
Biochemical and Structural Analysis of Substrate Promiscuity in Plant Mg 2+Dependent...
11
Biochemical and Structural Analysis of Substrate Promiscuity in Plant Mg 2+ -Dependent O-Methyltransferases Jakub G. Kopycki 1 †, Daniel Rauh 1 †, Alexander A. Chumanevich 1 , Piotr Neumann 1 , Thomas Vogt 2 ⁎ and Milton T. Stubbs 1,3 ⁎ 1 Institut für Biotechnologie, Martin-Luther-Universität Halle-Wittenberg, Kurt-Mothes- Straße 3, D-06120 Halle (Saale), Germany 2 Leibniz-Institute of Plant Biochemistry, Department of Secondary Metabolism, Weinberg 3, D-06120 Halle (Saale), Germany 3 Mitteldeutsches Zentrum für Struktur und Dynamik der Proteine, Kurt-Mothes-Straße 3, D-06120 Halle (Saale), Germany Received 12 November 2007; received in revised form 5 February 2008; accepted 7 February 2008 Available online 20 February 2008 Plant S-adenosyl-L-methionine-dependent class I natural product O- methyltransferases (OMTs), related to animal catechol OMTs, are depen- dent on bivalent cations and strictly specific for the meta position of aromatic vicinal dihydroxy groups. While the primary activity of these class I enzymes is methylation of caffeoyl coenzyme A OMTs, a distinct subset is able to methylate a wider range of substrates, characterized by the promis- cuous phenylpropanoid and flavonoid OMT. The observed broad substrate specificity resides in two regions: the N-terminus and a variable insertion loop near the C-terminus, which displays the lowest degree of sequence conservation between the two subfamilies. Structural and biochemical data, based on site-directed mutagenesis and domain exchange between the two enzyme types, present evidence that only small topological changes among otherwise highly conserved 3-D structures are sufficient to differentiate between an enzymatic generalist and an enzymatic specialist in plant nat- ural product methylation. © 2008 Elsevier Ltd. All rights reserved. Edited by M. Guss Keywords: crystal structure; substrate specificity; hybrid protein; loop exchange; plant secondary metabolism Introduction Methylation by S-adenosyl-L-methionine (SAM)- dependent O-methyltransferases (OMTs) (EC 2.1.1) is a common modification in natural product bio- synthesis. Site-specific O-methylation modulates the physiological properties and the chemical reactivity of phenolic hydroxy groups. 1 Class I plant cation- dependent OMTs (also known as caffeoyl coenzyme A OMTs or CCoAOMTs) constitute a group of low molecular mass (23–27 kDa) enzymes that play an important role in methylation of guaiacyl residues present in CCoA or 5-hydroxy feruloyl CoA, both precursors of monolignols. 2 In angiosperms, mem- bers of a second class of metal-independent OMTs (caffeic acid OMTs) are able to methylate the mono- lignol precursors caffeic acid, caffeyl aldehyde, or caffeyl alcohol in the meta position. 3–5 The action of *Corresponding authors. M. T. Stubbs is to be contacted at Institut für Biotechnologie, Martin-Luther-Universität Halle-Wittenberg, Kurt-Mothes-Straße 3, D-06120 Halle (Saale), Germany. E-mail addresses: [email protected]; [email protected]. † J.G.K. and D.R. contributed equally to this work. Present address: D. Rauh, Chemical Genomics Centre of the Max Planck Society, Otto-Hahn-Straße 15, D-44227 Dortmund, Germany. Abbreviations used: OMT, O-methyltransferase; PFOMT, phenylpropanoid and flavonoid OMT; CoA, coenzyme A; CCoA, caffeoyl CoA; rOMT, rat catechol OMT; SAM, S-adenosyl-L-methionine; SAH, S-adenosyl-L- homocysteine; SeMet, selenomethionine. doi:10.1016/j.jmb.2008.02.019 J. Mol. Biol. (2008) 378, 154–164 Available online at www.sciencedirect.com 0022-2836/$ - see front matter © 2008 Elsevier Ltd. All rights reserved.
-
Upload
uni-giessen -
Category
Documents
-
view
0 -
download
0
Transcript of Biochemical and Structural Analysis of Substrate Promiscuity in Plant Mg 2+Dependent...

doi:10.1016/j.jmb.2008.02.019 J. Mol. Biol. (2008) 378, 154–164
Available online at www.sciencedirect.com
Biochemical and Structural Analysis ofSubstrate Promiscuity in PlantMg2+-Dependent O-Methyltransferases
Jakub G. Kopycki1†, Daniel Rauh1†, Alexander A. Chumanevich1,Piotr Neumann1, Thomas Vogt2⁎ and Milton T. Stubbs1,3⁎
1Institut für Biotechnologie,Martin-Luther-UniversitätHalle-Wittenberg, Kurt-Mothes-Straße 3, D-06120 Halle (Saale),Germany2Leibniz-Institute of PlantBiochemistry, Department ofSecondary Metabolism,Weinberg 3, D-06120 Halle(Saale), Germany3Mitteldeutsches Zentrum fürStruktur und Dynamik derProteine, Kurt-Mothes-Straße 3,D-06120 Halle (Saale),Germany
Received 12 November 2007;received in revised form5 February 2008;accepted 7 February 2008Available online20 February 2008
*Corresponding authors.M. T. StubbInstitut für Biotechnologie, Martin-LHalle-Wittenberg, Kurt-Mothes-Stra(Saale), Germany. E-mail addresses:[email protected].† J.G.K. and D.R. contributed equPresent address: D. Rauh, Chemic
the Max Planck Society, Otto-Hahn-Dortmund, Germany.Abbreviations used: OMT, O-met
PFOMT, phenylpropanoid and flavocoenzyme A; CCoA, caffeoyl CoA; rOMT; SAM, S-adenosyl-L-methioninhomocysteine; SeMet, selenomethio
0022-2836/$ - see front matter © 2008 E
Plant S-adenosyl-L-methionine-dependent class I natural product O-methyltransferases (OMTs), related to animal catechol OMTs, are depen-dent on bivalent cations and strictly specific for themeta position of aromaticvicinal dihydroxy groups. While the primary activity of these class Ienzymes is methylation of caffeoyl coenzyme A OMTs, a distinct subset isable to methylate a wider range of substrates, characterized by the promis-cuous phenylpropanoid and flavonoid OMT. The observed broad substratespecificity resides in two regions: the N-terminus and a variable insertionloop near the C-terminus, which displays the lowest degree of sequenceconservation between the two subfamilies. Structural and biochemical data,based on site-directed mutagenesis and domain exchange between the twoenzyme types, present evidence that only small topological changes amongotherwise highly conserved 3-D structures are sufficient to differentiatebetween an enzymatic generalist and an enzymatic specialist in plant nat-ural product methylation.
© 2008 Elsevier Ltd. All rights reserved.
Keywords: crystal structure; substrate specificity; hybrid protein; loopexchange; plant secondary metabolism
Edited by M. Gusss is to be contacted atuther-Universitätße 3, D-06120 [email protected];
ally to this work.al Genomics Centre ofStraße 15, D-44227
hyltransferase;noid OMT; CoA,OMT, rat catechole; SAH, S-adenosyl-L-nine.
lsevier Ltd. All rights reserve
Introduction
Methylation by S-adenosyl-L-methionine (SAM)-dependent O-methyltransferases (OMTs) (EC 2.1.1)is a common modification in natural product bio-synthesis. Site-specific O-methylation modulates thephysiological properties and the chemical reactivityof phenolic hydroxy groups.1 Class I plant cation-dependent OMTs (also known as caffeoyl coenzymeA OMTs or CCoAOMTs) constitute a group of lowmolecular mass (23–27 kDa) enzymes that play animportant role in methylation of guaiacyl residuespresent in CCoA or 5-hydroxy feruloyl CoA, bothprecursors of monolignols.2 In angiosperms, mem-bers of a second class of metal-independent OMTs(caffeic acid OMTs) are able to methylate the mono-lignol precursors caffeic acid, caffeyl aldehyde, orcaffeyl alcohol in the meta position.3–5 The action of
d.

Fig. 1. Structures of OMT substrates methylated bypromiscuous CCoAOMT-like enzymes such as PFOMTfrom the ice plant M. crystallinum.18 1, quercetin; 2,quercetagetin; 3, caffeic acid; 4, CCoA; 5, caffeoyl glucose.The positions of the methyl group acceptor hydroxylgroups are marked; note the two methylation positions forquercetagetin.
155Substrate Promiscuity in Plant OMTs
both enzyme classes in a metabolic grid appears tobe crucial for determining the structural integrity ofthe lignin polymer.6,7
Detailed structural information on the cation-independent enzymes is available, including thechalcone and isoflavoneOMTs fromalfalfa (Medicagosativa), a flavonoid OMT from Arabidopsis thaliana, aswell as an OMT involved in methyl ester formationin Clarkia breweri.8–11 These enzymes are obligatorydimers, with each active site formed from residues ofboth monomeric polypeptide chains. Either generalacid/base catalysis (in the case of phenolic etherformation) or proper substrate positioning (in thecase of the methyl ester formation) is essential forefficient methyl transfer in the individual systems.Although a major goal of these investigations on theclass II enzymes has been the tailoredmodification ofmethylation specificity, which would enable theengineering of biochemical pathways for the pro-duction of transgenic plants with altered lignin com-position or alternative accumulation of diversenatural products, mutagenesis of individual aminoacids has until now only resulted in lower substratespecificity.10
In contrast, less structural information is availablefor cation-dependent OMTs. The 2.0-Å-resolutionstructure of rat catechol OMT (rOMT)12 exhibits atypical Rossmann fold within the coenzyme-bindingdomain of the enzyme, suggesting that SAMmethy-lases share a common structural motif.13 Methyl
Fig. 2. Overall structure of the PFOMT dimer in cartoon repin blue and the ‘insertion loop’ in orange; the secondary structucolor ramped from the N-terminus (blue) to the C-terminus (bound calcium ion (gray sphere) and the cofactor SAH (stick
transfer proceeds via an Sn2-like transition state, withone ionized hydroxyl group, supported by the Mg2+
ion, and a conserved lysine as a catalytic base, toattack the electron-deficient methyl group of SAM.14
In vertebrates, the enzyme is responsible for theinactivation of neurotransmitters such as dopaminein the brain and has therefore received particularattention as an attractive target to cure degeneratebrain diseases.13,15 However, high concentrations of
resentation. Monomer A is in yellow, with the N-terminusre elements of PFOMTare shown for monomer B, which isred). The positions of the active sites are indicated by therepresentation).
156 Substrate Promiscuity in Plant OMTs
catechol OMT are also found in the liver, kidney, orplacenta, where the enzyme inactivates potentiallyreactive or mutagenic phenolics such as quercetin.1Recently, the 3-D structure of the CCoA-specific
CCoAOMT fromM. sativawas solved at a resolutionof 2.4 Å.16 This enzyme, which almost exclusivelymethylates the CoA ester of caffeic acid, displaysvery low affinity for caffeic acid itself, with a directimpact on lignin polymer biosynthesis in gymno-and angiosperms.7 The structure of the alfalfaCCoAOMTprovides insights into themode of actionand specificity of this class of enzymes. While theoverall fold and the active site of the CCoAOMTdimer resemble those of the animal catechol OMT,significant differences in the positioning of the N-terminus and intervening loops in the vicinity of theactive site contribute to specificity for CCoA.In addition to the CCoA-specific enzymes, a second
type of cation-dependent OMT from the ice plantMesembryanthemum crystallinum [phenylpropanoidand flavonoid OMT (PFOMT)] with promiscuoussubstrate specificity has been described.17 Phyloge-netic analyses indicate that plant cation-dependent
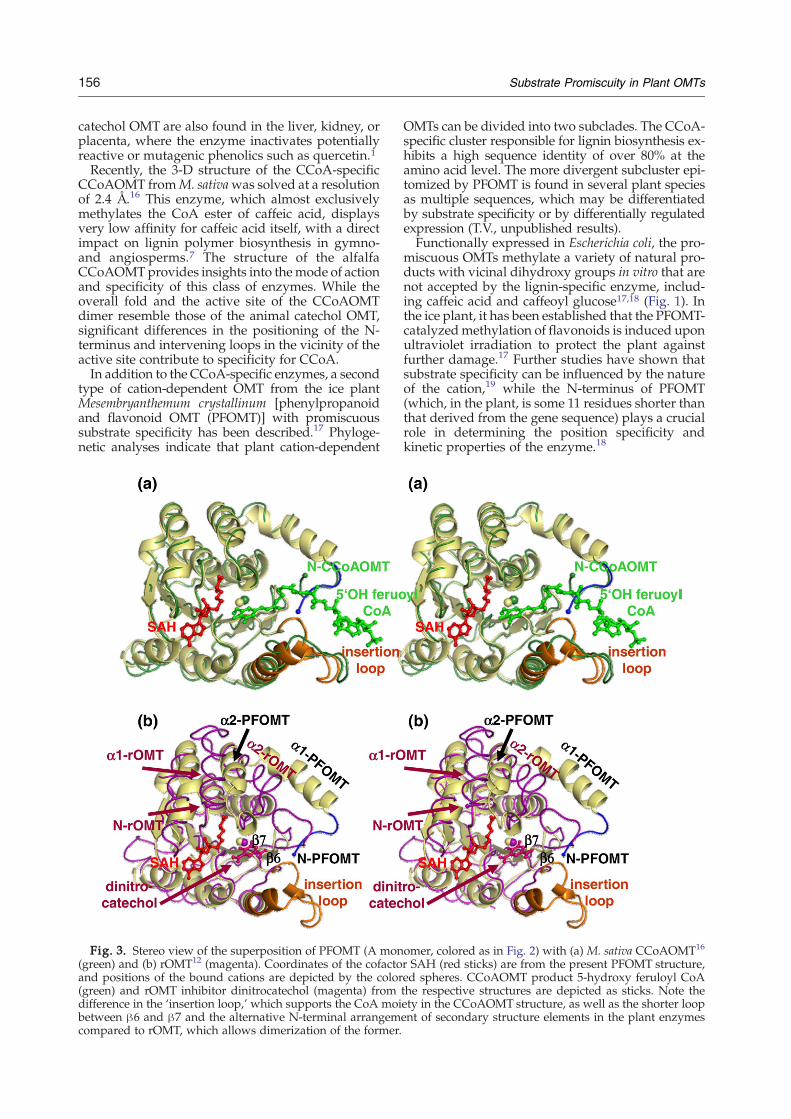
Fig. 3. Stereo view of the superposition of PFOMT (A mon(green) and (b) rOMT12 (magenta). Coordinates of the cofactoand positions of the bound cations are depicted by the color(green) and rOMT inhibitor dinitrocatechol (magenta) fromdifference in the ‘insertion loop,’ which supports the CoA moibetween β6 and β7 and the alternative N-terminal arrangemcompared to rOMT, which allows dimerization of the former.
OMTs can be divided into two subclades. The CCoA-specific cluster responsible for lignin biosynthesis ex-hibits a high sequence identity of over 80% at theamino acid level. The more divergent subcluster epi-tomized by PFOMT is found in several plant speciesas multiple sequences, which may be differentiatedby substrate specificity or by differentially regulatedexpression (T.V., unpublished results).Functionally expressed in Escherichia coli, the pro-
miscuous OMTs methylate a variety of natural pro-ducts with vicinal dihydroxy groups in vitro that arenot accepted by the lignin-specific enzyme, includ-ing caffeic acid and caffeoyl glucose17,18 (Fig. 1). Inthe ice plant, it has been established that the PFOMT-catalyzedmethylation of flavonoids is induced uponultraviolet irradiation to protect the plant againstfurther damage.17 Further studies have shown thatsubstrate specificity can be influenced by the natureof the cation,19 while the N-terminus of PFOMT(which, in the plant, is some 11 residues shorter thanthat derived from the gene sequence) plays a crucialrole in determining the position specificity andkinetic properties of the enzyme.18
omer, colored as in Fig. 2) with (a) M. sativa CCoAOMT16
r SAH (red sticks) are from the present PFOMT structure,ed spheres. CCoAOMT product 5-hydroxy feruloyl CoAthe respective structures are depicted as sticks. Note theety in the CCoAOMT structure, as well as the shorter loopent of secondary structure elements in the plant enzymes

157Substrate Promiscuity in Plant OMTs
Here, we report the crystallization and 3-D struc-ture determination of the cation-dependent PFOMTfrom the ice plant with promiscuous specificity at1.4 Å resolution. Catalytic properties of N-termin-ally truncated variants and hybrids containing loopsfrom the CCoA-specific alfalfa OMT demonstratetheir significant influence on substrate specificity.Simultaneous replacement of two regions from thealfalfa into the ice plant enzyme converts the pro-miscuous enzyme to a CCoA-specific OMT, demon-strating the important contribution of substratebinding by the N-terminus and the loop domain.
Fig. 4. The active site of PFOMT. (a) Representativeelectron density (2Fo−Fc, contoured at 0.9σ) showing thecofactor SAH and the bound cation (gray sphere), inter-preted as Ca2+. (b) Docking of the substrate quercetagetin inthe PFOMT active site. Quercetagetin is methylated in twodistinct positions (see Fig. 1), requiring opposing orienta-tions of the substrate (yellow and pink). Both orientationsmake use of aromatic stacking interactions between thesubstrate and the side chains of Tyr51, Trp184, and Phe198from the ‘insertion loop,’ positioning the pertinent hydroxylgroups in the vicinity of the SAM methyl donor and theactive-site Mg2+ ion (gray sphere).
Results and Discussion
PFOMT is a symmetric dimer of approximatedimensions 70×40×50 Å3 (Fig. 2). As expected,each monomer exhibits an α/β fold similar to themonomeric animal rOMT,13 with eight α-helices andseven β-strands, including the nucleotide-bindingmotif or Rossmann fold. In contrast to rOMT butin commonwith CCoAOMT,16 however, the first twohelices of PFOMT show a distinctly different topolo-gical arrangement. The position of helix α1 in theplant enzymes is in part accomplished by a deletionbetween strands β6 and β7 with respect to themammalian enzyme and contributes significantly todimer formation (Fig. 3). Despite cocrystallizationwith a methyl group donor SAM and the substratequercetin, only the resultant S-adenosyl-L-homocys-teine (SAH) could be detected in the active center(Fig. 4a). Although no electron density could beobserved for the catalytically important N-terminalresidues, the positioning of the first visible aminoacids indicates that the N-termini would be wellplaced to influence catalytic activity. The two activesites, demarcated by the presence of SAH and boundCa2+ ions (Fig. 2), could act independently of oneanother; it is conceivable, however, that a mobileamino-terminus of one monomer might communi-cate with the catalytic apparatus of the second mo-nomer, as has been observed recently for a dimericbacterial OMT from Leptospira interrogans of un-known specificity.20 This is in contrast to the plantcation-independent OMTs, whose active site consistsof residues from both monomers of the dimer, lead-ing to the strong influence of catalytic properties ofthe corresponding second subunit.8The catalytic apparatus and residues in the im-
mediate vicinity are almost identical in each of thethree metal-dependent enzymes, the only signifi-cant difference here being the replacement of rOMTTrp143, which partially covers the SAM moiety andforms a lower boundary to the substrate bindingpocket, by Asp165. All attempts at soaking crystalsor cocrystallization with a variety of substratesfailed to yield any electron density to establish thebinding modes within the active site of PFOMT(data not shown). Nevertheless, superposition of theactive sites of PFOMTwith ligand-bound structuresof CCoAOMT and rOMT allows modeling of thebinding of quercetin and quercetagetin, whose
extended aromatic moieties would be sandwichedbetween the side chains of Tyr51, Trp184, and Phe198(Fig. 4b).In silico docking experiments were carried out to
obtain semiquantitative data for the relative specifi-cities of the two plant enzymes (Table 1). Despite theabsence of structural data for the amino-terminalresidues for both proteins, the results are in generalagreement with the observed specificities. For quer-cetin, a substrate-like orientation of the flavonoidcould be determined in the active center of PFOMTwith reasonable binding energies, whereas no enzy-
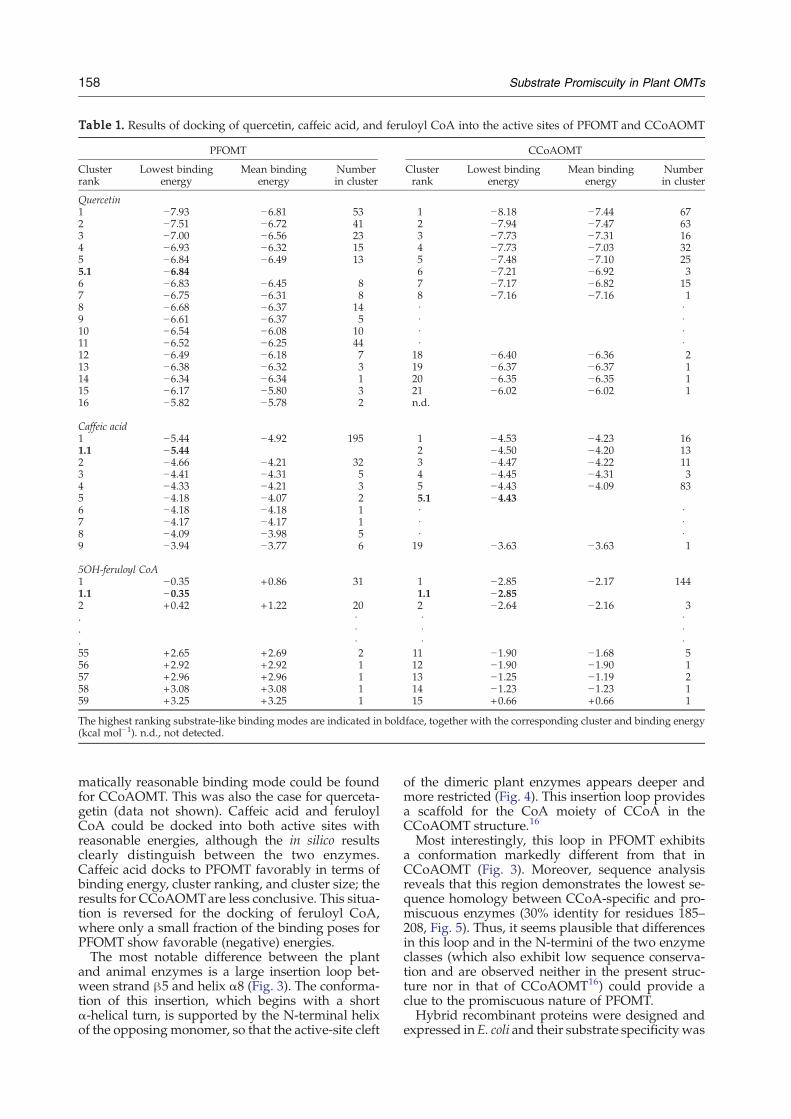
Table 1. Results of docking of quercetin, caffeic acid, and feruloyl CoA into the active sites of PFOMT and CCoAOMT
PFOMT CCoAOMT
Clusterrank
Lowest bindingenergy
Mean bindingenergy
Numberin cluster
Clusterrank
Lowest bindingenergy
Mean bindingenergy
Numberin cluster
Quercetin1 −7.93 −6.81 53 1 −8.18 −7.44 672 −7.51 −6.72 41 2 −7.94 −7.47 633 −7.00 −6.56 23 3 −7.73 −7.31 164 −6.93 −6.32 15 4 −7.73 −7.03 325 −6.84 −6.49 13 5 −7.48 −7.10 255.1 −6.84 6 −7.21 −6.92 36 −6.83 −6.45 8 7 −7.17 −6.82 157 −6.75 −6.31 8 8 −7.16 −7.16 18 −6.68 −6.37 14 · ·9 −6.61 −6.37 5 · ·10 −6.54 −6.08 10 · ·11 −6.52 −6.25 44 · ·12 −6.49 −6.18 7 18 −6.40 −6.36 213 −6.38 −6.32 3 19 −6.37 −6.37 114 −6.34 −6.34 1 20 −6.35 −6.35 115 −6.17 −5.80 3 21 −6.02 −6.02 116 −5.82 −5.78 2 n.d.
Caffeic acid1 −5.44 −4.92 195 1 −4.53 −4.23 161.1 −5.44 2 −4.50 −4.20 132 −4.66 −4.21 32 3 −4.47 −4.22 113 −4.41 −4.31 5 4 −4.45 −4.31 34 −4.33 −4.21 3 5 −4.43 −4.09 835 −4.18 −4.07 2 5.1 −4.436 −4.18 −4.18 1 · ·7 −4.17 −4.17 1 · ·8 −4.09 −3.98 5 · ·9 −3.94 −3.77 6 19 −3.63 −3.63 1
5OH-feruloyl CoA1 −0.35 +0.86 31 1 −2.85 −2.17 1441.1 −0.35 1.1 −2.852 +0.42 +1.22 20 2 −2.64 −2.16 3. · · ·. · · ·. · · ·55 +2.65 +2.69 2 11 −1.90 −1.68 556 +2.92 +2.92 1 12 −1.90 −1.90 157 +2.96 +2.96 1 13 −1.25 −1.19 258 +3.08 +3.08 1 14 −1.23 −1.23 159 +3.25 +3.25 1 15 +0.66 +0.66 1
The highest ranking substrate-like binding modes are indicated in boldface, together with the corresponding cluster and binding energy(kcal mol−1). n.d., not detected.
158 Substrate Promiscuity in Plant OMTs
matically reasonable binding mode could be foundfor CCoAOMT. This was also the case for querceta-getin (data not shown). Caffeic acid and feruloylCoA could be docked into both active sites withreasonable energies, although the in silico resultsclearly distinguish between the two enzymes.Caffeic acid docks to PFOMT favorably in terms ofbinding energy, cluster ranking, and cluster size; theresults for CCoAOMTare less conclusive. This situa-tion is reversed for the docking of feruloyl CoA,where only a small fraction of the binding poses forPFOMT show favorable (negative) energies.The most notable difference between the plant
and animal enzymes is a large insertion loop bet-ween strand β5 and helix α8 (Fig. 3). The conforma-tion of this insertion, which begins with a shortα-helical turn, is supported by the N-terminal helixof the opposingmonomer, so that the active-site cleft
of the dimeric plant enzymes appears deeper andmore restricted (Fig. 4). This insertion loop providesa scaffold for the CoA moiety of CCoA in theCCoAOMT structure.16
Most interestingly, this loop in PFOMT exhibitsa conformation markedly different from that inCCoAOMT (Fig. 3). Moreover, sequence analysisreveals that this region demonstrates the lowest se-quence homology between CCoA-specific and pro-miscuous enzymes (30% identity for residues 185–208, Fig. 5). Thus, it seems plausible that differencesin this loop and in the N-termini of the two enzymeclasses (which also exhibit low sequence conserva-tion and are observed neither in the present struc-ture nor in that of CCoAOMT16) could provide aclue to the promiscuous nature of PFOMT.Hybrid recombinant proteins were designed and
expressed in E. coli and their substrate specificitywas

Fig. 5. Sequence alignment of promiscuous OMTs (from M. crystallinum and Stellaria longipes) and diverse, specificCCoAOMTs, together with the mammalian rOMT. Secondary structure elements of PFOMT are shown, colored as in Fig.2 (monomer B). Boxed are the variable N-terminal residues (in blue) and the ‘insertion loop’ (in orange), which showsstructural and sequence divergence between the promiscuous and specific enzymes.
159Substrate Promiscuity in Plant OMTs
characterized to investigate the influence of thesetwo regions on substrate preference (Fig. 6). We havepreviously shown that the N-terminus of PFOMTplays a crucial role in determining the position spe-cificity and kinetic properties of the enzyme.18 A newconstruct wasmadewith a cleavable His tag in orderto study the influence of the N-terminal His tag.Removal of the His tag from native PFOMT (ΔHis-PFOMT, Table 2) resulted in a significant loss ofcatalytic efficiency toward the substrates caffeic acidand CCoA, largely due to a reduction in Km app,while turnover of the flavonoids was largelyunaffected. In contrast, replacement of the PFOMTN-terminus with that of the specific CCoAOMTenzyme from alfalfa (N-term hybrid) resulted in areduced catalytic efficiency toward flavonoids, aneffect that is even more pronounced for the sub-strates caffeic acid, CCoA, and caffeoyl glucose. In allcases, a 10- to 100-fold increase in Km app reflects areduction in substrate affinity, which is the majorfactor contributing to the strongly reduced catalyticefficiency kcat/Km app. As with PFOMT, removal ofthe His tag from the N-terminal hybrid (ΔHis–N-termhybrid) resulted in a strong reduction in activitytoward caffeic acid and CCoA; interestingly, theΔHis–N-term hybrid exhibited an increased activity
toward quercetagetin, such that this enzyme has analmost exclusive flavonoid activity.Substitution of the insertion loop from the specific
alfalfa CCoAOMT in the nonspecific PFOMT (loophybrid) leads to a stark reduction in soluble proteinexpression (1 mg/ml instead of the usual 10 mg/ml), with most of the protein depositing as insolubleinclusion bodies (data not shown). The loop hybridexhibited a 10-fold increase in affinity for and acti-vity against CCoA and a marked reduction in affi-nity for the flavonoids; that is, the loop-hybridprotein behaves more like CCoAOMT than PFOMT.On the other hand, replacement of the loop resultedin little change in Km app for caffeoyl glucose,although a considerable loss in activity against thissubstrate was observed. Despite the strongly re-duced Km app for CCoA, the high affinity for caffeoylglucose continues to distinguish the loop hybridfrom the corresponding alfalfa protein.Variants in which both the N-terminal peptide and
the insertion loop of PFOMTwere replaced simulta-neously by those of the corresponding CCoAOMTsequence (double hybrid) failed to increase the affi-nity for the CoA ester; instead, they led to a smallreduction in affinity for all substrates. This is incontrast to the pronounced decrease in affinity ob-

Fig. 6. (a) Cartoon illustrating hybrid enzymes of thepromiscuous cation-dependent PFOMT and the M. sativaCCoAOMT. 1, PFOMT; 2, PFOMTafter removal of the Histag using Factor Xa (ΔHis-PFOMT); 3, PFOMT with N-terminal domain (blue) fromM. sativaCCoAOMT (N-termhybrid); 4, same as 3 after Factor Xa cleavage of His tag(ΔHis–N-term hybrid); 5, PFOMT with ‘insertion loop’(orange) from M. sativa CCoAOMT (loop hybrid); 6,PFOMT with combined M. sativa N-terminal domain and‘insertion loop’ (double hybrid); 7, M. sativa CCoAOMT.NheI indicates the restriction site used for the generationof the loop and double hybrids (see Materials andMethods) (b) SDS/PAGE showing purified recombinantcation-dependent OMT hybrids.
160 Substrate Promiscuity in Plant OMTs
served toward caffeic acid, CCoA, and caffeoylglucose for the N-terminal-alone hybrids.Based on the present results, the N-terminal resi-
dues of PFOMT would appear to be primarily res-ponsible for the activity toward flavonoids, whilethe intermediate loop plays a role in the positioningand methylation of CCoA. While none of the hybridvariants created exhibit the desired specificity pro-file of CCoAOMT, the catalytic efficiency of the loophybrid toward CCoA closely approaches this goal.The relaxed specificity of PFOMT appears to comefrom a novel combination of structural elements inthe neighborhood of the active site, in particularthrough the use of a flexible N-terminal peptide.The modest change in specificity achieved through
recombining surface loops is in line with otherattempts to redesign enzymes by rational means.21
While we do not know the reasons for this forPFOMT at present, possible explanations couldcome from unpredictable changes in the scaffoldconformation, changes in protein stability, and/orchanges in the dynamic behavior of the hybrid en-
zymes. We have previously observed all these phe-nomena for variants of trypsin designed to behavelike the closely related serine proteinase Factor Xa.22,23
Clearly, more experimental data are required tounderstand the complex mechanisms driving pro-tein–ligand affinity and the evolution of substratespecificity. Results from directed evolution mayyield important clues for future tailoring of enzy-matic reactions.24,25
Conclusion and Summary
The structure determination of the promiscuousmetal-dependent OMT PFOMT reveals a dimericprotein with a high three-dimensional homology tothe CCoA-specific enzyme OMT CCoAOMT. Basedon the structures, hybrid proteins were designedthrough the exchange of N-terminal residues and ofa loop region representing an insertion with respectto the mammalian catechol OMT. The resulting loophybrids exhibited greater catalytic efficiency towardCCoA, while the N-terminal hybrids possessedlower affinity for nonflavonoid substrates. Althoughthe overall structures of the PFOMTand CCoAOMTappear highly similar, our results show that combi-nations of two variable domains can afford novelenzymes of distinct kinetic properties. With thecompletion of several genome projects, it is apparentthat plants contain several members of apparentlyredundant, but conserved CCoAOMT-like genes.The results presented here suggest that other mem-bers of this class may serve functions distinct tomethylation of the single lignin precursor, CCoA.
Materials and Methods
Crystallization of the recombinant full-lengthHis-tagged protein
Recombinant PFOMT was produced and affinity puri-fied as described previously.18 The protein was concen-trated and rebuffered into 10 mM Tris/HCl, pH 7.5,centrifuged to remove any precipitant, and diluted to aconcentration of 6 mg/ml with 10 mM Tris/HCl, pH 7.0.Crystals were obtained after optimization with 20%polyethylene glycol 4000 and 0.2 M CaCl2 in 100 mMHepes/NaOH, pH 7.0, using a concentration of 3 mg/mlPFOMT, 250 μM MgCl2, 250 μM SAM, 25 μM quercetin,and 2.5% dimethyl sulfoxide through the sitting dropmethod. For the production of selenomethionine (SeMet)-labeled protein for phase determination, bacteria weregrown in the presence of SeMet (50 mg/l) medium asdescribed previously26 with a yield of 10.5 mg/l of cul-ture. SeMet crystals were obtained under the same con-ditions as wild type using a concentration of 2 mg/mlSeMet-PFOMT.
Structure determination of PFOMT
Crystals of the native PFOMT were measured at theDeutsches Elektronen-Synchrotron facility (BW6Hamburg,Germany). Initial PFOMT crystals with the orthorhombic
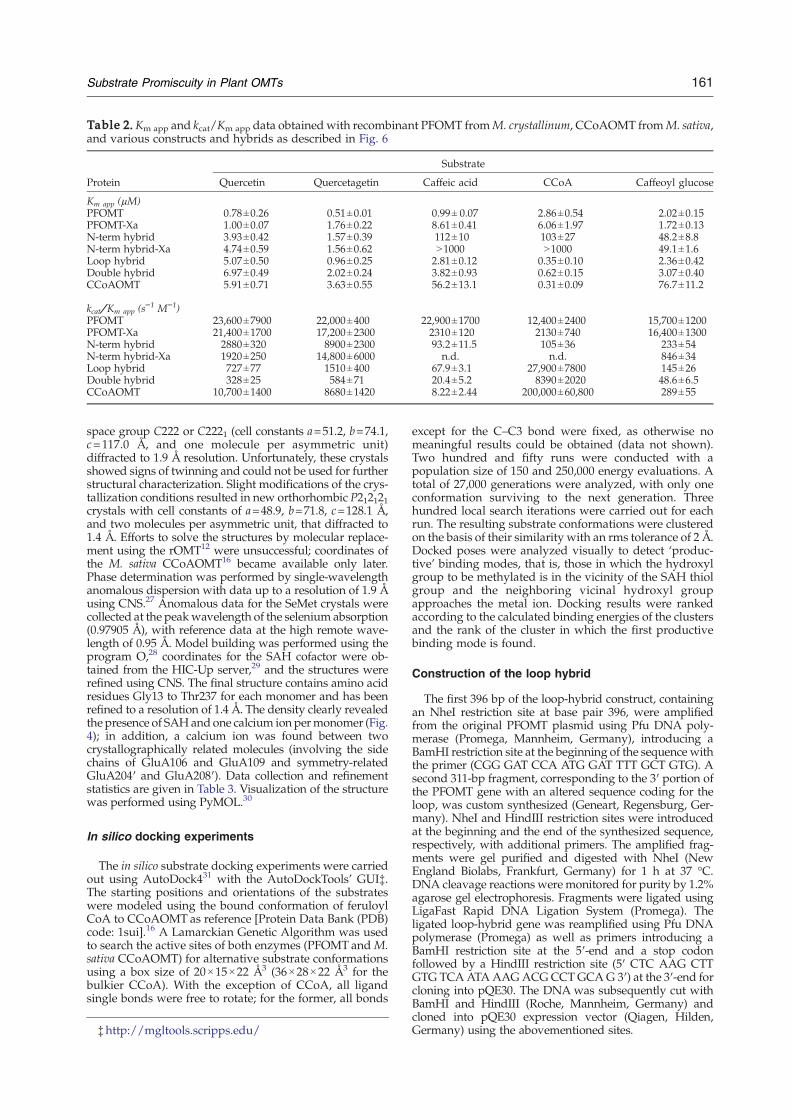
Table 2. Km app and kcat/Km app data obtained with recombinant PFOMT fromM. crystallinum, CCoAOMT fromM. sativa,and various constructs and hybrids as described in Fig. 6
Protein
Substrate
Quercetin Quercetagetin Caffeic acid CCoA Caffeoyl glucose
Km app (μM)PFOMT 0.78±0.26 0.51±0.01 0.99± 0.07 2.86±0.54 2.02±0.15PFOMT-Xa 1.00±0.07 1.76±0.22 8.61±0.41 6.06±1.97 1.72±0.13N-term hybrid 3.93±0.42 1.57±0.39 112±10 103±27 48.2±8.8N-term hybrid-Xa 4.74±0.59 1.56±0.62 N1000 N1000 49.1±1.6Loop hybrid 5.07±0.50 0.96±0.25 2.81±0.12 0.35±0.10 2.36±0.42Double hybrid 6.97±0.49 2.02±0.24 3.82±0.93 0.62±0.15 3.07±0.40CCoAOMT 5.91±0.71 3.63±0.55 56.2±13.1 0.31±0.09 76.7±11.2
kcat/Km app (s−1 M−1)
PFOMT 23,600±7900 22,000±400 22,900±1700 12,400±2400 15,700±1200PFOMT-Xa 21,400±1700 17,200±2300 2310±120 2130±740 16,400±1300N-term hybrid 2880±320 8900±2300 93.2±11.5 105±36 233±54N-term hybrid-Xa 1920±250 14,800±6000 n.d. n.d. 846±34Loop hybrid 727±77 1510±400 67.9±3.1 27,900±7800 145±26Double hybrid 328±25 584±71 20.4±5.2 8390±2020 48.6±6.5CCoAOMT 10,700±1400 8680±1420 8.22±2.44 200,000±60,800 289±55
161Substrate Promiscuity in Plant OMTs
space group C222 or C2221 (cell constants a=51.2, b=74.1,c=117.0 Å, and one molecule per asymmetric unit)diffracted to 1.9 Å resolution. Unfortunately, these crystalsshowed signs of twinning and could not be used for furtherstructural characterization. Slight modifications of the crys-tallization conditions resulted in new orthorhombic P212121crystals with cell constants of a=48.9, b=71.8, c=128.1 Å,and two molecules per asymmetric unit, that diffracted to1.4 Å. Efforts to solve the structures by molecular replace-ment using the rOMT12 were unsuccessful; coordinates ofthe M. sativa CCoAOMT16 became available only later.Phase determination was performed by single-wavelengthanomalous dispersion with data up to a resolution of 1.9 Åusing CNS.27 Anomalous data for the SeMet crystals werecollected at the peakwavelength of the selenium absorption(0.97905 Å), with reference data at the high remote wave-length of 0.95 Å. Model building was performed using theprogram O,28 coordinates for the SAH cofactor were ob-tained from the HIC-Up server,29 and the structures wererefined using CNS. The final structure contains amino acidresidues Gly13 to Thr237 for each monomer and has beenrefined to a resolution of 1.4 Å. The density clearly revealedthe presence of SAHand one calcium ion permonomer (Fig.4); in addition, a calcium ion was found between twocrystallographically related molecules (involving the sidechains of GluA106 and GluA109 and symmetry-relatedGluA204′ and GluA208′). Data collection and refinementstatistics are given in Table 3. Visualization of the structurewas performed using PyMOL.30
In silico docking experiments
The in silico substrate docking experiments were carriedout using AutoDock431 with the AutoDockTools' GUI‡.The starting positions and orientations of the substrateswere modeled using the bound conformation of feruloylCoA to CCoAOMT as reference [Protein Data Bank (PDB)code: 1sui].16 A Lamarckian Genetic Algorithm was usedto search the active sites of both enzymes (PFOMT andM.sativa CCoAOMT) for alternative substrate conformationsusing a box size of 20×15×22 Å3 (36×28×22 Å3 for thebulkier CCoA). With the exception of CCoA, all ligandsingle bonds were free to rotate; for the former, all bonds
‡http://mgltools.scripps.edu/
except for the C–C3 bond were fixed, as otherwise nomeaningful results could be obtained (data not shown).Two hundred and fifty runs were conducted with apopulation size of 150 and 250,000 energy evaluations. Atotal of 27,000 generations were analyzed, with only oneconformation surviving to the next generation. Threehundred local search iterations were carried out for eachrun. The resulting substrate conformations were clusteredon the basis of their similarity with an rms tolerance of 2 Å.Docked poses were analyzed visually to detect ‘produc-tive’ binding modes, that is, those in which the hydroxylgroup to be methylated is in the vicinity of the SAH thiolgroup and the neighboring vicinal hydroxyl groupapproaches the metal ion. Docking results were rankedaccording to the calculated binding energies of the clustersand the rank of the cluster in which the first productivebinding mode is found.
Construction of the loop hybrid
The first 396 bp of the loop-hybrid construct, containingan NheI restriction site at base pair 396, were amplifiedfrom the original PFOMT plasmid using Pfu DNA poly-merase (Promega, Mannheim, Germany), introducing aBamHI restriction site at the beginning of the sequence withthe primer (CGG GAT CCA ATG GAT TTT GCT GTG). Asecond 311-bp fragment, corresponding to the 3′ portion ofthe PFOMT gene with an altered sequence coding for theloop, was custom synthesized (Geneart, Regensburg, Ger-many). NheI and HindIII restriction sites were introducedat the beginning and the end of the synthesized sequence,respectively, with additional primers. The amplified frag-ments were gel purified and digested with NheI (NewEngland Biolabs, Frankfurt, Germany) for 1 h at 37 °C.DNA cleavage reactions were monitored for purity by 1.2%agarose gel electrophoresis. Fragments were ligated usingLigaFast Rapid DNA Ligation System (Promega). Theligated loop-hybrid gene was reamplified using Pfu DNApolymerase (Promega) as well as primers introducing aBamHI restriction site at the 5′-end and a stop codonfollowed by a HindIII restriction site (5′ CTC AAG CTTGTG TCAATAAAGACGCCTGCAG 3′) at the 3′-end forcloning into pQE30. The DNA was subsequently cut withBamHI and HindIII (Roche, Mannheim, Germany) andcloned into pQE30 expression vector (Qiagen, Hilden,Germany) using the abovementioned sites.
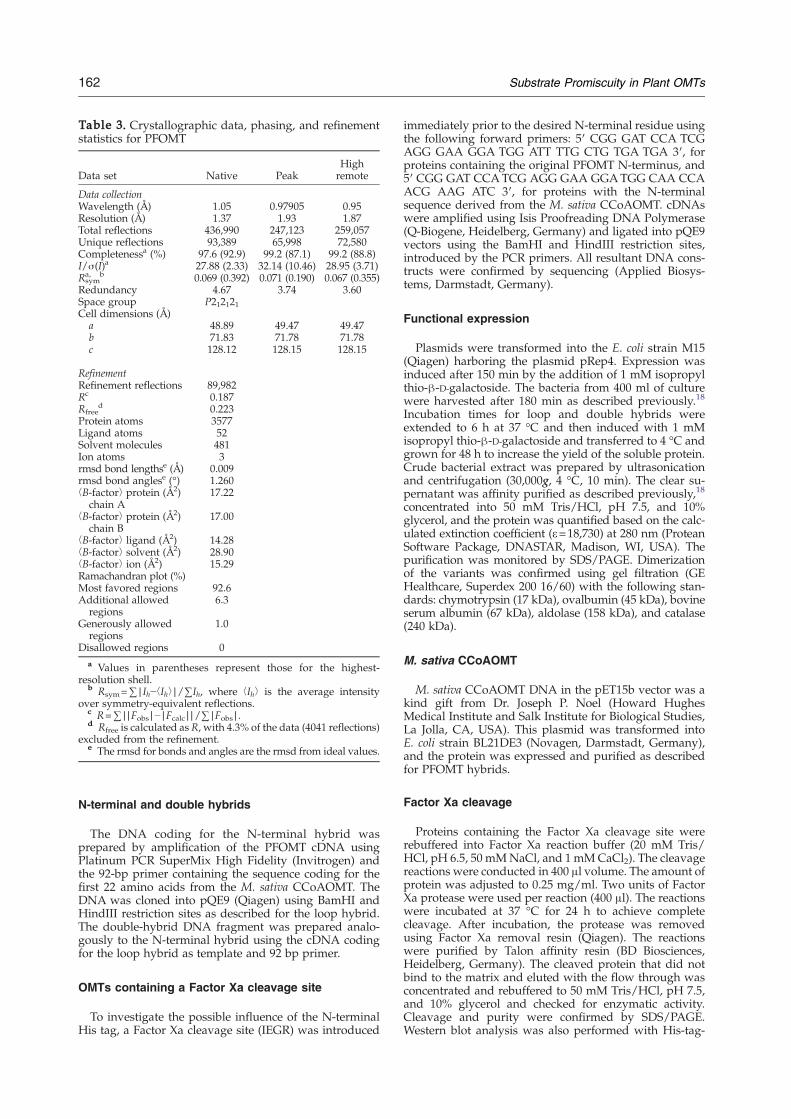
Table 3. Crystallographic data, phasing, and refinementstatistics for PFOMT
Data set Native PeakHighremote
Data collectionWavelength (Å) 1.05 0.97905 0.95Resolution (Å) 1.37 1.93 1.87Total reflections 436,990 247,123 259,057Unique reflections 93,389 65,998 72,580Completenessa (%) 97.6 (92.9) 99.2 (87.1) 99.2 (88.8)I/σ(I)a 27.88 (2.33) 32.14 (10.46) 28.95 (3.71)Rsyma, b 0.069 (0.392) 0.071 (0.190) 0.067 (0.355)
Redundancy 4.67 3.74 3.60Space group P212121Cell dimensions (Å)
a 48.89 49.47 49.47b 71.83 71.78 71.78c 128.12 128.15 128.15
RefinementRefinement reflections 89,982Rc 0.187Rfree
d 0.223Protein atoms 3577Ligand atoms 52Solvent molecules 481Ion atoms 3rmsd bond lengthse (Å) 0.009rmsd bond anglese (°) 1.260⟨B-factor⟩ protein (Å2)
chain A17.22
⟨B-factor⟩ protein (Å2)chain B
17.00
⟨B-factor⟩ ligand (Å2) 14.28⟨B-factor⟩ solvent (Å2) 28.90⟨B-factor⟩ ion (Å2) 15.29Ramachandran plot (%)Most favored regions 92.6Additional allowed
regions6.3
Generously allowedregions
1.0
Disallowed regions 0a Values in parentheses represent those for the highest-
resolution shell.b Rsym=∑|Ih−⟨Ih⟩|/∑Ih, where ⟨Ih⟩ is the average intensity
over symmetry-equivalent reflections.c R=∑||Fobs|−|Fcalc||/∑|Fobs|.d Rfree is calculated as R, with 4.3% of the data (4041 reflections)
excluded from the refinement.e The rmsd for bonds and angles are the rmsd from ideal values.
162 Substrate Promiscuity in Plant OMTs
N-terminal and double hybrids
The DNA coding for the N-terminal hybrid wasprepared by amplification of the PFOMT cDNA usingPlatinum PCR SuperMix High Fidelity (Invitrogen) andthe 92-bp primer containing the sequence coding for thefirst 22 amino acids from the M. sativa CCoAOMT. TheDNA was cloned into pQE9 (Qiagen) using BamHI andHindIII restriction sites as described for the loop hybrid.The double-hybrid DNA fragment was prepared analo-gously to the N-terminal hybrid using the cDNA codingfor the loop hybrid as template and 92 bp primer.
OMTs containing a Factor Xa cleavage site
To investigate the possible influence of the N-terminalHis tag, a Factor Xa cleavage site (IEGR) was introduced
immediately prior to the desired N-terminal residue usingthe following forward primers: 5′ CGG GAT CCA TCGAGG GAA GGA TGG ATT TTG CTG TGA TGA 3′, forproteins containing the original PFOMT N-terminus, and5′ CGG GAT CCATCG AGG GAA GGATGG CAA CCAACG AAG ATC 3′, for proteins with the N-terminalsequence derived from the M. sativa CCoAOMT. cDNAswere amplified using Isis Proofreading DNA Polymerase(Q-Biogene, Heidelberg, Germany) and ligated into pQE9vectors using the BamHI and HindIII restriction sites,introduced by the PCR primers. All resultant DNA cons-tructs were confirmed by sequencing (Applied Biosys-tems, Darmstadt, Germany).
Functional expression
Plasmids were transformed into the E. coli strain M15(Qiagen) harboring the plasmid pRep4. Expression wasinduced after 150 min by the addition of 1 mM isopropylthio-β-D-galactoside. The bacteria from 400 ml of culturewere harvested after 180 min as described previously.18
Incubation times for loop and double hybrids wereextended to 6 h at 37 °C and then induced with 1 mMisopropyl thio-β-D-galactoside and transferred to 4 °C andgrown for 48 h to increase the yield of the soluble protein.Crude bacterial extract was prepared by ultrasonicationand centrifugation (30,000g, 4 °C, 10 min). The clear su-pernatant was affinity purified as described previously,18
concentrated into 50 mM Tris/HCl, pH 7.5, and 10%glycerol, and the protein was quantified based on the calc-ulated extinction coefficient (ε=18,730) at 280 nm (ProteanSoftware Package, DNASTAR, Madison, WI, USA). Thepurification was monitored by SDS/PAGE. Dimerizationof the variants was confirmed using gel filtration (GEHealthcare, Superdex 200 16/60) with the following stan-dards: chymotrypsin (17 kDa), ovalbumin (45 kDa), bovineserum albumin (67 kDa), aldolase (158 kDa), and catalase(240 kDa).
M. sativa CCoAOMT
M. sativa CCoAOMT DNA in the pET15b vector was akind gift from Dr. Joseph P. Noel (Howard HughesMedical Institute and Salk Institute for Biological Studies,La Jolla, CA, USA). This plasmid was transformed intoE. coli strain BL21DE3 (Novagen, Darmstadt, Germany),and the protein was expressed and purified as describedfor PFOMT hybrids.
Factor Xa cleavage
Proteins containing the Factor Xa cleavage site wererebuffered into Factor Xa reaction buffer (20 mM Tris/HCl, pH 6.5, 50 mMNaCl, and 1 mMCaCl2). The cleavagereactions were conducted in 400 μl volume. The amount ofprotein was adjusted to 0.25 mg/ml. Two units of FactorXa protease were used per reaction (400 μl). The reactionswere incubated at 37 °C for 24 h to achieve completecleavage. After incubation, the protease was removedusing Factor Xa removal resin (Qiagen). The reactionswere purified by Talon affinity resin (BD Biosciences,Heidelberg, Germany). The cleaved protein that did notbind to the matrix and eluted with the flow through wasconcentrated and rebuffered to 50 mM Tris/HCl, pH 7.5,and 10% glycerol and checked for enzymatic activity.Cleavage and purity were confirmed by SDS/PAGE.Western blot analysis was also performed with His-tag-

163Substrate Promiscuity in Plant OMTs
specific antibodies (Qiagen) in order to verify the absenceof any His tag.
Enzyme activity tests and kinetic data
Enzyme assays were performed in a buffer containing100 mM K2PO4, pH 7.5, and 10% glycerol with 10 μMsubstrate (dissolved in 30% dimethyl sulfoxide), 0.5–2 μgof total protein, and 400 μMSAM in a total volume of 50 μl.The assays were incubated at 30 °C for 60 to 3600 s(dependent on the protein and substrate tested) and thenstopped by the addition of 20 μl of 7% trichloroacetic acidin 50% acetonitrile/water. The reaction products wereanalyzed by HPLC as described previously.17 Caffeoylglucose was prepared as described previously from caffeicacid and UDP-glucose with the purified recombinantsinapic acid glucosyltransferase from Brassica napus.32
CCoA was prepared based on published methods.33,34
Caffeic acid was obtained from Serva (Heidelberg, Ger-many), quercetagetin was obtained from Extrasynthese(Genay, France), and quercetin was obtained from Roth(Karlsruhe, Germany). The reaction products were ana-lyzed by reversed-phase liquid chromatography on aNucleosil 5-μm C18 column (50 mm length×4 mm innerdiameter; Macherey and Nagel, Düren, Germany), asdescribed previously.17 Compounds were analyzed withlinear gradients from 10% B (100% acetonitrile) in A (1.5%aqueous phosphoric acid) to 70% B in A (for phenolics),from 5% B to 50% B in A (for free acids and CoA esters),from 5% B to 30% B (for glucose esters), and from 20% Bin A to 80% B in A in 4 min (for flavonoids) at a flow rateof 1 ml min−1. Detection of flavonoids, catechol, coumar-ins, and hydroxycinnamic acid esters was performedbetween 260 and 400 nm. Identification and quantificationwas achieved with reference compounds from our Ins-titute collection. For Km determination of methyl groupacceptors, acceptor concentrations were chosen between 2and 20 μM, while SAM was kept constant at 1.5 mM. Km,Vmax, and kcat values were calculated by nonlinear curvefitting, assuming Michaelis–Menten steady state kinetics.All enzyme assays were recorded in triplicate for eachexperiment.
PDB accession code
Coordinates and structure factors have been depositedin the PDB with accession code 3C3Y.
Acknowledgements
We thank Jean-Luc Ferrer (Grenoble) and JosephP. Noel (San Diego) for providing the M. sativaCCoAOMT plasmid and information prior to pub-lication and Norbert Sträter (Leipzig) for the use ofhis X-ray facilities at the initial stages of this project.Diffraction data were measured on the MPG/GBFbeamline BW6 at Deutsches Elektronen-Synchro-tron, where we gratefully acknowledge the assis-tance of Gleb Bourenkov and Hans Bartunik. Thiswork was in part supported by grants VO719/5and STU297/4 from the Deutsche Forschungsge-meinschaft as well as the SchwerpunktprogrammSPP1152 ‘Evolution metabolischer Diversität.’
References
1. Zhu, B. T., Ezell, E. L. & Liehr, J. G. (1994). Catechol-O-methyltransferase-catalyzed rapid O-methylation ofmutagenic flavonoids. Metabolic inactivation as apossible reason for their lack of carcinogenicity in vivo.J. Biol. Chem. 269, 292–299.
2. Ye, Z. H., Kneusel, R. E., Matern, U. & Varner, J. E.(1994). An alternative methylation pathway in ligninbiosynthesis in zinnia. Plant Cell, 6, 1427–1439.
3. Maury, S., Geoffroy, P. & Legrand, M. (1999). TobaccoO-methyltransferases involved in phenylpropanoid meta-bolism. The different caffeoyl coenzyme A/5-hydroxy-feruloyl–coenzyme A 3/5-O-methyltransferase andcaffeic acid/5-hydroxyferulic acid 3/5-O-methyl-transferase classes have distinct substrate specificitiesand expression patterns. Plant Physiol. 121, 215–224.
4. Osakabe, K., Tsao, C. C., Li, L., Popko, J. L., Umezawa,T., Carraway, D. T. et al. (1999). Coniferyl aldehyde 5-hydroxylation and methylation direct syringyl ligninbiosynthesis in angiosperms. Proc. Natl Acad. Sci. USA,96, 8955–8960.
5. Inoue, K., Sewalt, V. J. H., Murray Ballance, G., Ni, W.,Sturzer, C. & Dixon, R. A. (1998). Developmentalexpression and substrate specificities of alfalfa caffeicacid 3-O-methyltransferase and caffeoyl coenzyme A3-O-methyltransferase in relation to lignification. PlantPhysiol. 117, 761–770.
6. Humphreys, J. M. & Chapple, C. (2002). Rewriting thelignin roadmap. Curr. Opin. Plant Biol. 5, 224–229.
7. Peter, G. & Neale, D. (2004). Molecular basis for theevolution of xylem lignification. Curr. Opin. Plant Biol.7, 737–742.
8. Zubieta, C., He, X. Z., Dixon, R. A. & Noel, J. P. (2001).Structures of two natural product methyltransferasesreveal the basis for substrate specificity in plant O-methyltransferases. Nat. Struct. Mol. Biol. 8, 271–279.
9. Zubieta, C., Kota, P., Ferrer, J. L., Dixon, R. A. & Noel,J. P. (2002). Structural basis for the modulation oflignin monomer methylation by caffeic acid/5-hydro-xyferulic acid 3/5-O-methyltransferase. Plant Cell, 14,1265–1277.
10. Zubieta, C., Ross, J. R., Koscheski, P., Yang, Y.,Pichersky, E. & Noel, J. P. (2003). Structural basis forsubstrate recognition in the salicylic acid carboxylmethyltransferase family. Plant Cell, 15, 1704–1716.
11. Yang, H., Ahn, J. H., Ibrahim, R. K., Lee, S. & Lim, Y.(2004). The three-dimensional structure of Arabidopsisthaliana O-methyltransferase predicted by homology-based modelling. J. Mol. Graphics Modell. 23, 77–87.
12. Vidgren, J., Svensson, L. A. & Liljas, A. (1994). Crystalstructure of catecholO-methyltransferase. Nature, 368,354–358.
13. Mannisto, P. T. & Kaakkola, S. (1999). Catechol-O-methyltransferase (COMT): biochemistry, molecularbiology, pharmacology, and clinical efficacy of thenew selective COMT inhibitors. Pharmacol. Rev. 51,593–628.
14. Schluckebier, G., O'Gara, M., Saenger, W. & Cheng, X.(1995). Universal catalytic domain structure of Ado-Met-dependent methyltransferases. J. Mol. Biol. 247,16–20.
15. Vidgren, J. (1998). X-ray crystallography of catecholO-methyltransferase: perspectives for target-baseddrug development. Adv. Pharmacol. 42, 328–330.
16. Ferrer, J. L., Zubieta, C., Dixon, R. A. & Noel, J. P.(2005). Crystal structures of alfalfa caffeoyl coenzymeA 3-O-methyltransferase. Plant Physiol. 137, 1009–1017.
17. Ibdah, M., Zhang, X. H., Schmidt, J. & Vogt, T. (2003).

164 Substrate Promiscuity in Plant OMTs
A novel Mg++-dependent O-methyltransferase in thephenylpropanoid metabolism of Mesembryanthemumcrystallinum. J. Biol. Chem. 278, 43961–43972.
18. Vogt, T. (2004). Regiospecificity and kinetic propertiesof a plant natural product O-methyl-transferase aredetermined by its N-terminal domain. FEBS Lett. 561,159–162.
19. Lukacin, R., Matern, U., Specker, S. & Vogt, T. (2004).Cations modulate the substrate specificity of bifunc-tional class I O-methyltransferase from Ammi majus.FEBS Lett. 577, 367–370.
20. Hou, X. W., Wang, Y. L., Zhou, Z. W., Bao, S. L., Lin,Y. J. & Gong, W. M. (2007). Crystal structure ofSAM-dependent O-methyltransferase from pathoge-nic bacterium Leptospira interrogans. J. Struct. Biol. 159,523–528.
21. Kohara, A., Nakajima, C., Yoshida, S. & Muranaka, T.(2007). Characterisation and engineering of glycosyl-transferases responsible for steroid saponin biosynth-esis in Solanaceous plants. Phytochemistry, 68, 478–486.
22. Reyda, S., Sohn, C., Klebe, G., Rall, K., Ullmann, D.,Jakubke, H.-D. & Stubbs, M. T. (2003). Reconstructingthe binding site of factor Xa in trypsin reveals ligand-induced structural plasticity. J. Mol. Biol. 325, 963–977.
23. Rauh, D., Klebe, G. & Stubbs, M. T. (2004). Under-standing protein–ligand interactions: the price ofprotein flexibility. J. Mol. Biol. 335, 1325–1341.
24. Aharoni, A., Gaidukov, L., Khersonsky, O., McQGould, S., Roodveldt, C. & Tawfik, D. S. (2005). The‘evolvability’ of promiscuous protein functions. Nat.Genet. 37, 73–76.
25. Williams, G. J., Zhang, C. & Thorson, J. S. (2007).Expanding the promiscuity of a natural product gly-cosyltransferase by directed evolution.Nat. Chem. Biol.3, 657–662.
26. Doublie, S. (1997). Preparation of selenomethionylproteins for phase determination. Methods Enzymol.276, 523–530.
27. Brünger, A., Adams, P., Clore, G., DeLano, W., Gros,P., Grosse-Kunstleve, R. et al. (1998). Crystallographyand NMR system: a new software suite for macro-molecular structure determination. Acta Crystallogr.,Sect. D: Biol. Crystallogr. 54, 905–921.
28. Jones, T. A., Zou, J. Y. & Cowan, S. W. (1991). Im-proved methods for building protein models in elec-tron density maps and the location of errors in thesemodels.Acta Crystallogr., Sect. A: Found. Crystallogr. 47,110–119.
29. Kleywegt, G. J. & Jones, T. A. (1998). Databases inprotein crystallography. Acta Crystallogr., Sect. D: Biol.Crystallogr. 54, 1119–1131.
30. DeLano, W. (2002). The PyMOL Molecular GraphicsSystem, DeLano Scientific, Palo Alto, CA.
31. Morris, G. M., Goodsell, D. S., Halliday, R. S., Huey,R., Hart, W. E., Belew, R. K. & Olson, A. J. (1998).Automated docking using a Lamarckian GeneticAlgorithm and empirical binding free energy func-tion. J. Comput. Chem. 19, 1639–1662.
32. Milkowski, C., Baumert, A. & Strack, D. (2000). Clon-ing and heterologous expression of a rape cDNAencoding UDP-glucose:sinapate glucosyltransferase.Planta, 211, 883–886.
33. Strack, D., Keller, H. & Weissenböck, G. (1987).Enzymatic synthesis of hydroxycinnamic acid estersof sugar acids and hydroaromatic acids by proteinpreparations of Raphanus sativus. J. Plant Physiol. 131,61–73.
34. Stöckigt, J. & Zenk, M. H. (1975). Chemical synthesisand properties of hydroxycinnamoyl coenzyme Aderivatives. Z. Naturforsch., C: Biosci. 30, 352–358.




















