
AGE, GROWTH AND SHELL MORPHOMETRICS OF THE TOP SHELL TECTUS DENTATUS (TROCHIDAE: PROSOBRANCHIA) ...
13
BLUE BIOTECHNOLOGY JOURNAL Volume 2, Issue 4, 2013 Table of Contents Bacterial Bioactive Compounds from Marine Resources 539 Hanan Abd-Elnaby Mass Culture of the Harpacticoid Copepod Tisbe biminiensis Fed Inert Diets 583 Aurelyanna C. B. Ribeiro and Lília P. Souza-Santos Conservation Measures and the Effect of Fluoride on an Important Common Bivalve Paphia textile gmelin, 1791 in Egyptian Waters 595 Neama A. Sallam, Ghada F. El-Said and Manal M. El-Sadaawy Age, Growth and Shell Morphometrics of the Top Shell Tectus dentatus (Trochidae: Prosobranchia) from Gemsha Bay, Red Sea 611 Fatma A. Abdel-Razek, Mostafa A. M. Mahmoud, Mohamed H. Yassien, Saad Z. Mohamed and Ali A. F. A. Gab-Alla New York
Transcript of AGE, GROWTH AND SHELL MORPHOMETRICS OF THE TOP SHELL TECTUS DENTATUS (TROCHIDAE: PROSOBRANCHIA) ...

BLUE BIOTECHNOLOGY JOURNAL
Volume 2, Issue 4, 2013
Table of Contents
Bacterial Bioactive Compounds from Marine Resources 539 Hanan Abd-Elnaby
Mass Culture of the Harpacticoid Copepod Tisbe biminiensis Fed Inert Diets 583 Aurelyanna C. B. Ribeiro and Lília P. Souza-Santos
Conservation Measures and the Effect of Fluoride on an Important Common Bivalve Paphia textile gmelin, 1791 in Egyptian Waters 595
Neama A. Sallam, Ghada F. El-Said and Manal M. El-Sadaawy
Age, Growth and Shell Morphometrics of the Top Shell Tectus dentatus (Trochidae: Prosobranchia) from Gemsha Bay, Red Sea 611
Fatma A. Abdel-Razek, Mostafa A. M. Mahmoud, Mohamed H. Yassien, Saad Z. Mohamed and Ali A. F. A. Gab-Alla
New York

Blue Biotechnology Journal
Volume 2, Issue 4
ISSN: 2163-3886
© Nova Science Publishers, Inc.
AGE, GROWTH AND SHELL MORPHOMETRICS
OF THE TOP SHELL TECTUS DENTATUS
(TROCHIDAE: PROSOBRANCHIA)
FROM GEMSHA BAY, RED SEA
Fatma A. Abdel-Razek1, Mostafa A. M. Mahmoud*1, Mohamed H. Yassien1, Saad Z. Mohamed2
and Ali A F. A Gab-Alla3
1National Institute of Oceanography and Fisheries, Invertebrates Aquaculture Lab. Egypt 2Suez Canal University, Faculty of Science, Marine Biology Department, Egypt
3Biological Sciences Department, Faculty of Applied Sciences, Umm Al-Qura
University, Makkah, Kingdom of Saudi Arabia
ABSTRACT
A total of 730 of the Top shell gastropod Tectus dentatus (Forskal 1775) from Gemsha Bay, Red Sea, were studied for shell morphometric, age, growth and mortality. The results showed that T. dentatus are dioecious. The sex ratio not varying significantly from 1:1 and no hermaphrodites or sex reverse had occurred. The decomposition of the composite distribution of pooled Tectus samples showed that it contains four age groups I, II, III and IV with mean lengths 31.11, 51.36, 68.58 and 77.50 mm, respectively. The Von Bertalanffy growth parameters were K= 0.31 year1, L = 101.53 mm. The growth performance index (Φ) was 3.51 and the exploitation rate (E) was 0.63.The values of total mortality coefficient (Z), natural mortality coefficient (M) and fishing mortality coefficient (F) were 0.84, 0.31 and 0.53 year1 respectively.
Keywords: Age, Growth, Morphometrics, Tectus dentatus
1. INTRODUCTION
T. dentatus is the largest and most important member of the family Trochidae in the Red
Sea. The meat is valued as sea food by the coastal population and tourism sector and the shell
is utilized for button manufacture, and is therefore of commercial importance (used in the
Corresponding author: Mostafa Ahmed Mohamed Mahmoud, National Institute of Oceanography and Fisheries, Red Sea Branch, Hurghada, Red Sea, Egypt. E-mail: [email protected]

612 Fatma A. Abdel-Razek, Mostafa A. M. Mahmoud, Mohamed H. Yassien et al.
manufacture of dental fillings and Candida, wood furniture inlays). It lives in shallow waters of coral reefs and also along rocks and coral patches in the Red Sea [1].
The natural distribution of Trochus, dependent as it is on the presence of reefs, occurs in the inter-tropical belt. The Trochus lives on coral reefs and more precisely on reef-flats formed by the debris of dead corals. Maximum density usually occurs on the great slabs of dead corals on which lie scattered chunks of corals covered with a fine film of small algae, diatoms and foraminifers. The large Trochus mainly inhabit the part of the reef that is exposed to wind; unless the topography of the bordering reef-flat is too uniform [2]. Their muscular foot enables them to cling firmly to the substrate and thus to resist any strong water turbulence while at the same time receiving optimum oxygenation. The depth range that is most suitable for this species is in the first ten meters from the surface [3], but Trochus may be found as deep as 24 m [4]. Gaha [5] found that there are two factors which may have considerable effect on the abundance of T. dentatus in Rabigh area on the Red Sea which are the natural conditions and the practices of fishing.
Studies on age and growth provide an insight into age class, structure of stock, changes in abundance of population, its relation to fishing, longevity and growth rate. Since growth is defined as an increase in body size, meat weight or volume, studying these could be the most appropriate method for measuring growth [6]. Methods of age determination in gastropods have not received much attention than those of other pelecypods since gastropods show fewer recognizable growth marks [7].
Relationships between various length and weight parameters are necessary in comparing growth parameters especially for commercially important species [8]. Length-weight relationships are used in the estimation of weight from length for individuals and for length classes [9] and conversion of growth-in-length equations to growth-in-weight for prediction of weight-at-age and subsequent use in stock assessment models [10]. Length-Weight relationship also allows comparisons of life history and morphology between species or between populations of species from different habitats or location [11].
Rajagopal [12] showed that the growth lines on the shells of temperate mollusks used to be the valuable pointers of age, but in tropical waters, on account of the lack of distinct seasons, variations in environmental parameters are limited and so, much difference in growth lines is not discernible.
There are many analytical tools that can be applied in assessing the exploitation level and stock status of marine fishery resources. Among them the FISAT (FAO-ICLARM Stock Assessment Tools) has been most frequently employed in estimating the population parameters [13-16], primarily because it requires only length-frequency data [17].
This study aims to investigate shell morphometrics, age, growth and mortalities of Tectus dentatus in Gemsha Bay, Red Sea and to provide an insight into age class, structure of stock, changes in abundance of population and its relation to fishing.
2. MATERIALS AND METHODS
Gemsha Bay is located in the Northern sector of the Red Sea (Figure 1), about 60 km north to Hurghada (27° 39` N: 33° 29` E). Gemsha Bay is one of the indentations dissecting the continuity of the Red Sea northwestern shoreline. It consists of two parts, Big Gemsha Bay and small Gemsha bay.

Age, Growth and Shell Morphometrics of the Top Shell Tectus dentatus … 613
Figure 1. Map of Gemsha Bay showing: A. Gemsha Bay as a part from the Red Sea, B. Big Gemsha Bay and C. Small Gemsha Bay.
A total of 730 of Tectus dentatus were collected (from May 2009 to May 2010) either by snorkeling or SCUBA diving, in depths ranging from 2 to 6 meters. The animals were preserved in 6% neutral formalin for subsequent biological investigation. Apart from these preserved samples (about 50 individual) were examined for the following measurements: Shell length (Sh.L.) and Maximum basal diameter (MBD) measured using vernier calipers, to the nearest 0.1 mm. The Maximum basal diameter was considered as an index of animal size.
The flesh parts were carefully removed from the shell, excess water was drained off and weighed then dried in an oven at 100ºC for 24 hours and then reweighed. The wet tissue weight, dry weight, hard shell (without tissues) weight and the total weight (hard shell and flesh) were measured using an analytical balance readout 0.01 g.
The sex was determined by eyes after removing the flesh parts. The gonad is white or creamy white in a male and green to dark green in a female with microscopic examination by smears of the gonads.
The data was examined for evidence of differential growth between the various size parameters measured by testing each pair of size variables y and x for their fit to the allometric equation y = Axb where A and b are constants.
The specimens of T. dentatus were analyzed using the FiSAT software [18] for applying ELEFAN-I (Electronic Length Frequency Analysis) to estimate the asymptotic length (L ) and growth coefficient (K) of the Von Bertalanffy Growth Function (VBGF). In order to identify the different modes (cohorts) and mean length of each cohort (age group) in the length-frequency distributions, the Bhattacharya‘s method [19], which was incorporated in the FISAT package, was applied [17].

614 Fatma A. Abdel-Razek, Mostafa A. M. Mahmoud, Mohamed H. Yassien et al.
The estimated L and K were used to calculate the growth performance index (Φ) using the equation of: Φ = 2 log10 L + log10K
The recruitment pattern was determined by backward projection on the length axis of the set of available length-frequency data, which determine the number of pulses per year and the relative strength of each pulse [17].
Total mortality coefficient (Z) of the natural population of T. dentatus from Gemsha Bay was determined by applying the linearized length-converted catch curve method of Pauly [10] through the FiSAT software package.
The natural mortalities coefficient (M) was estimated according to the procedure of Taylor (1960), who gives an expression for (M) as a function of 95% of the asymptotic length, A 0.95 , as :M = 2.996 / A 0.95
The fishing mortality coefficient (F) can also be obtained directly by subtracting the natural mortality from the total mortality rate, where; F = Z - M.
With the known values of F‘ and Z‘ the exploitation rate E‘ was calculated according to [20] where E = F/Z.
3. RESULTS
3.1. Sex Ratio
It is impossible to differentiate morphologically between both male and female T. dentatus because there is no secondary external sexual feature by which the sexes can be distinguished. It is therefore necessary to break the apex of the shell to reveal the gonad which, when mature, is a deep green color in the female and milky white in the male.
730 animals examined by eyes (microscopically in small one) from May 2009 to May 2010, the results showed that 50.41 % were males and 49.59 % were females as shown in Table (1). The departure from equal proportions of males and females was tested by a Chi-square (X2) test with one degree of freedom and found to be insignificant (X2 value was nearly equal to zero). This strongly indicates that the sex ratio in T. dentatus is even (1:1). No hermaphroditic individuals were found among the animals examined during the period of investigation.
3.2. Morphometric Relationships
A series of morphometric relationships were investigated by fitting regressions. Length-length, length-weight and weight-weight relationships have been determined for T. dentatus. The regression equations, coefficients of correlation and size ranges of different sexes are summarized in Table (2). The curves show linear model for length-length and weight-weight relationships and power models for length-weight relationships. The following were examined: MBD on shell length; MBD on all total weight; tissue weight; shell weight and dry weight; Shell length on all total weight; tissue weight; shell weight and dry weight; total weight on all tissue weight; shell weight; dry weight; tissue weight on both shell weight and dry weight and shell weight on tissue weight. The regression was determined separately.
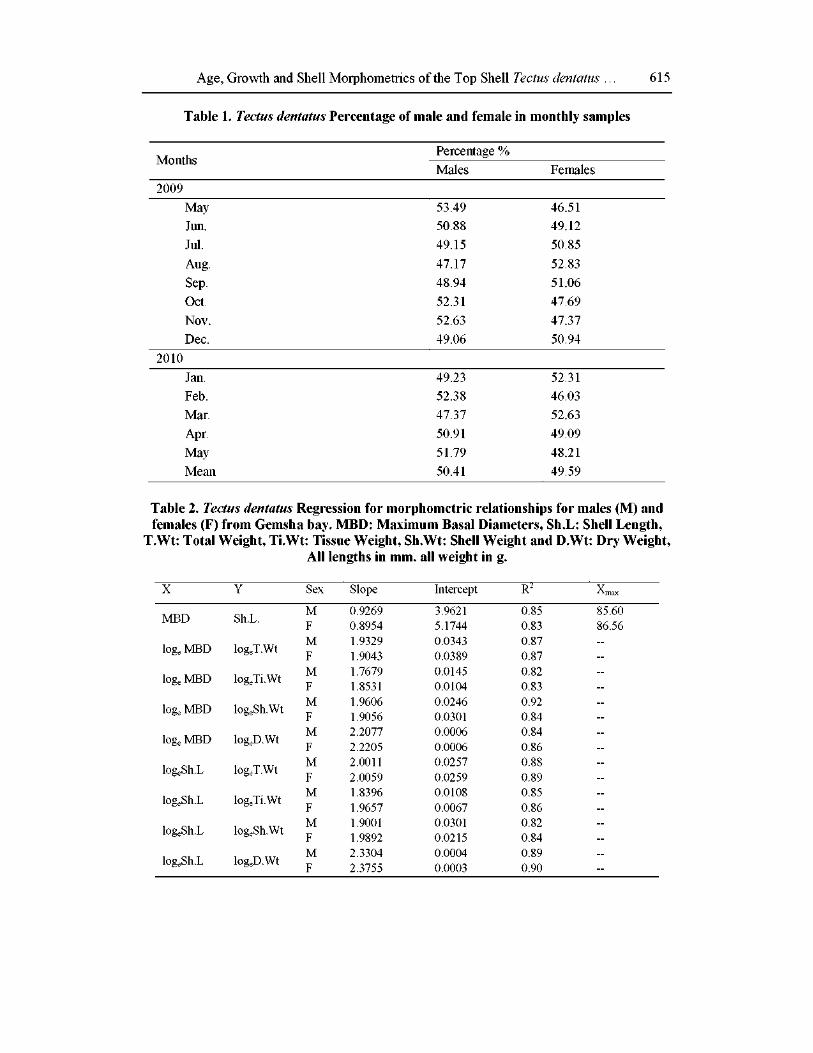
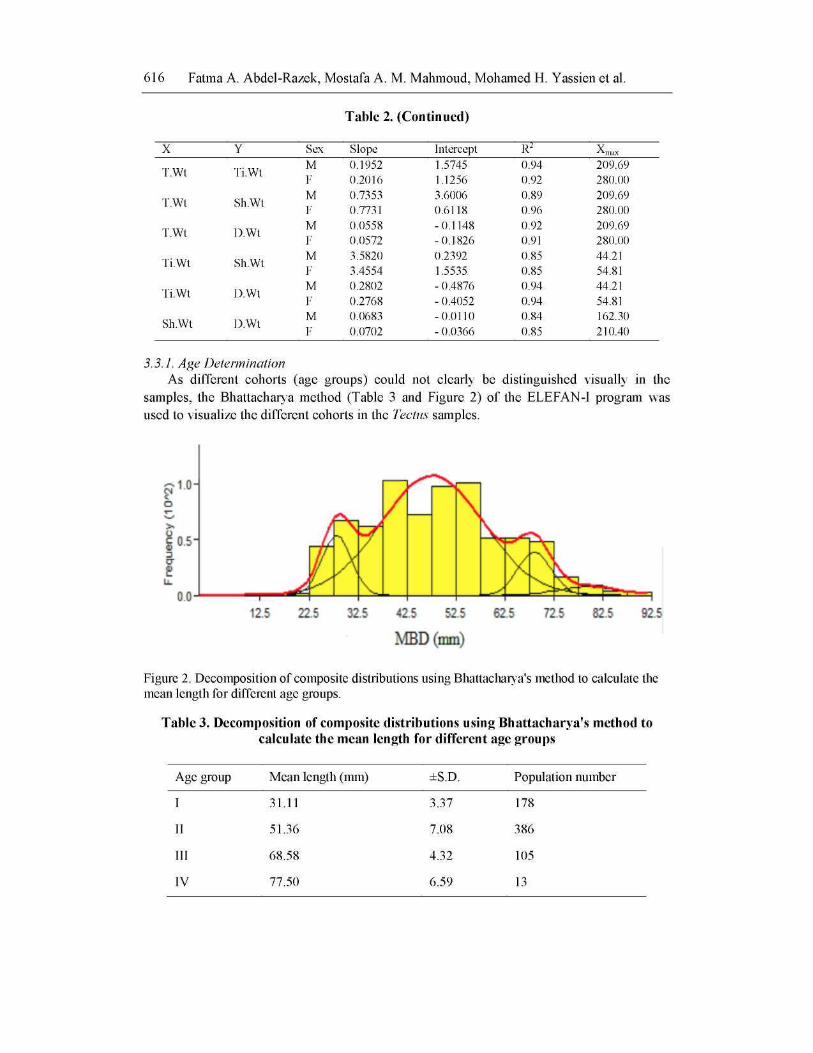
Age, Growth and Shell Morphometrics of the Top Shell Tectus dentatus … 617
Decomposition of the composite distribution of pooled Tectus samples showed that it
contains four age groups I, II, III and IV with mean lengths 31.11, 51.36, 68.58 and 77.50
mm, respectively. The population number of each age group revealed that age group (II)
represented the bulk of the animal population in Gemsha Bay and age group (IV) was the
least abundant one.
3.3.2. Estimation of Growth Parameters (Lx &K)
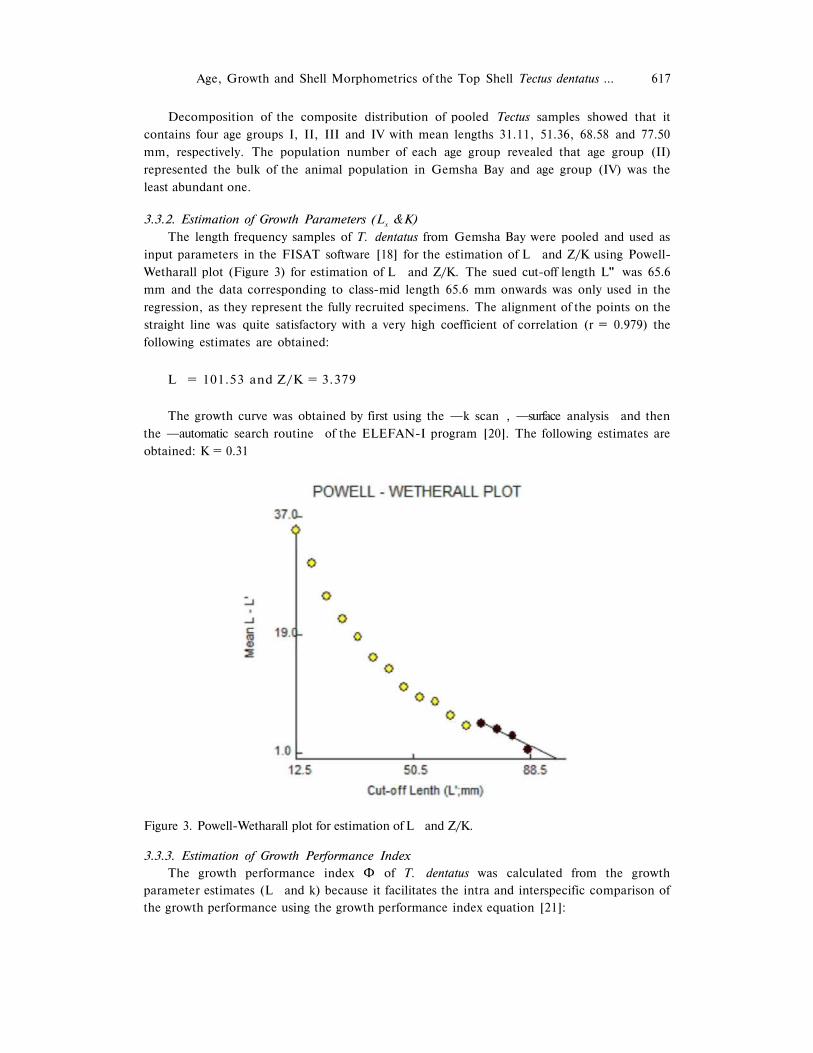
The length frequency samples of T. dentatus from Gemsha Bay were pooled and used as
input parameters in the FISAT software [18] for the estimation of L and Z/K using Powell-
Wetharall plot (Figure 3) for estimation of L and Z/K. The sued cut-off length L" was 65.6
mm and the data corresponding to class-mid length 65.6 mm onwards was only used in the
regression, as they represent the fully recruited specimens. The alignment of the points on the
straight line was quite satisfactory with a very high coefficient of correlation (r = 0.979) the
following estimates are obtained:
L = 101.53 and Z/K = 3.379
The growth curve was obtained by first using the ―k scan , ―surface analysis and then
the ―automatic search routine of the ELEFAN-I program [20]. The following estimates are
obtained: K = 0.31
Figure 3. Powell-Wetharall plot for estimation of L and Z/K.
3.3.3. Estimation of Growth Performance Index
The growth performance index Φ of T. dentatus was calculated from the growth
parameter estimates (L and k) because it facilitates the intra and interspecific comparison of
the growth performance using the growth performance index equation [21]:

618 Fatma A. Abdel-Razek, Mostafa A. M. Mahmoud, Mohamed H. Yassien et al.
Φ = log10 K + 2log10 L
While L was 101.53 and K was 0.31 so Φ = 3.51
3.3.4. Total Mortality Coefficient (Z) The total mortality coefficient (Z) of T. dentatus was estimated by applying the pooled
length-converted catch curve method [10], depending on the available length frequency data. The calculation was done with the FiSAT program.
The following estimates are obtained: Z = 0.84.
3.3.5. Natural Mortality Coefficient (M) The natural mortalities coefficient (M) was estimated according to the procedure of
Taylor (1960), who gives an expression for (M) as a function of 95% of the asymptotic length, A 0.95, as:
From the equation: M= 2.996 / A 0.95
While L was 101.53 so M = 0.311
3.3.6. Fishing mortality coefficient (F)
M value was subtracted from the Z- value in order to get the fishing mortality (F=Z-M).
So, from the equation: F=Z-M,
While Z was 0.84 and M was 0.311 then F = 0.529
3.3.7. Exploitation Rate (E) With the known values of F and Z the exploitation rate (E) was calculated according to
Sparre and Venema [20]:
From the equation: E = F/Z
While F was 0.529 and Z was 0.84 then E = 0.63
4. DISCUSSION
Age and growth information is extremely important in almost every aspect of fisheries aquaculture [8, 22]. Age refers to the quantitative description of length of time that an organism has lived, while growth is simply defined by von Bertalanffy [23] as the measurable increase in body weight or length. Growth provides an integrated assessment of the environmental (e.g., temperature) and endogenous conditions (e.g., genetic factors), and a useful metric to evaluate habitat suitability, prey availability or the influence of management activities on target species [22]. Growth occurs when a part of assimilated food or energy is retained and incorporated into the biomass of the organism [8, 24]. As a result, in gastropod mollusks, they increase their shell sizes and weights. Shell growth could either be

Age, Growth and Shell Morphometrics of the Top Shell Tectus dentatus … 619
indeterminate, continues throughout the life span or determinate, it may stop at some stage, usually when the gastropod reaches sexual maturity [8, 25].
A linear relationship exists in the T. dentatus population between MBD and shell length either in males or females. The slopes are 0.9269 and 0.8954 for male and female respectively. R2 is 0.8534 for male and 0.8314 for female.
The length-weight relationships showed that the pattern of growth (allometric) was verified from the value of b‘ in the equation, mean b values were 1.9329 for male and 1.9043 for female (negatively allometric) which means that the weight increases faster than the length. Regression equations derived from the logarithmically transformed data of shell length and weight were Log TWt= Log 0.343+1.9329 Log MBD and Log TWt= Log 0.0389+1.9043 Log MBD with a high correlation, R2 was 0.8672 and 0.8653 for male and female, respectively.
There was a negative allometric growth also between MBD and tissue weight, shell weight and dry weight. The increase in total weight is mostly associated to shell weight increase. Shell weight considers the heavier part in the animal and forms more than 78 % from total weight.
From the present study, it is obvious that there is further reflection in the higher shell/Tissue ratios in snails from the high shore, which show a general increase with increasing size suggesting that, as the Tectus grow, their shells become progressively heavier in relation to their tissue. There was a positive allometric growth between tissue weight and shell weight, b values were 3.5820 and 3.4554, and R2 was 0.8542 and 0.8511 for male and female, respectively.
Information regarding the biology, ecology and fishery of the species is very limited and currently has no regulation concerning the population structure of T. dentates in the Red Sea. Since, it has long been exploited; there is an urgent need to assess the status of the population stock, which is crucial for fishery management and sustainable utilization of the resource. Knowledge of various population parameters such as asymptotic length (L ), growth coefficient (K), mortality (total, natural and fishing) and exploitation rates (E) are therefore extremely important and urgently needed.
The software of FiSAT had identified four different cohorts in the wild population of T. dentatus in Gemsha and the mean lengths of these cohorts were 31.11, 51.36, 68.58 and 77.50 mm (MBD), respectively. The growth increments of these cohorts were: 20.25, 17.22 and 8.92 mm, respectively. It can be seen that the growth rate was rapid at the first two cohorts then decreased in the third cohorts. The rate of growth slows down as ageing proceeds and this is the normal growth rate [26].
The parameters of the Von Bertalanffy growth formula (VBGF) of T. dentatus from Gemsha using Powell-Wetharall plot for estimation of L and Z/K, through the FiSAT software package, was L = 101.53 mm (MBD), Z/K = 3.379, the used cut-off length L = 65.6 mm. It may be speculated that the growth rates obtained in this study are low (K = 0.31).
The values of estimated growth parameters (K and L ) of T dentatus from Gemsha indicate the longevity and slow growth rate of this species. Similar results were obtained by Bour and Grandperrin [27] as they estimated the following von Bertalanffy parameters on T.niloticus at Vanuatu: k = 0.32, L = 16.3. In Green Island on the Great Barrier Reef, Nash [28] estimated the following Von Bertalanffy parameters: (1) k = 0.25, L = 15.0, (slow); (2) k =0.49; L = 14.3 (fast) for the same species.

620 Fatma A. Abdel-Razek, Mostafa A. M. Mahmoud, Mohamed H. Yassien et al.
Growth performance index (Φ) estimated for T. dentatus can be usefully compared via
the growth performance index of Pauly [10]. The values of Φ quantifies the energy of a given
habitat where Φ is directly related to the growth and hence the metabolism and food
consumption by the animal.
When the estimated growth performance index (Φ = 3.51) was compared to the range of
values compiled [29] for marine gastropods of all latitudes (Φ = 2.2 - 4.7) and the tropics
(Φ = 3.6 - 4.7), it was found that the growth performance of T. dentatus lies before the low
end of the range observed for most other tropical marine gastropods (Φ = 3.6 - 4.7), but more
than that of the West Indian Topshell Cittarium pica (Φ = 3.31-3.48) [29].
In the present study, the value of total mortality (Z) was 0.84 year1. The fishing mortality
rate (F) was 0.53 year 1 and the natural mortality rate (M) was 0.31 year1. The fishing
mortality rate was higher than the natural mortality rates, thus indicating an unbalanced
position of the stock. Similar cases were obtained by Cob et al. [30] for Strombuscanarium in
the Johor Straits, Malaysia.
Schmidt et al. [29] studied the population structure of the West Indian top shell Cittarium
pica in the Caribbean Coast of Costa Rica at an unexploited and two exploited sites to
determine the present impact of the fishery on the resource. Length-frequency histograms
showed a strong shift towards smaller specimens at the exploited sites, which is also reflected
in significantly higher rates of total mortality (Z = 4.05 and 4.47) when compared to the
unexploited site (Z = 1.47).
The natural mortality rate (M) is usually difficult to measure and appears to vary a great
deal within species even over a small geographical range. Some work on Tasmanian abalone
has revealed rates of Maround 1.0 at some sites [31]. Cittarium pica, in the Caribbean Coast
of Costa Rica, has revealed rates of M from 1.34 to 1.60 [29] that lie in the range of values
(1.0-1.7) for marine gastropods given by several authors [32].
Other works done on marine gastropods showed that, many published natural mortality
rates (M) lie in the range 0.1 to 0.3 [28, 33- 36].
In this study, the natural mortality rate (M) was 0.31 per year for the population of T.
dentatus from Gemsha using the procedure of Taylor [37] which is in agreement with 0.1 to
0.3 supported by the above mentioned Scientists.
The exploitation rate (E) (= 0.63) was higher than the maximum allowable limit of
exploitation (>0.5) [29, 30] which was a further indication of overexploitation. For
sustainable utilization of the resource, serious efforts should immediately be taken in reducing
the exploitation rates of T. dentatus in the study area.
REFERENCES
[1] A. M. Eisawy. The spawning and development of Trochus (Tectus) dentatus Forskal.
Bull. Inst. Ocean. Fish, Cairo, 1 (1970) 379-394.
[2] B. D. Smith. Growth rate, abundance and distribution of the top shell Trochus niloticus
on Guam. PhD thesis, Univ. of Guam. (1979) 240p.
[3] R. Gail. Trochus fishing. SPC Q. Bull., 7(1) (1957) 48-49.
[4] J. A. McGowan. Current status of the trochus fishery in Micronesia. Rep. to the High
Commissioner, U.S. Trust Territory of the Pacific Islands, Saipan. (1956) 23 pp.

Age, Growth and Shell Morphometrics of the Top Shell Tectus dentatus … 621
[5] M. S. A. Gaha. Study of Tectus dentatus in Rabigh area on the Red Sea. M.Sc. Thesis, Faculty Fac. of Marine Science King Abdulaziz Univ. (2008).
[6] B. L. Bayne. In: Physiological ecology of estuarine organisms, (Eds.) F.J. Vernberg University of South Carolina press, Columbia, South Carolina. (1975) pp: 25.
[7] M. H. Chen and K. Soong. Estimation of age in the sex-changing, coral-inhabiting snail Coralliophila violacea from the growth striae on opercula and a mark-recapture experiment. Mar. Biol.140 (2002) 337-342.
[8] A. S. Ilano, A. Ito, K. Fujinaga and S. Nakao. Age determination of Buccinum isaotakii (Gastropoda: Buccinidae) from the growth striae on operculum and growth under laboratory conditions. Aquaculture, 242 (2004) 181-195.
[9] R. Anderson and S. Gutreuter. Length, weight, and associated structural indices. In: Nielsen, L., Johnson, D. (eds.), Fisheries Techniques. Am. Fish. Soc., (1983) 283–300.
[10] D. Pauly. Some simple methods for the assessment package of tropical fish stocks. FAO Fisheries Technical Paper No. 234 (1983) 52.
[11] M.B. Gaspar, M.N. Santos and P. Vasconcelos. Weight-length relationships of 25 bivalve species (mollusca: bivalvia) from the Algarve coast (southern Portugal). J. Mar. Biol. Assoc. U.K., 81 (2001) 805-807.
[12] S. Rajagopal. Studies on the intertidal mollusk Umboniumvestiarium (Archeogastropoda. Trochidae: Umbonidae) of the south-east coast of India, Ph.D. Thesis, Annamalai Univ., India, pp: (1982) 85.
[13] E. Mancera and J. Mendo. Population dynamics of the oyster Crassostrea rhizophorae from the Cienaga Grande de Santa Marta, Colombia. Fish. Res., 26 (1996) 139-148.
[14] S. J. M. Blaber, J. Staunton-Smith, D.A. Milton, G. Fry and T.V. Veldev. The biology and life history strategies of Ilisha (Teleostei: Pristigasteridae) in the coastal waters and estuaries of Sarawak. Estuar.Coast. Shelf Sci., 47 (1998) 499-511.
[15] S. M. Al-Barwani, A. Arshad, S.M.N. Amin, S.B. Japar, S.S. Siraj and C.K. Yap. Population dynamics of the green mussel Pernaviridis from the high spat-fall coastal water of Malacca, Peninsular Malaysia. Fish. Res., 84 (2007) 147-152.
[16] S. M. Amin, M. Zafar, and A. Halim. Age, growth, mortality and population structure of the oyster, Crassostrea madrasensis, in the Moheskhali Channel (Southeastern coast of Bangladesh). J. Applied Ichth., 24 (2008) 18-25.
[17] F. C. Gayanilo and D. Pauly. FAO-ICLARM stock assessment tools (FiSAT) Reference manual. FAO Computerized Information Series (Fisheries) No. 8. FAO, Rome (1997) 262.
[18] F. C. Gayanilo, P. Sparre and D. Pauly. The FiSAT user's guide.FAO computerized information series fisheries 99, ICLARM, DIFMAR, Rome (1993).
[19] C. G. Bhattacharya. A simple method of resolution of a distribution into Gaussian components. Biometrics, 23 (1967) 115-135.
[20] P. Sparre and S.C. Venema. Introduction to tropical fish stock assessment, Manual. FAO Fisheries Technical Paper No. 306/1. (1998) 407.
[21] D. Pauly and J.L. Munro. Once more on the comparison of growth in fish and invertebrate. ICLARM Fishbyte, 2 (1984) 21-33.
[22] R. D. Devries and R. V. Frie. Determination of age and growth. In: Murphy, B.R., Willis, D.W. (eds.), Fisheries Techniques. American Fisheries Society, (1996) 483-512.
[23] L. Von. Bertalanffy. A quantitative theory of organic growth (inquiries on growth laws. II). Human Biol., 10 (1938) 181-213.

622 Fatma A. Abdel-Razek, Mostafa A. M. Mahmoud, Mohamed H. Yassien et al.
[24] A. E. Kideys and R.G. Hartnoll. Energetics of mucus production in the common whelk Buccinum undatum L. J. Exp. Mar. Ecol., 150 (1991) 91-105.
[25] R. N. Hughes. A functional biology of marine gastropods Croom helm. London, (1986) 245.
[26] D. J. Tranter. Reproduction in Australian pearl oysters (lamellibranchia). III Pinctada margaritifera (L). Aust. J. Mar. Freshwat. Res., 9(4) (1958) 509-523.
[27] W. Bour and R. Grandperrin. Croissance des trocas a Vanuatu. Notes Doc. d'Oceanogr. ORSTOM.Port Vila.14: 31 pp. In: Foale S and Day R. 1997 Stock assessment of Trochus (Trochusniloticus) (Gastropoda: Trochidae) fisheries at West Nggela, Solomon Islands. Fish. Res., 33 (1985) 1-16.
[28] W. J. Nash. An evaluation of egg-per-recruit analysis as a means of assessing size limits for blacklip abalone (Haliotisrubra) in Tasmania. In Abalone of the World: Biology, Fisheries and Culture (S. Shepherd, M. Tegner and S. Guzman delProo, eds.). (1992) 318-338. Oxford: Blackwell Scientific Publications.
[29] S. Schmidt, M. Wolff and J.A. Vargas. Population ecology and fishery of Cittarium pica (Gastropoda: Trochidae) on the Caribbean coast of Costa Rica, Rev. Biol. Trop., 50(3/4) (2002) 1079-1090.
[30] Z. C. Cob, A. Arshad, J.S. Bujang and M.A. Ghaffar. Age, growth, mortality and population structure of Strombu scanarium (Gastropoda: Strombidae): Variations in male and female sub-populations. J. Applied. Sci., 9 (2009) 3287-3297.
[31] S. Foale and R. Day, Stock assessment of trochus (Trochus niloticus) (Gastropoda: Trochidae) fisheries at West Nggela, Solomon Islands. Fish. Res., 33 (1997) 1-16.
[32] A.O. Debrot. Survival, growth, and fecundity of the West Indian Topshell, Cittarium pica (Linnaeus), in various rocky intertidal habitats of the Exuma Cays, Bahamas. Veliger, 33 (1990) 363-371.
[33] P. R. Sluczanowski. A management oriented model of an abalone fishery whose substocks are subject to pulse fishing. Can. J. Fish.Aquat. Sci., 41 (1984) 1008-1014.
[34] M. J. Tegner, P.A. Breen and C.E. Lennert. Population biology of red abalones, Haliotis rufescens, in southern California and management of the red and pink, H. corrugata, abalone fisheries. Fish. Bull., 87 (1989) 313-339.
[35] S. A. Shepherd, S.A. Guzman del Proo, J. Turrubiates and J. Belmar. Growth, size at sexual maturity, and egg-per-recruit analysis of the abalone Haliotis fulgens in Baja California. Veliger, 34 (1991) 324-330.
[36] P. E. McShane and J.R. Naylor. Depth can affect post-settlement survival of Haliotis iris (Mollusca: Gastropoda). J Exp. Mar. Biol. Ecol., 187 (1995) 1-12.
[37] C. C. Taylor. Temperature, growth and mortality. The Pacific cockle. J. Cons. Cons. Int. Explor. Mer., 26 (1960) 117-124.
Received 4 November 2013; accepted in final form 22 December 2013.




















