
Barrier to gene flow between two ecologically divergent Populus species, P. alba (white poplar) and...
13
Molecular Ecology (2005) 14, 1045–1057 doi: 10.1111/j.1365-294X.2005.02469.x © 2005 Blackwell Publishing Ltd Blackwell Publishing, Ltd. Barrier to gene flow between two ecologically divergent Populus species, P. alba (white poplar) and P. tremula (European aspen): the role of ecology and life history in gene introgression C. LEXER,* M. F. FAY,* J. A. JOSEPH,* M.-S. NICA † and B. HEINZE † *Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3DS, UK, †Department of Genetics, Federal Office and Research Centre for Forests, Hauptstrasse 7, A-1140 Vienna, Austria Abstract The renewed interest in the use of hybrid zones for studying speciation calls for the identification and study of hybrid zones across a wide range of organisms, especially in long-lived taxa for which it is often difficult to generate interpopulation variation through controlled crosses. Here, we report on the extent and direction of introgression between two members of the ‘model tree’ genus Populus: Populus alba (white poplar) and Populus tremula (European aspen), across a large zone of sympatry located in the Danube valley. We geno- typed 93 hybrid morphotypes and samples from four parental reference populations from within and outside the zone of sympatry for a genome-wide set of 20 nuclear micros- atellites and eight plastid DNA restriction site polymorphisms. Our results indicate that introgression occurs preferentially from P. tremula to P. alba via P. tremula pollen. This unidirectional pattern is facilitated by high levels of pollen vs. seed dispersal in P. tremula (pollen/seed flow = 23.9) and by great ecological opportunity in the lowland floodplain forest in proximity to P. alba seed parents, which maintains gene flow in the direction of P. alba despite smaller effective population sizes (N e ) in this species (P. alba N e c. 500 – 550; P. tremula N e c. 550 – 700). Our results indicate that hybrid zones will be valuable tools for studying the genetic architecture of the barrier to gene flow between these two ecologically divergent Populus species. Keywords: admixture, dispersal, ecological divergence, hybridization, microsatellites, plastid DNA Received 30 September 2004; revision received 10 December 2004; accepted 10 December 2004 Introduction The genetic architecture of barriers to gene flow plays a central role in molecular ecology and evolutionary genetics. It holds the key to reconstructing the sequence of genetic changes that accompany or facilitate speciation (Bradshaw et al . 1995; Rieseberg et al . 1999; Orr 2001). Genetic architec- ture also allows assessment of the role of hybrid zones as evolutionary filters: hybrid zones may permit introgression of beneficial genes into a new genetic background, while preventing the movement of detrimental genes across species boundaries (Barton & Hewitt 1985; Barton & Gale 1993; Martinsen et al . 2001). At the within-species level, studying genetic architecture provides information about the potential of gene flow and selection to facilitate the spread of advantageous alleles across populations and thereby to maintain species as cohesive units (Rieseberg & Morjan 2004), and about the role of natural selection in promoting population divergence (Wilding et al . 2001; Luikart et al . 2003; Campbell and Bernatchez 2004). Hybrid zones have long been viewed as ‘natural labor- atories’ for studying barriers to gene flow (Barton & Hewitt 1985; Harrison 1990; Barton & Gale 1993). The fascination of molecular ecologists for hybrid zones stems, in part, from the fact that hybrid zones are often composed of a wide variety of genotypes resulting from many generations of recombination. Moreover, linkage disequilibrium (LD) induced by hybridization is thought to facilitate studies of microevolutionary processes (Rieseberg et al . 1999; Rogers Correspondence: Christian Lexer, Fax: +44 (0)20 83325310; E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Barrier to gene flow between two ecologically divergent Populus species, P. alba (white poplar) and...

Molecular Ecology (2005)
14
, 1045–1057 doi: 10.1111/j.1365-294X.2005.02469.x
© 2005 Blackwell Publishing Ltd
Blackwell Publishing, Ltd.
Barrier to gene flow between two ecologically divergent
Populus
species,
P. alba
(white poplar) and
P. tremula
(European aspen): the role of ecology and life history in gene introgression
C . LEXER,
*
M. F . FAY,
*
J . A . JOSEPH,
*
M. -S . NICA
†
and B . HEINZE
†
*
Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3DS, UK,
†
Department of Genetics, Federal Office and Research Centre for Forests, Hauptstrasse 7, A-1140 Vienna, Austria
Abstract
The renewed interest in the use of hybrid zones for studying speciation calls for theidentification and study of hybrid zones across a wide range of organisms, especially inlong-lived taxa for which it is often difficult to generate interpopulation variation throughcontrolled crosses. Here, we report on the extent and direction of introgression between twomembers of the ‘model tree’ genus
Populus
:
Populus alba
(white poplar) and
Populus tremula
(European aspen), across a large zone of sympatry located in the Danube valley. We geno-typed 93 hybrid morphotypes and samples from four parental reference populationsfrom within and outside the zone of sympatry for a genome-wide set of 20 nuclear micros-atellites and eight plastid DNA restriction site polymorphisms. Our results indicate thatintrogression occurs preferentially from
P. tremula
to
P. alba
via
P. tremula
pollen. Thisunidirectional pattern is facilitated by high levels of pollen vs. seed dispersal in
P. tremula
(pollen/seed flow = 23.9) and by great ecological opportunity in the lowland floodplainforest in proximity to
P. alba
seed parents, which maintains gene flow in the direction of
P. alba
despite smaller effective population sizes (
N
e
) in this species (
P. alba N
e
c.
500–550;
P. tremula N
e
c.
550–700). Our results indicate that hybrid zones will be valuable tools forstudying the genetic architecture of the barrier to gene flow between these two ecologicallydivergent
Populus
species.
Keywords
: admixture, dispersal, ecological divergence, hybridization, microsatellites, plastid DNA
Received 30 September 2004; revision received 10 December 2004; accepted 10 December 2004
Introduction
The genetic architecture of barriers to gene flow plays acentral role in molecular ecology and evolutionary genetics.It holds the key to reconstructing the sequence of geneticchanges that accompany or facilitate speciation (Bradshaw
et al
. 1995; Rieseberg
et al
. 1999; Orr 2001). Genetic architec-ture also allows assessment of the role of hybrid zones asevolutionary filters: hybrid zones may permit introgressionof beneficial genes into a new genetic background, whilepreventing the movement of detrimental genes acrossspecies boundaries (Barton & Hewitt 1985; Barton & Gale1993; Martinsen
et al
. 2001). At the within-species level,
studying genetic architecture provides information aboutthe potential of gene flow and selection to facilitate thespread of advantageous alleles across populations andthereby to maintain species as cohesive units (Rieseberg &Morjan 2004), and about the role of natural selection inpromoting population divergence (Wilding
et al
. 2001;Luikart
et al
. 2003; Campbell and Bernatchez 2004).Hybrid zones have long been viewed as ‘natural labor-
atories’ for studying barriers to gene flow (Barton & Hewitt1985; Harrison 1990; Barton & Gale 1993). The fascinationof molecular ecologists for hybrid zones stems, in part,from the fact that hybrid zones are often composed of awide variety of genotypes resulting from many generationsof recombination. Moreover, linkage disequilibrium (LD)induced by hybridization is thought to facilitate studies ofmicroevolutionary processes (Rieseberg
et al
. 1999; Rogers
Correspondence: Christian Lexer, Fax: +44 (0)20 83325310; E-mail:[email protected]

1046
C . L E X E R
E T A L .
© 2005 Blackwell Publishing Ltd,
Molecular Ecology
, 14, 1045–1057
et al
. 2001; Rieseberg & Buerkle 2002), reminiscent of geneticmapping studies in admixed human populations (Briscoe
et al
. 1994). An important aspect is that hybrid zones poten-tially allow the fitness of adult hybrid genotypes to beevaluated under natural conditions, which is often difficultto achieve otherwise in long-lived or experimentally lesstractable species (Rieseberg & Buerkle 2002; Lexer
et al
. 2003,2004). Also, hybrid zones represent promising venues forstudying interactions between heritable genetic variationand diversity at the community or ecosystem level (Whitham1989; Whitham
et al
. 1999).The potential usefulness of natural hybrid zones for
addressing important evolutionary questions calls for theidentification and study of hybrid populations across awide range of animals and plants including different lifehistories and mating systems. A well-developed concep-tual framework exists for characterizing newly describedhybrid zones. Particularly important questions include:(i) What is the genomic composition of hybrids, i.e. is thehybrid zone dominated by F
1
s (e.g. Milne
et al
. 2003), or arelater generation hybrids frequent (e.g. Rieseberg
et al
. 1999;cases reviewed by Arnold
et al
. 2004)? (ii) Is the hybridzone maintained by a balance between dispersal and selec-tion against hybrids (Barton & Hewitt 1985; Barton & Gale1993), or does its structure also depend on adaptation to theexternal environment (Moore 1977; Harrison 1990)? (ii) Howsymmetric or asymmetric are patterns of gene exchange,and to what extent are they caused by intrinsic geneticvs. ecological factors (Arnold 1993; Bacilieri
et al
. 1996)?Answering these questions informs molecular ecologistsabout which components of the barrier to gene flow arelikely to be most important in maintaining the hybrid zone,and which analytical tools are most appropriate for study-ing its evolution.
The identification and study of hybrid zones is a par-ticularly timely topic in long-lived forest trees. Althoughconventional quantitative trait locus (QTL) mapping hasfacilitated studies of the genetic architecture of interspecificdifferences in some taxa (e.g. Wu
et al
. 1997; Frewen
et al
.2000; Saintagne
et al
. 2004), for most species the necessaryadvanced generation crosses are not available. This isunfortunate, because trees exhibit numerous features thatdistinguish them from annual model plants and likelyaffect the way in which their populations evolve, e.g. con-trol of leaf and flower phenology, seasonal reallocationof nutrients, extreme plasticity in stress response-relatedcharacters, juvenile–mature phase change, and differentdynamics of colonization into open habitats (Austerlitz
et al
. 2000; Bradshaw
et al
. 2000). Also, there is considerableinterest in the question concerning the conditions underwhich trees are most likely to be able to adapt
in situ
to theexpected rate of climate change (Davis & Shaw 2001). Nat-ural hybrid zones may hold important clues regarding thegenetic architecture of adaptively important traits in trees,
in particular in cases in which species barriers are at leastpartly ecological, i.e. composed of genes or chromosomalsegments conferring differential adaptation. Numerouswell-defined examples exist for ecological species barriersamong temperate trees (e.g. Van Valen 1976; Eckenwalder1996; McKinnon
et al
. 2001; reviewed by Lexer
et al
. 2004),and no group of taxa may be better suited for studying themthan the genus
Populus (
poplars, aspens, cottonwoods).
Populus
species are diploids (2
n
= 38) with relativelysmall genome sizes (550 Mb; 2C = 1.1 pg), and the numer-ous favourable attributes of this genus have made
Populus
a ‘model forest tree’ (Bradshaw
et al
. 2000) and led to aninternational effort to sequence its complete genome (Tuskan
et al
. 2004a; see http://www.ornl.gov/sci/ipgc). Speciesbarriers in
Populus
are often porous, allowing for widespreadhybridization, and genomic segments carrying neutral orfavourable genes appear to be able to cross the species bound-aries in many cases (Rajora & Dancik 1992; Eckenwalder1996; Martinsen
et al
. 2001; Floate 2004). Comparative link-age mapping in three different species (
Populus deltoides,Populus trichocarpa
, and
Populus nigra
) revealed widespreadhomologies for most of the 19 linkage groups of the
Populus
genome (Cervera
et al
. 2001), suggesting that species barriers in
Populus
are genic rather than chromosomal. Further evidencefor partially conserved gene order stems from cytogeneticdata for ribosomal sequences (Prado
et al
. 1996). Taken together,this suggests that barriers are composed primarily of singlegenes and their interactions, although it is too early to rule outa role for chromosomal rearrangements in species isolation.
Here, we study the species barrier between
Populus alba
(white poplar) and
Populus tremula
(European aspen), twoecologically divergent species that hybridize frequently inEurope. We ask the following questions regarding hybridzones of
P. alba
and
P. tremula
, using data from a wide zoneof sympatry along the Danube as well as parental referencesamples from outside the sympatric zone: (i) What is thegenomic composition of hybrids when estimated with agenome-wide set of codominant nuclear microsatellites,and what do these estimates tell us about the extent anddirection of introgression? (ii) How do patterns of nucleargene exchange relate to the species origin of the plastid DNAmolecule in the same set of individuals? In addition, weassess the role of effective population size (
N
e
) and pollen/seed flow in gene introgression and comment on the poten-tial usefulness of hybrid zones among
P. alba
and
P. tremula
for genomic studies of microevolutionary processes.
Materials and methods
Hybrid zones between
Populus alba
and
Populus tremula
The existence of hybrids between
Populus alba
and
Populustremula
in the lowland floodplain forests of the Danube

B A R R I E R T O G E N E F L O W I N
P O P U L U S
1047
© 2005 Blackwell Publishing Ltd,
Molecular Ecology
, 14, 1045–1057
near Vienna, Austria, has been suspected for severaldecades (Krembs 1956; Adler
et al
. 1994). Based on morph-ological evidence, some authors have argued that the hybridform, known as
Populus
×
canescens
or grey poplar, may beeven more common than pure
P. alba
(Lazowski 1997). Toour knowledge, only two molecular studies exist on hybridsamong the two species (Rajora & Dancik 1992; Fossati
et al
.2004). Although neither of them was carried out within apopulation genetic framework (between one and 10 treeswere sampled for each of several populations and comparedto controlled crosses), both studies indicate that
P. alba
and
P. tremula
are well differentiated for allele frequencies atdominant and codominant genetic markers and that
P.
×
canescens
hybrids from different European river valleysappear to be a mixture of early generation hybrids andbackcrosses (Rajora & Dancik 1992; Fossati
et al
. 2004).
P. alba
and
P. tremula
differ strongly in their habitatrequirements. In the Danube valley,
P. alba
favours floodedareas adjacent to the river and often extends to the activezone of the floodplain, where it plays a key role duringearly stages of succession (Lazowski 1997). Tolerance toflooding in
P. alba
may be mediated by a number of differ-ent characters, including life history traits (e.g. lack ofdormancy, rapid germination and seedling growth, rapidroot growth; Karrenberg
et al
. 2002), and physiologicaladaptations related to the prevention of cell damage causedby ethanol, a by-product of anaerobic processes in floodedplant tissue (Kreuzwieser
et al
. 1999). The preferred habitatsof
P.
×
canescens
hybrids are located in close proximity to
P. alba
in the floodplain forest (Krembs 1956; Adler
et al
. 1994;Lazowski 1997), although fine-scale spatial patterns andniche widths of the two taxa have not yet been compared.
The second parental species,
P. tremula
, does not grow inflooded areas, or in close proximity of the Danube or itstributaries. Instead, its preferred habitats are located sev-eral kilometres away, reaching into the submontane andmontane zones of the Alps (Adler
et al
. 1994). In addition tothese divergent habitat requirements, numerous morpho-logical traits distinguish
P. alba
and
P. tremula
(Adler
et al
.1994), but their adaptive significance is unknown. It isalso an open question whether the Danube valley hybridzone follows a tension zone model with a balance of dis-persal and selection against hybrids, or whether ecologyplays a role in maintaining the zone (Barton & Hewitt 1985;Harrison 1990; Arnold 1997). One of the primary pieces ofevidence for or against a role for ecological selection, thegeographical width of the zone of hybridization, is difficultto assess because present-day floodplain forests representonly fragments of their historical distribution.
Population and within-genome sampling
The sampling scheme used here was chosen to best meetthe trade-off between sampling gene copies in populations
and sampling loci in the genome. Twenty independentnuclear microsatellite loci (chosen from different linkagegroups) were assayed in a total of 93
P.
×
canescens
hybridmorphotypes from the Danube valley hybrid zone and tworeference populations of each parental species,
P. alba
and
P. tremula
. Reference populations had an average samplesize of
N
= 20 and included, for each species, one populationfrom ‘within’ and one adjacent parental population from‘outside’ the zone of sympatry. In addition, eight plastidDNA restriction site polymorphisms were typed in 46 ofthe sampled hybrids and in an average of 19 plantsfrom each of the reference populations. The 20 nuclearmicrosatellites (Table 1), developed by Tuskan
et al
. (2004b)and Van der Schoot
et al
. (2000), are available at the follow-ing web site: http://www.ornl.gov/sci/ipgc/ssr_resource.htm. The four plastid DNA markers (described in thesucceeding section) were identified based on Grivet
et al
.(2001).
Based on the highly outcrossing breeding system ofthe study species (wind-dispersed pollen and seeds;Petit
et al
. 2003) and a lack of population differentiationin preliminary microsatellite studies, populations assampled here covered large geographical areas of up to60 km
2
(i.e. populations typically included numerous‘forest stands’ as usually defined by silviculturists). Thefollowing populations were sampled:
P. × canescens/vie(P. × canescens hybrid zone in the Danube valley nearVienna, Austria; sampling mid-point: 48.26°N, 16.23°E),P. alba/vie (reference sample of P. alba from the zone ofsympatry in the Danube valley near Vienna, Austria;sampling mid-point: 48.26°N, 16.27°E), P. alba/rom (P. albafrom outside the zone of sympatry in the Danube valleyin Romania; sampling mid-point: 43.77°N, 23.96°E), P.tremula/vie (reference sample of P. tremula from the zoneof sympatry in the Danube valley near Vienna, Austria;sampling mid-point: 48.28°N, 15.89°E), P. tremula/car (P.tremula from outside the sympatric zone in the EasternAlps in Carinthia, Austria; sampling mid-point: 46.62°N,13.85°E).
Molecular marker assays
Total genomic DNA was extracted using either the DNeasyPlant Mini Kit (QIAGEN) or a modified approach based onDoyle & Doyle (1987), and DNA was quantified usingeither a TKO-100 fluorometer (Hoefer Scientific Instruments)or an Eppendorf BIO photometer. The 20 nuclear micro-satellites were polymerase chain reaction (PCR)-amplifiedfollowing methods described previously by Burke et al.(2002), making use of a standard touchdown cyclingprogram with an annealing temperature (Ta) of 48 °C anda three-primer protocol including unlabelled M13-taggedforward and unlabelled/untagged reverse primers foreach marker, and a third ‘universal’ M13-primer labelled

1048 C . L E X E R E T A L .
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
with one of the fluorescent dyes, FAM, HEX, NED or JOE(Applied Biosystems). Microsatellite genotypes wereresolved either on an AB3700 or AB3100 automatedsequencer (Applied Biosystems), making use of the differentfluorescent dyes and size differences between loci formultiplexing. Molecular sizes in base pairs were deter-mined using either the GENSIZE-500 ROX (GENPAK)or the GENESCAN-500 ROX size standard (AppliedBiosystems), and result files from the sequencers wereanalysed using genescan and genotyper software (AppliedBiosystems).
Four plastid DNA fragments were screened for restric-tion site polymorphisms, yielding a total of eight polymorph-isms that were used for identifying and characterizingplastid DNA haplotypes. The PCR primers/restrictionenzymes used were: ccmp10R-trnHM/MspI, atpBSAM-rbcLSAM/EcoRI, trnTP-trnDP/HinfI, ccmp10R-rpl16R1516/EcoRI-HhaI. The nomenclature of the primers follows Grivetet al. (2001) and the web-based resource at http://bfw.ac.at/200/1859.html, where the reaction conditions can also befound. The restriction fragments were resolved on 2%agarose gels and visualized by UV/ethidium bromidestaining.
Data analysis
Genetic diversity of sampled loci and populations. In order tocharacterize the microsatellite loci in the two study speciesand their hybrids, the number of alleles (A), variance inallele size (Var), expected heterozygosity (HE), and observedheterozygosity (HO) were calculated for each locus usingthe program msa (Dieringer & Schloetterer 2003). For easeof presentation, populations of each parental species werecombined in the analysis, which is justified by low levels ofgenetic divergence between them (succeeding discussion).All genetic diversity estimates obtained in msa were correctedfor differences in sample size. In addition, departures fromHardy–Weinberg equilibrium (HWE) in each studied taxonwere computed using exact tests in genepop 1.2 (Raymond& Rousset 1995), in order to test the possibility thatparticular loci may deviate from HWE because of null alleles(= allele nonamplification) stemming from cross-speciesamplification of the microsatellites. Subsequently, eachpopulation studied was characterized using the variancein allele size, HE, and HO calculated by msa, allelic richnesscorrected for sample size in fstat (Goudet 1995), and thewithin-population inbreeding coefficient f estimated for
Table 1 Genetic variability at 20 nuclear microsatellite loci in Populus alba, Populus tremula, and their hybrids (P. × canescens), including locusname, repeat type, number of alleles (A), variance in allele size (Var), expected heterozygosity (HE), and observed heterozygosity (HO) foreach locus
LocusRepeat type
P. alba P. × canescens P. tremula
A Var HE HO A Var HE HO A Var HE HO
PMGC 2852‡ di 10 24.2 0.715 0.667 16 54.9 0.805 0.701** 12 42.1 0.834 0.692WMPS 15§ tri 7 20.5 0.773 0.657 8 26.4 0.788 0.819 7 19.8 0.731 0.784ORPM 312† tri 9 28.3 0.789 0.737 11 25.6 0.756 0.701 7 36.8 0.726 0.718ORPM 344† di 4 0.8 0.320 0.34 8 3.5 0.219 0.198 5 22.0 0.662 0.514ORPM 206† tri 2 3.7 0.023 0.023 6 5.1 0.136 0.129 4 6.9 0.580 0.500ORPM 127† di 3 2.1 0.118 0.122 3 3.6 0.122 0.128 5 12.8 0.663 0.281**ORPM 202† tri 3 1.5 0.571 0.595 5 1.5 0.595 0.719 4 3.8 0.565 0.750ORPM 29† di 2 1.4 0.028 0.028 7 9.2 0.243 0.066** 8 27.2 0.676 0.579ORPM 30–1†¶ di 2 7.4 0.088 0.091 4 15.7 0.215 0.239 3 0.5 0.370 0.316ORPM 30–2†¶ di 14 17.7 0.828 0.886 20 58.0 0.903 0.899 8 28.7 0.805 0.947ORPM 220† tetra 1 0.0 0.000 0.000 3 1.1 0.109 0.114 6 14.9 0.633 0.763ORPM 28† di 3 1.3 0.514 0.415 4 1.1 0.390 0.400 4 6.9 0.241 0.263ORPM 137† di 7 13.4 0.630 0.326** 6 4.2 0.517 0.433 8 31.1 0.611 0.355ORPM 14† tetra 1 0.0 0.000 0.000 2 0.0 0.011 0.011 2 0.5 0.122 0.128ORPM 21† di 2 0.1 0.204 0.227 2 0.1 0.129 0.092 1 0.0 0.000 0.000ORPM 60† tri 7 28.5 0.675 0.727 9 41.2 0.796 0.700 4 28.1 0.192 0.205ORPM 149† di 4 12.6 0.600 0.634 6 10.1 0.545 0.318** 4 8.1 0.581 0.649ORPM 167† di 2 0.2 0.024 0.024 4 5.9 0.262 0.292 2 0.2 0.026 0.026ORPM 214† di 3 1.9 0.371 0.415 4 1.3 0.338 0.318 2 0.5 0.263 0.306WPMS 5§ di 7 31.7 0.830 0.857 11 35.8 0.740 0.703* 12 148.0 0.832 0.722
†Markers developed by Tuskan et al. (2004b); ‡markers from http://www.ornl.gov/sci/ipgc/ssr_resource.htm. (2000); §markers by Van der Schoot et al. 2000); ¶two independent loci with nonoverlapping allele size ranges amplified; *P < 0.05, **P < 0.005, exact tests for departure from Hardy–Weinberg equilibrium.

B A R R I E R T O G E N E F L O W I N P O P U L U S 1049
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
microsatellite loci by genepop. The significance of f wastested through tests for departures from HWE for eachpopulation using exact tests. In addition, each populationwas characterized for levels of diversity at plastid DNAmarkers, using the number of haplotypes detected in eachsample and HE estimated by arlequin (Schneider et al.2000).
Effective population sizes and migration rates. Theta (4Neµ, withNe = effective population size and µ = mutation rate) for popu-lations of P. alba and P. tremula, and the effective number ofmigrants (Nem) between populations of each species, wereestimated following a coalescent theory and maximum-likelihood-based approach using migrate 1.5 (Beerli &Felsenstein 1999). Genetic divergence between populationsof each species (FST; Weir & Cockerham 1984) was used toobtain initial start values for the estimation of theta and Nem.The computations were carried out under both the infiniteallele model (IAM; Kimura & Crow 1964) and the stepwisemutation model (SMM; Kimura & Ohta 1978), and effectivepopulation sizes were estimated from theta values byassuming a microsatellite mutation rate of 10−3 per gameteper generation.
Nuclear admixture analysis and assignment tests. Nuclearadmixture proportions (Q) were estimated using the Bayesianapproach implemented in structure version 2 (Pritchardet al. 2000). The calculations were carried out under theadmixture model allowing for correlated allele frequencies,using the burn-in and simulation settings recommendedby the user manual. We did not estimate the number ofpopulations K from the data because this would assume thatpopulations are in HWE, which is not expected in hybridzones. Instead, populations of each parental specieswere combined in accordance with high levels of gene flow(Nem) between them, and admixture proportions Q forP. × canescens hybrids were estimated based on pooledallele frequency data from each parental species. Priorpopulation information was used for all parental genotypesoriginating from outside the zone of sympatry (plants frompopulations P. alba/rom and P. tremula/car), which allowedus to make optimal use of the allopatric reference samples,and to estimate the probability that any of these plants hadreceived genes from the other species within the last threegenerations.
For ease of presentation, patterns of admixture betweenP. alba and P. tremula were depicted in the form of two-dimensional population assignments rather than one-dimensional admixture proportions. Simple likelihood-basedassignment tests (sensu Rannala & Mountain 1997) werecarried out in arlequin, and the results were verified usinga genetic distance-based method which requires evenfewer assumptions about population structure (Cornuetet al. 1999).
Genetic structure analysis. Analysis of molecular variance(amova) in arlequin was used to obtain F-statistics fornuclear microsatellite and plastid DNA markers in overlapp-ing sets of individuals, hence allowing us the disentanglethe effects of pollen and seed dispersal on patterns of geneticvariability. We tested the hierarchies ‘among species’, ‘amongpopulations within species’, and ‘within populations’, usingtwo different models, one that contained populations ofthe two parental species only, and one that contained P. ×canescens hybrids nested within P. alba. All calculationswere carried out both under the IAM and SMM. To depictgenetic relationships between populations in a graphical way,an unrooted neighbour-joining (NJ) tree (Saitou & Nei 1987)was constructed based on the chord distance of Cavalli-Sforza& Edward (1967) using microsatellites. One thousandbootstrap replicates of the distance matrix were obtainedin msa (Dieringer & Schloetterer 2003), and the resultingNJ trees were analysed in phylip 3.6 (Felsenstein 2004).
Plastid DNA haplotype network. A median-joining (MJ)network (Bandelt et al. 1999) was constructed based onplastid DNA haplotypes using the program network(www.fluxus-engineering.com). This method is capableof resolving even complex haplotype phylogenies (Posada &Crandall 2001). It uses parsimony criteria to identify medianvectors, i.e. consensus sequences of mutually close sequencesof markers which are biologically equivalent to possibleunsampled or extinct ancestral haplotypes. Initial runsusing all haplotypes detected by the eight restriction-sitepolymorphisms yielded a highly complex structure withnumerous reticulations. Hence, following the user manual,we excluded singleton haplotypes and downweightedrapidly mutating characters in subsequent runs. The defaultsettings were used for all other parameters. To test for apossible role of cyto-nuclear incompatibilities in determiningpatterns of introgression across the hybrid zone (Arnold1993), nuclear admixture proportions and plastid DNAhaplotype data were compared in a simple way: nonpara-metric Spearman rank correlations were used to test for apossible association between nuclear and plastid genomiccomposition using spss (SPSS Inc.).
Results
Variability at microsatellite loci
All 20 nuclear microsatellites were variable with up to 20alleles segregating in each taxon, expected heterozygosities(HE) of up to 0.903, and observed heterozygosities (HO) ashigh as 0.947 (Table 1). Variance in allele size, a simplediversity parameter under the SMM, followed a similartrend as diversity estimates under the IAM. One locusdisplayed significant deviations from HWE in Populus albaand another did so in Populus tremula, possibly because

1050 C . L E X E R E T A L .
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
of population subdivision. In contrast, four loci deviatedfrom HWE in P. × canescens, consistent with more frequentdepartures from random mating in the hybrid population. Nosingle locus deviated from HWE across all three taxa (Table 1).Allele frequency tables for each locus/taxon in Table 1 areavailable from the corresponding author on request.
Genetic diversity, effective population size, and gene flow
Microsatellite-based diversity estimates for each of thefive populations studied (Table 2) revealed higher HE, HO,and variance in allele size for populations of P. tremulacompared to P. alba, while Populus × canescens hybrids weregenerally intermediate between the two parental species,and HE for plastid DNA markers followed a similar patternacross populations (Table 2). In contrast, allelic richness atmicrosatellite loci, the most sensitive diversity parameterused in this study, revealed increased variability in P. ×canescens, probably because of the admixture of allelesfrom each parental species (Table 2).
In accordance with higher levels of diversity, effectivepopulation size was higher in P. tremula than in P. alba(Fig. 1). The estimates reported here were obtained underthe SMM, although the IAM estimates followed the same pat-tern across populations and species. Estimates of Nemobtained from the same analysis (Fig. 1) revealed highlevels of gene flow and connectedness between populationsof each parental species (P. alba: Nem = 3.1 and 3.4 dependingon direction of gene exchange, P. tremula: Nem = 14.7 and6.9, respectively). The between-species differences in Nemlikely reflect different geographical distances betweenpopulations – c. 780 km for the reference populations ofP. alba and 240 km for populations of P. tremula. Based onhigh levels of gene exchange, populations from eachparental species were pooled for their use as referencesamples in admixture analyses.
Nuclear admixture analysis
Bayesian admixture analysis indicated that the sympatricpopulations of P. alba and P. tremula were pure (admixture
coefficients for each individual Q > 0.950 or Q < 0.050,respectively), with the exception of two individuals of P.alba which were reclassified as P. × canescens based on theirQ values (all results reported here are for populations afterreclassification). The probability that any of the allopatricpopulations (P. alba/rom and P. tremula/car) had receivedgenes by interspecific introgression within the last threegenerations before sampling did not reach the P = 0.05level for any of the individuals. In effect, Bayesianadmixture analysis suggested that the error rate of parentalspecies identification in the field was low. In contrast, awide range of admixture proportions was observed for P.× canescens hybrid morphotypes (95% confidence intervalof Q = 0.468–0.998, range = 0.072–0.998), indicating thepresence of a wide range of hybrid generations andpreferential backcrossing to P. alba. Hybrid morphotypeswith P. alba-like genotypes (Q > 0.950) were kept in
Species/population
Microsatellites Plastid DNA
VarAllelic richness HE HO f
No. haplotypes (sample size) HE
P. alba/rom 12.2 3.23 0.368 0.341 +0.075 5 (14) 0.758P. alba/vie 8.3 3.27 0.406 0.419 −0.033 5 (27) 0.789P. × canescens/vie 15.2 3.97 0.431 0.399 +0.074* 11 (46) 0.814P. tremula/vie 23.4 3.86 0.512 0.466 +0.092* 9 (16) 0.858P. tremula/car 20.5 3.89 0.492 0.483 +0.018 6 (17) 0.802
*Exact tests for departures from Hardy–Weinberg equilibrium P < 0.005.
Table 2 Genetic variability in populationsof Populus alba, Populus tremula, and P. ×canescens, including the variance in allelesize (Var), allelic richness corrected for sam-ple size, expected heterozygosity (HE),observed heterozygosity (HO), and the within-population inbreeding coefficient f formicrosatellites, the number of plastid DNAhaplotypes found in each sample, andHE for plastid DNA markers
Fig. 1 Effective population sizes (Ne) in Populus alba and Populustremula, and effective migration rates (Nem) between pairs ofpopulations of each species, estimated by a coalescent theory andmaximum-likelihood-based approach. For population abbrevia-tions see Materials and methods. Ninety-five percent confidenceintervals (CI) for the Ne estimates were: P. alba/vie: CI = 463–529,P. alba/rom: CI = 524–586, P. tremula/vie: CI = 545–613, P. tremula/car: CI = 658–762.

B A R R I E R T O G E N E F L O W I N P O P U L U S 1051
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
the P. × canescens category in this study, because, bynecessity, admixture analysis underestimates allelic con-tributions from the donor species to highly backcrossedindividuals unless many loci in the genome have beensampled.
Likelihood-based and genetic distance-based genotypeassignments gave comparable results, and the former areplotted in Fig. 2. The tendency of P. × canescens hybrids to
backcross towards P. alba is clearly visible in Fig. 2a. Inaddition, Fig. 2b reveals assignment probabilities amongP. alba and P. x canescens that would not have been discern-ible in Fig. 2a because of the high differentiation of allelefrequencies between the two parental species (scalingeffects). Hybrid plants that are differentiated from bothparental species in plots a and b are recombinant earlygeneration hybrids, as revealed by an inspection of indi-vidual microsatellite genotypes, except for four plants whichappear to be F1s.
Genetic structure at nuclear and plastid DNA markers
Genetic divergence statistics obtained by amova weresignificant at all hierarchical levels, when only populationsof the two parental species were considered or when theanalysis was carried out with P. × canescens nested withinP. alba. For simplicity, only the results obtained under theIAM are summarized here (Table 3). Most importantly, theproportion of variation among populations within eachspecies was small for nuclear microsatellites (2%; FSC =0.038) and clearly larger for plastid DNA (15%; FSC = 0.246),while the proportion of variation within populations followeda complementary pattern (Table 3). Our data inform usabout the role of pollen vs. seed dispersal in maintainingpatterns of gene introgression. As expected from gene flowand admixture analyses, populations of each parental spe-cies grouped together with high bootstrap support in theneighbour-joining tree, and P. × canescens had a strongeraffinity to P. alba than to P. tremula (Fig. 3).
Fig. 2 Log-likelihoods of microsatellite-based genotype assign-ments to Populus alba vs. Populus tremula (Fig. 2a) or P. alba vs. P. ×canescens hybrids (Fig. 2b). Taxon designations used to performthe analyses were based on leaf morphology and were testedby Bayesian admixture analysis prior to the assignment tests.Open circles, P. alba; filled circles, P. tremula; crosses, P. × canescenshybrids.
Fig. 3 Unrooted neighbour-joining tree based on Cavalli Sforza& Edward’s (1967) chord distance for microsatellites, includingbootstrap support values. For population abbreviations seeMaterials and methods.
1052 C . L E X E R E T A L .
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
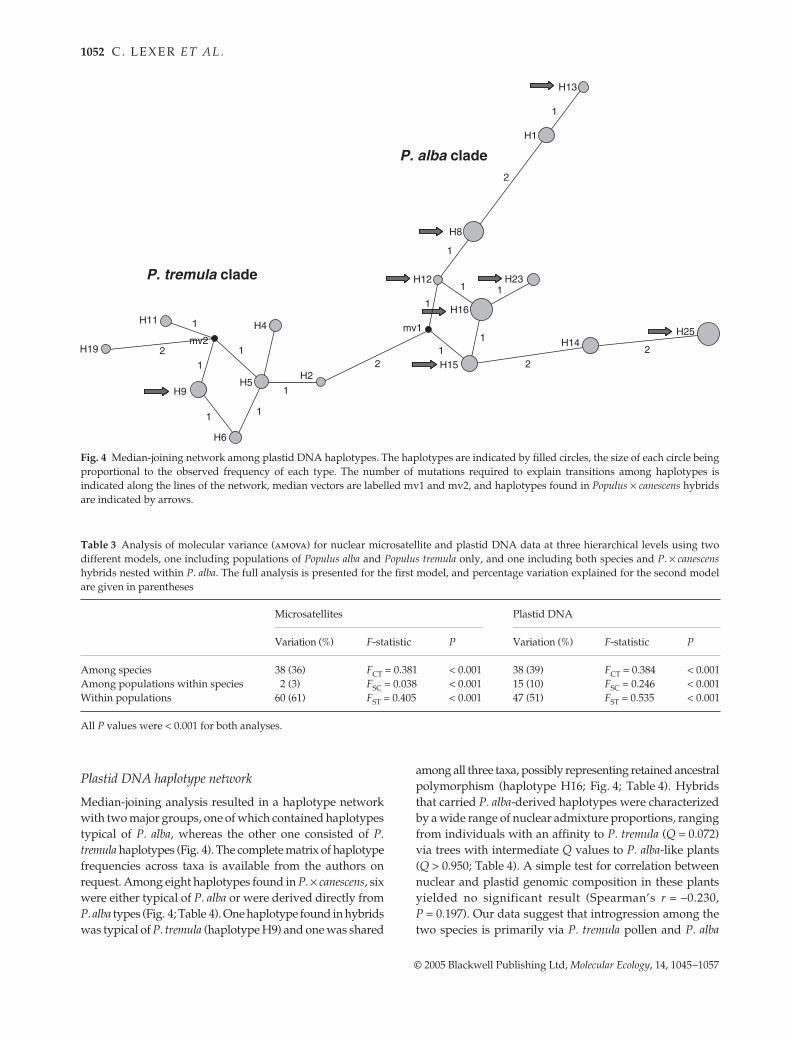
Plastid DNA haplotype network
Median-joining analysis resulted in a haplotype networkwith two major groups, one of which contained haplotypestypical of P. alba, whereas the other one consisted of P.tremula haplotypes (Fig. 4). The complete matrix of haplotypefrequencies across taxa is available from the authors onrequest. Among eight haplotypes found in P. × canescens, sixwere either typical of P. alba or were derived directly fromP. alba types (Fig. 4; Table 4). One haplotype found in hybridswas typical of P. tremula (haplotype H9) and one was shared
among all three taxa, possibly representing retained ancestralpolymorphism (haplotype H16; Fig. 4; Table 4). Hybridsthat carried P. alba-derived haplotypes were characterizedby a wide range of nuclear admixture proportions, rangingfrom individuals with an affinity to P. tremula (Q = 0.072)via trees with intermediate Q values to P. alba-like plants(Q > 0.950; Table 4). A simple test for correlation betweennuclear and plastid genomic composition in these plantsyielded no significant result (Spearman’s r = −0.230,P = 0.197). Our data suggest that introgression among thetwo species is primarily via P. tremula pollen and P. alba
Fig. 4 Median-joining network among plastid DNA haplotypes. The haplotypes are indicated by filled circles, the size of each circle beingproportional to the observed frequency of each type. The number of mutations required to explain transitions among haplotypes isindicated along the lines of the network, median vectors are labelled mv1 and mv2, and haplotypes found in Populus × canescens hybridsare indicated by arrows.
Table 3 Analysis of molecular variance (amova) for nuclear microsatellite and plastid DNA data at three hierarchical levels using twodifferent models, one including populations of Populus alba and Populus tremula only, and one including both species and P. × canescenshybrids nested within P. alba. The full analysis is presented for the first model, and percentage variation explained for the second modelare given in parentheses
Microsatellites Plastid DNA
Variation (%) F-statistic P Variation (%) F-statistic P
Among species 38 (36) FCT = 0.381 < 0.001 38 (39) FCT = 0.384 < 0.001Among populations within species 2 (3) FSC = 0.038 < 0.001 15 (10) FSC = 0.246 < 0.001Within populations 60 (61) FST = 0.405 < 0.001 47 (51) FST = 0.535 < 0.001
All P values were < 0.001 for both analyses.

B A R R I E R T O G E N E F L O W I N P O P U L U S 1053
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
seed parents, and that this asymmetric pattern is shaped byfactors other than intrinsic cytonuclear incompatibilities.
Discussion
Recently, the role of natural selection in population diver-gence and speciation has been hotly debated (Merila &Crokrak 2001; Wilding et al. 2001; McKay & Latta 2002;Rieseberg et al. 2002; Campbell and Bernatchez 2004), andthis has triggered renewed interest in hybrid zones as venuesfor studying the factors that create or maintain barriers togene flow between divergent populations (Rieseberg et al.1999; Martinsen et al. 2001; Rogers et al. 2001; Remington &Perugganan 2003; Vines et al. 2003). The identification andcharacterization of hybrid zones in diverse taxa is thereforean important task with numerous potential spin-offs inmolecular ecology. This is particularly true for organismswith existing genome programs such as Populus (Tuskanet al. 2004a), because the generation of complete genomicsequences, expressed sequence tag (EST) libraries, or othersources of genetic markers make it possible to study hybridzones from a genomic perspective. Our results on the extentand direction of introgression between the ecologicallydivergent species, Populus alba and Populus tremula, directlycontribute to the development of this approach in membersof a genus with considerable ecological importance(Whitham 1989; Eckenwalder 1996; Bradshaw et al. 2000).
Extent and direction of introgression
Our results confirm earlier reports of introgression betweenP. tremula and P. alba which were based on less extensivesampling of dominant and/or codominant nuclear markers(Rajora & Dancik 1992; Fossati et al. 2004), and they alsoallow us to suggest possible mechanisms responsible forthe observed patterns of hybridization. In the present study,20 microsatellite loci, representative of a large proportionof the nuclear genome of Populus, and eight plastid DNA
restriction site polymorphisms allow us to estimate effectivepopulation size (Ne) and gene flow (Nem) in the two parentalspecies (Fig. 1), study nuclear admixture (Fig. 2) and thespecies origin of plastid DNA variants (Fig. 4), and assessgenetic relationships and structure within and amongpopulations from both within and outside a large zone ofsympatry (Fig. 3; Table 3). Our plastid DNA data reveal thatintrogression between the two species occurs preferentiallyvia P. tremula pollen and P. alba seed parents (Fig. 4; Table 4),and our nuclear microsatellite data indicate that almostall hybrids in the studied hybrid zone are recombinantbackcrosses towards P. alba (Fig. 2). Our data not only informus about the genetic architecture of the Danube valleyhybrid zone, they also contain important information aboutthe factors that may be responsible for maintaining theobserved genetic structure.
The role of ecological and life history factors in gene introgression
Effective population sizes (Ne) were large in both speciesand were larger in P. tremula than in P. alba (Fig. 1), inagreement with the community ecology of the species. Inthe Danube valley, P. tremula occurs in mixed uplandforest communities covering huge geographical areas,whereas P. alba is mainly restricted to patchy, fragmentedfloodplain forests adjacent to the Danube (Krembs 1956;Adler et al. 1994; Lazowski 1997). This, together with thefact that P. tremula probably recolonized the area earlierthan the warmth-loving P. alba (Huntley & Birks 1983), mayaccount for larger Ne in the former. Note that the differencesin Ne (95% confidence intervals do not overlap; legend ofFig. 1) are unlikely to be artefacts arising from differencesin cross-transferability of the microsatellites, since geneticdiversity was higher in P. tremula for both nuclear micro-satellite and plastid DNA markers (Table 2). Large effectivepopulation sizes in the two species indicate that selec-tion should be more important in promoting evolutionary
Table 4 Plastid DNA haplotypes found in P. x canescens hybrids after exclusion of three singletons, including indication about haplotypesharing among taxa, group position in the plastid DNA network, frequencies in both species and their hybrids, and range of nuclearadmixture proportions (Q) for plants carrying each haplotype
Haplotype TaxonHaplotype group
FrequenciesNuclear admixture (Q)alba (N = 41) can (N = 46) trem (N = 33)
H8 alba/can alba 8 4 0 0.992–0.998H9 trem/can trem 0 1 7 0.592H12 can alba 0 2 0 0.985–0.997H13 can alba 0 3 0 0.996–0.998H15 alba/can alba 5 2 0 0.822–0.998H16 alba/can/trem alba 6 9 1 0.472–0.998H23 can alba 0 5 0 0.440–0.998H25 alba/can alba 4 17 0 0.072–0.998

1054 C . L E X E R E T A L .
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
change than drift. However, the direction of the interspecificdifference in Ne leaves us with a puzzle — why is introgressionin the direction of the species with the smaller effective popu-lation size, P. alba? As outlined in the succeeding discussion,a joint genetic structure analysis of nuclear and plastid DNAmay inform us about the likely causes for this pattern.
Analysis of molecular variance indicates that a much largerproportion of the plastid DNA variation resides amongpopulations of each parental species compared to nuclearmicrosatellites (15% vs. 2%, respectively; Table 3). If, forexample, only P. tremula populations are considered ingenetic divergence analyses, this results in FST estimates of0.310 for plastid DNA and 0.017 for nuclear microsatellites.In a finite island model under migration-drift equilibrium,this translates into a pollen/seed flow ratio of 23.9, followingequation 5 in Ennos (1994). Hence, pollen/seed flow is similarto that found in other long-lived forest trees with wind-dispersed pollen and seeds (e.g. Pinus muricata; pollen/seed ratio c. 24; Pinus radiata, pollen/seed ratio c. 31; Strausset al. 1993), but much lower than in trees with wind-dispersedpollen and heavy animal-dispersed fruits (e.g. Quercus robur/petraea complex, pollen/seed ratio c. 196; Kremer et al. 1991).Our results confirm that pollen dispersal in P. tremula ismuch more efficient than seed dispersal although both aremoved by wind, and this has important implications forinterpreting patterns of hybridization between the twospecies.
It has long been known that hybridization betweenP. alba and P. tremula is tightly connected to the disturbanceregime of the floodplain forest (Krembs 1956; Adler et al.1994; Lazowski 1997; Karrenberg et al. 2002), the habitat‘mosaic’ in which P. alba also occurs. This is where P. × canes-cens hybrid morphotypes were found and sampled in thepresent work. In contrast, the preferred upland habitats ofP. tremula are separated by at least several kilometres andextend far into the Alps. Hence, considering the low mobil-ity of seeds compared to pollen in P. tremula, it makes sensethat established hybrids on the floodplain preferentiallycarry the plastid DNA of their geographically closer parent,P. alba (Fig. 4). Note also that poplars are dioecious (male andfemale sex expressed on different trees). Hence, chances forestablishment of female F1 hybrids with P. tremula-derivedplastids, capable of passing P. tremula plastid DNA on toP. alba via backcrossing, are reduced even further (50% fora sex ratio of 1:1). The close geographical proximity ofP. alba and P. × canescens, well documented by ecologists(Krembs 1956; Adler et al. 1994; Lazowski 1997), also presentsa simple and plausible explanation for unidirectionalintrogression as an alternative to ‘intrinsic’ genetic factors.Of course, differences in flowering phenology must beexpected to contribute to the observed pattern as well —P. tremula tends to start flowering several weeks earlier thanP. alba (Senghas & Seybold 1993), i.e. P. tremula flowers mayalready be fertilized when P. alba pollen is spread.
Although studying locus-specific effects was not thegoal of this study, our nuclear and plastid DNA datalead us to speculate about the role of cytonuclear incom-patibilities in gene introgression. In the Danube valley,Populus × canescens hybrids preferentially carry plastid DNAhaplotypes from P. alba and are characterized by a widerange of nuclear admixture proportions (Table 4). In effect,there is no apparent incompatibility between P. alba plastidDNA and P. tremula nuclear genomes. We were not able totest for incompatibilities in the opposite direction, sinceasymmetric dispersal and location of ‘hybrid’ habitats actlike a filter, effectively preventing P. tremula plastid DNAfrom introgressing into P. alba as outlined earlier. To ourknowledge, historical crossing experiments (e.g. Wettstein1929) do not contain sufficient information to solve thispuzzle either, e.g. offspring from laboratory crosses neednot be viable under field conditions. Likewise, the fact thatintrogression is in the same backcross direction (to P. alba)in other European river valleys (Rajora & Dancik 1992;Fossati et al. 2004) does not resolve the question either,because ecological and spatial patterns in these places aresimilar. Nevertheless, studies of North American Populushybrid zones report a lack of cytonuclear incompatibilitiesin both crossing directions (Paige et al. 1991). This providesweak evidence against a role for cytonuclear incompatibil-ities in Populus, although we caution against prematuregeneralizations.
Regardless of the outcome of this debate, the wide dis-tribution of backcrossed hybrid genotypes (Fig. 2) indicatesthat the species barrier between P. alba and P. tremula isextremely porous. Continued backcrossing and introgressioncreate a wealth of interspecific recombination products,resulting in increased levels of genetic variability comparedto pure P. alba (Table 1) and notoriously large variances inphenotypic characters (Adler et al. 1994; Eckenwalder et al.1996). Given sufficient heritability in the relevant traits, thismay allow hybrids to take advantage of the highly dynamicsetting provided by the floodplain forest. To put it in EdgarAnderson’s words, ‘around a thousand different kinds ofhabitat would be needed to permit the various recombina-tions to find a niche somewhere’ (Anderson 1948). We arguethat the ‘mosaic’ of the floodplain forest provides thesehabitats, and therefore the ecological opportunity requiredfor the establishment of recombinant hybrid backcrosses.
Implications for studies of microevolutionary processes
Hybrid zones are important tools for studying the evolutionof reproductive barriers in sympatry, the role of selectionin creating or maintaining population/species differences,the role of hybridization in evolution, or the genetic archi-tecture of important phenotypic traits that differ betweenthe hybridizing populations (Barton & Hewitt 1985; Mallet& Barton 1989; Nuernberger et al. 1995; Rieseberg & Buerkle

B A R R I E R T O G E N E F L O W I N P O P U L U S 1055
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
2002; Vines et al. 2003). The last topic is particularly relevantfor hybridizing taxa with divergent ecological preferences,such as the two Populus species studied here (floodplainpioneer vs. upland species). The studied hybrid zone shouldbe suitable for QTL mapping-type studies of interspecificcharacter differences (sensu Rieseberg & Buerkle 2002),because (i) numerous diagnostic phenotypic charactersdiffer between P. alba and P. tremula (Adler et al. 1994), (ii)numerous ecological differences separate the two species(iii) the two species are strongly differentiated for nuclearmicrosatellite allele frequencies, and (iv) the proportion ofrecombinant genotypes in the hybrid zone is high.
Although the wide and continuous distribution of back-crossed genotypes (Fig. 2) speaks against a typical tensionzone model (balance between dispersal and selection againsthybrids), the question remains as to what extent ecologicalselection associated with the habitat ‘mosaic’ of the flood-plain forest accounts for the persistence of the hybrid zone(Harrison 1990; Barton & Gale 1993; Vines et al. 2003).In the absence of reliable data on historical hybrid zonewidth, this question may be addressed by studying spatialpatterns of hybridization, or associations between hybridgenomic composition and environmental variables (Vineset al. 2003; Dodd & Afzal-Rafii 2004). Also, the role of clonalreproduction on hybrid zone persistance (Schweitzer et al.2002) deserves special attention in these species, since clonalpropagation may allow P. × canescens hybrids to persist in thehybrid zone even in the absence of sexual recombination.
Finally, an important aspect is that backcrossed (recom-binant) hybrids between P. alba and P. tremula are found inseveral European river valleys (Rajora & Dancik 1992;Fossati et al. 2004). This, combined with the high potentialof nuclear microsatellites indicated by the present study,suggests the possibility of comparative genomic studiesamong several hybrid zones in Europe, equivalent to ‘rep-licated natural experiments’ across largely independentevolutionary trajectories.
Acknowledgements
We wish to thank Loren Rieseberg and Keith Gardner for numer-ous helpful discussions during the early phases of the project,Christian Fraissl and Franz Kovacs of the Danube FloodplainNational Park, Austria, the Forstverwaltung Lobau of the ViennaCity Council, Herbert Tiefenbacher and several other privatelandowners in Austria for their continued support during samplecollection, and Alex Buerkle for helpful discussions about dataanalyses in hybrid zones. This work was supported by award No.NE/C507037/1 of the Natural Environment Research Council(NERC) in the United Kingdom to CL.
References
Adler W, Oswald K, Fischer R (1994) Exkursionsflora von Oesterreich.Verlag Eugen Ulmer, Stuttgart und Wien.
Anderson E (1948) Hybridization of the habitat. Evolution, 2, 1–9.Arnold J (1993) Cytonuclear disequilibria in hybrid zones. Annual
Review of Ecology and Systematics, 24, 521–554.Arnold ML (1997) Natural Hybridization and Evolution. Oxford
University Press, New York.Arnold ML, Bouck AC, Cornman RS (2004) Verne Grant and
Louisiana irises: is there anything new under the sun? NewPhytologist, 161, 143–149.
Austerlitz F, Mariette S, Machon N, Gouyon P-H, Bernard G (2000)Effects of colonization processes on genetic diversity: differencesbetween annual plants and tree species. Genetics, 154, 1309–1321.
Bacilieri R, Ducousso A, Petit RJ, Kremer A (1996) Mating systemand asymetric hybridization in a mixed stand of European oaks.Evolution, 50, 900–908.
Bandelt H-J, Forster P, Roehl A (1999) Median-joining networksfor inferring intraspecific phylogenies. Molecular Biology andEvolution, 16, 37–48.
Barton NH, Gale KS (1993) Genetic analysis of hybrid zones.In: Hybrid Zones and the Evolutionary Process, pp. 13–45. OxfordUniversity Press, Oxford.
Barton NH, Hewitt GM (1985) Analysis of hybrid zones. AnnualReview of Ecology and Systematics, 16, 113–148.
Beerli P, Felsenstein J (1999) Maximum-likelihood estimation ofmigration rates and effective population numbers in two popu-lations using a coalescent approach. Genetics, 152, 763–773.
Bradshaw HD, Ceulemans R, Davis J, Stettler R (2000) Emergingmodel systems in plant biology: poplar (Populus) as a model foresttree. Journal of Plant Growth and Regulation, 19, 306–313.
Bradshaw HD, Wilbert SM, Otto KG, Schemske DW (1995) Geneticmapping of floral traits associated with reproductive isolationin monkeyflowers (Mimulus). Nature, 376, 762–765.
Briscoe D, Stephens JC, O’Brien SJ (1994) Linkage disequilibriumin admixed populations: applications in gene mapping. Journalof Heredity, 85, 59–63.
Burke JM, Tang S, Knapp SJ, Rieseberg LH (2002) Genetic analysisof sunflower domestication. Genetics, 161, 1257–1267.
Campbell D, Bernatechez L (2004) Genomic scan using AFLPmarkers as a means to assess the role of directional selection in thedivergence of sympatric whitefish ecotypes. Molecular Biologyand Evolution, 21, 945–956.
Cavalli-Sforza LL, Edwards AWF (1967) Phylogenetic analysis:models and estimation procedures. Evolution, 21, 550–570.
Cervera M-T, Storme V, Ivens B et al. (2001) Dense genetic linkagemaps of three Populus species (Populus deltoides, P. nigra, andP. trichocarpa) based on AFLP and microsatellite markers. Genetics,158, 787–809.
Cornuet J-M, Piry S, Luikart G, Estoup A, Solignac M (1999) Newmethods employing multilocus genotypes to select or excludepopulations as origins of individuals. Genetics, 153, 1989–2000.
Davis MB, Shaw RG (2001) Range shifts and adaptive responses toquaternary climate change. Science, 292, 673–679.
Dieringer D, Schlötterer C (2003) Microsatellite analyser (msa): aplatform independent analysis tool for large microsatellite datasets. Molecular Ecology Notes, 3, 167–169.
Dodd RS, Afzal-Rafii Z (2004) Selection and dispersal in a multi-species oak hybrid zone. Evolution, 58, 261–269.
Doyle JJ, Doyle JA (1987) A rapid DNA isolation procedure for smallquantities of fresh leaf tissue. Phytochemical Bulletin, 19, 11–15.
Eckenwalder JE (1996) Systematics and evolution of Populus. In:Biology of Populus, and its Implications for Management and Conser-vation (eds Stettler RF, Bradshaw HD, Heilman PE, HinckleyTM). NRC Research Press, Ottawa.

1056 C . L E X E R E T A L .
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
Ennos RA (1994) Estimating the relative rates of pollen and seedmigration among plant populations. Heredity, 72, 250–259.
Felsenstein J (2004) PHYLIP (Phylogeny Inference Package), Version 3.6.Department of Genome Sciences and Department of Biology,University of Washington, Seattle.
Floate KD (2004) Extent and patterns of hybridisation among thethree species of Populus that constitute the riparian forest ofsouthern Alberta, Canada. Canadian Journal of Botany, 82, 253–264.
Fossati T, Patrignani G, Zapelli I et al. (2004) Development ofmolecular markers to assess the level of introgression of P. tremulainto P. alba natural populations. Plant Breeding, 123, 382–385.
Frewen BE, Chen THH, Howe GT et al. (2000) Quantitative traitloci and candidate gene mapping of bud set and bud flush inPopulus. Genetics, 154, 837–845.
Goudet J (1995) fstat Version 1.2: a computer program to calculateF-statistics. Journal of Heredity 86, 485–486.
Grivet D, Heinze B, Vendramin GG, Petit RJ (2001) Genome walk-ing with consensus primers: application to the large single copyregion of chloroplast DNA. Molecular Ecology Notes, 1, 345–349.
Harrison RG (1990) Hybrid zones: windows on evolutionary proc-esses. Oxford Surveys in Evolutionary Biology, 7, 69–128.
Huntley B, Birks H (1983) An Atlas of Past and Present Pollen Mapsof Europe: 0–13 000 Years Ago. Cambridge University Press,Cambridge.
Karrenberg S, Edwards PJ, Kollmann J (2002) The life history ofSalicaceae living in the active zone of floodplains. FreshwaterBiology, 47, 733–748.
Kimura M, Crow JF (1964) The number of alleles that can be main-tained in a finite population. Genetics, 49, 725–738.
Kimura M, Ohta T (1978) Stepwise mutation model and distribu-tion of allelic frequencies in a finite population. Proceedings of theNational Academy of Sciences of the Unites States of America, 75,2868–2872.
Krembs O (1956) Die Graupappel in den Donau-Auen. AllgemeineForstzeitschrift, 11, 345–347.
Kremer A, Petit RJ, Zanetto A et al. (1991) Nuclear and organellegene diversity in Quercus robur and Q. petraea. In: Genetic Vari-ation in European Populations of Forest Trees (eds Mueller-Starck G,Ziehe M), pp. 141–166. Sauerlaender Verlag, Frankfurt, Germany.
Kreuzwieser J, Scheerer U, Rennenberg H (1999) Metabolic originof acetaldehyde emitted by poplar (Populus tremula × alba) trees.Journal of Experimental Botany, 50, 757–765.
Lazowski W (1997) Auen in Oesterreich — vegetation, landschaft undnaturschutz. In: Monographien Band 81. Federal EnvironmentAgency — Vienna, Austria.
Lexer C, Heinze B, Alia R, Rieseberg LH (2004) Hybrid zones as atool for identifying adaptive genetic variation in outbreedingforest trees: lessons from wild annual sunflowers (Helianthusspp.). Forest Ecology and Management, 197, 49–64.
Lexer C, Randell RA, Rieseberg LH (2003) Experimental hybrid-ization as a tool for studying selection in the wild. Ecology, 84,1688–1699.
Luikart G, England PR, Tallmon D, Jordan S, Taberlet P (2003) Thepower and promise of population genomics: from genotypingto genome typing. Nature Reviews Genetics, 4, 981–994.
Mallet J, Barton NH (1989) Strong natural selection in a warning-color hybrid zone. Evolution, 43, 421–431.
Martinsen GD, Whitham TG, Turek RJ, Keim P (2001) Hybridpopulations selectively filter gene introgression between species.Evolution, 55, 1325–1335.
McKay JK, Latta RG (2002) Adaptive population divergence: markers,QTL and traits. Trends in Ecology and Evolution, 17, 285–291.
McKinnon GE, Vaillancourt RE, Jackson HD, Potts BM (2001)Chloroplast sharing in the Tasmanian eucalypts. Evolution, 55,703–711.
Merila J, Crnokrak P (2001) Comparison of genetic differentiationat marker loci and quantitative traits. Journal of EvolutionaryBiology, 14, 892–903.
Milne RI, Terzioglu S, Abbott RJ (2003) A hybrid zone dominatedby fertile F1s: maintenance of species barriers in Rhododendron.Molecular Ecology, 12, 2719–2729.
Moore WS (1977) An evaluation of narrow hybrid zones in verte-brates. Quarterly Review of Biology, 52, 263–267.
Nuernberger B, Barton NH, McCallum C, Gilchrist J, Appleby M(1995) Natural selection on quantitative traits in the Bombinahybrid zone. Evolution, 49, 1224–1238.
Orr HA (2001) The genetics of species differences. Trends in Ecologyand Evolution, 16, 343–350.
Paige KN, Capman WC, Jennetten P (1991) Mitochondrial inheritancepatterns across a cottonwood hybrid zone: cytonuclear dis-equilibria and hybrid zone dynamics. Evolution, 45, 1360–1369.
Petit RJ, Aguinagalde I, de Beaulieu JL et al. (2003) Glacial refugia:hotspots but not melting pots of genetic diversity. Science, 300,1563–1565.
Posada D, Crandall KA (2001) Intraspecific gene genealogies: treesgrafting into networks. Trends in Ecology and Evolution, 16, 37–45.
Prado EA, Faivre-Rampant P, Schneider C, Darmency MA (1996)Detection of a variable number of ribosomal DNA loci byfluorescent in situ hybridisation in Populus species. Genome, 39,1020–1026.
Pritchard JK, Stephens M, Donnelly P (2000) Inference of popula-tion structure using multilucus genotype data. Genetics, 155,945–959.
Rajora OP, Dancik BP (1992) Genetic characterisation and relation-ships of Populus alba, P. tremula, and P. × canescens, and their clones.Theoretical and Applied Genetics, 84, 291–298.
Rannala B, Mountain JL (1997) Detecting immigration by usingmultilocus genotypes. Proceedings of the National Academy ofSciences of the Unites States of America, 94, 9197–9201.
Raymond M, Rousset F (1995) genepop (Version 1.2.): populationgenetics software for exact tests and ecumenicism. Journal ofHeredity 86, 248–249.
Remington DL, Perugganan MD (2003) Candidate genes, quantit-ative trait loci, and functional trait evolution in plants. InternationalJournal of Plant Sciences, 164, S7–S20.
Rieseberg LH, Buerkle CA (2002) Genetic mapping in hybridzones. American Naturalist, 159, S37–S49.
Rieseberg LH, Morjan CL (2004) How species evolve collectively:implications of gene flow and selection for the spread of advant-ageous alleles. Molecular Ecology, 13, 1341–1356.
Rieseberg LH, Whitton J, Gardner K (1999) Hybrid zones and thegenetic architecture of a barrier to gene flow between twosunflower species. Genetics, 152, 713–727.
Rieseberg LH, Widmer A, Arntz MA, Burke JM (2002) Directionalselection is the primary cause of phenotypic diversification.Proceedings of the National Academy of Sciences of the United Statesof America, 99, 12242–12245.
Rogers SM, Campbell D, Baird SJE, Danzmann RG, Bernatchez L(2001) Combining the analyses of introgressive hybridizationand linkage mapping to investigate the genetic architecture of

B A R R I E R T O G E N E F L O W I N P O P U L U S 1057
© 2005 Blackwell Publishing Ltd, Molecular Ecology, 14, 1045–1057
population divergence in the lake whitefish (Coregonus clupae-formis, Mitchell). Genetica, 111, 25–41.
Saintagne C, Bodenes C, Barreneche T et al. (2004) Distribution ofgenomic regions differentiating oak species by QTL detection.Heredity, 92, 20–30.
Saitou N, Nei M (1987) The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Molecular Biologyand Evolution, 4, 406–425.
Schneider S, Roessli D, Excoffier L (2000) ARLEQUIN Version 2.000. Asoftware for population genetics data analysis. Available at http://anthro.unige.ch/arlequin.
Van der Schoot J, Pospiskova M, Vosman B, Smulders MJM (2000)Development and characterization of microsatellite markers inblack poplar (Populus nigra L.). Theoretical and Applied Genetics,101, 317–322.
Schweitzer JA, Martinsen GD, Whitham TG (2002) Cottonwoodhybrids gain fitness traits of both parents: a mechanism for theirlong-term persistence? American Journal of Botany, 89, 981–990.
Senghas K, Seybold S (1993) ‘Schmeil-Fitschen’, Flora von Deutsch-land und angrenzender Laender. Quelle & Meyer Verlag, Hei-delberg-Wiesbaden.
Strauss SH, Hong Y-P, Hipkins VD (1993) High levels of popula-tion differentiation for mitochondrial DNA haplotypes in Pinusradiata, muricata, and attenuata. Theoretical and Applied Genetics,86, 605–611.
Tuskan GA, DiFazio SP, Teichmann T (2004a) Poplar genomics isgetting popular: the impact of the poplar genome project on treeresearch. Plant Biology, 6, 2–4.
Tuskan GA, Gunter LE, Yang ZK, Yin T, Sewell MM (2004b)Microsatellite discovery and genetic mapping in Populus tri-chocarpa × deltoides hybrids. Canadian Journal of Forestry Research,34, 85–93.
Van Valen L (1976) Ecological species, multispecies and oaks.Taxon, 25, 233–239.
Vines TH, Kohler SC, Thiel A et al. (2003) The maintenance ofreproductive isolation in a mosaic hybrid zone between thefire-bellied toads Bombina bombina and B. variegata. Evolution, 57,1876–1888.
Weir BS, Cockerham CC (1984) Estimating F-statistics for the ana-lysis of population structure. Evolution, 38, 1358–1370.
Wettstein W (1929) Die Zuechtung von Pappeln (Populus). DerZuechter, 1. Jahrg, 219–220.
Whitham TG (1989) Plant hybrid zones as sinks for pests. Science,244, 1490–1493.
Whitham TG, Martinsen GD, Floate KD, Dungey HS, Potts BM,Keim P (1999) Plant hybrid zones affect biodiversity: tools for agenetic-based understanding of community structure. Ecology,80, 416–428.
Wilding CS, Butlin RK, Grahame J (2001) Differential gene exchangebetween parapatric morphs of Littorina saxatilis detected usingAFLP markers. Journal of Evolutionary Biology, 14, 611–619.
Wu R, Bradshaw HD, Stettler RF (1997) Molecular genetics ofgrowth and development in Populus (Salicaceae). 5. Mappingquantitative trait loci affecting leaf variation. American Journal ofBotany, 84, 143–153.
This work was carried out within a larger collaboration betweenthe Genetics section at Kew and the Federal Office and ResearchCentre for Forests in Vienna, Austria. Christian Lexer, the P. I. atKew, is a population geneticist in Mike Fay’s Genetics section, andJeffrey Joseph is a research assistant there. The whole Geneticsgroup studies population genetic and cytogenetic aspects of plantevolution with a focus on conservation. Berthold Heinze’s groupis interested in the application of genomic tools to the functionalcharacterization, conservation, and breeding of forest trees, andMarius-Sorin Nica was a student in his lab.




















