
Patch Testing for Contact Allergy and Allergic Contact Dermatitis
Upload
khangminh22Category
view
2download
0
�����������������
Citation: Kim, E.-A.; Han, E.-J.; Kim,
J.; Fernando, I.P.S.; Oh, J.-Y.; Kim,
K.-N.; Ahn, G.; Heo, S.-J.
Anti-Allergic Effect of
3,4-Dihydroxybenzaldehyde Isolated
from Polysiphonia morrowii in
IgE/BSA-Stimulated Mast Cells and
a Passive Cutaneous Anaphylaxis
Mouse Model. Mar. Drugs 2022, 20,
133. https://doi.org/10.3390/
md20020133
Academic Editor: Hee Jae Shin
Received: 28 December 2021
Accepted: 7 February 2022
Published: 10 February 2022
Publisher’s Note: MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations.
Copyright: © 2022 by the authors.
Licensee MDPI, Basel, Switzerland.
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https://
creativecommons.org/licenses/by/
4.0/).
marine drugs
Article
Anti-Allergic Effect of 3,4-Dihydroxybenzaldehyde Isolatedfrom Polysiphonia morrowii in IgE/BSA-Stimulated Mast Cellsand a Passive Cutaneous Anaphylaxis Mouse ModelEun-A Kim 1,†, Eui-Jeong Han 2,3,†, Junseong Kim 1, Ilekuttige Priyan Shanura Fernando 4, Jae-Young Oh 5,Kil-Nam Kim 6 , Ginnae Ahn 3,4,* and Soo-Jin Heo 1,7,*
1 Jeju Marine Research Center, Korea Institute of Ocean Science & Technology (KIOST), Jeju 63349, Korea;[email protected] (E.-A.K.); [email protected] (J.K.)
2 Research Center for Healthcare and Biomedical Engineering, Chonnam National University,Yeosu 59626, Korea; [email protected]
3 Department of Food Technology and Nutrition, Chonnam National University, Yeosu 59626, Korea4 Department of Marine Bio-Food Sciences, Chonnam National University, Yeosu 59626, Korea;
[email protected] Food Safety and Processing Research Division, National Institute of Fisheries Science, Busan 46083, Korea;
[email protected] Chuncheon Center, Korea Basic Science Institute (KBSI), Chuncheon 24341, Korea; [email protected] Department of Marine Biology, University of Science and Technology, Daejeon 34113, Korea* Correspondence: [email protected] (G.A.); [email protected] (S.-J.H.); Tel.: +82-61-659-7213 (G.A.);
+82-64-798-6039 (S.-J.H.)† These authors contributed equally to this study.
Abstract: In this study, we investigated the anti-allergic effects of 3,4-dihydroxybenzaldehyde (DHB)isolated from the marine red alga, Polysiphonia morrowii, in mouse bone-marrow-derived culturedmast cells (BMCMCs) and passive cutaneous anaphylaxis (PCA) in anti-dinitrophenyl (DNP) im-munoglobulin E (IgE)-sensitized mice. DHB inhibited IgE/bovine serum albumin (BSA)-inducedBMCMCs degranulation by reducing the release of β-hexosaminidase without inducing cytotoxicity.Further, DHB dose-dependently decreased the IgE binding and high-affinity IgE receptor (FcεRI)expression and FcεRI-IgE binding on the surface of BMCMCs. Moreover, DHB suppressed thesecretion and/or the expression of the allergic cytokines, interleukin (IL)-4, IL-5, IL-6, IL-13, andtumor necrosis factor (TNF)-α, and the chemokine, thymus activation-regulated chemokine (TARC),by regulating the phosphorylation of IκBα and the translocation of cytoplasmic NF-κB into thenucleus. Furthermore, DHB attenuated the passive cutaneous anaphylactic (PCA) reaction reducingthe exuded Evans blue amount in the mouse ear stimulated by IgE/BSA. These results suggest thatDHB is a potential therapeutic candidate for the prevention and treatment of type I allergic disorders.
Keywords: Polysiphonia morrowii; 3,4-dihydroxybenzaldehyde; immunoglobulin E; bone marrow-derived cultured mast cells; passive cutaneous anaphylaxis
1. Introduction
The rapidly increasing incidence of allergic diseases, including asthma, allergic rhinitis,and atopic eczema, has become a significant public health problem [1,2]. Among allergicdiseases, type I allergic reactions are the most common in allergic patients and are causedby immunoglobulin E (IgE)/antigen-mediated mast cell activation [3]. Therefore, the inhi-bition of mast cell activation is a crucial factor in the development of treatment strategiesfor type I allergic diseases [3]. Mast cells may be activated through the crosslinking ofhigh-affinity receptors to IgE (FcεRI) by the IgE/antigen complex expressed on the surfaceof cells [4–6]. The activated mast cells are sensitized to release allergic mediators such asgranular contents (β-hexosaminidase) as well as allergic cytokines (interleukin (IL)-4, IL-5,
Mar. Drugs 2022, 20, 133. https://doi.org/10.3390/md20020133 https://www.mdpi.com/journal/marinedrugs

Mar. Drugs 2022, 20, 133 2 of 13
IL-6, IL-13), and tumor necrosis factor (TNF)(-α) and the chemokine, thymus activation-regulated chemokine (TARC), which induces allergic disease [6,7]. In particular, the majoreffectors of allergic symptoms are the allergic cytokines by regulated pathological problemson mast cells, as overexpression may cause pathological problems [5]. Recent studiesreported that NF-κB signaling plays a key role in the regulation of the expression andproduction levels of immune, inflammatory, and allergic cytokines and mediators [7–10].This suggests that the NF-κB signaling pathway is crucial in the allergic response. Variousmedications such as antihistamines or corticosteroids have been commonly used to inhibitallergic reactions; however, their use is limited owing to their severe or partial adverseeffects such as damage to the immune system, cardiac toxicity, and bone loss [6,7,11]. Inview of this, many studies have focused on the use of safe and natural materials as alterna-tives with lower toxicity and fewer side effects [9,10]. Among these natural materials, algaehave various biological activities. Algae contain a variety of functional compounds withan intriguing structural diversity [12–15]. Polysiphonia morrowii Harvey (P. morrowii), a redalga, produces several bioactive substances such as 5-bromo-3,4-dihydroxybenzaldehyde(BDB), 3,4-dihydroxybenzaldehyde (DHB), 3-bromo-5-(ethoxymethyl)-1,2-benzenediol, 3-bromo-4,5-dihydroxybenzyl methyl ether, 3-bromo-4,5-dihydroxybenzaldehyde, and bis(3-bromo-4,5-dihydroxybenzyl) ether [12–19]. These bio-active substances have diversephysiological effects including anti-inflammatory, antiviral, anti-bacterial, antioxidant, an-tiobesity, and antiallergic activities [12–19]. In our previous study, we reported that BDBisolated from P. morrwii possess antiallergic effects in IgE-induced mast cell activation andpassive cutaneous anaphylactic (PCA) reaction in mice model [12]. Additionally, recentstudies reported that the DHB, a compound without a bromo group from BDB, has theantioxidant, anti-inflammation and antiallergic effects [17,19,20]. However, there are noreports elucidating the plausible biological mechanism related to the antiallergic effects ofDHB in IgE-induced mast cell activation and PCA mice model. Therefore, in this study,we evaluated the antiallergic effects of DHB by inhibiting allergic factors and attemptedto unveil the underlying biological mechanism in IgE/BSA-stimulated a bone marrowcultured mast cells (BMCMCs) and a PCA mouse model.
2. Results2.1. DHB Effectively Decreased β-Hexosaminidase Release without Cytotoxicity in BMCMCs
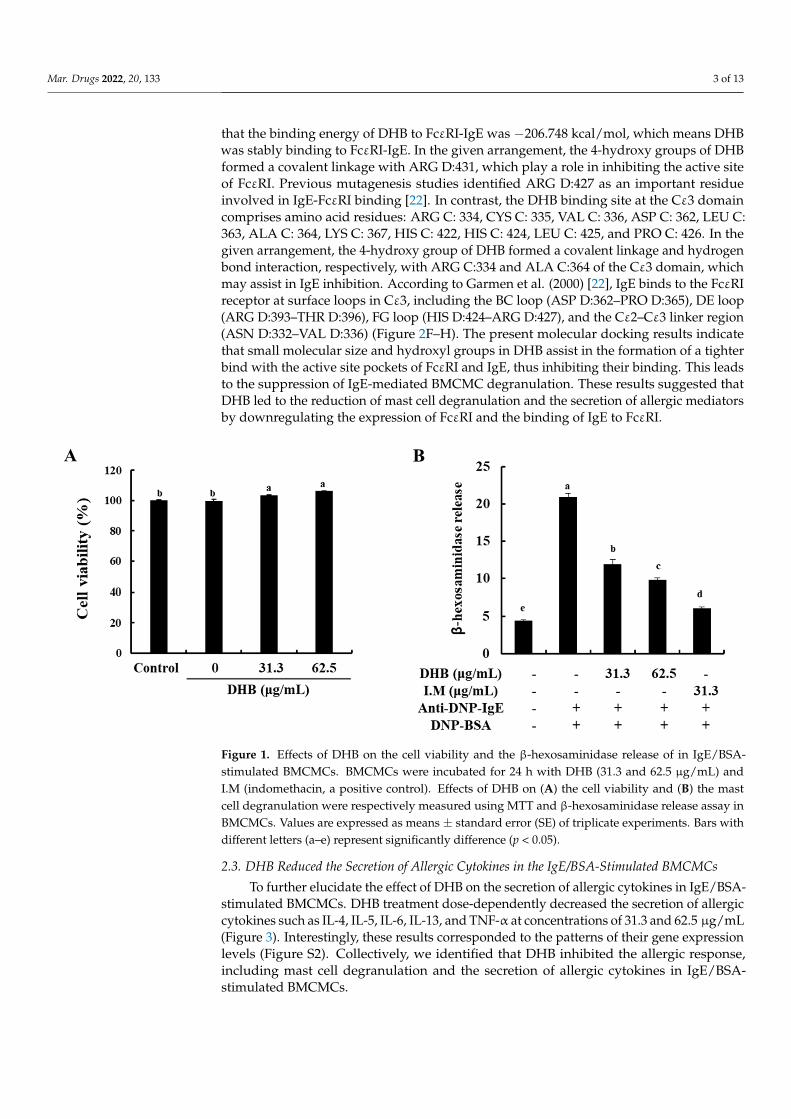
To identify the cytotoxicity of DHB on BMCMCs, the MTT assay was performed. Atall of the studied concentrations, DHB did not show any cytotoxicity in the BMCMCs com-pared to the untreated control cells (Figure 1A). On the basis of this result, we determinedthe concentration range of DHB for further experiments. Next, we investigated the effectof DHB on the degranulation of BMCMCs by evaluating the release of β-hexosaminidasein the IgE/BSA-stimulated BMCMCs. β-hexosaminidase release is a widely investigatedbiomarker of mast cell degranulation [21]. As shown in Figure 1B, DHB significantly re-duced the release of β-hexosaminidase in the IgE/BSA-stimulated BMCMCs. These resultsindicated that DHB suppressed the degranulation of the IgE/BSA-stimulated BMCMCs byreducing the β-hexosaminidase release.
2.2. DHB Inhibited the Expression of FcεRI and the Binding of IgE to FcεRI
To identify whether DHB inhibits the expression of FcεRI and the FcεRI-IgE bind-ing on the surface of BMCMCs, we performed flow cytometric analysis. DHB dose-dependently reduced the expression of FcεRI compared to that in the untreated control cells(Figure 2A). Typically, the binding of IgE on the surface of FcεRI initiates the activation ofthe mast cells and finally induces allergic reactions [5]. Based on the flow cytometry results(Figure 2B), IgE binding showed a marked increase in the IgE/BSA-stimulated cells com-pared with that of the control. Pretreatment with DHB significantly and dose-dependentlyreduced FcεRI expression at the investigated concentrations. DHB energy to FcεRI was−182.5245 kcal/mol. The binding site at the α-chain domain of DHB comprises amino acidresidues, ARG D:427, ARG D:431, and LEU D:429 (Figure 2C–E). In addition, we confirmed
Mar. Drugs 2022, 20, 133 3 of 13
that the binding energy of DHB to FcεRI-IgE was −206.748 kcal/mol, which means DHBwas stably binding to FcεRI-IgE. In the given arrangement, the 4-hydroxy groups of DHBformed a covalent linkage with ARG D:431, which play a role in inhibiting the active siteof FcεRI. Previous mutagenesis studies identified ARG D:427 as an important residueinvolved in IgE-FcεRI binding [22]. In contrast, the DHB binding site at the Cε3 domaincomprises amino acid residues: ARG C: 334, CYS C: 335, VAL C: 336, ASP C: 362, LEU C:363, ALA C: 364, LYS C: 367, HIS C: 422, HIS C: 424, LEU C: 425, and PRO C: 426. In thegiven arrangement, the 4-hydroxy group of DHB formed a covalent linkage and hydrogenbond interaction, respectively, with ARG C:334 and ALA C:364 of the Cε3 domain, whichmay assist in IgE inhibition. According to Garmen et al. (2000) [22], IgE binds to the FcεRIreceptor at surface loops in Cε3, including the BC loop (ASP D:362–PRO D:365), DE loop(ARG D:393–THR D:396), FG loop (HIS D:424–ARG D:427), and the Cε2–Cε3 linker region(ASN D:332–VAL D:336) (Figure 2F–H). The present molecular docking results indicatethat small molecular size and hydroxyl groups in DHB assist in the formation of a tighterbind with the active site pockets of FcεRI and IgE, thus inhibiting their binding. This leadsto the suppression of IgE-mediated BMCMC degranulation. These results suggested thatDHB led to the reduction of mast cell degranulation and the secretion of allergic mediatorsby downregulating the expression of FcεRI and the binding of IgE to FcεRI.
Mar. Drugs 2021, 19, x FOR PEER REVIEW 3 of 13
Figure 1. Effects of DHB on the cell viability and the β-hexosaminidase release of in IgE/BSA-stim-ulated BMCMCs. BMCMCs were incubated for 24 h with DHB (31.3 and 62.5 µg/mL) and I.M (in-domethacin, a positive control). Effects of DHB on (A) the cell viability and (B) the mast cell degran-ulation were respectively measured using MTT and β-hexosaminidase release assay in BMCMCs. Values are expressed as means ± standard error (SE) of triplicate experiments. Bars with different letters (a–e) represent significantly difference (p < 0.05).
2.2. DHB Inhibited the Expression of FcεRI and the Binding of IgE to FcεRI To identify whether DHB inhibits the expression of FcεRI and the FcεRI-IgE binding
on the surface of BMCMCs, we performed flow cytometric analysis. DHB dose-de-pendently reduced the expression of FcεRI compared to that in the untreated control cells (Figure 2A). Typically, the binding of IgE on the surface of FcεRI initiates the activation of the mast cells and finally induces allergic reactions [5]. Based on the flow cytometry re-sults (Figure 2B), IgE binding showed a marked increase in the IgE/BSA-stimulated cells compared with that of the control. Pretreatment with DHB significantly and dose-de-pendently reduced FcεRI expression at the investigated concentrations. DHB energy to FcεRI was −182.5245 kcal/mol. The binding site at the α-chain domain of DHB comprises amino acid residues, ARG D:427, ARG D:431, and LEU D:429 (Figure 2C–E). In addition, we confirmed that the binding energy of DHB to FcεRI-IgE was −206.748 kcal/mol, which means DHB was stably binding to FcεRI-IgE. In the given arrangement, the 4-hydroxy groups of DHB formed a covalent linkage with ARG D:431, which play a role in inhibiting the active site of FcεRI. Previous mutagenesis studies identified ARG D:427 as an im-portant residue involved in IgE-FcεRI binding [22]. In contrast, the DHB binding site at the Cε3 domain comprises amino acid residues: ARG C: 334, CYS C: 335, VAL C: 336, ASP C: 362, LEU C: 363, ALA C: 364, LYS C: 367, HIS C: 422, HIS C: 424, LEU C: 425, and PRO C: 426. In the given arrangement, the 4-hydroxy group of DHB formed a covalent linkage and hydrogen bond interaction, respectively, with ARG C:334 and ALA C:364 of the Cε3 domain, which may assist in IgE inhibition. According to Garmen et al. (2000) [22], IgE binds to the FcεRI receptor at surface loops in Cε3, including the BC loop (ASP D:362–PRO D:365), DE loop (ARG D:393–THR D:396), FG loop (HIS D:424–ARG D:427), and the Cε2–Cε3 linker region (ASN D:332–VAL D:336) (Figure 2F–H). The present molecular docking results indicate that small molecular size and hydroxyl groups in DHB assist in the formation of a tighter bind with the active site pockets of FcεRI and IgE, thus inhibiting their binding. This leads to the suppression of IgE-mediated BMCMC degranulation. These results suggested that DHB led to the reduction of mast cell degranulation and the secretion of allergic mediators by downregulating the expression of FcεRI and the binding of IgE to FcεRI.
Figure 1. Effects of DHB on the cell viability and the β-hexosaminidase release of in IgE/BSA-stimulated BMCMCs. BMCMCs were incubated for 24 h with DHB (31.3 and 62.5 µg/mL) andI.M (indomethacin, a positive control). Effects of DHB on (A) the cell viability and (B) the mastcell degranulation were respectively measured using MTT and β-hexosaminidase release assay inBMCMCs. Values are expressed as means ± standard error (SE) of triplicate experiments. Bars withdifferent letters (a–e) represent significantly difference (p < 0.05).
2.3. DHB Reduced the Secretion of Allergic Cytokines in the IgE/BSA-Stimulated BMCMCs
To further elucidate the effect of DHB on the secretion of allergic cytokines in IgE/BSA-stimulated BMCMCs. DHB treatment dose-dependently decreased the secretion of allergiccytokines such as IL-4, IL-5, IL-6, IL-13, and TNF-α at concentrations of 31.3 and 62.5 µg/mL(Figure 3). Interestingly, these results corresponded to the patterns of their gene expressionlevels (Figure S2). Collectively, we identified that DHB inhibited the allergic response,including mast cell degranulation and the secretion of allergic cytokines in IgE/BSA-stimulated BMCMCs.

Mar. Drugs 2022, 20, 133 4 of 13Mar. Drugs 2021, 19, x FOR PEER REVIEW 4 of 13
Figure 2. Effects of DHB on FcεRI expression and the binding of the IgE to FcεRI expression in IgE/BSA-stimulated BMCMCs and molecular docking analysis DHB binding. (A) Cell surface FcεRI expression and (B) IgE binding to FcεRI in BMCMCs were performed by flow cytometry. Prediction of stability and intermolecular interactions of DHB at (C–E) the active site of IgE and (F–H) IgE-FcεRI complex were identified by molecular docking analysis. Values are expressed as means ± standard error (SE) of triplicate experiments. Bars with different letters (a–c) represent significantly difference (p < 0.05).
2.3. DHB Reduced the Secretion of Allergic Cytokines in the IgE/BSA-Stimulated BMCMCs To further elucidate the effect of DHB on the secretion of allergic cytokines in
IgE/BSA-stimulated BMCMCs. DHB treatment dose-dependently decreased the secretion of allergic cytokines such as IL-4, IL-5, IL-6, IL-13, and TNF-α at concentrations of 31.3 and 62.5 µg/mL (Figure 3). Interestingly, these results corresponded to the patterns of their gene expression levels (Figure S2). Collectively, we identified that DHB inhibited the al-lergic response, including mast cell degranulation and the secretion of allergic cytokines in IgE/BSA-stimulated BMCMCs.
Figure 2. Effects of DHB on FcεRI expression and the binding of the IgE to FcεRI expression inIgE/BSA-stimulated BMCMCs and molecular docking analysis DHB binding. (A) Cell surface FcεRIexpression and (B) IgE binding to FcεRI in BMCMCs were performed by flow cytometry. Predictionof stability and intermolecular interactions of DHB at (C–E) the active site of IgE and (F–H) IgE-FcεRIcomplex were identified by molecular docking analysis. Values are expressed as means ± standarderror (SE) of triplicate experiments. Bars with different letters (a–c) represent significantly difference(p < 0.05).
Mar. Drugs 2021, 19, x FOR PEER REVIEW 5 of 13
Figure 3. Effects of DHB on the secretion of allergic cytokines in IgE/BSA-stimulated BMCMCs. The secretion of (A) IL-4, (B) IL-5, (C) IL-6, (D) IL-13, (E) TNF-α in cell supernatants were measured by mouse cytokine ELISA kits. Values are expressed as means ± standard error (SE) of triplicate exper-iments. Bars with different letters (a–d) represent significantly difference (p < 0.05).
2.4. DHB Inhibited the Activation of NF-κB Signaling Pathway in the IgE/BSA-Stimulated BMCMCs
The suppression of various cytokines expression via NF-κB inhibition is a key signal-ing pathway in the antiallergic effect [8]. Therefore, we investigated the effect of DHB on the IgE/BSA-induced action of NF-κB in the BMCMCs. As indicated in Figure 4, the stim-ulation of IgE/BSA induced the phosphorylation and degranulation of IκBα in the cytosol as well as the translocation of cytosolic NF-κB p65 to the nucleus. However, they were markedly modulated by the pretreatment of DHB in IgE/BSA-stimulated BMCMCs. Also, I.M, a positive control considerably reduced the activation of NF-κB p65. These results suggest that DHB improved the allergic response via inhibition of NF-κB signaling path-way in the IgE/BSA-stimulated BMCMCs.
Figure 4. Effects of DHB on NF-κB signaling pathway in IgE/BSA-stimulated BMCMCs. (A) Cyto-solic and (B) nucleic proteins obtained from BMCMCs were used for western blot analysis. The densitometry analysis of the western blot results was performed using (C) the ImageJ software to
Figure 3. Effects of DHB on the secretion of allergic cytokines in IgE/BSA-stimulated BMCMCs.The secretion of (A) IL-4, (B) IL-5, (C) IL-6, (D) IL-13, (E) TNF-α in cell supernatants were measuredby mouse cytokine ELISA kits. Values are expressed as means ± standard error (SE) of triplicateexperiments. Bars with different letters (a–d) represent significantly difference (p < 0.05).

Mar. Drugs 2022, 20, 133 5 of 13
2.4. DHB Inhibited the Activation of NF-κB Signaling Pathway in the IgE/BSA-StimulatedBMCMCs
The suppression of various cytokines expression via NF-κB inhibition is a key signalingpathway in the antiallergic effect [8]. Therefore, we investigated the effect of DHB on theIgE/BSA-induced action of NF-κB in the BMCMCs. As indicated in Figure 4, the stimulationof IgE/BSA induced the phosphorylation and degranulation of IκBα in the cytosol as wellas the translocation of cytosolic NF-κB p65 to the nucleus. However, they were markedlymodulated by the pretreatment of DHB in IgE/BSA-stimulated BMCMCs. Also, I.M, apositive control considerably reduced the activation of NF-κB p65. These results suggestthat DHB improved the allergic response via inhibition of NF-κB signaling pathway in theIgE/BSA-stimulated BMCMCs.
Mar. Drugs 2021, 19, x FOR PEER REVIEW 5 of 13
Figure 3. Effects of DHB on the secretion of allergic cytokines in IgE/BSA-stimulated BMCMCs. The secretion of (A) IL-4, (B) IL-5, (C) IL-6, (D) IL-13, (E) TNF-α in cell supernatants were measured by mouse cytokine ELISA kits. Values are expressed as means ± standard error (SE) of triplicate exper-iments. Bars with different letters (a–d) represent significantly difference (p < 0.05).
2.4. DHB Inhibited the Activation of NF-κB Signaling Pathway in the IgE/BSA-Stimulated BMCMCs
The suppression of various cytokines expression via NF-κB inhibition is a key signal-ing pathway in the antiallergic effect [8]. Therefore, we investigated the effect of DHB on the IgE/BSA-induced action of NF-κB in the BMCMCs. As indicated in Figure 4, the stim-ulation of IgE/BSA induced the phosphorylation and degranulation of IκBα in the cytosol as well as the translocation of cytosolic NF-κB p65 to the nucleus. However, they were markedly modulated by the pretreatment of DHB in IgE/BSA-stimulated BMCMCs. Also, I.M, a positive control considerably reduced the activation of NF-κB p65. These results suggest that DHB improved the allergic response via inhibition of NF-κB signaling path-way in the IgE/BSA-stimulated BMCMCs.
Figure 4. Effects of DHB on NF-κB signaling pathway in IgE/BSA-stimulated BMCMCs. (A) Cyto-solic and (B) nucleic proteins obtained from BMCMCs were used for western blot analysis. The densitometry analysis of the western blot results was performed using (C) the ImageJ software to
Figure 4. Effects of DHB on NF-κB signaling pathway in IgE/BSA-stimulated BMCMCs. (A) Cy-tosolic and (B) nucleic proteins obtained from BMCMCs were used for western blot analysis. Thedensitometry analysis of the western blot results was performed using (C) the ImageJ software toquantify the protein levels relative to corresponding housekeeping proteins. Values are expressedas means ± standard error (SE) of triplicate experiments. Bars with different letters (a–e) representsignificantly difference (p < 0.05).
2.5. DHB Improves the IgE/BSA Stimulation-Induced the PCA Reaction in Mice
To confirm the effect of DHB on the IgE/BSA stimulation-induced PCA reaction inmice, we monitored the amount of Evans blue dye extracted from the mice ear tissues.As shown in Figure 5A,B, stimulation by IgE/BSA markedly increased the PCA reactionby increasing the amount of Evans blue dye in mice ear tissue, compared to the normalmice group. Interestingly, it was significantly inhibited by the application of DHB with thereduced Evans blue amount. Next, we examined the degranulation of mast cells in miceear tissues by histological analysis. The topical application of DHB dose-dependently sup-pressed the epidermal thickness and degranulation of mast cells in ear tissues (Figure 5C,D).This result indicates that DHB effectively improved allergic reactions.

Mar. Drugs 2022, 20, 133 6 of 13
Mar. Drugs 2021, 19, x FOR PEER REVIEW 6 of 13
quantify the protein levels relative to corresponding housekeeping proteins. Values are expressed as means ± standard error (SE) of triplicate experiments. Bars with different letters (a–e) represent significantly difference (p < 0.05).
2.5. DHB Improves the IgE/BSA Stimulation-Induced the PCA Reaction in Mice To confirm the effect of DHB on the IgE/BSA stimulation-induced PCA reaction in
mice, we monitored the amount of Evans blue dye extracted from the mice ear tissues. As shown in Figure 5A,B, stimulation by IgE/BSA markedly increased the PCA reaction by increasing the amount of Evans blue dye in mice ear tissue, compared to the normal mice group. Interestingly, it was significantly inhibited by the application of DHB with the re-duced Evans blue amount. Next, we examined the degranulation of mast cells in mice ear tissues by histological analysis. The topical application of DHB dose-dependently sup-pressed the epidermal thickness and degranulation of mast cells in ear tissues (Figure 5C,D). This result indicates that DHB effectively improved allergic reactions.
Figure 5. Effects of DHB on the IgE/BSA stimulation-induced PCA reaction in mice. (A) The photo-graphic images of mice ear, (B) the amount of Evans blue dye extracted from mice ear tissues. (C,D) the degradation of mast cells was analyzed by H&E and toluidine blue staining assays in affected ear tissues. Values are expressed as means ± standard error (SE) of triplicate experiments. Bars with different letters (a,b) represent significantly difference (p < 0.05).
3. Discussion DHB, known to be a protocatechuic aldehyde and a water-soluble compound, is a
phenolic aldehyde. Generally, phenolic compounds represent the main type of secondary metabolite in plants and show multiple biological effects [23]. Therefore, these are used as pharmaceuticals and/or functional food components as antioxidants, anti-inflammatory agents, and antiproliferation and antiallergic agents [24–26]. Several studies have reported the biological activities of DHB, particularly antioxidant activity, anti-cardiac hypertro-phy, antiallergic effect, anti-inflammation, anti-atherosclerotic lesions, and anti-hepatitis B virus [17,19,24–26]. According to Kono et al. (2018), it was reported that DHB isolated from Prunus mume seed has antiallergy effect by suppressing the β-hexosaminidase re-
Figure 5. Effects of DHB on the IgE/BSA stimulation-induced PCA reaction in mice. (A) Thephotographic images of mice ear, (B) the amount of Evans blue dye extracted from mice ear tissues.(C,D) the degradation of mast cells was analyzed by H&E and toluidine blue staining assays inaffected ear tissues. Values are expressed as means ± standard error (SE) of triplicate experiments.Bars with different letters (a, b) represent significantly difference (p < 0.05).
3. Discussion
DHB, known to be a protocatechuic aldehyde and a water-soluble compound, is aphenolic aldehyde. Generally, phenolic compounds represent the main type of secondarymetabolite in plants and show multiple biological effects [23]. Therefore, these are used aspharmaceuticals and/or functional food components as antioxidants, anti-inflammatoryagents, and antiproliferation and antiallergic agents [24–26]. Several studies have reportedthe biological activities of DHB, particularly antioxidant activity, anti-cardiac hypertro-phy, antiallergic effect, anti-inflammation, anti-atherosclerotic lesions, and anti-hepatitisB virus [17,19,24–26]. According to Kono et al. (2018), it was reported that DHB iso-lated from Prunus mume seed has antiallergy effect by suppressing the β-hexosaminidaserelease in antigen-stimulated RBL-2H3 cells and BMCMCs [20]. However, the specificbiological mechanism underlying the effects of DHB has not yet been studied [27]. Inthe present study, we analyzed the antiallergic effect of DHB isolated from P. morrowii, ared alga, in IgE/BSA-stimulated BMCMCs and a PCA mouse model and evaluated itsbiological mechanism.
The annual increase in allergic diseases is caused by the inappropriate response ofimmune cells to allergens characterized by the excessive production of IgE [2,28]. IgE-allergenic stimulation is predisposed to produce histamine from stimulated mast cellsthrough a degranulation process. With the releases of histamine, β-hexosaminidase, agranule-associated exoglycosidase, is simultaneously degranulated in the secretory gran-ules of the mast cells and has been used as a biomarker of mast cell degranulation [5,28].Here, we observed DHB reduced the release of β-hexosaminidase in IgE/BSA-stimulatedBMCMCs. Treatment with 62.5 µg/mL DHB markedly suppressed the release of β-hexosaminidase compared with that of the only IgE/BSA-stimulated BMCMCs, without

Mar. Drugs 2022, 20, 133 7 of 13
cytotoxicity. Therefore, we suggest that DHB suppresses mast cell degranulation by reduc-ing β-hexosaminidase release. In a previous study, it was reported that DHB derived fromP. mume seed inhibited β-hexosaminidase release upon antigen-induced degranulation ofRBL-2H3 cells at the half maximal inhibitory concentration (IC50) values 0.6 mM, and itwas similar to the effects of DHB on the degranulation of BMCMCs [20].
Generally, mast cells express FcεRI on their membranes, which are important for theproinflammatory allergic response, and when an IgE-antigen binds with FcεRI, the receptoris activated and complex biological reactions occur, causing allergic reactions, includinginflammatory disorders [28]. Inhibiting IgE and FcεRI binding and FcεRI expression aresome of the possible approaches to prevent effector cell activation [9]. Drugs that couldachieve several or one of the above functions may be potential candidates for developingtherapeutics [27]. Recently, pharmacological research has been interested in the isolationof natural active compounds showing antiallergic activity with demonstrated inhibitionof β-hexosaminidase release as well as the expression of FcεRI and IgE binding to FcεRI,in marine algae [27,28]. Therefore, we chose the FcεRI and IgE as major target moleculesand confirmed the inhibition activity of DHB on mast cell degranulation through flowcytometry and molecular docking analysis. Flow cytometric analysis is one of the bestavailable methods to quantify the expression levels of specific surface receptor in livecells [9,10]. Molecular docking analysis is used in many natural product studies, because itcan confirm the stable binding ability for interaction of natural compounds with variousmolecular targets [9]. Our results revealed that DHB dose-dependently reduced the levelsof FcεRI expression on the surface of BMCMCs as well as IgE binding to FcεRI, comparedto the IgE-stimulated BMCMCs. The IgE antibody Fc comprises domains (Cε2-Cε3-Cε4),whereas IgE readily binds with the FcCε3 domains of the receptor (the α-chain domain ofFcεRI) [29]. Our previous study indicated BDB, a marine algal bromophenol of P. morrowiiexerted anti-allergic effect as reducing the formation of IgE and FcεRI complex by bindingto the active site of both FcεRI and IgE [9]. The outcomes of molecular docking analysisrevealed that DHB led to the feasible values of −182.525 kcal/mol and −206.748 kcal/molas free binding energies, provided that DHB could be a potent inhibitor of both FcεRI andIgE. DHB readily bound with the Cε3 domain of IgE than with the α-chain domain of FcεRI.These results clarify that the strong binding of the DHB to both FcεRI and IgE might reducemast cell degranulation caused by the IgE/BSA stimulation.
Furthermore, the activation of mast cells leads to a potential source of many cytokinesand a chemokine that might affect IgE-mediated allergic inflammation. These cytokines andchemokines include IL-1α, IL-2, IL-3, IL-4, IL-5, IL-6, IL-13, GM-CSF, TGF-β, TNF-α, IFN-γ,TARC, and several others [5,30]. IL-4 and IL-13 play an important role in mediating allergicreactions, which can promote the induction of IgE syntheses and the development of mastcells [5]. Particularly, IL-4 is the initial cytokine secreted from mast cells after IgE/BSAstimulation and activates the secondary cytokines, such as IL-5, IL-6, and IL-13 [31,32].IL-5 is also known to have a specific action in the development, priming, and survivalof eosinophils [33]. IL-6 had been defined as a modulate the adaptive immune responseduring early T-cell activation and TNF-α, which is a major effector cytokine in allergicreaction and improves mediator expression and cytokine in mast cells [2,34]. So, inhibitionof these allergic cytokines and chemokines is one of the major indicators of relieved allergicresponses. Thus, we confirmed that the secretion of allergic cytokines such as IL-4, IL-5,IL-6, IL-13, and TNF-α were down-regulated by the pretreatment of DHB in IgE/BSA-stimulated BMCMCs. With these results, we predict that DHB exerts an antiallergic effectby downregulating the gene expression and/or the protein secretion of various allergiccytokines and a chemokine as well as degranulation in IgE/BSA-stimulated BMCMCs.
The expression of cytokines is regulated by several intracellular signaling pathways,especially NF-κB, a transcription factor that plays a central role in the induction of geneexpression and is considered as a promising target for the treatment of disease [5,35]. NF-κB represents a family of a prime transcription factors which regulate a variety of genesincluding different processes of the immune and inflammatory responses [36]. Activated

Mar. Drugs 2022, 20, 133 8 of 13
NF-κB signaling results in the nuclear localization of NF-κB p65 and production of inflam-matory cytokines (IL-1β, IL-6, and TNF-α) is increased. Especially, TNF-α activates theNF-κB signaling to amplify the response of inflammation [8]. Therefore, the inhibition ofNF-κB is a major mechanism in mediating antiallergic effect. DHB significantly reducedthe phosphorylation of IκBα and the translocation of cytoplasmic NF-κB p65 to the nu-cleus in IgE/BSA-stimulated BMCMCs. These results prove that DHB suppressed mastcell activation, cytokines production, and binding of the IgE to surface FcεRI via NF-κBsignaling pathway.
Finally, we examined the anti allergic effects of DHB in a localized IgE-mediatedallergic response based in vivo study. PCA reaction, which evaluate by increased vascularpermeability and ear swelling, is that responses through binding of the IgE and FcεRI inmast cells [10,20]. Therefore, this model is generally used as the allergic animal model.Treatment of DHB inhibited the intensity of Evans blue dye in the ears. Thus, our resultssuggest that DHB exerts antiallergic effects through suppression of mast cell activation.
4. Materials and Methods4.1. Materials
Minimum Essential Medium α (α-MEM), penicillin-streptomycin, fetal bovine serum(FBS), and dimethylsulfoxide (DMSO) were purchased from Thermo Fisher Scientific(Waltham, MA, USA). Pokeweed mitogen (PWM), 2-mercaptoethanol (2-ME), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), Evans blue reagent, mono-clonal anti-dinitrophenyl (DNP)-IgE (clone SPE-7), and β-actin antibody were purchasedfrom Sigma-Aldrich (St. Louis, MO, USA). DNP-BSA was purchased from LSL Japan Inc.(Tokyo, Japan). Primary and secondary antibodies were purchased from Cell SignalingTechnology Inc. (Danvers, MA, USA).
4.2. Purification of DHB from P. morrowii
DHB was purified from P. morrowii harvested from the coast of Jeju Island, accordingto the method described by Ko et al. (2019) [13]. Briefly, an 80% aqueous methanol crudeextract of P. morrowii dry powder was suspended in water and fractionated into chloroform.The chloroform fraction was further separated on a silica open column via stepwise elutionof chloroform and methanol mixtures of different ratios. The fractions with the highestbioactivity were pooled and further separated on a Sephadex LH-20 column with 100%methanol elution followed by reversed-phase (C18) HPLC purification (Alliance 2690;Waters Corp., Milford, MA, USA). The identity of the purified compound was confirmedusing NMR. The purified DHB was diluted using PBS (2000-fold) for biological experiments(Figure S1).
4.3. Preparation of BMCMCs
C57BL/6 male mice (8-week-old), reared under specific pathogen-free conditions, werepurchased from Orient Bio (Gwangju, Korea). BMCMCs were obtained from 8-week-oldmale C57BL/6 mice according to a previous method [10]. The cells were sub-cultured inα-MEM media (10% FBS, 1% penicillin-streptomycin) with 10% PWM-stimulated spleencell-conditioned medium (PWM-SCM) and 0.2% 2-ME once a week for 7 weeks; and above98% of the mast cells recognized as Giemsa-staining positive were used for the experi-ments. The institutional ethical committee has approved these experiments (Approval No.CNU IACUC-YS-2019-4) and was complying with “Principles of Laboratory Animal Care”guidelines (NIH publication No. 80-23, revised 1996).
4.4. Measurement of Cytotoxicity
The effect of DHB treatment on cell viability was evaluated using an MTT assay [9,10].Accordingly, BMCMCs (2 × 104/well) were treated with different concentrations of DHB(31.3 and 62.5 µg/mL) and incubated at 37 ◦C for 24 h. Subsequently, 15 µL of 5 mg/mLMTT solution was added, and the cells were further incubated for 4 h. The formazan crystals

Mar. Drugs 2022, 20, 133 9 of 13
formed within the cells were dissolved using DMSO, and the absorbance was measured at540 nm using a microplate reader (SpectraMax® M2/M2e, Sunnyvale, CA, USA).
4.5. Measurement of β-Hexosaminidase Release
To assess the inhibitory effect of DHB on mast cell degranulation, BMCMCs (2 × 105/well)were first incubated for 1 h with different DHB concentrations and sensitized using mono-clonal anti-dinitrophenyl (DNP)-IgE (clone SPE-7) (1 µg/mL) for 4 h [25,26]. BSA-Tyrode’sbuffer (0.1%) was used as a vehicle control where appropriate. Next, the BMCMCs sen-sitized with IgE were incubated for 1 h with the synthetic allergen, DNP-BSA, at 37 ◦C.After incubation, each supernatant was obtained, and the remaining cells were washedwith 500 µL of 0.1% BSA-Tyrode’s buffer. The cells were then lysed in 50 µL of 0.5% TritonX-100-Tyrode’s buffer for 20 min. The respective supernatants and cell lysates were mixedwith the substrate buffer (1.3 mg/mL 4-p-nitrophenyl-N-acetyl-β-D-glucosaminide and0.1 M sodium citrate, pH 4.5) for 1 h. The reaction was terminated by adding 0.2 M glycine(pH 10.7), and the absorbance of the mixtures was measured at 405 nm using a microplatereader. β-hexosaminidase release from BMCMCs was calculated according to previouslyreported methods [26].
4.6. RT-PCR Analysis
Reverse transcription polymerase chain reaction (RT-PCR) was carried out usingtotal RNA isolated from BMCMCs using Trizol reagent [37]. The cDNA was synthesizedusing PrimeScript RT Reagent Kit (TaKaRa Bio Inc., Shiga, Japan). PCR was carried outfor 35 cycles using the respective primers indicated in our previous study [9]. The PCRconditions were set as follows: denaturation at 94 ◦C for 5 min, annealing at 55–60 ◦Cfor 1 min, and an extension phase at 72 ◦C for 20 min in a TaKaRa PCR Thermal Cycler(TaKaRa Bio Inc., Otsu, Japan). The PCR amplification products were electrophoresed on1.5% ethidium bromide/agarose gels and visualized under UV transillumination (VilberLourmat, Marne la Uallee, France).
4.7. Measurement of Cytokines Levels
The BMCMCs (2 × 106/well) were incubated for 1 h with different DHB concentrationsand then sensitized with anti-DNP-IgE (1 µg/mL) for 4 h. Next, the sensitized BMCMCswere incubated in DNP-BSA at 37 ◦C for 24 h with a CO2 humidified atmosphere. The cellsuspension supernatants were analyzed using mouse cytokine ELISA kits (Biolegend Inc.,San Diego, CA, USA) according to the manufacturer’s instructions.
4.8. Flow Cytometric Analysis of Mast Cells Expressing FcεRI and FcεRI-IgE (IgE Binding)
To prepare the cells for the measurement of the surface expression levels of FcεRI,the BMCMCs (4 × 105/well) were incubated with DHB for 6 h at 37 ◦C. To measure thesurface expression levels of IgE- FcεRI binding, the cells (4 × 105/well) were reacted withDHB for 2 h and sensitized using DNP-IgE (1 µg/mL) for 4 h at 37 ◦C. The prepared cellswere blocked using anti-CD16/CD32 monoclonal antibodies (Thermo Fisher Scientific,Rockford, USA) for 20 min to prevent nonspecific binding of the antibodies. To detect theexpression of FcεRI, the cells were incubated with FITC-conjugated anti-FcεRI antibodies(clone: MAR-1; Thermo Fisher Scientific, Rockford, IL, USA); to detect the expression of IgE-bound FcεRI, the cells were incubated with FITC-conjugated anti-mouse IgE monoclonalantibodies (clone: 23G3; Thermo Fisher Scientific, Rockford, IL, USA). The fluorescentlystained cells were measured using flow cytometry (Beckman coulter, Brea, CA, USA).The total BMCMCs were gated on a forward scatter (FSC-A)/side scatter (SSC-A) plot toomit outliers.
4.9. Western Blot Analysis
The BMCMCs (2 × 105/well) were incubated with DHB for 1 h before anti-DNP-IgE (1 µg/mL) sensitization, which was carried out for 4 h. Indomethacin (I.M) was

Mar. Drugs 2022, 20, 133 10 of 13
used as a positive control. Then, the cells were treated with DNP-BSA (100 ng/mL) for30 min. Cytosolic and nuclear proteins were separately obtained from the cells using aNE-PER® nuclear and cytoplasmic extraction kit (Thermo Scientific, Waltham, MA, USA).The protein concentrations of the cell lysates were measured using a BCA protein assaykit (Thermo Scientific, Waltham, MA, USA) according to the manufacturer’s instructions.IκBα, phospho (p)- IκBα, p65, and p-p65 (1:1000 dilution, Cell Signaling Technology) andβ-actin (1:3000 dilutions, Cell Signaling Technology) were used as the primary antibodies,and the secondary antibodies used were HRP-conjugated anti-mouse IgG and anti-rabbitIgG (1:3000 dilution, Cell Signaling Technology). The densitometry analysis of the Westernblot results was performed using the ImageJ software to quantify the protein levels relativeto corresponding housekeeping proteins [10].
4.10. In Silico Docking of DHB with IgE and FcεRI
The binding energies and types of intermolecular interactions between DHB withthe active sites of IgE (PDB ID; 4GRG) and the IgE-FcεRI complex (PDB ID: 1F6A) wereanalyzed via molecular docking. The crystal structures of the respective proteins wereobtained from the Protein Data Bank (PDB, http://www.pdb.org, accessed on 5 July 2017).The ligand (DHB) structure was optimized to a stable geometry via Gaussian semiempiricalPM6 calculations using Gaussian 09 software. Ligand charge and torsional bonds wereanalyzed, and hydrogen atoms were added to both proteins and ligand before dockingsimulations. Flexible ligand docking and docking refinements were carried out usingthe CDOCKER plugin (default settings) in Accelrys Discovery Studio 3.5 (Accelrys, Inc.,San Diego, CA, USA), which utilizes a molecular dynamics simulated-annealing-basedalgorithm. Potential binding sites for DHB were identified in the known binding pockets ofthe proteins.
4.11. PCA Test
Balb/C male mice (8-week-old), reared under specific pathogen-free conditions, werepurchased from Orient Bio (Gwangju, Korea). The in vivo effects of DHB were analyzed ina PCA mouse model induced using DNP-IgE together with DNP-BSA. First, anti-DNP-IgE(500 ng) was intradermally injected into the dorsal skin of both ears. Then, using a 1 mLsyringe, DHB was topical applicated to the mice 2 h before the induction of anaphylaxis.Subsequently, the mice were intravenously treated with 30 µL of DNP-BSA (10 mg) salinesolution containing 4% Evans blue dye. The control group was treated with saline insteadof DHB. After 30 min, the mice were euthanized by anesthetizing them with isofluranefollowed by cervical dislocation, and skin tissues collected from the dorsal ear were soakedin 1 mL of formamide and left overnight (24 h) at 64 ◦C. The absorbance intensity at 620 nmwas taken as a measure of the amount of extravagated dye. Histological analysis wasconducted according to the method described previously [38]. The count of degranulatedmast cells in ear tissues was obtained as described previously [39].
4.12. Statistical Analysis
The data were compared one-way ANOVA and Duncan’s multiple range test using theSPSS Statistics V20 software package (SPSS, Chicago, IL, USA). The values are presented asthe means ± standard error (SE), and p values < 0.05 were considered significant.
5. Conclusions
In conclusion, the present findings demonstrated that DHB attenuated the antial-lergic response through the inhibition of the mast cell degranulation, FcεRI expression,the crosslinking of IgE to FcεRI, and allergic cytokine secretion via the NF-κB activa-tion in IgE/BSA-stimulated BMCMCs, and suppressed the allergic reactions in IgE/BSA-stimulation induced PCA model. DHB isolated from P. morrowii may be used as an agentfor functional foods and pharmaceuticals to suppress allergic reactions.

Mar. Drugs 2022, 20, 133 11 of 13
Supplementary Materials: The following are available online at: https://www.mdpi.com/article/10.3390/md20020133/s1, Figure S1: Effects of DHB on FcεRI expression and the crosslinking of IgEto FcεRI expression in IgE/BSA-stimulated BMCMCs. (A) cell surface FcεRI expression and (B) IgEbinding to FcεRI in BMCMCs, Figure S2: Effects of DHB on the mRNA expression levels of allergiccytokines and chemokine in IgE/BSA-stimulated BMCMCs. The expression levels of (A) cytokinesand (B) chemokine using RT-PCR analysis. Figure S3: The 1H (A) and 13 ◦C (B) NMR spectrumof DHB.
Author Contributions: Conceptualization, E.-A.K., E.-J.H. and G.A.; investigation, E.-A.K., E.-J.H.,J.K., I.P.S.F., J.-Y.O. and K.-N.K.; data curation—formal analysis, E.-A.K. and E.-J.H.; project admin-istration/oversight, G.A. and S.-J.H.; writing—original draft, E.-A.K. and E.-J.H.; writing—review,editing, and revision, G.A. and S.-J.H. All authors have read and agreed to the published version ofthe manuscript.
Funding: This research received no external funding.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Acknowledgments: This work was supported by a research grant from the Korea Institute ofOcean Science and Technology (PEA0022). And supported by Basic Science Research Programthrough the National Research Foundation of Korea (NRF) funded by the Ministry of Education(2017R1D1A1B04035921).
Conflicts of Interest: The authors declare no conflict of interest.
References1. Min, K.-D.; Yi, S.-J.; Kim, H.-C.; Leem, J.-H.; Kwon, H.-J.; Hong, S.; Kim, K.S.; Kim, S.-Y. Association between exposure to
traffic-related air pollution and pediatric allergic diseases based on modeled air pollution concentrations and traffic measures inSeoul, Korea: A comparative analysis. Environ. Health 2020, 19, 6. [CrossRef]
2. Dhakal, H.; Lee, S.; Choi, J.K.; Kwon, T.K.; Khang, D.; Kim, S.-H. Inhibitory effects of orientin in mast cell-mediated allergicinflammation. Pharmacol. Rep. 2020, 72, 1002–1010. [CrossRef] [PubMed]
3. Liu, F.-T.; Goodarzi, H.; Chen, H.-Y. IgE, Mast Cells, and Eosinophils in Atopic Dermatitis. Clin. Rev. Allergy Immunol. 2011,41, 298–310. [CrossRef] [PubMed]
4. Kalesnikoff, J.; Huber, M.; Lam, V.; Damen, J.E.; Zhang, J.; Siraganian, R.P.; Krystal, G. Monomeric IgE Stimulates SignalingPathways in Mast Cells that Lead to Cytokine Production and Cell Survival. Immunity 2001, 14, 801–811. [CrossRef]
5. Kim, M.-J.; Choi, Y.-A.; Lee, S.; Choi, J.K.; Kim, Y.-Y.; Kim, E.-N.; Jeong, G.-S.; Shin, T.-Y.; Jang, Y.H.; Kim, S.-H. Prunus serrulatavar. spontanea inhibits mast cell activation and mast cell-mediated anaphylaxis. J. Ethnopharmacol. 2019, 250, 112484. [CrossRef][PubMed]
6. Ye, J.; Piao, H.; Jiang, J.; Jin, G.; Zheng, M.; Yang, J.; Jin, X.; Sun, T.; Choi, Y.H.; Li, L.; et al. Polydatin inhibits mast cell-mediatedallergic inflammation by targeting PI3K/Akt, MAPK, NF-κB and Nrf2/HO−1 pathways. Sci. Rep. 2017, 7, 11895. [CrossRef][PubMed]
7. Ahn, G.; Amagai, Y.; Matsuda, A.; Kang, S.-M.; Lee, W.; Jung, K.; Oida, K.; Jang, H.; Ishizaka, S.; Matsuda, K.; et al. Dieckol, aphlorotannin ofEcklonia cava, suppresses IgE-mediated mast cell activation and passive cutaneous anaphylactic reaction. Exp.Dermatol. 2015, 24, 968–970. [CrossRef]
8. Wee, J.H.; Zhang, Y.-L.; Rhee, C.-S.; Kim, D.-Y. Inhibition of allergic response by intranasal selective NF-κB decoy oligodeoxynu-cleotides in a murine model of allergic rhinitis. Allergy Asthma Immunol. Res. 2017, 9, 61. [CrossRef]
9. Han, E.J.; Fernando, I.P.S.; Kim, E.-A.; Kim, J.; Jung, K.; Kim, S.-Y.; Cha, S.-H.; Kim, K.-N.; Heo, S.-J.; Ahn, G. 5-Bromo-3,4-dihydroxybenzaldehyde from Polysiphonia morrowii attenuate IgE/BSA-stimulated mast cell activation and passive cutaneousanaphylaxis in mice. Biochem. Pharmacol. 2020, 178, 114087. [CrossRef]
10. Han, E.J.; Kim, H.-S.; Sanjeewa, K.; Herath, K.; Jeon, Y.-J.; Jee, Y.; Lee, J.; Kim, T.; Shim, S.-Y.; Ahn, G. Eckol from Ecklonia cavaSuppresses Immunoglobulin E-mediated Mast Cell Activation and Passive Cutaneous Anaphylaxis in Mice (Running Title:Anti-Allergic Activity of Ecklonia cava). Nutrients 2020, 12, 1361. [CrossRef]
11. Park, J.H.; Lee, B.; Kim, H.K.; Kim, E.-Y.; Kim, J.-H.; Min, J.-H.; Kim, S.; Sohn, Y.; Jung, H.-S. Peimine inhibits the production ofproinflammatory cytokines through regulation of the phosphorylation of NF-κB and MAPKs in HMC-1 Cells. Pharmacogn. Mag.2017, 13, S359. [PubMed]
12. Han, E.J.; Shin, E.-J.; Kim, M.J.; Heo, S.-J.; Kim, E.-A.; Ahn, G. Anti-allergic Effect of Methanol Extract from Polysiphonia morrowiiHarveyin IgE/BSA-stimulated Bone Marrow-derived Cultured Mast Cells anda Passive Cutaneous Anaphylaxis Mouse Model. J.Chitin Chitosan 2019, 24, 99–104. [CrossRef]

Mar. Drugs 2022, 20, 133 12 of 13
13. Ko, E.-Y.; Heo, S.-J.; Cho, S.-H.; Lee, W.; Kim, S.-Y.; Yang, H.-W.; Ahn, G.; Cha, S.-H.; Kwon, S.-H.; Jeong, M.S.; et al. 3-Bromo-5-(ethoxymethyl)-1, 2-benzenediol inhibits LPS-induced pro-inflammatory responses by preventing ROS production anddownregulating NF-κB in vitro and in a zebrafish model. Int. Immunopharmacol. 2019, 67, 98–105. [CrossRef] [PubMed]
14. Cho, S.-H.; Heo, S.-J.; Yang, H.-W.; Ko, E.-Y.; Jung, M.S.; Cha, S.-H.; Ahn, G.; Jeon, Y.-J.; Kim, K.-N. Protective Effect of 3-Bromo-4,5-Dihydroxybenzaldehyde from Polysiphonia morrowii Harvey against Hydrogen Peroxide-Induced Oxidative Stress In Vitroand In Vivo. J. Microbiol. Biotechnol. 2019, 29, 1193–1203. [CrossRef]
15. Ko, S.-C.; Ding, Y.; Kim, J.; Ye, B.-R.; Kim, E.-A.; Jung, W.-K.; Heo, S.-J.; Lee, S.-H. Bromophenol (5-bromo-3,4-dihydroxybenzaldehyde)isolated from red alga Polysiphonia morrowii inhibits adipogenesis by regulating expression of adipogenic transcription factorsand AMP-activated protein kinase activation in 3T3-L1 adipocytes. Phytother. Res. 2018, 33, 737–744. [CrossRef] [PubMed]
16. Kang, N.-J.; Han, S.-C.; Kang, H.-J.; Ko, G.; Yoon, W.-J.; Kang, H.-K.; Yoo, E.-S. Anti-Inflammatory Effect of 3-Bromo-4,5-Dihydroxybenzaldehyde, a Component of Polysiphonia morrowii, In Vivo and In Vitro. Toxicol. Res. 2017, 33, 325–332. [CrossRef]
17. Kim, S.-Y.; Kim, S.R.; Oh, M.-J.; Jung, S.-J.; Kang, S.Y. In Vitro antiviral activity of red alga, Polysiphonia morrowii extract andits bromophenols against fish pathogenic infectious hematopoietic necrosis virus and infectious pancreatic necrosis virus. J.Microbiol. 2011, 49, 102–106. [CrossRef]
18. Choi, Y.K.; Ye, B.-R.; Kim, E.-A.; Kim, J.; Kim, M.-S.; Lee, W.W.; Ahn, G.-N.; Kang, N.; Jung, W.-K.; Heo, S.-J. Bis (3-bromo-4,5-dihydroxybenzyl) ether, a novel bromophenol from the marine red alga Polysiphonia morrowii that suppresses LPS-inducedinflammatory response by inhibiting ROS-mediated ERK signaling pathway in RAW 264.7 macrophages. Biomed. Pharmacother.2018, 103, 1170–1177. [CrossRef]
19. Kang, S.Y.; Lee, S.-Y.; Choi, J.-H.; Jung, S.-J. In vitro Anti-bacterial and Anti-scuticociliate Activities of Extract and Bromophenolsof the Marine Red Alga Polysiphonia morrowii with Structure-activity Relationships. Korean J. Fish. Aquat. Sci. 2014, 47, 45–51.[CrossRef]
20. Kono, R.; Nakamura, M.; Nomura, S.; Kitano, N.; Kagiya, T.; Okuno, Y.; Inada, K.-I.; Tokuda, A.; Utsunomiya, H.; Ueno, M.Biological and epidemiological evidence of anti-allergic effects of traditional Japanese food ume (Prunus mume). Sci. Rep. 2018,8, 11638. [CrossRef]
21. Fukuishi, N.; Murakami, S.; Ohno, A.; Yamanaka, N.; Matsui, N.; Fukutsuji, K.; Yamada, S.; Itoh, K.; Akagi, M. Does β-Hexosaminidase Function Only as a Degranulation Indicator in Mast Cells? The Primary Role of β-Hexosaminidase in Mast CellGranules. J. Immunol. 2014, 193, 1886–1894. [CrossRef] [PubMed]
22. Garman, S.C.; Wurzburg, B.A.; Tarchevskaya, S.S.; Kinet, J.-P.; Jardetzky, T.S. Structure of the Fc fragment of human IgE bound toits high-affinity receptor FcεRIα. Nature 2000, 406, 259–266. [CrossRef] [PubMed]
23. Šterbová, D.; Matejícek, D.; Vlcek, J.; Kubán, V. Combined microwave-assisted isolation and solid-phase purification proceduresprior to the chromatographic determination of phenolic compounds in plant materials. Anal. Chim. Acta 2004, 513, 435–444.[CrossRef]
24. Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623.[CrossRef] [PubMed]
25. Mahdi, J.; Al-Musayeib, N.; Mahdi, E.; Pepper, C. Pharmacological importance of simple phenolic compounds on inflammation,cell proliferation and apoptosis with a special reference to β-D-salicin and hydroxybenzoic acid. Eur. J. Inflamm. 2013, 11, 327–336.[CrossRef]
26. Tan, J.W.; Israf, D.A.; Harith, H.H.; Hashim, N.F.M.; Ng, C.H.; Shaari, K.; Tham, C.L. Anti-allergic activity of 2,4,6-trihydroxy-3-geranylacetophenone (tHGA) via attenuation of IgE-mediated mast cell activation and inhibition of passive systemic anaphylaxis.Toxicol. Appl. Pharmacol. 2017, 319, 47–58. [CrossRef] [PubMed]
27. Zhou, Z.; Zhang, Y.; Ding, X.-R.; Chen, S.-H.; Yang, J.; Wang, X.-J.; Jia, G.-L.; Chen, H.-S.; Bo, X.-C.; Wang, S.-Q. Protocatechuicaldehyde inhibits hepatitis B virus replication both in vitro and in vivo. Antivir. Res. 2007, 74, 59–64. [CrossRef]
28. Guo, R.H.; Park, J.U.; Jo, S.J.; Ahn, J.H.; Park, J.H.; Yang, J.Y.; Lee, S.S.; Park, M.J.; Kim, Y.R. Anti-allergic Inflammatory Effects ofthe Essential Oil From Fruits of Zanthoxylum coreanum Nakai. Front. Pharmacol. 2018, 9, 1441. [CrossRef]
29. Vo, T.-S.; Ngo, D.-H.; Kim, S.-K. Marine algae as a potential pharmaceutical source for anti-allergic therapeutics. Process Biochem.2012, 47, 386–394. [CrossRef]
30. Williams, C.M.; Galli, S.J. The diverse potential effector and immunoregulatory roles of mast cells in allergic disease. J. AllergyClin. Immunol. 2000, 105, 847–859. [CrossRef]
31. Gessner, A.; Mohrs, K.; Mohrs, M. Mast Cells, Basophils, and Eosinophils Acquire Constitutive IL-4 and IL-13 Transcripts duringLineage Differentiation That Are Sufficient for Rapid Cytokine Production. J. Immunol. 2005, 174, 1063–1072. [CrossRef]
32. McLeod, J.J.; Baker, B.; Ryan, J.J. Mast cell production and response to IL-4 and IL-13. Cytokine 2015, 75, 57–61. [CrossRef]33. Robinson, D.S. Th-2 cytokines in allergic disease. Br. Med. Bull. 2000, 56, 956–968. [CrossRef] [PubMed]34. A Neveu, W.; Allard, J.L.; Raymond, D.M.; Bourassa, L.M.; Burns, S.M.; Bunn, J.Y.; Irvin, C.G.; Kaminsky, D.A.; Rincon, M.
Elevation of IL-6 in the allergic asthmatic airway is independent of inflammation but associates with loss of central airwayfunction. Respir. Res. 2010, 11, 28. [CrossRef]
35. Debnath, T.; Lee, Y.M.; Lim, J.H.; Lim, B.O. Anti-allergic and anti-atopic dermatitis effects of Gardenia Fructus extract. Food Agric.Immunol. 2018, 29, 665–674. [CrossRef]
36. Sohn, E.-H.; Jang, S.-A.; Joo, H.; Park, S.; Kang, S.-C.; Lee, C.-H.; Kim, S.-Y. Anti-allergic and anti-inflammatory effects of butanolextract from Arctium Lappa L. Clin. Mol. Allergy 2011, 9, 4. [CrossRef]

Mar. Drugs 2022, 20, 133 13 of 13
37. Syafni, N.; Putra, D.P.; Arbain, D. 3,4-dihydroxybenzoic acid and 3,4-dihydroxybenzaldehyde from the fern Trichomanes ChinenseL.; Isolation, antimicrobial and antioxidant Properties. Indones. J. Chem. 2012, 12, 273–278. [CrossRef]
38. Jung, K.; Tanaka, A.; Fujita, H.; Matsuda, A.; Oida, K.; Karasawa, K.; Okamoto, N.; Ohmori, K.; Jee, Y.; Shin, T.; et al. Peroxisomeproliferator–activated receptor γ–mediated suppression of dendritic cell function prevents the onset of atopic dermatitis inNC/Tnd mice. J. Allergy Clin. Immunol. 2011, 127, 420–429.e6. [CrossRef]
39. Kim, D.K.; Kim, H.S.; Kim, A.-R.; Kim, J.H.; Kim, B.; Noh, G.; Kim, H.S.; Beaven, M.A.; Kim, Y.M.; Choi, W.S. DJ-1 regulates mastcell activation and IgE-mediated allergic responses. J. Allergy Clin. Immunol. 2012, 131, 1653–1662.e1. [CrossRef]
Copyright © 2022 FDOKUMEN
















![1,10Dihydro3,9-dimethylpyrazolo[3,4- a ]carbazole](https://static.fdokumen.com/doc/165x107/63225bc364690856e1092115/110dihydro39-dimethylpyrazolo34-a-carbazole.jpg)



