
Determination of Natural and Artificial Radionuclides in Moss ...
Upload
independentCategory
view
2download
0
BRIEF COMMUNICATION
doi:10.1111/j.1558-5646.2009.00787.x
AND IF ENGLER WAS NOT COMPLETELYWRONG? EVIDENCE FOR MULTIPLEEVOLUTIONARY ORIGINS IN THE MOSS FLORAOF MACARONESIADelphine A. Aigoin,1,2,3 Nicolas Devos,1,4 Sanna Huttunen,5,6 Michael S. Ignatov,7,8
Juana M. Gonzalez-Mancebo,9,10 and Alain Vanderpoorten1,11
1Institute of Botany, University of Liege, 27 Blvd du Rectorat, B22, Sart Tilman, 4000 Liege, Belgium2E-mail: [email protected]
3Institute of Evolutionary Sciences, University of Montpellier II, Place Eugene Bataillon, 34095 Montpellier Cedex 5, France4E-mail: [email protected]
5Laboratory of Genetics, Department of Biology, University of Turku, 20014 Turku, Finland6E-mail: [email protected]
7Main Botanical Garden of Russian Academy of Sciences, Botanicheskaya 4, 127276 Moscow, Russia8E-mail: [email protected]
9Department of Botany, University of La Laguna, 38271 La Laguna, Tenerife, Canary Islands, Spain10E-mail: [email protected]: [email protected]
Received March 27, 2009
Accepted June 23, 2009
The Macaronesian endemic flora has traditionally been interpreted as a relict of a subtropical element that spanned across Europe
in the Tertiary. This hypothesis is revisited in the moss subfamily Helicodontioideae based on molecular divergence estimates
derived from two independent calibration techniques either employing fossil evidence or using an Monte Carlo Markov Chain
(MCMC) to sample absolute rates of nucleotide substitution from a prior distribution encompassing a wide range of rates docu-
mented across land plants. Both analyses suggest that the monotypic Madeiran endemic genus Hedenasiastrum diverged of other
Helicodontioideae about 40 million years, that is, well before Macaronesian archipelagos actually emerged, in agreement with
the relict hypothesis. Hedenasiastrum is characterized by a plesiomorphic morphology, which is suggestive of a complete morpho-
logical stasis over 40 million years. Macaronesian endemic Rhynchostegiella species, whose polyphyletic origin involves multiple
colonization events, evolved much more recently, and yet accumulated many more morphological novelties than H. percurrens.
The Macaronesian moss flora thus appears as a complex mix of ancient relicts and more recently dispersed, fast-evolving taxa.
KEY WORDS: Bryophytes, fossils, macronesian endemism, molecular dating, morphological evolution.
Macaronesia is a string of North Atlantic volcanic islands (the
Azores, Madeira, Canaries, and Cape Verde) that emerged 0.4–
20 million years ago and are characterized by high rates of en-
demism (see Juan et al. 2000 for review). Engler (1879), fol-
lowed by many biogeographers (see Vanderpoorten et al. 2007
for review), proposed that Macaronesian endemics are the relics
3 2 4 8C© 2009 The Author(s). Journal compilation C© 2009 The Society for the Study of Evolution.Evolution 63-12: 3248–3257

BRIEF COMMUNICATION
of biota that were widespread across Europe during the Tertiary
and decimated on the continent during the glaciations. In con-
trast with the expectations of the refugium concept, however,
several lines of evidence from analyses of moss species dis-
tributions (Vanderpoorten et al. 2007) and molecular evolution
rates in angiosperms (Carine 2005) have recently questioned En-
gler’s hypothesis. In mosses, the hypothesis of extinction of a
Tertiary flora in all but Macaronesian areas is solely supported
by extremely limited fossil evidence in the genus Echinodium
(Frahm 2004). This interpretation is, however, weakened by the
polyphyletic origin of the genus (Stech et al. 2008), thereby
raising doubts about the actual sister relationship between fos-
sil and extant Macaronesian Echinodium species. By contrast,
the nested phylogenetic position of the Azorean endemic liver-
wort Leptoscyphus azoricus within a Neotropical clade (Devos
and Vanderpoorten 2009); the close biogeographic affinities of
several Macaronesian groups with the North and South Ameri-
can continents (see Vanderpoorten et al. [2007] for review); and
the polyphyletic origin among Macaronesian populations of the
moss Grimmia montana, which are genetically identical or most
closely related to those of different continents (Vanderpoorten
et al. 2008) all point to recurrent migrations between the latter
and the Macaronesian archipelagos, potentially followed by in situ
speciation.
The continental extinction and recent speciation hypothe-
ses make different predictions regarding the rates of speciation
and morphological evolution. In fact, as opposed to neoendemics,
which originated from local speciation processes and often under-
went spectacular adaptive radiations involving a sudden burst of
morphological diversification (see Gillepsie and Roderick [2002]
for review), paleoendemics, which survived continental extinc-
tions on the islands, have most often retained a highly conserved
morphology for millions, or tens of million years, to such an extent
that extant taxa appear conspecific with fossil species (Sunding
1979).
In this article, we revisit hypotheses on the origin of Mac-
aronesian endemism in the mosses from the Brachytheciaceae,
subfamily Helicodontioideae. This group includes several Mac-
aronesian endemics, namely the monotypic Madeiran Hedenasi-
astrum percurrens, and three Rhynchostegiella species: Rhyn-
chostegiella bourgeana and R. trichophylla, which are restricted
to the Canary Islands, and R. macilenta, whose distributions
span Madeira and the Canaries. In addition, and unlike most
bryophytes, a fairly well-documented fossil record is available
(Miller 1984). We produced a molecular phylogeny of the Heli-
codontioideae to test the hypothesis of a radiation within Rhyn-
chostegiella and date the origin of its Macaronesian endemic lin-
eages, contrasting the results derived from a fossil calibration and
an analysis employing absolute rates of molecular evolution. We
then used the phylogeny to test the hypothesis of a long mor-
phological stasis since the Tertiary period among Macaronesian
endemics.
Material and MethodsTAXONOMIC AND MORPHOLOGICAL CHARACTER
SAMPLING
The 17 genera of Helicodontioideae as circumscribed by Aigoin
et al. (2009) were sampled (Table 1). Each genus was represented
by one to three species, with a special emphasis on Rhynchoste-
giella, for which all eight species were sampled. Three other
species, namely R. papuensis, R. leptoneura, and R. muriculata,
clearly do not belong to the genus, and the appropriate taxonomic
changes will be presented elsewhere. Aerobryidium filamentosum,
a species of the sister family Meteroriaceae, as well as representa-
tive taxa of each of the three subfamilies of Brachytheciaceae was
sampled as outgroups (Table 1). Forty-three morphological char-
acters that are variable across the Helicodontioideae, including
34 gametophytic and nine sporophytic ones, were scored for each
of the sampled taxa (Appendices S1 and S2) (see Huttunen and
Ignatov [2004] for a thorough account on character significance
and description).
MOLECULAR PROTOCOLS AND PHYLOGENETIC
ANALYSES
Four chloroplast regions (trnL-trnF, atpB-rbcL, psbT-psbH, and
psbA-trnH) were selected for exhibiting the appropriate level of
variation at the genus level in the Brachytheciaceae (Huttunen and
Ignatov 2004). DNA extraction, PCR and sequencing protocols,
sequence editing, alignment, indel scoring, and selection of mod-
els for DNA substitutions and indel evolution follow Aigoin et al.
(2009). Phylogenetic reconstruction was conducted with MrBayes
3.1.2. Four independent Monte Carlo Markov Chains (MCMCs)
of 2,000,000 iterations each were run and trees and model param-
eters were sampled every 10,000 generations. The convergence
of the MCMCs was verified visually from the likelihood values,
and trees of the “burn-in” were discarded.
A significant departure of alternative topologies involving a
monophyletic origin of the three Macaronesian endemic Rhyn-
chostegiella species, namely R. bourgeana, R. macilenta, and R.
trichophylla, was tested by constrained analyses. The MCMC
analysis described above was rerun under the constraint that only
trees fitting with a monophyletic origin of the Macaronesian en-
demic Rhynchostegiella species were sampled. We then deter-
mined whether the constraint induced a significant loss of like-
lihood by means of the Bayes factors, as assessed by twice the
difference in the log marginal likelihood between the two runs.
MOLECULAR DATING
Times of divergence were calculated to determine the origin
of the most recent common ancestor (hereafter, MRCA) of H.
EVOLUTION DECEMBER 2009 3 2 4 9

BRIEF COMMUNICATION
Ta
ble
1.
Taxo
nsa
mp
ling
,vo
uch
erin
form
atio
n(f
or
seq
uen
ces
pro
du
ced
for
the
pre
sen
tst
ud
y),a
nd
Gen
Ban
kac
cess
ion
nu
mb
ers.
Spec
ies
trnL
/trn
Fat
pB/r
bcL
psbT
/psb
Hps
bA/t
rnH
Vou
cher
spec
imen
for
sequ
ence
s
Aer
obry
idiu
mfil
amen
tosu
m(H
ook.
)M
.Fle
isch
.A
F397
789
–A
F417
347
–A
erol
indi
gia
capi
llac
ea(H
orns
ch.)
M.M
enze
lFJ
2624
14FJ
2624
41FJ
2624
74FJ
2624
99E
cuad
or,T
oapa
nta
&C
aran
qui1
437
(MO
)B
rach
ythe
cias
trum
coll
inum
(Sch
leic
h.ex
Mul
l.H
al.)
AY
1847
76A
Y66
3296
AY
1847
57–
Igna
tov
and
Hut
tune
nB
rach
ythe
cium
sale
bros
um(H
offm
.ex
F.W
eber
and
AF3
9785
7A
Y66
3309
AF4
1744
8A
Y31
2896
D.M
ohr)
Schi
mp.
Bry
hnia
nova
e-an
glia
e(S
ull.
and
Les
q.)
Gro
utA
F161
122
AJ2
8839
7A
F417
405
–B
ryoa
nder
soni
ail
lece
bra
(Hed
w.)
H.R
ob.
AF3
9781
9–
AF4
1736
5FJ
2625
01U
SA,B
ower
s22
214
(MA
)C
irri
phyl
lum
cras
sine
rviu
m(T
aylo
r)L
oesk
ean
dM
.Fle
isch
.FJ
2624
15FJ
2624
43FJ
2624
76FJ
2625
02Fr
ance
,Van
derp
oort
en41
3(L
G)
Cir
riph
yllu
mko
pone
nii(
Igna
tov)
Igna
tov
and
Hut
tune
nFJ
2624
16FJ
2624
44A
F417
446
FJ26
2503
Papu
aN
ewG
uine
a,K
opon
en32
122
(H)
Cir
riph
yllu
mpi
life
rum
(Hed
w.)
Gro
utFJ
2624
17FJ
2624
45A
F417
403
FJ26
2504
Finl
and,
Kop
onen
&H
uttu
nen
1324
(H)
Cla
smat
odon
parv
ulus
(Ham
pe)
Sull.
FJ26
2418
FJ26
2446
FJ26
2477
FJ26
2505
USA
,Van
derp
oort
en47
48(L
G)
Don
rich
ards
iam
acro
neur
on(G
rout
)H
.A.C
rum
and
L.E
.And
erso
nA
Y00
9848
AF3
2232
3–
–D
onri
char
dsia
patu
lifo
lia
(Car
dota
ndT
her.)
Igna
tov
and
Hut
tune
nFJ
2624
19FJ
2624
47FJ
2624
78FJ
2625
06C
hina
,Kop
onen
etal
.539
20(H
)E
urhy
nchi
astr
umpu
lche
llum
(Hed
w.)
Igna
tov
and
Hut
tune
nFJ
2624
20FJ
2624
48FJ
2624
79FJ
2625
07Fr
ance
,Sot
iaux
&So
tiau
x14
670
(her
b.So
tiaux
)E
urhy
nchi
ella
zeyh
eri(
Spre
ng.e
xM
ull.
Hal
.)M
.Fle
isch
.FJ
2624
21FJ
2624
49FJ
2624
80FJ
2625
30So
uth
Afr
ica,
Hyl
ande
rB
7851
8(S
)H
eden
asia
stru
mpe
rcur
rens
(Hed
enas
)Ig
nato
van
dV
ande
rpoo
rten
–FJ
2624
42FJ
2624
75FJ
2625
00M
adei
ra,H
eden
asB
4288
3(S
)H
elic
odon
tium
capi
llar
e(H
edw
.)A
.Jae
ger
FJ26
2422
FJ26
2450
FJ26
2481
FJ26
2512
Ven
ezue
la,V
ande
rpoo
rten
V18
0(L
G)
Hom
alot
heci
ella
subc
apil
lata
(Hed
w.)
Bro
th.
FJ26
2423
FJ26
2451
F417
462
FJ26
2513
U.S
.A.,
All
en&
Red
fear
nsn
(NY
)H
omal
othe
cium
fulg
esce
ns(M
itt.e
xM
ull.
Hal
.)A
.Jae
ger
AF3
9787
7E
F530
978
AF4
1746
6–
Met
eori
dium
rem
otif
oliu
m(M
ull.
Hal
.)M
ull.
Hal
.A
Y30
6783
–A
F417
418
FJ26
2514
Suri
nam
e,N
ewto
n43
99(B
M)
Mic
roeu
rhyn
chiu
mpu
mil
um(W
ilson
)Ig
nato
van
dV
ande
rpoo
rten
FJ26
2434
FJ26
2466
FJ26
2493
FJ26
2509
Fran
ce,V
ande
rpoo
rten
4415
(LG
)O
kam
urae
abr
achy
dict
yon
(Car
dot)
Nog
.FJ
2624
24FJ
2624
52A
Y18
4771
FJ26
2515
Chi
na,K
opon
enet
al.4
8969
(H)
Oxy
rrhy
nchi
umhi
ans
(Hed
w.)
Loe
ske
FJ26
2425
FJ26
2453
FJ26
2482
FJ26
2508
Fran
ce,V
ande
rpoo
rten
401
(LG
)O
xyrr
hync
hium
sava
tier
i(Sc
him
p.ex
Bes
ch.)
Bro
th.
AF3
9785
9FJ
2624
54A
F397
859
FJ26
2510
Chi
na,K
opon
enet
al.5
1775
(H)
Oxy
rrhy
nchi
umva
gans
(A.J
aege
r)Ig
nato
van
dH
uttu
nen
AF3
9786
2FJ
2624
55A
F417
450
FJ26
2511
Chi
na,K
opon
enet
al.4
9717
(H)
Pla
tyhy
pnid
ium
aust
rinu
m(H
ook.
f.an
dW
ilson
)M
.Fle
isch
.A
Y18
4791
–FJ
2624
83FJ
2625
16A
ustr
alia
,Str
eim
ann
4954
4(H
)P
laty
hypn
idiu
mri
pari
oide
s(H
edw
.)D
ixon
AY
8575
73A
Y85
7595
––
Pse
udor
hync
host
egie
lla
duri
aei(
Mon
t.)Ig
nato
van
dV
ande
rpoo
rten
FJ26
2428
FJ26
2459
FJ26
2486
FJ26
2519
Mad
eira
,Hed
enas
B90
50(S
)P
seud
oscl
erop
odiu
mpu
rum
(Hed
w.)
M.F
leis
ch.
AF3
9779
7A
F233
567
AF4
1747
0–
Rem
yell
abr
achy
podi
a(M
.Fle
isch
.)Ig
nato
van
dH
uttu
nen
AF3
9785
4FJ
2624
56A
F417
423
–Pa
pua
New
Gui
nea,
Kop
onen
3300
7(H
)R
hync
host
egie
lla
bour
gean
a(M
itt.)
Bro
th.
FJ26
2426
FJ26
2457
FJ26
2484
FJ26
2517
ElH
ierr
o,D
irks
e18
82(L
)R
hync
host
egie
lla
curv
iset
a(B
rid.
)L
impr
.FJ
2624
27FJ
2624
58FJ
2624
85FJ
2625
18Fr
ance
,Van
derp
oort
en42
1(L
G)
Rhy
ncho
steg
iell
aho
lsti
i(B
roth
.)B
roth
.FJ
2624
29FJ
2624
60FJ
2624
87FJ
2625
20So
uth
Afr
ica,
Hyl
ande
rB
7853
4(S
)R
hync
host
egie
lla
lept
oneu
raC
ardo
tFJ
2624
31FJ
2624
62FJ
2624
89FJ
2625
22C
hina
,He
3607
4(M
O)
Rhy
ncho
steg
iell
ali
tore
a(D
eN
ot.)
Lim
pr.
FJ26
2432
FJ26
2463
FJ26
2490
FJ26
2523
Mad
eira
,Hed
enas
B90
57(S
)
Co
nti
nu
ed.
3 2 5 0 EVOLUTION DECEMBER 2009

BRIEF COMMUNICATION
Ta
ble
1.
Co
nti
nu
ed.
Spec
ies
trnL
/trn
Fat
pB/r
bcL
psbT
/psb
Hps
bA/t
rnH
Vou
cher
spec
imen
for
sequ
ence
s
Rhy
ncho
steg
iell
am
acil
enta
(Ren
auld
and
Car
dot)
Car
dot
FJ26
2433
FJ26
2464
FJ26
2491
FJ26
2524
Mad
eira
,Hed
enas
B45
03(S
)R
hync
host
egie
lla
mur
icul
ata
(Hoo
k.f.
and
Wils
on)
Bro
th.
DQ
2082
14FJ
2624
65FJ
2624
92FJ
2625
25A
ustr
alia
,Str
eim
ann
4962
8(H
)R
hync
host
egie
lla
papu
ensi
sE
.B.B
artr
am–
–A
F417
439
–R
hync
host
egie
lla
tene
lla
(Dic
ks.)
Lim
pr.
FJ26
2436
FJ26
2468
FJ26
2495
FJ26
2527
Rus
sia,
Igna
tov
&Ig
nato
va05
-614
3(M
W)
Rhy
ncho
steg
iell
ate
neri
ffae
(Mon
t.)D
irks
ean
dB
oum
an#1
FJ26
2430
FJ26
2461
FJ26
2488
FJ26
2521
Mad
eira
,Hed
enas
B90
96(S
)R
hync
host
egie
lla
tene
riffa
e(M
ont.)
Dir
kse
and
Bou
man
#2FJ
2624
37FJ
2624
69FJ
2624
96FJ
2625
28Fr
ance
,Van
derp
oort
en36
2(L
G)
Rhy
ncho
steg
iell
ate
neri
ffae
(Mon
t.)D
irks
ean
dB
oum
an#3
FJ26
2435
FJ26
2467
FJ26
2494
FJ26
2526
Switz
erla
nd,H
eden
asB
1189
5(S
)R
hync
host
egie
lla
tric
hoph
ylla
Dir
kse
and
Bou
man
FJ26
2438
FJ26
2470
FJ26
2497
FJ26
2529
Gra
nC
anar
ia,D
irks
e13
843
(L)
Rhy
ncho
steg
ium
psil
opod
ium
Igna
tov
and
Hut
tune
nFJ
2624
39FJ
2624
71FJ
2624
98FJ
2625
31C
hina
,Kop
onen
etal
.518
03Sc
orpi
uriu
mci
rcin
atum
(Bri
d.)
M.F
leis
ch.a
ndL
oesk
eA
F397
834
FJ26
2472
AF4
1741
0–
Fran
ce,V
ande
rpoo
rten
M16
(LG
)Sq
uam
idiu
mbr
asil
iens
e(H
orns
ch.)
Bro
th.
FJ26
2440
FJ26
2473
AF4
1739
3FJ
2625
32Ta
nzan
ia,P
ocs
etal
.881
61(R
)Z
elom
eteo
rium
patu
lum
(Hed
w.)
Man
uel
AF3
9778
7–
AF4
1736
2–
percurrens and of the three Macaronesian endemics of Rhyn-
chostegiella. Examination of rate variation among branches of the
phylogeny suggested a strong departure of the sequence data from
the molecular clock (see below). Divergence times were therefore
estimated using a Bayesian MCMC method under a relaxed-clock
model employing an uncorrelated lognormal model of rate vari-
ation among branches in the tree as implemented by BEAST
1.4.8. Nonequivocal Miocene fossils of Cirriphyllum piliferum
and Oxyrrhynchium hians, and Pliocene fossils of Clasmatodon
parvulus (Miller 1984) were used for calibration at three internal
nodes. In bryophytes, wherein the probability of fossilization is
very low owing to the absence of lignified tissues and much re-
duced cuticle, the presence of a species is likely to be recorded
in the sediment only well after its actual phylogenetic origin. We
therefore used asymmetric distribution priors with a mode at the
beginning of the period considered and asymptotically skewed
toward the end of it (Fig. 1).
In a second set of analyses, absolute nucleotide substitu-
tion rates documented for a large number of plant lineages (in-
cluding bryophytes) were used. Accordingly, a normal distribu-
tion, with a mean and standard deviation of 5.0 × 10−10 and
10−10 substitutions/site/year, respectively, which corresponds to
the average absolute substitution rate of cpDNA across land plants
and largely encompasses their variation range (see Schnabel and
Wendel [1998] for review), was used as a prior on the absolute
rates of evolution for the four cpDNA regions combined.
Although BEAST allows for topologies, branch lengths, and
rates to be coestimated, we chose to perform the dating analyses
onto topologies that were derived independently from the kind of
calibration used. Therefore, we constrained the topology to match
the 50% majority-rule consensus of the trees sampled from the
posterior probability distribution generated by MrBayes, but used
the MCMC implemented by BEAST to sample branch lengths
and substitution rates depending on their posterior probabilities.
A Yule prior on branching rates was employed and two indepen-
dent MCMC analyses were each run for 10,000,000 steps. Param-
eter values were sampled every 1000 cycle over the 10,000,000
MCMC steps. Convergence and acceptable mixing of the sam-
pled parameters was checked using Tracer 1.4. After discarding
the burn-in steps, the two runs were combined to obtain an es-
timate of the posterior probability distribution of the divergence
dates of the ancestral nodes.
ANCESTRAL CHARACTER STATE RECONSTRUCTIONS
Ancestral character state reconstructions were performed to con-
trast the hypotheses that Macaronesian endemics are characterized
by plesiomorphic or derived character states. All reconstructions
were performed after pruning the outgroups from the trees. The
probabilities of a change in a branch were calculated by esti-
mating the instantaneous forward and backward rates among the
EVOLUTION DECEMBER 2009 3 2 5 1
BRIEF COMMUNICATION
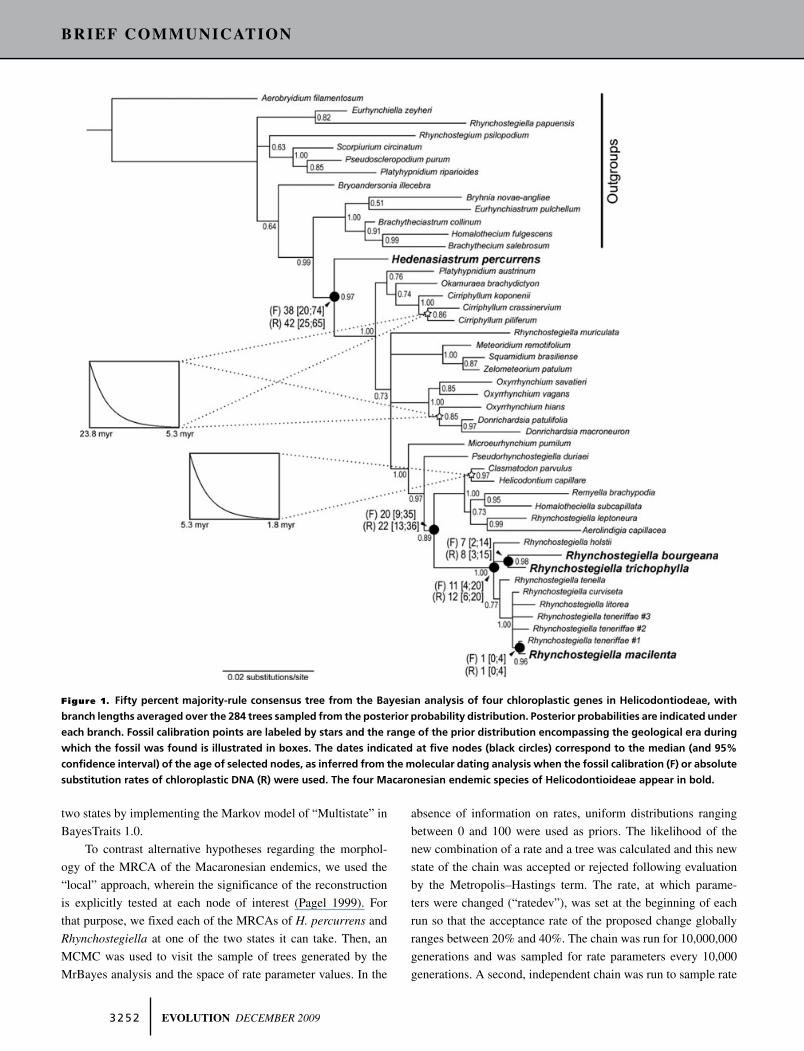
Figure 1. Fifty percent majority-rule consensus tree from the Bayesian analysis of four chloroplastic genes in Helicodontiodeae, with
branch lengths averaged over the 284 trees sampled from the posterior probability distribution. Posterior probabilities are indicated under
each branch. Fossil calibration points are labeled by stars and the range of the prior distribution encompassing the geological era during
which the fossil was found is illustrated in boxes. The dates indicated at five nodes (black circles) correspond to the median (and 95%
confidence interval) of the age of selected nodes, as inferred from the molecular dating analysis when the fossil calibration (F) or absolute
substitution rates of chloroplastic DNA (R) were used. The four Macaronesian endemic species of Helicodontioideae appear in bold.
two states by implementing the Markov model of “Multistate” in
BayesTraits 1.0.
To contrast alternative hypotheses regarding the morphol-
ogy of the MRCA of the Macaronesian endemics, we used the
“local” approach, wherein the significance of the reconstruction
is explicitly tested at each node of interest (Pagel 1999). For
that purpose, we fixed each of the MRCAs of H. percurrens and
Rhynchostegiella at one of the two states it can take. Then, an
MCMC was used to visit the sample of trees generated by the
MrBayes analysis and the space of rate parameter values. In the
absence of information on rates, uniform distributions ranging
between 0 and 100 were used as priors. The likelihood of the
new combination of a rate and a tree was calculated and this new
state of the chain was accepted or rejected following evaluation
by the Metropolis–Hastings term. The rate, at which parame-
ters were changed (“ratedev”), was set at the beginning of each
run so that the acceptance rate of the proposed change globally
ranges between 20% and 40%. The chain was run for 10,000,000
generations and was sampled for rate parameters every 10,000
generations. A second, independent chain was run to sample rate
3 2 5 2 EVOLUTION DECEMBER 2009

BRIEF COMMUNICATION
parameters and derive overall likelihoods of the reconstructions
when the node of interest was fixed at its alternative state. Bayes
factors were then used to determine the support for alternative
state at each node of interest.
ResultsPHYLOGENETIC RELATIONSHIPS
The chloroplast matrix contains 19% variable sites. The 50%
majority-rule consensus of the 284 trees sampled from the poste-
rior probability distribution is presented in Figure 1. Macaronesian
endemics within the Helicondontioideae appear at the two extrem-
ities of the phylogeny. The Madeiran endemic Hedenasiastrum
percurrens is sister to all other genera of the Helicodontioideae,
and this relationship is supported with a posterior probability
(hereafter, p.p.) of 1.00. By contrast, Rhynchostegiella is one of
the most recently diverging groups of species of the Helicodon-
tioideae. Within Rhynchostegiella, the sub-Saharan African R.
holstii is part of a polytomy with two other clades. The first clade
includes the Canarian endemics R. bourgeana and R. trichophylla
with a p.p. of 0.98. Within the second clade, the Macaronesian
endemic R. macilenta is resolved as sister to a Madeiran accession
of R. teneriffae with a p.p. of 0.96. The hypothesis of a mono-
phyletic origin of the three Macaronesian endemic species can be
significantly rejected. In fact, constraining all three Macaronesian
endemic species to monophyly resulted in a significantly lower
log marginal likelihood by comparison with the unconstrained
analysis (log marginal likelihood of −6884.07 and −6861.42 for
the constrained and unconstrained analyses, respectively).
MOLECULAR DIVERGENCE DATING
The coefficient of variation of rates among branches of the phy-
logeny was 0.63 (95% highest posterior density 0.44–0.83) for the
first analysis (fossil calibration) and 0.60 (95% highest posterior
density 0.43–0.77) for the second one (rate sampling from a prior
distribution). This suggests a strong departure of the data from a
molecular clock, a condition in which the coefficient of variation
equals zero. The two kinds of calibration used for molecular dat-
ing by a relaxed-clock model provided congruent results regard-
ing both the medians of the posterior probability distributions and
their 95% confidence intervals (Fig. 1). Hedenasiastrum percur-
rens emerged 38 [20;74] million years with the fossil calibration
and 42 [25;65] when absolute substitution rates were employed.
The divergence and diversification of Rhynchostegiella, respec-
tively, date back to 20 [9;35] and 11 [4;20] million years with
the fossil calibration and 22 [13;36] and 12 [6;20] with absolute
rates of molecular evolution. The MRCA of the Macaronesian
endemics R. bourgeana and R. trichophylla was dated back to 7
[2;14] million years with the fossil calibration and 8 [3;15] when
a distribution of absolute substitution rates was used as a prior,
whereas R. macilenta is estimated to have diverged more recently,
1 [0;4] million year according to the fossil calibration, and 1 [0;4]
when absolute rates were used (Fig. 1).
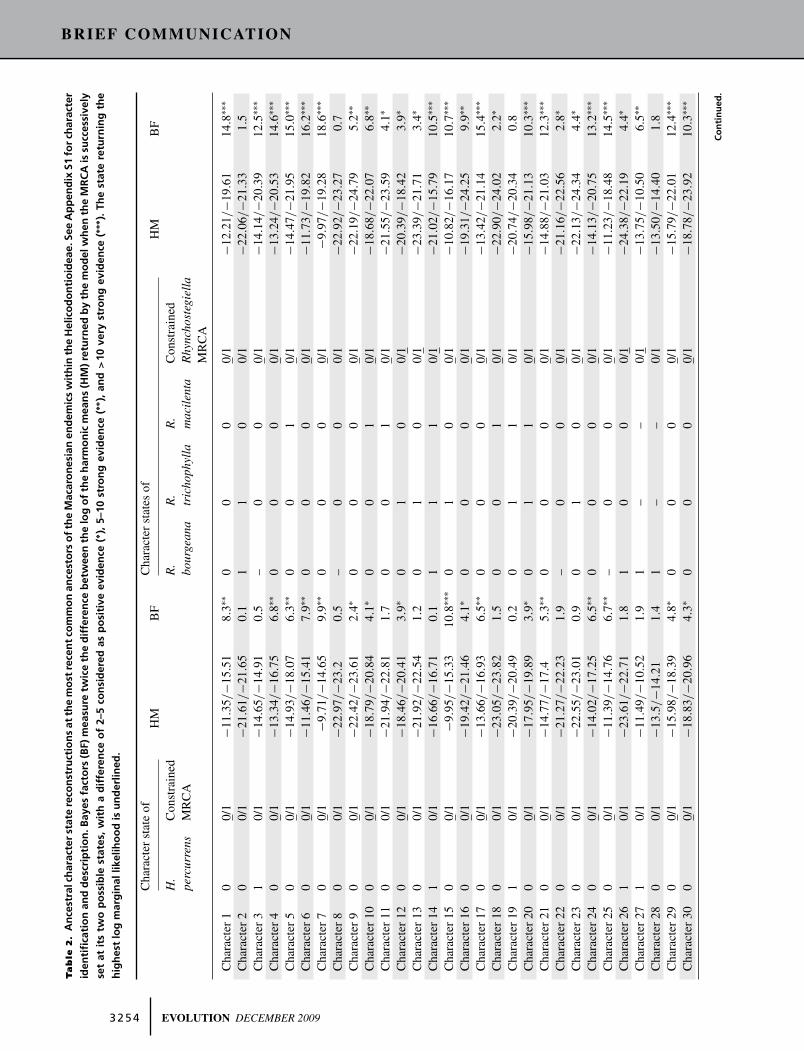
ANCESTRAL CHARACTER STATE RECONSTRUCTIONS
The marginal log-likelihoods of the reconstruction of ancestral
morphological character states at the MRCA of the Macaronesian
endemics are presented in Table 2. When the MRCA of the Heli-
codontioideae was constrained to be morphologically identical to
the Madeiran endemic H. percurrens, the marginal log-likelihoods
of the reconstruction were significantly higher for 20 of the 32
investigated gametophytic characters. For the remaining 12 char-
acters, the marginal log-likelihood obtained after fixing the root
of the Helicodontioideae at one or the alternative state were not
significantly different (i.e., the Bayes factors were <2), so that the
state at the root was ambiguous for those characters. Altogether,
the two Canarian endemics R. bourgeana and R. trichophylla dif-
fer from the MRCA of the genus by six character states (#12, 13,
15, 20, 23, 26). Finally, R. macilenta, which is the phylogeneti-
cally most recently diverging Macaronesian endemic within the
Helicodontioideae, differs from the MRCA of Rhynchostegiella
by nine character states (#5, 10, 11, 12, 13, 18, 20, 26, and 35).
DiscussionThe two calibration techniques used in the present study, that
is the use of fossils and the sampling of absolute substitution
rates from a prior distribution encompassing a wide range of
rates documented across land plants, rely on completely different
assumptions. Yet, they returned highly congruent results, which
strongly reinforces the confidence in the molecular dating in-
ferences. The MRCA of Rhynchostegiella dates back to about
10 million years. This timing is compatible with both a paleo-
and a neo-endemic origin of its Macaronesian endemic species.
The polyphyletic origin of the latter, however, does not lend sup-
port to Engler’ refugium hypothesis. Like in some angiosperms
(see Carine et al. [2004] for review), the colonization of Mac-
aronesia by Rhynchostegiella species seems to have involved at
least two dispersal events. Rhynchostegiella was one of the few
bryophyte genera, wherein several Macaronesian endemics had
been described, and therefore was one of the few candidates for
adaptive radiations. The polyphyletic origin of its Macaronesian
endemics invalidates that hypothesis, although the sister relation-
ship between R. bourgeana and R. trichophylla suggests that local
speciation may, to a minimal extent, have happened. Furthermore,
and although this needs to be confirmed by additional sampling,
the Macaronesian endemic R. macilenta is resolved as a sister to
a sympatric accession of R. teneriffae, rendering the latter species
paraphyletic. Such a situation has been recently increasingly de-
scribed in other bryophyte species and has been interpreted in
EVOLUTION DECEMBER 2009 3 2 5 3
BRIEF COMMUNICATION
Ta
ble
2.
An
cest
ralc
har
acte
rst
ate
reco
nst
ruct
ion
sat
the
mo
stre
cen
tco
mm
on
ance
sto
rso
fth
eM
acar
on
esia
nen
dem
ics
wit
hin
the
Hel
ico
do
nti
oid
eae.
See
Ap
pen
dix
S1fo
rch
arac
ter
iden
tifi
cati
on
and
des
crip
tio
n.B
ayes
fact
ors
(BF)
mea
sure
twic
eth
ed
iffe
ren
ceb
etw
een
the
log
of
the
har
mo
nic
mea
ns
(HM
)ret
urn
edb
yth
em
od
elw
hen
the
MR
CA
issu
cces
sive
ly
set
atit
stw
op
oss
ible
stat
es,
wit
ha
dif
fere
nce
of
2–5
con
sid
ered
asp
osi
tive
evid
ence
(∗),
5–10
stro
ng
evid
ence
(∗∗ )
,an
d>
10ve
ryst
ron
gev
iden
ce(∗
∗∗).
The
stat
ere
turn
ing
the
hig
hes
tlo
gm
arg
inal
likel
iho
od
isu
nd
erlin
ed.
Cha
ract
erst
ate
ofC
hara
cter
stat
esof
HM
BF
HM
BF
H.
Con
stra
ined
R.
R.
R.
Con
stra
ined
perc
urre
nsM
RC
Abo
urge
ana
tric
hoph
ylla
mac
ilen
taR
hync
host
egie
lla
MR
CA
Cha
ract
er1
00/
1−1
1.35
/−1
5.51
8.3∗∗
00
00/
1−1
2.21
/−1
9.61
14.8
∗∗∗
Cha
ract
er2
00/
1−2
1.61
/−2
1.65
0.1
11
00/
1−2
2.06
/−2
1.33
1.5
Cha
ract
er3
10/
1−1
4.65
/−1
4.91
0.5
–0
00 /
1−1
4.14
/−2
0.39
12.5
∗∗∗
Cha
ract
er4
00/
1−1
3.34
/−1
6.75
6.8∗∗
00
00/
1−1
3.24
/−2
0.53
14.6
∗∗∗
Cha
ract
er5
00/
1−1
4.93
/−1
8.07
6.3∗∗
00
10/
1−1
4.47
/−2
1.95
15.0
∗∗∗
Cha
ract
er6
00/
1−1
1.46
/−1
5.41
7.9∗∗
00
00/
1−1
1.73
/−1
9.82
16.2
∗∗∗
Cha
ract
er7
00/
1−9
.71/
−14.
659.
9∗∗0
00
0/1
−9.9
7/−1
9.28
18.6
∗∗∗
Cha
ract
er8
00/
1−2
2.97
/−2
3.2
0.5
–0
00/
1−2
2.92
/−2
3.27
0.7
Cha
ract
er9
00 /
1−2
2.42
/−2
3.61
2.4∗
00
00/
1−2
2.19
/−2
4.79
5.2∗∗
Cha
ract
er10
00/
1−1
8.79
/−2
0.84
4.1∗
00
10/
1−1
8.68
/−2
2.07
6.8∗∗
Cha
ract
er11
00/
1−2
1.94
/−2
2.81
1.7
00
10/
1−2
1.55
/−2
3.59
4.1∗
Cha
ract
er12
00/
1−1
8.46
/−2
0.41
3.9∗
01
00/
1−2
0.39
/−1
8.42
3.9∗
Cha
ract
er13
00/
1−2
1.92
/−2
2.54
1.2
01
00/
1−2
3.39
/−2
1.71
3.4∗
Cha
ract
er14
10/
1−1
6.66
/−1
6.71
0.1
11
10/
1−2
1.02
/−1
5.79
10.5
∗∗∗
Cha
ract
er15
00/
1−9
.95/
−15.
3310
.8∗∗
∗0
10
0/1
−10.
82/−1
6.17
10.7
∗∗∗
Cha
ract
er16
00/
1−1
9.42
/−2
1.46
4.1∗
00
00/
1−1
9.31
/−2
4.25
9.9∗∗
Cha
ract
er17
00/
1−1
3.66
/−1
6.93
6.5∗∗
00
00/
1−1
3.42
/−2
1.14
15.4
∗∗∗
Cha
ract
er18
00/
1−2
3.05
/−2
3.82
1.5
00
10/
1−2
2.90
/−2
4.02
2.2∗
Cha
ract
er19
10/
1−2
0.39
/−2
0.49
0.2
01
10/
1−2
0.74
/−2
0.34
0.8
Cha
ract
er20
00/
1−1
7.95
/−1
9.89
3.9∗
01
10/
1−1
5.98
/−2
1.13
10.3
∗∗∗
Cha
ract
er21
00/
1−1
4.77
/−1
7.4
5.3∗∗
00
00/
1−1
4.88
/−2
1.03
12.3
∗∗∗
Cha
ract
er22
00/
1−2
1.27
/−2
2.23
1.9
–0
00/
1−2
1.16
/−2
2.56
2.8∗
Cha
ract
er23
00/
1−2
2.55
/−2
3.01
0.9
01
00/
1−2
2.13
/−2
4.34
4.4∗
Cha
ract
er24
00/
1−1
4.02
/−1
7.25
6.5∗∗
00
00/
1−1
4.13
/−2
0.75
13.2
∗∗∗
Cha
ract
er25
00/
1−1
1.39
/−1
4.76
6.7∗∗
–0
00/
1−1
1.23
/−1
8.48
14.5
∗∗∗
Cha
ract
er26
10/
1−2
3.61
/−2
2.71
1.8
10
00/
1−2
4.38
/−2
2.19
4.4∗
Cha
ract
er27
10/
1−1
1.49
/−1
0.52
1.9
1–
–0/
1−1
3.75
/−1
0.50
6.5∗∗
Cha
ract
er28
00/
1−1
3.5/
−14.
211.
41
––
0/1
−13.
50/−1
4.40
1.8
Cha
ract
er29
00 /
1−1
5.98
/−1
8.39
4.8∗
00
00/
1−1
5.79
/−2
2.01
12.4
∗∗∗
Cha
ract
er30
00/
1−1
8.83
/−2
0.96
4.3∗
00
00/
1−1
8.78
/−2
3.92
10.3
∗∗∗
Co
nti
nu
ed.
3 2 5 4 EVOLUTION DECEMBER 2009

BRIEF COMMUNICATION
Ta
ble
2.
Co
nti
nu
ed.
Cha
ract
erst
ate
ofC
hara
cter
stat
esof
HM
BF
HM
BF
H.
Con
stra
ined
R.
R.
R.
Con
stra
ined
perc
urre
nsM
RC
Abo
urge
ana
tric
hoph
ylla
mac
ilen
taR
hync
host
egie
lla
MR
CA
Cha
ract
er31
00/
1−1
7.11
/−1
9.02
3.8∗
00
00/
1−1
6.87
/−2
2.17
10.6
∗∗∗
Cha
ract
er32
00/
1−1
0.21
/−1
4.69
9.0∗∗
00
00/
1−1
0.63
/−1
9.37
17.5
∗∗∗
Cha
ract
er33
–0/
1–
–0
00
0/1
−12.
19/−1
7.84
11.3
∗∗∗
Cha
ract
er34
–0/
1–
–0
00
0/1
−10.
22/−1
7.89
15.3
∗∗∗
Cha
ract
er35
–0/
1–
–0
01
0/1
−18.
08/−2
0.17
4.2∗
Cha
ract
er36
–0/
1–
–0
00
0/1
−11.
73/−1
7.15
10.8
∗∗∗
Cha
ract
er37
–0/
1–
–0
00
0/1
−14.
79/−1
8.41
7.2∗∗
Cha
ract
er38
–0/
1–
–0
00
0/1
−15.
89/−1
6.93
2.1∗
Cha
ract
er39
–0/
1–
–0
00
0/1
−12.
97/−1
6.73
7.5∗∗
Cha
ract
er40
–0/
1–
–0
00
0/1
−13.
20/−1
6.78
7.2∗∗
Cha
ract
er41
–0/
1–
–0
00
0/1
−11.
62/−1
5.76
8.3∗∗
Cha
ract
er42
–0/
1–
–0
00
0/1
−11.
85/−1
6.44
9.2∗∗
Cha
ract
er43
–0/
1–
–0
00
0/1
−10.
71/−1
7.63
13.8
∗∗∗ terms of recent speciation and incomplete lineage sorting (see
Sotiaux et al. [2009] for review), thereby reinforcing the idea of
a neoendemic origin of R. macilenta.
Although the polyphyletic origin and recent evolution of the
Macaronesian endemic Rhynchostegiella species is thus sugges-
tive of multiple colonization events followed by rather recent di-
vergence, the Madeiran endemic moss H. percurrens diverged
about 40 million years, that is well before Madeira actually
emerged, 5.2 million years. This time frame, together with the
fairly long branch leading to this unique species, is suggestive
of numerous extinctions and a relictual origin. Hedenasiastrum
percurrens actually becomes the only clear-cut documented case
of relict in the Macaronesian bryophyte flora. In the context of a
“dispersalist counter-revolution” fueled by the outcome of recent
molecular phylogenies and molecular dating studies (de Queiroz
2005), and of a reassessment of the recent origin of emblematic is-
land endemics (e.g., Goldberg et al. 2008; Grandcolas et al. 2008),
including Macaronesia (e.g., Jordal and Hewitt 2004; Emerson
and Oromı 2005; Dimitrov et al. 2008), the present study argues
for a pluralistic view of Macaronesian biogeography. Hedenasias-
trum percurrens thus falls within the increasingly limited category
of confirmed Tertiary relicts. The acceptance of a relictual origin
of H. percurrens, together with its morphological identity with
the MRCA of the Helicodontioideae, dated at 40 million years,
lends support to the traditional view of bryophytes as “unmoving
sphinxes of the past” (see Devos and Vanderpoorten [2009] for
review). In fact, the morphological similarity between extant and
fossil species suggests that some groups of mosses have persisted
with little morphological change for at least 80 million years
(see Devos and Vanderpoorten [2009] for review). The question
of whether this similarity is indeed due to the direct sister rela-
tionship or homoplasy remains, however, open. For example, the
oldest liverwort fossil, the Devonian Pallaviciinites devonicus, ex-
hibits a striking resemblance with the unrelated Pallavicinia and
Symphyogyna (Heinrichs et al. 2007), suggesting that the similar-
ity between fossil and extant taxa likely results from homoplasy.
Thus, although evidence for cryptic speciation in bryophytes
is mounting (see Devos and Vanderpoorten [2009] for review),
morphological stasis over periods of time of a range of tens of
million years actually appears as an extremely rare phenomenon.
In contrast, fast rates of morphological evolution or diversification
have been repeatedly reported in bryophytes (Shaw et al. 2003;
Wall 2005; Devos and Vanderpoorten 2009; Sotiaux et al. 2009).
In the case of H. percurrens, however, although character evolu-
tion might be constrained, so that all the space of morphologies
cannot be necessarily explored (Beldade et al. 2002), convergence
or parallelism occurring to develop what is the same taxon two
or more times is, arguably, less likely than plesiomorphy (Zander
2008). The retention of so many plesiomorphic traits during a 40-
million-year period is remarkable and can be compared to other
EVOLUTION DECEMBER 2009 3 2 5 5

BRIEF COMMUNICATION
spectacular examples of living fossils (see Lee et al. [2006] for
review). Surprisingly, and despite their much more recent origin,
all three Macaronesian endemic Rhynchostegiella species accu-
mulated many more morphological novelties than H. percurrens.
Although the remarkable stasis of H. percurrens might originate
from stabilizing selection, the reasons for such striking differences
in evolutionary rates within the same group of mosses remain
completely unexplained.
ACKNOWLEDGMENTSDA, ND, and AV acknowledge financial support from the Belgian Fundsfor Scientific Research (F.R.S.–FNRS) and the Leopold III Funds, andMI the RFBR 07-04-00013. Many thanks are due to M. Carine, P.-H.Fabre, R. G. Gillepsie, F. Kjellberg, and three anonymous reviewers fortheir constructive comments on an earlier draft of this article.
LITERATURE CITEDAigoin, D. A., S. Huttunen, M. S. Ignatov, G. M. Dirkse, and A.
Vanderpoorten. 2009. Rhynchostegiella (Brachytheciaceae): molecularre-circumscription of a convenient taxonomic repository. J. Bryol. Inpress.
Beldade, P., K. Koops, and P. M. Brakefield. 2002. Developmental constraintsversus flexibility in morphological evolution. Nature 416:844–847.
Carine, M. A. 2005. Spatio-temporal relationships of the Macaronesian en-demic flora: a relictual series or window of opportunity? Taxon 54:895–903.
Carine, M. A., S. J. Russel, A. Santos-Guerra, and J. Francisco-Ortega. 2004.Relationships of the Macaronesian and Mediterranean floras: molec-ular evidence for multiple colonizations into Macaronesia and back-colonization of the continent in Convolvulus (Convolvulaceae). Am. J.Bot. 91:1070–1085.
de Queiroz, A. 2005. The resurrection of oceanic dispersal in historic-biogeography. Trends Ecol. Evol. 20:68–73.
Devos, N., and A. Vanderpoorten. 2009. Range disjunctions, speciation, andmorphological transformation rates in the liverwort genus Leptoscyphus.Evolution 63:779–792.
Dimitrov, D., M. A. Arnedo, and C. Ribera. 2008. Colonization and diversi-fication of the spider genus Pholcus Walckenaer, 1805 (Araneae, Phol-cidae) in the Macaronesian archipelagos: evidence for long-term occu-pancy yet rapid recent speciation. Mol. Phylogenet. Evol. 48:596–614.
Emerson, B. C., and P. Oromı. 2005. Diversification of the forest beetle genusTarphius on the Canary islands, and the evolutionary origins of islandendemics. Evolution 59:586–598.
Engler, A. 1879. Versuch einer Eintwicklungsgeschichte, insbesondere derFlorengebiete seit der Tertiarperiode: I. Die extra-tropischen Gebieteder nordlischen Hemisphare. W. Engelmann, Leipzig.
Frahm, J.-P. 2004. A new contribution to the moss flora of Baltic and Saxonamber. Rev. Paleobot. Palynol. 129:81–101.
Gillepsie, R. G., and G. K. Roderick. 2002. Arthropods on islands: coloniza-tion, speciation, and conservation. Annu. Rev. Entomol. 47:595–632.
Goldberg, J., S. A. Trewick, and A. M. Paterson. 2008. Evolution of NewZealand’s terrestrial fauna: a review of molecular evidence. Philos.Trans. R. Soc. Lond. B. 363:3319–3334.
Grandcolas, P., J. Murienne, T. Robillard, L. Desutter-Grandcolas, H.Jourdan, E. Guilbert, and L. Deharveng. 2008. New Caledonia: avery old Darwinian island? Philos. Trans. R. Soc. Lond B. 363:3309–3317.
Heinrichs, J., J. Hentschel, R. Wilson, K. Feldberg, and H. Schneider.2007. Evolution of leafy liverworts (Jungermanniidae, Marchantio-phyta): estimating divergence times from chloroplast DNA sequencesusing penalized likelihood with integrated fossil evidence. Taxon 56:31–44.
Huttunen, S., and M. S. Ignatov. 2004. Phylogeny of the Brachytheciaceae(Bryophyta) based on morphology and sequence level data. Cladistics20:151–183.
Jordal, B. H., and G. M. Hewitt. 2004. The origin and radiation of Macarone-sian beetle breeding in Euphorbia: the relative importance of multipledata partitions and population sampling. Syst. Biol. 53:711–734.
Juan, C., B. C. Emerson, P. Oromı, and G. M. Hewitt. 2000. Colonizationand diversification: towards a phylogeographic synthesis for the CanaryIslands. Trends Ecol. Evol. 15:104–109.
Lee, J., J. Alrubaian, and R. M. Dores. 2006. Are lungfish living fossils?Observation on the evolution of the opioid/orphanin gene family. Gen.Comp. Endocrinol. 148:306–314.
Miller, N. G. 1984. Tertiary and Quaternary fossils. Pp. 1194–1232 in R.Schuster ed. New Manual of Bryology, Vol. 2. Hattori Botanical Labo-ratory, Nichinan.
Pagel, M. 1999. The maximum likelihood approach to reconstructing ances-tral character states of discrete characters on phylogenies. Syst. Biol.48:612–622.
Schnabel, A., and J. F. Wendel. 1998. Cladistic biogeography of Gleditsia(Leguminosae) based on ndhF and rpl16 chloroplast gene sequences.Am. J. Bot. 85:1753–1765.
Shaw, A. J., C. J. Cox, B. Goffinet, W. R. Buck, and S. B. Boles. 2003.Phylogenetic evidence of a rapid radiation of pleurocarpous mosses(Bryophyta). Evolution 57:2226–2241.
Sotiaux, A., J. Enroth, S. Olsson, D. Quandt, and A. Vanderpoorten. 2009.When morphology and molecules tell us different stories: a case-in-pointwith Leptodon corsicus, a new and unique endemic moss species fromCorsica. J. Bryol. 31 In press.
Stech, M., M. Sim-Sim, M. G. Equivel, S. Fontinha, R. Tangney, C. Lobo, R.Gabriel, and D. Quandt. 2008. Explaining the ‘anomalous’ distributionof Echinodium (Bryopsida: Echinodiaceae): independent evolution inMacaronesia and Australasia. Org. Divers. Evol. 8:282–292.
Sunding, P. 1979. Origin of the Macaronesian flora. Pp. 13–40 in D. Bramwelled. Plants and Island. Academic Press, London.
Vanderpoorten, A., M. A. Carine, and F. Rumsey. 2007. Does Macaronesiaexist? Conflicting signal in the bryophyte and pteridophyte floras. Am.J. Bot. 94:625–639.
Vanderpoorten, A., N. Devos, B. Goffinet, O. Hardy, and A. J. Shaw. 2008.The barriers to oceanic island radiation in bryophytes: insights from thephylogeography of the moss Grimmia montana. J. Biogeogr. 35:654–663.
Wall, D. P. 2005. Origin and rapid diversification of a tropical moss. Evolution59:1413–1424.
Zander, R. H. 2008. Evolutionary inferences from nonmonophyly of tradi-tional taxa on molecular trees. Taxon 57:1182–1188.
Associate Editor: J. Vamosi
3 2 5 6 EVOLUTION DECEMBER 2009

BRIEF COMMUNICATION
Supporting InformationThe following supporting information is available for this article:
Appendix S1. List of 43 morphological binary characters scored on the sample of taxa from the Helicodontioideae.
Appendix S2. Matrix of 43 morphological characters (see Appendix S1) scored for taxa of the Helicodontioideae used in the
present study.
Supporting Information may be found in the online version of this article.
(This link will take you to the article abstract).
Please note: Wiley-Blackwell is not responsible for the content or functionality of any supporting information supplied by the
authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
EVOLUTION DECEMBER 2009 3 2 5 7
Copyright © 2022 FDOKUMEN




















