
Anusa et al, 2012 AJAS
12
This article was downloaded by: [INASP - ZIMBABWE ] On: 03 April 2012, At: 00:14 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK African Journal of Aquatic Science Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/taas20 The influence of pool size on species diversity and water chemistry in temporary rock pools on Domboshawa Mountain, northern Zimbabwe A Anusa a b , HGT Ndagurwa a b & CHD Magadza a a Tropical Resources Ecology Programme (TREP), Department of Biological Sciences, Faculty of Science, University of Zimbabwe, PO Box MP 167, Mount Pleasant, Harare, Zimbabwe b National University of Science and Technology (NUST), PO Box AC 939 Ascot, Bulawayo, Zimbabwe Available online: 02 Apr 2012 To cite this article: A Anusa, HGT Ndagurwa & CHD Magadza (2012): The influence of pool size on species diversity and water chemistry in temporary rock pools on Domboshawa Mountain, northern Zimbabwe, African Journal of Aquatic Science, 37:1, 89-99 To link to this article: http://dx.doi.org/10.2989/16085914.2012.666378 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of Anusa et al, 2012 AJAS

This article was downloaded by: [INASP - ZIMBABWE ]On: 03 April 2012, At: 00:14Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
African Journal of Aquatic SciencePublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/taas20
The influence of pool size on species diversityand water chemistry in temporary rock pools onDomboshawa Mountain, northern ZimbabweA Anusa a b , HGT Ndagurwa a b & CHD Magadza aa Tropical Resources Ecology Programme (TREP), Department of Biological Sciences,Faculty of Science, University of Zimbabwe, PO Box MP 167, Mount Pleasant, Harare,Zimbabweb National University of Science and Technology (NUST), PO Box AC 939 Ascot, Bulawayo,Zimbabwe
Available online: 02 Apr 2012
To cite this article: A Anusa, HGT Ndagurwa & CHD Magadza (2012): The influence of pool size on species diversity andwater chemistry in temporary rock pools on Domboshawa Mountain, northern Zimbabwe, African Journal of AquaticScience, 37:1, 89-99
To link to this article: http://dx.doi.org/10.2989/16085914.2012.666378
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss, actions,claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

African Journal of Aquatic Science 2012, 37(1): 89–99Printed in South Africa — All rights reserved
Copyright © NISC (Pty) LtdAFRICAN JOURNAL OF
AQUATIC SCIENCEISSN 1608-5914 EISSN 1727-9364
http://dx.doi.org/10.2989/16085914.2012.666378
African Journal of Aquatic Science is co-published by NISC (Pty) Ltd and Taylor & Francis
The influence of pool size on species diversity and water chemistry in temporary rock pools on Domboshawa Mountain, northern Zimbabwe
A Anusa1,2, HGT Ndagurwa1,2* and CHD Magadza1
1 Tropical Resources Ecology Programme (TREP), Department of Biological Sciences, Faculty of Science, University of Zimbabwe, PO Box MP 167 Mount Pleasant, Harare, Zimbabwe2 Current address: National University of Science and Technology (NUST), PO Box AC 939 Ascot, Bulawayo, Zimbabwe * Corresponding author, e-mail: [email protected]
The effect of pool size (area and depth) on species diversity and physicochemical characteristics of rock pool habitats on Domboshawa Mountain, northern Zimbabwe, was studied from December 2006 to May 2007. Pools were categorised based on maximum depth. Pool duration was a key factor structuring pool communities, driving their species diversity and nutrient content. Active predatory insects (Coleoptera, Odonata, Hemiptera) and zooplank-ton (Cladocera, Copepoda, Rotifera species) were associated with long-lived pools. As pool duration increased, early phytoplankton communities dominated by short-residence green algae were replaced by blue-green algae. The number of species present increased as pool area increased. Using depth as a proxy for disturbance, species composition in rock pools was influenced by the duration of inundation. A unique rock pool community with a filter-feeding component dominated by Cladocera, and from which large branchiopods were absent, is described. Nutrient status and community diversity in rock pools are determined by pool size, and pool depth, a proxy for habitat duration, is a major structuring factor in these temporary aquatic habitats.
Keywords: community structure, disturbance, macroinvertebrates, morphometry, nutrients, phytoplankton, pool duration, zooplankton
Introduction
Temporary water bodies occur worldwide but are most abundant in arid and semi-arid regions (Williams 2006). These habitats are characterised by aquatic phases varying in length from a few days to several months (Blaustein and Schwartz 2001) and recurrent dry phases of varying duration (Williams 1987). The duration of inundation in temporary water bodies determines abiotic and biotic interactions which influence species richness, predator (Skelly 1996) and community structure (Ward and Blaustein 1994, Schneider and Frost 1996). Freshwater rock pools include depressions on rocky substrata, fed by precipitation, rivers and groundwater, which periodically hold fresh water (Jocqué et al. 2010). However, organisms in groundwater-fed rock pools and riverbed rock pools are brought in with the water, whereas precipitation-dependent rock pool communities contain organisms able to survive varying periods of desiccation (Jocqué et al. 2010). This paper describes the more typical rain-fed freshwater rock pools characterised by dynamic and unpredictable flooding cycles. Rock pools around the world originate through weathering and erosion (Campbell 1997) resulting in very similar geomorphological structures and abiotic environments, usually being pan- or bucket-shaped with variable dimensions. The similarity in rock pool environment and comparability enables analysis of patterns across different spatial scales between pool systems in different regions and even across continents (Brendonck et al. 2010).
Temporary freshwater rock pool habitats have been studied in South Africa on the Korannaberg (Vanschoenwinkel et
al. 2007, 2008, 2010) and in Botswana at Thamaga and Kgale Siding pools (Brendonck et al. 1998, Brendonck et al. 2000, Jocqué et al. 2007a, Hulsmans et al. 2008). The length of the inundation cycle (hydroperiod) and its variation, frequency and periodicity (hydroregime) were shown to be key structuring factors in the pools (Hulsmans et al. 2008) and many other studies report on the relation between hydroperiod and life-history characteristics such as growth rate, maturation rate and hatching phenology (Hathaway and Simovich 1996, Marcus and Weeks 1997, Brendonck and Riddoch 2000, 2001). Hydroperiod and pool basin depth have different individual effects on biota, but there is often a strong correlation between maximum depth and average hydroperiod (Brooks and Hayashi 2002, Vanschoenwinkel et al. 2009). Pool size has also been shown to influence physico chemical parameters (Williams 1987, Scholnick 1994) in temporary water bodies. Jocqué et al. (2010) describe variation in physicochemical parameters in temporary rock pools, and pool chemistry can fluctuate widely over short periods due to the small size of these water bodies. Nutrients may be abundant immediately after filling, and enrichment occurs mainly through decomposition of dead aquatic organisms, faeces from large terrestrial vertebrates and organic material that gets blown in (McLachlan 1981, Osborne and McLachlan 1985).
Time is probably the underlying environmental factor affecting community composition through influence on coloni-sation process and taxa replacement (Jocqué et al. 2007a).
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12

Anusa, Ndagurwa and Magadza90
This results in rock pool community composition changes determined by habitat persistence. As in other types of temporary water bodies, organisms inhabiting rock pools fall into two categories, passive and active dispersers (Wiggins et al. 1980). The former survive the dry periods in situ via drought-resistant resting stages, whereas active dispersers such as insects are usually only present in the rock pools during the wet phase, migrating away before they dry up (Williams 2006). Taxon richness follows different trajectories in pools of different hydroperiods, and the colonisation processes may also differ. Generally, slow-maturing organisms (i.e. active dispersers) and fast-maturing organisms associate with long-lived and short-lived pools, respectively (Schenk et al. 2004).
In Zimbabwe, rock pools are locally abundant on rocky outcrops on Domboshawa Mountain and these temporary habitats facilitate the quantification of important population and community structuring processes which are often hard or impossible to quantify in larger, more complex systems (Brendonck et al. 2010). Also, because they are structurally simple, small and pristine, they represent convenient systems in which to study ecological concepts (Srivastava et al. 2004). In rock pools the use of morphometric basin properties (e.g. depth) as proxies for aspects of pool hydroregime, such as hydroperiod, can be justified (Jocqué et al. 2006, Altermatt et al. 2009). Therefore we tested the influence of pool morphom-etry on species diversity in rock pools. We also investigated the temporal variation of physicochemical parameters and the influence of pool duration on pool communities (i.e. macroin-vertebrates, zooplankton and phytoplankton). We hypothe-sised that temporal variation in physicochemical parameters differed with pond hydroperiod (Williams 1987, Scholnick 1994) and expected community composition to be influenced by the duration of the pools (Vanschoenwinkel et al. 2010).
Materials and methods
Study areaDomboshawa Mountain (17°35′ N, 31°10′ E) is located 25 km north of Harare, in the Mashonaland East province at an elevation of 1 638 m. Domboshawa lies in the Chinamora porphyritic granite complex, which covers an area of 600 km2. The Chinamora granite is exposed in bare dome-shaped hills and steep-walled dwalas that characterise the Domboshawa- Chinamora uplands (Baldock 1999). Domboshawa Mountain is a mushroom-shaped intrusive, with a broad-domed top formed from exfoliated pophyritic granite (Baldock 1999). The soils are sandy, often with shiny feldspar grains, and support a sparse broom of resurrection plants (Myrothamnus flabel-lifolios) and grassland vegetation. Most of the rock surface is covered by lichens and mosses (Baldock 1999). The rainy season lasts from October/November to April, when most of the rain falls as sporadic heavy convectional storms with intermittent showers, followed by a dry season from May to September. The seasonal rainfall at Domboshawa ranges between 400 and 1 150 mm (Nyamapfene 1991). During the study period the area received 519 mm of rainfall and the air temperature averaged 27.94 °C (SE 0.86).
Data collection Eleven rock pools were selected on rocky outcrops of Domboshawa Mountain and were sampled once every week from 16 December 2006 to 5 May 2007. No aquatic
macrophytes grew in any of the pools. Depth was measured conventionally at the center of the pool basin to the nearest 5 mm using a measuring rod (Brendonck et al. 2000). Pool area was determined by layering 25 cm × 25 cm grids on a scale drawing of the pool and summing the number of grids covering the pool. For truncated grids the area of a trapezium was used to calculate the area. We used a hierarchical cluster analysis (HCA) to categorise pools based on water level (maximum depth) using Euclidian distances as the measure between clusters (SPSS 16 for Windows). The pools were grouped into three duration categories using the maximum depth (dmax) of each pool: short-lived (S), 24.21 cm (SE 1.74); medium-lived (M), 35.62 cm (SE 2.02); and long-lived (L) 77.82 cm (SE 2.45). We used maximum depth because it gives a reasonable indication of the maximum duration of the inundation period (Jocqué et al. 2006, Vanschoenwinkel et al. 2009). The number of samples collected varied among the pools as some dried and refilled during the study period, S (n = 12), M (n = 15) and L (n = 21). The short-lived pools dried up during the 13th week and the medium-lived ones during the 15th week. The long-lived pools had declined to <50% of their maximum depth by the 22nd week.
Dissolved oxygen concentration (mg l–1), water tempera-ture (°C), conductivity (μS cm–1) and pH were measured on site using WTW meters (conductivity meter 330i, O2 meter 315i, pH meter 340; WTW, Weilheim, Germany). Samples of water were collected in 750 ml polythene bottles and refrig-erated at 4 °C for preservation before analysis (Mackereth 1978). The TNT Persulfate Digestion Method 10072 was used to determine total phosphorus and total nitrogen was determined using the TNT Phosver 3 Persulfate Digestion Method as described by HACH (1996–2000). Plankton was collected using plankton nets of 20 μm and 64 μm mesh for phytoplankton and zooplankton, respectively. Phytoplankton samples were fixed on site with 10% formalin for later microscopic examination (Bartram and Balance 1996). We determined phytoplankton composition quantitatively by pouring a 1 ml sample into a counting chamber, identi-fying, and counting under the microscope (Prescott 1970). Primary production was determined by incubating clear and dark bottles in the pools for six hours from 09:00 to 15:00 at each sampling. Gross primary production was estimated as the difference between dissolved oxygen concentra-tion in clear and dark bottles, following Marra (2002). All zooplankton samples were quantitative and were taken using a hand-held 64 μm mesh zooplankton net in all the microhabitats of the pool. All microhabitats were covered so as to obtain a representative composite sample for each pool. The samples were fixed with 10% formalin on site and later washed into a 250 ml bottle with 70% alcohol for preservation. The composition was examined quantitatively by pouring the sample through a 3-tiered sieve system, and identification and counting followed Jacobs (1986). We identi-fied Coleoptera, Diptera, Ephemeroptera, Hemiptera and Odonata to family level, Cladocera, Copepoda, Cyclopoida and Rotifera to species level, and Oligochaeta to order level.
Data analysisSpecies diversity was calculated using the Shannon-Wiener index (H) (Krebs 1999) for the pooled invertebrates and phytoplankton species observed in each pool. We used one-way analysis of variance (ANOVA) to test for differences
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12

African Journal of Aquatic Science 2012, 37(1): 89–99 91
in species diversity among the pools. When F-tests indicated significant differences, means were compared using post hoc Tukey HSD (p < 0.05). A regression analysis was used to establish the relationship between the pools’ morpho-metric parameters (area and depth) and abiotic variables (pH, conductivity, dissolved oxygen, total nitrogen, and total phosphorus). The relationship between species diversity and primary production was also tested using a regres-sion analysis. Abiotic variables were log-transformed, with the exception of pH. Stepwise multiple regression analysis was used to determine the significant predictors of species diversity in the pools. We used a correspond-ence analysis biplot to visualise temporal variation in
community composition (macroinvertebrates, zooplankton and phytoplankton) during inundations in pools with different durations (Vanschoenwinkel et al. 2010). All abundance data were logarithmically transformed (log x+1) prior to analysis. The relationship between all groups (phytoplankton, macroin-vertebrates and zooplankton) and between groups and duration classes were explored using principal component analysis (PCA). We also performed a PCA to establish the relationship between abiotic and morphometric variables. In all our tests α = 0.05. We conducted statistical tests using XLSTAT (Addinsoft 2010, Germany) and SPSS 16 for Windows (SPSS Inc., 2007, Chicago, Illinois).
Results
Community structureA total of 12 macroinvertebrate taxa belonging to five orders were recorded from the rock pools (Table 1). Abundance was higher in L pools than in S and M pools. Aeshnidae, Notonectidae and Naucoridae occurred commonly in all pools (Table 1). A total of 19 zooplankton species from four orders were recorded in all the pools (Table 2). Rotifers
TaxonPool longevity
Short Medium Long Copepoda
Diaptomus castor* 1 2 3Cladocera
Chyodorus gibbus* 1 2 1Daphnia pulex 9 8 15Moina dubia* 1 2 1Pleuroxus chappuissi* 2 2 1Pleuroxus procurvatus* 1 2 1
CyclopoidaCyclopoid nauplii 6 5 13Ectocyclops hirsutus* 1 2 1Eucyclops leuckarti* 1 2 3Eucyclops serrulatus* 1 2 1Halicyclops magniceps* 1 2 3
Rotifera Asplanchna girodi 17 12 39Brachionus caudatus 3 3 10Brachionus paludosa 4 6 11Conochilus unichornis 1 7 23Epiphanes clavulata 12 8 20Lepadella patella* 4 2 3Rotaria citrine* 1 2 1Synchaeta pedinetum 1 5 3
* Denotes species present at <2 samplings
Table 2: Zooplankton abundance (individuals l–1) in rock pools of short, medium and long longevity on Domboshawa Mountain from December 2006 to May 2007
Order Family Pool longevity
Short Medium Long Odonata Aeshnidae 44 7 135
Libelliludae 10 3 25Coleoptera Dytiscidae 15 12 28
Hydranidae 7 9 21Diptera Chironomidae 11 44 18
Diptera larva 1 2 25Hemiptera Corixidae 9 8 24
Naucoridae 41 26 93Notonectidae 36 17 115
Oligochaeta – 6 13 3Ephemeroptera Prosopistomatidae 14 7 39Anura Tadpole 23 12 44
Table 1: Macroinvertebrate abundance in rock pools of short, medium and long longevity on Domboshawa Mountain from December 2006 to May 2007
Table 3: The abundance (individuals l–1) of phytoplankton species in rock pools of short, medium and long duration on Domboshawa Mountain from December 2006 to May 2007
TaxonPool longevity
Short Medium LongAnkyra judayi 6 15 29Arthrodesmus mucronulatus* 1 1 1Asterocystis smargadina* 1 2 2Batrachospermum monilliforme* 2 3 2Chlamydomonas polypyredenium* 1 2 3Closterium aciculare* 1 2 5Cylindrocystis brebissonni 4 9 12Ceratium hirudinella* 1 4 3Chladophora holsatica* 2 3 2Chlorococcum humicola* 1 2 5Chrysarachnian insidans 3 5 11Chlamydomyxa labrynthoides* 1 8 2Cosmarium margariatum* 2 3 9Coelastrum microporum* 1 2 8Closterium parvulum* 6 1 1Euglena viridis* 5 1 2Microcystis aeruginosa 93 253 2 032Oocystis lacustris* 1 2 9Poriphyridium cruentum* 8 9 6Perone dimorpha 2 3 16Peridinium willei* 1 2 1Stigochlonium aestivale* 4 7 2Synechocystis aquatilis 44 100 610Schizogonium murale* 1 2 2Staurastrum pingue 1 2 27Spirogyra rhizobrachiales 4 14 25Stipitococcus vasiformis* 1 2 5Tetraedron caudatum* 1 2 5Tabellaria flocculosa 10 23 16Tetraspora gelatinosa 8 12 19Trachelomonas lafvrei* 1 2 4Uronema elongatum* 1 2 3Urococcus insignis* 1 2 7Ulothrix zonata 6 10 18* Denotes species that were present at <2 samplings
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12

Anusa, Ndagurwa and Magadza92
DIMENSION 1: 22.7%−1.0 −0.5 0
−1.0
−0.5
0
0.5
−1.5
DIM
EN
SIO
N 2
: 16.
0%
0.5 1.0 1.5 2.0
1.0
1.5
L
L
L
L
LM
M
M
MM
S
S
S
S
Ashn
A. gir
B. cau
B. pal
Chiron
Corix
C. unic
C. nau
DystD. pul
E. clav
Hyran
Lib Noto Nauc
Oligo
Pros
S. ped
Tadpole
t1
t2
t3
t4
t1
t2
t3
t1
t2
t3
t4
Figure 1: Correspondence analysis biplot illustrating the relationships among invertebrate communities in pools with different duration on Domboshawa Mountain from December 2006 to May 2007 (S = short-lived, M = medium long-lived, L = long-lived pools) at different phases of community assembly (time since inundation: t1 = 35 d, t2 = 70 d, t3 = 105 d, t4 = 147 d). Variation explained by each axis is given after the axis title
S
S
S M
M
M
L
L
LL
MA. jud
C. breb
C. ins
M. aer
P. dim
S. aes
S. aqu
S. pin
S. rhiz
T. flocU. zon
DIMENSION 1: 52.8%. .
1.
1.
2.
2.
2.
DIM
EN
SIO
N 2
: 17.
5%
1.
1.
.LMS
t1
t1
t1t2
t2
t2 t
tt
t
t
Figure 2: Correspondence analysis biplot illustrating the relationships among phytoplankton communities in pools of different duration on Domboshawa Mountain from December 2006 to May 2007 (S = short-lived, M = medium long-lived, L = long-lived pools) at different phases of community assembly (time since inundation: t1 = 35 d, t2 = 70 d, t3 = 105 d, t4 = 147 d). Variation explained by each axis is given after the axis title
Ashn = AeshnidaeA. gir = Asplanchna girodi B. cau = Brachionus caudatus B. pal = Brachionus paludosa Chiron = Chironomidae Corix = CorixidaeC. unic = Conochilus unichonis C. nau = Cyclopoid nauplii Dyst = DytiscidaeD. pul = Daphnia pulex E. clav = Epiphanes clavulataHyran = HyraenidaeLib = Libelliludae Noto = NotonectidaeNauc = Naucoridae Oligo = Oligochaetae Pros = Prosopistomatidae S. ped = Synchaeta pedinetum
A. jud = Ankyra judayi C. breb = Cylindrocystis brebissonniC. ins = Chrysarachnian insidansM. aer = Microcystis aeruginosa P. dim = Perone dimorphaS. aes = Stigochlonium aestivale S. aqu = Synechocystis aquatilisS. pin = Staurastrum pingueS. rhiz = Spirogyra rhizobrachiales T. floc = Tabellaria flocculosaU. zon = Ulothrix zonata
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12
African Journal of Aquatic Science 2012, 37(1): 89–99 93
such as Epiphanes clavulata and Asplanchna girodi were commoner in all the pools than Copepoda, Cyclopoida and Cladocera. We recorded a total of 34 phytoplankton species in all the pools (Table 3). Zooplankton and phytoplankton abundances were positively related (F1,86 = 14.85, r2 = 0.15, p < 0.001). Generally, macroinvertebrate, zooplankton and phytoplankton abundance increased as pool longevity increased.
Temporal analysis of macroinvertebrate and zooplankton communities showed a separation of S pools from M and L pools throughout their inundation cycles (Figure 1). Short-lived pools were associated with Chironomidae and Oligochaeta, whereas Corixidae, Dytiscidae, Hydranidae, Libelleludae, Naucoridae and Notonectidae were associ-ated with the M and L pools. Macroinvertebrate and zooplankton composition in M and L pools followed a more or less similar trajectory 105 d and 147 d after filling (Figure 1). The zooplankton species Asplanchna girodi, Brachionus caudatus, Conochilus unichonis and Epiphanes clavulata, a well as cyclopoid nauplii, were dominant in L pools. However, Daphnia pulex was associated with S pools and the initial communities of L pools. Phytoplankton composi-tion in L and S pools followed a similar trend 70 d and 105 d after filling. Most of the phytoplankton, except Spirogyra
rhizobrachiales, Tabellaria flocculosa and Ulothrix zonata, maintained high abundances in L pools driven by the nutrient load (Figure 2). Short-lived pools were different from M and L pools 35 d after filling, being dominated by Spirogyra rhizobrachiales, Tabellaria flocculosa and Ulothrix zonata (Figure 2).
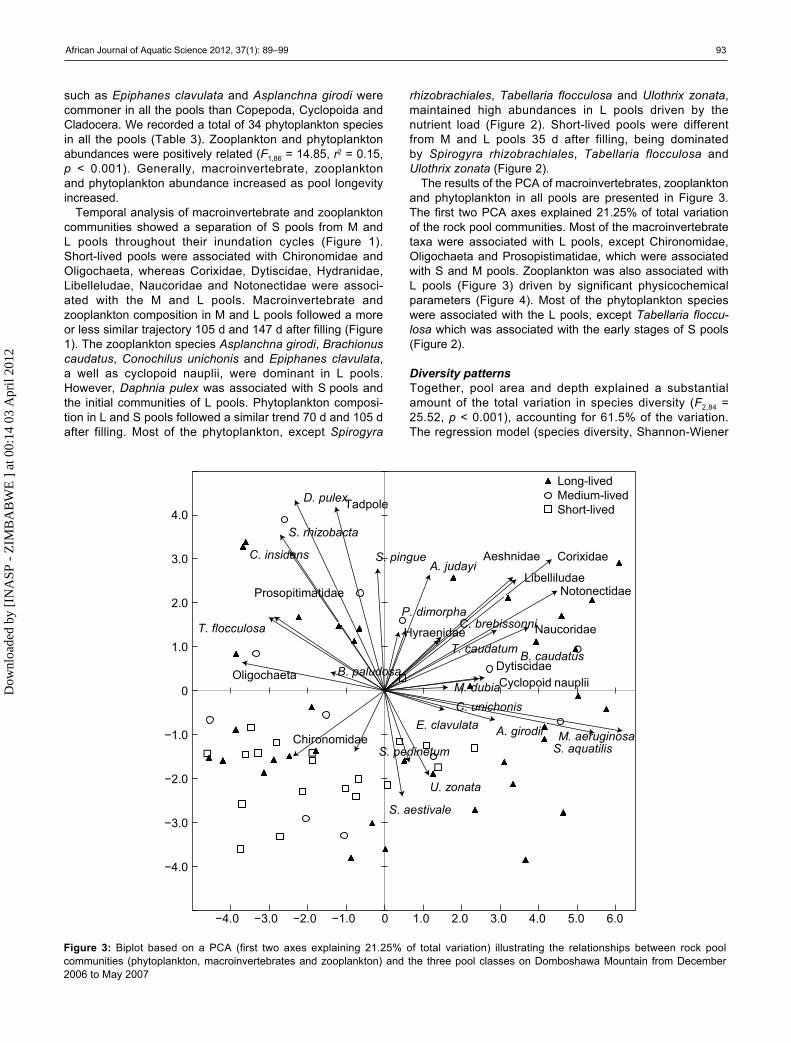
The results of the PCA of macroinvertebrates, zooplankton and phytoplankton in all pools are presented in Figure 3. The first two PCA axes explained 21.25% of total variation of the rock pool communities. Most of the macroinvertebrate taxa were associated with L pools, except Chironomidae, Oligochaeta and Prosopistimatidae, which were associated with S and M pools. Zooplankton was also associated with L pools (Figure 3) driven by significant physicochemical parameters (Figure 4). Most of the phytoplankton species were associated with the L pools, except Tabellaria floccu-losa which was associated with the early stages of S pools (Figure 2).
Diversity patternsTogether, pool area and depth explained a substantial amount of the total variation in species diversity (F2,84 = 25.52, p < 0.001), accounting for 61.5% of the variation. The regression model (species diversity, Shannon-Wiener
Libelliludae
Tadpole
Dytiscidae
Notonectidae
Corixidae
Oligochaeta
Chironomidae
Hyraenidae
Prosopitimatidae
Naucoridae
Aeshnidae
C. brebissonni
C. insidans
S. rhizobacta
P. dimorphaT. flocculosa
S. pingue
S. aquatilis
T. caudatum
M. aeruginosa
S. aestivale
U. zonata
A. judayi
D. pulex
B. paludosa
E. clavulata
M. dubia
A. girodii
C. unichonis
S. pedinetum
Cyclopoid nauplii
B. caudatus
−4.0
−4.0
−3.0
−3.0
0
−1.0
−1.0
0
2.0
2.0
−2.0
−2.0 1.0
1.0
3.0
3.0
4.0
4.0 5.0 6.0
Long-livedMedium-livedShort-lived
Figure 3: Biplot based on a PCA (first two axes explaining 21.25% of total variation) illustrating the relationships between rock pool communities (phytoplankton, macroinvertebrates and zooplankton) and the three pool classes on Domboshawa Mountain from December 2006 to May 2007
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12
Anusa, Ndagurwa and Magadza94
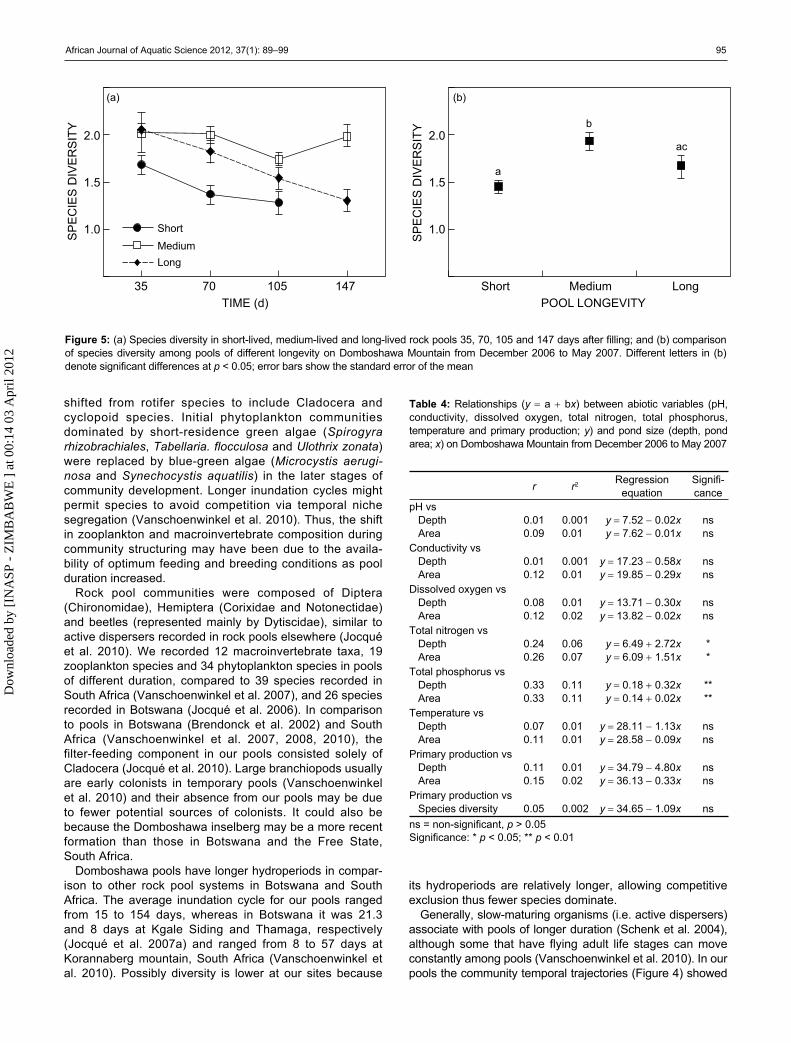
index [H] = 0.15Area – 2.10Depth + 1.01) included pool area first because of its high correlation to species diversity (r = 0.49, n = 87, p < 0.001). The model indicated that shallow rock pools with a large surface area (medium-lived pools) contained the highest diversity of species. Species diversity declined with time since inundation in the long-lived and short-lived pools (Figure 5a). A comparison of the pools indicated that medium-lived pools were signif-icantly (F2,84 = 7.75, p < 0.001) more diverse than short-lived and long-lived pools (Figure 5b). Primary production in the pools was not dependent on pool size and species composition (Table 4).
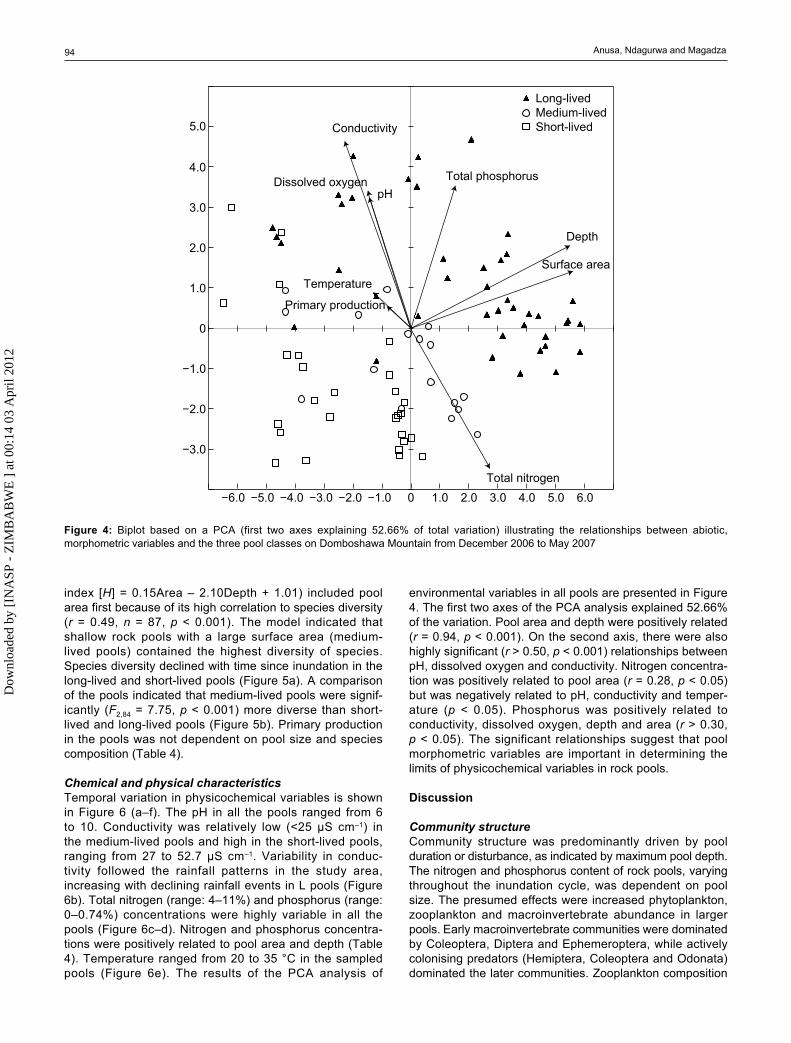
Chemical and physical characteristicsTemporal variation in physicochemical variables is shown in Figure 6 (a–f). The pH in all the pools ranged from 6 to 10. Conductivity was relatively low (<25 μS cm–1) in the medium-lived pools and high in the short-lived pools, ranging from 27 to 52.7 μS cm–1. Variability in conduc-tivity followed the rainfall patterns in the study area, increasing with declining rainfall events in L pools (Figure 6b). Total nitrogen (range: 4–11%) and phosphorus (range: 0–0.74%) concentrations were highly variable in all the pools (Figure 6c–d). Nitrogen and phosphorus concentra-tions were positively related to pool area and depth (Table 4). Temperature ranged from 20 to 35 °C in the sampled pools (Figure 6e). The results of the PCA analysis of
environmental variables in all pools are presented in Figure 4. The first two axes of the PCA analysis explained 52.66% of the variation. Pool area and depth were positively related (r = 0.94, p < 0.001). On the second axis, there were also highly significant (r > 0.50, p < 0.001) relationships between pH, dissolved oxygen and conductivity. Nitrogen concentra-tion was positively related to pool area (r = 0.28, p < 0.05) but was negatively related to pH, conductivity and temper-ature (p < 0.05). Phosphorus was positively related to conductivity, dissolved oxygen, depth and area (r > 0.30, p < 0.05). The significant relationships suggest that pool morphometric variables are important in determining the limits of physicochemical variables in rock pools.
Discussion
Community structureCommunity structure was predominantly driven by pool duration or disturbance, as indicated by maximum pool depth. The nitrogen and phosphorus content of rock pools, varying throughout the inundation cycle, was dependent on pool size. The presumed effects were increased phytoplankton, zooplankton and macroinvertebrate abundance in larger pools. Early macroinvertebrate communities were dominated by Coleoptera, Diptera and Ephemeroptera, while actively colonising predators (Hemiptera, Coleoptera and Odonata) dominated the later communities. Zooplankton composition
Conductivity
Dissolved oxygen Total phosphorus
Depth
Surface area
Total nitrogen
Temperature
Primary production
pH
−4.0
5.0
−3.0−6.0 −5.0
−3.0
0
−1.0
−1.0
0
2.0
2.0
−2.0
−2.0 1.0
1.0
3.0
3.0
4.0
4.0 5.0 6.0
Long-livedMedium-livedShort-lived
Figure 4: Biplot based on a PCA (first two axes explaining 52.66% of total variation) illustrating the relationships between abiotic, morphometric variables and the three pool classes on Domboshawa Mountain from December 2006 to May 2007
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12
African Journal of Aquatic Science 2012, 37(1): 89–99 95
Figure 5: (a) Species diversity in short-lived, medium-lived and long-lived rock pools 35, 70, 105 and 147 days after filling; and (b) comparison of species diversity among pools of different longevity on Domboshawa Mountain from December 2006 to May 2007. Different letters in (b)denote significant differences at p < 0.05; error bars show the standard error of the mean
shifted from rotifer species to include Cladocera and cyclopoid species. Initial phytoplankton communities dominated by short-residence green algae (Spirogyra rhizobrachiales, Tabellaria. flocculosa and Ulothrix zonata) were replaced by blue-green algae (Microcystis aerugi-nosa and Synechocystis aquatilis) in the later stages of community development. Longer inundation cycles might permit species to avoid competition via temporal niche segregation (Vanschoenwinkel et al. 2010). Thus, the shift in zooplankton and macroinvertebrate composition during community structuring may have been due to the availa-bility of optimum feeding and breeding conditions as pool duration increased.
Rock pool communities were composed of Diptera (Chironomidae), Hemiptera (Corixidae and Notonectidae) and beetles (represented mainly by Dytiscidae), similar to active dispersers recorded in rock pools elsewhere (Jocqué et al. 2010). We recorded 12 macroinvertebrate taxa, 19 zooplankton species and 34 phytoplankton species in pools of different duration, compared to 39 species recorded in South Africa (Vanschoenwinkel et al. 2007), and 26 species recorded in Botswana (Jocqué et al. 2006). In comparison to pools in Botswana (Brendonck et al. 2002) and South Africa (Vanschoenwinkel et al. 2007, 2008, 2010), the filter-feeding component in our pools consisted solely of Cladocera (Jocqué et al. 2010). Large branchiopods usually are early colonists in temporary pools (Vanschoenwinkel et al. 2010) and their absence from our pools may be due to fewer potential sources of colonists. It could also be because the Domboshawa inselberg may be a more recent formation than those in Botswana and the Free State, South Africa.
Domboshawa pools have longer hydroperiods in compar-ison to other rock pool systems in Botswana and South Africa. The average inundation cycle for our pools ranged from 15 to 154 days, whereas in Botswana it was 21.3 and 8 days at Kgale Siding and Thamaga, respectively (Jocqué et al. 2007a) and ranged from 8 to 57 days at Korannaberg mountain, South Africa (Vanschoenwinkel et al. 2010). Possibly diversity is lower at our sites because
its hydro periods are relatively longer, allowing competitive exclusion thus fewer species dominate.
Generally, slow-maturing organisms (i.e. active dispersers) associate with pools of longer duration (Schenk et al. 2004), although some that have flying adult life stages can move constantly among pools (Vanschoenwinkel et al. 2010). In our pools the community temporal trajectories (Figure 4) showed
r r2 Regression equation
Signifi-cance
pH vs Depth Area
0.010.09
0.0010.01
y = 7.52 − 0.02xy = 7.62 − 0.01x
nsns
Conductivity vs Depth Area
0.010.12
0.0010.01
y = 17.23 − 0.58xy = 19.85 − 0.29x
nsns
Dissolved oxygen vs Depth Area
0.080.12
0.010.02
y = 13.71 − 0.30xy = 13.82 − 0.02x
nsns
Total nitrogen vs Depth Area
0.240.26
0.060.07
y = 6.49 + 2.72xy = 6.09 + 1.51x
**
Total phosphorus vs Depth Area
0.330.33
0.110.11
y = 0.18 + 0.32xy = 0.14 + 0.02x
****
Temperature vs Depth Area
0.070.11
0.010.01
y = 28.11 − 1.13xy = 28.58 − 0.09x
nsns
Primary production vs Depth Area
0.110.15
0.010.02
y = 34.79 − 4.80xy = 36.13 − 0.33x
nsns
Primary production vs Species diversity 0.05 0.002 y = 34.65 − 1.09x nsns = non-significant, p > 0.05Significance: * p < 0.05; ** p < 0.01
Table 4: Relationships (y = a + bx) between abiotic variables (pH, conductivity, dissolved oxygen, total nitrogen, total phosphorus, temperature and primary production; y) and pond size (depth, pond area; x) on Domboshawa Mountain from December 2006 to May 2007
35 70 105 147 Short Medium Long
ShortMediumLong
TIME (d) POOL LONGEVITY
SP
EC
IES
DIV
ER
SIT
Y
SP
EC
IES
DIV
ER
SIT
Y
2.0
1.5
1.0
2.0
1.5
1.0
(a) (b)
a
b
ac
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12

Anusa, Ndagurwa and Magadza96
that macroinvertebrates (i.e. active dispersers) associated with long-lived pools. The prominence of actively dispersing predatory insects (Odonata, Hemiptera and Coleoptera) in the later stages of M and L pools could be because zooplankton-rich pools might provide excellent feeding conditions for adult Hemiptera and Coleoptera (Vanschoenwinkel et al. 2010). Longer inundation cycles also provide a refuge for actively dispersing insects with an adult aquatic phase, such as several Hemiptera (Jocqué et al. 2007b). In addition, longer-lived pools remove the developmental constraints which could prevent some organisms (e.g. Odonata) occurring in short-lived pools, allowing for a transition of taxa (succession) and successful reproduction (Jocqué et al. 2007b).
Predation pressure is predicted to increase toward the end of inundations (Spencer et al. 1999). We found that
predatory insects appeared in the later successional stages of M and L pools. This was likely to reduce the importance of the abiotic environment in community structuring. However, zooplankton abundances were also high in L pools, indicating a weak link between predation and community structure (Vanschoenwinkel et al. 2010). Zooplankton enters the active community via in situ dispersal from the dormant propagule bank, and is expected to dominate initial pool communities. However, in contrast, zooplankton was abundant in the later communities of M and L pools. This may be linked to species specific responses to hatching cues (reviewed in Brendonck and De Meester 2003), or perhaps optimal feeding conditions immediately after inundation were absent due to lower densities of live food sources such as bacteria and algae (Boven et al. 2008). Perhaps, optimal feeding conditions
102030405060708090
2
4
6
8
1 3 5 7 9
pH
102030405060708090
90
2
4
6
8
10
12
102030405060708090
10
20
30
40
50
102030405060708090
0.1
0.2
0.3
0.4
0.5
0.6
0.7
1020304050607080
5
10
15
20
25
30
102030405060708090
2468
10121416
ShortMediumLongRainfall
11 13 15 17 19 21 1 3 5 7 9 11 13 15 17 19 21
1 3 5 7 9 11 13 15 17 19 21 1 3 5 7 9 11 13 15 17 19 21
1 3 5 7 9 11 13 15 17 19 21 1 3 5 7 9 11 13 15 17 19 21
10(a) (b)
(c) (d)
(e) (f)
CO
ND
UC
TIVI
TY
RAI
NFA
LL (m
m)
RAI
NFA
LL (m
m)
RAI
NFA
LL (m
m)
RAI
NFA
LL (m
m)
RAI
NFA
LL (m
m)
RAI
NFA
LL (m
m)
TOTA
L N
ITR
OG
EN (m
g l−1
)
TOTA
L PH
OSP
HO
RU
S (m
g l−1
) D
ISSO
LVED
OXY
GEN
(mg
l−1)
WEEK WEEK
TEM
PER
ATU
RE
(°C
)
Figure 6: Temporal variation of (a) pH, (b) conductivity, (c) total nitrogen, (d) total phosphorus, (e) temperature and (f) dissolved oxygen in short-, medium- and long-lived pools in relation to rainfall events on Domboshawa Mountain from December 2006 to May 2007
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12

African Journal of Aquatic Science 2012, 37(1): 89–99 97
were responsible for structuring the zooplankton community because phytoplankton and zooplankton abundance were positively related.
Blue-green algae develop abundantly when water residence times are longer (Wetzel 2001). In a similar manner Microcystis aeruginosa and Synechocystis aquatilis persisted as major components of the long-lived pool systems. Large pools tend to provide more stable physic-ochemical conditions than smaller pools (Vanschoenwinkel et al. 2009). Rock pools are classified as oligotrophic systems open to nutrient inputs and outputs (Jocqué et al. 2010). Our analysis revealed high nutrient load (nitrogen and phosphorus) in larger deeper pools than smaller pools (Figure 2). This may have resulted from the accumulation of animal dung from livestock in the study area. It may also be because large pools have a larger catchment area than smaller pools. Green algae and diatoms usually dominate in habitats of short residence times (Wetzel 2001). We consist-ently found green algae Spirogyra rhizobrachiales, Tabellaria flocculosa and Ulothrix zonata occurring abundantly in short-lived pools. These findings indicate the importance of pool duration and nutrient load in structuring the phytoplankton community in temporary rock pools.
Community species diversity patternsThe island biogeography theory states that the number of species captured increases with the size of the geographical area (MacArthur and Wilson 1967). In our analysis species diversity was positively related to pool area. Larger pools had a more diverse community than smaller pools. The controlling effect may not be area itself, but area is positively related to habitat diversity, particularly because different types of habitats are included when area is increased (Browne 1981, Meintjes 1996). Larger pools tend to provide more microhabitats than smaller pools (March and Bass 1995). As in our study, Meintjes (1996), studying crustacean-dominated pans in South Africa, also found that larger pans supported more species than smaller pans.
We found that the species composition of M pools was more diverse, followed by L pools, and that S pools were the least diverse. This follows the intermediate disturbance hypothesis (IDH) which states that diversity will be highest at sites that have had an intermediate frequency of distur-bance that prevents competitive exclusion, and will be lower at sites that have experienced very high or very low distur-bance frequencies (Connell 1978, Huston 1979). However, contrary to our findings, Vanschoenwinkel et al. (2009) reported higher taxon richness in large deep pools with long hydro periods. We used rock pool basin depth as a proxy for disturbance because Vanschoenwinkel et al. (2009) showed that there was a correlation between depth and disturbance-related parameters e.g. inundation frequency. Long-lived pools are predicted to offer a more stable habitat than smaller ones (Jocqué et al. 2007a). This may have allowed communities to build up towards some dominant species, resulting in lower species numbers. Additionally, in long-lived pools some species may finish their life cycle before the pools dry out. Collecting at this time could also result in lower species numbers. On the other hand, in short-lived pools, community development is truncated by a shorter inundation period that limits the time available for
a community to stabilise (Jocqué et al. 2007a). This time window is too short to allow several species to colonise and/or reproduce. It is expected that short-lived pools experi-ence frequent disturbances that may prohibit the addition or replacement of species. As in our study, other studies also reported lower diversity in habitats experiencing more severe disturbance regimes (Therriault and Kolosa 2001, Urban 2004, Serrano and Fahd 2005, Vanschoenwinkel et al. 2010).
Chemical and physical characteristicsPhysicochemical variables are significant in local processes that control species tolerances and distributions (Corkum 1989). Pool morphometry (depth and area) emerged as the most important variables determining the nitrogen and phosphorus content of the pools, which in turn had an effect on community structure and composition. We suggest that increased pool size (depth and area), may have resulted in additional feeding and spatial niches, allowing more species to occur in the larger pools. Boven and Brendonck (2009) also reported that, in pools with a longer filling cycle, feeding and growth conditions improved with time.
Overall, physicochemical variables except pH and dis-solved oxygen exhibited high variability among the rock pools. Water temperature in pools is dependent on climate and, in our study, temperature ranged from 20 to 35 °C. Rock pools are characterised by low conductivity immedi-ately after filling, typically fluctuating between 10 and 30 μS cm–1 (Jocqué et al. 2010). Conductivity in our pools ranged from 6 to 50 μS cm–1, similar to observations made elsewhere (Hamer and Martens 1998, Jocqué et al. 2007a). Consistent with observations from other studies, pH ranged from 6 to 10, from acidic to alkaline (Jocqué et al. 2010). Algal photosynthesis is an equilibrium process that is also coupled to dissolved oxygen production and any shifts can result in changes in pH (Wetzel 2001). The observed association between pH and dissolved oxygen suggests the possible contribution of algal photosynthesis.
Conclusions
Pool depth and area can be useful predictors of species diversity and nutrients in rock pools. Pool duration, a key structuring factor in temporary aquatic habitats, was the major determinant of rock pool communities (Vanschoenwinkel et al. 2010). However, we remain conservative in our interpre-tations, since habitat duration was not specifically analysed. This is largely because habitat size and hydroregime explain both shared and unique variation in community structure and diversity, indicating that their individual effects on biota can be different (Vanschoenwinkel et al. 2009). Further, our study describes a different type of temporary rock pool habitat inhabited by different species in comparison to those described from Botswana and South Africa. Particularly so, because of the absence of large branchiopods which may suggest that the Domboshawa inselberg may be a relatively recent formation or that the study area has fewer potential sources for colonists. Thus, the uniqueness of our rock pool systems indicates that the conservation value of temporary aquatic habitats may not be limited to habitat size but should also include variation across spatial scales.
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12

Anusa, Ndagurwa and Magadza98
Acknowledgements — The authors are indebted to the National Museums and Monuments of Zimbabwe for allowing research on Domboshawa Mountain, and to staff at the Tropical Resources Ecology Programme, University of Zimbabwe. Special thanks go to Bram Vanschoenwinkel and Luc Brendonck for their attention to earlier versions of the manuscript.
References
Altermatt F, Pajunen VI, Ebert D. 2009. Desiccation of rock pool habitats and its influence on population persistence in a Daphnia metacommunity. PLoS ONE 4(3): e4703. doi:10.1371/journal.pone.0004703.
Baldock JW. 1999. The geology of the Harare Greenstone Belt and surrounding granitic terrain. Zimbabwe Geological Survey Bulletin 4: 68–69.
Bartram J, Balance R. 1996. Water quality monitoring: a practical guide to the design and implementation of fresh water quality studies and monitoring programmes. London: Chapman and Hall.
Blaustein L, Schwartz SS. 2001. Why study ecology in temporary pools? Israel Journal of Zoology 47: 303–312.
Boven L, Brendonck L. 2009. Impact of hydroperiod on seasonal dynamics in temporary pool cladoceran communities. Fundamental and Applied Limnology 174: 147–157.
Boven L, Stoks R, Forro L, Brendonck L. 2008. Seasonal dynamics in water quality and vegetation cover in temporary pools with variable hydroperiods in Kiskunsag (Hungary). Wetlands 28: 401–410.
Brendonck L, De Meester L. 2003. Egg banks in freshwater zooplankton: evolutionary and ecological archives in the sediment. Hydrobiologia 491: 65–84.
Brendonck L, De Meester L, Riddoch B. 2000. Regional structuring of genetic variation in short-lived rock pool populations of Branchipodopsis wolfi (Crustacea: Anostraca). Oecologia 123: 506–515.
Brendonck L, Jocqué M, Hulsmans A, Vanschoenwinkel B. 2010. Pools ‘on the rocks’: freshwater rock pools as model system in ecological and evolutionary research. Limnetica 29: 25–40.
Brendonck L, Michels E, De Meester L, Riddoch B. 2002. Temporary pools are not ‘enemy-free’. Hydrobiologia 486: 147–159.
Brendonck L, Riddoch BJ. 2000. Egg bank dynamics in anostracan desert rock pool populations (Crustacea: Branchiopoda). Archiv für Hydrobiologie 148: 71–84.
Brendonck L, Riddoch BJ. 2001. Hatching characteristics of the fairy shrimp Branchipodopsis wolfi in relation to the stochastic nature of its habitat, desert rock pools. Verhandlungen des Internationalen Verein Limnologie 27: 3931–3935.
Brendonck L, Riddoch BJ, Van de Weghe V, Van Dooren T. 1998. The maintenance of egg banks in very short lived pools – a case study with anostracans (Branchiopoda). Archiv für Hydrobiologie. Special Issues on Advanced Limnology 52: 141–161.
Brooks RT, Hayashi M. 2002. Depth-area-volume and hydroperiod relationships of ephemeral (vernal) forest pools in southern New England. Wetlands 22: 247–255.
Browne RA. 1981. Lakes as islands: biogeographic distribution, turnover rates and species composition in the lakes of central New York. Journal of Biogeography 8: 75–83.
Campbell EM. 1997. Granite landforms. Journal of the Royal Society of Western Australia 80: 101–112.
Connell JH. 1978. Diversity in tropical rainforests and coral reefs. Science 199: 1302–1310.
Corkum LD. 1989. Patterns of benthic invertebrate assemblages in rivers of northwestern North America. Freshwater Biology 21: 191–205.
HACH. 1996–2000. Water analysis handbook. Loveland: HACH Company. pp 491–709.
Hamer ML, Martens K. 1998. The large Branchiopoda (Crustacea)
from temporary habitats of the Drakensberg region, South Africa. Hydrobiologia 384: 151–165.
Hathaway SA, Simovich MA. 1996. Factors affecting the distribution and co-occurrence of two southern Californian anostracans (Branchiopoda), Branchinecta sandiegonensis and Streptocephalus woottoni. Journal of Crustacean Biology 16: 669–677.
Hulsmans A, Vanschoenwinkel B, Pyke C, Riddoch BJ, Brendonck L. 2008. Quantifying the hydroregime of a temporary pool habitat: a modelling approach for ephemeral rock pools in SE Botswana. Ecosystems 11: 89–100.
Huston M. 1979. A general hypothesis of species diversity. American Naturalist 113: 81–101.
Jacobs DH. 1986. Order Hemiptera. In: Scholtz I, Holm E (eds), Insects of southern Africa. Durban: Butterworths. pp 112–175.
Jocqué MT, Martens K, Riddoch BJ, Brendonck L. 2006. Faunistics of ephemeral rock pools in southeastern Botswana. Archiv für Hydrobiologie 165: 415– 431.
Jocqué MT, Riddoch BJ, Brendonck L. 2007a. Successional phases and species replacements in freshwater rock pools: towards a biological definition of ephemeral systems. Freshwater Biology 52: 1734–1744.
Jocqué M[T], Timms BV, Brendonck L. 2007b. A contribution on the biodiversity and conservation of the freshwater fauna of rocky outcrops in the central wheatbelt of Western Australia. Journal of the Royal Society of Western Australia 90: 137–142.
Jocqué MT, Vanschoenwinkel B, Brendonck L. 2010. Anostracan monopolisation of early successional phases in temporary waters? Fundamental and Applied Limnology 176: 127–132.
Krebs CJ. 1999. Ecological methodology. Menlo Park, California: Benjamin Cummings.
MacArthur RH, Wilson EO. 1967. The theory of island biogeography. New Jersey: Princeton University Press. pp 13–17.
Mackereth FJH. 1978. Water analysis. Ambleside: Freshwater Biological Science Association, Publication No. 36.
March F, Bass D. 1995. Application of island biogeography theory to temporary pools. Journal of Freshwater Ecology 10: 83–85.
Marcus V, Weeks SC. 1997. The effects of pond duration on the life history traits of an ephemeral pond crustacean, Eulimnadia texana. Hydrobiologia 359: 213–221.
Marra J. 2002. Primary production. In: Williams PJ, Le B, Thomas DN, Reynolds CS (eds), Phytoplankton productivity: carbon assimilation in marine and freshwater ecosystems. Oxford: Blackwell Scientific Publishers. pp 78–108.
McLachlan AJ. 1981. Food sources and foraging tactics in tropical rain pools. Zoological Journal of the Linnean Society 71: 265–277.
Meintjes S. 1996. Seasonal changes in the invertebrate community of small shallow ephemeral pans at Bain`s Vlei, South Africa. Hydrobiologia 317: 51–64.
Nyamapfene K. 1991. The soils of Zimbabwe. Harare: Nehanda Publishers. pp 71–74.
Osborne PL, McLachlan AJ. 1985. The effect of tadpoles on algal growth in temporary rain-filled rock pools. Freshwater Biology 15: 77–88.
Prescott JW. 1970. How to know freshwater algae (2nd edn). Iowa: William C Brown Publishers.
Schenk K, Suhling F, Martens A. 2004. Egg distribution, mate-guarding intensity and offspring characteristics in dragonflies (Odonata). Animal Behaviour 68: 599–606.
Schneider WD, Frost TM. 1996. Habitat duration and community structure in temporary ponds. Journal of the North American Benthological Society 15: 64–86.
Scholnick DA. 1994. Seasonal variation and diurnal fluctuations in ephemeral desert pools. Hydrobiologia 294: 111–116.
Serrano L, Fahd K. 2005. Zooplankton communities across a hydroperiod gradient of temporary ponds in the Doñana National Park (SW Spain). Wetlands 25:101–111.
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12

African Journal of Aquatic Science 2012, 37(1): 89–99 99
Skelly DK. 1996. Pond drying, predators, and the distribution of Pseudcaris tadpoles. Copeia 1996: 599–605.
Spencer M, Blaustein L, Schwartz SS, Cohen JE. 1999. Species richness and the proportion of predatory animal species in temporary freshwater pools: relationships with habitat size and permanence. Ecology Letters 2: 157–166.
Srivastava DS, Kolasa J, Bengtsson J, Gonzalez A, Lawler SP, Miller TE, Munguia P, Romanuk T, Schneider DC, Trzcinski MK. 2004. Are natural microcosms useful model systems for ecology? Trends in Ecology and Evolution 19: 379–384.
Therriault TW, Kolasa J. 2001. Desiccation frequency reduces species diversity and predictability of community structure in coastal rock pools. Israel Journal of Zoology 47: 477–489.
Urban MC. 2004. Disturbance heterogeneity determines freshwater metacommunity structure. Ecology 85: 2971–2978.
Vanschoenwinkel B, De Vries C, Seaman M, Brendonck L. 2007. The role of metacommunity processes in shaping invertebrate rock pool communities along a dispersal gradient. Oikos 116: 1255–1266.
Vanschoenwinkel B, Gielen S, Vandewaerde H, Seaman M, Brendonck L. 2008. Relative importance of different dispersal vectors for small aquatic invertebrates in a rock pool meta-community. Ecography 31: 567–577.
Vanschoenwinkel B, Hulsmanss A, De Roeck ER, De Vries C, Seaman M, Brendonck L. 2009. Community structure in temporary freshwater pools: disentangling effects of habitat size and hydroregime. Freshwater Biology 54: 1487–1500.
Vanschoenwinkel B, Waterkeyn A, Jocqué M, Boven L, Seaman M, Brendonck L. 2010. Species sorting in space and time — the impact of disturbance regime on community assembly in a temporary pool metacommunity. Journal of North American Benthological Society 29: 1267–1278.
Ward D, Blaustein L. 1994. The overriding influence of flash floods on species-area curves in ephemeral Negev desert pools: a consideration of the value of island biogeography. Journal of Biogeography 21: 595–603.
Wetzel RG. 2001. Limnology: lake and river ecosystems (3rd edn). California: Academic Press.
Wiggins GB, Mackay RJ, Smith IM. 1980. Evolutionary and ecological strategies of animals in annual temporary pools. Archives of Hydrobiology 58: 97–206.
Williams DD. 1987. The ecology of temporary waters. London: Croom Helm.
Williams DD. 2006. The biology of temporary waters. New York: Oxford University Press.
Received 12 February 2011, accepted 4 October 2011
Dow
nloa
ded
by [
INA
SP -
ZIM
BA
BW
E ]
at 0
0:14
03
Apr
il 20
12




















