Assessing the trophic state of Linhos lake: a first step towards ecological rehabilitation
13
Journal of Environmental Management (2002) 64, 285–297 doi:10.1006/jema.2001.0521, available online at http://www.idealibrary.com on Assessing the trophic state of Linhos lake: a first step towards ecological rehabilitation R. Pereira * , A. M. V. M. Soares ‡ , R. Ribeiro ‡ and F. Gon ¸ calves Departamento de Biologia, Universidade de Aveiro, Campus Universitario de Santiago, 3810-193 Aveiro, Portugal ‡ Instituto do Ambiente e Vida, Departamento de Zoologia, Universidade de Coimbra, Portugal Received 1 June 2000; accepted 8 October 2001 Lack of recognition of the value of wetlands has led to the loss of considerable areas of these ecosystems in the past. Linhos lake (Figueira da Foz, Portugal) is a good example of one of these ecosystems, in which human intervention was responsible for its environmental degradation and led to its precocious terrestrialization. Physico-chemical conditions and zooplankton community structure were studied in Linhos lake, in order to evaluate ecosystem functioning and to acquire baseline information. The system is characterised by high oxygen depletion. Spatial heterogeneity was confirmed by the existence of significant differences in total densities for the three zooplanktonic groups. Rotifers were the most abundant group attaining their maximum density in April (2251Ð1 ind/l). Keratella quadrata, K. cochlearis, Polyarthra vulgaris, Filinia terminalis and Hexarthra mira were the main abundant species. Correspondence analysis suggested temperature as the main controlling factor in species seasonality. In order to prevent the precocious disappearance of the lake some restoration measures were proposed based on zooplankton community structure. 2002 Elsevier Science Ltd. Keywords: zooplankton community structure, lake terrestrialization, ecological rehabilitation adaptative management. Introduction At the beginning of the last century numer- ous small Portuguese freshwater lakes, between Oporto and Lisbon, near the coast, were drained in an attempt to eradicate the malaria vector that enjoyed, in central Portugal, ideal breeding habitats. One of these lakes, Linhos lake, almost disappeared in the process. More recently, the lake has been affected by the removal of groundwater by a paper mill. These activities contributed to a significant reduction of its surface area and depth and led to the development of a dense macrophyte community. Additionally, the enrichment of the sediments with organic matter provided annually by senescent macrophytes and their subsequent decomposition, depleted the oxygen and enhanced L Corresponding author. Email: [email protected] nutrient loading, particularly phosphorus, within the overlying water which thus became eutrophic (Pokorn ´ y, 1994). The loss of submerged vegeta- tion and reduction in biodiversity (Michaud et al., 1979; Hosper and Jagtman, 1990; Br¨ onmark and Weisner, 1992; Wetzel, 1993; Klinge et al., 1995), the occurrence of algal blooms (Michaud et al., 1979; Brix and Schierup, 1989; Hosper and Jagt- man, 1990; Klinge et al., 1995), and increase in turbidity (Klinge et al., 1995) are some of the characteristics associated with eutrophic shallow lakes, which seriously compromise their wildlife and recreational use. The eutrophication process remains a major problem in Linhos, as in most of the freshwater lakes of Portugal, and improvement strategies are required (e.g. Vasconcelos, 1994; Silva et al., 1997; Gon¸ calves et al., 1996). The understanding of ecosystem functioning is essential to decide which restoration measures are appropriate (Wetzel, 1993). However, planning and 0301–4797/02/030285C13 $35.00/0 2002 Elsevier Science Ltd.
Transcript of Assessing the trophic state of Linhos lake: a first step towards ecological rehabilitation

Journal of Environmental Management (2002) 64, 285–297doi:10.1006/jema.2001.0521, available online at http://www.idealibrary.com on
Assessing the trophic state of Linhos lake: afirst step towards ecological rehabilitation
R. Pereira*†, A. M. V. M. Soares†‡, R. Ribeiro‡ and F. Goncalves†
† Departamento de Biologia, Universidade de Aveiro, Campus Universitario de Santiago, 3810-193Aveiro, Portugal‡ Instituto do Ambiente e Vida, Departamento de Zoologia, Universidade de Coimbra, Portugal
Received 1 June 2000; accepted 8 October 2001
Lack of recognition of the value of wetlands has led to the loss of considerable areas of these ecosystems in the past.Linhos lake (Figueira da Foz, Portugal) is a good example of one of these ecosystems, in which human intervention wasresponsible for its environmental degradation and led to its precocious terrestrialization. Physico-chemical conditions andzooplankton community structure were studied in Linhos lake, in order to evaluate ecosystem functioning and to acquirebaseline information. The system is characterised by high oxygen depletion. Spatial heterogeneity was confirmed by theexistence of significant differences in total densities for the three zooplanktonic groups. Rotifers were the most abundantgroup attaining their maximum density in April (2251Ð1 ind/l). Keratella quadrata, K. cochlearis, Polyarthra vulgaris, Filiniaterminalis and Hexarthra mira were the main abundant species. Correspondence analysis suggested temperature asthe main controlling factor in species seasonality. In order to prevent the precocious disappearance of the lake somerestoration measures were proposed based on zooplankton community structure. 2002 Elsevier Science Ltd.
Keywords: zooplankton community structure, lake terrestrialization, ecological rehabilitation adaptativemanagement.
Introduction
At the beginning of the last century numer-ous small Portuguese freshwater lakes, betweenOporto and Lisbon, near the coast, were drainedin an attempt to eradicate the malaria vectorthat enjoyed, in central Portugal, ideal breedinghabitats. One of these lakes, Linhos lake, almostdisappeared in the process. More recently, the lakehas been affected by the removal of groundwaterby a paper mill. These activities contributed to asignificant reduction of its surface area and depthand led to the development of a dense macrophytecommunity. Additionally, the enrichment of thesediments with organic matter provided annuallyby senescent macrophytes and their subsequentdecomposition, depleted the oxygen and enhanced
Ł Corresponding author. Email: [email protected]
nutrient loading, particularly phosphorus, withinthe overlying water which thus became eutrophic(Pokorny, 1994). The loss of submerged vegeta-tion and reduction in biodiversity (Michaud et al.,1979; Hosper and Jagtman, 1990; Bronmark andWeisner, 1992; Wetzel, 1993; Klinge et al., 1995),the occurrence of algal blooms (Michaud et al.,1979; Brix and Schierup, 1989; Hosper and Jagt-man, 1990; Klinge et al., 1995), and increase inturbidity (Klinge et al., 1995) are some of thecharacteristics associated with eutrophic shallowlakes, which seriously compromise their wildlifeand recreational use. The eutrophication processremains a major problem in Linhos, as in most ofthe freshwater lakes of Portugal, and improvementstrategies are required (e.g. Vasconcelos, 1994;Silva et al., 1997; Goncalves et al., 1996).
The understanding of ecosystem functioning isessential to decide which restoration measures areappropriate (Wetzel, 1993). However, planning and
0301–4797/02/030285C13 $35.00/0 2002 Elsevier Science Ltd.

286 R. Pereira et al.
restoration of a natural area need not be inhibitedby a lack of scientific information, if an adaptativemanagement process is adopted. This is a processin which resource managers plan managementactivities continually, monitor and adjust them tomeet their goals (Marcin, 1995).
Adequate restoration measures for many lakesshould focus on the control of emergent macrophytestanding crops. Wetlands dominated by perennialmacrophytes such as Phragmites, are one of theworld’s most productive ecosystems (Bjork, 1994a).Its overgrowth is continually stimulated by therecycling of nutrients from the sediments, whichconstrains the number of trophic levels and top-down forces that in turn regulate their standingcrops (Power, 1992; Bjork, 1994a). Commonly,additional measures should also be taken toreduce phosphorus internal loading. Phosphorusis a key element in freshwater lakes (Hosper andJagtman, 1990; Seip, 1994) since its concentrationin those systems is lower than its biologicaldemand (Wetzel, 1993). Zooplankton constitutesa sensitive tool for monitoring eutrophication(Pejler, 1983; Magadza, 1994; Silva et al., 1997),because populations react immediately to changesin trophic status. The information obtained fromthis study will allow the monitoring of the effectsof intended restoration measures.
Eutrophication usually affects both the physicaland chemical environment and can lead to signif-icant changes in the phytoplankton and zooplank-ton communities’ structure (Flores and Barone,1994; Uku and Mavuti, 1994). Generally, this pro-cess results in a zooplankton community dominatedby rotifers and small bodied cladocerans (Gliwicz,1990; Matsubara, 1993; Ejsmont-Karabin, 1995)which are less vulnerable to feeding interferenceby large inedible algae species (Gliwicz, 1990; Aueret al., 1990) and capable of feeding on detritus andbacterial size particles (Bogdan and Gilbert, 1982).The aim of the present study was to develop thefirst set of measures for the ecological rehabilita-tion of Linhos lake. In order to meet this goal, thezooplankton community structure was character-ized and inferences made about how the physicaland chemical conditions of the lake affect it.
Material and methods
Study site
The Lake is a small (10 ha), shallow (1Ð43 m meandepth; 2Ð3 m maximum depth) and eutrophic water
Oporto
Figueira daFoz
Linhos lake
SPAIN
ATLANTICOCEAN
Lisbon
12
3
N
Figure 1. Location of Linhos lake.
body, located 15 km South of Figueira da Foz (Por-tugal) (Figure 1). A large community of emergentmacrophytes was present, with Phragmites aus-tralis (CAV.) Trin. Ex. Steudel and Typha latifoliaL. as the dominant species. At the end of the grow-ing season, senescent macrophytes fall into the lakeforming a completely anoxic organic layer (approxi-mately 1Ð5 m thickness, personal observation). Thebrownish coloured water of the Lake is indicativeof the presence of high concentrations of dissolvedorganic compounds. Submerged macrophytes wereabsent. A dense forest consisting largely of Pinuspinaster Aiton and Eucalyptus globulus Labill anda marsh zone, characterised by high species diver-sity, surround the lake, isolating it from humanactivity, hence offering good conditions for wildlife,especially migratory birds.
Sampling procedures
Three sampling sites were defined in the Lake.Sites 1 and 3 were located near the emergent vege-tation and site 2 in open water. Samples for waterchemistry and zooplankton determinations werecollected monthly from the surface, on each sam-pling site, during 12 months. Dissolved oxygen,Secchi disk transparency, conductivity, tempera-ture and pH were recorded in situ every month.Temperature (š0Ð5°C) and pH (š0Ð02 pH) weremeasured by Jenway 3150 pH meter. The conduc-tivity was measured by a WTW LF92 conductivity

Assessing the trophic state of Linhos lake 287
meter. Dissolved oxygen (mg/l,š0Ð1 mg/l) was mea-sured by a WTW OXI92 oxygen meter. From Marchto December 1996 vertical profiles for dissolvedoxygen were obtained measuring this parameterat 25 cm intervals. Ammonium (mg/l) was deter-mined by the Nessler method, nitrate (mg/l) bycadmium reduction, nitrite (mg/l) by a colorimet-ric method and phosphorus (mg/l) following theascorbic acid method (APHA et al., 1989). Forchlorophyll-a analysis, water samples were filteredthrough Whatman GF/C filters (0Ð45µm porosity;47 mm diameter). Filters were ground in 90% ace-tone and stored at 4°C for 24 h. After centrifugation,absorbency was measured at 664 nm and 750 nmbefore adding HCl and at 750 nm and 665 nm afteracidification by a Jenway 6100 spectrophotome-ter. Chlorophyll-a concentration was calculatedand total suspended solids determined accordingto the methods of the APHA, AWWA and WPCF(1989).
Zooplankton samples were collected in a 1Lbottle. Ten liters of water were filtered, per sample,through a plankton net (20 mm mesh). Each samplewas immediately preserved in borate buffered with4% formalin. Three replicates per sampling sitewere collected for rotifer observation, and six forcladocerans and copepods.
In the laboratory, copepods (Dussart, 1969) andcladocerans (Amoros, 1984; Scourfield and Hard-ing, 1966) were identified and counted under adissection microscope (50ð). For rotifers, sedimen-tation chambers were used, under an invertedmicroscope (Ruttner-Kolisko, 1974).
Statistical analysis
One-way ANOVA was used to detect the existenceof significant differences among sites for environ-mental parameters and in total densities of rotifers,cladocerans and copepods, during each month.In order to meet the basic assumptions requiredfor a valid analysis of variance (Zar, 1996), totaldensities of the three zooplankton groups weretransformed by the equation: x0DLog.xC1/.
The zooplankton assemblages were examinedfor their taxonomic diversity using the Shan-non–Wiener index (H0). An equitability index (J0)was also calculated to complement the informationprovided by the diversity index, since the J0 is use-ful to detect the presence of new species in thesystem (Legendre and Legendre, 1979; Washing-ton, 1984). These indices were applied to generasince in some cases identification to species levelwas not possible (Washington, 1984).
Resemblances between the zooplankton assem-blages were analysed by correspondence analysis(CA) using NT-SYS 1Ð8 (Rohlf, 1992). Months andspecies were plotted together to ascertain out theexistence of gradients leading to species distribu-tion.
To assess positive links between the most abun-dant taxa and environmental parameters, simpleregression analysis was performed with the trans-formed data. Regression models were compared(regression coefficients and intercepts) by analy-sis of covariance. Whenever null hypothesis wasrejected, a multiple comparison test (Tukey test)was used to determine which slopes and interceptsdiffered from each other (Zar, 1996).
Results
Environmental parameters
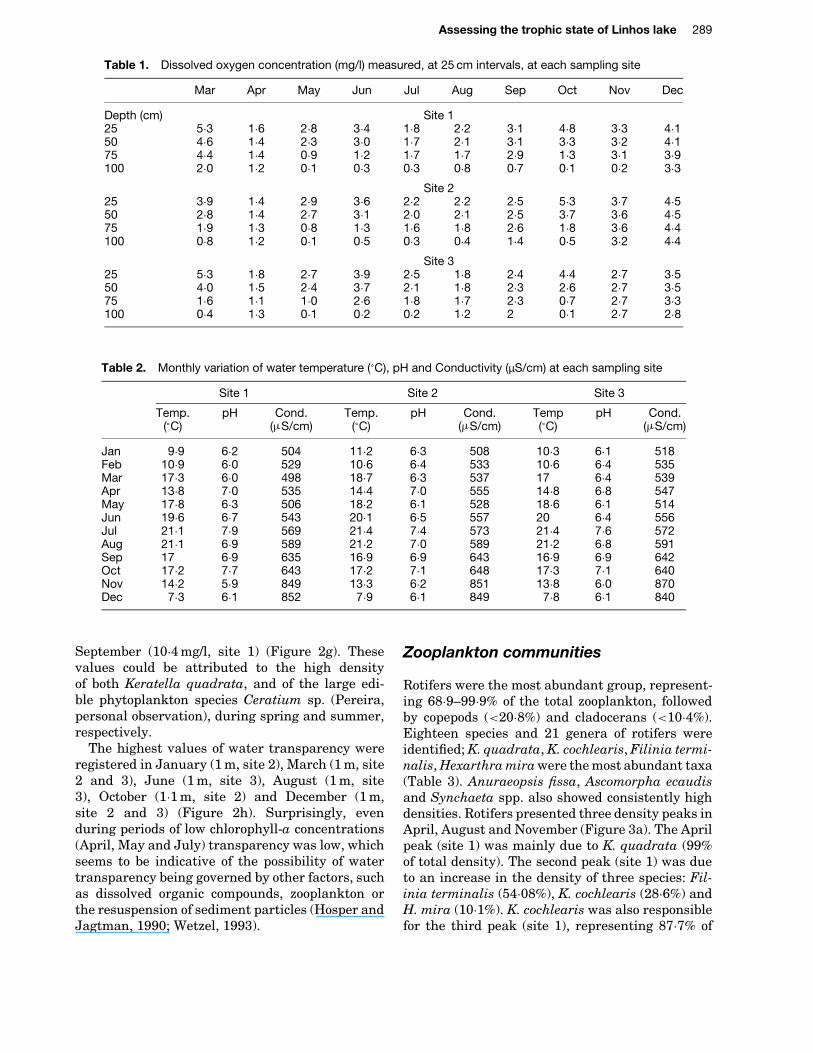
Linhos lake (henceforth the Lake) is characterisedby strong dissolved oxygen depletion. Dissolvedoxygen concentrations vary between 0Ð3 mg/l and5Ð3 mg/l (Figure 2a). The highest values wererecorded in March (5Ð3 mg/l, sites 1 and 3) andOctober (5Ð3 mg/l, site 3). Dissolved oxygen profilesdeclined with increasing depth, indicative of thewidespread anoxia in the deep water and surfacesediments (Table 1).
Water temperature ranged between 7Ð3°C(December) and 21Ð4°C (July) (Table 2). The Lakedid not exhibit temperature stratification duringthe study period (Table 2).
The pH values ranged between 5Ð9 and 7Ð9. Thehigher values were registered in July (7Ð93) andOctober (7Ð65) (Table 2).
Specific conductance in lake water averaged�870 µS/cm (Table 2); a gradual increase from thebeginning to the end of the study was observed.
Chlorophyll-a concentrations peaked in January(11Ð0 µ/l, site 1). High values were also registered inMarch (9Ð6 µ/l, site 2; 7Ð6 µg/l, site 3), June (7Ð4 µg/l,site 1; 8Ð0 µg/l, site 2), and November (10Ð4 µg/l, site1; 10Ð0 µg/l, site 3) (Figure 2b). Seasonal variationin chlorophyll was not observed.
Nitrites were the least abundant nitrogen com-pound. Their concentrations were higher in thefirst four months, especially in February (15 µg/l;site 1). Values of 0 µg/l were obtained during latespring and summer (Figure 2c).
As with nitrites, nitrates showed highestconcentrations during winter, decreasing graduallyin spring, reaching lowest values in June (0Ð2 mg/l,site 2). During the final six months

288 R. Pereira et al.
0D
6
Month
(a)
Dis
solv
ed o
xyge
n (
mg/
l)
J
12345
F M A M J J A S O N0
D
12
Month
(b)
Ch
loro
phyl
l-α
(ng/
m3 )
J
2468
10
F M A M J J A S O N
0D
16
Month
(c)
Nit
rite
s (µ
g/l)
J
2468
12
F M A M J J A S O N0
D
1.6
Month
(d)
Nit
rate
s (m
g/l)
J
0.20.40.60.8
1.2
F M A M J J A S O N
0D
1.8
Month
(e)
Am
mon
ia (
mg/
l)
J
0.20.40.6
1.0
1.4
F M A M J J A S O N0
D
1.4
Month
(f)
Ph
osph
oru
s (m
g/l)
J
0.20.40.60.8
1
F M A M J J A S O N
0D
12
Month
(g)
T.S.
S. (
mg/
l)
J
2468
10
F M A M J J A S O N0
D
1.2
Month
(h)
Wat
er t
ran
spar
ency
(m
)
J
0.20.40.60.8
1
F M A M J J A S O N
14
10
1.4
1
1.6
1.2
0.8
1.2
Figure 2. Annual variation, within each sampling site of: (a) dissolved oxygen; (b) chlorophyll-a; (c) nitrite concentrations;(d) nitrate concentrations; (e) ammonia concentrations; (f) phosphorus concentrations; (g) total suspended solidsconcentrations; (h) water transparency. ( ) Site 1; ( ) Site 2; ( ) Site 3.
(July–December), nitrates displayed almost novariation (Figure 2d).
Ammonia concentrations showed small varia-tions throughout the year, with the exception oftwo peak values: July (1Ð7 mg/l, site 1) and Decem-ber (0Ð91 mg/l, site 1) (Figure 2e).
Phosphorus concentrations were higher inFebruary (xD1Ð07 mg/l) and March (xD1Ð04 mg/l).Throughout the last six months of the year,concentrations were very low, varying between0Ð06 mg/l and 0Ð001 mg/l (Figure 2f); suggestingpossible removal from the water through biological
uptake by macrophytes and phytoplankton. Thelast group was characterised in those monthsby a great abundance of large edible algae(Pereira, personal observation). Bacteria andmicroplankton could also be responsible, throughrapid immobilization of phosphorus from surfacewaters, during the growing season (Freedman,1989).
Four peaks of Total Suspended Solids (TSS) wererecorded in March (10Ð4 mg/l, site 1; 10Ð3 mg/l,site 3), April (9Ð2 mg/l, site 1; 10Ð1 mg/l site 2),August (9Ð1 mg/l, site 1; 10Ð9 mg/l, site 3) and
Assessing the trophic state of Linhos lake 289
Table 1. Dissolved oxygen concentration (mg/l) measured, at 25 cm intervals, at each sampling site
Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Depth (cm) Site 125 5Ð3 1Ð6 2Ð8 3Ð4 1Ð8 2Ð2 3Ð1 4Ð8 3Ð3 4Ð150 4Ð6 1Ð4 2Ð3 3Ð0 1Ð7 2Ð1 3Ð1 3Ð3 3Ð2 4Ð175 4Ð4 1Ð4 0Ð9 1Ð2 1Ð7 1Ð7 2Ð9 1Ð3 3Ð1 3Ð9100 2Ð0 1Ð2 0Ð1 0Ð3 0Ð3 0Ð8 0Ð7 0Ð1 0Ð2 3Ð3
Site 225 3Ð9 1Ð4 2Ð9 3Ð6 2Ð2 2Ð2 2Ð5 5Ð3 3Ð7 4Ð550 2Ð8 1Ð4 2Ð7 3Ð1 2Ð0 2Ð1 2Ð5 3Ð7 3Ð6 4Ð575 1Ð9 1Ð3 0Ð8 1Ð3 1Ð6 1Ð8 2Ð6 1Ð8 3Ð6 4Ð4100 0Ð8 1Ð2 0Ð1 0Ð5 0Ð3 0Ð4 1Ð4 0Ð5 3Ð2 4Ð4
Site 325 5Ð3 1Ð8 2Ð7 3Ð9 2Ð5 1Ð8 2Ð4 4Ð4 2Ð7 3Ð550 4Ð0 1Ð5 2Ð4 3Ð7 2Ð1 1Ð8 2Ð3 2Ð6 2Ð7 3Ð575 1Ð6 1Ð1 1Ð0 2Ð6 1Ð8 1Ð7 2Ð3 0Ð7 2Ð7 3Ð3100 0Ð4 1Ð3 0Ð1 0Ð2 0Ð2 1Ð2 2 0Ð1 2Ð7 2Ð8
Table 2. Monthly variation of water temperature (°C), pH and Conductivity (µS/cm) at each sampling site
Site 1 Site 2 Site 3
Temp. pH Cond. Temp. pH Cond. Temp pH Cond.(°C) (mS/cm) (°C) (mS/cm) (°C) (mS/cm)
Jan 9Ð9 6Ð2 504 11Ð2 6Ð3 508 10Ð3 6Ð1 518Feb 10Ð9 6Ð0 529 10Ð6 6Ð4 533 10Ð6 6Ð4 535Mar 17Ð3 6Ð0 498 18Ð7 6Ð3 537 17 6Ð4 539Apr 13Ð8 7Ð0 535 14Ð4 7Ð0 555 14Ð8 6Ð8 547May 17Ð8 6Ð3 506 18Ð2 6Ð1 528 18Ð6 6Ð1 514Jun 19Ð6 6Ð7 543 20Ð1 6Ð5 557 20 6Ð4 556Jul 21Ð1 7Ð9 569 21Ð4 7Ð4 573 21Ð4 7Ð6 572Aug 21Ð1 6Ð9 589 21Ð2 7Ð0 589 21Ð2 6Ð8 591Sep 17 6Ð9 635 16Ð9 6Ð9 643 16Ð9 6Ð9 642Oct 17Ð2 7Ð7 643 17Ð2 7Ð1 648 17Ð3 7Ð1 640Nov 14Ð2 5Ð9 849 13Ð3 6Ð2 851 13Ð8 6Ð0 870Dec 7Ð3 6Ð1 852 7Ð9 6Ð1 849 7Ð8 6Ð1 840
September (10Ð4 mg/l, site 1) (Figure 2g). Thesevalues could be attributed to the high densityof both Keratella quadrata, and of the large edi-ble phytoplankton species Ceratium sp. (Pereira,personal observation), during spring and summer,respectively.
The highest values of water transparency wereregistered in January (1 m, site 2), March (1 m, site2 and 3), June (1 m, site 3), August (1 m, site3), October (1Ð1 m, site 2) and December (1 m,site 2 and 3) (Figure 2h). Surprisingly, evenduring periods of low chlorophyll-a concentrations(April, May and July) transparency was low, whichseems to be indicative of the possibility of watertransparency being governed by other factors, suchas dissolved organic compounds, zooplankton orthe resuspension of sediment particles (Hosper andJagtman, 1990; Wetzel, 1993).
Zooplankton communities
Rotifers were the most abundant group, represent-ing 68Ð9–99Ð9% of the total zooplankton, followedby copepods (<20Ð8%) and cladocerans (<10Ð4%).Eighteen species and 21 genera of rotifers wereidentified; K. quadrata, K. cochlearis, Filinia termi-nalis, Hexarthra mira were the most abundant taxa(Table 3). Anuraeopsis fissa, Ascomorpha ecaudisand Synchaeta spp. also showed consistently highdensities. Rotifers presented three density peaks inApril, August and November (Figure 3a). The Aprilpeak (site 1) was mainly due to K. quadrata (99%of total density). The second peak (site 1) was dueto an increase in the density of three species: Fil-inia terminalis (54Ð08%), K. cochlearis (28Ð6%) andH. mira (10Ð1%). K. cochlearis was also responsiblefor the third peak (site 1), representing 87Ð7% of

290 R. Pereira et al.
Table 3. Rotifers, cladocerans and copepods identified for Linhos lake
J F M A M J J A S O N D
RotatoriaBrachionus patulus Muller
Brachionus angularis Gosse
Brachionus falcatus Zacharias
Brachionus bidentatus Andersen
Brachionus calyciflorus (Pallas)
Platyas quadricornis Ehrenberg
Keratella quadrata Muller C CC
Keratella valga Carlin
Keratella cochlearis Gosse C C CC C C CC CCAnuraeopsis fissa Gosse C
Euchlanis spp.
Lepadella spp. C
Squatinella sp.
Lecane quadridentata Ehrenberg
Lecane spp.
Mytilina sp.
Trichotria sp.
Monommata sp.
Scaridium sp.
Trichocerca elongata Gosse
Trichocerca spp.
Ascomorpha ecaudis Perty C
Asplancna sp.
Synchaeta spp. C C C C C C C
Polyarthra vulgaris Carlin C C C
Testudinella sp.
Pompholix sp.
Filinia hofmanni Koste
Filinia terminalis Plate C CC CC C
Filinia opoliensis Zacharias
Hexarthra mira Hudson C
CladoceraDiaphanosoma brachyurum Lieven
Ceriodaphnia pulchella Sars
Bosmina longirostris (O. F. Muller)
Alona protzi Hartwing
Alona rectangula Sars
Alona tenuicaudis Sars
Alona weltneri Keilhack
Alona costata Sars
Alonella nana (Baird)
Alonella exigua Lilljeborg
Chydorus sphaericus Muller
Ilyocryptus sordidus (Lieven)
CopepodaAcanthocyclops robustus G.O. Sars
Copidodiaptomus numidicus Gurney
Copepodits
Nauplii C C C C C C C CHarpaticoids
( �ind/l<10; C �10<ind/l<100; CC � ind/l>100).
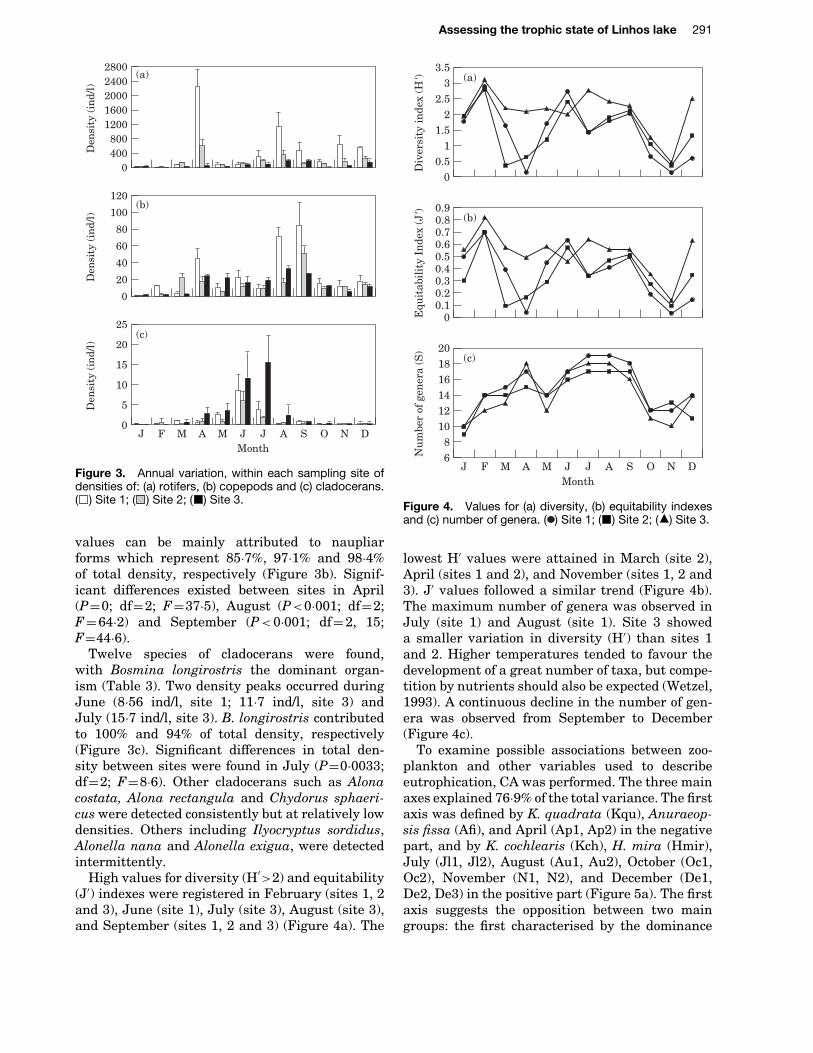
total density (Figure 3a). Significant differencesin density between sites were found in April(PD0Ð0002; dfD2; FD48Ð1), August (PD0Ð0008;dfD2; FD29Ð2), and November (PD0Ð0008; dfD2;FD29Ð3).
Copepods were represented by two species: Acan-thocyclops robustus and Copidodiaptomus numidi-cus (Table 3). Maximum densities were recordedin April (42Ð6 ind/l, site 1), August (70Ð4 ind/l,site 1), and September (83Ð5 ind/l, site 1). These
Assessing the trophic state of Linhos lake 291
0
2800
Den
sity
(in
d/l)
400800
1200160020002400
0
120
Den
sity
(in
d/l)
20
40
60
80
100
0D
25
Month
Den
sity
(in
d/l)
J
5
10
15
20
F M A M J J A S O N
(c)
(b)
(a)
Figure 3. Annual variation, within each sampling site ofdensities of: (a) rotifers, (b) copepods and (c) cladocerans.( ) Site 1; ( ) Site 2; ( ) Site 3.
values can be mainly attributed to naupliarforms which represent 85Ð7%, 97Ð1% and 98Ð4%of total density, respectively (Figure 3b). Signif-icant differences existed between sites in April(PD0; dfD2; FD37Ð5), August (P<0Ð001; dfD2;FD64Ð2) and September (P<0Ð001; dfD2, 15;FD44Ð6).
Twelve species of cladocerans were found,with Bosmina longirostris the dominant organ-ism (Table 3). Two density peaks occurred duringJune (8Ð56 ind/l, site 1; 11Ð7 ind/l, site 3) andJuly (15Ð7 ind/l, site 3). B. longirostris contributedto 100% and 94% of total density, respectively(Figure 3c). Significant differences in total den-sity between sites were found in July (PD0Ð0033;dfD2; FD8Ð6). Other cladocerans such as Alonacostata, Alona rectangula and Chydorus sphaeri-cus were detected consistently but at relatively lowdensities. Others including Ilyocryptus sordidus,Alonella nana and Alonella exigua, were detectedintermittently.
High values for diversity (H0>2) and equitability(J0) indexes were registered in February (sites 1, 2and 3), June (site 1), July (site 3), August (site 3),and September (sites 1, 2 and 3) (Figure 4a). The
6D
20
Month
(c)
Nu
mbe
r of
gen
era
(S)
J
810121416
F M A M J J A S O N
18
0
0.9(b)
Equ
itab
ilit
y In
dex
(J')
0.10.20.3
0.5
0.70.8
0
3.5(a)
Div
ersi
ty in
dex
(H')
0.51
1.52
2.53
0.4
0.6
Figure 4. Values for (a) diversity, (b) equitability indexesand (c) number of genera. ( ) Site 1; ( ) Site 2; ( ) Site 3.
lowest H0 values were attained in March (site 2),April (sites 1 and 2), and November (sites 1, 2 and3). J0 values followed a similar trend (Figure 4b).The maximum number of genera was observed inJuly (site 1) and August (site 1). Site 3 showeda smaller variation in diversity (H0) than sites 1and 2. Higher temperatures tended to favour thedevelopment of a great number of taxa, but compe-tition by nutrients should also be expected (Wetzel,1993). A continuous decline in the number of gen-era was observed from September to December(Figure 4c).
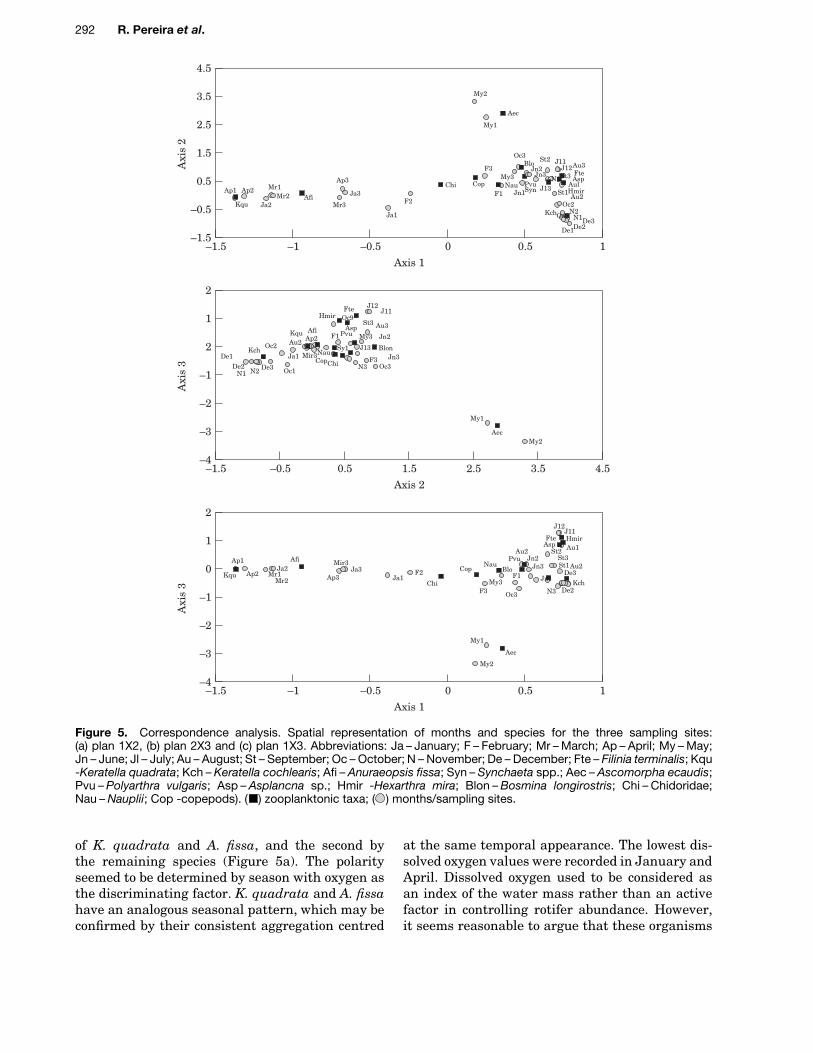
To examine possible associations between zoo-plankton and other variables used to describeeutrophication, CA was performed. The three mainaxes explained 76Ð9% of the total variance. The firstaxis was defined by K. quadrata (Kqu), Anuraeop-sis fissa (Afi), and April (Ap1, Ap2) in the negativepart, and by K. cochlearis (Kch), H. mira (Hmir),July (Jl1, Jl2), August (Au1, Au2), October (Oc1,Oc2), November (N1, N2), and December (De1,De2, De3) in the positive part (Figure 5a). The firstaxis suggests the opposition between two maingroups: the first characterised by the dominance
292 R. Pereira et al.
–1.51
4.5
Axis 1
Axi
s 2
–1.5
–0.5
0.5
1.5
2.5
3.5
0.50–0.5–1
Afi
–44.5
2
Axis 2
Axi
s 3
–1.5
–3
–2
–1
2
1
2.51.50.5–0.5
De1
–41
2
Axis 1
Axi
s 3
–1.5
–3
–2
–1
0
1
0.50–0.5–1
Afi
3.5
Mr2Mr1
Ja2Kqu
Ap2Ap1
Mr3
Ja3
Ap3
Ja1
F2
Chi Cop
F3My3
NauJn1Syn
PvuJn3
Jn2Blo
Oc3 St2
F1J13
J11
My1
Aec
My2
J12Au3
St3 FteAsp
AulSt1Hmir
Au2Oc2
N2N1De3De2De1
Oc3Kch
N2
De2N1 N2 De3
KchOc2
Oc1
Ja1
Au2Kqu Afi
Ap2
Mir3NauCopChi
Sy1
F1 Pvu
Hmir Oc9
Asp
Fte J12J11
St3 Au3
My3 Jn2
J13 Blon
Oc3F3
N3Jn3
My1
AecMy2
Mir3
Ap3Ja2
Mr1Mr2
Ap2
Ap1
KquJa3
Ja1F2
Chi
CopNau
My3
Oc3
F1Blo
Pvu
F3
Au2Jn2
Jn3
Jn1
N3 De2
De3St1St3
Au1
Au2
Kch
HmirJ11
J12
FteAsp
St2
My1
Aec
My2
Figure 5. Correspondence analysis. Spatial representation of months and species for the three sampling sites:(a) plan 1X2, (b) plan 2X3 and (c) plan 1X3. Abbreviations: Ja – January; F – February; Mr – March; Ap – April; My – May;Jn – June; Jl – July; Au – August; St – September; Oc – October; N – November; De – December; Fte – Filinia terminalis; Kqu-Keratella quadrata; Kch – Keratella cochlearis; Afi – Anuraeopsis fissa; Syn – Synchaeta spp.; Aec – Ascomorpha ecaudis;Pvu – Polyarthra vulgaris; Asp – Asplancna sp.; Hmir -Hexarthra mira; Blon – Bosmina longirostris; Chi – Chidoridae;Nau – Nauplii; Cop -copepods). ( ) zooplanktonic taxa; ( ) months/sampling sites.
of K. quadrata and A. fissa, and the second bythe remaining species (Figure 5a). The polarityseemed to be determined by season with oxygen asthe discriminating factor. K. quadrata and A. fissahave an analogous seasonal pattern, which may beconfirmed by their consistent aggregation centred
at the same temporal appearance. The lowest dis-solved oxygen values were recorded in January andApril. Dissolved oxygen used to be considered asan index of the water mass rather than an activefactor in controlling rotifer abundance. However,it seems reasonable to argue that these organisms

Assessing the trophic state of Linhos lake 293
Table 4. Regression analysis of temperature, total suspended solids and chlorophyll-a on taxa densities [log(DENC1/DaCblog.EFC1/] (DEN – taxa densities; EF – environmental factor)
N r2 r F P a b
TemperatureH. mirasites 1, 3 71 18Ð89 0Ð44 16Ð401 0Ð000 �4Ð136š1Ð164 1Ð689š0Ð417site 2 35 30Ð04 0Ð55 14Ð596 0Ð001 �4Ð430š1Ð294 1Ð761š0Ð461
F. terminalissites 1, 2 71 36Ð75 0Ð61 40Ð675 0Ð000 �10Ð947š1Ð956 4Ð461š0Ð699site 3 35 18Ð44 0Ð43 7Ð686 0Ð009 �5Ð894š2Ð457 2Ð433š0Ð878
B.longirostrissites 1, 2 143 18Ð99 0Ð44 33Ð296 0Ð000 �2Ð361š0Ð490 1Ð011š0Ð175site 3 71 34Ð29 0Ð59 36Ð527 0Ð000 �3Ð804š0Ð740 1Ð597š0Ð264
Naupliisites 1, 2 143 11Ð34 0Ð34 18Ð166 0Ð000 �1Ð086š0Ð844 1Ð287š0Ð302site 3 71 30Ð94 0Ð56 31Ð367 0Ð000 �2Ð738š0Ð917 1Ð836š0Ð328
Total suspended solids
Naupliisite 1, 2 143 50Ð10 0Ð71 142Ð562 0Ð000 �0Ð113š0Ð228 1Ð500š0Ð126site 3 71 9Ð68 0Ð31 7Ð505 0Ð008 1Ð072š0Ð487 0Ð745š745Ð272
H.mirasites 1, 2 71 8Ð61 0Ð29 6Ð594 0Ð012 �0Ð530š0Ð461 0Ð653š0Ð254site 3 35 21Ð21 0Ð46 9Ð155 0Ð005 �1Ð252š0Ð554 0Ð938š0Ð310
Chlorophyll-a
Naupliisites 1, 3 143 18Ð01 �0Ð42 31Ð188 0Ð000 3Ð896š0Ð262 �0Ð782š0Ð140site 2 71 28Ð15 0Ð53 27Ð424 0Ð000 0Ð874š0Ð298 0Ð970š0Ð185
are influenced by dissolved oxygen, being tolerantto low concentrations.
The second axis was defined by K. cochlearis(Kch) and December (De1, De2, De3), in the neg-ative part, and by Ascomorpha ecaudis (Aec) andMay (My1, My2), in the positive part (Figure 5b).The third axis was defined by Ascomorpha ecaudis(Aec) and May (My1, My2), in the negative part,and by Filinia terminalis (Fte) and July (Jl1, Jl2)in the positive part (Figure 5c).
Statistically significant links (P<0Ð05) betweendensities of the most abundant taxa and envi-ronmental factors were found only for H. mira,F. terminalis, Bosmina longirostris and Nauplii.Analysis of covariance (ANCOVA) also showed sig-nificant differences between slopes for the threesampling sites.
Temperature accounted for the total variabilityin the distribution of those taxa, namely: 19Ð0%(P<0Ð001, sites 1, 3) and 30Ð0% (P<0Ð001, site2) for Hexarthra mira; 36Ð8% (P<0Ð001, sites 1,2) and 18Ð4 (PD0Ð00897, site 3) for Filinia ter-minalis; 19Ð0% (P<0Ð001, sites 1, 2) and 34Ð3%(P<0Ð001, site 3) for Bosmina longirostris; 11Ð3%
(P<0Ð001, sites 1, 2) and 30Ð9% (P<0Ð001, site3) for naupliar forms (Table 4). However, thedistribution of naupliar forms was mainly asso-ciated with total suspended solids in site 1 and2 (r2D50Ð1%, P<0Ð001). Chlorophyll-a also for18Ð01% (P<0Ð001, sites 1, 3) and 28Ð2% (P<0Ð001,site 2) in the overall variation of naupliar densi-ties (Table 4).
The abundance of Hexarthra mira was lessstrongly related to total suspended solids (r2D21Ð2%, PD0Ð0047, site 3; r2D8Ð6%, PD0Ð0124, sites1, 2) (Table 4).
Discussion
According to the classification proposed by Freed-man (1989) for the trophic category of lakes andinland waters, the Lake could be considered tobe hypereutrophic, based on its mean annualphosphorus concentration (282 µg/l) and mini-mum annual transparency (0Ð5 m). However, meanannual chlorophyll-a concentration (4Ð89 mg/m3)places the Lake in the mesotrophic category.

294 R. Pereira et al.
These observations suggested that, despite appar-ent nutrient availability, phytoplankton growthwas inhibited in the Lake. This suggests a shiftfrom a primary productivity based on phytoplank-ton to one based on macrophytes in natural ageinglakes (Bjork, 1994a). Macrophytes could be themain factor responsible for phytoplankton inhibi-tion in the Lake, through competition for lightand nutrients (Moss, 1990; Wetzel, 1993; Jasser,1995). Dissolved organic compounds resulting fromorganic matter mineralization may also createshade conditions and could also be a limiting factorto phytoplanktonic productivity (Carpenter et al.,1998). Chlorophyll-a concentrations recorded inthis study were lower than those registered byGoncalves et al. (1996) in Mira lake, and by Vas-concelos (1990) (27 mg/m3 in June), in Bracas lake,two eutrophic freshwater coastal lakes, located afew kilometres from Linhos. In the Lake, the densecoverage of macrophytes played an important rolein water quality degradation. The large demandfor oxygen to sustain the decomposition of thisorganic material accounted for the low values fordissolved oxygen obtained at depth. The decom-position of particulate organic matter is the mainoxygen consuming process in deeper water (Wetzel,1993).
Rotifers were by far the most dominant zoo-planktonic group. A similar observation wasrecorded for some Sicilian mesotrophic water bod-ies (Flores and Barone, 1994). Their dominancein the Lake probably arose from their wide toler-ance to variable dissolved oxygen concentrations(Galkovskaya, 1995) and from their capability tofeed on bacteria and detritus (Ruttner-Kolisko,1974; Habdija et al., 1993). The most abundantspecies – Keratella quadrata, K. cochlearis, Pol-yarthra vulgaris, Filinia terminalis and Hexarthramira – are abundant in water bodies with a widerange of physico-chemical factors and are able touse bacteria and detritus in suspension as foodresources (Bogdan and Gilbert, 1982; Zankai, 1989;Habdija et al., 1993; Ooms-Wilms, 1997). Macro-phyte source detritus is likely the most importantfood source for rotifers in the Lake, since no cor-relations were found between these species andchlorophyll-a concentrations. In natural conditionsbacteria are difficult to assimilate; their food qual-ity is poor as they lack some essential compoundsfor animal nutrition (Ooms-Wilms, 1997). The rel-ative abundance of Filinia and Hexarthra, giventheir ability to feed on detritus, has been sug-gested as an indicator for mesotrophic water bodies(Maemets, 1983). The representation of the cope-pod group almost exclusively by naupliar forms
may be explained by a significant extended devel-opment times of these stages. This phenomenon isusually observed to be the result of a lack of goodfood resources (Hart, 1990).
The CA biplot of species and months showedthree main groupings, each species grouped withits highest density month. However, species distri-bution follows a temperature gradient along axis 2(Figure 5a), suggesting that temperature was themost important factor in determining seasonality.Regression analysis also indicated the influenceof temperature on the abundance of Hexarthramira, Filinia terminalis, Bosmina longirostris andnaupliar forms, throughout the year.
B. longirostris (the most abundant cladoceran)displayed higher densities in June–July, afterwhich it started to decrease. This could beexplained by lower levels of dissolved oxygen andalso by the high densities of Ceratium sp., whichwere recorded in zooplankton samples from Augustto September. This large edible species could notbe eaten by small cladocerans, such as Bosmina(Auer et al., 1990).
The pattern of occurrence of F. terminalis wassomewhat surprising. This species, consideredas a cold stenothermic and hypolimnetic form(Ruttner-Kolisko, 1974), showed a density peakduring June–July. This could be explained bythe conjugation of high food availability with lowoxygen concentrations.
Despite its small area, significant differences intotal densities between sites for the three zooplank-tonic groups were found in the Lake during densitypeaks. However, this could not be attributed to theenvironmental parameters measured since signif-icant differences among sampling sites were notfound. The efficiency with which different micro-crustacean species can utilise non algal resourcessuch as bacteria, protozoan (ciliates) and detritusmay contribute to how the species selects the habi-tat (Smiley and Tessier, 1998). This was confirmedonly for naupliar forms, whose distribution wasrelated to that of chlorophyll-a (Table 4) and totalsuspended solids (Table 4). A positive correlationwas also found between H. mira densities and totalsuspended solids (Table 4) suggesting that detrituscould be the main nutrient source for those organ-isms. Total densities of zooplanktonic groups alsotended to increase from site 3 to site 1, which maybe due to passive drift within the small water area,as an increase in water level induces flow fromsite 3 to site 1. Wind can also contribute to thisspatial distribution, concentrating zooplankton insome areas.

Assessing the trophic state of Linhos lake 295
With regard to species diversity, values regis-tered for the Lake were low, as expected. Reducedbiodiversity is one of the features of eutrophica-tion and subsequent terrestrialisation, as physico-chemical conditions change (Brix and Schierup,1989; Wetzel, 1993). The highest values, observedin February during this study, were not due to ahigh number of taxa. However, as shown by thehigh value of J0, diversity can increase without anincrement in taxon number, if evenness increases(Hulbert, 1971). On the other hand, despite thehigh number of genera, diversity values registeredin April were low, due to a density peak presentedby Keratella quadrata.
The information provided by this first character-isation of environmental factors and zooplanktoncommunity structure in the Lake could be usedfor monitoring the effects of future restorationactions. Several restoration measures have beenapplied in shallow eutrophic lakes and the reportsof their results can be used as theoretical founda-tions to derive and support restoration measuresto be implemented in the Lake (Bettinetti et al.,1996; Moss et al., 1996; Søndergaard et al., 2000).
Perennial macrophytes such as Phragmites aus-tralis and Typha latifolia have morphological andphysiological characteristics giving them ecologi-cal primacy in eutrophic systems. They can actas a pump pushing nutrients from the sediments(Odum, 1971; Bjork, 1994b) which are incorporatedand preserved in their structure during the grow-ing season (Brix and Schierup, 1989; Bjork, 1994b).Moreover, they have large internal air spaces fortransporting oxygen to the roots and rhizomes, andcan stimulate the decomposition of organic matterand the growth of nitrifying bacteria, creating oxi-dised conditions in a zone which is usually anoxic(Brix and Schierup, 1989). On this basis, annualcutting and removing of emergent macrophytes atthe end of the growing season could be an importantmeasure in reducing the internal load of nutrientsin the Lake. The use of fire could be an alternativeprocedure, although the roots and rhizomes are notdamaged by fire an increase in biomass in followingyears could be expected (Bjork, 1994b). In addition,the following measures could be proposed:
(1) Cutting arboreal vegetation, mainly Eucalyp-tus globulus, near the banks, this will helpreduce the exogenous input of organic mat-ter due to falling leaves and increase wateroxygenation by wind exposure and improveconditions for waterfowl flight.
(2) Increasing water area by controlling existingsluices and constructing an additional sluice toregulate water level.
(3) Periodic cleaning of ditches to reduce the inputof organic matter due to surface runoff.
(4) Periodic clearing of undergrowth in the sur-rounding forest.
Additional measures, such as dredging of detri-tus covering the bottom of the Lake, shouldbe planned if a system with sufficient longerterm resilience capacity is to be obtained. Thewidespread anoxia in the sediment–water inter-face decreases the efficiency of phosphorus trappingin surface sediments (Quiros, 1990; Caraco, 1993);sediments could therefore be an important nutrientsource to control. The removal of the detritus layerand underlying nutrient-rich sediments is proba-bly the most effective method to improve waterquality (Hosper and Jagtman, 1990; Nielsen, 1991;Bjork, 1994c). Silva et al. (1997) described a partialdredging of lake-bottom sediments in Bracas whichimproved water trophic conditions. Moss et al.(1996) reported a reduction in total phosphorus andchlorophyll concentrations, a colonization of macro-phytes and an increase in Daphnia populations(abundance and body size) after sediment removalby suction dredging in a shallow eutrophic lake.However, the small water area of the Lake and itsshallow depth may make mechanical interventiondifficult. Phillips et al. (1994) have suggested thatthe high organic content and total phosphorus con-tent of the remaining sediment after dredging mayresult in a large release of phosphorus in subse-quent years. Other methods, such as the additionof phosphorus binding nutrients (aluminium sul-phate, iron or calcium nitrate) (Nielsen, 1991; Romoand Becares, 1994; Ripl, 1994; Welch and Schrieve,1994; Søndergaard et al., 2000), coupled with thecontrol of growing macrophytes, could be used as apossibly simple and cheaper alternative. However,though Søndergaard et al. (2000; unpublished data)have reported that the hypolimnetic addition of cal-cium nitrate was effective in the reduction of theinternal release of phosphorus, permanent effectscould not be obtained with a single dose. Romoand Becares (1994) also confirmed that despitecontrolling nutrients in Madrid’s urban lakes bythe application of aluminium sulphate, additionalmeasures were considered essential for long-termreduction in phytoplankton biomass. Additionally,aluminium sulphate and other chemicals may accu-mulate in the top sediment layer and may impairbenthic communities (Bettinetti et al., 1996; Klee-berg et al., 2000).
Finally, hypolimnetic oxidation was tried as arestoration process in some Danish lakes, with thepurpose of increasing the sediments’ phosphorusbinding capacity, thus enhancing the survival of

296 R. Pereira et al.
aquatic organisms (Søndergaard et al., 2000). Theresults obtained suggested that this was not a cost-effective process as it needs to be conducted formore than 12 years in order to produce permanenteffects (Søndergaard et al., 2000).
In conclusion, zooplankton community structureproved to be a useful tool for characterizing thetrophic state of a shallow water lake and iden-tifying factors responsible for its water qualitydegradation. Further research is required to char-acterize the phytoplankton community structureand phytoplankton–zooplankton interactions. Webelieve that Linhos lake has reached the highestaquatic trophic state, characterised by enhancedphytoplanktonic primary production, and is noweffectively a ‘dead’ ecosystem. We have proposedsome restoration measures which we believe shouldbe immediately implemented following the collec-tion of further baseline information if the disap-pearance of the Lake is to be prevented, and itsecological quality restored.
Acknowledgements
The authors are very grateful to Pedro Raposo, M. J.Ferreira, P. C. Silva and other colleagues that helpedwith the fieldwork. This study was partially supportedby Programa PRAXIS XXI in the form of a M.Sc. grantto R. Pereira.
References
Amoros, C. (1984). Introduction pratique a lasystematique des organismes des eaux continentalesfrancaises. Crustaces cladoceres. Bulletin Mensuel dela Societe Linneenne de Lyon 4, 120–145.
APHA, AWWA and WPCF (1989). Standard Methodsfor Examination of Water and Wastewater. 17th ed.Washington: American Public Health Association.
Auer, M. T., Storey, M. L., Effler, S. W., Auer, N. A. andSze, P. (1990). Zooplankton impacts on chlorophylland transparency in Onondaga Lake, New York, USA.Hydrobiologia 200/201, 603–617.
Bettinetti, A., Pypaert, P. and Sweerts, J.-P. (1996).Application of an integrated management approach tothe restoration project of the Lagoon of Venice. Journalof Environmental Management 46, 207–227.
Bjork, S. (1994a). The evolution of lakes and wetlands.In Restoration of Lake Ecosystem. A Holistic Approach(Martina Eiseltova, ed.), pp. 6–15. Slimbridge: IWRB.
Bjork, S. (1994b). Macrophyte control. In Restoration ofLake Ecosystem. A Holistic Approach (Martina Eisel-tova, ed.), pp. 89–96. Slimbridge: IWRB.
Bjork, S. (1994c). Sediment removal. In Restoration ofLake Ecosystem. A Holistic Approach (Martina Eisel-tova, ed.), pp. 82–88. Slimbridge: IWRB.
Bogdan, K. G. and Gilbert, J. J. (1982). Seasonalpatterns of feeding by natural populations of Keratella,
Polyarthra and Bosmina: clearance rates, selectivitiesand contributions to community grazing. Limnologyand Oceanography 27(5), 918–934.
Brix, H. and Schierup, H.-H. (1989). The use of aquaticmacrophytes in water pollution control. Ambio 18,100–107.
Bronmark, C. and Weisner, S. E. B. (1992). Indirecteffects of fish community structure on submergedvegetation in shallow eutrophic lakes: an alternativemechanism. Hydrobiologia 234/244, 293–301.
Caraco, N. F. (1993). Disturbance of the phosphoruscycle: a case of indirect effects of human activity.Trends in Ecological Evolution 8(2), 51–54.
Carpenter, S. R., Cole, J. J., Kitchell, J. F. and Pace, M. L.(1998). Impact of dissolved organic carbon, phosphorusand grazing on phytoplankton biomass and productionin experimental lakes. Limnology and Oceanography43(1), 73–80.
Dussart, B. H. (1969). Les copepodes des eaux conti-nentales d’Europe occidentale. Cyclopoıdes et biologie.Edition N. Boubee & Cie.
Ejsmont-Karabin, J. (1995). Rotifer occurrence in rela-tion to age, depth and trophic state of quarry lakes.Hydrobiologia 313/314, 21–28.
Flores, L. N. and Barone, R. (1994). Relationship betweentrophic state and plankton community structure in21 Sicilian dam reservoirs. Hydrobiologia 275/276,197–205.
Freedman, B. (1989). Environmental Ecology. TheImpacts of Pollution and other Stresses on Ecosys-tem Structure and Function. San Diego, CA: AcademicPress.
Galkovskaya, G. A. (1995). Oxygen consumption rate inrotifers. Hydrobiologia 313/314, 147–156.
Gliwicz, Z. M. (1990). Why do cladocerans fail to controlalgal blooms? Hydrobiologia 200/201, 83–97.
Goncalves, F., Ribeiro, R., Vasconcelos, V. and Soares,A. M. V. M. (1996). Anthropogenic influences onseasonal changes of nutrients, physical and chemicalfactors in the three coastal freshwater shallow lakes(Portugal). Limnetica 12(2), 47–52.
Habdija, I., Primc-Habdija, B., Erben, R. and Belinic, I.(1993). Trophic role of rotifers in plankton of lakeKozjak (Plitvice Lakes). Hydrobiologia 257, 101–106.
Hart, R. C. (1990). Copepod post-embryonic durations:pattern, conformity and predictability. The reali-ties of isochronal and equiproportional developmentand trends in the copepodid-naupliar duration ratio.Hydrobiologia 206, 175–206.
Hosper, S. H. and Jagtman, E. (1990). Biomanipulationadditional to nutrient control for restoration of shallowlakes in the Netherlands. Hydrobiologia 200/201,523–534.
Hulbert, S. H. (1971). The non concept of species diver-sity: a critique and alternative parameters. Ecology52, 577–586.
Jasser, I. (1995). The influence of macrophytes on aphytoplankton community in experimental conditions.Hydrobiologia 306, 21–32.
Kleeberg, A., Nixdorf, B. and Mathes, J. (2000). Lakejabel restoration project: phosphorus status and possi-bilities and limitations of diversion of its nutrient-richmain inflow. Lakes & Reservoirs: Research and Man-agement 5, 23–33.
Klinge, M., Grimm, M. P. and Hosper, S. H. (1995).Eutrophication and ecological rehabilitation of dutch

Assessing the trophic state of Linhos lake 297
lakes: conceptual framework. Water Science and Tech-nology 31(8), 207–218.
Legendre, L. and Legendre, P. (1979). EcologieNumerique. Le Traitement Multiple des DonnesEcologiques. Tome 1, 1st ed. Paris: Masson.
Maemets, A. (1983). Rotifers as indicators of lake typesin Estonia. Hydrobiologia 104, 357–361.
Magadza, C. H. D. (1994). Evaluation of eutrophicationcontrol in Lake Chivero, Zimbabwe, by multivariateanalysis of zooplankton. Hydrobiologia 272, 277–292.
Marcin, T. C. (1995). Integrating social sciences intoforest ecosystem management research. Journal ofForestry 29–33.
Matsubara, T. (1993). Rotifer community structure inthe south basin of lake Biwa. Hydrobiologia 271, 1–10.
Michaud, M. T., Atchison, G. J., McIntosh, A. W.,Mayes, R. A. and Nelson, D. W. (1979). Changesin phosphorus concentrations in a eutrophic lakeas a result of macrophyte-kill following herbicideapplication. Hydrobiologia 66(2), 105–111.
Moss, B. (1990). Engineering and biological approachesto the restoration from eutrophication of shallow lakesin which aquatic plant communities are importantcomponents. Hydrobiologia 200/201, 367–377.
Moss, B., Stansfield, J., Irvine, K., Perrows, M. andPhillips, G. (1996). Progressive restoration of a shallowlake: a 12-year experiment in isolation, sedimentremoval and biomanipulation. Journal of AppliedEcology 33, 71–86.
Nielsen, L. K. (1991). Water pollution. In Introductionto Environmental Management (P. E. Hansen andJorgensen, eds), pp. 115–175. New York: Elsevier.
Odum, E. P. (1971). Fundamentos de Ecologia. 2nd ed.Lisboa: Fundacao Calouste Gulbenkian.
Ooms-Wilms, A. L. (1997). Are bacteria an importantfood source for rotifers in eutrophic lakes? Journal ofPlankton Research 19, 1124–1141.
Pejler, B. (1983). Zooplanktonic indicators of trophy andtheir food. Hydrobiologia 101, 11–114.
Phillips, G., Jackson, R., Bennett, C. and Chilvers, A.(1994). The importance of sediment phosphorusrelease in the restoration of very shallow lakes (TheNorfolk Broads, England) and implications for bioma-nipulation. Hydrobiologia 275/276, 445–456.
Pokorny, J. (1994). Development of aquatic macrophytesin shallow lakes and ponds. In Restoration of LakeEcosystem. A Holistic Approach (Martina Eiseltova,ed.), pp. 36–43. Slimbridge: IWRB.
Power, M. E. (1992). Top-down and bottom-up forcesin food webs: do plants have primacy? Ecology 73(3),733–746.
Quiros, R. (1990). Factors related to variance of residualsin chlorophyll – total phosphorus regressions in lakesand reservoirs of Argentina. Hydrobiologia 200/201,343–355.
Ripl, W. (1994). Sediment treatment. In Restoration ofLake Ecosystem. A Holistic Approach (Martina Eisel-tova, ed.), pp. 75–81. Slimbridge: IWRB.
Rohlf, F. J. (1992). Numerical Taxonomy and Multivari-ate Analysis System. New York: Exeter Software.
Romo, S. and Becares, E. (1994). Water management oftwo shallow urban eutrophic lakes. Water Science andTechnology 30, 299–302.
Ruttner-Kolisko, A. (1974). Plankton rotifers. Biologyand taxonomy. Suppl. Die Binnengewasser, pp. 1–145.Stuttgart: Schweizerbart’sch Verlagsbuchhandlung.
Scourfield, D. J. and Harding, J. P. (1966). A key to theBritish freshwater cladocera. Freshwater BiologicalAssociation Scientific Publication 5.
Seip, K. L. (1994). Phosphorus and nitrogen limitationof algal biomass across trophic gradients. AquaticSciences 56(1), 16–27.
Silva, P. C. L. D., Goncalves, F., Ribeiro, R. andSoares, A. M. V. M. (1997). First evaluation ofthe restoration of Bracas lagoon (Figueira da Foz-Portugal). Archives Hydrobiology 141, 109–125.
Smiley, E. A. and Tessier, A. J. (1998). Environmentalgradients and the horizontal distribution of microcrus-taceans in lakes. Freshwater Biology 39, 397–409.
Søndergaard, M., Jeppesen, E., Jensen, J. P. andLauridsen, T. (2000). Lake restoration in Denmark.Lakes & Reservoirs: Research and Management 5,151–159.
Uku, J. N. and Mavuti, K. M. (1994). Comparativelimnology, species diversity and biomass relationshipof zooplankton and phytoplankton in five freshwaterlakes in Kenya. Hydrobiologia 272, 251–258.
Vasconcelos, V. (1990). Ecotoxicologia de Cianobacterias:Impacte Dos seus ‘Blooms’ nas Comunidades Animaise Sua Aplicacao na Gestao da Qualidade da agua.Provas de aptidao cientıfica e pedagogica. Porto:Faculdade de Ciencias do Porto.
Vasconcelos, V. (1994). Toxic cyanobacteria (blue-greenalgae) in Portuguese fresh waters. Archives Hydrobi-ology 130, 439–451.
Washington, H. G. (1984). Diversity, biotic and similarityindices. A review with special relevance to aquaticecosystems. Water Research 1, 164–172.
Welch, E. B. and Schrieve, G. D. (1994). Alum treatmenteffectiveness and longevity in shallow lakes. Hydrobi-ologia 275/276, 423–431.
Wetzel, R. G. (1993). Limnologia. 2nd ed. Lisboa:Fundacao Calouste Gulbenkian.
Zankai, N. P. (1989). Horizontal distribution of rotiferplankton along a trophic gradient in lake Balaton:changes of community structure and abundance dur-ing the past 20 years. Archives Hydrobiologia 115,111–123.
Zar, J. H. (1996). Biostatistical Analysis, 3rd ed. Engle-wood Cliffs, NJ: Prentice-Hall.