
Analysis of genetically modified red-fleshed apples reveals effects on growth and consumer...
12
Analysis of genetically modified red-fleshed apples reveals effects on growth and consumer attributes Richard V. Espley 1, *, Arnaud Bovy 2 , Christina Bava 1 , Sara R Jaeger 1 , Sumathi Tomes 1 , Cara Norling 3 , Jonathan Crawford 3 , Daryl Rowan 3 , Tony K. McGhie 3 , Cyril Brendolise 1 , Jo Putterill 4 , Henk J. Schouten 2 , Roger P. Hellens 1 and Andrew C. Allan 1,4 1 The New Zealand Institute for Plant & Food Research Limited (PFR), Auckland, New Zealand 2 Plant Research International, Wageningen, The Netherlands 3 PFR, Palmerston North, New Zealand 4 School of Biological Sciences, University of Auckland, Auckland, New Zealand Received 22 June 2012; revised 12 September 2012; accepted 25 September 2012. *Correspondence (fax + 64 9925 7001; email [email protected]) Keywords: anthocyanin, apple, biotechnology, metabolites, red flesh, sensory. Summary Consumers of whole foods, such as fruits, demand consistent high quality and seek varieties with enhanced health properties, convenience or novel taste. We have raised the polyphenolic content of apple by genetic engineering of the anthocyanin pathway using the apple transcription factor MYB10. These apples have very high concentrations of foliar, flower and fruit anthocyanins, especially in the fruit peel. Independent lines were examined for impacts on tree growth, photosynthesis and fruit characteristics. Fruit were analysed for changes in metabolite and transcript levels. Fruit were also used in taste trials to study the consumer perception of such a novel apple. No negative taste attributes were associated with the elevated anthocyanins. Modification with this one gene provides near isogenic material and allows us to examine the effects on an established cultivar, with a view to enhancing consumer appeal independently of other fruit qualities. Introduction The consumption of fresh fruits and vegetables is an important component of a healthy diet. Initiatives such as ‘5-a-day’ are aimed at increasing the number of fruit and vegetable servings in the human diet in response to mounting evidence for their health benefits (National Cancer Institute, 2000). The effectiveness of such campaigns would be enhanced by the introduction of novel fruit and vegetable varieties that are both attractive to the consumer and provide more health properties. In the majority of plant species, pigmentation is controlled by the relative amounts of anthocyanins, chlorophyll and carote- noids. These compounds are essential for plant health and performance, but are also considered as phytonutrients or markers for dietary health (Harborne and Williams, 2000; Mayne, 1996). Because of their potential to provide a novel appearance, pigments are often the target of plant-breeding programmes. Breeding programmes for fruits exploit germplasm collections to develop new varieties with improved production or consumer traits. An example is the effort to breed red-fleshed apples with elevated concentrations of anthocyanins (Volz et al., 2009). This involves crossing red-fleshed wild apples, which originate from the centre of origin of apple in Central Asia, with modern white- fleshed commercial varieties. Many subsequent generations of crosses are then required to exclude unwanted characteristics caused by linkage drag, such as poor taste, texture and storage. An alternative approach would be the direct integration of the dominant red-flesh MYB allele (Espley et al., 2009) into a modern high-quality commercial apple cultivar via a transgenic or cisgenic approach (Schouten and Jacobsen, 2008). Previous examples of genetic modification of apples have largely targeted production traits such as disease resistance, dwarfing and fruit storage (reviewed in Bulley et al., 2007). Several crops have been genetically modified (GM) to increase dietary health benefits, such as potato (Rommens et al., 2008) and tomato (Butelli et al., 2008; Rein et al., 2006). Engineering of fruit pigmentation in tomato has demonstrated success in pigment manipulation. This includes the transformation of tomato with the maize transcription factors (TFs) Lc (bHLH) and C1 (MYB, Bovy et al., 2002). Fruit showed an elevation of flavonoids, but not anthocyanins. Silencing of the DE-ETIO- LATED1 (DET1) gene in tomato also showed elevation in flavonoids (including gains in anthocyanins) and carotenoids (Davuluri et al., 2005). To date, the largest gain in anthocyanins in tomato has been from the simultaneous over-expression of the Antirrhinum TFs Delila (bHLH) and Rosea1 (MYB, Butelli et al., 2008). This resulted in a large increase in anthocyanins, producing purple-coloured fruit with a high degree of antioxi- dant activity. Anthocyanins are elevated in response to developmental, environmental and stress stimuli, including temperature, nutri- ent status, wounding, pathogen infection, water stress, and UV, visible and far-red light (Chalker-Scott, 1999; Christie et al., 1994; Dixon and Paiva, 1995; Steyn et al., 2002). The biosynthesis of anthocyanin pigments is transcriptionally regu- lated by the MYB-bHLH-WD40 (MBW) TF complex (reviewed in Ramsay and Glover (2005) and Feller et al. (2011)). The first isolated MYB member of this complex was the maize C1 gene (Paz-Ares et al., 1987) and homologues have subsequently been isolated from many flower and fruit species (Allan et al., 2008; Martin and Paz-Ares, 1997). In Arabidopsis, the PRO- DUCTION OF ANTHOCYANIN PIGMENTS 1 (PAP1) gene (also called MYB75; At1g56650) was identified by activation tagging (Borevitz et al., 2000). Over-expression of PAP1 in Arabidopsis results in purple-coloured leaves with increased biosynthesis of anthocyanins and quercetin glycosides. Microarray analysis of 408 ª 2012 The Authors Plant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd Plant Biotechnology Journal (2013) 11, pp. 408–419 doi: 10.1111/pbi.12017
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Analysis of genetically modified red-fleshed apples reveals effects on growth and consumer...

Analysis of genetically modified red-fleshed applesreveals effects on growth and consumer attributesRichard V. Espley1,*, Arnaud Bovy2, Christina Bava1, Sara R Jaeger1, Sumathi Tomes1, Cara Norling3,Jonathan Crawford3, Daryl Rowan3, Tony K. McGhie3, Cyril Brendolise1, Jo Putterill4, Henk J. Schouten2,Roger P. Hellens1 and Andrew C. Allan1,4
1The New Zealand Institute for Plant & Food Research Limited (PFR), Auckland, New Zealand2Plant Research International, Wageningen, The Netherlands3PFR, Palmerston North, New Zealand4School of Biological Sciences, University of Auckland, Auckland, New Zealand
Received 22 June 2012;
revised 12 September 2012;
accepted 25 September 2012.
*Correspondence (fax + 64 9925 7001;
email [email protected])
Keywords: anthocyanin, apple,
biotechnology, metabolites, red flesh,
sensory.
SummaryConsumers of whole foods, such as fruits, demand consistent high quality and seek varieties withenhanced health properties, convenience or novel taste. We have raised the polyphenoliccontent of apple by genetic engineering of the anthocyanin pathway using the appletranscription factorMYB10. These apples have very high concentrations of foliar, flower and fruitanthocyanins, especially in the fruit peel. Independent lines were examined for impacts on treegrowth, photosynthesis and fruit characteristics. Fruit were analysed for changes in metaboliteand transcript levels. Fruit were also used in taste trials to study the consumer perception of sucha novel apple. No negative taste attributes were associated with the elevated anthocyanins.Modification with this one gene provides near isogenic material and allows us to examine theeffects on an established cultivar, with a view to enhancing consumer appeal independently ofother fruit qualities.
Introduction
The consumption of fresh fruits and vegetables is an importantcomponent of a healthy diet. Initiatives such as ‘5-a-day’ areaimed at increasing the number of fruit and vegetable servings inthe human diet in response to mounting evidence for their healthbenefits (National Cancer Institute, 2000). The effectiveness ofsuch campaigns would be enhanced by the introduction of novelfruit and vegetable varieties that are both attractive to theconsumer and provide more health properties.
In the majority of plant species, pigmentation is controlled bythe relative amounts of anthocyanins, chlorophyll and carote-noids. These compounds are essential for plant health andperformance, but are also considered as phytonutrients ormarkers for dietary health (Harborne and Williams, 2000; Mayne,1996). Because of their potential to provide a novel appearance,pigments are often the target of plant-breeding programmes.Breeding programmes for fruits exploit germplasm collections todevelop new varieties with improved production or consumertraits. An example is the effort to breed red-fleshed apples withelevated concentrations of anthocyanins (Volz et al., 2009). Thisinvolves crossing red-fleshed wild apples, which originate fromthe centre of origin of apple in Central Asia, with modern white-fleshed commercial varieties. Many subsequent generations ofcrosses are then required to exclude unwanted characteristicscaused by linkage drag, such as poor taste, texture and storage.An alternative approach would be the direct integration of thedominant red-flesh MYB allele (Espley et al., 2009) into a modernhigh-quality commercial apple cultivar via a transgenic or cisgenicapproach (Schouten and Jacobsen, 2008). Previous examples ofgenetic modification of apples have largely targeted productiontraits such as disease resistance, dwarfing and fruit storage(reviewed in Bulley et al., 2007).
Several crops have been genetically modified (GM) to increasedietary health benefits, such as potato (Rommens et al., 2008)and tomato (Butelli et al., 2008; Rein et al., 2006). Engineeringof fruit pigmentation in tomato has demonstrated success inpigment manipulation. This includes the transformation oftomato with the maize transcription factors (TFs) Lc (bHLH) andC1 (MYB, Bovy et al., 2002). Fruit showed an elevation offlavonoids, but not anthocyanins. Silencing of the DE-ETIO-LATED1 (DET1) gene in tomato also showed elevation inflavonoids (including gains in anthocyanins) and carotenoids(Davuluri et al., 2005). To date, the largest gain in anthocyaninsin tomato has been from the simultaneous over-expression ofthe Antirrhinum TFs Delila (bHLH) and Rosea1 (MYB, Butelliet al., 2008). This resulted in a large increase in anthocyanins,producing purple-coloured fruit with a high degree of antioxi-dant activity.
Anthocyanins are elevated in response to developmental,environmental and stress stimuli, including temperature, nutri-ent status, wounding, pathogen infection, water stress, andUV, visible and far-red light (Chalker-Scott, 1999; Christie et al.,1994; Dixon and Paiva, 1995; Steyn et al., 2002). Thebiosynthesis of anthocyanin pigments is transcriptionally regu-lated by the MYB-bHLH-WD40 (MBW) TF complex (reviewed inRamsay and Glover (2005) and Feller et al. (2011)). The firstisolated MYB member of this complex was the maize C1 gene(Paz-Ares et al., 1987) and homologues have subsequentlybeen isolated from many flower and fruit species (Allan et al.,2008; Martin and Paz-Ares, 1997). In Arabidopsis, the PRO-DUCTION OF ANTHOCYANIN PIGMENTS 1 (PAP1) gene (alsocalled MYB75; At1g56650) was identified by activation tagging(Borevitz et al., 2000). Over-expression of PAP1 in Arabidopsisresults in purple-coloured leaves with increased biosynthesis ofanthocyanins and quercetin glycosides. Microarray analysis of
408ª 2012 The Authors
Plant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd
Plant Biotechnology Journal (2013) 11, pp. 408–419 doi: 10.1111/pbi.12017

35S:PAP1 Arabidopsis lines have shown increased expression of38 genes, including genes of the phenylpropanoid pathway andof anthocyanin-specific biosynthetic genes (Tohge et al., 2005).In apple, the anthocyanin pathway has been shown to becontrolled by the MYB transcription factor, MYB10 (Espleyet al., 2007). MYB10 shows a high degree of homology withanthocyanin regulators from other species, including Petunia,tomato and maize, and is closely related to Arabidopsis PAP1,with 77% amino acid identity at the R2R3 domain and 58% tothe entire protein (Espley et al., 2007).
MYB10 shows an expression pattern that correlates withanthocyanin pathway genes and is able to activate the promotersof a number of these genes. When fused to a 35S promoter, it iscapable of driving anthocyanin accumulation in both heterolo-gous systems and in apple (Espley et al., 2007). When MYB10was isolated from diverse apple germplasm, such as the red-fleshed Malus domestica var. Niedzwetzkyana and white-fleshed‘Royal Gala’ or ‘Granny Smith’, no difference in the proteinsequence was found. Differences in expression of MYB10,between apple cultivars are due to differences in the promoterof MYB10 (Espley et al., 2009). The MYB10 gene is allelic to apeel-expressed gene, MdMYB1 or MYBA (Ban et al., 2007;Lin-Wang et al., 2010; Takos et al., 2006a). Genetic evidencesuggests that these alleles are the major controllers of apple peelcolour (Ban et al., 2007; Zhu et al., 2010) and flesh and foliagecolour (Chagne et al., 2007). MdMYB1/MYBA expression isinduced by light and cold temperature, while MYB10 transcriptlevel is suppressed at higher temperatures (Ban et al., 2007;Lin-Wang et al., 2010; Takos et al., 2006a).
In this study, we show the effect of engineering a largeincrease in anthocyanins in apple and the outcome that this hason fruit qualities and consumer perceptions. The results indicatethat a red-fleshed apple would meet current consumer expecta-tions for flavour, with the added appeal of enhanced novelty.
Results
Analysis of MYB10 trees and fruit
‘Royal Gala’ apple was transformed with a binary vectorcontaining MYB10 cDNA under the control of the CaMV35Spromoter as previously described (Espley et al., 2007) to createtransgenic MYB10 apple plants (Figure 1a). After tissue culture,transformed ‘Royal Gala’ plantlets were micrografted onto ‘M.9’rootstock. Once growth was established, the trees were trans-ferred to the National Climate Laboratory (Plant & Food Researchin Palmerston North, NZ) and grown in conditions to hastenmaturity and fruiting. This process, from transformation tofruiting, took approximately 2 years (Figure S1 and Table S1).These trees showed ectopic accumulation of anthocyanin,resulting in deep pigmentation in the leaves, particularly evidentin and around veins. There was considerable anthocyaninaccumulation in areas of the trunk, including the outer phloemlayers and vasculature rays. Microscopic analysis of leavesrevealed anthocyanin accumulation predominantly in the spongymesophyll and lower epidermis and in tissue close to orcomprising the vascular rays (Figure 1c,d). The palisade mesophylldid not appear to accumulate anthocyanins (Figure 1e). Nosignificant changes to the rate of growth or the tree architecturewere observed from a comparison of wild-type (WT) ‘Royal Gala’and the MYB10 trees (Figure S2).
Ectopic leaf anthocyanin did not appear to compromisephotosynthetic electron transport capacity (Figure 2). Data onchlorophyll fluorescence were gathered using a pulse amplitude–modulated (PAM) photosynthetic yield analyser for leaves oftransgenic and control plants. The electron transport rate (ETR)versus photosynthetically active radiation (PAR) was plotted, andthe curves were used to estimate maximum photosyntheticcapacity (PSmax) and photosynthetic efficiency (initial slope of theETR versus PAR, a). A statistical analysis (F-test, statistical
(a) (b)
(c)
(d)
(e)
Figure 1 Over-expression of MYB10 in apple
produces highly anthocyanic plants. The three
panels in (a) show representative independent
lines of MYB10 (left), 1, 3 and 4 and control
wild-type (WT) ‘Royal Gala’ (right). Intense
pigmentation was seen in the trunk (b, left) and
leaves (c, top) of MYB10 lines. Back-lit images of
leaf tissue (d) further highlight the difference in
colour and show the accumulation of
anthocyanins in MYB10 tissue (left) in and around
leaf veins. (e) Microscopic images of leaf cross-
sections of ‘Royal Gala’, left, and MYB10, right.
Scale bar = 100 lm.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Genetically modified high-anthocyanin apples 409
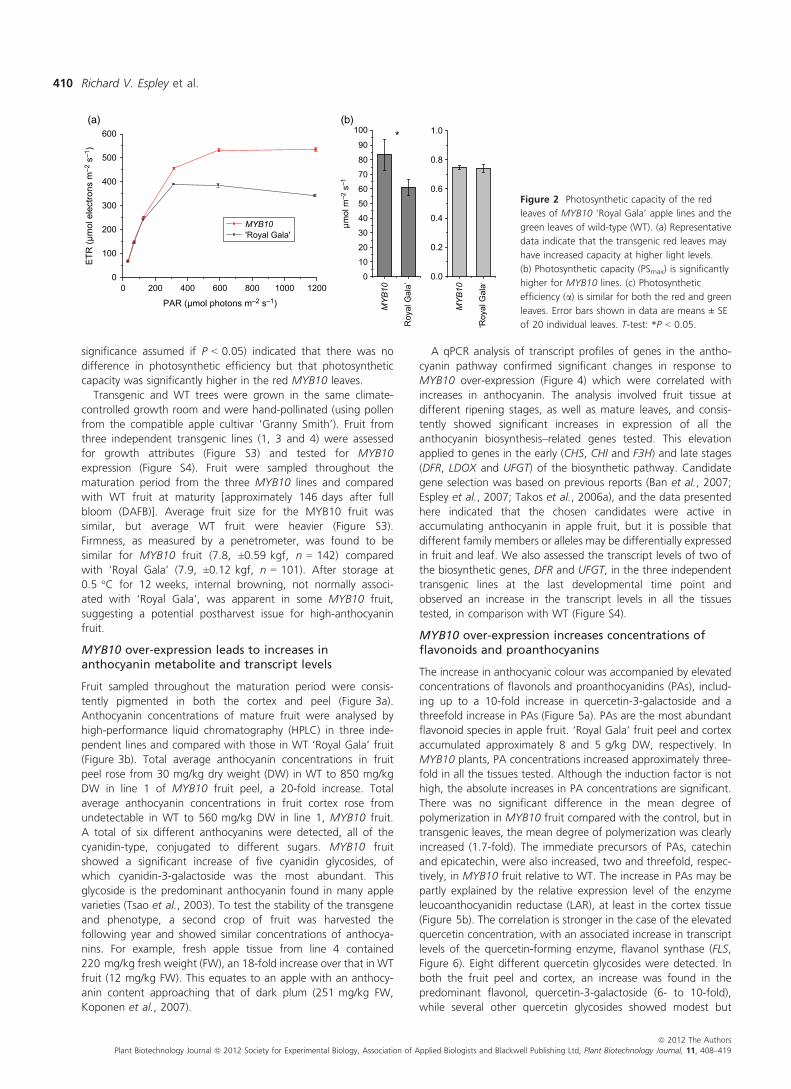
significance assumed if P < 0.05) indicated that there was nodifference in photosynthetic efficiency but that photosyntheticcapacity was significantly higher in the red MYB10 leaves.
Transgenic and WT trees were grown in the same climate-controlled growth room and were hand-pollinated (using pollenfrom the compatible apple cultivar ‘Granny Smith’). Fruit fromthree independent transgenic lines (1, 3 and 4) were assessedfor growth attributes (Figure S3) and tested for MYB10expression (Figure S4). Fruit were sampled throughout thematuration period from the three MYB10 lines and comparedwith WT fruit at maturity [approximately 146 days after fullbloom (DAFB)]. Average fruit size for the MYB10 fruit wassimilar, but average WT fruit were heavier (Figure S3).Firmness, as measured by a penetrometer, was found to besimilar for MYB10 fruit (7.8, ±0.59 kgf, n = 142) comparedwith ‘Royal Gala’ (7.9, ±0.12 kgf, n = 101). After storage at0.5 °C for 12 weeks, internal browning, not normally associ-ated with ‘Royal Gala’, was apparent in some MYB10 fruit,suggesting a potential postharvest issue for high-anthocyaninfruit.
MYB10 over-expression leads to increases inanthocyanin metabolite and transcript levels
Fruit sampled throughout the maturation period were consis-tently pigmented in both the cortex and peel (Figure 3a).Anthocyanin concentrations of mature fruit were analysed byhigh-performance liquid chromatography (HPLC) in three inde-pendent lines and compared with those in WT ‘Royal Gala’ fruit(Figure 3b). Total average anthocyanin concentrations in fruitpeel rose from 30 mg/kg dry weight (DW) in WT to 850 mg/kgDW in line 1 of MYB10 fruit peel, a 20-fold increase. Totalaverage anthocyanin concentrations in fruit cortex rose fromundetectable in WT to 560 mg/kg DW in line 1, MYB10 fruit.A total of six different anthocyanins were detected, all of thecyanidin-type, conjugated to different sugars. MYB10 fruitshowed a significant increase of five cyanidin glycosides, ofwhich cyanidin-3-galactoside was the most abundant. Thisglycoside is the predominant anthocyanin found in many applevarieties (Tsao et al., 2003). To test the stability of the transgeneand phenotype, a second crop of fruit was harvested thefollowing year and showed similar concentrations of anthocya-nins. For example, fresh apple tissue from line 4 contained220 mg/kg fresh weight (FW), an 18-fold increase over that in WTfruit (12 mg/kg FW). This equates to an apple with an anthocy-anin content approaching that of dark plum (251 mg/kg FW,Koponen et al., 2007).
A qPCR analysis of transcript profiles of genes in the antho-cyanin pathway confirmed significant changes in response toMYB10 over-expression (Figure 4) which were correlated withincreases in anthocyanin. The analysis involved fruit tissue atdifferent ripening stages, as well as mature leaves, and consis-tently showed significant increases in expression of all theanthocyanin biosynthesis–related genes tested. This elevationapplied to genes in the early (CHS, CHI and F3H) and late stages(DFR, LDOX and UFGT) of the biosynthetic pathway. Candidategene selection was based on previous reports (Ban et al., 2007;Espley et al., 2007; Takos et al., 2006a), and the data presentedhere indicated that the chosen candidates were active inaccumulating anthocyanin in apple fruit, but it is possible thatdifferent family members or alleles may be differentially expressedin fruit and leaf. We also assessed the transcript levels of two ofthe biosynthetic genes, DFR and UFGT, in the three independenttransgenic lines at the last developmental time point andobserved an increase in the transcript levels in all the tissuestested, in comparison with WT (Figure S4).
MYB10 over-expression increases concentrations offlavonoids and proanthocyanins
The increase in anthocyanic colour was accompanied by elevatedconcentrations of flavonols and proanthocyanidins (PAs), includ-ing up to a 10-fold increase in quercetin-3-galactoside and athreefold increase in PAs (Figure 5a). PAs are the most abundantflavonoid species in apple fruit. ‘Royal Gala’ fruit peel and cortexaccumulated approximately 8 and 5 g/kg DW, respectively. InMYB10 plants, PA concentrations increased approximately three-fold in all the tissues tested. Although the induction factor is nothigh, the absolute increases in PA concentrations are significant.There was no significant difference in the mean degree ofpolymerization in MYB10 fruit compared with the control, but intransgenic leaves, the mean degree of polymerization was clearlyincreased (1.7-fold). The immediate precursors of PAs, catechinand epicatechin, were also increased, two and threefold, respec-tively, in MYB10 fruit relative to WT. The increase in PAs may bepartly explained by the relative expression level of the enzymeleucoanthocyanidin reductase (LAR), at least in the cortex tissue(Figure 5b). The correlation is stronger in the case of the elevatedquercetin concentration, with an associated increase in transcriptlevels of the quercetin-forming enzyme, flavanol synthase (FLS,Figure 6). Eight different quercetin glycosides were detected. Inboth the fruit peel and cortex, an increase was found in thepredominant flavonol, quercetin-3-galactoside (6- to 10-fold),while several other quercetin glycosides showed modest but
0 200 400 600 800 1000 12000
100
200
300
400
500
600
ETR
(�m
ol e
lect
rons
m–2
s–1
)
PAR (�mol photons m–2 s–1)
MYB10'Royal Gala'
0
10
20
30
40
50
60
70
80
90
100
�mol
m–2
s–1
0.0
0.2
0.4
0.6
0.8
1.0*
MY
B10
Roy
al G
ala’
MY
B10
‘Roy
al G
ala ’
(a) (b)
Figure 2 Photosynthetic capacity of the red
leaves of MYB10 ‘Royal Gala’ apple lines and the
green leaves of wild-type (WT). (a) Representative
data indicate that the transgenic red leaves may
have increased capacity at higher light levels.
(b) Photosynthetic capacity (PSmax) is significantly
higher for MYB10 lines. (c) Photosynthetic
efficiency (a) is similar for both the red and green
leaves. Error bars shown in data are means ± SE
of 20 individual leaves. T-test: *P < 0.05.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Richard V. Espley et al.410

significant increases (two to threefold). In both tissues, totalquercetin concentrations increased up to 5-fold compared withthe control.
Further metabolite analysis showed that the cortex of MYB10fruit accumulated 1.5-and 3.6-fold higher concentrations offructose and glucose and 0.44-fold lower concentrations ofsucrose than WT fruit (Table 1), although these differences inconcentration were not statistically significant (P > 0.05). Malicacid concentrations were not significantly changed.
The analysis also revealed that b-carotene concentrations wereincreased in the peel and cortex of MYB10 fruit by 9.8- and 6.5-fold in the skin and cortex, respectively (Table 1). Apple fruit isgenerally not considered a rich source of carotenoids, but variedconcentrations of b-carotene, lutein, violaxanthin and anthera-xanthin and others are often present (Ampomah-Dwamenaet al., 2012; Felicetti and Schrader, 2009; Knee, 1972). The maincarotenoid, b-carotene, was present at approximately 15 mg/kgDW in the skin of MYB10 fruit. Concentrations of a-tocopherolwere not changed in transgenic MYB10 plants.
A study of gene transcript levels in the carotenoid pathwayshowed transcript elevation of the early biosynthetic steps,phytoene synthase and phytoene desaturase, especially at fruitmaturity for the peel (Figure S5 and Table S2).
High anthocyanins do not adversely affect appleconsumer qualities
Anthocyanins appear to have little or no taste, but it has beensuggested that they are astringent (Brossaud et al., 2001).
Research with wine has shown that PAs or condensed tanninscontribute to astringency and bitterness, but also suggests thatanthocyanins can modulate this effect by interacting with variousphenolic compounds (Vidal et al., 2004a,b). We conducted asensory panel to evaluate the effect on taste and astringency, andultimately on consumer acceptability, of the increased anthocy-anins, flavan-3-ols (catechin and epicatechin), PAs and quercetinglycosides in MYB10 fruit. The consumer panel consisted of 50screened participants who were asked to determine whetherMYB10 apples had similar or different taste attributes to the WTapples that had been grown under similar conditions. The panelconsisted of four sets for fruit, two samples per set; (i) A trainingset using American-sourced ‘Gala’ and ‘Pink Lady’ apples; (ii) TheNew Zealand-grown MYB10 and WT apples; (iii) Two samples,both from American-sourced ‘Gala’ and (iv) Two samples, bothfrom American-sourced ‘Gala’, but with a prompt that onesample might be GM.
Following the tasting of both apples in each set, consumerswere asked to provide comments on the sensory attributes of thefruit. Texture was a frequent descriptor used, with 39/50consumers commenting on texture attributes, including crisp-ness/crunchiness and juiciness. In terms of taste, sweetness wasagain most commonly mentioned and second to this was ‘nice’ or‘flavourful’. Eleven descriptors relating to texture were identifiedacross the four sets. There was a noticeable drop in the numberof consumers who related the terms crisp/crunchy to the setcontaining the WT and MYB10 apples, compared with therelative consistency across the other sets. Sweetness was less
(a)
Fruit peel
Ret. Ɵme (min)
‘Royal Gala’
MYB10 Line 1
MYB10 Line 3
MYB10 Line 4
InducƟonfactor
T-test
Ret. Ɵme (min)
‘Royal Gala’
MYB10 Line 1
MYB10 Line 3
MYB10 Line 4
InducƟonfactor
T-test
(b)
Cyanidin-3-galactoside 18.4 48.7 728.7 590.0 480.0 12.3 **
Cyanidin-3-glucoside 17.8 4.0 63.6 52.1 44.2 13.3 **
Cyanidin-glycoside 1 20.4 4.6 17.1 15.2 12.7 3.3 **
Cyanidin-glycoside 2 22.6 0.5 17.4 12.7 7.4 24 **
Cyanidin-glycoside 3 22.9 0.4 26.6 21.4 16.9 50 **
Cyanidin glycoside 4 23 7 nd 2 4 0 5 ndCyanidin-glycoside 4 23.7 2.4 0.5 nd - -
2.1659.1968.5582.85latoT
Fruit cortex
Cyanidin-3-galactoside 18.4 nd 473.3 252.0 168.7 >199 -
Cyanidin 3 glucoside 17 8 nd 46 9 24 9 20 4 > 21Cyanidin-3-glucoside . . . . -
Cyanidin-glycoside 1 20.4 nd 11.8 9.1 7.3 > 6.3 -
Cyanidin-glycoside 2 22.6 nd 10.0 4.3 1.7 > 3.6 -
Cyanidin-glycoside 3 22.9 nd 21.3 12.8 9.9 > 9.8 -
Cyanidin-glycoside 4 23.7 nd 1.9 nd nd - -
0.8021.3032.565latoT
Figure 3 Anthocyanin concentrations were greatly increased in MYB10 ‘Royal Gala’ apple fruit. (a) Left, whole wild-type (WT) and MYB10 fruit with
MYB10 slice and right, MYB10 (Line 4) and WT cut fruit. (b) Anthocyanin concentrations in pooled samples of peel and cortex of MYB10 and WT fruit in
mg/kg dry weight (DW). T-test: *P < 0.05; **P < 0.01; ***P < 0.001. Induction factor: average concentrations of transgenic lines (1, 3 and 4)/average
concentrations of control lines (1 and 2). nd, not detected <1.5 mg/kg DW.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Genetically modified high-anthocyanin apples 411

frequently used to describe WT (n = 15) and MYB10 (n = 5),compared with almost half the participants doing so in the othersets. More importantly, in the set containing the WT and MYB10apples, some consumers specifically described these as being ‘notsweet’, an attribute that barely rated in the other sets.
In terms of the texture and taste descriptors, there was littledifference between sets 3 and 4 (i.e. American ‘Gala’ applesversus American ‘Gala’ with a GM prompt). This suggests that the
GM prompt did not negatively influence their opinions. In termsof the preference data, however, it was noticed that a largernumber of participants identified a preference in Set 4 (whenthere was a GM prompt), whereas in the previous set a largernumber of participants identified the two samples as the same.Again, by collecting data for apples that were known to be thesame or different, we were better able to understand thecomments for the WT and MYB10 apples.
10
12
14
4
6
8
10
6
PAL
CHS F3H
p-Courmaroyl-CoA 3 x Malonyl-CoA0
2
4
6
8
Rel
ativ
e E
xpre
ssio
n
1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
0
2
Rel
ativ
e E
xpre
ssio
n
Peel Peel Cortex1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
Cortex
MY
B10
Lea
fR
G L
eaf
0
2
4
Rel
ativ
e E
xpre
ssio
n
1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
OH
OH
OH
O
OH
O
HO
HO
CHI
CHS
Chalcone
Naringenin
CHI
CHS
Peel Peel Cortex Cortex
MY
B10
Lea
fR
G L
eaf
6
8
10
12Peel Peel Cortex Cortex
MY
B10
Lea
fR
G L
eaf
15
20
25
30
35
40
CHI DFR
OH O
OH
OH
O
O
OH
HO
F3H
Dihydrokaempferol
OH
O
OH
OH
HO
F3’H
Dihydroquercetin
0
2
4
Rel
ativ
e E
xpre
ssio
n
Peel Peel Cortex1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
Cortex
MY
B10
Lea
fR
G L
eaf
0
5
10Rel
ativ
e E
xpre
ssio
nPeel Peel Cortex
1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5Cortex
RG
Lea
fM
YB
10 L
eaf
2020
OH
OH
OH
O
OH
OH
OH
O
OH
OH
HO
HO
DFR
LDOX
Leucoanthocyanidin
Anthocyanidin
OH O
5
10
15
Rel
ativ
e E
xpre
ssio
n
4
8
12
16
Rel
ativ
e E
xpre
ssio
n
LDOX UFGT
OH OH
OH
OH
O
OH
O-Gal
HO
UFGT
Cyanidin 3-galactoside
0
Peel Peel Cortex1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
Cortex
MY
B10
Lea
fR
G L
eaf
0
Peel Peel Cortex1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
CortexM
YB
10 L
eaf
RG
Lea
f
‘Royal Gala’
MYB10
1 2 3 4 5Key
Figure 4 Anthocyanin gene expression in MYB10 ‘Royal Gala’ apple tissue shows an increase for all anthocyanin structural genes tested. Schematic of
the anthocyanin pathway in apple and the relative expression profiles of associated genes in the peel, cortex and leaf of MYB10 and wild-type (WT).
Three fruit from each of the three lines (1, 3 and 4) were pooled and compared with pooled WT fruit. The key shows the five developmental stages
at (1) 40, (2) 67, (3) 102, (4) 130 and (5) 146 days after full bloom (DAFB), as per the x-axis labelling. Photographs for stages 1–5 are representative
of cortex colour and are not shown to scale. Data are presented as means (±SE) of four technical qPCR replicates.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Richard V. Espley et al.412

The results indicated that the potential negative flavourattributes of the elevated flavonoids and PAs did not affectsweetness, bitterness and acidity, although it is possible that theapparent increase in sugars (Table 1) may have offset anydetectable difference in astringency. Individual fruit for the tastetrial were assessed for sugar content by refractometer, butvariation between individual fruit was too great for significantvariation between MYB10 and WT (WT, 10.4 °Brix ± 1.3compared with MYB10, 10.1 °Brix ± 0.6). In the absence ofvisual cues, consumers did not detect changes in flavour.Analysis of the preference scores revealed that more panellists(n = 29) preferred the WT apple to the MYB10 apple (n = 8,Figure 7), but this was related to the texture of the fruit.Twenty-six per cent of consumers preferred both apples equally(n = 13). While there was little difference in measured fruitfirmness, sensorial fruit texture did appear to be compromised(i.e. softer fruit), leading to a preference for the WT fruit.Response to the idea of a red-fleshed apple was positivelyreceived by all the consumers. Overall, taste and texture werethe key attributes in the minds of consumers. When presentedwith the information that the red-fleshed apple may have anadditional health benefit, the consumer response remainedpositive (see Appendix S1 online).
A separate analysis of the aroma volatile profiles ofMYB10 andWT fruit was measured by headspace GCMS after cold storage(24 days) and further periods (3 and 7 days) of ripening at roomtemperature. The volatile profiles of MYB10 and WT, consistingof measurements of around 45 compounds, were remarkablysimilar at both maturities. Only two minor compounds, identifiedtentatively by MS as a hexenoic acid and as methyl iso-eugenol,appeared higher in control fruit measured after 7 days ripening(Figure S6).
Discussion
Here, we show the effect of over-expressing an MYB TF in appleand the subsequent effects on the trees, the fruit and humansensory perceptions. Over-expression of the TF MYB10 in appleleads to highly pigmented red-fleshed fruit with elevatedconcentrations of total anthocyanin. This phenotype wasachieved using one member of the MBW complex, the MYBcomponent, and has produced trees and fruit that are similar tothose of red-fleshed apple varieties that originate from CentralAsia. It is conceivable that using more than one member of thecomplex, such as the MYB and the bHLH, may produce an evenmore extreme phenotype, as has been demonstrated in tomato
‘Royal Gala’
MYB10Key 1 2 3 4 5
1 2 3 4 50
2
4
6
Rel
ativ
e E
xpre
ssio
n
Peel1 2 3 4 5
Peel1 2 3 4 5
Cortex1 2 3 4 5
Cortex
MY
B10
Lea
fR
G L
eaf
(a)
(b) LAR
Fruit peel‘Royal Gala'
1‘Royal Gala'
2MYB10 Line 1
MYB10 Line 3
MYB10Line 4
Inductionfactor
T-test
Epicatechin 715 678 2572 2815 2457 3.8 ***Catechin 123 152 274 391 322 2.4 *Total PA 8328 9525 26952 35517 30968 3.5 **
Fruit cortex‘Royal Gala'
1‘Royal Gala'
2MYB10Line 1
MYB10Line 3
MYB10Line 4
Inductionfactor
T-test
Epicatechin 287 463 1341 1305 1365 3.6 **Catechin 73.8 129 192 192 242 2.1 *Total PA 2816 4603 10884 11551 12511 3.1 **
Figure 5 Concentrations of flavan-3-ols and gene expression of LAR in MYB10 ‘Royal Gala’ apple fruit. (a) Concentration of flavan-3-ols in transgenic
MYB10 lines in mg/kg dry weight. Induction factor: average levels of transgenic lines (1, 3 and 4)/average levels of control lines (1 and 2). T-test:
*P < 0.05; **P < 0.01; ***P < 0.001. (b) Relative expression levels of LAR in MYB10 and wild-type (WT). Key as Figure 4. Data are presented as
means (±SE) of four technical replicates.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Genetically modified high-anthocyanin apples 413

(Butelli et al., 2008). At the cellular level, ectopic expression ofthe MYB10 gene led to anthocyanin accumulation in a range ofcell types including leaf, mesophyll and cambial layers. Theseelevated anthocyanin concentrations had little or no influence onleaf size or internode length. The older transgenic trees appearedarchitecturally indistinguishable from control plants.
No detrimental effect was observed on photosynthesis due tothe accumulation of anthocyanins in MYB10 leaves. Althoughleaves appeared to be uniformly red, microscopic sectionsshowed that cyanic cells were largely confined to the lowerepidermis and vasculature. In leaves where anthocyanins
accumulate exclusively in the upper epidermis, there can be asubstantial loss in photosynthetic capacity (Burger and Edwards,1996), whilst accumulation in the palisade and spongy mesophylllayers results in a smaller decrease (Gould et al., 2002). Whenanthocyanins are accumulated in the lower epidermis, there maybe little reduction in photosynthetic efficiency, and, indeed, theseleaves may demonstrate higher maximum yields (Gould et al.,1995). Cyanic leaves have also been shown to recover morequickly than acyanic leaves when exposed to strong light (Feildet al., 2001; Neill and Gould, 2003). At high light levels, theanthocyanin pigments may act as filters, absorbing potentially
1 2 3 4 50
5
10
15
20
25
Rel
ativ
e ex
pres
sion
Peel1 2 3 4 5
Cortex1 2 3 4 5
Cortex
MY
B10
Lea
fR
G L
eaf1 2 3 4 5
Peel
‘Royal Gala’
MYB10Key 1 2 3 4 5
(a)
(b)Fruit peel
‘Royal Gala' 1
Royal Gala' 2
MYB10 Line 1
MYB10 Line 3
MYB10 Line 4
Inductionfactor
T-test
Quercetin pentose 14.9 28.5 65.5 116 90.9 4.2 *Quercetin pentose 27.3 63.6 115 167 139 3.1 *Quercetin-3-glucoside 5.4 7.6 15.1 33.8 26.6 3.9 *Quercetin-3-galactoside 36.7 35.8 263 473 360 10.1 **Quercetin-3-rhamnoside 16.0 29.8 30.3 50.8 49.0 1.9Chlorogenic acid 1169 1153 5299 1643 2703 2.8
Phloridzin 83.3 283 530 413 253 2.2Phloridzin xyloside 5.7 13.1 24.7 31.3 25.7 2.9 *
Fruit cortex‘Royal Gala'
1Royal Gala'
2MYB10 Line 1
MYB10 Line 3
MYB10 Line 4
Inductionfactor
T-test
Quercetin pentose 1.6 nd 13.2 11.9 13.9 5.7 **Quercetin pentose 2.9 nd 6.0 8.4 12.7 3.0Quercetin-3-glucoside 2.7 nd 3.3 nd 3.4 1.1 **Quercetin-3-galactoside 12.2 2.9 47.6 47.7 54.5 6.6 *Quercetin-3-rhamnoside 3.5 5.1 12.3 9.6 15.3 2.9 *Chlorogenic acid 1881 1934 2913 2234 4860 1.7Phloridzin 296 166 131 119 180 0.6Phloridzin xyloside 0.2 nd 4.3 4.2 5.1 2.8
nd not detected <3.1 mg/kg DW.
FLS
Figure 6 Gene expression of flavanol synthase (FLS) and levels of quercetin glycosides in MYB10 ‘Royal Gala’ apple fruit. (a) Relative expression levels
of FLS in MYB10 and wild-type (WT) tissues. Key as Figure 4. Data are presented as means (±SE) of four technical replicates. (b) Concentration of
quercetin glycosides in transgenic MYB10 lines in mg/kg dry weight. Induction factor: average levels of transgenic lines (1, 3 and 4)/average levels of
control lines (1 and 2). T-test: *P < 0.05; **P < 0.01; ***P < 0.001.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Richard V. Espley et al.414

damaging wavelengths of light and so reducing photoinhibition.This may explain the apparent increase in photosynthetic capacityfor the red leaves of MYB10 at higher light levels, while thelocation of leaf anthocyanin may explain the similarity inphotosynthetic efficiency in both red and green leaves. It shouldbe noted that the light levels used in the containment growthfacility (900 lmol/m2/s) is less than the possible maximum in fieldconditions (2000 lmol/m2/s).
While the red phenotype for both the leaves and fruit ofMYB10 lines was very apparent, we have not determined whatthe effect of different light levels would be on the phenoptype. Ithas been shown that the MYB/MYBA1 allele is light-induced andso it is possible that different light levels may further influence thisphenotype.
The enhanced anthocyanin concentrations were associatedwith fruit browning. This browning was confined to the fruitcortex and is contrary to the elevated antioxidant capacity of thefruit. Internal membrane instability at low temperature could leadto anthocyanin leakage from the vacuole and interaction withcellular oxidation elements, such as polyphenol oxidase (PPO).Anthocyanin degradation in planta is often induced duringdevelopment or pathogen attack (Pourcel et al., 2007). In
isolation, anthocyanins may be quite stable in the presence ofPPO, but anthocyanin degradation can readily occur in thepresence of both PPO and other phenolics, as has been shown inlychee and blueberry (Jiang, 2000; Kader et al., 1998). Browningin MYB10 apples may be due to increases of both anthocyaninsand, for example, flavan 3-ols.
Analysis of the transcript levels of anthocyanin-related biosyn-thetic genes from pooled transgenic and WT fruit showed largeincreases in the MYB10 samples. The specific fold changes ofthese genes varied but were consistently higher than WT invirtually all tissues and at all time points. Similarly, we tested twoof these genes in the individual lines (1, 3 and 4) for thesubsequent season’s fruit and found high elevation of transcriptabundance (Figure S4). While these do not always tightlycorrelate with MYB10 expression levels (Figure S4), the patternof relative expression does appear to confirm the likelihood of co-ordinated regulation.
The increased flux in the anthocyanin pathway was alsoassociated with increases in metabolite concentrations of otherpolyphenols, including up to a 10-fold increase in quercetin-3-galactoside, a threefold increase in flavan-3-ols (twofold incatechins and fourfold in epicatechins) and a threefold increasein PAs. Epicatechin has been implicated in the beneficialcardiovascular effects of a flavonoid-rich diet and represents auseful target for health improvement (Schroeter et al., 2006).Previous studies into the control of polyphenolics in tomato haveshown increases in flavonoid concentrations, either by targetedover-expression of the flavonoid pathway, although apparentlythis did not produce a simultaneous increase in anthocyanins(Muir et al., 2001), or through flavonoid-regulating TFs, whichdid appear to produce an increase in flavonols but not anthocy-anins (Bovy et al., 2002). An increase in flavonoids was reportedin highly anthocyanic tomato transformed with the anthocyanin-related MYB and bHLH TFs from Antirrhinum (Butelli et al., 2008).Rutin and other quercetin derivatives were elevated, particularlyin the tomato peel. In Arabidopsis, PAP1 over-expression resultedin highly pigmented plants and a 10-fold increase in quercetinglycosides (Tohge et al., 2005). In apple, the increase in theflavan-3-ols, catechin and epicatechin, and in PAs is partlycorrelated with an increase in the relative expression level of theenzyme LAR1, at least in the cortex tissue. This correlation ofmetabolite concentration and gene expression was more appar-ent in the case of elevated quercetin and the increase in transcriptlevels of the enzyme, FLS, which leads to the formation ofquercetin. Here, we report an increase in FLS transcript levels overthe entire fruit development series (peel and cortex) in the MYB10tissue compared with WT (Figure 6a). In contrast, previous studiesin apple have shown that FLS transcript levels tend to be at theirhighest early and then late in fruit development, although thisdoes not necessarily follow flavonol concentrations and appearedto correlate more with anthocyanin synthesis than flavonolsynthesis (Takos et al., 2006b). This might suggest a possiblerole for MYB10, or simply relate to an increased pathway flux.
Metabolite and transcript analysis of the fruit revealed changesgenerally limited to the precursors or members of the anthocyaninpathway. A notable exception to this was a significant increase inb-carotene. While the elevation of anthocyanins and flavonoidsmay be expected in the transcriptionally coordinated phenylprop-anoid pathways, the simultaneous elevation in carotenoidconcentrations is surprising. An interaction between the phenyl-propanoid and carotenoid pathways has been observed in theRNAi-mediated suppression of the light-signalling pathway
Table 1 Fold change in primary and non-anthocyanic metabolite
concentrations (mean ± SE) between three independent transgenic
apple MYB10 lines (1, 3 and 4) and wild type (WT) fruit samples
(transgenic/WT)
Peel Cortex
Chlorogenic acid 2.77 ± 1.15 1.75 ± 0.83
Phloridzin 2.18 ± 0.54 0.62 ± 0.12
ß-carotene 9.81 ± 2.15** 6.50 ± 0.73*
a-carotene 3.08 ± 1.19 0.46 ± 0.33
a-tocopherol 0.85 ± 0.06 0.78 ± 0.05
Malic acid 0.74 ± 0.12 0.94 ± 0.14
Fructose 1.62 ± 0.17* 1.49 ± 0.26
Glucose 3.47 ± 0.99* 3.57 ± 1.54
Sucrose 0.31 ± 0.14 0.44 ± 0.17
Aspartic acid 0.39 ± 0.21 0.44 ± 0.21
T test (log transformed) of differences between transgenic and WT metabolite
concentrations.
*P < 0.05; **P < 0.01.
27%56%
17%
MYB10WT ('Royal Gala')The same
Figure 7 Preference for the sensory qualities of MYB10 and wild-type
(WT) ‘Royal Gala’ apple fruit. Preferences are shown as percentages from a
double-blind sensory trial with 50 consumers.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Genetically modified high-anthocyanin apples 415

regulatory gene, DET1 (Davuluri et al., 2005). Subsequent analysishas demonstrated the widespread effect of DET1 disruption, aplausible outcome considering the putative global nature of DET1as a core regulator of essential plant processes (Enfissi et al., 2010).The light hyper-responsiveness observed in DET1 may result fromthe coordinated regulation of pathways related to high lightconditions, such as carotenoid and flavonoid-type antioxidants.This regulation does not explain the increase in carotenoids seen inMYB10 fruit. MYBs in a diverse range of species, such asArabidopsis, have been shown to be specific regulators of theflavonoid pathway without any apparent links to carotenoidbiosynthesis (Tohge et al., 2005). However, our results suggestthat this elevated anthocyanin pathway flux, or the highlypigmented phenotype, somehow stimulates carotenoid produc-tion. While there were some increases in transcript levels ofcarotenoid biosynthetic genes, particularly PDS and PSY, there islittle evidence for the coordinated regulation of the wholepathway, or those components that lead to b-carotene production.For tomatoDET1 lines that also show an increase in carotenoids butnot in associated gene transcripts, it has been suggested that thismay be explained by posttranscriptional regulation (Enfissi et al.,2010). In apple, the pathway flux and corresponding accumulationof carotenoids appear to be controlled by three enzymeswithin thepathway (Ampomah-Dwamena et al., 2012). Using germplasmwith varying amounts of anthocyanins, the three enzymes, 15-cis-zeta-carotene isomerase (ZISO), carotenoid isomerase (CRTISO)and lycopene epsilon cyclase (LYC-e), were shown to correlate withand predict carotenoid concentrations in apple peel and flesh.Further work is required to explain the possible link, such astranscriptional control or changes in light perception, betweenthese two diverse pathways.
The changed metabolite profile of the MYB10 fruit did notappear to alter the flavour and aroma qualities of the fruit. Thisincluded an apparent increase in sugar concentrations in MYB10fruit, where fructose and glucose were increased by 1.5, and3.6-fold, respectively. However, any difference in taste may havebeen partly dissipated by a reduction in sucrose concentrationsby approximately 50%. In the absence of visual cues in a blindtrial, consumers did not detect a difference in taste. During thetest, prompts as to what sensory characteristics may be found inthe apples were not given to consumers. After answering thepreference question, they were simply asked for commentsabout the sensory aspects of the fruit that they liked/disliked. Ifastringency was a feature in the MYB10 fruit, we would haveexpected to see higher frequency of use of this term or relatedconsumer terms (bitter, aftertaste, unpleasant taste) relative tothe control apples, but this was not the case. The qualitative datamay help to explain why MYB10 apples were less preferred thanthe control apples, as they were described less frequently ascrisp/crunchy. There was also a tendency for them to bedescribed less frequently as sweet. Apples that are crisp/crunchyare often preferred to those with a mushy/floury texture, andsweetness is generally desired by consumers.
Volatile concentrations were also similar for both MYB10 fruitand WT. Recently, it has been shown that over-expression ofArabidopsis PAP1 in rose increases terpenoid scent compounds asa result of enhanced metabolic flux in both the phenylpropanoidand isoprenoid pathways (Zvi et al., 2012). The concentrations ofthe two minor compounds that changed in the MYB10 apples,hexenoic acid and methyl iso-eugenol, would be unlikely to bedetected by consumers. While there was little difference inmeasured fruit firmness, fruit texture did appear to be compro-
mised, leading to a preference for the control fruit. It suggeststhat the anthocyanin-rich fruit may be maturing more quickly,leading to a reduction in cell wall rigidity and perceived softness.From the structured discussion, the panellists in the consumer trialwere unanimously attracted to the concept of a red-fleshedapple. Our analysis of both sensory and volatile profiles of applesuggested no detectable difference in flavour and aromabetween MYB10 and WT fruit.
Many plant-breeding programmes have the aim of enhancingnovelty and food crop nutritional status. Previously, we haveshown that in apple, naturally occurring alleles of MYB10 areresponsible for anthocyanin accumulation in both vegetative andfruit tissue. These alleles are responsible for the red flesh of theancient wild apple accessions (Chagne et al., 2007). Harnessingthese naturally occurring alleles in a breeding programme willproduce high-anthocyanin apples (Espley et al., 2009). Our resultsindicate that red-fleshed apples, produced either through con-ventional breeding or by cisgenics (Schouten et al., 2006) of eliteapple cultivars, are likely to retain all the current consumerexpectations of flavour, with the added appeal of elevated colourand potential health enhancement.
Experimental procedures
Generation of MYB10 ‘Royal Gala’ lines and fruit
MYB10 ‘Royal Gala’ lines were generated by Agrobacterium-mediated transformation (Espley et al., 2007). Stringent growthconditions in a controlled environment were designed to hastenmaturity and flowering (Figure S1). After tissue culture, trans-formed plantlets of transgenic lines were micro-grafted onto‘M.9’ rootstock, adapted from a previously described protocol(Lane et al., 2003). The scions were tissue cultured transgenicshoots, approximately 3 cm long and prepared by cutting asection at the base of the stem and cutting back most of theexpanded leaves. ‘M.9’ rootstocks were prepared by pruninggrowing shoots to 10 cm from the base, making two short cutsthrough the bark and peeling the bark back for scion insertion.The scion was inserted under the bark of the rootstock, held inplace with a short strip of masking tape and covered with apolythene bag to prevent excessive moisture loss. The graftedplants were placed in a shaded area for 10–14 days until leafexpansion was detected on the scions. The bags were removedand the plants gradually exposed to higher light levels. Oncegrowth was established, the trees were transferred to theNational Climate Laboratories at Plant & Food Research inPalmerston North, New Zealand. Growth conditions, whichincluded stages representing fast growth, enforced winter chillingand optimum spring and summer-like conditions to hasten timeto flowering, are shown in Table S1. In particular, initial vegetativegrowth was encouraged with high light (900 lmol/m2/s), longdays (18 h), high CO2, and high temperature (25 °C). During thespring and summer phases, the light and temperature werereduced (550 lmol/m2/s and 20 °C respectively).
Photosynthetic capacity
The fluorescence yield (F) and maximal yield (F 0m) were measuredusing a PAM Chlorophyll Fluorometer (Walz GmbH, Effeltrich,Germany), according to the manufacturer’s instructions. Truelight levels were measured with a light metre (Li-COR, Lincoln,NE) and calculations adjusted accordingly, using a correctionfactor of 0.345. After the plants were dark-adapted, fluorescencemeasurements were taken at six levels of increasing irradiance.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Richard V. Espley et al.416

Yield was calculated from fluorescence (F) using the equation(F 0m ! FÞ=F 0m, where F 0m is the maximal fluorescence yield of theilluminated leaf sample (with all PS II centres closed) and F is themaximal fluorescence yield of the leaf sample just prior toapplication of the light pulse (Kooten and Snel, 1990). The ETRwas then calculated using the yield multiplied by the PAR andmultiplied by 0.5 (a single electron requires the absorption of twoquanta) and 0.84 (the fraction of incident light absorbed by aleaf, Genty et al., 1989). ETR versus PAR was plotted and thecurves were used to estimate maximum photosynthetic capacity(PSmax) and photosynthetic efficiency (initial slope of the ETRversus PAR, a).
Metabolite analysis
All samples were ground under liquid nitrogen using an IKA (IKA,Staufen, Germany) grinder. Powders were stored at !80 °C. Forflavonoid extraction and analysis, 50 mg frozen material wasextracted with 75% aqueous methanol and sonicated for 10 min.Subsequent HPLC of flavonoids species was carried out on a LunaC18-reversed phase column (Phenomenex, Torrance, CA) columnwith a gradient of 5–50% acetonitrile in 0.1% trifluoroaceticacid, followed by photodiode array (PDA) detection. PAs weredetected using the phloroglucinol method and expressed as (epi)catechin units (mg/kg DW). Different compounds were identifiedand quantified using authentic standards. Metabolite data werelog transformed before statistical analysis.
For carotenoid extraction and analysis, 50 mg frozen powderwas extracted with chlorophorm, dried and taken up in 1 mLethyl acetate. Carotenoids were separated with a YMC-Packreverse-phase C30 column and eluted compounds were moni-tored continuously with a PDA detector. A Shimadzu RF551spectrofluorometric detector, set at 296 nm excitation and340 nm emission, was used online after the PDA detector toanalyse tocopherols. Absorbance spectra and retention times ofeluting peaks were compared with those of commerciallyavailable carotenoid and tocopherol standards.
Plant gene expression analysis
Apple fruit were collected at five time points: 40, 67, 102, 130 and146 DAFB. RNA was isolated from the fruit peel and cortexseparately (six fruit from three individual transformed lines andWT)and leaves by a method adapted from Chang et al. (1993).FollowingDNase treatment, first strand cDNA synthesis was carriedout using oligo dT according to the manufacturer’s instructions(Invitrogen, Carlsbad, CA). Gene specific primers were designed toa stringent set of criteria to enable application of universal reactionconditions. The sequence of each primer pair and the relevantaccession number are shown in Table S3. qPCR DNA amplificationand analysis was carried out using the LightCycler® 480 System(Roche Diagnostics, Mannheim, Germany), with LightCycler 480software v1.5. All reactions were performed using the LightCycler480 SYBR Green I Master Mix (Roche). Reactions were 5 min at95 °C, followed by 40 cycles of 5 s at 95 °C, 5 s at 60 °C, and 10 sat 72 °C, followed by a 65–95 °C melting curve detection. Fourreplicates were performed using 2.5 lL Master Mix, 0.25 lL eachprimer (10 lM), 1.25 lL diluted cDNA (1 : 50) and nuclease-freewater to a final volumeof 5 lL. Ano-template controlwas includedin each run. Standard curves for each gene were generated using acDNA serial dilution, and resultant PCR efficiency calculations wereimported into the expression analysis. For relative quantification,Actin (CN938023)was selected as the reference genebecause of itsconsistent transcript level throughout fruit tissues and leaves.
Consumer trial
The preference test study was conducted in Sacramento,California. The aim was to determine whether consumers coulddetect differences between New Zealand-grown WT (‘RoyalGala’) and MYB10 apples, by asking them to indicate apreference between the two apples. The limited number of fruitavailable meant that only one set of WT and MYB10 apples couldbe presented per person. An additional three sets were addedwith locally sourced fruit to provide validation for the preferenceratings. The first of these was designed to be a training set, to‘walk’ the consumers through the tasting procedures with theAmerican ‘Gala’ and Pink Lady® apples. This was followed by theWT and MYB10 apples. The third set was designed to have twopieces of apple from the same segment of fruit, to understandhow consumers were rating fruit that were the same taste andtexture. The final set used the same segment of fruit with a verbalprompt that one sample would probably be GM. The aim of thiswas to determine whether receiving information that the applemay be GM before consumption would influence the responses.
Each session was held in two parts. The first part involved atasting component, and the second part involved a groupdiscussion. Four sessions were held with 11–14 consumers in eachgroup. The study involved a preference test where consumerstasted two samples consecutively and indicatedwhether they had apreference for one sample over the other, or liked (or disliked) bothsamples equally. On the same ballot, the consumers were requiredto provide comments about the sensory attributes of the fruit (e.g.taste, texture), including comments about how they thought theapples were similar or different. Since the MYB10 apple had aneasily distinguished appearance, a strict protocol to prevent theparticipants knowing the colour of the apple was deployed, wherethe panellists wore black-out glasses and all fruit were bagged.
Volatile analysis
Fruit (nine MYB10 or seven WT) were stored for 24 days at 4 °C,then held at room temperature until volatiles were measuredafter 3 and 7 days by dynamic headspace. Fruit were packed into1.5-L glass jars and charcoal-filtered air was drawn into thebottom of each jar for 3 h at 40 mL/min. Volatile compoundswere absorbed onto Tenax-TA® adsorbent traps attached to thetop of each jar. After volatile collection, the adsorbent traps wereeluted with diethyl ether and the eluate analysed by GC-MS(Rowan et al., 2009).
Acknowledgements
We thank Tim Holmes for photography and Minna Pesonen forgraphics and Paul Sutherland for help with microscopy. We thankInova Fruit B.V for project support. This work was supported bythe New Zealand Ministry for Science and Innovation (contractCOX0207).
References
Allan, A.C., Hellens, R.P. and Laing, W.A. (2008) MYB transcription factors that
colour our fruit. Trends Plant Sci. 13, 99–102.
Ampomah-Dwamena, C., Dejnoprat, S., Lewis, D., Sutherland, P., Volz, R.K.
and Allan, A.C. (2012) Metabolic and gene expression analysis of apple
(Malus 9 domestica) carotenogenesis. J. Exp. Bot. 63, 4497–4511.
Ban, Y., Honda, C., Hatsuyama, Y., Igarashi, M., Bessho, H. and Moriguchi, T.
(2007) Isolation and functional analysis of a MYB transcription factor gene
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Genetically modified high-anthocyanin apples 417

that is a key regulator for the development of red coloration in apple skin.
Plant Cell Physiol. 48, 958–970.
Borevitz, J.O., Xia, Y., Blount, J., Dixon, R.A. and Lamb, C. (2000) Activation
tagging identifies a conserved MYB regulator of phenylpropanoid
biosynthesis. Plant Cell, 12, 2383–2394.
Bovy, A., de Vos, R., Kemper, M., Schijlen, E., Almenar Pertejo, M., Muir, S.,
Collins, G., Robinson, S., Verhoeyen, M., Hughes, S., Santos-Buelga, C. and
van Tunen, A. (2002) High-flavonol tomatoes resulting from the heterologous
expression of the maize transcription factor genes LC and C1. Plant Cell, 14,
2509–2526.
Brossaud, F., Cheynier, V. and Noble, A.C. (2001) Bitterness and astringency of
grape and wine polyphenols. J. Grape Wine Res. 7, 33–39.
Bulley, S.M., Malnoy, M., Atkinson, R.G. and Aldwinckle, H.S. (2007)
Transformed apples: traits of significance to growers and consumers.
Transgenic Plant J. 1, 267–279.
Burger, J. and Edwards, G.E. (1996) Photosynthetic efficiency, and
photodamage by UV and visible radiation, in red versus green leaf Coleus
varieties. Plant Cell Physiol. 37, 395–399.
Butelli, E., Titta, L., Giorgio, M., Mock, H.-P., Matros, A., Peterek, S., Schijlen,
E.G.W.M., Hall, R.D., Bovy, A.G., Luo, J. and Martin, C. (2008) Enrichment of
tomato fruit with health-promoting anthocyanins by expression of select
transcription factors. Nat. Biotech. 26, 1301–1308.
Chang, S., Puryear, J. and Cairney, J. (1993) A simple and efficient method for
isolating RNA from pinetrees. Plant Mol. Biol. Rep. 11, 113–116.
Chagne, D., Carlisle, C., Blond, C., Volz, R., Whitworth, C., Oraguzie, N.,
Crowhurst, R., Allan, A., Espley, R., Hellens, R. and Gardiner, S. (2007)
Mapping a candidate gene (MdMYB10) for red flesh and foliage colour in
apple. BMC Genomics, 8, 212.
Chalker-Scott, L. (1999) Environmental significance of anthocyanins in plant
stress responses. Photochem. Photobiol. 70, 1–9.
Christie, P.J., Alfenito, M.R. and Walbot, V. (1994) Impact of low-temperature
stress on general phenylpropanoid and anthocyanin pathways—
enhancement of transcript abundance and anthocyanin pigmentation in
maize seedlings. Planta, 194, 541–549.
Davuluri, G.R., van Tuinen, A., Fraser, P.D., Manfredonia, A., Newman, R.,
Burgess, D., Brummell, D.A., King, S.R., Palys, J., Uhlig, J., Bramley, P.M.,
Pennings, H.M.J. and Bowler, C. (2005) Fruit-specific RNAi-mediated
suppression of DET1 enhances carotenoid and flavonoid content in
tomatoes. Nat. Biotechnol. 23, 890–895.
Dixon, R.A. and Paiva, N.L. (1995) Stress-induced phenylpropanoid metabolism.
Plant Cell, 7, 1085–1097.
Enfissi, E.M.A., Barneche, F., Ahmed, I., Lichtle, C., Gerrish, C., McQuinn, R.P.,
Giovannoni, J.J., Lopez-Juez, E., Bowler, C., Bramley, P.M. and Fraser, P.D.
(2010) Integrative transcript and metabolite analysis of nutritionally enhanced
DE-ETIOLATED1 downregulated tomato fruit. Plant Cell, 22, 1190–1215.
Espley, R.V., Hellens, R.P., Putterill, J., Stevenson, D.E., Kutty-Amma, S. and
Allan, A.C. (2007) Red colouration in apple fruit is due to the activity of the
MYB transcription factor, MdMYB10. Plant J. 49, 414–427.
Espley, R.V., Brendolise, C., Chagne, D., Kutty-Amma, S., Green, S., Volz, R.,
Putterill, J., Schouten, H.J., Gardiner, S.E., Hellens, R.P. and Allan, A.C. (2009)
Multiple repeats of a promoter segment causes transcription factor
autoregulation in red apples. Plant Cell, 21, 168–183.
Feild, T.S., Lee, D.W. and Holbrook, N.M. (2001) Why leaves turn red in
autumn. The role of anthocyanins in senescing leaves of red-osier dogwood.
Plant Physiol. 127, 566–574.
Felicetti, D.A. and Schrader, L.E. (2009) Changes in pigment concentrations
associated with sunburn browning of five apple cultivars. I. Chlorophylls and
carotenoids. Plant Sci. 176, 78–83.
Feller, A., Machemer, K., Braun, E.L. and Grotewold, E. (2011) Evolutionary and
comparative analysis of MYB and bHLH plant transcription factors. Plant J. 66,
94–116.
Genty, B., Briantais, J.M. and Baker, N.R. (1989) The relationship between the
quantum yield of photosynthetic electron-transport and quenching of
chlorophyll fluorescence. Biochim. Biophys. Acta, 990, 87–92.
Gould, K.S., Kuhn, D.N., Lee, D.W. and Oberbauer, S.F. (1995) Why leaves are
sometimes red. Nature, 378, 241–242.
Gould, K.S., Vogelmann, T.C., Han, T. and Clearwater, M.J. (2002) Profiles of
photosynthesis within red and green leaves of Quintinia serrata. Physiol.
Plant. 116, 127–133.
Harborne, J.B. and Williams, C.A. (2000) Advances in flavonoid research since
1992. Phytochemistry, 55, 481–504.
Jiang, Y.M. (2000) Role of anthocyanins, polyphenol oxidase and phenols in
lychee pericarp browning. J. Sci. Food Agric. 80, 305–310.
Kader, F., Haluk, J.P., Nicolas, J.P. and Metche, M. (1998) Degradation of
cyanidin 3-glucoside by blueberry polyphenol oxidase: Kinetic studies and
mechanisms. J. Agric. Food Chem. 46, 3060–3065.
Knee, M. (1972) Anthocyanin, carotenoid, and chlorophyll changes in the peel
of Cox’s orange pippin apples during ripening on and off the tree. J. Exp. Bot.
23, 184–196.
Kooten, O. and Snel, J.F.H. (1990) The use of chlorophyll fluorescence
nomenclature in plant stress physiology. Photosynth. Res. 25, 147–150.
Koponen, J.M., Happonen, A.M., Mattila, P.H. and Torronen, A.R. (2007)
Contents of anthocyanins and ellagitannins in selected foods consumed in
Finland. J. Agric. Food Chem. 55, 1612–1619.
Lane, W.D., Bhagwat, B., Wahlgren, S. and Armstrong, J.D. (2003) Apple
micrografting protocol to establish transgenic clones on field ready rootstock.
HortTechnology, 13, 641–646.
Lin-Wang, K., Bolitho, K., Grafton, K., Kortstee, A., Karunairetnam, S., McGhie,
T., Espley, R., Hellens, R. and Allan, A. (2010) An R2R3 MYB transcription
factor associated with regulation of the anthocyanin biosynthetic pathway in
Rosaceae. BMC Plant Biol. 10, 50.
Martin, C. and Paz-Ares, J. (1997) MYB transcription factors in plants. Trends
Genet. 13, 67–73.
Mayne, S.T. (1996) Beta-carotene, carotenoids, and disease prevention in
humans. FASEB J. 10, 690–701.
Muir, S.R., Collins, G.J., Robinson, S., Hughes, S., Bovy, A., Ric De Vos, C.H.,
van Tunen, A.J. and Verhoeyen, M.E. (2001) Overexpression of petunia
chalcone isomerase in tomato results in fruit containing increased levels of
flavonols. Nat. Biotechnol. 19, 470–474.
National Cancer Institute (2000) A day for better health program evaluation
report. http://dccps.nci.nih.gov/5ad_exec.html (last accessed 1 March
2006).
Neill, S.O. and Gould, K.S. (2003) Anthocyanins in leaves: light attenuators or
antioxidants? Funct. Plant Biol. 30, 865–873.
Paz-Ares, J., Ghosal, D., Wienand, U., Peterson, P.A. and Saedler, H. (1987) The
regulatory c1 locus of Zea mays encodes a protein with homology to myb
proto-oncogene products and with structural similarities to transcriptional
activators. EMBO J. 6, 3553–3558.
Pourcel, L., Routaboul, J.M., Cheynier, V., Lepiniec, L. and Debeaujon, I. (2007)
Flavonoid oxidation in plants: from biochemical properties to physiological
functions. Trends Plant Sci. 12, 29–36.
Ramsay, N.A. and Glover, B.J. (2005) MYB-bHLH-WD40 protein complex and
the evolution of cellular diversity. Trends Plant Sci. 10, 63–70.
Rein, D., Schijlen, E., Kooistra, T., Herbers, K., Verschuren, L., Hall, R.,
Sonnewald, U., Bovy, A. and Kleemann, R. (2006) Transgenic flavonoid
tomato intake reduces C-reactive protein in human C-reactive protein
transgenic mice more than wild-type tomato. J. Nutr. 136, 2331–2337.
Rommens, C.M., Richael, C.M., Yan, H., Navarre, D.A., Ye, J., Krucker, M. and
Swords, K. (2008) Engineered native pathways for high kaempferol and
caffeoylquinate production in potato. Plant Biotechnol. J. 6, 870–886.
Rowan, D.D., Hunt, M.B., Alspach, P.A., Whitworth, C.J. and Oraguzie, N.C.
(2009) Heritability and genetic and phenotypic correlations of Apple
(Malus 9 domestica) fruit volatiles in a genetically diverse breeding
population. J. Agric. Food Chem. 57, 7944–7952.
Schouten, H.J. and Jacobsen, E. (2008) Cisgenesis and intragenesis, sisters in
innovative plant breeding. Trends Plant Sci. 13, 260–261.
Schouten, H.J., Krens, F.A. and Jacobsen, E. (2006) Do cisgenic plants warrant
less stringent oversight? Nat. Biotech. 24, 753–753.
Schroeter, H., Heiss, C., Balzer, J., Kleinbongard, P., Keen, C.L., Hollenberg,
N.K., Sies, H., Kwik-Uribe, C., Schmitz, H.H. and Kelm, M. (2006) (–)-
Epicatechin mediates beneficial effects of flavanol-rich cocoa on vascular
function in humans. Proc. Natl Acad. Sci. USA 103, 1024–1029.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Richard V. Espley et al.418

Steyn, W.J., Wand, S.J.E., Holcroft, D.M. and Jacobs, G. (2002) Anthocyanins in
vegetative tissues: a proposed unified function in photoprotection. New
Phytol. 155, 349–361.
Takos, A.M., Jaffe, F.W., Jacob, S.R., Bogs, J., Robinson, S.P. and Walker, A.R.
(2006a) Light-induced expression of a MYB gene regulates anthocyanin
biosynthesis in red apples. Plant Physiol. 142, 1216–1232.
Takos, A.M., Ubi, B.E., Robinson, S.P. and Walker, A.R. (2006b) Condensed
tannin biosynthesis genes are regulated separately from other flavonoid
biosynthesis genes in apple fruit skin. Plant Sci. 170, 487–499.
Tohge, T., Nishiyama, Y., Hirai, M.Y., Yano, M., Nakajima, J., Awazuhara, M.,
Inoue, E., Takahashi, H., Goodenowe, D.B., Kitayama, M., Noji, M.,
Yamazaki, M. and Saito, K. (2005) Functional genomics by integrated
analysis of metabolome and transcriptome of Arabidopsis plants over-
expressing an MYB transcription factor. Plant J. 42, 218–235.
Tsao, R., Yang, R., Young, J.C. and Zhu, H. (2003) Polyphenolic profiles in eight
apple cultivars using n (HPLC). J. Agric. Food Chem. 51, 6347–6353.
Vidal, S., Courcoux, P., Francis, L., Kwiatkowski, M., Gawel, R., Williams, P.,
Waters, E. and Cheynier, V. (2004a) Use of an experimental design approach
for evaluation of key wine components on mouth-feel perception. Food Qual.
Pref. 15, 209–217.
Vidal, S., Francis, L., Noble, A., Kwiatkowski, M., Cheynier, V. and Waters, E.
(2004b) Taste and mouth-feel properties of different types of tannin-like
polyphenolic compounds and anthocyanins in wine. Anal. Chim. Acta, 513,
57–65.
Volz, R., Oraguzie, N., Whitworth, C., How, N., Chagne, D., Carlisle, C. and
Gardiner, S. (2009) Red flesh breeding in apple—progress and challenges.
Acta Hort. 814, 337–342.
Zhu, Y., Evans, K. and Peace, C. (2010) Utility testing of an apple skin color
MdMYB1 marker in two progenies. Mol. Breed., 27, 1–8.
Zvi, M.M.B., Shklarman, E., Masci, T., Kalev, H., Debener, T., Shafir, S., Ovadis,
M. and Vainstein, A. (2012) PAP1 transcription factor enhances production of
phenylpropanoid and terpenoid scent compounds in rose flowers. New
Phytol. 195, 335–345.
Supporting informationAdditional Supporting information may be found in the onlineversion of this article:
Figure S1 The workflow from transformation to fruit harvest.Figure S2 Measurements of leaf width, leaf length and internodelength from MYB10 and control lines.Figure S3 Characteristics of MYB10 fruit.Figure S4 Relative gene expression of DFR, UFGT and MYB10 in‘Royal Gala’ WT andMYB10 lines 1, 3 and 4, leaf, peel and cortextissue.Figure S5 Relative expression of carotenoid pathway genes.Figure S6 Comparison of headspace volatiles produced byMYB10 and WT apple fruit.Table S1 Growth regime for the MYB10 lines and WT ‘RoyalGala’.Table S2 Carotenoid levels in WT ‘Royal Gala’ and MYB10transgenic apples.Table S3 Primer sequences used in qPCR analysis of fruit tissue.Appendix S1 Supplemental experimental procedures.
Please note: Wiley-Blackwell are not responsible for the contentor functionality of any supporting materials supplied by theauthors. Any queries (other than missing material) should bedirected to the corresponding author for the article.
ª 2012 The AuthorsPlant Biotechnology Journal ª 2012 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd, Plant Biotechnology Journal, 11, 408–419
Genetically modified high-anthocyanin apples 419




















