
Antisense expression of the 20-hydroxyecdysone receptor (EcR) in transfected mosquito cells uncovers...
8
In VitroCell. Dev.Biol. Animal 37:522-529, September 2001 2001 Society tbr In VitroBiology 1071-2690/01 $10.00+0.00 ANTISENSE EXPRESSION OF THE 20-HYDROXYECDYSONE RECEPTOR (EcR) IN TRANSFECTED MOSQUITO CELLS UNCOVERS A NEW EcR ISOFORM THAT VARIES AT THE C-TERMINAL END G. JAYACHANDRAN 1 AND A. M. FALLON Department of Entomology, University of Minnesota, 1980 Folwell Avenue, St. Paul, Minnesota 55108 (Received 13 October 2000; accepted 21 March 2001) SUMMARY The insect steroid hormone 20-hydroxyecdysone initiates a cascade of regulatory events in a temporal and tissue- specific manner by first binding to a complex of an ecdysone receptor (EcR) protein and a ultraspiracle protein. Using an antisense (As) ribonucleic acid approach, we show that disruption of EcR expression in transfected C7-10 cells from the mosquito Aedes albopictus affects survival and growth. From stably transfected cells, we recovered a new isoform of A. albopictus AalEcRa, which is named AalEcRb. The deduced amino acid sequence of AalEcRb was almost identical to that of AalEcRa, with the exception of a seven amino acid sequence near the C-terminus. Using polymerase chain reaction followed by restriction enzyme analysis, we found that AalEcRa is the predominant species expressed by wild- type C7-10 cells, while cells transfected with As-EcR expressed both isoforms at approximately equal levels. Key words: Aedes albopictus; C7-10 cell line; ecdysone receptor; antisense ribonucleic acid; transfection. INTRODUCTION We have recently reported that Aedes albopictus C7-10 mosquito ceils in tissue culture express an ecdysone receptor (EeR) (Jaya- chandran and Fallon, 2000) that had 97% amino acid sequence identity to an EcR from Aedes aegypti (Wang et al., 1998). The EcR is an integral part of the steroid hormone-mediated transcriptional cascade, and its presence in C7-10 cells is consistent with the evidence that these cells mount a physiological response to 20- hydroxyecdysone (20E) (Gerenday and Fallon, 1996). The temporal and spatial regulation of three distinct EcR isoforms has been de- scribed in Drosophila melanogaster (Talbot et al., 1993), and the single A. aegypti EcR has been shown to he present in two distinct subpopulations in vitellogenic mosquitoes (Miura et al., 1999). To gain an insight into the physiological importance of EcR in C7-10 cells in culture we first studied whether EcR transcripts are translated into detectable levels of protein product in these ceils. Secondly, we also expressed antisense (As) ribonucleic acid (RNA) in transfectcd cells to examine the effect of inhibiting EcR expres- sion on the properties of the cells. The As strategy can be used to inactivate gene expression in transfected cells by directing synthesis of RNA complementary to the gene of interest. In an early application, for example, Rosenberg et al. (1985) demonstrated that transcription of the noncoding strand resulted in the production of phenocopies of Krtippel mutant em- bryos in D. melanogaster. Later, it was shown that stable transfor- mation of Drosophila tissue culture cells with a gene encoding heat- inducible RNA complementary to the hsp26 message was highly specific for hsp26 itself, and did not affect other heat shock proteins To whom correspondence should be addressed at E-mail: jayac001@tc. umn.edu (McGarry and Lindquist, 1986). More recently, this targeted gene silencing technique has been used to show that the sensitivity of mosquito cells to methotrexate is affected by the expression of As constructs encoding the mosquito dihydrofolate reductase (dhfr) gene (Shotkoski and Fallon, 1994). In the present study, we show that interference with the normal expression of the EcR compro- mises growth and survival of transfected C7-10 cells. Moreover, analysis of the expression of the As construct in the most severely aftected cells uncovered a second mosquito EcR isoform. MATFRIALS AND METItOI)S Cell culture and tran,~'~ction. The A. albopictas (C7-10) mosquito cells were maintained in E-5 medium containing 5% t~tal bovine serum essen- tially as described previously (Falhm and Stollar, 1987). E-0 is a serum- free medium, and E-10 contained 10% serum. For transt~ction, plasmid deoxyribonucleic acids (DNAs) were prepared using the Qiagen Maxi plas- mid isolation kit (Valencia, CA) as per the manufacturer's instructions. C7- 10 cells were cotransfected with pDHFR9 (Shotkoski and Fallon, 1993) and EeR constructs at a molar ratio of 1:1 using Lipofectamine (Life Tech- nologies, Roekville, MD) as a transt~ction agent. Total DNA concentrations ranged from 2 to 30 ~g per plate. Transl~ctions were carried out in E-0 medium, which was replaced by E-10 8 h alter transt~ction. E-10 was replaced with E-5 24 h after transfection. Seventy-two hours after trans- fection, the transformed cells were selected in 1 t~M methntrexate (Shot- koski and Fallon, 1993). After recove~ 7 of stable transformants, three in- dividual clones transfected with As-DNA, and one clone transfected with sense (S)-DNA were chosen for analysis. Clones were maintained in E-5 medium containing 1 p~M methotrexate. Isolation of nuclear and cytoplasmicproteinfractions. Nuclei were obtained from C7-10 cells as described by Wu et al. (1979) with slight modifications. Cells grown in monolayer culture to approximately 80% confluency were quick chilled at - 2 0 ~ C for 1 min. The ceils were scraped and collected by centrifugation at 2000 x g for 4 rain at 4~ C. The pellet was washed twice with phosphate-buffered saline (PBS) (Dulbecco and Vogt, 1954) and resuspended in 3 volumes of buffer A (60 mM KC1, 15 mM NaC1, 1 mM 522
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Antisense expression of the 20-hydroxyecdysone receptor (EcR) in transfected mosquito cells uncovers...

In Vitro Cell. Dev. Biol. Animal 37:522-529, September 2001 �9 2001 Society tbr In Vitro Biology 1071-2690/01 $10.00+0.00
ANTISENSE EXPRESSION OF THE 20-HYDROXYECDYSONE RECEPTOR (EcR) IN TRANSFECTED MOSQUITO CELLS UNCOVERS A NEW EcR ISOFORM THAT VARIES AT
THE C-TERMINAL END
G. JAYACHANDRAN 1 AND A. M. FALLON
Department of Entomology, University of Minnesota, 1980 Folwell Avenue, St. Paul, Minnesota 55108
(Received 13 October 2000; accepted 21 March 2001)
SUMMARY
The insect steroid hormone 20-hydroxyecdysone initiates a cascade of regulatory events in a temporal and tissue- specific manner by first binding to a complex of an ecdysone receptor (EcR) protein and a ultraspiracle protein. Using an antisense (As) ribonucleic acid approach, we show that disruption of EcR expression in transfected C7-10 cells from the mosquito Aedes albopictus affects survival and growth. From stably transfected cells, we recovered a new isoform of A. albopictus AalEcRa, which is named AalEcRb. The deduced amino acid sequence of AalEcRb was almost identical to that of AalEcRa, with the exception of a seven amino acid sequence near the C-terminus. Using polymerase chain reaction followed by restriction enzyme analysis, we found that AalEcRa is the predominant species expressed by wild- type C7-10 cells, while cells transfected with As-EcR expressed both isoforms at approximately equal levels.
Key words: Aedes albopictus; C7-10 cell line; ecdysone receptor; antisense ribonucleic acid; transfection.
INTRODUCTION
We have recently reported that Aedes albopictus C7-10 mosquito ceils in tissue culture express an ecdysone receptor (EeR) (Jaya- chandran and Fallon, 2000) that had 97% amino acid sequence identity to an EcR from Aedes aegypti (Wang et al., 1998). The EcR is an integral part of the steroid hormone-mediated transcriptional cascade, and its presence in C7-10 cells is consistent with the evidence that these cells mount a physiological response to 20- hydroxyecdysone (20E) (Gerenday and Fallon, 1996). The temporal and spatial regulation of three distinct EcR isoforms has been de- scribed in Drosophila melanogaster (Talbot et al., 1993), and the single A. aegypti EcR has been shown to he present in two distinct subpopulations in vitellogenic mosquitoes (Miura et al., 1999).
To gain an insight into the physiological importance of EcR in C7-10 cells in culture we first studied whether EcR transcripts are translated into detectable levels of protein product in these ceils. Secondly, we also expressed antisense (As) ribonucleic acid (RNA) in transfectcd cells to examine the effect of inhibiting EcR expres- sion on the properties of the cells.
The As strategy can be used to inactivate gene expression in transfected cells by directing synthesis of RNA complementary to the gene of interest. In an early application, for example, Rosenberg et al. (1985) demonstrated that transcription of the noncoding strand resulted in the production of phenocopies of Krtippel mutant em- bryos in D. melanogaster. Later, it was shown that stable transfor- mation of Drosophila tissue culture cells with a gene encoding heat- inducible RNA complementary to the hsp26 message was highly specific for hsp26 itself, and did not affect other heat shock proteins
To whom correspondence should be addressed at E-mail: jayac001@tc. umn.edu
(McGarry and Lindquist, 1986). More recently, this targeted gene silencing technique has been used to show that the sensitivity of mosquito cells to methotrexate is affected by the expression of As constructs encoding the mosquito dihydrofolate reductase (dhfr) gene (Shotkoski and Fallon, 1994). In the present study, we show that interference with the normal expression of the EcR compro- mises growth and survival of transfected C7-10 cells. Moreover, analysis of the expression of the As construct in the most severely aftected cells uncovered a second mosquito EcR isoform.
MATFRIALS AND METItOI)S
Cell culture and tran,~'~ction. The A. albopictas (C7-10) mosquito cells were maintained in E-5 medium containing 5% t~tal bovine serum essen- tially as described previously (Falhm and Stollar, 1987). E-0 is a serum- free medium, and E-10 contained 10% serum. For transt~ction, plasmid deoxyribonucleic acids (DNAs) were prepared using the Qiagen Maxi plas- mid isolation kit (Valencia, CA) as per the manufacturer's instructions. C7- 10 cells were cotransfected with pDHFR9 (Shotkoski and Fallon, 1993) and EeR constructs at a molar ratio of 1:1 using Lipofectamine (Life Tech- nologies, Roekville, MD) as a transt~ction agent. Total DNA concentrations ranged from 2 to 30 ~g per plate. Transl~ctions were carried out in E-0 medium, which was replaced by E-10 8 h alter transt~ction. E-10 was replaced with E-5 24 h after transfection. Seventy-two hours after trans- fection, the transformed cells were selected in 1 t~M methntrexate (Shot- koski and Fallon, 1993). After recove~ 7 of stable transformants, three in- dividual clones transfected with As-DNA, and one clone transfected with sense (S)-DNA were chosen for analysis. Clones were maintained in E-5 medium containing 1 p~M methotrexate.
Isolation of nuclear and cytoplasmic protein fractions. Nuclei were obtained from C7-10 cells as described by Wu et al. (1979) with slight modifications. Cells grown in monolayer culture to approximately 80% confluency were quick chilled at -20 ~ C for 1 min. The ceils were scraped and collected by centrifugation at 2000 x g for 4 rain at 4 ~ C. The pellet was washed twice with phosphate-buffered saline (PBS) (Dulbecco and Vogt, 1954) and resuspended in 3 volumes of buffer A (60 mM KC1, 15 mM NaC1, 1 mM
522

MOSQUITO EcR ISOFORMS 5 2 3
5' 5' UTR
[ NB
EcR12
EcR domains
IC I D [ E ] F
40 amino acids
EcR- 13 ?
EcR-4
T EcR-5
PCR
EcR-2 3' UTR
3'
EcR-3 EcR11
200 bp
EcR-4 EcR (1.0 kb)
I EcR-3
Klenow End-Filling
Parent Plasmid
~ S V 4 0 (1598 bp)
Hsp70 (1191 bp) ~ A T (773 bp)
XhoI (-195)
,l Linearize (Xhol)
,l Klenow End-Filling
Sense
I Blunt-end ligation
Antisense
S (-195)
I
Hsp70 EcR (1.0 kb) Hsp70
As (-195)
I
Hsp70 EcR (1.0 kb) Hsp70
FIG. 1. Cloning strategy for EcR constructs. A schematic of the AalEcR (Jayachandran and Fatlon, 2000) is shown in the left. Open boxes indicate domains of the nuclear hormone receptor and the solid box directly below indicates the coding region in the AalEcR transcript. PCR primers used in the present study are marked, with open symbols indicating forward primers, andfilled symbols indicating reverse primers. In the right, the plasmid hspCAT-1 (Di Nocera and Dawid, 1983), was linearized at a unique XhoI site (marked by a solid arrowhead), end-filled, and blunt-end ligated to the 1-kb PCR-generated fragment from the A. albopictus EcR. Clones in S and As orientation were confirmed by sequencing.
ethylenediamine tetraacetic acid, 0.1 mM ethylene glycol- bis[aminoethylether]-tetraacetic acid, 0.1 mM phenylmethylsulfonyl fluo- ride [PMSF], 0.15 mM spermine, 0.15 mM spermidine, 15 mM Tris-HC1 [pH 7.4], and 0.25 M sucrose). After addition of NP-40 (100% stock) to a final concentration of 0.25%, the cells were vortexed and checked under a microscope to confirm the lysis of ceils and presence of intact nuclei. Two volumes of buffer B (60 mM KC1, 15 nLM NaC1, 0.1 mM PMSF, 0.15 mM spermine, 0.15 mM spermidine, 15 mM Tris-HC1 [pH 7.4], 0.5 mM dithiothreitol, and 1.8 M sucrose) was added, and the nuclei were pelleted by centrifugation in a swinging bucket rotor at 20,000 • g for 15 min. The supernatant (cytoplasmic fraction) and nuclear pellet were stored separately at - 8 0 ~ C. Samples were sonicated in PBS, centrifuged at 5000 • g for 5 min and the supernatant was loaded onto a 10% polyacrylamide gel (Lae- mmli, 1970).
Western blotting. Western blotting was carried out using the Immuno-Blot assay kit fi'om BioRad Laboratories (Hercules, CA) as per the manufacturer's instructions. Protein was transferred to Protran nitrocellulose membranes (Schleicher and Schuell, Keene, NH) in Tris-glycine buffer using a BioRad transfer apparatus. After transfer, membranes were incubated overnight with the primary antibody (anti-AaEcR), which was kindly provided by Dr. A. S. Raikhel (Department of Entomology and Program in Genetics, Michigan State University, East Lansing, MI), in blocking solution (5% nonfat milk in PBS) at 1:500 dilution. The next d the membranes were washed with 20 mM Tris- HC1, pH 7.5, containing 0.5 M NaC1 and 0.05% Tween-20, as per the man- ufacturer's instructions. The membranes were incubated with diluted goat anti-rabbit secondary antibody conjugated to alkaline phosphatase (BioRad)
for 2 h at room temperature and developed according to the manufacturer's instructions.
Col~struction of As-EcR plasmids. The 1-kb DNA fragment spanning the hormone-binding domain E from the C7-10 EeR complementary DNA (eDNA) was obtained by polymerase chain reaction (PCR), using EcR3 and EcR4 primers (Fig. 1) and Taq polymerase (Promega, Madison, WI). As out- lined in Fig. 1, the PCR product was end filled using Klenow DNA poty- merase (Stratagene, La Jolla, CA). The parent plasmid HspCAT-1 (Di Noeera and Dawid, 1983) was linearized with XhoI at a unique restriction site in the Hsp70 promoter region, 195 nucleotides (nt) upstream of the CAT transcrip- tion start site. The linearized plasmid was end filled and ligated to the blunt- ended EcR fragment using T4 DNA ligase (Promega). The orientation of insertion was confirmed by PCR and sequencing. DNA was sequenced using the ABI PRISM automatic sequencing apparatus (Model 377, version 2.1.1) and analyzed using the Wisconsin Package ~ (Version 9.0) (Genetic Computer Group Inc., Madison, WI).
RESULTS
EcR protein is expressed in C7-10 cells. In a previous invest iga-
tion, we u sed reverse t r ansc r ip ta se -po lymerase cha in react ion (RT-
PCR) to obta in a hornolog of the A. aegypti EcR receptor from A.
albopictus C7-10 cells (Fig. 1). The s u c c e s s of this approach sug-
ges ted tha t the EcR gene is t r ansc r ibed in this cell cul ture sys tem,

524 JAYACHANDRAN AND FALLON
FIG. 2. Subcellular localization of EcR protein. Extracts (20 txg protein) from nuclei and cytoplasm were analyzed separately with antibody to A. aegypti EcR, generously provided by A. S. Raikhel. Panel A is the Western blot and panel B is the corresponding Coomassie blue-stained gel. Stars designate putative EcR isoforms.
TABLE 1
PROPERTIES OF STABLE EcR TRANSFORMANTS
Parameter S-3 As- 1 As-3
Total EcR DNA introduced (txg) 15 1.5 Appearance, 48 h Vacuolated Vacuolated Appearance, 1 wk Transformants begin to appear Vacuolated First clones appear (d) 8 15 Number of clones 10 d after transfection 52 - - Number of final successful stable transformants (30 d) 52 26 Appearance after two passages Normal Vacuolated Appearance after 10 passages Normal Normal Appearance after 15 passages Normal Normal Initial frequency of passage Weekly Weekly Later frequency of passage Weekly Weekly
15 Vacuolated Severe vacuolation 27
11 Vacuolated and appear sick Appear normal, but slow growth Normal Bimonthly Weekly
consistent with earlier evidence for the synthesis of ecdysone- inducible proteins (Lan et al., 1993). To extend this analysis, we used an antibody to the A. aegypti EcR to evaluate whether EcR transcripts are translated into protein in A. albopictus C7-10 ceils. Based on relative band intensity in Western blots (Fig. 2A), 20- txg protein samples from both the nucleus and the cytoplasm of C7-10 ceils contained a predominant EcR isoform that migrated at - 8 0 kDa. In the nuclear fraction, we also detected minor bands at 75, 74, and 72 kDa (Fig. 2A, lane 1), while only the 74-kDa band was present in the cytoplasmic fraction (Fig. 2A, lane 2). As expected, the abundance of these proteins was low, and these par- ticular bands were not identifiable on a corresponding stained gel (Fig. 2B).
To test whether the relative ratios of these EcR isoforms could be perturbed using As-RNA, C7-10 cells were cotransfected with pDHFR9 plasmid encoding a selectable dhfr gene (Shotkoski and Fallon, 1993) and S- or As-EcR constructs. In Table 1, the prop- erties of clones resulting from transfection with S-DNA were typ- ified by clone S-3, whereas representative clones recovered from
transformation with As-EcR are named As-1 and As-3. After transformation with S-DNA, methotrexate-resistant clones ap- peared after 8 d. In contrast, recovery of resistant clones after cotransfection with As-EcR DNA was delayed until 15 d when cells were exposed to 1.5 ~g of EeR DNA, and to 27 d when 15 txg of EcR DNA was used for transfection. In total, alter transfee- tion with 15 ~g of EcR DNA, the recovery of S clones was fivefold higher than that of As clones, and even with a 10-fold reduction in DNA input, only 26 clones were recovered after transformation with As-DNA. Moreover, the S transformants grew more quickly and had a normal appearance relative to wild-type C7-10 cells, while the As transformants had a vaeuolated appearance and grew more slowly. With continued passage, however, these differences gradually diminished, suggesting that eel1 populations derived from clones transfeeted with As-EcR were unstable, generating faster growing variants during continued maintenance in culture. These differences in the properties of the transfected cells sug- gested that disruption of normal EcR expression has a deleterious effect on C7-10 cells.

MOSQUITO EcR ISOFORMS 525
FIG. 3. Western blot of transfected cells using anti-EcR antibody. Samples contained approximately 20/xg of protein. C7-10 cells are nontransfected controls. S-3 represents a clone recovered after transfection with S-DNA, and As-3, As-2, and As-1 represent clones recovered after transfection with As-DNA. The plus (+) and minus (-) signs designate the presence or absence of a heat shock (2 h at 41 ~ C). Cells were harvested after a 1-h recovery period at 28 ~ C.
Expression of EcR isoforms in transfected cells. When expression of EcR protein in the transfected clones was examined by Western blotting, we found that the predominant 80-kDa EcR protein present in untransfected C7-10 control cells (Fig. 3, lanes 1 and 2) was also present in the S-3 cells (Fig. 3, lanes 3 and 4). As expected, from the location of the EcR insert upstream of the heat shock control elements and downstream of a conserved -150-bp regula- tory region 'X' (Karch et al., 1981), heat shock had no measurable effect on the overall level of EcR expression in S-3 cells (Fig. 3, lanes 1-4). In contrast, clones transfected with As-EcR DNA ex- pressed proportionately higher levels of the 75- and 72-kDa EcR isoforms in the absence of heat shock, and with the exception of clone As-3 (Fig. 3, lane 5) only the 80-kDa band persisted after heat shock (Fig. 3, lanes 7 and 9). We note, however, that with the present protocols these data are qualitative, and we have not un- dertaken a rigorous quantitative assessment at this time. We further note that relative to clones As-1 and As-2, clone As-3 was derived from cells that had been transfected with a relatively high amount of input DNA (Table 1), and that disruption of cell metabolism by heat shock appeared to abolish expression of EcR isoforms. To a first approximation, this differential response in clones transfected with S versus As-EcR plasmids suggested that expression of As- EcR leads to compensatory changes in those cells that can be re- covered as viable, stably transformed clones.
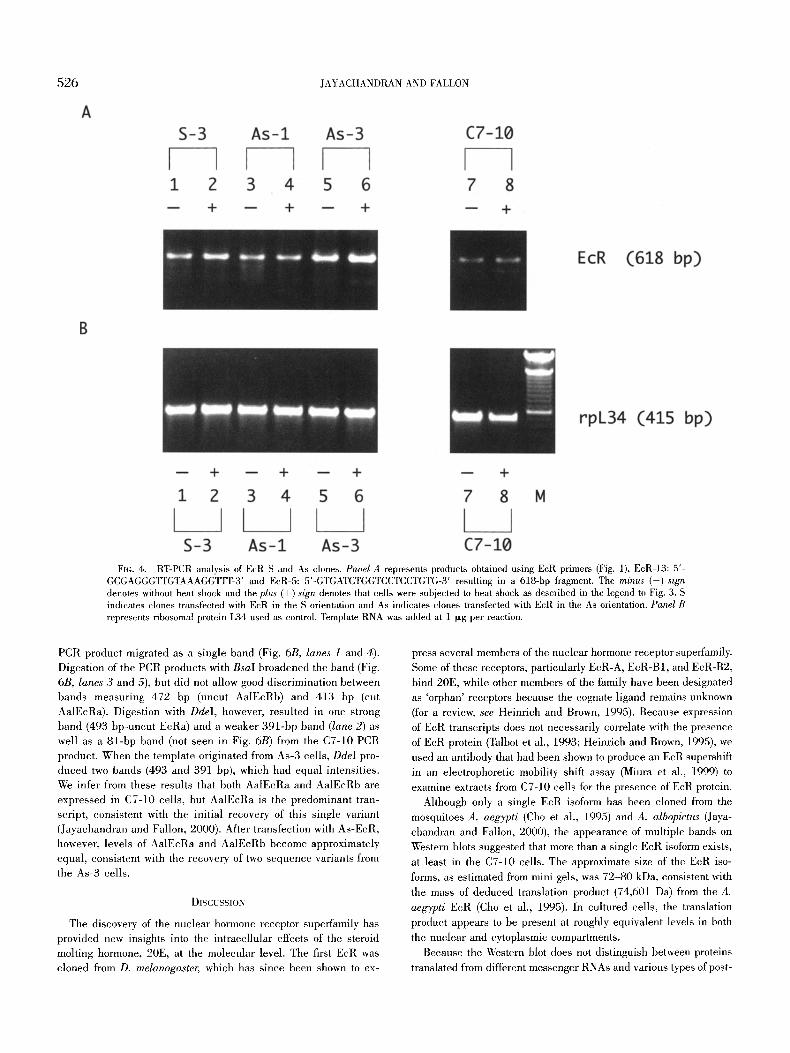
In contrast to its effects on relative proportions of EcR protein in clones containing As-EcR, disruption of cell metabolism by heat shock had no apparent effect on the overall abundance of EcR transcripts as detected by RT-PCR. This result, however, would not discriminate among potentially different transcripts of nearly iden- tical length. However, the As-3 cells in particular appeared to make more EcR transcript than As-1 and S-3 cells, which in turn pro- duced more product than C7-10 ceils (Fig. 4A, compare lanes 1-6 with lanes 7 and 8). In contrast, amounts of rpL34 PCR products were comparable in each sample (Fig. 4B). To examine the PCR products more closely, primer E c R l l (Fig. 1) was used with reverse transcriptase to make full length cDNA copies of EcR transcripts from C7-10 ceils, S-3 cells, As-l, and As-3 cells. From C7-10 and As-1 cells, amplification of the resulting cDNA using EcR12 and E c R l l primer pairs yielded only a single band. After sequence analysis, we found that with the exception of As-3 ceils, this band
was identical to the EcR isoform described previously (Jayachan- dran and Fallon, 2000). The PCR from As-3 cells, however, yielded a single band containing two distinct sequences, AalEcRa and a new EcR sequence variant called AalEcRb. These PCR results were replicated with three independent RNA preparations.
Among the first 600 amino acids, AalEcRa and AalEcRb were nearly identical (Fig. 5). Isolated nucleotide differences resulted in six amino acid substitutions, one of which was common to the EcR from A. aegypti (Cho et al., 1995). This region of virtual identity among mosquito EcRs included the DNA-binding domain (set be- tween brackets) and the hormone-binding domain (underlined). Downstream of amino acid 610, both A. albopictus EcRs had a six amino acid gap, relative to the AaEcR described by Cho et al. (1995). Slightly fm~ther downstream was another region of variability in an amino acid stretch characterized by an abundance of gluta- mine, serine, and glycine. In this region, all three mosquito EcRs showed differences, particularly among the final 20-30 amino acids. We note that the abundance of serines in this portion of the se- quence provides potential sites for posttranslational modification by phosphorylation.
C7-10 cells express EcRa and EcRb. Because we recovered only AalEcRa from C7-10 cells, it was of interest to learn whether only AalEcRa was expressed by these cells. We used RT-PCR followed by restriction endonuclease digestion of PCR products to differ- entiate between AalEcRa and AalEcRb transcripts. Although the span of 21 nt corresponding to the QQQPPHP deletion in Aal- EcRb lacked sites that could readily distinguish full length PCR products, we noted that the nucleotide sequence corresponding to the isoform AalEcRa has a BsaI site (nt 1997, where +1 corrres- ponds to the A of the ATG start site) absent from AalEcRb. Like- wise, at nt 1976, AalEcRb has a DdeI site absent from AalEcRa. When PCR products were cut with these enzymes, diagnostic frag- ments predicted on the basis of nucleotide sequence are shown in Fig. 6A, while Fig. 6B shows corresponding experimental results. First, we used primers common to both isoforms to produce from C7-10 and As-3 cells, separately, relatively short ( - 5 0 0 bp) PCR products. The template for this reaction was generated by RT- PCR, using EcR-11, and the subsequent PCR reaction used prim- ers EcR-2 and EcR-11 (Fig. 1). The PCR products were digested with BsaI and DdeI. In both C7-10 and As-3 cells, the undigested
526 JAYACHANDRAN AND FALLON
FIG. 4. RT-PCR analysis of EcR S and As clones. Panel A represents products obtained using EcR primers (Fig. 1), EcR-13: 5'- GCGAGGGTYGTAAAGGTTT-3' and EcR-5: 5'-GTGATCTGGTCCTCCTGTG-3' resulting in a 618-bp fragment. The minus (-) sign denotes without heat shock and the plus (+) sign denotes that cells were subjected to heat shock as described in the legend to Fig. 3. S indicates clones transfected with EcR in the S orientation and As indicates clones transfected with EcR in the As orientation. Panel B represents ribosomal protein L34 used as control. Template RNA was added at 1 I-~g per reaction.
PCR product migrated as a single band (Fig. 6B, lanes 1 and 4). Digestion of the PCR products with BsaI broadened the band (Fig. 6B, lanes 3 and 5), but did not allow good discrimination between bands measuring 472 bp (uncut AalEcRb) and 413 bp (cut AalEcRa). Digestion with DdeI, however, resulted in one strong band (493 bp-uncut EcRa) and a weaker 391-bp band (lane 2) as well as a 81-bp band (not seen in Fig. 6B) from the C7-10 PCR product. When the template originated from As-3 ceils, DdeI pro- duced two bands (493 and 391 bp), which had equal intensities. We infer from these results that both AalEcRa and AalEcRb are expressed in C7-10 cells, but AalEcRa is the predominant tran- script, consistent with the initial recovery of this single variant (Jayachandran and Fallon, 2000). After transfection with As-EcR, however, levels of AalEcRa and AalEcRb become approximately equal, consistent with the recovery of two sequence variants from the As-3 cells.
DISCUSSION
The discovery of the nuclear hormone receptor supeffamily has provided new insights into the intracellular effects of the steroid molting hormone, 20E, at the molecular level. The first EcR was cloned from D. melanogaster, which has since been shown to ex-
press several members of the nuclear hormone receptor superfamily. Some of these receptors, particularly EcR-A, EcR-B1, and EcR-B2, bind 20E, while other members of the family have been designated as 'orphan' receptors because the cognate ligand remains unknown (for a review, see Heinrich and Brown, 1995). Because expression of EcR transcripts does not necessarily correlate with the presence of EcR protein (Talbot et al., 1993; Heiurich and Brown, 1995), we used an antibody that had been shown to produce an EcR supershift in an electrophoretic mobility shift assay (Miura et al., 1999) to examine extracts from C7-10 cells for the presence of EcR protein.
Although only a single EcR isoform has been cloned from the mosquitoes A. aegypti (Cho et al., 1995) and A. albopictus (Jaya- chandran and Fallon, 2000), the appearance of multiple bands on Western blots suggested that more than a single EcR isoform exists, at least in the C7-10 cells. The approximate size of the EcR iso- forms, as estimated from mini gels, was 72-80 kDa, consistent with the mass of deduced translation product (74,601 Da) from the A. aegypti EcR (Cho et al., 1995). In cultured cells, the translation product appears to be present at roughly equivalent levels in both the nuclear and cytoplasmic compartments.
Because the Western blot does not distinguish between proteins translated from different messenger RNAs and various types of post-

MOSQUITO EcR ISOFORMS 527
AaIEcRo AalEcRb
AaEcR
AalEcRa AalEcRb
AaEcR
1 50 MMKRRWSNNG GFTALRMLDD SSSEVTSSSA ALGMTMSPNS LGSPNYDELE
51 100 LWSSYEDNAY NGHSVLSNGN NNLGGCGAAN NLLMNGIVGN NNLNG~@4NMA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . F . . . . . . . . . . . .
AalEcRo AalEcRb
AaEcR
AaIEcRa AalEcRb
AaEcR
AalEcRa AalEcRb
AaEcR
AalEcRa AalEcRb
AaEcR
AalEcRa AalEcRb
AaEcR
AalEcRa AalEcRb
AaEcR
AalEcRa AalEcRb
AaEcR
101 150 SQAVQANANS IQHIVGNLIN GVNPNQTLIP PLPSIIQNTL MNTPRSESVN G . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
151 " I - - 200 SISSGREDLS PSSSLNGYTD GSDAKKQKKG PTPRQQEEL~LVCGDRASGY
1211121222 2221211122 2222222112 122121122 121222 221 201 250 HYNALTCEGC KGFFRRSVTK NAVYCCKFGH ACEMDMYMRR KCQECRLKKC . . . . . . . . . . . . . . . . . . . . . . . . . . . . . P . . . . . . . . . . . . . . . . . . . .
301 350 PILMKCDPPP HQAIPLLPEK LLQENRLRNI PLLTAN(~AV IYKLIWYQDG
351 400 YE~PSEEDLK RINIGSPNEE ED(~HDVHFRH ITEITILTV~ LIVEFAKGLP
. . . . . . . . . . . . . . . . . , . . . . . . . . . . . . . . . . . ~ . . . . . . . . . . , , . .
401 450 AFTKIP~EDQ ITLLKACSSE VI~I4LP, MARRY DAATDSILFA NNRSYTRDSY
. . . . . . . . . . . . . . . . . . . . . . . . . . . . o . . . . . . . . . . . . . . . . . . . . .
451 500 AaIEcRa RMAGMADTIE DLLHFCRQMF SLTVDNVEYA LLTAIVIFSD RPGLEqAELV AalEcRb
AaEcR
AalEcRa 501 55@ EHIQSYYIDT LRIYILNRHA GDPKCSVlFA KLLSILTELR TLGNQNSEMC
AoIEcRb AaEcR
AoIEcRo AoIEcRb
AoEcR
AaIEcRa AoIEcRb
AaEcR
AaIEcRa AaIEcRb
AaEcR
F I G . 5 .
. . . . . . . . . . . . . . . . �9 . . . . . . . . ~ . . . . . . ~ 1 7 6 1 7 6 . . . . , . ~ . . . . .
0 . . . . . . . . . ~ . . . . . . . . . . , ~ 1 7 6 1 7 6 1 7 6 1 7 6 1 7 6 o . . . . . . . . . . . . . . . . . . .
551 6 ~ FSLKLKNRKL PRFLEEIWDV QDIPPSMQAQ MHSHGTPQSS SSSSSSSSSS , . . . . . . . . . . . . . . . . . . . . . . . . . . . , , , . . , , , . , . . . . . . . . . . . .
, . , . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
601 650 SSNGSSNGNS NSN . . . . . . G PHPHPHGQQL TPNQQQPPHP QQQQHSQLQQ
. . . . . . . . . . S..SNSSQH . . . . . . . . . . . . . . . . . - - - - G1111111
651 690 VHANGSGSGG ASSNNSSSSG GLGVPVGL-- GGGALDHV* . . . . . . . . . . . . . . . . . . . . . . S . . . . . GG . . ,V . .q . * . . . . . . . . . . -G...N . . . . . V- . . - . . . . .L.M.,Q.*
Amino acid sequence alignment of mosquito EcRs. AalEcRa (re- named 'a' from that previously published [Jayachandran and Fallon, 2000]) and AalEcRb were obtained from the cell line C7-10 ofA. albopictus (Aal). AaEcR is from A. aegypti (Cho et al., 1995). The DNA-binding domain is marked with square brackets. The hormone-binding domain is underlined. Dots indicate identity to AatEcRa; letters designate amino acid dift~rences, and the asterisk represents the stop codon.
translational modification of a single EcR protein, we infer from these results that at least one EcR gene is both transcribed and translated in C7-10 cells. The observation that transfected cells exhibit changes in relative abundance of EcR isoforms on Western blots is consistent with the expectation that regulation of EcR ex- pression, even at the level of a single cell type, may be complex. In addition, the slow growth and relatively poor recovel~r of cells transfected with As-EcR suggests that As expression in some way interfered with the "normal" gene expression.
The typical nuclear hormone receptor is organized into a series of functional motifs designated, from the amino to the carboxyl ter- mini, as A/B, C, D, E, and F (Fig. 1). Among insect EcR proteins, Domains C and E, which are responsible for DNA binding and ligand binding, respectively, are highly conserved. Domain A/B, however, mediates contact with other transcription factors, and in those cases in which a particular insect species has been shown to express more than a single EcR isoform such as D. melanogaster (Talbot et al., 1993), the moths Bombyx mori (Swevers et al., 1995; Kamimura et al., 1997), Manduca sexta (Jindra et al., 1996), and Choristoneura fumiferana (Perera et al., 1999), and the beetle Te- nebrio molitor (Mouillet et al., 1997) the variability between iso- forms occurs in the A/B domain. Between insect species, consid- erable variation has been noted in C-terminal domain F of the var- ious EcRs that have been described. For example, the 106 amino acid Domain F of A. aegypti has only 28% similarity to a corre- sponding 227 amino acid domain F of Drosophila EcRs. Among themselves, however, the Drosophila isoforms EcR-A, EcR-B1, and EcR-B2 are similar. In an alignment of six EcRs from different species, Mouillet et al. (]997) show that region F is longer in D. melanogaster and A. aegypti, relative to M. sexta, B. mori, and C. fumiJ~rana. In Chironomus tentans, Domain F contains only 24 ami- no acids (Imhof et al., 1993), and in T. molitor, only five amino acids (Mouillet et al., 1997).
In the present study, the differences between AalEcRa and AalEcRb are substantial only at the extreme C-terminal domain, in a region that is rich in glutamine, serine, and glycine. The nucle- otide differences between the EcR isoforms delete a stretch of seven additional amino acids from the C-terminus of AalEcRb, relative to AalEcRa. This short motif is meaningful in the sense that it is present in several transcription factors, including Drosophila sev- enless protein (Michael et al., 1990), human AP-1 (Hattori et al., 1988), Drosophila homeotic distal-less protein (Vachon et ah, 1992), human nfineralocorticoid receptor (Arriza et al., 1987), and human sterol regulatory element binding protein-1 (Yokoyama et ah, 1993).
Because Domain F is so variable among species, it is not sur- prising that its function is poorly understood (Heinrich and Brown, 1995). This consideration led us to identify restriction enzyme sites that would allow us to distinguish between the two transcripts using a PCR-based approach. The predicted presence of a unique BsaI site in a - 5 0 0 - b p PCR product from AalEcRa, and a unique DdeI site in AalEcRb, allowed us to conclude that both transcripts are expressed by C7-10 cells, and by transformants containing the As- EcR construct. However, AalEcRa seems to predominate in C7-10 cells, while the two isoforms appeared to be represented equally in As-3 cells. We note, however, that this analysis was done several wk after the recovery of the As-3 clone, and it remains to be learned whether this ratio changes as the stably transformed cells overcome the deleterious effect of transformation with As-EcR.
Our interest in mosquito EcRs evolved from early efforts to ex-

528 JAYACHANDRAN AND FALLON
FIG. 6. Expression of AalEcRa and AalEcRb transcripts. A. Schematic represen- tation of the experimental design to examine the relative expression of EcR isoforms in C7- 10 and transfected cells. The recognition sites of the different enzymes are denoted by ar- rows, and sizes of expected products are shown. Values indicate nucleotide positions, relative to the A of translational start site as +1. B. Restriction endonuclease analysis of EcR isoforms obtained using primers EcR-2 and EcR-11 (Fig. 1). Lanes ] and 4 are uncut (U) PCR products from C7-10 and As-3 ceils, respectively. Lanes labeled D contain PCR products digested with DdeI and lanes labeled B contain PCR products digested with BsaI. Note that DdeI does not cut AalEcRa, and BsaI does not cut AalEcRb. The -80-bp product does not show in the figure.
A AalEcRa
1584
AalEcRb
(1997) B
I 2O76
2055 L
D (1976)
J J
J J
J
Ddel (D)
493 nt
472 nt
Bsal
(B)
AalEcRa [ I
AalEcRb
493
m
391
Im
81
AalEcRa
AalEcRb
I I
413
472
[]
8O

MOSQUITO EcR ISOFORMS 529
press 20E-inducible DNA constructs in transfected cells. Well-
characterized ecdysone response elements (EcREs) have been mapped upstream of the A. aegypti vitellogenin gene (Romans et al., 1995) but we have not succeeded in showing 20E-inducibil i ty of the vitellogenin promoter in transfected C7-10 cells. We reasoned
that failure to demonstrate a 20E response in transfected cells might result from the absence of EcR in the cell line, or the need for
specialized transcription factors that may be specific to fat body. Because introduction of constructs with a synthetic 23-bp EcRE
upstream of a reporter gene was also unsuccessful , we tested for the expression of EcR and ultraspiracle protein homologs in these cells. As was the case in the present study, the mere introduction
of DNA containing synthetic EcREs interfered with the recovel 7 of stably transformed clones in a manner that was dependent on DNA concentration (Jayachandran and Fallon, pers. comm.). Thus, al- though the mechanisms remain unclear, perturbation of 20E metab-
olism in cultured cells appears to have significant physiological
effects that may eventually provide an insight into the role of 20E in the intact organism.
ACKNOWLEDGMENTS
This work was supported by grant AI 43971 from the National Institutes of Health, Bethesda, Maryland, and by the University of Minnesota Experi- ment Station, St. Paul, Minnesota.
REFERENCES
An'iza, J. L.; Weinberger, C.; Cerelli, G.; Glaser, T. M.; Handelin, B. L.; Housman, D. E.; Evans, R. M. Cloning of human mineralocorticoid receptor complementary DNA: structural and functional kinship with the glucocorticoid receptor. Science 237:268-275; 1987.
Cho, W. L.; Kapitskaya, M. Z.; Raikhel, A. S. Mosquito ecdysteroid receptor: analysis of the cDNA and expression during vitellogenesis. Insect Biochem. Mol. Biol. 25:19-27; 1995.
Di Nocera, P. P.; Dawid, I. B. Transient expression of genes introduced into cuhured cells of Drosophila. Proc. Natl. Acad. Sci. USA 80:7095-7098; 1983.
Dulbecco, R.; Vogt, M. Plaque formation and isolation of pure lines of polio- myletis virus. J. Exp. Med. 99:167-182; 1954.
Fallon, A. M.; Stollar, V. The biochemistry and genetics of mosquito cells in culture. In: Maramomsch, K., ed. Advances in cell culture. Vol. 5. New York: Academic Press; 1987:97-137.
Gerenday, A.; Fallon, A. M. Cell cycle parameters in Aedes albopictus mos- quito cells. In Vitro Cell. Dev. Biol. 32A:307-312; 1996.
Hattori, K.; Angel, P.; Le Beau, M. M.; Karin, M. Structure and chromosomal localization of the functional intronless human JUN protooncogene. Proc. Natl. Acad. Sci. USA 85:9148-9152; 1988.
Heinrich, V. C.; Brown, N. E. Insect nuclear receptors: a developmental and comparative perspective. Insect Biochem. Mol. Biol. 25:881-897; 1995.
hnhof, M. O.; Rusconi, S.; Lezzi, M. Cloning of a Chironomus tentans cDNA encoding a protein (cEcRH) homologous to the Drosophila melano- gaster ecdysteroid receptor (dEcR). Insect Biochem. Mol. Biol. 23:115-124; 1993.
Jayachandran, G.; Fallon, A. M. Evidence for expression of EcR and USP components of the 20-hydroxyecdysone receptor by a mosquito cell line. Arch. Insect Biochem. Physiol. 43:87-96; 2000.
Jindra, M.; Malone, E; Hiruma, K.; Riddiford, L. M. Developmental profiles and ecdysteroid regulation of the mRNAs for two ecdysone receptor isoforms in the epidermis and wings of the tobacco horuworm, Man- duca sexta. Dev. Biol. 180:258-272; 1996.
Kamimura, M.; Tomita, S.; Kiuchi, M.; Fujiwara, H. Tissue-specific and stage- specific expression of two silkworm ecdysone receptor isofm~ns--ec- dysteroid-dependent transcription in cultured anterior silk glands. Eur. J. Biochem. 248:786-793; 1997.
Karch, E; Torok, I.; Tissieres, A. Extensive regions of homology in front of the two hsp70 heat shock variant genes in Drosophila melanogaster. J. Mol. Biol. 148:219-230; 1981.
Laemmli, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-885; 1970.
Lan, Q.; Gerenday, A.; Fallon, A. M. Cultured Aedes albopictus mosquito cells synthesize hormone-inducible proteins. In Vitro Cell. Dev. Biol. 29A:813-818; 1993.
McGaiTy, T. J.; Lindquist, S. Inhibition of heat shock protein synthesis by heat-inducible antisense RNA. Proc. Natl. Acad. Sci. USA 83:399-403; 1986.
Michael, W. M.; Bowtell, D. D.; Rubin, G. M. Comparison of the sevenless genes of Drosophila virilis and Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 87:5351-5353; 1990.
Miura, K.; Wang, S. E; Raikhel, A. S. Two distinct subpopulations of ecdy- sone receptor complex in the female mosquito during vitellogenesis. Mol. Cell. Endocrinol. 156:111-120; 1999.
Mouillet, J. E; Delbecque, J. P.; Quennedy, B.; Delachambre, J. Cloning of two putative ecdysteroid receptor isoforms from Tenebrio molitor and their developmental expression in the epidermis during metamorpho- sis. Eur. J. Biochem. 248:856-863; 1997.
Perera, S. C.; Ladd, T. R.; Dhadialla, T. S.; Krell, P. J.; Sohi, S. S.; Retnak- aran, A.; Palli, S. R. Studies on two ecdysone receptor isoforms of the spruce budwman, Choristoneura fumiferana. Mol. Cell. Endocri- nol. 152:73-84; 1999.
Romans, P.; Tu, Z.; Ke, P.; Hagedorn, H. H. Analysis of a vitellogenin gene of the mosquito, Aedes aegypti and comparisons to vitellogenins from other organisms. Insect Biochem. Mol. Biol. 25:939-958; 1995.
Rosenberg, U. B.; Preiss, A.; Seifert, E.; Jackle, H.; Knipple, D. C. Produc- tion of phenocopies by Krtippel antisense RNA injection into Dro- sophila embryos. Nature 313:703-706; 1985.
Shotkoski, E A.; Fallon, A. M. The mosquito dihydrofolate reductase gene functions as a dominant selectable marker in transfected cells. Insect Biochem. Mol. Biol. 23:883-893; 1993.
Shotkoski, A. E; Fallon, A. M. Expression of an antisense dihydrofolate re- ductase transcript in transfected mosquito ceils: effects on growth and plating efficiency. Am. J. Trop. Med. Hyg. 50:433439; 1994.
Swevers, L.; Drevet, J. R.; Lunke, M. D.; Iatrou, K. The silkmoth homolog of the Drosophila ecdysone receptor (B1 isoform): cloning and anal- ysis of expression during follicular cell differentiation. Insect Bioch- era. Mol. Biol. 25:857-866; 1995.
Talbot, W. S.; Swyryd, E. A.; Hogness, D. S. Drosophila tissues with different metamorphic responses to ecdysone express different ecdysone re- ceptor isoforms. Cell 73:1323-1337; 1993.
Vachon, G.; Cohen, B.; Pfeifle, C.; McGuffin, M. E.; Botas, J.; Cohen, S. M. Homeotic genes of the Bithorax complex repress limb development in the abdomen of the Drosophila embryo through the target gene distal-less. Cell 71:437-450; 1992.
Wang, S. E; Miuru, K.; Miksicek, R. J.; Segraves, W. A.; Raikhel, A. S. DNA binding and transactivation characteristics of the mosquito ecdysone receptor-ultraspiracle complex. J. Biol. Chem. 273:27,531-27,540; 1998.
Wu, C.; Bingham, P. M.; Livak, K. J.; Holmgren, R.; Elgin, S. C. R. The chromatin structure of specific genes: evidence for higher order do- mains of defined DNA sequence. Cell 16:797-806; 1979.
Yokoyama, C.; Wang, X.; Briggs, M. R.; Admon, A.; Wu, J.; Hua, X.; Gold- stein, J. L.; Brown, M. S. SREBP-1, a basic helix-loop-helix-helix- leucine zipper protein that controls transcription of the low density lipoprotein receptor gene. Cell 75:187-197; 1993.




















