
Attractiveness of common insectary and harvestable floral resources to beneficial insects
9
Attractiveness of common insectary and harvestable floral resources to beneficial insects Brian N. Hogg a,⇑ , Robert L. Bugg b , Kent M. Daane a a Department of Environmental Science Policy and Management, University of California, 137 Mulford Hall, Berkeley, CA 94720-3114, USA b Consulting Biologist, 1555 Pinnacles Place, Davis, CA 95616-6660, USA article info Article history: Received 10 May 2010 Accepted 9 September 2010 Available online 17 September 2010 Keywords: Syrphidae Toxomerus Conservation biological control Floral provisioning Insectary plants Sweet alyssum abstract Resources for natural enemies are often lacking in agricultural fields. The provisioning of floral resources in crop fields can ameliorate this problem by providing nectar and pollen to natural enemies. To select an appropriate floral resource, plants must be screened for their flowering times and attractiveness to nat- ural enemies and pests. We tested the attractiveness of nine species of annual flowering plants to hover- flies, which are important predators of aphids in California lettuce fields. We also sampled arthropods in the foliage of tested plants, to assess the abundances of other natural enemy and pest species that were present. Tested plants included three commonly-used insectary plants and six flowering plant species that showed potential as either harvestable herbs or cut flowers. Harvestable insectary plants may pro- vide additional economic incentive for growers to set aside land for floral resources. The commonly-used insectary plant sweet alyssum consistently attracted the most hoverflies and the least bees, while poten- tially harvestable plants attracted few hoverflies. Competition with bees may have reduced hoverfly visits to several of the tested plant species. Sweet alyssum also stayed in bloom the longest, and contained the highest numbers of predatory hemipterans. Results suggest that plants should be screened for their attractiveness to not only the target biological control agent, but also to other potential competitors for floral resources. While this initial study focused on a limited selection of harvestable annual plants, a wide variety of other marketable plant species, particularly perennials, remain to be tested for their attractiveness to hoverflies or other beneficial arthropods. Ó 2010 Elsevier Inc. All rights reserved. 1. Introduction Nectar and pollen are critical for the survival and reproductive success of many natural enemy species such as hoverflies and par- asitoids (Cowgill et al., 1993; Lavandero et al., 2005), and can be a supplemental food source for spiders (Taylor and Pfannenstiel, 2008). These resources are often lacking in agricultural monocul- tures, where farming practices like tilling and herbicide applica- tions reduce plant diversity (Tilman et al., 2001). The addition of floral resources to crop fields can improve the survival, develop- ment and fecundity of natural enemy species (Irvin et al., 1999; Lee and Heimpel, 2008; Tylianakis et al., 2004), resulting in lower herbivore densities and crop damage (Gurr et al., 2000). Including non-crop plants in agricultural fields can also moderate environ- mental conditions such as temperature and humidity and provide alternative habitat for natural enemies (Thomas et al., 1991). Selecting flowering species that both provide resources to spe- cific natural enemies and meld into the target crop’s management program is not always a clear process (Bianchi and Wackers, 2008). Flowering plant species can differ widely in their attractiveness to natural enemies (Ambrosino et al., 2006; Colley and Luna, 2000; Cowgill et al., 1993). One approach involves screening candidate plants for traits that are appropriate for the targeted natural ene- mies and the crop system (Baggen and Gurr, 1998; Pontin et al., 2006). Ideally, the flowering plant should bloom early in the crop cycle to attract natural enemies before pest damage occurs (Colley and Luna, 2000). For crops with a short duration from planting to harvest, only plants that flower quickly may be appropriate. In addition, the selected flowering plant should attract fewer pests than the crop itself (Baggen et al., 1999). In this study we screened six species of potentially marketable herbs or cut flowers for their attractiveness to hoverflies (Diptera: Syrphidae). Adult hoverflies depend upon floral resources as adults (Gilbert, 1981), while the larvae are highly effective predators of aphids (Tenhumberg and Poehling, 1995; Rojo and Marcos-Garcia, 1997; Müller and Godfray, 1999; Wyss et al., 1999; Michaud and Belliure, 2001). The primary aim of this study was to compare the attractiveness of several harvestable flowering plants with floral resource species that are more commonly used in California organic row crops, particularly in lettuce fields, where hoverflies are a key element in the biological control of the lettuce aphid, 1049-9644/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.biocontrol.2010.09.007 ⇑ Corresponding author. Fax: +1 559 646 6593. E-mail address: [email protected] (B.N. Hogg). Biological Control 56 (2011) 76–84 Contents lists available at ScienceDirect Biological Control journal homepage: www.elsevier.com/locate/ybcon
Transcript of Attractiveness of common insectary and harvestable floral resources to beneficial insects

Biological Control 56 (2011) 76–84
Contents lists available at ScienceDirect
Biological Control
journal homepage: www.elsevier .com/ locate/ybcon
Attractiveness of common insectary and harvestable floral resourcesto beneficial insects
Brian N. Hogg a,⇑, Robert L. Bugg b, Kent M. Daane a
a Department of Environmental Science Policy and Management, University of California, 137 Mulford Hall, Berkeley, CA 94720-3114, USAb Consulting Biologist, 1555 Pinnacles Place, Davis, CA 95616-6660, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 May 2010Accepted 9 September 2010Available online 17 September 2010
Keywords:SyrphidaeToxomerusConservation biological controlFloral provisioningInsectary plantsSweet alyssum
1049-9644/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.biocontrol.2010.09.007
⇑ Corresponding author. Fax: +1 559 646 6593.E-mail address: [email protected] (B.N. Hogg
Resources for natural enemies are often lacking in agricultural fields. The provisioning of floral resourcesin crop fields can ameliorate this problem by providing nectar and pollen to natural enemies. To select anappropriate floral resource, plants must be screened for their flowering times and attractiveness to nat-ural enemies and pests. We tested the attractiveness of nine species of annual flowering plants to hover-flies, which are important predators of aphids in California lettuce fields. We also sampled arthropods inthe foliage of tested plants, to assess the abundances of other natural enemy and pest species that werepresent. Tested plants included three commonly-used insectary plants and six flowering plant speciesthat showed potential as either harvestable herbs or cut flowers. Harvestable insectary plants may pro-vide additional economic incentive for growers to set aside land for floral resources. The commonly-usedinsectary plant sweet alyssum consistently attracted the most hoverflies and the least bees, while poten-tially harvestable plants attracted few hoverflies. Competition with bees may have reduced hoverfly visitsto several of the tested plant species. Sweet alyssum also stayed in bloom the longest, and contained thehighest numbers of predatory hemipterans. Results suggest that plants should be screened for theirattractiveness to not only the target biological control agent, but also to other potential competitorsfor floral resources. While this initial study focused on a limited selection of harvestable annual plants,a wide variety of other marketable plant species, particularly perennials, remain to be tested for theirattractiveness to hoverflies or other beneficial arthropods.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
Nectar and pollen are critical for the survival and reproductivesuccess of many natural enemy species such as hoverflies and par-asitoids (Cowgill et al., 1993; Lavandero et al., 2005), and can be asupplemental food source for spiders (Taylor and Pfannenstiel,2008). These resources are often lacking in agricultural monocul-tures, where farming practices like tilling and herbicide applica-tions reduce plant diversity (Tilman et al., 2001). The addition offloral resources to crop fields can improve the survival, develop-ment and fecundity of natural enemy species (Irvin et al., 1999;Lee and Heimpel, 2008; Tylianakis et al., 2004), resulting in lowerherbivore densities and crop damage (Gurr et al., 2000). Includingnon-crop plants in agricultural fields can also moderate environ-mental conditions such as temperature and humidity and providealternative habitat for natural enemies (Thomas et al., 1991).
Selecting flowering species that both provide resources to spe-cific natural enemies and meld into the target crop’s managementprogram is not always a clear process (Bianchi and Wackers, 2008).
ll rights reserved.
).
Flowering plant species can differ widely in their attractiveness tonatural enemies (Ambrosino et al., 2006; Colley and Luna, 2000;Cowgill et al., 1993). One approach involves screening candidateplants for traits that are appropriate for the targeted natural ene-mies and the crop system (Baggen and Gurr, 1998; Pontin et al.,2006). Ideally, the flowering plant should bloom early in the cropcycle to attract natural enemies before pest damage occurs (Colleyand Luna, 2000). For crops with a short duration from planting toharvest, only plants that flower quickly may be appropriate. Inaddition, the selected flowering plant should attract fewer peststhan the crop itself (Baggen et al., 1999).
In this study we screened six species of potentially marketableherbs or cut flowers for their attractiveness to hoverflies (Diptera:Syrphidae). Adult hoverflies depend upon floral resources as adults(Gilbert, 1981), while the larvae are highly effective predators ofaphids (Tenhumberg and Poehling, 1995; Rojo and Marcos-Garcia,1997; Müller and Godfray, 1999; Wyss et al., 1999; Michaud andBelliure, 2001). The primary aim of this study was to comparethe attractiveness of several harvestable flowering plants withfloral resource species that are more commonly used in Californiaorganic row crops, particularly in lettuce fields, where hoverfliesare a key element in the biological control of the lettuce aphid,

B.N. Hogg et al. / Biological Control 56 (2011) 76–84 77
Nasonovia ribis-nigri Mosley (Pascual-Villalobos et al., 2006; Smithet al., 2008). Land devoted to floral resources is typically removedfrom crop production, and harvestable floral resources were inves-tigated as a means of offsetting this loss, as suggested by Bugg et al.(2008). As secondary aims of this study, we examined the role ofcandidate plant species in providing habitat for potential naturalenemies and pests, and the attractiveness of plants to bees thatwere in high numbers at the study site. Bees may reduce hoverflyvisits to flower species through competition (Ambrosino et al.,2006; Morse, 1981).
2. Materials and methods
2.1. Site description
The experiment took place during a single growing season fromMay to September 2009 at an 18 ha organic vegetable and cutflower farm in Sonoma County, California. The research plot wasapproximately 80 � 20 m, and was bordered on one side by a smallpond and on all other sides by crop fields. Several honey bee, Apismellifera L., hives were present in the field on the opposite side ofthe pond, �200 m from the research plot. The plot was irrigatedusing overhead, high-volume sprinklers every 1–2 d during germi-nation and early growth, and every 3–4 d thereafter (equivalent to�3 cm of water per week). Compost was applied to the plot beforethe experiment began at a rate of 2–3.6 tons/ha. After consultationwith the farm manager, kale (Brassica oleracea var. acephela) waschosen as the focal crop since it had higher aphid densities thanlettuce or other vegetable crops at the site and in Sonoma Countyin general.
Flowering plant species were selected based on the followingcriteria: (1) annual growth cycle and fast growth, (2) ability to startfrom seed, (3) general hardiness, (4) tolerance for full sun, (5) highlevel of flower production, and (6) the collaborating farm man-ager’s input on the marketability of harvestable floral species.The tested species included: sweet alyssum (Lobularia maritima),buckwheat (Fagopyrum esculentum), tansy phacelia (Phacelia tan-acetifolia), wild mustard (Brassica sp.), wild arugula (Diplotaxismuralis), Thai basil (Ocimum basilicum var. thyrsiflora), white bor-age (Borago officinalis ‘Alba’), cosmos (Cosmos sulphureus), and nas-turtium (Tropaeolum majus ‘Milkmaid’). Sweet alyssum, buckwheatand tansy phacelia were included to provide a comparison withcommonly-used insectary plants. Mustard, wild arugula and basilare harvestable herbs, while borage, cosmos and nasturtium havevalue as cut flowers. These plants represent only a small propor-tion of the possible harvestable insectary species suggested byBugg et al. (2008); some of the suggested plants, e.g. culinary sage(Salvia officinalis), lovage (Levistica officinale), oregano (Origanumvulgare), and rosemary (Rosmarinus officinalis), are perennial anddid not lend themselves well to use in lettuce fields or this short-term experiment. With the exception of tansy phacelia, the flowersof all tested species were white or yellow, which has been shownto be preferred by hoverflies (Cowgill et al., 1993). Kale was in-cluded as the crop treatment.
All species were planted from seed. The research plot was tilledon 15 May and seeds were planted the following day. Five repli-cates of each flower treatment were planted in a completely ran-domized design. Treatments were blocked by position in relationto the pond, to control for potential differences in moisture andproximity to the bee hives. Each treatment plot was 3 � 6 m andconsisted of two strips of the same flower species (with two rowsper strip) and two rows of kale. Rows ran parallel to the pond.Rows of kale were planted �1.5 m apart, and flower strips werethen planted in between the kale rows, so that one row of kale inthe middle of each plot was surrounded on both sides by two strips
of the same flower species, while the other row of kale at the edgeof each plot served as a buffer between plots. Rows within eachflower strip were �0.35 m apart, while rows of kale were �0.5 mfrom flower strips. On the advice of the farm manager, kale andborage were thinned on 5 June to 1–2 plants per �40 cm, to pre-vent overcrowding. All other plant species were left unthinned.Weeds (primarily amaranth, Amaranthus deflexus) were removedfrom plots every 21 d from 12 June to 22 July; after this time,weeds no longer posed a threat to maturing plants. To preventplants from reseeding, flowers were cut back when no open flow-ers remained in any one of the five plots for each species.
2.2. Sampling methods
Visual observations were conducted every 5–9 d between 24June and 23 September. For each plant species, observations beganonce plants started flowering, and ended when no open flowers re-mained in any one of the five treatment plots. Samples were takenon clear windless days between 1000 and 1400 h; observers al-ways faced the sun during observations, to avoid casting a shadowon flower strips. For each sample, observers randomly selected a1 m section (flower rows were �0.25 m wide) within the central4 m of one flower row, leaving a 1 m buffer on either side. Numbersof hoverflies and bees (honey bees, bumble bees, and native bees)visiting flower strips were counted. Both flower strips in each plotwere observed for 3 min each, for a total of 6 min per plot per plantspecies. A visit was recorded only if the fly or bee landed on thetreatment plant. The number of open flowers and weeds werecounted before and after each observation, respectively, within a0.25 � 0.25 m area (equivalent to a 0.25 m length of one flowerrow).
To sample the arthropod community inhabiting flower strips,the foliage of flowering plants was vacuumed every 12–15 d, from2 July to 26 August, using the vacuum function of a blower-vacuum(model RY09907, Ryobi Ltd., Hiroshima-Ken, Japan) with a piece oforgandy fitted over the intake tube to collect arthropods. A25 � 50 cm area of each flower strip was vacuumed for 15 s. Thecentral 4 m of each flower strip was divided into sections, and dif-ferent sections within each plot were sampled on consecutive sam-pling dates to prevent oversampling. Vacuum sampling focused onthe non-flowering parts of the plants. Samples were stored at�4 �Cbefore sorting in the lab. Collected arthropods were sorted to thehighest taxonomic level required for determination of feedingmode. Numbers of aphids were extremely high in some kale sam-ples on the last sample date, 26 August, and their total numberswere estimated by spreading aphids evenly over a Petri dish thathad been divided into quadrates of equal area, counting the num-ber of aphids in one quadrate, and multiplying by the total numberof quadrates in the Petri dish.
To assess the hoverfly species composition at the site and toexamine whether hoverfly species observed at flowers were aphi-dophagous, hoverflies were collected after observations from 1 Julyto 2 September using standard census walks. The collector walkedat an even pace for 15 min through each block, collecting everyhoverfly that was seen with a butterfly net. The plant species thateach fly was collected on was recorded. The timer was stoppedwhile each fly was collected, so that the total 15 min was spentsearching. Flies were stored at �4 �C and later identified to species.
2.3. Statistical analyses
Data from the two flower strips in each block were summed forall analyses. To assess the role of plant species as habitat for natu-ral enemies or potential pests, numbers of abundant (representedin samples by >100 individuals) beneficial arthropod groups (Ara-neae, predatory Hemiptera, and parasitic Hymenoptera) and insect

78 B.N. Hogg et al. / Biological Control 56 (2011) 76–84
families containing known agricultural pests (Chrysomelidae[Coleoptera] and herbivorous Aphididae, Cicadellidae, Lygaeidae[Hemiptera]) in vacuum samples were compared between flower-ing plant species. Since plants can function as habitat for naturalenemies even when not in bloom (Frank et al., 2008), data for eachplant species were averaged across the five sample dates. Numbersof arthropods in each group were compared among plant speciesusing ANOVA, with separation of means by pairwise comparison(Tukey HSD, P < 0.05); data were log(x + 1) or square root(x + 0.5) transformed, as needed, to improve homogeneity ofvariances.
To assess the attractiveness of flowering plant species to hover-flies and bees, numbers of hoverflies and bees (honey bees, otherApoidea) visiting plant species with the same peak bloom timeswere compared. Prior to analysis, numbers of visits for each plantspecies were averaged across the 3-week peak bloom period, theweek of peak bloom plus 1 week on either side (after Fiedler andLandis, 2007). Numbers of hoverflies and bees visiting flower stripswere compared between plant species using ANOVA, with separa-tion of means by pairwise comparison (Tukey HSD, P < 0.05); datawere log(x + 1) or square root (x + 0.5) transformed, as needed, toimprove homogeneity of variances. Sweet alyssum bloomed con-tinuously throughout the entire experiment and was included inall comparisons, while tansy phacelia experienced two peak bloomperiods and was included in two comparisons. The kale controltreatment was included in all comparisons.
In all plots, some unintended plant species (e.g., weeds) werepresent. To ensure that weeds did not influence the attractivenessor habitat structure of plant species, ANOVA was used to comparenumbers of weed stems between treatments based on week ofpeak bloom. This analysis showed that weeds did not differ signif-icantly between treatments (29 June–10 July: F = 2.74, P = 0.08; 10July–22 July: F = 0.21, P = 0.89; 5 August–19 August: F = 0.34,P = 0.88; 9 September–23 September: F = 3.05, P = 0.09).
Fig. 1. Mean numbers of open flowers across time on treatment plants that reachedpeak bloom (A) early, (B) in mid-season, and (C) late.
3. Results
3.1. Flowering times and duration
Tested plant species were clearly differentiated by periods ofbloom (Fig. 1). Buckwheat and mustard both reached peak bloombetween 24 June and 10 July (weeks 1–3), less than 6 weeks afterplanting (Fig. 1A). Although they bloomed before the other plantspecies, they both bloomed for short periods and had gone to seedby 22 July. Sweet alyssum, wild arugula and tansy phacelia also ini-tiated bloom between 24 June and 10 July, reached peak bloom be-tween 10 July and 22 July (weeks 3–5), and remained in flower forlong periods: sweet alyssum bloomed continuously throughout theentire study, tansy phacelia showed two peak bloom times (thesecond occurred between 5 August and 19 August) and wild aru-gula produced flowers until 2 September (Fig. 1B). Basil, borageand nasturtium reached peak bloom between 5 August and 19 Au-gust (weeks 7–9), and went quickly to seed by 9 September(Fig. 1C). Cosmos did not begin to produce flowers until 5 August,11 weeks after planting, and was still in bloom on the last observa-tion date, 23 September (Fig. 1C).
3.2. Plant species as habitat for herbivorous pests and natural enemies
A total 16,344 arthropods from 11 orders were collected andidentified in vacuum samples (Table 1). Potential natural enemiesrepresented 4.6% of the overall arthropod composition. The mostabundant natural enemies in plants were predatory hemipterans(1.8%; primarily the anthocorid Orius tristicolor), hymenopterousparasitoids (1.9%) and spiders (0.8%). Individuals from predatory
or parasitic families in the orders Coleoptera, Diptera, Neuropteraand Pseudoscorpiones were present in low numbers. Syrphids rep-resented only 0.24% of arthropod composition in vacuum samples,with a 1–2.5 ratio of adults to larvae. Aphids constituted the major-ity of the arthropods collected (73.7%); 99.5% of aphids were col-lected on kale and 98.9% of these were collected on the lastsample date (26 August). Other abundant herbivores includedCicadellidae (8.0%), herbivorous Lygaeidae (4.3%, represented so-lely by a Nysius sp.), Chloropidae (3.6%) and Chrysomelidae (0.8%).
Numbers of herbivorous pest insects differed among plant spe-cies. Aphids had the clearest separation, with almost 500 times moreaphids found on kale (the tested crop plant) than on any of the insec-tary or floral plant species (F9,39 = 33.093, P < 0.001, Fig. 2A). Whenexcluding kale from the analysis, aphid densities did not vary amongthe tested plant species (F8,35 = 1.015, P = 0.443). Plant species af-fected chrysomelids (F9,39 = 5.216, P < 0.001), with higher numbers
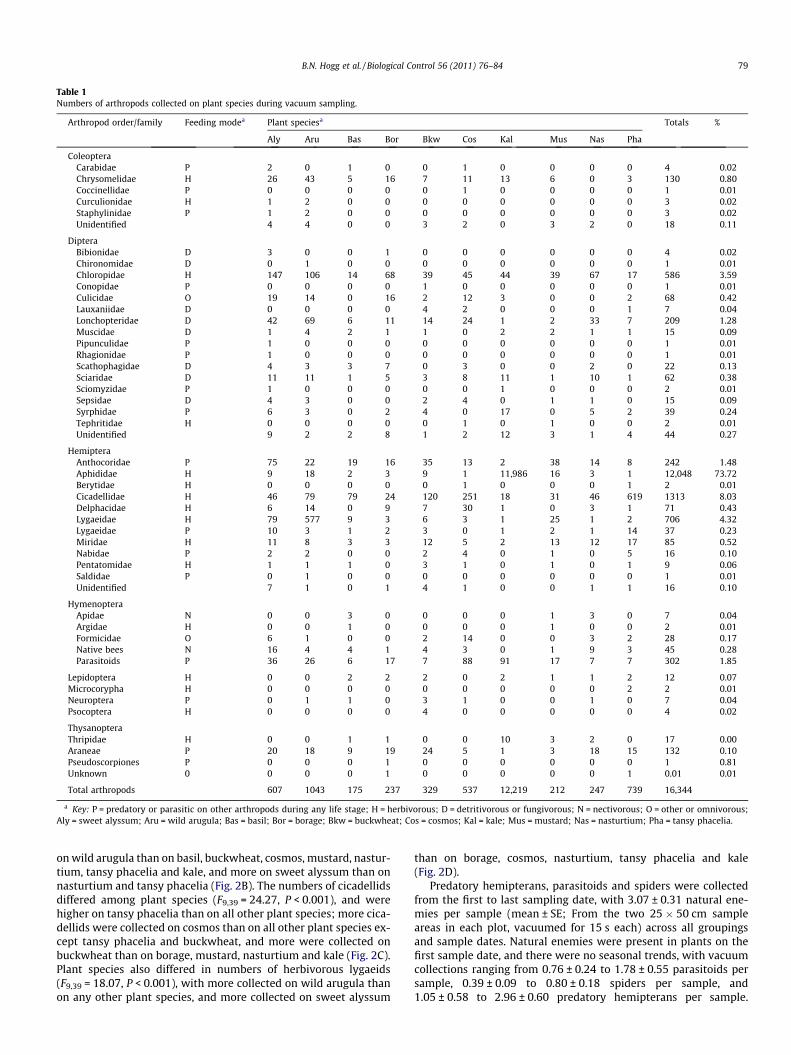
Table 1Numbers of arthropods collected on plant species during vacuum sampling.
Arthropod order/family Feeding modea Plant speciesa Totals %
Aly Aru Bas Bor Bkw Cos Kal Mus Nas Pha
ColeopteraCarabidae P 2 0 1 0 0 1 0 0 0 0 4 0.02Chrysomelidae H 26 43 5 16 7 11 13 6 0 3 130 0.80Coccinellidae P 0 0 0 0 0 1 0 0 0 0 1 0.01Curculionidae H 1 2 0 0 0 0 0 0 0 0 3 0.02Staphylinidae P 1 2 0 0 0 0 0 0 0 0 3 0.02Unidentified 4 4 0 0 3 2 0 3 2 0 18 0.11
DipteraBibionidae D 3 0 0 1 0 0 0 0 0 0 4 0.02Chironomidae D 0 1 0 0 0 0 0 0 0 0 1 0.01Chloropidae H 147 106 14 68 39 45 44 39 67 17 586 3.59Conopidae P 0 0 0 0 1 0 0 0 0 0 1 0.01Culicidae O 19 14 0 16 2 12 3 0 0 2 68 0.42Lauxaniidae D 0 0 0 0 4 2 0 0 0 1 7 0.04Lonchopteridae D 42 69 6 11 14 24 1 2 33 7 209 1.28Muscidae D 1 4 2 1 1 0 2 2 1 1 15 0.09Pipunculidae P 1 0 0 0 0 0 0 0 0 0 1 0.01Rhagionidae P 1 0 0 0 0 0 0 0 0 0 1 0.01Scathophagidae D 4 3 3 7 0 3 0 0 2 0 22 0.13Sciaridae D 11 11 1 5 3 8 11 1 10 1 62 0.38Sciomyzidae P 1 0 0 0 0 0 1 0 0 0 2 0.01Sepsidae D 4 3 0 0 2 4 0 1 1 0 15 0.09Syrphidae P 6 3 0 2 4 0 17 0 5 2 39 0.24Tephritidae H 0 0 0 0 0 1 0 1 0 0 2 0.01Unidentified 9 2 2 8 1 2 12 3 1 4 44 0.27
HemipteraAnthocoridae P 75 22 19 16 35 13 2 38 14 8 242 1.48Aphididae H 9 18 2 3 9 1 11,986 16 3 1 12,048 73.72Berytidae H 0 0 0 0 0 1 0 0 0 1 2 0.01Cicadellidae H 46 79 79 24 120 251 18 31 46 619 1313 8.03Delphacidae H 6 14 0 9 7 30 1 0 3 1 71 0.43Lygaeidae H 79 577 9 3 6 3 1 25 1 2 706 4.32Lygaeidae P 10 3 1 2 3 0 1 2 1 14 37 0.23Miridae H 11 8 3 3 12 5 2 13 12 17 85 0.52Nabidae P 2 2 0 0 2 4 0 1 0 5 16 0.10Pentatomidae H 1 1 1 0 3 1 0 1 0 1 9 0.06Saldidae P 0 1 0 0 0 0 0 0 0 0 1 0.01Unidentified 7 1 0 1 4 1 0 0 1 1 16 0.10
HymenopteraApidae N 0 0 3 0 0 0 0 1 3 0 7 0.04Argidae H 0 0 1 0 0 0 0 1 0 0 2 0.01Formicidae O 6 1 0 0 2 14 0 0 3 2 28 0.17Native bees N 16 4 4 1 4 3 0 1 9 3 45 0.28Parasitoids P 36 26 6 17 7 88 91 17 7 7 302 1.85
Lepidoptera H 0 0 2 2 2 0 2 1 1 2 12 0.07Microcorypha H 0 0 0 0 0 0 0 0 0 2 2 0.01Neuroptera P 0 1 1 0 3 1 0 0 1 0 7 0.04Psocoptera H 0 0 0 0 4 0 0 0 0 0 4 0.02
ThysanopteraThripidae H 0 0 1 1 0 0 10 3 2 0 17 0.00Araneae P 20 18 9 19 24 5 1 3 18 15 132 0.10Pseudoscorpiones P 0 0 0 1 0 0 0 0 0 0 1 0.81Unknown 0 0 0 0 1 0 0 0 0 0 1 0.01 0.01
Total arthropods 607 1043 175 237 329 537 12,219 212 247 739 16,344
a Key: P = predatory or parasitic on other arthropods during any life stage; H = herbivorous; D = detritivorous or fungivorous; N = nectivorous; O = other or omnivorous;Aly = sweet alyssum; Aru = wild arugula; Bas = basil; Bor = borage; Bkw = buckwheat; Cos = cosmos; Kal = kale; Mus = mustard; Nas = nasturtium; Pha = tansy phacelia.
B.N. Hogg et al. / Biological Control 56 (2011) 76–84 79
on wild arugula than on basil, buckwheat, cosmos, mustard, nastur-tium, tansy phacelia and kale, and more on sweet alyssum than onnasturtium and tansy phacelia (Fig. 2B). The numbers of cicadellidsdiffered among plant species (F9,39 = 24.27, P < 0.001), and werehigher on tansy phacelia than on all other plant species; more cica-dellids were collected on cosmos than on all other plant species ex-cept tansy phacelia and buckwheat, and more were collected onbuckwheat than on borage, mustard, nasturtium and kale (Fig. 2C).Plant species also differed in numbers of herbivorous lygaeids(F9,39 = 18.07, P < 0.001), with more collected on wild arugula thanon any other plant species, and more collected on sweet alyssum
than on borage, cosmos, nasturtium, tansy phacelia and kale(Fig. 2D).
Predatory hemipterans, parasitoids and spiders were collectedfrom the first to last sampling date, with 3.07 ± 0.31 natural ene-mies per sample (mean ± SE; From the two 25 � 50 cm sampleareas in each plot, vacuumed for 15 s each) across all groupingsand sample dates. Natural enemies were present in plants on thefirst sample date, and there were no seasonal trends, with vacuumcollections ranging from 0.76 ± 0.24 to 1.78 ± 0.55 parasitoids persample, 0.39 ± 0.09 to 0.80 ± 0.18 spiders per sample, and1.05 ± 0.58 to 2.96 ± 0.60 predatory hemipterans per sample.

Fig. 2. Mean numbers (+SE) of (A) aphid, (B) chrysomelid, (C) cicadellid, and (D) lygaeid pest species collected in vacuum samples. Data were averaged across the five sampledates. Different lower-case letters above means indicate significant differences (Tukey HSD, P < 0.05).
80 B.N. Hogg et al. / Biological Control 56 (2011) 76–84
Numbers of predatory hemipterans showed the greatest variationamong plant species (F9,39 = 7.243, P < 0.001), with higher numberscollected in sweet alyssum than in all other plant species exceptbuckwheat and mustard, and higher numbers in buckwheat andmustard than in kale (Fig. 3A). The numbers of hymenopteran par-asitoids were also affected by plant species (F9,39 = 4.615, P < 0.001)with most collected on kale and cosmos (Fig. 3B). While there wasa treatment impact on the numbers of spiders collected(F9,39 = 2.197, P = 0.043), their numbers did not differ among plantspecies in pairwise comparisons (Fig. 3C).
Fig. 3. Mean numbers of beneficial (A) hemipterans, (B) hymenopterans, and (C) spideDifferent lower-case letters above means indicate significant differences (Tukey HSD, P
3.3. Attractiveness of plant species to hoverflies and bees
Sweet alyssum consistently attracted the most hoverflies duringall periods of the study (Fig. 4). From 19 June to 10 July, numbers ofhoverfly visits were affected by plant species (F3,16 = 15.90,P < 0.001), with more hoverflies visiting sweet alyssum than mus-tard or the kale control treatment (Fig. 4A). Numbers of hoverflyvisits did not differ between sweet alyssum and buckwheat, how-ever. Numbers of hoverfly visits also differed among plant speciesfrom 10 July to 22 July (F3,16 = 46.52, P < 0.001), with more hoverfly
rs collected in vacuum samples. Data were averaged across the five sample dates.< 0.05).

Fig. 4. Mean numbers of visits (+SE) from hoverflies, honey bees, and other Apoidea (combined bumble bees and native bees) to plants that reached peak bloom between (A)24 June and 10 July (weeks 1–3); (B) 10 July and 22 July (weeks 3–5); (C) 5 August and 19 August (weeks 7–9); (D) 9 September and 23 September (weeks 12–14). Prior toanalysis data were averaged across the 3-week peak bloom period for each plant species (the week of peak bloom and each week on either side). Different lower-case lettersabove means indicate significant differences (Tukey HSD, P < 0.05).
B.N. Hogg et al. / Biological Control 56 (2011) 76–84 81
visits to all the flowering plant species than to kale and with morehoverfly visits to sweet alyssum than to tansy phacelia or wild aru-gula (Fig. 4B). From 5 August to 19 August, hoverflies were moreoften observed on sweet alyssum than on basil, borage, nasturtium,tansy phacelia or kale (F5,23 = 8.64, P < 0.001, Fig. 4C). Results weresimilar between 9 September and 23 September; hoverflies visitedsweet alyssum more than cosmos or kale (F2,12 = 33.28, P < 0.001,Fig. 4D). In kale, numbers of hoverflies visits per observation (fromtwo 25 � 50 cm areas, observed for 30 s each) ranged from 0 (onfour dates, on 22 July, 29 July, 16 September and 23 September)to a peak of 3.80 ± 1.31 (mean ± SE) on 2 September, coinciding
with the increase in aphids at this time. Numbers of hoverfly visitsto sweet alyssum showed a more complicated pattern across time,and ranged from 1.8 ± 0.58 hoverflies per observation on 24 June toa peak of 6.20 ± 0.97 on 17 July, followed by a second peak on 9September, when and 5.40 ± 1.21 hoverflies per observation wererecorded.
Honey bees, in contrast to hoverflies, were not attracted tosweet alyssum, although the number of honey bee visits differedamong plant species from 24 June to 10 July (F3,16 = 29.61,P < 0.001). All three flowering plant species attracted more beesthan kale, and mustard received more bee visits than sweet alyssum

82 B.N. Hogg et al. / Biological Control 56 (2011) 76–84
or buckwheat (Fig. 4A). Honey bee visitation was also affected byplant species from 10 July to 22 July (F3,16 = 39.35, P < 0.001), withmore visits were observed on wild arugula and tansy phacelia thanon sweet alyssum or kale (Fig. 4B). Between 5 August and 19 August,more honey bee visits were recorded on borage than sweet alyssum,wild arugula, nasturtium, tansy phacelia or kale and more visitswere observed on wild arugula, nasturtium and tansy phacelia thanon sweet alyssum or kale (F5,23 = 33.58, P < 0.001, Fig. 4C). Visitationalso differed among plant species from 9 September to 23 Septem-ber (F2,12 = 124.41, P < 0.001), and more honey bees were recordedon cosmos than sweet alyssum or kale (Fig. 4D).
Other Apoidea pollinator species included bumblebees (Bombusspp.) and various native bees (families Apidae, Haliticidae, Megac-hilidae), which have been grouped together as there was no clearlydominant species throughout the study and their numbers were al-ways lower than A. mellifera. From 24 June to 10 July and from 9September to 23 September, when tansy phacelia was not includedin analysis, bee visits were low and there was little separation be-tween treatments. From 24 June to 10 July, there were more beesrecorded on buckwheat than kale (F3,16 = 3.99, P = 0.027, Fig. 4A).As tansy phacelia began to flower more between 10 July and 22July, bee visits were affected by plant species (F3,16 = 16.25,P < 0.001), with more visits to tansy phacelia than all other species,and more visits to sweet alyssum that to kale (Fig. 4B). Plant spe-cies also affected numbers of bee visits between 5 August and 19August (F5,23 = 27.04, P < 0.001), with more visits to tansy phaceliathan all other treatments and more visits to sweet alyssum than tobasil or kale (Fig. 4C). In the final period of the study, bee visitationdid not differ among plant species (F2,12 = 2.86, P = 0.096, Fig. 4D).
During standard census walks, 157 hoverfly individuals repre-senting nine hoverfly species or morphospecies were collected (Ta-ble 2). Species composition was dominated by three species:Toxomerus marginatus (Say) (60.5%), Syrphus opinator Osten Sacken(11.5%), and Allograpta obliqua Say (8.3%). All of these species havebeen implicated as key aphid predators in California (Smith andChaney, 2007). Most hoverflies were collected on sweet alyssum;a large proportion was also collected on kale, after aphids arrivedat the site in mid-August. Corresponding to the increase in aphidnumbers, the highest numbers of syrphids were collected in lateAugust and early September; 69 (44.0%) of syrphid adults were col-lected on three samples dates between 19 August and 2 Septem-ber. There was considerable species turnover across time. In thefive collection dates between 1 and 29 July, T. marginatus repre-sented 52 out of the 71 (73.2%) hoverflies collected, but made uponly 40 of the 95 (42.1%) hoverflies collected in the five dates be-tween 5 August and 9 September. Several species, such as S. opin-ator, A. obliqua and Platycheirus stegnus Say only appeared incollections in late July or August.
Table 2Numbers of hoverfly species collected on plant species during standard census walks.
Hoverfly species Plant speciesa
Aly Aru Bas Bor Bkw
Allograpta obliqua 4 0 1 0 0Chrysotoxum sp. 2 0 0 0 0Paragus haemorrhous 7 0 0 0 0Platycheirus stegnus 0 0 0 0 1Sphaerophoria sp. 1 4 0 0 1 5Sphaerophoria sulphuripes 1 0 0 0 2Syrphus sp. 1 1 0 0 0 0Syrphus opinator 2 0 1 0 1Toxomerus marginatus 32 3 7 9 12Unidentified hoverflies 0 0 0 1 0
Total hoverflies 53 3 9 11 21
a Key: Aly = sweet alyssum; Aru = wild arugula; Bas = basil; Bor = borage; Bkw = buphacelia.
4. Discussion
Mustard and buckwheat started flowering earlier than anyother plant species. Other studies have noted the extremely shortinterval between planting and flowering for buckwheat (Stephenset al., 1998). Both mustard and buckwheat had gone to seed withinthe first 4 weeks, however. Sweet alyssum, on the other hand,starting blooming just one week after mustard and buckwheat,and then continued blooming for longer than any of the otherplants tested. Other plants that bloomed relatively early and con-tinuously were wild arugula and tansy phacelia.
Differences also emerged between plants in numbers of poten-tial pests. Floral resources should be viewed as a potential sourcefor pests if they harbor more pests than the crop itself (Winkleret al., 2006). While numbers of aphids, the primary pest, were farhigher on kale than on any of the treatment plants, the other her-bivores collected in this study were likely to have been generalists,and therefore also pose a threat to a variety of crops. Wild arugulacontained the highest numbers of Nysius sp. (Lygaeidae), which canbe a pest in grain and vegetable crops. Numbers of chrysomelidswere also high in wild arugula. A key chrysomelid in this studywas the western spotted cucumber beetle, Diabrotica undecimpunc-tata undecimpunctata Mannerheim, which is a serious agriculturalpest, particularly of corn and legumes. Tansy phacelia, on the otherhand, contained high numbers of cicadellid leafhoppers. While wedid not sort leafhoppers to species, collected leafhoppers werelikely to have been generalists, since tansy phacelia is uncommonin the Sonoma area and the same leafhopper species was collectedon all plants.
The natural enemy group that showed the clearest differencebetween treatment plants was the predatory Hemiptera, repre-sented primarily by the anthocorid O. tristicolor (White), whichwere highest in number in sweet alyssum. These results corrobo-rate those of Chaney (1998), who also found higher numbers ofan unidentified Orius sp. in sweet alyssum than in other testedplants. Orius species are predators of small soft-bodied pest insectssuch as aphids and thrips, and may have economic importance in avariety of crop systems. Fiedler and Landis (2007) similarly re-ported that O. insidiosus (Say) was the most abundant predator inthe perennial plants they tested. Although the annual growth cycleof sweet alyssum limits its role as an overwintering habitat for nat-ural enemies, significant numbers of predatory hemipterans werepresent on the first sample date. Numbers of parasitic Hymenop-tera, on the other hand, were higher in cosmos than in other treat-ment plant species. The high numbers of parasitoids in kale likelyresulted from the presence of aphids. Aphid parasitoids are knownto respond to cues such as aphid honeydew and plant volatiles (Duet al., 1998; Powell et al., 1998; Storeck et al., 2000). Although we
Totals %
Cos Kal Mus Nas Pha
1 5 0 2 0 13 8.30 0 0 0 0 2 1.30 0 0 0 0 7 4.50 4 0 0 0 5 3.20 0 0 0 0 10 6.40 2 0 0 0 5 3.20 0 0 0 0 1 0.61 9 0 0 4 18 11.52 14 2 6 8 95 60.50 0 0 0 0 1 0.6
4 34 2 8 12 157
ckwheat; Cos = cosmos; Kal = kale; Mus = mustard; Nas = nasturtium; Pha = tansy

B.N. Hogg et al. / Biological Control 56 (2011) 76–84 83
did not sort parasitoids to species, most appeared to be aphid par-asitoids (Aphidiidae); however, parasitoids are highly mobile andmay have been only transient inhabitants of treatment plots. Spi-ders did not differ between plant species. The plants tested byFrank et al. (2008) also did not show any differences in numbersof spiders. As generalist predators, spiders may not be sensitiveto slight differences in plant architecture, although an increase inplant biomass and/or diversity may increase overall spider num-bers in crop fields (Frank et al., 2008).
Sweet alyssum consistently attracted more hoverflies than allother treatment plants except buckwheat. Other studies have re-ported that hoverflies prefer buckwheat over other floweringplants (Lovei et al., 1998; Stephens et al., 1998). Flower architec-ture may have played a role in determining the attractiveness ofsweet alyssum and buckwheat to hoverflies. Both plant specieshave flowers with short corollae (Vattala et al., 2006), which in-creases the availability of nectar to beneficial insects, while thenectaries of flower species with deep corollae, such as tansy phac-elia and nasturtium (Baggen et al., 1999), may be inaccessible tohoverflies (Colley and Luna, 2000; Lovei et al., 1998). Although bor-age does not have particularly deep corollae (Baggen et al., 1999),its nectar may still have been inaccessible to hoverflies.
We also suspect that the presence of bees on treatment plantsinfluenced hoverfly preferences. Aggressive interactions were ob-served between bees and hoverflies on several occasions duringthe study, but were not frequent enough to quantify (Hogg, pers.observ.). Honey bees visited all other treatment plants more thansweet alyssum, with the notable exception of buckwheat, whileother Apoidea strongly preferred tansy phacelia. The extraordi-narily high numbers of honey bees at the research site may explaindiscrepancies between the current study and other studies thathave shown different preferences among hoverflies than those re-ported here (Colley and Luna, 2000) or no preferences (Pontin et al.,2006). Other researchers have implicated bees in excluding hover-flies from flowers; the mere presence of bees may discourage hov-erflies from visiting flowers (Ambrosino et al., 2006). Morse (1981)concluded that T. marginatus, the dominant hoverfly species in thecurrent study, avoided flowers that contained bumble bees.
In fact, T. marginatus may have had an inordinate influence onthe results. The high relative abundance of T. marginatus, particu-larly early in the summer, may explain why hoverfly visits tosweet alyssum peaked before aphids arrived in significant numbersat the site. T. marginatus may be most successful in crop fields withlow to moderate aphid densities (Smith and Chaney, 2007), andmay not show a strong response to increases in aphid numbers.The second peak in hoverfly visits that occurred when high num-bers of aphids were present was more in line with expectations,since aphid colonies and associated chemicals are likely to attractgravid female hoverflies (Budenberg and Powell, 1992; Bargenet al., 1998; Almohamad et al., 2007). By this time, hoverfly speciessuch as P. stegnus, A. obliqua and S. opinator had arrived at the site,apparently in response to the dramatic rise in aphid numbers.
It should be stressed that the combination of plants we testedmay have shaped our results; in particular, the presence ofsweet alyssum may have diverted hoverflies from nearby plantsthat they otherwise would have visited. Nonetheless, the poten-tially harvestable crops tested were clearly less attractive to hover-flies than sweet alyssum. Furthermore, many of these annualharvestable species, particularly the herbs, may not be marketableif they are allowed to flower. The leaves and stems of wild arugula,basil and mustard became woody and essentially inedible at flow-ering time. The cut flowers may be somewhat forgiving in this re-gard, although the flowers would have been marketable only asbuds or very young flowers. Harvesting a limited proportion ofthe harvestable plant strips before they flower may be a partialsolution to this problem, particularly for the cut flowers, which
would continue to produce new flowers after each cutting. Itshould be noted that we only tested six potentially harvestableplants in this study, however. Testing more potentially harvestablespecies may be a fruitful avenue for future research. In particular,many perennials show promise as attractants for hoverflies (Bugget al., 2008), and unlike many annuals, are still marketable afterthey come to flower.
Thus, we conclude that sweet alyssum is the most appropriateplant species of those tested for the attraction of syrphid fliesand natural enemies, particularly where bees are in high numbers,and may have the additional benefit of attracting few pests. Buck-wheat also showed potential for attracting hoverflies. One strategymay be to use combined plantings of flowers with different phe-nologies or nutritional attributes (Pontin et al., 2006). To increasenumbers of hoverflies early in the crop cycle, it may be possibleto plant buckwheat alongside sweet alyssum; buckwheat will pro-duce large amounts of flowers early in the crop cycle, whilesweet alyssum will continue to flower and attract hoverfliesthroughout the season. A harvestable alternative would be prefer-able to sweet alyssum or buckwheat, however, especially sinceboth species, particularly buckwheat, have the potential to becomeweedy (Daane, pers. observ.). It should be noted that the resultspresented here may have been skewed by the close proximity ofthe research site to bee hives. Further studies are needed thatexamine the potential role of competition in influencing the effec-tiveness of flowering strips in attracting hoverflies and other natu-ral enemies.
Acknowledgments
This work was funded by EPA grant X8-96982401 and USDA-NRI (NIFA) grant 2008-35302-04677. We thank Oak Hill Farm forthe use of their land, and David Cooper for help in planting andmaintaining the research plot. We thank Monica Cooper for helpwith the initial experimental design. We also thank Erik Nelsonfor input during initial planning for the study, and Eve Roubinet,Brianne Crabtree, Korey Kassir, Addie Abrams and John Hutchinsfor help in the field.
References
Almohamad, R., Verheggen, F.J., Francis, F., Haubruge, E., 2007. Predatory hoverfliesselect their oviposition site according to aphid host plant and aphid species.Entomologia Experimentalis et Applicata 125, 13–21.
Ambrosino, M.D., Luna, J.M., Jepson, P.C., Wratten, S.D., 2006. Relative frequencies ofvisits to selected insectary plants by predatory hoverflies (Diptera: Syrphidae),other beneficial insects, and herbivores. Environmental Entomology 35, 394–400.
Baggen, L.R., Gurr, G.M., 1998. The influence of food on Copidosoma koehleri(Hymenoptera: Encyrtidae), and the use of flowering plants as a habitatmanagement tool to enhance biological control of potato moth, Phthorimaeaopercullela (Lepidoptera: Gelechiidae). Biological Control 11, 9–17.
Baggen, L.R., Gurr, G.M., Meats, A., 1999. Flowers in tri-trophic systems:mechanisms allowing selective exploitation by insect natural enemies forconservation biological control. Entomologia Experimentalis et Applicata 91,155–161.
Bargen, H., Saudhof, K., Poehling, H., 1998. Prey finding by larvae and adult femalesof Episyrphus balteatus. Entomologia Experimentalis et Applicata 87, 245–254.
Bianchi, F., Wackers, F.L., 2008. Effects of flower attractiveness and nectaravailability in field margins on biological control by parasitoids. BiologicalControl 46, 400–408.
Budenberg, W.J., Powell, W., 1992. The role of honeydew as an ovipositionalstimulant for two species of syrphids. Entomologia Experimentalis et Applicata64, 57–61.
Bugg, R.L., Colfer, R.G., Chaney, W.E., Smith, H.A., Cannon, J., 2008. Flower flies(Syrphidae) and other biological control agents for aphids in vegetable crops.University of California Agriculture and Natural Resources, Publication 8285,Oakland, CA.
Chaney, W.E., 1998. Biological control of aphids in lettuce using in-field insectaries.In: Pickett, C.H., Bugg, R.L. (Eds.), Enhancing Biological Control. University ofCalifornia Press, Berkeley, California, pp. 73–83.
Colley, M.R., Luna, J.M., 2000. Relative attractiveness of potential beneficial insectaryplants to aphidophagous hoverflies (Diptera: Syrphidae). Environmental Ento-mology 29, 1054–1059.

84 B.N. Hogg et al. / Biological Control 56 (2011) 76–84
Cowgill, S.E., Wratten, S.D., Sotherton, N.W., 1993. The selective use of floralresources by the hoverfly Episyrphus balteatus (Diptera: Syrphidae) on farmland.Annals of Applied Biology 122, 223–231.
Du, Y., Poppy, G.M., Powell, W., Pickett, J.A., Wadhams, L.J., 1998. Identification ofsemiochemicals released during aphid feeding that attract parasitoid Aphidiuservi. Journal of Chemical Ecology 24, 1355–1368.
Fiedler, A.K., Landis, D.A., 2007. Attractiveness of Michigan native plants to arthropodnatural enemies and herbivores. Environmental Entomology 36, 751–765.
Frank, S.D., Shrewsbury, P.M., Esiekpe, O., 2008. Spatial and temporal variation innatural enemy assemblages on Maryland native plant species. EnvironmentalEntomology 37, 478–486.
Gilbert, F.S., 1981. Foraging ecology of hover flies: morphology of the mouthparts inrelation to feeding on nectar and pollen in some urban species. EcologicalEntomology 6, 245–262.
Gurr, G.M., Wratten, S.D., Barbosa, P., 2000. Success in conservation biologicalcontrol of arthropods. In: Gurr, G.M., Wratten, S.D. (Eds.), Biological Control:Measures of Success. Kluwer Academic Publishers, Dordrecht, The Netherlands,pp. 105–124.
Irvin, N.A., Wratten, S.D., Frampton, C.M., Bowie, M.H., Evans, A.M., Moar, N.T., 1999.The phenology and pollen feeding of three hover fly (Diptera: Syrphidae) speciesin Canterbury, New Zealand. New Zealand Journal of Zoology 26, 105–115.
Lavandero, B., Wratten, S., Shishehbor, P., Worner, S., 2005. Enhancing theeffectiveness of the parasitoid Diadegma semiclausum (Helen): movementafter use of nectar in the field. Biological Control 34, 152–158.
Lee, J.C., Heimpel, G.E., 2008. Floral resources impact longevity and oviposition rateof a parasitoid in the field. Journal of Animal Ecology 77, 565–572.
Lovei, G.L., MacLeod, A., Hickman, J.M., 1998. Dispersal and effects of barriers on themovement of the New Zealand hover fly Melanostoma fasciatum (Dipt.:Syrphidae) on cultivated land. Journal of Applied Entomology 122, 115–120.
Michaud, J.P., Belliure, B., 2001. Impact of syrphid predation on production ofmigrants in colonies of the brown citrus aphid, Toxoptera citricida (Homoptera:Aphididae). Biological Control 21, 91–95.
Morse, D.H., 1981. Interactions among syrphid flies and bumble bees on flowers.Ecology 62, 81–88.
Müller, C.B., Godfray, H.C.J., 1999. Predators and mutualists influence the exclusionof aphid species from natural communities. Oecologia 119, 120–125.
Pascual-Villalobos, M.J., Lacasa, A., Gonzalez, A., Varo, P., Garcia, M.J., 2006. Effect offlowering plant strips on aphid and syrphid populations in lettuce. EuropeanJournal of Agronomy 24, 182–185.
Pontin, D.R., Wade, M.R., Kehrli, P., Wratten, S.D., 2006. Attractiveness of single andmultiple species flower patches to beneficial insects in agroecosystems. Annalsof Applied Biology 148, 39–47.
Powell, W., Pennacchio, L.F., Poppy, G.M., Tremblay, E., 1998. Strategies involved inthe location of hosts by the parasitoid Aphidius ervi Haliday (Hymenoptera:Braconidae: Aphidiinae). Biological Control 11, 104–112.
Rojo, S., Marcos-Garcia, M.A., 1997. Syrphid predators (Dipt.: Syrphidae) of gallforming aphids (Hom.: Aphididae) in Mediterranean areas: implications forbiological control of fruit trees pests. Entomophaga 42, 269–276.
Smith, H.A., Chaney, W.E., 2007. A survey of syrphid predators of Nasonovia ribisnigriin organic lettuce on the Central Coast of California. Journal of EconomicEntomology 100, 39–48.
Smith, H.A., Chaney, W.E., Bensen, T.A., 2008. Role of syrphid larvae and otherpredators in suppressing aphid infestations in organic lettuce on California’sCentral Coast. Journal of Economic Entomology 101, 1526–1532.
Stephens, M.J., France, C.M., Wratten, S.D., Frampton, C., 1998. Enhancing biologicalcontrol of leafrollers (Lepidoptera: Tortricidae) by sowing buckwheat(Fagopyrum esculentum) in an orchard. Biocontrol Science and Technology 8,547–558.
Storeck, A., Poppy, G.M., van Emden, H.F., Powell, W., 2000. The role of plantchemical cues in determining host preference in the generalist aphidparasitoid Aphidius colemani. Entomologia Experimentalis et Applicata 97,41–46.
Taylor, R.M., Pfannenstiel, R.S., 2008. Nectar feeding by wandering spiders on cottonplants. Environmental Entomology 37, 996–1002.
Tenhumberg, B., Poehling, H., 1995. Syrphids as natural enemies of cereal aphids inGermany: aspects of their biology and efficacy in different years and regions.Agriculture, Ecosystems and Environment 52, 39–43.
Thomas, M.B., Wratten, S.D., Sotherton, N.W., 1991. Creation of island habitats infarmland to manipulate populations of beneficial arthropods: predatordensities and emigration. Journal of Applied Ecology 28, 906–917.
Tilman, D., Fargione, J., Wolff, B., D’Antonio, C., Dobson, A., Howarth, R., Schindler, D.,Schlesinger, W.H., Simberloff, D., Swackhamer, D., 2001. Forecasting agricul-turally driven global environmental change. Science 292, 281–284.
Tylianakis, J.M., Didham, R.K., Wratten, S.D., 2004. Improved fitness of aphidparasitoids receiving resource subsidies. Ecology 85, 658–666.
Vattala, H.D., Wratten, S.D., Phillips, C.B., Wäckers, F.L., 2006. The influence of flowermorphology and nectar quality on the longevity of a parasitoid biologicalcontrol agent. Biological Control 39, 179–185.
Winkler, K., Wackers, F., Bukovinszkine-Kiss, G., van Lenteren, J., 2006. Sugarresources are vital for Diadegma semiclausum fecundity under field conditions.Basic and Applied Ecology 7, 133–140.
Wyss, E., Villager, M., Müller-Schärer, H., 1999. The potential of three native insectpredators to control the rosy apple aphid, Dysaphis plantaginea. Biocontrol 44,171–182.




















