
Antioxidant defense system induced by a methanol extract of Caesalpinia bonducella in rat liver. ...
9
Antioxidant Defense System Induced by a Methanol Extract of Caesalpinia bonducella in Rat Liver M. Gupta, U.K. Mazumder, R. Sambath Kumar, Thangavel Sivakumar, P. Gomathi, and Y. Rajeshwar Department of Pharmaceutical Technology, Jadavpur University, Kolkata, India Abstract The antioxidant defense system dramatically controls hepa- tocellular carcinoma induced by N-nitrosodiethylamine (NDEA). In order to assess the anticarcinogenic activity of a methanol extract of Caesalpinia bonducella (MECB) leaves containing flavonoids and triterpenoids, the antiox- idant defense system has been evaluated. The effect of MECB on lipid peroxidation end-product malondialde- hyde (MDA), enzymatic antioxidants such as superoxide dismutase (SOD) and catalase (CAT), and nonenzymatic antioxidants glutathione (GSH), vitamin E, and vitamin C levels were analyzed in the liver of control and experi- mental animals. Serum was also analyzed for transaminase activity (SGOT and SGPT), alkaline phosphatase (SALP), bilirubin, total protein, and uric acid. Depletion of all these antioxidants was recorded in cancer conditions. These del- eterious effects are controlled by the administration of MECB. After drug administration, there was a marked increase in antioxidant levels and a dramatic decrease in lipid peroxidation levels. MECB also produced a protective effect by decreasing the activity of serum enzymes, biliru- bin, and increased the protein and uric acid levels. From the above results, it can be concluded that the observed anticarcinogenic properties of MECB may also be explained by its strong antioxidant capacity and capability to induce an in vivo antioxidant system. Keywords: Antioxidants, Caesalpinia bonducella, hepato- cellular carcinoma, lipid peroxidation, N-nitrosodiethy- lamine, serum enzymes. Introduction Hepatocarcinogenesis is a multistep process (Farber, 1987; Farber & Sarma, 1987). N-Nitrosodiethylamine (NDEA), a chemical carcinogen, is widely used to initiate hepatocarcinogenesis in rats. Hepatocarcinogenesis can also be induced by phenobarbital, carbon tetrachloride, dichlorodiphenyl trichloroethane, partial hepatectomy (PH), and so on (Farber, 1996). Hepatocellular carci- noma is one of the most common cancers and one of the most lethal (Schaff et al., 1993). Chemoprevention involving the use of synthetic or natural products to inhibit or reverse the carcinogenic process is an effective approach to control cancer (Metlin, 1997). It is well- known that N-nitroso compounds act as strong carcino- gens in various mammals including primates (Leoppky & Li, 1991). Foodstuffs such as milk and meat products, salted fish, alcoholic beverages, and a few varieties of vegetables are the principal sources of nitroso compounds (Tricker et al., 1991). NDEA has been suggested to cause oxidative stress and cellular injury due to the enhanced formation of free radicals. NDEA has been shown to be metabolized to its active ethyl radical metabolite, and reactive products interact with DNA causing mutation that leads to carcinogenesis (Anis et al., 2001). Experi- mental, clinical, and epidemiological studies have pro- vided evidence supporting the role of reactive oxygen species in the etiology of cancer (Ray et al., 2000). Several biochemical markers have been suggested for biomonitoring the action of anticancer agents. Esti- mation of circulatory lipid peroxidation end-product malondialdehyde (MDA), enzymatic antioxidant super- oxide dismutase (SOD) and catalase (CAT), and nonenzymatic antioxidants such as glutathione (GSH), vitamin E, and vitamin C is a reliable method for scree- ning the action of chemopreventive agents. In recent years, there has been considerable emphasis on the identification of plant products as possible anti- carcinogens with antioxidant properties. Caesalpinia Accepted: March 1, 2005 Address correspondence to: M. Gupta, Department of Pharmaceutical Technology, Jadavpur University, Kolkata 700 032, India. Tel.: 91-33-24404123; Fax: 91-33-24146967; E-mail: [email protected] DOI: 10.1080/13880200590963439 # 2005 Taylor & Francis Ltd. Pharmaceutical Biology 2005, Vol. 43, No. 5, pp. 411–419
-
Upload
srcpwarangal -
Category
Documents
-
view
2 -
download
0
Transcript of Antioxidant defense system induced by a methanol extract of Caesalpinia bonducella in rat liver. ...

Antioxidant Defense System Induced by a Methanol Extract of
Caesalpinia bonducella in Rat Liver
M. Gupta, U.K. Mazumder, R. Sambath Kumar, Thangavel Sivakumar, P. Gomathi, and Y. Rajeshwar
Department of Pharmaceutical Technology, Jadavpur University, Kolkata, India
Abstract
The antioxidant defense system dramatically controls hepa-tocellular carcinoma induced by N-nitrosodiethylamine(NDEA). In order to assess the anticarcinogenic activityof a methanol extract of Caesalpinia bonducella (MECB)leaves containing flavonoids and triterpenoids, the antiox-idant defense system has been evaluated. The effect ofMECB on lipid peroxidation end-product malondialde-hyde (MDA), enzymatic antioxidants such as superoxidedismutase (SOD) and catalase (CAT), and nonenzymaticantioxidants glutathione (GSH), vitamin E, and vitaminC levels were analyzed in the liver of control and experi-mental animals. Serum was also analyzed for transaminaseactivity (SGOT and SGPT), alkaline phosphatase (SALP),bilirubin, total protein, and uric acid. Depletion of all theseantioxidants was recorded in cancer conditions. These del-eterious effects are controlled by the administration ofMECB. After drug administration, there was a markedincrease in antioxidant levels and a dramatic decrease inlipid peroxidation levels. MECB also produced a protectiveeffect by decreasing the activity of serum enzymes, biliru-bin, and increased the protein and uric acid levels. Fromthe above results, it can be concluded that the observedanticarcinogenic properties of MECB may also beexplained by its strong antioxidant capacity and capabilityto induce an in vivo antioxidant system.
Keywords: Antioxidants, Caesalpinia bonducella, hepato-cellular carcinoma, lipid peroxidation, N-nitrosodiethy-lamine, serum enzymes.
Introduction
Hepatocarcinogenesis is a multistep process (Farber,1987; Farber & Sarma, 1987). N-Nitrosodiethylamine
(NDEA), a chemical carcinogen, is widely used to initiatehepatocarcinogenesis in rats. Hepatocarcinogenesis canalso be induced by phenobarbital, carbon tetrachloride,dichlorodiphenyl trichloroethane, partial hepatectomy(PH), and so on (Farber, 1996). Hepatocellular carci-noma is one of the most common cancers and one ofthe most lethal (Schaff et al., 1993). Chemopreventioninvolving the use of synthetic or natural products toinhibit or reverse the carcinogenic process is an effectiveapproach to control cancer (Metlin, 1997). It is well-known that N-nitroso compounds act as strong carcino-gens in various mammals including primates (Leoppky& Li, 1991). Foodstuffs such as milk and meat products,salted fish, alcoholic beverages, and a few varieties ofvegetables are the principal sources of nitroso compounds(Tricker et al., 1991). NDEA has been suggested to causeoxidative stress and cellular injury due to the enhancedformation of free radicals. NDEA has been shown to bemetabolized to its active ethyl radical metabolite, andreactive products interact with DNA causing mutationthat leads to carcinogenesis (Anis et al., 2001). Experi-mental, clinical, and epidemiological studies have pro-vided evidence supporting the role of reactive oxygenspecies in the etiology of cancer (Ray et al., 2000).
Several biochemical markers have been suggested forbiomonitoring the action of anticancer agents. Esti-mation of circulatory lipid peroxidation end-productmalondialdehyde (MDA), enzymatic antioxidant super-oxide dismutase (SOD) and catalase (CAT), andnonenzymatic antioxidants such as glutathione (GSH),vitamin E, and vitamin C is a reliable method for scree-ning the action of chemopreventive agents.
In recent years, there has been considerable emphasison the identification of plant products as possible anti-carcinogens with antioxidant properties. Caesalpinia
Accepted: March 1, 2005
Address correspondence to: M. Gupta, Department of Pharmaceutical Technology, Jadavpur University, Kolkata 700 032, India. Tel.:91-33-24404123; Fax: 91-33-24146967; E-mail: [email protected]
DOI: 10.1080/13880200590963439 # 2005 Taylor & Francis Ltd.
Pharmaceutical Biology2005, Vol. 43, No. 5, pp. 411–419

bonducella (L.) Flem. (Caesalpiniaceae) [fever nut; bun-duc nut; commonly known as nata karanja (Hindi)], aprickly shrub, is found throughout the hotter parts ofIndia, Myanmar, and Sri Lanka. The leaves of this plantare traditionally used for the treatment of liver disorders,inflammation, and tumors (Kirtikar & Basu, 1975). Ithas also been reported to possess multiple therapeuticproperties such as antipyretic, antidiuretic, anthelmintic,antibacterial (Neogi & Nayak, 1958) anticonvulsant(Adesina, 1982), anti-anaphylactic and antidiarrheal,antiviral (Dhar et al., 1968), antiasthmatic (Gayarajaet al., 1978), and antiamoebic and antiestrogenic activi-ties (Raghunathan et al., 1982). Currently, the hepato-protective and antioxidant role of paracetamol-inducedliver damage in rats (Gupta et al., 2003a) and anti-inflammatory, analgesic, and antipyretic activities(Gupta et al., 2003b) of Caesalpinia bonducella have beenreported from our laboratory. Also, antitumor activityand antioxidant status of Caesalpinia bonducella againstEhrlich ascites carcinoma in Swiss albino mice werereported from our laboratory (Gupta et al., 2004).
The current study deals with the antioxidant defensesystem induced by the methanol extract of Caesalpiniabonducella (MECB) in NDEA-induced oxidative damagein the liver of Wistar albino rats.
Materials and Methods
Plant material and extraction
The plant Caesalpinia bonducella Flem. (Caesalpiniaceae)was collected from the Kolli Hills of Tamilnadu, India.The plant material was taxonomically identified by theBotanical Survey of India, Kolkata. A voucher specimen(no. GMS-2) has been preserved in our laboratory forfuture reference. The leaves were shade-dried withoccasional shifting and then powdered with a mechanicalgrinder and stored in an air-tight container. The driedpowder material of the leaf (500 g) was defatted with pet-roleum ether (60–80�C) in a Soxhlet apparatus. Thedefatted powder material thus obtained was furtherextracted with methanol for 72 h in the Soxhlet. The sol-vent was removed by distillation under suction, and theresulting semisolid mass was vacuum-dried using rotaryflash evaporator to yield a solid residue (8.78%w=w).Phytochemical screening of the extract revealed thepresence of alkaloids, saponins, flavonoids, triterpenes,tannins, and steroids.
Animals
Studies were carried out using Wistar albino rats(150–180 g) of either sex. They were obtained from theanimal house, Indian Institute of Chemical Biology(IICB), Kolkata, India. The animals were grouped andhoused in polyacrylic cages (38� 23� 10 cm) with not
more than six animals per cage and maintained understandard laboratory conditions (temperature 25� 2�C)with dark and light cycle (14=10 h). They were allowedfree access to standard dry pellet diet (Hindustan Lever,Kolkata, India) and water ad libitum. The rats were accli-matized to laboratory conditions for 10 days before com-mencement of the experiment. All procedures describedwere reviewed and approved by the university animalethical committee.
Chemicals
N-Nitrosodiethylamine (NDEA), 1-chloro-2,4-dinitro-benzene (CDNB), bovine serumalbumin (SigmaChemicalCo., St. Louis, MO, USA), thiobarbituric acid, nitrobluetetrazolium chloride (NBT) (Loba Chemie, Bombay,India), 5,50-dithio bis-2-nitrobenzoic acid (DTNB) (SISCOResearch Laboratory, Bombay) were used in the study.All the reagents used were of analytical reagent grade.
Experimental design
Male Wistar albino rats were divided into 5 groups(n ¼ 6). All the groups received a single intraperitonealinjection of NDEA (200mg=kg, b.w.) followed by carbontetrachloride (3ml=kg, b.w.), once a week, subcutaneousinjection for 8 weeks (Sundaresan & Subramanian, 2003),except group 1 (control group), group 2 (MECB200mg=kg b.w. treated), and group 3 NDEA (200mg=kg,kg, b.w) (NDEA control group). From the first day,normal saline, 5ml=kg, and propylene glycol, 5ml=kg,were administered to control and NDEA control groups,respectively, for 8 weeks on alternate days, orally. MECBat different doses (100 and 200mg=kg. b.w.) was adminis-tered on alternate days orally to groups 4 and 5, respect-ively, for 8 weeks. After administration of the last dosefollowed by 18 h fasting, 6 rats from each group weresacrificed for the study of anticarcinogenic activity byestimation of serum and liver biochemical parameters.
Biochemical studies
Blood was obtained from all animals by puncturingretro-orbital plexus. The blood samples were allowed toclot for 45min at room temperature. Serum was separatedby centrifugation at 2500 rpm at 30�C for 15min and usedfor the estimation of various biochemical parameters,namely, serum glutamyl pyruvate transaminase (SGPT),serum glutamyl oxalacetic acid transaminase (SGOT)(Bergmeyer et al., 1978), serum alkaline phosphatase(SALP) (King, 1965), and serum bilirubin by the methodof Malloy and Evelyn (1937), and protein content wasmeasured by the method of Lowry et al. (1951). The plasmauric acid was estimated by the method of Caraway (1963).
After collection of blood samples, the rats were sacri-ficed and the livers were excised, rinsed in ice-cold normal
412 M. Gupta et al.
saline followed by 0.15 M Tris-HCl (pH 7.4), blotted dryand weighed. A 10% w=v of homogenate was preparedin 0.15M Tris-HCl buffer and processed for the estimationof lipid peroxidation by the method of Ohkawa et al.(1979). A part of the homogenate, after precipitating pro-teins with trichloroacetic acid (TCA), was used for esti-mation of glutathione by the method of Ellman (1959).Vitamin E, by Quaife and Dju (1948) with slight modifi-cation by Baker and Frank (1951), and vitamin C, byOmaye et al. (1979), were also estimated. The rest of thehomogenate was centrifuged at 15,000 rpm for 15min at4�C. The supernatant thus obtained was used for the esti-mation of SOD by the method of Kakkar et al. (1984),and CAT activity was measured by the method of Aebi(1974).
Statistical analysis
The experimental results were expressed as mean� SEM.Data were assessed by ANOVA followed by Student’sunpaired t-test. p values of <0.05 were considered asstatistically significant.
Results
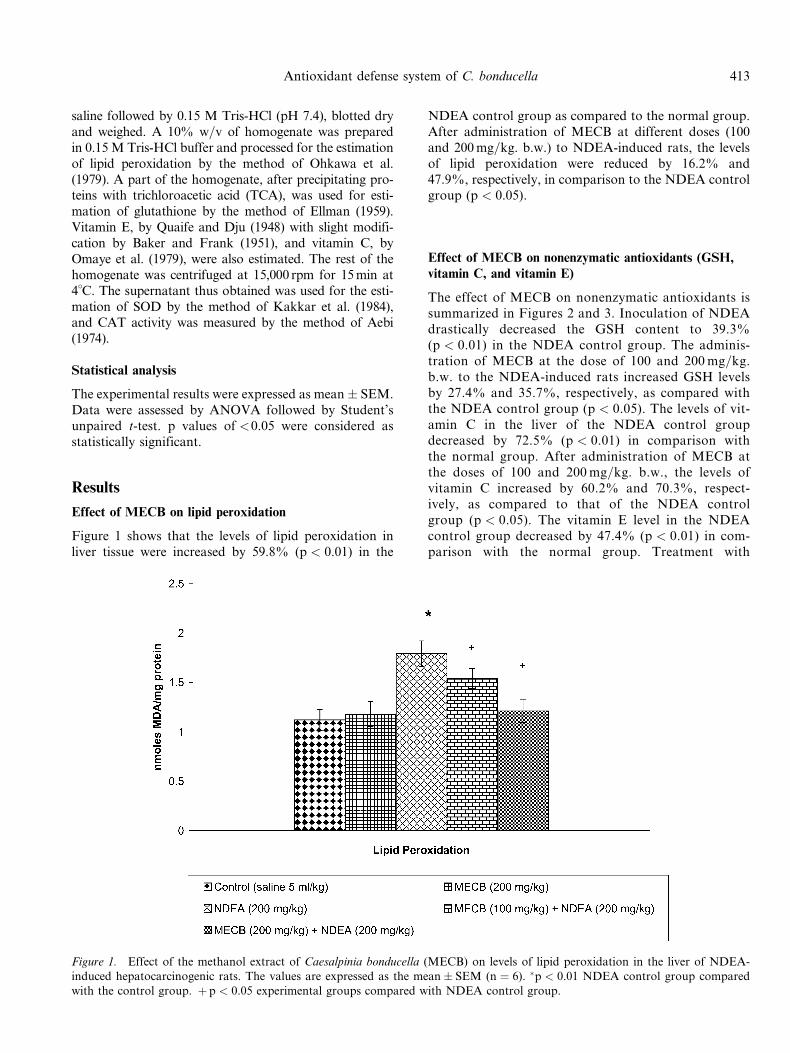
Effect of MECB on lipid peroxidation
Figure 1 shows that the levels of lipid peroxidation inliver tissue were increased by 59.8% (p < 0.01) in the
NDEA control group as compared to the normal group.After administration of MECB at different doses (100and 200mg=kg. b.w.) to NDEA-induced rats, the levelsof lipid peroxidation were reduced by 16.2% and47.9%, respectively, in comparison to the NDEA controlgroup (p < 0.05).
Effect of MECB on nonenzymatic antioxidants (GSH,
vitamin C, and vitamin E)
The effect of MECB on nonenzymatic antioxidants issummarized in Figures 2 and 3. Inoculation of NDEAdrastically decreased the GSH content to 39.3%(p < 0.01) in the NDEA control group. The adminis-tration of MECB at the dose of 100 and 200mg=kg.b.w. to the NDEA-induced rats increased GSH levelsby 27.4% and 35.7%, respectively, as compared withthe NDEA control group (p < 0.05). The levels of vit-amin C in the liver of the NDEA control groupdecreased by 72.5% (p < 0.01) in comparison withthe normal group. After administration of MECB atthe doses of 100 and 200mg=kg. b.w., the levels ofvitamin C increased by 60.2% and 70.3%, respect-ively, as compared to that of the NDEA controlgroup (p < 0.05). The vitamin E level in the NDEAcontrol group decreased by 47.4% (p < 0.01) in com-parison with the normal group. Treatment with
Figure 1. Effect of the methanol extract of Caesalpinia bonducella (MECB) on levels of lipid peroxidation in the liver of NDEA-induced hepatocarcinogenic rats. The values are expressed as the mean� SEM (n ¼ 6). �p < 0.01 NDEA control group comparedwith the control group. þ p < 0.05 experimental groups compared with NDEA control group.
Antioxidant defense system of C. bonducella 413

MECB at doses of 100 and 200mg=kg. b.w. increasedvitamin E levels by 31.3% and 45.3%, respectively,when compared to that of the NDEA control(p < 0.05).
Effect of MECB on antioxidant enzymes (SOD and CAT)
The effect of MECB on antioxidant enzymes is givenin Figure 4. The levels of SOD in the liver of theNDEA control group decreased by 32.4% (p < 0.01).
Figure 2. Effect of the methanol extract of Caesalpinia bonducella (MECB) on levels of glutathione content in the liver of NDEA-induced hepatocarcinogenic rats. The values are expressed as the mean� SEM (n ¼ 6). �p < 0.01 NDEA control group comparedwith the control group. þ p < 0.05 experimental groups compared with NDEA control group.
Figure 3. Effect of the methanol extract of Caesalpinia bonducella (MECB) on levels of vitamin E and vitamin C in the liver ofNDEA-induced hepatocarcinogenic rats. The values are expressed as the mean� SEM (n ¼ 6). �p < 0.01 NDEA control groupcompared with the control group. þ p < 0.05 experimental groups compared with NDEA control group.
414 M. Gupta et al.

After administration of MECB at doses of 100 and200mg=kg, levels of SOD increased by 15.4% and28.7%, respectively, as compared to that of theNDEA control group (p < 0.05). The CAT level in
the NDEA control group decreased by 62.5%(p < 0.01) in comparison with the normal group.Treatment with MECB at doses of 100 and200mg=kg. b.w. increased CAT levels by 41.2% and
Figure 4. Effect of the methanol extract of Caesalpinia bonducella (MECB) on levels of SOD and CAT in the liver of NDEA-inducedhepatocarcinogenic rats. The values are expressed as the mean� SEM (n ¼ 6). �p < 0.01 NDEA control group compared with thecontrol group. þ p < 0.05 experimental groups compared with NDEA control group.
Figure 5. Effect of the methanol extract of Caesalpinia bonducella (MECB) on levels of GPT, GOT, and ALP in the serum of NDEA-induced hepatocarcinogenic rats. The values are expressed as the mean� SEM (n ¼ 6). �p < 0.01 NDEA control group comparedwith the control group. þ p < 0.05 experimental groups compared with NDEA control group.
Antioxidant defense system of C. bonducella 415

52.2%, respectively, when compared to that of theNDEA control (p < 0.05).
Effect of MECB on serum enzymes, bilirubin, protein,
and uric acid
Changes in the activities of serum enzymes (GPT, GOT,and ALP), bilirubin and protein content, and uric acid inthe serum of NDEA-induced hepatocellular carcinomain rats are summarized in Figures 5 and 6. The level ofserum marker enzymes GPT, GOT, ALP, and bilirubinwere found to be significantly increased, and the proteincontent and uric acid levels were significantly decreasedin NDEA control rats when compared with the normalgroup (p < 0.01). Treatment with MECB at doses of100 and 200mg=kg b.w. decreased the activity of serumtransaminase, ALP, and bilirubin in a dose-dependentmanner and increased the protein content and uric acidlevels in NDEA-induced hepatocellular carcinoma com-pared to that of control groups (p < 0.05).
Discussion
The results showed that administration of MECB couldprevent NDEA-induced oxidative damage in the rat. Ithas been reported that NDEA is metabolized to alkylat-ing reactants, which could interact with DNA andinitiate carcinogenesis (Anis et al., 2001). Although manyrisk factors have been reported, lipid peroxidation plays
an important role in hepatic carcinogenesis. Further,enhanced lipid peroxidation associated with depletionof antioxidants is a characteristic finding in a variety ofmalignancies. The current study indicates that theincreased levels of MDA in the rats of the NDEA controlgroup could be ascribed to the excessive generation offree radicals. Szatrowski and Nathan (1991) have sug-gested that tumor cells produce a substantial amountof hydrogen peroxide and reactive oxygen metabolitesthat are released into the circulation. Therefore, theincreased susceptibility of plasma and liver of NDEA-treated rats could be due to the production of reactiveoxygen metabolites during the metabolism of NDEAor during the process of carcinogenesis. Decreased levelsof MDA in MECB-treated rats suggest the extracts couldscavenge free radicals produced by NDEA. MECB hasbeen reported to inhibit lipid peroxidation and scavengereactive oxygen species (Gupta et al., 2003, 2004).
SOD and CAT provide the first defense against oxygentoxicity by catalyzing the dismutation of superoxide anionto hydrogen peroxide and decomposition of hydrogen per-oxide to water and molecular oxygen. Earlier reportsshowed the decreased activities of glutathione peroxidase(GPx), SOD, and CAT in hepatoma (Corrocher et al.,1986). Decreased activities of SOD and CAT in NDEA-treated rats could be due to overuse of these nonenzymaticand enzymatic antioxidants to scavenge the productsof lipid peroxidation. Tumor cells have been reported tosequester essential antioxidants from the circulationin order to meet the demands of the growing tumor (Buzby
Figure 6. Effect of the methanol extract of Caesalpinia bonducella (MECB) on levels of bilirubin, protein, and uric acid in the serumof NDEA-induced hepatocarcinogenic rats. The values are expressed as the mean� SEM (n ¼ 6). �p < 0.01 NDEA control groupcompared with the control group. þ p < 0.05 experimental groups compared with the NDEA control group.
416 M. Gupta et al.

et al., 1980). MECB did not enhance antioxidant enzymes.It prevented NDEA-induced inactivation of antioxidantenzymes by an unknown mechanism. Enhanced levelsand activities of nonenzymatic and enzymatic antioxidantsin MECB-treated rats indicate the chemopreventive poten-tial of the extract.
GSH plays a critical role in important cellular functions,which includes the maintenance of thiol status of proteins,the destruction ofH2O2, lipid peroxides, free radicals, trans-location of amino acids across cell membranes, the detoxifi-cation of foreign compounds, and the biotransformation ofdrugs (James & Hrabison, 1982). The decreased level ofGSH in NDEA control group animals may be due to itsuse by the excessive amount of free radicals. Increased lipidperoxidation (LPO) also correlates with depletion of GSH,and this effect is expectedat a stepprior to alterationof poly-unsaturated fatty acids (Burk, 1983). GSH depletion maysensitize tumor cells to cytostatic effects without promotingtoxicity toward normal host cells (Romine & Kessel, 1986).The administration of NDEA also depletes GSH, and thisdepletion not only compromises cellular defenses againstattack by reactive molecules but also has profound effectson normal hepatocellular function. The GSH levels wereincreased to near normal in drug-treated rats, which maybe due to the antioxidant activity of MECB.
Vitamin E is the most significant antioxidant of itskind in animal cells, and it can protect against carcino-genesis and tumor growth (Das, 1994). Significantlydecreased vitamin E levels in NDEA control animalsmight be due to the excessive use of this antioxidantfor quenching enormous free radicals produced in theseconditions. The increased level of vitamin E in drug-treated rats reveals the antioxidative nature of MECB.The extract may scavenge the free radicals and thusmaintains the normal level of vitamin E. This effectcan be attributed to the presence of flavonoids that canserve as a double antioxidant, as free radical scavengersand as excited species quencher (Fraga et al., 1988).
The availability of vitamin C is a determining factor incontrolling and potentiating many aspects of host resist-ance to cancer. The decreased level of vitamin C wasfound in NDEA control animals. Vitamin C can protectcell membranes and lipoprotein particles from oxidativedamage by regenerating the antioxidant form of vitaminE (Buettner, 1993). Thus, vitamins C and E act synergis-tically in scavenging a wide variety of reactive oxygenspecies (ROS). The recovery of vitamin C to near normallevels in drug-treated rats was found to be due to thepotent anticancer activity of nut extract that may inducethe regeneration of ascorbic acid.
In the assessment of liver damage by hepatotoxin, thedetermination of enzyme levels such as SGPT and SGOTis widely used. Necrosis or membrane damage releases theenzyme into circulation. High levels of SGOT indicate theliver damage, such as that due to viral hepatitis, as well ascardiac infarction and muscle injury. Therefore, SGPT is
more specific to the liver and is thus a better parameterfor detecting liver injury (Willianson et al., 1996). MECBat different doses caused a significant decrease in SGPTand SGOT levels in NDEA-treated rats. Serum ALPand bilirubin levels, on the other hand, are related tothe function of hepatic cell. Increases in the serum levelof ALP are due to increased synthesis in presence ofincreasing biliary pressure (Moss & Butterworth, 1974).MECB at different doses caused significant inhibition ofSALP and bilirubin levels in NDEA-induced hepatocellu-lar carcinoma in rats. Effective control of bilirubin leveland alkaline phosphatase activity points toward an earlyimprovement in the secretory mechanism of the hepaticcell.
Uric acid, the metabolic end-product of purine metab-olism, has proved to be a selective antioxidant, capableespecially of reacting with free radicals and hypochlorousacid (Hasugawa & Kuroda, 1989). The reduced level ofuric acid in hepatocarcinogenesis conditions may bedue to the increased use of uric acid against increasedproduction of the free radicals, which is a characteristicfeature of cancer condition. The reversal of altered uricacid level to near normal in MECB-treated rats couldbe due to strong antioxidant property of MECB, whichcontributes to its anticarcinogenic potency.
As already stated, Caesalpinia bonducella contain flavo-noid (bonducellin-A) (Purushothaman et al., 1982), triter-penoids (a-amyrin, b-amyrin, lupeol, and lupeol acetate)(Saeed & Sabir, 2003), and diterpenoids (bonducin, bondu-cellpin A–C, caesaldekarin A–F, and caesalpin A–C) (Peteret al., 1997, 1999; Lyder et al., 1998; Kinoshita, 2000), andsteroid (b-sitosterol) (Lai et al., 1977). Plant-derived naturalproducts such as flavonoids, terpenoids and steroids, andso forth, have received considerable attention in recentyears due to their diverse pharmacological propertiesincluding antioxidant and antitumor activity (DeFeudiset al., 2003; Takeoka & Dao, 2003). Antioxidants play animportant role in inhibiting and scavenging radicals, thusproviding protection to humans against infection anddegenerative diseases. Therefore, the presence of flavo-noids, triterpenoids, and steroids in MECBmay be respon-sible for the antioxidant and anticarcinogenic activity.
In conclusion, biochemical alterations observed in hepa-tocarcinogenesis status seems to be mainly due to an oxy-radical–mediated mechanism, involving lipid peroxidation,under conditions of reduced antioxidant defenses. Theevidence comprises loss of enzymatic (Premalatha et al.,1997) and nonenzymatic antioxidants that scavenge super-oxide, hydrogen peroxide, and lipid peroxides. The resultsyet available are overwhelming, which suggests thatMECBadministration may be helpful in quenching free radicalsand induction of an in vivo antioxidant defense system bythe flavonoids and triterpenoids present in it. These obser-vations indicate that with regard to NDEA-induced hepa-tocarcinogenesis, MECB administration may be protectiveand therapeutic. Thus, considering free radical mediation in
Antioxidant defense system of C. bonducella 417

NDEA-induced hepatocarcinogenesis, the current findingssuggest the anticarcinogenic capability of MECB throughits antioxidant properties and induction of in vivo antioxi-dant defense system.
Further investigations are in progress in our labora-tory to identify the active principles involved in this anti-oxidant and anticarcinogenic activity.
Acknowledgment
The authors are grateful to AICTE, New Delhi, India,for providing financial support to this work.
References
Adesina SK (1982): Studies on some plants used as anticon-
vulsant in Amerindian and African traditional medi-
cine. Fitoterapia 53: 147–162.
Aebi H (1974): Catalase. In: Bergmeyer HV, ed., Methods in
Enzymatic Analysis, Vol 2. New York, Academic Press,
pp. 674–684.
Anis KV, Rajesh Kumar NV, Kuttan R (2001): Inhibition
of chemical carcinogenesis by biberine in rats and mice.
J Pharm Pharmacol 53: 763–768.
Baker AF, Frank G (1951): Estimation of vitamin E in
tissues. In: Bollinger G, ed., Chromaeographic in Labor-
atorium Handbuch. Berlin, Springer-Verlag, pp. 41–52.
Bergmeyer HU, Scheibe P, Wahlefeld AW (1978): Optimiza-
tion of methods for aspartate aminotransferase and
alanine aminotransferase. Clin Chem 24: 58–61.
Buettner GR (1993): The pecking order of free radicals and
antioxidants: lipid peroxidation, a-tocopherol and
ascorbate. Arch Biochem Biophys 300: 535–543.
Burk RF (1983): Glutathione-dependent protection by rat
liver microsomal protein against lipid peroxidation.
Biochim Biophys Acta 757: 21–28.
Buzby GP, Mullen JH, Stein TP, Roasto EF (1980): Host
tumor interactions and nutrient supply. Cancer 45:
2940–2947.
Caraway WT (1963): Uric acid. In: Seligson D, ed., Stan-
dard Methods of Clinical Chemistry, Vol 4. New York,
Academic Press, pp. 239–247.
Corrocher R, Casani M, Bellisola GB, Nicoli N, Guidi GC,
Sandre G (1986): Severe impairment of antioxidants
system in human hepatoma. Cancer 58: 11658–11662.
Das S (1994): Vitamin E in the genesis and prevention of
cancer. A review. Acta Oncol 33: 615–619.
DeFeudis FV, Papadopoulos V, Drieu K (2003): Ginkgo
biloba extracts and cancer: A research area in its
infancy. Fundam Clin Pharmacol 17: 405–417.
Dhar ML, Dhar MM, Dhawan BN, Mehrotra BN, Roy C
(1968): Screening of Indian plants for biological
activity. Indian Exp Biol 6: 232–247.
Ellman GL (1959): Tissue sulphydryl groups. Arch Biochem
Biophys 82: 70–77.
Farber E (1987): Pathogenesis of experimental liver cancer:
Comparison with humans. Arch Toxicol (Suppl) 100:
281–288.
Farber E (1996): The step by step development of epithelial
cancer: from genotype to phenotype. In: Van de Woude
GF, Klien G, eds., Advances in Cancer Research.
San Diego, Academic Press, 70: pp. 21–48.
Farber E, Sarma DS (1987): Hepatocarcinogenesis: A
dynamic cellular perspective. Lab Invest 56: 4–22.
Fraga CG, Leibovitz BE, Toppel AL (1988): Lipid peroxi-
dation measured as TBARS in tissue slices: Character-
ization and comparison with homogenates and
microsomes. Free Radic Biol Med 4: 155–161.
Gayaraja S, Shinde S, Agarwal SL (1978): Antiasthmatic
properties ofCaesalpinia bonduc leaves. Indian JPharmacol
10: 86–89.
Gupta M, Mazumder UK, Sambath Kumar R (2003a):
Hepatoprotective and antioxidant role of Caesalpinia
bonducella on paracetamol-induced liver damage in
rats. Nat Prod Sci 9: 186–191.
Gupta M, Mazumder UK, Sambath Kumar R, Siva Kumar
T (2003b): Studies on antiinflammatory, analgesic and
antipyretic properties of methanol extract of Caesalpi-
nia bonducella leaves in experimental animal models.
Iranian J Pharmacol Thera 2: 30–34.
Gupta M, Mazumder UK, Sambath Kumar R, Siva Kumar
T, Vamsi MLM (2004): Antitumor activity and antiox-
idant status of Caesalpinia bonducella against Ehrlich
ascites carcinoma in Swiss albino mice. J Pharmacol
Sci 94: 177–184.
Hasugawa T, Kuroda M (1989): A new role of uric acid as
antioxidant in human plasma. Japanese J Clin Pathol
37: 1020–1027.
James RC, Hrabison RD (1982): Hepatic glutathione and
hepatotoxicity. Biochem Pharmacol 31: 1829–1835.
Kakkar P, Das B, Viswanathan PN (1984): A modified
spectrophotometric assay of superoxide dismutase.
J Biochem Biophys 21: 130–132.
King J (1965): The hydrolases-acid and alkaline phospatase.
In: Van D, ed., Practical Clinical Enzymology. London,
Nostrand Company Ltd, pp. 191–208.
Kinoshita T (2000): Chemical studies on the Philippine crude
drug calumbibit; The isolation of new cassane diterpenes
fused with alpha. Chem Pharm Bull 48: 1375–1377.
Kirtikar KR, Basu BD (1975): Indian Medicinal Plants, Vol
2. Dehradum, India, Bishen Mahendra Pal Singh, pp.
842–844.
Lai MS, Shameel S, Ahmad VU, Ushmanghani K (1977):
Chemical constituents of Caesalpinia bonduc. Pak J
Sci Ind Res 40: 20–22.
Leoppky RN, Li YE (1991): Nitrosamine activation and
detoxification through free radicals and their derived
cations. In: Neill IKO, Chem J, Bartch H, eds., Rel-
evance to Human Cancer of Nitrosoamine Compounds,
Tobacco and Mycotoxins. Lyan, LARC Scientific Publi-
cation. No. 1005, pp. 375–382.
418 M. Gupta et al.

Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951):
Protein measurement with the Folin-phenol reagent.
J Biol Chem 193: 265–275.
Lyder Dl, Peter SR, Tinto WF (1998): Minor cassane
diterpenoids of Caesalpinia bonduc. J Nat Prod 61:
1462–1465.
Malloy HT, Evelyn KA (1937): The determination of bilir-
ubin with the photoelectric colorimeter. J Biol Chem
119: 481–490.
Metlin C (1997): Chemoprevention: Will it work? Int J
Cancer 10: 18–21.
Moss DW, Butterworth PJ (1974): Enzymology and Medi-
cine. London, Pitman Medical, p. 139.
Neogi NC, Nayak KP (1958): Biological investigation of
Caesalpinia bonducella F. Indian J Pharmacol 20:
95–100.
Ohkawa H, Onishi N, Yagi K (1979): Assay for lipid peroxi-
dation in animal tissue by thiobarbituric acid reaction.
Anal Biochem 95: 351–358.
Omaye ST, Turun Ball TD, Sauberlich HE (1979):
Selected method for the determination of ascorbic acid in
animal cells, tissues and fluids. Meth Enzymol 62: 1–11.
Peter SR, Tinto WF, Lean S, Reynolds WF, Yu M (1997):
Bonducellpins A–D new cassane furanoditerpenes of
Caesalpinia bonduc. J Nat Prod 60: 1219–1221.
Peter SR, Tinto WF, Lean S, Reynolds WF, Yu M (1999):
Cassane diterpenes from Caesalpinia bonducella. Phyto-
chemistry 47: 1153–1155.
Premalatha B, Sujatha V, Sachdanandam P (1997): Modu-
lating effect of Semecarpus anacardium Linn. nut
extract on glucose metabolising enzymes in aflatoxin
B1-induced experimental hepatocellular carcinoma.
Pharmacol Res 36: 187–192.
Purushothaman KK, Kalyani K, Subramaniam K,
Shanmughanathan SP (1982): Structure of bonducellin-
A new homoisoflavone from Caesalpinia bonducella.
Indian J Chem 21: 383–386.
Quaife MC, Dju MY (1948): Chemical estimation of vit-
amin E in tissue and a-tocopherol content of normal
tissues. J Biol Chem 180: 263–272.
Raghunathan K, Mitra R, Karanja (1982): In: Raghu-
nathan K, Mitra R, eds., Pharmacognosy of Indigenous
Drugs, Part I. New Delhi, India, Central Council for
Research in Ayurveda and Siddha, pp. 484–510.
Ray G, Batra S, Shkula NK, Deo S, Raina V, Asok S,
Husain SA (2000): Lipid peroxidation, free radicals
production and antioxidant status in breast cancer.
Breast Cancer Res Treat 59: 163–170.
Romine MT, Kessel D (1986): Intracellular glutathione as a
determinant of responsiveness to antitumour drugs.
Biochem Pharmacol 35: 3323–3326.
Saeed MA, Sabir AW (2003): Irritant potential of some con-
stituents from seeds of Caesalpinia bonducella (L.)
Flaming. J Asian Nat Prod Res 5: 35–41.
Schaff ZS, Lapis K, Henson DE (1993): Liver. In: Henson
DE, Albores Saavedra J, eds., Pathology of Incipient
Neoplasia. Philadelphia, Saunders, pp. 151–166.
Sundaresan S, Subramanian P (2003): s-Allyl cysteine inhibits
circulatory lipid peroxidation and promotes antioxidants
in N-nitrosodiethylamine-induced carcinogenesis. Pol J
Pharmacol 55: 37–42.
Szatrowski TP, Nathan CF (1991): Production of large
amounts of hydrogen peroxide by human tumor cells.
Cancer Res 51: 794–798.
Takeoka GR, Dao LT (2003): Antioxidant constituent of
almond [Prunus dulcis (Mill.) D.A. Webb.] hulls.
J Agric Food Chem 51: 496–501.
Tricker AR, Pfundstein B, Theobald E, Preussman R,
Spiegehalder B (1991): Mean daily intake of volatile
N-nitrosamines from foods and beverages in West
Germany in 1989–90. Food Chem Toxicol 29: 729–732.
Willianson EM, Okpako DT, Evans FJ (1996): Selection,
Preparation and Pharmacological Evaluation of Plant
Material. Chichester, England, John Wiley.
Antioxidant defense system of C. bonducella 419




![KÜLTÜR ve TURİZM BAKANLIĞI UZMANLIK TEZLERİ (#419] LİSTESİ](https://static.fdokumen.com/doc/165x107/632009cd01d52108cc019437/kueltuer-ve-turizm-bakanligi-uzmanlik-tezleri-419-listesi.jpg)















