
Assessing the effectiveness of a hunting moratorium on target and non-target species
9
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Assessing the effectiveness of a hunting moratorium on target and non-target species

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Assessing the effectiveness of a hunting moratorium on targetand non-target species
Alejandro Martínez-Abraín a,b,⇑, Covadonga Viedma c, Juan Antonio Gómez c, Miguel Angel Bartolomé c,Juan Jiménez c, Meritxell Genovart b, Simone Tenan b,d
a Universidade da Coruña, Departamento de Bioloxía Animal, Bioloxía Vexetal e Ecoloxía, Campus da Zapateira s/n, E-15071 A Coruña, Spainb IMEDEA (CSIC-UIB), Population Ecology Group, Miquel Marquès 21, 07190 Esporles, Mallorca, Spainc Generalitat Valenciana, Servicio de Espacios Naturales y Biodiversidad, Conselleria de Infraestructuras, Territorio y Medio Ambiente, Francisco Cubells 7, 46011 Valencia, Spaind MUSE - Museo delle Scienze, Sezione Zoologia dei Vertebrati, Corso del Lavoro e della Scienza 3, IT-38123 Trento, Italy
a r t i c l e i n f o
Article history:Received 7 December 2012Received in revised form 8 May 2013Accepted 4 June 2013
Keywords:Hunting moratoriumCommon cootCrested cootSouthern EuropeManagement effectivenessWaterfowlSurvival probability
a b s t r a c t
Information on the effectiveness of wildlife management actions is scarce, despite the great relevance ofthis type of information for maximizing conservation goals while minimizing resource expenditure. Herewe assess the management effectiveness of a four-year hunting moratorium, addressed to protecting adeclining waterbird game species: common coot Fulica atra. We also studied the indirect benefits thatthis management action could have had on a non-target endangered species (crested coot Fulica cristata),currently being reintroduced in the study region (Comunidad Valenciana, eastern Spain). We found thatwintering common coots interrupted their marked negative trend coinciding with the hunting morato-rium, and Before-After-Control-Impact modelling confirmed this fact. However breeding common cootscontinued their negative trend in numbers. We also found that crested coots increased their winteringnumbers during the hunting moratorium years but not during breeding. We detected a strong and timevariant cost of release on survival probability of crested coots, but annual survival probability was foundto be constant and low for experienced birds, with no clear effects of the hunting moratorium on survivalprobability. We conclude that the moratorium had some positive effect on both species, but we suggestthat lack of enforcement during a traditional hunting practice at the end of each hunting season, mostlikely precluded the moratorium having a long-lasting effect on the breeding numbers and probablyon survival, of both species. Hence, when fully-enforced hunting moratoria are difficult to implement,we recommend the creation of hunting preserves of high habitat quality to attract coots during the win-ter, allowing its subsequent reproduction during the breeding season.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The effectiveness of management actions addressed to preserv-ing wildlife is seldom assessed, despite large money expenses maybe involved (e.g. in developing compulsory mitigation measuresassociated to infrastructure construction), and the growing needof basing decision-making on solid scientific evidence (see e.g.Stewart et al., 2005). The biological effects of some types of Euro-pean wildlife exploitation moratoria, such as those applied to traw-ler fishing in the Mediterranean, have been well explored (e.g. Oroet al., 1995; Arcos and Oro, 1996; Jiménez and Martínez-Vilalta,1998; Oro et al., 2004; Laneri et al., 2010) as well as the effective-ness of temperate marine reserves worldwide (Stewart et al.,
2009). However, the literature on the biological effects of birdhunting moratoria is scarce in Europe probably because of the dif-ficulty of having proper information (i.e. mainly counts and esti-mates of survival probability from individually-marked animals),before, during and after the hunting moratorium. Information onadaptive management of waterfowl, including effects of huntingregulations on migratory bird populations, is more common inNorth America (see e.g. Nichols et al. 2007; Williams and Johnson1995).
Here we assess the management effectiveness of a four-year(2005–2008) hunting moratorium implemented in southern Eur-ope (i.e. Comunidad Valenciana, eastern Spain), addressed to pro-tecting a declining game species, and the possible indirect effectson a non-target endangered species. This is interesting becausethe management of threatened species is typically approached bymeans of direct conservation measures aiming to improvedemographic parameters of the target species. However, speciesare integrated in assemblages, and constituent parts of animal
0006-3207/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.biocon.2013.06.009
⇑ Corresponding author at: Universidade da Coruña, Departamento de BioloxíaAnimal, Bioloxía Vexetal e Ecoloxía, Campus da Zapateira s/n, E-15071 A Coruña,Spain. Tel.: +34 652817285.
E-mail address: [email protected] (A. Martínez-Abraín).
Biological Conservation 165 (2013) 171–178
Contents lists available at SciVerse ScienceDirect
Biological Conservation
journal homepage: www.elsevier .com/ locate /biocon

Author's personal copy
assemblages interact among them in complex ways so thatconservation actions addressed to one species can have side effectson others. For example many gregarious bird species experienceboth conspecific and heterospecific attraction (Mínguez, 1994;Martínez-Abraín, 1998, 1999; Martínez-Abraín et al., 2001).However, heterospecific attraction can sometimes lead indirectlyto the formation of ecological traps (see Delibes et al., 2001;Schalaepfer et al., 2002; Battin, 2004; Robertson and Hutto,2006), when the species acting as a lure is for example a facultativepredator or a game species (Dwernychuk and Boag, 1972; Abramset al., 2012). The latter situation was identified for crested coots(Fulica cristata), a critically endangered waterbird species in Europethat gathers in mixed flocks during the winter with common coots(Fulica atra), a game species, in southern European wetlands(Martínez-Abraín et al., 2007). Owing to this, crested coots arekilled accidentally when common coots are hunted, because bothspecies are indistinguishable under hunting conditions. Hence ahunting moratorium addressed to improving the status of commoncoots can be an indirect procedure to help improving the currentlow success of the ongoing reintroduction program for crestedcoots in eastern Spain (Martínez-Abraín et al., 2011). Crested cootshave been subjected to two reintroduction programs in Spain, theabove-cited one (launched in 2000), and a second one in Andalusia,southern Spain (Varo and Amat, 2008). For the first one Tavecchiaet al. (2009) found that crested coots suffered a high cost in termsof probability of survival immediately after release. This is emerg-ing as a property of many reintroduction programs (see e.g. Oroet al., 2010; Champagnon et al., 2012). In fact immediately-released crested coots had an annual survival probability of0.14 ± 0.03 (mean ± SE), compared to 0.20 ± 0.04 for individualssurviving in the wild more than one month after release but lessthan one year, and 0.66 ± 0.20 for coots surviving at least one yearafter release (Tavecchia et al., 2009). The cause of this high local orapparent mortality during their first year of life has not beenproperly identified yet, but it could be at least partially relatedwith incidental mortality during the winter hunting season,together with low habitat quality for the species at release sites,and lack of experience in the wild (op. cit.). We explore herewhether the implementation of the four-years hunting morato-rium fulfils the expectancy of wildlife managers of improving thedemography of common wintering and breeding common coots,as well as that of the associated crested coots, or not.
2. Methods
2.1. Effects of the moratorium on common coots
Firstly, the change over time in common coot numbers duringwintering and breeding periods were modelled as a smooth non-linear function of time using generalized additive models (GAM;Hastie and Tibshirani, 1990) and the corresponding mixed mod-els (GAMM; Wood, 2006). More specifically we used NegativeBinomial GA(M)M to deal with overdispersion (Zuur et al.,2009). GAMs are useful for modelling trends because they incor-porate smoothing procedures into the model fitting process, andallow a range of curves to be considered (linear trends to annualfluctuations). GAMs also allow for a statistical test of changes indirection of the index trajectory (Fewster et al., 2000), and therelationship between response and time is assumed to besmooth but not necessarily linear (O’Brien et al., 2010). For theselection of the degree of smoothing, controlled by specifyingdegrees of freedom, we followed Fewster et al. (2000). Temporalautocorrelation was tested on the basis of AIC, and taken intoaccount with an auto-regressive model of order 1 (AR-1; Zuuret al., 2009).
We followed Fewster et al. (2000) and Buckland et al. (2005) toanalyze direction, magnitude, and timing of changes in breedingand wintering common coot abundance. The rate of change inthe number of breeding pairs or wintering birds was measuredby the slope (i.e. first derivative), of the smoothed trend. Non-lin-ear trends allow for changes in the rate of change over time. Fluc-tuations over time in the rate of change were measured bycalculating the second derivative of the trend at each time point,and the (non-parametric) bootstrapped 95% confidence intervalaround the second derivative. If the second derivative is greaterthan zero, the curve is turning upward, while if it is less than zero,the curve is turning downward. The magnitude of the secondderivative quantifies the tightness of the curvature. Values ofapproximately zero indicate that the model curve is roughly linear,and the population trajectory is changing at a steady rate. Years inwhich the second derivative is markedly different from zero arethose in which something is altering that rate of populationchange. In conjunction with other temporally-referenced data,the timing of the change points might help to suggest causes ofthe change (Fewster et al., 2000). All results in this paper were ob-tained from 3000 bootstrap replicates. Analyses were performedusing software R (R Development Core Team, 2010) with packagesmgcv (Wood, 2006), nlme (Pinheiro et al.,2013), and boot (Davisonand Hinkley, 1997). Data consisted on single counts performed inthe month of January every year.
Secondly, to analyze the effects of the hunting moratorium oncommon coots we performed a BACI analysis (BA = before/afterand CI = control/impact) (Underwood 1994). We modelled thenumber of common coots counted, on an annual basis, duringthe wintering and breeding seasons, in 10 wetlands of ComunidadValenciana (Fig. 1) during the period 1990–2012 as a function of BAand CI. We controlled for time as a continuous variable to accountfor possible variations in the number of coots due to annual varia-tions not related to the moratorium. Factors before-after and con-trol-impact were introduced as dummy variables (0–1) in eachcase. Our design included 15 pre-moratorium years, 4 years duringmoratorium, and 3 years after the moratorium for wintering cootsand 9, 4 and 3 years, respectively, for breeding coots. However, forour modelling we grouped the years during and after the huntingmoratorium to avoid power problems. The study sites included 7wetlands which were affected by the hunting moratorium (‘‘treat-ment or impact’’ hereon): Cabanes, Mijares, Almenara, Moro,Albufera, Xeresa-Xeraco and Pego-Oliva, and 3 sites which werenot (‘‘control’’ hereon): El Hondo, Santa Pola and Galvany (Fig. 1).Hence we had information on the abundance of common coots be-fore, during and after the moratorium for treatment and controlareas. For our BACI analysis we used Generalized Linear Modelswith negative binomial errors and log link to deal with the largeoverdispersion detected when modelling our coot counts withPoisson errors, after empirically discarding the quasi-Poisson fam-ily as a good alternative (see Crawley, 2007; Zuur et al., 2009). Webuilt multiple models, corresponding to biologically-meaningfulhypotheses, and compared and selected them by means of AIC-based criteria (Burnham and Anderson 2002; Anderson 2008). Ifthe moratorium had an effect on coot abundance we would expectto select as the most parsimonious model the one containing aninteraction term between the factors BA and CI, meaning that thenumber of coots had changed in a different way in control and im-pact sites in relation to the hunting moratorium. BACI analyseswere performed using R library MASS (Venables and Ripley, 2002).
2.2. Effects of the moratorium on crested coots
Since crested coots are not present in the control area (southernAlicante province wetlands, bottom part in Fig. 1), except for a fewoccasional individuals in some winters, we were not able to
172 A. Martínez-Abraín et al. / Biological Conservation 165 (2013) 171–178

Author's personal copy
analyze the management effectiveness of the moratorium bymeans of a BACI design. Hence we compared the average numberof coots wintering in the treatment areas before (2000–2004), dur-ing (2005–2008) and after (2009–2011) the hunting moratorium,distinguishing between marked and unmarked individuals. Thecomparisons were made by means of overdispersed Poisson GLM(breeding) and Negative Binomial GLM (wintering data).Overdispersed models were selected by means of quasi-AIC (QAIC;Anderson et al., 1994; Richards, 2008) with AICcmodavg R package(Mazerolle, 2013).
We also estimated the annual survival probability of n = 779captive-bred crested coots. The effects of hunting on survival ofaquatic bird species have been studied previously by means of cap-ture–recapture models (see e.g. Juillet et al., 2010; Juillet et al.,2012). From 2000 to 2012, within the framework of the reintroduc-tion program of the species in the region, juveniles of crested cootswere annually released bearing a darvic neck collar with an alpha-numeric code. Birds were resighted from a distance almost all overthe year but for estimating annual local survival (i.e. a survival thatcannot distinguish mortality from permanent emigration) we onlyincluded May resightings (i.e. the month in which a higher field ef-fort is performed to resighting birds annually). We used U-CARE(Lebreton et al., 1992; Choquet et al. 2009) to assess the fit of theCormack–Jolly–Seber model to data (Pradel et al., 1997). We testedthe effects of time and hunting moratorium (three time-periodsconsidered: before, during and after the hunting moratorium) onlocal survival probabilities of newly-released and already-released(i.e. experienced birds). We also tested a time-variant recaptureprobability. Modelling was performed by means of software M-SURGE (Choquet et al., 2004). We compared and selected models
by means of AIC-based criteria (Burnham and Anderson 2002;Anderson 2008).
3. Results
3.1. Common coots
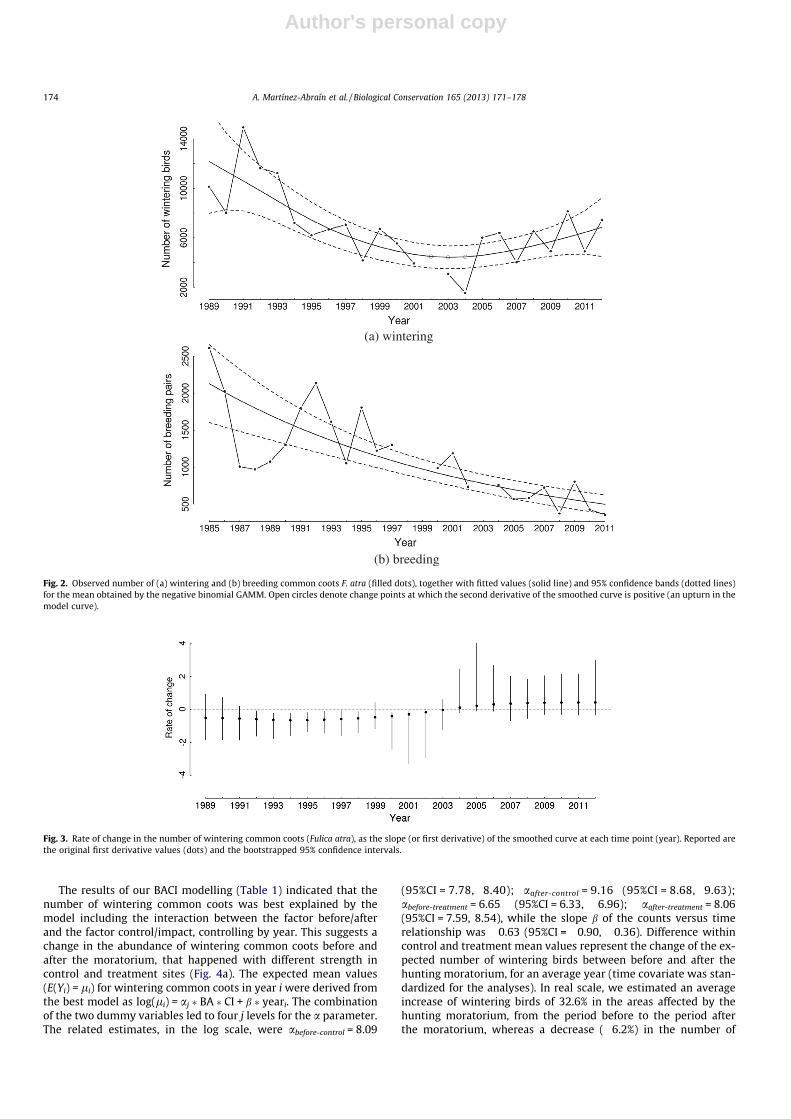
The trend in wintering common coots showed a clear non-lin-ear pattern (Fig. 2a). A decreasing trend was estimated from thebeginning of the survey up to year 2003 (Fig. 3). In this first period,the statistically significant rate of decrease in the number of win-tering birds ranged from �0.66 (1994) to �0.30 (2001). From year2004 to the end of the survey period a positive, but statisticallynon-significant, trend was estimated (Fig. 3). The statistical signif-icance of absolute changes in abundance trend was evaluated bycalculating the second derivatives for the curve at each time step(year). For years 2002, 2003, and 2004 95% bootstrap confidenceintervals for the second derivative did not include zero, suggestinga statistically significant positive change in trend of wintering birdsin these years (Fig. 2a).
A steady negative trend was estimated for the number of com-mon coot breeding pairs (Fig. 2b). The slopes, estimated throughthe first derivative of model curve, ranged from �0.30 to �0.58,although only the slope for the year 2004 was found to be differentfrom zero (�0.54; 95%CI = �0.01, �0.89). Values for the secondderivative of the trend curve were close to zero, and the related95%CIs encompassed this value, suggesting an approximately lin-ear model curve and a population trajectory that was changing atan almost steady rate.
Fig. 1. Location of the ten study sites within the western European context. Solid black dots are treatment areas (affected by the common coot hunting ban) and solid blacksquares are control areas (unaffected by the ban). CAB = Cabanes; MIJ = Mijares; ALM = Almenara; MOR = Moro; ALB = Albufera; XER = Xeresa; PEG-OLI = Pego-Oliva;GAL = Galvany; POL = Santa Pola; HON = Hondo.
A. Martínez-Abraín et al. / Biological Conservation 165 (2013) 171–178 173
Author's personal copy
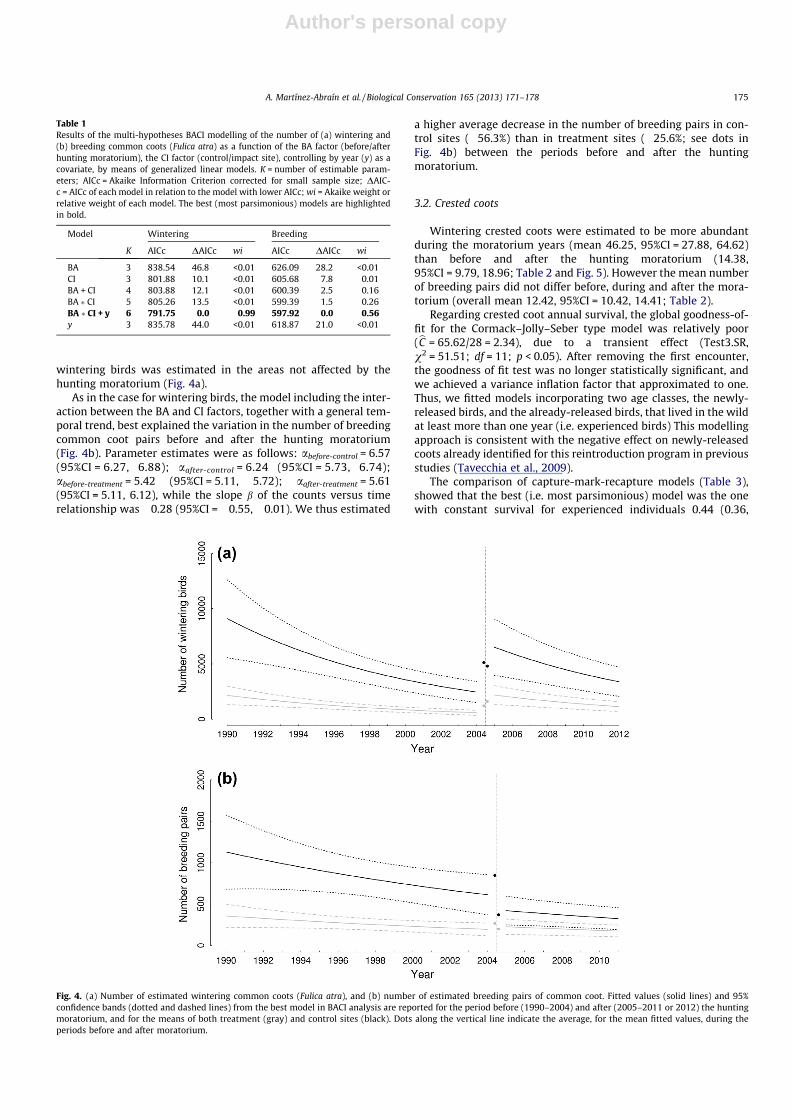
The results of our BACI modelling (Table 1) indicated that thenumber of wintering common coots was best explained by themodel including the interaction between the factor before/afterand the factor control/impact, controlling by year. This suggests achange in the abundance of wintering common coots before andafter the moratorium, that happened with different strength incontrol and treatment sites (Fig. 4a). The expected mean values(E(Yi) = li) for wintering common coots in year i were derived fromthe best model as log(li) = aj � BA � CI + b � yeari. The combinationof the two dummy variables led to four j levels for the a parameter.The related estimates, in the log scale, were abefore-control = 8.09
(95%CI = 7.78, 8.40); aafter-control = 9.16 (95%CI = 8.68, 9.63);abefore-treatment = 6.65 (95%CI = 6.33, 6.96); aafter-treatment = 8.06(95%CI = 7.59, 8.54), while the slope b of the counts versus timerelationship was �0.63 (95%CI = �0.90, �0.36). Difference withincontrol and treatment mean values represent the change of the ex-pected number of wintering birds between before and after thehunting moratorium, for an average year (time covariate was stan-dardized for the analyses). In real scale, we estimated an averageincrease of wintering birds of 32.6% in the areas affected by thehunting moratorium, from the period before to the period afterthe moratorium, whereas a decrease (�6.2%) in the number of
(a) wintering
(b) breeding
Fig. 2. Observed number of (a) wintering and (b) breeding common coots F. atra (filled dots), together with fitted values (solid line) and 95% confidence bands (dotted lines)for the mean obtained by the negative binomial GAMM. Open circles denote change points at which the second derivative of the smoothed curve is positive (an upturn in themodel curve).
Fig. 3. Rate of change in the number of wintering common coots (Fulica atra), as the slope (or first derivative) of the smoothed curve at each time point (year). Reported arethe original first derivative values (dots) and the bootstrapped 95% confidence intervals.
174 A. Martínez-Abraín et al. / Biological Conservation 165 (2013) 171–178
Author's personal copy
wintering birds was estimated in the areas not affected by thehunting moratorium (Fig. 4a).
As in the case for wintering birds, the model including the inter-action between the BA and CI factors, together with a general tem-poral trend, best explained the variation in the number of breedingcommon coot pairs before and after the hunting moratorium(Fig. 4b). Parameter estimates were as follows: abefore-control = 6.57(95%CI = 6.27, 6.88); aafter-control = 6.24 (95%CI = 5.73, 6.74);abefore-treatment = 5.42 (95%CI = 5.11, 5.72); aafter-treatment = 5.61(95%CI = 5.11, 6.12), while the slope b of the counts versus timerelationship was �0.28 (95%CI = �0.55, �0.01). We thus estimated
a higher average decrease in the number of breeding pairs in con-trol sites (�56.3%) than in treatment sites (�25.6%; see dots inFig. 4b) between the periods before and after the huntingmoratorium.
3.2. Crested coots
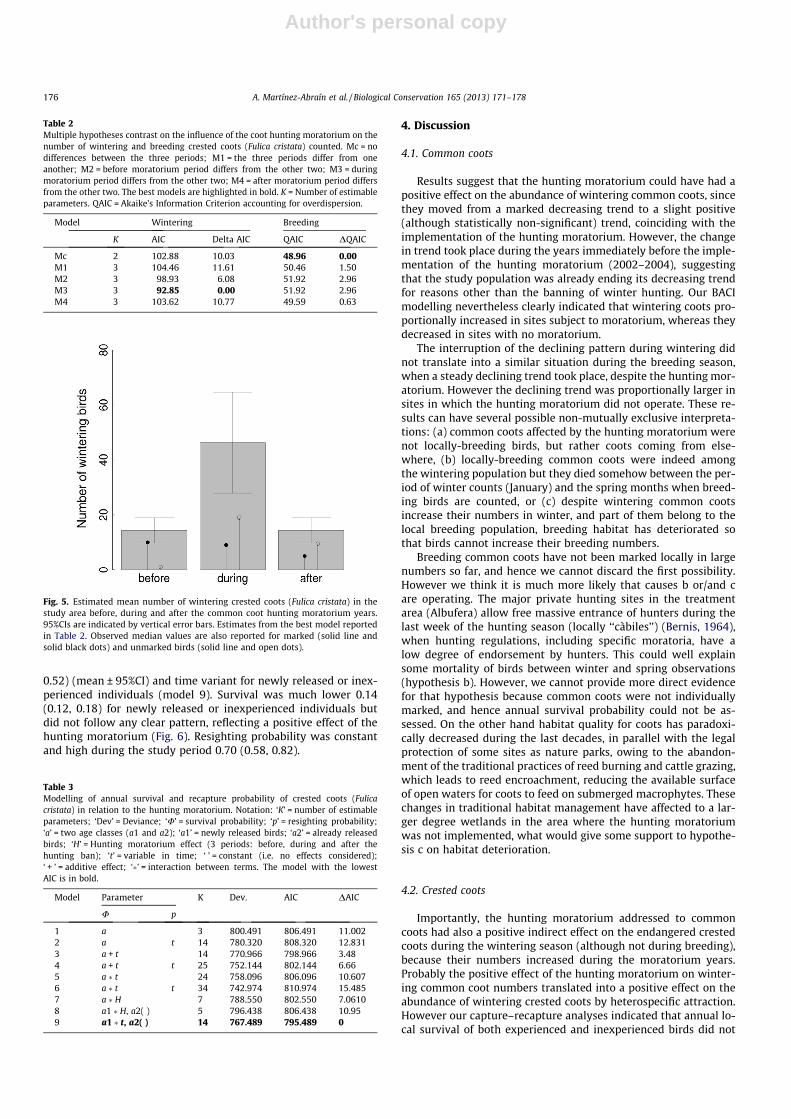
Wintering crested coots were estimated to be more abundantduring the moratorium years (mean 46.25, 95%CI = 27.88, 64.62)than before and after the hunting moratorium (14.38,95%CI = 9.79, 18.96; Table 2 and Fig. 5). However the mean numberof breeding pairs did not differ before, during and after the mora-torium (overall mean 12.42, 95%CI = 10.42, 14.41; Table 2).
Regarding crested coot annual survival, the global goodness-of-fit for the Cormack–Jolly–Seber type model was relatively poor(bC = 65.62/28 = 2.34), due to a transient effect (Test3.SR,v2 = 51.51; df = 11; p < 0.05). After removing the first encounter,the goodness of fit test was no longer statistically significant, andwe achieved a variance inflation factor that approximated to one.Thus, we fitted models incorporating two age classes, the newly-released birds, and the already-released birds, that lived in the wildat least more than one year (i.e. experienced birds) This modellingapproach is consistent with the negative effect on newly-releasedcoots already identified for this reintroduction program in previousstudies (Tavecchia et al., 2009).
The comparison of capture-mark-recapture models (Table 3),showed that the best (i.e. most parsimonious) model was the onewith constant survival for experienced individuals 0.44 (0.36,
Table 1Results of the multi-hypotheses BACI modelling of the number of (a) wintering and(b) breeding common coots (Fulica atra) as a function of the BA factor (before/afterhunting moratorium), the CI factor (control/impact site), controlling by year (y) as acovariate, by means of generalized linear models. K = number of estimable param-eters; AICc = Akaike Information Criterion corrected for small sample size; DAIC-c = AICc of each model in relation to the model with lower AICc; wi = Akaike weight orrelative weight of each model. The best (most parsimonious) models are highlightedin bold.
Model Wintering Breeding
K AICc DAICc wi AICc DAICc wi
BA 3 838.54 46.8 <0.01 626.09 28.2 <0.01CI 3 801.88 10.1 <0.01 605.68 7.8 0.01BA + CI 4 803.88 12.1 <0.01 600.39 2.5 0.16BA � CI 5 805.26 13.5 <0.01 599.39 1.5 0.26BA � CI + y 6 791.75 0.0 0.99 597.92 0.0 0.56y 3 835.78 44.0 <0.01 618.87 21.0 <0.01
Fig. 4. (a) Number of estimated wintering common coots (Fulica atra), and (b) number of estimated breeding pairs of common coot. Fitted values (solid lines) and 95%confidence bands (dotted and dashed lines) from the best model in BACI analysis are reported for the period before (1990–2004) and after (2005–2011 or 2012) the huntingmoratorium, and for the means of both treatment (gray) and control sites (black). Dots along the vertical line indicate the average, for the mean fitted values, during theperiods before and after moratorium.
A. Martínez-Abraín et al. / Biological Conservation 165 (2013) 171–178 175
Author's personal copy
0.52) (mean ± 95%CI) and time variant for newly released or inex-perienced individuals (model 9). Survival was much lower 0.14(0.12, 0.18) for newly released or inexperienced individuals butdid not follow any clear pattern, reflecting a positive effect of thehunting moratorium (Fig. 6). Resighting probability was constantand high during the study period 0.70 (0.58, 0.82).
4. Discussion
4.1. Common coots
Results suggest that the hunting moratorium could have had apositive effect on the abundance of wintering common coots, sincethey moved from a marked decreasing trend to a slight positive(although statistically non-significant) trend, coinciding with theimplementation of the hunting moratorium. However, the changein trend took place during the years immediately before the imple-mentation of the hunting moratorium (2002–2004), suggestingthat the study population was already ending its decreasing trendfor reasons other than the banning of winter hunting. Our BACImodelling nevertheless clearly indicated that wintering coots pro-portionally increased in sites subject to moratorium, whereas theydecreased in sites with no moratorium.
The interruption of the declining pattern during wintering didnot translate into a similar situation during the breeding season,when a steady declining trend took place, despite the hunting mor-atorium. However the declining trend was proportionally larger insites in which the hunting moratorium did not operate. These re-sults can have several possible non-mutually exclusive interpreta-tions: (a) common coots affected by the hunting moratorium werenot locally-breeding birds, but rather coots coming from else-where, (b) locally-breeding common coots were indeed amongthe wintering population but they died somehow between the per-iod of winter counts (January) and the spring months when breed-ing birds are counted, or (c) despite wintering common cootsincrease their numbers in winter, and part of them belong to thelocal breeding population, breeding habitat has deteriorated sothat birds cannot increase their breeding numbers.
Breeding common coots have not been marked locally in largenumbers so far, and hence we cannot discard the first possibility.However we think it is much more likely that causes b or/and care operating. The major private hunting sites in the treatmentarea (Albufera) allow free massive entrance of hunters during thelast week of the hunting season (locally ‘‘càbiles’’) (Bernis, 1964),when hunting regulations, including specific moratoria, have alow degree of endorsement by hunters. This could well explainsome mortality of birds between winter and spring observations(hypothesis b). However, we cannot provide more direct evidencefor that hypothesis because common coots were not individuallymarked, and hence annual survival probability could not be as-sessed. On the other hand habitat quality for coots has paradoxi-cally decreased during the last decades, in parallel with the legalprotection of some sites as nature parks, owing to the abandon-ment of the traditional practices of reed burning and cattle grazing,which leads to reed encroachment, reducing the available surfaceof open waters for coots to feed on submerged macrophytes. Thesechanges in traditional habitat management have affected to a lar-ger degree wetlands in the area where the hunting moratoriumwas not implemented, what would give some support to hypothe-sis c on habitat deterioration.
4.2. Crested coots
Importantly, the hunting moratorium addressed to commoncoots had also a positive indirect effect on the endangered crestedcoots during the wintering season (although not during breeding),because their numbers increased during the moratorium years.Probably the positive effect of the hunting moratorium on winter-ing common coot numbers translated into a positive effect on theabundance of wintering crested coots by heterospecific attraction.However our capture–recapture analyses indicated that annual lo-cal survival of both experienced and inexperienced birds did not
Table 2Multiple hypotheses contrast on the influence of the coot hunting moratorium on thenumber of wintering and breeding crested coots (Fulica cristata) counted. Mc = nodifferences between the three periods; M1 = the three periods differ from oneanother; M2 = before moratorium period differs from the other two; M3 = duringmoratorium period differs from the other two; M4 = after moratorium period differsfrom the other two. The best models are highlighted in bold. K = Number of estimableparameters. QAIC = Akaike’s Information Criterion accounting for overdispersion.
Model Wintering Breeding
K AIC Delta AIC QAIC DQAIC
Mc 2 102.88 10.03 48.96 0.00M1 3 104.46 11.61 50.46 1.50M2 3 98.93 6.08 51.92 2.96M3 3 92.85 0.00 51.92 2.96M4 3 103.62 10.77 49.59 0.63
Fig. 5. Estimated mean number of wintering crested coots (Fulica cristata) in thestudy area before, during and after the common coot hunting moratorium years.95%CIs are indicated by vertical error bars. Estimates from the best model reportedin Table 2. Observed median values are also reported for marked (solid line andsolid black dots) and unmarked birds (solid line and open dots).
Table 3Modelling of annual survival and recapture probability of crested coots (Fulicacristata) in relation to the hunting moratorium. Notation: ‘K’ = number of estimableparameters; ‘Dev’ = Deviance; ‘U’ = survival probability; ‘p’ = resighting probability;‘a’ = two age classes (a1 and a2); ‘a1’ = newly released birds; ‘a2’ = already releasedbirds; ‘H’ = Hunting moratorium effect (3 periods: before, during and after thehunting ban); ‘t’ = variable in time; ‘�’ = constant (i.e. no effects considered);‘ + ’ = additive effect; ‘�’ = interaction between terms. The model with the lowestAIC is in bold.
Model Parameter K Dev. AIC DAIC
U p
1 a � 3 800.491 806.491 11.0022 a t 14 780.320 808.320 12.8313 a + t � 14 770.966 798.966 3.484 a + t t 25 752.144 802.144 6.665 a � t � 24 758.096 806.096 10.6076 a � t t 34 742.974 810.974 15.4857 a � H � 7 788.550 802.550 7.06108 a1 � H, a2(�) � 5 796.438 806.438 10.959 a1 � t, a2(�) � 14 767.489 795.489 0
176 A. Martínez-Abraín et al. / Biological Conservation 165 (2013) 171–178

Author's personal copy
increase with the hunting moratorium. To the best of our knowl-edge of the study system we think that these two apparently con-tradictory results could be coupled due to the fact that somecrested-coots probably die during the week of ‘‘càbiles’’, whenmany hunting regulations are oversighted. In fact the increase inwintering crested coots during the moratorium years was mostlydue to unmarked birds which are invisible for the capture–recap-ture analysis, probably coming from outside the study system.
Experienced captive-bred crested coots from the same projectanalyzed by us were found to have a survival of 0.66 ± 0.20 afterone year in the wild, with data from 2000 to 2006 (Tavecchiaet al., 2009). Our present analyses however reflect an even lowerannual survival of experienced birds (0.44 ± 0.04) with data from2000 to 2012. This low and constant survival is probably relatedwith accidental mortality in hunting sites (despite the huntingmoratorium), where crested coots join common coot flocks, andare hence difficult to distinguish by hunters. Mortality of birdswhich have survived one year in the wild or more cannot be as-signed to food availability problems or lack of expertise to avoidpredators. Evidence of additive direct and indirect effects of hunt-ing on survival and recruitment, while controlling for relevantenvironmental factors, has been recently provided for anotherwaterfowl species (Juillet et al., 2012). An alternative explanationfor this apparent low survival would be a permanent emigrationof coots outside the study area, but a very low percentage ofcrested coots have been resighted far from the study area, despitethe large network of wetland birdwatchers in Spain. In addition,the extremely low and variable survival of inexperienced birds isprobably a consequence of their lack of habituation to the wildenvironment after captive breeding, where hunting would play alikely lower role. This incapacity of inexperienced birds to live bytheir own during the first months after release is probably a keycomponent of the failure of the reintroduction program in thestudy region, together with low habitat quality for breeding(Tavecchia et al. 2009; Martínez-Abraín et al. 2011).
5. Applied conservation implications
The hunting moratorium was effective to halt the decline in thenumbers of wintering, but not breeding, common coots, and indi-rectly had a positive effect on wintering (but not breeding) num-bers of the endangered crested coots. However, if hunting sitesfail to respect the regulations, or additionally the effectiveness ofthe ban is difficult to assess (e.g. no-individually marked birdsavailable to obtain estimates of survival probability as in the casewith common coots), hunting bans become of little conservation
use. When fully enforcing hunting moratoria is not possible, dueto logistic or socio-political reasons, alternative conservation strat-egies could be applied, such as creating hunting preserves withhigh quality habitat, run by the environmental authorities, whichcan attract coots better than hunting sites. Coots are relativelylong-lived animals (and hence quite sensitive to adult survival),but they can also produce a large number of offspring annuallywhen food is abundant. In this sense coots could qualify as bet-hedging species sensu Saether et al. (1996). Hence, both survivaland reproduction need to be improved for its conservation. Thecreation of hunting preserves, which remain flooded all year round,can have benefits both for winter survival and successful reproduc-tion during spring-summer. This would be the case if sites aremanaged to have abundant submerged macrophytes, open waterareas for large wintering flocks and helophyte rich areas (i.e. reed-beds) where coots can build their nests and protect offspring frompredation. To achieve this goal, traditional practices, such as reedburning or cattle grazing, should be restored to protected siteswhere they have been abandoned.
Acknowledgements
We are grateful to Giacomo Tavecchia and Marc Kéry for pro-viding advice in relation to the capture–recapture analysis. Thisstudy has been funded by the Spanish Ministry of Science (GrantRef. CGL2009-08298), the Regional Government of Balearic Islands(FEDER funding) and by the regional government of ComunidadValenciana (Generalitat Valenciana). AMA was funded by an IsidroParga Pondal postdoctoral contract from Xunta de Galicia. We aregrateful to Daniel Oro and Jorge Crespo for providing fruitful com-ments to drafts of the manuscript.
References
Abrams, P.A., Ruokolainen, L., Shuter, B.J., McCann, K.S., 2012. Harvesting createsecological traps: consequences of invisible mortality risks in predator–preymetacommunities. Ecology 93, 281–293.
Anderson, D.R., Burnham, K.P., White, G.C., 1994. AIC model selection inoverdispersed capture–recapture data. Ecology 75, 1780–1793.
Anderson, D.R., 2008. Model Based Inference in the Life Sciences: a Primer onEvidence. Springer, New York.
Arcos, J.M., Oro, D., 1996. Changes in foraging range of Audouin’s Gulls Larusaudouinii in relation to a trawler moratorium in the Western Mediterranean.Colon. Waterbirds 19, 128–131.
Battin, J., 2004. When good animals love bad habitats: ecological traps and theconservation of animal populations. Conserv. Biol. 18, 1482–1491.
Bernis, F., 1964. Información Española Sobre Anátidas y Fochas: Época Invernal.SEO, Madrid.
Fig. 6. Annual survival probability of inexperienced crested coots (Fulica cristata) during the study period. The hunting moratorium for common coots (F. atra) run from 2005to 2008.
A. Martínez-Abraín et al. / Biological Conservation 165 (2013) 171–178 177

Author's personal copy
Buckland, S.T., Magurran, A.E., Green, R.E., Fewster, R.M., 2005. Monitoring changein biodiversity through composite indices. Philos. Trans. Royal Soc.B 360, 243–254.
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Multimodel Inference: APractical Information-Theoretic Approach. Springer, New York.
Champagnon, J., Guillemain, M., Elmberg, J., Massez, G., Cavallo, F., Gauthier-Clerc,M., 2012. Low survival after release into the wild: assessing « the burden ofcaptivity » on Mallard physiology and behaviour. Eur. J. Wildlife Res. 58, 255–267.
Choquet, R., Reboulet, A.-M., Pradel, R., Jiménez, O., Lebreton, J.-D., 2004. M-SURGE:new software specifically designed for multistate capture–recapture models.Anim. Biodivers. Conserv. 27, 207–215.
Choquet, R., Lebreton, J.-D.-., Giménez, O., Reboulet, A.M., Pradel, R., 2009. U-CARE:utilities for performing goodness of fit tests and manipulating capture–recapture data. Ecography 32, 1071–1074.
Crawley, M.J., 2007. The R book. John Wiley and Sons, Ltd., West Sussex.Davison, A.C., Hinkley, D.V., 1997. Bootstrap Methods and Their Applications.
Cambridge University Press, Cambridge.Delibes, M., Gaona, P., Ferreras, P., 2001. Effects of an attractive sink leading into
maladaptive habitat selection. Am. Nat. 158, 277–285.Dwernychuk, L.W., Boag, D.A., 1972. Ducks nesting in association with gulls: an
ecological trap? Can. J. Zool. 50, 559–563.Fewster, R.M., Buckland, S.T., Siriwardena, G.M., Baillie, S.R., Wilson, J.D., 2000.
Analysis of population trends for farmland birds using generalized additivemodels. Ecology 81, 1970–1984.
Hastie, T.J., Tibshirani, R.J., 1990. Generalized Additive Models. CRC Press, Chapman& Hall.
Jiménez, J., Martínez-Vilalta, A., 1998. The effects of fishing moratorium on clutchsize and breeding success in two Audouin’s gull Larus audouinii colonies ineastern Spain. In: Association les amis des oiseaux and Medmaravis. (Eds.),Ecologie des Oiseaux Marins et Gestion Intégrée du Litoral en Mediterranée,Arcs ed., Tunis, pp. 234–248.
Juillet, C., Choquet, R., Gauthier, G., Pradel, R., 2010. A capture–recapture model withdouble-marking, live and dead encounters, and heterogeneity of reporting dueto auxiliary mark loss. J. Agr., Biol., Environ. Stat. 16, 88–104.
Juillet, C., Choquet, R., Gauthier, G., Lefebvre, J., Pradel, R., 2012. Carry-over effects ofspring hunt and climate on recruitment to the natal colony in a migratoryspecies. J. Appl. Ecol. 49, 1237–1246.
Laneri, K., Louzao, M., Martínez-Abraín, A., Arcos, J.M., Belda, E.J., Guallart, J.,Sánchez, A., Giménez, M., Maestre, R., Oro, D., 2010. Trawling regime influenceslongline seabird bycatch in the mediterranean: new insights from a small-scalefishery. Mar. Ecol. Prog. Ser. 420, 241–252.
Lebreton, J.D., Burnham, K.P., Clobert, J., Anderson, D.R., 1992. Modeling survival andtesting biological hypotheses using marked animals: a unified approach withcase studies. Ecol. Monogr. 62, 67–118.
Mazerolle, J.M., 2013. AICcmodavg: Model selection and multimodel inferencebased on (Q)AIC(c). R package version 1.28. http://CRAN.R-project.org/package=AICcmodavg.
Martínez-Abraín, A., 1998. Asociación trófica de ardeidas en el arrozal de la albuferade Valencia. Ardeola 45, 29–34.
Martínez-Abraín, A., 1999. Patrones de asociación de anátidas durante la invernadaen un dormidero del este de España. Ardeola 46, 163–169.
Martínez-Abraín, A., Viedma, C., Ramón, N., Oro, D., 2001. A note on the potentialrole of philopatry and conspecific attraction as conservation tools in Audouin’sGull Larus audouinii. Bird Conserv. Int. 11, 143–147.
Martínez-Abraín, A., Viedma, C., Bartolomé, M.A., Gómez, J.A., Oro, D., 2007. Huntingsites as ecological traps for coots in southern Europe: implications for theconservation of a threatened species. Endangered Sp. Res. 3, 69–76.
Martínez-Abraín, A., Regan, H., Viedma, C., Villuendas, E., Bartolomé, M.A., Gómez,J.A., Oro, D., 2011. Cost-effectiveness of translocation options for a threatenedwaterbird. Conserv. Biol. 25, 726–735.
Mínguez, E., 1994. Attraction of the European storm-petrel to heterospecific calls ofother north Atlantic Procellariiformes. Ardeola 41, 141–149.
Nichols, J.D., Runge, M.C., Johnson, F.A., Williams, B.K., 2007. Adaptive harvestmanagement of North American waterfowl populations: a brief history andfuture prospects. J. Ornithol. 148, S343–S349.
O’Brien, T.G., Baillie, J.E.M., Krueger, L., Cuke, M., 2010. The wildlife picture index:monitoring top trophic levels. Anim. Conserv. 13, 335–343.
Oro, D., Bosch, M., Ruiz, X., 1995. Effects of a trawling moratorium on the breedingsuccess of the yellow-legged gull, Larus cachinnans. Ibis 137, 547–549.
Oro, D., Cam, E., Pradel, R., Martínez-Abraín, A., 2004. Influence of food availabilityon demography and local population dynamics in a long-lived seabird. Proc.Royal Soc. London B 271, 387–396.
Oro, D., Martínez-Abraín, A., Villuendas, E., Sarzo, B., Mínguez, E., Carda, J.,Genovart, M., 2010. Lessons from a failed translocation program with aseabird species: determinants of success and conservation value. Biol.Conserv. 144, 851–858.
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., the R Development Core Team,2013. nlme: Linear and Nonlinear Mixed Effects Models. R package version3.1-108.
Pradel, R., Hines, J.E., Lebreton, J.-D., Nichols, J.D., 1997. Capture–recapture survivalmodels taking account of transients. Biometrics 53, 60–72.
R Development Core Team., 2010. R: A language and environment for statisticalcomputing, reference index version 2.11.1.R foundation for StatisticalComputing, Vienna, Austria. ISBN3-900051-07-0, URL <http://www.R-project.org>.
Richards, S.A., 2008. Dealing with overdispersed count data in applied ecology. J.Appl. Ecol. 45, 218–227.
Robertson, B.A., Hutto, R.L., 2006. A framework for understanding ecological trapsand an evaluation for existing evidence. Ecology 87, 1075–1085.
Saether, B.E., Ringsby, T.H., Roskaft, E., 1996. Life history variation, populationprocesses and priorities in species conservation: towards a reunion of researchparadigms. Oikos 77, 217–226.
Schalaepfer, M.A., Runge, M.C., Sherman, P.W., 2002. Ecological and evolutionarytraps. Trends Ecol. Evolut. 17, 474–480.
Stewart, G.B., Coles, C.F., Pullin, A.S., 2005. Applying evidence-based practice inconservation management: lessons from the first systematic review anddissemination projects. Biol. Conserv. 126, 270–278.
Stewart, G., Kaiser, M.J., Côté, I.M., Halpern, B.J., Lester, S.E., Bayliss, H.R., Pullin, A.S.,2009. Temperate marine reserves: global ecological effects and guidelines forfuture networks. Conserv. Lett. 2, 245–253.
Tavecchia, G., Viedma, C., Martínez-Abraín, A., Bartolomé, M.A., Gómez, J.A., Oro, D.,2009. Maximizing re-introduction success: assessing the immediate cost ofrelease in a threatened waterfowl. Biol. Conserv. 142, 3005–3012.
Underwood, A.J., 1994. On beyond BACI: sampling designs that might reliably detectenvironmental disturbances. Ecol. Appl. 4, 4–15.
Varo, N., Amat, J., 2008. Differences in foraging behaviour of sympatric coots withdifferent conservation status. Wildlife Res. 35, 612–616.
Venables, W.N., Ripley, B.D., 2002. Modern Applied Statistics with S. Fourth Edition.Springer, New York.
Williams, B.K., Johnson, F.A., 1995. Adaptive management and the regulation ofwaterfowl harvests. Wildlife Soc. Bull. 23, 430–436.
Wood, S., 2006. Generalized Additive Models: an Introduction with R. Chapman andHall/CRC Press.
Zuur, A., Ieno, E.N., Walker, N., Saveliev, A.A., Smith, G.M., 2009. Mixed EffectsModels and Extensions in Ecology with R. Springer.
178 A. Martínez-Abraín et al. / Biological Conservation 165 (2013) 171–178




















