
Biochemical characterization of the monoclonal antibody-defined ovarian carcinoma-associated antigen...
8
Int. J. Cancer: 37, 705-712 (1986) 0 1986 Alan R. Liss, Inc. BIOCHEMICAL CHARACTERIZATION OF THE MONOCLONAL ANTIGEN SGA ANTIBODY-DEFINED OVARIAN CARCINOMA-ASSOCIATED Theonne A. DE KRETSER’, Heather J. THORNE, Diana PICONE and David G. JOSE Imrnunogenetics Research Unit. 7he Cancer Institute. 481 Little Lonsdale Street, Melbourne, Victoria 3000, Australia. The molecular nature of SGA, the ovarian-carci- noma-associated antigen defined by the MAb OM-I, has been determined. The cell-surface form of the SGA molecule is a glycoprotein with pl <4.2, which on PAGE analysis has an apparent MW of approxi- mately 360 kDa. This was the only OM-I-reactive spe- cies found on the cell surface. The apparent MW was unaffected by reducing conditions. The predominant cytoplasmic form of SGA is a non-glycosylated 170- kDa molecule with pl 6.5. Pulse-chase experiments were complicated by the extremely slow rate of SGA synthesis. However, the data indicate that the SGA molecule is synthesized as a 190-kDa protein, cleaved to yield a 170-kDa non-glycosylated intracellular form which is slowly glycosylated to the 360-kDa cell-sur- face species. Western blotting experiments revealed the presence of the 360-kDa glycosylated molecule in human ovarian cell culture supernatants. For more than a decade, the development of immu- nological markers for ovarian epithelial cancer has been of interest to those searching for the means to detect, diagnose, monitor and treat this disease. Since the first demonstration of ovarian tumour-associated antigens using heteroantisera (Levi et al., 1969), a number of such antigens have been described. Promi- nent amongst these are the ovarian cancer antigen (OCA) of Knauf and colleagues (Knauf and Urbach, 1977, 1980; Knauf et al., 1984) and the ovarian-carci- noma-associated antigen (OCAA) of Bhattacharya and Barlow (1978). The advent of MAb technology has seen the devel- opment of the OC125 antibody which defines the CA125 antigen (Bast et al., 1981; Kabawat et al., 1983). The use of this antibody for monitoring the progression of ovarian carcinomas has been reported (Bast et al., 1983; Klug et al., 1984). Recently, MAbs reacting with glycolipid antigens associated with ovar- ian mucinous carcinomas have been described (Taglia- bue et al., 1985; Miotti et al., 1985). MAb-defined antigens with wider tissue distribution, such as placen- tal alkaline phosphatase (Goldstein et al., 1982; Sun- derland et al., 1984; Travers and Bodmer, 1984), HMFG2 (Arklie et al., 1981; Taylor-Papadimitriou et al., 1981) and F36/22 (Croghan et al., 1983), may also be of value in the diagnosis of ovarian cancers (Sunderland et al., 1984; Nouwen et al., 1985; van de Voorde et al., 1985; Epenetos et al., 1982; Singer et al., 1985; Croghan et al., 1984). Previous studies established that the MAb OM-1 identified a cell-surface antigen which, while present at low levels in normal “Mullerian” epithelium, was expressed at high levels by ovarian serous and endo- metrioid carcinoma cells (de Kretser et al., 1985). Extensive analysis showed readily detectable SGA expression in normal tissues to be restricted to 3 highly- specialized cell types-sebaceous gland cells, lung type-XI pneumocytes and placental syncytial tropho- blasts. Amongst tumour tissues, an extremely high level of SGA expression was a consistent feature of malignant ovarian serous and endometrioid tumours. No increase in basal levels of SGA expression was detectable in malignant ovarian mucinous tumours, or in benign ovarian tumours. No SGA expression was observed in other epithelium-derived tumours, includ- ing breast and colon adenocarcinomas. As the differ- entiation between metastatic breast or colon carcinoma and ovarian adenocarcinoma is often difficult, SGA expression may prove valuable as a marker for ovarian serous and endometrioid malignancies. The capacity to distinguish between benign and malignant ovarian tumours on the basis of SGA expression further in- creases its potential usefulness (de Kretser et al., 1985). The biochemical nature of SGA was determined by electrophoretic analysis of molecules immunoprecipi- tated from biosynthetically-labelled ovarian carcino- ma-derived cell lines using the MAb OM-1. MATERIAL AND METHODS Cell lines The ovarian carcinoma-derived cell lines Colo 3 16 (gift of Dr. G. Moore, CO) and 80.13A (Bertonocello et al., 1985) were grown as monolayer cultures in RPMI 1640 medium (Flow, Melbourne, Australia) supplemented with 10% foetal calf serum (Flow) and 40 pg/ml gentamycin (Roussell, Castle Hill, NSW, Australia). Biosynthetic labelling of cells Colo 316 and 80.13A cells were biosynthetically labelled with 35S-methionine by 24 hr incubation at 37°C of semi-confluent cells in 75-cm2 flasks (approx- imately 6 X lo6 cells) with 10 ml methionine-free RPMI 1640 (Gibco, Melbourne) containing 10% di- alyzed foetal calf serum and 1 mCi 35S-methionine (Amersham, Australia). Cells were harvested by lysis and cell lysates prepared (de Kretser et al., 1982). Independently, Colo 3 16 cells (semi-confluent 25- cm2 flask; approximately 2 x lo6 cells) were labelled by addition of 50 pCi 14C-mannose (Amersham) fol- Abbreviations: kDa, kilodaltons; SDS, sodium dodecyl sul- phate; MW, molecular weight; SGA, sebaceous gland antigen; PAGE, polyacrylamide gel electrophoresis; NEPHGE, non-equi- librium PH gradient electrophoresis; lD, one-dimensional;MAb, monoclonal antibody. ‘To whom reprint requests should be sent. Received: June 19, 1985 and in revised form December 5, 1985.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Biochemical characterization of the monoclonal antibody-defined ovarian carcinoma-associated antigen...

Int. J. Cancer: 37, 705-712 (1986) 0 1986 Alan R. Liss, Inc.
BIOCHEMICAL CHARACTERIZATION OF THE MONOCLONAL
ANTIGEN SGA ANTIBODY -DEFINED OVARIAN CARCINOMA-ASSOCIATED
Theonne A. DE KRETSER’, Heather J. THORNE, Diana PICONE and David G. JOSE Imrnunogenetics Research Unit. 7he Cancer Institute. 481 Little Lonsdale Street, Melbourne, Victoria 3000, Australia.
The molecular nature of SGA, the ovarian-carci- noma-associated antigen defined by the MAb OM-I, has been determined. The cell-surface form of the SGA molecule is a glycoprotein with pl <4.2, which on PAGE analysis has an apparent MW of approxi- mately 360 kDa. This was the only OM-I-reactive spe- cies found on the cell surface. The apparent MW was unaffected by reducing conditions. The predominant cytoplasmic form of SGA is a non-glycosylated 170- kDa molecule with pl 6.5. Pulse-chase experiments were complicated by the extremely slow rate of SGA synthesis. However, the data indicate that the SGA molecule is synthesized as a 190-kDa protein, cleaved to yield a 170-kDa non-glycosylated intracellular form which is slowly glycosylated to the 360-kDa cell-sur- face species. Western blotting experiments revealed the presence of the 360-kDa glycosylated molecule in human ovarian cell culture supernatants.
For more than a decade, the development of immu- nological markers for ovarian epithelial cancer has been of interest to those searching for the means to detect, diagnose, monitor and treat this disease. Since the first demonstration of ovarian tumour-associated antigens using heteroantisera (Levi et al., 1969), a number of such antigens have been described. Promi- nent amongst these are the ovarian cancer antigen (OCA) of Knauf and colleagues (Knauf and Urbach, 1977, 1980; Knauf et al., 1984) and the ovarian-carci- noma-associated antigen (OCAA) of Bhattacharya and Barlow (1978).
The advent of MAb technology has seen the devel- opment of the OC125 antibody which defines the CA125 antigen (Bast et al., 1981; Kabawat et al., 1983). The use of this antibody for monitoring the progression of ovarian carcinomas has been reported (Bast et al., 1983; Klug et al., 1984). Recently, MAbs reacting with glycolipid antigens associated with ovar- ian mucinous carcinomas have been described (Taglia- bue et al., 1985; Miotti et al., 1985). MAb-defined antigens with wider tissue distribution, such as placen- tal alkaline phosphatase (Goldstein et al., 1982; Sun- derland et al., 1984; Travers and Bodmer, 1984), HMFG2 (Arklie et al., 1981; Taylor-Papadimitriou et al . , 1981) and F36/22 (Croghan et al., 1983), may also be of value in the diagnosis of ovarian cancers (Sunderland et al., 1984; Nouwen et al., 1985; van de Voorde et al., 1985; Epenetos et al., 1982; Singer et al., 1985; Croghan et al., 1984).
Previous studies established that the MAb OM-1 identified a cell-surface antigen which, while present at low levels in normal “Mullerian” epithelium, was expressed at high levels by ovarian serous and endo- metrioid carcinoma cells (de Kretser et al., 1985). Extensive analysis showed readily detectable SGA expression in normal tissues to be restricted to 3 highly- specialized cell types-sebaceous gland cells, lung
type-XI pneumocytes and placental syncytial tropho- blasts. Amongst tumour tissues, an extremely high level of SGA expression was a consistent feature of malignant ovarian serous and endometrioid tumours. No increase in basal levels of SGA expression was detectable in malignant ovarian mucinous tumours, or in benign ovarian tumours. No SGA expression was observed in other epithelium-derived tumours, includ- ing breast and colon adenocarcinomas. As the differ- entiation between metastatic breast or colon carcinoma and ovarian adenocarcinoma is often difficult, SGA expression may prove valuable as a marker for ovarian serous and endometrioid malignancies. The capacity to distinguish between benign and malignant ovarian tumours on the basis of SGA expression further in- creases its potential usefulness (de Kretser et al., 1985).
The biochemical nature of SGA was determined by electrophoretic analysis of molecules immunoprecipi- tated from biosynthetically-labelled ovarian carcino- ma-derived cell lines using the MAb OM-1.
MATERIAL AND METHODS
Cell lines The ovarian carcinoma-derived cell lines Colo 3 16
(gift of Dr. G. Moore, CO) and 80.13A (Bertonocello et al., 1985) were grown as monolayer cultures in RPMI 1640 medium (Flow, Melbourne, Australia) supplemented with 10% foetal calf serum (Flow) and 40 pg/ml gentamycin (Roussell, Castle Hill, NSW, Australia). Biosynthetic labelling of cells
Colo 316 and 80.13A cells were biosynthetically labelled with 35S-methionine by 24 hr incubation at 37°C of semi-confluent cells in 75-cm2 flasks (approx- imately 6 X lo6 cells) with 10 ml methionine-free RPMI 1640 (Gibco, Melbourne) containing 10% di- alyzed foetal calf serum and 1 mCi 35S-methionine (Amersham, Australia). Cells were harvested by lysis and cell lysates prepared (de Kretser et al., 1982).
Independently, Colo 3 16 cells (semi-confluent 25- cm2 flask; approximately 2 x lo6 cells) were labelled by addition of 50 pCi 14C-mannose (Amersham) fol-
Abbreviations: kDa, kilodaltons; SDS, sodium dodecyl sul- phate; MW, molecular weight; SGA, sebaceous gland antigen; PAGE, polyacrylamide gel electrophoresis; NEPHGE, non-equi- librium PH gradient electrophoresis; lD, one-dimensional; MAb, monoclonal antibody.
‘To whom reprint requests should be sent.
Received: June 19, 1985 and in revised form December 5 , 1985.

706 DE KRETSE
lowed by incubation at 37°C for 20 hr. Cell lysates were prepared as above. Preparation of glycoprotein fractions
Glycoprotein fractions were prepared from 35S-me- thionine-labelled cell lysates using a column contain- ing equal volumes of Lens culinaris (lentil) lectin- conjugated and Ricinis communis (ricin)-conjugated Sepharose 4B (de Kretser et al., 1982). Monoclonal antibody OM-1
Stocks of purified antibody at a concentration of 1 mg/ml protein were prepared by protein-A affinity chromatography of the ammonium-sulphate-precipi- tated immunoglobulin fraction of ascitic fluids col- lected from mice inoculated intraperitoneally with the hybridoma OM-1 (de Kretser et al., 1985\ The pooled purified antibody had a titre of >lo- by ELISA against Colo 316 cells (de Kretser et al., 1985), and was stored in small aliquots at -70°C. The antibody OM-1 has been shown to be of the IgM class (de Kretser et al., 1985).
Immunoprecipitation SGA was precipitated from 200-pl aliquots of cell
lysates, glycoprotein fractions or culture supernatants by incubation with 5 pl of purified OM-1 antibody plus 1 p1 of rabbit anti-mouse immunoglobulin antiserum (Miles, Melbourne) for 30 min at 20°C. Formalin- fixed Staphylococcus aureus (SaCI; Cowan I strain; 15 pl 20% v/v) was added to precipitate the antigen- antibody complexes (de Kretser et al., 1982). Antigen- antibody-Sac1 pellets were stored at -70°C prior to analysis. One-dimensional, sodium dodecyl sulphate polyacrylamide gel electrophoretic analysis ( I D-SDS-PAGE)
1D-SDS-PAGE analyses of immunoprecipitated pro- teins using 7 % acrylamide slab gels were performed (de Kretser et al., 1982). Two-dimensional gel electrophoresis
Two-dimensional analyses of immunoprecipitated proteins were performed (de Kretser et al., 1982) us- ing non-equilibrium PH gradient electrophoresis (NEPHGE) for first-dimension separation and SDS- PAGE using 7% acrylamide slab gels in the second dimension. Pulse-chase experiments
Aliquots of lo6 Colo 316 cells were seeded into multiple 25-cm2 tissue culture flasks and allowed to attach and grow overnight. The resulting monolayers were rinsed once with methionine-free RPMI 1640 and pulse-labelled by incubation at 37°C for 1 hr in 2.5 mi methionine-free RPMI 1640 supplemented with 200 pCi 35S-methionine. After incubation, radioactive me- dium was withdrawn and replaced by complete RPMI 1640 containing 10% foetal calf serum (FCS), and incubation at 37°C was continued. At 0, 1, 2, 6, 8 and 12 hr thereafter, one flask was withdrawn from incu- bation and a cell lysate prepared. Preliminary experi- ments were performed to determine the optimal du- ration of both pulse and chase. Identification of cell-su@ace-exposed proteins
Confluent monolayers of Colo 316 cells (25-cm2 flasks; approximately 3 x lo6 cells) were treated for
IR ET AL
1 hr at 37°C with 50 pg/ml OM-1 antibody, 20 pg/ml anti-HLA MAb or 50 pgiml normal mouse immuno- globulin fraction in RPMI 1640 supplemented with 10% FCS and 0.02% sodium azide. Unbound antibody was removed by rinsing in phosphate-buffered saline containing 0.02% sodium azide and cell lysates pre- pared as described above. Aliquots of 200 pl of this lysate were immunoprecipitated as described above, but without the addition of extra primary MAb. Im- munoprecipitated proteins were separated by 1D-SDS- PAGE as described above and probed with the OM-1 antibody by Western blotting as detailed below. Western blotting experiments
Proteins separated by 1 D-SDS-PAGE as described above were blotted onto nitrocellulose (Schleicher- Schull, BA85; Bartelt, Melbourne) (Towbin et al., 1979; Bittner et al., 1980; Burnette, 1981). Immuno- blotted proteins were probed with OM-1 MAb at a dilution of 1 : 100. Bound OM-1 antibody was detected by reaction with biotinylated sheep anti-mouse immu- noglobulin antiserum (1 :200 dilution; Amersham), fol- lowed by incubation with pre-formed streptavidin- peroxidase complexes (1 : 500 dilution; Amersham). The nitrocellulose-bound complexes were visualized by reaction with 4-chloro-1-naphthol (Sigma, St. Louis, MO). Purijkation of SGA from culture supernatants
SGA was partially purified from culture superna- tants by concentration and fractionation on a Sephacryl S200 superfine column (Pharmacia, Australia; 56 cm X 1.5 cm diam.) at 50 pl/min. Two major peaks were eluted from this column, the first containing the gly- coprotein form of SGA together with sundry protein contaminants. The second peak contained lower mo- lecular weight proteins such as albumin. The first peak, concentrated to 10 mg/ml protein, was treated with the protease inhibitor phenyl methyl sulphonyl fluoride (Sigma; 1 mM final concentration) and sodium azide to 0.01% and stored at 4°C. Aliquots of 5 1 1 were analysed by 1D-PAGE and Western blotting.
RESULTS
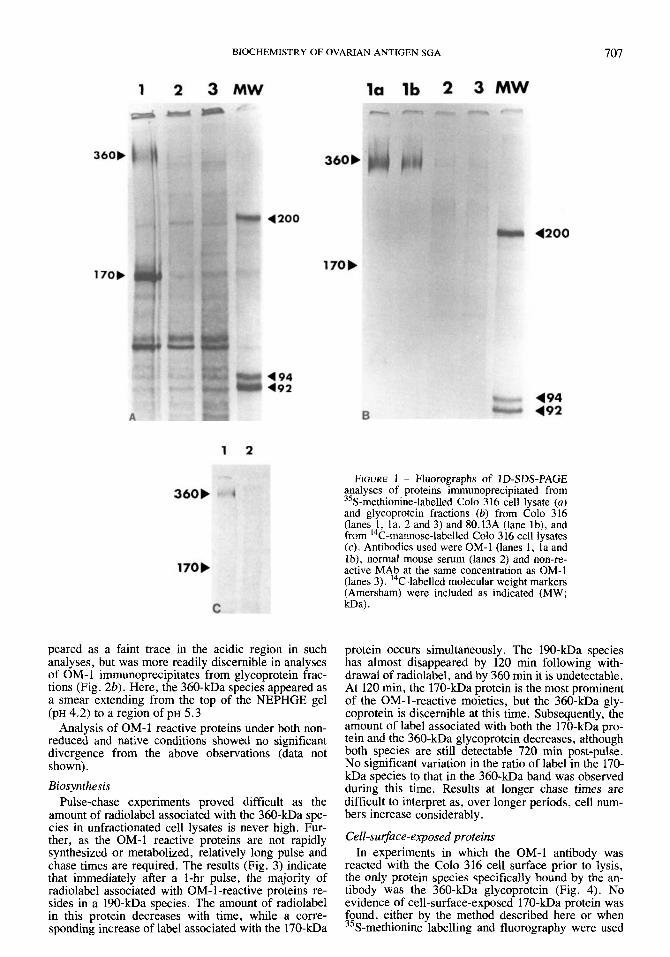
Molecular weight, glycosylation and p l 1 D-SDS-PAGE analyses of proteins immunoprecip-
itated from 35S-methionine-labelled Colo 3 16 cell ly- sates by the MAb OM-1 consistently revealed the presence of a definite band at 170 kDa, together with a less distinct band at 360 kDa (Fig. la). Identical results were obtained using the 80.13A cell line (data not shown). Similar analysis of material immunopre- cipitated from glycoprotein fractions of both Colo 3 16 and 80.13A cell lysates indicated the presence of the 360-kDa species, but not the 170 kDa protein, in these samples (Fig. lb). The 360-kDa band was not well resolved by the electrophoresis system used, a phe- nomenon characteristic of highly glycosylated proteins.
1D-SDS-PAGE analyses of proteins immunoprecip- itated from ''C-mannose-labelled cell lysates revealed the presence of the 360-kDa species only (Fig. lc). No evidence of the 170-kDa protein could be detected, even after prolonged exposures.
In two-dimensional NEPHGE-PAGE analyses of the OM-1-reactive proteins in Colo 316 cell lysates, the 170-kDa species appeared as a discrete spot with ap- parent PI of 6.5 (Fig. 2a). The 360-kDa species ap-
BIOCHEMISTRY OF OVARIAN ANTIGEN SGA 707
peared as a faint trace in the acidic region in such analyses, but was more readily discernible in analyses of OM- 1 immunoprecipitates from glycoprotein frac- tions (Fig. 2b). Here, the 360-kDa species appeared as a smear extending from the top of the NEPHGE gel (PH 4.2) to a region of PH 5.3
Analysis of OM-1 reactive proteins under both non- reduced and native conditions showed no significant divergence from the above observations (data not shown). Biosynthesis
Pulse-chase experiments proved difficult as the amount of radiolabel associated with the 360-kDa spe- cies in unfractionated cell lysates is never high. Fur- ther, as the OM-l reactive proteins are not rapidly synthesized or metabolized, relatively long pulse and chase times are required. The results (Fig. 3) indicate that immediately after a 1-hr pulse, the majority of radiolabel associated with OM- 1-reactive proteins re- sides in a 190-kDa species. The amount of radiolabel in this protein decreases with time, while a corre- sponding increase of label associated with the 170-kDa
FIGURE 1 - Fluorographs of ID-SDS-PAGE analyses of proteins immunoprecipitated from 35S-methionine-labelled Colo 3 16 cell lysate (a) and glycoprotein fractions (b) from Colo 316 (lanes 1, la, 2 and 3) and 80.13A (lane lb), and from ''C-mannose-labelled Colo 316 cell lysates (c) . Antibodies used were OM-1 (lanes 1, la and lb), normal mouse serum (lanes 2) and non-re- active MAb at the same concentration as OM-1 (lanes 3). ''C-labelled molecular weight markers (Amersham) were included as indicated (MW; m a ) .
protein occurs simultaneously. The 190-kDa species has almost disappeared by 120 min following with- drawal of radiolabel, and by 360 min it is undetectable. At 120 min, the 170-kDa protein is the most prominent of the OM-1-reactive moieties, but the 360-kDa gly- coprotein is discernible at this time. Subsequently, the amount of label associated with both the 170-kDa pro- tein and the 360-kDa glycoprotein decreases, although both species are still detectable 720 min post-pulse. No significant variation in the ratio of label in the 170- kDa species to that in the 360-kDa band was observed during this time. Results at longer chase times are difficult to interpret as, over longer periods, cell num- bers increase considerably.
Cell-&ace-exposed proteins In experiments in which the OM-1 antibody was
reacted with the Colo 316 cell surface prior to lysis, the only protein species specifically bound by the an- tibody was the 360-kDa glycoprotein (Fig. 4). No evidence of cell-surface-exposed 170-kDa protein was found, either by the method described here or when 35S-methionine labelling and fluorography were used

DE KRETSER ET AL 708
FIGURE 2 - Fluorographs of NEPHGE-PAGE analyses of proteins immunoprecipitated by MAb OM-1 from 35S-methi- onine-labelled Colo 316 cell lysate (a) and glycoprotein frac- tion (b). The PH was determined at various points along the first dimensional gel and two points are shown, together with MW in kDa, determined by comparison with 14-labelled molecular weight markers (Amersham).
to detect precipitated proteins (data not shown). Con- versely, when cell lysates were analyzed for OM-l- reactive proteins by Western blotting procedures, the 170-kDA molecule was the most prominent species detected (see below; Fig. 5). Presence in extracellular jluids
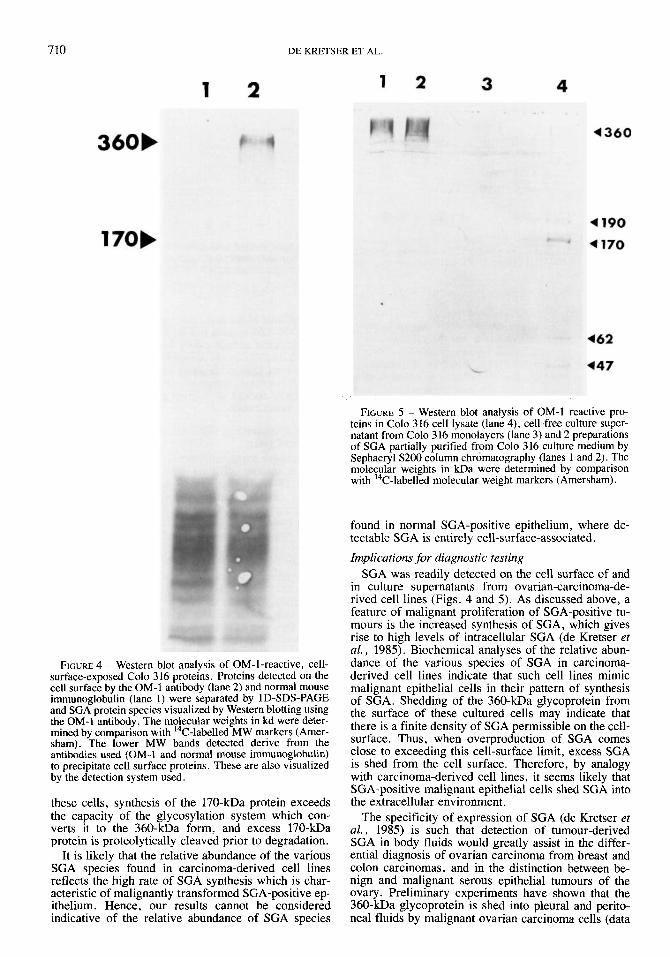
The SGA protein species present in culture fluids were examined by Western blotting using the OM-1 antibody (Fig. 5). When unconcentrated culture fluid
was analysed, the 360-kDa glycoprotein was barely detectable while the 170-kDa species was apparently absent (lane 3). However, analysis of SGA size-frac- tionated from concentrated culture fluids showed large amounts of the 360 kDa glycoprotein (lanes 1 and 2). No evidence of the 170-kDa species was found in these partially purified preparations of SGA. In comparison, in cell lysates the 170 kDa protein is the most promi- nent of the OM-1-reactive species present (lane 4). In these samples, the 190 kDa precursor polypeptide, the 360 kDa glycoprotein and lower molecular weight species of 62 and 47 kDa, presumably proteolytic cleavage products, were also detected.
DISCUSSION
Characteristics of SGA protein species Our analyses have shown that, in ovarian carcinoma
cells, SGA exists in 2 major forms with apparent molecular weights of 170 and 360 kDa (Fig. la). The ability of lectins to bind these proteins was assessed (Fig. lb). Lentil lectin binds with high affinity to core carbohydrate (high mannose-containing) residues which are the first carbohydrate structures to be added to protein during glycosylation. The lectin ricin binds to galactose residues often added during subsequent modification of core structures to form complex car- bohydrate residues. With few exceptions, these lectins used together will bind glycoproteins synthesized by mammalian cells. The results indicated that the 170- kDa protein did not react with the lectins, suggesting that this molecule is non-glycosylated. The 360-kDa protein was bound by the lectins, indicating that this protein is glycosylated. These conclusions were con- firmed by the results obtained using I4C-mannose as radiolabel (Fig. lc). In this case, no incorporation into the 170-kDa protein could be detected while incorpo- ration into the 360-kDa species was readily observed. Hence, the major forms of SGA in ovarian carcinoma cells are a 170-kDa non-glycosylated protein and a 3 60- kDa gl y coprotein.
The apparent MW on PAGE of the 360-kDa glyco- protein may not reflect its true MW, as many glycopro- teins do not behave ideally in this system. Hence, the figure of 360 kDa should be considered strictly as the apparent MW observed under the particular experi- mental conditions employed here. Nevertheless, the figure given for apparent MW on SDS-PAGE is of value for purposes of comparison with other molecules similarly analysed. The poor resolution observed for the 360-kDa glycoprotein on 1D-SDS-PAGE is not a function of the gel system used, as other glycoproteins of MW > 360 kDa seen in analyses of total glycopro- tein fractions are well resolved. Further, the appear- ance of the 360-kDa species on 1D-SDS-PAGE does not alter with acrylamide concentration (5-12% acry- lamide; data not shown). The highly glycosylated gly- coproteins HMFGl and HMFG2 are similarly, poorly resolved by ID-SDS-PAGE (Burchell et al., 1983). It is probable that the poor resolution observed for the 360-kDa glycoprotein reflects a high degree of glyco- sylation.
The only SGA protein detected on the surface of ovarian carcinoma cells was the 360-kDa glycoprotein (Fig. 4). The 360-kDa glycoprotein was also detected in culture supernatants from ovarian carcinoma-de- rived cell lines (Fig. 5). No evidence of the presence

BIOCHEMISTRY OF OVARIAN ANTIGEN SGA 709
FIGURE 3 - Fluorographs of ID-SDS-PAGE analysis of proteins immunoprecipitated from Colo 316 cell lysates prepared at the times indicated after a 1-hr pulse of ”S-methionine, as described in “Methods”. Antibodies used were normal mouse serum (lanes 1) and MAb OM-1 (lanes 2). 14C-labelled MW markers (Amersham) are included (MW; ma).
of the 170-kDa non-glycosylated protein was found either on the cell surface or in culture supernatants. Conversely, the 170-kDa protein was the predominant SGA species detected in cell lysates. Neither the 360- kDa nor the 170-kDa molecule could be dissociated into subunits, nor was any evidence of association of either of these proteins with any other protein detected. It therefore seems likely that the 170-kDa non-glyco- sylated protein is the intracellular precursor of the 360- kDa cell-surface-exposed glycoprotein. Biosynthesis of SGA
The results obtained indicate that SGA is synthesized as a 190-kDa polypeptide. Subsequent proteolytic cleavage of this molecule presumably removes a signal sequence of 20 kDa to yield a 170-kDa protein which is the major intracellular form of SGA. It seems likely that this protein then undergoes substantial glycosyla- tion, ultimately giving rise to the 360-kDa cell-surface species. In the cell lines studied, this cell-surface mol- ecule is apparently shed from the cell and is found in the extracellular fluid as a 360-kDa glycoprotein.
The absence of the 170-kDa protein from the cell surface and extracellular environment, its non-glyco- sylated state, the existence of a glycosylated form of SGA in the same cells and the PI of the 170-kDa protein relative to the PI of the 360-kDa glycoprotein are all consistent with the hypothesis that the 170-kDa protein may be the intracellular precursor of the 360- kDa glycoprotein. The results of pulse-chase experi- ments were inconclusive in this respect. Nevertheless, the results obtained do not rule out the existence of a precursor-product relationship between the 170-kDa protein and the 360-kDa glycoprotein.
The pulse-chase results illustrate the difficulty of biosynthetic labelling experiments on very slowly syn- thesized molecules. Thus, the 190-kDa precursor pro- tein was the only SGA species detectably labelled after a l-hr pulse (Fig. 3). This problem was compounded by the shedding of relatively large quantities of the 360-kDa glycoprotein into the culture fluid. Thus, in these experiments, the cell-surface-associated 360-kDa molecule may well have behaved as a biosynthetic intermediate between the 170-kDa protein and the 360- kDa extracellular species. Such a hypothesis would account for the relatively constant ratio of radiolabel in the 170-kDa protein and the 360-kDa cell-associated glycoprotein.
On the basis of the data reported here, the possibility that the 170-kDa protein and the 360-kDa glycoprotein are independent entities cannot be entirely discounted. However, the likelihood of finding two unrelated mol- ecules which have no features in common other than the OM- l-defined antigenic determinant but which nevertheless coexist in the two tumour cell lines ex- amined seems remote.
Relative abundance of 170- and 360-kDa species in tumour-cell lysates
The relative amounts of the various SGA species in lysates of tumour cell lines was assessed by Western blotting (Fig. 5). It seems unlikely that the OM-1 antibody differentiates between the 170- and 360-kDa molecules, as the results of the Western blot of Colo 316 cell lysate (Fig. 5) show the same relative amounts of these two proteins as was detected by silver staining of samples of CoIo 316 cell lysates similarly separated on 1D-SDS-PAGE (data not shown).
Immunoperoxidase studies show that SGA is re- stricted to the cell surface in normal epithelium, and this cell-surface expression is increased in proliferat- ing epithelium (de Kretser et al., 1985). Generally accepted notions of biosynthesis therefore suggest that the 360-kDa cell-surface product should be the most abundant SGA species present. However, in metastatic carcinoma cells, the cellular distribution of SGA is altered, with considerable SGA activity being detected in the cytoplasm of malignant cells (de Kretser et al., 1985). As the cell lines used in this study were derived from metastatic carcinoma cells, it is likely that they maintain the pattern of SGA biosynthesis characteristic of such cells. This has been confirmed by immunoper- oxidase staining experiments (data not shown). Our present results are in accord with this finding, showing a high level of the intracellular, 170-kDa SGA protein relative to its presumptive product, the 360-kDa cell- surface form (Fig. la; Fig. 5). The presence of readily detectable 190-kDa precursor polypeptide in tumour- cell lysates also points to abundant SGA syn- thesis. The 62- and 47-kDa polypeptides observed in cell lysates (Fig. 5) are most likely derived through proteolytic cleavage of either the 190- or 170-kDa proteins as they are not glycosylated but must contain the OM-l-defined antigenic determinant in order to be visualized in the Western blotting experiments. Their presence in tumour-cell lysates may indicate that, in
710 D E KRETSER ET AL
FIGURE 4 - Western blot analysis of OM-1-reactive, cell- surface-exposed Colo 316 proteins. Proteins detected on the cell surface by the OM-1 antibody (lane 2) and normal mouse immunoglobulin (lane 1) were separated by ID-SDS-PAGE and SGA protein species visualized by Western blotting using the OM-1 antibody. The molecular weights in kd were deter- mined by comparison with 14C-labelled MW markers (Amer- sham). The lower MW bands detected derive from the antibodies used (OM-I and normal mouse immunoglobulin) to precipitate cell surface proteins. These are also visualized by the detection system used.
these cells, synthesis of the 170-kDa protein exceeds the capacity of the glycosylation system which con- verts it to the 360-kDa form, and excess 170-kDa protein is proteolytically cleaved prior to degradation.
It is likely that the relative abundance of the various SGA species found in carcinoma-derived cell lines reflects the high rate of SGA synthesis which is char- acteristic of malignantly transformed SGA-positive ep- ithelium. Hence, our results cannot be considered indicative of the relative abundance of SGA species
FIGURE 5 - Western blot analysis of OM-1 reactive pro- teins in Colo 316 cell lysate (lane 4), cell-free culture super- natant from Colo 3 16 monolayers (lane 3) and 2 preparations of SGA partially purified from Colo 316 culture medium by Sephacryl S 2 0 0 column chromatography (lanes 1 and 2). The molecular weights in kDa were determined by comparison with ''C-labelled molecular weight markers (Amersham).
found in normal SGA-positive epithelium, where de- tectable SGA is entirely cell-surface-associated. Implications for diagnostic testing
SGA was readily detected on the cell surface of and in culture supernatants from ovarian-carcinoma-de- rived cell lines (Figs. 4 and 5). As discussed above, a feature of malignant proliferation of SGA-positive tu- mours is the increased synthesis of SGA, which gives rise to high levels of intracellular SGA (de Kretser et al., 1985). Biochemical analyses of the relative abun- dance of the various species of SGA in carcinoma- derived cell lines indicate that such cell lines mimic malignant epithelial cells in their pattern of synthesis of SGA. Shedding of the 360-kDa glycoprotein from the surface of these cultured cells may indicate that there is a finite density of SGA permissible on the cell- surface. Thus, when overproduction of SGA comes close to exceeding this cell-surface limit, excess SGA is shed from the cell surface. Therefore, by analogy with carcinoma-derived cell lines, it seems likely that SGA-positive malignant epithelial cells shed SGA into the extracellular environment.
The specificity of expression of SGA (de Kretser et al., 1985) is such that detection of tumour-derived SGA in body fluids would greatly assist in the differ- ential diagnosis of ovarian carcinoma from breast and colon carcinomas, and in the distinction between be- nign and malignant serous epithelial tumours of the ovary. Preliminary experiments have shown that the 360-kDa glycoprotein is shed into pleural and perito- neal fluids by malignant ovarian carcinoma cells (data

BIOCHEMISTRY OF OVARIAN ANTIGEN SGA 711
not shown). At present, it is not clear whether SGA is shed at low levels from normal SGA-positive epithelial cells, nor at what stage of malignant transformation extracellular SGA becomes detectable. Similarly, it is not known if the levels of extracellular SGA will re- flect tumour mass. Experiments are currently under- way to devise suitable methods for the measurement of SGA levels in patient serum. Relationship to other ovarian-carcinoma- associated antigens
On the basis of its tissue distribution, SGA has been shown to be distinct from all other ovarian-carcinoma- associated antigens (de Kretser et al., 1985). The mo- lecular description of SGA supports this assertion. Hence, as a 360-kDa glycoproteinl170-kDa protein, SGA is unlike OCA which has been described as a 70- kDa protein (Knauf and Urbach, 1980). The Movl and Mov2 antigens are glycolipids (Miotti et al., 1985) and, in common with the high-molecular-weight gly- coprotein OCAA (Bhattacharya and Barlow, 1978), are found on mucinous ovarian tumours from which SGA is absent (de Kretser et al., 1985). The CA125 antigen has been defined as a dimer comprising pro- teins of 180 and >20-kDa in trypsinized cells, but apparently exists as a larger, >200-kDa protein in intact cells (Masuho et al., 1984). This finding sug- gests that the CA125 antigen is somewhat smaller than SGA, as is placental alkaline phosphatase which is a homodimer of a 67-kDa protein (Sunderland et al., 1984; Travers and Bodmer, 1984).
The antigens detected by the MAbs HMFG2 and F36122 have been described as highly glycosylated, high-molecular-weight glycoproteins of > 400 and > 700 kDa respectively (Burchell et al., 1983; Croghan et al., 1984). In the case of HMFG2, the antigenic determinant recognized appears to be carbohydrate in
nature. Further, a degree of variation in the molecular weight of HMFG2-reactive glycoproteins detected in different breast cancer cell lines has been observed (Burchell et al., 1983). Similarly, F36122 has been reported to react with a family of high-molecular- weight glycoproteins in ovarian carcinoma cells (Croghan et al., 1984). As precise determination of the molecular weight of highly glycosylated glycopro- teins >200 kDA is extremely difficult, it is possible that, in ovarian carcinomas, the molecule detected by either of these antibodies may be SGA. It should be emphasized that the OM-1 MAb reacts with a protein- based determinant and its reaction with SGA is unaf- fected by the degree of glycosylation (Fig. 1). The tissue distribution of staining obtained with the OM-1 antibody is more restricted than those reported for the HMFG2 and F36122 antibodies (de Kretser et al., 1985; Arklie et al., 1981; Taylor-Papadimitriou et al., 1981; Croghan et al., 1983). It may be that SGA is one of a family of highly glycosylated, high-molecu- lar-weight glycoproteins comprising molecules with different protein moieties but similar carbohydrate structures, which are differentially expressed on the surface of many types of epithelial cell. The possibility of such a family relationship existing between HMFG2 and SGA is currently under investigation.
ACKNOWLEDGEMENTS
We thank Mr. C. Frewen, Ms. S. Trewartha and Mr. J . Bartholomeusz for their assistance with photog- raphy, Dr. T. Kreigler for help and advice on column purification of proteins and Mr. P. Dempsey, Dr. T. Kreigler, Dr. K. Savin, Dr. R. Bradley and Dr. G. Hodgson for their comments. This work was sup- ported by the Research Committee of the Cancer Institute.
REFERENCES
ARKLIE, J., TAYLOR-PAPADIMITRIOU, J., BODMER, W.F., EGAN, M., and MILLIS, R., Differentiation antigens expressed by epi- thelial cells in the lactating breast are also detectable in breast cancers. Inr. J . Cancer, 28, 23-29 (1981). BAST, R.C., FEENEY, M., LAZARUS, H., NADLER, L.M., COL- VIN, R.B., and KNAPP, R.C., Reactivity of a monoclonal anti- body with human ovarian carcinoma. J. clin. Invest., 68, 1331- 1337 (1981). BAST, R.C., KLUG, T.L., ST. JOHN, E., JENISON, E. , NILOFF, J.M., LAZARUS, H., BERKOWITZ, R.S., LEAVITT, T., GRIFFITHS, T., PARKER, L., ZURAWSKI, V.R., and KNAPP, R.C., A radioim- munoassay using a monoclonal antibody to monitor the course of epithelial ovarian cancer. N. Engl. J. Med., 309, 883-887 (1983). BHATTACHARYA, M., and BARLOW, J.J., Ovarian tumour anti- gens. Cancer, 42, 1616-1620 (1978). BERTONOCELLO, I., BRADLEY, T.R., WEBBER, L.M., HODGSON, G.S., and CAMPBELL, J.J., Human tumour cell lines established using dona1 agar culture. Aust. J. exp. Biol. med. Sci. , 63,241- 248 (1985). BITTNER, M., KUPFERER, P., and MORRIS, C.F., Electrophoretic transfer of proteins and nucleic acids from slab gels to diazoben- zyloxymethyl cellulose or nitrocellulose sheets. Anal. Biochem. , 102,459-471 (1980). BURCHELL, J., DURBIN, H., and TAYLOR-PAPADIMITRIOU, J., Complexity of expression of antigenic determinants recognized by monoclonal antibodies HMFG-1 and HMFG-2 in normal and malignant human mammary epithelial cells. J. Immunol. , 131,
BURNETTE, W.N., “Western blotting”: electrophoretic transfer of proteins from sodium dodecyl sulphilte-polyacrylamide gels to
508-513 (1983).
unmodified nitrocellulose and radiographic detection with anti- body and radioiodinated protein-A. Anal. Biochem., 112, 195- 203 (1981). CROGHAN, G.A., PAPSIDERO, L.D., VALENZUELA, L.A., NEM- OTO, T., PENETRANTE, R., and CHU, T.M., Tissue distribution of an epithelial and tumor-associated antigen recognised by mono- clonal antibody F36/22. Cancer Res., 43,4980-4988 (1983). CROGHAN, G.A., WINGATE, M.B., GAMARRA, M., JOHNSON, E., MINGCHU, T., ALLEN, H., VALENZUELA, L., TSUKADA, Y., and PAPSIDERO, L.D., Reactivity of monoclonal antibody F36/22 with human ovarian adenocarcinomas. Can Res., 44, 1954-1962 (1984). DE KRETSER, T.A., CRUMFTON, M.C., BODMER, J.G., and BOD- MER, W.F., Demonstration of two distinct light chains in HLA- DR associated antigens by two-dimensional gel electrophoresis. Europ. J. Immunol., 12,214-221 (1982). DE KRETSER, T.A., THORNE, H.J., JACOBS, D. J., and JOSE, D.G., The sebaceous gland antigen defined by the OM-1 antibody is expressed at high density on the surface of ovarian carcinoma cells. Europ. J . Cancer din. Oncol., 21, 1019-1035 (1985).
ING, M., and BODMER, W.F., Use of two epithelium-specific monoclonal antibodies for diagnosis of malignancy in serous effusions. Lancet, 11, 1004-1006 (1982). GOLDSTEIN, D.J., ROGERS, C., and HARRIS, H. , A search for trace expression of placental-like alkaline phosphatase in non- malignant human tissues: demonstration of its occurrence in lung, cervix, testis and thymus. Clin. chim. Actn, 125, 63-75 (1982). KABAWAT, S.E., BAST, R.C., WELCH, W.R., KNAPP, R.C., and
EPENETOS, A.A., C4NT1, G., TAYLOR-PAPADIMITRIOU, J., CURL-

7 12 DE KRETSER ET AL.
COLVIN, R.C., Immunopathologic characterization of a monoclo- nal antibody that recognizes common surface antigens of human ovarian tumors of serous, endometrioid, and clear cell types. Amer. J. din. Parhol., 79, 98-104 (1983). KLUG, T.L., BAST, R.C., NILOFF, J.M., KNAPP, R.C., and Zu- RAWSKI, V.R., Monoclonal antibody immunoradiometric assay for an antigenic determinant (CA125) associated with human epithelial ovarian carcinomas. Cancer Res., 44, 1048-1053 (1984). KNAUF, S., and URBACH, G.I., Purification of human ovarian tumour-associated antigen and demonstration of circulating tu- mour antigen in patients with advanced ovarian cancer. Amer. J. Obstet. Gynec., l27,705-710 (1977). KNAUF, S., and URBACH, G.I., Identification, purification and radioimmunoassay of NBI7OK, a human ovarian tumor-associ- ated antigen. Cancer Res., 41, 1351-1357 (1980). KNAUF, S . , TAILLON-MILLER, P., HELMKAMP, B.F., BONFIGLIO, T.A., and BEECHAM, J.B., Selectivity for ovarian cancer of an improved serum radioimmunoassay for human ovarian tumor- associated antigen NB170K. Gynec. Oncol., 17, 347-355 (1984). LEVI, M.M., KELLER, S., and MANDL, I, Antigenicity of a papillary serous cystadenocarcinoma tissue homogenate and its fractions. Amer. J. Obstet. Gynec., 105, 856-861 (1969). MASUHO, Y., ZALUTSKY, M., KNAPP, R.C., and BAST, R.C., Interaction of monoclonal antibodies with cell surface antigens of human ovarian carcinomas. Cancer Res., 44, 2813-2819 (1984). MIOTTI, S., AGUANNO, S . , CANEVARI, S., DIOTTI, A., ORLANDI, R., SONNINO, S., and COLNAGHI, M.I., Biochemical analysis of human ovarian cancer-associated antigens defined by murine monoclonal antibodies. Cancer Res., 45, 826-832 (1985). NOUWEN, E.J., POLLET, D.E., SCHELSTRAETE, J.B., EERDEKENS, M.W., HANSCH, C., VAN DE VOORDE, A., and DE BROE, M.E.,
Human placental alkaline phosphatase in benign and malignant ovarian neoplasia. Cancer Res., 45, 892-902 (1985). SINGER, S. , BODDINGTON, M.M., and HUDSON, E.A., Immuno- cytochemical reaction of Cal and HMFGZ monoclonal antibod- ies with cells from serous effusions. J. clin. Path., 38, 180-184 (1985). SUNDERLAND, C.A., DAVIES, J.O., and STIRRAT, G.M., Immu- nohistology of normal and ovarian cancer tissue with a monoclo- nal antibody to placental alkaline phosphatase. Cancer Res., 44,
TAGLIABUE, E., MENARD, S., DELLA TORRE, G., BARBANTI, P., MARIANI-CONSTATINI, R., PORRO, G., and COLNAGHI, M.I., Gen- eration of monoclonal antibodies reacting with human epithelial ovarian cancer. Cancer Rex, 45,379-385 (1985). TAYLOR-PAPADIMITRIOIJ, J., PETERSON, J.A., ARKLIE, J., BUR- CHELL, J., CERIANI, R.L., and BODMER, W.F., Monoclonal anti- bodies to epithelium-specific components of the human milk fat globule membrane: production and reaction with cells in culture. Znt. J. Cancer, 28, 17-21 (1981). TOWBIN, H., STAEHELIN, T., and GORDON, J., Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. nut. Acad. Sci.
TRAVERS, P., and BODMER, W., Preparation and characterization of monoclonal antibodies against placental alkaline phosphatase and other human trophoblast-associated determinants. Int. J . Cancer, 33,633-641 (1984). VAN DE VOORDE, A., DE GROOTE, G., DE WAELE, P., DE BROE, M.E., POLLET, D., DE BOEVER, J., VANDEKERCKHOVE, D., and FIERS, W., Screening of sera and tumour extracts of cancer patients using a monoclonal antibody directed against human placental alkaline phosphatase. Europ. J . Cancer clin. Oncol.,
4496-4502 (1984).
p u s h . ) , 76,4350-4354 (1979).
21, 65-71 (1985).




















