
Altered DNA binding specificity of Arnt by selection of partner bHLH-PAS proteins
11
Altered DNA binding specificity of Arnt by selection of partner bHLH–PAS proteins Koshi Kinoshita, Yasuo Kikuchi, Yukie Sasakura, Masashi Suzuki 1 , Yoshiaki Fujii-Kuriyama and Kazuhiro Sogawa* Department of Biomolecular Sciences, Graduate School of Life Sciences, Tohoku University, Sendai, 980–8578, Japan and 1 AIST-NIBHT CREST Centre of Structural Biology, Higashi 1–1, Tsukuba 305–0046, Japan Received February 26, 2004; Revised May 3, 2004; Accepted May 14, 2004 ABSTRACT The Ah receptor (AhR) and HLF are transcription factors involved in xenobiotic metabolism and hypoxic response, respectively. AhR and HLF heterodimerize with Arnt as the common partner, and bind to asymmetric E-boxes termed XRE and HRE, respectively. In order to investigate nucleotide preference of the heterodimers, reporter plasmids with oligonucleotides for XREs or HREs with systematic mutations were constructed and their activity was determined. Comparison of the activity revealed that DNA length and nucleotide preference recognized by Arnt subunit in the two heterodimers were largely different between XRE and HRE. We expressed AhR–Arnt and HLF–Arnt in Escherichia coli and used them for DNA binding. The dissociation constant of HLF–Arnt–HRE was 10.4 6 1.6 nM. Competition activity of mutated XREs or HREs with wild type was consistent with their tran- scription activity. Bending of XRE and HRE induced by binding of the relevant heterodimers was observed with stronger bending of XRE than of HRE. By deletional and mutational analyses, an alanine and three arginine (Ala 8, Arg 9, Arg 11 and Arg 12) residues in the basic sequence of HLF were found to be indispensable for the transcriptional activity. INTRODUCTION The basic helix–loop–helix motif (bHLH) is a frequently used domain structure in transcription factors. This motif mediates dimerization that results in the formation of both homodimers and heterodimers binding to the E-box site CANNTG to regulate transcription (1,2). The bHLH motif consists of the basic sequence followed by two amphipathic a-helices separated by a loop of variable length. The N-terminal basic sequence is rich in basic amino acids and directly contacts bases in the major groove of DNA. It is believed that conformation of the basic region is altered from a random coil to an a-helical structure when it binds to DNA (3,4). Members of the bHLH–PAS (Per-Arnt-Sim) family consti- tute a subgroup of the HLH protein family of transcription factors and play key roles in several physiological phenomena including xenobiotic metabolism [Ah receptor (AhR)](5), hypoxic responses such as angiogenesis and erythropoiesis (HIF-1a and HLF) (6,7), maintenance of circadian rhythms (Clock) (8) and neurogenesis (Sim 1 and Sim 2) (9). These transcription factors, except for Clock, form heterodimers with the AhR nuclear translocator (Arnt), which is also a bHLH–PAS domain protein and acts as the common partner protein. Both HLH and PAS domains function to mediate protein–protein interaction for dimerization and the N-terminal basic sequence of each subunit contacts a half- site of the asymmetric E-box (10). AhR is localized in the cytosol in association with Hsp90 in quiescent cells. When xenobiotics such as 2,3,7,8-tetrachloro- dibenzo-p-dioxin (TCDD) and 3-methylcholanthrene (MC) enter the cells, AhR binds to them as ligands and translocates into the nucleus. In the nucleus, AhR dimerizes with Arnt and the AhR–Arnt complex binds to the xenobiotic responsive element (XRE), which is localized in the upstream region of genes for mainly MC-inducible drug metabolizing enzymes such as CYP1A1, glutathione S-transferase Ya subunit and a form of UDP-glucronosyl transferase (11). HIF-1a and HLF (HIF-2a) are closely related transcription factors that are activated by the reduction of oxygen (12). At normoxia, these two proteins are rapidly degraded through a ubiquitin–proteasome pathway. Under hypoxic conditions, the HIF-1a and HLF stabilize, form heterodimers with Arnt and upregulate genes, such as those for erythropoietin (EPO), vascular endothelial growth factor (VEGF) and glycolytic enzymes, by binding to the hypoxia response element (HRE) in the promoter region. Since HRE has a core sequence different from that of XRE, the two heterodimers, AhR–Arnt and HIF-1a–Arnt or HLF–Arnt, exhibit no cross-binding. Sim 1 and Sim 2 function negatively on transcription (13), while the Drosophila counterpart dSim is a positive transcrip- tion factor. dSim is a master regulatory gene that controls the development of the neurons and glia localized along the midline of the central nervous system (CNS). Drosophila Arnt, a tango gene product (Tgo) that is closely related to *To whom correspondence should be addressed. Tel: +81 22 217 6590; Fax +81 22 217 6594; Email: [email protected] Present address: Y. Fujii-Kuriyama, Center for Tsukuba Advanced Reserch Alliance, University Tsukuba, Tsukuba, Ibaraki 305–8577, Japan Nucleic Acids Research, 2004, Vol. 32, No. 10 3169–3179 DOI: 10.1093/nar/gkh637 Nucleic Acids Research, Vol. 32 No. 10 ª Oxford University Press 2004; all rights reserved Published online June 9, 2004 by guest on November 30, 2013 http://nar.oxfordjournals.org/ Downloaded from
Transcript of Altered DNA binding specificity of Arnt by selection of partner bHLH-PAS proteins

Altered DNA binding speci®city of Arnt by selectionof partner bHLH±PAS proteinsKoshi Kinoshita, Yasuo Kikuchi, Yukie Sasakura, Masashi Suzuki1,
Yoshiaki Fujii-Kuriyama and Kazuhiro Sogawa*
Department of Biomolecular Sciences, Graduate School of Life Sciences, Tohoku University, Sendai, 980±8578,Japan and 1AIST-NIBHT CREST Centre of Structural Biology, Higashi 1±1, Tsukuba 305±0046, Japan
Received February 26, 2004; Revised May 3, 2004; Accepted May 14, 2004
ABSTRACT
The Ah receptor (AhR) and HLF are transcriptionfactors involved in xenobiotic metabolism andhypoxic response, respectively. AhR and HLFheterodimerize with Arnt as the common partner,and bind to asymmetric E-boxes termed XRE andHRE, respectively. In order to investigate nucleotidepreference of the heterodimers, reporter plasmidswith oligonucleotides for XREs or HREs withsystematic mutations were constructed and theiractivity was determined. Comparison of the activityrevealed that DNA length and nucleotide preferencerecognized by Arnt subunit in the two heterodimerswere largely different between XRE and HRE. Weexpressed AhR±Arnt and HLF±Arnt in Escherichiacoli and used them for DNA binding. Thedissociation constant of HLF±Arnt±HRE was 10.4 61.6 nM. Competition activity of mutated XREs orHREs with wild type was consistent with their tran-scription activity. Bending of XRE and HRE inducedby binding of the relevant heterodimers wasobserved with stronger bending of XRE than ofHRE. By deletional and mutational analyses, analanine and three arginine (Ala 8, Arg 9, Arg 11 andArg 12) residues in the basic sequence of HLF werefound to be indispensable for the transcriptionalactivity.
INTRODUCTION
The basic helix±loop±helix motif (bHLH) is a frequently useddomain structure in transcription factors. This motif mediatesdimerization that results in the formation of both homodimersand heterodimers binding to the E-box site CANNTG toregulate transcription (1,2). The bHLH motif consists of thebasic sequence followed by two amphipathic a-helicesseparated by a loop of variable length. The N-terminal basicsequence is rich in basic amino acids and directly contactsbases in the major groove of DNA. It is believed that
conformation of the basic region is altered from a random coilto an a-helical structure when it binds to DNA (3,4).
Members of the bHLH±PAS (Per-Arnt-Sim) family consti-tute a subgroup of the HLH protein family of transcriptionfactors and play key roles in several physiological phenomenaincluding xenobiotic metabolism [Ah receptor (AhR)](5),hypoxic responses such as angiogenesis and erythropoiesis(HIF-1a and HLF) (6,7), maintenance of circadian rhythms(Clock) (8) and neurogenesis (Sim 1 and Sim 2) (9). Thesetranscription factors, except for Clock, form heterodimerswith the AhR nuclear translocator (Arnt), which is also abHLH±PAS domain protein and acts as the common partnerprotein. Both HLH and PAS domains function to mediateprotein±protein interaction for dimerization and theN-terminal basic sequence of each subunit contacts a half-site of the asymmetric E-box (10).
AhR is localized in the cytosol in association with Hsp90 inquiescent cells. When xenobiotics such as 2,3,7,8-tetrachloro-dibenzo-p-dioxin (TCDD) and 3-methylcholanthrene (MC)enter the cells, AhR binds to them as ligands and translocatesinto the nucleus. In the nucleus, AhR dimerizes with Arnt andthe AhR±Arnt complex binds to the xenobiotic responsiveelement (XRE), which is localized in the upstream region ofgenes for mainly MC-inducible drug metabolizing enzymessuch as CYP1A1, glutathione S-transferase Ya subunit and aform of UDP-glucronosyl transferase (11).
HIF-1a and HLF (HIF-2a) are closely related transcriptionfactors that are activated by the reduction of oxygen (12). Atnormoxia, these two proteins are rapidly degraded through aubiquitin±proteasome pathway. Under hypoxic conditions, theHIF-1a and HLF stabilize, form heterodimers with Arnt andupregulate genes, such as those for erythropoietin (EPO),vascular endothelial growth factor (VEGF) and glycolyticenzymes, by binding to the hypoxia response element (HRE)in the promoter region. Since HRE has a core sequencedifferent from that of XRE, the two heterodimers, AhR±Arntand HIF-1a±Arnt or HLF±Arnt, exhibit no cross-binding.
Sim 1 and Sim 2 function negatively on transcription (13),while the Drosophila counterpart dSim is a positive transcrip-tion factor. dSim is a master regulatory gene that controls thedevelopment of the neurons and glia localized along themidline of the central nervous system (CNS). DrosophilaArnt, a tango gene product (Tgo) that is closely related to
*To whom correspondence should be addressed. Tel: +81 22 217 6590; Fax +81 22 217 6594; Email: [email protected] address:Y. Fujii-Kuriyama, Center for Tsukuba Advanced Reserch Alliance, University Tsukuba, Tsukuba, Ibaraki 305±8577, Japan
Nucleic Acids Research, 2004, Vol. 32, No. 10 3169±3179DOI: 10.1093/nar/gkh637
Nucleic Acids Research, Vol. 32 No. 10 ã Oxford University Press 2004; all rights reserved
Published online June 9, 2004 by guest on N
ovember 30, 2013
http://nar.oxfordjournals.org/D
ownloaded from

mammalian Arnt, dimerizes with dSim to bind the CNSmidline element (CME) and activates CNS midline transcrip-tion (14). Sim 1 and Sim 2 also function in the development ofCNS. Because CME closely resembles HRE in sequence,HIF-1a±Arnt or HLF±Arnt heterodimer could bind to CME.However, the target genes of the dSim±Arnt heterodimer arecompletely different from those of the HIF-1a±Arnt orHLF±Arnt heterodimer. It is reported that the speci®city oftarget gene selection was performed through the PAS domainsof dSim and HIF-1a (15). To date, genes located downstreamof mammalian Sims are unknown.
Since the core sequences of XRE (CACGC) and HRE(CACGTA) contain CAC half-sites in common, the CAC half-site was expected to be recognized by Arnt. This expectationwas con®rmed by DNA±protein cross-linking experiments(16) and the ®nding that Arnt was able to form a homodimer tobind to a symmetric type 2 E-box, CACGTG (17).
For binding of bHLH±PAS proteins to cognate responseelements, ¯anking sequence is equally important to the coresequence of the response elements. However, the nucleotidepreference in the ¯anking sequence has not been studied so farand is largely unknown. In this work, we have investigatedDNA recognition of the AhR±Arnt and HLF±Arnt hetero-dimers to the ¯anking sequence of XRE and HRE coresequences, and elucidated the preference for base pairs bymonitoring transcription activity of XREs and HREs withmutations in the ¯anking sequence. Nucleotide preference andlength recognized by Arnt in the ¯anking of XRE core elementwere strikingly different from those of HRE. DNA bindinganalyses of bacterially expressed AhR±Arnt and HLF±Arnt toXRE and HRE supported the ®nding. Furthermore, inaccordance with the results, bending of XRE induced bybinding of AhR±Arnt was clearly shown in the circularpermutation assay.
MATERIALS AND METHODS
Construction of plasmids
Reporter plasmids containing mutated HRE and XRE wereconstructed by inserting the synthetic oligonucleotides(88mer) which consist of four-tandem repeats of a variety ofXRE (18mer; TGAGTTCTCACGCTAGCA) and HRE(18mer; 5¢-TGAGACAGCACGTAGGGC-3¢) sequencesspaced with four nucleotides of GATC and four nucleotidesfor ligation (CGCG for the coding strand and GATC for thenon-coding strand) into the MluI±BglII site of pGL3 PromoterVector (Promega).
The bacterial expression plasmid pRSETB-hAhR-hArnt,which encodes amino acids 27±436 of AhR and 82±464 ofArnt, was constructed as described previously (18). pRSETB-mHLF-hArnt was constructed similarly. A DNA fragment ofmouse HLF cDNA encoding amino acids 15±360 inclusive ofbHLH±PAS regions was ampli®ed by PCR and cloned into theNheI±HindIII of pRSETB vector (Invitrogen). The resultantplasmid, pRSETB-mHLF, was digested with HaeII, and theblunt-ended fragment containing cDNA was cloned into theA¯III site of pRSETB-hArnt (amino acids 82±464) plasmid(18) to generate HLF pRSETB-mHLF-hArnt. pRSETB-mHLF(D1±2)-hArnt and pRSETB-mHLF(K1A)-hArnt wereconstructed by inserting oligonucleotides for mutated basic
sequences into the NdeI±BsmBI site of pRSETB-mHLF.Construction of an expression plasmid of Flag-tagged HLF,pBOSFlagmHLF, was performed by replacing DNA (from theEcoRI to the BsmBI site) coding for the N-terminal basicsequence of HLF in pBOSmHLF (12) with a syntheticoligonucleotide for Flag-tag and the basic sequence of HLF. Inthe oligonucleotide, an NdeI site was introduced just after theC-terminus to the Flag-tag to facilitate construction of HLFmutants. The Flag-tagged N-terminal basic sequence of HLFis as follows: MDYKDDDDKHRKEKSRDAARCRR±.Effector plasmids for Flag-tagged HLF mutant plasmidswere constructed by inserting oligonucleotides for mutatedbasic sequences into the NdeI±BsmBI site ofpBOSFlagmHLF.
pBend-HRE and pBend-XRE used in the circular permu-tation assay were constructed by inserting oligonucleotides ofXRE 5¢-CTAGATTGATGCTAGCGTGAGAACTCAAATG-3¢, 5¢-TCGACATTTGAGTTCTCACGCTAGCATCAAT-3¢and of HRE 5¢-CTAGATTGAGCCCTACGTGCTGTCT-CAAATG-3¢, 5¢-TCGACATTTGAGACAGCACGTAGGG-CTCAAT-3¢ between the XbaI and SalI sites of pBend2(generously provided by Dr S. Adhya). All plasmids con-structed were con®rmed by sequencing.
Cell culture and reporter assay
Human hepatoma Hep3B cells were maintained in Dulbecco'smodi®ed Eagle's medium supplemented with 10% fetalbovine serum and 40 mg/ml kanamycin. The cells weretransfected using the calcium phosphate method as describedpreviously (19). Cells in a 60 mm dish were transientlytransfected with 5 mg of DNA (2 mg of reporter plasmidtogether with 3 mg pBOS-LacZ, or 2 mg each of effector andreporter plasmids together with 1 mg pBOS-LacZ). Four hoursafter transfection, the cells received fresh medium. For assaysof reporter activity using plasmids containing HRE and XRE,cells were incubated for 18 h in normoxia and for 40 h in thepresence or absence of 1 mM MC, respectively, beforeharvesting. The luciferase activity was determined using aluciferase detection kit (Promega) and normalized to b-galactosidase activity, which was used as an internal control.
Expression of bHLH±PAS proteins in Escherichia coli
pRSETB-hAhR-hArnt, pRSETB-mHLF-hArnt, pRSETB-mHLF(D1±2)-hArnt and pRSETB-mHLF(K1A)-hArnt wereexpressed as described previously (18). In brief, the E.colistrain BL21(DE3), pLysS containing pRSETB-hAhR-hArnt,pRSETB-mHLF-hArnt, pRSETB-mHLF(D1±2)-hArnt orpRSETB-mHLF(K1A)-hArnt was grown in 10 ml of L-Broth (50 mg/ml ampicillin, 25 mg/ml chloramphenicol) at37°C until OD600 > 1.0. Con®rming suf®cient multiplication,they were grown in 100 ml L-Broth at 20°C, and isopropyl-b-D-thiogalactopyranoside (IPTG) (0.1 mM) was added atOD600 = 0.5 to induce protein expression. After overnightcultivation at 20°C, the bacteria were recovered by centrifuga-tion and lysed by sonication in extract buffer (20 mM HEPESpH 7.9, 500 mM NaCl, 5 mM imidazole, 10% glycerol, 0.1%NP-40, 1mM b-mercaptoethanol). Then the sample wascentrifuged at 18 000 r.p.m. for 30 min and the supernatantwas added to 0.1 ml Ni-NTA gel (Qiagen). After gentlerotating at 4°C for 1 h, the sample was centrifuged at1000 r.p.m. for 1 min and the precipitating gel was washed
3170 Nucleic Acids Research, 2004, Vol. 32, No. 10
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from
®ve times with extract buffer. Co-expressed proteins wereeluted with elution buffer (20 mM HEPES pH 7.9, 500 mMNaCl, 200 mM imidazole, 10% glycerol, 0.1% NP40, 1 mM b-mercaptoethanol). The puri®ed hAhR-hArnt, mHLF-hArnt,mHLF(D1±2)-hArnt and mHLF(K1A)-hArnt proteins wereestimated to amount to 30±40% of total protein by SDS±PAGE and subsequent densitometric determination (data notshown).
Gel mobility shift assay (GMSA)
Bacterially expressed HLF±Arnt and AhR±Arnt heterodimerswere used for GMSA. The proteins were preincubated at 4°Cfor 30 min in a total volume of 10 ml of binding buffer (10 mMHEPES pH 7.9, 100 mM NaCl, 0.1 mM EDTA, 3 mM MgCl21 mM dithiothreitol, 0.1 mg/ml bovine serum albumin,0.01mg/ml salmon sperm DNA, 10% glycerol), followed byincubation with 2 ml of 32P-labeled oligonuculeotide probe(10 000 c.p.m.) at 20°C for 25 min. The reaction mixture was
electrophoresed in a non-denaturing 4.5% polyacrylamide geland the gel was dried and autoradiographed.
DNase I footprinting
A DNA fragment containing a single copy of the XRE or HREsequence was prepared by cleaving pBend-XRE or pBend-HRE with XhoI and Asp718. The fragment was labeled at the5¢ end of the coding or non-coding strand by treating it with[32P]ATP and T4 polynucleotide kinase and then received asecond digestion by BglII and BamHI, respectively. Thelabeled fragment was puri®ed on agarose gels and incubatedwith puri®ed AhR±Arnt or HLF±Arnt in a total volume of40 ml of binding buffer (10 mM HEPES pH 7.9, 100 mMNaCl, 0.1 mM EDTA, 3 mM MgCl2, 1 mM dithiothreitol,0.1 mg/ml bovine serum albumin, 10% glycerol, 4 mg/mlpoly[dI-dC]) at 4°C for 20 min. Next, 2 ml of a DNase Isolution (5 mM CaCl2, 10 mM MgCl2, 17.5 U/ml DNase I)were added to the mixture and incubated for 5 s. Reactions
Nucleic Acids Research, 2004, Vol. 32, No. 10 3171
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from
were terminated by the addition of 40 ml of phenol saturatedwith 3% NaCl. Digested DNA was fractionated through an8% polyacrylamide±8 M urea sequencing gel (acrylamide:bisacrylamide = 19:1). Chemical cleavage of another portionof the labeled DNA at purine residues (A + G) was performedaccording to the procedure described by maxam and Gilbert(20), and the resultant DNA was run in parallel with thesamples of digested DNA. After electrophoresis, gels weredried and autoradiographed.
Circular permutation assay
A circular permutation assay was performed as describedpreviously (21). The plasmids pBend-HRE and pBend-XREwere each separately digested with ®ve restriction enzymeslisted in the legend to Figure 5, and DNA fragments werepuri®ed by agarose gel electrophoresis and labeled with 32P-ATP. Using these DNA fragments and purifed proteins
expressed in E.coli, gel mobility shift assay was conductedas described above.
RESULTS
Mutational analysis of HRE and XRE
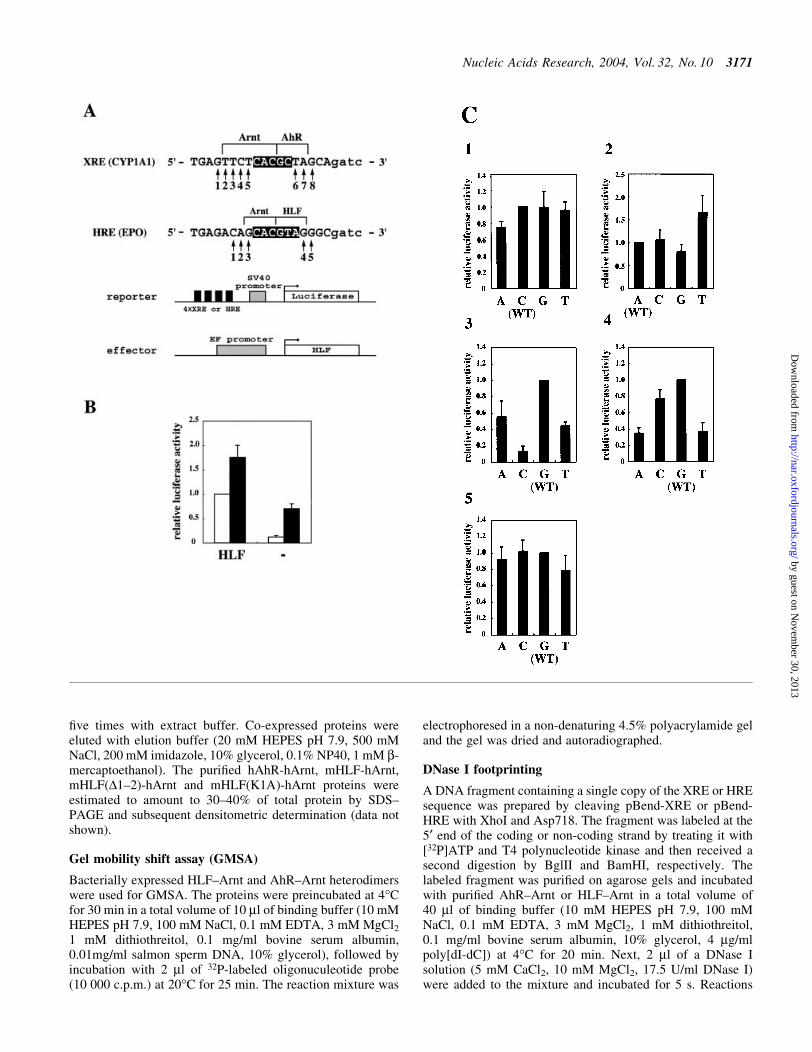
In order to estimate the nucleotide preference that AhR±Arntand HLF±Arnt heterodimers exhibit in the ¯anking region ofXRE and HRE, respectively, systematic mutations wereintroduced to the response elements in the promoter ofreporter plasmids as shown in Figure 1A. As parental responseelements, XRE1 in the rat CYP1A1 gene and the HREsequence in the human erythropoietin gene that are known toact as potent enhancers were selected and used as wild types.Each nucleotide in the ¯anking sequence of wild-type XREand HRE core sequence changed to three other nucleotides(positions 1±8 for XRE and 1±5 for HRE). These reporter
Figure 1. Mutational analysis of HRE and XRE. (A) Sequence of wild-type XRE and HRE and structure of reporter and effector plasmids. A 22mer ofoligonucleotides for XRE from XRE 1 of the CYP1A1 gene and HRE originating from the EPO gene is shown. Mutated nucleotides are shown by 1±8 forXRE and 1±5 for HRE. Regions contacted with Arnt and AhR or Arnt and HLF are shown by bars above the sequences. Reporter plasmids contain fourtandemly repeated XREs or HREs upstream of the SV40 promoter and luciferase structural gene. The effector plasmid contains elongation factor enhancerand promoter to induce expression of HLF in Hep3B cells. (B) Effect of coexpression of HLF on reporter activity. The effector plasmid (2 mg) shown in(A) was cotransfected with 2 mg of HRE reporter plasmid in (A) and 1 mg of pBOSlacZ, and expressed luciferase activity was determined. Open and ®lledboxes show luciferase activity from cells incubated in normoxic and hypoxic conditions, respectively. The activity was shown relative to the activity fromcells transfected with HLF effector plasmid and incubated in normoxia. The luciferase activity was normalized with b-galactosidase activity used as aninternal control. The activity is the average of at least three independent experiments with SD. (C) Transcriptional activity of plasmids with mutated HRE.The position of the mutated nucleotide was shown above each diagram. A, C, G and T below diagrams denote nucleotides changed in HRE. The originalnucleotide used in wild-type HRE is shown as (WT). Luciferase activity was determined using cells incubated in normoxic conditions, and is shown relativeto the activity of wild type. The luciferase activity was normalized with b-galactosidase activity used as an internal control. The activity is the average of atleast three independent experiments with SD. (D) Transcriptional activity of plasmids with mutated XRE. The position of the mutated nucleotide is shownabove each diagram. A, C, G and T below diagrams denote nucleotides changed in XRE. The original nucleotide used in wild-type HRE is shown as (WT).Luciferase activity was determined from cells incubated in the presence or absence of MC for 40 h after transfection and is shown relative to the activity ofthe wild type induced by MC. The luciferase activity was normalized with b-galactosidase activity used as an internal control. The activity is the average ofat least three independent experiments with SD.
3172 Nucleic Acids Research, 2004, Vol. 32, No. 10
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from
plasmids were introduced into Hep3B cells which expressendogenous AhR, HLF and HIF-1a and show a strongresponse to xenobiotic and hypoxic stimuli. In the case ofreporter assays of plasmids containing HRE sequences, wild-type HLF was coexpressed to minimize the effect ofendogenous HIF-1a. As shown in Figure 1B, the expressed
luciferase activity in normoxic cells was increased about eighttimes by the coexpression of HLF. The activity was furtherincreased 2.5-fold in hypoxic conditions. As the effect ofexogeneous HLF emerged more strongly in the normoxiccondition than the hypoxic condition, we chose the normoxiccondition to determine reporter activity of mutated HREsdriven by HLF. We ®rst introduced mutations at position 3 ofwild-type HRE (Fig. 1C). When compared with wild type, thethree mutants exhibited decreased luciferase activity. Inparticular, mutation to C largely decreased the activity ofwild type to 13.1%, suggesting the presence of an interactionbetween the base pair at this position and the HLF±Arntheterodimer. Mutations at position 2 were then introduced andluciferase activity was assayed. Mutations of A in wild type toC, G and T showed activity similar to wild type, suggestingthat the heterodimer did not recognize the base pair at position2. Since a weak augmentation in activity was found whenmutated to T at position 2, we further introduced similarmutations at position 1. Nearly the same amounts of activity ofmutants as for the wild type was found, suggesting nointeraction between DNA and the heterodimer at this position.The mutation at position 4 from G of wild type to A and Tdecreased its activity to 35% and 37% of wild type,respectively, suggesting interaction of the heterodimer withHRE at this position. Mutations at position 5 to othernucleotides did not affect the activity, suggesting no inter-action between HRE and the dimer at position 5. Takentogether, these results suggest that the HLF±Arnt heterodimercontacted the eight base pairs 5¢-GCACGTAG-3¢ in HRE. Forrecognition of HRE by HLF±Arnt, four base pairs wererecognized by each subunit as shown in Figure 1A.
We performed similar experiments to investigate recogni-tion of the AhR±Arnt heterodimer to XRE, using the luciferasereporter plasmids containing mutated XRE sequences(Fig. 1D). Strict preference for base-pair recognition byAhR±Arnt was observed in the ¯anking region. Selection of Tand C at positions 4 and 5 in XRE greatly decreased theactivity. At position 3, changes to A or G led to a largedecrease in activity. Furthermore, mutation to A or C atposition 2 also led to a slight decrease in activity. These resultsstrongly suggest that recognition of Arnt to XRE extends toposition 2. Mutations at position 6 did not affect activity, andmutations to C, G or T at position 7 led to a large reduction inactivity. In particular, mutation to G diminished activitynearly to the background level. Taken together, the AhR±Arntheterodimer recognized 10 base pairs of XRE (positions 2±7,excluding position 6), and at positions 1 and 8 the heterodimerdid not show any base-pair preference.
In vitro DNA binding of HLF±Arnt and AhR±Arnt
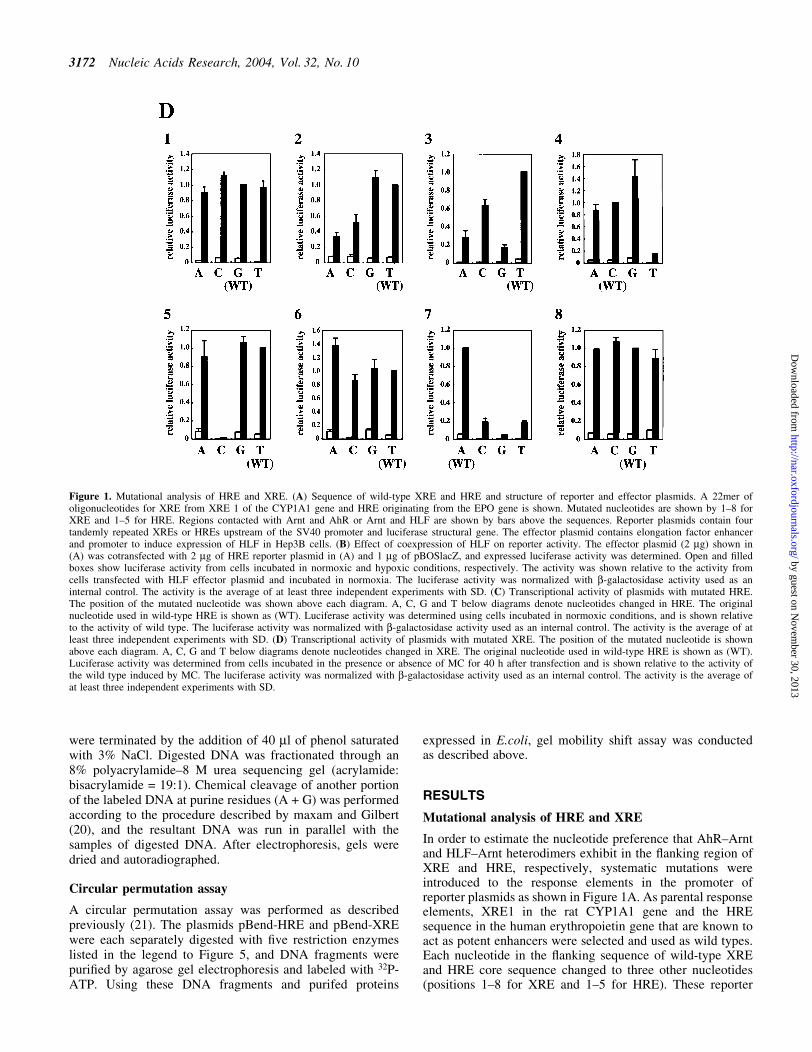
Before doing experiments on DNA binding of heterodimers,the dissociation constant of the bacterially expressed HLF±Arnt heterodimer±HRE complex was determined. As shownin Figure 2A, the Kd value was 10.4 6 1.6 nM. This value wasslightly lower than that of the AhR±Arnt to XRE (18). Thisresult strongly suggests that the recombinant HLF±Arntheterodimer has a DNA binding property quite similar tothat of cellular origin, although to date no Kd values using acellular HLF±Arnt heterodimer have been reported.Methylation of the CpG sequence in the oligonucleotides ofHRE greatly reduced the ability to compete with unmethylated
Figure 2. Binding of the bacterially expressed HLF±Arnt heterodimer toHRE. (A) The dissociation constant was determined using the HLF±Arntexpressed in E.coli and synthetic oligonucleotides of HRE in GMSA. Arepresentative diagram of a Scatchard plot and a typical autoradiographicresult of GMSA are shown on the left and right, respectively. The HREconcentrations in lanes 1±6 were 25, 78.8, 14, 10.5, 7.9 and 5.9 nM,respectively. Protein concentration in the reaction mixture was 35 ng/ml.The ®lled and open arrowheads show the bound and free probes,respectively. The bands indicated by an arrow were not reproducible.(B) Effect of CpG methylation in the HRE sequence on binding. Theposition of 5-methyl cytosine in HRE is marked with arrows. Allcombinations of oligonucleotides with methylated and unmethylated strandsof HRE were used as competitors in GMSA where unmethylated HRE wasused as the probe. Numbers with an asterisk above autoradiography showmethylated strands of HRE. Increasing amounts of competitors of 12.5-fold(lanes 2, 5, 8 and 11), 25-fold (lanes 3, 6, 9 and 12) and 50-fold (lanes 4, 7,10 and 13) over probe were added to the pre-incubation mixture.
Nucleic Acids Research, 2004, Vol. 32, No. 10 3173
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from

HRE (Fig. 2B). A similar decrease in the binding activity ofthe AhR±Arnt heterodimer to XRE has already been reported(18).
Recombinant AhR±Arnt heterodimer was incubated with32P-labeled probe for wild-type XRE, and increasing amountsof competitors were added (Fig. 3). Similar experiments wereperformed using a probe for HRE and bacterially expressedHLF and Arnt. We chose three different types of competitorsin af®nity with mutations in the region mainly contacted byArnt: in the ®rst type the transcriptional activity of thesequence was similar to or higher than wild type (m1, m6), inthe second it was lower (m2, m4, m5, m7 and m8) and in thethird it was much lower (m3). We could not ®nd mutantcompetitors with extreme low activity for HRE. We used m18(22) as a negative control in competition with XRE.Competition with m1 and m6 showed more intense compe-tition than that with wild type (Fig. 3A and B, lanes 5±7). Thecompetitive activity of m2, m4, m5, m7 and m8 was lowerthan wild type and correlated roughly with their transcriptionalactivity. No or very weak competitive activity, similar to thatof m18, was observed with m3 which showed essentially notranscriptional activity. These results demonstrate that thebinding activity of AhR±Arnt and HLF±Arnt heterodimer tothe mutated XRE and HRE, respectively, is highly correlatedwith the transcriptional activity shown in Figure 1B and C.
DNase I footprinting analysis of XRE and HRE
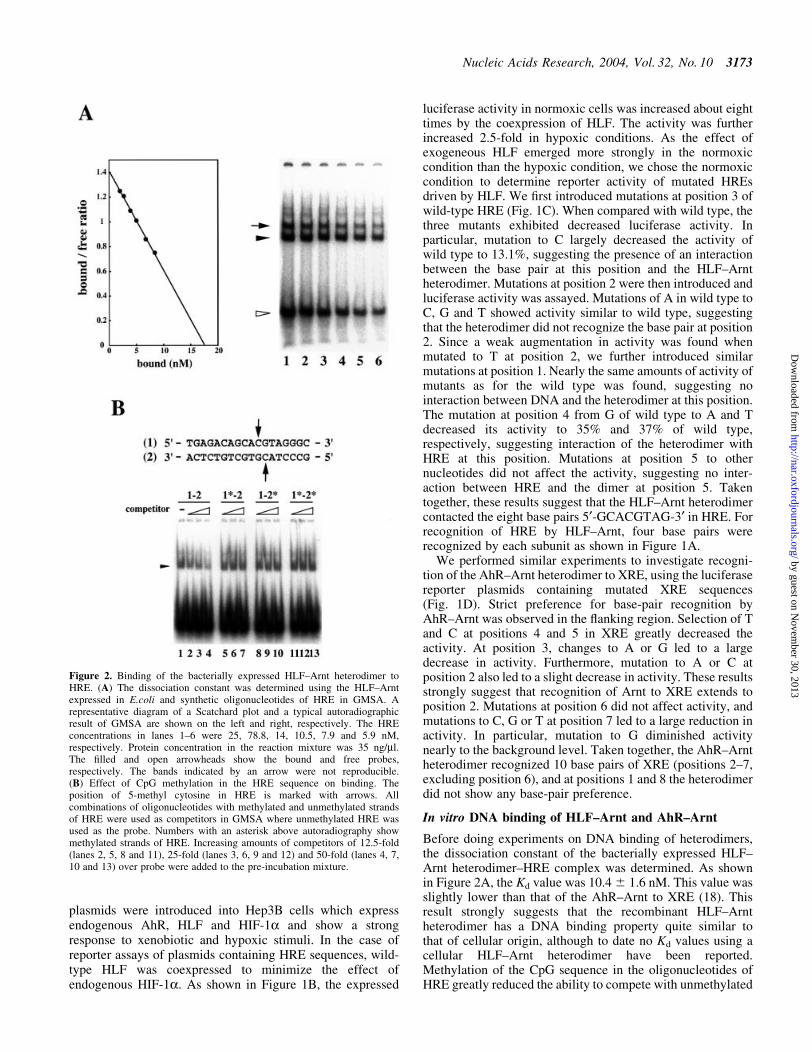
To further investigate the region recognized by HLF±Arnt orAhR±Arnt, DNase I footprinting was performed using the
puri®ed proteins expressed in E.coli and DNA fragments,including a single copy of HRE or XRE. When the protectedregion of HRE and XRE was compared, we found that thecoding strand of XRE, especially the 3¢-¯anking region ofXRE, was more protected than that of HRE from DNase Idigestion as shown in Figure 4. Hypersensitive sites werefound 5¢ upstream of XRE and HRE in the coding strand and 3¢downstream of XRE and HRE in the non-coding strand. Weaksignals for hypersensitive sites were also observed 3¢ down-stream of XRE and HRE. Weak protection of bases was foundin the 5¢ region in the coding sequence of XRE and HRE. Thisprotection was probably due to non-speci®c binding ofprotein.
Bending of HRE and XRE
As the recognized sequence of XRE by the AhR±Arntheterodimer was unexpectedly long, as shown in Figure 1C,the DNA fragments containing XRE were expected to bendwhen interacting with the heterodimer. To investigate theDNA bending of XRE and HRE, we conducted a circularpermutation assay. The structures of the probes, includingXRE and HRE, are shown in Figure 5A. Five types of probesin which the binding site of the protein was moved from oneend to the other were prepared for each response element.As shown in Figure 5B, the two heterodimers bent thecorresponding DNA and the migration pattern of the AhR±Arnt±XRE complex was more progressive than that of HLF±Arnt±HRE. This result shows that XRE is more strongly bentthan HRE when interacting with protein. The DNA fragments
Figure 3. Competition of mutated XRE or HRE with wild type. (A) Competitive GMSA was performed using representative mutated XRE as competitorsand bacterially expressed AhR±Arnt heterodimer. Puri®ed protein (40 ng) was mixed with buffer A containing mutated oligonucleotides on ice for 30 min.About 133 pg of 32P-labeled (10 000 c.p.m.) XRE (wild type) was added to the mixture which was incubated at 20°C for 25 min. The mutated XRE sequencewas shown, and only mutated nucleotides in mutants were represented. The transcriptional activity obtained in Figure 1 is also shown. A representativeautoradiograph of GMSA is shown. Increasing amounts of competitors [30-fold (lanes 2, 5, 8 and 11), 100-fold (lanes3, 6, 9 and 12) and 250-fold (lanes 4, 7,10 and 13)] were used for competition. The arrow shows the retarded band. (B) Competitive GMSA was performed using representative mutated HRE ascompetitors. The experiment was performed using the same procedure as in (A) using bacterial expressed HLF±Arnt heterodimer and mutated HRE.Increasing amounts of competitors [100-fold (lanes 2, 5, 8 and 11), 250-fold (lanes 3, 6, 9 and 12) and 500-fold (lanes 4, 7, 10 and 13)] were used forcompetition assay. The arrow shows the retarded band.
3174 Nucleic Acids Research, 2004, Vol. 32, No. 10
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from
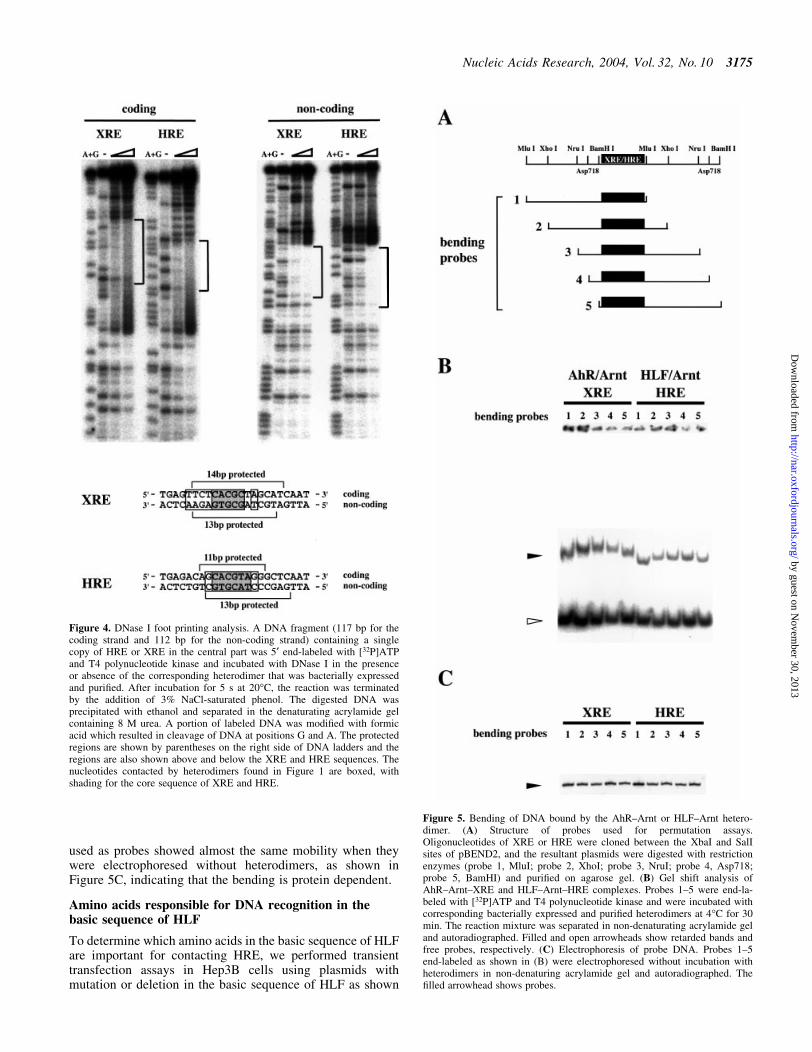
used as probes showed almost the same mobility when theywere electrophoresed without heterodimers, as shown inFigure 5C, indicating that the bending is protein dependent.
Amino acids responsible for DNA recognition in thebasic sequence of HLF
To determine which amino acids in the basic sequence of HLFare important for contacting HRE, we performed transienttransfection assays in Hep3B cells using plasmids withmutation or deletion in the basic sequence of HLF as shown
Figure 4. DNase I foot printing analysis. A DNA fragment (117 bp for thecoding strand and 112 bp for the non-coding strand) containing a singlecopy of HRE or XRE in the central part was 5¢ end-labeled with [32P]ATPand T4 polynucleotide kinase and incubated with DNase I in the presenceor absence of the corresponding heterodimer that was bacterially expressedand puri®ed. After incubation for 5 s at 20°C, the reaction was terminatedby the addition of 3% NaCl-saturated phenol. The digested DNA wasprecipitated with ethanol and separated in the denaturating acrylamide gelcontaining 8 M urea. A portion of labeled DNA was modi®ed with formicacid which resulted in cleavage of DNA at positions G and A. The protectedregions are shown by parentheses on the right side of DNA ladders and theregions are also shown above and below the XRE and HRE sequences. Thenucleotides contacted by heterodimers found in Figure 1 are boxed, withshading for the core sequence of XRE and HRE.
Figure 5. Bending of DNA bound by the AhR±Arnt or HLF±Arnt hetero-dimer. (A) Structure of probes used for permutation assays.Oligonucleotides of XRE or HRE were cloned between the XbaI and SalIsites of pBEND2, and the resultant plasmids were digested with restrictionenzymes (probe 1, MluI; probe 2, XhoI; probe 3, NruI; probe 4, Asp718;probe 5, BamHI) and puri®ed on agarose gel. (B) Gel shift analysis ofAhR±Arnt±XRE and HLF±Arnt±HRE complexes. Probes 1±5 were end-la-beled with [32P]ATP and T4 polynucleotide kinase and were incubated withcorresponding bacterially expressed and puri®ed heterodimers at 4°C for 30min. The reaction mixture was separated in non-denaturating acrylamide geland autoradiographed. Filled and open arrowheads show retarded bands andfree probes, respectively. (C) Electrophoresis of probe DNA. Probes 1±5end-labeled as shown in (B) were electrophoresed without incubation withheterodimers in non-denaturing acrylamide gel and autoradiographed. The®lled arrowhead shows probes.
Nucleic Acids Research, 2004, Vol. 32, No. 10 3175
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from
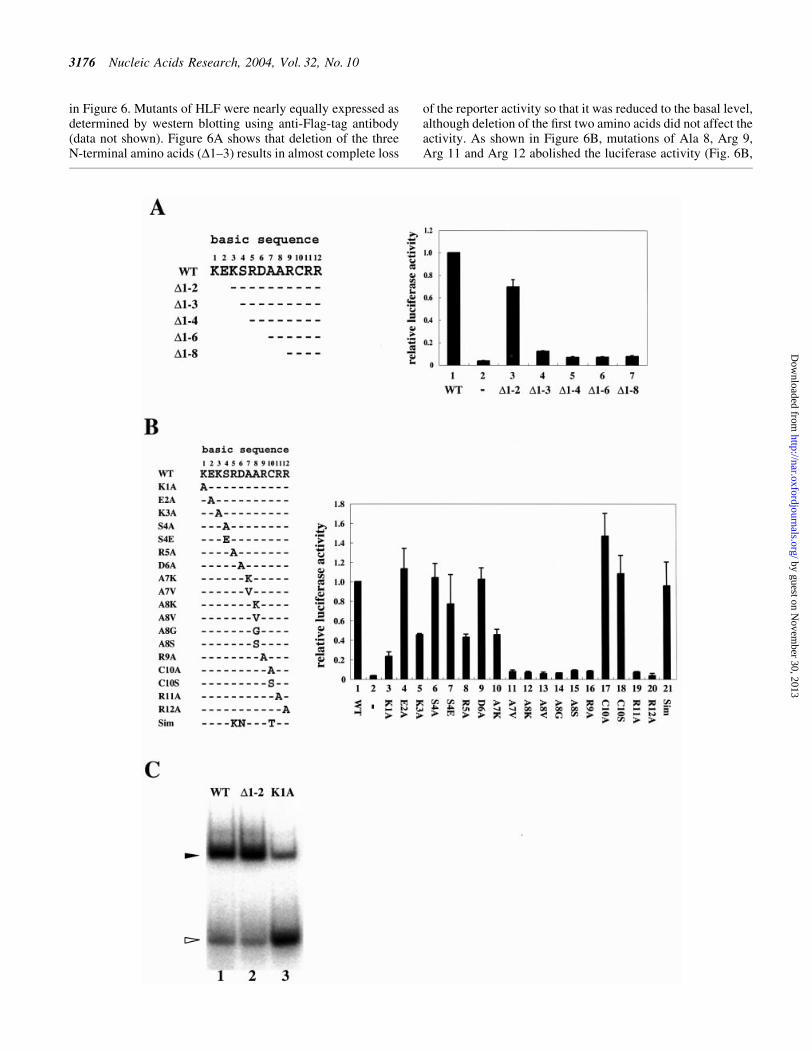
in Figure 6. Mutants of HLF were nearly equally expressed asdetermined by western blotting using anti-Flag-tag antibody(data not shown). Figure 6A shows that deletion of the threeN-terminal amino acids (D1±3) results in almost complete loss
of the reporter activity so that it was reduced to the basal level,although deletion of the ®rst two amino acids did not affect theactivity. As shown in Figure 6B, mutations of Ala 8, Arg 9,Arg 11 and Arg 12 abolished the luciferase activity (Fig. 6B,
3176 Nucleic Acids Research, 2004, Vol. 32, No. 10
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from

lanes 12±17 and 20), suggesting that these amino acids areresponsible for DNA binding. Although mutation of Ala 7 toVal led to a large decrease in activity, mutation to Lys showedsigni®cant luciferase activity (Figure 6B, lanes 10 and 11).When basic amino acids Lys 3 and Arg 5 were mutated to Ala,the luciferase activity decreased by about 55%, suggesting aconsiderable role in the interaction of these amino acids withDNA. Mutation of Lys 1 to Ala caused a considerablereduction in luciferase activity, although Lys 1 was shown tobe dispensable for the activity by the analysis of a deletionmutant (D1±2). To solve the discrepancy, we expressed twomutants of HLF, D1±2 and K1A, with Arnt in E.coli and usedthe expressed heterodimers to examine the DNA bindingactivity to HRE. As shown in Figure 6C, the DNA bindingactivity of the mutants was roughly associated with thetranscription activity. From the result, we concluded that the®rst two amino acids in the basic sequence of HLF were notinvolved in direct DNA binding, and that mutation of Lys 1 toAla indirectly damaged the DNA binding.
The sequence of CME is known to be closely related to, ornearly the same as, that of HRE (15), and three amino acids inthe basic sequence are different between HLF and Sim 1(Fig. 6B). We mutated the three amino acids in HLF to that ofSim 1, and the expressed luciferase activity was determined.As shown in Figure 6B, the mutated HLF with basic sequenceof Sim 1 showed activity similar to wild-type HLF (Fig. 6B,lane 21). This result suggests that the amino acid substitutionbetween HLF and Sim 1 did not affect DNA binding activity,and that the two factors have quite similar DNA bindingspeci®city.
DISCUSSION
bHLH±PAS proteins constitute a subgroup in the bHLHsuperfamily and bind to the asymmetric E-box. Of the bHLH±PAS proteins, AhR and HLF have key roles in xenobioticmetabolism and hypoxic response, respectively, by activatingtranscription. AhR and HLF form heterodimers with Arnt andbind to DNA elements termed XRE and HRE, respectively.These elements have the core sequence CACGC (XRE) orCACGTA (HRE). Although the occurrence of the coresequence in the DNA is necessary for speci®c binding, it isnot enough to assure ef®cient binding of the factors, but its¯anking sequence must be considered. However, a detailedanalysis for ¯anking sequences has not been performed. Wesystematically mutated XRE and HRE in their ¯anking region(Fig. 1), and found that the AhR±Arnt and HLF±Arnt
heterodimers recognize 10 bp and 8 bp DNA, respectively.Sequence speci®city of Arnt subunits was clearly differentbetween AhR±Arnt and HLF±Arnt.
Flanking sequences of the XRE core have been analyzedaccording to the method of DNA selection and ampli®cation(23). The DNA sequences selected by several rounds ofselection were to some extent compatible with those shown inFigure 1D. However, a closer comparison showed cleardifferences probably due to overly repeated selection andampli®cation and/or a bias rising from the strategy used. In theliterature, certain base pairs were not used for XRE even in thesites distant from the core sequence. Extended contact of Arntin the AhR±Arnt to the DNA has been reported (24) with alimited number of mutated XRE sequences, consistent withthe results of this study. A survey of known enhancersequences containing XRE or HRE found in various genesfor drug metabolism and hypoxia satis®ed the preferablesequences elucidated in this study (25±27).
DNA recognition of the AhR±Arnt heterodimer wasunexpectedly long, and it exceeded one helical turn of DNA.This long DNA recognition by AhR±Arnt was mainlyperformed by the Arnt subunit. Presumably, Arnt in theheterodimer with AhR contacts the DNA beyond the majorgroove by bending DNA. Rather strong DNA bending by theAhR±Arnt was observed (Fig. 5). It is not known why thedifference in DNA recognition of Arnt between XRE and HREarose. A subtle conformational change or positioning of thebasic region caused by partner proteins could generate thedifference. In the bHLH-zip protein family, MAX has asimilar function to Arnt. MAX can heterodimerize to c-Mycand MAD, and also form a homodimer. However, unlike Arnt,the recognition sequence of MAX seems identical in hetero-dimers or homodimers (4), and no DNA bending was reported.
The three-dimensional structures of the DNA±proteincomplexes of several bHLH-domain-containing transcriptionfactors, including MAX, USF, MyoD and PHO4 homodimers,have been elucidated (28±31). The DNA recognition helix ofthe basic region was inserted into the corresponding majorgroove of DNA. The liganded DNA was essentially B formand was not bent by the binding of proteins. The discovery ofthe bending of XRE and HRE, as shown in this study, was anextraordinary ®nding and may be characteristic of an asym-metric E-box. The X-ray crystallographic studies demon-strated that conserved amino acids His, Glu and Arg atpositions 4, 8 and 12, respectively, directly interacted in basicsequences with bases in the major groove (31). These aminoacids were not conserved in the basic sequence of HLF except
Figure 6. Deletional and mutational analyses of the basic sequence of HLF. (A) Deletional analysis of the HLF basic sequence. The amino acid sequence(1±12) of the basic region of HLF and the deleted amino acids are shown on the left. Effector plasmids (2 mg) with corresponding deletions werecotransfected into Hep3B cells together with 2 mg of the HRE reporter plasmid shown in Figure 1A and 1 mg of pBOSlacZ using the same procedure asshown in Figure 1. The cells were incubated in normoxic conditions for 18 h after transfection. The luciferase activity was normalized with b-galactosidaseactivity used as an internal control. The activity is the average of at least three independent experiments with SD. Effector plasmids with or without full-length HLF cDNA were used as positive and negative controls, respectively, and the activity is shown in lanes 1 and 2. (B) Mutational analysis of the HLFbasic sequence. Mutated amino acids are shown on the left. The amino acids of basic sequence were replaced with Ala. The alanine residues in Ala 7 and Ala8 in the wild type were changed to K or V for Ala 7 and K, V, G or S for Ala 8. The structure of the basic sequence of Sim 1 is also shown. The transfectionprocedures and culture conditions of cells were the same as that in (A). Effector plasmids with or without full-length HLF cDNA were used as positive andnegative controls, respectively, and the activity is shown in lanes 1 and 2. The luciferase activity was normalized with b-galactosidase activity used as aninternal control. The activity is the average of at least three independent experiments with SD. (C) Binding activity of HLF mutants to HRE. Mutants of HLF,D1±2 and K1A were generated with Arnt in E.coli and puri®ed. The DNA binding activity of the mutants was examined in GMSA according to the sameprocedure as shown in Figure 3B. Filled and open arrowheads show retarded bands and free probes, respectively.
Nucleic Acids Research, 2004, Vol. 32, No. 10 3177
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from

for Arg 12. Mutational analysis of the amino acids in positions4, 8 and 12 of HLF clari®ed that mutation of Ser 4 to Ala wasnot effective in the transcriptional activity. Instead, mutationor deletion of the surrounding basic amino acids at positions 3and 5 reduced the transcriptional activity. Mutation of Ala 8 toLys, Val, Gly and Ser showed extensive reduction intranscription activity, suggesting the necessary interaction ofAla 8 with DNA albeit with the charge changed from negativeto neutral when compared with Glu 8 in bHLH-zip proteins. Ithas been shown that mutations of Ser 4 in the HIF-1a basicregion showed no effect on acticity (32). Although the reportdid not show the importance of basic amino acids surroundingSer 4, the contact between the amino acids and the DNA mightbe conserved between HLF and HIF-1a.
To date, no data have been available on dissociationconstant of HLF±Arnt±HRE. Thus HLF and Arnt were co-expressed in E.coli, puri®ed and used for determination of thedissociation constant of the HLF±Arnt±HRE complex. Thedissociation constant of HLF±Arnt±HRE was similar to that ofAhR±Arnt±XRE (18). Essentially no different dissociationconstants were exhibited between cellular and recombinantAhR±Arnt heterodimers [see (18) and references cited there-in]. CpG methylation of each strand of HRE largely reducedthe af®nity, suggesting a regulation mechanism of transcrip-tion via methylation of DNA. Reduction of expression ofCYP1A1 presumably through CpG methylation of XRE wasreported in senescent model animals (33).
In expectation of the occurrence of XRE or HRE inthe promoter of newly identi®ed target genes, sequenceinformation carried by the core sequences was, to some extent,useful for a rough estimation of presence of the responseelements, but the information was not enough for accurateidenti®cation and estimation of strength of the responseelements. The base-pair preference in the ¯anking sequence ofXRE and HRE elucidated in this study, would help to identifythe response elements in the genes, although estimated XREand HRE consensus sequences are not complete becauseinteractions between neighboring amino acids in the basicsequence were not considered in the experiments.
ACKNOWLEDGEMENTS
We thank Dr O. Gotoh and Dr K. Yasumoto for helpfuldiscussions and Dr S. Adhya for the kind gift of pBEND2vector. This work was supported in part by Grant-in-Aid forresearch on priority area (A) from the Ministry of Education,Science, Sports and Culture of Japan, and research for theFuture Program of the Japan Society for the Promotion ofScience.
REFERENCES
1. Baxevanis,A.D. and Vinson,C.R. (1993) Interactions of coiled coils intranscription factors: where is the speci®city? Curr. Opin. Genet. Dev., 3,278±285.
2. Blackwell,T.K. and Weintraub,H. (1990) Differences and similarities inDNA-binding preferences of MyoD and E2A protein complexes revealedby binding site selection. Science, 250, 1104±1110.
3. Murre,C., McCaw,P.S. and Baltimore,D. (1989) A new DNA binding anddimerization motif in immunoglobulin enhancer binding, daughterless,MyoD and myc proteins. Cell, 56, 777±783.
4. Ferre-D'Amare,A.R., Prendergast,G.C., Ziff,E.B. and Burley,S.K. (1993)Recognition by Max of its cognate DNA through a dimeric b/HLH/Zdomain. Nature, 363, 38±45.
5. Whitlock,J.P.,Jr (1999) Induction of cytochrome P4501A1. Annu. Rev.Pharmacol., 39, 103±125.
6. Hochachka,P.W., Buck,L.T., Doll,C.J. and Land,S.C. (1996) Unifyingtheory of hypoxia tolerance: molecular/metabolic defense and rescuemechanisms for surviving oxygen lack. Proc. Natl Acad. Sci. USA, 93,9493±9498.
7. Guillemin,K. and Krasnow,M.A. (1997) The hypoxic response: huf®ngand HIFing. Cell, 89, 9±12.
8. King,D.P., Zhao,Y., Sangoram,A.M., Wilsbacher,L.D., Tanaka,M.,Antoch,M.P., Steeves,T.D., Vitaterna,M.H., Kornhauser,J.M.,Lowrey,P.L., et al. (1997) Positional cloning of the mouse circadianclock gene. Cell, 16, 641±653.
9. Stark,K.A., Yee,G.H., Roote,C.E., Williams,E.L., Zusman,S. andHynes,R.O. (1997) A novel alpha integrin subunit associates with betaPSand functions in tissue morphogenesis and movement during Drosophiladevelopment. Development, 124, 4583±4594.
10. Swanson,H.I. (2002) DNA binding and protein interactions of the AHR/ARNT heterodimer that facilitate gene activation. Chem. Biol. Interact.,141, 63±76.
11. Mimura,J. and Fujii-Kuriyama,Y. (2003) Functional role of AhR in theexpression of toxic effects by TCDD. Biochim. Biophys. Acta, 1619,263±268.
12. Ema,M., Taya,S., Yokotani,N., Sogawa,K., Matsuda,Y. and Fujii-Kuriyama,Y. (1997) Proc. Natl Acad. Sci. USA, 94, 4273±4278.
13. Ema,M., Morita,M., Ikawa,S., Tanaka,M., Matsuda,Y., Gotoh,O.,Saijoh,Y., Fujii,H., Hamada,H., Kikuchi,Y. et al. (1996) Two newmembers of the murine Sim gene family are transcriptional repressorsand show different expression patterns during mouse embryogenesis.Mol. Cell. Biol., 16, 5865±5875.
14. Wharton,K.A.,Jr, Franks,R.G., Kasai,Y. and Crews,S.T. (1994) Controlof CNS midline transcription by asymmetric E-box-like elements:similarity to xenobiotic responsive regulation. Development, 120, 3563±3569.
15. Zelzer,E., Wappner,P. and Shilo BZ. (1997) The PAS domain conferstarget gene speci®city of Drosophila bHLH/PAS proteins. Genes Dev.,11, 2079±2089.
16. Bacsi,S.G., Reisz-Porszasz,S. and Hankinson,O. (1995) Orientation ofthe heterodimeric aryl hydrocarbon (dioxin) receptor complex on itsasymmetric DNA recognition sequence. Mol. Pharmacol., 47, 432±438.
17. Sogawa,K., Nakano,R., Kobayashi,A., Kikuchi,Y., Ohe,N.,Matsushita,N. and Fujii-Kuriyama,Y. (1995) Possible function of Ahreceptor nuclear translocator (Arnt) homodimer in transcriptionalregulation. Proc. Natl Acad. Sci. USA, 92, 1936±1940.
18. Kikuchi,Y., Ohsawa,S., Mimura,J., Ema,M., Takasaki,C., Sogawa,K. andFujii-Kuriyama,Y. (2003) Heterodimers of bHLH±PAS proteinfragments derived from AhR, AhRR and Arnt prepared by co-expressionin Escherichia coli: characterization of their DNA binding activity andpreparation of a DNA complex. J. Biochem. (Tokyo), 134, 83±90.
19. Fujisawa-Sehara,A., Sogawa,K., Nishi,C. and Fujii-Kuriyama,Y. (1986)Regulatory DNA elements localized remotely upstream from the drug-metabolizing cytochrome P-450c gene. Nucleic Acids Res., 14, 1465±1477.
20. Maxam,A.M. and Gilbert,W. (1980) Sequencing end-labeled DNA withbase-speci®c chemical cleavages. Methods Enzymol., 65, 499±560.
21. Kim,J., Zwieb,C., Wu,C. and Adhya,S. (1989) Bending of DNA by gene-regulatory proteins: construction and use of a DNA bending vector.Gene, 85,15±23.
22. Semenza,G.L. and Wang,G.L. (1992) A nuclear factor induced byhypoxia via de novo protein synthesis binds to the human erythropoietingene enhancer at a site required for transcriptional activation. Mol. Cell.Biol., 12, 5447±5454.
23. Swanson,H.I., Chan,W.K. and Brad®eld,C.A. (1995) DNA bindingspeci®cities and pairing rules of the Ah receptor, ARNT and SIMproteins. J. Biol. Chem., 270, 26292±26302.
24. Shen,E.S. and Whitlock,J.P.,Jr (1992) Protein±DNA interactions at adioxin-responsive enhancer. Mutational analysis of the DNA-binding sitefor the liganded Ah receptor. J. Biol. Chem., 267, 6815±6819.
25. Kress,S., Reichert,J. and Schwarz,M. (1998) Functional analysis of thehuman cytochrome P4501A1 (CYP1A1) gene enhancer. Eur. J.Biochem., 258, 803±812.
3178 Nucleic Acids Research, 2004, Vol. 32, No. 10
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from

26. Camenisch,G., Stroka,D.M., Gassmann,M. and Wenger,R.H. (2001)Attenuation of HIF-1 DNA-binding activity limits hypoxia-inducibleendothelin-1 expression. P¯ugers Arch., 443, 240±249.
27. Baba,T., Mimura,J., Gradin,K., Kuroiwa,A., Watanabe,T., Matsuda,Y.,Inazawa,J., Sogawa,K. and Fujii-Kuriyama,Y. (2001) Structure andexpression of the Ah receptor repressor gene. J. Biol. Chem., 276, 33101±33110.
28. Nair,S.K. and Burley,S.K. (2003) X-ray structures of Myc-Max and Mad-Max recognizing DNA: molecular bases of regulation by proto-oncogenic transcription factors. Cell, 112, 193±205.
29. Ferre-D'Amare,A.R., Pognonec,P., Roeder,R.G. and Burley,S.K. (1994)Structure and function of the b/HLH/Z domain of USF. EMBO J., 13,180±189.
30. Ma,P.C., Rould,M.A., Weintraub,H. and Pabo,C.O. (1994) Crystalstructure of MyoD bHLH domain-DNA complex: perspectives on DNA
recognition and implications for transcriptional activation. Cell, 77, 451±459.
31. Shimizu,T., Toumoto,A., Ihara,K., Shimizu,M., Kyogoku,Y., Ogawa,N.,Oshima,Y. and Hakoshima,T. (1997) Crystal structure of PHO4 bHLHdomain±DNA complex: ¯anking base recognition. EMBO J., 16, 4689±4697.
32. Michel,G., Minet,E., Ernest,I., Roland,I., Durant,F., Remacle,J. andMichiels,C. (2000) A model for the complex between the hypoxia-inducible factor-1 (HIF-1) and its consensus DNA sequence. J. Biomol.Struct. Dyn., 18, 169±179.
33. Takahashi,Y., Suzuki,C. and Kamataki,T. (1998) Silencing of CYP1A1expression in rabbits by DNA methylation. Biochem. Biophys. Res.Commun., 247, 383±386.
Nucleic Acids Research, 2004, Vol. 32, No. 10 3179
by guest on Novem
ber 30, 2013http://nar.oxfordjournals.org/
Dow
nloaded from




















