
Biodegradation of chlorinated biphenyls (Fenclor 42) in batch cultures with mixed and pure aerobic...
12
Chemosphere, gol. 22, Nos 1-2, pp. 3-14, 1991 (X)45~535/91 $3.00 + 0.00 Printed in Great Britain Pergamon Press plc BIODEGRADATION OF CHLORINATED BIPHENYLS (FENCLOR 42) IN BATCH CULTURES WITH MIXED AND PURE AEROBIC CULTURES. 1" Fabio Fava , Sergio Zappoli 2, Leonardo Marchetti 1, Luciano Morselli 2. (1) Department of Applied Chemistry and Materials Science, University of Bologna, Viale Risorgimento 2, 40136 Bologna, Italy. (2) Department of lndustrial Chemistry and Materials, University of Bologna, Viale Risorgimento 4, 40136 Bolo- gna, Italy. ABSTRACT Two consortia of bacterial strains belonging to the genus Pseudomonas, and a pure strain of Pseudomonas cepacia were isolated from river sediments by using biphenyl and 4- chlorobiphenyl as a sole carbon source. Metabolic de- gradation of various polychlorinated biphenyls (PCBs) by these organisms was studied. The biodegradation pro- ducts of 4-chlorobiphenyl were analyzed in Pseudomonas. Several low chlorine PCB congeners of Fenclor 42 mixture can be easily degraded by these mixed and pure cultures, especially in growing cell systems. Chlorine was not produced during the degradation of all the chlorobiphenyls tested, including 4-chlorobiphenyl. INTRODUCTION Mixtures of polychlorinated biphenyls (PCBs) have been extensively used for many years in a wide variety of industrial applications, such as dielecNc fluids, solvents and plasticizers (1). Although the use of these substan- ces has been greatly curtailed in most parts of the world, considerable quantities of PCBs still affect environmental conditions. Because of their hydrophobic nature, PCBs have accumulated primarily in soil and aquatic sediments, where they are strongly adsorbed by organic matter (2). Many bacterial strains capable of degrading low chlorina- ted biphenyls have already been described in the literature (3-11). Ahmed and Focht (3) postulated that the degra- dation of low-chlorine-content PCBs in two species of Achromobacter proceeds via the formation of a 2,3-dihydroxy derivative which is open in a meta-l,2 position to ultimately generate the corresponding chloroben- zoate. Furukawa et al. (5, 6, 7) showed that single, pure congeners of commercial Aroclor 1242, 1254 mixtures
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Biodegradation of chlorinated biphenyls (Fenclor 42) in batch cultures with mixed and pure aerobic...

Chemosphere, gol. 22, Nos 1-2, pp. 3-14, 1991 (X)45~535/91 $3.00 + 0.00 Printed in Great Britain Pergamon Press plc
BIODEGRADATION OF CHLORINATED BIPHENYLS (FENCLOR 42) IN BATCH
CULTURES WITH MIXED AND PURE AEROBIC CULTURES.
1" Fabio Fava , Sergio Zappoli 2, Leonardo Marchetti 1, Luciano Morselli 2.
(1) Department of Applied Chemistry and Materials Science, University of Bologna, Viale Risorgimento 2, 40136 Bologna, Italy. (2) Department of lndustrial Chemistry and Materials, University of Bologna, Viale Risorgimento 4, 40136 Bolo- gna, Italy.
ABSTRACT
Two consortia of bacterial strains belonging to the genus Pseudomonas, and a pure strain of Pseudomonas cepacia were isolated from river sediments by using biphenyl and 4- chlorobiphenyl as a sole carbon source. Metabolic de- gradation of various polychlorinated biphenyls (PCBs) by these organisms was studied. The biodegradation pro- ducts of 4-chlorobiphenyl were analyzed in Pseudomonas. Several low chlorine PCB congeners of Fenclor 42 mixture can be easily degraded by these mixed and pure cultures, especially in growing cell systems. Chlorine was not produced during the degradation of all the chlorobiphenyls tested, including 4-chlorobiphenyl.
INTRODUCTION
Mixtures of polychlorinated biphenyls (PCBs) have been extensively used for many years in a wide variety
of industrial applications, such as dielecNc fluids, solvents and plasticizers (1). Although the use of these substan-
ces has been greatly curtailed in most parts of the world, considerable quantities of PCBs still affect environmental
conditions. Because of their hydrophobic nature, PCBs have accumulated primarily in soil and aquatic sediments,
where they are strongly adsorbed by organic matter (2). Many bacterial strains capable of degrading low chlorina-
ted biphenyls have already been described in the literature (3-11). Ahmed and Focht (3) postulated that the degra-
dation of low-chlorine-content PCBs in two species of Achromobacter proceeds via the formation of a
2,3-dihydroxy derivative which is open in a meta-l,2 position to ultimately generate the corresponding chloroben-
zoate. Furukawa et al. (5, 6, 7) showed that single, pure congeners of commercial Aroclor 1242, 1254 mixtures

and of different commercial Kanaclors, may be degraded at a molar percentage of 15% by Acinetobacter sp. in re-
sting cells conditions. Bedard et al. classified a contaminant of Furukawa's strain as Corynebacterium sp., and
found it to be less active towards other strains with respect to the transformation of PCBs. Moreover, Bedard et
al.(8, 9, 10) described a new type of 3,4 dioxygenase attack on PCB congeners by Alcaligenes eutrophus H850, in
contrast to the more common 2,3-dioxygenase attack originally shown by Ahmed and Focht (3) and successfully
observed by Furukawa et al. The degrading capability varies substantially from species to species. Complete mine-
ralization of monochlorinated biphenyls by single strains has been reported (12, 13). But up to now no culture ca-
pable of completely mineralizing more highly chlorinated biphenyls, or of growing on congeners having more than
one chlorine has been unequivocally described. Research by Kong and Sayler (14) has shown the existence of sui-
table mixed cultures which are capable of completely degrading 4-chloro biphenyl into carbon dioxide, water, and
chlorine ions (14).
Procaryotic organisms were selected in order to degrade the different congeners into related alogenated ben-
zoic acid and linear halogenated carboxylic acids. The genetic approach has also been recently tested. Furukawa et
al. (15) have cloned and sequenced an entire gene from P.putida which encodes degradation of biphenyl and poly-
chlorinated biphenyls into benzoate and chlorobenzoates.
Some key questions provided the framework for our experiments: was it possible to find other bacteria with a
degrading competence equal or superior to that of the strains studied until now ? How can resting and growing
cells be compared with regards the transformation of Fenclor 42 ? What is the efficacy of Fenclor 42 transforma-
tion by pure and mixed cultures, respectively? What are the specific degrading pathways adopted by pure strains?
MATERIALS AND METHODS
Isolation and culture methods.
All mixed and pure cultures were selected from river PCB- contaminated sediments after several cycles of
enriching growth. Approximately 5 g of soil were incubated in 50 mL of the culture medium. Each trial employed
a medium made up of phosphate- buffered mineral salts (PAS), containing biphenyl as the carbon source, which
in some cases was supplemented with 0.02% yeast extract; PAS was prepared by adding 77.5 mL of concentrated
phosphate buffer (PA) to 910 mL of distilled water (medium follows Bedard et al.(8)). After autoclaving and coo-
ling, 10 mL of sterile PAS 100 X salts were added. Biphenyl was supplied by addition of 2 mL of a 50% stock so-
lution in acetone (the final biphenyl concentration was 0.1% ) to the autoclaved medium; this concentration of
acetone did not affect the growth curve, as compared to a control condition where the biphenyl was directly added
to the broth medium. PA concentrate was composed of K2HPO4 (56.77 g/L), KH2PO4 (21.94 g/L), and NH4C1
(27.61 g/L). PAS 100 X salts contained MgSO4 (19.5 g/L), MnSO4xH20 (5g/L), FeSO4x7H20 (lg/L), and
CaC12x2H20 (0.3 g/L), together with several drops of concentrated H2SO4 per liter to avoid precipitation of basic
salts. When necessary, this medium was solidified with 2% purified agar (Biolife, Italy). During growth in the
medium containing 1 g/L of biphenyl, all cultures were maintained aerobically in the dark at 28°C in a gyratory

shaker at 150 rpm. After five cultural cycles in this medium, four mixed cultures were obtained. In order to assess
their ability to degrade PCBs, these mixed cultures were tested on 4 chloro-biphenyl (4CB), which is a good mo-
lecular model for our study. Among these active mixed cultures, some pure cultures were selected by means of the
simple Cook strategy (16), that is by streaking the growth enrichment culture to single colonies on solid, non-se-
lective medium (Medium TSA: trypticase 1.5%, Phytone 0.5%, NaC1 0.5%, Agar 1.5%). A specimen of each type
of colony was then transferred to a sterile selective liquid medium. The liquid cultures showing growth were strea-
ked again on a solid, non selective medium (TSA). After three cycles of alternating growth on non-selective plates
and in selective liquid medium, attainment of the desired degree of purity was demonstrated by the homogeneous
morphology of colonies and cells. All the pure cultures thus isolated were examined by rapid screening in order to
evaluate 4CB degrading capability. Single pure cultures grew on PAS-agar plates containing 0.1% of biphenyl as
the only carbon source. After three days of incubation at 28°C, the developed colonies were sprayed with a 4%
(wt/vol) 4CB solution in hexane in order to deposit a film of lipophilic 4CB on the surface of the plates. The plates
were immediately incubated at 28°C for 48 hrs; positive colonies were recognized as those surrounded by a clear,
yellow zone in the 4CB film. This concentration of hexane did not affect the cometabolic enzymatic attack. The
ability of these isolates to grow on 4CB as the only carbon source in liquid PAS medium was also tested. The abili-
ty of degrading both 4CB and Fenclor 42 was tested in resting cell and growing cell assays.
Microorganisms.
Of the strains with PCB degrading capability, the most effective, CP9, was shown to be generally motile,
gram negative, catalase and oxidase positive and strictly aerobic, and to have double or single rods, no production
of water-soluble pigment, no nitxate production, nitrite reduction capability as well as no agar hydrolysis, and to
grow in mineral medium with biphenyl and benzoic acid. Colonies grown on biphenyl-mineral agar were circu-
lar, smooth, courvex, light yellow and traslucent. The results suggest that these organisms belong to the Pseudomo-
na__ss genus ( Bergey's Manual of Determinative Bacteriology). The exact species identification (Pseudomonas
cepacia) was carried out by the API system 20, specific for no-Enterobacteriaceae.
Preparation of Resting Cells.
After overnight growth of cells on biphenyl, the crystals of biphenyl were removed by filtration through glass
wool. After harvesting by centrifugation at 7000 rpm for 15 minutes, the cells were washed twice in a 0.05M so-
dium phosphate buffer (pH 7.5), and resuspended in the same buffer in order to obtain an optical density of 1.0 at
615 nm. Fenclor 42 (25 ppm) and the mixture of PCB congeners (2 ppm for each congeners) were then added in
the form of concentrated acetone solutions. Incubation was carried out in 25 mL bottles, which were sealed with
screw teflon caps. These bottles were incubated for two days on a rotary platform shaker (150 rpm) at 28°C. Con-
trois were autoclaved at 110°C for 10 minutes before being added to the solution of PCBs being analyzed.
Assays on growing cells.
For assays of growing cells, acetonic solution of biphenyl and acetonic solutions of Fenclor 42 or of PCB

mixtures were added, at the same concentration as be-
fore, to 50 mL of PAS medium in 250 Erlenmeyer
flasks. The flasks were fitted with a specific chemical
trap filled with macroreticular resin, XAD-2, as shown
in figure 1. After inoculation (1-5 % vol/vol), the
flasks were incubated for several days (5 days for
4CB degradation studies and 15 days for Fenclor 42
degradation studies) at 28°C on a rotary shaker at 150
rpm. Controls were prepared by heat inactivation
(110°C for 10 minutes) of cell suspensions prior to in-
cubation with PCBs or 4CB. All inoculated cultural
media were placed in two different glass containers.
These containers were then wrapped in aluminum foil
to protect them from light and finally sealed with te-
flon foil.
FIG.1 Growing cell culture apparatus: Erlenmeyer flask with amberlite XAD-2 sterile chemical trap.
PCB extraction.
After incubation, cell growth of each assay was stopped either by refrigeration of single cultures or by addi-
tion of perchloric acid (final concentration 0.7%). Each cultural broth was extracted three times with 10 mL of a
hexane-acetone mixture (2:1). In order to reduce the loss of PCBs due to evaporation or absorption into the vessel,
extraction procedures were carried out in the same incubation vessel previously silanized and sealed with alumi-
num-lined or teflon caps. In the case of growing cell assays, extraction was carried out as follows: 20 mL of hexa-
ne-acetone (2:1) were first eluted through the XAD-2 column into the Erlenmeyer flask, which was then left in an
ultrasonic bath for ten minutes. After extraction the liquid was transferred into a separatory funnel. The aqueous la-
yer was then collected in the flask and the above procedure was repeated twice while all the organic layers were
anhydrified with Na2SO4 and collected in a 100 mL volumetric flask.
In order to check the recovery of the extraction procedure, which could have been affected by PCB adsor-
ption on cells, a stronger extraction test was carried out. After centrifugation of the exhausted aqueous layer, the
cell portion was extracted with 80% hot ethanol (80°C) for 20 minutes. The cell suspension was then centrifuged to
remove an 80% ethanol layer from the cells. The cells were once again extracted with 20% ethanol (20°C) for 20
minutes and then centrifuged so as to obtain a 20% ethanol layer. No significant concentration of PCBs was reco-
vered in these ethanol cell-free extracted fractions. The recovery of PCB from broth by means of ultrasonic extrac-
tion was about 95%.
GC analysis.
The samples collected after anhydrification were analyzed by capillary gas chromatography using an elec-

TABLE 1. Assignement of congeners to chromatographic peak and their re- tention time relative to octachloro naphtalene.
Retention time lupac No Substitution pattern relative to OCN (Ref.20)
0.4390 3 4 0.4634 9 2,5 0.4879 5 2,3 0.5365 15 4,4' 0.5366 17 2,2',4 0.5367 16,32 2,2',3; 2,4',6 0.5609 26 2,3 ',5 0.5610 25 2,3',4 0.5611 31,28 2,4',5; 2,4,4' 0.5853 33 2',3,4 0.5854 22 2,3,4' 0.5854 45,46 2,2',3,6; 2,2',3,6' 0.5855 52 2,2',5,5' 0.6097 49 2,2',4,5' 0.6098 44 2,2',3,5' 0.6099 37,42,59 3,4,4'; 2,2',3,4'; 2,3,3',6 0.6341 41,64,71 2,2',3'4; 2,3,4',6; 2,3',4',6 0.6342 40 2,2',3,3' 0.6584 74 2,4,4',5 0.6585 70 2,3 ',4',5 0.6586 88 2,2',3,4,6 0.6828 56,60 2,3,3 ',4'; 2,3,4,4' 0.6829 101 2,2'4,5,5' 0.6830 99 2,2',4,4',5 0.6831 97 2,2',3' ,4,5 0.7073 87 2,2',3,4,5' 0.7074 110 2,3,3,'4,4' 0.7317 82 2,2',3,3 '4 0.7318 108,118,143 2,3,3,'4,5'; 2,3',4,4',5; 2,2',3,4,5, 0.7561 146 2,2',3,4',5,5' 0.7562 153 2,2',4,4',5,5,' 0.7805 138 2,2,',3,4,4',5' 0.7806 178 2,2,' ,3,3',5,5',6 0.8536 180 2,2',3,4,4',5,5' 0.8782 170 2,2,',3,3',4,4',5
tron capture detector (ECD). All
samples were run on a Nordion Mi-
cromat HRGC Mod 471 equipped
with a 50 m x 0.25 mm I.D. fused si-
lica column (SE 54 type phase). Test
runs were conducted under the follo-
wing conditions: initial temperature,
40°C; isothermal for 2 min; first tem-
perature rate 5°C/rain; final tempera-
ture 280°C; isothermal for 16 rain;
injector (splitless mode) 240°C; ECD
320°C; argon carrier gas flow-rate, 2
mL/min; sample size 2 laL/column.
Peak integration was carried out with
the Baseline package by Dynamic
Solution. As described by Morselli et
al. (18), the identification of single
PCB congeners was performed using
congeners standards when available,
or by comparing the data reported in
previous studies (19,20) and the re-
tention times relative to octachloro-
naphtalene (OCN) calculated by
Mullin et al (21). Amount of degra-
dation was determined by using the
peak intensity of congener 56 (see ta-
bIe 1) which is not subject to degradation by the bacteria used in our experiment. By using this method it is thus
possible to overcome many of the problems arising from the recovery of analytes from bacterial cultures. Table 1
lists the identified congeners with the corresponding Iupac No. (as per Ballschmiter and Zell (20)) and the chlorine
substitution pattern.
Spectrophotometric measurements.
A Perkin Elmer 402 UV-visible spectrophotometer was used for all UV-visibile measurements. Infrared
spectra were obtained on KBr pellets of purified compound by using a Bmker IFS 48 IR spectrophotometer.
Chemicals.
2-chlorobiphenyl (2CB), 3-chlorobiphenyl (3CB), 4CB and all PCB congeners used for the specific mixtures

were supplied by Ultra Scientific, North Kingstown (RI), while Fenclor 42 (a laboratory sample of an old commer-
cial mixture containing primarily tetra- and pentha-chlorobiphenyls) was kindly provided by Caffaro (Italy). Biphe-
nyl and all chlorobenzoic acid isomers were obtained from Merck (Germany). Pesticide grade hexane, methanol
and acetone were purchased from Baker (Holland).
Amberlite XAD-2 resin was supplied by Carlo Erba (Italy).
RESULTS
The two mixed cultures grew readily on the biphenyl added to the PAS minimal medium as the sole carbon
source. MIX 1, obtained after 5 cycles of enrichement in PAS supplemented with biphenyl and 0.02% yeast ex-
tract, grew quite well on 4CB, but did not grow on 2CB and 3CB. MIX 3, selected without yeast extract, grew
well on biphenyl but not on 2CB, 3CB, 4CB. Among the pure cultures isolated, only one, CP15, grew quite well on
4CB. The other pure cultures were only able to grow on biphenyl substrates, even if they could cometabolize
4CB. The ability of these cultures to utilize higher chlorinated PCB congeners as growth substrates was not tested.
All pure cultures obtained from the mixed ones were screened by means of a rapid spray test in order to determine
their ability to cometabolize 4CB. In order to evaluate microbial ability to degrade the 4CB, mixed and pure cul-
tures were tested. After 5 days of incubation, mixed cultures degraded about 75% of the initial 1000 ppm of 4CB
added to the medium. After the same incubation period and under similar conditions, pure cultures degraded 80-
85% of the initial 4CB. CP15, which uses this molecule as a carbon source, degraded 95% of the initial 4CB. 4-
chlorobenzoic acid (4CBA) was extracted, purified and identified from all broths as the last stable metabolite of the
C 0 N C E N T R A T I O N
250
4-CB (ppm°lO*-l) OFU'lO*6/rnl
200 ~ j ~ ' - , ~ - 4-OBA (ppm'lO*-l)
0 20 40 60 80 100 120 140 INCUBATION PERIOD (HOURS)
160
FIG.2 Degradation of 4CB, production of 4CBA and cellular growth in CP9 growing cell batch culture.
degradation pathway by means of IR
spectroscopy. An intermediate metabo-
lite, which appeared quite late in the
growth medium, consisted of a yellow-
colored compound with an absorbance
peak at 400 nm. With time, the spectral
peak was seen to shift to 430 nm and
the medium color changed to bright yel-
low. After further incubation, the inter-
mediate metabolite with absorbance
peak at 430 nm disappeared, and a new
intermediate metabolite with a UV ab-
sorbance peak at 230-235 nm (typical of
4CBA) accumulated. As reported by
other Authors (4), the color and absor-
ption peak at 430 nm was found to be
removed by acidification, and a new pe-
ak at 320-330 nm was formed. The kinetic pattern of 4CB cometabolization by Pseudomonas cepacia (CP9 strain)
on PAS mineral medium supplemented with yeast extract (0.02%) and biphenyl (0.1%) was also studied.
Figure 2 shows both the related 4-chlorobenzoic acid formation and the rate of cellular growth in relation to
time for the CP9 strain.
D e g r a d a t i o n o f F e n c l o r 42.
Each pure and mixed culture was incubated for 40 days on Fenclor 42 in batch culture. Subsequently, the abi-
lity of both mixed and pure cultures to degrade the commercial mixtures of PCBs in resting-cell and growing-cell
conditions was assayed. Degradation was carried out on a 25 ppm acetonic solution of Fenclor 42 for the growing
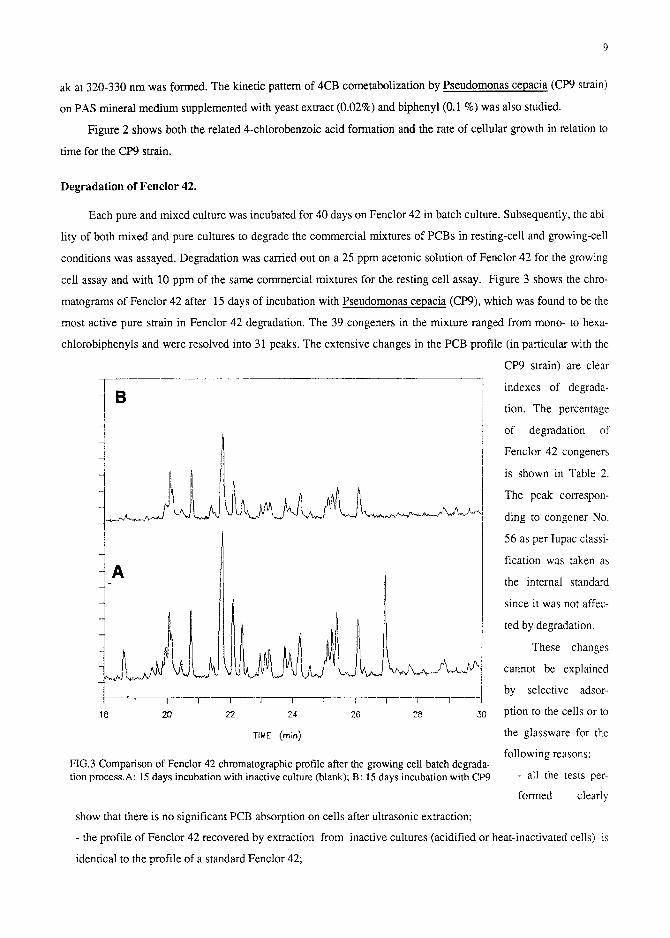
cell assay and with 10 ppm of the same commercial mixtures for the resting cell assay. Figure 3 shows the chro-
matograms of Fenclor 42 after 15 days of incubation with Pseudomonas cepacia (CP9), which was found to be the
most active pure strain in Fenclor 42 degradation. The 39 congeners in the mixture ranged from mono- to hexa-
chlorobiphenyls and were resolved into 31 peaks. The extensive changes in the PCB profile (in particular with the
B
A
18 20 22 24 26 28 30
"rIME (mln)
FIG.3 Comparison of Fenclor 42 chromatographic profile after the growing cell batch degrada- tion process.A: 15 days incubation with inactive culture (blank); B: 15 days incubation with CP9
CP9 strain) are clear
indexes of degrada-
tion. The percentage
of degradation of
Fenclor 42 congeners
is shown in Table 2.
The peak correspon-
ding to congener No.
56 as per Iupac classi-
fication was taken as
the internal standard
since it was not affec-
ted by degradation.
These changes
cannot be explained
by selective adsor-
ption to the cells or to
the glassware for the
following reasons:
- all the tests per-
formed clearly
show that there is no significant PCB absorption on cells after ultrasonic extraction;
- the profile of Fenclor 42 recovered by extraction from inactive cultures (acidified or heat-inactivated cells) is
identical to the profile of a standard Fenclor 42;
10
- each unmodified peak has the same area as that of the related peak of the control culture.
In resting cell conditions, a technique preferred by most researchers, no significant degradation of any
PCB congeners was observed after three or more days of incubation. In addition, the degradation results were
usually not reproducible especially for low chlorinated congeners, which are the easiest to degrade. However, the
results were found to range within the experimental error percentage which is generally admitted for these experi-
mental conditions.
Degradation of a synthetic mixture of PCB congeners.
Following on the above findings, an assay was attempted by using a synthetic mixture of eight pure PCB
congeners. The purpose of this experiment was to study the specific degradation pathways of our active pure cultu-
res as well as to compensate for the risk of obscure results which can occur owing to frequently incomplete gas
chromatographic resolution and to the uncertain identification of the Fenclor peaks. Congeners were thus selected
for our defined mixtures to satisfy the following criteria:
- congeners preferably present in commercial mixtures of PCBs;
- congeners capable of being completely resolved by capillary column GC;
- congeners representing several different structural classes of PCB s.
Our assay mixture included eight PCBs congeners ranging from dichloro- to penthachlorobiphenyls, repre-
senting various structural classes such as:
- congeners chlorinated on a single ring (2,5 dichlorobiphenyl);
- congeners blocked at 2-3 sites to prevent 2,3-dioxygenase attack (2,2',5,6'-tetrachlorobiphenyl, 2,2',4,5,5'-
TABLE 2. Percentage of degradation of PCB congeners afterl5 days of incubation with mixed cultures and CP-9 strain in growing cells condition.
Iupac DEG% DEG % DEG % Iupac DEG % DEG % DEG°k number MIX 1 MIX 3 CP-9 number MIX 1 MIX 3 CP-9
3 100 I00 99 9 100 100 53 5 100 100 100 15 9 0 62 17 20 17 44
16/32 15 7 44 26 I0 9 46 25 90 90 59
31/28 45 42 48 33 74 69 64 22 100 72 72
45/46 9 0 32 52 12 15 " 49 49 7 15 41 44 15 35 44
37/59/42 16 100 34
71/41/64 14 17 41 40 22 100 33 74 16 20 37 70 15 22 39 88 17 23 41
56(*) IS IS IS 101 0 0 60 87 0 0 32 110 0 0 55 82 0 0 0
108/118 0 0 0 146 0 0 0 178 0 0 0 180 0 0 0 170 0 0 0
TOTAL 18 76 50
(*) congener 56 was used as internal standard.

11
penthachloro biphenyl);
- congeners blocked at 3-4 sites inaccessible to the 3,4- dioxygenase (2,3',4,4'-tetrachlorobiphenyl, 2,4,4',6-te-
trachlorobiphenyl, 4,4'- dichlorobiphenyl);
- congeners usually not degraded by PCB degrading bacteria; these congeners were taken as internal stand-
ards (2,2',6,6'- tetrachlorobiphenyl, 3,3',5,5'-tetrachlorobiphenyl).
The study was carried out in growing cell conditions on the P. cepacia strain (CP9) which had been isolated
by us.
Each congener was added to the medium at a concentration of 2 and 3 ppm following congener chlorination.
Table 3 shows the percentage of PCB-congener degradation of the synthetic mixture after incubation for 10 days in
CP9 growing cell conditions. The two dichlorobiphe-
nyls congeners, Nos. 9 and 15, were completely de-
graded, whereas the heaviest chlorinated congeners
showed degrees of degradation almost tending to zero.
The only noticeable exception was the 29% degrada-
tion of congener No. 66. However, care should be ta-
ken in measuring the absolute rates of degradation. In
fact, as there is evidence that the degradation of any
congener in a mixture may be influenced by the other
congeners contained in the same commercial or syn-
thetic mixture, absolute rates of degradation should be
determined with individual congeners in the presence
of an appropriate internal standard.
TABLE 3. Degradation percentage of selected PCB congeners after 15 days of incubation with CP-9 strain.
Iupac Substitution Degradation % No pattern CP-9
9 2,5 100 15 4,4' 100 54 2,2',6,6' 4 53 2,2' ,5,6' 8 75 2,4,4',6 7 66 2,3',4,4' 29 80 3,3',5,5' 3
101 2,2',4,5,5' 0
DISCUSSION
Improved growing cell assays with strictly aerobic strains for degradation studies were made possible thanks
to the special equipment available in our laboratory. In particular, the vertical XAD-2 silanized trap, which can be
completely sealed with aluminium foil, allows constant sterile aeration of shaken cultures while avoiding any pos-
sible loss of the PCBs added to the medium. Consequently, after incubation and before extraction of the cultural
broth, all PCBs reversibly adsorbed onto the resin were eluted into the assay silanized vessels by subsequent addi-
tions of the extraction mixture. The solution thus obtained prevents any false-positive results in degradation stu-
dies due to volatilization, adsorption or absorption losses. A species of Pseudomonas (P. cepacia CP9 strain)
capable of degrading some low chlorinated members of the PCB family, was found. The 4CB was easily decom-
posed into 4-chlorobenzoic acid (4CBA) by both the mixed and pure culture microorganisms employed. Yellow
compounds were found to be the intermediate metabolites leading to the production of 4CBA. The kinetic of de-
gradation, however, was so fast that the intermediate metabolites could not be isolated during incubation. The yel-

12
low pigment was probably 2-hydroxy-6-oxo-6-(4-chlorophenyl)-hexa-2,4-dienoic acid as already reported by Fum-
kawa et a1.(4,6). In addition to the degrading ability of pure culture, that of mixed cultures was also studied as these
were judged to be more representative of the conditions actually existing in nature. Pure culture, however, was
found to have a higher degradation capability on 4CB and on Fenclor 42 than mixed cultures, both in resting cell
and in growing cell systems. Fenclor 42, a commercial mixture which had never been tested before, was employed
for these experiments.
Until now, the resting cell system has not given any satisfactory results: in fact, in a resting cell assay with
cells harvested at the end of the logarithmic growth phase, both mixed and pure cultures degraded an insignificant
percentage of Fenclor congeners. Moreover, different cell densities in the adopted range had no significant influen-
ce on these results. It is still open to question whether growing or resting cell conditions may be the better assay
condition to test PCB biodegradation (11). Contrasting with the results obtained by Furukawa and Bedard under re-
sting cell conditions, Kohler et al. showed that Acinetobacter sp. P6 strain and Arthrobacter sp. B 1B strain were ca-
pable of better degradation in growing cell condition. In this case, PCB degradation of Aroclor 1254 was more
intensive as well as extensive (32 and 23 %) than the degradation obtained in resting cell conditions (respectively
17 and 8%). The study conducted by us was aimed at comparing degradation results under growing and resting
cell conditions. Growing cell assays provided more reliable results, while resting cell conditions gave both uncon-
stant and only slightly significant results. It should be pointed out that for our cultures we did not check the optimi-
zation of the resting cell methodology used by other researchers. Better results, therefore, may be obtained even if
in our opinion it can be safely predicted that the percentage of degradation will always be lower in resting cell
conditions as compared to growing cell conditions. In fact, the good results obtained in growing cell assays with
both pure and mixed cultures show that growing cells are more active than resting cells with respect to transforma-
tion of PCBs. In growing cell assays Pseudomonas cepacia always outperformed MIX 1 and MIX 3 in their ability
to transform Fenclor 42 . More specifically, mixed cultures transformed some congeners to a greater extent than
what the CP9 strain was capable of doing, whereas the P. cepacia pure strain transformed congeners that were not
attacked by MIX 1 and MIX 3 cultures. It is worth noting that MIX 3 culture degrades heavy chlorinated conge-
ners more extensively than what MIX 1 and the CP9 strain do. In fact, it is possible that other species of microorga-
nisms, present in the MIX 3 culture but not involved in the degradation pathway, could be responsible for
facilitating the global process by removing potentially inhibitory intermediates.
The results of degradation of synthetic mixtures given above furnish information obtainable from defined
congener assays. It has been postulated that the initial enzyme attack is 2,3-dioxygenase, and that subsequently me-
ta-cleavage occurs between carbon atoms C-1 and C-2 (3,5). Theoretically, a dioxygenase could also attack posi-
tion 3,4: as a matter of fact, previous studies have shown that several microorganisms with degrading ability
follow the 3,4 pathway (9). The ability of the CP9 strain to metabolize 4,4'-dichlorobiphenyl and 2,3',4,4'-tetra-
chlorobiphenyl clearly indicates that this strain only attacks congeners with open 2,3 sites (see Table 3).
It can thus be hypothesized that this strain probably uses a 2,3-dioxygenase as is shown in the more common
metabolic sequence illustrated in figure 4. The 2,5-dichlorobiphenyl that we included as positive degradation inter-
nal standard was readily degraded. Our results also tend to confirm that congeners with heavy orto-substitution

13
Ol Ol Cl Cl Cl
Ol Ol OI OI
FIG.4 Proposed pathway for degradation of PCBs by a 2,3-dioxygenase (3,6).
(2,2',6,6'-tetrachlorobiphenyl) are the most resistant to attack by aerobic bacteria, independently of the availability
of other sites for attack. In addition, it was found that 3,5,3',5'-tetrachlorobiphenyl, unavailable for 2,3 and 3,4
enzymatic attacks, was resistant to degradation by P. cepacia (CP9). The data obtained from the degradation of
Fenclor 42 and of the synthetic mixture of PCB congeners are in line with the results reported by Furukawa with
regards the following points: a decrease of degradation occurs as chlorine substitution increases; PCBs containing
two chlorine atoms in the orto positions of a single ring (i.e. 2,6-) and/or in each ring (i.e. 2,2'-) show a strong resi-
stance to degradation; PCBs containing all chlorine atoms on a single ring are generally degraded faster than those
containing the same number of chlorine on both rings; preferential ring cleavage of molecules occurs at non-chlori-
nated or less chlorinated ring. On the basis of these observations, it can be concluded that the lower percentage of
2,3',4,4'-tetrachlorobiphenyl degradation compared to that of 4,4'- dichlorobiphenyl may be due to the relatively
high chlorination of the latter.
ACKNOWLEDGMENTS
We thank Prof. Diego Matteuzzi for his helpful suggestions and Mr. P. Luigi Tonetti of Caffaro Itaiy who
kindly supplied us with old laboratory samples of Apirolio R and Fenclor R.
REFERENCES
1) Fishbeing L. Chromatographic and biological aspects of polychlorinated biphenyls. 1972. J. Chromatogr. 68:345-350.
2) Hutzinger O., Safe S. and Zikto V..1974. The chemistry of PCBs. CRC press, Boca Raton, Fla.
3) Ahmed M., D.D Focht. 1973. Degradation of polychlorinated biphenyls by two species of Achromobacter. Can. J. Microbioi.19:49-52.

14
4) Furukawa K. and F. Matsumura. 1976. Microbial metabolism of polychlorinated biphenyls. Studies and the rela- tive degradability of polychlorinated biphenyl components by Alkaligenes sp.. J. Agric. Food Chem.. 24:251-256.
5) Furukawa K., F, Matsumura, and K. Tonomura. 1978 .Alcaligenes and Acinetobacter strains capable of degra- ding polychlorinated biphenyls. Agric. Biol. Chem. 42:543-548.
6) Furukawa K., N. Tomizuka and A. Kamibayashi. 1979. Effect of chlorine substitution on bacterial metabolism of various polychlorinated biphenyls. Appl. Environ. Microbiol. 38:301-310.
7) Furukawa K.,N. Tomizuka and A. Kamibayashi. 1983. Metabolic breakdown of Kanaclors (polychlorobiphe- nyls) and their products by Acinetobacter sp.. Appl. Environ. Microbiol. 46:140-145.
8) Bedard D.L.,R.Unterman, L.H. Bopp, M.J. Brennan, M.L. Haberl and C. Johnson. 1986. Rapid assay for scree~ ning and characterizing microorganisms for the ability to degrade polychlorinated biphenyls. Appl. Environ. Mi- crobiol. 51:761-768.
9) Bedard D.L, M.L. Haberl, R. May and M.J. Brennan. 1987. Evidence for novel mechanisms of polychlorinated biphenyl metabolism in Alcaligenes eutrophus H850. Appl. Environ. Microbiol. 53:1103-1112.
10) Bedard D.L., R.E. Wagner, M.J. Brennan, M.L. Haberl and J.F. Brown. 1987. Extensive degradation of Aro- clots and environmentally transformed polychlorinated biphenyls by Alcaligenes eutroDhus H850. Appl. Environ. Microbiol. 53:1094-1102.
11) Kohler H.P.E., D. Kohler-Staub and D.D. Focht. 1988. Cometabolism of polychlorinated biphenyls: enhanced transformation of Aroclor 1254 by growing bacterial cells. Appl. Environ. Microbiol. 5_._44:1940-1945.
12) Shields, M.S.,S.W. Hopper, and G.S. Sayler.1985. Plasmid mediated mineralization of 4-chlorobiphenyl. J. Bacteriol. 163:882-889.
13) Furukawa K. and A. M. Chakrabarty. 1982. Involvement of plasmids in total degradation of chlorinated biphe- nyls. Appl. Environ. Microbiol. 44:619-626.
14) Kong H.L., G.S. Sayler. 1983. Degradation and total mineralization of monohalogenated biphenyls in natural sediment and mixed bacterial culture. Appl. Environ. Microbiol. 46:666-672.
15) Hayase N., K. Taira and K. Furukawa. 1990. Pseudomonos putida K F 715 bph ABCD operon encoding biphe- nyl and polychlorinated biphenyl degradation: cloning, analysis, and expression in soil bacteria. J. Bact. 172:1160- 1164.
16) Cook A.M., H. Grossenbacher and R. Hutter. 1983. Isolation and cultivation of microbes with biodegradative potential. Experientia. 39:1191-1198.
17) Chakrabarty A.R.. 1982. Genetic mechanisms in the dissimulation of chlorinated compounds, pp 127-139. In A.M. Chakrabarty, (Ed.), Biodegradation and detoxification of environmental pollutants. CRC Press, Inc, Boca Ra- ton.
18) L.Morselli, S.Zappoli, A.Donati. 1989. Identification, quantification and distribution of Polychlorinated biphe- nyls (PCB) in an Antarctic marine environment: Terranova Bay, Ross Sea. Ann. Chim. 79:677-688.
19) P.D.Capel, R.A.Rapaport, S.J. Eisenreich, B.B. Looney. 1985. PCBQ: computerized quantification of total PCB and congeners in environmental samples. Chemosphere 14:439-450.
20) K.Ballschmiter, M.Zell. 1980. Analysis of PCB by glass-capillary GC: composition of technical Aroclor- and Clophen- PCB mixtures. Fresen ius Z. Anal.Chem., 302:20-31.
21) M.D.Mullin, C.M.Pochini., S.MC.Crindle, M.Rornkes, S.H.Safe, L.M.Sa fe. 1984. High resolution PCB analy- sis: synthesis and chromatogra phic properties of all 209 PCB congeners. Env.Sci.Technol.:l_~ 468.
(Received 27 September 1990)




















