
Applications of stable isotope ratio mass spectrometry in cattle dung carbon cycling studies
6
Applications of stable isotope ratio mass spectrometry in cattle dung carbon cycling studies y Jennifer A. J. Dungait 1 * , Roland Bol 1 , Elisa Lopez-Capel 2 , Ian D. Bull 3 , David Chadwick 1 , Wulf Amelung 4 , Steven J. Granger 1 , David A. C. Manning 2 and Richard P. Evershed 3 1 Soil Cross Institute Programme, North Wyke Research, Okehampton EX20 2SB, UK 2 School of Civil Engineering and Geosciences, Drummond Building, University of Newcastle, Newcastle upon Tyne NE1 7RU, UK 3 Organic Geochemistry Unit, Bristol Biogeochemistry Research Centre, School of Chemistry, University of Bristol, Cantock’s Close, Bristol BS8 1TS, UK 4 INRES-Bodenwissenschaften, University of Bonn, Nusallee-13, D-53115 Bonn, Germany Received 31 July 2009; Revised 7 October 2009; Accepted 8 October 2009 Understanding the fate of dung carbon (C) in soils is challenging due to the ubiquitous presence of the plant-derived organic matter (OM), the source material from which both dung-derived OM and soil organic matter (SOM) predominantly originate. A better understanding of the fate of specific components of this substantial source of OM, and thereby its contribution to C cycling in terrestrial ecosystems, can only be achieved through the use of labelled dung treatments. In this short review, we consider analytical approaches using bulk and compound-specific stable carbon isotope analysis that have been utilised to explore the fate of dung-derived C in soils. Bulk stable carbon isotope analyses are now used routinely to explore OM matter cycling in soils, and have shown that up to 20% of applied dung C may be incorporated into the surface soil horizons several weeks after application, with up to 8% remaining in the soil profile after one year. However, whole soil d 13 C values represent the average of a wide range of organic components with varying d 13 C values and mean residence times in soils. Several stable 13 C isotope ratio mass spectrometric methods have been developed to qualify and quantify different fractions of OM in soils and other complex matrices. In particular, thermo- gravimetry-differential scanning calorimetry-isotope ratio mass spectrometry (TG-DSC-IRMS) and gas chromatography-combustion-IRMS (GC-C-IRMS) analyses have been applied to determine the incorporation and turnover of polymeric plant cell wall materials from C 4 dung into C 3 grassland soils using natural abundance 13 C isotope labelling. Both approaches showed that fluxes of C derived from polysaccharides, i.e. as cellulose or monosaccharide components, were more similar to the behaviour of bulk dung C in soil than lignin. However, lignin and its 4-hydroxypropanoid monomers were unexpectedly dynamic in soil. These findings provide further evidence for emerging themes in biogeochemical investigations of soil OM dynamics that challenge perceived concepts of recalcitrance of C pools in soils, which may have profound implications for the assessment of the potential of agricultural soils to influence terrestrial C sinks. Copyright # 2010 John Wiley & Sons, Ltd. Natural abundance stable 13 C isotope labelling is one of the few proven techniques available for the examination of soil C dynamics in naturally functioning ecosystems. Compared with 14 C radioisotope labelling, it is a safe and cost effective method that gives an accurate interpretation of processes in complex systems, such as soils. Isotope ratio mass spec- trometry (IRMS) is widely used to determine the difference in natural abundance of 13 C between C 3 (d 13 C ¼32 to 20%) and C 4 (d 13 C ¼9 to 17%) vegetation which provides the basis for estimating the contribution of 13 C- enriched C 4 sources to soil organic matter (SOM) in ecosystems otherwise dominated by C 3 vegetation. 1 Follow- ing changes in land use from a C 3 to C 4 vegetation, bulk SOM d 13 C values have been used to investigate organic matter (OM) turnover in soils, 2 particle size fractions 3,4 and soil aggregates. 5 Naturally, the d 13 C values of cattle products reflect the stable isotope values of their feed; 6 therefore, cattle that are fed naturally 13 C-enriched C 4 species forage, i.e. Zea mays, produce a convenient source of natural abundance 13 C- labelled dung that can be applied as a treatment to soils in C 3 ecosystems to explore the cycling of dung C. Thus, by using bulk and compound-specific IRMS, the spatiotemporal dynamics of whole dung C cycling, and that of specific biochemical components of dung in the soil, can be determined. The mean residence times of C pools have been shown to increase with the application of manure. 7 In the Hoosfield Classical Experiment at Rothamsted, annual applications of manure at a rate of 35 t ha 1 over 140 years resulted in a three- RAPID COMMUNICATIONS IN MASS SPECTROMETRY Rapid Commun. Mass Spectrom. 2010; 24: 495–500 Published online in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/rcm.4332 *Correspondence to: J. A. J. Dungait, Soil Cross Institute Pro- gramme, North Wyke Research, Okehampton EX20 2SB, UK. E-mail: [email protected] y Presented at SIMSUG 2009, held 14–15 January 2009 at the University of Glasgow. Copyright # 2010 John Wiley & Sons, Ltd.
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of Applications of stable isotope ratio mass spectrometry in cattle dung carbon cycling studies

RAPID COMMUNICATIONS IN MASS SPECTROMETRY
Rapid Commun. Mass Spectrom. 2010; 24: 495–500
) DOI: 10.1002/rcm.4332
Published online in Wiley InterScience (www.interscience.wiley.comApplications of stable isotope ratio mass spectrometry in
cattle dung carbon cycling studiesy
Jennifer A. J. Dungait1*, Roland Bol1, Elisa Lopez-Capel2, Ian D. Bull3, David Chadwick1,
Wulf Amelung4, Steven J. Granger1, David A. C. Manning2 and Richard P. Evershed3
1Soil Cross Institute Programme, North Wyke Research, Okehampton EX20 2SB, UK2School of Civil Engineering and Geosciences, Drummond Building, University of Newcastle, Newcastle upon Tyne NE1 7RU, UK3Organic Geochemistry Unit, Bristol Biogeochemistry Research Centre, School of Chemistry, University of Bristol, Cantock’s Close, Bristol BS8
1TS, UK4INRES-Bodenwissenschaften, University of Bonn, Nusallee-13, D-53115 Bonn, Germany
Received 31 July 2009; Revised 7 October 2009; Accepted 8 October 2009
*Correspogramme,E-mail: jeyPresenteUniversit
Understanding the fate of dung carbon (C) in soils is challenging due to the ubiquitous presence of the
plant-derived organic matter (OM), the source material from which both dung-derived OM and soil
organic matter (SOM) predominantly originate. A better understanding of the fate of specific
components of this substantial source of OM, and thereby its contribution to C cycling in terrestrial
ecosystems, can only be achieved through the use of labelled dung treatments. In this short review, we
consider analytical approaches using bulk and compound-specific stable carbon isotope analysis that
have been utilised to explore the fate of dung-derived C in soils. Bulk stable carbon isotope analyses
are now used routinely to explore OM matter cycling in soils, and have shown that up to 20% of
applied dung C may be incorporated into the surface soil horizons several weeks after application,
with up to 8% remaining in the soil profile after one year. However, whole soil d13C values represent
the average of a wide range of organic components with varying d13C values andmean residence times
in soils. Several stable 13C isotope ratio mass spectrometric methods have been developed to qualify
and quantify different fractions of OM in soils and other complex matrices. In particular, thermo-
gravimetry-differential scanning calorimetry-isotope ratio mass spectrometry (TG-DSC-IRMS) and
gas chromatography-combustion-IRMS (GC-C-IRMS) analyses have been applied to determine the
incorporation and turnover of polymeric plant cell wall materials from C4 dung into C3 grassland soils
using natural abundance 13C isotope labelling. Both approaches showed that fluxes of C derived from
polysaccharides, i.e. as cellulose or monosaccharide components, were more similar to the behaviour
of bulk dung C in soil than lignin. However, lignin and its 4-hydroxypropanoid monomers were
unexpectedly dynamic in soil. These findings provide further evidence for emerging themes in
biogeochemical investigations of soil OMdynamics that challenge perceived concepts of recalcitrance
of C pools in soils, which may have profound implications for the assessment of the potential of
agricultural soils to influence terrestrial C sinks. Copyright # 2010 John Wiley & Sons, Ltd.
Natural abundance stable 13C isotope labelling is one of the
few proven techniques available for the examination of soil C
dynamics in naturally functioning ecosystems. Compared
with 14C radioisotope labelling, it is a safe and cost effective
method that gives an accurate interpretation of processes in
complex systems, such as soils. Isotope ratio mass spec-
trometry (IRMS) is widely used to determine the difference
in natural abundance of 13C between C3 (d13C¼�32 to
�20%) and C4 (d13C¼�9 to �17%) vegetation which
provides the basis for estimating the contribution of 13C-
enriched C4 sources to soil organic matter (SOM) in
ecosystems otherwise dominated by C3 vegetation.1 Follow-
ndence to: J. A. J. Dungait, Soil Cross Institute Pro-North Wyke Research, Okehampton EX20 2SB, [email protected] at SIMSUG 2009, held 14–15 January 2009 at they of Glasgow.
ing changes in land use from a C3 to C4 vegetation, bulk SOM
d13C values have been used to investigate organic matter
(OM) turnover in soils,2 particle size fractions3,4 and soil
aggregates.5 Naturally, the d13C values of cattle products
reflect the stable isotope values of their feed;6 therefore, cattle
that are fed naturally 13C-enriched C4 species forage, i.e. Zea
mays, produce a convenient source of natural abundance 13C-
labelled dung that can be applied as a treatment to soils in C3
ecosystems to explore the cycling of dung C. Thus, by using
bulk and compound-specific IRMS, the spatiotemporal
dynamics of whole dung C cycling, and that of specific
biochemical components of dung in the soil, can be
determined.
The mean residence times of C pools have been shown to
increase with the application of manure.7 In the Hoosfield
Classical Experiment at Rothamsted, annual applications of
manure at a rate of 35 t ha�1 over 140 years resulted in a three-
Copyright # 2010 John Wiley & Sons, Ltd.

496 J. A. J. Dungait et al.
fold increase in soil organic C levels over that in unfertilised
plots, and about 50% higher than in unfertilised plots 104
years after manure addition had been discontinued.8 This
and other long-term studies (20–120 years) in the UK,
Denmark, USA and Canada have indicated that manure can
play a positive role in increasing soil C stocks in soils.9
However, other studies have concluded that dung appli-
cation may have a limited effect10,11 or indeed a negative
effect on soil C sequestration.12 Clearly, the dynamics of
incorporation of dung C into soil are not well understood,
but they are known to be highly variable in both space and
time.13,14 The time taken for 75% of cow dung to disappear is
reported to range from 32 to 450 days;15 the initial quality of
dung affects rates of decay,16 and environmental variables,
such as temperature and rainfall, strongly influence
decomposition.13
Although many studies have determined the concen-
trations of inorganic components in dung and dung-treated
soils that directly affect plant growth, e.g. N, P, K, Mg and
Ca,17–20 or act as pollutants,21–23 e.g. heavy metals,24,25
relatively little work has been carried out to identify and
quantify the biochemical components that contribute to
SOM. Cow dung is a complex mixture of biochemical
components that are likely to decompose at different rates
(Fig. 1). Gravimetric procedures are the most widely applied
to estimate the contribution of different biochemical fractions
to dung and studies have shown that the major component is
undigested lignocellulosic plant cell wall material.26 Haynes
and Williams27 reported that cattle dung comprised 47–68%
‘fibre’ (the sum of hemicellulose, cellulose and lignin), and
Dungait et al.28,29 estimated that cow dung derived from
Lolium perenne and Z. mays forages was 20–30% hemicellu-
lose, 20–30% cellulose, 7% lignin, 12% crude protein and
3–5% fats using the ‘Forage Fibre Analysis’ procedure
described by Van Soest.26 Currently, no reliable estimates
exist for the concentration of dung lignin due to the
challenges presented by extraction of monomers with
subsequent quantification against an internal standard.
Analyses of dung carbohydrates analysed as alditol acetates
using gas chromatography (GC) determined concentrations
of up to 80% dry weight as sugars in dung due to the
inclusion of the soluble components, which may also derive
from microbial debris, lost during Forage Fibre Analysis.30
Figure 1. Ruminant digestion and the origin of faecal matter,
showing major components of dung (adapted from Van
Soest26).
Copyright # 2010 John Wiley & Sons, Ltd.
Bull et al.31 investigated the lipid components of farmyard
manure (FYM) in order to assess the effect that variable
manure application has on the free lipid content of the soil.
The lipids in dung were dominated by 5b-stanols and C26 n-
alkanol, with relatively minor contributions from carboxylic
acids, wax esters and n-alkanes.
Different components of plant-derivedOM are also known
to have a range of d13C values due to fractionations against
the heavier isotope during biosynthesis. Lignin and lipids are
depleted compared with bulk plant tissue, while cellulose
and hemicellulose are 1–2% more 13C-enriched.1,32 Dungait
et al.33 used off-line pyrolysis to extract lignin monomers
from dung and dung-treated soils for stable 13C-isotope
analysis using GC-combustion-IRMS (GC-C-IRMS). The m/z
44 ion current and the instantaneous ratio of m/z 45/44 ions
recorded for the off-line pyrolysate of lipid-extracted C4
dung, displaying base-line resolved pyrolysis products for
which compound-specific d13C values could be obtained, are
shown in Fig. 2. Dung lignin-derived moieties in the
pyrolysate were up to 7% 13C-depleted relative to bulk
dung, although syringol and 4-vinylguaiacol were 13C-
enriched by up to 4% and 2%, respectively. The 13C-depleted
values are in agreement with earlier studies, which indicated
that plant tissues yield lignin products 2–7% depleted in 13C
comparedwith whole tissue, although Kracht and Gleixner34
determined up to 5% 13C-enrichment of lignin moieties
compared with bulk values in lignin pyrolysates of peat. The
d13C values determined for lipids as n-carboxylic acids
(iC14:0–C30:0) extracted from C3 and C4 dung (Fig. 3) also
showed depletion of up to ca. 6% compared with bulk dung,
with increasing depletion with hydrocarbon chain length in
very long chain fatty acids (VLCFA) >C20.28 Carbohydrate
d13C values determined for arabinose, xylose, galactose and
glucose extracted from C4 dung and analysed as their alditol
acetates were �10.4� 0.5%, �10.4� 0.4%, �8.3� 1.6% and
�11.5� 0.6%, respectively (bulk d13C value �12.6� 0.3%),
which are similar to values reported for Z. mays leaf by
Derrien et al.,35 and values for C3 dung �26.8� 1.1%,
�28.7� 0.9, �27.4� 2.6% and �29.9� 0.7% (bulk d13C value
�31.3� 0.1%) were similar to those derived from ryegrass
(L. perenne28). The variability in concentration and d13C
values between individual components of dung-derived OM
drives the need to develop and apply sensitive tools to
determine their contribution to bulk values in order to
understand C cycling in soils treated with manures as soil
improvers or under livestock management.
Bulk stable 13C isotope IRMSNatural abundance 13C-labelled C4 plant matter can be
applied to restricted areas of experimental plots, and is
especially useful for short-term bulk decomposition studies.
Amelung et al.36 and Bol et al.37 developed a method using the
difference in natural abundance 13C values between C4 dung
(d13C¼�15.4%) andC3 dung (d13C¼�25.6%) applied to a C3
soil (d13C¼�27.9%) in trace bulk dung C incorporation into
soils using continuous flow (CF)-IRMS. Bulk d13C values of C3
and C4 dung-treated soil were used in a simple mixing model
to estimate dung C incorporation after dung deposition. This
approachwas also used to trace dung-derived C into soils,29,37
soil particulate size fractions,38,39 and leachates.37,40 The
Rapid Commun. Mass Spectrom. 2010; 24: 495–500
DOI: 10.1002/rcm
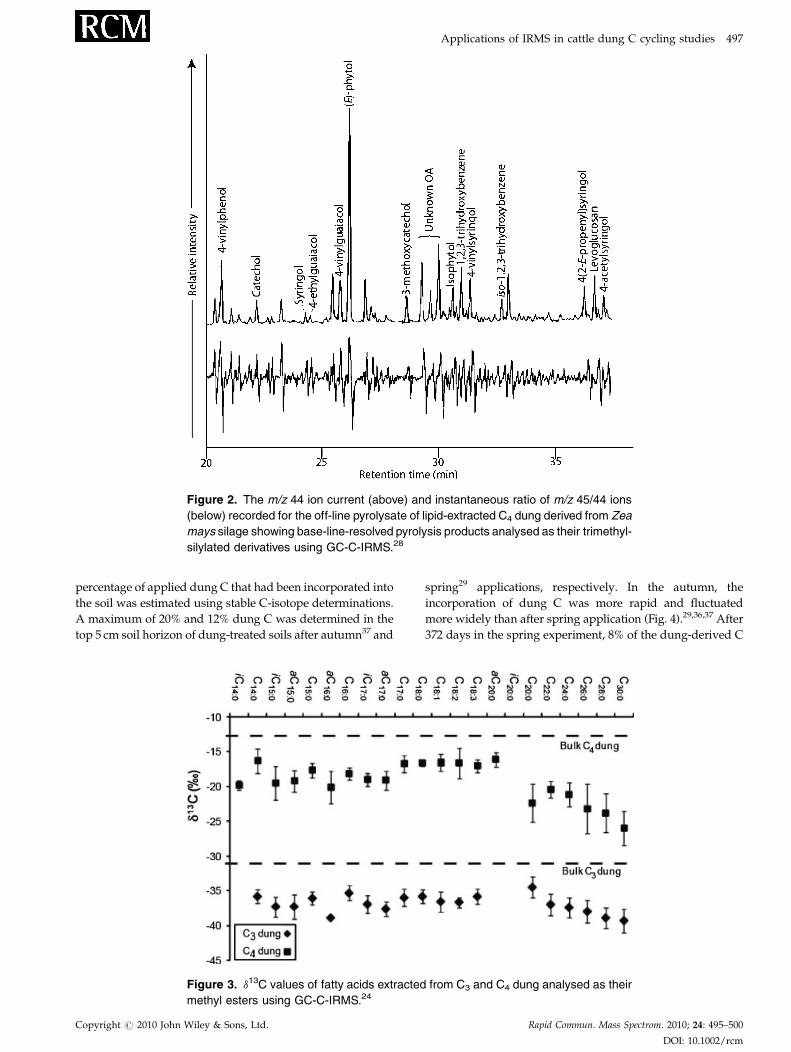
Figure 2. The m/z 44 ion current (above) and instantaneous ratio of m/z 45/44 ions
(below) recorded for the off-line pyrolysate of lipid-extracted C4 dung derived from Zea
mays silage showing base-line-resolved pyrolysis products analysed as their trimethyl-
silylated derivatives using GC-C-IRMS.28
Applications of IRMS in cattle dung C cycling studies 497
percentage of applied dung C that had been incorporated into
the soil was estimated using stable C-isotope determinations.
A maximum of 20% and 12% dung C was determined in the
top 5 cm soil horizon of dung-treated soils after autumn37 and
Figure 3. d13C values of fatty acids extracted
methyl esters using GC-C-IRMS.24
Copyright # 2010 John Wiley & Sons, Ltd.
spring29 applications, respectively. In the autumn, the
incorporation of dung C was more rapid and fluctuated
more widely than after spring application (Fig. 4).29,36,37 After
372 days in the spring experiment, 8% of the dung-derived C
from C3 and C4 dung analysed as their
Rapid Commun. Mass Spectrom. 2010; 24: 495–500
DOI: 10.1002/rcm

Figure 4. Comparison of data from dung application exper-
iments after autumn and spring deposition showing differential
incorporation of percentage applied dung C between seasons
calculated using bulk d13C stable isotope determinations ana-
lysed using CF-IRMS (adapted from Dungait et al.33).
Figure 5. Proportion of dung-derived C incorporation in C3
and C4 dung-treated soils at various temperature intervals
during thermal decomposition estimated using TG-DSC-
IRMS.49
498 J. A. J. Dungait et al.
remained in the soil, providing direct evidence for the
mechanism for increasing C stocks in soils treated annually
with manures. Amelung et al.36 traced dung-derived C into
particle-size fractions in the top 1–5 cm of soil. As the particle
size diameter increased, the d13C values increased. The
proportion of dung C in fine clay (<0.2mm) maximised after
42 days, coinciding with a maximum in the bulk soil d13C
values and leaching rate. Thepercentage of particulateC in the
coarse sand fraction (250–2000mm) increased until the end of
the experiment, and it was suggested that undigested
polyphenolic substances, such as lignin, contributed an
increasing proportion to dung-derived C.
Due to the diversity in the decomposition dynamics and
the d13C values of individual biochemical components of
dung, fluxes in bulk d13C values in C4 dung-treated soil may
not imply a total loss or gain of dung C. For instance,
depletion in bulk d13C values may simply provide evidence
for the decline of labile, relatively 13C-enriched dung-derived
components, such as carbohydrates, from the soils. Indeed,
initial characterisation of components (NDF, ADF, ADF-
lignin, Kjedahl N and fats) of C4 and C3 dung using forage
fibre analysis29 and GC analysis of polysaccharides,30
lignin33 and lipid28 fractions of C3 and C4 dung revealed
differences between the composition of the two dung types
which may have affected the parity of their incorporation
into soil. Therefore, the later study of dung incorporation
after spring application29 used the d13C values of bulk soil
and individual compounds extracted from C4 dung-treated
and untreated (control) soils as end members in the mixing
model to estimate dung C incorporation and turnover. Thus,
it is essential that the contributions of individual biochemical
components to bulk dung C, and the residence times of those
individual components in the soil, are investigated in order
to adequately understand spatiotemporal fluxes of C in
dung-treated soils.
Compound-specific stable 13C-isotope IRMSThe most abundant constituents of soil C are carbohydrates
derived from the endogenous plant litter that comprises up
to 50% of SOM. Recent studies have questioned both the
Copyright # 2010 John Wiley & Sons, Ltd.
perceived lability of carbohydrates in soils35,41,42 and the
recalcitrance of lignin,33,43–46 suggesting that carbohydrates
have a considerable role to play in C sequestration in soils.
Plant cell walls are the major components of dung and SOM
in grassland systems, but their identification and quantifi-
cation pose analytical challenges due to the chemically
intractable nature of lignin and polysaccharide polymers,
and their ubiquitous nature in edaphic systems. Here, the
application of two complementary compound-specific stable
isotope approaches: (1) thermogravimetry-differential scan-
ning calorimetry-IRMS (TG-DSC-IRMS) and (2) GC-C-IRMS
to the investigation of dung C dynamics in grassland soils,
are considered.
TG-DSC-IRMSThermogravimetry (TG) combined with differential scan-
ning calorimetry (DSC) has been applied to explore the
chemical changes in OM during decomposition, especially
with regard to the composition of the bulk constituents of
degrading plant material.47–49 TG-DSC analysis involves
continuous and simultaneous measurement of weight-loss
(TG) and energy change (DSC) during heating. Exothermic
decomposition of aliphatic and carboxyl groups predomi-
nates at temperatures of 300–3508C, with more refractory
aromatic C being lost at higher temperatures of around 400–
4508C. Coupled to IRMS this analytical approach allows the
d13C values of volatile organic components evolved during
heating to be determined during a single heating experiment,
providing valuable insights into the dynamics of labile and
recalcitrant components of SOM.
Lopez-Capel et al.50 applied TG-DSC-IRMS to describe and
characterise the changes in C4 dung-derived C over 70 days
after autumn application of dung in the same grassland soil
previously analysed using CF-IRMS (see above). d13C values
were more depleted with increasing combustion tempera-
tures, with cellulosic components being around 4% more13C-enriched than more refractory components, i.e. lignin.
Using the dung C incorporation equation,29 the percentage of
dung C incorporated into soil using the exothermic peak
between 400–4508C suggestedmajor incorporation of dung C
after 40 days in line with the bulk isotope study described
(Fig. 4). However, Fig. 5 also illustrates that aromatic dung C
Rapid Commun. Mass Spectrom. 2010; 24: 495–500
DOI: 10.1002/rcm
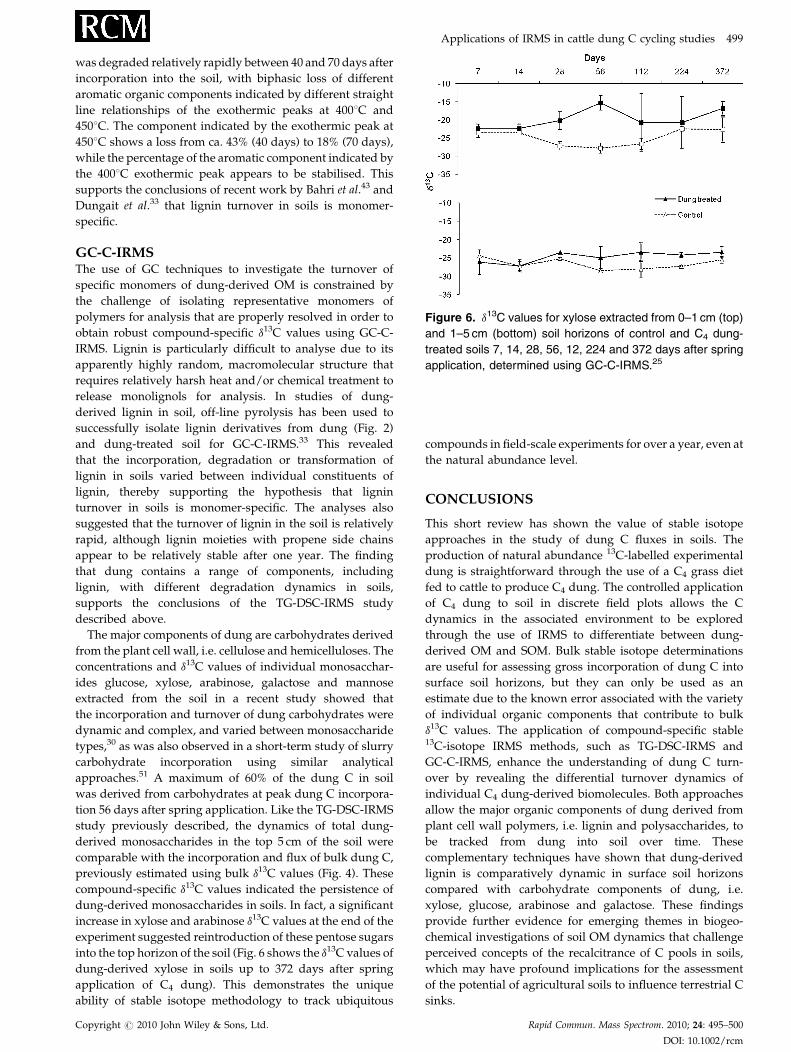
Figure 6. d13C values for xylose extracted from 0–1 cm (top)
and 1–5 cm (bottom) soil horizons of control and C4 dung-
treated soils 7, 14, 28, 56, 12, 224 and 372 days after spring
application, determined using GC-C-IRMS.25
Applications of IRMS in cattle dung C cycling studies 499
was degraded relatively rapidly between 40 and 70 days after
incorporation into the soil, with biphasic loss of different
aromatic organic components indicated by different straight
line relationships of the exothermic peaks at 4008C and
4508C. The component indicated by the exothermic peak at
4508C shows a loss from ca. 43% (40 days) to 18% (70 days),
while the percentage of the aromatic component indicated by
the 4008C exothermic peak appears to be stabilised. This
supports the conclusions of recent work by Bahri et al.43 and
Dungait et al.33 that lignin turnover in soils is monomer-
specific.
GC-C-IRMSThe use of GC techniques to investigate the turnover of
specific monomers of dung-derived OM is constrained by
the challenge of isolating representative monomers of
polymers for analysis that are properly resolved in order to
obtain robust compound-specific d13C values using GC-C-
IRMS. Lignin is particularly difficult to analyse due to its
apparently highly random, macromolecular structure that
requires relatively harsh heat and/or chemical treatment to
release monolignols for analysis. In studies of dung-
derived lignin in soil, off-line pyrolysis has been used to
successfully isolate lignin derivatives from dung (Fig. 2)
and dung-treated soil for GC-C-IRMS.33 This revealed
that the incorporation, degradation or transformation of
lignin in soils varied between individual constituents of
lignin, thereby supporting the hypothesis that lignin
turnover in soils is monomer-specific. The analyses also
suggested that the turnover of lignin in the soil is relatively
rapid, although lignin moieties with propene side chains
appear to be relatively stable after one year. The finding
that dung contains a range of components, including
lignin, with different degradation dynamics in soils,
supports the conclusions of the TG-DSC-IRMS study
described above.
The major components of dung are carbohydrates derived
from the plant cell wall, i.e. cellulose and hemicelluloses. The
concentrations and d13C values of individual monosacchar-
ides glucose, xylose, arabinose, galactose and mannose
extracted from the soil in a recent study showed that
the incorporation and turnover of dung carbohydrates were
dynamic and complex, and varied between monosaccharide
types,30 as was also observed in a short-term study of slurry
carbohydrate incorporation using similar analytical
approaches.51 A maximum of 60% of the dung C in soil
was derived from carbohydrates at peak dung C incorpora-
tion 56 days after spring application. Like the TG-DSC-IRMS
study previously described, the dynamics of total dung-
derived monosaccharides in the top 5 cm of the soil were
comparable with the incorporation and flux of bulk dung C,
previously estimated using bulk d13C values (Fig. 4). These
compound-specific d13C values indicated the persistence of
dung-derived monosaccharides in soils. In fact, a significant
increase in xylose and arabinose d13C values at the end of the
experiment suggested reintroduction of these pentose sugars
into the top horizon of the soil (Fig. 6 shows the d13C values of
dung-derived xylose in soils up to 372 days after spring
application of C4 dung). This demonstrates the unique
ability of stable isotope methodology to track ubiquitous
Copyright # 2010 John Wiley & Sons, Ltd.
compounds in field-scale experiments for over a year, even at
the natural abundance level.
CONCLUSIONS
This short review has shown the value of stable isotope
approaches in the study of dung C fluxes in soils. The
production of natural abundance 13C-labelled experimental
dung is straightforward through the use of a C4 grass diet
fed to cattle to produce C4 dung. The controlled application
of C4 dung to soil in discrete field plots allows the C
dynamics in the associated environment to be explored
through the use of IRMS to differentiate between dung-
derived OM and SOM. Bulk stable isotope determinations
are useful for assessing gross incorporation of dung C into
surface soil horizons, but they can only be used as an
estimate due to the known error associated with the variety
of individual organic components that contribute to bulk
d13C values. The application of compound-specific stable13C-isotope IRMS methods, such as TG-DSC-IRMS and
GC-C-IRMS, enhance the understanding of dung C turn-
over by revealing the differential turnover dynamics of
individual C4 dung-derived biomolecules. Both approaches
allow the major organic components of dung derived from
plant cell wall polymers, i.e. lignin and polysaccharides, to
be tracked from dung into soil over time. These
complementary techniques have shown that dung-derived
lignin is comparatively dynamic in surface soil horizons
compared with carbohydrate components of dung, i.e.
xylose, glucose, arabinose and galactose. These findings
provide further evidence for emerging themes in biogeo-
chemical investigations of soil OM dynamics that challenge
perceived concepts of the recalcitrance of C pools in soils,
which may have profound implications for the assessment
of the potential of agricultural soils to influence terrestrial C
sinks.
Rapid Commun. Mass Spectrom. 2010; 24: 495–500
DOI: 10.1002/rcm

500 J. A. J. Dungait et al.
REFERENCES
1. Boutton TW. Stable isotope ratios of natural materials:I. atmospheric, terrestrial, marine and freshwater environ-ments. InCarbon Isotope Techniques, Coleman DC, Fry B (eds).Academic Press: San Diego, 1991; 219–242.
2. Boutton TW, Archer SR, Midwood AJ, Zitzer SF, Bol R.Geoderma 1998; 82: 5.
3. Balesdent J, Mariotti A, Guillet B. Soil Biol. Biochem. 1987; 19:25.
4. Poirier N, Derenne S, Balesdent J, Chenu C, Bardoux G,Mariotti A, Largeau C. Org. Geochem. 2000; 31: 813.
5. Puget P, Angers DA, Chenu C. Soil Biol. Biochem. 1999; 31: 55.6. Deniro MJ, Epstein S.Geochim. Cosmochim. Acta 1978; 42: 495.7. Fortuna A, Harwood R, Kizilkaya K, Paul EA. Soil Biol.
Biochem. 2003; 35: 1005.8. Haynes RJ, Naidu R. Nutr. Cyc. Agroecosys. 1998; 51: 123.9. Edmeades DC. Nutr. Cyc. Agroecosys. 2003; 66: 165.
10. Bol R, Eriksen J, Smith P, Garnett MH, Coleman K, Chris-tensen BT. Rapid Commun. Mass Spectrom. 2005; 19: 3216.
11. Gulde S, Chung H, Amelung W, Chang C, Six J. Soil Sci. Soc.Am. J. 2008; 72: 605.
12. Gerzabek MH, Pichlmayer F, Kirchmann H, Haberhauer G.Eur. J Soil Sci. 1997; 48: 273.
13. Dickinson CH, Craig G. New Phytol. 1990; 115: 139.14. MacDiarmid BN, Watkin BR. J. Br. Grass. Soc. 1972; 27: 48.15. Dickinson CH, Underhay VXH, Ross V.New Phytol. 1981; 88:
129.16. Weeda WC. NZ J. Agric. Res. 1967; 10: 150.17. Aarons SR, Hosseini HM, Dorling L, Gourley CJP.Aus. J. Soil
Res. 2004; 42: 115.18. Aarons SR, O’Connor CR, Gourley CJP. Aus. J. Soil Res. 2004;
42: 107.19. Chadwick DR, John F, Pain BF, Chambers BJ, Williams J.
J. Agric. Sci. 2000; 134: 159.20. Sheldrick W, Syers JK, Lingard J.Nutr. Cyc. Agroecosys. 2003;
66: 119.21. Brookman SKE, Chadwick DR, Retter AR. Environ. Technol.
2005; 26: 1189.22. Chadwick D, Fish R, Oliver DM, Heathwaite L, Hodgson C,
Winter M. Trends Food Sci. Technol. 2008; 19: 240.23. Chadwick DR, Pain BF, Brookman SKE. J. Env. Qual. 2000;
29: 277.24. Hoekstra NJ, Bosker T, Lantinga EA. Agric. Ecosys. Environ.
2002; 93: 189.25. Wilcke W, Bol R, Amelung W. Geoderma 2002; 106: 273.26. Van Soest PJ. Nutritional Ecology of the Ruminant. Cornell
University: Ithaca, NY, 1994.27. Haynes RJ, Williams PH. Adv. Agron. 1993; 49: 119.
Copyright # 2010 John Wiley & Sons, Ltd.
28. Dungait JAJ. Molecular and compound-specific stable car-bon isotope investigation of the fate of dung carbon in atemperate grassland soil. PhD thesis, University of Bristol,Bristol, UK. 2005.
29. Dungait JAJ, Bol R, Evershed RP. Isot. Environ. Health Stud.2005; 41: 3.
30. Dungait JAJ, Bol R, Bull ID, Evershed RP.Org. Geochem. 2009;DOI 10.1016/j.orggeochem.08.001.
31. Bull ID, Van Bergen PF, Poulton PR, Evershed RP. Org.Geochem. 1998; 28: 11.
32. Schweizer M, Fear J, Cadisch G. Rapid Commun. Mass Spec-trom. 1999; 13: 1284.
33. Dungait JAJ, Stear NA, van Dongen BE, Bol R, Evershed RP.Rapid Commun. Mass Spectrom. 2008; 22: 1631.
34. Kracht O, Gleixner G. Org. Geochem. 2000; 31: 645.35. Derrien D, Marol C, Balabane M, Balesdent J. Eur. J. Soil Sci.
2006; 57: 547.36. Amelung W, Bol R, Friedrich C. Rapid Commun. Mass Spec-
trom. 1999; 13: 1291.37. Bol R, Amelung W, Friedrich C, Ostle N. Soil Biol. Biochem.
2000; 32: 1337.38. Bol R, Amelung W, Friedrich C. Eur. J. Soil Sci. 2004; 55: 71.39. Fangueiro D, Chadwick D, Dixon L, Bol R. Soil Biol. Biochem.
2007; 39: 2608.40. Bol R, Ostle NJ, Friedrich C, Amelung W, Sanders I. Isot.
Environ. Health Stud. 1999; 35: 97.41. Gleixner G, Bol R, Balesdent J.Rapid Commun.Mass Spectrom.
1999; 13: 1278.42. Gleixner G, Poirier N, Bol R, Balesdent J.Org. Geochem. 2002;
33: 357.43. Bahri H, DignacMF, Rumpel C, Rasse DP, Chenu C,Mariotti
A. Soil Biol. Biochem. 2006; 38: 1977.44. Bahri H, Rasse DP, Rumpel C, Dignac MF, Bardoux G,
Mariotti A. Soil Biol. Biochem. 2008; 40: 1916.45. Dignac M-F, Bahri H, Rumpel C, Rasse DP, Bardoux G,
Balesdent J, Girardin C, Chenu C, Mariotti A. Geoderma2005; 128: 3.
46. Rasse DP, DignacMF, Bahri H, Rumpel C,Mariotti A, ChenuC. Eur. J. Soil Sci. 2006; 57: 530.
47. Lopez-Capel E, Abbott GD, Thomas KM, Manning DAC.J. Anal. Appl. Pyrol. 2006; 75: 82.
48. Manning DAC, Lopez-Capel E, Barker S.Min. Mag. 2005; 69:425.
49. Lopez-Capel E, Sohi SP, Gaunt JL, Manning DAC. Soil Sci.Soc. Am. J. 2005; 69: 136.
50. Lopez-Capel E, Bol R, Manning DA. Rapid Commun. MassSpectrom 2005; 19: 3192.
51. Sauheitl L, Glaser B, Bol R. Rapid Commun. Mass Spectrom2005; 19: 1437.
Rapid Commun. Mass Spectrom. 2010; 24: 495–500
DOI: 10.1002/rcm




















