-gleichbare Analyse erlauben würde.
36 © Schumannia 7 2015
1 - 23 - 56 - 1213 - 2223 - 73
© Barthlott et al. 2013
Diversity centres of Cactaceae
Species per 2500 km²
Map 8
1500 km
(defined as the top 5%most species rich areas)
9444402717
MammillariaEchinocereusCoryphanthaOpuntiaCylindropuntia
Species rich genera:
367 species, 43 genera
Chihuahua centre
2810666
MammillariaOpuntiaFerocactusPachycereusSelenicereus
Species rich genera:
107 species, 27 genera
Puebl -Oaxaca centre
Jalisco centre
654
MammillariaStenocereusOpuntia
Species rich genera:
35 species, 17 genera654333
MammillariaEchinocereusFerocactusCylindropuntiaOpuntiaStenocereus
Species rich genera:
33 species, 13 genera
Sonora-Sinaloan centre
1687555
MelocactusPilosocereusRhipsalisArrojadoaMicranthocereusPereskia
Species rich genera:
74 species, 23 genera
Caatinga centre
24533
RhipsalisSchlumbergeraLepismiumPilosocereus
Species rich genera:
47 species, 13 genera
Mata Atlantica centre
5928231914
EchinopsisRebutiaParodiaGymnocalyciumCleistocactus
Species rich genera:
214 species, 37 genera
Southern Central Andes centre
Map 8: Diversity centres of Cactaceae.Karte 8: Diversitätszentren der Cactaceae (definiert als die oberen 5 % artenreichsten Gebiete; Arten pro 2500 km²; species rich genera = artenreiche Gattungen).

5.1 General overview
Except for the high latitude boreal forests andTundra vegetation of North America and thetemperate rain forests of southern Chile, atleast some cactus species can be found any-where in the Americas (map 7).Rhispsalis bac-cifera is the only species naturally found also inthe old World, ranging from tropical WestAfrica via madagascar,mauritius,and Réunionto Sri Lanka.Highest species richness of cacti (≥ 23 speciesper 2,500 km², the 5% most species rich areas;maps 8 & 9) can be found in seven distinct cen-tres in mexico, Brazil, Argentina, and Bolivia.At the genus level, additional centres are locatedin Peru and in northern Venezuela (map 10)—these are covered more in detail in chapter 5.6.The four North American centres of highspecies richness in mexico and the southernuSA are located in dry ecoregions (Table 1,map 11): 1) the Chihuahua and the mesetaCentral matorral, 2) the Puebla-oaxaca centrewith a diversity of different climates and habi-tats, as well as two minor areas 3) near the Pa-cific coast of Jalisco (Jalisco centre), and 4) atthe border of the southern Sonora desert to theSinaloan dry forests (Sonora-Sinaloan centre).In South America, there are three main cen-tres of cactus species richness in 1) the south-ern central Andes in Bolivia and northern Ar-gentina, 2) the Caatinga, and 3) the coastal for-est area of south-east Brazil. The latter centreis mainly dominated by epiphytic species of theRhipsalideae (compare Table 2, map 12).Intermediate species richness can be found allover south-east Brazil,which is linked via a cor-ridor of intermediate species richness inuruguay and Paraguay to the Bolivian centreof diversity. Coastal Peru and parts of the cen-tral Andean dry valleys show intermediatespecies richness, but an especially high con-centration of species with extremely restricteddistribution ranges. Northern Venezuela andmost parts of the Caribbean also show inter-mediate levels of species richness.Low species richness (≤ 5 species per 2,500km²) occur in large parts of the Amazon Basin,many fire-prone habitats, e.g., of the Brazilian
Cerrado, the Llanos Savannah of Colombiaand Venezuela, mediterranean climate areas inCalifornia and Chile,and in the temperate east-ern uSA.
5.2 Centres of diversity
For the delineation of the centres of cactusspecies richness (map 8,Tables 1 & 2), we usedthe top 5% areas with highest species numbers(≥ 23 species per 2,500 km²). To qualify as oneof the centres listed below, at least four adja-cent grid cells of 50 x 50 km² have to fulfil thiscriterion.We distinguish seven diversity centres.
The four north/central American centresChihuahua centre: The mexican centre of di-versity with the largest area and by far the high-est species numbers is the large complex ofdeserts and xeric shrublands of the Chihuahuadesert, including the mexican meseta Centralmatorral (compareTable 1). It includes parts ofthe north-eastern Sonora desert as well. High-est species richness can be found in the south-eastern part of the centre with a diverse mosaicof matorral and, e.g., the Sierra madre orien-tal Pine-oak Forests. map 11 shows the finescale patterns of species richness within themexican centres in smaller grid cells of 10 x 10km.The centre is characterized especially by ahigh diversity of globular species of the tribeCacteae and is the main centre of species rich-ness of the opuntioideae. most other tribes areabsent or poorly represented. The growthforms of shrubby and arborescent cacti alsohave their centres of species richness in theChihuahua, whereas columnar cacti are almostabsent.Puebla-Oaxaca centre: This mexican centremainly includes the Tehuacán-Cuicatlán valley.It is geographically isolated from the nearbyand much larger Chihuahua centre by the Trans-mexican volcanic mountain belt. Its topogra-phic diversity (geodiversity in a wider sense)results in a variety of vegetation types such astropical dry forests, xeric shrublands, thornshrubs, and deserts.The Puebla-oaxaca centrehas the highest concentration of columnar
species of the tribe Cacteae in mexico. In con-trast to the Chihuahua centre, this centre ishome to a high number of cacti pollinated bybats, most of them columnar. The mean rangesize of the species occurring in this region is muchsmaller compared to the Chihuahua centre. manyof the species are restricted to this region.Sonora-Sinaloan centre:This centre comprisesa small area of high species richness at thesouthern transition of the Sonora desert to theSinaloan dry forest dominated by deciduousthorn forest. The species rich north-easternpart of the Sonora desert, in contrast, is in-cluded in our larger Chihuahua centre (seeabove). unlike the situation in those northernparts of the Sonoran desert, frost is rare in thesouthern Sonora-Sinaloan centre. The regionis characterised by a mixture of temperate andtropical species.The growth forms of cacti pres-ent in the southern Sonora include globosespecies (e.g. Mammillaria, Ferocactus) and co-lumnar species (e.g.Stenocereus) of the Cacte-ae, columnar species of the Phyllocacteae sub-tribus Echinocereinae and shrubby Opuntia.Jalisco centre: This small centre of high cactusdiversity at the pacific coast of the mexican stateJalisco has a more humid climate compared tothe other northern diversity centres resulting intropical dry forest vegetation. As a result, thiscentre is home not only to globularMammilariaspecies and abundant columnar and arborescentcacti like Stenocereus, Cephalocereus (all Cac-teae), andOpuntia, but also epiphytic species ofEpiphyllum and Selenicereus.
The three South American centresSouthern central Andes centre: The largestSouth American centre of diversity also har-bours the highest species number in SouthAmerica and is characterized by the highestphylogenetic and morphological diversity. 10out of the 11 tribes of cacti accepted here oc-cur in the southern central Andes centre.Witheight tribes occurring per 2,500 km² grid cell inlarge parts of this centre, this is twice the num-ber found in most parts of the Chihuahua cen-tre. The centre is located in the Semiarid andarid valleys of central and southern Bolivia andnorthern Argentina. In the northern part of the
37© Schumannia 7 2015
5 Patterns of diversity and endemism
Jens Mutke, Kirsten Burstedde, Jan Laurens Geffert, Andrea Miebach,M. Daud Rafiqpoor, Anke Stein & Wilhelm Barthlott

39© Schumannia 7 2015
Map 10: Generic diversity centres of Cactaceae (highlighting the 5% most genus rich areas, compare map 26)Karte 10: Diversitätszentren der Cactaceae, Gattungen (definiert als die oberen 5 % gattungsreichsten Gebiete; Arten pro 2500 km²).
1500 km© Barthlott et al. 2013
Diversity centres of Cactaceae: genera
Genera per 2500 km²
1 - 23 - 56 - 89 - 1213 - 22
Map 10
(defined as the top 5% most rich areas)

centre, original vegetation comprises decidu-ous forests with scattered naturally forest freesteep slopes with rock outcrops. In the south,in the prepuna ecoregion, Andean shrublandsand semideserts are found. This centre har-bours the highest number of globular cacti inSouth America, but is also a centre of colum-nar cacti. It is the only South American regionwith significant numbers of cushion formingcacti. In addition to mainly bee-pollinatedspecies, it has a considerable number of sphin-gophilous species. Especially the western An-dean part of the Bolivian province Tarija andneighbouring areas of Salta and Jujuy in north-ern Argentina are extremely diverse whenlooking at fine scale patterns of species richnessin 10 x 10 km grid cells (map 12).
Caatinga centre: This Brazilian centre is lo-cated in the southern part of the Caatingaecoregion and the Campos Rupestres linked tomountain ranges of the Serra Espinhaco andChapada Diamantina.While the dominant veg-etation types of the Caatinga region are xericshrubland and thorn forest, highest species di-versity of cacti can be found in azonal habitatislands such as rock outcrops (“inselbergs”) ornutrient poor sandy soils.Although species di-versity is much lower compared to the Chi-huahua or the southern central Andes centre,the Caatinga centre is home to a high numberof genera—especially of the tribe Cereeae.Ac-cordingly, this centre is characterized by onlymoderate species numbers of globose cacti, buta high diversity of columnar species. The Caa-
tinga centre has the highest numbers of cactipollinated by bats and birds and only low num-bers of bee-pollinated species.Mata Atlântica centre: This centre in south-east Brazil is ecologically quite different fromall the others, as it is characterized by humid,evergreen rain forest. Important taxa here arethe arborescent species Pereskia grandifolia,and particularly a large number of epiphyticspecies of Rhipsalis and related genera. Someother species in this centre grow in isolatedazonal habitats such as coastal sand dunes orrock outcrops (“inselbergs”, Figs. 5 & 6) (mostlyCereeae, same genera as in the Caatinga cen-tre). only small patches of the original vegeta-tion of the south-east Brazil Atlantic rainforestare left today.
40 © Schumannia 7 2015
Centre Chihuahua Puebla-Oaxaca Sonora-Sinaloan Jalisco
area size (km²) 1,020,000 47,500 12,500 12,500
vegetation deserts and xeric tropical dry forests, transition of Sonoran tropical dry forestshrublands deserts and xeric desert and Sinaloan
shrublands dry forests
mainWWF Chihuahuan desert (NA1303); Tehuacán valley Sonoran-Sinaloan Jalisco dry forestsecoregions Central mexican matorral matorral (NT1316), transition subtropical (NT0217)
(NA1302); Sierra madre Balsas dry forests dry forest (NA0201)oriental pine-oak forests (NT0205)(NA0303)
dominating Cacteae Cacteae Phyllocacteae – Phyllocacteae –taxa Echinocereinae, Cacteae, Echinocereinae &
Opuntia Hylocereinae, Cacteae,Opuntia
most species globular globular globular globular, epiphyticrich growth form
dominating bee pollination bee pollination bee pollination bee pollinationpollination syndrome
Table 1: The North/Central American diversity centres
Centre Southern Central Andes Caatinga Mata Atlântica
area size (km²) 297,500 157,500 40,000
vegetation inter-Andean dry forests and xeric shrubland and thorn atlantic rain forests, coastalshrublands with steep rocky forest with rock outcrops sand dunes, granit rockslopes outcrops
mainWWF ecoregions Bolivian montane dry forests Caatinga (NT1304), Campos Serra do mar coastal forests(NT0206), central Andean puna Rupestres montane savanna (NT0160); Bahia coastal(NT1002), southern Andean (NT0703), Atlantic dry forests forests (NT0103)yungas (NT0165) (NT0202)
dominating taxa Trichocereinae Cereinae Rhipsalideae
most species rich growth form globular columnar epiphytic
dominating pollination syndrome bee pollination bird pollination, bat pollination bee pollination
Table 2: The South American diversity centres

In addition to those seven centres presentedabove, very small areas in Rio Grande do Sul(south-east Brazil) and in south-east Paraguayreach more than 23 species per 2,500 km². Es-pecially south-east Paraguay is an importantarea with high genus diversity (map 10, seechapter 5.6 for a more detailed discussion of di-versity patterns at higher taxonomic level).Even though the Atacama Desert and thewestern slopes of the Andes in Peru harbouronly moderate species diversity, they are hometo a) a high diversity of species with extremelysmall distribution ranges (compare chapter 5.3)and b) intermediate to high numbers of genera(compare chapter 5.6).The Andes of northernPeru are one of the few regions were at least 7out of the 11 tribes of the cacti occur (map 27).Additional centres of high taxonomic diversityare the coastal mountains of northern Vene-zuela with high numbers of genera and tribes,and parts of the Greater Antilles, especiallysouth-east Cuba and the islands of Hispaniolaand Puerto Rico (maps 26 & 27).
5.3 Range-sizes of Cacti
most species of cacti have extremely small dis-tribution ranges. As a comparison: whereas
“Birdlife International” uses a threshold of50,000 km² range size for the definition of en-demic bird areas, almost half of the cactusspecies have range sizes smaller than 10,000km² (map 13). more than 300 species haveranges even smaller than 2,000 km². Examplesof species with extremely small ranges, re-stricted to one or a few populations, are Dis-cocactus horstii in minas Gerais (Brazil),Aztekium ritteri and Geohintonia mexicana inthe Sierra madre oriental in Nuevo Leon(mexico), andCereus insularis on the islands ofFernando de Noronha (Brazil) (compare Figs.35–37).Looking at the distribution patterns of therange-restricted species (“narrow endemics”with range sizes smaller than 10,000 km², map13), only some of them occur in one of theseven diversity centres listed above. Narrowendemic species are found only in the southerncentral Andes centre (e.g. species of Cleisto-cactus, Parodia, Rebutia), the Puebla-oaxacacentre (many Mammillaria species) as well asin the (south-)eastern parts of the Chihuahuacentre (many species of Mammillaria, Thelo-cactus, Turbinicarpus, and others). Additional‘centres’ of range-restricted cacti are parts ofthe coastal Atacama Desert and the westernslopes of theAndes, especially in northern Peru
(e.g. Espostoa, Haageocereus, Matucana, andmany smaller genera), and the northern partsof the Chilean matorral (mainly Copiapoa,Eriosyce). Small numbers of range-restrictedspecies occur all over south-east Brazil, north-ern and central Argentina and Chile, and largeareas of mexico and Costa Rica.At least some widespread species occur in al-most all parts of the overall distribution rangeof the Cactaceae.As a result, the median rangesizes for the species of a given place is higherthan 100,000 km² in most grid cells (map 14)—even though most cactus species have distri-bution ranges of less than 10,000 km²! How-ever, those 50% species with small ranges arescattered over many places and only few ofthem co-occur in one place.The species with the largest range size withinthe Cactaceae is the tricontinental Rhipsalisbaccifera. Also within the Americas, Rhipsalisbaccifera has the largest distribution range ofall cacti species. many species with very largerange sizes are either epiphytic Rhipsalideaeor Hylocereinae (e.g.,Hylocereus,Lepismium)or shrubby opuntioideae. Nevertheless, evenwithin the epiphytic growth form, most speciesof the genera Weberocereus in Central Amer-ica or Schlumbergera in south-east Brazil havevery small known distribution ranges. In the
41© Schumannia 7 2015
250 kmSpecies per 100 km²
Diversity centres in Central America
1 - 1011 - 2324 - 2829 - 3536 - 4041 - 5051 - 6061 - 73Diversity centres
Map 11
© Barthlott et al. 2013
(defined as the top 5% most species rich areas)
Map 11: Diversity centres in Central America.Karte 11: Diversitätszentren in MIttelamerika (definiert als dieoberen 5 % artenreichsten Gebiete; Arten pro 100 km²).
500 km
Diversity centresin South America
Species per 100 km²
1 - 1011 - 2324 - 2829 - 3536 - 4041 - 4546 - 5051 - 73Diversity centres
Map 12
© Barthlott et al. 2013
(defined as the top5% most speciesrich areas)
Map 12: Diversity centres in South America.Karte 12: Diversitätszentren in Südamerika (definiert als dieoberen 5 % artenreichsten Gebiete; Arten pro 100 km²).

42 © Schumannia 7 2015
1500 km
Range-restricted species('endemics')Species per 2500 km²with ranges under 10,000 km²
© Barthlott et al. 2013
123 - 45 - 67 - 89 - 1011 - 1516 - 25
Map 13
Map 13: Range-restricted species (‘endemics’).Karte 13: Arten mit kleinem Areal (‚Endemiten’; Arten pro 2500 km² mit Arealen kleiner als 10.000 km²).
Range-restricted species('endemics')Species per 2500 km²with ranges under 10,000 km²
©
123 - 45 - 67 - 89 - 1011 - 1516 - 25
M

genus Rhipsalis, more than one third of thespecies have distribution ranges smaller than10,000 km². Looking at the largest genera ofthe family, the range size distribution is veryuneven. In the large genusMammillaria, whichis mostly restricted to mexico and southernuSA, more than half of the species have dis-tribution ranges smaller than 2,000 km² (2/3less than 10,000 km²), whereas some taxa suchas M. heyderi, M. lasiacantha, or M. grahamiiare really widespread. A similar situation isfound in Parodia, where 70% of the species—mainly in Rio Grande do Sul (Brazil) and uru-guay on the one hand and the southern centralAndes centre in Bolivia and Argentina on theother hand—have range sizes smaller than10,000 km². In contrast, most species of thegenus Opuntia are widespread, and only a fewhave restricted ranges of less than 10,000 km².Echinopsis, which as a genus is restricted main-ly to the (southern) central Andes, has speciesof all range sizes, similar to Echinocereus inmexico and the southern uSA. Gymnocaly-cium, which is mainly from Argentina, is an ex-ample for a genus where most species havevery small range sizes—similar, e.g., toEriosycein Chile.
There are other genera, which as a genus havelarge ranges, but include many restricted rangespecies. In the genus Pilosocereus, which oc-curs from south-east Brazil to northern mex-ico, almost half of the species have distributionranges smaller than 10,000 km², and only fewhave ranges of several hundred thousand km².A somewhat similar situation can be found inthe genus Melocactus, which has many re-stricted range species especially in south-eastBrazil but can be found even in mexico andthe Caribbean.
5.4 Spatial distribution of growthforms
There is a strong difference in the diversity pat-terns of the two most species rich growth formswithin cacti: the globular and the columnarspecies (maps 15 & 16). By far the highestspecies numbers of globular cacti occur in theChihuahua centre (almost exclusively tribeCacteae). only moderate species numbers arefound in the southern central Andes centre(mainly Cereeae subtribes Trichocereinae andNotocacteae), and in Rio Grande do Sul
44 © Schumannia 7 2015
Fig. 33: An extraordinary life-form within theOpuntieae: liana Tacinga braunii at its typelocality in Brazil (near Itaobim, state MinasGerais). Photo: W. Barthlott.Abb. 33: Eine außergewöhnliche Lebensforminnerhalb der Opuntieae: die Liane Tacingabraunii an ihrem Typfundort in Brasilien(bei Itaobim, Staat Minas Gerais).
1500 km
Growth form Globular:Species diversity (692 sp.)Species per 2500 km²
1 - 23 - 56 - 1011 - 1516 - 2526 - 3536 - 4546 - 56
Map 15
© Barthlott et al. 2013
1500 km
Growth form Columnar:Species diversity (205 sp.)Species per 2500 km²
1 - 23 - 45 - 67 - 89 - 1011 - 1213 - 1415 - 16
Map 16
© Barthlott et al. 2013
Map 15: Growth form globular, species diversity (692 sp).Karte 15: Wuchsform kugelig, Artendiversität (692 Arten;Arten pro 2500 km²).
Map 16: Growth form columnar, species diversity (205 sp).Karte 16: Wuchsform säulig, Artendiversität (205 Arten;Arten pro 2500 km²).

46 © Schumannia 7 2015
Fig. 35: Discocactus horstii, extremelyrange-restricted species (near Diamantina,Minas Gerais, Brazil). Photo: W. Barthlott.Abb. 35: Discocactus horstii, eine Art mitextrem kleinem Verbreitungsgebiet(bei Diamantina, Minas Gerais, Brasilien).
Fig. 36: Highly endemic Pelecyphora asellifor-mis in the Mexican state San Luis Potosí. Photo:W. Barthlott.Abb. 36: Pelecyphora aselliformis ist im mexi-kanischen Staat San Luis Potosí endemisch.
Fig. 37: Highly endemic Geohintonia mexicanain Nuevo León, Mexico. Photo: W. Barthlott.Abb. 37: Geohintonia Mexicana, endemisch inNuevo León, Mexiko.
Fig. 39: Cereus jamacaru, an example for an arborescent growth formwith columnar branches, with Werner Rauh (right) and Diedrich J. Supthut(left) in the Brazilian state Bahia. Photo: W. BarthlottAbb. 39: Ein Beispiel für eine baumartige Wuchsform mit säuligen Ästen:Cereus jamacaru, mit Werner Rauh (rechts) und Diedrich J. Supthut (links)im brasilianischen Staat Bahia.
Fig. 38: Micranthocereus polyanthus in a field of white quartz sand nearCaetité (Bahia, Brazil). Photo: W. Barthlott.Abb. 38: Micranthocereus polyanthus in weißem Quarzsand bei Caetité(Bahia, Brasilien).
pischen Trockenwaldvegetation. In der Folgeist dieses Zentrum nicht nur Heimat kugeligerMammillaria-Arten und häufiger säuliger undbaumförmiger Kakteen wie Stenocereus, Ce-phalocereus (alle Cacteae) und Opuntia, son-dern auch von epiphytischen Arten der Gat-tungen Epiphyllum und Selenicereus.
Die drei südamerikanischen ZentrenSüdliches Zentralanden-Zentrum: Das größte
südamerikanische Diversitätszentrum beher-bergt auch die höchste Artenanzahl in Süd-amerika und ist durch die höchste phylo-genetische und morphologische Diversität ge-kennzeichnet. Zehn der hier akzeptierten elfKakteen-Triben kommen in dem südlichenZentralanden-Zentrum vor. mit in großen Tei-len dieses Zentrums acht vorkommenden Tri-ben pro 2.500-km²-Rasterfeld ist die Anzahldoppelt so groß wie in den meisten Gebieten
des Chihuahua-Zentrums. Das Zentrum be-findet sich in den semiariden und ariden Tälerndes zentralen und südlichen Boliviens sowiedes nördlichen Argentiniens. Im nördlichenTeil des Zentrums umfasst die Vegetation laub-werfende Wälder mit dispersen, natürlichwaldfreien Steilhängen und felsigen Lichtun-gen. Im Süden, der Präpuna-Ökoregion, domi-nieren Andenbuschländer und Halbwüsten.Dieses Zentrum beherbergt die höchste An-zahl kugeliger Kakteen in Südamerika, ist aberauch ein Zentrum der Säulenkakteen. Es istdie einzige südamerikanische Region mit einersignifikanten Anzahl polsterbildender Kak-teen. Zusätzlich zu den vorwiegend bienen-bestäubten Arten hat es eine beachtliche An-zahl schwärmerblütiger Arten. Besonders derwestliche Teil der bolivianischen Provinz Ta-rija und die benachbarten Gebiete Saltas undJujuys in Nordargentinien sind extrem vielfäl-tig, wenn man den Artenreichtum in einer fei-nen Skala von 10 x 10 km-Rasterzellen be-trachtet (Karte 12).Caatinga-Zentrum:Dieses brasilianische Zen-trum liegt im südlichen Teil der Caatinga-Öko-region und den Campos Rupestres, die an dieBergketten der Serra Espinhaço und ChapadaDiamantina anknüpfen. Während trockenesBuschland und Dornwald die vorherrschendenVegetationstypen der Caatinga-Region sind, istdie höchste Kakteenartendiversität in azona-len Habitaten wie Felskuppen (sog. Inselberge)oder auf nährstoffarmen sandigen Böden zufinden. obwohl die Artenvielfalt im Vergleichzu dem Chihuahua- oder südlichem Zentra-landen-Zentrum viel niedriger ist, kommt hiereine hohe Anzahl von Gattungen – besondersder Tribus Cereeae – vor. Dementsprechend istdieses Zentrum durch eine mittlere Artenzahlkugeliger Kakteen, aber eine hohe Diversitätsäuliger Arten charakterisiert. Das Caatinga-Zentrum hat die höchsten Zahlen von durchFledermäuse und Vögel bestäubten Arten undnur geringe Zahlen bienenbestäubter Arten.Mata Atlântica-Zentrum: Dieses Zentrum inSüdostbrasilien ist von allen anderen Zentrenökologisch recht deutlich unterschieden, da esdurch feuchten, immergrünen Regenwald ge-kennzeichnet ist. Wichtige Taxa sind hier diebaumförmige Art Pereskia grandifolia und be-sonders eine hohe Anzahl epiphytischer Artenvon Rhipsalis und verwandten Gattungen. Ei-nige andere Arten dieses Zentrums wachsenin isolierten azonalen Habitaten wie Küsten-dünen oder Felskuppen (Inselberge, Abb. 5 &6) (meist Cereeae, die gleichen Gattungen wieim Caatinga-Zentrum). Nur kleine Flecken derursprünglichen Vegetation des südostbrasilia-nischen Regenwaldes sind heute noch übrig.Außer den sieben, oben vorgestellten Zentrenerreichen sehr kleine Gebiete in Rio Grande
56 © Schumannia 7 2015
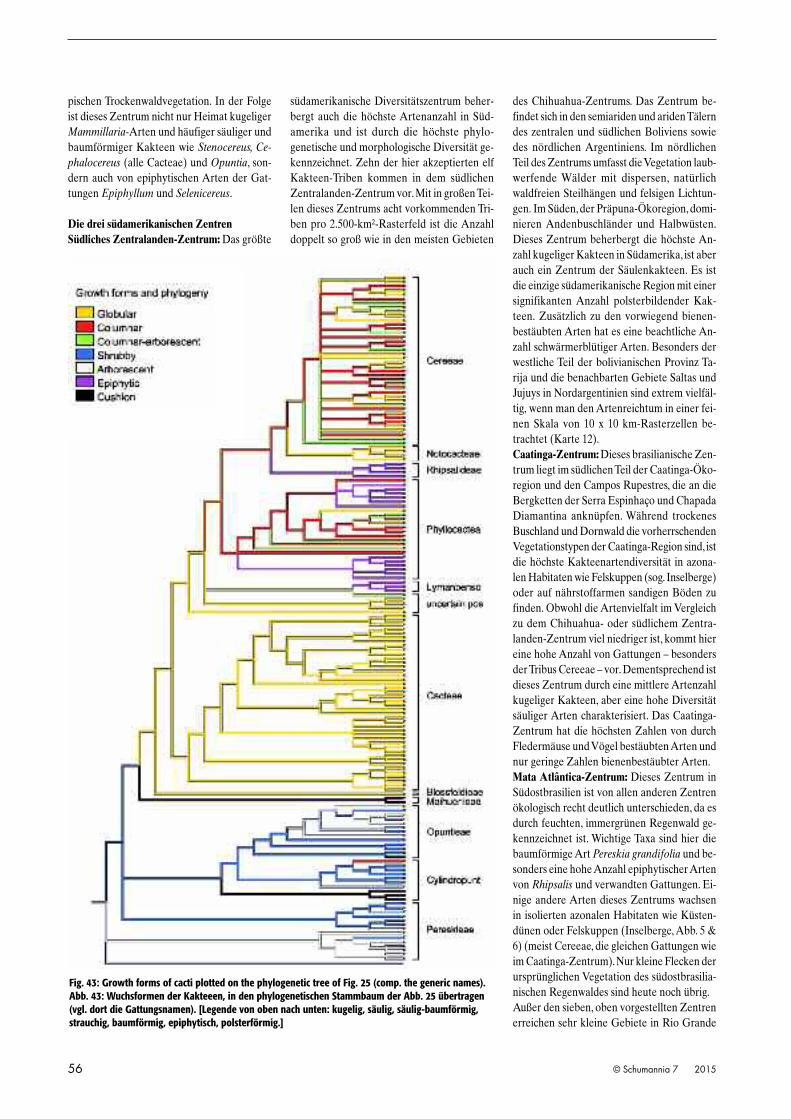
Fig. 43: Growth forms of cacti plotted on the phylogenetic tree of Fig. 25 (comp. the generic names).Abb. 43: Wuchsformen der Kakteeen, in den phylogenetischen Stammbaum der Abb. 25 übertragen(vgl. dort die Gattungsnamen). [Legende von oben nach unten: kugelig, säulig, säulig-baumförmig,strauchig, baumförmig, epiphytisch, polsterförmig.]

meter große Verbreitungsgebiete. Eine etwasähnliche Situation ist in der Gattung Meloca-ctus zu finden,die viele kleinräumig verbreiteteArten besonders in Südostbrasilien hat, abersogar in mexiko und der Karibik vorkommt.
5.4 Räumliche Verbreitung derWuchsformen
Die Verbreitungsmuster der zwei artenreich-sten Wuchsformen innerhalb der Kakteen (diekugeligen und die säuligen Arten) zeigen deut-liche unterschiede (Karten 15 & 16). Bei wei-tem die höchsten Artenzahlen der Kugelkak-teen kommen im Chihuahua-Zentrum vor(fast ausschließlich der Tribus Cacteae). Nurmittlere Artenzahlen findet man im südlichenZentralanden-Zentrum (vorwiegend Cereeaeder Subtriben Trichocereinae und Notoca-cteae) und in Rio Grande do Sul (Südost-Bra-silien). Geringe Artenzahlen findet man in derCaatinga (vorwiegendMelocactus). Im Gegen-satz dazu gibt es fast keine Säulenkakteen imzentralen Teil des Chihuahua-Zentrums; derenhöchste Diversität ist in der Caatinga zu finden(besonders Cereeae Subtribus Cereinae, Abb.39), im südlichen Zentralanden-Zentrum (be-sonders Cereeae Subtribus Trichocereinae)und in der Atacama-Wüste und den westlichenHängen der Anden in Peru (Cereeae Subtri-bus Trichocereinae). Das kleine Puebla-oa-xaca-Zentrum in mexiko hat, im Gegensatz zudem benachbarten Chihuahua-Zentrum,eben-falls eine signifikante Anzahl säuliger Kakteen,hauptsächlich aus der Phyllocacteae SubtribusEchinocereinae.Die strauchigen und baumförmigen Arten vonPereskia und den opuntioideae (Karten 18 &19,Abb. 40) weisen ihre höchsten Artenzahlenim Chihuahua-Zentrum und dem Puebla-oa-xaca-Zentrum auf, und nur eine geringe Ar-tenvielfalt in Südamerika.Epiphytische Kakteen haben, verglichen mitdem Rest der Familie, deutlich verschiedeneökologische Präferenzen (Karte 20). Die epi-phytischen Rhipsalideae (Abb. 41, vorwiegendsüdamerikanisch, besonders Südost-Brasilien)und Hylocereinae (vorwiegend mittelamerika)sind am artenreichsten in den Tieflandregen-wäldern der mata Atlântica und mittelameri-kas. Jedoch kommen nur wenige Arten beiderTriben (z. B. Disocactus amazonicus und Stro-phocactus witii) in dem großen Regenwaldge-biet des Amazonasbeckens vor – vorwiegendim westlichen Amazonien.Polsterbildende Kakteen (Abb. 42, hauptsäch-lich Phyllocacteae-Echinocereinae und -Cylin-dropuntieae, sowieMaihuenia) sind an speziel-len Standorten mit Frost oder stark trocknen-den Winden, wie in den Hochanden Süd-
62 © Schumannia 7 2015
Fig. 45: Convergent evolution of red hummingbird flowers in South and North America in unrelatedgenera of small globular cacti: Mammillaria poselgeri from Mexico (Baja California) (left), Matucanaformosa from N-Peru (right). It is almost typical that a different flower syndrome within the predo-minantly entomophilous genus Mammillaria led to creation of a separate genus (Cochemiea posel-geri) in the past. Photos: W. Rauh.Abb. 45: Konvergente Entstehung roter Kolibriblumen in Süd- und Nordamerika bei nicht verwand-ten Gattungen kleiner Kugelkakteen: links Mammillaria poselgeri aus Mexiko (Baja California),rechts Matucana formosa aus N-Peru. Es ist beinahe typisch, dass ein abweichendes Blütensyndrominnerhalb der vorwiegend entomophilen Gattung Mammillaria früher zur Abtrennung als eigeneGattung (Cochemiea poselgeri) führte.
Fig. 46: Convergent evolution of night-blooming, white, hawkmoth pollinated flowers with extremelylong flower tubes in not related species: the dwarf columnar cactus Echinopsis mirabilis from Argen-tina (left), Epiphyllum phyllanthus (right), a widespread (Mexico to N-Argentina) rainforest epiphyte.E. phyllanthus is one of the extremely spingophilous species with a flower tube of up to about 20 cmlength and can probably be pollinated only by the moth Cocytius cruentus. Photos: W. Barthlott.Abb. 46: Konvergente Entstehung nachtblühender weißer Schwärmerblumen mit extrem langen Blüten-röhren bei nicht verwandten Arten: links der zwergige Säulenkaktus Echinopsis mirabilis aus Argenti-nien, rechts Epiphyllum phyllanthus, ein weitverbreiteter (Mexiko bis Nordargentinien) Regenwald-Epiphyt. E. phyllanthus ist einer der extrem spingophilen Arten mit einer Blütenröhre von bis über20 cm Länge und kann möglicherweise nur von dem Schwärmer Cocytius cruentus bestäubt werden.

6.1 Introduction
As cacti represent popular plants of great or-namental value, for a long time they have beenthe subject of conservationist concerns (e.g.,BENSON 1982, OLDFIELD 1984a, b). Due to a rel-atively better knowledge of many species, butalso as a consequence of early overexploita-tion for horticulutural uses, cacti, since the1970s, were among the first plants to becomeprotected under international law. The wholefamily Cactaceae is included in theAppendicesI and II of the “Convention on internationaltrade in endangered species of wild fauna andflora” (CITES).This protection was especiallyrelated to the international trade in endan-gered species and to high demands in the or-namental plant market, but it did not takethreats from other sources into account.Withincreasing global conservation efforts aimingat the establishment of a more effective pro-tected area network,numerous species of Cac-taceae have also benefitted from in-situ con-servation measures. Still, the distributionranges of 19% of the species ranges do notoverlap with any protected area and 40% ofthe species ranges are not covered by protectedareas of the IUCN categories I to IV. Most ofthese have distribution ranges smaller than50,000 km². Clearly, the representation withina protected area does not automatically guar-antee effective conservation, and the existenceof a species outside protected areas is not in-dicating imminent threat.The geographical assessment of cacti pre-sented here can be a starting point for a moreinformed evaluation of their threat and con-servation status. Clearly, many threats to bio-diversity in general, and to cacti in particular,have been dynamically unfolding and chang-ing over the last decades. Still relevant treat-ments of cacti conservation have been pro-vided by TAYLOR & al. (1997) and TAYLOR &ZAPPI (2004), which, however can be comple-mented by integral and systematic assess-ments. The systematic discussion of threats tocacti according tomodern threat classificationapproaches (SALAFSKY& al. 2008) is a first step,
which has also been adopted in the course ofthe elaboration of the IUCN Red List (IUCN2012).Additionally, spatial analyses allow fora certain differentiation of conservation prob-lems in the different regions of the Cactaceaerange. In the future, these spatial analyses canbe refined further, e.g. including detailed gapanalyses.This is urgently required as the IUCNRed List is far from being complete and up todate.The IUCN Red List still shows a strong biastowards a few well investigated countries suchas Brazil and Mexico. About 48% of all cactispecies,which have been assigned to one of theRed List categories Vulnerable, Endangered,Critically endangered, Extinct in the wild orExtinct, areMexican taxa, and 49% are Brazil-ian. The analysis also might underestimate thethreat status of species living outside arid re-gions. According to the current Red List(IUCN 2012), about 60% of the most endan-gered species (categories as above) would livein deserts or rocky areas, and only about 12%are forest taxa. However, in the case of manyepiphyte species from forest habitats (e.g., in
the very highly fragmented and deforestedMata Atlantica region, Brazil) it seems likelythat the conservation status of many cactusspecies is poor. IBISCH & al. (2000) indicatedthat a few Bolivian epiphytic cacti would be atleast vulnerable to extinction,and the situationis likely to have worsened.According to IUCN(2012) however, the only epiphyte species thatare classified as vulnerable or more critical areRhipsalis cereoides, R. crispata, R. pilocarpa,R. russellii, and Schlumbergera kautskyi (allfromBrazil). Intensemonitoring efforts are re-quired to detect significant population declines.These are often affordable only for rich coun-tries, where even single populations and vari-eties are carefully observed (e.g., compare iso-lated populations in the Sonoran desert ofsouth-central Arizona of Nichol’s turk’s headcactus Echinocactus horizonthalonius var.nicholii with a dramatic decline from 1995 to2008 (MCINTOSH & al. 2011).In the followingwe discuss the principal threatsto cacti adopting the standard classification bySALAFSKY & al. (2007), which is also applied byIUCN (2012).
68 © Schumannia 7 2015
6 Conservation and hotspots:cactus diversity in change
Pierre L. Ibisch & Jens Mutke
Fig. 51: Shield inselberg in Bahia, Brazil, with Melocactus ferreophilus; agricultural not usable relicthabitat in a destroyed landscape. Photo: W. Barthlott.Fig. 51: Schildinselberg in Bahia, Brasilien, mit Melocactus ferreophilus; ein landwirtschaftlich nichtnutzbarer Relikt-Standort inmitten einer zerstörten Landschaft.

6.2 Threats to Cactaceae
Residential and commercial development, hu-man settlements or other nonagricultural landuses with a substantial footprintMany fast growing cities exist in (sub)tropicalarid areas, where agriculture-related land usechange tends to be less important. With in-creasing human population growth, develop-ment, and urbanization, area consumption forresidential and commercial infrastructure be-comes ever more relevant also for cacti. Anexample is Cleistocactus acanthurus, which isthreatened by the development of Lima,Chosica and satellite cities (MOTTRAM 1998). InLatin America, a significant number of largecities are located in more or less narrow aridvalleys with relevant occurrences of cacti. Forexample, in Bolivia, the rapid growth of the cityof Cochabamba and satellite settlements drivesthe conversion of habitats both of the bottomand the slopes of the valley system affectingcacti populations of Cleistocactus parviflorus,Echinopsis obrepanda, Parodia schwebsianaamong others (previously even benefitted bydeforestation, IBISCH 2004b) (Fig. 55). In an as-sessment of the Cactaceae of the Querétanosemi-desert it was concluded that fewer specieswere threatened by residential developmentthan other land-use changes; however, the cor-responding impact was consideredmore severe(CHÁVEZ & al. 2007). While agricultural land-use impacts mainly zonal ecosystems withmore or less fertile soils, residential develop-ment also affects azonal sites such as rock out-crops or steep slopes, especially in the case ofmegacities suffering from decreasing space forexpansion (Figs. 51–52). Non-agricultural landconsumption must not be neglected as a prob-lem especially for specialist plants in azonalhabitats. Currently, 14% of the cacti recordedby IUCN (2012), which have been assigned toone of the Red List categoriesVulnerable,En-dangered, Critically endangered,Extinct in thewild or Extinct suffer from threats belonging tothis category.
Agriculture and aquaculture threats fromfarming and ranching as a result of agriculturalexpansion and intensification, including silvi-culture, mariculture, and aquacultureAbout 14% of all cacti species, which havebeen assigned to one of theRedList categoriesVulnerable, Endangered, Critically endan-gered, Extinct in thewild or Extinct, are threat-ened by agriculture driven land-use change(IBISCH & al. 2000, IUCN 2012,MÜLLER 2007).For a long time, many cacti, in many regionsbenefited from extensive land-use. Deforesta-tion and the subsequent grazing in (semi-)aridshrub-lands often led to the expansion of suit-
able habitat for species that were stress-toler-ant and resistant to herbivory, but poor com-petitors, includingmany terrestrial cacti (IBISCH2004a). Indeed,grazing can benefit cactus pop-ulations (e.g., in the case of someMammillariaspecies in Tehuacán Valley, MARTORELL & PE-TERS 2009). However, with increasing intensityof grazing the degradation of ground vegeta-tion and soils leads to severe hydric erosiontriggering loss of habitat, and the intensifica-tion of agricultural land-use e.g. applying mas-sive mechanical clearing, burning and irriga-tion the elimination of cactus habitat is pro-moted. With rapidly growing demand foragricultural commodities and arable land, ad-vancement of industrial agriculture even un-der rather marginal conditions, as well as pro-gressing desertification, land conversion is be-coming an ever more important threat tospecies of Cactaceae. This is also increasinglyreflected in reports about principal threats: ex-cessive pasturing and exploitation of thedesert, habitat fragmentation and land-use
changes, and anthropogenic fire are increas-ingly named as relevant conservation problems(e.g.,BRAILOVSKY SIGNORET &HERNÁNDEZ 2010,CHÁVEZ& al. 2007,CLARK-TAPIA& al. 2005,EM-MING 2005, GOLUBOV & al. 2009, LEON DE LALUZ 2005). In some cases, the compositionalchange of plant communities is causing threats;e.g., Opuntia humifusa, originally inhabitingopenQuercus savanna vegetationwas reportedto suffer from shading by an encroachingcanopy ofQuercus and Pinus, and smotheringby leaf litter (ABELLA & JAEGER 2004). Af-forestation activities can threaten habitats ofCactaceae, such as in the case of Neoporteriaaspillagae being affected byPinus radiata plan-tations in Chile (NOVOA 2002).
Energy production and mining threats fromproduction of nonbiological resourcesMining and extraction of rocks can very specif-ically threaten species of Cactaceae at azonalsites otherwise not affected by land-use changes.For instance, this has been reported for Turbi-
69© Schumannia 7 2015
Fig. 52: The highly mimetic Ariocarpus kotschoubeyanus (red circle) next to a flowering Proboscideain the Mexican state San Luis Potosí. Photo: W. Barthlott.Abb. 52: Der hoch mimetische Ariocarpus kotschoubeyanus (roter Kreis) neben einer blühendenProboscidea im mexikanischen Staat San Luis Potosí.

71© Schumannia 7 2015
1000 kmCanada: species diversity and protected areasSpecies per 100 km²
M
© Barthlott et al. 2013
Protected areas
123
Map 47
©
Map 47: Canada. Species diversity and protected areas. Country area: 9.093.507 km²; 3 species (0 endemic); 2 genera (0 endemic); 0 range-restrictedspecies; 0 species in CITES Appendix I; 0 species in the IUCN Red List.Karte 47: Kanada. Artendiversität und Schutzgebiete (Arten pro 100 km²). Staatsgebiet: 9.093.507 km²; 3 Arten (0 endemisch); 2 Gattungen (0 endemisch);0 Arten mit begrenztem Areal; 0 Arten im CITES-Anhang I; 0 Arten in der IUCN-Roten-Liste.
Map 48: USA. Species diversity and protected areas. Country area: 9,476,600 km²; 173 species (32 endemic); 33 genera (1 endemic); 24 range-restrictedspecies; 18 species in CITES Appendix I; 6 species in the IUCN Red List.Karte 48: USA. Artendiversität und Schutzgebiete (Arten pro 100 km²). Staatsgebiet: 9.476.600 km²; 173 Arten (32 endemisch); 33 Gattungen(1 endemisch); 24 Arten mit begrenztem Areal; 18 Arten im CITES-Anhang I; 6 Arten in der IUCN-Roten-Liste.
1000 kmUSA: species diversity and protected areasSpecies per 100 km²
Map 48
1 - 56 - 1011 - 1516 - 2021 - 2526 - 3031 - 4041 - 52
© Barthlott et al. 2013
Protected areas

7.1. Subfamily: Pereskioideae
PereskiaMill. (Maps 64–66)17 species.Woody climbers, shrubs or trees,nor-mally 4–5 m; stems not conspicuously succu-lent, spiny; with broad persistent leaves. Flow-ers rotate white, orange, red or purplish. Fruitsfleshy globose, red, yellow or black. Distribu-tion: Caribbean, Central America, northernColombia,Venezuela (Guyanas, coastal regions),northern Peru (Andes), Bolivia, Paraguay,northern Argentina (Andes and lowlands),Brazil to northern Uruguay (Atlantic region).Centres of diversity:Caribbean (Haiti),Bolivia(eastern slopes of the Andes), eastern Brazil.Main references: LEUENBERGER (1986), AN-DERSON (2001), TAYLOR & ZAPPI (2004), HUNT(2006).
7.2. Subfamily: Maihuenioideae
Maihuenia Phil. (Map 67)2 species. Low cushion-forming, with persist-ing leaves. Flowers rotate,white, yellow or red.Fruits fleshy.Distribution: central Chile to cen-tral Argentina. Main references: KIESLING(1999),ANDERSON (2001),HOFFMANN&WALTER
(2004), HUNT (2006).
7.3. Subfamily: Opuntioideae
Austrocylindropuntia Backeb. (Map 68)6 species. Small to large shrubs, cylindric, withmore or less tuberous, finally deciduous leaves.Flowers rotate, yellow,pink or red.Fruits juicy.Distribution: Ecuador to northern Argentina(Andes). Main references: JØRGENSEN (1999),ANDERSON (2001), TAYLOR & ZAPPI (2004),HUNT (2006).
BrasiliopuntiaA. Berger (Map 69)1 species.Lage tree up to 25m,erect,with clus-ters of spines. Flowers near apex of leader orterminal segment or borne by proliferation.
Flowers yellow.Fruits solitary or clustered,glo-bose to ovoid. Distribution: Peru, Bolivia,northernArgentina, southern Paraguay and incoastal regions of Brazil.Main references:KIES-LING (1999), ANDERSON (2001), TAYLOR & ZAPPI(2004).
Consolea Lem. (Map 70)3 species. Treelike with trunk and primarybranches cylindric, not articulated, ultimate;segments flattened as in Opuntia. Flowerssmall, diurnal. Fruits juicy, ovoid.Distribution:Caribbean islands (mostly coastal rocks).Mainreferences:ANDERSON (2001), HUNT (2006).
Corynopuntia F.M. Knuth (Maps 71–72)14 species.Dwarf caespitose opuntioid shrubs;branches cylindric-clavate. Flowers yellow orrose to purple. Fruits ellipsoid fleshy at first,later dry, yellow to brownish, smooth, some-times spiny.Distribution: from central Mexicoto southern USA. Centre of diversity: Chi-huahuan Desert. Main references: GUZMÁN &al. (2003), HUNT (2006), USDA & NRCS(2008).
Cumulopuntia F. Ritter (Map 73)5 species. Low clumps. Flowers yellow, some-times red; young shoots with small, cylindrical,deciduous leaves. Fruits thick-walled andfleshy.Distribution: southern Peru, south-west-ern Bolivia, northern Chile and north-westernArgentina. Centre of diversity: south-easternBolivia. Main references: BRAKO & ZARUCCHI(1993), ANDERSON (2001), HUNT (2006).
Cylindropuntia (Engelm.) F.M.Knuth (Maps74–79)33 species.Shrubby or treelike,branched.Flow-ers variously coloured.Fruits fleshy or dry.Dis-tribution: south-western USA to Mexico,Caribbean islands. Centre of diversity: north-westernMexico to south-westernUSA (south-ern California to Arizona). Main references:ANDERSON (2001),GUZMÁN& al. (2003),USDA& NRCS (2008).
GrusoniaRchb. f. ex Britton &Rose (Map 80)1 species. Low shrubs. Flowers variouslycoloured.Fruits cylindric, various, fleshy or dry.Distribution: northern Mexico (Coahuila).Main references: GONZALEZ ELIZONDO & al.(1991), VILLARREAL QUINTANILLA (2001), GUZ-MÁN & al. (2003).
Maihueniopsis Speg. (Map 81)8 species. Low compact cushions; roots tuber-ous. Flowers yellow, orange, pink or red. Fruitsjuicy, thick-walled, indehiscent. Distribution:southern Bolivia,Argentina and eastern Chile.Main references: KIESLING (1999), ANDERSON(2001), HOFFMANN & WALTER (2004), HUNT(2006).
Miqueliopuntia Fric ex F. Ritter (Map 82)1 species. Shrubby, forming clumps up to 1.4 m,Flowers almost white, fruits globose,pale greento whitish, juicy. Distribution: central Chile(coastal regions). Main references: ANDERSON(2001), HOFFMANN &WALTER (2004).
Nopalea Salm-Dyck (Map 83)3 species.Tree-like,with a cylindric trunk.Flow-ers purplish pink or red. Fruits ovoid. Distri-bution: north-easternMexico toCentralAmer-ica. Main references: KIESLING (1999), ANDER-SON (2001), GUZMÁN & al. (2003), TAYLOR &ZAPPI (2004), HUNT (2006).
OpuntiaMill. (Maps 84–95)64 species.Trees and shrubs, some low to cush-ion-forming; stem normally segmented; seg-ments cylindric, clavate, subglobose, or moreor less flattened. Flowers rotate, yellow, pink,red or whitish. Fruits umbilicate, fleshy or dry.Distribution: southern Canada (temperateforests),most parts of theUSA,CentralAmer-ica, Mexico, Caribbean, north-eastern coast ofColombia and Venezuela, Galápagos islands,western Colombia,Ecuador,Peru,Bolivia,Ar-gentina, Paraguay,Uruguay and south-easternBrazil. Centres of diversity: central Mexico(southernChihuahuanDesert); southernMex-
82 © Schumannia 7 2015
7 Distribution maps of Cactaceae
Wilhelm Barthlott, Kerstin Burstedde, Jan Laurens Geffert, Pierre L. Ibisch, Nadja Korotkova,Andrea Miebach, M. Daud Rafiqpoor, Anke Stein & Jens Mutke
Additional data and revisions contributed by Ralf Bauer, Graham Charles, Paul Hoxey,David Hunt, Martin Lowry & Nigel Taylor

98 © Schumannia 7 2015
Map/Karte 68: Austrocylindropuntia.
Austrocylindropuntiacylindrica floccosapachypusshaferisubulatavestita 500 km
© Barthlott et al. 2013
Map 68
Pereskiaaureifloranemorosasacharosastenanthaweberiana1000 km
Map 66
© Barthlott et al. 2013
Map/Karte 66: Pereskia.
Maihueniapatagonicapoeppigii 500 km
© Barthlott et al. 2013
Map 67
Map/Karte 67: Maihuenia.

109© Schumannia 7 2015
Opuntiaaureabasilarishuajuapensismicrodasyspinkavaepycnantharufidastreptacantha 500 km
Map 94
© Barthlott et al. 2013
Map/Karte 94: Opuntia.
Opuntiaaciculataatrispinaaureispinabravoanachloroticaengelmanniilagunaelittoralismacrocentraoricola 500 km
Map 93
© Barthlott et al. 2013
Map/Karte 93: Opuntia.
Opuntiafuliginosahyptiacanthalasiacanthaleucotrichapiliferarobustastenopetalatehuacana 250 km
Map 95
© Barthlott et al. 2013
Map/Karte 95: Opuntia.

120 © Schumannia 7 2015
1000 kmCleistocactus: Diversity patternSpecies per 2500 km²
123456
Map 127
© Barthlott et al. 2013
Cleistocactusbaumanniibrookeaebuchtieniicandelillachrysocephalushyalacanthuslanicepsluribayensisparapetiensisstrausii 250 km
Map 128
© Barthlott et al. 2013
Map 127: Cleistocactus, diversity pattern.Karte 127: Cleistocactus, Diversitätsmuster (Arten pro 2500 km²).
Map/Karte 128: Cleistocactus.

AAU (2008): AAU Herbarium Database. –www.aubot.dk/search_form.php [accessed02.09.2008].
ABELLA, S. R. & J. F. JAEGER (2004): Ecology of eas-tern prickly pear cactus (Opuntia humifusa) inOak Openings Preserve, northwestern Ohio.– Michigan Bot. 43: 1–11.
ANDERSON, C. L., K. BREMER & E. M. FRIIS (2005): Da-ting phylogenetically basal eudicots using rbcLsequences and multiple fossil reference points.– Amer. J. Bot. 92: 1737–1748.
ANDERSON, E. F. (2001): The cactus family. – Port-land: Timber Press.
ANDERSON, E. F. (2005): Das große Kakteen-Lexi-kon. – Stuttgart: E. Ulmer
ANDERSON, E. F. (2011): Das große Kakteen-Lexi-kon. 2. Aufl. – Stuttgart: E. Ulmer.
ANDERSON, E. F. & W. A. FITZ MAURICE (1997): Ario-carpus revisited. – Haseltonia 5: 1–20.
ANDERSON, E. F., S. ARIAS MONTES & N. P. TAYLOR
(1994): Threatened cacti of Mexico. – Succ.Pl. Res. 2: 5–116.
ANDERSON, E. F., W. A. FITZ MAURICE, B. FITZ MAU-RICE, A. HOFER, M. SOTOMAYOR, A. G. ARRENDONDO
& B. SÁNCHEZ (2002): Turbinicarpus gielsdorfi-anus. – In: IUCN: Red List of threatened spe-cies. Version 2012.1. www.iucnredlist.org[accessed 13.09.2012].
ANDERSON, E. P., J. MARENGO, R. VILLALBA, S. HALLOY,B. YOUNG, D. CORDERO, F. GAST, E. JAIMES & D. RUIZ
(2011): Consequences of climate change forecosystems and ecosystem services in the Tro-pical Andes. – In: R. M. S. K. HERZOG, P. M. JOR-GENSEN & H. TIESSEN: Climate change and bio-diversity in the Tropical Andes: 1–18. Sao Josedos Campos & Paris: MacArthur Foundation,Inter-American Institute of Global Change Re-search & Scientific Committee on Problems ofthe Environment.
ANTONELLI, A. & I. SANMARTÍN (2011): Why are thereso many plant species in the Neotropics? – Ta-xon 60: 403–414.
APPLEQUIST, W. L. & R. S. WALLACE (2001): Phylo-geny of the portulacaceous cohort based onndhF sequence data. – Syst. Bot. 26: 406–419.
APPLEQUIST, W. L. & R. S. WALLACE (2002): Deletionsin the plastid trnT-trnL intergenic spacer defineclades within Cactaceae subfamily Cactoi-deae. – Pl. Syst. Evol. 231: 153–162.
ARAKAKI, M., P.-A. CHRISTIN, R. NYFFELER, A. LENDEL, U.EGGLI, R. M. OGBURN, E. SPRIGGS, M. J. MOORE &E. J. EDWARDS (2011): Contemporaneous andrecent radiations of the world’s major succulentplant lineages. – Proc. Natl. Acad. Sci. USA108: 8379–8384.
ARIAS MONTES, S., S. GAMA LÓPEZ & L. ULISES GUZMÁN
CRUZ (1997): Cactaceae A. L. Juss. – MexicoCity: Universidad Nacional Autonoma de Me-xico.
BACKEBERG, C. (1942a): Cactaceae Lindley. Syste-matische Übersicht (Neubearbeitung) mit Be-stimmungsschlüssel. – Cactaceae (Berlin)1941(2): 1–80.
BACKEBERG, C. (1942b): Zur Geschichte der Kak-teen im Verlauf der Entwicklung des amerika-nischen Kontinentbildes. – Cactaceae (Berlin)1942(2): 1–72.
BACKEBERG, C. (1944): Verbreitung und Vorkom-men der Cactaceae. – Cactaceae (Berlin)1943/1944(2): 1–111, 44 maps.
BACKEBERG, C. (1958): Die Cactaceae. Handbuchder Kakteenkunde 1: Einleitung und Beschrei-bung der Pereskioideae und Opuntioideae. –Jena: G. Fischer.
BACKEBERG, C. (1959): Die Cactaceae. Handbuchder Kakteenkunde 2: Cereoideae (Hylocereeae– Cereeae [Austrocerinae]). – Jena: G. Fischer.
BACKEBERG, C. (1959): Die Cactaceae. Handbuchder Kakteenkunde 3: Cereoideae (Austroca-ctinae). – Jena: G. Fischer.
BACKEBERG, C. (1960): Die Cactaceae. Handbuchder Kakteenkunde 4: Cereoideae (Boreocere-eae). – Jena: G. Fischer.
BACKEBERG, C. (1961): Die Cactaceae. Handbuchder Kakteenkunde 5: Cereoideae (Boreoca-ctinae). – Jena: G. Fischer.
BACKEBERG, C. (1962): Die Cactaceae. Handbuchder Kakteenkunde 6: Nachträge und Index. –Jena: G. Fischer.
BALICK, M. J., M. H. NEE & D. E. ATHA (2000):Checklist of the vascular plants of Belize, withcommon names and uses. – St. Louis: MissouriBotanical Garden.
BÁRCENAS, R. T., C. YESSON & J. A. HAWKINS (2011):Molecular systematics of the Cactaceae. – Cla-distics 27: 1–20.
BARTHLOTT, W. (1977): Kakteen. – Stuttgart: Belser.BARTHLOTT, W. (1983): Biogeography and evolu-
tion in neo- and palaeotropical Rhipsalinae(Cactaceae). – Sonderb. Naturwiss. VereinsHamburg 7: 241–248.
BARTHLOTT, W. (1988): Über die systematischenGliederungen der Cactaceae. – Beitr. Biol.Pflanzen 63: 17–40.
BARTHLOTT, W. & D. HUNT (1993): Cactaceae. – In:K: KUBITZKI: The families and genera of vascu-lar plants 2: 161–197. Berlin [& al.]: Springer.
BARTHLOTT, W. & N. P. TAYLOR (1995): Notes to-wards a monograph of Rhipsalideae (Cacta-ceae). – Bradleya 13: 43–79.
BARTHLOTT, W., W. LAUER & A. PLACKE (1996): Glo-bal distribution of species diversity in vascularplants: towards a global world map of phyto-diversity. – Erdkunde 50: 317–327.
BARTHLOTT, W., S. POREMBSKI, M. KLUGE, J. HOPKE & L.SCHMIDT (1997): Selenicereus wittii (Cacta-ceae): an epiphyte adapted to AmazonianIgapo inundation forests. – Pl. Syst. Evol. 206:175–185.
BARTHLOTT, W., J. MUTKE, M. D. RAFIQPOOR, G. KIER &H. KREFT (2005): Global centres of vascularplant diversity. – Nova Acta Leop. 92(342):61–83.
BARTHLOTT, W., W. R. ERDELEN & M. D. RAFIQPOOR
(2014): Biodiversity and technical innovations:bionics. – In: D. LANZERATH & M. FRIELE: Conceptand value in biodiversity. Routledge Studies inBiodiversity Politics and Management: 300–315. New York: Routledge.
BAUER, R. (2003): A synopsis of the tribe Hyloce-reeae F.Buxb. – Cactaceae Syst. Init. 17: 3–63.
BENSON, L. (1982): The cacti of the United Statesand Canada. – Stanford: Stanford UniversityPress.
BRAILOVSKY SIGNORET, D. & H. M. HERNÁNDEZ (2010):Mazapil, Zacatecas: diversity and conserva-tion of cacti in a poorly-known arid region innorthern Mexico. – Cact. Succ. J. (Los Ange-les) 82: 197–202.
BRAKO, L. & J. L. ZARUCCHI (1993): Catalogue of theflowering plants and gymnosperms of Peru. –St. Louis: Missouri Botanical Garden.
BRAUN, P. J. & E. ESTEVES PEREIRA (2001): Die Kak-teen Brasiliens mit Anmerkungen zu anderenSukkulenten und xeromorphen Bromelien. –Schumannia 3: 6–235.
BRAUN, P. J. & E. ESTEVES PEREIRA (2004): Pilosoce-reus mollispinus – a new species of Cactaceaefrom the state of Goias and comments on ot-her Pilosocereus species from Central Brazil. –Cact. Succ. J. (Los Angeles) 76: 73–82.
BREGMAN, R. (1996): The genus Matucana: bio-logy and systematics of fascinating Peruviancacti. – Rotterdam: Balkema.
BURR, B. & D. SUPTHUT (eds.) (1994): Artenschutzbei Sukkulenten. – Schumannia 1: 1–52.
BUTTERWORTH, C. A. (2006): Resolving “Nyffeler’spuzzle”—the intriguing taxonomic position ofBlossfeldia. – Haseltonia 3: 3–10.
BUTTERWORTH, C. A. & R. S. WALLACE (2005): Mo-lecular phylogenetics of the leafy cactus genusPereskia (Cactaceae). – Syst. Bot. 30: 800–808.
BUTTERWORTH, C. A., J. H. COTA-SÁNCHEZ & R. S. WAL-LACE (2002): Molecular systematics of Tribe Ca-cteae (Cactaceae: Cactoideae): a phylogenybased on rpl16 intron sequence variation. –Syst. Bot. 27: 257–270.
CHARLES, G. (2009): Micranthocereus. – Cact.Succ. J. (Los Angeles) 81: 150–155.
CHÁVEZ, R. J., J. G. HERNÁNDEZ & E. SÁNCHEZ (2007):Documentación de factores de amenaza parala flora cactológica del semidesierto Queret-ano. – Bol. Nakari 18: 89–95.
CHUK, M. (2010): Invasive cacti – a threat to therangelands of Australia, Bourke. – In: E. A. C.WATERS: Proceedings of the 16th Biennial Con-ference of the Australian Rangeland Society. 5pp. Perth: Australian Rangeland Society.
198 © Schumannia 7 2015
8 References/Literatur

Acanthocereus 83, 112Acharagma 83, 112Argentina 16, 36, 37, 80Ariocarpus 69, 70, 72, 83, 112Armatocereus 83, 113Arrojadoa 63, 83, 113Arthrocereus 83, 113Astrophytum 72, 83, 114Austrocactus 83, 114Austrocylindropuntia 49, 82, 98Aylostera = RebutiaAztekium 41, 83, 114Bergerocactus 83, 114biogeographic regions 22biomes 14Blossfeldia 15, 20, 25, 28, 83, 115Blossfeldieae 23, 26Bolivia 36, 37–41, 47, 79Brachycereus 20, 84, 115Brasilicereus 84, 115Brasiliopuntia 82, 99Brazil 36, 37–41, 78Browningia 84, 115–116Caatinga centre 36, 40, 41, 65Cacteae 23, 25, 26, 57, 58, 60Cactoideae 26, 83, 55, 58Calymmanthium 84, 116Canada 71Caribbean 72Carnegiea 72, 73, 84, 116Castellanosia 84, 116Central America 35, 37–41, 76Cephalocereus 84, 117Cereeae 23, 26, 66Cereineae 26Cereus 41, 46, 84, 117–119Chihuahua centre 36, 37–40, 65Chile 41, 81Cipocereus 84, 119Cleistocactus 41, 69, 84, 120–121climate factors 65Coleocephalocereus 15, 84, 122Colombia 13, 76Conservation 68–73Consolea 82, 99Copiapoa 17, 28, 41, 84, 122–123Corryocactineae 26Corryocactus 84, 123–123Corynopuntia 82, 100Coryphantha 84, 124–126Cuba 19, 49, 72Cumarinia 84, 126Cumulopuntia 16, 82, 101
Cylindropuntia 18, 82, 101–103Cylindropuntieae 23, 26, 59Dendrocereus 19, 70, 84, 127Denmoza 84, 127Discocactus 41, 46, 63, 84, 127–128Disocactus 45, 63, 84, 128–129Echinocactus 20, 60, 84, 129Echinocereinae 26Echinocereus 21, 85, 130–134Echinopsis 16, 62, 69, 85, 135–138Ecuador 77Epiphyllum 62, 85, 138–139Epithelantha 85, 139Eriosyce 41, 85, 139–140Escobaria 85, 141Escontria 85, 142Espostoa 41, 85, 142Espostoopsis 85, 142Eulychnia 85, 143Facheiroa 85, 143Ferocactus 16, 85, 143–145Frailea 85, 145–146Geohintonia 41, 46, 85, 146growth form 44–45, 47, 49, 56Grusonia 82, 104Guyanas 77Gymnocalycium 85, 147–150Haageocereus 41, 85, 150Harrisia 85, 151Hatiora 85, 151Hylocereinae 25, 26Hylocereus 86, 151–152Jalisco centre 36, 37, 40, 41, 65Jasminocereus 86, 152Lasiocereus 86, 152latitudinal gradient 64Leocereus 86, 153Lepismium 86, 154Leptocereus 86, 154Leuchtenbergia 86, 155Lobivia = EchinopsisLophophora 86, 155Lymanbensonia 29, 86, 155Lymanbensonieae 23, 25, 26Maihuenia 25, 45, 82, 98Maihuenieae 23Maihuenioideae 25, 26, 82Maihueniopsis 82, 104Mammillaria 41, 62, 68, 70, 72, 86, 156–162Mammilloydia 86, 162MataAtlantica centre 36, 40, 41, 65Matucana 41, 62, 86, 163Melocactus 68, 70, 86, 163–165
Mexico 36, 37–40, 74Micranthocereus 46, 86, 165Mila 86, 166Miqueliopuntia 82, 104Myrtillocactus 86, 166Neobuxbaumia 86, 167Neolloydia 86, 167Neoraimondia 86, 167Neowerdermannia 86, 168Nopalea 82, 104Notocacteae 23, 25, 26, 61Notocactus = ParodiaObregonia 87, 168Opuntia 19, 48, 69, 72, 82, 105–109Opuntieae 23, 26, 59Opuntioideae 15, 20, 26, 31–33, 55, 82Oreocereus 87, 168Oroya 87, 169Ortegocactus 87, 169Pachycereeae 25, 26Pachycereus 18, 87, 169–170Paraguay 47, 49, 80Parodia 41, 69, 87, 170–173Pediocactus 70, 72, 86, 174Pelecyphora 46, 87, 174Peniocereus 87, 174–175Pereskia 13, 20, 25, 82, 97–98Pereskieae 23Pereskioideae 13, 20, 24, 26, 55, 82Pereskiopsis 83, 110Peru 41, 78Pfeiffera 87, 176Phyllocacteae 15, 23, 25, 26, 60Pierrebraunia 87, 176Pilosocereus 70, 87, 176–179Polaskia 87, 179pollination 47–50, 51, 57Praecereus 87, 179Pseudoacanthocereus 87, 180Pseudorhipsalis 87, 180Pterocactus 83, 110Puebla-Oaxaca centre 36, 37, 40, 41, 65Punotia 83, 110Pygmaeocereus 87, 181Quiabentia 83, 111range size 41–44Rauhocereus 87, 181Rebutia 41, 87, 181–183Rebutiinae 26Rhipsalideae 10, 23, 26, 30, 61Rhipsalidopsis 88, 184Rhipsalinae 30Rhipsalis 19, 20, 41, 48, 68, 88, 184–187
202 © Schumannia 7 2015
Index
The index covers genera, tribes, subfamilies, important synonyms, the country names of the ‘country profiles’ and some key words.




















