
Alternative Female Choice Tactics in the Scorpionfly ...
18
AMER. ZOOL., 24:367-383 (1984) Alternative Female Choice Tactics in the Scorpionfly Hylobittacus apicalis (Mecoptera) and Their Implications 1 RANDY THORNHILL Department of Biology, University of New Mexico, Albuquerque, New Mexico 87131 SYNOPSIS. Females of the scorpionfly Hylobittacus apicalis choose mates on the basis of material benefits (nuptial arthropod prey size) and probably on the basis of genetic benefits males deliver at mating. Females feed on the male's prey throughout copulation. They prefer males with large prey as mates and often refuse males who present small prey. That females may value male genetic quality is suggested by differences in ability of males to obtain large prey, which if inherited would influence offspring fitness, and by females often terminating mating with males with small prey before they transfer any sperm or a complete ejaculate. Females hunt only when males with prey are not available because hunting exposes individuals to predators. Female Hylobittacus apicalis exhibit alternative mate choice tactics, which are condition-dependent in expression and probably comprise a conditional strategy. Body size, recent feeding history, and male availability determine how discriminating an individual female actually is, and these conditions may determine the value of material and genetic benefits in mate choice decisions. The results suggest that female choice controls male behavior. When females become choosy, males are forced to obtain rare large prey despite the increased risks to males associated with this behavior. The implications of the findings on H. apicalis are discussed in relation to condition- dependent female choice patterns in other species and the evolutionary maintenance of female choice. INTRODUCTION The aspect of sexual selection involving female choice is poorly understood and controversial (Williams, 1975; Maynard Smith, 1978; Thornhill, 1980a; Taylor and Williams, 1981; Lambert et ai, 1982). Some authors deny that female choice has been demonstrated (Lambert et ai, 1982). Others question its importance, at least when females only value male genetic qual- ity in choice decisions (and not male-pro- vided material benefits), because the selec- tive maintenance of adaptive choice is often difficult to envision (see discussion in ref- erences immediately above). Two major problems in studies during the 1970s of the role of female choice in evolution have been 1) the difficulty of sep- arating male-male competition and female choice as agents causing differential suc- cess of males (see review in Thornhill, 1980a) and 2) the cryptic nature of female choice when it occurs during or after mat- ing (Thornhill, 1983). Bittacid scorpion- 1 From the Symposium on Alternative Reproductive Tactics presented at the Annual Meeting of the Amer- ican Society of Zoologists, 27-30 December 1982, at Louisville, Kentucky. flies (Mecoptera: Bittacidae) have been use- ful in solving these problems: males provide females with a nuptial arthropod gift dur- ing mating, and even after direct male-male competition for prey has ended, choosy females operate to influence which males secure copulations, pass complete ejacu- lates, and (probably) fertilize eggs (Thorn- hill, 1976, 1977, 1979a, 19806, c, 1983; Alcock, 1979a; D. T. Gwynne, personal communication). There has been little empirical work directed at elucidating the evolution of within species variation in female choice and its adaptive significance. Recent research on the evolution of alternative reproductive behaviors has focused on males almost entirely (e.g., see reviews in Alcock, 19796; Cade, 1980; Dawkins, 1980; Rubenstein, 1980; Thornhill, 1981; Thornhill and Alcock, 1983; Eberhard, 1982; Gross, 1982; but see Brockmann et al., 1979; Noonan, 1981; Strassmann, 1981; West-Eberhard, 1981). Further- more, in much of the theoretical work on mate choice, females are viewed as pos- sessing one population-wide or species-spe- cific choice pattern, which may be based on an absolute or a relative criterion (or 367 Downloaded from https://academic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Alternative Female Choice Tactics in the Scorpionfly ...

AMER. ZOOL., 24:367-383 (1984)
Alternative Female Choice Tactics in the ScorpionflyHylobittacus apicalis (Mecoptera) and Their Implications1
RANDY THORNHILL
Department of Biology, University of New Mexico,Albuquerque, New Mexico 87131
SYNOPSIS. Females of the scorpionfly Hylobittacus apicalis choose mates on the basis ofmaterial benefits (nuptial arthropod prey size) and probably on the basis of genetic benefitsmales deliver at mating. Females feed on the male's prey throughout copulation. Theyprefer males with large prey as mates and often refuse males who present small prey.That females may value male genetic quality is suggested by differences in ability of malesto obtain large prey, which if inherited would influence offspring fitness, and by femalesoften terminating mating with males with small prey before they transfer any sperm ora complete ejaculate. Females hunt only when males with prey are not available becausehunting exposes individuals to predators. Female Hylobittacus apicalis exhibit alternativemate choice tactics, which are condition-dependent in expression and probably comprisea conditional strategy. Body size, recent feeding history, and male availability determinehow discriminating an individual female actually is, and these conditions may determinethe value of material and genetic benefits in mate choice decisions. The results suggestthat female choice controls male behavior. When females become choosy, males are forcedto obtain rare large prey despite the increased risks to males associated with this behavior.
The implications of the findings on H. apicalis are discussed in relation to condition-dependent female choice patterns in other species and the evolutionary maintenance offemale choice.
INTRODUCTION
The aspect of sexual selection involvingfemale choice is poorly understood andcontroversial (Williams, 1975; MaynardSmith, 1978; Thornhill, 1980a; Taylorand Williams, 1981; Lambert et ai, 1982).Some authors deny that female choice hasbeen demonstrated (Lambert et ai, 1982).Others question its importance, at leastwhen females only value male genetic qual-ity in choice decisions (and not male-pro-vided material benefits), because the selec-tive maintenance of adaptive choice is oftendifficult to envision (see discussion in ref-erences immediately above).
Two major problems in studies duringthe 1970s of the role of female choice inevolution have been 1) the difficulty of sep-arating male-male competition and femalechoice as agents causing differential suc-cess of males (see review in Thornhill,1980a) and 2) the cryptic nature of femalechoice when it occurs during or after mat-ing (Thornhill, 1983). Bittacid scorpion-
1 From the Symposium on Alternative ReproductiveTactics presented at the Annual Meeting of the Amer-ican Society of Zoologists, 27-30 December 1982, atLouisville, Kentucky.
flies (Mecoptera: Bittacidae) have been use-ful in solving these problems: males providefemales with a nuptial arthropod gift dur-ing mating, and even after direct male-malecompetition for prey has ended, choosyfemales operate to influence which malessecure copulations, pass complete ejacu-lates, and (probably) fertilize eggs (Thorn-hill, 1976, 1977, 1979a, 19806, c, 1983;Alcock, 1979a; D. T. Gwynne, personalcommunication).
There has been little empirical workdirected at elucidating the evolution ofwithin species variation in female choiceand its adaptive significance. Recentresearch on the evolution of alternativereproductive behaviors has focused onmales almost entirely (e.g., see reviews inAlcock, 19796; Cade, 1980; Dawkins, 1980;Rubenstein, 1980; Thornhill, 1981;Thornhill and Alcock, 1983; Eberhard,1982; Gross, 1982; but see Brockmann etal., 1979; Noonan, 1981; Strassmann,1981; West-Eberhard, 1981). Further-more, in much of the theoretical work onmate choice, females are viewed as pos-sessing one population-wide or species-spe-cific choice pattern, which may be basedon an absolute or a relative criterion (or
367
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

368 RANDY THORNHILL
criteria) (e.g., Fisher, 1930; Zahavi, 1975).However, aspects of mating system theory,particularly the original outline of the evo-lution of resource-defense polygyny in birds(Verner and Willson, 1966; Orians, 1969)and its recent extensions (Bradbury andVehrencamp, 1977; Emlen and Oring,1977; Borgia, 1979; Searcy, 1979; Brown,1981; Downhower and Brown, 1981; Wit-tenberger, 1981), credit females with adap-tive flexibility of mate choice (also see Bur-ley and Moran, 1979;Janetos, 1980;Burley,1981). In this paper I focus on the scor-pionfly Hylobittacus apicalis (Mecoptera:Bittacidae) in order to address the evolu-tion of intra- and interpopulation variationin female choice behavior, and I brieflyreview literature on other taxa pertainingto this topic.
BACKGROUND INFORMATION
The scorpionfly (also called hangingfly)Hylobittacus apicalis is an ideal species forstudies of behavior under field conditions.It is diurnally active, mating and catchingprey arthropods (mainly insects) fre-quently throughout the day. Adult H. api-calis spend their life among the low herbs.Adults are medium-sized insects (bodylength about 1.9 cm). The sexes are distin-guishable from a distance of a few metersby differences in the terminal abdominalsegments and in flight pattern. Both sexesare slow flyers and make only short flightsamong the herbs.
My published work on H. apicalis wasconducted from 1971-1979 on large pop-ulations in southeastern Michigan (Thorn-hill, 1976, 1977, 1979a, b, 19806, c). Inthese high density settings, females exhibita large degree of mate choice. In summaryof my published work: the sequence of sex-ual behavior in H. apicalis begins when amale catches for himself or steals fromanother male a prey insect and begins tofeed on it. After feeding about 10 min,males usually discard the prey if the preyis small (<16 mm2 in surface area—length x width or height whichever isgreatest) or contains little nutrient due toits use in a previous copulation. Only about10 percent of males retain small prey (< 16mm2) prior to premating flights and usethese prey as nuptial gifts. If the prev is
large (^16 mm2), after briefly feeding andwhile still holding the prey in his hind tarsi,the male initiates short premating flightsamong the herbs. After each flight the maleexposes his abdominal pheromone-dis-persing vesicles. Females respond to malesfrom a distance by olfaction. The male pre-sents the nuptial prey to arriving females,who feed as the male attempts to couple.If the prey is large, the female will couplewith the male and copulation will be ter-minated by the male after a duration of 20min or more, a time sufficient for completeinsemination of the female. If the prey issmall the female may depart after samplingit but before mating (50 percent of obser-vations involving small prey). If the femaleallows mating it will be terminated by thefemale in less than 20 min (x = 6 min).About 5 min of mating are required forany sperm transfer. This is roughly theaverage mating duration at which femalesterminate mating with males possessingsmall prey when such males are allowed tomate. Between 5 and 20 min of copulationthere is a direct positive relationshipbetween the number of sperm transferredand mating duration. Only the female feedson the prey during copulation and she feedsthroughout copulation. Only after matingwith males bearing large prey do femalesbecome sexually nonreceptive (i.e., do notrespond to male pheromone and thereforedo not mate) and begin laying eggs. Theperiod of oviposition and nonreceptivity is3-4 hr, after which females become recep-tive and mate again, preferring a mate witha large prey.
Although H. apicalis males are usuallyselective of prey used as nuptial offerings,hunting females do not discard small prey.However when males become abundantseasonally, females rarely catch their ownprey but depend on prey provided by males.
The adaptive significance of femalechoice is expected to relate to two classesof reproductive benefits gained by discrim-inating females: material and genetic.Material benefits to choosy females includeresources (e.g., nuptial prey) and protec-tion. Genetic benefits include increased fit-ness resulting when they mate with maleswith superior genetic constitutions.
Choosy female H. apicalis clearly receive
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

FEMALE CHOICE TACTICS IN A SCORPIONFLY 369
material benefits from preferred mates.Females are more fecund when they selectmales with large, as opposed to small, prey(Thornhill, 1976, 19806). Also, femaleswho prefer males with large prey probablyhave enhanced survival via reduction intheir exposure to web-building spiders, themost important predators on H. apicalis.Spider webs are encountered during for-aging flights; thus the additional huntingby males leads to greater exposure to spi-ders and higher mortality (Thornhill,19796, 19806). Discrimination against cer-tain males may result in some increase infemale exposure to web-building spidersbecause they have to fly to another male.However, males with large prey are usuallyavailable, females can locate a second malerapidly, and the flight path of a femaletoward a second male is probably verydirect because of pheromone emission bythe male. Females move around littlebetween matings.
Choosy female H. apicalis also mayreceive genetic benefits from preferredmates. This suggestion derives from theobservation that females who mate with amale possessing small prey often terminatecopulation before sperm transfer. If hunt-ing prowess and the ability to obtain largeprey are heritable in bittacid scorpionflies,which may be the case (Thornhill, 1983),discriminating females may produce suc-cessful offspring of both sexes. The sonsof discriminating females would be attrac-tive to females and perhaps also reproducemore because of increased survival asso-ciated with greater prey securing ability;their daughters might have better survival(and hence reproduction) when requiredto catch prey on their own.
In principle, female animals may choosemales before, during or after mating(Thornhill and Alcock, 1983; Thornhill,1983). Premating female choice includescircumstances in which females do notrespond to certain males or refuse to matewith them, preferring others. Femalechoice during mating is female termina-tion of mating with certain males beforeinsemination or complete insemination.Female insects store sperm. In insect speciesin which females mate multiple times, ejac-ulates from more than one male overlap
temporally in the sperm storage structure(Parker, 1970; Gwynne, 1984a). When thenumber of sperm transferred is related tomating duration (as in H. apicalis, Thorn-hill, 1976), females can bias fertilizationrates toward preferred mates by control ofmating duration. Postmating female choiceoccurs when females use primarily or onlysperm of preferred mates for fertilizingeggs. Postmating female choice may resultfrom a female's ability to reduce or elim-inate completely the fertilizing potential ofejaculates from inferior mates by digestingthem, moving them to a location away fromthe site of fertilization, remating whenthere is sperm precedence and last maleadvantage, or by the pattern of egg layingwhen fertilization occurs during oviposi-tion as in insects (Lloyd, 1979; Sivinski,1980; Walker, 1980; Thornhill and Alcock,1983).
All three forms of female choice (pre-mating, postmating and during mating)occur in H. apicalis. Females discriminatebefore mating, preferring males with largeprey. They discriminate during mating byadjusting copulation duration. Mating isterminated when the female ceases feed-ing, pulls free from the male, and flies away.Female H. apicalis who receive small nup-tial gifts do not lay eggs, remate immedi-ately and continue to remate until they matewith a male with a large prey. The remat-ing behavior of females is postmatingfemale choice, and it will result in differ-ential reproduction among males if the lastmale to mate before egg laying fertilizesmost of the eggs of the female. Last malemating advantage in fertilization is thegeneral pattern in insects (Parker, 1970),especially in those species in which malesprovide females with material benefits dur-ing mating (Gwynne, 1984a) as in scorpion-flies.
WITHIN POPULATION VARIATION INFEMALE CHOICE IN HYLOBITTACUS APICALIS
Hypotheses
Fifty percent of the time female H. api-calis in the large populations in southeast-ern Michigan do not allow a male with smallprey to copulate, and in the remaining casesthey mate only for a brief time and not
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

370 RANDY THORNHILL
long enough for any sperm or a completeejaculate to be transferred. I examined thepossibility of predictable differencesbetween females behaving in these alter-native ways toward males, hypothesizingthat large females may be less discriminat-ing in their mate choice. The benefit asso-ciated with large body size in female H.apicalis is higher potential fecundity (num-ber of eggs/ovary: n = 39 females, r =0.82, P < 0.001; Thornhill, 19806). Thecontribution of size to survival (and hencereproduction) depends on food resourcelevels, and the greater cost of maintenancecaused by large size is met only when suf-ficient food is available (see Downhower,1976 for evidence and discussion). For largefemales, the benefit of receiving food viacopulations with males with small preymight override the presumed cost ofreceiving some sperm from an inferiorhunter.
Female age may be an important factorinfluencing the extent of female choice. Afemale might be relatively nondiscriminat-ing of mates early in her adult life in orderto receive male gametes and begin repro-duction early. As a female ages she maybecome more discriminating in order toobtain 1) a mate different from the com-mon male phenotype (e.g., rare male phe-notype) if her choice is directed at diver-sifying the gene combinations of her broodmembers or 2) a genetically fit male in someother sense. On the other hand, if femalechoosiness is positively related to femalereproductive value, an age pattern differ-ent (probably often the reverse) from thatdescribed is expected.
Premating female choice on the basis ofnuptial prey size in H. apicalis under highdensity is an important form of sexualselection on males (Thornhill, 1977, 19806,c). But in normally high density popula-tions, when male numbers are low (eitherearly or late in the flight season), anecdotalobservations in 1971-1974 suggested thatfemales show less premating choice thanunder high male density (Thornhill, 1976,1977). Also at low density females fre-quently catch their own prey, indicatingthat they incur foraging risks when male-provided food is less available. These
observations led to the hypothesis thatpotential mate availability is an importantfactor in premating female choice deci-sions in H. apicalis. Males emerge slightlyearlier and live a shorter time than females,and almost all adult emergence occurs overa period of several days at the beginningof the flight period (seasonal period inwhich adults are on the wing) (Thornhill,1974; Byers and Thornhill, 1983). Thuslate and early in the flight period femalesshould have reduced mate choice optionsbecause density of males with prey is lowerthan at peak population density. Even atpeak density only a fraction of males areholding prey at any one time (2-12% ofmales) (Thornhill, 19806), but females canstill find a second male with prey rapidly(x = 18 min) after rejecting a male withsmall prey. I felt that females would relaxmate choice standards with a reduction inavailable mates to lower risks and energyexpenditure associated with finding maleswith large prey or finding their own prey.
Methods
I studied female body size and its effecton mate choice via field observations andlaboratory experiments. The effect of ageon female choice behavior was examinedin the laboratory.
Females (n = 120) were observed as theyinteracted with males with small prey dur-ing 18 June-10 July 1980 and 19 June-1July 1981 at the University of Michigan'sEdwin S. George Reserve near Pinckney,MI. After sighting a male releasing pher-omone and holding a small prey I observedhim until he attracted a female and duringhis interaction with the female. In 80% ofthe interactions I retrieved the prey aftermating and measured its surface area. Inthe remaining cases the prey was droppedat the termination of interaction and it wasnecessary to estimate its size (see Thorn-hill, 19806 for techniques of observationand obtaining or estimating prey size).Because these field observations were madeduring the seasonal peak in adult numbersand observations were made at several siteson the George Reserve it is likely that mostobservations represent different individu-als. For each male-female interaction I
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

FEMALE CHOICE TACTICS IN A SCORPIONFLY 371
recorded whether the male was rejected(i.e., the female did not allow copulation)or accepted (i.e., female allowed copula-tion), the duration of copulation and thesex terminating copulation (see Thornhill,1977 for unambiguous technique fordetermining which sex of H. apicalis ter-minates a copulation). At the terminationof all interactions I captured the pair withan insect net and measured the left fore-wing (right forewing when left was incom-plete) and hind femur of each sex with ver-nier calipers. Forewing length is an accuratemeasure of body size in bittacids (Thorn-hill, 1983). Since forewing length and hindfemur length are highly correlated (r =0.92, n = 40, P < 0.001) I use only fore-wing measurements in subsequent discus-sion.
Experiments were used to further exam-ine the influence of female size on femalechoice and to study the relation betweenfemale age and mate choice. Recentlyemerged, incompletely-sclerotized femaleswere collected from the field and housedin laboratory cages containing abundantfood (house crickets of various sizes) andwater for one day prior to experiments.Females become fully sclerotized and readyto mate within 12-16 hr after emergence(Thornhill, 1976). Thus, the females usedin experiments were approximately thesame age and had no prior mating expe-rience. Males of unknown age and historywere collected from the field and cagedseparately from females.
Experiment 1 addresses female body sizeand mate choice by food-deprived females.Previously well-fed females (see immedi-ately above) were placed in large terrariaand provided with water, but not food, forthe night (about 7 PM-7 AM). In the labo-ratory adult H. apicalis will feed during thenight if food is abundantly available in theircage, but in nature both males and femalesare quiescent at night and do not feed ormate. Thus the prevention of females fromfeeding for a 12 hr night is a normal expe-rience. The forewings of females weremeasured and each female was distinctlymarked using model airplane paint (seeThornhill, 1977 for discussion of markingtechnique). Female size (forewing length)
in natural populations ranges from about12-20 mm (see below). I used nonoverlap-ping size ranges for designating small andlarge females in experiments: large =17—20 mm; small = 12-15. In the morning (7-9 AM) experimental terraria were set up asfollows: three individually-marked maleswere placed in each of 30, 52 x 25 x 31cm terraria and given small house cricketsas nuptial offerings (range of surface areasof prey = 9-11 mm2). When all three malesbegan release of pheromone, a small anda large female were added to each terrar-ium (see Thornhill, 1976 for further dis-cussion of methods). Three males were usedin each terrarium in order to mimic andkeep constant an environment of high mateavailability for females, a factor that influ-ences female choice (see below). Male-female interactions in each terrarium wereobserved for one hour after placing malesand females together. I recorded whetherfemales rejected or accepted a male(s),whether copulations were male or femaleterminated, copulation duration, and num-ber of males females interacted with.
Experiment II was set up identical to thefirst experiment in every detail but one:the 30 small and 30 large females wereeach given a large cricket (range of surfaceareas: 40-50 mm2) after their quiescentovernight period without food. Followingthe feeding, females were placed withmales. Females fed on their large cricketsfor 40-75 min. This experiment was con-ducted to determine if recent feeding wouldinfluence a female's decision to mate witha male with a small prey and if the decisionis related to female body size.
I tested the hypothesis that density ofmales is a factor influencing the extent offemale choice via observations of seasonalvariation in premating and mating behav-ior of males and females in 1980 and 1981at a site near the Edwin S. George Reserve(site is 4.5 mi west of Ann Arbor, MI onScio-Church Road; see Thornhill, 1974 forsite description). Observations were col-lected during approximately the first fivedays of adult emergence of H. apicalis andduring peak population density at the sitein 1980 and 1981. The first five day periodcorresponds to 10-14 June in 1980 and
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

372 RANDY THORNHILL
11-15 June in 1981. The second period ofobservation was 18-22 June 1980 and 19-23 June 1981. During the first five days ofadult emergence, population density is low(one to two individuals/3 x 3 m section ofhabitat). By 18 June in 1980 and 19 Junein 1981 population size had climbed todensities similar to peak population sizesin previous years (10—18 individuals/3 x3 m section of habitat; x = 12.4 indi-viduals). In the first five days of emergencethe population is male-biased (2 or 3 males/female), but absolute male density is stilllow. At the time of population peak sexratio is about unity. Emergence is very syn-chronous and the great majority of indi-viduals (95 + %) emerge within the first 10days. Thus my observations addresschanges in female choice behavior offemales at two stages in their adult life,which corresponds to early and midadultlife. (See Thornhill [1974] for data on pop-ulation size, sex ratio and seasonality of H.apicalis at this site.)
Male-female interactions were observedon 10, 14, 18, and 22 June in 1980 and11, 15, 19, and 23 June in 1981. Each dateequals a sample. Two replicates of obser-vations were recorded per sample date.Replicates of each sample were obtainedfrom different sections of the habitat inorder to reduce the likelihood of observingthe same individual more than once.
Each replicate consisted of the followingthree sets of information: 1) the surfacearea of prey carried by 30 males duringpremating flights (i.e., prey that was to beused by males as nuptial offerings), 2) thesurface area of nuptial prey in 25 copulat-ing pairs, and 3) the recording of the occur-rence and duration of copulation between15 pairs of males with small prey andfemales, located at the beginning of theirpremating interaction.
Two-tailed /-tests are used to comparemean differences. In all cases the variancesof the means compared are not signifi-cantly different.
Results and discussion
Females allowed males to copulate in 62%(75 of 120) of field observations of male-female interactions involving small prey
(<15 mm2) under high population densityat the George Reserve in 1980 and 1981.(This is similar to the 50% copulation fre-quency observed in my previous study basedon a smaller sample size [n = 50; Thornhill,19806].) In the remainder of cases (n = 45)females pulled away from the male aftertasting his prey and flew away without mat-ing. All copulations were terminated byfemales (x ± SD copulation duration = 8.5min ± 2.4, n = 75).
Female body size influences a female'sdecision to mate with a male with smallprey. Females who did not allow males tomate were significantly smaller (x ± SDforewing length = 14.7 ± 1.5, n = 45) thanthose who did (17.0 ± 1.2, n = 75:/ = 8.74,P < 0.001). Furthermore, there is a posi-tive relationship between female body sizeand duration of copulation (n = 75, r =0.62, P < 0.001). There is also a positiverelationship between prey size and copu-lation duration (n = 75, r = 0.52, P <0.001), as is common in my studies on thisand other bittacids (see Thornhill, 1980ft,1983). But there is no significant rela-tionship between female body size andnuptial prey size in the 75 copulations (r =0.12, P> 0.1). Thus, large females aremore likely to copulate with a male withsmall prey and to copulate for a longer timethan small females, and the increase in cop-ulation time of large females is not due totheir mates possessing larger prey thanpossessed by mates of small females. Thesefindings suggest that large females requiremore food which they take from males withsmall prey at the potential cost of receiptof some of their gametes. About 50% ofthe 75 copulations in this sample of obser-vations extended beyond 5 min duration,which corresponds to the time when spermbegins to flow into the female.
That the female makes the decision tocopulate or not with a male possessing smallprey and for how long is indicated by mypublished work (Thornhill, 1976, 1977,19806). This interpretation is further sup-ported by the present study. There is nosignificant difference in male size (fore-wing length in mm) between interactionsinvolving copulation (x ± SD = 15.0 ± 1.7,n = 75) and those without copulation (x ±
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

FEMALE CHOICE TACTICS IN A SCORPIONFLY 373
SD = 16.2 ± 1.8, n = 45) (t = 0.9, 0.2 <P < 0.5). Also there is no significant rela-tionship between male size and matingduration (n = 75, r = 0.12, P > 0.1). Thus,the results on female behavior cannot beexplained by larger males forcing femalesto mate or mate longer.
The above findings on variation in femalebehavior in H. apicalis are further sup-ported by the two laboratory experiments,which were designed to factor out femaleage and specifically examine the role ofbody size in female mating decisions. InExperiment I only four of 30 small femalesmated (13.3%) but 21 of 30 large femalesmated (70.0%) (Yates' x
2 = 17.6, P <0.001), and large females mated for sig-nificantly longer (Table 1) (/ = 4.1, P <0.001). The four small females that mateddid so with the third male encountered inthree cases and the second male encoun-tered in the remaining case. One smallfemale mated twice. Each of the 26 smallfemales that did not mate interacted withall three males within the 60 min periodof observation, and seven of these femalesinteracted with a male (n = 4) or males (n =3) more than once. But the 21 large femalesthat mated did so with the first maleencountered in 19 cases and the secondmale in two cases. All the 21 large femalesmated with at least two males and 17 ofthem mated with all three males during theobservation period. The nine large femalesthat did not mate interacted with all threemales in their environment; five of thesefemales visited one male more than once.
On the other hand, both large and smallfemales infrequently mated with males withsmall prey when fed immediately prior toexposure to such males (Table 1, Experi-ment II), and there is no significant differ-ence in premating choice behavior betweenfemale size categories (Yates' x2 — 0.10,P > 0.1). Large fed-females copulatedlonger than small fed-females but samplesize is too small for analysis (Table 1,Experiment II). Although the distributionsof mating and nonmating small females arenot significantly different between the twoexperiments (Yates' x2 = 0, P > 0.1), feed-ing radically modified the behavior of largefemales: 21 of 30 large females mated in
TABLE 1. Results of experiments with H. apicalis to ex-amine whether a female's body size influences her willingnessto mate with a male possessing a small cricket (9-11 mm2
surface area) as a nuptial gift.*
Femalesize
SmallLarge
SmallLarge
Female behavior
Accept amate(s)
421
57
Rejectall mates
Expt. I
269
Expt. II2523
Mating duration(5c + SD min)when mating
occurred
5.8 ± 0.912.9 ± 3.2
4.1 ± 1.06.1 ± 1.8
* The experiments differ only in that females inExperiment II were fed a large cricket (40-50 mm2
surface area) immediately prior to exposure to maleswith prey whereas females in Experiment I were not.All females were virgins and approximately the sameage. Large female = 17-20 mm forewing length. Smallfemale = 12-15 mm forewing length. Each experi-ment involved 30 replicates. Each replicate involvedthree males with small prey and one large and onesmall female.
Experiment I, but only 7 of 30 large femalesmated in Experiment II (Yates' x2 = 11.32,P < 0.001). Feeding also affected the dura-tion of mating, especially for large females.Average mating duration for large femalessignificantly declined from 12.9 to 6.1 minbetween experiments (t = 5.31,P < 0.001);as did mating duration of small females (t =2.66, 0.02 < P < 0.05). Furthermore,small females in Experiment II behavedsimilar to small females in Experiment I:all who mated did so with the third maleencountered, and the small females whodid not mate interacted with male aftermale. But the large females in ExperimentII behaved like small females: they matedonly after first interacting with and refus-ing two males.
The experimental results clearly showthat small females are more discriminatingof mates than large females after 12 hr offood deprivation. Since all females werethe same age (approximately) and virgins,age and mating history do not confuseresults. Although I have not directly testedwhether a female's prior mating history(virgin vs. nonvirgin) is important, thechoosiness of small virgin females indicatesthat females do not mate indiscriminatelyearly in life in order to obtain sperm for
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

374 RANDY THORNHILL
quick initiation of reproduction. More-over, the results reveal that large femalesfed prior to placement with males behavelike small females in that they become morediscriminating at two levels: 1) they are lesslikely to accept a male with small prey as amate and 2) if they do accept a male withsmall prey they mate for only a brief dura-tion, terminating copulation at about thetime (6 min) when sperm begins to be deliv-ered.
The results of the field observations andlaboratory experiments discussed thus farindicate that large females because of ahigher maintenance cost may require morefood. In copulating with males with smallprey females risk receipt of sperm frommales who may be inferior genetically.However, females, especially large ones,seem to modify copulation duration in rela-tion to the trade-off between obtaining foodand avoiding sperm from males with smallprey. The interpretation that females incura cost via receipt of inferior sperm whenthey mate for more than 5 min with a malewith small prey assumes that ejaculate com-petition does not completely favor the malewith a large prey who completely insemi-nates a female in a lengthy copulation (20min or more mating time). As discussed,females remain sexually receptive and donot lay eggs until they mate with a malewith a large prey; this is the case even whenfemales mate with several males with smallprey (see Background Information). Theassumption is reasonable because completesperm precedence by the last male to matebefore a female lays eggs is very rare ininsects (Parker, 1970; Gwynne, 1984a).
Field observations near the GeorgeReserve in 1980 and 1981 suggest thatadult population density affects whethermales retain or discard small prey beforerelease of pheromone (A. in Table 2, 1980;A. in Table 3, 1981). The two replicatesfor each sample within each density are notsignificantly different (50% probability test,P > 0.1 in all cases) in either year, and ingeneral, samples within a density are uni-form for each year (the exception is sam-ples for low density in 1981, 50% proba-bility test, P = 0.05). A comparison of thetwo densities for each year reveals highly
significant differences (50% probabilitytests, P < 0.001 for both years): a greaterproportion of males are seen carrying smallprey (<16 mm2) under low adult density.In both years the proportion of males car-rying small prey dropped from over 50%to less than 19% as population densityincreased.
It is not completely clear at this timewhether male decisions about nuptial preysize use reflect changes with age (e.g., thelearning of better hunting techniques) and/or stem from changes in female choicebehavior seasonally—that is, femalesbecome more discriminating with anincrease in mate availability (see below).My previous work reveals that males,immediately after becoming fully sclero-tized, are capable of catching large prey inthe field and laboratory and show a pref-erence for large prey (i.e., discard smallprey after feeding on it) when prey areabundant in the laboratory. This suggeststhat changes in male behavior with age arenot the major factor explaining the greateruse of small nuptial prey when adult num-bers are low. Also, sweep sample studiesconducted in 1977 and 1978 reveal thatalthough the abundance of large prey inthe habitat of H. apicalis in southeasternMichigan varies from 5 to 37% of totalarthropods from sample date to sampledate, there is no seasonal pattern to changesin abundance of large prey; e.g., large preyare not less available in the early portionof the flight period of H. apicalis than atpeak density (Thornhill, 19806, unpub-lished). Thus, I feel that the difference inprey size use by males under low us. highdensity is a function of changes in femalechoice behavior (see below).
Females radically modify choice behav-ior in relation to changes in populationdensity (i.e., mate availability) (B. and C. inTables 2 and 3). First consider the obser-vations of total matings that involved smallprey (B. in Tables 2 and 3). The two rep-licates for each sample in each density arenot significantly different (50% probabilitytest, P > 0.1 in all cases) in either year.Also, samples within each density are sim-ilar for each year (50% probability test,P > 0.1 in all cases) with the exception of
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

FEMALE CHOICE TACTICS IN A SCORPIONFLY 375
TABLE 2. Data on prey sizes in possession of single males and on male-female interactions during low and high populationdensities of Hylobittacus apicalis in 1980.*
Type ofobservation
A. 6S with smallprey/sample
S6 with smallprey/density
B. Of total matings,those involvingsmall prey/sample
Of total matings,those involvingsmall prey/density
C. Of matings involvingsmall prey, thoseterminated by femalebefore 6 min/sample
Of matings involvingsmall prey,those terminated byfemale before 6 min/density
Number {%) observations
Low density-
Sample 1 Sample 2
36(60) 25(41.7)
61 (50.8)
38 (76.0) 25 (50.0)
63 (63.0)
3 (10.0) 2 (6.7)
5 (8.3)
High density
Sample 1 Sample 2
12(20.0) 8(13.3)
20(16.7)
12(24.0) 2(4.0)
14(14.0)
14(46.7) 15(50.0)
29 (48.3)
* Small prey = < 16 mm2. Each sample is two replicates (see Methods).
the two samples for high density in 1980(50% probability test, 0.05 < P < 0.01).However for both years significantlygreater proportions of females (50% prob-ability test, P < 0.001 for each year) wereobserved mating with males with small preyunder low density. Mate availability alsoaffects the point at which females termi-nate matings with males with small prey ifthey allow mating (C. in Tables 2 and 3).There are no significant differencesbetween replicates or samples of observa-tions of matings involving small prey whichwere terminated by the female before 6min for each density in each year (50%probability test, P > 0.1 in all cases). Buthighly significant differences in this femalebehavior were found between densities ineach year (50% probability test, P < 0.001).Under low density only about eight per-cent of females terminated matings withmales possessing small prey before 6 minin both years, but close to 50% did so athigh density in both years. The change infemale copulation duration with density incopulations involving small prey is evenmore dramatic when average mating dura-tions are considered (Table 4). Mean mat-ing durations are similar between repli-
cates of each sample and between samplesof each density in both years. But the effectof density on mean mating duration is sig-nificant (1980, t = 8.0, P < 0.001; 7957,t = 7.3, P < 0.001); mean mating durationdeclines by about 5 min from low to highdensity in each year.
Observations of female choice and nup-tial prey size use indicate major changes inconnection with relative availability ofmates. The observations at low and highdensity reflect the same cohort of individ-uals because emergence is seasonally verysynchronous. The fact that the patterns ofyoung female behavior depicted in theexperimental results (Table 1) are similarto those obtained by observations of olderfemales at high density at the GeorgeReserve and the site near the GeorgeReserve indicates that the low density(young females) and high density (olderfemales) observational comparisons inTables 2, 3 and 4 do not reflect changesin female choice behavior with age or mat-ing experience, but instead reflect femalebehavioral modifications to cope with dif-ferent mate availabilities. Under low maledensity, females are less selective—bothpremating choice and choice during mat-
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

376 RANDY THORNHILL
TABLE 3. Data on prey sizes in possession of single males and on male-female interactions during low and high populationdensities o/Hylobittacus apicalis in 1981*
Type ofobservation
A. $i with smallprey/sample
S6 with smallprey/density
B. Of total matings,those involvingsmall prey/sample
Of total matings,those involvingsmall prey/density
C. Of matings involvingsmall prey, thoseterminated by femalebefore 6 min/sample
Of matings involvingsmall prey,those terminated byfemale before 6 min/density
Number (5
Low density
Sample 1 Sample 2
40 (66.7) 23 (38.3)
66 (55.0)
27 (54.0) 36 (72.0)
63 (63.0)
2 (6.7) 3 (10.0)
5(8.3)
I) observations
High density
Sample 1 Sample 2
11 (18.3) 11 (18.3)
22(18.3)
10 (20.0) 14 (28.0)
24 (24.0)
14(46.7) 14(46.7)
28 (46.7)
* Small prey = < 16 mm2. Each sample is two replicates (see Methods).
ing are relaxed—because this allows them ability on female remating behavior or eggto acquire more food without hunting on laying (i.e., postmating female choice) in H.their own, and presumably they are willing apicalis. My published results on postmat-to pay the cost of receiving sperm from ing female choice in the form of rematinginferior males. Under high density, females behavior were obtained with high densityare very selective in terms of choice in the females of assorted sizes, and althoughcontexts of premating and during mating, there was considerable variation in the data,because they have more mates to evaluate most females show the pattern of only ini-and mates can be evaluated quickly. But dating sexual nonreceptivity and egg lay-under high density large females still cop- ing following mating with a male with largeulate longer than small females and prob- prey (Thornhill, 1976). Future work willably receive more sperm from inferior examine the prediction that under low matemates as a result. availability females will not require a mat-
I have not directly examined the influ- ing with a male possessing large prey forence of female body size or mate avail- egg laying, and this relaxation of postmat-
TABLE 4. Mating durations (x ± SD minutes) in H. apicalis: Field observations on pairs mating with a small prey(<16 mm2 surface area) under low and high adult population density in 1980 and 1981.*
Low density High densityDatasummary Sample 1 Sample 2 Sample 1 Sample 2
1980Sample 10.6 ± 3.3 11.2 ±3 .8 6.1 ± 3.2 6.0 ± 3.1Density 10.9 ± 3.5 6.0 ± 3.1
1981Sample 11.4 ± 3 . 3 10.5 + 3.7 6.3 ± 3.0 6.4 ± 3.8Density 11.0 ± 3.5 6.3 ± 3.4
Each sample is two replicates (see Methods).
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

FEMALE CHOICE TACTICS IN A SCORPIONFLY 377
ing female choice will primarily be mani-fested by large females.
I suggest above that changes in malebehavior in relation to population size aredictated largely by female behavior: whenfemales become more choosy males haveto comply despite greater risks to malesassociated with obtaining large prey (i.e.,greater exposure to spiders). Previously Ihypothesized (Thornhill, 1976, 19806) thatnuptial feeding evolved in H. apicalis as ameans of reducing the amount of hunting,a risk-taking activity, that a female partic-ipates in during the time she is laying eggsfertilized by the male that fed her. Thisinterpretation of the selective advantage ofnuptial feeding for males assumes that thelast male successfully completing a longcopulation with a female (resulting in fullinsemination and a female period of non-receptivity and egg laying) fertilizes a highproportion of her eggs until she matesagain. Although this hypothesis is reason-able for the early evolutionary steps towardnuptial feeding in H. apicalis, it does noteasily fit the findings presented here onchanges in male behavior. Data on abun-dance of web-building spiders at five sitesin southeastern Michigan reveal that num-bers of spiders do not change significantlyduring the adult season of H. apicalis(Thornhill, unpublished), which suggeststhat hunting females would be equallyexposed to spider predation under bothlow and high population densities. Thusmales are not using large prey more fre-quently under high density to reduce thehunting their mates engage in; nor are largeprey more common in the habitat underhigh density. Therefore I am now inclinedto view the use of large prey by males asbehavior dictated by females.
When female demands become greaterit is in the interest of males to meet thosedemands, because this is the only way malescan achieve sexual access and insure thatfemales will use their sperm. Under mostcircumstances females control the repro-ductive process to a greater extent thanmales because they are gamete recipientsrather than donors (Alexander and Borgia,1979). This asymmetry in control of repro-duction sets the stage for male exploitation
by females and for changes in female choicecriteria that males must meet in order toreproduce. It seems that male H. apicalisunder high density are forced by femalesto obtain large prey despite great risks, andby providing a large prey a male is deliv-ering enough food to change the female'sbehavior from trading sexual access forfood to sexual nonresponsiveness and egglaying.
Selection on males to circumvent femalechoice may in some cases be stronger thanselection on females to choose (Parker,1979; Thornhill, 1980d, 1984). Unlikepanorpid scorpionflies, male bittacids can-not easily force copulation (Thornhill,1980d, 1981, 1983, 1984). More work isneeded to untangle the nature and extentof sexual conflict in//, apicalis. Indeed, thisand other scorpionfly species provideinteresting systems for analyzing dispari-ties in male and female reproductive inter-ests and their evolved outcomes (Thorn-hill, 1983, 1984).
Regardless of the precise proximate andevolutionary explanations for the behaviorof female H. apicalis I have outlined, theimportant point for the focus of this paperis the demonstration of intrapopulationvariation in female choice behavior and theindication that the variation has an adap-tive explanation. In the next section I out-line my study of between population vari-ation in female choice behavior in H.apicalis.
BETWEEN POPULATION VARIATION INFEMALE CHOICE IN HYLOBITTACUS APICALIS
Methods
During 12-30 July 1980 and 5-30 July1981 I conducted field studies of a rela-tively small H. apicalis adult population nearMt. Lake in Giles County, Virginia (site is1.3 mi west of intersection of state roads613 and 700). I first visited this site in 1977.My estimates of peak population sizes(average number of individuals per 3 x 3m habitat section, n = 12 sections per year)in 1977 and in the two years of field studyare: 1977 (x = 4, range 1-7), 1980 (x =3.5, range 2-8), 1981 (x = 3.3, range 1-9). This is considerably lower than peak
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022
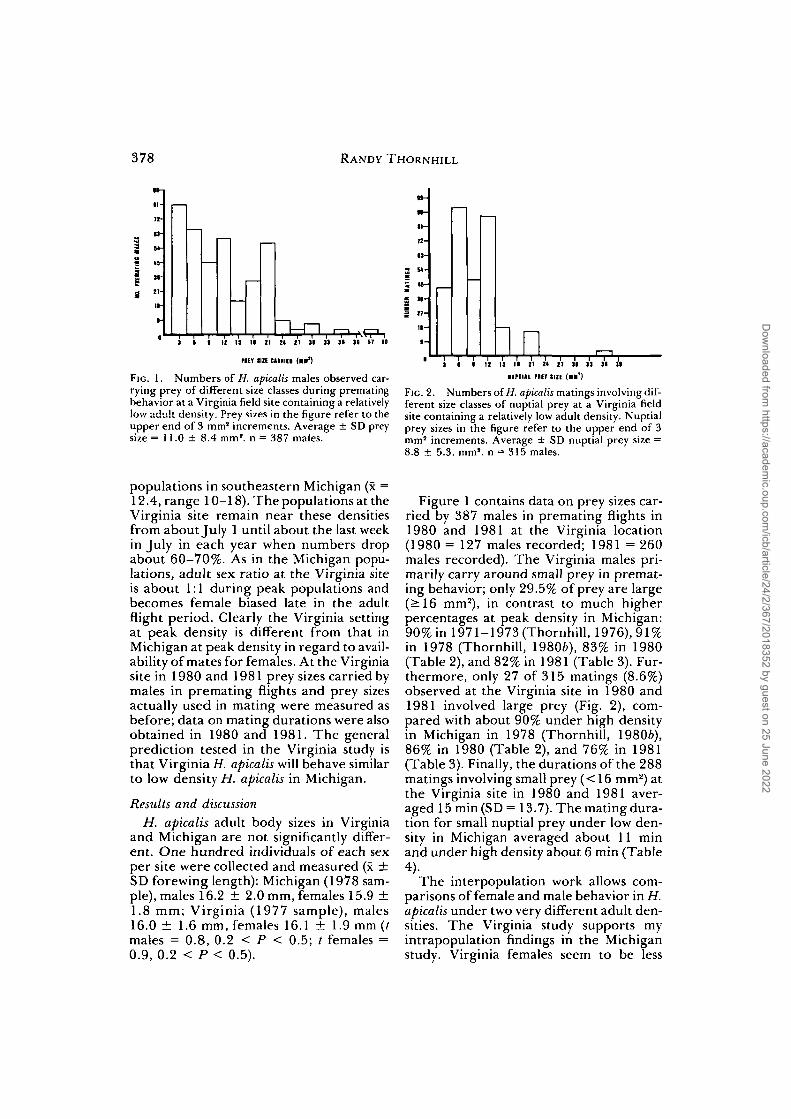
378 RANDY THORNHILL
II-
7 1 -
i :s uI I.s "•
ii-
^
1 I I 1J IS I I 21 !l 11 I I 1) 31 31 57 It
mi mi MMIEI (••')
FIG. 1. Numbers of H. apicalis males observed car-rying prey of different size classes during prematingbehavior at a Virginia field site containing a relativelylow adult density. Prey sizes in the figure refer to theupper end of 3 mm2 increments. Average ± SD preysize = 11.0 ± 8.4 mm2, n = 387 males.
ii-
i- in3 I • It 15 II Jl » >T II II 31 31
NPTIM MET till {»»')
FIG. 2. Numbers of H. apicalis matings involving dif-ferent size classes of nuptial prey at a Virginia fieldsite containing a relatively low adult density. Nuptialprey sizes in the figure refer to the upper end of 3mm2 increments. Average ± SD nuptial prey size =8.8 ± 5.3. mm2, n = 315 males.
populations in southeastern Michigan (x =12.4, range 10-18). The populations at theVirginia site remain near these densitiesfrom about July 1 until about the last weekin July in each year when numbers dropabout 60-70%. As in the Michigan popu-lations, adult sex ratio at the Virginia siteis about 1:1 during peak populations andbecomes female biased late in the adultflight period. Clearly the Virginia settingat peak density is different from that inMichigan at peak density in regard to avail-ability of mates for females. At the Virginiasite in 1980 and 1981 prey sizes carried bymales in premating flights and prey sizesactually used in mating were measured asbefore; data on mating durations were alsoobtained in 1980 and 1981. The generalprediction tested in the Virginia study isthat Virginia H. apicalis will behave similarto low density H. apicalis in Michigan.
Results and discussion
H. apicalis adult body sizes in Virginiaand Michigan are not significantly differ-ent. One hundred individuals of each sexper site were collected and measured (x ±SD forewing length): Michigan (1978 sam-ple), males 16.2 ± 2.0 mm, females 15.9 ±1.8 mm; Virginia (1977 sample), males16.0 ± 1.6 mm, females 16.1 ± 1.9 mm (tmales = 0.8, 0.2 < P < 0.5; t females =0.9, 0.2 < P < 0.5).
Figure 1 contains data on prey sizes car-ried by 387 males in premating flights in1980 and 1981 at the Virginia location(1980 = 127 males recorded; 1981 = 260males recorded). The Virginia males pri-marily carry around small prey in premat-ing behavior; only 29.5% of prey are large(^16 mm2), in contrast to much higherpercentages at peak density in Michigan:90% in 1971-1973 (Thornhill, 1976), 91%in 1978 (Thornhill, 19806), 83% in 1980(Table 2), and 82% in 1981 (Table 3). Fur-thermore, only 27 of 315 matings (8.6%)observed at the Virginia site in 1980 and1981 involved large prey (Fig. 2), com-pared with about 90% under high densityin Michigan in 1978 (Thornhill, 19806),86% in 1980 (Table 2), and 76% in 1981(Table 3). Finally, the durations of the 288matings involving small prey (< 16 mm2) atthe Virginia site in 1980 and 1981 aver-aged 15 min (SD =13.7). The mating dura-tion for small nuptial prey under low den-sity in Michigan averaged about 11 minand under high density about 6 min (Table4).
The interpopulation work allows com-parisons of female and male behavior in H.apicalis under two very different adult den-sities. The Virginia study supports myintrapopulation findings in the Michiganstudy. Virginia females seem to be less
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

FEMALE CHOICE TACTICS IN A SCORPIONFLY 379
selective of males in terms of both pre-mating choice (i.e., they more frequentlyaccept males with small prey as mates) andfemale choice during mating (i.e., they cop-ulate longer with males with small prey)than high density Michigan females, andVirginia females behave similarly to lowdensity Michigan females in terms of theirmate choice pattern. The relaxation offemale choice criteria by Virginia femalesmay influence male nuptial prey size selec-tion in the same fashion as under low den-sity in Michigan. The abundance of largenuptial prey may differ between Michiganand Virginia, but given the comparisons Ihave outlined, the most likely explanationfor the similarity in high frequency utili-zation of small nuptial prey by males inVirginia and in low density settings inMichigan is similar female choice stan-dards, especially since males (at least inMichigan) are capable of finding and will-ing to provide large nuptial prey when theyare rare but females demand them.
VARIATION IN FEMALE CHOICE IN SPECIESOTHER THAN HYLOB/TTACUS APICALIS
AND CONCLUSIONS
Female choice behavior is sometimesviewed by theorists and empiricists as a rel-atively inflexible species and/or popula-tional trait. This is not the case in H. api-calis. Species of bittacids other thanHylobittacus apicalis may also exhibit con-siderable within species flexibility in matechoice behavior (Thornhill, 1983). HereI briefly mention a few studies which sug-gest that mate choice behavior may beadaptively flexible in a variety of organ-isms.
An interesting example of possible adap-tive flexibility in female choice derives fromMcLain's (1981) study of the soldier beetleChauliognathus pennsylvanicus. The adultbeetles compete with wasps for food in theform of flowers of various species of weedyplants. The wasps attack beetles in orderto drive them from the flowers. They aremore likely to attack single beetles thanpairs, resulting in single females often beingforced from food and losing foraging time.In the presence and absence of wasps thesizes of single males are about the same,
but mating males are significantly smallerin the former circumstance. These resultssuggest that females relax mate choicestandards from a preference of large maleswhen such a modification is in their repro-ductive interests.
The early models of female behavior inresource-based mating systems of birds andtheir later extensions and refinements (seeIntroduction for references) require adap-tive flexibility in female choice behavior.Females are assumed to make decisionsabout pairing on the basis of payoff toreproduction. For example, they areexpected to choose to pair with an alreadymated male when there is a greater fitnessreturn from becoming a second mate of amale than from pairing with a matelessmale. In particular Orians' (1980) long termwork on decision making during matechoice by female redwing and yellow-headed blackbirds has revealed that femalesprobably value territory quality more thancharacteristics of the territory owner andthat females select territories such thatfledging success among females is nothighly variable over a range of ecologicalconditions. This suggests considerableadaptive flexibility in female choice in thesebirds. It seems that the female blackbirdsemploy ecological information in decisionmaking that reflects average ecologicalconditions.
Recent work on the mottled sculpin fish(Coitus bairdi) reveals flexibility in femalechoice behavior (Brown, 1981; Down-hower and Brown, 1981). Male sculpinsguard egg-deposition sites and tend the fryafter the eggs hatch. Females prefer largemales and large males are better parents—i.e., there is a positive relationship betweenmale body size and hatching success. Butthe timing of breeding is as important tofemale reproductive success as the size ofa female's mate, and females strive to breedearly in the season. Thus females trade offtime in searching for a very large male forearly breeding. The patterns of femalesculpin behavior found by Brown andDownhower indicate considerable adap-tive decision making by females in associ-ation with this trade off. Brown and Down-hower argue that timing of female
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

380 RANDY THORNHILL
reproduction is likely to be an importantparameter influencing compromises ofchoosing females in many vertebrates.
Male katydids (Orthoptera: Tettigoni-idae) produce a spermatophore that isattached to the female's genitalia duringmating. The spermatophore is quite largein some species, making up as much as 30%of male body weight. The female eats thespermatophore, and evidence indicates thatspermatophore nutrients are important tofemale fecundity (Morris, 1980; Gwynne,1982). Male choice is expected when malesbecome a limiting resource for females(Trivers, 1972; Gwynne, 1981, 1983;Thornhill and Alcock, 1983). Gwynne'sstudy of mate choice in the Mormon cricket(actually a katydid, Anabrus simplex) pro-vides one of the most striking examples ofadaptive shifts in mate choice behavior byindividuals (Gwynne, 1981, 19846). Underhigh density Mormon crickets form rov-ing bands somewhat similar to those inmigratory locusts. In this context food islimited, and the large nutritious spermato-phore of the male (up to 27% of male bodyweight) becomes very valuable to females,which results in a sex role reversal in com-petitive and choice behavior. A male whois ready to mate ascends a plant and beginsto sing. Females in the vicinity quicklyrespond and approach the calling male.Females often interact aggressively bygrappling as they converge upon a singingmale. The winner of direct female-femalecompetition is not guaranteed the male'slarge spermatophore because of the male'sdiscrimination against about two-thirds offemales that fight their way to his singingperch.
But in low density populations the sexrole reversal is not evident. In this casemales are evenly spaced due to aggressionand fights between neighboring males arealso seen. There is no evidence of materejection by males and males eagerlydescended from their calling perches tomeet responsive females.
Gwynne (19846) feels that the role re-versal in behavior may only be expressed inMormon crickets at high density becauseonly under this condition do males become
a limiting resource for females. In the highdensity populations individuals competevigorously for food. Optimum food plants,seeds, and dead insects are contested byindividuals in the band and are eatenquickly. Females in low density populationsseem to have a ready supply of food (sweet-clover seeds). Males in low density popu-lations, having access to more food, areprobably able to accrue more quickly theresources necessary to produce spermato-phores. Nutritious spermatophores, there-fore, may be more critical to female repro-duction at high densities. Gwynne'sinterpretation of the interpopulation dif-ferences in behavior is supported by hisobservations on one high density popula-tion in which the roving band encountereda section of habitat containing an abundantsupply of sweet clover. As the band movedinto this area and began to feed a tempo-rary sex role reversal occurred: malesshowed correlates of competitive behaviorand females became noncompetitive formales and less eager to mate.
How female choice on the basis of fea-tures that confer only genetic benefits tothe female is maintained is not understoodfor any organism exhibiting it (MaynardSmith, 1978; Taylor and Williams, 1981,1983; Thornhill, 1980a, 1983) but it isclear that in some such organisms femalesenhance their direct fitness by mate choice(Drosophila: greater competitive ability oflarvae, Partridge, 1980; Campsis: increasedseed number and weight, Bertin, 1982;Scaphiopus toads: greater offspring sur-vival, Woodward, manuscript), whichargues strongly for the existence of suffi-cient genetic variation to maintain adap-tive female choice. It is clear that the extentof female choice may vary from very littleto very extreme within a season under someconditions in Hylobittacus apicalis. Also Ber-tin's (1982) work on Campsis suggests withingeneration changes in the extent of femalechoice. Female choice itself could be animportant means for maintaining geneticvariation relevant to the maintenance ofadaptive choice.
Adoption of alternative tactics of femalechoice in Hylobittacus is condition depen-
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

FEMALE CHOICE TACTICS IN A SCORPIONFLY 381
dent. There is suggestion that condition-dependent switches from one tactic toanother influence female choice decisionsin the organisms other than Hylobittacusdiscussed in this section. Female choicebefore and during mating in H. apicalisdepends on conditions of body size, mateavailability and feeding history of thefemale. It is likely that female choicebehavior in H. apicalis represents a con-ditional strategy (for discussion of types ofbehavioral polymorphisms see West-Eber-hard, 1979, 1981; Cade, 1980; Dawkins,1980; Thornhill, 1981). Selection hasfavored a flexible female choice phenotypein Hylobittacus. An adult female keepsoptions for greater reproductive successopen by adoption of condition-dependentalternatives. For example, when food ormates are scarce females focus on materialbenefits (food and reduction of risk-takingassociated with hunting) rather than geneticbenefits provided by males. The focus onmaterial benefits coincides with a switch toa lower standard in terms of the male afemale mates with and is inseminated by.But when food or mates are abundant,females switch to a preference of males whocan deliver the best genetic benefits.
ACKNOWLEDGMENTS
Steve Austad, Luther Brown, Gary Dod-son, Larry Marshall and Bruce Woodwardprovided helpful criticisms on the manu-script. The study was supported by grants(BNS-7912208, DEB-7910193) from theNational Science Foundation. MichelleGrenier's salary was provided by the PrattFellowship Fund of the Department ofBiology, University of Virginia. MikeBrown, Michelle Grenier and NancyThornhill helped with aspects of theresearch. Drs. Francis Evans and RobertStorer provided facilities at the Edwin S.George Reserve. Drs. Jim Riopel and JimMurray, Jr. provided facilities and supportat Mountain Lake Biological Station.
This paper is dedicated to Dr. GeorgeW. Byers in the year of his 60th birthday.He has generously provided me with infor-mation about taxonomy, geographical dis-tributions and general biology of Mecop-
tera during the 11 yr in which I have studiedthese insects.
REFERENCES
Alcock, J. 1979a. Selective mate choice by femalesof Harpobittacus australis (Mecoptera: Bittacidae).Psyche 86:213-217.
Alcock, J. 1979ft. The evolution of intraspecificdiversity in male reproductive strategies in somebees and wasps. In M. S. Blum and N. A. Blum(eds.), Sexual selection and reproductive competitionin insects, pp. 381-402. Academic Press, NewYork.
Alexander, R. D. and G. Borgia. 1979. On the originand basis of the male-female phenomenon. In M.S. Blum and N. A. Blum (eds.), Sexual selectionand reproductive competition in insects, pp. 417-440.Academic Press, New York.
Bertin, R. I. 1982. Paternity and fruit production intrumpet creeper (Campsis radicans). Am. Nat. 119:694-709.
Borgia, G. 1979. Sexual selection and the evolutionof mating systems. In M. S. Blum and N. A. Blum(eds.), Sexual selection and reproductive competitionin insects, pp. 19-80. Academic Press, New York.
Bornemissza, G. E. 1966. Observations on the hunt-ing and mating behavior of two species of scor-pion flies (Bittacidae: Mecoptera). Aust. J. Zool.14:371-382.
Bradbury.J. W. and S. L. Vehrencamp. 1977. Socialorganization and foraging in emballonurid bats.III. Mating systems. Behav. Ecol. Sociobiol. 2:1-17.
Brockmann, H. J., A. Grafen, and R. Dawkins. 1979.Evolutionary stable nesting strategy in a diggerwasp. J. Theoret. Biol. 77:473-496.
Brown, L. 1981. Patterns of female choice in mottledsculpins (Cottidae, Teleostei). Anim. Behav. 29:375-382.
Burley, N. 1981. Mate choice by multiple criteria ina monogamous species. Am. Nat. 1 17:515-527.
Burley, N. and N. Moran. 1979. The significance ofage and reproductive experience in the matepreferences of feral pigeons, Columba livia. Anim.Behav. 27:686-698.
Byers, G. W. and R. Thornhill. 1983. The biologyof Mecoptera. Annual Rev. Ent. 28:203-228.
Cade, W. 1980. Alternative male reproductivebehaviors. Fla. Ent. 63:30-44.
Dawkins, R. 1980. Good strategy or evolutionarilystable strategy? In G. W. Barlow andj. Silverberg(eds.), Soaobiology: Beyond nature/nurture?, pp.331-370. Westview Press, Boulder, Colorado.
Downhower, J. F. 1976. Darwin's finches and theevolution of sexual dimorphism in body size.Nature 263:558-563.
Downhower, J. F. and L. Brown. 1981. The timingof reproduction and its behavioral consequencesfor mottled sculpins, Cottus bairdi. In R. D. Alex-ander and D. W. Tinkle (eds.), Natural selectionand social behavior: Recent research and new theory,pp. 78—95. Chiron Press, Portland, Oregon.
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

382 RANDY THORNHILL
Eberhard, W. G. 1982. Beetle horn dimorphism:Making the best of a bad lot. Am. Nat. 119:420-426.
Emlen, S. T. and L. W. Oring. 1977. Ecology, sexualselection, and the evolution of mating systems.Science 197:215-223.
Falconer, D. 1960. Introduction to quantitative genetics.Ronald Publ. Co., New York.
Fisher, R. A. 1930. The genetical theory of natural selec-tion. Clarendon Press, Oxford (2nd ed., 1958;Dover, New York).
Gross, M. R. 1982. Sneakers, satellites and parentals:Polymorphic mating strategies in North Ameri-can sunfishes. Z. Tierpsychol. 60:1-26.
Gwynne, D. T. 1981. Sexual difference theory: Mor-mon crickets show role reversal in mate choice.Science 213:779-780.
Gwynne, D. T. 1982. Mate selection by female katy-dids (Orthoptera: Tettigoniidae, Conocephalusnigropleurum). Anim. Behav. 30:734-738.
Gwynne, D. T. 1983. Male nutritional investmentand the evolution of sexual differences in theOrthoptera. In D. T. Gwynne and G. K. Morris(eds.), Orthopteran mating systems: Sexual competitionin a diverse group of insects, pp. 337-366. WestviewPress, Boulder, Colorado.
Gwynne, D. T. 1984a. Male mating effort, confi-dence of paternity and insect sperm competition.In R. L. Smith (ed.), Sperm competition and theevolution of animal mating systems. Academic Press,New York. (In press)
Gwynne, D. T. 1984fc. Male nutrient investment,population density and sexual selection in Mor-mon crickets (Anabrus simplex, Orthoptera: Tetti-goniidae). Evolution. (In press)
Janetos, A. C. 1980. Strategies of female mate choice:A theoretical analysis. Behav. Ecol. Sociobiol. 7:107-112.
Lambert, D. M., P. D. Kingett, and E. Slooten. 1982.Intersexual selection: The problem and a discus-sion of the evidence. Evol. Theory 6:67-78.
Lloyd, J. E. 1979. Mating behavior and natural selec-tion. Fla. Ent. 62:17-34.
Maynard Smith, J. 1978. The evolution of sex. Cam-bridge Univ. Press, Cambridge.
McLain, D. K. 1981. Interspecific interference com-petition and mate choice in the soldier beetleChauliognathus pennsylvanicus. Behav. Ecol.Sociobiol. 9:65-66.
Morris, G. K. 1980. Calling display and matingbehaviour of Copiphora rhinoceros Pictet (Orthop-tera: Tettigoniidae). Anim. Behav. 28:42-51.
Noonan, K. M. 1981. Individual strategies of inclu-sive-fitness-maximizing in Polistesfuscatus' found-resses. In R. D. Alexander and D. W. Tinkle (eds.),Natural selection and social behavior: Recent researchand new theory,^. 18-44. Chiron Press, Portland,Oregon.
Orians, G. H. 1969. On the evolution of mating sys-tems in birds and mammals. Am. Nat. 103:589—603.
Orians, G. H. 1980. Some adaptations of mar\h-netfmgblackbirds. Princeton University Press, Princeton,New Jersey.
Parker, G. A. 1970. Sperm competition and its evo-lutionary consequences in the insects. Biol. Rev.45:525-567.
Parker, G. A. 1979. Sexual selection and sexual con-flict. In M. S. Blum and N. A. Blum (eds.), Sexualselection and reproductive competition in insects, pp.123-166. Academic Press, New York.
Partridge, L. 1980. Mate choice increases a com-ponent of offspring fitness in fruit flies. Nature283:290-291.
Rubenstein, D. I. 1980. On the evolution of alter-native mating strategies. In J. I. E. R. Staddon(ed.), Limits to action: The allocation of individualbehavior, pp. 65-100. Academic Press, New York.
Searcy, W. A. 1979. Female choice of mates: A gen-eral model for birds and its application to red-winged blackbirds {Agelaius phoeniceus). Am. Nat.114:77-100.
Sivinski,J. 1980. Sexual selection and insect sperm.Fla. Ent. 63:99-111.
Strassmann, J. E. 1981. Kin selection and satellitenests in Polistes exclamans. In R. D. Alexander andD. W. Tinkle (eds.), Natural selection and socialbehavior: Recent research and new theory, pp. 45—60. Chiron Press, Portland, Oregon.
Taylor, P. D. and G. C. Williams. 1981. On themodelling of sexual selection. Q. Rev. Biol. 56:305-313.
Thornhill, R. 1974. Evolutionary ecology of Mecop-tera (Insecta). Ph.D. Diss., University of Michi-gan.
Thornhill, R. 1976. Sexual selection and nuptialfeeding behavior in Bittacus apicahs (Insecta:Mecoptera). Am. Nat. 110:529-548.
Thornhill, R. 1977. The comparative predatory andsexual behavior of hangingflies (Mecoptera: Bit-tacidae). Misc. Pub. Mus. Zool., University ofMichigan, Ann Arbor, No. 69, pp. 1-49.
Thornhill, R. 1979a. Male and female sexual selec-tion and the evolution of mating strategies ininsects. In M. S. Blum and N. A. Blum (eds.),Sexual selection and reproductive competition in insects,pp. 81-121. Academic Press, New York.
Thornhill, R. 19796. Adaptive female-mimickingbehavior in a scorpionfly. Science 205:412-414.
Thornhill, R. 1980a. Competitive, charming malesand choosy females: Was Darwin correct? Fla.Ent. 63:5-30.
Thornhill, R. 19806. Mate choice in Hylobittacus api-calis (Insecta: Mecoptera) and its relation to somemodels of female choice. Evolution 34:519-538.
Thornhill, R. 1980c Sexual selection in the black-tipped hangingfly. Sci. Am. 242:162-172.
Thornhill, R. 1980d. Rape in Panorpa scorpionfliesand a general rape hypothesis. Anim. Behav. 28:52-59.
Thornhill, R. 1981. Panorpa scorpionflies: Systemsfor understanding resource-defense polygyny andthe evolution of alternative male reproductiveefforts. Ann. Rev. Ecol. Syst. 12:355-386.
Thornhill, R. 1983. Cryptic female choice in thescorpionH) Harpnhtttacus nignreps and its impli-cations. Am. Nat. 122:765-788.
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

FEMALE CHOICE TACTICS IN A SCORPIONFLY 383
Thornhill, R. 19846. Alternative hypotheses for traitspresumed to have evolved in the context of spermcompetition. In R. L. Smith (ed.), Sperm competi-tion and the evolution of animal mating systems. Aca-demic Press, New York. (In press)
Thornhill, R. and J. Alcock. 1983. The evolution ofinsect mating systems. Harvard University Press,Cambridge, Massachusetts.
Trivers, R. L. 1972. Parental investment and sexualselection. In B. Campbell (ed.), Sexual selection andthe descent of man, 1871-1971, pp. 136-179.Aldine, Chicago.
Verner, J. and M. F. Willson. 1966. The influencesof habitats on mating systems of North Americanpasserine birds. Ecology 47:143-147.
Walker, W. F. 1980. Sperm utilization strategies innonsocial insects. Am. Nat. 115:780-799.
West-Eberhard, M. J. 1979. Sexual selection, socialcompetition, and evolution. Proc. Phil. Soc. Amer.123:222-234.
West-Eberhard, M.J. 1981. Intragroup selection andthe evolution of insect societies. In R. D. Alex-ander and D. W. Tinkle (eds.), Natural selectionand social behavior: Recent research and new theory,pp. 3-17. Chiron Press, Portland, Oregon.
Williams, G. C. 1975. Sex and evolution PrincetonUniv. Press, Princeton, New Jersey
Wittenberger.J. F. 1981. Animal social behavior. Dux-bury Press, Boston.
Woodward, B. Manuscript. Paternal effects on off-spring survival in Scaphiopus couchi (Couch'sspadefoot).
Zahavi, A. 1975. Mate selection—A selection for ahandicap. J. Theor. Biol. 53:205-214.
Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022

Dow
nloaded from https://academ
ic.oup.com/icb/article/24/2/367/2018352 by guest on 25 June 2022




















